
Modelling tissues in 3D: the next future of pharmaco-toxicology and food research
Upload
independentCategory
view
0download
0
1768 (2007) 1784–1795www.elsevier.com/locate/bbamem
Biochimica et Biophysica Acta
The mitochondrial ryanodine receptor in rat heart: A pharmaco-kinetic profile
Beth A. Altschafl a, Gisela Beutner b, Virendra K. Sharma b,Shey-Shing Sheu b, Héctor H. Valdivia a,⁎
a Department of Physiology, University of Wisconsin Medical School, 601 Science Drive, Madison, WI 53711, USAb Department of Pharmacology and Physiology, Box 711, University of Rochester, School of Medicine and Dentistry,
601 Elmwood Avenue, Rochester, NY 14642, USA
Received 28 November 2006; received in revised form 9 April 2007; accepted 10 April 2007Available online 19 April 2007
Abstract
A protein discovered within inner mitochondrial membranes (IMM), designated as the mitochondrial ryanodine receptor (mRyR), has beenrecognized recently as a modulator of Ca2+ fluxes in mitochondria. The present study provides fundamental pharmacological andelectrophysiological properties of this mRyR. Rat cardiac IMM fused to lipid bilayers revealed the presence of a mitochondrial channel withgating characteristics similar to those of classical sarcoplasmic reticulum RyR (SR-RyR), but a variety of other mitochondrial channels obstructedclean recordings. Mitochondrial vesicles were thus solubilized and subjected to sucrose sedimentation to obtain mRyR-enriched fractions.Reconstitution of sucrose-purified fractions into lipid bilayers yielded Cs+-conducting, Ca2+-sensitive, large conductance (500–800 pS) channelswith signature properties of SR-RyRs. Cytosolic Ca2+ increased the bursting frequency and mean open time of the channel. Micromolarconcentrations of ryanodine induced the appearance of subconductance states or inhibited channel activity altogether, while Imperatoxin A(IpTxa), a specific activator of RyRs, reversibly induced the appearance of distinct subconductance states. Remarkably, the cardiac mRyRdisplayed a Ca2+ dependence of [3H]ryanodine binding curve similar to skeletal RyR (RyR1), not cardiac RyR (RyR2). Overall, the mRyRdisplayed elemental attributes that are present in single channel lipid bilayer recordings of SR-RyRs, although some exquisite differences werealso noted. These results therefore provide the first direct evidence that a unique RyR occurs in mitochondrial membranes.© 2007 Published by Elsevier B.V.
Keywords: Mitochondria; Ryanodine receptor; Planar lipid bilayer; Single channel recording; Imperatoxin A; Calcium
1. Introduction
Extracellular Ca2+ influx through the dihydropyridinereceptor induces the opening of Ca2+ release channels incardiac muscle, giving rise to the Ca2+-induced-Ca2+ release(CICR) mechanism that triggers muscle contraction [1,2]. TheCa2+ release channel that is essential for excitation–contrac-tion (E–C) coupling is a ∼2 million Da homotetramericprotein of the sarcoplasmic reticulum (SR) that bindsryanodine with high specificity and affinity; hence it isnamed the ryanodine receptor (RyR) [3–5]. A variety ofendogenous substances have been described to regulate RyRactivity, including Ca2+, Mg2+, ATP, H+, calmodulin, severalprotein kinases [6,7], and accessory proteins like sorcin and
⁎ Corresponding author. Tel.: +1 608 265 5960; fax: +1 608 265 7821.E-mail address: [email protected] (H.H. Valdivia).
0005-2736/$ - see front matter © 2007 Published by Elsevier B.V.doi:10.1016/j.bbamem.2007.04.011
FKBP12 [8]. Likewise, several exogenous regulators of RyRactivity, although not directly associated with the CICRmechanism, play an important role in defining the pharmaco-logical profile of the RyR [9]. These modulators include:ryanodine [10], caffeine [11], dantrolene [12], and scorpionpeptide toxins [13], among others. Overall, the role of the RyRin E–C coupling, its pharmacological properties, and itsmolecular identity are fairly well characterized.
Recently, studies have shown that an inner mitochondrialmembrane (IMM) protein runs with identical mobility to SR-RyRs in SDS-PAGE, cross-reacts with RyR antibodies, andmediates ryanodine- and dantrolene-sensitive Ca2+ fluxes[14,15]. It is possible, therefore, that a mitochondrial channelprotein with signature features of the RyR (i.e. a mitochon-drial RyR, mRyR) may be an integral component of the Ca2+
transport system of mitochondria. If verified, this wouldundoubtedly be a surprising topological location for a RyR

1785B.A. Altschafl et al. / Biochimica et Biophysica Acta 1768 (2007) 1784–1795
that would prompt the notion that there is heterogeneity ofRyR localization within cardiomyocytes. In this study, wefused purified mitochondrial membranes into planar lipidbilayers and determined the existence of a mitochondrialchannel that behaves remarkably similar to SR-RyRs. Weshow that the unitary channel conductance and openprobability of this presumed mRyR in the presence of someof the most classical modulators of RyR activity, including:Ca2+, ryanodine, and Imperatoxin A (IpTxa, is a 3.7 kDapeptide toxin, isolated from the venom of the scorpion Pan-dinus imperator, with high affinity to and exquisite selectivityfor RyRs) are remarkably similar to those of the SR-RyR.However, a unique identifying feature of this mRyR was itsCa2+-dependence of activity curve, which was similar to thatof skeletal RyR (RyR1), instead of that of cardiac RyR(RyR2), as would be expected from its tissue localization.These results were in concordance with those of Beutner et al.[14,15] and reinforce the notion that a mRyR is an integralcomponent of mitochondrial membranes and not a residualcomponent of SR. Several studies support the idea thatmitochondria sequester significant amounts of mobilizedcytosolic Ca2+ in cardiac muscle cells, thereby shaping cyto-solic Ca2+ transients [16–21], and also controlling cellularenergy production on a beat-by-beat basis [22]. Thus, apotential role of the mRyR may be to modulate Ca2+ fluxes inthe mitochondria through Ca2+-induced Ca2+ uptake, perhapssimultaneously functioning as a transducer for excitation–metabolism coupling in cardiac cells [15].
2. Materials and methods
2.1. Materials
[3H]ryanodine was purchased from NEN Life Science Products (Shelton,CT). The antibody against the voltage-dependent anion channel (VDAC) wasobtained from Calbiochem (San Diego, CA) and the antibodies against thesarco- and endoplasmic reticulum Ca2+-ATPase (SERCA2) and calsequestrinwere from Santa Cruz Biotechnology (Santa Cruz, CA). Porcine brainphosphatidylethanolamine and phosphatidylserine were purchased from AvantiPolar Lipids (Birmingham, AL). Precast SDS-polyacrylamide gels were fromBio-Rad (Hercules, CA). All other chemicals were purchased from Sigma (St.Louis, MO) unless otherwise noted.
2.2. Methods
2.2.1. Isolation of rat heart mitochondriaHeart mitochondria were isolated in isotonic ice-cold mannitol/sucrose
buffer (M/S buffer; in mM: 225 mannitol, 75 sucrose, 0.5 EGTA, 1 glutathione,10 HEPES, pH 7.4) by differential centrifugation and subsequent purification ona Percoll gradient as described previously [23].
2.2.2. Preparation of mitochondrial subfractionsMitochondrial subfractions were prepared as described previously [14,24].
Isolated mitochondria from rat hearts were osmotically shocked in 10 mMNa2HPO4/NaH2PO4 for 20 min on ice followed by the addition of sucrose(20% final concentration). Mitochondrial membranes were disrupted bysonication (two times for 30 s), and eventually intact mitochondria wereremoved by centrifugation at 7000×g. The supernatant was transferred as afinal layer onto a continuous sucrose gradient (from 60% to 30% sucrose in10 mM HEPES, pH 7.4, plus protease inhibitor cocktail (Roche MolecularBiochemicals, Indianapolis, IN)) and centrifuged for at least 8 h at 70,000×gto separate mitochondrial membrane vesicles. The mitochondrial subfractions
were tested for specific marker proteins: succinate dehydrogenase (SDH) forIMM (detailed below), creatine kinase for contact sites (CS) between IMMand outer mitochondrial membrane (OMM), and the VDAC for OMM. Thecharacterized CS and IMM fractions were diluted 2-fold with M/S buffer,which contained protease inhibitor cocktail, and centrifuged for 90 min at300,000×g to sediment the mRyR. The pellet was resuspended in a smallvolume of M/S buffer and was stored in liquid nitrogen until needed. TheIMM samples used for these experiments tested negative for SERCA2,calsequestrin, and VDAC (marker proteins for the SR and OMM,respectively). Likewise, the CS samples tested negative for SERCA, butwere slightly contaminated with VDAC.
2.2.3. Succinate dehydrogenase testSuccinate dehydrogenase (SDH) was measured enzymatically according to
the procedure of Pette [25]. Two to 5 μl sample volume (for IMM) or 50 to100 μl sample volume (for other mitochondrial subfractions) was added to anassay mixture containing (in mM): 1 sodium phosphate, 1 potassium cyanide,0.1 phenazine methosulfate and 0.1 cytochrome c. The reaction was started with10 mM succinate. The increase of absorbance was measured with a Genesis 5spectrophotometer at 550 nm.
2.2.4. Denaturing SDS-PAGE and Western blot analysisFor the determination of SERCA and calsequestrin contamination within the
mitochondrial preparations, aliquots (50 μg and 30 μg, respectively) ofmitochondrial subfractions were loaded in 10% SDS-polyacrylamide gels. Theseparated proteins were transferred onto nitrocellulose membranes for 90 min at100 V. Western blots were performed using the Amersham enhancedchemiluminescence assay. To determine the validity of the RyR purifications(described below), some of the collected fractions were stained with silver saltsby modifications of a previously described procedure [26]. Briefly, 40 μg ofprotein was subjected to SDS-polyacrylamide gel electrophoresis in a 7.5%polyacrylamide precast mini-gel. The gel was incubated for 1 h in 50 ml offixing solution (ethanol: glacial acetic acid: water; 15:5:30). After discarding thefixing solution, the gel was incubated two times in 50 ml of 30% ethanol for30 min with gentle shaking. The gel was washed 3 times, 10 min each, in 100 mlof water and then incubated for 30 min in a silver solution (AgNO3:H2O; 1:20).After washing with deionized water, the gel was treated with 50 ml of anaqueous solution containing 2.5% sodium carbonate and 0.02% formaldehydeuntil stained bands were apparent. The reaction was quenched by washing thegel in 1% acetic acid for a few minutes followed by washing with deionizedwater.
2.2.5. Preparation of SR-enriched microsomes from skeletal and cardiacmuscle
Skeletal SR was isolated from frog hind leg muscles and cardiac SR wasisolated from rat hearts by differential centrifugation as described [27,28].Briefly, dissected muscle was placed in ice-cold saline solution (0.9% NaCl,10 mM Tris–HCl, pH 6.8), minced to small pieces, weighed, and homo-genized with 3 volumes of saline solution in a Waring blender for 2 min athigh speed. The saline solution contained the following protease inhibitors:leupeptin (12 μM), phenylmethylsulphonyl fluoride (PMSF; 100 μM),benzamidine (10 μM), and aprotinin (10 μM). The blended tissue washomogenized more thoroughly using a Brickmann Polytron (three times for15 s each on setting 2). The homogenate was centrifuged at 4000×g for20 min, and the resultant supernatant was filtered through 4 layers of cheesecloth and centrifuged again at 8000×g for 20 min. The supernatant from the8000×g spin was centrifuged at 40,000×g for 30 min. The 40,000×g pellet,containing the SR-microsomes, was resuspended in a sucrose buffer(containing 0.3 M sucrose, 0.1 M KCl, 5 mM Na-PIPES, pH 6.8), aliquoted,quickly frozen, and stored at −80 °C before use.
2.2.6. Purification of SR-RyR and mRyRThe frog skeletal microsomes and isolated IMM and CS subfractions were
subsequently purified via sucrose gradient centrifugation [5,29]. The thawedsamples were diluted 1:1 with solubilization buffer (final concentrations: 1 MNaCl, 40 mM Na-PIPES, 100 μM CaCl2, 0.5% CHAPS, 1:5000 dilution ofprotease inhibitor cocktail (Sigma), pH 7.2) and incubated for 1 h on ice. Thesolubilized preparations were sedimented by centrifugation at 4 °C for 40 min in

1786 B.A. Altschafl et al. / Biochimica et Biophysica Acta 1768 (2007) 1784–1795
a Beckman Ti45 rotor at 100,000×g. The RyR-containing supernatant from thesolubilized samples was layered on top of 32 ml continuous sucrose gradients(containing 5–20% sucrose, 0.3 M NaCl, 40 mM Na-PIPES, 300 μM CaCl2,0.1% CHAPS, pH 7.5). Following centrifugation in a Beckman SW28 rotor forat least 16 h at 76,000×g at 4 °C, fractions were collected (automatic pump set at1.5 ml/tube) from the bottom of the tube upwards (with fraction #1 containing20% sucrose and the final fraction containing 5% sucrose). This fractionationresulted in 20 tubes per gradient, and each fraction was silver-stained (see above)and tested for [3H]ryanodine binding activity, as performed by Valdivia et al.[30], in order to localize the RyR in the gradients. The first 4 fractions of eachgradient contained the highest counts of RyR activity. To facilitate fusion of themRyR into lipid bilayers, these fractions were aliquoted together andreconstituted into proteoliposomes using the procedure of Lee et al. [31].
2.2.7. [3H]ryanodine binding assay[3H]ryanodine binding to mitochondrial membrane vesicles was carried out as
described previously [14]. Briefly, 100 μg of rat heart mitochondrial membraneswere incubated in 0.17 M KCl, 10 mM MOPS (pH 7.0), 10 nM [3H]ryanodine,and with various concentrations of CaCl2 to set free Ca2+ concentration in therange of 100 nM to 1 mM (total volume 500 μl). The incubation occurred in thepresence (+IpTxa) or absence (control) of 1 μM IpTxa and lasted for 4 h at 37 °C.Likewise, [3H]ryanodine binding to rat cardiac SR was carried out as describedpreviously [32]. Briefly, the standard incubation medium contained 0.2 M KCl,30 mM Mops (pH 6.8), 100 μg of cardiac microsomes, 10 nM [3H]ryanodine,1 mMEGTA, and CaCl2 necessary to set free concentration of Ca
2+ in the range of10 nM to 10 mM (total volume 100 μl). The incubation occurred in the presence(+IpTxa) or absence (control) of 1 μM IpTxa and lasted 90 min at 37 °C. Sampleswere always run in duplicate, filtered onto glass fiber filters (Whatman GF/C), andwashed three times with 5 ml of cold water using a Brandel M-24R cell harvester(Gaithersburg, MD). The filters were placed in scintillation vials, 7 ml of liquidscintillation mixture was added, and the retained radioactivity was measured in aBeckman LS-6500 β-counter. The specific binding was defined as the differencebetween the binding in the absence (total binding) and presence (nonspecificbinding) of 10 μM unlabeled ryanodine.
2.2.8. Planar bilayer recordings of RyRsOur single channel recordings in lipid bilayers were performed as described
previously for the SR-RyR [32,33]. The mRyRs or purified frog skeletal RyRswere reconstituted into planar lipid bilayers composed of phosphatidylethano-lamine and phosphatidlyserine (1:1) dissolved in decane at a concentration of20 mg/ml. Single channel data were collected at steady negative voltages (whichvaried depending on the experiment) in symmetrical 300 mM cesium (Cs+)methanesulfonate, 20 mM Na–HEPES, pH 7.2. Proteoliposomes or purifiedRyRs were added to the cis (cytosolic) side of the chamber, which contained thevoltage command electrode. The trans chamber contained the reference electrodeand corresponded to the luminal side of the channel. In this configuration, Cs+
flows from the luminal to the cytosolic side at negative holding potentials. Cs+
was chosen as the charge carrier to increase the channel conductance and to avoidinterference from K+ channels present in the SR and inner mitochondrialmembranes. Chloride channels were mostly blocked by replacing chloride withthe impermeant methanesulfonate anion. Contaminant Ca2+ (∼3–5 μM, asdetermined by a Ca2+ electrode) served to activate RyRs. In some experiments,50 μMCaCl2, 10 μM ryanodine, or 500 nM IpTxa was added to the cis side of thechannel, as detailed in the text. Some of the recordings were also made with20 mM KCl added to both sides of the chamber in order to stimulate channelactivity [34], as specified in the text. Channel activity was filtered with an 8-polelow pass Bessel filter set at 1.5 kHz and digitized at a rate of 4 kHz using aDigidata 1200 AD/DA interface. Data acquisition and analysis were performedwith Axon Instruments (Burlingame, CA) hardware and software (pClamp 8).
3. Results
3.1. The mRyR is distinct from the SR-RyR
Calsequestrin and SERCA are specific marker proteins forthe SR. To determine if the rat heart mitochondrial subfractions
had any SR contamination, Western blots performed with anantibody against the cardiac isoform of SERCA (SERCA2a)showed that the rat cardiac SR-containing cytosol (Fig. 1A, lane1), but not mitochondrial subfractions (Fig. 1A, lanes 2–6) hadimmunoreactivity against this antibody. Likewise, cardiac SRand mitochondrial vesicles from rat hearts retained the presenceof calsequestrin (Fig. 1B, lanes 1–2); however, furtherpurification and separation of mitochondrial vesicles intoOMM, CS, and IMM completely removed calsequestrin fromthese preparations. All IMM fractions were free of the VDAC, amarker protein for the OMM (not shown, see [14,15]). The CSfraction, consisting of attachments of the OMM and IMM, wasmostly SERCA2a-free (Fig. 1A, lane 3). To further ensure thequality of the IMM fraction, we measured the activity ofsuccinate dehydrogenase, an IMM-resident enzyme. During thepurification of IMM vesicles the activity of this enzyme shouldincrease because matrix proteins or proteins of the OMM areremoved. As indicated in Table 1, the specific activity forsuccinate dehydrogenase increased from 1.35±1.06 mU/mgprotein in the OMM to 2598.6±318.6 mU/mg protein in theIMM. We used IMM vesicles in most lipid bilayer experimentsafter verifying the lack of detection of OMM and SR proteins.Thus, we can safely ascertain that the channels we labeled“mRyR” in this study are localized in the mitochondria and donot correspond to SR channels.
To further demonstrate that the IMM fractions were notcontaminated with SR components, we conducted Ca2+-dependence of [3H]ryanodine binding curves to IMM and SRvesicles from rat hearts. The rationale was based on thesurprising finding that the mRyR cross-reacts with an anti-RyR1(skeletal isoform) antibody, not an anti-RyR2 antibody [15].Further, if IMM contained RyRs from SR vesicles, then bothpreparations would show a RyR2-like response to Ca2+, whichdiffers from a RyR1-like response in the modest Ca2+-dependent inactivation occurring at high [Ca2+]. Finally, weconducted these experiments in the absence and the presence ofIpTxa, a high-affinity peptide ligand of RyRs [32,35] that affectsdifferently the Ca2+-dependence of [3H]ryanodine bindingcurve to RyR1 and RyR2 channels. IpTxa increases [3H]ryanodine binding to RyR1 at all [Ca2+], but it produces dualeffect on RyR2, initially increasing (pCa 7–5) and thendecreasing (pCa 4-2) [3H]ryanodine binding. The result is inan augmented bell-shaped curve in RyR1 and a sigmoidal curvein RyR2 [32,35]. Thus, IpTxa could exaggerate RyR isoform-dependent differences in [3H]ryanodine binding, which couldhelp estimate the level of SR contamination in our mitochon-drial membranes. Fig. 1C shows the Ca2+ dependence of [3H]ryanodine binding to the isolated cardiac mitochondrial vesiclesand the effect of IpTxa. Specific binding in the absence of IpTxa(control) had a threshold for detection at∼100 nM Ca2+ (pCa 7)and was maximal at 10 μM Ca2+ (pCa 5). Higher Ca2+
concentrations decreased [3H]ryanodine binding, resulting in abell-shaped curve remarkably similar to that of RyR1 [32,35]. Inthe presence of IpTxa (+IpTxa) the binding curve was also bell-shaped but significantly augmented by ∼2-fold (see figurelegend). By contrast, the Ca2+-dependence of [3H]ryanodinebinding to SR vesicles displayed a different profile (Fig. 1D,

Fig. 1. Western blot of SERCA and calsequestrin in mitochondrial subfractions and the effect of IpTxa on the Ca2+ dependence of [3H]ryanodine binding.Mitochondrial subfractions from isolated and osmotically shocked rat heart mitochondria were probed with SERCA2a (A) and calsequestrin (B) antibodies to test forcontamination of the IMM and CS with SR or cytosolic membrane fragments (50 μg and 30 μg of protein per lane in each blot, respectively). The arrows indicate thepositions and approximate molecular weights of the SERCA2a and calsequestrin proteins, respectively. Each numbered lane corresponds to the cellular preparationlisted to the right of each immunoblot. Rat cardiac SR vesicles were used as positive controls for each blot (lane 1). The binding of [3H]ryanodine to rat mitochondrialvesicles (C) and rat cardiac SR (D) was performed as described in Materials and methods. IpTxa (1 μM) was either present (+IpTxa, dashed line) or absent (control,solid line) throughout the incubation period. Values are normalized to the Bmax of the control samples for each graph. Data points are the mean (±S.E.) of threeindependent binding assays using mitochondrial vesicles and five independent binding assays using cardiac SR microsomes. Lines linking data points have notheoretical meaning.
1787B.A. Altschafl et al. / Biochimica et Biophysica Acta 1768 (2007) 1784–1795
control) and was unevenly affected by IpTxa (+IpTxa). In themost extreme example, that of the Ca2+-dependent inactivationof binding at pCa 3 in control conditions, binding is almost nullin RyR1 [32,35] and mRyR (Fig. 1C), but robust in cardiac SR(Fig. 1D). Thus, mRyR behaved like RyR1 in [3H]ryanodinebinding assays, and not as RyR2 as expected if themitochondrial samples used in this study were contaminatedby SR proteins. Our results therefore suggest that the mRyRisolated from mitochondrial membrane vesicles indeed origi-nates from a separate pool of RyRs.
3.2. Diversity of ion channels in mitochondrial vesicles
In an attempt to characterize the single channel kinetics of themRyR, we reconstituted rat IMM or CS vesicles into planar lipidbilayers as described [32,33]. The conditions used for these
Table 1Succinate dehydrogenase (SDH) activity is enriched in the IMM fraction
Compartment SDH activity [mU/mg]±SE
Mitochondria 704.837±68.351Cytosol 10.138±2.364OMM 1.355±1.062CS 337.629±55.646IMM 2598.589±318.596
experiments (see Materials and methods) theoretically shouldfavor recording of large currents through RyRs while silencingother unrelated channels. In some experiments, 20 mMKCl wasadded to the recording solution, as Cl− ions have provedeffective in enhancing RyR activity [34]. A variety of substatesfrom individual or multiple types of channels were generallyseen, obstructing the identification of unitary conductances (Fig.2A, B), but even then, there were instances in which a potentialmRyR was conspicuous within the recording. In Fig. 2A forexample, the expanded section of the recording contained singlechannel fluctuations that had gating kinetics and unitaryconductance remarkably similar to SR-RyR. The full, unitaryCs+ conductance of this 500 ms segment was 661 pS; inaddition, the channel was unresponsive to 2 μM CsA, aconcentration that effectively blocks the mitochondrial perme-ability transition pore (PTP) [36]. These brief occurrences ofRyR-like channel gating were promising and demonstrated thepossibility that, in the absence of other unrelated currents, achannel from the IMM with similar conductance as the SR-RyRmay fuse into the planar lipid bilayer.
Given the heterogeneity of channels in the mitochondrialmembranes [37,38], other channels could be able to conductCs+ as well and may have been enhanced by the presence ofKCl in the recording medium. Fig. 2B shows traces of steady-state activity from several channels or multiple substates of an

Fig. 3. Silver staining analysis of a RyR from purified IMM and CS subfractions.Following CHAPS solubilization and sucrose gradient centrifugation, severalmembrane fractions were collected, samples (40 μg) were run on an SDS-PAGEgel, and stained with silver salts to test the validity of the purification method.The arrow indicates the position of a 200-kDa molecular weight myosin markermolecule. The silver salts stained a protein band of approximately 500 kDa.MW, kaleidoscope prestained standard markers (Bio-Rad); Frog somes, frogskeletal SR-enriched microsomes 40 k pellet; Frog F1, purified frog skeletalfraction 1 (20% sucrose); Pig F1, purified pig cardiac fraction 1 (20% sucrose);IMM F1, purified IMM fraction 1 (20% sucrose); CS F3, purified CS fraction 3.
Fig. 2. Detection of mitochondrial channels in planar lipid bilayers. (A) Single mitochondrial channel recorded from IMM vesicle reconstituted into a planar lipidbilayer. The enlarged portion of the trace represents a potential mRyR. Channel openings are represented as downward deflections of the base-line current. Solutions inboth chambers bathing the planar bilayer were (in mM) 300 Cs+ methanesulfonate, 20 KCl, and 20 MOPS (pH 7.2) along with nominal levels of Ca2+ (∼3–5 μM).After channel fusion, 2 μMCsAwas added to the cis side in each experiment. With a holding potential of −30 to −40 mV, Cs+ current flows from the trans (luminal) tocis (cytosolic) side. All records were low pass filtered at 1.5 kHz using an 8-pole Bessel filter and digitized at 4 kHz. (B) Representative single channel traces of avariety of ion channels/channel substates from IMM vesicles under the same conditions as described above. Traces b, c, and d have the same current/time scale astrace e. c: closed state of the channels.
1788 B.A. Altschafl et al. / Biochimica et Biophysica Acta 1768 (2007) 1784–1795
individual channel recorded from IMM vesicles at a holdingpotential of −30 to −40 mV in the presence of nominal levels ofCa2+ (∼3–5 μM). Interestingly, several mitochondrial currentsrecorded such as traces 2B-b, c, and e, had extremely fast gatingand transitioned between a variety of conductance states. Thesechannels/channel substates most likely correspond to the PTPand/or VDAC, or translocases of the inner membrane, whencompared with published electrophysiological characterizationsof these channels (for review see [36]). Trace 2B-c, for example,was inhibited by the addition of 2 μM CsA as shown in Fig. 2(see the last 1.2 s) suggesting that this channel is indeed the PTP.
3.3. Purification of mitochondrial subfractions by sucrosegradient centrifugation
Single channel recordings such as those depicted in Fig. 2substantiated the necessity for additional purification of themitochondrial subfractions in order to isolate the mRyR. Weused sucrose-gradient sedimentation of solubilized IMM or CSvesicles to purify the mRyR, as originally described for SR-RyR[5,29]. The sedimentation coefficient for the RyR is 30 S [39],thus the RyR localizes in the heavier sucrose fractions with thistechnique. Frog skeletal and pig cardiac SR-enriched micro-somes were subjected to the same treatment as the mitochon-drial subfractions and were used as positive controls (Fig. 3,
Frog-F1 and Pig-F1). The occurrence of a high molecularweight band (∼500 kDa) in the purified IMM or CS fractionswith migration similar to SR-RyR monomers was confirmed bysilver staining (Fig. 3). Fig. 3 verifies that the mRyR localizedwithin the heavier, 20% sucrose fractions (IMM-F1, CS-F3).The detection of the mRyR within the heavier sucrose fractionsconfirmed that the mRyR may be isolated in an analogous

1789B.A. Altschafl et al. / Biochimica et Biophysica Acta 1768 (2007) 1784–1795
manner as the SR-RyR; we thus obtained a more purifiedpreparation to use in lipid bilayer experiments.
3.4. Single channel kinetics of the mRyR
In order to facilitate fusion of mRyR into planar lipidbilayers, we reconstituted the solubilized IMM and CS proteinsinto proteoliposomes using the procedure described by Lee et al.[31]. We used single channel conductance as the first of a seriesof biophysical and pharmacological criteria to designatemitochondrial channels as mRyRs [40,41]. Accordingly,profiles of single channels likely corresponding to mRyRssolely based on their unitary channel conductances wererecorded (n=21). Fig. 4A shows six representative channelsreconstituted from the IMM or CS purified fractions that displayconductance values similar to Cs+-conducting SR-RyRs. Tracesa, b, and c represent channels fused from purified IMMproteoliposomes, while traces d, e, and f are channels fusedfrom purified CS proteoliposomes. All channels depicted in Fig.4A were recorded at −20 to −50 mV, thus downward de-flections of the baseline current correspond to channel open-ings. In all traces there are discrete, fully-resolved openings thatare characteristic of classical RyRs. Some of the openings arelonger lasting (traces c and d) and some are fast-flickering
Fig. 4. A kinetic profile of the mitochondrial ryanodine receptor. (A) RyRs from purifinto planar lipid bilayers. Traces a, b, and c represent single channel recordings of threcordings from purified CS proteoliposomes. Channels are activated by steady appdeflections of the base-line current represent the open state of the channel. Themethanesulfonate, 20 KCl and 20 MOPS (pH 7.2). All recordings were made in theaddition of 2 μMCsA. (B) Single channel current–voltage relationship for trace a frostate of the RyRs.
(trace f). In some of the recordings (traces a and e), the channelsenter into periods of half-conductance states that are reminiscentof the intrinsic subconductance states present in SR-RyRs [39].Thus, most kinetic states previously reported for SR-RyRs [7]are represented in our mRyR recordings. Fig. 4B displays arepresentative single channel current–voltage relationship ofthese mitochondrial channels (trace 4A-a). In the range of −30to +20 mV, the slope conductance was 616±41 pS whenmeasured in symmetrical 300 mM cesium methanesulfonatesolution. Overall, the unitary conductance for the purified IMMor CS channels presented in Fig. 4 fluctuated between 550 and790 pS, values which are close to those expected for SR-RyRunder these recording conditions [42]. The slight but significantdeviation of these values from the average Cs+-conducting SR-RyR is most likely due to modifications of the recordingsolution (See Discussion).
3.5. Calcium activation of the mRyR
Next, we investigated the effects of some classical RyRmodulators on mitochondrial channels that had conductancemeasurements appropriate for RyRs as described above. TheRyRs in the SR of both cardiac and skeletal muscle are activatedby nanomolar to micromolar concentrations of Ca2+ [34]. We
ied IMM and CS subfractions were reconstituted into proteoliposomes and fusede mRyR from purified IMM proteoliposomes, while traces d, e, and f are mRyRlied negative recording potentials of between −20 to −50 mV, thus downwardrecording solution for all of the experiments contained (in mM) 300 Cs+
presence of nominal levels of Ca2+ (∼3–5 μM) and were unresponsive to them panel Awith a linear slope conductance measurement of 616±41 pS. c: closed
1790 B.A. Altschafl et al. / Biochimica et Biophysica Acta 1768 (2007) 1784–1795
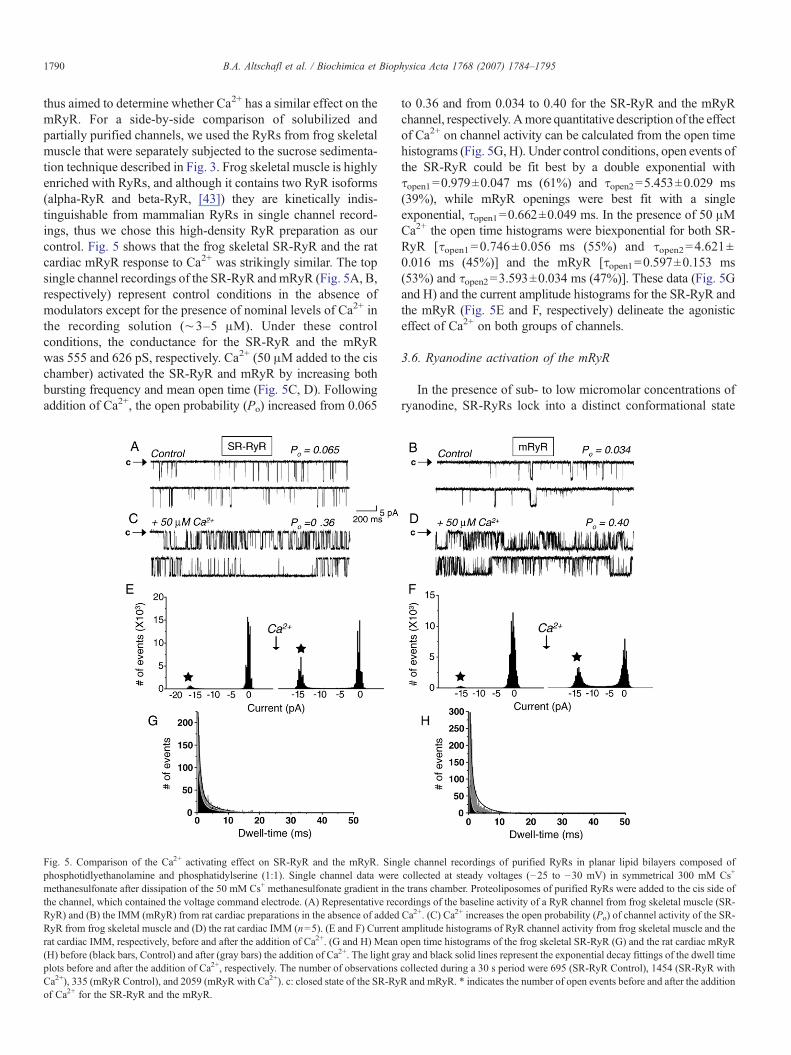
thus aimed to determine whether Ca2+ has a similar effect on themRyR. For a side-by-side comparison of solubilized andpartially purified channels, we used the RyRs from frog skeletalmuscle that were separately subjected to the sucrose sedimenta-tion technique described in Fig. 3. Frog skeletal muscle is highlyenriched with RyRs, and although it contains two RyR isoforms(alpha-RyR and beta-RyR, [43]) they are kinetically indis-tinguishable from mammalian RyRs in single channel record-ings, thus we chose this high-density RyR preparation as ourcontrol. Fig. 5 shows that the frog skeletal SR-RyR and the ratcardiac mRyR response to Ca2+ was strikingly similar. The topsingle channel recordings of the SR-RyR andmRyR (Fig. 5A, B,respectively) represent control conditions in the absence ofmodulators except for the presence of nominal levels of Ca2+ inthe recording solution (∼3–5 μM). Under these controlconditions, the conductance for the SR-RyR and the mRyRwas 555 and 626 pS, respectively. Ca2+ (50 μM added to the cischamber) activated the SR-RyR and mRyR by increasing bothbursting frequency and mean open time (Fig. 5C, D). Followingaddition of Ca2+, the open probability (Po) increased from 0.065
Fig. 5. Comparison of the Ca2+ activating effect on SR-RyR and the mRyR. Singphosphotidlyethanolamine and phosphatidylserine (1:1). Single channel data weremethanesulfonate after dissipation of the 50 mM Cs+ methanesulfonate gradient in ththe channel, which contained the voltage command electrode. (A) Representative recRyR) and (B) the IMM (mRyR) from rat cardiac preparations in the absence of addedRyR from frog skeletal muscle and (D) the rat cardiac IMM (n=5). (E and F) Currentrat cardiac IMM, respectively, before and after the addition of Ca2+. (G and H) Mean(H) before (black bars, Control) and after (gray bars) the addition of Ca2+. The light grplots before and after the addition of Ca2+, respectively. The number of observationsCa2+), 335 (mRyR Control), and 2059 (mRyR with Ca2+). c: closed state of the SR-Ryof Ca2+ for the SR-RyR and the mRyR.
to 0.36 and from 0.034 to 0.40 for the SR-RyR and the mRyRchannel, respectively. Amore quantitative description of the effectof Ca2+ on channel activity can be calculated from the open timehistograms (Fig. 5G, H). Under control conditions, open events ofthe SR-RyR could be fit best by a double exponential withτopen1=0.979±0.047 ms (61%) and τopen2=5.453±0.029 ms(39%), while mRyR openings were best fit with a singleexponential, τopen1=0.662±0.049 ms. In the presence of 50 μMCa2+ the open time histograms were biexponential for both SR-RyR [τopen1=0.746±0.056 ms (55%) and τopen2=4.621±0.016 ms (45%)] and the mRyR [τopen1=0.597±0.153 ms(53%) and τopen2=3.593±0.034 ms (47%)]. These data (Fig. 5Gand H) and the current amplitude histograms for the SR-RyR andthe mRyR (Fig. 5E and F, respectively) delineate the agonisticeffect of Ca2+ on both groups of channels.
3.6. Ryanodine activation of the mRyR
In the presence of sub- to low micromolar concentrations ofryanodine, SR-RyRs lock into a distinct conformational state
le channel recordings of purified RyRs in planar lipid bilayers composed ofcollected at steady voltages (−25 to −30 mV) in symmetrical 300 mM Cs+
e trans chamber. Proteoliposomes of purified RyRs were added to the cis side ofordings of the baseline activity of a RyR channel from frog skeletal muscle (SR-Ca2+. (C) Ca2+ increases the open probability (Po) of channel activity of the SR-amplitude histograms of RyR channel activity from frog skeletal muscle and theopen time histograms of the frog skeletal SR-RyR (G) and the rat cardiac mRyRay and black solid lines represent the exponential decay fittings of the dwell timecollected during a 30 s period were 695 (SR-RyR Control), 1454 (SR-RyR withR and mRyR. * indicates the number of open events before and after the addition

Fig. 6. Ryanodine induces subconducting states in mRyR channels. Single channel recordings of purified mRyRs recorded using the same conditions as described inMaterials and methods and in Fig. 5 legend. (A) a: representative recording of the baseline activity of a mRyR from the IMM in the absence of ryanodine. b and c: Po
and current amplitude histograms, respectively, of RyR channel activity before the addition of modulators. (B) a: representative recording of mRyR single channelactivity after the addition of 10 μM ryanodine to the cis chamber (n=3). b and c: Po and current amplitude histograms, respectively, demonstrating the initial activatingeffect of ryanodine because of prolonged subconducting activity. c: closed state of the mRyR.
1791B.A. Altschafl et al. / Biochimica et Biophysica Acta 1768 (2007) 1784–1795
characterized by long-lasting subconducting openings repre-senting ∼50% of full conductance [3]. We therefore tested theeffect of ryanodine on mitochondrial channels. Fig. 6 shows anIMM channel reconstituted in a lipid bilayer in the absence(control, Fig. 6A-a), and the presence (Fig. 6B-a) of 10 μMryanodine added to the cis chamber. Under control conditions(Fig. 6A) the channel displayed quick, intermittent openingsthat resulted in overall low Po of 0.023 for the duration of therecording. The diary of activity for the first 20 s of channelrecording (Fig. 6A-b) emphasizes the quiescent nature of thischannel in the absence of external modulators. By contrast,addition of 10 μM ryanodine to the channel (Fig. 6B) drama-tically altered its activity by inducing the appearance of a half-conducting state that lasted for several seconds as depicted inthe diary of openings (Fig. 6B-b) and the current amplitudehistogram (Fig. 6B-c). Even then, the channel also showed anuncharacteristic response to ryanodine by alternating betweenperiods of sub- and full-conducting openings for the first ten s,followed by complete inhibition throughout the remainder ofthe recording (Fig. 6B-b). As a result of this bimodal effect, thePo initially increased to 0.97, presumably while ryanodineexerted its activating effect, and then fell to zero as ryanodineexerted a blocking effect. The normalized Po for the first 20 swas therefore 0.13 and zero thereafter. Interestingly, we foundno mitochondrial channels that irreversibly locked into
subconducting states by ryanodine. There were channels com-pletely inhibited by 10 μM ryanodine (n=2), while otherchannels displayed alternating periods of sub- and full-conducting states (n=3) in response to ryanodine.
3.7. Imperatoxin A induces mRyR subconductance
Unlike its differential effects on the Ca2+ dependence of [3H]ryanodine binding, IpTxa affects the single channel kinetics ofall RyR isoforms (RyR1, RyR2 and RyR3) in a similar manner[35,44]. Like ryanodine, IpTxa at nanomolar concentrationsinduces the appearance of long-lasting subconductance states inboth SR-RyR isoforms, RyR1 and RyR2. At negative recordingpotentials (such as those used for these experiments), thesubstate conductance represents approximately 43% of the fullconductance openings [35]. Unlike ryanodine, however, thereversible binding of IpTxa allows the channel to fluctuatebetween sub- and full-conducting states. Fig. 7 shows that,remarkably, IpTxa exhibited its classical activating effect ona reconstituted mRyR channel purified from mitochondrialsubfractions.
The mRyR depicted in Fig. 7A was recorded under controlconditions (in a nominally-free Ca2+ solution without additionalmodulators) and had gating kinetics characteristic of SR-RyRs,i.e., numerous fast openings alternating with few long-lasting

1792 B.A. Altschafl et al. / Biochimica et Biophysica Acta 1768 (2007) 1784–1795
openings, both giving rise to sporadic bursts of channel activity.The overall activity and conductance of the channel aresummarized in the current amplitude histogram (Fig. 7A, rightpanel). Conversely, the same channel after addition of 500 nM
IpTxa (Fig. 7B, left panel) and the resulting amplitudehistogram (Fig. 7B, right panel) displayed the signature effectof IpTxa on the mRyR. The toxin induced long-lastingsubconducting states interspersed between periods of full-

1793B.A. Altschafl et al. / Biochimica et Biophysica Acta 1768 (2007) 1784–1795
conducting channel activity. As expected, the substate con-ductance approximated 40% of the full conductance openings.The amplitude histogram of mRyR channel activity (Fig. 7B,right panel) emphasizes that modulation by IpTxa increased thePo of the mRyR by increasing the frequency of channelopenings and decreasing the duration of the closed events.Indeed, mRyR Po augmented from 0.13 in control to 0.86 in thepresence of IpTxa (Fig. 7A and B, respectively). In Fig. 7C wecompared the gating of the presumed mRyR with that of frogSR-RyR recorded under identical experimental conditions. Asshown in the compressed and expanded traces, the activity ofboth channels oscillated between periods of long-lastingsubconductance states, representing occupation of the channelby IpTxa [35], and normal full conductance openings,representing gating of IpTxa-free channels. Notice the remark-able resemblance in the gating characteristics of both channels.
4. Discussion
The availability of techniques that segregate mitochondrialsubfractions from SR and other membrane particulates [14,15]has made it possible to identify a channel protein with physicalcharacteristics similar to those of SR-RyRs in an unusuallocation, the mitochondria. The differential effects of IpTxa onthe Ca2+ dependence of [3H]ryanodine binding (Fig. 1C, D)reinforce the notion that mRyR has exquisite functionaldifferences with the SR-RyR from cardiac tissue. Likewise,Western blots of the mitochondrial subfractions recognize ahigh molecular weight (∼500 kDa) band within the IMM thatcross-reacts with RyR1 (skeletal isoform) but not RyR2 (cardiacisoform) specific antibodies, again strengthening the assertionthat this protein is a unique mitochondrial component and not anSR contaminant (which would otherwise yield a strong RyR2signal) [15]. [3H]Ryanodine, a specific ligand of the SR-RyR,binds to mitochondrial vesicles with high affinity, and in thepresence of ryanodine, Ca2+ uptake into isolated heartmitochondria is substantially suppressed [14]. Thus, a proteinsimilar to SR-RyR appears to be an important functional Ca2+
transporter within the IMM.The present study demonstrates that the mitochondria
contain a functional RyR within the IMM that shares severalpharmacological and electrophysiological characteristics withthe SR-RyR at the single channel level. We reasoned that thepresently described mRyR is of mitochondrial origin and freefrom RyR2 contamination on the following grounds: (1) IMMand CS subfractions were free from detectable amounts ofSERCA and calsequestrin protein (Fig. 1A and B, respectively),indicative for SR; (2) some of the channels fused into lipidbilayers were blocked by CsA, indicating that these vesiclescontained the mitochondrial PTP; (3) ryanodine exerted
Fig. 7. Imperatoxin A achieves its classical RyR effect by inducing periodic, prolongedrecorded using the same conditions as described inMaterials andmethods. (A) Represewithout the presence of RyR modulators. The right panel represents a current amplitudepicted in panel A recorded after the addition of 500 nM IpTxa to the cis chamber (nafter IpTxa took effect. (C) The effects of IpTxa activity on a frog skeletal SR-RyR vs. tpresence of IpTxa. The enlarged trace represents a 2-s subsection of the channel recordsubfractions in the presence of IpTxa. The enlarged trace represents a 2-s subsection
distinctive effects on mRyR that differ from those exerted bythe alkaloid on RyR2 (Fig. 6), suggesting that mRyR representsa RyR isoform that retains the bulk of RyR properties butmaintains unique characteristics, akin to the different Ca2+-dependence of channel activity seen with RyR1 and RyR2[34,45]; and (4) the Ca2+ dependence of [3H]ryanodine bindingin the presence and absence of IpTxa to IMM vesicles wasanalogous to that expected for RyR1 (Fig. 1C, D), consistentwith Western blot data [15], and different from that of RyR2, aswould be expected if cardiac SR vesicles contaminated ourIMM preparations.
The IMM contains a variety of proteins, which exhibitchannel activity in bilayer membranes, including the Ca2+
uniporter [46], the ANT [47–49], and proteins of themitochondrial protein transport system [50]. Several studiessupport the hypothesis that the PTP is a protein complexcontaining components of the IMM, such as the ANT, and theOMM, such as VDAC [51,52]. Given the high sensitivity of ourelectrophysiological assay, it was not surprising that wereconstituted a multiplicity of ion channels from the IMM orCS subfractions (Fig. 2). Some of the traces depicted in Fig. 2were from channels of unknown molecular identity, so it islikely that we recorded IMM transport proteins that have not yetbeen fully characterized in lipid bilayer experiments. Unfortu-nately, our single channel lipid bilayer assay does not allow usto determine the precise location of the mRyR within thedistinct mitochondrial substructures. However, VDAC, amarker protein for the OMM, was below detection levels inWestern blot characterization of IMM vesicles, which mostlikely excludes the localization of mRyR in OMM. Importantly,we had the most success fusing mRyR channels from the IMMpreparations (the mitochondrial subfraction used for themajority of the experiments described in this study). Therefore,if we make the reasonable assumption that the likelihood ofdetecting a protein in a given vesicular preparation is directlyproportional to the relative abundance of the protein in the samepreparation, then the higher frequency of mRyR detection inIMM vesicles favors the notion that IMM is the preferredlocation of this channel, in agreement with supporting dataobtained by electron microscopy and Western blot analysis withRyR antibodies [14,15].
Traditionally, the enormous mass (∼2 million Da/channeltetramer) and sedimentation coefficient (30 S) of the SR-RyRhave been exploited for separation of this channel from other SRproteins [29,39]. Purification by sucrose gradient centrifugationalso proved successful to separate the mRyR from othermitochondrial channels (Fig. 3). This provided further evidencethat the mRyR is a channel complex that can be purified with thesame methods used for the SR-RyR. Nevertheless, in spite ofmRyR enrichment, we continued to detect “contaminating”
substates of mRyR channel activity. Single channel recordings of purified mRyRsntative single channel profile of mRyR recorded in a nominally-free Ca2+ solutionde histogram for 30 s of the trace represented on the left. (B) The same channel=5). The right panel represents a current amplitude histogram for 30 s of the tracehe rat cardiac mRyR. Top: a 1-min recording of a RyR from frog skeletal SR in theing. Bottom: a 1-min recording of a RyR from purified rat cardiac mitochondrialof the mRyR channel recording. c: closed state of the SR-RyR and mRyR.

1794 B.A. Altschafl et al. / Biochimica et Biophysica Acta 1768 (2007) 1784–1795
channels from purified subfractions that did not appear tocorrespond to the mRyR. The predominant types of unrelatedchannels were blocked by CsA, suggesting that the PTP mayhave co-segregated with the mRyR in our sucrose sedimentationprocedure. It has been hypothesized that the PTP is a multi-protein complex, which may consist of VDAC, the ANT,hexokinases [51], and the benzodiazepine receptor [53] allhaving a total molecular weight of ∼600 kDa. Given the largemolecular weight of the PTP multi-protein complex and that ofthe mRyR, it is probable that both of these protein complexescould colocalize within the heavier sucrose fractions.
In order to characterize the mRyR at the single channel level,we established a set of criteria to designate certain channels asmRyR candidates. The first index of activity to categorize amitochondrial channel as potential mRyR was single channelconductance. Smith et al. [42] found that RyR1 conducts Cs+ athigher rate than other monovalent cations. For example, inrecording solution containing 50 mM cis and 250 mM trans[Cs+], the unitary conductance of RyR1 is ∼500 pS [42]. Forthe present study, we used 50 mM cis and 300 mM trans [Cs+](and then dissipated the gradient following channel fusion sothat both sides of the mRyR contained 300 mM Cs+), predictingmRyR single channel unitary conductances of 500–600 pS.Nevertheless, the addition of KCl to the recording solution mayhave caused the mRyR conductance to deviate from thosevalues expected for a Cs+-conducting RyR1. When millimolarconcentrations of KCl were added to the recording solution tofavor channel activity, we expected an increase of RyRconductance to above 600 pS [42], which we observed in thesingle channel mRyR experiments recorded under theseconditions (Fig. 4A, traces a–e). This was possibly the singlemost significant factor in the unitary conductance variation weobserved (Fig. 4), followed perhaps by Cs+ leak from cis totrans chambers during bilayer re-painting and the intrinsic noiseof the high-frequency recording bandwidth (1.5 kHz).
The second criterion to designate channel activity as thatcorresponding to mRyR was the lack of response to CsA(∼2 μM), excluding the possibility that the channel recordingsrepresented substates of the PTP, which can rapidly flicker fromits fully closed state to a substate of 600–700 pS [37]. Finally,we expected the candidate channels to be sensitive to Ca2+,ryanodine, and IpTxa (Figs. 5–7) because, although with smallvariations, these modulators produce signature effects on allRyR isoforms. The profile of the single channel recordings andthe unitary Cs+ conductance measurements of the channelsdepicted in Fig. 4A, as well as their lack of modulation by CsA,confirmed that these channels are indeed representative of themRyR. Likewise, as seen in Figs. 5–7, the mRyR displayed theelemental attributes that are present in single channel lipidbilayer recordings of a SR-RyR and responded similarly toclassical modulators of SR-RyR activity.
Using these criteria, we have determined some of the mostsalient pharmaco-kinetic characteristics of the mRyR. ThemRyR described in this study displays gating propertiesreminiscent of those seen in the classical SR-RyR, includingan analogous conductance for monovalent cations and Ca2+
sensitivity. The mRyR conforms to a ryanodine-altered substate
in the presence of the ligand, and IpTxa binds reversibly to themRyR allowing for the oscillation of channel activity fromsubconducting to full-conducting states. Future experimentsshould determine whether the typical SR-RyR accessoryproteins are associated with this intriguing mRyR and if theirmodulatory effects are similar in a novel environment. Equallyimportant, the present functional assays should provide apractical platform to test the role of the mRyR as a modulator ofCa2+ fluxes in mitochondria and as a transducer for excitation–metabolism coupling in cardiac cells.
Acknowledgements
This investigation was supported by NIH grant NRSA-T32-HL07936 (University of Wisconsin Cardiovascular ResearchTraining grant to B.A.A.); American Heart Association grant0335425T (to G.B.); NIH grant HL-33333 and AHA grant0050839T (to S.S.S), and NIH grant HL-55438 (to H.H.V.).
References
[1] A. Fabiato, Calcium-induced release of calcium from the cardiacsarcoplasmic reticulum, Am. J. Physiol. 245 (1983) C1–C14.
[2] M.D. Stern, E.G. Lakatta, Excitation–contraction coupling in the heart: thestate of the question, FASEB J. 6 (1992) 3092–3100.
[3] T. Imagawa, J.S. Smith, R. Coronado, K.P. Campbell, Purified ryanodinereceptor from skeletal muscle sarcoplasmic reticulum is the Ca2+-permeable pore of the calcium release channel, J. Biol. Chem. 262(1987) 16636–16643.
[4] M. Inui, A. Saito, S. Fleischer, Purification of the ryanodine receptor andidentity with feet structures of junctional terminal cisternae of sarcoplasmicreticulum from fast skeletal muscle, J. Biol. Chem. 262 (1987) 1740–1747.
[5] F.A. Lai, H.P. Erickson, E. Rousseau, Q.Y. Liu, G. Meissner, Purificationand reconstitution of the calcium release channel from skeletal muscle,Nature 331 (1988) 315–319.
[6] J.L. Sutko, J.A. Airey, Ryanodine receptor Ca2+ release channels: doesdiversity in form equal diversity in function? Physiol. Rev. 76 (1996)1027–1071.
[7] M. Fill, J.A. Copello, Ryanodine receptor calcium release channels,Physiol. Rev. 84 (2002) 893–922.
[8] D.M. Bers, Macromolecular complexes regulating cardiac ryanodinereceptor function, J. Mol. Cell. Cardiol. 37 (2004) 417–429.
[9] R. Zucchi, S. Ronca-Testoni, The sarcoplasmic reticulum Ca2+ channel/ryanodine receptor: modulation by endogenous effectors, drugs anddisease states, Pharmacol. Rev. 49 (1997) 1–51.
[10] G. Meissner, A. el-Hashem, Ryanodine as a functional probe of the skeletalmuscle sarcoplasmic reticulum Ca2+ release channel, Mol. Cell. Biochem.114 (1992) 119–123.
[11] E. Rousseau, G. Meissner, Single cardiac sarcoplasmic reticulum Ca2+-release channel: activation by caffeine, Am. J. Physiol. 256 (1989)H328–H333.
[12] C. Franzini-Armstrong, F. Protasi, Ryanodine receptors of striatedmuscles: a complex channel capable of multiple interactions, Physiol.Rev. 77 (1997) 699–729.
[13] G.B. Gurrola, X. Zhu, H.H. Valdivia, Scorpion peptides as high-affinityprobes of RyR function, in: X.T. Wehrens, A.R. Marks (Eds.), RyanodineReceptors: Structure, Function andDysfunction in Clinical Disease, Springer,New York, 2005, pp. 191–200.
[14] G. Beutner, V.K. Sharma, D.R. Giovannucci, D.I. Yule, S.S. Sheu,Identification of a ryanodine receptor in rat heart mitochondria, J. Biol.Chem. 276 (2001) 21482–21488.
[15] G. Beutner, V.K. Sharma, L. Lin, S.Y. Ryu, R.T. Dirksen, S.S. Sheu, Type 1ryanodine receptor in cardiac mitochondria: transducer of excitationmetabolism coupling, Biochim. Biophys. Acta 1717 (2005) 1–10.

1795B.A. Altschafl et al. / Biochimica et Biophysica Acta 1768 (2007) 1784–1795
[16] G. Hajnóczky, L.D. Robb-Gaspers, M. Seitz, A.P. Thomas, Decoding ofcytosolic calcium oscillations in the mitochondria, Cell 82 (1995)415–424.
[17] E. Chacon, H. Ohata, I.S. Harper, D.R. Trollinger, B. Herman, J.J. Lemasters,Mitochondrial free calcium transients during excitation–contractioncoupling in rabbit cardiac myocytes, FEBS Lett. 382 (1996) 31–36.
[18] P.B. Simpson, J.T. Russell, Mitochondria support inositol 1,4,5-trispho-sphate-mediated Ca2+ waves in cultured oligodendrocytes, J. Biol. Chem.271 (1996) 33493–33501.
[19] M.R. Duchen, A. Leyssens, M. Crompton, Transient mitochondrialdepolarizations reflect focal sarcoplasmic reticular calcium release insingle rat cardiomyocytes, J. Cell Biol. 142 (1998) 975–988.
[20] V. Sharma, V. Ramesh, C. Franzini-Armstrong, S.S. Sheu, Transport ofCa2+ from sarcoplasmic reticulum to mitochondria in rat ventricularmyocytes, J. Bioenerg. Biomembranes 32 (2000) 97–104.
[21] P. Pacher, A.P. Thomas, G. Hajnóczky, Ca2+ marks: miniature calciumsignals in single mitochondria driven by ryanodine receptors, Proc. Natl.Acad. Sci. U. S. A. 99 (2002) 2380–2385.
[22] J. Huser, L.A. Blatter, S.S. Sheu, Mitochondrial calcium in heart cells:beat-to-beat oscillations or slow integration of cytosolic transients?J. Bioenerg. Biomembranes 32 (2000) 27–33.
[23] S. Rehncrona, L. Mela, B.K. Siesjo, Recovery of brain mitochondrialfunction in the rat after complete and incomplete cerebral ischemia, Stroke10 (1979) 437–446.
[24] K. Ohlendieck, I. Riesinger, V. Adams, J. Krause, D. Brdiczka, Enrichmentand biochemical characterization of boundary membrane contact sitesfrom rat-liver mitochondria, Biochim. Biophys. Acta 860 (1986) 672–689.
[25] D. Pette, Microphotometric measurement of initial maximum reaction ratesin quantitative enzyme histochemistry in situ, Histochem. J. 13 (1981)19–27.
[26] E.J. Schoenle, L.D. Adams, D.W. Sammons, Insulin-induced rapiddecrease of a major protein in fat cell plasma membranes, J. Biol. Chem.259 (1984) 12112–12116.
[27] M.L. Entman, T.R. Snow, D. Freed, A. Schwartz, Analysis of calciumbinding and release by canine cardiac relaxing system (sarcoplasmicreticulum). The use of specific inhibitors to construct a two-componentmodel for calcium binding and transport, J. Biol. Chem. 248 (1973)7762–7772.
[28] A.J. Lokuta, T.B. Rogers, W.J. Lederer, H.H. Valdivia, Modulation ofcardiac ryanodine receptors of swine and rabbit by a phosphorylation–dephosphorylation mechanism, J. Physiol. 487 (1995) 609–622.
[29] X. Zhu, J. Ghanta, J.W. Walker, P.D. Allen, H.H. Valdivia, The calmodulinbinding region of the skeletal ryanodine receptor acts as a self-modulatorydomain, Cell Calcium 35 (2004) 165–177.
[30] H.H. Valdivia, M.S. Kirby, W.J. Lederer, R. Coronado, Scorpion toxinsspecifically targeted against the calcium release channel of cardiac andskeletal muscle, Proc. Natl. Acad. Sci. U. S. A. 89 (1992) 12185–12189.
[31] H.B. Lee, L. Xu, G. Meissner, Reconstitution of the skeletal muscleryanodine receptor-Ca2+ release channel protein complex into proteolipo-somes, J. Biol. Chem. 269 (1994) 13305–13312.
[32] R. El-Hayek, A.J. Lokuta, C. Arevalo, H.H. Valdivia, Peptide probe ofryanodine receptor function. Imperatoxin A, a peptide from the venom ofthe scorpion Pandinus imperator, selectively activates skeletal-typeryanodine receptor isoforms, J. Biol. Chem. 270 (1995) 28696–28704.
[33] R.P. Xiao, H.H. Valdivia, K. Bogdanov, C. Valdivia, E.G. Lakatta, H.Cheng, The immunophilin FK506-binding protein modulates Ca2+ releasechannel closure in rat heart, J. Physiol. 500 (1997) 343–354.
[34] G. Meissner, E. Rios, A. Tripathy, D.A. Pasek, Regulation of skeletalmuscle Ca2+ release channel (ryanodine receptor) by Ca2+ and monovalentcations and anions, J. Biol. Chem. 272 (1997) 1628–1638.
[35] A. Tripathy, W. Resch, L. Xu, H.H. Valdivia, G. Meissner, Imperatoxin Ainduces subconductance states in Ca2+ release channels (ryanodinereceptors) of cardiac and skeletal muscle, J. Gen. Physiol. 111 (1998)679–690.
[36] M. Zoratti, I. Szabó, The mitochondrial permeability transition, Biochim.Biophys. Acta 1241 (1995) 139–176.
[37] M. Zoratti, I. Szabó, Electrophysiology of the inner mitochondrialmembrane, J. Bioenerg. Biomembranes 26 (1994) 543–553.
[38] B. O'Rourke, Mitochondrial ion channels, Annu. Rev. Physiol. 69 (2007)19–49.
[39] F.A. Lai, H. Erickson, B.A. Block, G. Meissner, Evidence for a junctionalfeet-ryanodine receptor complex from sarcoplasmic reticulum, Biochem.Biophys. Res. Commun. 143 (1987) 704–709.
[40] J.S. Smith, R. Coronado, G. Meissner, Single channel measurements of thecalcium release channel from skeletal muscle sarcoplasmic reticulum.Activation by Ca2+ and ATP and modulation by Mg2+, J. Gen. Physiol. 88(1986) 573–588.
[41] Q.Y. Liu, F.A. Lai, E. Rousseau, R.V. Jones, G. Meissner, Multipleconductance states of the purified calcium release channel complex fromskeletal sarcoplasmic reticulum, Biophys. J. 55 (1989) 415–424.
[42] J.S. Smith, T. Imagawa, J. Ma, M. Fill, K.P. Campbell, R. Coronado,Purified ryanodine receptor from rabbit skeletal muscle is the calcium-release channel of sarcoplasmic reticulum, J. Gen. Physiol. 92 (1988)1–26.
[43] Y. Ogawa, T. Murayama, N. Kurebayashi, Ryanodine receptor isoforms ofnon-mammalian skeletal muscle, Front. Biosci. 7 (2002) d1184–d1194.
[44] T. Nabhani, X. Zhu, I. Simeoni, V. Sorrentino, H.H. Valdivia, J. Garcia,Imperatoxin A enhances Ca2+ release in developing skeletal musclecontaining ryanodine receptor type 3, Biophys. J. 82 (2002) 1319–1328.
[45] A. Chu, M. Fill, E. Stefani, M.L. Entman, Cytoplasmic Ca2+ does notinhibit the cardiac muscle sarcoplasmic reticulum ryanodine receptor Ca2+
channel, although Ca2+-induced Ca2+ inactivation of CA2+ release isobserved in native vesicles, J. Membr. Biol. 135 (1993) 49–59.
[46] Y. Kirichok, G. Krapivinsky, D.E. Clapham, The mitochondrial calciumuniporter is a highly selective ion channel, Nature 427 (2004) 360–364.
[47] A. Ruck, M. Dolder, T. Wallimann, D. Brdiczka, Reconstituted adeninenucleotide translocase forms a channel for small molecules comparable tothe mitochondrial permeability transition pore, FEBS Lett. 426 (1998)97–101.
[48] N. Brustovetsky, M. Klingenberg, Mitochondrial ADP/ATP carrier can bereversibly converted into a large channel by Ca2+, Biochemistry 35 (1996)8483–8488.
[49] N. Brustovetsky, A. Becker, M. Klingenberg, E. Bamberg, Electricalcurrents associated with nucleotide transport by the reconstitutedmitochondrial ADP/ATP carrier, Proc. Natl. Acad. Sci. U. S. A. 93(1996) 664–668.
[50] C. Muro, S.M. Grigoriev, D. Pietkiewicz, K.W. Kinnally, M.L. Campo,Comparison of the TIM and TOM channel activities of the mitochondrialprotein import complexes, Biophys. J. 84 (2003) 2981–2989.
[51] G. Beutner, A. Ruck, B. Riede, D. Brdiczka, Complexes between porin,hexokinase, mitochondrial creatine kinase and adenylate translocatordisplay properties of the permeability transition pore. Implication forregulation of permeability transition by the kinases, Biochim. Biophys.Acta 1368 (1998) 7–18.
[52] M. Crompton, The mitochondrial permeability transition pore and its rolein cell death, Biochem. J. 341 (1999) 233–249.
[53] K.W. Kinnally, D.B. Zorov, Y.N. Antonenko, S.H. Snyder, M.W.McEnery,H. Tedeschi, Mitochondrial benzodiazepine receptor linked to innermembrane ion channels by nanomolar actions of ligands, Proc. Natl. Acad.Sci. U. S. A. 90 (1993) 1374–1378.
Copyright © 2022 FDOKUMEN




















