
The mRNA Surveillance Protein hSMG-1 Functions in Genotoxic Stress Response Pathways in Mammalian...
14
Molecular Cell, Vol. 14, 585–598, June 4, 2004, Copyright 2004 by Cell Press The mRNA Surveillance Protein hSMG-1 Functions in Genotoxic Stress Response Pathways in Mammalian Cells The most recently described PIKK family member was identified in human cells and named hSMG-1, based on its homology to the Caenorhabditis elegans protein CeSMG-1 (Denning et al., 2001; Yamashita et al., 2001). In the worm, CeSMG-1 functions in a RNA surveillance Kathryn M. Brumbaugh, 1,4 Diane M. Otterness, 1,4 Christoph Geisen, 1 Vasco Oliveira, 1 John Brognard, 1 Xiaojie Li, 2 Fabrice Lejeune, 2 Randal S. Tibbetts, 3 Lynne E. Maquat, 2 and Robert T. Abraham 1, * 1 Program in Signal Transduction Research pathway termed nonsense-mediated mRNA decay (NMD) (Cali and Anderson, 1998; Hodgkin et al., 1989; The Burnham Institute La Jolla, California 92037 Maquat, 2003; Page et al., 1999; Pulak and Anderson, 1993). NMD mediates the rapid degradation of mRNAs 2 Department of Biochemistry and Biophysics School of Medicine and Dentistry bearing premature termination codons (PTCs), which threaten cell survival by producing truncated, potentially University of Rochester Rochester, New York 14642 dominant-interfering proteins (Frischmeyer and Dietz, 1999; Maquat, 2003; Maquat and Carmichael, 2001). Ya- 3 Department of Pharmacology University of Wisconsin mashita et al. (2001) provided evidence that inhibition of hSMG-1 activity in mammalian cells leads to suppres- Madison, Wisconsin 53706 sion of NMD, suggesting that hSMG-1 is indeed a func- tional homolog of CeSMG-1. Although the exact mecha- nism remains undefined, hSMG-1 regulates NMD in part Summary through the phosphorylation of hUpf1, a conserved component of the NMD machinery in metazoan cells Members of the PI 3-kinase-related kinase (PIKK) fam- ily function in mitogenic and stress-induced signaling (Denning et al., 2001; Page et al., 1999; Yamashita et al., 2001). hSMG-1 phosphorylates at least four sites in pathways in eukaryotic cells. Here, we characterize the newest PIKK family member, hSMG-1, as a genotoxic the carboxy-terminal, S/T-Q-rich domain of hUpf1, and mutational analysis indicated that these phosphoryla- stress-activated protein kinase that displays some functional overlap with the related kinase, ATM, in hu- tion events were important for NMD activity (Yamashita et al., 2001). man cells. Both ATM and hSMG-1 phosphorylate Ser/ Thr-Gln-containing target sequences in the checkpoint During a search for PIKK family members, we indepen- dently isolated a human cDNA that was nearly identical protein p53 and the nonsense-mediated mRNA decay (NMD) protein hUpf1. Expression of hSMG-1 is required to hSMG-1. Based on initial biochemical and functional characterizations of the encoded protein, we named this for optimal p53 activation after cellular exposure to ge- notoxic stress, and depletion of hSMG-1 leads to spon- PIKK “ATX” to acknowledge its similarity to the genome surveillance kinases ATM and ATR (Shiloh, 2003). To taneous DNA damage and increased sensitivity to ion- izing radiation (IR). Moreover, IR exposure triggers maintain consistency with the precedent literature (Den- ning et al., 2001; Yamashita et al., 2001), we have re- hUpf1 phosphorylation at Ser/Thr-Gln motifs, and both ATM and hSMG-1 contribute to these phosphorylation named our cDNA hSMG-1. In this report, we present evidence that, in addition to its critical role in NMD, events. Finally, NMD is suppressed in hSMG-1- but not ATM-deficient cells. These results indicate that hSMG-1 hSMG-1 is centrally involved in the regulation of the genotoxic stress response machinery in mammalian cells. plays important roles in the maintenance of both ge- nome and transcriptome integrity in human cells. Cellular exposure to ultraviolet (UV) light or IR stimulates hSMG-1 kinase activity, and hSMG-1 function is required for maximal p53 phosphorylation and stabilization in Introduction radiation-damaged cells. Moreover, hSMG-1-deficient The phosphoinositide 3-kinase-related kinases (PIKKs) cells are sensitized to IR-induced stress. These results comprise a family of high molecular mass signaling pro- indicate that hSMG-1, like ATM and ATR, is required for teins that function in diverse cellular processes, includ- the maintenance of genomic stability in mammalian ing cell growth, genome surveillance, and gene expres- cells. sion (Abraham, 2001; Tibbetts and Abraham, 2000). Mammalian cells express six PIKK family members, three of which (ATM, ATR, and DNA-dependent protein Results kinase) participate in cell-cycle checkpoint and DNA repair pathways. These three genome surveillance ki- Cloning of the hSMG-1 cDNA nases transmit signals through the phosphorylation of During a BLAST search for novel PIKKs, we noted an proteins on Ser and Thr residues and share a strong expressed sequence tag (EST) (KIAA0421) bearing ho- preference for protein substrates containing core Ser/ mology to a conserved region in the catalytic domains Thr-Gln (S/T-Q) motifs, which are frequently found in of PIKK family members. The full-length hSMG-1 cDNA multiple copies localized in “S/T-Q-rich domains.” was assembled from overlapping cDNA fragments ob- tained by PCR amplification of human brain and Jurkat T cell cDNA libraries. This cDNA encodes a 3521 amino *Correspondence: [email protected] 4 These authors contributed equally to this work. acid polypeptide with a deduced molecular mass of
-
Upload
independent -
Category
Documents
-
view
2 -
download
0
Transcript of The mRNA Surveillance Protein hSMG-1 Functions in Genotoxic Stress Response Pathways in Mammalian...

Molecular Cell, Vol. 14, 585–598, June 4, 2004, Copyright 2004 by Cell Press
The mRNA Surveillance Protein hSMG-1Functions in Genotoxic Stress ResponsePathways in Mammalian Cells
The most recently described PIKK family member wasidentified in human cells and named hSMG-1, basedon its homology to the Caenorhabditis elegans proteinCeSMG-1 (Denning et al., 2001; Yamashita et al., 2001).In the worm, CeSMG-1 functions in a RNA surveillance
Kathryn M. Brumbaugh,1,4 Diane M. Otterness,1,4
Christoph Geisen,1 Vasco Oliveira,1
John Brognard,1 Xiaojie Li,2 Fabrice Lejeune,2
Randal S. Tibbetts,3 Lynne E. Maquat,2
and Robert T. Abraham1,*1Program in Signal Transduction Research pathway termed nonsense-mediated mRNA decay
(NMD) (Cali and Anderson, 1998; Hodgkin et al., 1989;The Burnham InstituteLa Jolla, California 92037 Maquat, 2003; Page et al., 1999; Pulak and Anderson,
1993). NMD mediates the rapid degradation of mRNAs2 Department of Biochemistry and BiophysicsSchool of Medicine and Dentistry bearing premature termination codons (PTCs), which
threaten cell survival by producing truncated, potentiallyUniversity of RochesterRochester, New York 14642 dominant-interfering proteins (Frischmeyer and Dietz,
1999; Maquat, 2003; Maquat and Carmichael, 2001). Ya-3 Department of PharmacologyUniversity of Wisconsin mashita et al. (2001) provided evidence that inhibition
of hSMG-1 activity in mammalian cells leads to suppres-Madison, Wisconsin 53706sion of NMD, suggesting that hSMG-1 is indeed a func-tional homolog of CeSMG-1. Although the exact mecha-nism remains undefined, hSMG-1 regulates NMD in partSummarythrough the phosphorylation of hUpf1, a conservedcomponent of the NMD machinery in metazoan cellsMembers of the PI 3-kinase-related kinase (PIKK) fam-
ily function in mitogenic and stress-induced signaling (Denning et al., 2001; Page et al., 1999; Yamashita etal., 2001). hSMG-1 phosphorylates at least four sites inpathways in eukaryotic cells. Here, we characterize the
newest PIKK family member, hSMG-1, as a genotoxic the carboxy-terminal, S/T-Q-rich domain of hUpf1, andmutational analysis indicated that these phosphoryla-stress-activated protein kinase that displays some
functional overlap with the related kinase, ATM, in hu- tion events were important for NMD activity (Yamashitaet al., 2001).man cells. Both ATM and hSMG-1 phosphorylate Ser/
Thr-Gln-containing target sequences in the checkpoint During a search for PIKK family members, we indepen-dently isolated a human cDNA that was nearly identicalprotein p53 and the nonsense-mediated mRNA decay
(NMD) protein hUpf1. Expression of hSMG-1 is required to hSMG-1. Based on initial biochemical and functionalcharacterizations of the encoded protein, we named thisfor optimal p53 activation after cellular exposure to ge-
notoxic stress, and depletion of hSMG-1 leads to spon- PIKK “ATX” to acknowledge its similarity to the genomesurveillance kinases ATM and ATR (Shiloh, 2003). Totaneous DNA damage and increased sensitivity to ion-
izing radiation (IR). Moreover, IR exposure triggers maintain consistency with the precedent literature (Den-ning et al., 2001; Yamashita et al., 2001), we have re-hUpf1 phosphorylation at Ser/Thr-Gln motifs, and both
ATM and hSMG-1 contribute to these phosphorylation named our cDNA hSMG-1. In this report, we presentevidence that, in addition to its critical role in NMD,events. Finally, NMD is suppressed in hSMG-1- but not
ATM-deficient cells. These results indicate that hSMG-1 hSMG-1 is centrally involved in the regulation of thegenotoxic stress response machinery in mammalian cells.plays important roles in the maintenance of both ge-
nome and transcriptome integrity in human cells. Cellular exposure to ultraviolet (UV) light or IR stimulateshSMG-1 kinase activity, and hSMG-1 function is requiredfor maximal p53 phosphorylation and stabilization inIntroductionradiation-damaged cells. Moreover, hSMG-1-deficient
The phosphoinositide 3-kinase-related kinases (PIKKs) cells are sensitized to IR-induced stress. These resultscomprise a family of high molecular mass signaling pro- indicate that hSMG-1, like ATM and ATR, is required forteins that function in diverse cellular processes, includ- the maintenance of genomic stability in mammalianing cell growth, genome surveillance, and gene expres- cells.sion (Abraham, 2001; Tibbetts and Abraham, 2000).Mammalian cells express six PIKK family members,three of which (ATM, ATR, and DNA-dependent protein Resultskinase) participate in cell-cycle checkpoint and DNArepair pathways. These three genome surveillance ki- Cloning of the hSMG-1 cDNAnases transmit signals through the phosphorylation of During a BLAST search for novel PIKKs, we noted anproteins on Ser and Thr residues and share a strong expressed sequence tag (EST) (KIAA0421) bearing ho-preference for protein substrates containing core Ser/ mology to a conserved region in the catalytic domainsThr-Gln (S/T-Q) motifs, which are frequently found in of PIKK family members. The full-length hSMG-1 cDNAmultiple copies localized in “S/T-Q-rich domains.” was assembled from overlapping cDNA fragments ob-
tained by PCR amplification of human brain and JurkatT cell cDNA libraries. This cDNA encodes a 3521 amino*Correspondence: [email protected]
4These authors contributed equally to this work. acid polypeptide with a deduced molecular mass of

Molecular Cell586
395 kDa and shows extensive sequence identity to the Yamashita et al., 2001). In the present studies, we com-pared the abilities of hSMG-1 and ATM to phosphorylatehSMG-1 cDNAs described in two earlier reports (Den-p53 at Ser-15 (a canonical phosphorylation site for ATM)ning et al., 2001; Yamashita et al., 2001). Our hSMG-1and the hUpf1 carboxy-terminal domain, which containscDNA is eight amino acids shorter at the amino terminusat least two hSMG-1 phosphorylation sites (Yamashitathan the cDNA studied by Yamashita et al. (2001). Thiset al., 2001). HEK 293T cells were transfected with adifference reflects alternative usage of exon four in ourplasmid vector encoding HA-SMG-1wt, HA-SMG-1KI, orhSMG-1 cDNA versus exon five in the Yamashita et al.HA-tagged wild-type ATM (HA-ATMwt), and �-HA immu-(2001) clone (see Supplemental Figure S1 at http://www.noprecipitates were assayed for kinase activity withmolecule.org/cgi/content/full/14/5/585/DC1). We alsoGST-p531–70 as the substrate (Tibbetts et al., 1999). Weisolated the hSMG-1 isoform described by Yamashitaobserved that HA-SMG-1wt displayed robust GST-p531–70et al. (2001); however, the clone that we chose for furtherkinase activity (Figure 1B, left panel). Based on the ratioanalysis was found most frequently in our cDNA librar-of substrate radiolabeling to the level of HA-tagged pro-ies. The biological significance of the sequence variationtein, the specific kinase activity of HA-SMG-1wt was ap-at the amino terminus of hSMG-1 remains unclear.proximately 3.5-fold higher than that of HA-ATMwt overthree independent trials. As observed in previous stud-
Expression of Endogenous and Recombinant hSMG-1 ies with ATM and ATR (Banin et al., 1998; Canman etAs previously reported (Denning et al., 2001; Yamashita al., 1998; Tibbetts et al., 1999), phosphorylation of GST-et al., 2001), multitissue Northern analysis revealed that p531–70 by HA-SMG-1wt was nearly abolished by a Ser→hSMG-1 mRNA transcripts were widely expressed in Ala substitution at Ser-15 (Figure 1B, right panel). Wehuman tissues, with the highest levels observed in heart also confirmed that the p53 (Ser-15) kinase activity ofand skeletal muscle. Immunoblot analyses indicated hSMG-1 was sensitive to wortmannin (Yamashita et al.,that the hSMG-1 protein was expressed at readily de- 2001) (Supplemental Figure S3 on Molecular Cell’s web-tectable levels in human hematopoietic, mesenchymal, site). The drug concentrations required for half-maximaland epithelial cell lines (our unpublished data). Database inhibition (IC50) of hSMG-1 activity in immune complexessearches revealed that the hSMG-1 showed the highest and in intact cells were approximately 80 nM and 2 �M,overall sequence homology to CeSMG-1 (Supplemental respectively, indicating that the wortmannin sensitivityFigure S2 on Molecular Cell’s website). Interestingly, of hSMG-1 was similar to that of ATM (Sarkaria et al.,hSMG-1 contains a conserved domain, termed NH2, 1998).which resembles the FKBP-12•rapamycin binding (FRB) While these studies were in progress, Yamashita et al.domain of mTOR. However, FKBP12•rapamycin neither (2001) demonstrated that the NMD pathway componentbinds the FRB-like domain nor inhibits hSMG-1 kinase hUpf1 was a substrate for hSMG-1. The amino acid se-activity in vitro (our unpublished data; Denning et al., quences surrounding the four carboxy-terminal phosphor-2001), indicating that hSMG-1 is not a relevant target ylation sites (LSQP, LSQD, LSQD, and LSQY) identifiedfor rapamycin in mammalian cells. in this study resemble the consensus site for phosphory-
To further characterize the translation product encoded lation by ATM (O’Neill et al., 2000). To comparativelyby our hSMG-1 cDNA, we transfected human embryonic evaluate the catalytic activities of hSMG-1 and ATM to-kidney (HEK) 293T cells with expression plasmids encod- ward the S/T-Q-rich domain of hUpf1, we constructed aing HA-tagged, wild-type hSMG-1 (HA-SMG-1wt) or kinase GST fusion protein containing the carboxyl terminus ofinactive hSMG-1 (HA-SMG-1KI). HA-SMG-1KI contains an hUpf1 (amino acids 1019–1118), including the four S-QAsp→Ala substitution at a conserved residue (Asp-2195) phosphorylation sites. The GST-hUpf11019–1118 protein
was phosphorylated by both HA-SMG-1wt and HA-ATMwtin the hSMG-1 catalytic domain. Similar mutations havein immune complex kinase assays, with HA-SMG-1wtbeen shown to inactivate the catalytic domains of vari-again showing higher specific activity under theseous PIKK family members (Canman et al., 1998; Clibyin vitro assay conditions (Figure 1B, left panel).et al., 1998; Hunter, 1995). Immunoblot analyses of
Exposure of cells to ionizing radiation (IR) stimulateswhole-cell extracts from nontransfected or HA-SMG-ATM kinase activity (Banin et al., 1998; Canman et al.,1-transfected HEK 293T cells revealed a single major1998). To determine whether the hSMG-1 activity wasimmunoreactive band migrating at the predicted molec-similarly responsive to radiation-induced stress, HA-ular mass of �395 kDa (Figure 1A, right panel). TheSMG-1wt-transfected HEK 293T cells were treated withmolecular mass of the recombinant HA-SMG-1 proteineither 10 Gy IR or 100 J/m2 UV-B light, and �-HA immuno-corresponded closely to that of the endogenousprecipitates were assayed for p53 (Ser-15) kinase activ-hSMG-1 (Figure 1A, left panel). Consistent with the pre-ity. Radiation exposure stimulated the protein kinasedicted size of hSMG-1, the band recognized by �-SMG-1activity of HA-SMG-1wt (Figure 1C). The maximal re-(Ab-1) migrated with a significantly lower electrophoreticsponse was observed at 1 hr postirradiation, and, inmobility than either ATM (molecular mass, 370 kDa) (Fig-repeated trials, IR proved to be a more effective stimulusure 1B) or ATR (molecular mass, 305 kDa) (see Supple-for hSMG-1 activation (Figure 1C, right panel). Interest-mental Figure S4 on Molecular Cell’s website).ingly, biochemical fractionation and immunofluores-cence microscopy revealed that hSMG-1 is present in
hSMG-1 Is a Stress-Activated Kinase that both the cytoplasmic and nuclear compartments (Sup-Phosphorylates p53 and hUpf1 In Vitro plemental Figure S4 on Molecular Cell’s website). Thus,Previous results indicated that the catalytic domain of this PIKK family member can potentially respond tohSMG-1, like that of ATM and ATR, preferentially targets stress signals generated in either the cytoplasm or nu-
cleus of irradiated cells.S/T-Q motifs for phosphorylation (Denning et al., 2001;
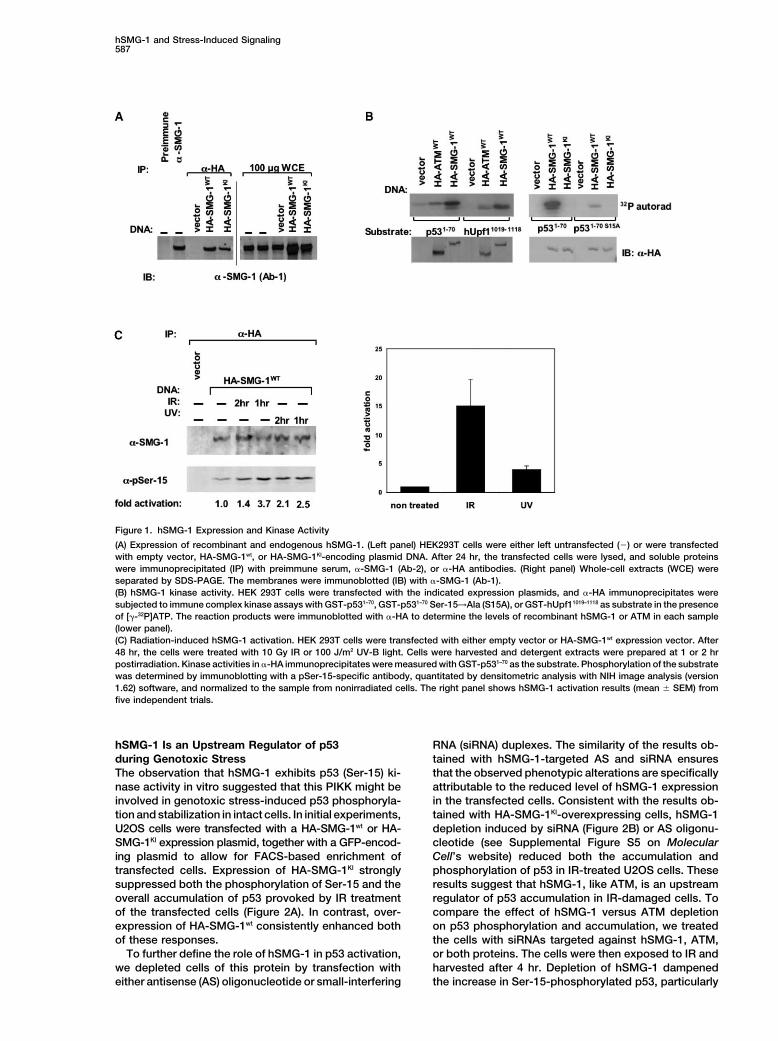
hSMG-1 and Stress-Induced Signaling587
Figure 1. hSMG-1 Expression and Kinase Activity
(A) Expression of recombinant and endogenous hSMG-1. (Left panel) HEK293T cells were either left untransfected (�) or were transfectedwith empty vector, HA-SMG-1wt, or HA-SMG-1KI-encoding plasmid DNA. After 24 hr, the transfected cells were lysed, and soluble proteinswere immunoprecipitated (IP) with preimmune serum, �-SMG-1 (Ab-2), or �-HA antibodies. (Right panel) Whole-cell extracts (WCE) wereseparated by SDS-PAGE. The membranes were immunoblotted (IB) with �-SMG-1 (Ab-1).(B) hSMG-1 kinase activity. HEK 293T cells were transfected with the indicated expression plasmids, and �-HA immunoprecipitates weresubjected to immune complex kinase assays with GST-p531–70, GST-p531–70 Ser-15→Ala (S15A), or GST-hUpf11019–1118 as substrate in the presenceof [�-32P]ATP. The reaction products were immunoblotted with �-HA to determine the levels of recombinant hSMG-1 or ATM in each sample(lower panel).(C) Radiation-induced hSMG-1 activation. HEK 293T cells were transfected with either empty vector or HA-SMG-1wt expression vector. After48 hr, the cells were treated with 10 Gy IR or 100 J/m2 UV-B light. Cells were harvested and detergent extracts were prepared at 1 or 2 hrpostirradiation. Kinase activities in �-HA immunoprecipitates were measured with GST-p531–70 as the substrate. Phosphorylation of the substratewas determined by immunoblotting with a pSer-15-specific antibody, quantitated by densitometric analysis with NIH image analysis (version1.62) software, and normalized to the sample from nonirradiated cells. The right panel shows hSMG-1 activation results (mean � SEM) fromfive independent trials.
hSMG-1 Is an Upstream Regulator of p53 RNA (siRNA) duplexes. The similarity of the results ob-tained with hSMG-1-targeted AS and siRNA ensuresduring Genotoxic Stress
The observation that hSMG-1 exhibits p53 (Ser-15) ki- that the observed phenotypic alterations are specificallyattributable to the reduced level of hSMG-1 expressionnase activity in vitro suggested that this PIKK might be
involved in genotoxic stress-induced p53 phosphoryla- in the transfected cells. Consistent with the results ob-tained with HA-SMG-1KI-overexpressing cells, hSMG-1tion and stabilization in intact cells. In initial experiments,
U2OS cells were transfected with a HA-SMG-1wt or HA- depletion induced by siRNA (Figure 2B) or AS oligonu-cleotide (see Supplemental Figure S5 on MolecularSMG-1KI expression plasmid, together with a GFP-encod-
ing plasmid to allow for FACS-based enrichment of Cell’s website) reduced both the accumulation andphosphorylation of p53 in IR-treated U2OS cells. Thesetransfected cells. Expression of HA-SMG-1KI strongly
suppressed both the phosphorylation of Ser-15 and the results suggest that hSMG-1, like ATM, is an upstreamregulator of p53 accumulation in IR-damaged cells. Tooverall accumulation of p53 provoked by IR treatment
of the transfected cells (Figure 2A). In contrast, over- compare the effect of hSMG-1 versus ATM depletionon p53 phosphorylation and accumulation, we treatedexpression of HA-SMG-1wt consistently enhanced both
of these responses. the cells with siRNAs targeted against hSMG-1, ATM,or both proteins. The cells were then exposed to IR andTo further define the role of hSMG-1 in p53 activation,
we depleted cells of this protein by transfection with harvested after 4 hr. Depletion of hSMG-1 dampenedthe increase in Ser-15-phosphorylated p53, particularlyeither antisense (AS) oligonucleotide or small-interfering

Molecular Cell588
Figure 2. Role of hSMG-1 in Stress-Induced p53 Phosphorylation and Accumulation
(A) hSMG-1 overexpression. U2OS cells were cotransfected with a GFP-NLS, together with empty vector, or with HA-SMG-1wtT- or HA-SMG-1KI-encoding plasmid. After 40 hr, the cells were exposed to 10 Gy IR, and GFP-positive cells were collected after 4 hr with a fluorescence-activated cell sorter. Whole-cell extracts were resolved by SDS-PAGE and sequentially immunoblotted with the indicated antibodies.(B) hSMG-1 depletion by siRNA. U2OS cells were mock transfected or transfected with hSMG-1 siRNA. After 48 hr, the cells were treatedwith 10 Gy IR or 200 J/m2 UV and were lysed after 4 hr. Soluble proteins were separated by SDS-PAGE and immunoblotted with theindicated antibodies.(C) ATM versus hSMG-1 depletion. U2OS cells were transfected with hSMG-1 or ATM siRNA, either singly or in combination. After 72 hr, thecells were treated with 3 or 10 Gy IR and harvested at 1 hr or 4 hr postirradiation. Detergent-soluble proteins were separated by SDS-PAGEand were immunoblotted with the indicated antibodies. LEVEL denotes p53 protein expression normalized to the nonirradiated, luciferasesiRNA-treated control sample.
at the lower (3 Gy) dose of IR (Figure 2C). After exposure transfected cells, treatment with 10 Gy IR or 200 J/m2
UV-B light triggered an increase in hChk2 phosphoryla-to 10 Gy IR, the defect in p53 expression was morepronounced in ATM-deficient cells, and approximately tion, as indicated by the increased intensity of the more
slowly migrating band in �-hChk2 immunoblots (Figureadditive effects were observed in cells treated with bothsiRNAs. The residual phosphorylation of p53 at Ser-15 2B). In contrast to the p53 results, hSMG-1 depletion
with siRNA (Figure 2B) or AS oligonucleotide (Supple-in the hSMG-1/ATM double-knockdown cells was likelydue at least in part to the activity of ATR, which is a mental Figure S5 on Molecular Cell’s website) did not
interfere with IR- or UV-induced phosphorylation ofknown p53 (Ser-15) kinase (Tibbetts et al., 1999). We alsonoted that hSMG-1-deficient cells displayed elevated hChk2 in U2OS cells. These results indicate that, while
ATM and hSMG-1 converge on p53 as a common target,levels of p53 phosphorylation and protein expression inthe absence of extrinsic genotoxic stress, suggesting hChk2 is regulated exclusively by ATM.that loss of hSMG-1 is sufficient to trigger a spontaneousDNA damage response (see below). DNA Damage Responses in hSMG-1-Deficient Cells
Closer examination of Figures 2B and 2C reveals thatA second target substrate for ATM in IR-damagedcells is the checkpoint kinase hChk2 (Bartek and Lukas, hSMG-1-deficent cells exhibit constitutive phosphoryla-
tion of hChk2 and p53, suggesting that these cells are2003). To determine whether hSMG-1 also participatedin hChk2 activation, we examined hChk2 phosphoryla- responding to DNA damage incurred in the absence of
extrinsic genotoxic agents. To further examine this possi-tion in hSMG-1 siRNA-treated U2OS cells. In mock-
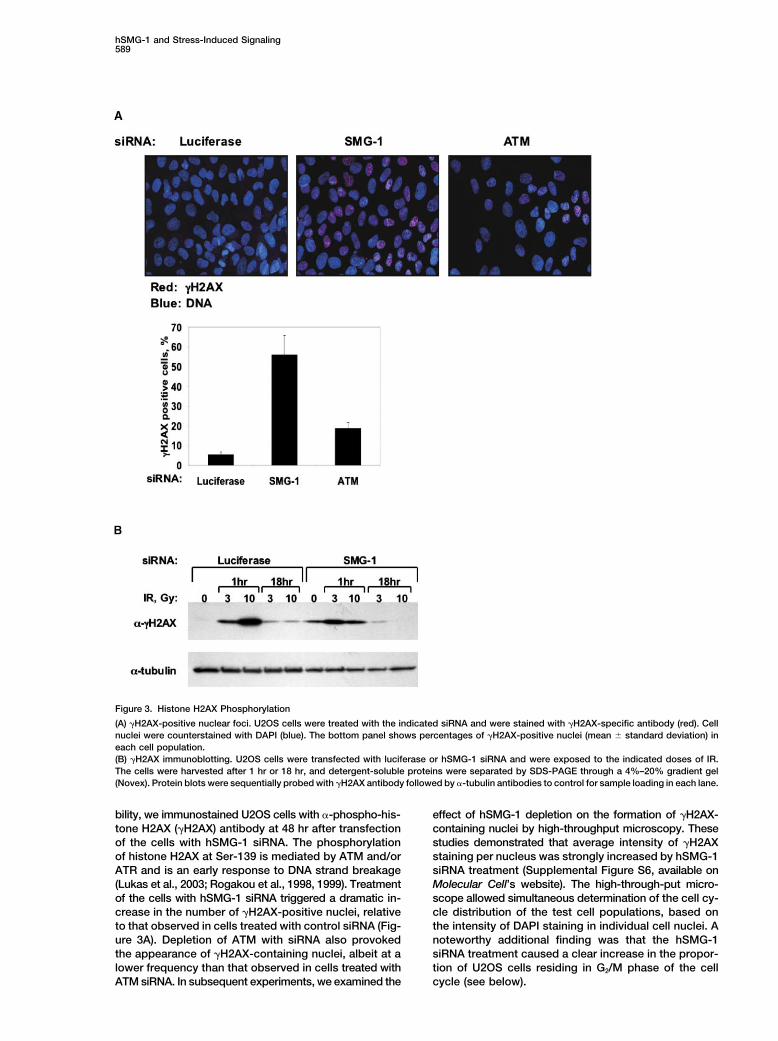
hSMG-1 and Stress-Induced Signaling589
Figure 3. Histone H2AX Phosphorylation
(A) �H2AX-positive nuclear foci. U2OS cells were treated with the indicated siRNA and were stained with �H2AX-specific antibody (red). Cellnuclei were counterstained with DAPI (blue). The bottom panel shows percentages of �H2AX-positive nuclei (mean � standard deviation) ineach cell population.(B) �H2AX immunoblotting. U2OS cells were transfected with luciferase or hSMG-1 siRNA and were exposed to the indicated doses of IR.The cells were harvested after 1 hr or 18 hr, and detergent-soluble proteins were separated by SDS-PAGE through a 4%–20% gradient gel(Novex). Protein blots were sequentially probed with �H2AX antibody followed by �-tubulin antibodies to control for sample loading in each lane.
bility, we immunostained U2OS cells with �-phospho-his- effect of hSMG-1 depletion on the formation of �H2AX-containing nuclei by high-throughput microscopy. Thesetone H2AX (�H2AX) antibody at 48 hr after transfection
of the cells with hSMG-1 siRNA. The phosphorylation studies demonstrated that average intensity of �H2AXstaining per nucleus was strongly increased by hSMG-1of histone H2AX at Ser-139 is mediated by ATM and/or
ATR and is an early response to DNA strand breakage siRNA treatment (Supplemental Figure S6, available onMolecular Cell’s website). The high-through-put micro-(Lukas et al., 2003; Rogakou et al., 1998, 1999). Treatment
of the cells with hSMG-1 siRNA triggered a dramatic in- scope allowed simultaneous determination of the cell cy-cle distribution of the test cell populations, based oncrease in the number of �H2AX-positive nuclei, relative
to that observed in cells treated with control siRNA (Fig- the intensity of DAPI staining in individual cell nuclei. Anoteworthy additional finding was that the hSMG-1ure 3A). Depletion of ATM with siRNA also provoked
the appearance of �H2AX-containing nuclei, albeit at a siRNA treatment caused a clear increase in the propor-tion of U2OS cells residing in G2/M phase of the celllower frequency than that observed in cells treated with
ATM siRNA. In subsequent experiments, we examined the cycle (see below).

Molecular Cell590
The increased accumulation of �H2AX in hSMG-1- assays with hSMG-1-depleted A549 cells after treatmentdepleted cells was confirmed by immunoblotting of with various doses of IR. Loss of hSMG-1 caused anuclear extracts with �H2AX antibody (Figure 3B). Inter- significant increase in the radiosensitivity of hSMG-1estingly, hSMG-1-deficient cells showed an IR dose- cells (Figure 5). The level of radiosensitization observeddependent defect in histone H2AX phosphorylation that in this setting was similar to that previously reportedwas evident at 1 hr after treatment with 10 Gy IR but for A549 cells treated with 10 �M wortmannin, a drugnot with the lower dose of 3 Gy IR. At 18 hr postirradia- concentration known to mediate substantial inhibitiontion, �H2AX levels had declined to near baseline in both of ATM, DNA-dependent protein kinase, and hSMG-1the luciferase and hSMG-1 siRNA-transfected cells, sug- (Sarkaria et al., 1998; Supplemental Figure S3 on Molec-gesting that loss of hSMG-1 function did not impair check- ular Cell’s website).point inactivation after the resolution of IR-induced DNAdamage. Similar results were obtained when the phos- hSMG-1 and ATM Jointly Contribute to Stress-phorylation of a second checkpoint protein, NBS1, was Induced hUpf1 Phosphorylationexamined in hSMG-1 siRNA-treated U2OS cells (data The hUpf1 protein, like its CeSMG-2 ortholog, is an es-not shown). These findings suggest that hSMG-1-defi- sential component of the NMD pathway (Culbertson,cient cells develop spontaneous DNA damage, leading 1999; Mendell et al., 2002; Sun et al., 1998; Wang et al.,to activation of ATM/ATR-mediated checkpoint re- 2002) and a physiologically relevant target for SMG-1sponses. kinases (see Figure 1B; Anders et al., 2003; Denning et
The checkpoint signaling defects observed in hSMG- al., 2001; Pal et al., 2001; Yamashita et al., 2001). In the1-depleted cells prompted a more detailed examination present study, we were interested in the possibility thatof their cell cycle profiles in the absence or presence of the phosphorylation state of hUpf1 might be modulatedIR-induced DNA damage. U2OS cells were transfected by IR-induced stress. Under normal conditions, hUpf1with hSMG-1 AS oligonucleotide or with the correspond- is subject to continuous phosphorylation-dephosphory-ing sense (S) oligonucleotide, and cell cycle distributions lation cycles (Anders et al., 2003; Chiu et al., 2003; Oh-were determined at 16 hr after IR exposure. In nonirradi- nishi et al., 2003), due in part to the action of a proteinated samples, AS treatment caused a clear reduction phosphatase 2A(PP2A)-like activity that is inhibited byin the percentage of G1 phase cells and a concomitant okadaic acid (OA) (Yamashita et al., 2001). We treatedaccumulation of G2/M phase cells (Figure 4A). The AS- FLAG-hUpf1-transfected U2OS cells with IR or OA, ortreated samples also contained an increased subpopu- both agents in combination, and harvested the cells atlation with �2 N DNA content, which suggested that 1 hr or 4 hr postirradiation. OA treatment provoked thethese cells were undergoing apoptotic cell death (see progressive accumulation of phosphate at S/T-Q sitessummary table in Figure 4A). After IR exposure, cells in FLAG-hUpf1 (Figure 6A), confirming that steady-statetreated with the S oligonucleotide accumulated in both phosphorylation at these sites is regulated by a PP2A-G1 and G2/M phases, a profile typical of p53-positive related phosphatase. More importantly, we observedcells that retain the G1 and G2 checkpoints. In contrast, that IR exposure also increased the phosphorylation ofthe AS-treated cells arrested primarily in G2/M phase
S/T-Q sites in FLAG-Upf1, consistent with earlier evi-after IR exposure, consistent with the notion that the
dence (see Figure 1) that this protein is a substrate forp53-dependent G1 checkpoint is compromised in these
at least two IR-stimulated kinases, hSMG-1 and ATM.cells (North and Hainaut, 2000; Waldman et al., 1995).
If hSMG-1 and ATM jointly participate in hUpf1 phos-Immunoblot analyses of the same cell populations con-phorylation in intact cells, then simultaneous depletionfirmed that hSMG-1 levels were profoundly reduced byof both proteins should suppress the phosphorylationthe AS treatment, with no significant change in the ex-of this protein at S/T-Q sites. To test this prediction, wepression of two control proteins, ATM and PLC-�1. Qual-transfected U2OS cells with hSMG-1 or ATM siRNA,itatively similar results were obtained with hSMG-1followed by the FLAG-Upf1 expression plasmid. Surpris-siRNA-treated cells (data not shown).ingly, depletion of either hSMG-1 or ATM increased theThe accumulation of hSMG-1-deficient cells in G2/Mbasal phosphorylation of hUpf1 at S/T-Q motifs andphase under basal culture conditions was consistentblunted the response to IR exposure (Figure 6B). In con-with the activation of a DNA damage-induced G2 check-trast, both basal and IR-inducible phosphorylation ofpoint. To address this possibility, we determinedhUpf1 was markedly suppressed in hSMG-1/ATM dou-whether treatment with caffeine, an inhibitor of the G2bly depleted cells. Taken together, these results suggestcheckpoint (Powell et al., 1995; Yao et al., 1996), antago-that loss of either hSMG-1 or ATM triggers a stressnized the increase in G2 and M phase cells observedresponse that leads to increased basal phosphorylationafter AS treatment. Caffeine treatment completely re-by the remaining PIKK family member and that elimina-versed the effect of AS treatment on the ratio of G2/Mtion of both kinases severely impairs hUpf1 phosphory-to G1 phase cells but had no effect on the cell cyclelation in both unstressed and stressed cells. Once again,distribution of the S oligonucleotide-transfected cellOA caused dramatic increases in the phosphorylation ofpopulation (Figure 4B). Interestingly, exposure of theS/T-Q sites in hUpf1, even in the hSMG-1/ATM double-hSMG-1-depleted cells to caffeine also increased theknockdown cells (Figure 6B). The latter result empha-percentage of hypodiploid cells, which suggests that G2
sizes the importance of protein phosphatases in thecheckpoint activation suppresses apoptotic cell death,regulation of the hUpf1 phosphorylation state, even un-possibly by blocking the progression of damaged cellsder conditions of severe depletion of both upstreaminto M phase.hUpf1 kinases.To gain further insights into the role of hSMG-1 in
stress responses, we performed clonogenic survival Finally, we determined whether IR or OA exposure

hSMG-1 and Stress-Induced Signaling591
Figure 4. Effect of hSMG-1 Depletion on Cell Cycle Progression
(A) Cell cycle distribution. U2OS cells were either nontransfected or transfected with sense (S) or antisense (AS) oligonucleotide targetedagainst hSMG-1. After 40 hr, cells were exposed to 20 Gy IR. Cell cycle distributions were examined after 16 hr by flow cytometry. The tableshows the percentage of cells in each cell cycle phase. The right panel shows immunoblot analyses from the same cell populations.(B) Effect of caffeine. U2OS cells were transfected with S or AS oligonucleotide as described above. The cells were treated for 8 hr with 2mM caffeine and were then harvested for cell cycle distribution analyses. The table shows percentages of cells in each cell cycle phase, plusthe ratio of G2/M to G1 cells for each sample. The right panel depicts immunoblotting results from the same cell populations.

Molecular Cell592
with �-phospho-S/T-Q antibody (Figure 6C, left panel).To confirm these results, we performed the converseexperiment in which cell extracts were immunoprecipi-tated with the �-phospho-S/T-Q antibody, followed by�-hUpf1. IR exposure increased the amounts of bothhUpf1 and the positive control protein NBS1 in theseimmunoprecipitates, indicating that both proteins werephosphorylated by hSMG-1 and/or ATM in response toIR exposure (Figure 6C, right panel).
Effect of Genotoxic Stress on NMDTo examine directly the requirement for hSMG-1 in NMD,we depleted cells of this protein kinase with AS oligonu-cleotide (see Figure 4) and determined NMD efficiencywith an established reporter assay (Ishigaki et al., 2001).
Figure 5. Radiation Sensitivity of hSMG-1-Depleted Cells The S or AS oligonucleotide-transfected cells were co-A549 cells were transfected with luciferase or hSMG-1 siRNA and transfected with either the normal human �-globin genereplated after 24 hr for clonogenic survival assays. After 48 hr, the (Norm) or a premature termination codon (PTC)-bearingcells were exposed to the indicated doses of IR, and surviving colo-
�-globin gene (Ter), plus a reference plasmid encodingnies counted after 12 days in culture. The immunoblot showsmouse urinary protein (MUP) to control for variations inhSMG-1 levels in the test cell populations at 72 hr posttransfection.transfection efficiency and RNA recovery. Transfectionof the cells with AS but not S suppressed NMD of theTer-encoded mRNA, indicating that hSMG-1 expressiontriggered the phosphorylation of endogenous hUpf1 in
nontransfected U2OS cells. Exposure of the cells to 10 is required for optimal NMD pathway function in non-damaged cells (Figure 7A).Gy IR or OA triggered a time-dependent increase in
hUpf1 phosphorylation as detected by immunoblotting In the final set of experiments, we investigated the
Figure 6. IR-Induced hUpf1 Phosphorylation
(A) Effect of IR or OA. U2OS cells were transfected with a FLAG-hUpf1, and, after 24 hr, cells were serum starved for 16 hr. Cells were thentreated with 250 nM OA and/or 10 Gy IR, and cellular extracts were immunoprecipitated with �-FLAG-agarose beads. Immunoprecipitatedproteins were separated by SDS-PAGE and sequentially probed with �-pS/T-Q antibody, followed by �-FLAG antibody. Immunoblots wereanalyzed by densitometry, and �-pS/T-Q immunoreactivity was normalized to the level of �-FLAG immunoreactivity for the same sample. Thesignal ratio was obtained by normalizing these values to the nonirradiated control.(B) Effect of hSMG-1 versus ATM depletion. U2OS cells were transfected with the indicated siRNA, and after 72 hr, the cells were exposedto IR or OA. The cells were lysed after 1 or 4 hr, and immunoblotting was done with the indicated antibodies. Signal ratios were calculatedas described in (A).(C) Phosphorylation of endogenous hUpf1. U2OS cells were treated for the indicated times with IR or OA, and �-hUpf1 immunoprecipitateswere sequentially immunoblotted with �-pS/T-Q and �-hUpf1 antibodies (left panel). Cell extracts were also immunoprecipitated with �-S/T-Q antibodies and immunoblotted with either �-hUpf1 or �-NBS1 antibodies (right panel).

hSMG-1 and Stress-Induced Signaling593
potential role of ATM, as well as the impact of genotoxic Anderson, 1993; Yamashita et al., 2001). Furthermore, wereport that hSMG-1 functions in genome surveillancestress on NMD. For technical reasons, we turned to aand cell viability maintenance in mammalian cells. Wedifferent assay, which measures the steady-state levelpropose that hSMG-1 teams with the related PIKK familyof an endogenous PTC-bearing mRNA in the Calu6 lungmembers ATM and ATR to form a triad of S/T-Q-directedcarcinoma line. These cells contain mutant TP53 alleleskinases that serve as proximal signal transducers in DNAthat express a PTC at codon 196, thereby rendering thedamage recognition and stress response pathways.resulting p53 (1–195) transcripts candidates for NMDBased on the finding that hUpf1 is phosphorylated by(Yamashita et al. 2001). We treated Calu6 cells withboth ATM and hSMG-1 in cells exposed to genotoxicsiRNAs directed against hSMG-1 or ATM or with bothstress, our results suggest that hUpf1 is also integratedsiRNAs and measured the steady-state levels of theinto the stress response network activated by DNA-p53 (1–195) transcripts by quantitative real-time PCRdamaging agents.(Q-PCR). Depletion of hSMG-1 leads to a 2-fold increase
The notion that genotoxic stress-induced signals im-in p53 (1–195) mRNA levels and to the appearance ofpinge on both the DNA damage response and NMDdetectable amounts of the truncated p53 (1–195) proteinmachinery is consistent with the fact that most geno-(Figure 7B, left panel). Depletion of ATM alone had notoxic agents, including IR and UV light, actually triggereffect on the level of the p53 (1–195) mRNA, and com-indiscriminate macromolecular damage in eukaryoticbined hSMG-1/ATM deficiency caused no greater stabi-cells (Herrlich et al., 1992, 1997). Recovery from geno-lization of this transcript than did hSMG-1 depletiontoxic stress therefore hinges not only on DNA repairalone. These results were consistent with findings thatbut also on the repair and/or replacement of damagedATM-deficient cells derived from ataxia telangiectasiaproteins, lipids, and other macromolecules, typicallypatients showed no impairment of NMD in the �-globinthrough induced gene transcription and mRNA transla-reporter assays described above (our unpublishedtion. Stress responses are also triggered by strong mito-data). We conclude from these studies that hSMG-1 butgenic stimuli, and it is noteworthy in this regard thatnot ATM is required for optimal functioning of the NMDgrowth factor stimulation modulates the overall phos-pathway under standard cell growth conditions. A strik-phorylation state of hUpf1 (Pal et al., 2001). Althoughing finding was that treatment of the cells with 5 �Mthe role of specific phosphorylation events in the regula-wortmannin, a drug concentration sufficient to causetion of hUpf1 function is only partially understood, it is�80% reduction in hSMG-1 kinase activity (Supplemen-tempting to speculate that the NMD pathway hastal Figure S3 on Molecular Cell’s website), elicited aevolved to respond to various environmental stress,higher level of p53 (1–195) mRNA and protein accumula-ranging from mitogenic stimuli, which activate explosivetion than did the hSMG-1 knockdown with siRNA. Theseincreases in gene transcription and translation, to geno-results suggest that the effect of wortmannin on NMDtoxic agents, which damage macromolecules and in-is not solely attributable to inhibition of hSMG-1 andcrease the risk that transcription yields abnormalthat other wortmannin-sensitive kinases participate inmRNAs.NMD pathway function in mammalian cells.
An important conclusion from this study is thatTo determine whether genotoxic stress affects NMD,hSMG-1 is a stress-responsive protein kinase. Cellularwe treated Calu6 cells with hSMG-1 and/or ATM siRNAsexposure to IR or UV light stimulates hSMG-1 kinaseand examined the steady-state level of p53 (1–195)activity, as measured in immune complex kinase assays.mRNA in cells exposed to 0 or 10 Gy IR (Figure 7C).The mechanism of hSMG-1 activation by radiation isAs reported above, treatment of the cells with hSMG-1unknown. However, an intriguing feature of this proteinsiRNA resulted in a �2.5-fold increase in p53 (1–195)is the large insert region (1000 amino acids) betweentranscript levels, whereas ATM siRNA treatment had nothe catalytic domain and the conserved FAT-C domain.effect. IR exposure consistently enhanced the expres-The hSMG-1 insert region was first identified as an inde-
sion of p53 (1–195) mRNA (to 3.7-fold over control) inpendent ORF in a yeast two-hybrid screen for protein
hSMG-1-depleted cells. The negative effect of IR onkinase C-/-interacting proteins (Diaz-Meco et al.,
NMD of the p53 (1–195) transcript was abrogated when 1996). However, current evidence argues that this puta-the hSMG-1-deficient cells were simultaneously de- tive /-interacting protein (LIP) is neither an alternativepleted of ATM. Qualitatively similar changes in p53 transcript nor a translation product of the hSMG-1 locus(1–195) expression were obtained by immunoblotting (our unpublished data; Denning et al., 2001). Given thatfor this protein in extracts from the same cell populations the insert region is not present in CeSMG-1, we hypothe-(Figure 7C, left panel). Thus, loss of hSMG-1 unmasks size that this region represents an evolutionary adapta-an ATM-dependent inhibitory effect of IR on NMD in tion that allows hSMG-1 to respond to a broadenedhuman cells. range of regulatory signals, including those triggered by
radiation-induced stress, in higher metazoans.Discussion The SMG-2/Upf1 proteins may also have undergone
evolutionary modifications that altered the regulationThe results of this study identify hSMG-1 as a stress- and function of this protein in different organisms. Theresponsive kinase that maintains the overall integrity of budding yeast homolog ScUpf1 lacks a carboxy-termi-gene expression in mammalian cells. Our results rein- nal S/T-Q-rich domain, consistent with the absence offorce earlier conclusions that hSMG-1 is a functional ho- a SMG-1 homolog in this single-celled organism (Cul-molog of the C. elegans SMG-1 protein, which was geneti- bertson, 1999). The worm homolog CeSMG-2 containscally defined as an essential participant in the NMD a S/T-Q-rich domain with 12 potential phosphorylation
sites for CeSMG-1. However, the sequences sur-pathway in the worm (Denning et al., 2001; Pulak and
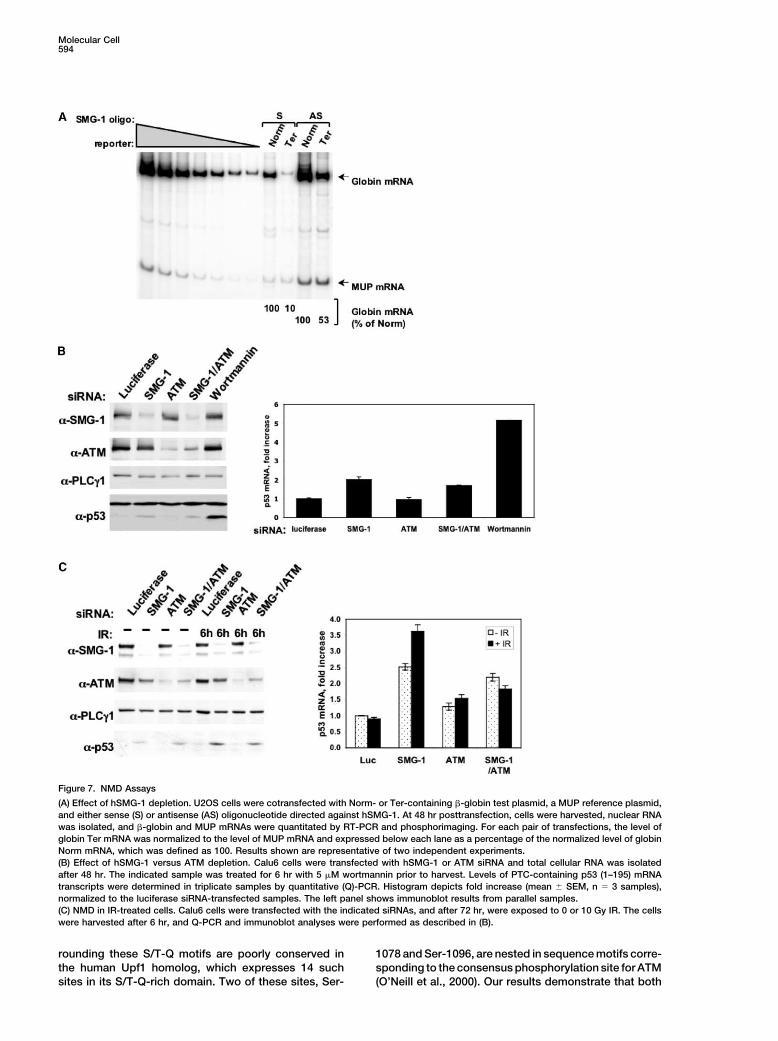
Molecular Cell594
Figure 7. NMD Assays
(A) Effect of hSMG-1 depletion. U2OS cells were cotransfected with Norm- or Ter-containing �-globin test plasmid, a MUP reference plasmid,and either sense (S) or antisense (AS) oligonucleotide directed against hSMG-1. At 48 hr posttransfection, cells were harvested, nuclear RNAwas isolated, and �-globin and MUP mRNAs were quantitated by RT-PCR and phosphorimaging. For each pair of transfections, the level ofglobin Ter mRNA was normalized to the level of MUP mRNA and expressed below each lane as a percentage of the normalized level of globinNorm mRNA, which was defined as 100. Results shown are representative of two independent experiments.(B) Effect of hSMG-1 versus ATM depletion. Calu6 cells were transfected with hSMG-1 or ATM siRNA and total cellular RNA was isolatedafter 48 hr. The indicated sample was treated for 6 hr with 5 �M wortmannin prior to harvest. Levels of PTC-containing p53 (1–195) mRNAtranscripts were determined in triplicate samples by quantitative (Q)-PCR. Histogram depicts fold increase (mean � SEM, n � 3 samples),normalized to the luciferase siRNA-transfected samples. The left panel shows immunoblot results from parallel samples.(C) NMD in IR-treated cells. Calu6 cells were transfected with the indicated siRNAs, and after 72 hr, were exposed to 0 or 10 Gy IR. The cellswere harvested after 6 hr, and Q-PCR and immunoblot analyses were performed as described in (B).
rounding these S/T-Q motifs are poorly conserved in 1078 and Ser-1096, are nested in sequence motifs corre-sponding to the consensus phosphorylation site for ATMthe human Upf1 homolog, which expresses 14 such
sites in its S/T-Q-rich domain. Two of these sites, Ser- (O’Neill et al., 2000). Our results demonstrate that both

hSMG-1 and Stress-Induced Signaling595
hSMG-1 and ATM phosphorylate the S/T-Q-rich domain phosphorylation in response to IR treatment (see Figure2B). This observation hinted that loss of hSMG-1 func-of hUpf1 in immune complex kinase assays. Interest-
ingly, the hUpf1 protein undergoes rapid nucleocy- tion triggers a spontaneous DNA damage response, aconclusion that was reinforced by the accumulation oftoplasmic shuttling (Mendell et al., 2002), which would
allow both hSMG-1 and ATM access to the S/T-Q-rich �H2AX-positive nuclear foci in the hSMG-1-deficientcells. The conversion of histone H2AX to �H2AX is de-domain of hUpf1 in intact cells. Yamashita et al. (2001)
performed a limited mutational analysis of the hUpf1 pendent on the phosphorylation of Ser139 by ATM orATR (Burma et al., 2001; Ward and Chen, 2001). Ourcarboxyl terminus and showed that Ala substitutions
at four S-Q motifs, including Ser-1078 and Ser-1096, observation that hSMG-1-depleted cells show defectivehistone H2AX phosphorylation after IR exposure indi-interfere with the functions of hUpf1 in NMD. Thus, phos-
phorylation of the S/T-Q-rich domain by hSMG-1 and/ cates that hSMG-1 is directly or indirectly involved in themodification of this protein in response to IR-inducedor ATM modulates the function of hUpf1 in RNA surveil-
lance. Recent studies provided evidence for rapid turn- stress. Studies are underway to more precisely deter-mine the contribution of hSMG-1 to histone H2AX phos-over of phosphate on both the worm and mammalian
SMG-2/Upf1 proteins and implicated a SMG-5/SMG-7- phorylation induced by different types of genotoxicagents.linked protein PP2A-like activity in the dephosphoryla-
tion of this protein (Anders et al., 2003; Chiu et al., 2003; In summary, our results indicate that hSMG-1 plays acontinuous role in genome maintenance, as well as NMD,Ohnishi et al., 2003). The present studies demonstrate
that full-length Upf1 accumulates phosphate at S/T-Q in cycling human cells. Like ATM, hSMG-1 is activated inIR-treated cells, and hUpf1 is a common target for bothsites in cells treated with PP2A-inhibitory concentra-
tions of OA (see Figure 6), indicating that the hUpf1- of these kinases in cells responding to IR-inducedstress. With its multiple S/T-Q sites, the carboxyl termi-linked protein phosphatase activity functions, at least
in part, to reverse the regulatory modifications imposed nus of hUpf1 is poised to receive both positive andnegative regulatory signals for NMD from hSMG-1 andby hSMG-1 and/or ATM.
A complete understanding of the roles of phosphory- ATM. We also cannot exclude the possibility that, inaddition to its primordial function in NMD, hUpf1 partici-lation in the regulation of hUpf1 function will require
detailed mapping of the S/T-Q phosphorylation sites pates more directly in the DNA damage response inmammalian cells. As appears to be the case for BRCA1after cellular exposure to IR and other stimuli. Our results
with Calu6 cells show that IR treatment increases the (Xu et al., 2002), differential modifications of specificS/T-Q residues by hSMG-1 versus ATM may directsuppression of NMD induced by loss of hSMG-1 (Figure
7C). We hypothesize that activation of ATM by IR (in the hUpf1 functions into different cellular response path-ways. The sharing of key upstream signaling elements,absence of hSMG-1) leads to phosphorylation of the
S/T-Q-rich domain at sites that inhibit the function of such as ATM, hSMG-1, and hUpf1, may allow the DNAdamage-response network to insure the overall qualityhUpf1 in NMD. This model is consistent with our finding
that siRNA-mediated depletion of hSMG-1 leads to of the transcriptome during cellular recovery fromstress-induced damage to DNA and other intracellularATM-dependent phosphorylation of hUpf1 (Figure 6B)
and concomitant suppression of NMD (Figure 7C). In macromolecules.human cells, the SMG-5/SMG-7-linked PP2A (Anders etal., 2003; Ohnishi et al., 2003) may act to reverse the Experimental Proceduresinhibition of hUpf1 function caused by phosphorylationat specific sites in the S/T-Q-rich domain. Cell Lines
U2OS osteosarcoma, Calu6, human embryonic kidney (HEK) 293,In addition to hUpf1, we identified p53 as a physiologi-and HEK 293T cells were cultured in low-glucose Dulbecco’s modi-cal target for hSMG-1 in intact cells. Based on thesefied Eagle’s medium (DMEM) supplemented with 10% fetal bovineresults, we conclude that the p53 (Ser-15) site repre-serum (FBS). A549 cells were cultured in DMEM/F12 supplemented
sents a convergent target for modification by three with 10% FBS. HeLa cells were cultured in high-glucose DMEMstress-responsive PIKKs—ATM, ATR, and hSMG-1. supplemented with 10% FBS. Where indicated, cells were �-irradi-However, we cannot rule out the possibility that other ated with a 137Cs source or UV irradiated with a UV-B source (max,
305 nm).targets in the p53 pathway play central roles in thehSMG-1-dependent stabilization of p53 in IR- or UV-damaged cells. For example, the hMDM2 protein, which Antibodiesfunctions as a suppressor of p53 expression, is a physio- hSMG-1-specific antibodies were raised by immunizing rabbits (Co-
calico Biologicals, Inc.) with bacterially expressed and purified GSTlogically important substrate for ATM in IR-treated cellsfusion proteins containing hSMG-1 amino acids 2281–2339 (Ab-1)(Maya et al., 2001). Future experiments will examine theor amino acids 1691–1790 (Ab-2). The �-SMG-1 (Ab-2) antiserumpossibility that hSMG-1-mediated hMDM2 phosphoryla-was affinity-purified over Affi-Gel 15 (Bio-Rad) coupled to the GST-
tion contributes to IR- or UV-induced p53 accumulation hSMG-11691–1790 fusion protein. The �-hUpf1 antibodies were pre-in human cells. pared by immunizing rabbits with a GST fusion protein containing
While our results identify p53 as a common target amino acids 557–612 of hUpf1. PLC-�1-specific antibodies havebeen described (Secrist et al., 1993). The �-ATM (Ab-3), �-ATRfor ATM, ATR, and hSMG-1, the checkpoint signaling(Ab-1), �-phospho-p53 (Ser-15), and �-p53 (Ab-6) reagents weredefects observed in hSMG-1-depleted cells do not over-obtained from Oncogene Science Research Products. Additionallap completely with those observed in cells lacking ATMantibodies were obtained from (sources in parentheses): �-HA (clone
or ATR. For example, hSMG-1-deficient cells display 12CA5; Covance Research Products), �-FLAG-M2 and �-tubulinconstitutive phosphorylation of hChk2 under normal cell (Sigma), �-Chk2 and �-�H2AX (Upstate Biotechnology), polyclonal
ATM/ATR substrate antibody (Cell Signaling Technology).culture conditions and a further enhancement of hChk2

Molecular Cell596
Isolation of hSMG-1 cDNA vitrogen). The sequence for the hSMG-1 siRNA (5�-CCAGGACACGAGGAAACUG-3�) corresponded to nucleotides 1753–1770 in theThe full-length hSMG-1 cDNA was isolated as a series of overlapping
fragments obtained by screening of a human cerebellum cDNA li- hSMG-1 cDNA. Cells were typically used in experiments after 48–72hr of siRNA treatment.brary (Clontech #HL1128a) and a Jurkat T cell cDNA library (Stra-
tagene #936219) and by 5� RACE and RT-PCR of human brain andJurkat cDNA. Additional details regarding the hSMG-1 cloning Clonogenic Assaysprocedure are available upon request. The hSMG-1 cDNA was ap- A549 cells were transfected with luciferase or hSMG-1 siRNA, andpended with an amino-terminal hemagglutinin (HA) epitope tag se- then were replated at predetermined cell numbers per 60 mm dishquence (CYPYDVPDYASL) and was cloned into the XhoI and NotI after 24 hr. After 48 hr, the cells were exposed to the indicatedsites of pcDNA3.1 (Invitrogen) to create the HA-SMG-1wt expression doses of IR, and 12 day clonogenic survival assays were performedplasmid. The kinase inactive hSMG-1 mutant (HA-SMG-1KI) contains as described (Sarkaria et al., 1998).an Asp→Ala substitution at residue 2195, generated by site-directedmutagenesis with the QuickChange kit (Stratagene). All plasmid con- Immunofluorescence Microscopystructs were sequenced to insure the fidelity of the PCR and clon- For immunofluorescence microscopy of endogenous hSMG-1 anding procedures. �H2AX, U2OS cells were plated in 4-well covered chamber slides
(Nalge Nunc). After the indicated treatments, the cells were fixedfor 15 min at room temperature in 2% paraformaldehyde. After wash-
Immunoprecipitation and Immunoblotting ing with phosphate-buffered saline (PBS) and quenching with 0.1 MFor immunoprecipitations, cell extracts were prepared by resus- glycine, the cells were permeabilized for 5 min at room temperaturepending washed cell pellets in lysis buffer (50 mM Tris-HCl [pH with 0.2% Triton X-100 containing 1% normal goat serum (NGS).
The permeabilized cells were blocked with PBS plus 1% NGS and7.4], 150 mM NaCl, 1% NP-40, 1 mM dithiothreitol) plus proteasethen stained with 3 �g per ml affinity-purified Ab-2 or with 2 �g perinhibitors. For standard immune complex kinase assays, the lysisml �H2AX-specific monoclonal antibody (Upstate). After 2 to 3 hrbuffer was modified to contain 1% Tween-20 detergent in placeat room temperature, the sample wells were washed with PBS plusof NP-40 and additional phosphatase inhibitors (20 mM �-glycerol1% NGS. The samples were then incubated with either Alexa Fluorphosphate and 50 nM microcystin). Samples were incubated on ice488-conjugated goat anti-rabbit immunoglobulin G (IgG) or Alexafor 15 min and then clarified by centrifugation. HA-tagged recombi-Fluor 594-conjugated goat anti-mouse IgG in the presence of 4 �gnant proteins were immunoprecipitated from cell extracts with 4 �gper ml 4�,6�-diamidino-2-phenylindole (DAPI). The slides wereof �-HA antibody. Endogenous hSMG-1 protein was immunoprecipi-mounted with Mowiol (Hoechst). Immunofluorescence microscopytated with 8 �g of Ab-2. After separation by SDS-PAGE, the proteinswas performed with a Leica DMIRE2 inverted automated digitalwere detected by autoradiography or immunoblotting. Antibody-imaging microscope. Image deconvolution was accomplished withbound proteins were immunoblotted with protein A-horseradishthe nearest-neighbor algorithm of the Simple PCI software (Compix,peroxidase (HRP) (Amersham) for rabbit antibodies or sheep anti-Inc.). Imaging of �-�H2AX-stained cells was also performed with anmouse IgG-HRP (Amersham) to detect mouse monoclonal antibod-Eidac 100� high-throughput microscopy system and the Cyto-ies. Immunoreactive proteins were illuminated with the Renaissanceshop� cell analysis suite (Q3DM, San Diego, CA).chemiluminescence system (NEN).
RNA Isolation and NMD AssaysTotal cellular or nuclear RNA was isolated using Trizol (Invitrogen)Immune Complex Kinase Assaysor the NE-PER kit (Pierce), respectively. The extent of NMD wasNative or recombinant hSMG-1 proteins were immunoprecipitateddetermined by RT-PCR quantitation of the levels of globin and MUPfrom cell extracts as described above, and the immunoprecipitatesmRNA as described previously (Ishigaki et al., 2001). In Calu6 cellwere washed twice in lysis buffer, once in high-salt buffer (100 mMexperiments, total cellular RNA was isolated with the RNeasy kitTris-HCl [pH 7.4], 500 mM LiCl) and once in kinase buffer (10 mM(Qiagen) and treated with DNase. First-strand cDNA was preparedHEPES [pH 7.4], 50 mM NaCl, 50 mM �-glycerophosphate). Kinasewith Superscript II (Stratagene) using a oligo (dT)12–18 primer. Q-PCRreactions were performed for 30 min at 30 C after adding 40 �lwas performed in triplicate with the brilliant SYBR green Q-PCRkinase buffer containing 10% glycerol, 1 mM DTT, 10 mM MnCl2,master mix (Stratagene). Two different p53-derived primer pairs20 nM microcystin, protease inhibitors, 1 �g of the indicated sub-were used in each assay, with qualitatively similar results. Q-PCRstrate, 10 �M ATP, and 10 �Ci [�-32P]ATP (6000 Ci/mmole) (NEN) towas performed with a Mx3000P system (Stratagene), and mutanteach sample.p53 mRNA transcript levels were normalized to levels of cyclophilinmRNA in each sample.
Cell TransfectionsAcknowledgmentsTo prepare recombinant HA-tagged hSMG-1 and ATM proteins,
9 � 105 HEK 293T cells were plated onto 60 mm dishes and wereThe authors thank Craig Hauser for assistance with the RT-PCRtransfected with 5 �g of the indicated HA-SMG-1 plasmid DNA usinganalyses. This work was supported by grants from the Nationalthe Fugene 6 transfection reagent (Roche). For NMD assays, U2OSInstitutes of Health (CA97950 to R.T.A.; DK33938 and GM59614cells were seeded onto 100 mm dishes (1 � 106 cells per dish). Afterto L.E.M.) and by grants to R.T.A. from the Ataxia-Telangiectasia20 hr, cells were transfected with 1.5 �g of pmCMV-Gl test plasmid,Children’s Project, Johnson & Johnson, and the Department of De-either Norm or 39Ter, and 1.5 �g of phCMV-MUP reference plasmidfense (DAMD17-02-1-0730). K.M.B. was supported by a postdoc-(Ishigaki et al., 2001).toral fellowship from the National Cancer Institute (CA83319).For S/AS oligonucleotide transfections, 1 � 105 U2OS cells were
plated in 60 mm dishes and were transfected after 30 hr with theReceived: August 8, 2003phosphorothioate oligonucleotide (Genset Oligos, La Jolla, CA). TheRevised: April 13, 2004S oligonucleotide spans hSMG-1 nucleotides 210–229 (5�-GAGAAccepted: April 16, 2004CAGGAGGGAGCTTGCT-3�), and the AS oligonucleotide is comple-Published: June 3, 2004mentary to this sequence. The cells were transfected with 8 �g/ml
oligonucleotide at a Fugene 6 to DNA ratio of 1.6:1. Forty hoursReferencesafter transfection, dishes were exposed to IR or UV-B light and then
harvested for immunoblotting, cell cycle distribution, or cell survivalAbraham, R.T. (2001). Cell cycle checkpoint signaling through theassays, as indicated in Results.ATM and ATR kinases. Genes Dev. 15, 2177–2196.For siRNA transfection experiments, U2OS cells were plated in
60 mm dishes (1 � 105 per dish) in complete medium. After 24 hr, Anders, K.R., Grimson, A., and Anderson, P. (2003). SMG-5, requiredcells were transfected with the indicated siRNA duplexes (Dhar- for C. elegans nonsense-mediated mRNA decay, associates with
SMG-2 and protein phosphatase 2A. EMBO J. 22, 641–650.macon) at a final concentration of 67 nM using Oligofectamine (In-

hSMG-1 and Stress-Induced Signaling597
Banin, S., Moyal, L., Shieh, S., Taya, Y., Anderson, C.W., Chessa, rent1/hUpf1 in altered splicing and decay of nonsense transcripts.Science 298, 419–422.L., Smorodinsky, N.I., Prives, C., Reiss, Y., Shiloh, Y., et al. (1998).
Enhanced phosphorylation of p53 by ATM in response to DNA dam- North, S., and Hainaut, P. (2000). p53 and cell-cycle control: a fingerage. Science 281, 1674–1677. in every pie. Pathol. Biol. 48, 255–270.Bartek, J., and Lukas, J. (2003). Chk1 and Chk2 kinases in check-
Ohnishi, T., Yamashita, A., Kashima, I., Schell, T., Anders, K.R.,point control and cancer. Cancer Cell 3, 421–429.Grimson, A., Hachiya, T., Hentze, M.W., Anderson, P., and Ohno,
Burma, S., Chen, B.P., Murphy, M., Kurimasa, A., and Chen, D.J. S. (2003). Phosphorylation of hUPF1 induces formation of mRNA(2001). ATM phosphorylates histone H2AX in response to DNA dou- surveillance complexes containing hSMG-5 and hSMG-7. Mol. Cellble-strand breaks. J. Biol. Chem. 276, 42462–42467. 12, 1187–1200.Cali, B.M., and Anderson, P. (1998). mRNA surveillance mitigates
O’Neill, T., Dwyer, A.J., Ziv, Y., Chan, D.W., Lees-Miller, S.P., Abra-genetic dominance in Caenorhabditis elegans. Mol. Gen. Genet.ham, R.H., Lai, J.H., Hill, D., Shiloh, Y., Cantley, L.C., et al. (2000).260, 176–184.Utilization of oriented peptide libraries to identify substrate motifs
Canman, C.E., Lim, D.S., Cimprich, K.A., Taya, Y., Tamai, K., Saka- selected by ATM. J. Biol. Chem. 275, 22719–22727.guchi, K., Appella, E., Kastan, M.B., and Siliciano, J.D. (1998). Activa-
Page, M.F., Carr, B., Anders, K.R., Grimson, A., and Anderson, P.tion of the ATM kinase by ionizing radiation and phosphorylation of(1999). SMG-2 is a phosphorylated protein required for mRNA sur-p53. Science 281, 1677–1679.veillance in Caenorhabditis elegans and related to Upf1p of yeast.
Chiu, S.Y., Serin, G., Ohara, O., and Maquat, L.E. (2003). Character-Mol. Cell. Biol. 19, 5943–5951.
ization of human Smg5/7a: a protein with similarities to Caenorhab-Pal, M., Ishigaki, Y., Nagy, E., and Maquat, L.E. (2001). Evidenceditis elegans SMG5 and SMG7 that functions in the dephosphoryla-that phosphorylation of human Upfl protein varies with intracellulartion of Upf1. RNA 9, 77–87.location and is mediated by a wortmannin-sensitive and rapamycin-Cliby, W.A., Roberts, C.J., Cimprich, K.A., Stringer, C.M., Lamb, J.R.,sensitive PI 3-kinase-related kinase signaling pathway. RNA 7, 5–15.Schreiber, S.L., and Friend, S.H. (1998). Overexpression of a kinase-
inactive ATR protein causes sensitivity to DNA-damaging agents Powell, S.N., DeFrank, J.S., Connell, P., Eogan, M., Preffer, F.,and defects in cell cycle checkpoints. EMBO J. 17, 159–169. Dombkowski, D., Tang, W., and Friend, S. (1995). Differential sensi-
tivity of p53(�) and p53(�) cells to caffeine-induced radiosensitiza-Culbertson, M.R. (1999). RNA surveillance. Unforeseen conse-tion and override of G2 delay. Cancer Res. 55, 1643–1648.quences for gene expression, inherited genetic disorders and can-
cer. Trends Genet. 15, 74–80. Pulak, R., and Anderson, P. (1993). mRNA surveillance by the Caeno-Denning, G., Jamieson, L., Maquat, L.E., Thompson, E.A., and Fields, rhabditis elegans smg genes. Genes Dev. 7, 1885–1897.A.P. (2001). Cloning of a novel phosphatidylinositol kinase-related
Rogakou, E.P., Pilch, D.R., Orr, A.H., Ivanova, V.S., and Bonner,kinase: characterization of the human SMG-1 RNA surveillance pro-W.M. (1998). DNA double-stranded breaks induce histone H2AXtein. J. Biol. Chem. 276, 22709–22714.phosphorylation on serine 139. J. Biol. Chem. 273, 5858–5868.
Diaz-Meco, M.T., Municio, M.M., Sanchez, P., Lozano, J., and Mos-Rogakou, E.P., Boon, C., Redon, C., and Bonner, W.M. (1999). Mega-cat, J. (1996). Lambda-interacting protein, a novel protein that spe-base chromatin domains involved in DNA double-strand breakscifically interacts with the zinc finger domain of the atypical proteinin vivo. J. Cell Biol. 146, 905–916.kinase C isotype lambda/iota and stimulates its kinase activity
in vitro and in vivo. Mol. Cell. Biol. 16, 105–114. Sarkaria, J.N., Tibbetts, R.S., Busby, E.C., Kennedy, A.P., Hill, D.E.,and Abraham, R.T. (1998). Inhibition of phosphoinositide 3-kinaseFrischmeyer, P.A., and Dietz, H.C. (1999). Nonsense-mediated
mRNA decay in health and disease. Hum. Mol. Genet. 8, 1893–1900. related kinases by the radiosensitizing agent wortmannin. CancerRes. 58, 4375–4382.Herrlich, P., Ponta, H., and Rahmsdorf, H.J. (1992). DNA damage-
induced gene expression: signal transduction and relation to growth Secrist, J.P., Burns, L.A., Karnitz, L., Koretzky, G.A., and Abraham,factor signaling. Rev. Physiol. Biochem. Pharmacol. 119, 187–223. R.T. (1993). Stimulatory effects of the protein tyrosine phosphatase
inhibitor, pervanadate, on T-cell activation events. J. Biol. Chem.Herrlich, P., Blattner, C., Knebel, A., Bender, K., and Rahmsdorf,268, 5886–5893.H.J. (1997). Nuclear and non-nuclear targets of genotoxic agents in
the induction of gene expression. Shared principles in yeast, ro- Shiloh, Y. (2003). ATM and related protein kinases: safeguardingdents, man and plants. Biol. Chem. 378, 1217–1229. genome integrity. Nat. Rev. Cancer 3, 155–168.Hodgkin, J., Papp, A., Pulak, R., Ambros, V., and Anderson, P. (1989).
Sun, X., Perlick, H.A., Dietz, H.C., and Maquat, L.E. (1998). A mutatedA new kind of informational suppression in the nematode Caeno-human homologue to yeast Upf1 protein has a dominant-negativerhabditis elegans. Genetics 123, 301–313.effect on the decay of nonsense-containing mRNAs in mammalian
Hunter, T. (1995). When is a lipid kinase not a lipid kinase? When it cells. Proc. Natl. Acad. Sci. USA 95, 10009–10014.is a protein kinase. Cell 83, 1–4.
Tibbetts, R.S., and Abraham, R.T. (2000). PI3K-related kinases—Ishigaki, Y., Li, X.J., Serin, G., and Maquat, L.E. (2001). Evidence for
roles in cell-cycle regulation and DNA damage responses. In Signal-a pioneer round of mRNA translation: mRNAs subject to nonsense-
ing Networks and Cell Cycle Control: The Molecular Basis of Cancermediated decay in mammalian cells are bound by CBP80 and
and Other Diseases, S.J. Gutkind, ed. (Bethesda, MD: HumanaCBP20. Cell 106, 607–617.
Press), pp. 267–301.Lukas, C., Falck, J., Bartkova, J., Bartek, J., and Lukas, J. (2003).
Tibbetts, R.S., Brumbaugh, K.M., Williams, J.M., Sarkaria, J.N.,Distinct spatiotemporal dynamics of mammalian checkpoint regula-Cliby, W.A., Shieh, S.Y., Taya, Y., Prives, C., and Abraham, R.T.tors induced by DNA damage. Nat. Cell Biol. 5, 255–260.(1999). A role for ATR in the DNA damage-induced phosphorylation
Maquat, L.E. (2003). Nonsense-mediated mRNA decay in mamma- of p53. Genes Dev. 13, 152–157.lian cells: from pre-mRNA splicing to mRNA translation and degrada-
Waldman, T., Kinzler, K.W., and Vogelstein, B. (1995). p21 is neces-tion. In Translation Mechanisms, J. Lapointe and L. Brakier-Gingras,sary for the p53-mediated G1 arrest in human cancer cells. Cancereds. (Austin, TX: Landes Biosciences), pp. 213–222.Res. 55, 5187–5190.Maquat, L.E., and Carmichael, G.G. (2001). Quality control of mRNA
function. Cell 104, 173–176. Wang, J., Chang, Y.F., Hamilton, J.I., and Wilkinson, M.F. (2002).Nonsense-associated altered splicing: a frame-dependent responseMaya, R., Balass, M., Kim, S.T., Shkedy, D., Leal, J.F.M., Shifman,distinct from nonsense-mediated decay. Mol. Cell 10, 951–957.O., Moas, M., Buschmann, T., Ronai, Z., Shiloh, Y., et al. (2001).
ATM-dependent phosphorylation of Mdm2 on serine 395: role in Ward, I.M., and Chen, J. (2001). Histone H2AX is phosphorylated inp53 activation by DNA damage. Genes Dev. 15, 1067–1077. an ATR-dependent manner in response to replicational stress. J.
Biol. Chem. 276, 47759–47762.Mendell, J.T., Rhys, C., and Dietz, H.C. (2002). Separable roles for

Molecular Cell598
Xu, B., O’Donnell, H., Kim, S.T., and Kastan, M.B. (2002). Phosphory-lation of serine 1387 in brca1 is specifically required for the Atm-mediated S-phase checkpoint after ionizing irradiation. Cancer Res.62, 4588–4591.
Yamashita, A., Ohnishi, T., Kashima, I., Taya, Y., and Ohno, S. (2001).Human SMG-1, a novel phosphatidylinositol 3-kinase-related pro-tein kinase, associates with components of the mRNA surveillancecomplex and is involved in the regulation of nonsense-mediatedmRNA decay. Genes Dev. 15, 2215–2228.
Yao, S.L., Akhtar, A.J., McKenna, K.A., Bedi, G.C., Sidransky, D.,Mabry, M., Ravi, R., Collector, M.I., Jones, R.J., Sharkis, S.J., et al.(1996). Selective radiosensitization of p53-deficient cells by caf-feine-mediated activation of p34cdc2 kinase. Nat. Med. 2, 1140–1143.
Accession Numbers
The GenBank accession number for the hSMG-1 sequence isAF395444.




















