
CHARGED DILATONIC BLACK HOLES: STRING FRAME VERSUS EINSTEIN FRAME
Upload
khangminh22Category
view
1download
0
The Open Reading Frame VI Product of Cauliflower mosaicvirus Is a Nucleocytoplasmic Protein: Its N TerminusMediates Its Nuclear Export and Formation ofElectron-Dense Viroplasms W
Muriel Haas,a,1 Angele Geldreich,a Marina Bureau,a Laurence Dupuis,a Veronique Leh,a,2 Guillaume Vetter,a
Kappei Kobayashi,b Thomas Hohn,b Lyubov Ryabova,a Pierre Yot,a and Mario Kellera,3
a Institut de Biologie Moleculaire des Plantes, Unite Propre de Recherche, Centre National de la Recherche
Scientifique 2357, Universite Louis Pasteur, 67084 Strasbourg Cedex, Franceb Friedrich Miescher Institute, CH-4002 Basel, Switzerland
The Cauliflower mosaic virus (CaMV) open reading frame VI product (P6) is essential for the viral infection cycle. It controls
translation reinitiation of the viral polycistronic RNAs and forms cytoplasmic inclusion bodies (viroplasms) where virus
replication and assembly occur. In this study, the mechanism involved in viroplasm formation was investigated by in vitro
and in vivo experiments. Far protein gel blot assays using a collection of P6 deletion mutants demonstrated that the
N-terminal a-helix of P6 mediates interaction between P6 molecules. Transient expression in tobacco (Nicotiana tabacum)
BY-2 cells of full-length P6 and P6 mutants fused to enhanced green fluorescent protein revealed that viroplasms are formed
at the periphery of the nucleus and that the N-terminal domain of P6 is an important determinant in this process. Finally, this
study led to the unexpected finding that P6 is a nucleocytoplasmic shuttle protein and that its nuclear export is mediated by
a Leu-rich sequence that is part of the a-helix domain implicated in viroplasm formation. The discovery that P6 can localize
to the nucleus opens new prospects for understanding yet unknown roles of this viral protein in the course of the CaMV
infection cycle.
INTRODUCTION
Cauliflower mosaic virus (CaMV) is the type member of the
Caulimovirus genus of the Caulimoviridae family. Its double-
stranded circular DNA (;8 kb) is replicated via an RNA in-
termediate by a virus-encoded reverse transcriptase, and the
virus is classified in the pararetrovirus supergroup, to which
animal viruses of the Hepadnaviridae family also belong (for
a review, see Haas et al., 2002). The CaMV genome possesses
six major open reading frames (ORFs I to VI), which are all located
on the same DNA strand. Many functions of the corresponding
gene products P1 to P6 have been elucidated, but the mecha-
nisms by which they operate during viral infection are not yet fully
understood.
The viral DNA is transcribed by the cellular RNA polymerase II
into two major capped and polyadenylated RNAs, a monocis-
tronic 19S mRNA and a pregenomic 35S RNA that serves as
a template both for reverse transcription and for translation into
P1 to P5. The 35S RNA undergoes alternative splicing events
leading to four mRNAs in which ORF I and part of ORF II are
deleted (Kiss-Laszlo et al., 1995). Currently, the mechanism
regulating the nuclear export of 35S RNA and its spliced forms is
unknown. These RNAs are translated by the cellular machinery
following two unconventional strategies, ribosomal shunt and
termination–reinitiation (for a review, see Ryabova et al., 2002).
The CaMV P6 protein (62 kD), which is expressed specifically
from the 19S RNA, is a multifunctional protein that represents
a key component in the CaMV infectious cycle. P6 is the major
determinant of host specificity and influences symptom severity
(Daubert et al., 1984; Daubert and Routh, 1990; Agama et al.,
2002). Inoculation of cruciferous and solanaceous plant species
with chimeric CaMV genomes bearing ORF VI derived from
different CaMV isolates showed that the N-terminal region of P6
is responsible for host specificity (Daubert et al., 1984; Schoelz
et al., 1986). Transgenic Arabidopsis thaliana plants expressing
P6 display disease symptoms whose severity is related to the
expression level of the transgene (Zijlstra et al., 1996). Compar-
ison of the cellular mRNA content of ORF VI-transgenic and
control Arabidopsis plants revealed that ORF VI expression
downregulates or upregulates several host genes (Geri et al.,
1 Current address: Unite Propre de Recherche, Centre National de laRecherche Scientifique 9050, Departement Recepteurs et ProteinesMembranaires, Ecole Superieure de Biotechnologie de Strasbourg,boulevard Sebastient Brant, 67412 Illkirch Cedex, France.2 Current address: Laboratoire de Dynamique, Evolution et Expressiondes Genomes de Microorganismes, Formation de Recherche enEvolution 2326, Centre National de la Recherche Scientifique, UniversiteLouis Pasteur, 28 rue Goethe, 67084 Strasbourg Cedex, France.3 To whom correspondence should be addressed. E-mail [email protected]; fax 33-3-88-61-44-42.The author responsible for distribution of materials integral to thefindings presented in this article in accordance with the policy describedin the Instructions for Authors (www.plantcell.org) is: Mario Keller([email protected]).W Online version contains Web-only data.Article, publication date, and citation information can be found atwww.plantcell.org/cgi/doi/10.1105/tpc.104.029017.
The Plant Cell, Vol. 17, 927–943, March 2005, www.plantcell.org ª 2005 American Society of Plant Biologists
Dow
nloaded from https://academ
ic.oup.com/plcell/article/17/3/927/6114488 by guest on 07 M
arch 2022

1999). Whether P6 plays a direct role in regulating expression of
these cellular genes (i.e., by regulating their transcription) has not
been determined. Finally, P6 from certain CaMV isolates can also
act as an avirulence gene product to promote a hypersensitive
response in some Nicotiana species (Palanichelvam et al., 2000;
Cole et al., 2001).
P6 trans-activates translation of the viral polycistronic 35S RNA
and its spliced versions and hence allows synthesis of a complete
set of viral proteins (for a review, see Ryabova et al., 2002). Park
et al. (2001) demonstrated that P6 is a translational reinitiation
factor that associates with the host translational machinery
and thus permits translation of downstream ORFs. This func-
tion is mediated by physical interactions between an internal
region, including the minimal sequence of P6 required for trans-
activation (the mini-TAV; De Tapia et al., 1993), the initiation factor
eIF3 (Park et al., 2001), and the ribosomal proteins L13 (Bureau
et al., 2004), L18 (Leh et al., 2000), and L24 (Park et al., 2001). The
observed interaction between P6 and the CaMV capsid protein
(P4) also suggests a role for P6 as a scaffolding protein in the
assembly of CaMV particles (Himmelbach et al., 1996).
P6 is an abundantly synthesized CaMV protein, and in the
cytoplasm of infected cells it forms amorphous, non-membrane-
limited, electron-dense inclusion bodies, also referred to as
viroplasms. These electron-dense viroplasms are distinct from
the electron-lucent inclusion bodies that are mainly composed of
CaMV P2 protein (Espinoza et al., 1991; Drucker et al., 2002). The
number and the size (2 to 10 mm in diameter) of the electron-
dense viroplasms depend on the stage of the viral cycle but also
on the CaMV isolate and the host plant (Shalla et al., 1980).
Electron-dense viroplasms are a hallmark of infection of plant
cells by caulimoviruses and soymoviruses. They probably play
an important role in the infectious cycle because they are the
sites of protein synthesis, viral genome replication, morphogen-
esis, and storage of the newly formed virions (Mazzolini et al.,
1989; Rothnie et al., 1994). Other CaMV proteins have been
detected in the electron-dense viroplasms (Drucker et al., 2002),
but none of them seems to be required for their formation
because transgenic Arabidopsis plants expressing P6 alone
contain inclusion body-like structures (Cecchini et al., 1997).
Previous data from several studies suggested that P6 self-
associates (Leh, 1999; Haas et al., 2000), and Li and Leisner
(2002) showed that multiple domains within P6 can interact with
the full-length protein; they proposed that these interactions
might be involved in the formation of viroplasms.
In this article, we demonstrate for the first time that P6 localizes
both in the cytoplasm and in the nucleus of plant cells and that
the N-terminal region of P6 has a dual function. It is a major
determinant for in vitro interaction between P6 molecules and
mediates the formation of viroplasms in vivo. It also contains
a Leu-rich nuclear export signal that prevents accumulation of
P6 molecules within the nucleus.
RESULTS
The N-Terminal Region Mediates P6–P6 Interactions in Vitro
In previously described far protein gel blot experiments (Leh,
1999), proteins from healthy and CaMV-infected turnip (Brassica
rapa) or Arabidopsis plants were fractionated by SDS-PAGE and
transferred to nitrocellulose. Using P6 expressed in Escherichia
coli and 32P-radiolabeled in vitro as overlay, a radioactive signal
was detected with proteins from infected plants at the level of
a polypeptide of 62 kD that also reacted with anti-P6 antibodies,
strongly suggesting that P6 interacts with itself. However,
because CaMV-P6 protein downregulates or upregulates the
expression of several host protein genes (Geri et al., 1999), it
could not be totally excluded that the blot-immobilized species
interacting with 32P-P6 in the above experiment was a host
protein of similar mobility to P6 that was expressed upon viral
infection. To rule out this possibility, we have performed an
identical far protein gel blot experiment except that the immo-
bilized proteins on the blot were obtained from an extract of
E. coli expressing P6. The 32P-P6 in the overlay again reacted
with a 62-kD species (Figure 1C, lane P6), which was also recog-
nized by anti-P6 antibodies (Figure 1B, lane P6), providing inde-
pendent confirmation that P6 can interact with itself. A similar
result was obtained using a pull-down assay (data not shown).
Because P6 contains several domains that can bind single-
and/or double-stranded RNA and RNA-DNA heteroduplexes (De
Tapia et al., 1993; Cerritelli et al., 1998), far protein gel blot assays
were also performed after treatment of both the overlay and
the membrane-bound proteins with RNase and DNase. These
treatments did not impair formation of the P6-P6 complex, de-
monstrating that neither RNA nor DNA mediates the P6–P6
interaction and consequently that one or more domains of P6
are directly involved.
To characterize the region(s) required for self-association of
P6, we tested the capacity of a series of P6 deletion mutants
(Figure 1A) to bind full-length P6. The mutants corresponded to
N- and C-terminal recurrent deletions and to P6 bearing internal
deletions of previously identified functional domains (i.e., the
mini-TAV and RNA binding sites) (De Tapia et al., 1993). The
deleted proteins were expressed in E. coli from pET3a recombi-
nant plasmids, separated by SDS-PAGE, blotted onto a nitrocel-
lulose membrane, and incubated in the presence of 32P-labeled
P6 in the overlay solution. As shown in Figure 1C, P6 interacted
with the C-terminal deletion mutants A, B, C, D, and E and with
the internally deleted mutants J, K, L, and M, but not with the
N-terminal deletion mutants F, G, H, and I. Because no radio-
active signals could be detected with the latter mutants even
though they were present in relatively high amounts on the mem-
brane (Figure 1B, lanes F to I), we can rule out the possibility
that the signals observed with the other P6 mutants correspond
to artifactual binding as a result of gross overloading of the
membrane with proteins. The minor signals observed in lanes J
to M (Figure 1C) correspond to interactions between the overlay
and P6 degradation products rather than to nonspecific inter-
actions with bacteria proteins as demonstrated by the control
experiment performed with bacteria transformed with empty
vector (Figure 1C, lane E. coli).
The smallest mutant able to interact with P6 was mutant A,
which corresponds to the 112 N-terminal amino acids of P6,
a region we will refer to as domain A. The latter was fused to the
N terminus of a protein from an unrelated virus, P42 of Beet
necrotic yellow vein virus, or to the C terminus of glutathione
S-transferase (GST) to analyze the ability of domain A to interact
928 The Plant Cell
Dow
nloaded from https://academ
ic.oup.com/plcell/article/17/3/927/6114488 by guest on 07 M
arch 2022
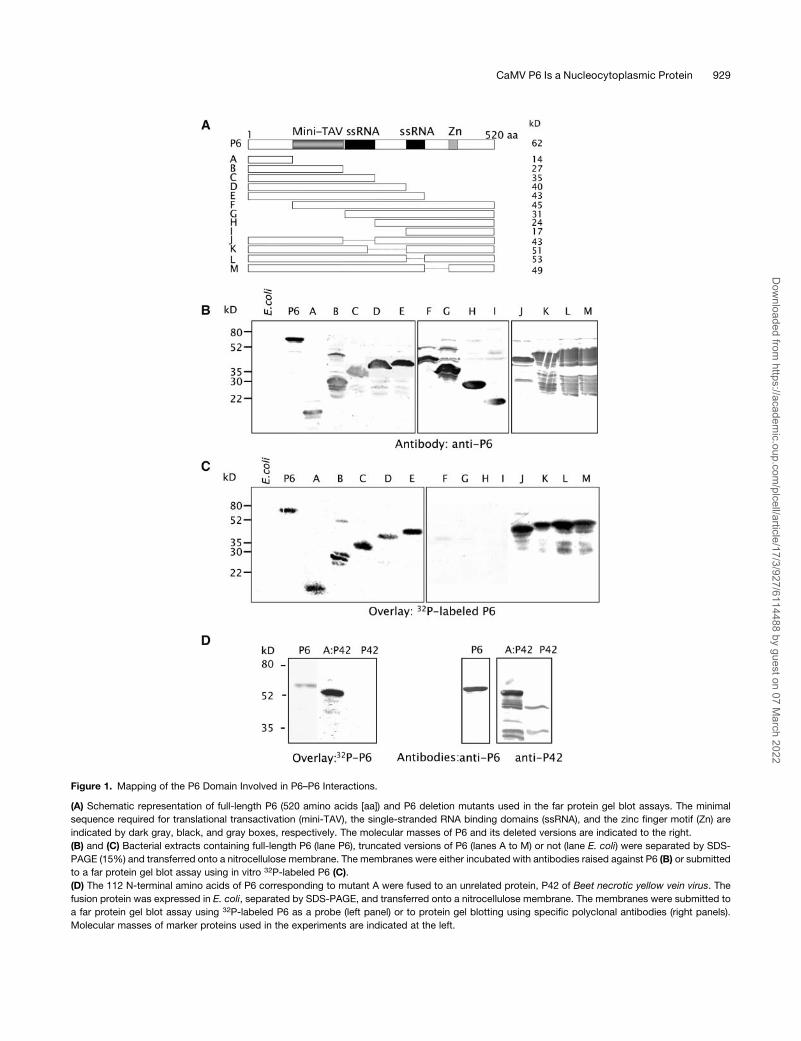
Figure 1. Mapping of the P6 Domain Involved in P6–P6 Interactions.
(A) Schematic representation of full-length P6 (520 amino acids [aa]) and P6 deletion mutants used in the far protein gel blot assays. The minimal
sequence required for translational transactivation (mini-TAV), the single-stranded RNA binding domains (ssRNA), and the zinc finger motif (Zn) are
indicated by dark gray, black, and gray boxes, respectively. The molecular masses of P6 and its deleted versions are indicated to the right.
(B) and (C) Bacterial extracts containing full-length P6 (lane P6), truncated versions of P6 (lanes A to M) or not (lane E. coli) were separated by SDS-
PAGE (15%) and transferred onto a nitrocellulose membrane. The membranes were either incubated with antibodies raised against P6 (B) or submitted
to a far protein gel blot assay using in vitro 32P-labeled P6 (C).
(D) The 112 N-terminal amino acids of P6 corresponding to mutant A were fused to an unrelated protein, P42 of Beet necrotic yellow vein virus. The
fusion protein was expressed in E. coli, separated by SDS-PAGE, and transferred onto a nitrocellulose membrane. The membranes were submitted to
a far protein gel blot assay using 32P-labeled P6 as a probe (left panel) or to protein gel blotting using specific polyclonal antibodies (right panels).
Molecular masses of marker proteins used in the experiments are indicated at the left.
CaMV P6 Is a Nucleocytoplasmic Protein 929
Dow
nloaded from https://academ
ic.oup.com/plcell/article/17/3/927/6114488 by guest on 07 M
arch 2022

with P6 when placed in an unrelated sequence context. P6
bound to A:P42 but not to P42 alone (Figure 1D) and to GST:A but
not to GST (see below), thus further demonstrating that domain A
can act independently of the rest of the amino acid sequence in
mediating P6 self-association in vitro. Taken together, the results
of the above far protein gel blot experiments provide evidence
that the N-terminal region encompasses the domain required for
P6 self-interaction in vitro.
The N Terminus of P6 Is an Essential Determinant for
Both the Formation of Viroplasms and Their
Localization in the Cytoplasm
To obtain further information about the importance of the
N-terminal region of P6 in the formation of inclusion bodies,
tobacco (Nicotiana tabacum) BY-2 cells were transfected with
recombinant pCK-EGFP plasmids coding for full-length P6 and
two deleted versions (A and P6DA) fused to the C terminus of
the enhanced green fluorescent protein (EGFP). The results de-
scribed below are representative of at least four independent
transfection experiments and observation by confocal laser
scanning microscopy (CLSM) at 20 h post-transfection.
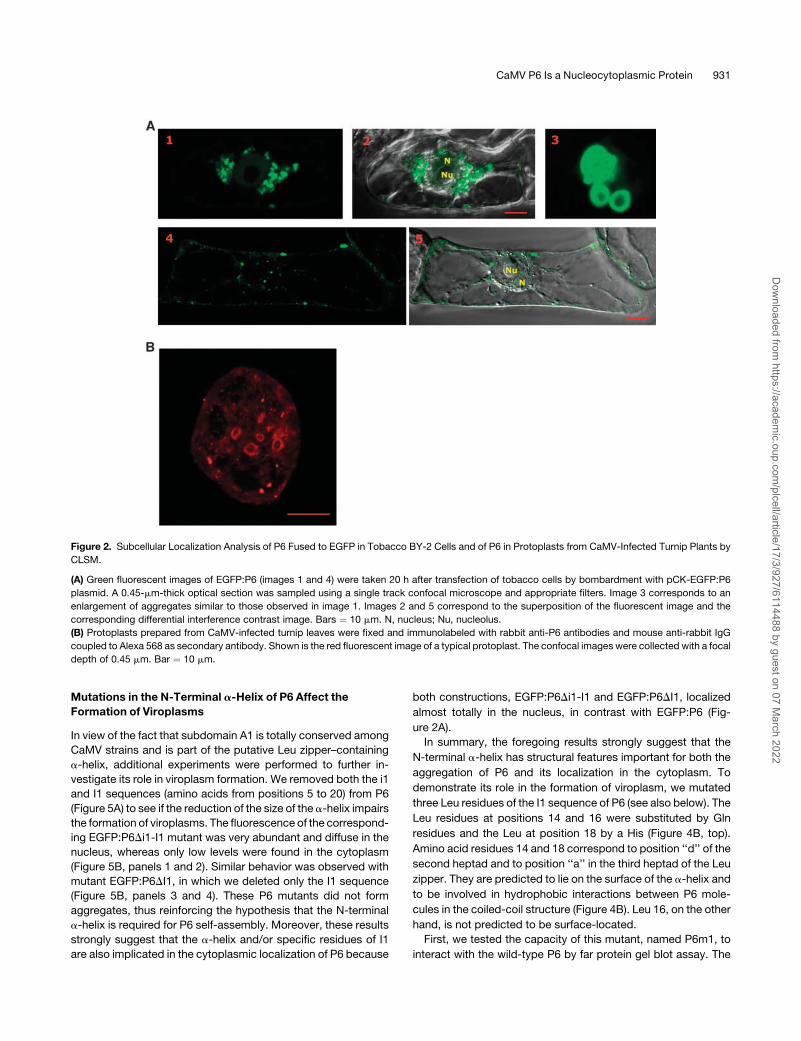
After bombardment of BY-2 cells with plasmids expressing the
EGFP:P6 fusion protein, ;80% of transfected cells contained
large cytoplasmic inclusion bodies (3 to 5 mm in diameter) with
pitted surfaces, generally in the proximity of the nucleus (Figure
2A, panels 1 and 2). The inclusion bodies were formed by
numerous smaller aggregates, most of which appeared as
hollow donut-like structures (Figure 2A, panel 3). To exclude
the possibility that their formation was artifactual (i.e., as a result
of the EGFP moiety fused to the P6), protoplasts were prepared
from CaMV-infected turnip plants as described by Kobayashi
et al. (1998), fixed and immunolabeled with anti-P6 and second-
ary antibodies coupled to the fluorochrome Alexa 568. Obser-
vations by CLSM revealed that the viroplasms thus produced in
the context of an authentic viral infection had a similar structure
(Figure 2B), demonstrating that the EGFP moiety has no pro-
nounced effect on the self-assembly of EGFP:P6 in tobacco
cells. Moreover these results also illustrate that P6 molecules
assemble properly and independently of the cellular context
because similarly shaped aggregates were formed in cells from
host (turnip) and non-host (tobacco) plants.
Approximately 20% of the tobacco cells expressing the
EGFP:P6 fusion protein contained aggregates of variable sizes
but <2 mm in diameter (Figure 2A, panels 4 and 5), which
probably correspond to early stages of viroplasm formation.
The smaller aggregates generally were scattered in the cyto-
plasm, although they were also sometimes found within the
nucleus when the cells were analyzed by CLSM. The presence of
such aggregates within the nucleus may indicate that EGFP:P6
molecules were transported to the nucleus (see below) and were
then unable to exit after their self-assembly because of the large
size of the resulting aggregates.
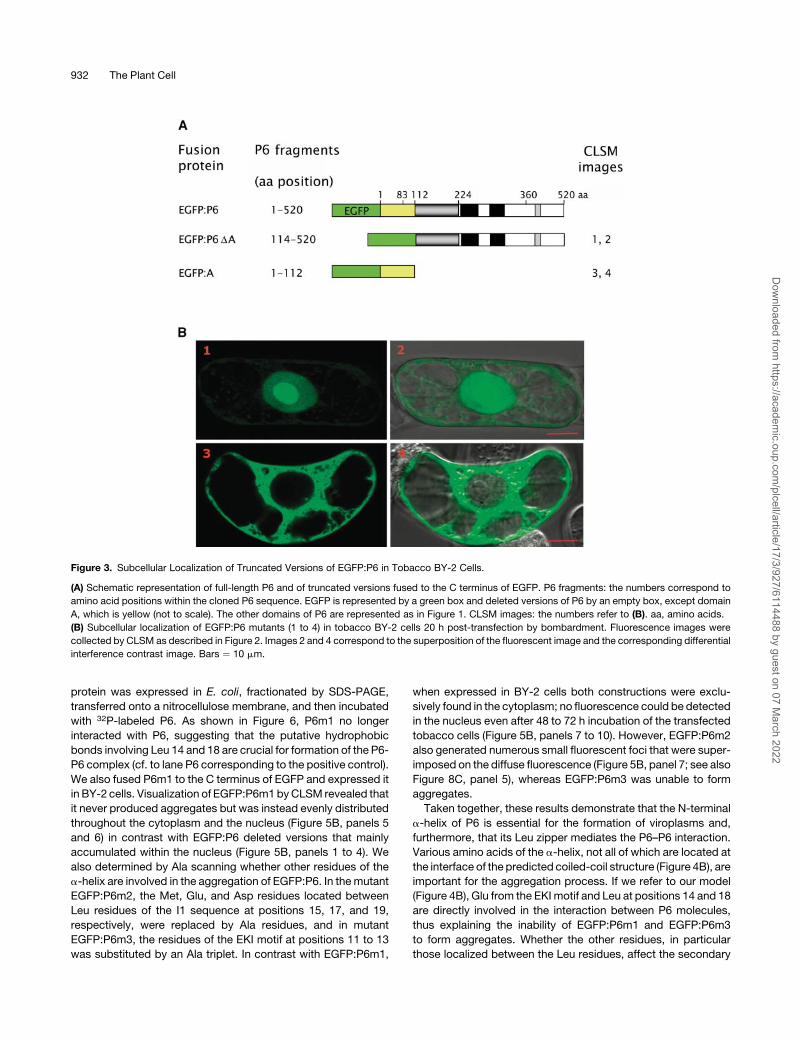
In contrast with EGFP:P6, EGFP:P6DA (Figure 3A) did not form
aggregates in tobacco cells (Figure 3B, panels 1 and 2) but was
mainly found in the nucleus and in particular within the nucleolus.
This result strongly suggests that the N-terminal region of P6 is
a determinant necessary for the formation of aggregates and that
it also contains signal(s) involved in the targeting and/or retention
of P6 within the cytoplasm. Similarly to EGFP:P6DA, expression
of EGFP:A never gave rise to aggregates of any size, but the
protein was instead distributed uniformly in the cytoplasm
(Figure 3B, panels 3 and 4). The failure of EGFP:A to accumulate
in the nucleus to a significant extent was somewhat surprising
because it is sufficiently small that it would be expected to be
able to traverse nuclear pores by passive diffusion.
Taken together, these data show that the N-terminal region of
P6 is necessary but apparently not sufficient for the formation of
viroplasms and, thus, that other region(s) of P6 contribute to this
process. Our failure to detect EGFP:A in the nucleus strongly
supports the idea that the cytosolic localization of P6 is governed
by domain A.
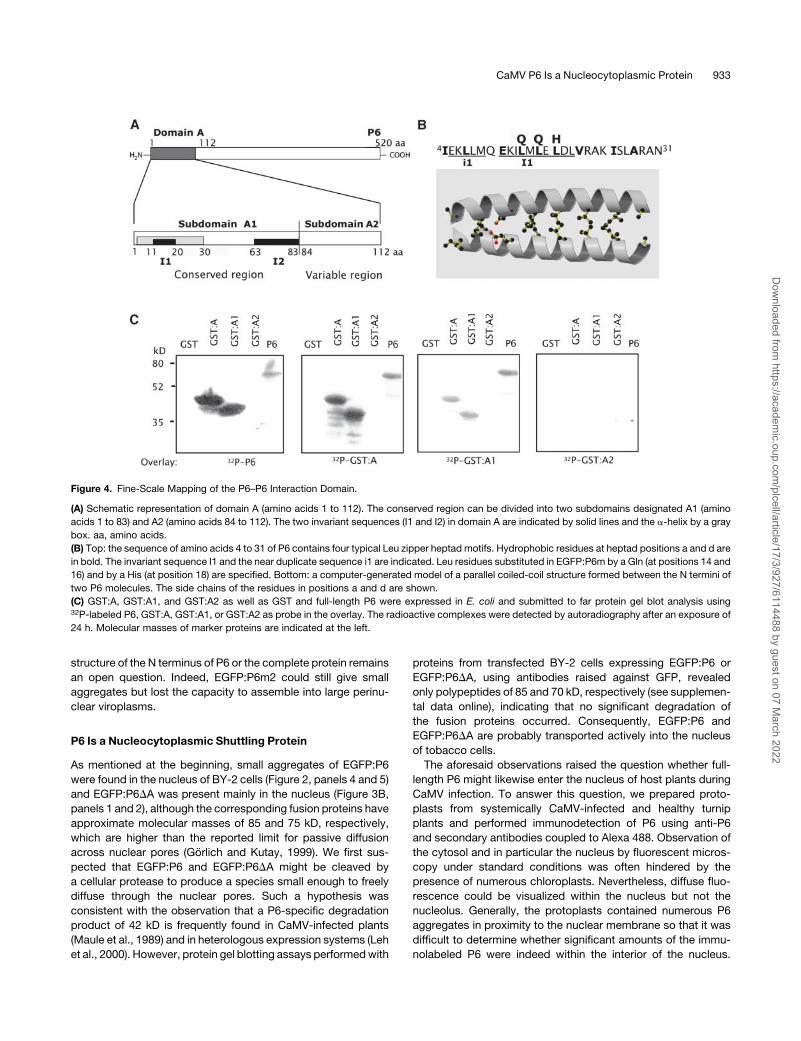
The N-Terminal P6–P6 Interaction Domain Is Conserved
among CaMV Strains
Comparison of the N-terminal sequence of P6 from CaMV strain
Cabb-B JI with its counterparts from other CaMV strains (Cabb
S, Cabb S-Japan, NY8153, CM1841, W260, D/H, D4, B29, Xin/
Jin, and Bari) showed identity ranging from 83 to 97%. The region
can be divided into two subdomains that we have designated A1
(amino acids 1 to 83) and A2 (amino acids 84 to 112), respectively
(Figure 4A). The former is the most conserved (87 to 99% of
identity) with two notable invariant sequences: I1 (amino acids 11
to 20) and I2 (amino acids 63 to 83). Note that I1 also contains
a pentapeptide motif EKILM (residues 11 to 15) that is identical at
four of five positions to the upsteam sequence EKLLM (motif i1;
Figure 4B, top). The sequence I1 forms part of a predicted a-helix
located near the N terminus (positions 4 to 31) that contains
a series of four successive heptad sequences (abcdefg), where
residues in positions a and d are hydrophobic as in Leu zipper
motifs (Figure 4B). This sequence is predicted by computer
analysis (Berger, 1995) to dimerize via a coiled-coil structure with
>98% probability. Subdomain A2 is more variable (62 to 93%
identity) and does not possess a conserved motif among CaMV
strains; it has a predicted b sheet configuration.
As a first step toward further defining the P6–P6 interaction
domain, the sequences coding for subdomains A1 and A2 were
amplified by PCR and cloned into the pGEX-2TK vector to
produce GST-tagged proteins. The capacity of these fusion
proteins to self-interact and to interact with P6 or GST:A were
tested by far protein gel blot experiments. Radiolabeled P6
interacted with GST:A and GST:A1 but not with GST:A2 (Figure
4C, left panel), showing that subdomain A1 is responsible for the
interaction between P6 molecules. This result was confirmed by
the finding that, when either GST:A or GST:A1 was used as
overlay, they interacted with P6 and with themselves but not with
GST:A2, whereas the latter was unable to bind either to P6 or to
any of the fusion proteins (Figure 4C, right panels). Note that none
of the fusion proteins interacted with GST, excluding the possi-
bility that the observed interactions were due to dimerization of
the tag. Taken together, these results provide evidence that the
83 N-terminal amino acids encompass the domain required for
P6 self-interaction in vitro and, hence, probably for the formation
of viroplasms.
930 The Plant Cell
Dow
nloaded from https://academ
ic.oup.com/plcell/article/17/3/927/6114488 by guest on 07 M
arch 2022
Mutations in the N-Terminal a-Helix of P6 Affect the
Formation of Viroplasms
In view of the fact that subdomain A1 is totally conserved among
CaMV strains and is part of the putative Leu zipper–containing
a-helix, additional experiments were performed to further in-
vestigate its role in viroplasm formation. We removed both the i1
and I1 sequences (amino acids from positions 5 to 20) from P6
(Figure 5A) to see if the reduction of the size of the a-helix impairs
the formation of viroplasms. The fluorescence of the correspond-
ing EGFP:P6Di1-I1 mutant was very abundant and diffuse in the
nucleus, whereas only low levels were found in the cytoplasm
(Figure 5B, panels 1 and 2). Similar behavior was observed with
mutant EGFP:P6DI1, in which we deleted only the I1 sequence
(Figure 5B, panels 3 and 4). These P6 mutants did not form
aggregates, thus reinforcing the hypothesis that the N-terminal
a-helix is required for P6 self-assembly. Moreover, these results
strongly suggest that the a-helix and/or specific residues of I1
are also implicated in the cytoplasmic localization of P6 because
both constructions, EGFP:P6Di1-I1 and EGFP:P6DI1, localized
almost totally in the nucleus, in contrast with EGFP:P6 (Fig-
ure 2A).
In summary, the foregoing results strongly suggest that the
N-terminal a-helix has structural features important for both the
aggregation of P6 and its localization in the cytoplasm. To
demonstrate its role in the formation of viroplasm, we mutated
three Leu residues of the I1 sequence of P6 (see also below). The
Leu residues at positions 14 and 16 were substituted by Gln
residues and the Leu at position 18 by a His (Figure 4B, top).
Amino acid residues 14 and 18 correspond to position ‘‘d’’ of the
second heptad and to position ‘‘a’’ in the third heptad of the Leu
zipper. They are predicted to lie on the surface of the a-helix and
to be involved in hydrophobic interactions between P6 mole-
cules in the coiled-coil structure (Figure 4B). Leu 16, on the other
hand, is not predicted to be surface-located.
First, we tested the capacity of this mutant, named P6m1, to
interact with the wild-type P6 by far protein gel blot assay. The
Figure 2. Subcellular Localization Analysis of P6 Fused to EGFP in Tobacco BY-2 Cells and of P6 in Protoplasts from CaMV-Infected Turnip Plants by
CLSM.
(A) Green fluorescent images of EGFP:P6 (images 1 and 4) were taken 20 h after transfection of tobacco cells by bombardment with pCK-EGFP:P6
plasmid. A 0.45-mm-thick optical section was sampled using a single track confocal microscope and appropriate filters. Image 3 corresponds to an
enlargement of aggregates similar to those observed in image 1. Images 2 and 5 correspond to the superposition of the fluorescent image and the
corresponding differential interference contrast image. Bars ¼ 10 mm. N, nucleus; Nu, nucleolus.
(B) Protoplasts prepared from CaMV-infected turnip leaves were fixed and immunolabeled with rabbit anti-P6 antibodies and mouse anti-rabbit IgG
coupled to Alexa 568 as secondary antibody. Shown is the red fluorescent image of a typical protoplast. The confocal images were collected with a focal
depth of 0.45 mm. Bar ¼ 10 mm.
CaMV P6 Is a Nucleocytoplasmic Protein 931
Dow
nloaded from https://academ
ic.oup.com/plcell/article/17/3/927/6114488 by guest on 07 M
arch 2022
protein was expressed in E. coli, fractionated by SDS-PAGE,
transferred onto a nitrocellulose membrane, and then incubated
with 32P-labeled P6. As shown in Figure 6, P6m1 no longer
interacted with P6, suggesting that the putative hydrophobic
bonds involving Leu 14 and 18 are crucial for formation of the P6-
P6 complex (cf. to lane P6 corresponding to the positive control).
We also fused P6m1 to the C terminus of EGFP and expressed it
in BY-2 cells. Visualization of EGFP:P6m1 by CLSM revealed that
it never produced aggregates but was instead evenly distributed
throughout the cytoplasm and the nucleus (Figure 5B, panels 5
and 6) in contrast with EGFP:P6 deleted versions that mainly
accumulated within the nucleus (Figure 5B, panels 1 to 4). We
also determined by Ala scanning whether other residues of the
a-helix are involved in the aggregation of EGFP:P6. In the mutant
EGFP:P6m2, the Met, Glu, and Asp residues located between
Leu residues of the I1 sequence at positions 15, 17, and 19,
respectively, were replaced by Ala residues, and in mutant
EGFP:P6m3, the residues of the EKI motif at positions 11 to 13
was substituted by an Ala triplet. In contrast with EGFP:P6m1,
when expressed in BY-2 cells both constructions were exclu-
sively found in the cytoplasm; no fluorescence could be detected
in the nucleus even after 48 to 72 h incubation of the transfected
tobacco cells (Figure 5B, panels 7 to 10). However, EGFP:P6m2
also generated numerous small fluorescent foci that were super-
imposed on the diffuse fluorescence (Figure 5B, panel 7; see also
Figure 8C, panel 5), whereas EGFP:P6m3 was unable to form
aggregates.
Taken together, these results demonstrate that the N-terminal
a-helix of P6 is essential for the formation of viroplasms and,
furthermore, that its Leu zipper mediates the P6–P6 interaction.
Various amino acids of the a-helix, not all of which are located at
the interface of the predicted coiled-coil structure (Figure 4B), are
important for the aggregation process. If we refer to our model
(Figure 4B), Glu from the EKI motif and Leu at positions 14 and 18
are directly involved in the interaction between P6 molecules,
thus explaining the inability of EGFP:P6m1 and EGFP:P6m3
to form aggregates. Whether the other residues, in particular
those localized between the Leu residues, affect the secondary
Figure 3. Subcellular Localization of Truncated Versions of EGFP:P6 in Tobacco BY-2 Cells.
(A) Schematic representation of full-length P6 and of truncated versions fused to the C terminus of EGFP. P6 fragments: the numbers correspond to
amino acid positions within the cloned P6 sequence. EGFP is represented by a green box and deleted versions of P6 by an empty box, except domain
A, which is yellow (not to scale). The other domains of P6 are represented as in Figure 1. CLSM images: the numbers refer to (B). aa, amino acids.
(B) Subcellular localization of EGFP:P6 mutants (1 to 4) in tobacco BY-2 cells 20 h post-transfection by bombardment. Fluorescence images were
collected by CLSM as described in Figure 2. Images 2 and 4 correspond to the superposition of the fluorescent image and the corresponding differential
interference contrast image. Bars ¼ 10 mm.
932 The Plant Cell
Dow
nloaded from https://academ
ic.oup.com/plcell/article/17/3/927/6114488 by guest on 07 M
arch 2022
structure of the N terminus of P6 or the complete protein remains
an open question. Indeed, EGFP:P6m2 could still give small
aggregates but lost the capacity to assemble into large perinu-
clear viroplasms.
P6 Is a Nucleocytoplasmic Shuttling Protein
As mentioned at the beginning, small aggregates of EGFP:P6
were found in the nucleus of BY-2 cells (Figure 2, panels 4 and 5)
and EGFP:P6DA was present mainly in the nucleus (Figure 3B,
panels 1 and 2), although the corresponding fusion proteins have
approximate molecular masses of 85 and 75 kD, respectively,
which are higher than the reported limit for passive diffusion
across nuclear pores (Gorlich and Kutay, 1999). We first sus-
pected that EGFP:P6 and EGFP:P6DA might be cleaved by
a cellular protease to produce a species small enough to freely
diffuse through the nuclear pores. Such a hypothesis was
consistent with the observation that a P6-specific degradation
product of 42 kD is frequently found in CaMV-infected plants
(Maule et al., 1989) and in heterologous expression systems (Leh
et al., 2000). However, protein gel blotting assays performed with
proteins from transfected BY-2 cells expressing EGFP:P6 or
EGFP:P6DA, using antibodies raised against GFP, revealed
only polypeptides of 85 and 70 kD, respectively (see supplemen-
tal data online), indicating that no significant degradation of
the fusion proteins occurred. Consequently, EGFP:P6 and
EGFP:P6DA are probably transported actively into the nucleus
of tobacco cells.
The aforesaid observations raised the question whether full-
length P6 might likewise enter the nucleus of host plants during
CaMV infection. To answer this question, we prepared proto-
plasts from systemically CaMV-infected and healthy turnip
plants and performed immunodetection of P6 using anti-P6
and secondary antibodies coupled to Alexa 488. Observation of
the cytosol and in particular the nucleus by fluorescent micros-
copy under standard conditions was often hindered by the
presence of numerous chloroplasts. Nevertheless, diffuse fluo-
rescence could be visualized within the nucleus but not the
nucleolus. Generally, the protoplasts contained numerous P6
aggregates in proximity to the nuclear membrane so that it was
difficult to determine whether significant amounts of the immu-
nolabeled P6 were indeed within the interior of the nucleus.
Figure 4. Fine-Scale Mapping of the P6–P6 Interaction Domain.
(A) Schematic representation of domain A (amino acids 1 to 112). The conserved region can be divided into two subdomains designated A1 (amino
acids 1 to 83) and A2 (amino acids 84 to 112). The two invariant sequences (I1 and I2) in domain A are indicated by solid lines and the a-helix by a gray
box. aa, amino acids.
(B) Top: the sequence of amino acids 4 to 31 of P6 contains four typical Leu zipper heptad motifs. Hydrophobic residues at heptad positions a and d are
in bold. The invariant sequence I1 and the near duplicate sequence i1 are indicated. Leu residues substituted in EGFP:P6m by a Gln (at positions 14 and
16) and by a His (at position 18) are specified. Bottom: a computer-generated model of a parallel coiled-coil structure formed between the N termini of
two P6 molecules. The side chains of the residues in positions a and d are shown.
(C) GST:A, GST:A1, and GST:A2 as well as GST and full-length P6 were expressed in E. coli and submitted to far protein gel blot analysis using32P-labeled P6, GST:A, GST:A1, or GST:A2 as probe in the overlay. The radioactive complexes were detected by autoradiography after an exposure of
24 h. Molecular masses of marker proteins are indicated at the left.
CaMV P6 Is a Nucleocytoplasmic Protein 933
Dow
nloaded from https://academ
ic.oup.com/plcell/article/17/3/927/6114488 by guest on 07 M
arch 2022
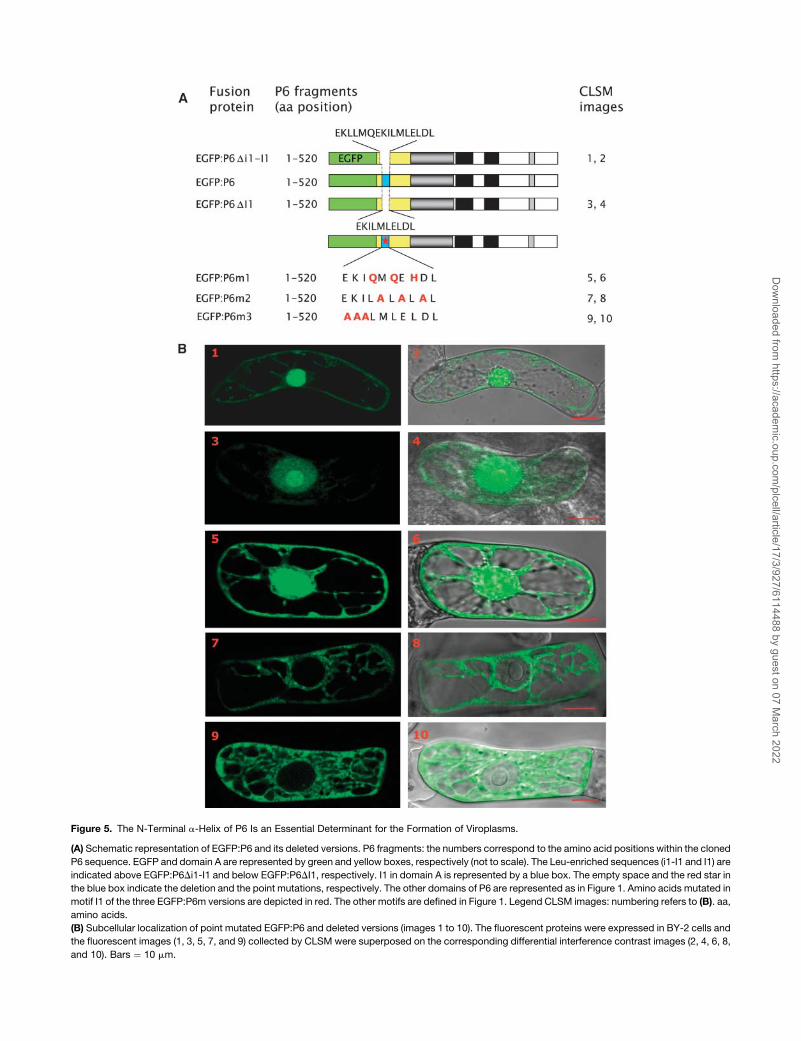
Figure 5. The N-Terminal a-Helix of P6 Is an Essential Determinant for the Formation of Viroplasms.
(A) Schematic representation of EGFP:P6 and its deleted versions. P6 fragments: the numbers correspond to the amino acid positions within the cloned
P6 sequence. EGFP and domain A are represented by green and yellow boxes, respectively (not to scale). The Leu-enriched sequences (i1-I1 and I1) are
indicated above EGFP:P6Di1-I1 and below EGFP:P6DI1, respectively. I1 in domain A is represented by a blue box. The empty space and the red star in
the blue box indicate the deletion and the point mutations, respectively. The other domains of P6 are represented as in Figure 1. Amino acids mutated in
motif I1 of the three EGFP:P6m versions are depicted in red. The other motifs are defined in Figure 1. Legend CLSM images: numbering refers to (B). aa,
amino acids.
(B) Subcellular localization of point mutated EGFP:P6 and deleted versions (images 1 to 10). The fluorescent proteins were expressed in BY-2 cells and
the fluorescent images (1, 3, 5, 7, and 9) collected by CLSM were superposed on the corresponding differential interference contrast images (2, 4, 6, 8,
and 10). Bars ¼ 10 mm.
Dow
nloaded from https://academ
ic.oup.com/plcell/article/17/3/927/6114488 by guest on 07 M
arch 2022

Therefore, immunolabeling with anti-P6 antibodies was per-
formed on purified nuclei isolated from healthy and CaMV-
infected turnip protoplasts; the nuclei were also stained with
propidium iodide (Figures 7A and 7B, panel 2). Nuclei prepared
from healthy plants never reacted with anti-P6 antibodies (Figure
7A), whereas ;50% of those from infected plants were immu-
nolabeled (Figure 7B). P6-Alexa 488 fluorescent foci were
observed in the nucleoplasm and/or the nucleolus from infected
turnip cells, thus demonstrating that P6 molecules do indeed
enter the nucleus during the CaMV infection cycle. The nuclei
were often contaminated with P6-containing viroplasms be-
cause the latter remained firmly attached to the outer surface
even when they were further purified through a composite
sucrose/Percoll gradient. Therefore, we realized a series of
photographs obtained by CLSM analysis of 0.5-mm-thick sec-
tions across purified and P6-immunolabeled nuclei. As shown in
Figure 7C, large contaminating viroplasms progressively disap-
peared from view in successive sections (panels 5 to 14),
whereas small P6-labeled aggregates progressively appeared
within the organelle (panels 13 to 18), thus providing additional
evidence for the presence of P6 within the nucleus. The fluores-
cent foci might correspond to P6 aggregates and/or to inter-
actions between P6 and specialized nuclear compartments such
as speckles (for a review, see Lamond and Spector, 2003) or
Cajal bodies (Ochs et al., 1994). The localization of P6 in the
nucleolus, observed in some cases, might be related to its
capacity to interact with the ribosomal subunits as shown by
Park et al. (2001).
To exclude the possibility that P6 entered the nuclei during
their purification (i.e., by diffusion through an altered nuclear
envelope), nuclei from healthy protoplasts were incubated during
30 min at 48C with ;100 mg of soluble P6 protein expressed in
E. coli. The preparation was immunolabeled as describe above.
No fluorescence was detectable within the organelles after this
treatment, thus reinforcing our conclusion that the P6 protein
found in nuclei from infected cells is actively transported there in
the course of the CaMV replication cycle.
In conclusion, our data clearly support the idea that P6 can
enter the nucleus during viral infection and also indicate that P6 is
a nucleocytoplasmic shuttling protein. Furthermore, our findings
also strongly suggest that the sequence downstream of domain
A is implicated in the nuclear localization of P6, whereas the
N-terminal region of P6 might contain a nuclear export signal
(NES).
The N Terminus of P6 Contains an NES That Is Recognized
by the CRM-1 Nuclear Export Pathway
The latter hypothesis is reinforced by the fact that deletion of the
conserved hydrophobic sequence I1 located in subdomain A1 or
mutation of the Leu residues at positions 14, 16, and 18 partially
abolished nuclear export of P6 (Figure 5B, panels 3 to 6).
Moreover, this Leu-rich sequence bears some resemblance to
the NES (EKDTLLIDL) found in the BR1 protein of the Squash leaf
curl virus, a geminivirus (Ward and Lazarowitz, 1999), and to the
NES sequence of several known rapidly shuttling nuclear pro-
teins, such as HIV Rev protein (for a review, see Pollard and
Malim, 1998).
To provide further evidence that the aforesaid sequence is an
NES, we deleted the sequence I1 or mutated Leu residues 14, 16,
and 18 in the EGFP:A fusion protein (Figure 8A) as described
above, and the behavior in tobacco BY-2 cells of the mutants
was compared with that of the nonmutated protein. These
experiments were performed with EGFP:A rather than with the
full-length P6 because nuclear accumulation of EGFP:A mutants
would be directly relevant to the impairment of the export
process, whereas the accumulation of EGFP:P6 mutants in the
nucleus does not permit discrimination between an export defect
and an active import of the protein. Indeed, the wild-type fusion
protein EGFP:A was never found in the nucleus, although it is of
a size that should permit it to diffuse freely through the nuclear
pore (Figure 3B, panels 3 and 4).
When sequence I1 was removed from domain A, EGFP:ADI1
was equally distributed in the cytoplasm and the nucleus except
for the nucleolus (Figure 8B, panels 1 and 2), whereas EGFP:A
localized exclusively to the cytoplasmic compartment (Figure
3B, panels 3 and 4). The subcellular localization in BY-2 cells of
mutant EGFP:Am (Figure 8B, panels 3 and 4), in which the three
Leu residues of I1 were replaced by polar residues, was similar
to that observed with EGFP:ADI1 (Figure 8B, panels 1 and 2);
EGFP:Am was found in both the cytoplasm and in the nucleus
but not in the nucleolus. The absence of both EGFP:A mutants in
the nucleolus, in contrast with the situation with EGFP:P6DI1 and
EGFP:P6 point mutated versions (Figure 5), might be due to the
fact that the N-terminal region of P6 is unable to interact with
ribosomal proteins, whereas the EGFP:P6 mutants still contain
the corresponding interaction domains (i.e., mini-TAV and RNA
binding domain A) (Leh et al., 2000; Park et al., 2001; Bureau
et al., 2004).
All these results support a model in which (1) both EGFP:A and
EGFP:A mutants can enter and exit the nucleus by diffusion, (2)
EGFP:A molecules but neither EGFP:ADI1 nor EGFP:Am are
rapidly exported to the cytoplasm, and (3) that the sequence I1
functions as an NES because point mutations of Leu residues in
I1 impair the export of EGFP:A. Thus, these Leu residues appear
Figure 6. Point Mutations in the Invariant Sequence I1 Impair P6–P6
Interactions
Protein extracts of bacteria expressing either wild-type P6 (lane P6),
point-mutated P6 (lane P6m1), or an extract of bacteria transformed with
empty vector (lane E. coli) were fractionated by SDS-PAGE (15%) and
transferred onto a nitrocellulose membrane. The membranes were
incubated either with antibodies raised against P6 (right panel) or
submitted to a far protein gel blot assay using in vitro 32P-labeled P6
(left panel). Molecular masses of marker proteins are indicated to the left.
CaMV P6 Is a Nucleocytoplasmic Protein 935
Dow
nloaded from https://academ
ic.oup.com/plcell/article/17/3/927/6114488 by guest on 07 M
arch 2022

to be essential residues for the nuclear export of P6 as already
described for nucleocytoplasmic shuttling proteins possessing
a Leu-rich NES and in particular for the BR1 protein of Squash
leaf curl virus (Ward and Lazarowitz, 1999). However, we cannot
totally exclude that the invariant sequence I1 also has properties
involved in the retention of fusion protein in the cytosol.
Therefore, the nuclear export of P6 was further investigated,
treating BY-2 cells transfected with the aforementioned re-
combinant plasmids with leptomycin B, which specifically inhib-
its the CRM-1 pathway involved in the nuclear export of many
proteins (Fornerod et al., 1997; Kudo et al., 1998). When BY-2
cells expressing EGFP:A were incubated with 100 nM lepto-
mycin B, 6 h after bombardment, the fluorescent protein accu-
mulated abundantly in the nucleus (Figure 8C, panel 2), whereas
no fluorescence was present in this compartment in untreated
control cells (Figure 8C, panel 1), thus confirming that EGFP:A
molecules are actively exported from the nucleus. The fusion
protein EGFP:P6 formed large aggregates in the cytoplasm and
was undetectable in the nucleus of transfected tobacco cells
(Figure 8C, panel 3), but when the latter were treated with
leptomycin B, it was found in both the cytoplasm and in the
nucleus (Figure 8C, panel 4). The nuclear compartment con-
tained diffuse fluorescence, accompanied as expected by many
small aggregates because P6 was able to self-interact. This
result proves that P6 is actively transported between the nucle-
ocytoplasmic compartments. Similar experiments were also
performed with point mutated EGFP:P6 versions to determine
the functional importance of different residues in the nuclear
export of P6. As expected, the mutant EGFP:P6m1 had the same
subcellular localization in BY-2 cells treated with leptomycin B
Figure 7. Nuclear Localization of P6.
Nuclei from healthy (A) and CaMV-infected turnip leaves (B) were fixed and immunolabeled with rabbit anti-P6 antibodies and mouse anti-rabbit IgG
coupled to Alexa 488 as secondary antibodies (image 1) and stained with propidium iodide (image 2). Panel 3 corresponds to differential interference
contrast images and the right-hand images (panel 4) to their superposition with the fluorescent and propidium-stained images. The confocal images
were collected with a focal depth of 0.45 mm (C). A series of such optical sections through a nucleus isolated from an infected plant, anti-P6
immunolabeled, and stained with propidium iodide. Bar ¼ 5 mm.
936 The Plant Cell
Dow
nloaded from https://academ
ic.oup.com/plcell/article/17/3/927/6114488 by guest on 07 M
arch 2022
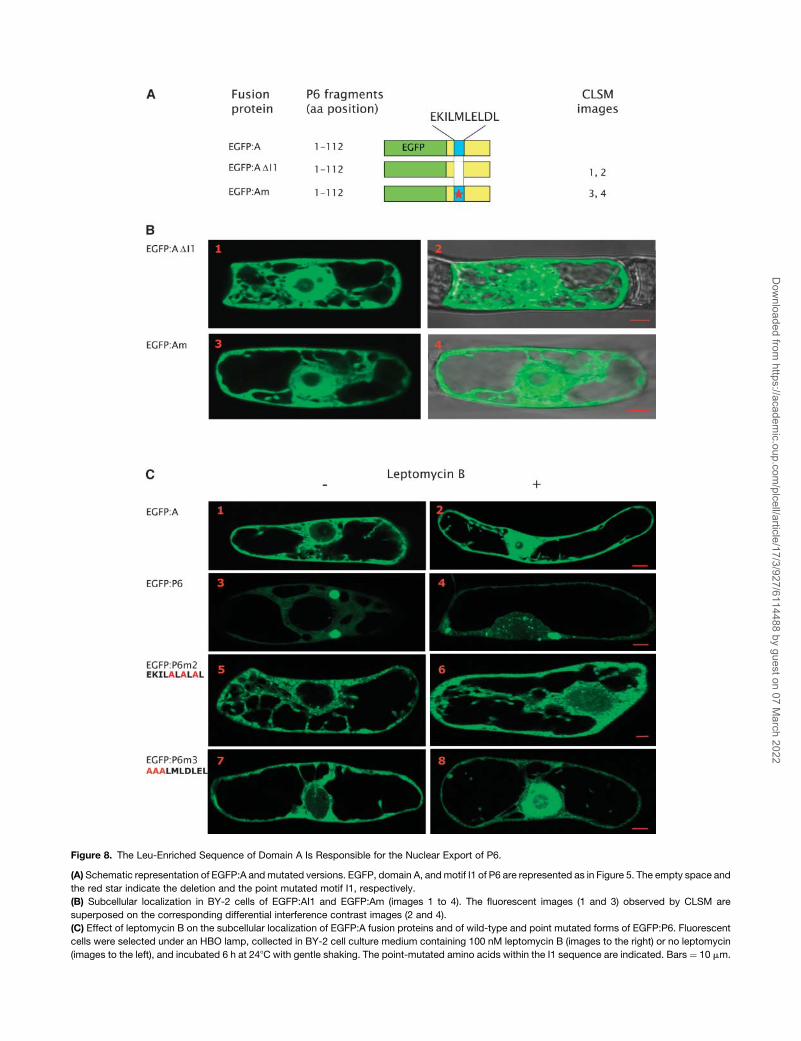
Figure 8. The Leu-Enriched Sequence of Domain A Is Responsible for the Nuclear Export of P6.
(A) Schematic representation of EGFP:A and mutated versions. EGFP, domain A, and motif I1 of P6 are represented as in Figure 5. The empty space and
the red star indicate the deletion and the point mutated motif I1, respectively.
(B) Subcellular localization in BY-2 cells of EGFP:AI1 and EGFP:Am (images 1 to 4). The fluorescent images (1 and 3) observed by CLSM are
superposed on the corresponding differential interference contrast images (2 and 4).
(C) Effect of leptomycin B on the subcellular localization of EGFP:A fusion proteins and of wild-type and point mutated forms of EGFP:P6. Fluorescent
cells were selected under an HBO lamp, collected in BY-2 cell culture medium containing 100 nM leptomycin B (images to the right) or no leptomycin
(images to the left), and incubated 6 h at 248C with gentle shaking. The point-mutated amino acids within the I1 sequence are indicated. Bars ¼ 10 mm.
Dow
nloaded from https://academ
ic.oup.com/plcell/article/17/3/927/6114488 by guest on 07 M
arch 2022

(data not shown) or not (Figure 5B, panels 5 and 6) because
mutations of the Leu residues are sufficient per se to impair the
export of the fusion protein. The fluorescence in the cytoplasm
even after leptomycin B treatment suggests that not all
EGFP:P6m1 molecules entered the nucleus because they were
retained in the cytoplasm or, alternatively, that other residues of
the I1 sequence might contribute to the export. In contrast with
EGFP:P6m1, leptomycin B had only little effect on the nuclear
export of EGFP:P6m2 because almost no fluorescence could be
detected within the nucleus (Figure 8C, panels 5 and 6) in most
BY-2 cells observed by CLSM; only few cells exhibited a highly
fluorescent nucleus. The difference observed in the response to
leptomycin B treatment might be due to the fact that (1) the
residues mutated in EGFP:P6m2 are indeed essential for export
as hypothesized previously or (2) that their mutations modify the
conformation of EGFP:P6 in a manner that interferes somehow
with the activity of the inhibitor. The EGFP:P6m3 mutant mainly
accumulated in the nucleoplasm when the cells were incubated
in the presence of leptomycin B, whereas it did not in untreated
cells; only low levels of EGFP:P6m3 were found in the cytoplasm
of treated cells (Figure 8C, panels 7 and 8). This result suggests
that the EKI motif is not part of the NES, or if it is, it has a minor
influence on the export of P6 by the CRM-1 pathway.
Together, these data lead to the conclusion that P6 contains at
its N terminus an NES and that its Leu residues involved in the
nuclear export of P6 are recognized by the CRM-1 pathway.
The activity of this export pathway limits the accumulation of P6
in the nucleus that could be deleterious for the CaMV infectious
cycle.
DISCUSSION
During the course of a CaMV infection, P6 forms electron-dense
viroplasms that are thought to be virion factories that serve as
a scaffold for virus replication and assembly. In this study, we
have investigated by in vitro and in vivo experimental approaches
the mechanism leading to the formation of these viroplasms. Our
results show that P6 self-interacts, as already suggested (Haas
et al., 2000), and further demonstrate that, at least in vitro, the
P6–P6 interactions are exclusively mediated by the N-terminal
region encompassing residues 1 to 83 (subdomain A1). No other
P6 sequence was able to interact in vitro with full-length P6. The
fact that fusion of domain A to unrelated proteins drove this
interaction in vitro demonstrates that the N terminus of P6 is
sufficient per se to promote self-interaction independently from
neighboring sequences. Our results only partially agree with
those obtained by Li and Leisner (2002), who used the yeast
double hybrid system to show that, in addition to the N terminus,
three other domains of P6 are able to bind full-length P6, namely
the mini-TAV domain, the downstream adjacent sequence, and
the C-terminal region except for a 90-amino-acid-long se-
quence. Possibly the additional interacting domains character-
ized in the double hybrid system bind to P6 with an affinity that is
too low for detection in far protein gel blot experiments. In-
complete renaturation of the domains in question could also
interfere with activity in the in vitro assays. On the other hand, we
cannot totally exclude the possibilities that some of the inter-
actions detected in yeast by Li and Leisner (2002) might be
mediated by yeast RNA or proteins. Indeed, domains of P6
identified by these authors to be involved in P6–P6 interactions
have nucleic acid binding properties and/or interact with cellular
proteins, in particular nuclear-localized proteins (Park et al.,
2001; Bureau et al., 2004).
The role of subdomain A1 in the formation of viroplasms was
investigated by transient expression in tobacco BY-2 cells of full-
length P6 and deleted versions of P6 fused to EGFP. Expression
of full-length P6 fusion protein gave rise to viroplasms located
in the proximity of the nucleus, which are structurally similar to
those found in CaMV-infected plants. Indeed, in both cases, the
viroplasms appeared by confocal microscopy analysis as ag-
gregates of multiple hollow macromolecular structures, and it is
suggested that they might be produced stepwise by the assem-
bly of the donut-like structures that could be visualized in some
transfected tobacco cells. Similar hollow structures were also
observed when NSP2 and NSP5 proteins, the two major viral
components of dense viroplasms induced by rotaviruses, were
coexpressed in cultured host cells in the absence of other
rotaviral proteins and of rotavirus replication (Fabbretti et al.,
1999). We assume that the hollow center of the P6 aggregates
corresponds to the electron-lucent holes of viroplasms observed
by electron microscopy in CaMV-infected cells (Xiong et al.,
1982).
The results obtained by transient expression of truncated
versions of P6 clearly demonstrate that, although the 83
N-terminal amino acids of P6 are necessary, they are not suffi-
cient for the formation of viroplasms and that consequently other
region(s) of P6 are implicated in this process. The domain
between amino acids 289 and 379, referred to as D3 by Li and
Leisner (2002), might be one of these sequences because
these authors have shown that it plays an important role in P6
self-association when tested in vivo using the yeast two hybrid
system. This region of P6 is also engaged in interactions needed
for translational transactivation (De Tapia et al., 1993; Park et al.,
2001). In any event, our findings suggest that the N-terminally
mediated P6–P6 interaction is a prerequisite for further inter-
actions between other P6 sequences and/or for stabilization of
macromolecular structures because formation of viroplasms
was totally impaired when an N-terminally truncated P6 (P6DA)
was expressed in tobacco cells.
Computer analysis of subdomain A1 indicated that it forms an
amphipathic a-helix at its N terminus (residues 4 to 31). The latter
contains a Leu zipper motif that could form a parallel coiled-coil
structure (Lupas, 1997), strongly suggesting that such a confor-
mation is implicated in the interaction between P6 molecules.
This hypothesis is confirmed by the results of far protein gel blot
assays showing that the intermolecular interaction is lost if key
hydrophobic amino acids of the Leu zipper motifs are substituted
by polar residues. Involvement of such an interaction in the
formation of viroplasms is evidenced by the failure of P6 carrying
these point mutations in the a-helix to form aggregates in
transfected tobacco cells. Currently, we hypothesize that the
coiled-coil formation between the N termini of interacting P6
molecules induces conformational changes that allow other
regions of P6 to participate in the aggregation process. Indeed,
mutations of amino acids that are not located at the coiled-coil
interacting surface did not prevent the formation of small
938 The Plant Cell
Dow
nloaded from https://academ
ic.oup.com/plcell/article/17/3/927/6114488 by guest on 07 M
arch 2022

aggregates; however, they impaired assembly of these aggre-
gates into viroplasms. It is evident that a further understanding of
the mechanism of viroplasm formation will also require charac-
terization of the other domains of P6 implicated in this process
as well as possible cellular structures and/or factors that might
be involved, such as endomembranes and/or the microtubular
network. Nevertheless, no host-specific factors seem to be
needed because viroplasms that formed in tobacco cells,
a non-host for CaMV, are similar to those found in host cells.
As noted earlier, membrane components may function in the
early steps of P6 self-assembly, characterized by formation of
small aggregates as the latter disappeared upon treatment of
tobacco cells with nonionic detergent, whereas the viroplasms
located at the nuclear periphery were unaffected (data not
shown). Further investigations will be required to determine
whether binding to endomembranes is essential for the self-
assembly of P6 and whether its hydrophobic N terminus, which
presents features of a peptide signal (Blobel and Dobberstein,
1975), plays a role. Because viroplasm formation does not
visually perturb the microtubule or the actin filament networks
in BY-2 cells, it will be of particular interest to determine whether
viroplasm formation involves the cytoskeleton, as described in
animal cells for aggresomes and for the viral factories induced by
large cytoplasmic DNA viruses such as African swine fever virus
(Kopito, 2000; Heath et al., 2001). Johnston et al. (1998) pro-
posed a model in which small aggregates are delivered by
retrograde transport along microtubules to the periphery of the
nucleus where they are assembled into large structures.
Transient expression of EGFP-tagged P6 led to the unex-
pected discovery that P6, considered until now as a cytoplasmic
protein, is actually a nucleocytoplasmic shuttling protein. The
presence of this viral protein within the nucleus was confirmed by
its immunodetection in nuclei prepared from CaMV-infected
turnip leaves and CLSM-generated optical serial sections
through the organelles. Our results, obtained with transfected
tobacco cells, indicate that only a small fraction of P6, probably
molecules that are not engaged in the aggregation process,
enters the nucleus. Indeed, EGFP:P6 mainly formed inclusion
bodies in the cytoplasm and was almost undetectable in the
nucleus of tobacco cells. Only inhibition of the export process
by leptomycin B systematically permitted observation of diffuse
EGFP:P6 and small aggregates within the nucleus. Our results
also indicate that the export of P6 probably occurs very rapidly
in infected cells, so that only low amounts are present in
the nucleus at any time. The strong P6 nuclear export activity
probably prevents accumulation of P6 within the nucleus, which
could be deleterious for CaMV infectivity. In addition, it is
more than likely that the nucleocytoplasmic transport is finely
Table 1. Oligonucleotides Used as Primers to Generate the PCR Products Cloned into pGEX-2TK, pCK-EGFP, pETP42,
and pETKaKS.6 Vectors
Name Sequence Restriction Site
A Cloning of sequences corresponding to CaMV P6 truncated versions
ABam(þ) gccggatccATGGAGAACGAAAAACTC BamHI
AEco(�) gatgaattcTCATGGAATTCCCTGATGAGG EcoRI
A1Eco(�) cacgaatccCTAAGCCATCAACGGATTTG EcoRI
A2Bam(þ) ggcggatccTCCAATATCTTGTCAAAAGAT BamHI
A2Eco(�) cacgaattcCTATTCTGCTCTGAGAGGAGC EcoRI
B Cloning of sequences corresponding to CaMV P6 protein and its
truncated versions
Bsr mut(þ) GACTGGGGTTGTACTAAGG
Bsr mut(�) CCGCCTTAGTACAACC
P6Bsr(þ) catgtacaagATGGAGAACATAGAAAAACTCATAGTACAACC BsrGI
PBsr(þ) gcatgtacaagCTCAAGATCAGAAGTACTATTC BsrGI
P6DABsr(þ) gcatgtacaagATCCCACAAAAATCTGAGCTTAA BsrGI
QBsr(þ) gcatgtacaagCCAATCCCACAAAAATCTGAG BsrGI
P6Xba(�) gattctagaTCAATCCACTTGCTTTGAAGAC XbaI
AXba(�) gattctagaTCATGGAATTCCCTGATGAGG XbaI
A1Xba(�) gattctagaTCAAGCCATCAACGGATTTGT XbaI
BXba(�) gattctagaTCAGGAGATCTCTTTTGGGGC XbaI
QXba(�) gattctagaTCAAAATATGTCTTTCTCTGTGTTCTTG XbaI
C Cloning of the sequence corresponding to the P6 N-terminal domain
(amino acids 1 to 112)
ANco(þ) gatccatggATGGAGAACATAGAAAAACTC NcoI
ANco(�) ctaccatggtAATTCCCTGATGAGGACG NcoI
D Site-directed mutagenesis
NESmut(þ) AAAATACAAATGCAAGAACACGATCTA
DNES(�) CTCTTGCATGAGGAGTTTTT
NES(þ) GTAAGAGCAAAAATAAGCTTA
Di1(�) TATGTTCTCCATCTTGTACAGC
Forward primer (þ) and reverse primer (�). Restriction sites in the primer sequences are represented by bold-faced lower-case letters. PCR products
were cloned into pGEX-2TK (A), pCK-EGFP (B and D), pETP42 (C), and pETKaKS.6 (D) vectors.
CaMV P6 Is a Nucleocytoplasmic Protein 939
Dow
nloaded from https://academ
ic.oup.com/plcell/article/17/3/927/6114488 by guest on 07 M
arch 2022

regulated during the viral cycle (in other words, that it occurs only
at specific stages). These would explain the difficulty we en-
countered to detect P6 in nuclei from infected plants.
The study of the behavior of P6 mutants in BY-2 cells revealed
that the nuclear export activity is associated with the Leu-rich
sequence (residues 11 to 20) at the N terminus of P6. Its in-
volvement in nuclear export was demonstrated by the incapacity
of P6 to exit the nucleus when Leu residues of the sequence were
point mutated and by the fact that mutated domain A of P6
accumulates in the nucleus, in contrast with the wild-type form.
The sequence EKI is not implicated in the export of P6 as
evidenced by the results of experiments performed with lepto-
mycin B, but it is an important determinant for the formation of
viroplasms. Concerning the involvement of the other residues of
the I1 invariant sequence, further investigations are necessary to
definitively answer this question. Interestingly, the NES is part of
the a-helix that is involved in P6 self-assembly and this fact might
explain why deletion of the first 90 nucleotides of the CaMV ORF
VI abolishes systemic infection and significantly reduces the
replication of the genome in single cells (Kobayashi and Hohn,
2003). The overlap between domains involved in P6 export and
self-assembly also raises the question of how these two activities
are regulated during the viral cycle. We hypothesize that P6
protein shuttling between the nuclear and cytoplasmic compart-
ments primarily involves a population of P6 monomers (or dimers)
that have escaped the aggregation process. Recent studies have
demonstrated that importins fulfill a dual function as a nuclear
import receptor and cytoplasmic chaperone for nuclear imported
proteins (Jakel et al., 2002). Such an antiaggregation mechanism
might also be involved for P6 molecules. This does not, however,
exclude the possibility that P6 could be incorporated into
viroplasms after their export from the nucleus.
The discovery that P6 is a nucleocytoplasmic shuttle protein
opens new prospects for understanding the mechanisms by
which this viral protein regulates the CaMV infectious cycle.
The function(s) of P6 in the nucleus can only be a matter for
speculation at present. P6 might have a role similar to the Rev
protein of HIV-1 (Pollard and Malim, 1998) in controlling export of
CaMV 35S RNA and its spliced versions because it also has the
capacity to bind single- and double-stranded RNA (De Tapia
et al., 1993; Cerritelli et al., 1998). The presence of P6 in the
nucleolus, where assembly of ribosomal subunits occurs, raises
the possibility that P6 might interact directly with ribosomes
before their export to render them competent for translation of
the CaMV polycistronic mRNA. The ribosomal proteins L18 and
L24, which interact with the mini-TAV (Leh et al., 2000) and RNA
binding domains (Park et al., 2001) of P6, respectively, could be
targets for P6 because they participate in the formation of the
60S subunit in the nucleolus (Andersen et al., 2002). Other
functions might also be associated with the nucleocytoplasmic
localization of P6 (i.e., inhibition of nonsense-mediated mRNA
decay to prevent degradation of the 35S RNA and its spliced
versions) (for a review, see Maquat and Carmichael, 2001). These
hypotheses are supported by the finding that P6 nuclear export is
mediated by the CRM-1 pathway (Kudo et al., 1998), which is
known to be specifically used for export of the ribosomal
subunits and of some cellular mRNAs (for a review, see Weis,
2002).
METHODS
Construction of Recombinant Plasmids
Recombinant plasmids were constructed by insertion of viral sequences
into the pET3a derivatives pETKaKS (Leh et al., 2000), pGEX-2TK
(Amersham-Pharmacia Biotech, Uppsala, Sweden), and pCK-EGFP
(Clontech, Palo Alto, CA). DNA fragments flanked by appropriate re-
striction cloning sites were generated using PCR; the oligonucleotides
used for PCR are listed in Table 1.
CaMV ORF VI and its derivatives were cloned either into the KpnI
and SacI sites or into the SacI site of the pETKaKS plasmid. Viral DNA
sequences were amplified from plasmid pMD324 containing the CaMV
Cabb-JI genome (Delseny and Hull, 1983) using two primers bearing at
their 59 termini KpnI and SacI sites, respectively, or SacI sites. The DNA
fragments were digested with the appropriate restriction enzymes and
introduced into pETKaKS cleaved with KpnI and SacI or with SacI. All
constructs were confirmed to be error free by sequencing. Expression of
the recombinant plasmids in Escherichia coli generates fusion proteins
containing at their N terminus the decapeptide Met-Arg-Arg-Ala-Ser-
Val-Gly-Ser-Gly-Thr, which can be phosphorylated in vitro by a protein
kinase from bovine heart muscle (the phosphorylation site is in bold-faced
type).
The DNA sequence encoding the N terminus of the CaMV P6 protein
(nucleotides 1 to 336 of ORF VI) was amplified from the pETKaKS.6
recombinant plasmid encompassing the complete ORF VI using primers
carrying an NcoI restriction site at their 59 end. The PCR fragment was
digested with NcoI and cloned into pETP42 (Lauber et al., 1998), which
had been cleaved with the same enzyme to produce pET-A:P42.
Plasmids pGST:A (ORF VI nucleotides 1 to 336), pGST:A1 (ORF VI
nucleotides 1 to 249), and pGST:A2 (ORF VI nucleotides 250 to 336),
coding for different regions of the P6 N terminus fused to GST, were
obtained by PCR amplification of different ORF VI sequences with
primers containing 59 terminal BamHI and EcoRI sites (Table 1). The
amplified DNA fragments were digested with the appropriate restriction
endonucleases and cloned into linearized pGEX-2TK.
The pCK-EGFP vector was used to construct the recombinant plas-
mids coding for fusion proteins between EGFP and wild-type CaMV P6 or
P6 mutants. The corresponding ORF VI sequences were amplified by
PCR from pMD324 using two primers carrying at their 59 ends BsrGI and
XbaI sites, respectively (Table 1).
Deletions and point mutations were introduced in EGFP:P6 and
EGFP:A by site-directed mutagenesis (Stratagene, La Jolla, CA). The
recombinant plasmids were amplified by PCR using pfu Turbo poly-
merase according to the manufacturer’s instructions and designed
internal oligonucleotides as primers (Table 1). The mixture was then
incubated with two units of DpnI for 2 h at 378C to destroy the
template. The 59 ends of PCR products were phosphorylated by T4
polynucleotide kinase in the presence of 1 mM ATP for subsequent
ligation. Error-free recombinant plasmids were identified by DNA
sequencing.
Production and Phosphorylation of Recombinant Proteins
E. coli BL21/DE3(pLysS) strain was transformed by electroporation with
pETKaKS and pGEX-2TK recombinant plasmids coding for full-length P6
or P6 mutants. Expression of the heterologous proteins was induced with
1 mM isopropyl b-thiogalactoside for 2 h once the bacterial culture had
reached the exponential phase. Bacteria were collected by centrifu-
gation at 5000g for 5 min, resuspended in HMK buffer (20 mM Tris-HCl,
pH 7.5, 100 mM NaCl, and 12 mM MgCl2), and lysed by sonication (three
pulses for 20 s at 50 W). After centrifugation at 12,000g for 10 min,
the supernatant was discarded and the inclusion bodies resuspended
in 500 mL of HMK buffer.
940 The Plant Cell
Dow
nloaded from https://academ
ic.oup.com/plcell/article/17/3/927/6114488 by guest on 07 M
arch 2022

Full-length P6 and the P6 fragments A, A1, and A2 expressed from
pETKaKS and pGEX-2TK vectors, respectively, were labeled in the
presence of [g-32P]ATP (3000 Ci/mmole) and bovine heart muscle protein
kinase (10 units) for 2 h at room temperature, according to the instructions
of the manufacturer (Sigma-Aldrich, St. Louis, MO). Excess radioactive
ATP was eliminated by filtration through a Sephadex G50 or G25 column
(Amersham-Pharmacia Biotech) depending on the molecular mass of the
labeled fusion proteins.
Protein Gel Blot Analysis
Proteins from recombinant bacteria were separated by SDS-PAGE
and electrophoretically transferred onto a nitrocellulose membrane
(Schleicher and Schuell, Dassel, Germany). The membranes were
blocked overnight in 5% nonfat dried milk in PBS buffer (140 mM NaCl,
2.7 mM KCl, and 8.1 mM Na2HPO4, pH 7.3) containing 0.1% Tween 20
and then incubated for 4 h at room temperature with specific rabbit or
sheep polyclonal antibodies raised against P6 (1:10,000 dilution) or GST
(1:5,000 dilution), respectively. The membranes were washed with PBS
buffer and treated with goat anti-rabbit IgG antibodies, respectively,
conjugated either to alkaline phosphatase or peroxidase, at the dilution
recommended by the manufacturer.
Far Protein Gel Blot Assays
A protein blotting overlay technique was used to detect interactions
between proteins. Proteins were resolved by SDS-PAGE and transferred
onto a nitrocellulose membrane. Membranes were washed several times
at 48C in HM buffer (10 mM Tris-HCl, pH 7.5, 100 mM NaCl, and 25 mM
MgCl2) containing 5% nonfat dried milk and incubated for 12 h at 48C with
gentle shaking in the same buffer containing the [32P]-labeled protein in
the overlay. After three washes in HM buffer, the membranes were dried
and radioactive complexes were detected by autoradiography.
Transient Expression in Tobacco BY-2 Cells
The CaMV P6 protein and its deleted versions fused to EGFP were
transiently expressed in BY-2 tobacco suspension cells (Nicotiana
tabacum cv Bright Yellow 2) maintained as described by Banjoko and
Trelease (1995). Cells were subcultured each 7 d and harvested 3 d
after medium renewal for biolistic transfection. Cells were filtered onto
Whatman disks and placed for 2 to 4 h on 0.8% agar MS media plates
supplemented with 0.1 M mannitol and 0.1 M sorbitol. Particle prepara-
tion and bombardment assays were performed as described by Hunold
et al. (1995) with modifications: 2 mg of 1.1 mm tungsten particles (Bio-
Rad, Hercules, CA) were immersed in 1 mL of absolute alcohol for 20 min.
Dried particles were then successively mixed with 10 mg of recombinant
plasmid DNA (pCK-EGFP vector) supplemented with 18% glycerol,
0.75 M CaCl2, and 90 mM spermidine in a final volume of 90 mL. The fir-
ing distance was 11 cm and the helium pressure was 7 bars. After bom-
bardment, cells were transferred to 0.8% agar MS media plates and
incubated in the dark at 288C. BY-2 transfected cells were collected under
HBO binoculars (excitation/emission wavelength 488/505 to 545 nm) 20 h
after bombardment and cultured in MS liquid medium before further
treatment and/or CLSM observations.
Virus and Host Plant
Turnips (Brassica rapa cv Just Right F1 hybrid, provided by Takii and
Co., Kyoto, Japan) were mechanically inoculated at the four leaf stage
(Jacquot et al., 1998) with CaMV Cabb-JI and grown in a greenhouse
at 228C for 5 weeks before preparation of protoplasts from infected
leaves.
Isolation of Turnip Nuclei
Protoplasts prepared from CaMV-infected and noninfected turnip leaves
(Kobayashi et al., 1998) were used to isolate nuclei. Approximately 8 3
106 protoplasts were washed twice in nuclei buffer (250 mM sucrose,
25 mM Mes, 0.5 mM EDTA, 1 mM MgCl2, 1 mM EGTA, pH 5.5, and
a complete cocktail of protease inhibitors [Roche, Indianapolis, IN]). After
centrifugation for 5 min at 100g, the protoplasts were resuspended in
50 mL of cold nuclei buffer containing 1 mM DTT, 0.025% Nonidet P-40,
and 1 mM phenylmethylsulfonyl fluoride and shaken slowly for 20 min at
48C. Nuclei were isolated by filtering the suspension through 50-mm
mesh nylon and collected at 48C by centrifugation for 5 min at 550g. They
were resuspended in the nuclei buffer and centrifuged at 48C through a
discontinuous gradient composed of 18% Ficoll and 85% Percoll for
15 min at 8000g. The band containing the nuclei, located between the
Ficoll and Percoll layers, was diluted threefold with 10 mM Pipes-KOH,
pH 7.0, and centrifuged at 600g during 10 min.
Fluorescence Analysis
Fluorescent BY-2 tobacco cells, transfected with EGFP or a protein fused
to EGFP, were observed between a slide and cover slip with a Zeiss
LSM510 confocal microscope (Jena, Germany). EGFP was viewed by
excitation at 488 nm with an argon laser using an appropriate emission
filter to collect the green signal from the optical section. Fluorescent cells
were also observed under the same conditions after incubation for 8 h at
248C with gentle shaking in the BY-2 cell culture medium containing
100 nM leptomycin B.
For immunofluorescence studies, protoplasts or nuclei prepared as
described above were harvested and fixed for 15 min with gentle shaking
in protoplast or nuclei-specific medium, respectively, containing 4%
glutaraldehyde. Thereafter, they were washed three times with the
appropriate medium, once with the medium diluted volume to volume
with PBS, then again with PBS and finally resuspended in PBS buffer. A
sample of protoplasts or nuclei was mounted on a poly-L-Lys–coated
cover slip, allowed to settle for 1 h at room temperature, and then treated
overnight at 48C in a 0.1% sodium borohydride solution. Protoplasts and
nuclei were incubated for 1 h in a blocking solution (5% acetylated BSA
[Aurion, Wageningen, The Netherlands], 5% normal goat serum, and
0.1% cold water fish skin gelatin prepared in PBS) and then overnight with
the polyclonal anti-P6 antibodies. After six washes with 0.1% BSAc in
PBS, protoplasts or nuclei were treated with goat anti-rabbit antibodies
coupled to Alexa 488 (Molecular Probes, Eugene, OR), respectively, for
12 h. After removal of excess secondary antibodies by six washes in 0.1%
BSAc in PBS, the protoplasts and nuclei were subsequently examined
with a Zeiss LSM510 confocal microscope.
ACKNOWLEDGMENTS
We thank Marc Bergdoll for the three-dimensional modeling of P6 and
John Stanley for providing us with the CaMV Cabb-JI genome se-
quence. We are most grateful to Christiane Garaud and Jerome
Mutterer for advice on CLSM and to Ken Richards for critical reading
of the manuscript. The Inter-Institute Confocal Microscopy Platform was
cofinanced by the Region Alsace, Centre National de la Recherche
Scientifique, the Universite Louis Pasteur, and the Association de la
Recherche pour le Cancer. This work was supported by the Centre
National de la Recherche Scientifique and by the Universite Louis
Pasteur of Strasbourg.
Received November 1, 2004; accepted December 9, 2004.
CaMV P6 Is a Nucleocytoplasmic Protein 941
Dow
nloaded from https://academ
ic.oup.com/plcell/article/17/3/927/6114488 by guest on 07 M
arch 2022

REFERENCES
Agama, K., Beach, S., Schoelz, J., and Leisner, S.M. (2002). The 59
third of Cauliflower mosaic virus Gene VI conditions resistance
breakage in Arabidopsis ecotype Tsu-0. Virology 92, 190–196.
Andersen, J.S., Lyon, C.E., Fox, A.H., Leung, A.K.L., Lam, Y.W.,
Steen, H., Mann, M., and Lamond, A.I. (2002). Directed proteomic
analysis of the human nucleolus. Curr. Biol. 12, 1–11.
Banjoko, A., and Trelease, R.N. (1995). Development and application
of an in vivo plant peroxisome import system. Plant Physiol. 107,
1201–1208.
Berger, B. (1995). Algorithms for protein structural motif recognition.
J. Comput. Biol. 2, 125–138.
Blobel, G., and Dobberstein, B. (1975). Transfer of proteins across
membranes. I. Presence of proteolytically processed and unpro-
cessed nascent immunoglobulin light chains on membrane-bound
ribosomes of murine myeloma. J. Cell Biol. 67, 835–851.
Bureau, M., Leh, V., Haas, M., Geldreich, A., Ryabova, L., Yot, P.,
and Keller, M. (2004). P6 protein of Cauliflower mosaic virus, a trans-
lational reinitiator, interacts with ribosomal protein L13 from Arabi-
dopsis thaliana. J. Gen. Virol. 85, 3765–3775.
Cecchini, E., Gong, Z., Geri, C., Covey, S.N., and Milner, J.J. (1997).
Transgenic Arabidopsis lines expressing gene VI from Cauliflower
mosaic virus variants exhibit a range of symptom-like phenotypes and
accumulate inclusion bodies. Mol. Plant Microbe Interact. 10, 1094–
1120.
Cerritelli, S., Fedoroff, O., Reid, B., and Crouch, R. (1998). A common
40 amino acid motif in eukaryotic RNases H1 and caulimovirus ORF VI
proteins binds to duplex RNAs. Nucleic Acids Res. 26, 1834–1840.
Cole, A.B., Kiraly, L., Ross, K., and Schoelz, J.E. (2001). Uncoupling
resistance from cell death in the hypersensitive response of Nicotiana
species to Cauliflower mosaic virus infection. Mol. Plant Microbe
Interact. 14, 31–41.
Daubert, S.D., and Routh, G. (1990). Point mutations in Cauliflower
mosaic virus gene VI confer host-specific symptom changes. Mol.
Plant Microbe Interact. 3, 341–345.
Daubert, S.D., Schoelz, J., Debao, L., and Shepherd, R.J. (1984).
Expression of disease symptoms in Cauliflower mosaic virus genomic
hybrids. J. Mol. Appl. Genet. 2, 537–547.
Delseny, M., and Hull, R. (1983). Isolation and characterization of
faithful altered clones of the genomes of Cauliflower mosaic virus
isolates Cabb B-JI, CM4-184 and Bari. Plasmid 9, 31–41.
De Tapia, M., Himmelbach, A., and Hohn, T. (1993). Molecular
dissection of the Cauliflower mosaic virus translation transactivator.
EMBO J. 12, 3305–3314.
Drucker, M., Froissart, R., Hebrard, E., Uzest, M., Ravallec, M.,
Esperandieu, P., Mani, J.-L., Pugniere, M., Roquet, F., Fereres, A.,
and Blanc, S. (2002). Intracellular distribution of viral gene products
regulates a complex mechanism of Cauliflower mosaic virus acquisi-
tion by its aphid vector. Proc. Natl. Acad. Sci. USA 99, 2422–2427.
Espinoza, A.M., Medina, V., Hull, R., and Markham, P.G. (1991).
Cauliflower mosaic virus gene II products forms distinct inclusion
bodies in infected plant cells. Virology 185, 337–344.
Fabbretti, E., Afrikanova, I., Vascotto, F., and Burrone, O.R.
(1999). Two non-structural rotavirus proteins, NSP2 and NSP5, form
viroplasm-like structures in vivo. J. Gen. Virol. 80, 333–339.
Fornerod, M., Ohno, M., Yoshida, M., and Mattaj, I.W. (1997). CRM1
is an export receptor for leucine-rich nuclear exports signals. Cell 90,
1051–1060.
Geri, C., Cecchini, E., Giannakou, M., Covey, S., and Milner, J.
(1999). Altered patterns of gene expression in Arabidopsis elicited by
Cauliflower mosaic virus (CaMV) infection and by a CaMV gene VI
transgene. Mol. Plant Microbe Interact. 12, 377–384.
Gorlich, D., and Kutay, U. (1999). Transport between the cell nucleus
and the cytoplasm. Annu. Rev. Cell Dev. Biol. 15, 607–660.
Haas, M., Bureau, M., Geldreich, A., Yot, P., and Keller, M. (2002).
Cauliflower mosaic virus: Still in the news. Mol. Plant Pathol. 3,
419–429.
Haas, M., Leh, V., Yot, P., and Keller, M. (2000). Caracterisation du
domaine de la proteine P6 du Virus de la mosaıque du chou-fleur
implique dans la formation des viroplasmes. Virologie 4, 165.
Heath, C.M., Windsor, M., and Wileman, T. (2001). Aggresomes
resemble sites specialized for virus assembly. J. Cell Biol. 153,
449–455.
Himmelbach, A., Chapdelaine, Y., and Hohn, T. (1996). Interaction
between Cauliflower mosaic virus inclusion body protein and capsid
protein: Implication for viral assembly. Virology 217, 147–157.
Hunold, R., Burrus, M., Bronner, R., Duret, J.P., and Hahne, G.
(1995). Transient gene expression in sunflower (Helianthus annuus L.)
following microprojectile bombardment. Plant Sci. 105, 95–109.
Jacquot, E., Geldreich, A., Keller, M., and Yot, P. (1998). Mapping
regions of the Cauliflower mosaic virus ORF III product required for
infectivity. Virology 242, 395–402.
Jakel, S., Mingot, J.-M., Schwarzmaier, P., Hartmann, E., and
Gorlich, D. (2002). Importins fulfil a dual function as nuclear import
receptors and cytoplasmic chaperones for exposed basic domains.
EMBO J. 21, 377–386.
Johnston, J.A., Ward, C.L., and Kopito, R.R. (1998). Aggresomes:
A cellular response to misfolded proteins. J. Cell Biol. 143, 1883–
1898.
Kiss-Laszlo, Z., Blanc, S., and Hohn, T. (1995). Splicing of Cauliflower
mosaic virus 35S RNA is essential for viral infectivity. EMBO J. 14,
3552–3562.
Kobayashi, K., and Hohn, T. (2003). Dissection of cauliflower mosaic
virus transactivator/viroplasmin reveals distinct essential functions in
basic virus replication. J. Virol. 77, 8577–8583.
Kobayashi, K., Nakayashiki, H., Tsuge, S., Mise, K., and Furusawa, I.
(1998). Accumulation kinetics of viral gene products in Cauliflower
mosaic virus-infected turnip protoplast. Microbiol. Immunol. 42, 65–69.
Kopito, R.R. (2000). Aggresomes, inclusion bodies and protein aggre-
gation. Trends Cell Biol. 10, 524–530.
Kudo, N., Wolff, B., Sekimoto, T., Schreiner, E.P., Yoneda, Y.,
Yanagida, M., Horinouchi, S., and Yoshida, M. (1998). Leptomycin
B inhibition of signal-mediated nuclear export by direct binding to
CRM1. Exp. Cell Res. 1, 540–547.
Lamond, A.I., and Spector, D.L. (2003). Nuclear speckles: A model for
nuclear organelles. Mol. Cell. Biol. 4, 605–611.
Lauber, E., Bleykasten-Grosshans, C., Erhardt, M., Bouzoubaa, S.,
Jonard, G., Richards, K., and Guilley, H. (1998). Cell-to-cell move-
ment of Beet necrotic yellow vein virus: I. Heterologous complemen-
tation experiments provide evidence for specific interactions among
the triple gene block proteins. Mol. Plant Microbe Interact. 11,
618–625.
Leh, V. (1999). Etude des Interactions entre Proteines Engageant les
Produits des ORFIII et ORFVI du Virus de la Mosaıque du Chou-Fleur.
PhD dissertation (Strasbourg, France: Universite Louis Pasteur).
Leh, V., Yot, P., and Keller, M. (2000). The Cauliflower mosaic virus
translational transactivator interacts with the 60S ribosomal subunit
protein L18 of Arabidopsis thaliana. Virology 266, 1–7.
Li, Y., and Leisner, S.C. (2002). Multiple domains within the Cauliflower
mosaic virus gene VI product interact with the full-length protein. Mol.
Plant Microbe Interact. 15, 1050–1057.
Lupas, A. (1997). Predicting coiled-coil regions in proteins. Curr. Opin.
Struct. Biol. 7, 388–393.
Maquat, L., and Carmichael, G. (2001). Quality control of mRNA
function. Cell 104, 173–176.
942 The Plant Cell
Dow
nloaded from https://academ
ic.oup.com/plcell/article/17/3/927/6114488 by guest on 07 M
arch 2022

Maule, A.J., Harker, C.L., and Wilson, I.G. (1989). The pattern of
accumulation of Cauliflower mosaic virus specific products in infected
turnips. Virology 169, 436–446.
Mazzolini, L., Dabos, P., Constantin, S., and Yot, P. (1989). Further
evidence that viroplasms are the site of Cauliflower mosaic virus
genome replication by reverse transcription during viral infection.
J. Gen. Virol. 70, 3439–3449.
Ochs, R.L., Stein, T.W., and Tan, E.M.M. (1994). Coiled bodies in the
nucleolus of breast cancer cells. J. Cell Sci. 107, 385–399.
Palanichelvam, K., Cole, A.B., Shababi, M., and Schoelz, J.E. (2000).
Agroinfiltration of Cauliflower mosaic virus gene VI elicits hypersen-
sitive response in Nicotiana species. Mol. Plant Microbe Interact. 13,
1275–1279.
Park, H.-S., Himmelbach, A., Browning, K.S., Hohn, T., and Ryabova,
L.A. (2001). A plant viral ‘‘reinitiation’’ factor interacts with the host
translational machinery. Cell 106, 723–733.
Pollard, V.W., and Malim, M.H. (1998). The HIV-1 Rev protein. Annu.
Rev. Microbiol. 52, 491–532.
Rothnie, H., Chapdelaine, Y., and Hohn, T. (1994). Pararetroviruses
and retroviruses: A comparative review of viral structure and gene
expression strategies. Adv. Virus Res. 44, 1–67.
Ryabova, L.A., Pooggin, M.M., and Hohn, T. (2002). Viral strategies of
translation initiation: Ribosomal shunt and reinitiation. Prog. Nucleic
Acid Res. Mol. Biol 272, 1–39.
Schoelz, J.E., Shepherd, R.J., and Daubert, S.D. (1986). Gene VI of
CaMV encodes a host range determinant. Mol. Cell. Biol. 6, 2632–
2637.
Shalla, T.A., Shepherd, R.J., and Petersen, L.J. (1980). Comparative
cytology of nine isolates of Cauliflower mosaic virus. Virology 102,
381–388.
Ward, B.M., and Lazarowitz, S.G. (1999). Nuclear export in plants. Use
of geminivirus movement proteins for a cell-based export assay. Plant
Cell 11, 1267–1276.
Weis, K. (2002). Nucleocytoplasmic transport: Cargo trafficking across
the border. Curr. Opin. Cell Biol. 14, 328–335.
Xiong, C., Balazs, E., Lebeurier, G., Hindenlang, C., Stoeckel, M.E.,
and Porte, A. (1982). Comparative cytology of two isolates of
Cauliflower mosaic virus. J. Gen. Virol. 61, 75–81.
Zijlstra, C., Scharer-Hernandez, N., Gal, S., and Hohn, T. (1996).
Arabidopsis thaliana expressing the Cauliflower mosaic virus ORF VI
transgene has a late flowering phenotype. Virus Genes 13, 5–17.
CaMV P6 Is a Nucleocytoplasmic Protein 943
Dow
nloaded from https://academ
ic.oup.com/plcell/article/17/3/927/6114488 by guest on 07 M
arch 2022
Copyright © 2022 FDOKUMEN




















