
The novel Cladosporium fulvum lysin motif effector Ecp6 is a virulence factor with orthologues in...
18
The novel Cladosporium fulvum lysin motif effector Ecp6 is a virulence factor with orthologues in other fungal species Melvin D. Bolton, 1,2†‡ H. Peter van Esse, 1,3† Jack H. Vossen, 1†§ Ronnie de Jonge, 1 Ioannis Stergiopoulos, 1 Iris J. E. Stulemeijer, 1 Grardy C. M. van den Berg, 1 Orlando Borrás-Hidalgo, 1,5 Henk L. Dekker, 4 Chris G. de Koster, 4 Pierre J. G. M. de Wit, 1,3 Matthieu H. A. J. Joosten 1 and Bart P. H. J. Thomma 1,3 * 1 Laboratory of Phytopathology, Wageningen University, Binnenhaven 5, 6709 PD Wageningen, the Netherlands. 2 Department of Plant Pathology, North Dakota State University, Fargo, ND 58105-5012, USA. 3 Centre for BioSystems Genomics (CBSG), PO Box 98, 6700 AB Wageningen, the Netherlands. 4 Swammerdam Institute for Life Sciences, Mass Spectrometry, Nieuwe Achtergracht 166, 1018 WV Amsterdam, the Netherlands. 5 Centre for Genetic Engineering and Biotechnology, PO Box 6162, Havana, 10600, Cuba. Summary During tomato leaf colonization, the biotrophic fungus Cladosporium fulvum secretes several effec- tor proteins into the apoplast. Eight effectors have previously been characterized and show no signifi- cant homology to each other or to other fungal genes. To discover novel C. fulvum effectors that might play a role in virulence, we utilized two-dimensional polyacrylamide gel electrophoresis (2D-PAGE) to visualize proteins secreted during C. fulvum–tomato interactions. Three novel C. fulvum proteins were identified: CfPhiA, Ecp6 and Ecp7. CfPhiA shows homology to proteins found on fungal sporogenous cells called phialides. Ecp6 contains lysin motifs (LysM domains) that are recognized as carbohydrate- binding modules. Ecp7 encodes a small, cysteine-rich protein with no homology to known proteins. Heter- ologous expression of Ecp6 significantly increased the virulence of the vascular pathogen Fusarium oxysporum on tomato. Furthermore, by RNA interfer- ence (RNAi)-mediated gene silencing we demonstrate that Ecp6 is instrumental for C. fulvum virulence on tomato. Hardly any allelic variation was observed in the Ecp6 coding region of a worldwide collection of C. fulvum strains. Although none of the C. fulvum effectors identified so far have obvious orthologues in other organisms, conserved Ecp6 orthologues were identified in various fungal species. Homology- based modelling suggests that the LysM domains of C. fulvum Ecp6 may be involved in chitin binding. Introduction Cladosporium fulvum (syn. Passalora fulva) is a biotrophic pathogen that causes leaf mold of tomato (Lycopersicon esculentum Mill. syn. Solanum esculentum) (Thomma et al., 2005). After germination of conidia, the fungus pro- duces runner hyphae that penetrate stomata predomi- nantly on the lower side of the leaf. Once inside the apoplast, C. fulvum does not penetrate host cells or develop haustoria but remains confined to the intercellular space between plant mesophyll cells (de Wit, 1977). Despite much research on the C. fulvum–tomato interac- tion, the molecular components that C. fulvum utilizes for infection and colonization are largely unknown (Thomma et al., 2005). Plant pathogens secrete molecules called effectors that contribute to the establishment of disease to their hosts. As the complete set of effectors of a potential pathogen determines the outcome of the interaction with a possible host, it is important to make an inventory of this effector catalogue. Many plant pathogenic bacteria inject effector proteins into the cytoplasm of host cells by means of the type III secretion system (TTSS) to subvert host cellular physiology to the bacterium’s advantage (Grant et al., 2006; Tang et al., 2006). This process is orchestrated by specific cis-elements in the promoters of genes encoding type III effector proteins, a feature that has been exploited to identify such effectors in genome-wide functional screens (Guttman et al., 2002; Chang et al., 2005). In a similar way, the discovery that several oomycete effector molecules enter the host cytoplasm through a specific host targeting RXLR-DEER motif (Whisson et al., 2007) Accepted 21 April, 2008. *For correspondence. E-mail bart.thomma@ wur.nl; Tel. (+31) 317 484536; Fax (+31) 317 483412. † These authors contributed equally. ‡ Present address: USDA – ARS, Northern Crop Science Laboratory, Fargo, ND 58105-5677, USA. § Present address: Plant Research International BV, PO Box 16, 6700 AA Wageningen, the Netherlands. Molecular Microbiology (2008) doi:10.1111/j.1365-2958.2008.06270.x © 2008 The Authors Journal compilation © 2008 Blackwell Publishing Ltd
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of The novel Cladosporium fulvum lysin motif effector Ecp6 is a virulence factor with orthologues in...

The novel Cladosporium fulvum lysin motif effector Ecp6 is avirulence factor with orthologues in other fungal species
Melvin D. Bolton,1,2†‡ H. Peter van Esse,1,3†
Jack H. Vossen,1†§ Ronnie de Jonge,1
Ioannis Stergiopoulos,1 Iris J. E. Stulemeijer,1
Grardy C. M. van den Berg,1
Orlando Borrás-Hidalgo,1,5 Henk L. Dekker,4
Chris G. de Koster,4 Pierre J. G. M. de Wit,1,3
Matthieu H. A. J. Joosten1 andBart P. H. J. Thomma1,3*1Laboratory of Phytopathology, Wageningen University,Binnenhaven 5, 6709 PD Wageningen, the Netherlands.2Department of Plant Pathology, North Dakota StateUniversity, Fargo, ND 58105-5012, USA.3Centre for BioSystems Genomics (CBSG), PO Box 98,6700 AB Wageningen, the Netherlands.4Swammerdam Institute for Life Sciences, MassSpectrometry, Nieuwe Achtergracht 166, 1018 WVAmsterdam, the Netherlands.5Centre for Genetic Engineering and Biotechnology, POBox 6162, Havana, 10600, Cuba.
Summary
During tomato leaf colonization, the biotrophicfungus Cladosporium fulvum secretes several effec-tor proteins into the apoplast. Eight effectors havepreviously been characterized and show no signifi-cant homology to each other or to other fungal genes.To discover novel C. fulvum effectors that might playa role in virulence, we utilized two-dimensionalpolyacrylamide gel electrophoresis (2D-PAGE) tovisualize proteins secreted during C. fulvum–tomatointeractions. Three novel C. fulvum proteins wereidentified: CfPhiA, Ecp6 and Ecp7. CfPhiA showshomology to proteins found on fungal sporogenouscells called phialides. Ecp6 contains lysin motifs(LysM domains) that are recognized as carbohydrate-binding modules. Ecp7 encodes a small, cysteine-richprotein with no homology to known proteins. Heter-ologous expression of Ecp6 significantly increased
the virulence of the vascular pathogen Fusariumoxysporum on tomato. Furthermore, by RNA interfer-ence (RNAi)-mediated gene silencing we demonstratethat Ecp6 is instrumental for C. fulvum virulence ontomato. Hardly any allelic variation was observed inthe Ecp6 coding region of a worldwide collection ofC. fulvum strains. Although none of the C. fulvumeffectors identified so far have obvious orthologuesin other organisms, conserved Ecp6 orthologueswere identified in various fungal species. Homology-based modelling suggests that the LysM domains ofC. fulvum Ecp6 may be involved in chitin binding.
Introduction
Cladosporium fulvum (syn. Passalora fulva) is a biotrophicpathogen that causes leaf mold of tomato (Lycopersiconesculentum Mill. syn. Solanum esculentum) (Thommaet al., 2005). After germination of conidia, the fungus pro-duces runner hyphae that penetrate stomata predomi-nantly on the lower side of the leaf. Once inside theapoplast, C. fulvum does not penetrate host cells ordevelop haustoria but remains confined to the intercellularspace between plant mesophyll cells (de Wit, 1977).Despite much research on the C. fulvum–tomato interac-tion, the molecular components that C. fulvum utilizes forinfection and colonization are largely unknown (Thommaet al., 2005).
Plant pathogens secrete molecules called effectors thatcontribute to the establishment of disease to their hosts.As the complete set of effectors of a potential pathogendetermines the outcome of the interaction with a possiblehost, it is important to make an inventory of this effectorcatalogue. Many plant pathogenic bacteria inject effectorproteins into the cytoplasm of host cells by means of thetype III secretion system (TTSS) to subvert host cellularphysiology to the bacterium’s advantage (Grant et al.,2006; Tang et al., 2006). This process is orchestrated byspecific cis-elements in the promoters of genes encodingtype III effector proteins, a feature that has been exploitedto identify such effectors in genome-wide functionalscreens (Guttman et al., 2002; Chang et al., 2005). In asimilar way, the discovery that several oomycete effectormolecules enter the host cytoplasm through a specifichost targeting RXLR-DEER motif (Whisson et al., 2007)
Accepted 21 April, 2008. *For correspondence. E-mail [email protected]; Tel. (+31) 317 484536; Fax (+31) 317 483412. †These authorscontributed equally. ‡Present address: USDA – ARS, Northern CropScience Laboratory, Fargo, ND 58105-5677, USA. §Present address:Plant Research International BV, PO Box 16, 6700 AA Wageningen,the Netherlands.
Molecular Microbiology (2008) � doi:10.1111/j.1365-2958.2008.06270.x
© 2008 The AuthorsJournal compilation © 2008 Blackwell Publishing Ltd

has been exploited to identify oomycete effectorcatalogues. It is currently predicted that the genomes ofoomycete plant pathogens contain hundreds of sucheffectors (Whisson et al., 2007; Jiang et al., 2008).
The effectors of extracellularly growing plant patho-genic fungi are usually very rich in cysteine residuesinvolved in disulphide bridges, thereby protecting themagainst proteinases that occur frequently in apoplasticspaces of their host plants (Joosten and de Wit, 1999;Rep, 2005; Thomma et al., 2005; Kamoun, 2006). Atpresent, relatively few whole-genome sequences of plantpathogenic fungi are available when compared with bac-teria (Xu et al., 2006). As most effector proteins fromextracellular pathogenic fungi are secreted, apoplasticextract from colonized plants is an important resource forthe discovery of molecular factors important in severalplant diseases (Joosten and de Wit, 1999; Rep, 2005;Thomma et al., 2005; Kamoun, 2006).
As C. fulvum is restricted to the tomato apoplast dur-ing colonization, all communication and exchange ofmolecular components between C. fulvum and its hostoccurs in the apoplastic space. So far, analysis of theprotein composition of the apoplastic space of C. fulvum-infected tomato leaves has mainly been focused on iden-tification of race-specific avirulence proteins (Avrs) thatare secreted by the fungus during infection and invokea resistance response in tomato genotypes carryingcognate C. fulvum resistance (Cf ) genes (van Kan et al.,1991; Joosten et al., 1994; Luderer et al., 2002; Wester-ink et al., 2004). In addition, a number of extracellularproteins (Ecps) secreted during infection by all strains ofC. fulvum have been identified (Joosten and de Wit,1988; Wubben et al., 1994; Laugé et al., 1998; 2000;Haanstra et al., 1999; 2000). Like Avrs, Ecps induce aresistance response in tomato accessions carrying asyet unidentified Cf-Ecp resistance genes. Collectively,the Avrs and Ecps are the secreted effecor proteins. Intotal, eight C. fulvum secreted effector proteins havebeen characterized in detail and their correspondinggenes have been cloned (van Kan et al., 1991; van denAckerveken et al., 1993; Joosten et al., 1994; Laugéet al., 2000; Luderer et al., 2002; Westerink et al., 2004;Thomma et al., 2005). All these secreted effector pro-teins are relatively small (ranging from 3 to 15 kDa) andcontain a high and even number of cysteine residuesthat appear to be involved in disulphide bridge formation(Kooman-Gersmann et al., 1997; van den Burg et al.,2003). These bridges provide a compact tertiary struc-ture that contributes to stability and activity of thesecreted effector proteins in the protease-rich tomatoapoplast (Joosten et al., 1997; Tornero et al., 1997;Jorda et al., 1999; Krüger et al., 2002; van Esse et al.,2006). All of these effector proteins elicit a defenceresponse in plants carrying the cognate Cf genes in a
‘gene-for-gene’ manner (Kruijt et al., 2005). The obser-vation that these proteins are maintained within thepopulation together with their abundance and specificaccumulation during pathogenesis suggests that theseproteins play an important role in fungal virulence(Thomma et al., 2005). Indeed, transformants containinggene knockouts of either Ecp1 or Ecp2 were shown tohave impaired aggressiveness in mature tomato plants(Laugé et al., 1997). Recent data show that also Avr2 isa genuine virulence factor of C. fulvum (H.P. van Esseet al., submitted). It has previously been shown that Avr2interacts with, and inhibits, the tomato cysteine proteaseRcr3 which, in compliance with the guard hypothesis,is required for Cf-2-mediated immunity (Rooney et al.,2005). In compatible interactions, however, Avr2 inhibitsseveral additional extracellular host cysteine proteasesthat are required for host basal defence (H.P. van Esseet al., submitted). Protection of chitin, a major constituentof fungal cell walls, against plant chitinases by the chitin-binding Avr4 effector protein (van den Burg et al., 2006)was recently shown to contribute to C. fulvum virulence(van Esse et al., 2007).
In addition to the secreted effectors, Nrf1 and Aox havebeen identified as virulence factors of C. fulvum (Segerset al., 2001; Thomma et al., 2006). The nitrogen responseregulator Nrf1 was found to control expression of Avr9 butno other known Avr or Ecp genes in planta (Pérez-Garcíaet al., 2001; Thomma et al., 2006). Interestingly, dis-ruption of the Nrf1 gene reduces C. fulvum virulencesignificantly (Thomma et al., 2006). Similarly, targeteddisruption of Aox1, a starvation-induced acetaldehydedehydrogenase, caused decreased colonization of thehost plant (Segers et al., 2001).
To visualize extracellular proteins present in compatibleand incompatible C. fulvum–tomato interactions, theapoplastic proteome of C. fulvum-infected tomato wasanalysed using two-dimensional polyacrylamide gelelectrophoresis (2D-PAGE). Several proteins that areproduced during infection were identified by mass spec-trometry (MS) and coding sequences for three novelC. fulvum proteins were obtained by reverse geneticsemploying PCR with degenerate primers based onMS/MS sequence tags and N-terminal sequencing. Weused RNA interference (RNAi) for functional analysis inC. fulvum and demonstrate that one of the identifiedsecreted effectors is crucial for C. fulvum virulence.
Results
Quantification of C. fulvum biomass in infectedtomato leaves
In a compatible interaction involving the susceptibleMoneyMaker Cf-0 (MM-Cf-0) tomato cultivar which lacks
2 M. D. Bolton et al. �
© 2008 The AuthorsJournal compilation © 2008 Blackwell Publishing Ltd, Molecular Microbiology
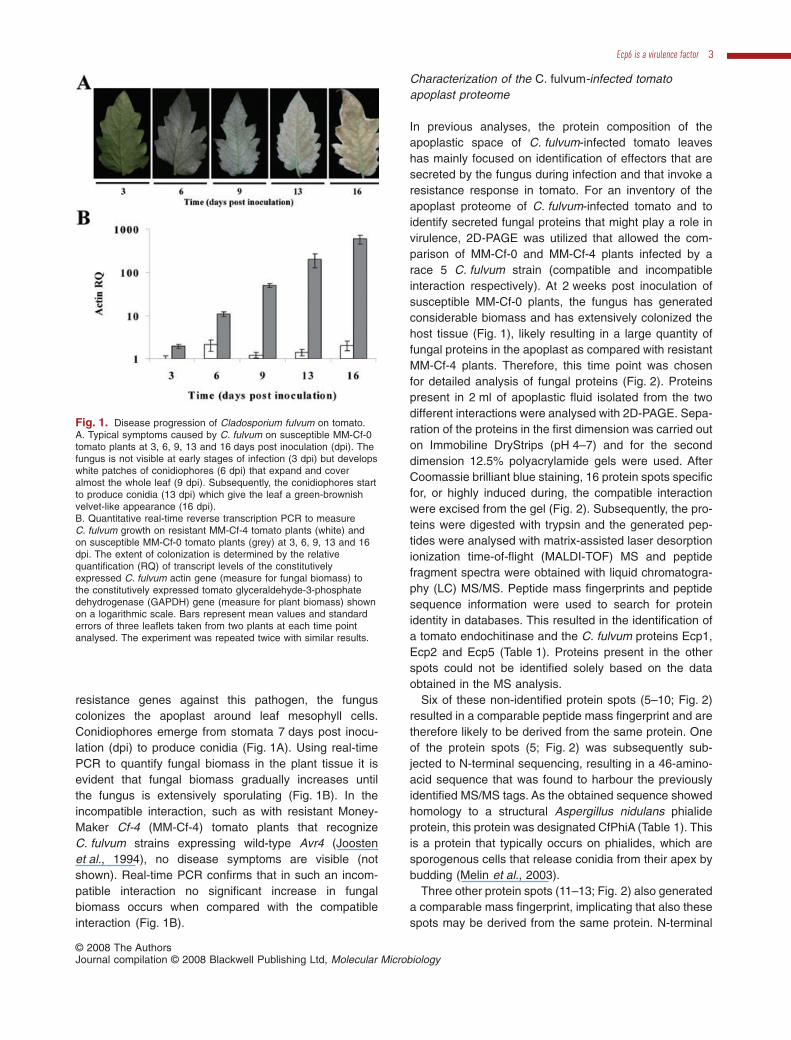
resistance genes against this pathogen, the funguscolonizes the apoplast around leaf mesophyll cells.Conidiophores emerge from stomata 7 days post inocu-lation (dpi) to produce conidia (Fig. 1A). Using real-timePCR to quantify fungal biomass in the plant tissue it isevident that fungal biomass gradually increases untilthe fungus is extensively sporulating (Fig. 1B). In theincompatible interaction, such as with resistant Money-Maker Cf-4 (MM-Cf-4) tomato plants that recognizeC. fulvum strains expressing wild-type Avr4 (Joostenet al., 1994), no disease symptoms are visible (notshown). Real-time PCR confirms that in such an incom-patible interaction no significant increase in fungalbiomass occurs when compared with the compatibleinteraction (Fig. 1B).
Characterization of the C. fulvum-infected tomatoapoplast proteome
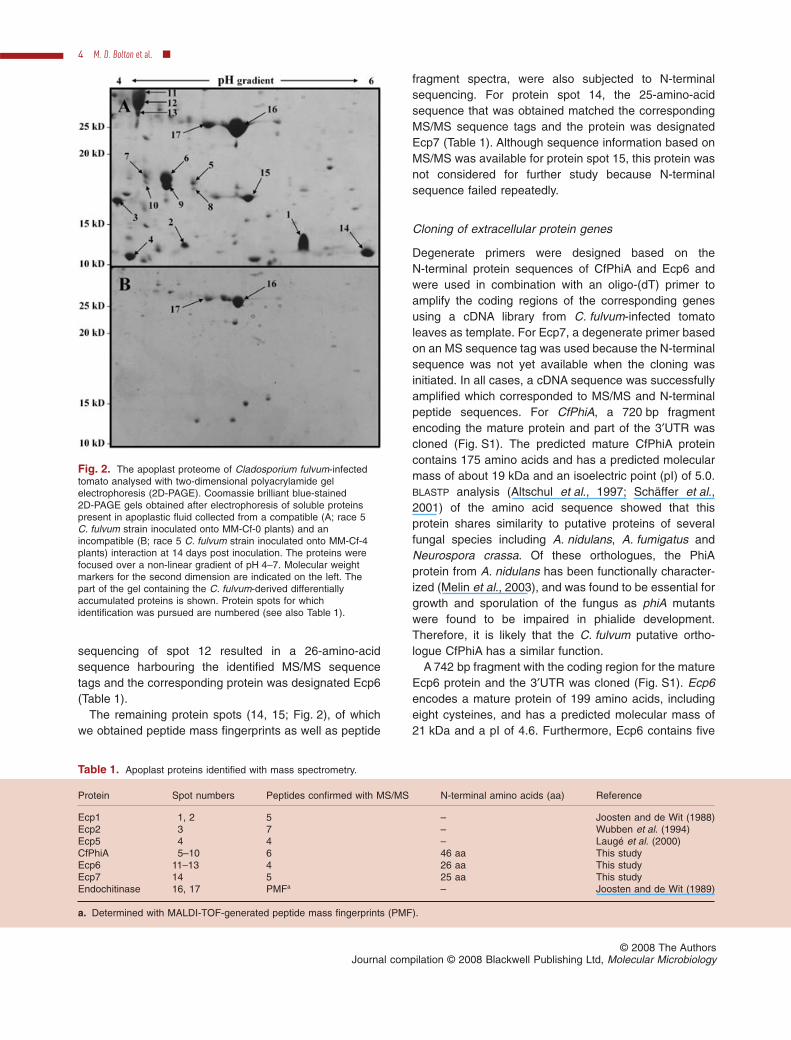
In previous analyses, the protein composition of theapoplastic space of C. fulvum-infected tomato leaveshas mainly focused on identification of effectors that aresecreted by the fungus during infection and that invoke aresistance response in tomato. For an inventory of theapoplast proteome of C. fulvum-infected tomato and toidentify secreted fungal proteins that might play a role invirulence, 2D-PAGE was utilized that allowed the com-parison of MM-Cf-0 and MM-Cf-4 plants infected by arace 5 C. fulvum strain (compatible and incompatibleinteraction respectively). At 2 weeks post inoculation ofsusceptible MM-Cf-0 plants, the fungus has generatedconsiderable biomass and has extensively colonized thehost tissue (Fig. 1), likely resulting in a large quantity offungal proteins in the apoplast as compared with resistantMM-Cf-4 plants. Therefore, this time point was chosenfor detailed analysis of fungal proteins (Fig. 2). Proteinspresent in 2 ml of apoplastic fluid isolated from the twodifferent interactions were analysed with 2D-PAGE. Sepa-ration of the proteins in the first dimension was carried outon Immobiline DryStrips (pH 4–7) and for the seconddimension 12.5% polyacrylamide gels were used. AfterCoomassie brilliant blue staining, 16 protein spots specificfor, or highly induced during, the compatible interactionwere excised from the gel (Fig. 2). Subsequently, the pro-teins were digested with trypsin and the generated pep-tides were analysed with matrix-assisted laser desorptionionization time-of-flight (MALDI-TOF) MS and peptidefragment spectra were obtained with liquid chromatogra-phy (LC) MS/MS. Peptide mass fingerprints and peptidesequence information were used to search for proteinidentity in databases. This resulted in the identification ofa tomato endochitinase and the C. fulvum proteins Ecp1,Ecp2 and Ecp5 (Table 1). Proteins present in the otherspots could not be identified solely based on the dataobtained in the MS analysis.
Six of these non-identified protein spots (5–10; Fig. 2)resulted in a comparable peptide mass fingerprint and aretherefore likely to be derived from the same protein. Oneof the protein spots (5; Fig. 2) was subsequently sub-jected to N-terminal sequencing, resulting in a 46-amino-acid sequence that was found to harbour the previouslyidentified MS/MS tags. As the obtained sequence showedhomology to a structural Aspergillus nidulans phialideprotein, this protein was designated CfPhiA (Table 1). Thisis a protein that typically occurs on phialides, which aresporogenous cells that release conidia from their apex bybudding (Melin et al., 2003).
Three other protein spots (11–13; Fig. 2) also generateda comparable mass fingerprint, implicating that also thesespots may be derived from the same protein. N-terminal
Fig. 1. Disease progression of Cladosporium fulvum on tomato.A. Typical symptoms caused by C. fulvum on susceptible MM-Cf-0tomato plants at 3, 6, 9, 13 and 16 days post inoculation (dpi). Thefungus is not visible at early stages of infection (3 dpi) but developswhite patches of conidiophores (6 dpi) that expand and coveralmost the whole leaf (9 dpi). Subsequently, the conidiophores startto produce conidia (13 dpi) which give the leaf a green-brownishvelvet-like appearance (16 dpi).B. Quantitative real-time reverse transcription PCR to measureC. fulvum growth on resistant MM-Cf-4 tomato plants (white) andon susceptible MM-Cf-0 tomato plants (grey) at 3, 6, 9, 13 and 16dpi. The extent of colonization is determined by the relativequantification (RQ) of transcript levels of the constitutivelyexpressed C. fulvum actin gene (measure for fungal biomass) tothe constitutively expressed tomato glyceraldehyde-3-phosphatedehydrogenase (GAPDH) gene (measure for plant biomass) shownon a logarithmic scale. Bars represent mean values and standarderrors of three leaflets taken from two plants at each time pointanalysed. The experiment was repeated twice with similar results.
Ecp6 is a virulence factor 3
© 2008 The AuthorsJournal compilation © 2008 Blackwell Publishing Ltd, Molecular Microbiology
sequencing of spot 12 resulted in a 26-amino-acidsequence harbouring the identified MS/MS sequencetags and the corresponding protein was designated Ecp6(Table 1).
The remaining protein spots (14, 15; Fig. 2), of whichwe obtained peptide mass fingerprints as well as peptide
fragment spectra, were also subjected to N-terminalsequencing. For protein spot 14, the 25-amino-acidsequence that was obtained matched the correspondingMS/MS sequence tags and the protein was designatedEcp7 (Table 1). Although sequence information based onMS/MS was available for protein spot 15, this protein wasnot considered for further study because N-terminalsequence failed repeatedly.
Cloning of extracellular protein genes
Degenerate primers were designed based on theN-terminal protein sequences of CfPhiA and Ecp6 andwere used in combination with an oligo-(dT) primer toamplify the coding regions of the corresponding genesusing a cDNA library from C. fulvum-infected tomatoleaves as template. For Ecp7, a degenerate primer basedon an MS sequence tag was used because the N-terminalsequence was not yet available when the cloning wasinitiated. In all cases, a cDNA sequence was successfullyamplified which corresponded to MS/MS and N-terminalpeptide sequences. For CfPhiA, a 720 bp fragmentencoding the mature protein and part of the 3′UTR wascloned (Fig. S1). The predicted mature CfPhiA proteincontains 175 amino acids and has a predicted molecularmass of about 19 kDa and an isoelectric point (pI) of 5.0.BLASTP analysis (Altschul et al., 1997; Schäffer et al.,2001) of the amino acid sequence showed that thisprotein shares similarity to putative proteins of severalfungal species including A. nidulans, A. fumigatus andNeurospora crassa. Of these orthologues, the PhiAprotein from A. nidulans has been functionally character-ized (Melin et al., 2003), and was found to be essential forgrowth and sporulation of the fungus as phiA mutantswere found to be impaired in phialide development.Therefore, it is likely that the C. fulvum putative ortho-logue CfPhiA has a similar function.
A 742 bp fragment with the coding region for the matureEcp6 protein and the 3′UTR was cloned (Fig. S1). Ecp6encodes a mature protein of 199 amino acids, includingeight cysteines, and has a predicted molecular mass of21 kDa and a pI of 4.6. Furthermore, Ecp6 contains five
Fig. 2. The apoplast proteome of Cladosporium fulvum-infectedtomato analysed with two-dimensional polyacrylamide gelelectrophoresis (2D-PAGE). Coomassie brilliant blue-stained2D-PAGE gels obtained after electrophoresis of soluble proteinspresent in apoplastic fluid collected from a compatible (A; race 5C. fulvum strain inoculated onto MM-Cf-0 plants) and anincompatible (B; race 5 C. fulvum strain inoculated onto MM-Cf-4plants) interaction at 14 days post inoculation. The proteins werefocused over a non-linear gradient of pH 4–7. Molecular weightmarkers for the second dimension are indicated on the left. Thepart of the gel containing the C. fulvum-derived differentiallyaccumulated proteins is shown. Protein spots for whichidentification was pursued are numbered (see also Table 1).
Table 1. Apoplast proteins identified with mass spectrometry.
Protein Spot numbers Peptides confirmed with MS/MS N-terminal amino acids (aa) Reference
Ecp1 1, 2 5 – Joosten and de Wit (1988)Ecp2 3 7 – Wubben et al. (1994)Ecp5 4 4 – Laugé et al. (2000)CfPhiA 5–10 6 46 aa This studyEcp6 11–13 4 26 aa This studyEcp7 14 5 25 aa This studyEndochitinase 16, 17 PMFa – Joosten and de Wit (1989)
a. Determined with MALDI-TOF-generated peptide mass fingerprints (PMF).
4 M. D. Bolton et al. �
© 2008 The AuthorsJournal compilation © 2008 Blackwell Publishing Ltd, Molecular Microbiology
predicted N-glycosylation sites, explaining the location ofthe Ecp6 protein spots on the 2D-gel. Based on BLASTP
analysis, Ecp6 was found to share significant homology tothe glycoprotein CIH1 identified in the plant pathogenicfungus Colletotrichum lindemuthianum (Perfect et al.,1998). Although the contribution of CIH1 to pathogenicityis unknown, it has been shown to accumulate duringinfection on bean in the walls of intracellular hyphae andthe interfacial matrix which separates the hyphae from theinvaginated host plasma membrane (Perfect et al., 1998).
For Ecp7, a 464 bp cDNA fragment was cloned contain-ing the coding region for 84 amino acids of the matureEcp7 protein. N-terminal sequencing of Ecp7 revealedthat a stretch of 16 amino acids precedes the peptide thatwas identified as an MS tag, and based on which thedegenerate primer for cloning the cDNA was designed(Fig. S1). Therefore it should be concluded that Ecp7encodes a mature protein of 100 amino acids whichincludes six cysteines and has a predicted molecularmass of 11 kDa and a pI of 6.0. BLASTP analysis of theamino acid sequence revealed no significant homology ofEcp7 to other protein sequences deposited in publicdatabases.
CfPhiA, Ecp6 and Ecp7 are expressed during infection
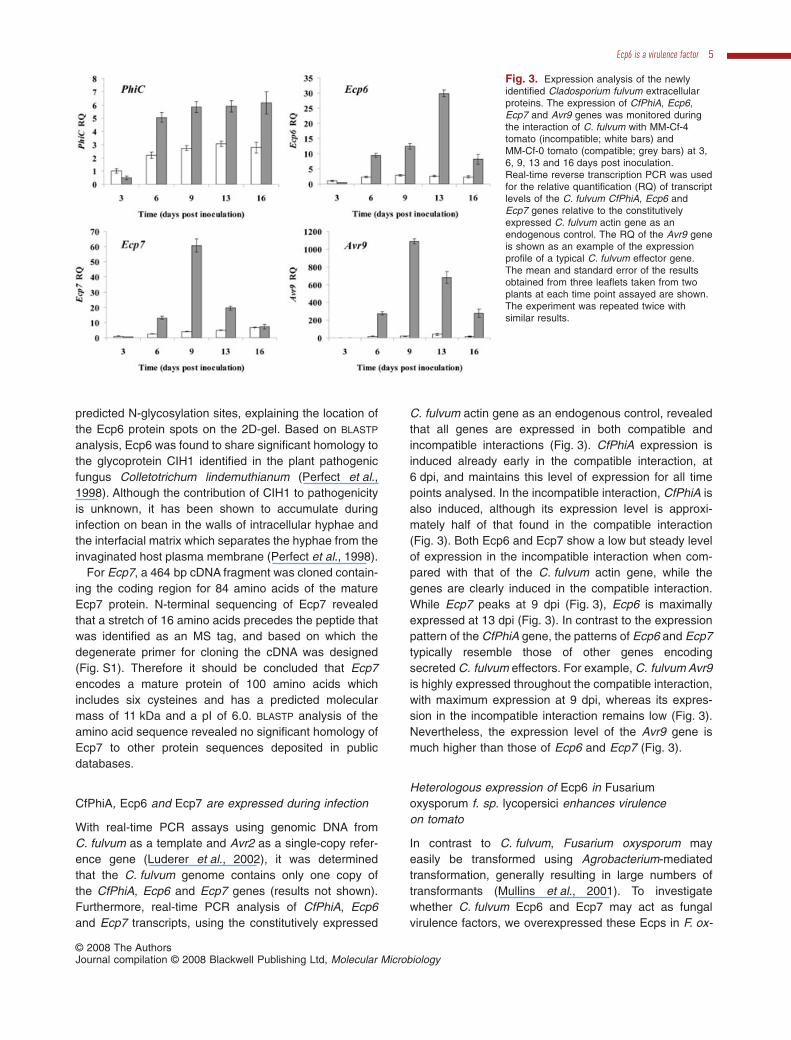
With real-time PCR assays using genomic DNA fromC. fulvum as a template and Avr2 as a single-copy refer-ence gene (Luderer et al., 2002), it was determinedthat the C. fulvum genome contains only one copy ofthe CfPhiA, Ecp6 and Ecp7 genes (results not shown).Furthermore, real-time PCR analysis of CfPhiA, Ecp6and Ecp7 transcripts, using the constitutively expressed
C. fulvum actin gene as an endogenous control, revealedthat all genes are expressed in both compatible andincompatible interactions (Fig. 3). CfPhiA expression isinduced already early in the compatible interaction, at6 dpi, and maintains this level of expression for all timepoints analysed. In the incompatible interaction, CfPhiA isalso induced, although its expression level is approxi-mately half of that found in the compatible interaction(Fig. 3). Both Ecp6 and Ecp7 show a low but steady levelof expression in the incompatible interaction when com-pared with that of the C. fulvum actin gene, while thegenes are clearly induced in the compatible interaction.While Ecp7 peaks at 9 dpi (Fig. 3), Ecp6 is maximallyexpressed at 13 dpi (Fig. 3). In contrast to the expressionpattern of the CfPhiA gene, the patterns of Ecp6 and Ecp7typically resemble those of other genes encodingsecreted C. fulvum effectors. For example, C. fulvum Avr9is highly expressed throughout the compatible interaction,with maximum expression at 9 dpi, whereas its expres-sion in the incompatible interaction remains low (Fig. 3).Nevertheless, the expression level of the Avr9 gene ismuch higher than those of Ecp6 and Ecp7 (Fig. 3).
Heterologous expression of Ecp6 in Fusariumoxysporum f. sp. lycopersici enhances virulenceon tomato
In contrast to C. fulvum, Fusarium oxysporum mayeasily be transformed using Agrobacterium-mediatedtransformation, generally resulting in large numbers oftransformants (Mullins et al., 2001). To investigatewhether C. fulvum Ecp6 and Ecp7 may act as fungalvirulence factors, we overexpressed these Ecps in F. ox-
Fig. 3. Expression analysis of the newlyidentified Cladosporium fulvum extracellularproteins. The expression of CfPhiA, Ecp6,Ecp7 and Avr9 genes was monitored duringthe interaction of C. fulvum with MM-Cf-4tomato (incompatible; white bars) andMM-Cf-0 tomato (compatible; grey bars) at 3,6, 9, 13 and 16 days post inoculation.Real-time reverse transcription PCR was usedfor the relative quantification (RQ) of transcriptlevels of the C. fulvum CfPhiA, Ecp6 andEcp7 genes relative to the constitutivelyexpressed C. fulvum actin gene as anendogenous control. The RQ of the Avr9 geneis shown as an example of the expressionprofile of a typical C. fulvum effector gene.The mean and standard error of the resultsobtained from three leaflets taken from twoplants at each time point assayed are shown.The experiment was repeated twice withsimilar results.
Ecp6 is a virulence factor 5
© 2008 The AuthorsJournal compilation © 2008 Blackwell Publishing Ltd, Molecular Microbiology

ysporum f. sp. lycopersici. To this end, the sequencesencoding the mature proteins were fused in frame withthe sequence encoding the C. fulvum Avr4 signalpeptide for extracellular targeting (Joosten et al., 1997)into a binary vector under control of the fungal constitu-tive ToxA promoter (Ciuffetti et al., 1997). UsingAgrobacterium-mediated transformation a large numberof transformants were obtained, and presence of thetransgene was confirmed by PCR (data not shown).Four transformants were randomly picked for each ofthe C. fulvum Ecps and tested in an inoculation assayon tomato. Upon inoculation of tomato plants with trans-formants that overexpress Ecp7, disease developmentwas indistinguishable from disease caused by the non-transformed progenitor strain (data not shown). In con-trast, on tomato plants that were inoculated with each ofthe four transformants that overexpress Ecp6, diseasesymptoms developed earlier and were more severecompared with the inoculation with the non-transformedprogenitor F. oxysporum f. sp. lycopersici strain (Fig. 4Aand B) or the transformants that overexpress Ecp7 (datanot shown). With reverse transcription PCR it was con-firmed that in each of the transformants, but not in theprogenitor F. oxysporum f. sp. lycopersici strain, Ecp6was expressed (Fig. 4C).
RNAi-mediated silencing of Ecp6 compromisesC. fulvum virulence on tomato.
RNAi has been successfully employed for gene func-tional analysis in filamentous fungi (Nakayashiki et al.,2005). This is particularly relevant for fungi like C. fulvum
for which homologous recombination is not straightfor-ward. Recent evidence has shown that PEG-mediatedtransformation may generate somaclonal variation thatmay be circumvented by Agrobacterium-mediated trans-formation which is, however, significantly less efficient(van Esse et al., 2007). Therefore, RNAi was recentlysuccessfully implemented to silence the expressionof C. fulvum effector genes (van Esse et al., 2007).
Based on the results obtained with heterologousexpression of C. fulvum Ecp6 in F. oxysporum f. sp.lycopersici, we applied RNAi-mediated silencing for func-tional analysis of the C. fulvum Ecp6 gene usingAgrobacterium-mediated transformation with constructsaimed at generating double-stranded RNA that targetsthese genes (RNAi). A pGREEN-based binary vector,carrying transfer DNA (T-DNA) that contains either anourseothricin resistance cassette or a hygromycin resis-tance cassette, and an inverted-repeat fragment of thetarget gene under control of the fungal constitutive ToxApromoter (Ciuffetti et al., 1997), was used to provokeRNAi-mediated gene silencing. To target the expressionof the Ecp6 gene, two RNAi constructs were generatedbased on different sections of the Ecp6 coding region.Agrobacterium-mediated transformation of the RNAi con-structs generated several antibiotic-resistant transfor-mants for each construct. Analysis of the transformantsindicated that their growth in vitro was indistinguishablefrom that of the progenitor race 5 isolate (data not shown).As C. fulvum effector genes show variable expressionwhen cultured in vitro (Thomma et al., 2006), 4-week-oldMM-Cf-0 tomato plants were inoculated with three trans-genic C. fulvum strains to determine whether the introduc-
Fig. 4. Symptoms caused by wild-type Fusarium oxysporum f. sp. lycopersici and heterologous Ecp6 overexpression transformants onsusceptible tomato.A and B. Side view (A) and top view (B) of the disease phenotype caused by F. oxysporum f. sp. lycopersici wild-type (WT) and fourindependent heterologous Ecp6 overexpression transformants (Ecp6-1 to Ecp6-4) on susceptible tomato MoneyMaker plants when comparedwith mock-inoculated tomato (mock) at 14 days post inoculation.C. Reverse transcription PCR to detect in planta transcription of heterologously expressed C. fulvum Ecp6 in F. oxysporum f. sp. lycopersiciwild-type and four independent heterologous Ecp6 overexpression transformants (Ecp6-1 to Ecp6-4) on susceptible tomato MoneyMakerplants when compared with mock-inoculated tomato (mock) at 14 days post inoculation.
6 M. D. Bolton et al. �
© 2008 The AuthorsJournal compilation © 2008 Blackwell Publishing Ltd, Molecular Microbiology
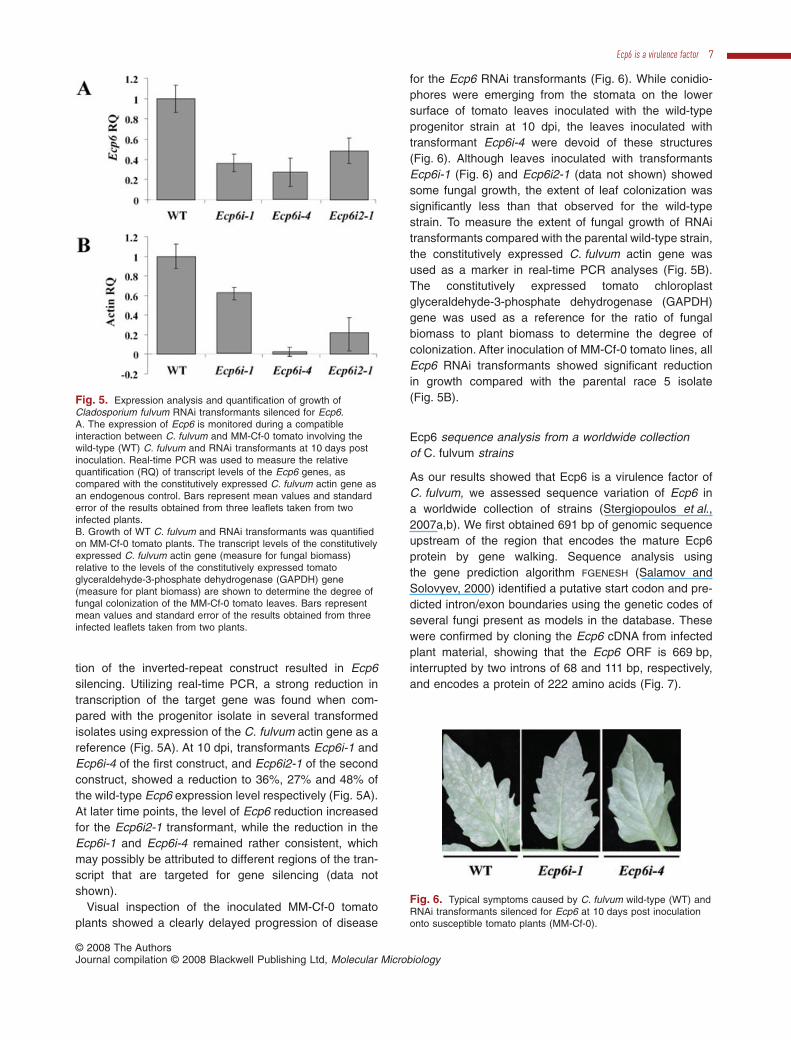
tion of the inverted-repeat construct resulted in Ecp6silencing. Utilizing real-time PCR, a strong reduction intranscription of the target gene was found when com-pared with the progenitor isolate in several transformedisolates using expression of the C. fulvum actin gene as areference (Fig. 5A). At 10 dpi, transformants Ecp6i-1 andEcp6i-4 of the first construct, and Ecp6i2-1 of the secondconstruct, showed a reduction to 36%, 27% and 48% ofthe wild-type Ecp6 expression level respectively (Fig. 5A).At later time points, the level of Ecp6 reduction increasedfor the Ecp6i2-1 transformant, while the reduction in theEcp6i-1 and Ecp6i-4 remained rather consistent, whichmay possibly be attributed to different regions of the tran-script that are targeted for gene silencing (data notshown).
Visual inspection of the inoculated MM-Cf-0 tomatoplants showed a clearly delayed progression of disease
for the Ecp6 RNAi transformants (Fig. 6). While conidio-phores were emerging from the stomata on the lowersurface of tomato leaves inoculated with the wild-typeprogenitor strain at 10 dpi, the leaves inoculated withtransformant Ecp6i-4 were devoid of these structures(Fig. 6). Although leaves inoculated with transformantsEcp6i-1 (Fig. 6) and Ecp6i2-1 (data not shown) showedsome fungal growth, the extent of leaf colonization wassignificantly less than that observed for the wild-typestrain. To measure the extent of fungal growth of RNAitransformants compared with the parental wild-type strain,the constitutively expressed C. fulvum actin gene wasused as a marker in real-time PCR analyses (Fig. 5B).The constitutively expressed tomato chloroplastglyceraldehyde-3-phosphate dehydrogenase (GAPDH)gene was used as a reference for the ratio of fungalbiomass to plant biomass to determine the degree ofcolonization. After inoculation of MM-Cf-0 tomato lines, allEcp6 RNAi transformants showed significant reductionin growth compared with the parental race 5 isolate(Fig. 5B).
Ecp6 sequence analysis from a worldwide collectionof C. fulvum strains
As our results showed that Ecp6 is a virulence factor ofC. fulvum, we assessed sequence variation of Ecp6 ina worldwide collection of strains (Stergiopoulos et al.,2007a,b). We first obtained 691 bp of genomic sequenceupstream of the region that encodes the mature Ecp6protein by gene walking. Sequence analysis usingthe gene prediction algorithm FGENESH (Salamov andSolovyev, 2000) identified a putative start codon and pre-dicted intron/exon boundaries using the genetic codes ofseveral fungi present as models in the database. Thesewere confirmed by cloning the Ecp6 cDNA from infectedplant material, showing that the Ecp6 ORF is 669 bp,interrupted by two introns of 68 and 111 bp, respectively,and encodes a protein of 222 amino acids (Fig. 7).
Fig. 5. Expression analysis and quantification of growth ofCladosporium fulvum RNAi transformants silenced for Ecp6.A. The expression of Ecp6 is monitored during a compatibleinteraction between C. fulvum and MM-Cf-0 tomato involving thewild-type (WT) C. fulvum and RNAi transformants at 10 days postinoculation. Real-time PCR was used to measure the relativequantification (RQ) of transcript levels of the Ecp6 genes, ascompared with the constitutively expressed C. fulvum actin gene asan endogenous control. Bars represent mean values and standarderror of the results obtained from three leaflets taken from twoinfected plants.B. Growth of WT C. fulvum and RNAi transformants was quantifiedon MM-Cf-0 tomato plants. The transcript levels of the constitutivelyexpressed C. fulvum actin gene (measure for fungal biomass)relative to the levels of the constitutively expressed tomatoglyceraldehyde-3-phosphate dehydrogenase (GAPDH) gene(measure for plant biomass) are shown to determine the degree offungal colonization of the MM-Cf-0 tomato leaves. Bars representmean values and standard error of the results obtained from threeinfected leaflets taken from two plants.
Fig. 6. Typical symptoms caused by C. fulvum wild-type (WT) andRNAi transformants silenced for Ecp6 at 10 days post inoculationonto susceptible tomato plants (MM-Cf-0).
Ecp6 is a virulence factor 7
© 2008 The AuthorsJournal compilation © 2008 Blackwell Publishing Ltd, Molecular Microbiology

The full-length sequence of Ecp6 was obtained from acollection of 50 C. fulvum strains (Table S1). Analysis ofthe sequence 62 bp upstream of the start codon to 91 bpdownstream of the stop codon revealed that variationwithin Ecp6 was very limited, resulting in a total of fivesingle-nucleotide polymorphisms (SNPs) within thesestrains (Fig. 7). One SNP (G > A at 494 bp downstream ofthe putative start codon) occurred inside the second intronof Ecp6, and was only detected in one Canadian strain(#34; Table S1). The other four SNPs all occurred inseven strains originating from North America (#31, #34,#40, #41; Table S1), and Japan (#67, #71, #74; Table S1).While one SNP (G > A at 128 bp) occurred in the firstintron, two other SNPs are silent mutations (C > T at335 bp and G > A at 662 bp). Only one SNP (C > A at142 bp) is predicted to result in an amino acid substitution(Thr25 > Asn; Fig. 7).
Orthologues of Ecp6 are found in several fungal species
Interrogation of the C. fulvum Ecp6 protein sequenceusing BLASTP (Altschul et al., 1997; Schäffer et al., 2001)and Pfam analysis (Finn et al., 2008) indicates that theEcp6 protein contains three lysin motif (LysM) domains.These domains are widespread protein modules ofapproximately 40 amino acids, originally identified in abacterial autolysin that degrades bacterial cell walls (Joriset al., 1992). LysM domains are also found in eukaryoticproteins, and presently LysM domains are implicated inbinding of diverse carbohydrates that occur in bacterialpeptidoglycan, fungal chitin and Nod factor signals thatare produced by Rhizobium bacteria during the initiationof root nodules on legumes (Butler et al., 1991; Amonet al., 1998; Ponting et al., 1999; Bateman and Bycroft,2000). We queried all available fungal genome sequencesand expressed sequence tag (EST) libraries (Table 2) forthe presence of Ecp6-like proteins using BLASTP orTBLASTN respectively. The retrieved sequences were sub-sequently analysed for predicted protein domains usingHMMER (http://hmmer.janelia.org/) loaded with thecurrent Pfam HMM library (http://pfam.sanger.ac.uk). Pre-diction of significant LysM domains (E-value cut-off 0.001)was used as a selection criterion for further analysis.Subsequently, all sequences containing predicted LysM
domains were aligned, permitting for the selection offungal proteins with high overall similarity to C. fulvumEcp6. In this way, a list of 16 putative C. fulvum Ecp6-likeproteins was generated, containing five Aspergillus nigerproteins, two Magnaporthe grisea proteins, and one fromeach Mycosphaerella fijiensis, M. graminicola, Botrytiscinerea, Sclerotinia sclerotiorum, A. nidulans, A. oryzae,A. flavus, C. lindemuthianum and Leptosphaeriamaculans. For these 17 proteins, using CLUSTALW
(Chenna et al., 2003) a multiple sequence alignmentanalysis was performed (Fig. S2). In addition to the LysMdomains, the positions of the cysteine residues that flankthe LysM domains, and the high abundance of proline,serine and threonine residues in the LysM linker regionsappear to be conserved (Fig. S2). Subsequently aneighbour-joining tree (Saitou and Nei, 1987) was con-structed to reveal evolutionary relationships (Fig. 8).Based on this tree, the 16 Ecp6-like proteins can bedivided into three groups. C. fulvum Ecp6 clusters withthree Ecp6-like proteins of M. graminicola, M. fijiensis andL. maculans that all contain three LysM domains (Group1, Fig. 8). The second group of Ecp6-like proteins encom-passes the two M. grisea Ecp6-like proteins and CIH1from C. lindemuthianum that are shorter than other Ecp6-like proteins and have only two LysM domains (Group 2,Fig. 8). The largest group of Ecp6-like proteins, encom-passing the five A. niger proteins in addition to those ofA. nidulans, A. oryzae, S. sclerotiorum and B. cinerea,contain two LysM domains and a weak, but not significant,signature of a third LysM domain (Group 3, Fig. 8).
Homology modelling of Ecp6 LysM domains
Although LysM domains have been identified in over 1500proteins, the three-dimensional (3D) structure of onlythree LysM domains has been reported. Two of these areof bacterial origin, the 3D structure of a LysM domain ofthe Escherichia coli membrane-bound lytic murein trans-glycosylase D (MltD; PDB code: 1EOG; Bateman andBycroft, 2000) and the LysM domain of the Bacillus sub-tilis spore protein ykuD of unknown function (PDB code:1Y7M; Bielnicki et al., 2006). Recently, the 3D structure ofthe LysM domain of the human hypothetical proteinSB145 was determined using nuclear magnetic reso-
Fig. 7. Allelic variation of the Cladosporiumfulvum Ecp6 gene. Open reading frames areshown as light grey boxes and introns asblack boxes. The predicted signal peptide isindicated as dark grey box. The white flagindicates a single-nucleotide polymorphism(SNP) that leads to an amino acid substitutionin the Ecp6 protein. Silent mutations areindicated by a T. The figure is drawn at scale.
8 M. D. Bolton et al. �
© 2008 The AuthorsJournal compilation © 2008 Blackwell Publishing Ltd, Molecular Microbiology

nance (NMR) imaging (PDB code: 2DJP). The structuralorganization of the three LysM domains from these differ-ent proteins is highly similar, and characterized by a baabfold, with the two helices stacking on one side of the plategenerated by a double-stranded antiparallel b-sheet.
Recently, the first characterization of an interaction of aLysM domain with its ligand was reported (Ohnuma et al.,2008). Binding of oligomers of N-acetylglucosamine[(GlcNAc)n], a monosaccharide derivative of glucose thatis a building block for bacterial peptidoglycan and fungalchitin, to the LysM domains of a chitinase from Pterisryukyuensis was monitored with NMR spectroscopy. Thestoichiometry of (GlcNAc)n/LysM binding was found tooccur in a 1:1 ratio. Furthermore, using (GlcNAc)5 it wasshown that binding of this oligomer to the LysM domainoccurs at a shallow groove formed by the N-terminal partof helix 1, the loop between strand 1 and helix 1, theC-terminal part of helix 2, and the loop between helix 2and strand 2.
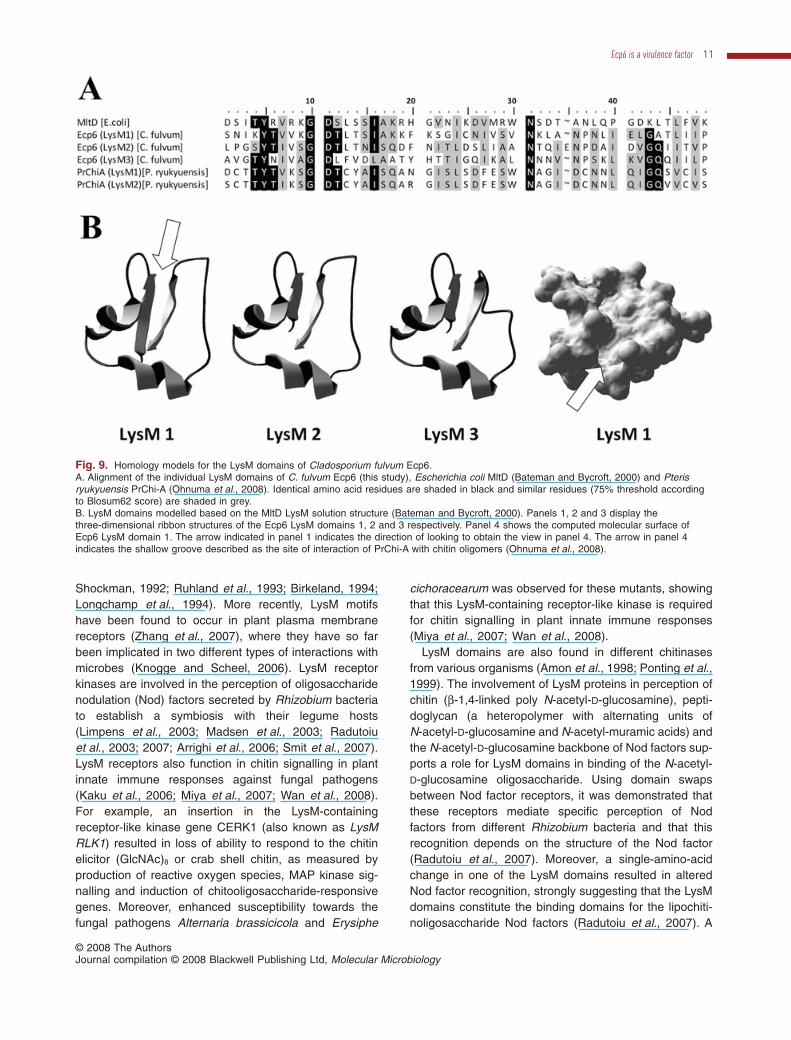
To predict the ligand binding site with correspondingbinding specificities of the C. fulvum Ecp6 LysM domains,homology-based modelling based on the 3D structure ofthe LysM domain of the MltD structure was performed.The MltD and Ecp6 LysM domains show moderate butsignificant overall sequence similarity (53%, 47% and33%, respectively, for LysM domains 1, 2 and 3; Fig. 9A).Moreover, by assessing local Kyte–Doolittle (KD) hydro-phobicity values (Kyte and Doolittle, 1982), the conservedsecondary structure could be predicted, which was sub-sequently used to predict the 3D structure. The predicted3D structure of the three individual Ecp6 LysM domains ishighly similar, with small changes in the position of thesecond loop of the third LysM domain (Fig. 9B). Moreover,due to sufficient similarity (52%) of LysM domain 1of C. fulvum Ecp6 to LysM domains 1 and 2 ofP. ryukyuensis PrChi-A (Fig. 9A), ligand binding can bemodelled according to the interaction between chitin oli-gomers and PrChi-A LysM domains. The molecularsurface of the first LysM domain of Ecp6 (Fig. 9B, panel 1)was computed and is shown in panel 4 of Fig. 9B. In thesurface of this LysM domain, a cavity is observed thatfulfils the requirements to act as binding site of chitinoligomers, based on the structural homology withPrChi-A.
Discussion
In this study, we employed a combined 2D-PAGE andproteomics approach to identify C. fulvum proteins pro-duced and accumulating in compatible as compared withincompatible interactions. It was anticipated that proteinsthat accumulated exclusively in the compatible interactionwere predominantly of pathogen origin. Indeed in this way,three novel extracellular C. fulvum proteins (CfPhiA, Ecp6and Ecp7) could be identified in addition to the previouslydescribed effector proteins Ecp1, Ecp2 and Ecp5(Table 1). CfPhiA was found to have homology to the PhiAprotein from A. nidulans which is important for phialideand conidium development (Melin et al., 2003). Several
Table 2. Fungal whole genome and EST sequence librariesscreened for Ecp6-like sequences.
Speciesa No. of hitsb LysMc
Cladosporium fulvum (Ecp6) 1 YesColletotrichum lindemuthianum (CIH1) 1 YesEST sequences
Alternaria brassicicola 0 N.a.Blumeria graminis 0 N.a.Colletotrichum gloeosporioides f. sp.
aeschynomene0 N.a.
Colletotrichum trifolii 0 N.a.Fusarium sporotrichioides 0 N.a.Leptosphaeria maculans 1 YesOphiostoma novo-ulmi 0 N.a.Phycomyces blakesleeanus 0 N.a.
Whole genome sequencesAspergillus flavus 2 Yes (1)Aspergillus nidulans 1 YesAspergillus niger 5 Yes (5)Aspergillus oryzae 1 YesBatrachochytrium dendrobatidis* 1 YesBotrytis cinerea 1 YesCandida sp. 0 N.a.Chaetomium globosum* 3 YesCryphonectria parasitica* 1 YesCryptococcus neoformans* 1 YesFusarium graminearum 0 N.a.Fusarium oxysporum 1 NoFusarium verticilliodes 0 N.a.Histoplasma capsulatum* 1 YesLaccaria bicolor 0 N.a.Lodderomyces elongisporus 0 N.a.Magnaporthe grisea 2 Yes (2)Mycosphaerella fijensis 1 YesMycosphaerella graminicola 1 YesNectria haematococca 0 N.a.Neurospora crassa 1 NoPhanerochaete chrysosporium 0 N.a.Pichia stipitis 0 N.a.Podospora anserina 1 NoPostia placenta 0 N.a.Sclerotinia sclerotiorum 1 YesStagonospora nodorum* 1 YesSporobolomyces roseus 0 N.a.Ustilago maydis 1 No
a. LysM-containing Ecp6-like proteins of the species indicated in boldare included in the alignment shown in Fig. 8. The asterisks indicatespecies for which a LysM-containing Ecp6-like protein is identified,but as the overall homology of these proteins to C. fulvum Ecp6 andC. lindemuthianum CIH1 is low they are not included in the alignmentshown in Fig. 8.b. BLASTP and TBLASTN searches (E-value < 0.001) were performedusing Cladosporium fulvum Ecp6 and Colletotrichum lindemuthianumCIH1 as queries.c. Sequences were analysed using HMMER (http://hmmer.janelia.org) loaded with the current Pfam HMM library (http://pfam.sanger.ac.uk) for the presence of LysM domains (E-value < 0.001). Numbersbetween brackets indicate how many of the hits contain a predictedLysM domain. N.a. is not applicable.
Ecp6 is a virulence factor 9
© 2008 The AuthorsJournal compilation © 2008 Blackwell Publishing Ltd, Molecular Microbiology

attempts to generate RNAi transformants for CfPhiA havefailed (data not shown), which suggests that silencing ofCfPhiA might be detrimental or even lethal. Furthermore,the expression pattern of CfPhiA compared with that ofAvr9 suggests that CfPhiA is likely not a genuine effectorof the fungus.
Of the newly identified fungal extracellular proteins,Ecp7 especially resembles the previously identified Avrsand Ecps. It is relatively small (the mature proteincontains 100 amino acid residues of which six are cys-teines) with a calculated molecular mass of approxi-mately 11 kDa. The even number of cysteine residuessuggests their involvement in disulphide bridges that aidin their stability and activity in the harsh protease-richapoplast (Joosten et al., 1997; Kooman-Gersmann et al.,1997; Thomma et al., 2005). In addition, the Ecp7expression profile during infection of tomato resemblesthat of other effector genes (Fig. 5). However, like formost of the previously identified Avrs and Ecps, theEcp7 amino acid sequence did not show significanthomology to sequences present in public databases.Despite the use of multiple transformants generated withtwo different RNAi constructs to target Ecp7 expression,we have not been able to obtain unambiguous evidenceshowing that Ecp7 is a virulence factor of C. fulvum(data not shown). This is in contrast to the findings forC. fulvum Ecp6.
The mature Ecp6 protein contains 199 amino acidsand has an estimated molecular mass of 21 kDa, makingit the largest of the abundantly secreted effector proteins
of C. fulvum identified so far. Previous studies onthe genes encoding secreted C. fulvum effectors haveshown that Avr genes accumulated considerably morepolymorphisms than Ecp genes (Stergiopoulos et al.,2007a). This was suggested to be due to the lack ofselection pressure imposed on the pathogen to over-come resistance mediated by R genes that recognizeEcps, as these have not been deployed yet in commer-cial tomato lines (Stergiopoulos et al., 2007a). In line withthese findings, polymorphisms in Ecp6 were only rarelyobserved. Of the 50 C. fulvum strains, only seven strainscontained allelic variants of Ecp6. All seven of thesestrains, which have previously been shown to be related(Stergiopoulos et al., 2007b), contained the same fourSNPs, while one strain contained an additional fifth SNP.Of these five SNPs, only one resulted in an amino acidchange, while the four others concerned silent or intronmutations. The occurrence of mostly synonymous modi-fications in Ecp genes was hypothesized to imply selec-tive constraints for maintaining Ecp protein sequencesor, alternatively, a recent common ancestor gene (Ster-giopoulos et al., 2007a). However, our finding that Ecp6markedly contributes to C. fulvum virulence, and thatEcp6 has orthologues in other fungal species, favoursthe second hypothesis.
The Ecp6 protein contains three lysin motifs (LysMdomains) that were originally found in a variety ofenzymes that bind to and hydrolyse peptidoglycanspresent in bacterial cell walls, of which lysozyme is thebest-known example (Joris et al., 1992; Kariyama and
Fig. 8. Homologues of Ecp6 in other fungalspecies. Neighbour-joining tree of 17Ecp6-like sequences from different fungalspecies. The evolutionary history of Ecp6-likeprotein sequences was inferred byneighbour-joining analysis (Saitou and Nei,1987) and bootstrap values (%) are indicatedat the nodes. The tree is drawn to scale, withbranch lengths representing evolutionarydistances. The positions containing alignmentgaps were eliminated in pair-wise sequencecomparisons. A total of 220 positions werecalculated in the final data set.
10 M. D. Bolton et al. �
© 2008 The AuthorsJournal compilation © 2008 Blackwell Publishing Ltd, Molecular Microbiology
Shockman, 1992; Ruhland et al., 1993; Birkeland, 1994;Longchamp et al., 1994). More recently, LysM motifshave been found to occur in plant plasma membranereceptors (Zhang et al., 2007), where they have so farbeen implicated in two different types of interactions withmicrobes (Knogge and Scheel, 2006). LysM receptorkinases are involved in the perception of oligosaccharidenodulation (Nod) factors secreted by Rhizobium bacteriato establish a symbiosis with their legume hosts(Limpens et al., 2003; Madsen et al., 2003; Radutoiuet al., 2003; 2007; Arrighi et al., 2006; Smit et al., 2007).LysM receptors also function in chitin signalling in plantinnate immune responses against fungal pathogens(Kaku et al., 2006; Miya et al., 2007; Wan et al., 2008).For example, an insertion in the LysM-containingreceptor-like kinase gene CERK1 (also known as LysMRLK1) resulted in loss of ability to respond to the chitinelicitor (GlcNAc)8 or crab shell chitin, as measured byproduction of reactive oxygen species, MAP kinase sig-nalling and induction of chitooligosaccharide-responsivegenes. Moreover, enhanced susceptibility towards thefungal pathogens Alternaria brassicicola and Erysiphe
cichoracearum was observed for these mutants, showingthat this LysM-containing receptor-like kinase is requiredfor chitin signalling in plant innate immune responses(Miya et al., 2007; Wan et al., 2008).
LysM domains are also found in different chitinasesfrom various organisms (Amon et al., 1998; Ponting et al.,1999). The involvement of LysM proteins in perception ofchitin (b-1,4-linked poly N-acetyl-D-glucosamine), pepti-doglycan (a heteropolymer with alternating units ofN-acetyl-D-glucosamine and N-acetyl-muramic acids) andthe N-acetyl-D-glucosamine backbone of Nod factors sup-ports a role for LysM domains in binding of the N-acetyl-D-glucosamine oligosaccharide. Using domain swapsbetween Nod factor receptors, it was demonstrated thatthese receptors mediate specific perception of Nodfactors from different Rhizobium bacteria and that thisrecognition depends on the structure of the Nod factor(Radutoiu et al., 2007). Moreover, a single-amino-acidchange in one of the LysM domains resulted in alteredNod factor recognition, strongly suggesting that the LysMdomains constitute the binding domains for the lipochiti-noligosaccharide Nod factors (Radutoiu et al., 2007). A
Fig. 9. Homology models for the LysM domains of Cladosporium fulvum Ecp6.A. Alignment of the individual LysM domains of C. fulvum Ecp6 (this study), Escherichia coli MltD (Bateman and Bycroft, 2000) and Pterisryukyuensis PrChi-A (Ohnuma et al., 2008). Identical amino acid residues are shaded in black and similar residues (75% threshold accordingto Blosum62 score) are shaded in grey.B. LysM domains modelled based on the MltD LysM solution structure (Bateman and Bycroft, 2000). Panels 1, 2 and 3 display thethree-dimensional ribbon structures of the Ecp6 LysM domains 1, 2 and 3 respectively. Panel 4 shows the computed molecular surface ofEcp6 LysM domain 1. The arrow indicated in panel 1 indicates the direction of looking to obtain the view in panel 4. The arrow in panel 4indicates the shallow groove described as the site of interaction of PrChi-A with chitin oligomers (Ohnuma et al., 2008).
Ecp6 is a virulence factor 11
© 2008 The AuthorsJournal compilation © 2008 Blackwell Publishing Ltd, Molecular Microbiology

high-affinity chitin-binding protein was isolated from theplasma membrane of suspension-cultured rice cells. Thisextracellular membrane-anchored protein, CEBiP, con-tains two LysM domains. Knockdown of CEBiP expres-sion diminished the elicitor-induced oxidative burst as wellas expression of chitin-induced genes. Moreover, bindingassays as well as affinity labelling showed that the plasmamembrane of knockdown lines for CEBiP carried lesselicitor binding sites (Kaku et al., 2006).
Chitin binding has also been demonstrated for theC. fulvum effector protein Avr4 which contains an inverte-brate chitin-binding domain (van den Burg et al., 2003).Through this chitin-binding activity, Avr4 was found toprotect C. fulvum hyphae from hydrolysis by plant chiti-nases (van den Burg et al., 2006; van Esse et al., 2007).It is tempting to speculate that C. fulvum Ecp6 is a chitin-binding protein too. To that end it is interesting to note thatthe Ecp6 homologue CIH1 of the plant pathogenic fungusC. lindemuthianum is found to be present at the surfaceof intracellularly growing fungal structures present ininfected plant tissue (Perfect et al., 1998). Ecp6 maypotentially act as a functional homologue of Avr4 throughthe ability to bind to chitin. Such functional redundancymight explain why the C. fulvum strain Can38, which har-bours a frameshift mutation in the Avr4 gene and as aconsequence does not produce Avr4, is still able to infecttomato (Joosten et al., 1997). Alternatively, Ecp6 may actas a ‘stealth factor’ by shielding fungal hyphae in a similarfashion as has been suggested for hydrophobins (White-ford and Spanu, 2002). Furthermore, the fungus mayavoid recognition by the plant by sequestering chitinmono- or oligomers that act as elicitors of defenceresponses once they are released by the activity of plantchitinases.
Using homology modelling, docking sites for the inter-action between LysM domains and their ligands havebeen predicted (Mulder et al., 2006; Radutoiu et al.,2007). We have used homology modelling to show thatthe Ecp6 LysM domains are likely to structurally resemblepreviously characterized LysM domains, and that basedon structural calculations GlcNAc oligomers may indeeddock to the LysM domains of Ecp6 in a similar fashion asto the LysM domains of PrChi-A (Ohnuma et al., 2008).Future experiments will reveal whether the LysM domainsof Ecp6 are able to bind chitin and, moreover, how Ecp6contributes to fungal virulence.
Experimental procedures
Fungal and plant materials, and infection assays
The wild-type race 5 strain of C. fulvum was stored in 50%glycerol at -80°C until revitalized on potato dextrose agar(PDA; Oxoid, Hampshire, England) and was grown at roomtemperature in the dark. Two-week-old C. fulvum PDA plate
cultures were used to harvest conidia by adding sterile waterto the plates and rubbing the surface with a sterile glass rodto release the conidia. Conidial suspensions were filteredthrough Miracloth (Calbiochem-Behring, La Jolla, CA), cen-trifuged at 4000 r.p.m. and washed twice with sterile waterafter which the conidial concentration was determined. Sub-sequently, the conidia were used for plant inoculations orAgrobacterium tumefaciens-mediated transformation.
All tomato plants were grown under standard greenhouseconditions: 21°C during the 16 h day period, 19°C at night,70% relative humidity (RH) and 100 Watt m-2 supplementallight when the sunlight influx intensity was below150 Watt m-2. The tomato (S. esculentum) cultivar Money-Maker, containing no resistance genes against C. fulvum(MM-Cf-0), and a MoneyMaker near isogenic line containingthe Cf-4 locus (MM-Cf-4) were used for all inoculations.C. fulvum was inoculated as described previously (de Wit,1977). Per 5-week-old tomato plant, 5 ml of conidial suspen-sion (1 ¥ 106 conidia per ml) was used for spray inoculationon the lower surface of the leaves until drop-off. Plants werekept at 100% RH under a plastic cover for 48 h afterinoculation. All experiments, starting from plant inoculations,were repeated at least twice.
Preparation of protein samples and 2D-PAGE
Leaves were harvested from C. fulvum-infected MM-Cf-0 andMM-Cf-4 lines at 14 dpi and apoplastic fluid (AF) was isolatedby vacuum infiltration (van Esse et al., 2006) usingde-mineralized water followed by centrifugation for 5 min andstored at -20°C until further analysis. AF from both interac-tions was freeze-dried and the residue was re-suspended in3.5 ml of water. After centrifugation (10 min at 4000 g)samples were desalted using a PD-10 desalting column (GEHealthcare, UK), freeze-dried again and stored at -20°C.Freeze-dried protein samples were dissolved in 340 ml ofRehydration Buffer [7 M urea, 2 M thiourea, 4% CHAPS,60 mM DTT, 0.002% (w/v) bromophenol blue] along with3.4 ml of IPG buffer pH 4–7 (GE Healthcare). The sampleswere vortexed briefly and centrifuged (10 min at 4000 g). Theprotein samples were applied to Immobiline DryStrips of18 cm with a non-linear pH 4–7 gradient (GE Healthcare),covered with paraffin oil and allowed to re-hydrate overnightat room temperature. Isoelectric focusing was performedusing the Ettan IPGphor electrophoresis apparatus (GEHealthcare) at 20°C maintaining 50 mA per strip. A total focus-ing of 70 k Vh was achieved by following a running protocolusing a step-n-hold gradient (1.5 h 0–3500 V, 6 h 3500 V).After first dimensional isoelectric focusing, the strips werestored at -20°C.
Subsequently, strips were placed in equilibration buffer[EB; 50 mM Tris, pH 8.8, 6 M urea, 30% (v/v) glycerol and 2%(w/v) SDS] supplemented with 65 mM DTT. After 15 min, thebuffer was replaced by EB supplemented with 135 mMiodoacetamide, and the strips were incubated for another15 min. The proteins were subsequently separated on 12.5%polyacrylamide gels; the gels were run at 70 V for the first30 min and subsequently at 200 V until the bromophenol bluereached the bottom of the gels. Gels were stained with Coo-massie brilliant blue overnight and de-stained with 10%ethanol and 7.5% HAc in water.
12 M. D. Bolton et al. �
© 2008 The AuthorsJournal compilation © 2008 Blackwell Publishing Ltd, Molecular Microbiology

Mass spectrometry
Protein spots were excised from the gel and digested withtrypsin with an in-gel method (Shevchenko et al., 1996). Thecollected extracts of the resulting tryptic peptides werefreeze-dried and stored at -20°C. The peptides werere-dissolved in 8 ml of 50% acetonitrile, 5% formic acid. MSand MS/MS information was acquired with a Q-Tof1 (Waters,Manchester, UK) coupled with a nano-LC Ultimate system(LC Packings Dionex, Sunnyvale, CA). After the dilutionof 1–2 ml of sample 12 times with water, peptides wereseparated on a nano-analytical column (75 mm insidediameter ¥ 15 cm C18 PepMap, LC Packings, Dionex) usinga gradient of 2–50% acetonitrile, 0.1% formic acid in 20 min.The flow of 300 nl min-1 was directly infused into the Q-Tof1,operating in data-dependent MS and MS/MS modes. Theresulting MS/MS spectra were processed with Masslynxsoftware (Waters, Manchester, UK) and used to search inMASCOT using the MSDB database. As sequence data ofboth C. fulvum and tomato are far from complete, MS/MSdata from un-assigned spectra were analysed by usingthe Masslynx Pepseq software for de novo sequenceinformation. Both BLAST (http://www.expasy.org/tools/blast)and MSBLAST were used to search for possible homologousproteins with the generated sequence information. ForMALDI-TOF analysis, a 1 ml volume was spotted on a targetplate after mixing the samples 1:1 (v/v) with a solution of10 mg ml-1 a-Cyano-4-hydroxycinnamic acid in 50% ethanol/50% acetonitrile/0.1% TFA. Reflectron MALDI-TOF spectrawere acquired on a TofSpec 2E (Waters, Manchester, UK).For peptide mass fingerprinting the resulting peptide masslists were used to search in MASCOT using the same MSDBdatabase.
Cloning of CfPhiA, Ecp6 and Ecp7
Based on the N-terminal CfPhiA sequence MDPIDVVWK, theforward degenerate primer Deg-PhiA along with an oligo-(dT)primer (Table S2) was used to isolate the CfPhiA codingsequence. Likewise, degenerate forward primers (Table S2)were designed matching the ETKATDCG and QITTQDFGsequences from the N-terminal sequences of Ecp6 and Ecp7respectively. Using the degenerate primers and a poly Tprimer PCR products were amplified from a cDNA libraryderived from a compatible interaction between C. fulvum andtomato using the high fidelity polymerase ExTaq (Takara,Shiga, Japan). Products were cloned into the pGEM-T Easyvector (Promega, Madison, WI) and sequenced.
Construction of plasmids for RNAi in C. fulvum
Two constructs for overexpression of inverted-repeat con-structs for RNAi based on two different parts of the Ecp6coding sequence were generated. For the first RNAi con-struct targeting the 3′ end of Ecp6, 218 bp of Ecp6 wasPCR-amplified from cDNA using a forward primer that addedan NcoI restriction site to the 5′ end (Ecp6i-F) and a reverseprimer that added EcoRI and NotI restriction sites to the 3′end (Ecp6i-R; Table S2). PCR reactions were carried outunder the following conditions: an initial denaturation step for
2 min followed by denaturation for 15 s at 94°C, annealing for30 s at 55°C and extension for 1 min at 72°C for 30 cycles,followed by a final elongation step at 72°C for 5 min. PCRproducts were separated on 1% agarose gels and were puri-fied using the DNeasy kit (Qiagen, Valencia, CA). Subse-quently, PCR products were cloned into the pGemT-Easyvector. Vectors were digested with NcoI and NotI or with NcoIand EcoRI. Both digested inserts were cleaned from gelusing the QIAquick gel extraction kit (Qiagen) and subse-quently ligated with a NotI- and EcoRI-digested 129 bpspacer segment from the Pichia pastoris Aox-1 gene into theNcoI-digested plasmid pFBB302 (Dr Brandwagt, Wagenin-gen University). The plasmid pFBB302 is constructed in thebackbone of the pGreen II binary vector (Hellens et al., 2000)and contains a nourseothricin resistance cassette (Maloneket al., 2004) to select for fungal transformants, and the UidAreporter gene flanked by the constitutive ToxA fungal pro-moter (Ciuffetti et al., 1997) and trpC terminator (Punt et al.,1987). Digestion with NcoI releases the UidA codingsequence and allows ligation of the inverted-repeat RNAisequence.
For the second RNAi construct targeting the 5′ end ofEcp6, two Ecp6 PCR products were generated of 250 and318 bp with the same forward primer that added an EcoRIrestriction site to the 5′ end (Ecp6i2-F) and two differentreverse primers that added a NotI restriction sites to the 3′end (Ecp6i2 k-R and Ecp6i2 l-R respectively; Table S2). PCRreactions and gel cleaning were performed similar as for thefirst RNAi construct. Subsequently, PCR products werecloned into the pGemT-Easy vector, digested with NotI andEcoRI, cleaned from gel and ligated into the EcoRI-digestedplasmid pFBT004. The plasmid pFBT004 is a modifiedversion of pFBB302, in which the nourseothricin resistancecassette is replaced by a hygromycin resistance cassette(Punt et al., 1987).
A. tumefaciens-mediated transformation of C. fulvum
RNAi plasmids were transformed into A. tumefaciens strainLBA1100 [containing the binary vector pSoup (Hellens et al.,2000)] by electroporation. A 3 ml culture of A. tumefacienswas grown overnight in 1¥ YT (Sambrook and Russell, 2001)supplemented with kanamycin (25 mg ml-1). The followingday, the culture was centrifuged and re-suspended in 50 ml offresh minimal medium (MM) (Hooykaas et al., 1979) supple-mented with kanamycin (25 mg ml-1) and grown overnight.The following day, the culture was centrifuged andre-suspended in 10 ml of fresh MM. One millilitre ofre-suspended bacteria was used to inoculate 50 ml ofinduction medium [IM; MM salts plus 40 mM 2-(N-morpholino)ethanesulphonic acid (MES), pH 5.3, 10 mMglucose and 0.5% (w/v) glycerol] supplemented with 200 mMacetosyringone (AS) and was grown for an additional 4–5 huntil the culture reached an optical density (OD600) of 0.25. Atthat point, the A. tumefaciens culture was centrifuged andre-suspended in 10 ml of sterile water. In addition, whileA. tumefaciens cultures were growing in IM+AS medium,C. fulvum conidia were harvested and subsequently sus-pended in 50 ml of B5 medium (Duchefa Biochemie BV,Haarlem, the Netherlands) at a concentration of approxi-mately 1 ¥ 106 conidia ml-1 and placed in a rotary shaker
Ecp6 is a virulence factor 13
© 2008 The AuthorsJournal compilation © 2008 Blackwell Publishing Ltd, Molecular Microbiology

(125 r.p.m.) at room temperature to induce germination ofconidia. After 4–5 h, germinated conidia were centrifugedtwice at 4000 r.p.m. and re-suspended in sterile water to afinal volume of 1 ¥ 107 conidia ml-1.
Five hundred microlitres from the induced A. tumefacienscell suspension was mixed with 10 ml of germinated conidiaand plated (200 ml per plate) on a 0.45-mm-pore, 45-mm-diameter nitrocellulose filter (Whatman, Hillsboro, OR) andplaced on co-cultivation medium (IM + 200 mM AS and 5 mMglucose and 1.5% technical agar). The co-cultivation mixturewas incubated at 22°C for 2 days. Following incubation, thefilter was transferred to PDA supplemented with 50 mg ml-1
nourseothricin (Werner BioAgents, Jena, Germany) or with100 mg ml-1 hygromycin B (Duchefa Biochemie BV, Haarlem,the Netherlands) as a selection agent for transformants and200 mg ml-1 cefotaxime (Duchefa Biochemie BV, Haarlem,the Netherlands) to kill A. tumefaciens cells. Individual trans-formants were transferred to new selection plates and incu-bated until conidiogenesis under normal growth conditions.Conidia from these plates were stored in 50% glycerol at-80°C until further analysis.
Real-time PCR analyses
Three leaflets were harvested from inoculated MM-Cf-0 andMM-Cf-4 plants at 3, 6, 9, 13 and 16 dpi. Leaf samples werecomposed of three leaflets from the second, third and fourthtomato leaves of two tomato plants taken at each time point,immediately frozen in liquid nitrogen and stored at -80°Cuntil used for RNA analysis. A similar procedure was usedfor RNAi transformant analysis. Ecp6 RNAi transformantsEcp6i-1 and Ecp6i-4 along with Ecp7 RNAi transformantsEcp7i-1, Ecp7i-3 and Ecp7i-7 were randomly chosen forinoculation and analysis with the progenitor race 5 wild-typestrain inoculated on MM-Cf-0 plants. Leaf samples weretaken at 10 dpi, immediately frozen in liquid nitrogen andstored at -80°C until used for RNA analysis.
Total RNA was isolated from infected leaf material usingthe RNeasy kit (Qiagen, Valencia, CA), including anin-column DNase treatment (Qiagen) according to manufac-turer’s instruction. Total RNA was used for cDNA synthesisusing an oligo-(dT) primer and the SuperScript II reversetranscriptase kit (Invitrogen, Carlsbad, CA) according tothe manufacturer’s instructions. Quantitative real-time PCRwas conducted using an ABI7300 PCR machine (AppliedBiosystems, Foster City, CA) with the GoldStar SYBRgreen PCR kit (Eurogentec, Seraing, Belgium). All primersequences are shown in Table S2. Expression primers weredesigned so that the reverse primer was not included in theRNAi construct to prevent detection of the constitutivelyexpressed RNAi construct. For the first RNAi construct,primer pair Ecp6-RNAi-RQ-F and Ecp6-RNAi-RQ-R wasused, and for the second RNAi construct primer pair Ecp6-RNAi2-RQ-F and Ecp6-RNAi2-RQ-R. Real-time PCR condi-tions were as follows: an initial 95°C denaturation step for10 min followed by denaturation for 15 s at 95°C, annealingfor 30 s at 60°C and extension for 30 s at 72°C for 40cycles, and analysed on the 7300 System SDS software(Applied Biosystems, Foster City, CA). To ensure nogenomic DNA contaminated RNA samples, real-time PCRwas also carried out on RNA without the addition of reverse
transcriptase. All experiments, including leaf inoculations,were repeated twice.
Heterologous expression of C. fulvum Ecp6 inF. oxysporum f. sp. lycopersici
For C. fulvum Ecp6, the cDNA corresponding to the matureprotein was amplified using primer Ecp6OE-F that also con-tained the sequence encoding the C. fulvum Avr4 signalpeptide for extracellular targeting (Table S2). For C. fulvumEcp7, the cDNA corresponding to the mature protein wasamplified in two steps. As the 5′ coding sequence was lackingfrom our cDNA clone, a primer was designed to add a 5′codon-optimized sequence stretch based on the N-terminalprotein sequence (Ecp7NtermF) and used in combinationwith the reverse primer Ecp7OE-R (Table S2). The resultingPCR product was used as template for a second PCR withprimer Ecp7OE-F that also contained the sequence encodingthe C. fulvum Avr4 signal peptide for extracellular targetingand a HindIII restriction site in combination with the reverseprimer Ecp7OE-R that contained a XmaI restriction site(Table S2). All PCR reactions were carried out under thefollowing conditions: an initial denaturation step for 2 minfollowed by denaturation for 15 s at 94°C, annealing for 30 sat 56°C and extension for 1 min at 72°C for 30 cycles, fol-lowed by a final elongation step at 72°C for 5 min. PCRproducts were separated on 1% agarose gels and purifiedusing the DNeasy kit (Qiagen, Valencia, CA). Subsequently,PCR products were cloned into the pGemT-Easy vector andsequenced. A correct clone was digested with EcoRI (forEcp6) or HindIII and XmaI (for Ecp7), cleaned from gel, andligated into the EcoRI- (for Ecp6) or HindIII- and XmaI- (forEcp7) digested plasmid pFBT004. The constructs were trans-formed into A. tumefaciens strain LBA1100 [containing thebinary vector pSoup (Hellens et al., 2000)] by electroporationessentially as described by Mersereau et al. (1990).Agrobacterium-mediated transformation of F. oxysporum f.sp. lycopersici was performed as described (Mullins et al.,2001).
Ecp6 gene walking
Three primers designed on the region encoding the matureEcp6 protein (TSP1, TSP2 and TSP3; Table S2) were used toamplify the genomic DNA sequence upstream of the regionthat encodes the mature Ecp6 protein using the DNA WalkingSpeedUpTM Premix Kit (Seegene, Rockville, MD) according tothe manufacturer’s instructions. Amplified products werecloned in the pGEM-T Easy vector (Promega, Madison,WI) and sequenced. Putative open reading frames (ORFs)were predicted using the FGENESH program (Salamov andSolovyev, 2000) of the MOLQUEST software package (avail-able at http://softberry.com/berry.phtml; Softberry, NY, USA)using the genetic codes of several fungi present in the data-base as models. ORFs were verified by cloning Ecp6 cDNA.For this purpose, total RNA was isolated from leaves ofMM-Cf-0 plants inoculated with a race 5 strain of C. fulvum at11 dpi and used for cDNA synthesis using an oligo-(dT)primer (Table S2) and the SuperScript II reverse tran-scriptase kit (Invitrogen, Carsbad, CA) as described previ-
14 M. D. Bolton et al. �
© 2008 The AuthorsJournal compilation © 2008 Blackwell Publishing Ltd, Molecular Microbiology

ously (van Esse et al., 2007). The generated cDNA was usedas template for the primers Ecp6_ChrWal_F1 and Ecp6_R(Table S2) to amplify the predicted Ecp6 ORF. The primersEcp6_F3, Ecp6_F2, Ecp6_R3, Ecp6_R2 (Table S2) thathybridized outside the predicted Ecp6 ORF were used asnegative controls. The 50 ml PCR reaction mixes contained5.0 ml of 10¥ SuperTaq PCR reaction buffer, 10 mM of eachdNTP (Promega Benelux bv, Leiden, the Netherlands), 20 mMof each primer, 1 unit of SuperTaq DNA polymerase (HTBiotechnology, Cambridge, UK) and approximately 100 ng ofcDNA as template. The PCR programme consisted of aninitial 5 min denaturation step at 94°C, followed by 35 cyclesof denaturation at 94°C (30 s), annealing at 55°C (30 s) andextension at 72°C (60 s). A final extension step at 72°C(7 min) concluded the reaction. Amplified products werecloned in the pGEM-T Easy vector (Promega, Madison, WI)and sequenced.
Ecp6 allelic variation
Allelic variation in Ecp6 was determined for 50 C. fulvumstrains (Table S1) that are part of a previously describedcollection (Stergiopoulos et al., 2007a,b). Strains were cul-tured on half-strength PDA (Oxoid, Hampshire, England) at22°C. Conidia were harvested from 15-day-old cultures andfreeze-dried prior to DNA extraction. Genomic DNA isolationswere performed using the DNeasy Plant Mini Kit (Qiagen,Valencia, CA) according to the manufacturer’s instructions.The forward primer Ecp6_F3, located 424 bp upstream of theEcp6 translation start codon, and the reverse primerEcp6_R3, located 99 bp downstream of the Ecp6 stop codon,were used to amplify Ecp6 (Table S2). The 50 ml PCR reac-tion mixes contained 5.0 ml of 10¥ SuperTaq PCR reactionbuffer, 10 mM of each dNTP (Promega Benelux bv, Leiden,the Netherlands), 20 mM of each primer, 1 unit of SuperTaqDNA polymerase (HT Biotechnology, Cambridge, UK) andapproximately 100 ng of genomic DNA as template. The PCRprogramme consisted of an initial 5 min denaturation step at94°C, followed by 35 cycles of denaturation at 94°C (30 s),annealing at 55°C (30 s) and extension at 72°C (60 s). A finalextension step at 72°C (7 min) concluded the reaction. Ampli-fied PCR products were excised from 0.8% agarose gels,purified using the GFXTM PCR DNA and Gel Band PurificationKit (Amersham Biosciences UK limited, Buckinghamshire,England), and sequenced using the forward primersEcp6_F2 and Ecp6_F in combination with the reverse primerEcp6_R3 (Table S2).
Bioinformatical analysis of Ecp6-like proteins
EST sequences from various fungal pathogens were down-loaded from the COGEME Phytopathogenic Fungi andOomycete EST Database version 1.6 (http://cogeme.ex.ac.uk) (Soanes and Talbot, 2006). The genome sequences ofvarious fungi listed in Table 2 were consulted at the websiteof Fungal Genome Initiative of the Broad Institute of MIT andHarvard (http://www.broad.mit.edu/annotation/fgi/) or at thewebsite of the USA Department of Energy Joint GenomeInstitute (http://genome.jgi-psf.org). The mining of Ecp6-like proteins was performed using NCBI BLAST, andthe Standalone-BLAST version 2.2.3 (Altschul et al., 1997;
Schäffer et al., 2001). Hmmpfam analysis of each identifiedcandidate was performed by running a customized Perl scriptfor Pfam HMM detection, available at ftp://ftp.sanger.ac.uk/pub/databases/Pfam, using Bioperl version 1.4 (http://bioperl.org) and HMMER version 2.3.2 (http://hmmer.janelia.org),which was loaded with the current Pfam ls and fs models(02.10.2007), for whole domain and fragment modelsrespectively. An E-value of 0.001 was used as cut-off. Theretained sequences were analysed in BioEdit version 7.0.5.3(http://www.mbio.ncsu.edu/BioEdit/bioedit.html). Multiplesequence alignment was performed by CLUSTALW version1.83 and for phylogenetic tree construction Molecular Evolu-tionary Genetic Analysis 4.0 (MEGA) was used (Kumar et al.,2001; Tamura et al., 2007). Phylogeny construction of fungalEcp6-like proteins was performed by neighbour-joininganalysis. We used p-distance as the distance parameter asspecified in the program MEGA. The inferred phylogeny wastested by 500 bootstrap replicates (Felsenstein, 1985).
Three-dimensional modelling was performed using theProtein Homology/analogY Recognition Engine (Phyre), aProtein fold recognition server (http://www.sbg.bio.ic.ac.uk/~phyre/; Kelley et al., 2000; Bennett-Lovsey et al., 2008).Estimated precision of generated models was used as anindication of significance. Subsequent analyses, visualizationand preparation of 3D figures were performed in the Swiss-PdbViewer version 3.7 (http://www.expasy.org/spdbv).
Acknowledgements
The authors thank Bert Essenstam and Henk Smid atUnifarm for excellent plant care. We acknowledge Bas Brand-wagt and John van’t Klooster for providing materials andassistance and three anonymous reviewers for usefulsuggestions. This project was carried out within the researchprogramme of the Centre of BioSystems Genomics (CBSG)which is part of the Netherlands Genomics Initiative/Netherlands Organization for Scientific Research. B.P.H.J.T.is supported by a Vidi grant of the Research Council for Earthand Life sciences (ALW) of the Netherlands Organization forScientific Research (NWO). I.S. is supported by ERA-PGproject ERA-PG 31855.00030.
References
van den Ackerveken, G.F.J.M., van Kan, J.A.L., Joosten,M.H.A.J., Muisers, J.M., Verbakel, H.M., and de Wit, P.J.G.M. (1993) Characterization of two putative pathogenicitygenes of the fungal tomato pathogen Cladosporiumfulvum. Mol Plant Microbe Interact 6: 210–215.
Altschul, S.F., Madden, T.L., Schäffer, A.A., Zhang, J.,Zhang, Z., Miller, W., and Lipman, D.J. (1997) GappedBLAST and PSI-BLAST: a new generation of protein databasesearch programs. Nucleic Acids Res 25: 3389–3402.
Amon, P., Haas, E., and Sumper, M. (1998) The sex-inducing pheromone and wounding trigger the same setof genes in the multicellular green alga Volvox. Plant Cell10: 781–789.
Arrighi, J.F., Barre, A., Ben Amor, B., Bersoult, A., Soriano,L.C., Mirabella, R., et al. (2006) The Medicago truncatulalysine motif-receptor-like kinase gene family includes NFP
Ecp6 is a virulence factor 15
© 2008 The AuthorsJournal compilation © 2008 Blackwell Publishing Ltd, Molecular Microbiology

and new nodule-expressed genes. Plant Physiol 142: 265–279.
Bateman, A., and Bycroft, M. (2000) The structure of a LysMdomain from E. coli membrane-bound lytic murein transg-lycosylase D (MltD). J Mol Biol 299: 1113–1119.
Bennett-Lovsey, R.M., Herbert, A.D., Sternberg, M.J.E., andKelley, L.A. (2008) Exploring the extremes of sequence/structure space with ensemble fold recognition in theprogram Phyre. Proteins: Structure, Function, Bioinformat-ics 70: 611–625.
Bielnicki, J., Devedjiev, Y., Derewenda, U., Dauter, Z.,Joachimiak, A., and Derewenda, Z.S. (2006) B. subtilisykuD protein at 2.0 A resolution: insights into the structureand function of a novel, ubiquitous family of bacterialenzymes. Proteins 62: 144–151.
Birkeland, N.K. (1994) Cloning, molecular characterization,and expression of the genes encoding the lytic functions oflactococcal bacteriophage phi LC3: a dual lysis system ofmodular design. Can J Microbiol 40: 658–665.
van den Burg, H.A., Westerink, N., Francoijs, K.J., Roth, R.,Woestenenk, E., Boeren, S., et al. (2003) Natural disulfidebond-disrupted mutants of AVR4 of the tomato pathogenCladosporium fulvum are sensitive to proteolysis, circum-vent Cf-4-mediated resistance, but retain their chitinbinding ability. J Biol Chem 278: 27340–27346.
van den Burg, H.A., Harrison, S.J., Joosten, M.H.A.J.,Vervoort, J., and de Wit, P.J.G.M. (2006) Cladosporiumfulvum Avr4 protects fungal cell walls against hydrolysis byplant chitinases accumulating during infection. Mol PlantMicrobe Interact 19: 1420–1430.
Butler, A.R., O’Donnell, R.W., Martin, V.J., Gooday, G.W., andStark, M.J. (1991) Kluyveromyces lactis toxin hasan essential chitinase activity. Eur J Biochem 199: 483–488.
Chang, J.H., Urbach, J.M., Law, T.F., Arnold, L.W., Hu, A.,Gombar, S., et al. (2005) Ahigh-throughput, near-saturatingscreen for type III effector genes from Pseudomonas syrin-gae. Proc Natl Acad Sci USA 102: 2549–2554.
Chenna, R., Sugawara, H., Koike, T., Lopez, R., Gibson, T.J.,Higgins, D.G., and Thompson, J.D. (2003) Multiplesequence alignment with the Clustal series of programs.Nucleic Acids Res 31: 3497–3500.
Ciuffetti, L.M., Tuori, R.P., and Gaventa, J.M. (1997) A singlegene encodes a selective toxin causal to the developmentof tan spot of wheat. Plant Cell 9: 135–144.
van Esse, H.P., Thomma, B.P.H.J., van’t Kooster, J.W., andde Wit, P.J.G.M. (2006) Affinity-tags are removed fromCladosporium fulvum effector proteins expressed in thetomato leaf apoplast. J Exp Bot 57: 599–608.
van Esse, H.P., Bolton, M.D., Stergiopoulos, I., de Wit, P.J.G.M., and Thomma, B.P.H.J. (2007) The chitin-bindingCladosporium fulvum effector protein Avr4 is a virulencefactor. Mol Plant Microbe Interact 20: 1092–1101.
Felsenstein, J. (1985) Confidence limits on phylogenies: anapproach using the bootstrap. Evolution 39: 783–791.
Finn, R.D., Tate, J., Mistry, J., Coggill, P.C., Sammut, S.J.,Hotz, H.R., et al. (2008) The Pfam protein familiesdatabase. Nucleic Acids Res 36: D281–D288.
Grant, S.R., Fisher, E.J., Chang, J.H., Mole, B.M., and Dangl,J.L. (2006) Subterfuge and manipulation: type III effectorproteins of phytopathogenic bacteria. Annu Rev Microbiol60: 425–449.
Guttman, D.S., Vinatzer, B.A., Sarkar, S.F., Ranall, M.V.,Kettler, G., and Greenberg, J.T. (2002) A functional screenfor the type III (Hrp) secretome of the plant pathogenPseudomonas syringae. Science 295: 1722–1726.
Haanstra, J.P.W., Laugé, R., Meijer-Dekens, F., Bonnema,G., de Wit, P.J.G.M., and Lindhout, P. (1999) The Cf-ECP2gene, conferring resistance to Cladosporium fulvum Cke.through recognition of the pathogenicity factor ECP2, islinked to, but not part of, the Cf-4/Cf-9 cluster on the shortarm of chromosome 1 of tomato. Mol Gen Genet 262:839–845.
Haanstra, J.P.W., Meijer-Dekens, F., Laugé, R., Seetanah,D.C., Joosten, M.H.A.J., de Wit, P.J.G.M., and Lindhout, P.(2000) Mapping strategy for resistance genes againstCladosporium fulvum on the short arm of chromosome 1of tomato: Cf-ECP5 near the Hcr9 Milky Way cluster. TheorAppl Genet 101: 661–668.
Hellens, R.P., Edwards, E.A., Leyland, N.R., Bean, S., andMullineaux, P.M. (2000) pGreen: a versatile and flexiblebinary Ti vector for Agrobacterium-mediated planttransformation. Plant Mol Biol 42: 819–832.
Hooykaas, P.J.J., Roobol, C., and Schilperoort, R.A. (1979)Regulation of the transfer of Ti-plasmids of Agrobacteriumtumefaciens. J Gen Microbiol 110: 99–109.
Jiang, R.H.Y., Tripathy, S., Govers, F., and Tyler, B.M. (2008)RXLR effector reservoir in two Phytophthora species isdominated by a single rapidly evolving super-family withmore than 700 members. Proc Natl Acad Sci USA 105:4874–4879.
Joosten, M.H.A.J., and de Wit, P.J.G.M. (1988) Isolation,purification and preliminary characterization of a proteinspecific for compatible Cladosporium fulvum (syn. Fulviafulva)–tomato interactions. Physiol Mol Plant Pathol 33:241–253.
Joosten, M.H.A.J., and de Wit, P.J.G.M. (1989) Identificationof several pathogenesis-related proteins in tomato leavesinoculated with Cladosporium fulvum (syn. Fulvia fulva) as1,3-beta-glucanases and chitinases. Plant Physiol 89:945–951.
Joosten, M.H.A.J., and de Wit, P.J.G.M. (1999) The tomato–Cladosporium fulvum interaction: a versatile experimentalsystem to study plant–pathogen interactions. Annu RevPhytopathol 37: 335–367.
Joosten, M.H.A.J., Cozijnsen, T.J., and de Wit, P.J.G.M.(1994) Host resistance to a fungal tomato pathogen lost bya single base-pair change in an avirulence gene. Nature367: 384–386.
Joosten, M.H.A.J., Vogelsang, R., Cozijnsen, T.J., Verberne,M.C., and de Wit, P.J.G.M. (1997) The biotrophic fungusCladosporium fulvum circumvents Cf-4-mediated resis-tance by producing unstable AVR4 elicitors. Plant Cell 9:367–379.
Jorda, L., Coego, A., Conejero, V., and Vera, P. (1999)Genomic cluster containing four differentially regulatedsubtilisin-like processing protease genes is in tomatoplants. J Biol Chem 274: 2360–2365.
Joris, B., Englebert, S., Chu, C.P., Kariyama, R., Daneo-Moore, L., Shockman, G.D., and Ghuysen, J.M. (1992)Modular design of the Enterococcus hirae muramidase-2and Streptococcus faecalis autolysin. FEMS Microbiol Lett70: 257–264.
16 M. D. Bolton et al. �
© 2008 The AuthorsJournal compilation © 2008 Blackwell Publishing Ltd, Molecular Microbiology

Kaku, H., Nishizawa, Y., Ishii-Minami, N., Akimoto-Tomiyama, C., Dohmae, N., Takio, K., et al. (2006) Plantcells recognize chitin fragments for defense signalingthrough a plasma membrane receptor. Proc Natl Acad SciUSA 103: 11086–11091.
Kamoun, S. (2006) A catalogue of the effector secretome ofplant pathogenic oomycetes. Annu Rev Phytopathol 44:41–60.
van Kan, J.A.L., van den Ackerveken, G.F.J.M., and de Wit,P.J.G.M. (1991) Cloning and characterization of cDNA ofavirulence gene Avr9 of the fungal pathogen Cladosporiumfulvum, causal agent of tomato leaf mold. Mol PlantMicrobe Interact 4: 52–59.
Kariyama, R., and Shockman, G.D. (1992) Extracellular andcellular distribution of muramidase-2 and muramidase-1 ofEnterococcus hirae ATCC 9790. J Bacteriol 174: 3236–3241.
Kelley, L.A., MacCallum, R.M., and Sternberg, M.J.E. (2000)Enhanced genome annotation using structural profiles inthe program 3D-PSSM. J Mol Biol 299: 501–522.
Knogge, W., and Scheel, D. (2006) LysM receptors recognizefriend and foe. Proc Natl Acad Sci USA 103: 10829–10830.
Kooman-Gersmann, M., Vogelsang, R., Hoogendijk, E.C.M.,and de Wit, P.J.G.M. (1997) Assignment of amino acidresidues of the AVR9 peptide of Cladosporium fulvum thatdetermine elicitor activity. Mol Plant Microbe Interact 10:821–829.
Krüger, J., Thomas, C.M., Golstein, C., Dixon, M.S., Smoker,M., Tang, S.K., et al. (2002) A tomato cysteine proteaserequired for Cf-2-dependent disease resistance and sup-pression of autonecrosis. Science 296: 744–747.
Kruijt, M., De Kock, M.J.D., and de Wit, P.J.G.M. (2005)Receptor-like proteins involved in plant disease resistance.Mol Plant Pathol 6: 85–97.
Kumar, S., Tamura, K., Jakobsen, I.B., and Nei, M. (2001)MEGA2: molecular evolutionary genetics analysis software.Bioinformatics 17: 1244–1245.
Kyte, J., and Doolittle, R.F. (1982) A simple method for dis-playing the hydropathic character of a protein. J Mol Biol157: 105–132.
Laugé, R., Joosten, M.H.A.J., van den Ackerveken, G.F.J.M.,van den Broek, H.W.J., and de Wit, P.J.G.M. (1997) The inplanta-produced extracellular proteins ECP1 and ECP2of Cladosporium fulvum are virulence factors. Mol PlantMicrobe Interact 10: 725–734.
Laugé, R., Joosten, M.H.A.J., Haanstra, J.P.W., Goodwin,P.H., Lindhout, P., and de Wit, P.J.G.M. (1998) Successfulsearch for a resistance gene in tomato targeted against avirulence factor of a fungal pathogen. Proc Natl Acad SciUSA 95: 9014–9018.
Laugé, R., Goodwin, P.H., de Wit, P.J.G.M., and Joosten,M.H.A.J. (2000) Specific HR-associated recognition ofsecreted proteins from Cladosporium fulvum occurs in bothhost and non-host plants. Plant J 23: 735–745.
Limpens, E., Franken, C., Smit, P., Willemse, J., Bisseling,T., and Geurts, R. (2003) LysM domain receptor kinasesregulating rhizobial Nod factor-induced infection. Science302: 630–633.
Longchamp, P.F., Mauel, C., and Karamata, D. (1994) Lyticenzymes associated with defective prophages of Bacillussubtilis: sequencing and characterization of the region
comprising the N-acetylmuramoyl-L-alanine amidase geneof prophage PBSX. Microbiology 140: 1855–1867.
Luderer, R., Takken, F.L.W., de Wit, P.J.G.M., and Joosten,M.H.A.J. (2002) Cladosporium fulvum overcomes Cf-2-mediated resistance by producing truncated AVR2 elicitorproteins. Mol Microbiol 45: 875–884.
Madsen, E.B., Madsen, L.H., Radutoiu, S., Olbryt, M.,Rakwalska, M., Szczyglowski, K., et al. (2003) A receptorkinase gene of the LysM type is involved in legume per-ception of rhizobial signals. Nature 425: 637–640.
Malonek, S., Rojas, M.C., Hedden, P., Gaskin, P., Hopkins,P., and Tudzynski, B. (2004) The NADPH-cytochromeP450 reductase gene from Gibberella fujikuroi is essentialfor gibberellin biosynthesis. J Biol Chem 279: 25075–25084.
Melin, P., Schnürer, J., and Wagner, E.G.H. (2003) Charac-terization of phiA, a gene essential for phialide develop-ment in Aspergillus nidulans. Fungal Genet Biol 40:234–241.
Mersereau, M., Pazour, G.J., and Das, A. (1990) Efficienttransformation of Agrobacterium tumefaciens by electro-portation. Gene 90: 149–151.
Miya, A., Albert, P., Shinya, T., Desaki, Y., Ichimura, K.,Shirasu, K., et al. (2007) CERK1, a LysM receptor kinase,is essential for chitin elicitor signaling in Arabidopsis. ProcNatl Acad Sci USA 104: 19613–19618.
Mulder, L., Lefebvre, B., Cullimore, J., and Imberty, A. (2006)LysM domains of Medicago truncatula NFP proteininvolved in Nod factor perception. Glycosylation state,molecular modeling and docking of chitooligosaccharidesand Nod factors. Glycobiology 16: 801–809.
Mullins, E.D., Chen, X., Romaine, P., Raina, R., Geiser, D.M.,and Kang, S. (2001) Agrobacterium-mediated transforma-tion of Fusarium oxysporum: an efficient tool for insertionalmutagenesis and gene transfer. Phytopathology 91: 173–180.
Nakayashiki, H., Hanada, S., Quoc, N.B., Kadotani, N., Tosa,Y., and Mayama, S. (2005) RNA silencing as a tool forexploring gene function in ascomycete fungi. Fungal GenetBiol 42: 275–283.
Ohnuma, T., Onaga, S., Murata, K., Taira, T., and Katoh, E.(2008) LysM domains from Pteris ryukyuensis chitinase-A:a stability study and characterization of the chitin-bindingsite. J Biol Chem 283: 5178–5187.
Pérez-García, A., Snoeijers, S.S., Joosten, M.H.A.J.,Goosen, T., and de Wit, P.J.G.M. (2001) Expression of theavirulence gene Avr9 of the fungal tomato pathogen Cla-dosporium fulvum is regulated by the global nitrogenresponse factor NRF1. Mol Plant Microbe Interact 14: 316–325.
Perfect, S.E., O’Connell, R.J., Green, E.F., Doering-Saad, C.,and Green, J.R. (1998) Expression cloning of a fungalproline-rich glycoprotein specific to the biotrophic interfaceformed in the Colletotrichum–bean interaction. Plant J 15:273–279.
Ponting, C.P., Aravind, L., Schultz, J., Bork, P., and Koonin,E.V. (1999) Eukaryotic signalling domain homologues inarchaea and bacteria. Ancient ancestry and horizontalgene transfer. J Mol Biol 289: 729–745.
Punt, P.J., Oliver, R.P., Digemans, M.A., Pouwels, P.H., andVan Den Hondel, C.A.M.J.J. (1987) Transformation of
Ecp6 is a virulence factor 17
© 2008 The AuthorsJournal compilation © 2008 Blackwell Publishing Ltd, Molecular Microbiology

Aspergillus based on the hygromycin B resistance markerfrom Escherichia coli. Gene 56: 117–124.
Radutoiu, S., Madsen, L.H., Madsen, E.B., Felle, H.H.,Umehara, Y., Gronlund, M., et al. (2003) Plant recognitionof symbiotic bacteria requires two LysM receptor-likekinases. Nature 425: 585–592.
Radutoiu, S., Madsen, L.H., Madsen, E.B., Jurkiewicz, A.,Fukai, E., Quistgaard, E.M., et al. (2007) LysM domainsmediate lipochitin-oligosaccharide recognition and Nfrgenes extend the symbiotic host range. EMBO J 26: 3923–39235.
Rep, M. (2005) Small proteins of plant-pathogenic fungisecreted during host colonization. FEMS Microbiol Lett253: 19–27.
Rooney, H.C.E., van’t Klooster, J.W., van der Hoorn, R.A.L.,Joosten, M.H.A.J., Jones, J.D.G. and de Wit, P.J.G.M.(2005) Cladosporium Avr2 inhibits tomato Rcr3 proteaserequired for Cf-2-dependent disease resistance. Science308: 1783–1786.
Ruhland, G.J., Hellwig, M., Wanner, G., and Fiedler, F.(1993) Cell-surface location of Listeria-specific proteinp60 – detection of Listeria cells by indirectimmunofluorescence. J Gen Microbiol 139: 609–616.
Saitou, N., and Nei, M. (1987) The neighbor-joining method:a new method for reconstructing phylogenetic trees. MolBiol Evol 4: 406–425.
Salamov, A.A., and Solovyev, V.V. (2000) Ab initio genefinding in Drosophila genomic DNA. Genome Res 10: 516–522.
Sambrook, J., and Russell, D. (2001) Molecular Cloning: ALaboratory Manual. Cold Spring Harbor, NY: Cold SpringHarbor Laboratory Press.
Schäffer, A.A., Aravind, L., Madden, T.L., Shavirin, S.,Spouge, J.L., Wolf, Y.I., et al. (2001) Improving theaccuracy of PSI-BLAST protein database searches withcomposition-based statistics and other refinements.Nucleic Acids Res 29: 2994–3005.
Segers, G., Bradshaw, N., Archer, D., Blissett, K., and Oliver,R.P. (2001) Alcohol oxidase is a novel pathogenicity factorfor Cladosporium fulvum, but aldehyde dehydrogenase isdispensable. Mol Plant Microbe Interact 14: 367–377.
Shevchenko, A., Wilm, M., Vorm. O., and Mann, M. (1996)Mass spectrometric sequencing of proteins from silver-stained polyacrylamide gels. Anal Chem 69: 850–858.
Smit, P., Limpens, E., Geurts, R., Fedorova, E., Dolgikh, E.,Gough, C., and Bisseling, T. (2007) Medicago LYK3, anentry receptor in rhizobial nodulation factor signaling. PlantPhysiol 145: 183–191.
Soanes, D.M., and Talbot, N.J. (2006) Comparative genomicanalysis of phytopathogenic fungi using expressedsequence tag (EST) collections. Mol Plant Pathol 7: 61–70.
Stergiopoulos, I., de Kock, M.J., Lindhout, P., and de Wit,P.J.G.M. (2007a) Allelic variation in the effector genes ofthe tomato pathogen Cladosporium fulvum reveals differ-ent modes of adaptive evolution. Mol Plant Microbe Inter-act 20: 1271–1283.
Stergiopoulos, I., Groenewald, M., Staats, M., Lindhout, P.,Crous, P.W., and de Wit, P.J.G.M. (2007b) Mating-typegenes and the genetic structure of a world-wide collectionof the tomato pathogen Cladosporium fulvum. FungalGenet Biol 44: 415–429.
Tamura, K., Dudley, J., Nei, M., and Kumar, S. (2007) MEGA4:Molecular Evolutionary Genetics Analysis (MEGA) software,version 4.0. Mol Biol Evol 24: 1596–1599.
Tang, X., Xiao, Y., and Zhou, J.M. (2006) Regulation of thetype III secretion system in phytopathogenic bacteria. MolPlant Microbe Interact 19: 1159–1166.
Thomma, B.P.H.J., van Esse, H.P., Crous, P.W., and de Wit,P.J.G.M. (2005) Cladosporium fulvum (syn. Passalorafulva), a highly specialized plant pathogen as a modelfor functional studies on plant pathogenic Mycos-phaerellaceae. Mol Plant Pathol 6: 379–393.
Thomma, B.P.H.J., Bolton, M.D., Clergeot, P.H., and de Wit,P.J.G.M. (2006) Nitrogen controls in planta expression ofCladosporium fulvum Avr9 but no other effector genes. MolPlant Pathol 7: 125–130.
Tornero, P., Conejero, V., and Vera, P. (1997) Identification ofa new pathogen-induced member of the subtilisin-like pro-cessing protease family from plants. J Biol Chem 272:14412–14419.
Wan, J., Zhang, X.C., Neece, D., Ramonell, K.M., Clough, S.,Kim, S.Y., et al. (2008) A LysM receptor-like kinase plays acritical role in chitin signaling and fungal resistance inArabidopsis. Plant Cell 20: 471–481.
Westerink, N., Brandwagt, B.F., de Wit, P.J.G.M., andJoosten, M.H.A.J. (2004) Cladosporium fulvum circum-vents the second functional resistance gene homologue atthe Cf-4 locus (Hcr9-4E) by secretion of a stable avr4Eisoform. Mol Microbiol 54: 533–545.
Whisson, S.C., Boevink, P.C., Moleleki, L., Avrova, A.O.,Morales, J.G., Gilroy, E.M., et al. (2007) A translocationsignal for delivery of oomycete effector proteins into hostplant cells. Nature 450: 115–118.
Whiteford, J.R., and Spanu, P.D. (2002) Hydrophobins andthe interactions between fungi and plants. Mol Plant Pathol3: 391–400.
de Wit, P.J.G.M. (1977) A light and scanning-electron micro-scopic study of infection of tomato plants by virulent andavirulent races of Cladosporium fulvum. Neth J PlantPathol 83: 109–122.
Wubben, J.P., Joosten, M.H.A.J., and de Wit, P.J.G.M.(1994) Expression and localization of two in planta inducedextracellular proteins of the fungal tomato pathogen Cla-dosporium fulvum. Mol Plant Microbe Interact 7: 516–524.
Xu, J.R., Peng, Y.L., Dickman, M.B., and Sharon, A. (2006)The dawn of fungal pathogen genomics. Annu Rev Phyto-pathol 44: 337–366.
Zhang, X.C., Wu, X., Findley, S., Wan, J., Libault, M.,Nguyen, H.T., et al. (2007) Molecular evolution of lysinmotif-type receptor-like kinases in plants. Plant Physiol144: 623–636.
Supplementary material
This material is available as part of the online article from:http://www.blackwell-synergy.com/doi/abs/10.1111/j.1365-2958.2008.06270.x(This link will take you to the article abstract).
Please note: Blackwell Publishing is not responsible for thecontent or functionality of any supplementary materials sup-plied by the authors. Any queries (other than missing material)should be directed to the corresponding author for the article.
18 M. D. Bolton et al. �
© 2008 The AuthorsJournal compilation © 2008 Blackwell Publishing Ltd, Molecular Microbiology




















