
The NADP-glutamate dehydrogenase of the cyanobacterium Synechocystis 6803: cloning, transcriptional...
16
Plant Molecular Biology 28: 173-188, 1995. © 1995 Kluwer Academic Publishers. Printed in Belgium. 173 The NADP-glutamate dehydrogenase of the cyanobacterium Synechocystis 6803: cloning, transcriptional analysis and disruption of the gdhA gene Sebastian Ch~tvez 1,3, Jose Carlos Reyes 1, Franck Chauvat 2, Francisco J. Florencio 1 and Pedro Candau 1,. 1Departamento de Bioquimica Vegetal y Biologia Molecular and Instituto de Bioquimica Vegetal y Fotos[ntesis, Universidad de Sevilla and CSIC, Apartado 1113, 41080 Sevilla, Spain (*author for correspondence); 2Service de Biochimie et G~n~tique Mol&ulaire, D~partement de Biologie Cellulaire et Mol&ulaire, C.E. Saclay, 91191 Gif sur Yvette cedex, France; 3present address: Institut ffir Molekularbiologie und Tumorforschung, Phillips-Universitgit, 3503 7 Marburg, Germany Received 4 August 1994; accepted in revised form 1 March 1995 Key words: cyanobacteria, gdhA mutant, glutamate dehydrogenase, nitrogen metabolism, growth-phase-specific regulation, Synechocystis PCC 6803 Abstract The gdhA gene of Synechocystis PCC 6803, which encodes an NADP-dependent glutamate dehydroge- nase (NADP-GDH), has been cloned by complementation of an Escherichia coli glutamate auxotroph. This gene was found to code for a polypeptide of 428 amino acid residues, whose sequence shows high identity with those of archaebacteria (42-47 ~o), some Gram-positive bacteria (40-44 ~) and mammals (37~o). The minimal fragment of Synechocystis DNA required for complementation (2 kb) carries the gdhA gene preceded by an open reading frame (ORF2) encoding a polypeptide of 130 amino acids. ORF2 and gdhA are co-transcribed as a 1.9 kb mRNA, but shorter transcripts including only gdhA were also detected. Two promoter regions were identified upon transcriptional fusion to the cat reporter gene of a promoter probe plasmid. Transcription from the promoter upstream of ORF2 was found to be regu- lated depending on the growth phase of Synechocystis, in parallel to NADP-GDH activity. This promoter is expressed in Escherichia coli too, in contrast to the second promoter, located between ORF2 and gdhA, which was silent in E. coli and did not respond to the stage of growth in Synechocystis. Disruption of the cyanobacterial gdhA gene with a chloramphenicol resistance cassette yielded a mutant strain totally lacking NADP-GDH activity, demonstrating that this gene is not essential to Synechocystis 6803 under our laboratory conditions. The nucleotide sequence data reported will appear in the EMBL, GenBank and DDBJ Nucleotide Sequence Databases under the accession number X77454.
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of The NADP-glutamate dehydrogenase of the cyanobacterium Synechocystis 6803: cloning, transcriptional...

Plant Molecular Biology 28: 173-188, 1995. © 1995 Kluwer Academic Publishers. Printed in Belgium. 173
The NADP-glutamate dehydrogenase of the cyanobacterium Synechocystis 6803: cloning, transcriptional analysis and disruption of the gdhA gene
Sebastian Ch~tvez 1,3, Jose Carlos Reyes 1, Franck Chauvat 2, Francisco J. Florencio 1 and Pedro Candau 1,. 1Departamento de Bioquimica Vegetal y Biologia Molecular and Instituto de Bioquimica Vegetal y Fotos[ntesis, Universidad de Sevilla and CSIC, Apartado 1113, 41080 Sevilla, Spain (*author for correspondence); 2 Service de Biochimie et G~n~tique Mol&ulaire, D~partement de Biologie Cellulaire et Mol&ulaire, C.E. Saclay, 91191 Gif sur Yvette cedex, France; 3present address: Institut ffir Molekularbiologie und Tumorforschung, Phillips- Universitgit, 3503 7 Marburg, Germany
Received 4 August 1994; accepted in revised form 1 March 1995
Key words: cyanobacteria, gdhA mutant, glutamate dehydrogenase, nitrogen metabolism, growth-phase-specific regulation, Synechocystis PCC 6803
Abstract
The gdhA gene of Synechocystis PCC 6803, which encodes an NADP-dependent glutamate dehydroge- nase (NADP-GDH), has been cloned by complementation of an Escherichia coli glutamate auxotroph. This gene was found to code for a polypeptide of 428 amino acid residues, whose sequence shows high identity with those of archaebacteria (42-47 ~o), some Gram-positive bacteria (40-44 ~ ) and mammals (37~o). The minimal fragment of Synechocystis DNA required for complementation (2 kb) carries the gdhA gene preceded by an open reading frame (ORF2) encoding a polypeptide of 130 amino acids. ORF2 and gdhA are co-transcribed as a 1.9 kb mRNA, but shorter transcripts including only gdhA were also detected. Two promoter regions were identified upon transcriptional fusion to the cat reporter gene of a promoter probe plasmid. Transcription from the promoter upstream of ORF2 was found to be regu- lated depending on the growth phase of Synechocystis, in parallel to NADP-GDH activity. This promoter is expressed in Escherichia coli too, in contrast to the second promoter, located between ORF2 and gdhA, which was silent in E. coli and did not respond to the stage of growth in Synechocystis. Disruption of the cyanobacterial gdhA gene with a chloramphenicol resistance cassette yielded a mutant strain totally lacking NADP-GDH activity, demonstrating that this gene is not essential to Synechocystis 6803 under our laboratory conditions.
The nucleotide sequence data reported will appear in the EMBL, GenBank and DDBJ Nucleotide Sequence Databases under the accession number X77454.

174
Introduction
Ammonium is an obligatory intermediate in the assimilation of inorganic sources of nitrogen by every plant and microorganism studied so far. One of the main products resulting from ammo- nium incorporation into carbon-skeletons is L-glutamate, which acts as a major distributor of amino groups in the biosynthetic metabolism. The assimilation of ammonium into L-glutamate is carried out either by the sequential action of glutamine synthetase (GS) and glutamate syn- thase (GOGAT) or by glutamate dehydrogenase (GDH). A dependence on the intracellular con- centration of ammonium has been proposed to explain the operativity of these two pathways, based on the weaker affinity of GDH to ammo- nium as compared to that of GS [40]. This is certainly the case in enterobacteria, where genetic evidence clearly shows that the GDH pathway is only functional at ammonium concentrations higher than 1 mM [62].
The GS-GOGAT cycle is also the main path- way for L-glutamate synthesis in higher plants, algae and photosynthetic prokaryotes [reviewed in 69]. In many of these organisms one or more enzymes with GDH activity are also present, and in some of them the reductive amination of 2-oxoglutarate by a GDH is predominant; such seems to be the role of the ammonium-inducible chloroplastic NADP-GDH of Chlorella sorokini- ana [4]. On the other hand, the NAD-dependent glutamate dehydrogenase (NAD-GDH) of higher plants plays a catabolic function, supplying car- bon skeletons to the Krebs cycle under carbon- limited conditions [63 ]. Even though the NADP- GDH-coding gene of Chlorella sorokiniana has been cloned [ 16], no null mutant for this activity is available in that alga, while genetic studies with plant GDHs are scarce and inconclusive due to secondary phenotypical features [45].
In cyanobacteria, the GS-GOGAT cycle is the general pathway of ammonium assimilation [50] but several species also show GDH activity [57]. This is the case ofSynechocystis PCC 6803, which, besides a fully active and regulated GS-GOGAT pathway [52], presents a significant level of
NADP-GDH [29] and a minor one of NAD- GDH [ 15]. The kinetic features of these enzymes [ 15, 29] and the lack of a complete Krebs cycle [58] argue against catabolic roles for cyanobac- terial GDHs. To study their role and since Syn- echocystis PCC 6803 is well suited for genetic ma- nipulation [76], we decided to initiate the genetic analysis of NADP-GDH in this organism. As a first step, we have cloned and sequenced its struc- tural gene (gdhA), which was then disrupted yielding the first NADP-GDH null mutant from a photosynthetic organism, as far as we know. In this article we also report the heterologous ex- pression of this gene in Escherichia coli, as well as the analysis of its two promoter regions.
Materials and methods
Strains and plasmids
Synechocystis PCC 6803 was obtained from In- stitut Pasteur (Paris, France). Cyanobacterial re- combinant strains were generated from Syn- echocystis SFCf~5 [14]. Escherichia coli DH5~ [33] was the host for plasmid constructions and E. coli CLR207recA [47] was the glutamate aux- otroph used for complementation experiments.
The E. coli-Synechocystis PCC6803 shuttle plasmids p F F l l , pFF11T and p F F l l L were de- scribed in [27]. pBG1 expresses the E. coli gdhA gene [47]. Other plasmids used in this work were derivatives of pBluescript II SK(+) (Stratagene, La Jolla, CA).
The minimal medium described in [55] was used for growing the CLR207recA strain. The concentration of supplementing amino acids and antibiotics, as well as the general conditions of E. coli growth were standard [64]. Synechocystis PCC 6803 was cultivated as described [13].
DNA manipulation and Southern blot hybridization
Isolation of total DNA of Synechocystis 6803 was essentially done as described [71]. Restriction and DNA-modifying enzymes were obtained from Boehringer Mannheim or Pharmacia, and

used according to the manufacturer's recommen- dations. Plasmid isolation from E. coli and re- combinant DNA procedures were standard [64]. For Southern hybridizations, 1 #g of restricted DNA was digested, electrophoresed in 0.7~o agarose gels, and transferred under vacuum to Z- Probe membranes (BioRad Laboratories). DNA probes labelled with digoxigenine were hybridized according to the protocol of the Dig-labelling and hybridization kit from Boehringer Mannheim.
Preparation of the libraries
Library A was constructed as follows: total DNA of Synechocystis 6803 was partially digested with Hind III and gel-purified fragments over 3 kb were cloned into pBluescript II SK(+) which had been opened with the same enzyme and treated with calf intestinal phosphatase to prevent self- ligation of the vector. The resulting ligation mix- ture was used to transform an mcrAB mutant of E. coli DH5c~, to avoid degradation of cyanobac- terial DNA by this restriction system [27]. A fur- ther plasmid isolation from more than 5000 in- dependent transformants constituted this library. Library B was constructed in a similar way, ex- cept that Sau 3AI partial digest of cyanobacterial DNA was inserted into the Barn HI site ofpBlue- script II SK(+).
DNA sequencing
A complete series of nested deletions was intro- duced into the cyanobacterial insert (pSCh4) that complements the glutamate auxotrophic mutant ofE. coli, using exonuclease III as described [35]. Sequence was determined in both directions by the dideoxy chain termination method [65 ], using [35S ]-dATP and a Sequenase 2.0 kit from United States Biochemical Corporation.
RNA isolation and northern blot analysis
Total RNA was isolated from late-exponential cultures of Synechocystis 6803 as described [54],
175
except that cells were frozen in liquid nitrogen and ground in a mortar before lysis. Total RNA aliquots containing 15/~g were electrophoresed in formaldehyde gels, transferred onto nylon mem- branes (Hybond N-plus, Amersham), and hybrid- ized according to instruction manuals from Amersham. To reuse the filter, the probe was re- moved by 5 min boiling in 0.1 ~o S D S.
Transformation procedures
Competent cells of E. coli were prepared accord- ing to Hanahan [33]. Transformation of Syn- echocystis 6803 was achieved following the method described [ 13]. Efficient plasmid-DNA transformation was obtained by using the strain SFCf~5 that harbours the recipient plasmid pFCf~5 [ 14] that is homologous to the incoming vector. This allowed for the rescue of the enter- ing plasmid by recombination with the resident one [27]. Plasmid DNA was then isolated from the transformants as described [13] and ampli- fied in E. coli DH5emcrAB prior to further mo- lecular analysis.
Enzymatic assay
NADP-GDH aminating activity was assayed in vitro as previously described in [29], except that 10 mM CaC12 was included in the reaction mix- ture. One unit of NADP-GDH activity corre- sponds to the amount of enzyme that catalyses the oxidation of 1 ~tmol of NADPH per minute Native electrophoresis gels (6.25~o acrylamide) were stained for NADP-GDH activity as follows: after incubating for 10 min in a 2 mM NADPH solution, gels were transferred to a Pyrex dish containing the additional components of the re- action mixture and exposed to UV light. Dark bands produced by oxidation of NADPH were photographically documented. Controls without ammonium were performed to ensure results specificity.
CAT activity was assayed in vitro at 37 °C by the spectrophotometric method of Shaw [66].

176
One unit of CAT activity corresponds to the amount of enzyme that catalyses the acetylation of 1/lmol of chloramphenicol per minute. To ob- tain cell-free extracts of E. coli for NADP-GDH assays and of Synechocystis 6803 for both NADP-GDH and CAT assays, cells were har- vested by centrifugation at 7000 x g for 10 min, resuspended in 50 mM HEPES-NaOH buffer (pH 7.0) containing 10 mM CaC12 and 1 mM PMSF, and disrupted by sonic treatment. The preparation of broken cells was centrifuged for 15 min at 10 000 × g and the resulting supernatant constituted the cell-free extract. Cell-free extracts of E. coli for CAT assays were obtained as de- scribed [27]. Protein concentration was mea- sured in cell-free extracts following [10] and in whole cells using the method described [46]. Ovalbumin was used as standard. Chlorophyll concentration in the cultures was quantified as described [42].
Computer programs
Homology searches in the EMBL database were performed using the FastA algorithm [59]. Mul- tiple alignment of protein sequences was made by using the CLUSTAL V program [36] with the PAM250 scoring matrix [21] and a gap penalty of 7 + 10, and the PileUp program of the GCG package [23] using standard gap penalties. For applying the CodonPreference algorithm [30] we previously calculated the average codon usage of Synechocystis 6803 by using the Codonfrequency program contained in the GCG package [23]. The following protein-coding sequences from this cyanobacterium contained in the EMBL data- base were utilized: agp, atpA, atpB, atpC, atpD, atpE, atpF, atpG, atpH, atpL cpn60, desA, dnaK, glcP, ictA, ndhA, ndhB, ndhC, ndhE, ndhG, ndhL ndhH, petA, petC, petD, petE, prk, psaA, psaB, psaC, psaD, psaE, psaF, psbA1, psbA2, psbA3, psbB, psbC, psbD, psbE, psbF, psbG l, psbG2, psbH, psbK, psbL, psbl, rpllO and rp112.
Results and discussion
Isolation of the gdhA gene from Synechocystis PCC 6803
The gdhA gene of Synechocystis 6803, encoding its NADP-GDH, was isolated from two gene librar- ies of this cyanobacterium by complementation of the E. coli CLR207recA strain, auxotrophic for glutamate [37]. Transformants were selected on medium containing ampicillin but lacking glutamate. Two different plasmids, pSChl from library A and pBE1 from library B, were isolated, based on their ability to confer prototrophy for glutamate to E. coli CLR207recA. The inserts of the plasmids were subjected to restriction analy- sis and showed to share a common region of at least 2 kb (Fig. 1A). One of the two plasmids iso- lated, pSChl, probably resulted from a triple ligation event. A Hind III-Eco RI fragment (3 kb long) of the pSChl plasmid, containing most of the common region, was subcloned into the pBluescript II SK(+) plasmid. As expected, the resulting pSCh4 plasmid retained the ability to complement the glutamate auxotrophy of the E. coli mutant. Indeed, similar levels of NADP- GDH activities were found in cell-free extracts from E. coli CLR207recA clones containing either pSChl, pBE1 or pSCh4 (see Table 1). To con- firm that these plasmids harbour the Synechocys- tis gdhA gene, we compared the electrophoretic mobility of the corresponding protein extracts to that of the Synechocystis purified enzyme. Fig- ure 2 shows that they were identical and, in ad-
Table 1. Specific NADP-GDH activities of cell-free extracts from E. coli CLR207recA carrying the isolated recombinant plasmids and from E. coli K12.
Strain NADP-GDH activity (U/mg protein)
CLR207recA (pBluescript II SK+) 0.00 CLR207recA (pBE1) 0.34 CLR207recA (pSChl) 0.67 CLR207recA (pSCh4) 0.54 K12 0.24
Cells were grown in LB medium.
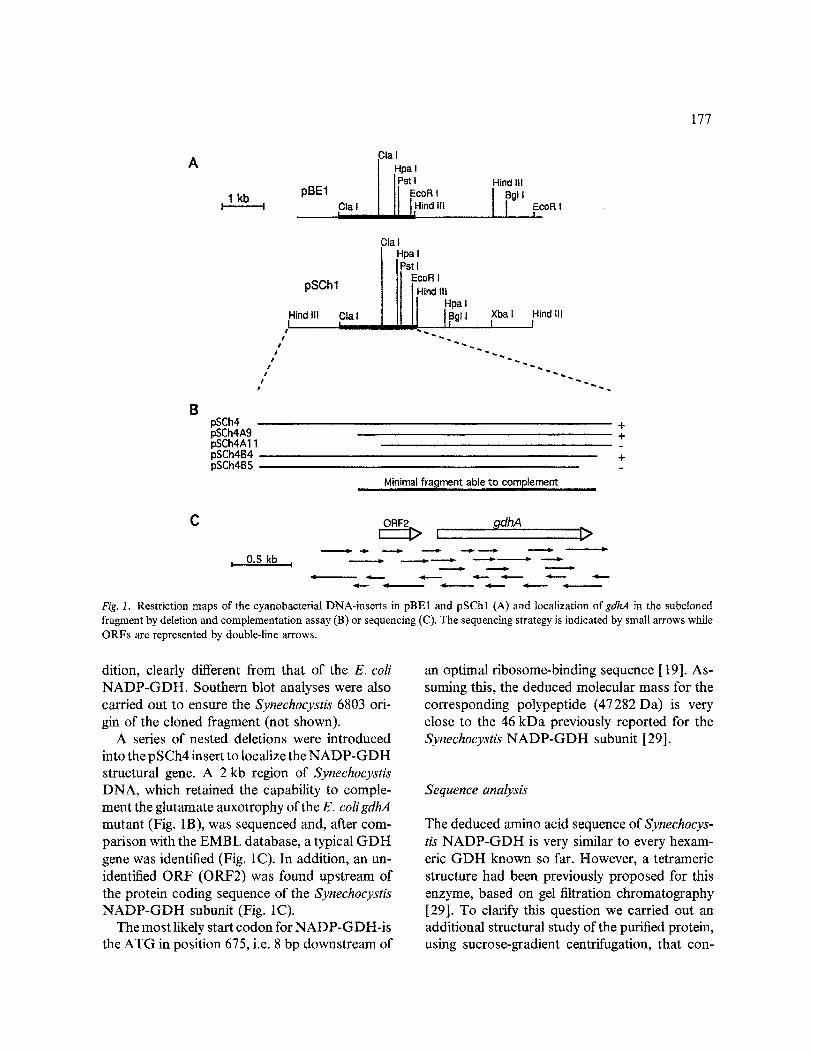
177
A
B
1 kb , pBE1
Cla I I
Cla I Hpa I I Pst I Hind III tI EcoRI [ ~gll II I, Hind Ill EcoRI
I
pSChl
Hind II1 Cla I I I,,,
I I
i I
I
l B
Cla I Hpa I I Pst I 11 Eco., II Hind Ill
II 1 Cl BpalgIl IXbaI lHindllI
pSCh4 + pSCh4A9 + pSCh4A11 pSCh4B4 + pSCh4B5
Minimal fragment able to complement
C ORF2[~ gdhA
I O . 5 k b , n " P P ~ ' - ~ . , ~ " . ,
~ 4 m ' l l 91 4==.-= qll
Fig. I. Restriction maps of the cyanobacterial DNA-inserts in pBE1 and pSChl (A) and localization ofgdhA in the subcloned fragment by deletion and complementation assay (B) or sequencing (C). The sequencing strategy is indicated by small arrows while ORFs are represented by double-line arrows.
dition, clearly different from that of the E. coli NADP-GDH. Southern blot analyses were also carded out to ensure the Synechocystis 6803 ori- gin of the cloned fragment (not shown).
A series of nested deletions were introduced into the pSCh4 insert to localize the NADP-GDH structural gene. A 2 kb region of Synechocystis DNA, which retained the capability to comple- ment the glutamate auxotrophy of the E. coligdhA mutant (Fig. 1B), was sequenced and, after com- parison with the EMBL database, a typical GDH gene was identified (Fig. 1C). In addition, an un- identified ORF (ORF2) was found upstream of the protein coding sequence of the Synechocystis NADP-GDH subunit (Fig. 1C).
The most likely start codon for NADP-GDH-is the ATG in position 675, i.e. 8 bp downstream of
an optimal ribosome-binding sequence [19]. As- suming this, the deduced molecular mass for the corresponding polypeptide (47282 Da) is very close to the 46 kDa previously reported for the Synechocystis NADP-GDH subunit [29].
Sequence analysis
The deduced amino acid sequence of Synechocys- tis NADP-GDH is very similar to every hexam- eric GDH known so far. However, a tetrameric structure had been previously proposed for this enzyme, based on gel filtration chromatography [29]. To clarify this question we carried out an additional structural study of the purified protein, using sucrose-gradient centrifugation, that con-

178
Fig. 2. Electrophoretic mobility of N A D P - G D H in clones of E. coli CLR207recA transformed with pBluescript II SK(+) (A), pBE1 (B), pSChl (C) or pBG1 (F), in E. coli DH5~ transformed with pSCh4 (D), and in a purified protein prepa- ration from Synechocystis 6803 (E). The PAGE was carried out under non-denaturing conditions and the gel was devel- oped by activity staining. E. coli DH5~ is a gdhA + strain and therefore a second band corresponding to E. coli N A D P - G D H is present in lane D. The slowest band in lane E is due to the formation of high molecular weight aggregates in the purified preparation [15]. pBG1 is a plasmid containing the E. coli gdhA gene.
firmed an hexameric configuration for Synechocys- tis N A D P - G D H (not shown).
Although Synechocystis N A D P - G D H can be aligned with every hexameric GDH, Table 2 shows that its level of similarity is higher to a subset of such sequences that includes those of the archaebacteria Halobacterium salinarium [5], Sulfolobus solfataricus [43] and the Archaeon ES4 [25], some Gram-positive bacteria such as Peptostreptococcus asaccharolyticus [67] and Clos- tridium difficile [41], and those of animals, repre- sented in Table 2 by the human sequence [1], since rat [20] and mouse [73] are almost identi- cal. In contrast, the Synechocystis sequence is much less similar to G D H sequences from other bacteria, both Gram-positive like Clostridum sym- biosum [72] and Corynebacterium glutamicum [8] and Gram-negative like Escherichia coli [49] and Salmonella typhimurium [ 31 ]; from fungi like Sac- charomyces cerevisiae [56], Schwanniomyces occi- dentalis [24], Neurospora crassa [38] and Aspergil-
Table 2. Amino acid sequences similarities among glutamate dehydrogenases from different sources)
Amino acid identity (%)
2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17
1 37 36 35 37 34 37 25 24 24 24 24 23 23 23 22 24 2 47 42 48 40 44 29 28 27 25 26 27 25 27 28 25 3 40 43 39 40 30 26 26 27 26 27 27 25 25 27 4 42 43 43 28 30 30 29 27 27 29 28 27 29 5 46 48 30 28 28 28 29 29 28 28 28 28 6 64 31 28 28 28 27 28 28 26 25 27 7 32 30 29 30 28 31 30 29 27 29 8 54 53 47 51 52 52 50 46 49 9 93 55 56 54 54 55 49 52
I0 55 57 55 54 53 48 51 11 52 50 49 48 49 49 12 59 52 51 53 53 13 52 50 48 48 14 71 64 65 15 64 67 16 71
G D H protein-coding sequences were aligned with the CLUSTAL V program [36], then the amino acid identity was calculated and expressed as a percentage of the shorter sequence. Sequences are the same as in Fig. 3. 1 1, human; 2, Synechocystis 6803; 3, Halobacterium salinarium; 4, Sulfolobus solfatanicus; 5, Archaeon ES4; 6, Peptostreptococcus asaccharolyticus; 7, Clostridium difficile; 8, Clostridium symbiosum; 9, Salmonella typhimurium; 10, Escherichia coli; 11, Corynebac- terium glutamicum; 12, Giardia lamblia; 13, Chlorella sorokiniana; 14, Neurospora crassa; 15, Aspergillus nidulans; 16, Schwanniomyces occidentalis; 17, Saccharomyces cerevisae.

lus nidulans [34]; from the chlorophyta Chlorella sorokiniana [16] and from Giardia lamblia, a primitive protozoan parasite [77]. In fact, two groups of GDHs can be defined according to the identity values shown in Table 2, in which the values resulting from pairwise comparisons inside each group are higher than those obtained by comparing any of its sequences to an outsider. In the group containing Synechocystis G D H all se- quences share at least 34~o identity among them. In the other group the minimal identity is 46~o, suggesting a higher homogeneity and, therefore, a closer evolutionary relationship of the corre- sponding GDHs. The existence of these two groups within the hexameric glutamate dehydro- genases reflects a non-common evolutive pattern, as recently pointed out [6]. It is worth noting that the only two GDHs from photosynthetic organ- isms ( Synechocystis and Chlorella) are not located in the same group and show a low degree of iden- tity (27~o), precluding a close kinship between these two genes. Both Synechocystis 6803 and Chlorella sorokiniana have another GDH, but these are NAD-dependent and, in the case of Chlorella, tetrameric and mitochondrial [51 ]. As the evolutive origin of chloroplasts seems to be associated to the endosymbiosis of a cyanobac- terial-like organism [26], a close relationship was expected to occur between the two genes com- pared in this work, both coding for enzymes that are hexameric, highly NADP-specific and, in the case of Chlorella, chloroplastic. On the contrary, Chlorella sorokiniana N A D P - G D H shows the highest identity (59~o) to that of Giardia lamblia, an organism lacking mitochondria and chloro- plasts that is considered to be a very primitive eukaryote [68 ]. Therefore the evolutionary origin of Chlorella sorokiniana N A D P - G D H is presum- ably not related to cyanobacteria and endosym- biosis. Unlike other nuclear genes encoding chlo- roplastic proteins that were transferred from the organelle genome to the nucleus [39], that encod- ing Chlorella sorokiniana N A D P - G D H was very probably always nuclear and the chloroplastic localization of its product should have been a late step in evolution.
Animal GDHs, although larger, are situated in
179
the same sequence group as Synechocystis G D H (Table 2). Animal GDHs are very well character- ized and several residues involved in the binding of metabolites that allosterically regulate their ac- tivity have been identified [17]. Most of these residues are not conserved in the cyanobacterial enzyme. Those are the cases of the histidine in- volved in ADP binding (see I in Fig. 3); the sev- eral residues acting in the allosteric binding of NADH, like the lysine marked as III in Fig. 3, and other residues implicated in the interaction with GTP (see VI in Fig. 3). This observation fits with the absence of allosteric regulation of Syn- echocystis N A D P - G D H by ADP, GTP or NADH (data not shown). The two archaebacterial GDHs (Sulfolobus solfataricus and Halobacterium sali- narium) included in that sequence group also lack allosteric regulations [7, 18]. We conclude that the higher similarity of this group of GDHs, in- cluding that of Synechocystis 6803 together with those of archaebacteria and some Gram-positive bacteria, to animal GDHs is not reflecting that the former group share also the regulatory prop- erties of the latter. It rather suggests that allosteric regulations have been acquired by animal GDHs through evolution.
The similarity of all the hexameric GDHs, in- cluding that of Synechocystis, is specially strong in the region close to the N-terminus that carries the three lysine residues (IIa, IIb and IIc in Fig. 3) which are involved in the binding of glutamate during catalysis [2]. By contrast, there is more variability in the coenzyme-binding domain, that is located in the C-terminus part of the protein [2]. While some of the residues involved in this binding are conserved, such as the regions around Asn-324 and Gly-351 (see IVb and IVc in Fig. 3), this is not the case of the region around posi- tion 257 that, in spite of being critical for the bind- ing of N A D P H to the enzymes of Neurospora crassa [ 11 ] and Salmonella typhimurium [3, 31 ], is different in the sequence of Synechocystis (see IVa in Fig. 3).
The residues around Gly-218 have been pro- posed to determine the coenzyme specificity of dehydrogenases: a G X G X X G motif would in- volve NAD dependence and a G X G X X A one

180 NADP Syn NAD Cdl NAD Pep
NADP ES4 NADP Hal
NAD/P Sul NAD/P H~ NADp Eco NADP Sal ~D Csy
NADP Cor NADp G~a NADP Chl NADP Neu NADP Asp NADP Sch NADP Sac
NADP Sy'n l~D Cd l NAD Pep
NADP ES4 NADP Hal
NAD/P Sul NAD/P Hum NADP Eco NADP Sal NAD Csy
NADP Cor NADP Gia NADP Chl NADP Neu NADp Asp NADP Sch NADP Sac
NA~P Syn ~AD Cdl NAD Pep
NADP ES4 NADP Hal
NAD/P Sul NADIP Hum NADP Eco NADP Sal ~qD Csy
NADP Cor NADP Gia NAD~ Chl I~D~ Neu l~DP Asp I~DP Sch NADP Sac
NADP Syn NAD Cdi NAD Pep
NADP ES4 NADP Hal
NAD/p Sul NADIP Smm NADP Eco NADP Sal NAD Csy
NADP Cor NADP Gia NADP Chl NADP Neu NADP Asp NADp Sch NADP Sac
NADP Syn NAD Cdl NAD Pep
NADP E$4 NADp Hal
NAD/p Sul NAD/P Hum NADP Eco NADP Sal NAD Csy
NADP Cor NADP Gia NADp Chl IqADP NeU NADP Asp NADp Sch NADP Sac
NADP Syn NAD Cdi I~D Pep
NADP E$4 NADP Fusl
NAD/P Sul I~D/P Hum NADP F.,CO NADP Sal ~qD Csy
NADP Cor NADP Gla NAOP Chl NADp Neu NADP Asp
• NADP Sch NADP Sac
~GSL FADASKRLEKALKk"/AI SDD ..... AGERLKYPKTSLSVSIPVRM 45 MSG KDVNV. EM. QSQVEN • CDKLG~PA----VY. L.. E .MRVIE ..... K. 48 MTDTLNPLVA. QEKVRI. C EKLGCDPA----VY • L. • E • QRVIEI .... K. 47 MVEQDP . EI .V.Q..R.AQ.~• . EE---- .L• F.. R. O/rIVE. T...E. 46
MTMASKSDST~ESGDEAADSTEPE.ALET.RRQ.YH.AS•LD.DQN-----IV ....... KVHE.T..IER 67 ~EVLSS..YTQOV.K.Y.VG~DNE . . . . TL.T.SQ.ERIIQ.K.QI.G 49
SEAVADREDDPNFF~VEGFFDRGASI~ELVEDLRTR.$EEOKRNRVRG-- .... ILRII.PCNHV..L.F.I.R 71 ~)QTYSLESFLNHVC~RDPNQTE..QoVREVMTT.--WPFLEOh~KYR(~ISLL...VE.ERVIQFRVV~FqD 69 MDQTCSLESFLNHV~PHQTE..Q.VREVMTT.--WPFLEQNPR~...VE.ERVIO~RVVWLD 69
MSKYVDRVIAEVE~4YADEPE•V~rfVEEVLSS.--GPVV.AHPEYEEVAIL• .MVI.ERVIEFRV.WED 67 MTVDEQV~AGEPE.HQ.VAEVLES.--ELVLEEDPHYADYGLIQ..CE.ERPVIFRV.WVD 69 ~AQTIEELIAVIKQRDG~E.RQ.VEEVVDS•--KV.FEREPKY--IPIF..MLE.ERVIIFRV•WMD 66
~ATTGDC~ALQKAVK~MATEAGTEGLVHGI ~qP ELRQLLTEI FMEDP E~E. MQ. VREVAVS. --QPVFEKRP EL--LP IFKQIVE. ERVIT F ~ 96 MSNLPSEPE. EQ° Y. E .AYT. ENSSLFQKHPEYRT--. LTVASI. ERVIQFRVVWED 55 MSNLPVEPE. EO .y • E .AST. ENSTLFEQ~EYRR--. LQVVSV. ERVIQFRVVWEN 55
MI~NGLPHEPE.QQ.YNE.VS..EESTLFTEEPEYKK--VIPVVSI.ERIIQFRVANEN 57 ° SEP E. QQ. YEEVVSS. EDSTLFE~EYRK--VLP IVSV. ERI IQFRVTWEN 51
DDGSLKI FPGYRVRYDDTRGPGKGGVRYHPN~II~DEVC~LAFh~4TFKCALLNLP FGGAKGGITLNP KELS RAELERLSH~'YI EAIAD--FIGPDIDILAP 143 .... I.T.K.F°SO~.AV..T... I.F.Q..SR...EA.SI ...... SVTGI .Y..G .... IVD. ST..QG .......... DG•YK--L.. EKV.VP . . 146 ... TV.V.K.W. SAHSSAV..S ..... F .... N .... EA.SL ..... GGA.G. •Y..G .... CVD .A... ER...Q....WVRG~YK--YL.DR... P. • 145 . ~ ~TV.V.T.F..O.NWA...T. °. I.W• .EE.LST.~q..A. • .W.T.~..Y..G .... IVD..K. oDR.K...A....R..Y.--V. S•YE..P.. 144 . . .TVEV.T• • •AQH.SV. • .Y. ° °L....D. •R..CVG.GM. • .W.. °~ ......... VAV ...... PE.K...T.RFT~E.R.--V...NQ. • P• . 165 S •. K..T .M°W°SO~qSAL. •Y ........... Q...EA. SMI° .W.NS. •L..Y..G...VRVD° .K.T.E...0...K..Q..YK--YL. SEL. °P.. 147 .... WEVIE...AQHSOH.T•C•.. I.. STD.SV...~%..SL..Y...WDV ...... A.VKI...NYTDN... KIT. RFTMEL .KKG .... G..VP.. 171 • RNOI~NRA~..QFSSAI..Y.. •M.F..S.NLSILEF.G.EQ...N..TT..M° •G...SDFD..GK.EG.~M.FC~qLMTELYR--F~.A.T.VP.G 167 • ENQV~..OFNSAI..Y...M.F..S.NLSILEF.G•EQ...N. •TT..M..G...SDFD..GK.EG.VM•FCQALMTELYR--~...T.VP.G 167. .N. k~/NT. °..QFNGAI..Y... L .FA. S .NLSIMEF.G. EQA..DS •TT..M ...... SDFD. NGK. DR .VM. FCQAFMTELYR--H ...... VP.G 165 .Q.QVHVNR.F..QFNSAL..Y.. •L.F•.S.NLGI•EF.G.GEI..NS.TG..I..G...SDFD..GK.DL•IM.FCESFMTELHR--H..EYR.VP.G 167 .A°RINVNR.F..Q.NSAL..Y...L•F. •S.NLSILEF.G.EQIL.NS.TT..M..G...SDFD..GK.DN.VM•FCQSFMTELQR--HV.A•T.VP•G 164 .A.N•QVNR.F•.Q.SSAI..Y...L•F. •S.NLSIMEF° °°EQI• •NS°TT..M..G. ° .CDFD..GK.D..VM•FCQSFMTEL~R--H•SYVQ.~'P.G 194 .N.NVQVNR.... ~FNSAL..y...L.L.. S .NLSILEF. G. EQI..N..TG. SM. • G°..ADFD..GK.D° .IR. F~HK--H..A. T .VP. G 153 .K.EV~•NR• • . •QFNSAL..y...L.F..S.NLSILEF.G.E~I..N..TG.~..G...SDFD..GK.DS.IR.FCTAFMTELCK--H..A.T.LP.G 153 .N.DVEVNN.F• •QFNSSL•.y...L.F..S.NLSILEF.G.EQI'..N..TG.SM..G...CDF...GR.DG.IR.~.R--Y..A.T.VP.G 155 .K.EQEVAQ.. ° .Q.NSAK..Y...L°F..S.NLSILKF°G.EOI..NS.TG.DM..G...LCVDL.GR.NN.IR.ICYAFMRELSR--H° °Q.T.VP.G 149
DVYTNEMM~S I I RRKI S p-A--VVTGKPVTMGGSQGRNTATGTGAFY IM~KFD . . . . QYP EN~I~rAVQGFGNAGMVVAE CLYQD- 231 ..N••GQI.$..V.E.NKLTGQ-.--SIG.I ..... EF...L. •TA...F.VAVT~EAAA.LGI-----D-MKKAE~ .... I..V.SYTVLN-CEEL 234 • .N..GQI .S.FV.E.V~LNGE-RM-DIGTF ..... AF. • .E...E. • .F.VAVVVRESA~.GI .... K-M. DAKI ....... V.TFTV~N-IERQ 234 H° . • .PQI.A .... E•EA.S.RKT.-.FGII .... LSI . . .L . . . E . .AR.. S .TIREARKVLGW---- .... ~LKGK.I. I . .Y .... YYL. N/MSE .Y 236 .MG.DPQT .A.L . .A..MQEGETT. ----G ...... PVV... E . . EE . P . RSVAI .T °LVCEYY . ----- .PLDE ....... X. SV.ANA. R- .LDKW 253 • .N.DSQT.A.FL.E.IK.TG.V---DFA.F ..... EL.. IGV.LYS..L•VAT.AKEEAANKFI-----G~V.EAR. II ..... V.YYAGKF.SE-M 236 .MS .G. RE. S. IA.T.ASTIGHYDIN. HAC ..... ISQ.. IH.. I S. o. R. V. HGIENFINEASYMSILG~TPGFGDK. FV ...... V. LHS~%Y. -HRF 270 . IGVGGREV.F.AGMqKELSNNT-A---C. F...G~SF...LI.PE...Y. LV. FrEA..K . . . . RHGMGF. G~A~. S. S. S.. VA~AI. K~/~-F 254 .IGVGGREV.F.AG~t~LS~S-S---C.F, ..GLSF...LI.PE...Y.LV.FTEA..K . . . . RHGLGF.G~R...S.S..VAQYAIoKA~q-F 254 .LGVGAREI.Y.YG..RK.VGGFYN--G.L...AR$Fo ° .LV.PE...Y.SV.YVEAV~( ...... HENDTLVGK...LA....VA~GA.KK.AE-L 253 .IGVGGREI.~'LFGH.RRMANQHES---G.L...GL.W...LV.TE...Y.CV°FVSE.IK------AKGESISGQKII.$°S. .VATYAI.EAQE-L 255 .IGVGAREI.YLYG..ERL.NEF-T--G.L...N°~...FI.PE...Y..V.FLEE.CK .... ---DNNTVIRGEN.LLS•S..VAO~AC.K.I.-L 251 . IGVGAREI. YLFG.. ER. T~Y-T---G. L...G~EY...EI.PE...Y..VLFVENV. K ..... DKGESLKGKRCL. S .A..VAQYC..L.LE-K 281 . IGVGGREI .Y . FGA. RKAANRFE----G .L... GLSW. ° .LI .PE . . . Y . LV. YVGH.. EY ..... SGAGSYAGER.. LS . S . . VAOYA. LK° IE-L 241 . IGVIGREV. FLFG.. RR.. N~WE----G.L... GGSW... LI .PE . . . Y.W°YV. H . IKHVT .... GGKESFAGKR.. I S . S . . VAOYA. LKVIE-L 243 . IGVGGREI .YLFGA. K~----G.L...GL .W... LI .PE...YVS. TTLKK. IE.AT .... NGKESFKGKR. ELS. S..VAQYA. LKVTE-L 245 . IGVGGREI .YLFGA. RSYENSWE----G.L...GLNW...LI .PE... Y.LV.YT .A. IDYAT-----NGKESF.GKR. TIE • S • .VAQYA.LKVIE-L 239
IW
GYKVVAI---SD SQGG IYNEQGI -----DI PAVIDYKQPd4RTLA~YCDQAICDLGENQQI SNAE .... LLALDVDVLIHAALENQITRDNADQV- 314 .GT...~EWCK.EGSYA° ...N.L-----.GQ.ML.•~E~E.-- .... GNLLNFpGAKR..LE.- .... FW.S...IV ....... S..KEV.E$I- 313 . G..C..AE~DRNEGNYAL.. °N.. -----. FKELLA.. EAN . . . . KTLIGFPGAER. TDE. -----FWTKEY. I IV ...... V..GER.KTI- 313 .M....V .... ...K ..... PD . L------NADE .L~ . . E . -- .... GSVK. FPGATN .T . E.- .... °.E.E....A...I.EV..KK...NI- 310 .ATI... ----..VN.AM. EPD.. ------ .TAS. PSHDEEP ..... E.VrTYADT~-... E.----..T ........... G.V..KE..EAI- 326 .A.I.GV .... . o oK..VI..K..-----.VGKA.EI.EK ....... TGSV~NYp ° GRKVT . E. ---- . . IS. C. I ........ V.NKF° .PK.- 310 .A.CI .V----GE .D . S . W.PD . • ---- .PKELE.F .LQ. -- .... GS. LGFPKAKPY-EGS .... I.EA.C.I ..... S.K.L.KS..PR.- 343 .AR. ITA---- . . . S . TVVD. S . --F-TKEELARL . EI . ASRDG ..... RV. DYAKEF-GLVTLEGQQPW---S . P . . IAL . C . TQ. ELDV-D .AHQL 335 . AR.. TA----... S. TVVD. S. --F-TP~EI .ASRDG------RV o DYARDF-GLTYLEG~---SVP.. IAL. C. TQ. ELDV-D .AR. L 335 . A. A. TL----. GPD. y.. DPE. --ITTEEK. NYML~RASG .N----KVQDYADEF-GVQFFPGEKPW--GQK. ° I IM. C. TQ. DVDL-E~ 335 .AT.IGF .... • ..S.WVHTPN.--V----.VAELREI.EVR.A .... RVSVYADEVEGATYHTDGSIW--D.KC.IAL.C.TQ.ELNG-ENAKTL 334 . A.. LTF----- ° , . N. T •VDED . --F-NEEKLAHLMYL. NEK. G------RVSEFEDKYPSVAYYEGKKPWECFEGQM. CIM. C • TQ. EVSG-DDATRL 336 .AI .LSL--- . .... YV. EPN. --FTREQLQAVQD~. KNNSA ..... RISEYKSD--TAVYV~RRKPWE o DCQ.. IAF. C. TQ. E. DE-F/)AELL 365 .AT..SL .... ... K.ALVATGESG-ITVE.. N..MAI. EARQS. ----TSFQH----AG~IEGARPWIRV-GK.. IAL. C. TQ. EVSK-EEAEGL 325 . GS.. SL---... K. SLIVEDESASFTpEE. ALIA. L. VAR~. SELATSS. F----AG~(FTYIPDARPWTNIPGKFE. AL. S. TQ. EVSG-EEAE~ 333 . GI . . SL---- ° . ° K. S .VSSN. ---IVP EQVLEIAAA. LKFKS . EEITKESVELFS . . . SVEYL . GVRPWAKV-GHF. °AL. S . TQKEVSGEEEA~AL 536 . GT.. SL---... K. C. IS. T. ---ITSEGVADIHSA .VNFKS. EQI~NEYSTF--S.. KVQYI . GARP~THV-QK.. IAL. C. TQ. EVSG-EE%EAL 327
ZW ~W
--- RARYI FEVANGPTTTAADD ILAS K . . . . GIYVFPDILVNAGGVTVSYFEWVQNRSGLYWS AKEVNDRLKEF~qE---- 384 ---K. KLVC.A ...... PE..EVF.ER- ..... ..VLT .... T ............... LY.Y... EE..EOKEEIA..K .... 383 ---N. KLVC.A ...... PEG. KV. TER ....... ..NLT .... T.S..oL...Y ..... QY.Y..T .......... EA.. EEKO~AD .~--- 383 ---K. KIVA ...... V.PE..E..FE.- ...... ..LQI..F.C ............... IT.Y..T 'LE.. REK.DK..TK---- 380 ---A.DLW° G ...... ST• o$.. oDR- . . . . DVA.I .... A ............ L.DINRRA.. LER. •. E. EAEoQA--- 396 ---K.EL.V.G....LoAD..E.~QR- ...... ..A.V....A ..... VG..V..AN.EM.EII. DEoAKELIVDR.NN---- 380 ---K.KI oAoG ...... PE• .K• FLER ....... N•M. I..LYL ............ LK. LNHVSYGRLTFKYERDSNYHLI/4S • QES • ER. FG~HGGT 431 IANGVKAVA. G.. M... IE. TELF-QQA ...... .VLFA. GKAA ..... AT .G~ .~%..A" -ARLGWK. EK. DA.. HHI. LD---- 408 IANGVKAVA. G..Mo.. IE. T. LF-LEA ...... . VLFA. G~A ..... AT.G~.~..A" -ARLSWK. EK. DA.. HHI. LI>---- 408 VANNVK. yI .... M...NE.LRF.M~ ..... NMV.A. SKA ...... L..G..MS..S- ERLSWT. E.. DSK. HQV. TD---- 409 ADNGC. FVA. G.. M. S. PE .VEVF-RER ..... D. RFG. G~A. V...AT. AL .MQ..A. H~FLE-----LRVHRR. PPGDSEEH ........ 408 VGLG~KFVA.G..M.S.AE.VHVY-HAK . . . . .~MYG.AEAS ..... S..G~.MS..S .......... VRLQWT.E..DOK.RGI.RG---- 409 IKHGCQ°W. G..M.S .NE. IHKY-N .A ...... .. IYC.GKAA ..... A..G~.MT...- ......... MSLNWTRE.. R. K. ERI. KD---- 438 LA~GCKFVA. GS. MGC .LE • IEVFENNRK--EKKGEAV~YA. GKAA. C... A.. G~. MA. ° S QRLNWTQA.. DEK.. DI. KN---- 405 IKSGV... A. GS. MGC. Q.. I •. FEAHRN--ANPGDA. ~YA. GKAA ..... A..c~ .MA..S" -ARLSWTSE.. DA... GI. ED---- 413 VEAGCK.. Ao GS . MGS . KE. I . VFEANR .... SNN%qqYA. GKAA. C...A..GL .MA.. $- QRVQWS. E.. DAK.. NI . YT-- 412 VAQGVKF .A. GS.~oS. pE. IAVFETARSTATGPSEA%q~YG. PKAA. L...A..GL.MA..S" QRITWTSER.DQ~.. RI. I:; 409
..... EAEHVWNITQELDV .................... NVRTAAY I F~qLNRLSEA~DAKGTRDYYQDS 428
..... AF.SI. K.KE.YN.- ................. TM.E...M. SIKKVA. o °KLRo WY 421
.... AIKG. FAVAD .YN.- ................ TL. E .V.MY. IKSIDV. oKLR.~Y 421 - AFYD .y .TAK.KNI. ~4. D.D.W.VQ.VYO..LDR. HVKH 420 ..... AhIRA. KDEYENR..- ................ TW.D....V..S.IA..HE. R.LWP 435 .... AFNTLYDYH . KKKLE- DHDL....MAL .VD .VVR..KoR.IL 421 I p IVP TA. FQDRI SGAS . K. IVHSGIAYT~RSARQIMRTAMEI'NLGLDL ..... VN . IEKVFKVYNEA.VTFT 505 ..... IHfiACVEF~G. -GE(~TN .................... YV~G. N. AGFV~VAD..L.Q.VZ 447
-IHHACVEYGGD-NEHTN- YVQG. N. AGFVKVAD..L.Q.VZ 447 ----IHDGSAAAAERYG~GYN- LVAG. N. VGFOKIAD..M.O. IAW 450 --- --LQDLCRDRSRVWTRERL- TLVG .N .HEFKKVAD..L .Q.VI 448 -- --- I FVACRDTAKKYGHP EN- YQMG. N .AGFLKVAD$. I EQ. CV 449 -----IYDSAMGPSR. YN- .VD .................. LAAG .N . AGFTKVAD .VK. Q. AV 476 .... AFFNGL. TAKTYVEAAEG-ELP ....... " ...... SLVAGSN. AGFVKV~.. HDQ. D~SEN 454 .... CFENGLETA. KFATPAKG-~/LP ............. SLV. GSN. AGFTKVA...KDQ. D~ 459 ..... CFDNCYDPAI KYSAEENADGLP ............. SLLKG °N. ASFIKVAD. • FDQ. DVY 459 ..... CFNECIDYAKKYT-. KDGKVLP ............... SLVKG .N. ASFIKV. D.. FDQ. DVF 454

181
would determine NADP specificity [75]. The motif found in the Synechocystis sequence (GF- GNAG) (see V in Fig. 3) clearly contradicts this hypothesis, since the cyanobacterial enzyme is highly specific for NADPH [29]. Similar incon- sistencies have been reported for Clostridium sym- biosum and Halobacterium salinarium GDH [72], and can also be seen for Archaeon ES4 (see Fig. 6 in [25] and V in Fig. 3).
uct of ORF2 was hardly detectable in E. coliwhen analysed with an expression system based on T7 RNA polymerase [70 ], whereas the gdhA product was simultaneously accumulated (not shown). This result suggests that the ORF2 product might be scarce, probably because of the rather deficient Shine-Dalgarno sequence of ORF2 (AAG at 10 bp from ATG), as compared to that of gdhA (AGGAG at 8 bp from ATG).
The open reading frame upstream ofgdhA gdhA is transcriptionally regulated by growth phase
As indicated above, the minimal DNA fragment from Synechocystis capable of complementing the gdhA mutation of E. coli includes an open read- ing frame (ORF2) located upstream of the gdhA gene. The deduced amino acid sequence of its coded protein shows no significant similarity to any sequence contained in the databases and no standard protein motifs were detected. Its de- duced molecular mass was 13,8kDa, and its composition, with a high content of alanine (14.6 ~o ) and valine (11.5 ~o ), determines a marked hydrophobic profile (not shown) and a theoreti- cal isoelectric point of 4.1.
ORF2 is very likely a protein-coding gene. That is the conclusion reached after applying the CodonPreference statistic [30], based on the comparison of its codon usage to the average of Synechocystis (not shown). Furthermore, the oc- currence of rare codons is even lower than in gdhA, what indicates that ORF2, as well as gdhA, has an optimal codon composition from a trans- lational point of view [22]. A similar positive con- clusion was obtained by the application of the TestCode method [28]. Nevertheless, the prod-
Although the deletion of ORF2 strongly de- creased the level of expression of the cyanobac- terial gdhA gene in E. coli, this gene could still be expressed in E. coli when cloned downstream of strong promoters such as the tac or the PR of 2 phage (see plasmids pSChl l and pSCh20 in Fig. 4). These two plasmids, lacking ORF2, were still able to complement the E. coli gdhA mutant, demonstrating that ORF2 is not needed for that phenomenon. Therefore, the apparent require- ment of ORF2 in the minimal fragment of cyano- bacterial DNA capable to complement the E. coli mutant, could result from the localization of ORF2 between the gdhA promoter that functions in E. coli and the gdhA coding sequence.
Using two specific probes for ORF2 and gdhA, we found that both genes are indeed cotrans- cribed in Synechocystis 6803 (1.9 kb mRNA in Fig. 5A and B). In addition, several shorter and more abundant transcripts were hybridized only by the gdhA probe (< 1.5 kb in Fig. 5B). These data suggest that two different promoter regions are involved in the expression of gdhA: one in front of ORF2 that promotes the long, bicistronic,
Fig. 3. Comparison of the predicted amino acid sequence for Syneehoeystis 6803 NADP-GDH with the sequences of other GDHs stored in the EMBL Sequence Data Library, by using the PileUp program [23]. The coenzyme specificity of each enzyme is in- dicated as follows: NAD, NAD-specific; NADP, NADP-specific; NAD/P, ambiguous specificity. The position of the residues involved in the allosteric activation by ADP in animals (I), in the binding of glutamate (IIa, lib, Ilc), in the allosteric inhibition by NADH in animals (III), in the coenzyme binding site (IVa, IVb, IVc), in the coenzyme specificity motif (V) as well as in the GTP-binding site (VI) are indicated. Amino acid residues identical to those of Synechocystis 6803 are shown as dots, while gaps are indicated by dashes. Syn, Synechocystis 6803; Cdi, Clostridium difficile; Pep, Peptostreptococcus asaccharolyticus; ES4, archaeon ES4; Hal, Halobacterium salinarium; Sul, Sulfolobus solfataricus; Hum, Homo sapiens; Eco, Escherichia coli; Sal, Salmonella typhi- murium; Csy, Ciostridium symbiosum; Cor, Corynebacterium glutamicum; Gia, Giardia lamblia; Chl, Chlorella sorokiniana; Neu, Neurospora crassa; Asp, Aspergillus nidulans; Sch, Schwanniomyces occidentalis; Sac, Saccharomyces cerevisiae.
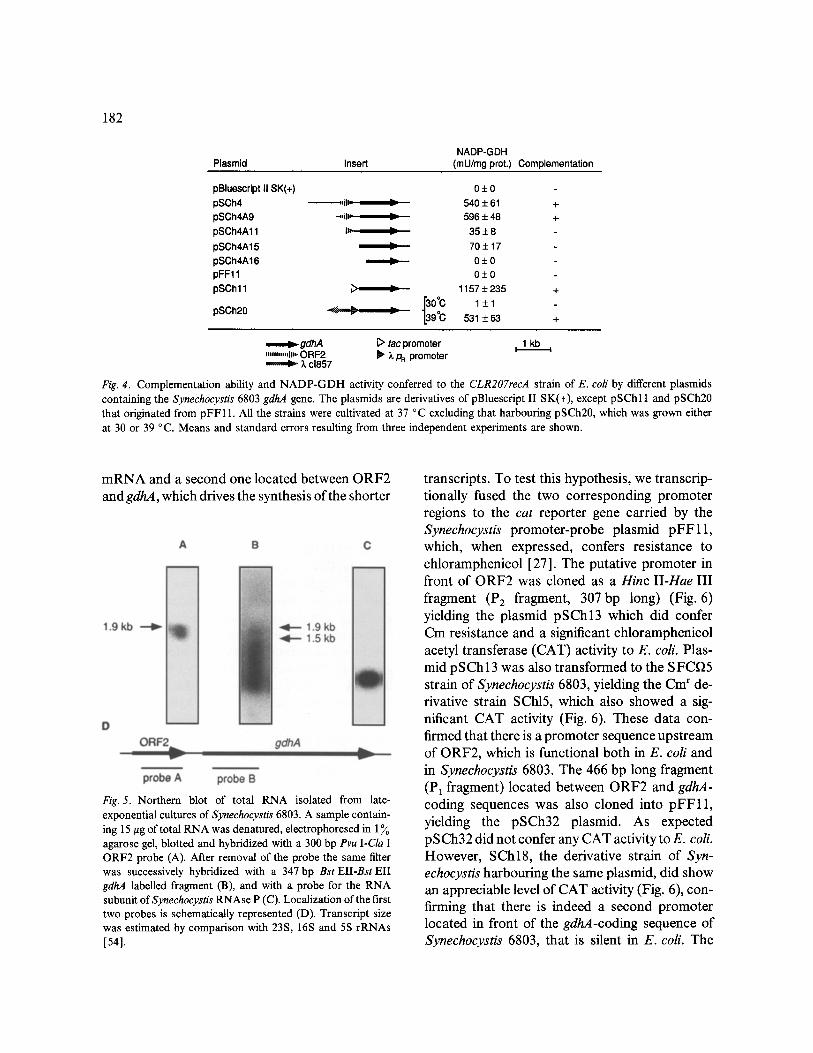
182
Plasmid Insert NADP-GDH
(mU/mg prot.) Complementation
pBluescript II SK(+) 0 + 0 pSCh4 ~ , , i l l = , I1 540 + 61 + pSCh4A9 ..,,~la, lit 596 + 48 +
pSCh4A11 ID, _= 35 + 8
pSCh4A15 I= 70 + 17
pSCh4A16 ~ 0 + 0 pFF11 0 + 0
pSChl I ~ _'~ 1157 + 235 +
L I 3 0 ° c 1 ± 1 pSCh20 ~ ~ 1.39°C 531 + 63 +
"'m"""lih' ORF2 ............ ,~- ;L cl857
C> tac promoter i 1 kb Z, Pa promoter
Fig. 4. Complementation ability and NADP-GDH activity conferred to the CLR207recA strain of E. coli by different plasmids containing the Synechocystis 6803 gdhA gene. The plasmids are derivatives of pBluescript II SK(+), except pSChl l and pSCh20 that originated from pFF11. All the strains were cultivated at 37 °C excluding that harbouring pSCh20, which was grown either at 30 or 39 °C. Means and standard errors resulting from three independent experiments are shown.
mRNA and a second one located between ORF2 and gdhA, which drives the synthesis of the shorter
Fig. 5. Northern blot of total RNA isolated from late- exponential cultures of Synechocystis 6803. A sample contain- ing 15/zg of total RNA was denatured, electrophoresed in 1 To agarose gel, blotted and hybridized with a 300 bp Pvu I-Cla I ORF2 probe (A). After removal of the probe the same filter was successively hybridized with a 347 bp Bst EII-Bst Eli gdhA labelled fragment (B), and with a probe for the RNA subunit ofSynechocystis RNAse P (C). Localization of the first two probes is schematically represented (D). Transcript size was estimated by comparison with 23S, 16S and 5S rRNAs [54].
transcripts. To test this hypothesis, we transcrip- tionally fused the two corresponding promoter regions to the cat reporter gene carried by the Synechocystis promoter-probe plasmid pFF11, which, when expressed, confers resistance to chloramphenicol [27]. The putative promoter in front of ORF2 was cloned as a Hinc II-Hae III fragment (P2 fragment, 307bp long) (Fig. 6) yielding the plasmid pSCh13 which did confer Cm resistance and a significant chloramphenicol acetyl transferase (CAT) activity to E. coil Plas- mid pSCh13 was also transformed to the SFCf~5 strain of Synechocystis 6803, yielding the Cm r de- rivative strain SChl5, which also showed a sig- nificant CAT activity (Fig. 6). These data con- firmed that there is a promoter sequence upstream of ORF2, which is functional both in E. coli and in Synechocystis 6803. The 466 bp long fragment (P1 fragment) located between ORF2 and gdhA- coding sequences was also cloned into pFF11, yielding the pSCh32 plasmid. As expected pSCh32 did not confer any CAT activity to E. coll. However, SCh18, the derivative strain of Syn- echocystis harbouring the same plasmid, did show an appreciable level of CAT activity (Fig. 6), con- firming that there is indeed a second promoter located in front of the gdhA-coding sequence of Synechocystis 6803, that is silent in E. coll. The

A ~ ir~ It SaU3A I I;1~ I I I H u |11 ,~,u3A I
"=°"ll ,~'"l "°°"11_"'l "1 [ / ORF2 gdhA
l Hint II [S Hae III au3A I
P2 -~ ~P P1
183
B E. coil Synechocystis 6803 CAT CAT
Plasmid Construct Cm r (mu/rngprot.) Strain Cm r (mu/mgprot.)
BamH I Hinc II pFF11 ~-- II ~ ._ 0 + 0 SFF16 0 + 0
BarnH IlHinc II
pSChl 3 ~- [- P~" ~ + 31 + 2 SChl 5 + 44 + 0
P1 . I Hint II pSCh32 !-- -. ~ 0 + 0 SCh18 + 21 + 1
t-- Transcription terminator
i ~>. catgene
• Insert
Fig. 6. Subcloning of two DNA fragments from the 5' region of the Synechocystis 6803 gdhA gene (A) in the pFFll promoter- probe plasmid. The ability of the resulting plasmids to confer chloramphenicol resistance and CAT activity was analysed in E. coli and Synechocystis 6803 (B). Means and standard errors calculated from two independent experiments are shown. Synechocystis cultures used for assaying CAT activity were grown up to a chlorophyll concentration of 25 #g/ml.
long exposure time required to detect the tran- scripts shown in Fig. 5 suggests that these mRNA are not very abundant, since sharp and fast ap- pearing bands were obtained when the same fil- ter was blotted with other probes, such as the RNA subunit of RNAseP (Fig. 5C). Therefore, rather than using northern experiments, we de- cided to use transcriptional fusions of the cat gene to study the regulation ofgdhA expression in Syn- echocystis 6803. Since the clearest regulation of NADP-GDH takes place in response to changes in the growth rate, we measured NADP-GDH and CAT activities in the strains SChl5 and S Ch 18 along the growth of the cultures. The pro- files of CAT and NADP-GDH are very similar in SChl5, with a clear increase in the late growth- phase of the culture (Fig. 7B), supporting that NADP-GDH activity is regulated at the level of transcription in Synechocystis 6803. However, CAT activity in SChl8 was almost invariable (Fig. 7C), suggesting that the regulation of gdhA expression along the growth phases is mainly ex-
erted through the promoter in front of ORF2, whereas the promoter upstream of gdhA is not regulated.
According to the levels of CAT activity shown in Fig. 7, the promoter region upstream of ORF2 is the main contributor to the expression ofgdhA at late growth phase. However, Fig. 5 shows that the bicistronic transcript is less abundant than the set of shorter ones only hybridizing gdhA. One possible cause of this discrepancy may lie in a reduced stability of the 5' region of the bicistronic transcript due to a higher sensitivity to nucleolytic degradation. A putative hairpin structure (CCT- A G G G C A A T G T C C G T C A T G G C C C C A G G ) located in the intercistronic region, partially overlapping ORF2, may be involved in this differential stability by increasing that of the mRNA region situated downstream of it. An analogous situation takes place in the rpsO-pnp operon of E. coli, where two coding regions with a secondary promoter and a steam-loop structure in between show different degrees of stability due
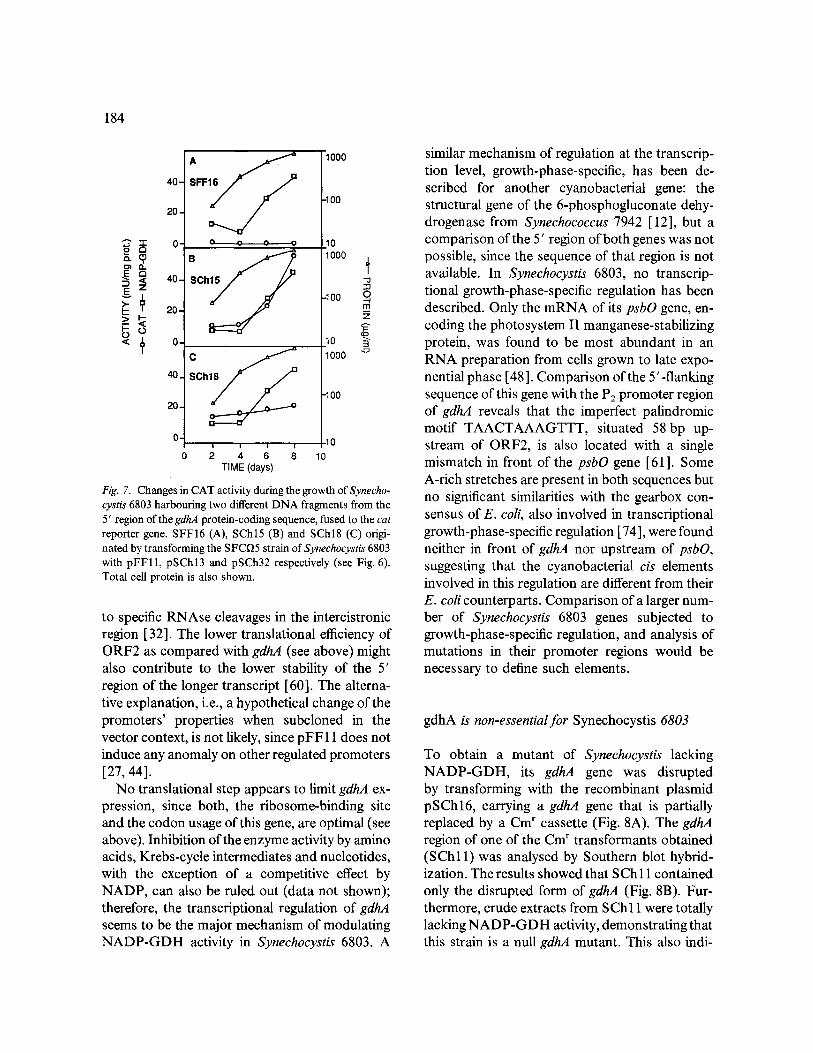
184
E
40-
20-
0"
40.
20-
0-
40.
20.
0-
n 0 0 0
:ch 10 i i i |
2 4 6 8 10 TIME (days)
1000
-I O0
10 1000 t
-100 m
T~
lO ~ 1000
"100
Fig. 7. Changes in CAT activity during the growth of Synecho- cystis 6803 harbouring two different DNA fragments from the 5' region of the gdhA protein-coding sequence, fused to the cat reporter gene. SFF16 (A), SChl5 (B) and SChl8 (C) origi- nated by transforming the SFCI)5 strain of Synechocystis 6803 with pFF11, pSChl3 and pSCh32 respectively (see Fig. 6). Total cell protein is also shown.
to specific RNAse cleavages in the intercistronic region [32]. The lower translational efficiency of ORF2 as compared with gdhA (see above) might also contribute to the lower stability of the 5' region of the longer transcript [60]. The alterna- tive explanation, i.e., a hypothetical change of the promoters' properties when subcloned in the vector context, is not likely, since pFF11 does not induce any anomaly on other regulated promoters [27, 44].
No translational step appears to limit gdhA ex- pression, since both, the ribosome-binding site and the codon usage of this gene, are optimal (see above). Inhibition of the enzyme activity by amino acids, Krebs-cycle intermediates and nucleotides, with the exception of a competitive effect by NADP, can also be ruled out (data not shown); therefore, the transcriptional regulation of gdhA seems to be the major mechanism of modulating NADP-GDH activity in Synechocystis 6803. A
similar mechanism of regulation at the transcrip- tion level, growth-phase-specific, has been de- scribed for another cyanobacterial gene: the structural gene of the 6-phosphogluconate dehy- drogenase from Synechococcus 7942 [12], but a comparison of the 5' region of both genes was not possible, since the sequence of that region is not available. In Synechocystis 6803, no transcrip- tional growth-phase-specific regulation has been described. Only the mRNA of its psbO gene, en- coding the photosystem II manganese-stabilizing protein, was found to be most abundant in an RNA preparation from cells grown to late expo- nential phase [48]. Comparison of the 5'-flanking sequence of this gene with the P2 promoter region of gdhA reveals that the imperfect palindromic motif TAACTAAAGTTT, situated 58bp up- stream of ORF2, is also located with a single mismatch in front of the psbO gene [61]. Some A-rich stretches are present in both sequences but no significant similarities with the gearbox con- sensus of E. coli, also involved in transcriptional growth-phase-specific regulation [74], were found neither in front of gdhA nor upstream of psbO, suggesting that the cyanobacterial cis elements involved in this regulation are different from their E. coli counterparts. Comparison of a larger num- ber of Synechocystis 6803 genes subjected to growth-phase-specific regulation, and analysis of mutations in their promoter regions would be necessary to define such elements.
gdhA is non-essential for Synechocystis 6803
To obtain a mutant of Synechocystis lacking NADP-GDH, its gdhA gene was disrupted by transforming with the recombinant plasmid pSChl6, carrying a gdhA gene that is partially replaced by a Cm r cassette (Fig. 8A). The gdhA region of one of the Cm r transformants obtained (SChl 1) was analysed by Southern blot hybrid- ization. The results showed that SCh 11 contained only the disrupted form of gdhA (Fig. 8B). Fur- thermore, crude extracts from SChl I were totally lacking NADP-GDH activity, demonstrating that this strain is a null gdhA mutant. This also indi-
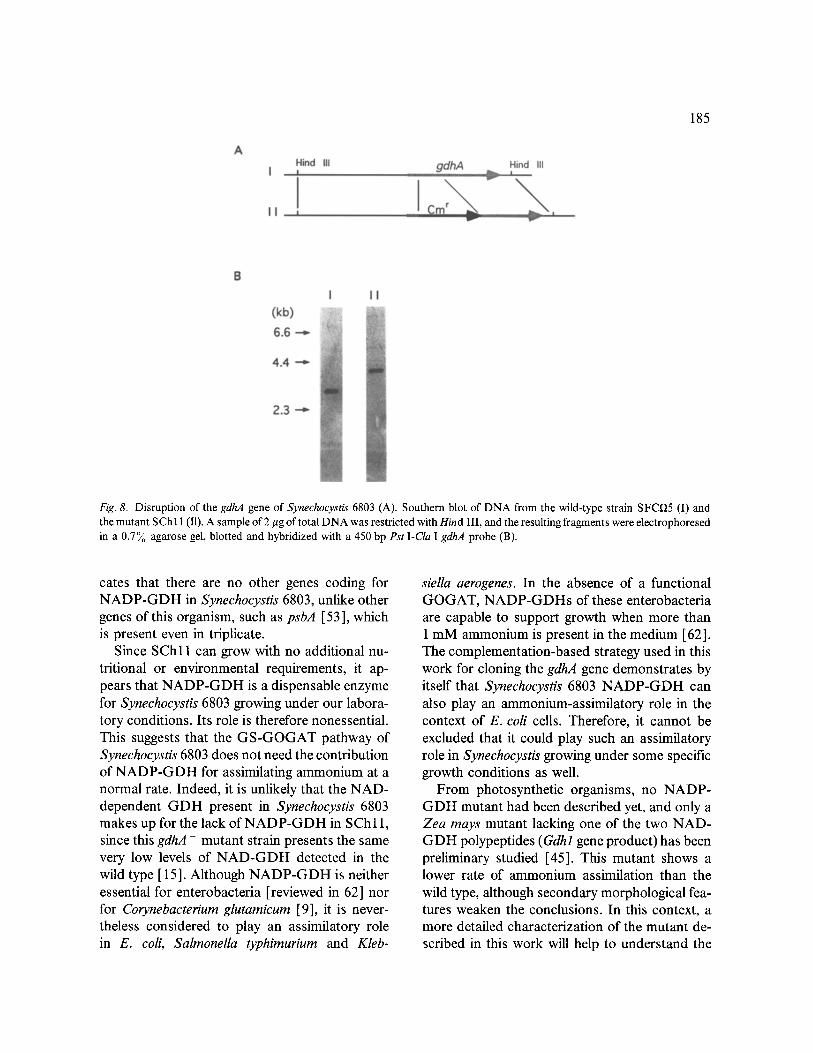
185
Fig. 8. Disruption of the gdhA gene of Synechocystis 6803 (A). Southern blot of DNA from the wild-type strain SFC~5 (I) and the mutant SCh 11 (I1). A sample of 2 #g of total DNA was restricted with Hind III, and the resulting fragments were electrophoresed in a 0.7% agarose gel, blotted and hybridized with a 450 bp Pst I-Cla I gdhA probe (B).
cates that there are no other genes coding for N A D P - G D H in Synechocystis 6803, unlike other genes of this organism, such as psbA [53 ], which is present even in triplicate.
Since SChl l can grow with no additional nu- tritional or environmental requirements, it ap- pears that N A D P - G D H is a dispensable enzyme for Synechocystis 6803 growing under our labora- tory conditions. Its role is therefore nonessential. This suggests that the G S - G O G A T pathway of Synechocystis 6803 does not need the contribution of N A D P - G D H for assimilating ammonium at a normal rate. Indeed, it is unlikely that the NAD- dependent G D H present in Synechocystis 6803 makes up for the lack of N A D P - G D H in SChl 1, since this gdhA - mutant strain presents the same very low levels of N A D - G D H detected in the wild type [15]. Although N A D P - G D H is neither essential for enterobacteria [reviewed in 62] nor for Corynebacterium glutamicum [9], it is never- theless considered to play an assimilatory role in E. coli, Salmonella typhimurium and Kleb-
siella aerogenes. In the absence of a functional GOGAT, N A D P - G D H s of these enterobacteria are capable to support growth when more than 1 mM ammonium is present in the medium [62]. The complementation-based strategy used in this work for cloning the gdhA gene demonstrates by itself that Synechocystis 6803 N A D P - G D H can also play an ammonium-assimilatory role in the context of E. coli cells. Therefore, it cannot be excluded that it could play such an assimilatory role in Synechocystis growing under some specific growth conditions as well.
From photosynthetic organisms, no NADP- G D H mutant had been described yet, and only a Zea mays mutant lacking one of the two NAD- G D H polypeptides (Gdhl gene product) has been preliminary studied [45]. This mutant shows a lower rate of ammonium assimilation than the wild type, although secondary morphological fea- tures weaken the conclusions. In this context, a more detailed characterization of the mutant de- scribed in this work will help to understand the

186
role of GDH in photosynthetic organisms. A careful analysis of the biochemical parameters re- lated to nitrogen metabolism under different physiological conditions is now being carried out using this mutant.
Acknowledgements
This work was funded by grants PB 88-0020 and PB 91-0127 from the Direcci6n General de In- vestigaci6n Cientifica y T6cnica (Spain) and by Junta de Andalucia. S. Ch. and J. C. R. were re- cipients of a predoctoral fellowship from Minis- terio de Educaci6n y Ciencia (Spain) and S. Ch. of a short-term fellowship from EMBO for stay- ing in C. E. Saclay. We thank Dr J. C. Wootton for providing E. coli CLR207recA and plasmid pBG1 and Dr A. Vioque for the RNase P probe. Critical reading by M. I. Muro-Pastor is grate- fully acknowledged.
References
1. Amuro N, Mayumi Y, Goto Y, Okazaki T: Molecular cloning and nucleotide sequence of the cDNA for human liver glutamate dehydrogenase precursor. Biochem Bio- phys Res Commun 152:1395-1400 (1988).
2. Baker PJ, Britton KL, Engel PC, Farrants GW, Lilley KS, Rice DW, Stillman TJ: Subunit assembly and active site location in the structure of glutamate dehydrogenase. Proteins 12:75-86 (1992).
3. Bansal A, Dayton MA, Zalkin H, Colman RF: Affinity labeling of a glutamyl peptide in the coenzyme binding site of NADP ÷ -specific glutamate dehydrogenase of Salmo- nelia typhimurium by 2-[(4-bromo-2,3-dioxobutyl)thio]-1, N6-ethenoadenosine 2', 5'-bisphosphate. J Biol Chem 264:9827-9835 (1989).
4. Bascomb NF, Schmidt RR: Purification and partial ki- netic and physical characterization of two chloroplast- localized NADP-specific glutamate dehydrogenase isoen- zymes and their preferential accumulation in Chlorella sorokiniana cells cultured at low or high ammonium levels. Plant Physiol 83:75-84 (1987).
5. Benachenhou N, Baldacci G: The gene for a halophilic glutamate dehydrogenase: sequence, transcription analy- sis and phylogenetic implications. Mol Gen Genet 230: 345-352 (1991).
6. Benachenhou-Lahfa N, Forterre P, Labedau B: Evolu- tion of glutamate dehydrogenase genes: evidence for two paralogous protein families and unusual branching pat-
terns of the Archaebacteria in the universal tree of life. J Mol Evol 36:335-346 (1993).
7. Bonete MJ, Camacho ML, Cadenas E: A new glutamate dehydrogenase from Halobacteriurn halobium with differ- ent coenzyme specificity. Int J Biochem 19:1149-1155 (1987).
8. BOrmann ER, Eikmanns B J, Sahm H: Molecular analy- sis of the Corynebacterium glutamicum gdh gene encoding glutamate dehydrogenase. Mol Microbiol 6:317-326 (1992).
9. BOrmann-EI Kholy ER, Eikmans B J, Gutmann M, Sahm H: Glutamate dehydrogenase is not essential for glutamate formation by Corynebacterium glutamicum. Appl Environ Microbiol 59:2329-2331 (1993).
10. Bradford M: A rapid and sensitive method for the quan- titation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal Biochem 72: 248- 254 (1976).
11. Brett M, Chambers GK, Holder AA, Fincham JRS, Wootton JC: Mutational amino acid replacements in Neurospora crassa NADP-specific glutamate dehydroge- nase. J Mol Biol 106:1-22 (1976).
12. Broedel Jr SE, Wolf Jr RE: Growth-phase-dependent induction of 6-phosphogluconate dehydrogenase and glu- cose 6-phosphate dehydrogenase in the cyanobacterium Synechococcus sp. PCC 7942. Gene 109:71-79 (1991).
13. Chauvat F, de Vries L, van der Ende A, van Arkel GA: A host-vector system for gene cloning in the cyanobac- terium Synechocystis PCC 6803. Mol Gen Genet 204: 185-191 (1986).
14. Chauvat F, Labarre J, Ferino F: Development of genes transfer system for the cyanobacterium Synechocystis PCC 6803. Plant Physiol Biochem 26:629-637 (1988).
15. Ch~vez S, Candau P: An NAD-specific glutamate dehy- drogenase from cyanobacteria. Identification and prop- erties. FEBS Lett 285:35-38 (1991).
16. Cock JM, Kim KD, Miller PW, Hutson RG, Schmidt RR: A nuclear gene with many introns encoding ammo- nium-inducible chloroplastic NADP-specific glutamate dehydrogenase(s) in Chlorella sorokiniana. Plant Mol Biol 17:1023-1044 (1991).
17. Colman RF: Glutamate dehydrogenase (bovine liver). In: Kuby SA (ed) A Study of Enzymes, vol II. Mechanism of Enzyme Action, pp. 173-192. CRC Press, Boca Raton, FL (1991).
18. Consalvi V, Chiaraluce R, Politi L, Gambacorta A, De Rosa M, Scandurra R: Glutamate dehydrogenase from the thermoacidophilic archaebacterium Sulfolobus solfa- taricus. Eur J Biochem 196:459-467 (1991).
19. Curry KA, Tomich CSC: Effect of ribosome binding site on gene expression in Escherichia coli. DNA 7:173-179 (1988).
20. Das AT, Moerer P, Charles R, Moorman AFM, Lamers WH: Nucleotide sequence of rat liver glutamate dehydro- genase cDNA. Nucl Acids Res 17:2355 (1989).
21. Dayhoff MO, Barker WC, Hunt LT: Establishing ho-

mologles in protein sequences. Meth Enzymol 91: 524- 545 (1983).
22. De Boer HA, Kastein RA: Biased codon usage: an ex- ploration of its role in optimization of translation. In: ReznikoffW, Gold L (eds) Maximizing Gene Expression, pp. 225-285. Butterworths, Boston (1986).
23. Devereux J, Haeberli P, Smithies O: A comprehensive set of sequences analysis programs for the VAX. Nucl Acids Res 12:387-395 (1984).
24. De Zoysa PA, Connerton IF, Watson DC, Johnston JR: Cloning, sequencing and expression of the Schwanniomy- ces occidentalis NADP-dependent glutamate dehydroge- nase gene. Curr Genet 20:219-224 (1991).
25. Di Rugglero J, Robb FT, Jagust R, Klump HH, Borges KM, Kessel M, Mal X, Adams MWW: Characterization, cloning, and in vitro expression of the extremely thermo- stable glutamate dehydrogenase from the hyperthermo- philic Archaeon, ES4. J Biol Chem 268:17767-17774 (1993).
26. Douglas SE, Turner S: Molecular evidence for the origin of plastids from a cyanobacterium-like ancestor. J Mol Evol 33:267-273 (1991).
27. Ferino F, Chauvat F: A promoter-probe vector-host sys- tem for the cyanobacterium Synechocystis PCC 6803. Gene 84:257-266 (1989).
28. Fickett JW: Recognition of protein coding regions in DNA sequences. Nucl Acids Res 10:5302-5319 (1982).
29. Florencio FJ, Marquts S, Candau P: Identification and characterization of a glutamate dehydrogenase in the uni- cellular cyanobacterium Synechocystis PCC 6803. FEBS Lett 223:37-41 (1987).
30. Gribskov M, Devereux J, Burguess RR: The codon pref- erence plot: graphic analysis of protein coding sequences and prediction of gene expression. Nucl Acids Res 12: 539-549 (1984).
31. Haeffner-Gormley L, Chen Z, Zalkin H, Colman RF: Evaluation of cysteine 238 and glutamic acid 284 in the coenzyme binding site of Salmonella typhimurium glutamate dehydrogenase by site-directed mutagenesis and reaction with the nucleotide analogue 2-((4-bromo- 2,3-dioxobutyl)thio) 1,N6-ethenoadenosine 2 ' ,5'-bisphos- phate. J Biol Chem 266:5388-5394 (1991).
32. Hajnsdorf E, Steier O, Coscoy L, Teysset L, Rtgnier P: Roles of RNase E, RNase II and PNPase in the degra- dation of the rpsO transcripts of Escherichia coli: stabiliz- ing function of RNase II and evidence for efficient deg- radation in an amspnp rnb mutant. EMBO J 13: 3368- 3377 (1994).
33. Hanahan D: Techniques for transformation ofE. coli. In: Glover DM (ed) DNA Cloning, vol. 1, pp. 109-135. IRL Press, Oxford (1985).
34. Hawkins AR, Gurr S J, Montague P, Kinghorn JR: Nucleotide sequence and regulation of expression of the Aspergillus nidulans gdhA gene encoding NADP depen- dent glutamate dehydrogenase. Mol Gen Genet 218: 105-111 (1989).
187
35. Henikoff S: Unidirectional digestion with exonuclease III creates targeted breakpoints for DNA sequencing. Gene 28:351-359 (1984).
36. Higgins DG, Bleasby AJ, Fuchs R: CLUSTAL V: im- proved software for multiple sequence alignment. CA- BI tS 8:189-191 (1992).
37. Jones KM, McPherson MJ, Baron AJ, Mattaj IW, Riordan CL, Wootton JC: The gdhA1 point mutation in Escherichia coli K12 CLR207 alters a key lysine residue of glutamate dehydrogenase. Mol Gen Genet 240: 286- 289 (1993).
38. Kinnaird JH, Fincham RS: The complete nucleotide se- quence of the Neurospora crassa am (NADP-specific glutamate dehydrogenase) gene. Gene 26:253-260 (1983).
39. Liaud M-F, Zhang DX, Cerff R: Differential intron loss and endosymbiotic transfer of chloroplast glyceraldehyde- 3-phosphate dehydrogenase gene to the nucleus. Proc Natl Acad Sci USA 87:8918-8922 (1990).
40. Lightfoot DA, Baron AJ, Wootton JC: Expression of the Escherichia coli glutamate dehydrogenase gene in the cy- anobacterium Synechococcus PCC 6301 causes ammo- nium tolerance. Plant Mol Biol 11:335-344 (1988).
41. Lyerly DM, Barroso LA, Wilkins TD: Identification of the latex test-reactive protein of Clostridium difficile as glutamate dehydrogenase. J Clin Microbiol 29: 2639- 2642 (1991).
42. Mackinney G: Absorption of light by chlorophyll solu- tions. J Biol Chem 140:315-322 (1941).
43. Maras B, Consalvi V, Chiaraluce R, Politi M, De Rosa L, Bossa F, Scandurra R, Barra D: The protein sequence of glutamate dehydrogenase from Sulfolobus solfataricus, a thermoacidophilic archaebacterium. Is the presence of N-e-methyllysine related to thermostability? Eur J Bio- chem 203:81-87 (1992).
44. Marraccini P, Cassier-Chauvat C, Bulteau S, Ch~tvez S, Chauvat F: Light-regulated promoters from Synechocystis PCC6803 share a consensus motif involved in photoregu- lation. Mol Microbiol 12:1005-1012 (1994).
45. Magalh,~tes JR, Ju GC, Rich PJ, Rhodes D: Kinetics of 15NH4+ assimilation in Zea mays: preliminary studies with a glutamate dehydrogenase (GDH1) null mutant. Plant Physiol 94:647-656 (1990).
46. Markwell MA, Haas SM, Bieber LL, Tolber NE: A modification of the Lowry procedure to simplify protein determination in membrane and lipoprotein samples. Anal Biochem 87:206-210 (1978).
47. Mattaj IW, McPherson MJ, Wootton JC: Localisation of a strongly conserved section of coding sequence in glutamate dehydrogenase genes. FEBS Lett 147:21-25 (1982).
48. Mayes SR, Cook KM, Self SJ, Zhang Z, Barber J: De- letion of the gene encoding the Photosystem II 33 kDa protein from Synechocystis sp. PCC 6803 does not inac- tivate water-splitting but increase vulnerability to photo- inhibition. Biochim Biophys Acta 1060:1-12 (1991).

188
49. McPherson MJ, Wootton JC: Complete nucleotide se- quence of the Escherichia coli gdhA gene. Nucl Acids Res 15:5257-5266 (1983).
50. Meeks JC, Wolk CP, Lockau W, Schilling N, Shaffer PW, Chien W-S: Pathways of assimilation of [13N]N2 and 13NH4 by cyanobacteria with and without hetero- cysts. J Bact 134:125-130 (1978).
51. Meredith MJ, Gronotajski RM, Schmidt RR: Physical and kinetic properties of the nicotinamide adenine dinucleotide-specific glutamate dehydrogenase purified from Chlorella sorokiniana. Plant Physiol 61:967-974 (1978).
52. M6rida A, Candau P, Florencio FJ: Regulation of glutamine synthetase activity in the unicellular cyanobac- terium Synechocystis sp. strain PCC 6803 by the nitrogen source: effect of ammonium. J Bact 173:4095-4100 (1991).
53. Metz J, Nixon PJ, Diner B: Nucleotide sequence of the psbA3 gene from the cyanobacterium Synechocystis PCC 6803. Nucl Acids Res 18:6715-6715 (1990).
54. Mohamed A, Jansson C: Influence of light on accumu- lation of photosynthesis-specific transcripts in the cyano- bacterium Synechocystis 6803. Plant Mol Biol 13: 693- 700 (1989).
55. Mountain A, McPherson MJ, Baron AJ, Wootton JC: The Klebsiella aerogenes (gdhA ) gene: cloning, high level expression and hybrid enzyme formation in Escherichia coil Mol Gen Genet 199:141-145 (1985).
56. Moye WS, Amuro N, Rao JKM, Zalkin H: Nucleotide sequence of yeast GDH1 encoding nicotinamide adenine dinucleotide phosphate-dependent glutamate dehydroge- nase. J Biol Chem 260:8502-8508 (1985).
57. Neilson AH, Doudoroff M: Ammonia assimilation in blue-green algae. Arch Mikrobiol 89:15-22 (1973).
58. Pearce J, Leach CK, Carr NG: The incomplete tricar- boxilic acid cycle in the blue-green alga Anabaena vari- abilis. J Gen Microbiol 55:371-378 (1969).
59. Pearson WR, Lipman DJ: Improved tools for biological sequence comparison. Proc Natl Acad Sci USA 85: 2444-2448 (1988).
60. Petersen C: Control of functional mRNA stability in bac- teria: multiple mechanisms ofnucleolytic and non-nucleo- lytic inactivation. Mol Microbiol 6:277-282 (1992).
61. Philbrick JB, Zilinskas BA: Cloning, nucleotide sequence and mutational analysis of the gene encoding the photo- system II manganese-stabilizing polypeptide of Syn- echocystis 6803. Mol Gen Genet 212:418-425 (1988).
62. Reitzer LJ, Magasanik B: Ammonia assimilation and the biosynthesis of glutamine, glutamate, aspartate, as- paragine, L-alanine and D-alanine. In: Neidhart FC, Ingraham JL, Low KB, Magasanik B, Schaechter M, Umbarger HE (eds) Escherichia coli and Salmonella typh- imurium. Cellular and Molecular Biology, vol I, pp. 302- 320. American Society for Microbiology, "Washington, DC (1987).
63. Robinson SA, Slade AP, Fox GG, Philips R, Ratcliffe
RG, Stewart GR: The role of glutamate dehydrogenase in plant nitrogen metabolism. Plant Physiol 95:509-516 (1991).
64. Sambrook J, Fritsch EF, Maniatis T: Molecular Cloning. A Laboratory Manual. Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY (1989).
65. Sanger F, Nicklen F, Coulson AR: DNA sequencing with chain-terminating inhibitors. Proc Nail Acad Sci USA 74:5463-5467 (1977).
66. Shaw WV: Chloramphenicol acetyltransferase from chloramphenicol-resistant bacteria. Meth Enzymol 43: 737-755 (1975).
67. Snedecor B, Chu H, Chen E: Selection, expression, and nucleotide sequencing of the glutamate dehydrogenase gene of Peptostreptococcus asaccharolyticus. J Bact 173: 6162-6167 (1991).
68. Sogin ML, Gunderson JH, Elwood HE, Alonso RA, Pe- attie DA: Phylogenetic meaning of the kingdom concept: an unusual ribosomal RNA from Giardia lamblia. Science 243:75-77 (1991).
69. Stewart GR, Mann AF, Fentem PA: Enzymes of glutamate formation: glutamate dehydrogenase, glutamine synthetase, and glutamate synthase. In: Stumpf K, Conn EE (eds) The Biochemistry of Plants, vol. 5, pp. 271-327. Academic Press, New York (1980).
70. Studier FW, Rosenberg AH, Dunn J J, Dubendorff JW: Use ofT7 RNA polymerase to direct expression of cloned genes. Meth Enzymol 185:60-89 (1990).
71. Tandeau de Marsac N, Borrias WE, Kuhlemeier CJ, Castets AM, van Arkel GA, van den Hondel CAMJJ: A new approach for molecular cloning in cyanobacteria: cloning of an Anacystis nidulans met gene using a Tn901- induced mutant. Gene 20:111-119 (1982).
72. Teller JK, Smith RJ, McPherson MJ, Engel PC, Guest JR: The glutamate dehydrogenase gene of Clostridium symbiosum. Cloning by polymerase chain reaction, se- quence analysis and over-expression in Escherichia coil Eur J Biochem 206:151-159 (1992).
73. Tzimagiorgis G, Moschonas NK: Molecular cloning, structure and expression analysis of a full-length mouse brain glutamate dehydrogenase cDNA. Biochim Biophys Acta 1089:250-253 (1991).
74. Vicente M, Kushner SR, Garrido T, Aldea M: The role of the 'gearbox' in the transcription of essential genes. Mol Microbiol 5 :2085-2091 (1991).
75. Wierenga RK, De Maeyer MCH, Hol WGJ: Interaction of pyrophosphate moieties with ~-helixes in dinucleotide binding proteins. Biochemistry 24:1346-1357 (1985).
76. Williams JGK: Construction of specific mutations in pho- tosystem II photosynthetic reaction center by genetic en- gineering methods in Synechocystis 6803. Meth Enzymol 167:766-778 (1988).
77. Yee J, Dennis PP: Isolation and characterization of a NADP-dependent glutamate dehydrogenase gene from the primitive eucaryote Giardia lamblia. J Biol Chem 267: 7539-7544 (1992).




















