
The Navigation System of the Brain - Indian Academy of ...
15
401 RESONANCE May 2015 GENERAL ARTICLE The Navigation System of the Brain 2014 Nobel Prize in Physiology or Medicine Prasanna Venkhatesh V Keywords Hippocampus, cognitive map, place cells, grid cells. “It is fair to say that, in general, no problems have been exhausted; instead, men have been exhausted by the prob- lems. Soil that appears impoverished to one researcher re- veals its fertility to another. Fresh talent approaching the analysis of a problem without prejudice will always see new possibilities – some aspect not considered by those who believe that a subject is fully understood. Our knowledge is so frag- mentary that unexpected findings appear in even the most fully explored topics. We must bear in mind that because science relentlessly differentiates, the minutiae of today often become important principles tomorrow.” – Santiago Ramon y Cajal The ability to navigate in space is one of the fundamental func- tions of the brain. It depends on the ability to have a sense of position which in turn is interlinked with the sense of direction, distance and the knowledge of the earlier positions through which one has travelled. You depend on it for your everyday activities ranging from finding your car in a parking lot to commuting to your workplace. The 2014 Nobel Prize in Physiology or Medicine was awarded to John O’Keefe, May-Britt Moser and Edvard Moser for their discoveries of nerve cells in the brain that consti- tute a positioning system that enables one to have a sense of position and navigation (Figure 1). Edward Tolman’s Idea of Cognitive Maps Some of the early experiments on spatial learning of animals were performed by psychologist Edward C Tolman during the late 1940’s. Skinner’s behaviourist paradigm dominated psychology during that period and most psychologists focussed on functional relations between stimulus and response, without considering the Prasanna Venkhatesh V is currently a graduate student at the Center for Neuroscience working with Aditya Murthy. He is working on voluntary control of reaching and pointing movements. His research interests include movement control, neural basis of animal behaviour, optogenetics and social neuroscience.
-
Upload
khangminh22 -
Category
Documents
-
view
2 -
download
0
Transcript of The Navigation System of the Brain - Indian Academy of ...

401RESONANCE May 2015
GENERAL ARTICLE
The Navigation System of the Brain
2014 Nobel Prize in Physiology or Medicine
Prasanna Venkhatesh V
Keywords
Hippocampus, cognitive map,
place cells, grid cells.
“It is fair to say that, in general, no problems have been
exhausted; instead, men have been exhausted by the prob-
lems. Soil that appears impoverished to one researcher re-
veals its fertility to another. Fresh talent approaching the
analysis of a problem without prejudice will always see new
possibilities – some aspect not considered by those who believe
that a subject is fully understood. Our knowledge is so frag-
mentary that unexpected findings appear in even the most
fully explored topics. We must bear in mind that because
science relentlessly differentiates, the minutiae of today often
become important principles tomorrow.”
– Santiago Ramon y Cajal
The ability to navigate in space is one of the fundamental func-
tions of the brain. It depends on the ability to have a sense of
position which in turn is interlinked with the sense of direction,
distance and the knowledge of the earlier positions through which
one has travelled. You depend on it for your everyday activities
ranging from finding your car in a parking lot to commuting to
your workplace. The 2014 Nobel Prize in Physiology or Medicine
was awarded to John O’Keefe, May-Britt Moser and Edvard
Moser for their discoveries of nerve cells in the brain that consti-
tute a positioning system that enables one to have a sense of
position and navigation (Figure 1).
Edward Tolman’s Idea of Cognitive Maps
Some of the early experiments on spatial learning of animals were
performed by psychologist Edward C Tolman during the late
1940’s. Skinner’s behaviourist paradigm dominated psychology
during that period and most psychologists focussed on functional
relations between stimulus and response, without considering the
Prasanna Venkhatesh V is
currently a graduate
student at the Center for
Neuroscience working with
Aditya Murthy. He is
working on voluntary
control of reaching and
pointing movements. His
research interests include
movement control, neural
basis of animal behaviour,
optogenetics and social
neuroscience.

402 RESONANCE May 2015
GENERAL ARTICLE
role of internal representations. Although Tolman was firmly
behaviourist in his methodology, he wanted to use behavioural
methods to understand the mental processes of humans and other
animals.
His experiment which was conducted over several days involved
three groups of rats running through a maze. For Group 1, food
was kept at the end of the maze. For Group 2, no food was kept,
and for Group 3, food was kept only on day 11. The Group 1 rats
quickly learned to rush to the end of the maze without making
many errors, while Group 2 rats wandered in the maze but did not
preferentially go to the end. Rats in Group 3 essentially showed
the same behaviour as Group 2 rats until food was introduced on
day 11; they quickly learned to run to the end of the maze and
made fewer errors like the Group 1 rats by the next day. This
showed that the Group 3 rats had learned about the organisation
of the maze even without the reward.
Tolman reasoned that animals did not passively react to the
external stimuli. Rather, they learnt facts about the world and
used it as and when required. This suggested that there was some
kind of ‘latent learning’, i.e., they were using the knowledge in
the preceding trials to build a map and utilize it when they were
motivated to do so. He concluded that spatial learning consists of
building cognitive maps in the nervous system and are not mere
stimulus response connections and that these maps can range
Figure 1. Winners of 2014
Nobel Prize in Physiology or
Medicine: John O’Keefe (left),
May-Britt Moser (middle),
Edvard Moser (right).
Photocredits: O’Keefe: David
Bishop/UCL; M.-B. and E.
Moser: Geir Mogen/Kavli Insti-
tute for Systems Neuroscience.

403RESONANCE May 2015
GENERAL ARTICLE
from a simple narrow strip to a broader comprehensive map. This,
however, did not address where in the brain these functions may
be localizedand howthebraincomputes suchcomplex behaviours.
Tolman’s cognitive map theory faced fierce criticism from the
behaviourists who believed that complex behaviours in animals
are achieved by chains of sensory-motor response relationships.
The following is an excerpt from Tolman’s paper in 1948 p.192
on cognitive maps in rats and men.
“We believe that in the course of learning, something like a field
map of the environment gets established in the rat’s brain… The
stimuli … are usually worked over ... into a tentative, cognitive-
like map of the environment. And it is this tentative map, indicat-
ing routes and paths and environmental relationships, which
finally determines what responses, if any, the animal will finally
release.”
The Hippocampus and its Function
During the late 1950s, the majority of the clinical studies sug-
gested that the hippocampus (an area in the brain deep inside the
temporal lobe) by and large plays a fundamental role in long-term
associative memory. This idea was derived from the famous
report in 1957 by William B Scoville and Brenda Milner describ-
ing the results of surgical removal of the hippocampi in patient ‘H
M’ in an attempt to alleviate his epileptic seizures. After the
surgery, H M suffered from a severe anterograde amnesia (inabil-
ity to form new episodic memories) and partial retrograde amne-
sia (inability to recall memories that occurred before the surgery).
This report played an important role in creating the link between
the hippocampus and memory.
For a long time investigations were directed towards understand-
ing the underlying mechanisms and the role of the hippocampus
in the formation of memory. Several patterns of electrical activity
have been recorded from the hippocampus, and they have been
correlated with behavioural or psychological states. During the
mid-1960s, C H Vanderwolf placed a large electrode into the
For a long time,
investigationswere
directed towards
understanding the
underlying
mechanisms and the
role of hippocampus
in the formation of
memory.

404 RESONANCE May 2015
GENERAL ARTICLE
hippocampus of a freely moving rat and recorded the EEG
activity during a wide range of behaviours shown by the animal.
He found that the theta rhythm in the EEGmostly correlated with
certain behaviours of the animal like orienting, sniffing and
walking or not at all, during eating, drinking, grooming, quiet
sitting, and slow wave sleep. It was also known by then that rats
with lesions in the hippocampus had ‘spatial problem solving’
deficits. These results indicated that there was some sort of
spatial processing happening in the hippocampus. Refer to Box 1
for more on the history of hippocampal anatomy and function.
Discovery of Place Cells in the Hippocampus
In the late 1960’s, single-cell neuronal recordings in awake,
unrestrained rats was a cutting-edge technology. With the help of
the brain atlas, tiny wire electrodes were guided to specific areas
of interest in the brain. When the electrode tip is close to a neuron,
the electrode can record the action potentials from that neuron.
During the experiment the action potentials were recorded along
with x, y coordinates of the rat’s location as viewed from above
with the help of a small light fixed to the rat’s head. John
O’Keefe, who was by then expert at recording neurons using this
technique in awake-behaving rats, recorded the hippocampus
neuronal activity when the animal was doing a variety of
behaviours. Anyconservativeneurophysiologist at that timewould
have considered this experiment radical because sensory and
motor areas were easy and reliable targets and decoding the
Figure 2. a) Place cell in the
hippocampus. b) Grid cell in
the medial entorhinal cortex.
Spike locations are repre-
sented as red dots and su-
perimposed on the animal’s
trajectory in the recording
enclosure as black. Whereas
most place cells have a single
firing location, the firing fields
of a grid cell form a periodic
triangular matrix tiling the
entire environment available
to the animal.
Reprinted from Trends Cogn
Sci., 14 (12), Derdikman D and
Moser EI, A manifold of spatial
maps in the brain, 562, 2010,
with permission from Elsevier.

405RESONANCE May 2015
GENERAL ARTICLE
Box 1. History of Hippocampal Anatomy and Function
The hippocampus lies deep within the medial temporal lobes of the human brain with a group of neuronal
networks quite distinctly organised compared to the other areas of the brain.The Bolognese anatomist Giulio
Cesare Aranzi (circa 1564) was credited for coining the name ‘hippocampus’ to this area of brain because
of its remarkable appearance similar to that of a small marine fish that belongs to the genus Hippocampus.
Hippocampus is the greek name for a mythical sea monster, hippos meaning ‘horse’ and kampos meaning
‘sea monster’ in ancient greek. When the hippocampus is cut in cross section, it resembles a ram’s horn.
So ancient anatomists named it cornu ammonis (abbreviated as CA in modern nomenclature) which in latin
literally means horn of the ram. CA (subdivided as CA1, CA2,CA3 and CA4) is one of the two interlocking
gyri composing the hippocampus, the other being the dentate gyrus (DG). The unique arrangement of all
the cell population into single layers had attracted the attention of many investigators of the central nervous
system of the late nineteenth century. One of the pioneers to illustrate the unique organization of the
hippocampus is the Italian anatomist Camilio Golgi in 1886 (Figure A, left). Santiago Ramon y Cajal in
1911 clearly illustrated the hippocampus of rodents (Figure A, right) with arrows showing his interpre-
tation of the likely direction of the information flow using the staining method developed by Golgi. The
early anatomical studies of the hippocampus were important during the nineteenth century controversy
between the neuron doctrine (by Ramon y Cajal) and the reticular theory (by Golgi). Golgi used his
observations of the hippocampal formation to support his arguments for the reticular theory. During the
ninteenth century and the early twentieth century, the hippocampus was believed to have several functions
like olfactory, emotion and attentional control. The idea that the hippocampal formation is intimately
associated with memory came into existence mainly due to the observations made on the patient ‘H M’ by
William Scoville and Brenda Milner in 1957.
Figure A. (Left) Golgi's schematic drawing summarizing
the structure of the hippocampus which was published in
the book, Golgi, C. Sulla fina anatomia degli organi centrali
del sistema nervoso (On the Fine Structure of the Central
Organs of the Nervous System). Milan,Ulrico Hoepli, 1886.
Courtesy: Wikipedia.
(Right) The famous drawing of the hippocampus by
Santiago Ramon y Cajal was published in his book, Cajal
SR. Histologie du système nerveux de l’homme & des
vertébré (Histology of the nervous system of humans and
vertebrates), Paris, Maloine, 1909–1911.
Courtesy: https://archive.org (Identifier: histologiedusyst01ram),
Wikipedia.
1. Chapter 2, P Andersen, R Morris, D Amaral, T Bliss, J O’Keefe, The hippocampus book, Oxford University
Press, Oxford, 2006.
2. C Golgi, M Bentivogli, L Swanson, On the fine structure of the pes hippocampi major, Brain Res. Bull., Vol.54,
pp.461–483, Translated from Golgi C, 1886.
3. Prasanna Venkhatesh V, Santiago Ramon y Cajal: The Father of Neuroscience, Resonance, Vol.15, pp.968–
976, 2010.

406 RESONANCE May 2015
GENERAL ARTICLE
neuronal data and correlating it with the behaviour from areas
deep inside the brain was like finding a needle in a hay stack.
However, this approach proved quite fruitful for John O’Keefe
and his student Dostrovsky in 1971, because they discovered
neurons in the rat hippocampus that showed activity correlated to
the rat’s location within its environment. They called these neu-
rons ‘place cells’ (Figure 2a).
Place cells fire when the animal is moving in a specific location in
the environment, which corresponds to the place field of that
particular neuron. These place cells were first found in the
pyramidal cell layer of area CA1 of the hippocampus. Later, other
groups showed that area CA3 of the hippocampus also has place
cells. With extensive research, he concluded the following in his
1976 paper. Place cells firing in a part of the animal’s environ-
ment is not because of something the animal does there or because
of its motivation for going to that place. Rather, they appear to be
a cognitive process, signalling the animal’s position within an
environment irrespective of its behaviour and motivational state
or the reward properties of that place. The discovery of place cells
by John O’Keefe in the hippocampus created a new surge of
interest in the field in spite of the skepticism from a few investi-
gators [18].
The Influential Book by O’Keefe and Nadel
O’Keefe and Lynn Nadel authored an influential book called the
The Hippocampus as a Cognitive Map in 1978, delineating the
theoretical foundation for the hippocampal function and the
importance of the relationship between place cells and cognitive
map theory. According to the cognitive map theory, there must be
an abstract representation of the environment in the brain and the
map may be used by the animal to calculate efficient paths. The
theory put forth by O’Keefe and Nadel was influenced by Edward
Tolman’s theory about ‘cognitive maps’ in rats.
There was, however, a fundamental conceptual difference be-
tween Tolman’s original formulation of cognitive maps and the
cognitive map theory formulated in the book. In rodents, Tolman
Place cells fire
when the animal is
moving in a
specific location in
theenvironment,
which corresponds
to the place field of
that particular
neuron.

407RESONANCE May 2015
GENERAL ARTICLE
linked the cognitive maps to spatial information; in humans, he
associated cognitive maps more to events and emotions without
referring to a spatial framework. On the other hand, O’Keefe and
Nadel’s cognitive maps ascribed increased importance to spatial
framework in all animals and also asserted that the hippocampus
is the core of a neural memory system providing an objective
spatial framework within which the items and events of an
organism’s experience are located and interrelated. That is to say,
the hippocampus not only provides a spatial map but also pro-
vides a platform for embedding context-dependent memory.
The figures in the book and in the early papers were marginally
convincing, and there was no quantitative analysis. Only in the
mid 1980’s, computer-based data acquisition produced a con-
vincing presentation of the robust phenomenon of place cells.
Several studies have tried to identify the cues which are respon-
sible for the selective firing of neurons in the place field. It was
clear that the notion of place constructed by the place cells was at
least in part due to the cues available to the animal at that location.
Experimental results showed that place cells do not respond to
particular stimuli but will continue to indicate the animal’s posi-
tion as long as subsets of spatial cues are available. Also, loca-
tions in the environment were not mapped onto the hippocampus
in any orderly topographic manner. O’Keefe and Nadel envi-
sioned that if all the place cells are connected appropriately, it
would form a spatial map of the environment.
Place-Cell Remapping
A couple of papers by Bob Muller and colleagues in 1987 left
little room for the place-cell skeptics who criticised the spatially
correlated activity of these cells as being confounded by spatially
correlated cues. The main evidence they showed in their articles
was that, minimization of local sensory cues did not diminish
spatial specificity of neurons. In fact, this drove the field to such
an extreme, that most of the experiments were conducted in cue-
poor environments long after the convincing demonstration of re-
presentation of ‘space’ by ‘place cells’ and not anything sensory.
Experimental results
showed that place
cells do not respond
to particular stimuli
but will continue to
indicate the animal’s
position as long as
subsets of spatial
cues are available.

408 RESONANCE May 2015
GENERAL ARTICLE
These two papers also answered the following questions: How
are different environments represented in the hippocampus?When
an animal goes from one environment to a second environment,
how will the spatial maps respond? Are there rules and regulari-
ties? These papers also contained the discovery that place-cell
maps are completely different in two environments: a phenom-
enon later called ‘remapping’. They found that, if a place cell has
firing fields in two environments, knowing the location of the
firing field in the first environment will not predict the location of
the firing field in the second environment. Changing the shape of
an enclosure invariably induces full remapping. To be brief, the
hippocampal map of space is sensitive to non-spatial factors and
understanding these factors will be essential in decoding the
relationship between hippocampal maps and memory.
Head Direction Cells and Boundary Cells
In order to maintain the spatial orientation and to guide naviga-
tion, an animal must have knowledge of its location, displacement
or distance and direction from that location. By now, we know
that place cells could be used to identify familiar environmental
locations. Also, therewas a speculation that the oscillations of the
theta rhythm1 of the LFP2 (local field potentials) that occurred
during running could measure the distance. The missing piece in
the puzzle is that if the hippocampal system were to guide
navigation, it needed a sense of direction, which was predicted by
O’Keefe and Nadel in their 1978 book. In 1984, Jim Ranck
reported that cells in the post-subiculum of the rat brain dis-
charges whenever the animal’s head points to a specific direction,
independent of their location or behaviour, thus providing a
compass signal. Head direction cells are found in many brain
regions like the anterior thalamic nuclei, post and parasubiculum
and also in the entorhinal cortex. Two papers in 1990 from Jim
Ranck’s group studied in detail the properties of the head direc-
tion (HD) cells. They found that HD cells are strongly modulated
by environmental cues such as visual landmarks.
The vestibular system and the self movement cues play a critical
2 The local field potential (LFP)
refers to the electric potential in
the extracellular space around
neurons. The LFP is a widely
available signal in many record-
ing configurations, ranging from
single-electrode recordings to
multi-electrode arrays.
1 Theta rhythm is an oscillatory
pattern in electroencephalo-
graphy (EEG) signals recorded
either from inside the brain or
from electrodes glued to the
scalp. Two types of theta rhythm
have been described. The ‘hip-
pocampal theta rhythm’ is a
strong oscillation that can be
observed in the hippocampus
and other brain structures. ‘Cor-
tical theta rhythms’ are low-fre-
quency components of scalp
EEG, usually recorded from hu-
mans. Cortical theta rhythms
observed in human scalp EEG
are a different phenomenon, with
no clear relationship to the hip-
pocampus. In human EEG stud-
ies, the term theta refers to fre-
quency components in the 4–7
Hz range, and the hippocampal
theta refers to a cycle of 6–8 Hz
range.

409RESONANCE May 2015
GENERAL ARTICLE
role in the generation of head direction signals. HD cells provide
a continuous signal that an animal will use to guide its navigation
and maintain orientation. Neil Burgess and O’Keefe in 1996
demonstrated that the boundaries of an environment can elicit
modulation in the location-specific firing of place cells. They
stretched/compressed the walls of the chamber and correspond-
ingly the place fields remapped. Burgess and colleagues formu-
lated a ‘boundary vector cell’ model that not only explained the
existing data but successfully predicted how cells would respond
to new manipulations. It is remarkable that the model predicted
the existence of boundary related cells and almost 10 years after
the prediction, boundary cells were discovered in a number of
brain regions.
Hippocampal Circuitry and the Entorhinal Cortex
The neuronal antecedents to the place cells were not understood
clearly. The obvious line of investigation was whether the map is
located within the hippocampus or it is constructed elsewhere and
merely transferred to the hippocampus. Anatomically entorhinal
cortex is the major source of cortical input to the hippocampal
place cells. This is substantiated by the findings that entorhinal
damage causes serious deficits in spatial problem-solving tasks as
hippocampal lesions do.
Important insights about the role of the entorhinal cortex as a
major input to the place cells came from lesion studies on the
dentate gyrus (DG) as reported in McNaughton et al, 1989. They
found that the place cells fired sharply even after a major lesion in
dentate gyrus, while the connections of the CA3 and the entorhinal
cortex were still intact. Clear evidence came from the Moser
group (led by EdwardMoser and May-Britt Moser) in 2002; they
showed that even when the connections of the CA3 and the CA1
were disrupted, CA1 still exhibited spatial firing. This negated
the possibility that the internal hippocampal circuitry was in-
volved in the computation of the spatial signals. This study
suggested that, the likely input to the CA1 place cells in the dorsal
hippocampus is from the entorhinal cortex. The connectivity of
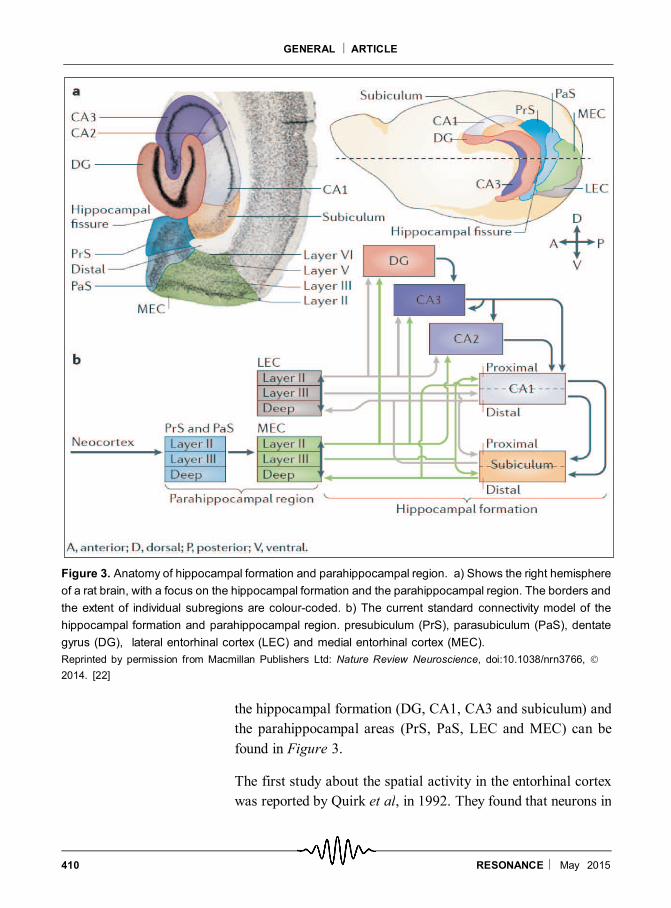
410 RESONANCE May 2015
GENERAL ARTICLE
the hippocampal formation (DG, CA1, CA3 and subiculum) and
the parahippocampal areas (PrS, PaS, LEC and MEC) can be
found in Figure 3.
The first study about the spatial activity in the entorhinal cortex
was reported by Quirk et al, in 1992. They found that neurons in
Figure 3. Anatomy of hippocampal formation and parahippocampal region. a) Shows the right hemisphere
of a rat brain, with a focus on the hippocampal formation and the parahippocampal region. The borders and
the extent of individual subregions are colour-coded. b) The current standard connectivity model of the
hippocampal formation and parahippocampal region. presubiculum (PrS), parasubiculum (PaS), dentate
gyrus (DG), lateral entorhinal cortex (LEC) and medial entorhinal cortex (MEC).
Reprinted by permission from Macmillan Publishers Ltd: Nature Review Neuroscience, doi:10.1038/nrn3766,
2014. [22]

411RESONANCE May 2015
GENERAL ARTICLE
the medial entorhinal cortex that project to intermediate hippoc-
ampus displayed weak spatial modulation compared to that of the
hippocampal cells. Anatomically, the entorhinal cortex contains
two major subdivisions – the medial and the lateral. By the late
90’s it became clear that the medial entorhinal cortex (MEC)
shows a dorsal-to-ventral organization in which the most dorsal
parts project exclusively to the dorsal hippocampus and the most
ventral parts only to theventral hippocampus. Thelateralentorhinal
cortex (LEC) shows a medial-to-lateral organization of projec-
tions to the hippocampus. Until 2004, most of the recordings of
spatial cells in the entorhinal cortex were from intermediate or
ventral parts of either LEC or MEC. These studies were not ideal
because, the ventral region projects primarily to the ventral
hippocampus, where place fields are large, and exhibit minimal
spatial variation in small environments. The cells from the most
dorsal parts of the entorhinal cortex, which provide the connec-
tions to the dorsal hippocampus where place cells were discov-
ered, were not investigated. With the help of the neuroanatomist,
Menno Witter, the Mosers in 2004 recorded from the dorso-caudal
region of the medial entorhinal cortex. They found that superficial
layers showed discrete firing fields (only the cell layers II and III of
the dorso-caudal medial entorhinal cortex (dMEC)) of the animal’s
current location as that of the place cells with each cell having
multiple firing fields. This study confirmed the hypothesis that the
spatial information is processed upstream of the hippocampus.
Discovery of Grid Cells in the Entorhinal Cortex
The Moser group noticed that the fields in the 2004 recordings
from the dMEC followed a regular organization with optimal
distance between them. In order to have a better understanding of
the spatial organization of the firing fields, the same cells were
subsequently recorded in larger environments, including boxes
with surface areas that were more than three times larger than
those of previous studies.
In 2005, the Mosers published a paper that provided a detailed
description of the spatial properties of the dMEC. The most
The most striking
and remarkable
observation in that
study was the
spatial regularity (a
tessellated
pattern) of the
neuronal firing

412 RESONANCE May 2015
GENERAL ARTICLE
striking and remarkable observation in that study was the spatial
regularity (a tessellated pattern) of the neuronal firing in the layer
II of the dMEC while the rat was walking in the enclosure (Figure
2b). The firing fields of the neurons in the layer II of dMEC
formed a beautiful grid like structure of tessellating equilateral
triangles; in other words the firing locations form the vertices of a
hexagonal grid, representing the entire space traversed by the animal.
The basic unit of the tessellated spatial firing pattern is called a grid
and the cells that elicit such a firing pattern are called ‘grid cells’. The
characteristic features of the grids are thespatial phase, spacingof the
grid and their orientation (Figure 4). The temporal and spatial auto
correlation analysis suggested that the recurring peaks in the
auto-correlograms did not reflect artefacts in the analytical proce-
dures.
Properties of Grid Cells
The questions that immediately arose are the following: how is
the grid cell population organized anatomically? Is the same
pattern of spatial firing seen in all layers? What are the factors
that determine the spacing, orientation and the size of the field?
The Moser group found that the spacing and the field size
increased from the dorsal to the ventral part of the dMEC. A key
question that pops up is, how much of external cues contribute to
the locations of the discharge by grid cells, and also whether the
cues are necessary to maintain the grid structure. Repeated tests
in the same environment showed that the vertices of the grids are
located at identical positions.
Figure 4. Parameters that
define grid cells. Cartoons of
firing patterns of pair grid cells
shown in green and blue.
Spatial phase is the position
of the gird vertices in the x–y
plane. (Left panel) Black and
red crosses represent the
phases of two different grid
cells. (Middle panel) Spacing
in the grids is the distance
between anyvertex of the grid
and the six adjacent vertices
in the rate map or in the
autocorrelogram. (Right
panel) Orientation of the grid
vertices is defined by the lines
that intersect the grid verti-
ces indicated in black and
red.
Reprinted by permission from
Macmillan Publishers Ltd: Na-
ture Review Neuroscience ,
doi:10.1038/nrn3766, 2014
[22].

413RESONANCE May 2015
GENERAL ARTICLE
However, when the landmarks are rotated, the grids rotate. Visual
cue deprivation studies showed that the grid was maintained even
in darkness and the grid space remains the same. Then the
question is – what determines the phase and orientation of the grid
cells? Although configuration of the environment may not be
essential for producing a grid pattern, what seem to be the
essential determinant of the phase and orientation are the land-
marks.
There are two conclusions which are derived from these results;
first, the external cues exert a significant influence over the phase
and orientation. Second, from the visual deprivation studies, the
authors concluded that self motion (vestibular-kinesthetic)3 is the
only likely source of maintained discharge in the grid cells in
moving animals. Also, the invariance of the firing pattern for
change in direction and velocity explains that the changes in
velocity and heading must be integrated over time to enable a
constant representation of spatial relationship between positions
which is called ‘path integration’4 [10,22].
Latest Findings
I am listing here some of the interesting recent findings in the grid
cell and place cell literature. The readers can look into the recent
reviews for more information. [19–23]
1. In 2011, Deshmukh and Knierim discovered that the Lateral
Entorhinal Cortex (LEC) represents both spatial and non-spatial
information. They found that some LEC neurons fire at the
location of the objects in the environment as well as showing that
LEC has ‘place cells’ in the presence of objects. This is an
important contribution because LEC and MEC may be providing
complementary spatial information to the hippocampus – MEC
provides path integration derived space while LEC provides
sensory derived space (as well as object related information).
2. Eichenbaum’s group, in 2011, reported that cells in the
hippocampal formation code for time and they called these ‘time
cells’.
4 Path integration is the process
of summing up information about
direction and distance travelled,
in order to keep track of one’s
relative position. Path integra-
tion is hypothesized to be the
basis for the formation of the
‘place code’ the hippocampus
uses to encode spatial memo-
ries. ‘Dead reckoning,’ tradition-
ally used by seafarers uses a
combination of path integration
(compass direction and speed
measured in ‘knots’) and taking
a fix on actual position when
‘landmarks’ are available. This
is akin to the emerging pattern
of MEC representing path inte-
gration derived space while LEC
representing landmark derived
space.
3 The vestibular senses (the sen-
sations of body rotation and of
gravitation and movement) arise
in the inner ear; the sense or-
gans are the hair cells that send
out signals over the auditory
nerve. Movement of the body’s
muscles, tendons, and joints is
also monitored by mechanore-
ceptors in these structures and
this process is called kinesthe-
sis.

414 RESONANCE May 2015
GENERAL ARTICLE
3. Recent work has shown that within a single animal, grid cells
can be clustered into 4 or 5 modules, such that grid scale increases
in a discrete fashion between one module to the other. The other
interesting finding is the fact that, when the grid spacing in each
module is defined by the dimensions of a fitted ellipse, the ratio
grows in leaps of ~ 2. The geometric progression defined by this
constant scale factor has been suggested to be optimal for repre-
senting environments at high spatial resolution.
4. There are several computational models that try to understand
the relationship between the grid cells, head direction cells,
border cells and place cells.
5. The grid and place cell systems are found in many mammalian
species including humans. Recently researchers have found place-
like cells in hippocampus and grid cells in the entorhinal cortex of
other mammalian systems like non-human primates, bats and
humans by directly recording from these areas. The fact that there
exists a similarity inmostmammals, in thehippocampal-entorhinal
structure (with navigational capacity) suggests that the place cell-
grid cell system is robust, functional and conserved in vertebrate
evolution.
Concluding Remarks
The discovery of place cells and grid cells by John O’Keefe,
Edvard Moser and May-Britt Moser has clearly revolutionised
our understanding of the cognitive functions of the brain, espe-
cially the spatial navigation. These findings open a lot of avenues
for future research in both memory and spatial navigation, since
the link between the two are yet to be understood.
Acknowledgements
I would like to thank
Sachin Deshmukh and
Kousik Sarathy for the
critical comments and for
the discussions. I also ac-
knowledge Divija Rao,
Namrata Iyer and Ranjani
Seshadri for their helpful
inputs.
Suggested Reading
[1] E C Tolman, Cognitive maps in rats and men, Psychol Bull., Vol.55, pp.189–208, 1948.
[2] W B Scoville, B Milner, Loss of recent memory after bilateral hippocampal lesions, J Neurol Neurosurg
Psychiatry, Vol.20, pp.11–21, 1957.
[3] J O’Keefe, J Dostrovsky, The hippocampus as a spatial map: preliminary evidence from unit activity in the
freely-moving rat, Brain Res., Vol.34, pp.171–175, 1971.
[4] J O’Keefe, Place units in the hippocampus of the freely moving rat, Exp Neurol., Vol.51, pp.78–109, 1976.

415RESONANCE May 2015
GENERAL ARTICLE
[5] J O’Keefe, L Nadel, The hippocampus as a cognitive map. Clarendon, OxfordUniversity Press, 1978. (Free online
Version: http://www.cognitivemap.net/)
[6] R U Muller, J L Kubie, The effects of changes in the environment on the spatial firing of hippocampal complex-
spike cells, J Neurosci, Vol.7, pp.1951–1968, 1987.
[7] J S Taube, R U Muller and J B Ranck Jr, Head-direction cells recorded from the postsubiculum in freely moving
rats, I. Effects of environmental manipulations, J Neurosci, Vol.10, pp.436–447, 1990a.
[8] V H Brun, M K Otnass, S Molden, H-A Steffenach, M P Witter M-B Moser and EI Moser, Place cells and place
recognition maintained by direct entorhinal-hippocampal circuitry, Science, Vol.296, pp.2243–2246, 2002.
[9] M Fyhn, SMolden, M PWitter, EI Moser andMBMoser, Spatial representation in the entorhinal cortex, Science,
Vol.305, pp.1258–1264, 2004.
[10] T Hafting, M Fyhn, S Molden, MB Moser, EI Moser, Microstructure of a spatial map in the entorhinal cortex,
Nature, Vol.436, pp.801–806, 2005.
[11] T Solstad, C Boccara, E Kropff, MB Moser, EI Moser, Representation of geometric borders in the entorhinal
cortex, Science, Vol.322, pp.1865–1868, 2008.
[12] S S Deshmukh, J J Knierim, Representation of non-spatial and spatial information in the lateral entorhinal
cortex, Front Behav Neurosci, Vol.5, p.69, 2011.
[13] M M Yartsev, M P Witter and N Ulanovsky, Grid cells without theta oscillations in the entorhinal cortex of bats,
Nature, Vol.479, pp.103–107, 2011.
[14] C JMacDonald, K QLepage, U T Eden and H Eichenbaum, Hippocampal “time cells” bridges the gap inmemory
for discontiguous events, Neuron, Vol.71, pp.737–749, 2011.
[15] N J Killian, M J Jutras and E A Buffalo, A map of visual space in the primate entorhinal cortex, Nature,Vol.491,
pp.761–764, 2012.
[16] J Jacobs, C T Weidemann, J F Miller, A Solway, J F Burke, X X Wei, N Suthana, M R Sperling, A D Sharan, I
Fried and M J Kahana, Direct recordings of grid-like neuronal activity in human spatial navigation, Nat
Neurosci, Vol.16, pp.1188–1190, 2013.
[17] M M Yartsev, N Ulanovsky, Representation of three-dimensional space in the hippocampus of flying bats,
Science, Vol.340, pp.367–372, 2013.
[18] P J Best, AMWhite, Placing hippocampal single-unit studies in a historical context, Hippocampus, Vol.9, pp.346–
351, 1999.
[19] E I Moser, E Kropff, M B Moser, Place cells, grid cells, and the brain’s spatial representation system, Annu Rev
Neurosci., Vol.31, pp.69–89, 2008.
[20] L M Giocomo, M Moser, E Moser, Computational models of grid cells, Neuron, Vol.71, pp.589–603, 2011.
[21] D Derdikman, J J Knierim (eds.) Space, Time andMemory in the Hippocampal Formation, Springer (Wien), 2014.
[22] E I Moser, Y Roudi, M PWitter, CKentros, T Bonhoeffer and MBMoser, Grid cells and cortical representation,
Nature Rev Neuroscience, Vol.15, pp.466–481, 2014.
[23] G S Maya, L Liora, Y Yossi and N Ulanovsky, Spatial cognition in bats and rats: from sensory acquisition to
multiscale maps and navigation, Nature Rev Neuroscience, Vol.16, pp.94–108, 2015.
Address for Correspondence
Prasanna Venkhatesh V
Center for Neuroscience
Indian Institute of Science
Bengaluru 560 012, India
Email: [email protected]




















