
The Miocene peat-forming vegetation of northwestern Germany: an analysis of wood remains and...
28
ELSEVIER Review of Palaeobotany and Palynology 104 (1999) 239–266 The Miocene peat-forming vegetation of northwestern Germany: an analysis of wood remains and comparison with previous palynological interpretations Isabel Figueiral a,b,L , Volker Mosbrugger a , Nick P. Rowe c , Abdul R. Ashraf a , Torsten Utescher d , Timothy P. Jones e a Institut fu ¨r Geologie und Pala ¨ontologie, Sigwartstr. 10, D-72076 Tu ¨bingen, Germany b UPRESA5059, CNRS, Institut de Botanique, 163, Rue A. Broussonet, 34090 Montpellier, France c Institut des Sciences de L’Evolution, UMR 5554, CNRS, Univ. Montpellier II, Place E. Bataillon, 34095 Montpellier, France d Institut fu ¨r Geologie, Nussallee 8, D-53115 Bonn, Germany e Department of Earth Sciences, University of Wales, Cardiff, CF1 3YE, UK Received 23 March 1998; accepted 26 August 1998 Abstract New methods of sampling, preparing and identifying fossil wood fragments preserved in browncoal are presented and used to aid in reconstructions of the vegetation from the Miocene Lower Rhine Basin (Germany). The technique involves laboratory charring wood specimens followed by routine identification with reflected light microscopy. Problems concerning the taxonomic identification of the fossil wood are discussed. A list of taxa is presented for the Garzweiler Seam and the implications for reconstructing the Miocene palaeoenvironment are discussed with reference to ecological comparisons of extant taxa: the nearest living relative approach. Results indicate a dominance of conifer wood types, mostly representing members of the Taxodiaceae and Cupressaceae; extensive sampling is required of both small (macroscopic) and large fragments to obtain results comparable with palynological assemblages. Up to now, twenty-one taxa of gymnosperms and angiosperms have been recognized from the Garzweiler seam. Data obtained are compared with those from pollen analysis and corroborate the previously identified trend consisting of increasing concentrations of Taxodium towards the upper section of the seam. 1999 Elsevier Science B.V. All rights reserved. Keywords: Lower Rhine Basin; Miocene; palaeoenvironment; fossil wood; charcoal; palynology 1. Introduction The Lower Rhine Basin is situated in the north- western headlands of the Rheinisches Schieferge- birge, in northwestern Germany. During the Tertiary, subsidence along NW- to SE-running faults formed a L Corresponding author. Fax: C33-467-543537; E-mail: [email protected] rift basin; this lead to transgression of the North Sea, during the Oligocene, and subsequent formation of an embayment open to the sea to the northwest (Scha ¨fer et al., 1996). During the Miocene, there were sev- eral marine transgressions, where higher sea levels resulted in the development of peat-forming coastal swamps and forests (Mosbrugger et al., 1994). Several palaeoecological studies have been car- ried out in this region. Miocene and Pliocene 0034-6667/99/$ – see front matter 1999 Elsevier Science B.V. All rights reserved. PII:S0034-6667(98)00059-1
-
Upload
independent -
Category
Documents
-
view
4 -
download
0
Transcript of The Miocene peat-forming vegetation of northwestern Germany: an analysis of wood remains and...

ELSEVIER Review of Palaeobotany and Palynology 104 (1999) 239–266
The Miocene peat-forming vegetation of northwestern Germany:an analysis of wood remains and comparison with previous
palynological interpretations
Isabel Figueiral a,b,Ł, Volker Mosbrugger a, Nick P. Rowe c, Abdul R. Ashraf a,Torsten Utescher d, Timothy P. Jones e
a Institut fur Geologie und Palaontologie, Sigwartstr. 10, D-72076 Tubingen, Germanyb UPRESA 5059, CNRS, Institut de Botanique, 163, Rue A. Broussonet, 34090 Montpellier, France
c Institut des Sciences de L’Evolution, UMR 5554, CNRS, Univ. Montpellier II, Place E. Bataillon, 34095 Montpellier, Franced Institut fur Geologie, Nussallee 8, D-53115 Bonn, Germany
e Department of Earth Sciences, University of Wales, Cardiff, CF1 3YE, UK
Received 23 March 1998; accepted 26 August 1998
Abstract
New methods of sampling, preparing and identifying fossil wood fragments preserved in browncoal are presentedand used to aid in reconstructions of the vegetation from the Miocene Lower Rhine Basin (Germany). The techniqueinvolves laboratory charring wood specimens followed by routine identification with reflected light microscopy. Problemsconcerning the taxonomic identification of the fossil wood are discussed. A list of taxa is presented for the GarzweilerSeam and the implications for reconstructing the Miocene palaeoenvironment are discussed with reference to ecologicalcomparisons of extant taxa: the nearest living relative approach. Results indicate a dominance of conifer wood types,mostly representing members of the Taxodiaceae and Cupressaceae; extensive sampling is required of both small(macroscopic) and large fragments to obtain results comparable with palynological assemblages. Up to now, twenty-onetaxa of gymnosperms and angiosperms have been recognized from the Garzweiler seam. Data obtained are comparedwith those from pollen analysis and corroborate the previously identified trend consisting of increasing concentrations ofTaxodium towards the upper section of the seam. 1999 Elsevier Science B.V. All rights reserved.
Keywords: Lower Rhine Basin; Miocene; palaeoenvironment; fossil wood; charcoal; palynology
1. Introduction
The Lower Rhine Basin is situated in the north-western headlands of the Rheinisches Schieferge-birge, in northwestern Germany. During the Tertiary,subsidence along NW- to SE-running faults formed a
Ł Corresponding author. Fax: C33-467-543537; E-mail:[email protected]
rift basin; this lead to transgression of the North Sea,during the Oligocene, and subsequent formation of anembayment open to the sea to the northwest (Schaferet al., 1996). During the Miocene, there were sev-eral marine transgressions, where higher sea levelsresulted in the development of peat-forming coastalswamps and forests (Mosbrugger et al., 1994).
Several palaeoecological studies have been car-ried out in this region. Miocene and Pliocene
0034-6667/99/$ – see front matter 1999 Elsevier Science B.V. All rights reserved.PII: S 0 0 3 4 - 6 6 6 7 ( 9 8 ) 0 0 0 5 9 - 1

240 I. Figueiral et al. / Review of Palaeobotany and Palynology 104 (1999) 239–266
plant remains (wood, roots, seeds, fruits) havebeen described (Kilpper, 1968a,b; Van der Burgh,1964, 1973, 1978a,b, 1984, 1987; Minnigerode andRichter, 1986; Pingen, 1990; Belz, 1992; Belz andMosbrugger, 1994; Mosbrugger et al., 1994; amongstothers), while floral successions have been based onthe analysis of pollen and spores (Utescher et al.,1992; Ashraf and Mosbrugger, 1995, 1996; Huhn etal., 1997).
Several estimates of the climate in this area, dur-ing the Miocene and Pliocene, have been attempted.Van der Burgh (1973) suggested a mean annual tem-perature of 11–16ºC for the Miocene, with a meanannual precipitation of at least 1000 mm, based on(a) the presence of palms, (b) the occurrence ofPicea=Larix on peat, (c) the presence of Sequoia, (d)frequencies of leaf impressions with entire marginsin floras of comparable age and (e) the climate ofareas of western Europe where similar species growat present. More recently, Van der Burgh (1984)suggested an equable climate with little seasonalvariation in temperature and precipitation. Accord-ing to Mosbrugger and Utescher (1997) the meanannual temperature might have reached its maximumduring the Middle Miocene (16–21ºC), followed bycooling trends during the Late Miocene (14–16ºC)and the Early Pliocene (13–14ºC). These climaticestimates are based on the coexistence approach ap-plied to the fruit and seed floras (Mosbrugger andUtescher, 1997). The palaeoclimatic calculations arederived from the climatic tolerances of the NearestLiving Relatives to the fossil plants.
Recent studies by Jones et al. (1996) suggest aclimate-related double cyclicity, observed from vari-ations in δ13C of the brown coal matrix through aprofile (Upper Seam, Upper Miocene) of 24.2 m.This study indicates a strong potential for correlatingfluctuations of stable carbon isotopes with changesin vegetation composition as observed from palyno-logical assemblages. Heavier isotopic compositionswere correlated with the presence of Sequoiapol-lenites polyformosus which is believed to representa species growing preferentially in humid climates.Following the palynological and isotopic descrip-tions of profiles through the browncoal, we havesampled wood remains preserved in browncoal (lig-nite), matrix and fossil charcoal, in an attempt tocorrelate identified taxa based on wood with the
patterns observed in the palynological and isotopeprofiles. Thorough and exhaustive sampling was in-tended to generate sufficient samples of wood taxato address the following questions: (1) are therepreservational biases on which taxa are present inthe browncoal matrix? (2) how is wood anatomypreserved, and is this preservation different betweenconifer and angiosperm wood? (3) Is it possibleto infer information concerning climatic conditionsbased on anatomical features preserved? (4) Doesthe inclusion of small fragments �5 mm in the woodidentification analysis, significantly improve the listof taxa? (5) How often do charcoal fragments oc-cur and are there taxa preferentially preserved ascharcoal as a result of wildfire? (6) How do theplant assemblages identified from the study of woodremains compare with those from palynology?
This paper aims to provide answers to these ques-tions as well as to describe the new ‘charcoalifi-cation’ method for the study and identification oflignite-rank fossil wood. We discuss the efficiencyof the method, present the results obtained so far,and comment on their palaeoenvironmental signifi-cance. A brief summary of palynological data is alsopresented.
2. Material and methods
The material studied was collected in the Ham-bach open cast mine, situated to the northwest ofthe Erft block, in the southern part of the LowerRhine Embayment (Fig. 1). The profile sampled in-cludes the Garzweiler Seam (horizon 6E in the localstratigraphy), which is split into units I, II and III.The study of this seam was particularly interesting aspalynological results were already available (Ashrafand Mosbrugger, 1995, 1996; Huhn et al., 1997).The precise age of this Seam is still debated; it ispossible that, at least, part of it was formed duringthe late Seravallian–early Tortonian, during the 2ndSciadopitys maximum identified from pollen analy-sis (Zagwijn and Hager, 1987). The Garzweiler Seamwas sampled vertically every 20 cm. The surface ofthe seam was first cleared of debris and weatheredmaterial. Fossil wood and charcoal samples alongwith matrix were hand-picked and chiselled manu-ally from the cleared vertical surface for up to 1

I. Figueiral et al. / Review of Palaeobotany and Palynology 104 (1999) 239–266 241
Fig. 1. Location of Garzweiler profile (6E) in the opencast mine at Hambach, Lower Rhine Embayment (Germany). *D wood sampling.
metre laterally and stored in plastic bags. The abun-dance of both large hand specimens and macroscopicfragments varied significantly through the profile.
A new method has been applied for the identifi-cation of fossil wood, which allows rapid processingand permits numerous, rapid identifications. Thismethod is currently used for identifying charcoal-ified wood from Quaternary environments; it hasbeen applied here on Tertiary fossil wood for the first
time as standard techniques of embedding and thin-sectioning are too time-consuming for the routineidentification of numerous samples.
Each fragment of fossil wood is wrapped in alu-minium foil and charcoalified in a laboratory furnaceat 400ºC, for 20–25 min. The temperature selectedminimizes the effects of volumetric shrinkage andthe formation of cracks, but nevertheless completelycharcoalifies the material (Slocum et al., 1978).

242 I. Figueiral et al. / Review of Palaeobotany and Palynology 104 (1999) 239–266
The specimens are left to cool slowly and thenfractured manually along the three anatomical planes(transversal, longitudinal tangential and longitudinalradial). Fractured parts are placed on the microscopestage in a small dish containing dried poppy (Pa-paver setigerum) seeds (Vernet, 1973). This providesan excellent medium for supporting and makingslight changes to the orientation of charcoal frag-ments for obtaining optimum focus (Figueiral, inpress). The fractured surfaces are viewed with in-cident light and the organization of the tissues andspecific details of cellular types can be viewed usingobjectives from ð5 toð80. Handling of fragments iscarried out with ‘flexible’ tweezers which can safelygrasp the specimens without shattering the charcoal.
This simple technique enables the identificationor ‘characterization’ of many wood samples in arelatively short time. Wood microstructure remainsmostly unchanged despite shrinkage and homoge-nization of cell walls during charcoalification, so thatimportant features such as pitting and ornamentationare retained. The changes observed during charcoal-ification of extant wood material appear to hold truefor the Miocene wood. According to Schweingruber(1978, p. 204), “The loss of 70–80% of the woodmass causes a shrinkage of 7–13% longitudinallyand 12–25% radially=tangentially. The cell wall isreduced to 1=5–1=4 of its original thickness”. Char-coalification also ‘cleans’ the fossil wood of fungalhyphae which often obscure anatomical characters(Plate I, 1 and 2).
The samples from the Garzweiler seam also in-cluded fragments of charcoal originating from wild-fire in the Miocene environment. Fossil charcoalcould be fractured and observed in exactly the sameway as the laboratory charred lignite. This facili-tated, especially, comparison of small-scale features
PLATE I
1. Transverse section of a fossil conifer illustrating the state of wood before charcoalification (scale bar D 16 µm).2. Transverse section of the same specimen from 1, after charcoalification. This process cleans the surface allowing the observation of
details which are otherwise obscured. Homogenisation of cell walls during charcoalification is also noticed (scale bar D 40 µm).3. ‘Chamaecyparis–Cupressus’, transverse section showing sucession of growth rings with similar width (scale bar D 95 µm).4. ‘Chamaecyparis–Cupressus’, cupressoid pitting observed in cross-fields. Uniseriate bordered pits are also observed in the tracheid
walls (scale bar D 19 µm).5. ‘Taxodium’, taxodioid pits (with large ovoid apertures wider than the border) in cross-fields (radial section) (scale bar D 19 µm).6. ‘Sciadopitys’, radial section. One large pit per cross-field, similar to those designated as ‘podocarpoid’ (Greguss, 1955). (scale barD
19 µm).
such as pitting and ornamentation which is otherwisemore difficult between different modes of preserva-tion and preparation.
Identification involves comparison of fossil mate-rial with wood atlases (Phillips, 1949; Metcalfe andChalk, 1950, 1983; Jacquiot, 1955; Greguss, 1955,1959; Jacquiot et al., 1973; Grosser, 1977; Detienneand Jacquet, 1983; Schweingruber, 1990; amongstothers) and, whenever possible, with reference col-lections of extant species (thin-sections of wood andcharred fragments). In our case, the reference col-lections included those of the ESA 5059 (CNRS)and of the Cirad-Foret in Montpellier (France). Thealteration and compaction of the wood samples hasmeant that automated means of identification viacomputerized programmes was not possible.
Wood identification has proved an extremely dif-ficult exercise because of problems associated withrelatively poor preservation of material and mostespecially, the varying degrees of compaction. Thelatter problem most seriously affects observation ofcharacters in the transverse plane with the major-ity of specimens showing significantly compressedtissues, particularly among angiosperms. Coniferwoods are less often compressed and it was of-ten possible to observe small sections of growthrings; this was rarely possible with fragments ofangiosperm wood.
In addition to the technical problems of preserva-tion, are the usual difficulties in attempting ‘iden-tifications’ based on fossil wood: developmentalvariations in wood anatomy within a single frag-ment; similarity=differences between wood anatomyof different genera and species; possible differencesbetween anatomical characters from fossil taxa andtheir putative and comparable modern analogues.In this study, which includes 708 fragments of

I. Figueiral et al. / Review of Palaeobotany and Palynology 104 (1999) 239–266 243
PLATE I

244 I. Figueiral et al. / Review of Palaeobotany and Palynology 104 (1999) 239–266
fossil wood and charcoal a conservative approachis employed when making identifications. Selectedanatomical characters are considered to distinguishdifferent ‘taxa’, which are later compared with char-acteristics present in modern species and with fossilwoods already described in the literature. Given thedifficulties of preservation and fragment size, po-tentially diagnostic characters that were taken intoaccount here, include:
Gymnosperms: presence=absence of resin ducts;presence=absence of tracheid wall thickenings; typeof cross-field pitting; intertracheid pitting and rayfeatures.
Angiosperms: size and distribution of vessels;presence=absence of spiral thickenings; type of per-foration plates; rays and parenchyma.
Another advantage of the charcoalification tech-nique is that the same specimen can be viewedunder SEM as well as reflected light with little extrapreparation. This is particularly useful for verifyingsmall-scale structures at the limit of resolution withthe incident light, or which require a larger depth offocus.
Descriptions presented here will be brief, and em-phasize only those features which enabled us to rec-ognize and characterize the different types of wood(TS D transverse section, LTS D longitudinal tan-gential section, LRS D longitudinal radial section).These different types may not correspond to fossilwood species previously identified by other authors,particularly concerning the identification of generabelonging to the Cupressaceae and Taxodiaceae. Thedifficulty in clearly distinguishing these two familieshas been discussed by Van der Burgh (1973). As-signment of our fossil material to one or the otheris based mostly on characters concerning number ofbordered pits in tracheids, tracheid width, majoritytype of cross-field pits and their size and distributionwithin each cross-field, ray height; these appear to bethe most significant characteristics, after observationof modern analogues.
Observation of contradictory anatomical features,in a single fragment, obliges us to be very careful.The number of specimens identified only to the levelof ‘conifer’ is therefore particularly high. Taxonomicclassification appears inside inverted commas to sig-nal the fact that identification of this type of materialshould always be considered as provisional.
The SEM micrographs were taken with a GEOLscanning electron microscope. In the following ac-count, and in further comparisons, reference willbe made, in particular to the important work of J.Van der Burgh on the Miocene of the Lower RhineEmbayment.
3. Qualitative results
Data obtained so far are based on the analysis of708 pieces of both fossil wood and charcoal (char-coal is relatively rare) from Seam Garzweiler III(lowermost), Garzweiler II (middle) and I (upperseam). Twenty-one different taxa have been distin-guished, covering fifteen families (Table 1). Most ofthe data were obtained from units II and I. SeamGarzweiler III yields only five fragments of fossilwood.
3.1. Gymnosperms
Family Cupressaceae
Three different taxa assigned to this family havebeen distinguished.
‘Chamaecyparis–Cupressus’ (Plate I, 3 and 4)
TS — growth rings present and distinct. Early=late wood transition abrupt.
LTS — rays uniseriate, rarely biseriate. Rays low,mostly 2–8, sometimes up to 12 cells high.
LRS — tracheid bordered pitting uniseriate, rarelybiseriate. Cross-field pitting mostly cupressoid, 1–3large pits per cross-field (mostly 1–2, one under theother), rarely taxodioid.
Anatomical features observed are similar to thosecharacterizing modern species of genera Cupressusand Chamaecyparis. Lack of cross-field pitting maybe observed in modern Chamaecyparis formosensisMast. (Greguss, 1955). The genus Juniperus wasconsidered initially but the nodular thickenings in theend walls of the parenchyma, often present in thisgenus, are not clearly observed in our specimens;this taxon is identified in both Garzweiler I and II.
Seeds of Cupressaceae, referred to as Cupresso-conus rhenanus Kilpper, have been identified by Vander Burgh (1987).

I. Figueiral et al. / Review of Palaeobotany and Palynology 104 (1999) 239–266 245
Table 1Taxa identified in Garzweiler Seam (units I, II and III) based on fossil wood remains and correlation with modern analogues
Correlation with Possible correlation with Garzweilerextant families extant genera=species
III II I
Taxodiaceae Taxodium (distichum) * *Taxodiaceae Taxodium ? *Cupressaceae Chamaecyparis=Cupressus * *Pinaceae Larix=Picea *Pinaceae Pinus(section Pinaster?) *Sciadopityaceae Sciadopitys verticillata * *Taxodiaceae Glyptostrobus pensilis ? * *Cupressaceae Actinostrobus=Juniperus=Cupressus *Cupressaceae Cupressus sempervirens *
Symplocaceae Symplocos * *Styracaceae Styrax * *Oleaceae Fraxinus *Oleaceae Linociera (D Chionanthus)=Olea *Myricaceae Myrica *Palmae *Hamamelidaceae *Angiosperm (Lauraceae?) *Fagaceae Quercus (deciduous) *Cornaceae *Lauraceae *Ericaceae Vaccinium *Ericaceae (?) *
Total of fragments 5 297 405
‘Actinostrobus–Cupressus–Juniperus’
Anatomical features are similar to those describedabove, except for the rays and cross-field pitting;Rays with tall marginal cells with 2–5 cupressoid=taxodioid pits distributed vertically.
A similar arrangement of pits in tall marginal cellsmay be observed in certain species of genera such asJuniperus (J. mexicana Schlechtendal, J. foetidissimaWill., J. distans Florin, J. conferta Parl., Cupressus(C. duclouxiana Hickel) and Actinostrobus (A. pyra-midalis Miquel).
Reference to this type of feature has not beenrecorded previously in fossil wood from the LowerRhine Embayment; fragments assigned to this taxonare identified in both Garzweiler I and II.
‘Cupressus’
Characters observed are very similar to those de-scribed above, except for the rays. In this case raysare high, up to, at least, 19 cells high. This feature
is observed in extant species of genus Cupressus.Species such as C. sempervirens L., and C. ducloux-iana Hickel have rays up to 20 cells high, while C.lusitanica v. Benthamii Carr., C. arizonica Greeneand C. torulosa Don. have rays exceeding 20 cellshigh. The single charcoal fragment identified comesfrom Garzweiler I.
Seeds of Cupressaceae, referred to as Cupresso-conus rhenanus Kilpper, have been identified by Vander Burgh (1987).
Family Taxodiaceae
‘Taxodium distichum’ (Plate I, 5)
TS — growth rings present and distinct.LTS — rays uniseriate (rarely biseriate) tall,
sometimes exceeding 25 cells high.LRS — ray tracheids absent. Multiseriate and
opposite, pitting present along the whole length oflarge tracheids. Cross-field pitting taxodioid (mostly)

246 I. Figueiral et al. / Review of Palaeobotany and Palynology 104 (1999) 239–266
to cupressoid, in horizontal rows of 2 to 6 pits.Prominent nodular end walls of parenchyma.
This taxon, present in both Garzweiler I and II isparticularly abundant towards the top of GarzweilerI. Features observed are similar to those of extantTaxodium distichum (L.) Richard. A probable fos-sil equivalent of Taxodium distichum has been sug-gested to be Taxodium dubium (Von Stenberg) Heer.Seeds and scales of this species have been frequentlyidentified in the Miocene browncoal sediments ofthe area (Van der Burgh, 1987; M. Pingen, pers.commun., 1997); this latter species has also beenidentified in the palynology (Ashraf and Mosbrug-ger, 1996).
A few specimens appear to show slight differ-ences from extant T. distichum in terms of pitting:the pits are more rounded than those previouslyobserved by us and there are only 1–3 pits, percross-field. We believe that these fragments belongto the same genus but most probably not to the samespecies and are thus referred to as ‘Taxodium’. Glyp-tostrobus may also possess almost circular pits butthis genus presents very distinctive bordered pits.
‘Glyptostrobus’
Anatomical features are close to those of the otherTaxodiaceae except for tracheid bordered pitting andcross-field pitting. The small size of bordered pits intracheids is particularly striking. Pits in cross-fieldsare also smaller than those from Taxodium and areof various types. Characters observed are similar tothose of modern Glyptostrobus pensilis (Staunton)K. Koch, and of fossil Glyptostroboxylon tenerum(Kraus) Conwentz (Van der Burgh, 1973); this taxonis present in both Garzweiler I and II.
Abundant cones and seeds of Glyptostrobus havebeen identified from lignite and sand deposits of theUpper Miocene in this area (Van der Burgh, 1987).
Similarities among genera and species of modernCupressaceae and Taxodiaceae have led specialiststo use the nominations Taxodioxylon, Juniperoxylonand Cupressinoxylon when referring to the differentgenera=species of these families preserved as fossils.
Wood species identified in the Lower Rhine Em-bayment include Juniperoxylon (Juniperoxylon ju-niperoides (Kownas) Huard, Juniperoxylon pachy-derma (Goppert) Krausel, Juniperoxylon rhenanum
Van der Burgh) Taxodioxylon (Taxodioxylon taxodiiGothan, Taxodioxylon gypsaceum (Goppert) Krausel,Taxodioxylon germanicum (Greguss), Taxodioxyloncryptomerioides Schonfeld.
Macrofossils preserved as compressions of Cu-pressus, Tetraclinis, Cryptomeria, Glyptostrobus, Se-quoia and Taxodium have also been identified in thebrowncoal of the Rhine Basin.
Family Sciadopityaceae
‘Sciadopitys’ (Plate I, 6)
TS — growth rings present but not very clear,always with abrupt transition of early=late wood.Latewood less than 1=4 width of growth ring.
LTS — rays very short, often only 2–3 cells high.LRS — tracheid bordered pitting mostly uniseri-
ate, rarely biseriate. Ray tracheids absent. Cross-fieldpits similar to those of Podocarpus.
Characters observed in the fragments present inGarzweiler I and II, are similar to those of extantSciadopitys verticillata. Fossil wood presenting thesame features has been previously identified in thisregion as Sciadopityoxylon wettsteinii Jurasky (Vander Burgh, 1973).
In Hambach, Sciadopitys stumps from an in situforest have also been identified by C. Gee (in Mos-brugger et al., 1994).
The taxonomic affinity of Sciadopitys has been amatter of debate for many years but recent phyloge-netic analyses have confirmed some recent authorssuggestions (e.g. Page, 1990) that the genus is notclosely related to taxodiacean conifers and that itis best considered as a monogeneric family, Sciado-pityaceae (Price et al., 1993; Price, 1995; Stefanovicet al., 1998).
Family Pinaceae
Two different elements can be identified withinthis family:
‘Larix=Picea’ (Plate II, 3 and 4)
TS — sections of growth rings observed do notshow vertical resin ducts.
LTS — rays uni-and multiseriate; one resin ductpresent in middle of multiseriate rays.

I. Figueiral et al. / Review of Palaeobotany and Palynology 104 (1999) 239–266 247
LRS — spiral thickenings in early wood tra-cheids; indentures in horizontal ray parenchyma;cross-field pitting piceoid to taxodioid.
The combined presence of resin ducts and spiralthickenings appears in a few extant species of Larix,Picea, Pseudotsuga and Cathaya. However, in thetwo last genera, the presence of spiral thickeningsis known in both early and latewood, which doesnot occur in these fossil specimens. We believe,therefore, that our specimens, identified in bothGarzweiler I and II, are closer to some species ofLarix=Picea. The distinction between modern Larixand Picea, based only on wood anatomy characters,has been hampered by the similarities observed inthese two genera. Some tendencies, however, havebeen noted (Schweingruber, 1990) concerning:
(1) heartwood colour: indistinct in Picea and red-dish in Larix;
(2) early-=latewood transition: gradual in Picea,generally abrupt in Larix;
(3) position of resin canals in rays: more-or-less inthe centre in Picea, more or less eccentric in Larix;
(4) number of rows of pits in radial section:mostly one in Picea, frequently two in Larix;
(5) type of ray tracheid bordered pits: more or lessangular in Picea, more or less smooth in Larix.
Other authors (Bartholin, 1979; Anagnost et al.,1994) consider that the type and proportion of raytracheid bordered pits (6) is the most reliable to dis-tinguish the two genera; four types of bordered pitscan be distinguished (distinct from Schweingruber,1990).
When attempting to apply all these considerationsto our material it became clear that: the heartwoodcolour, of course, can not be taken into account;compaction prevents us from unequivocally distin-guishing slight differences in early=latewood transi-tions; and a lack of statistically reliable observationsprevents us from using the third character identi-fied. The fourth feature is not conclusive, while thefifth=sixth could not be clearly observed in our spec-imens. Therefore, as a matter of caution we preferto use the nomination ‘Larix=Picea ’ (Larix beingmentioned first based on alphabetic order).
Several species of Piceoxylon have been distin-guished: P. arcticum (Schmalhausen) Krausel, P.rhenanum Van der Burgh, P. megapolitanum (Kobbe)Van der Burgh, P. schoenfeldii Van der Burgh and P.
pseudotsugae Gothan emend. Van der Burgh. Conesof Picea omorikoides Weber have also been identi-fied (Van der Burgh, 1973, 1987) from sand deposits.
‘Pinus’ (Plate II, 1 and 2)
TS — growth rings present and distinct; latewoodconspicuous; normal vertical resin ducts, small andpresent in some growth rings.
LTS — rays uni- and multiseriate; uniseriate raysare very low and 2–7 cells high; multiseriate rays areup to 12 cells high. Horizontal resin ducts with thinwalls, in multiseriate rays.
LRS — tracheid bordered pitting uniseriate.Ray tracheids present and with dentate walls; rayparenchyma with rather large simple (pinoid) pits.
Anatomical features are close to those of modernMediterranean pines such as Pinus pinaster Aitonand Pinus pinea L. Several species of Pinuxylonhave been identified from the lignite beds of theLower Rhine basin, based mostly on the structure ofwood and cones (Van der Burgh, 1964, 1973, 1987).These different species have been compared withmodern sections of Pinus:
Fossil wood Modern section
P. parryoides (Gothan) Krauselemend. Van der Burgh Parrya
P. tarnocziense (Tuzson) Greguss StrobusP. halepensoides Van der Burgh PineaP. pinastroides (Kraus)
Stockmans et Williere SulaP. paxii Krausel PinusP. eschweilerense Van der Burgh PinusP. taedioides Krausel PinasterP. arjuzanxianum Huard PinasterP. ponderosoides Van der Burgh Pinaster
Features observed in our fragments of Pinus,identified only in Garzweiler II, appear to be similarto those of pines belonging to the section Pinaster.
Other species of Pinus have been identified basedon the study of cones, seeds and scales from sanddeposits: Pinus leitzii Kirchheimer, Pinus spinosaHerbst, Pinus thomasiana (Goeppert) Reichenbachand Pinus uranii (Unger) Schimper (Van der Burgh,1987).

248 I. Figueiral et al. / Review of Palaeobotany and Palynology 104 (1999) 239–266
PLATE II

I. Figueiral et al. / Review of Palaeobotany and Palynology 104 (1999) 239–266 249
3.2. Angiosperms
3.2.1. Monocotyledons
Family Palmae (Plate II, 5)
TS — 3 or 4 large metaxylem vessels surroundedby parenchymatous sheath.
LRS — Metaxylem vessel walls with reticulateand annular thickenings.
Features observed are similar to those of modernpalms. The size of the fragments, identified in Garz-weiler II, did not permit a more accurate identification.
Two different species of palm have been describedby Van der Burgh (1984), based on the morphologyof stem compressions with petioles and leaves at-tached. The first species is Sabal maior Unger; asecond species is still unidentified. The fossil ma-terial was recovered from the sands overlying seamFrimmersdorf at the opencast mine of Zukunft West(Eschweiler). Stumps of Palmae from an in-situ for-est (at the top of the Frimmersdorf Seam) have alsobeen identified by C. Gee (Mosbrugger et al., 1994).
3.2.2. Dicotyledons
Family Cornaceae
TS — diffuse porous. Pores mostly isolated, ca.30–50 µm diameter.
LTS — rays 2–5 cells wide and at least up to 15cells high.
LRS — ray tissue heterogeneous; vessels withscalariform perforation plates with more than 20bars; large vessel element pitting.
Anatomical features observed lead us to includethis taxon in the family Cornaceae. It is recordedin the Garzweiler II Seam. Endocarps of Cornusbugloviana (Negru) Van der Burgh have been found
PLATE II
1. ‘Pinus’, transverse section showing the presence of vertical resin ducts in late wood of two growth rings. Variable width of growthring increments is also noticed, probably due to variable environmental conditions (scale bar D 190 µm).
2. ‘Pinus’, presence of ray tracheids with slightly dentate walls observed in radial section. Cross-fields with large pinoid pits (scale barD 19 µm).
3. ‘Larix=Picea’, the presence of spiral thickenings is noticed in the tracheid walls (radial section) (scale bar D 23 µm).4. ‘Larix=Picea’, small, abundant, piceoid to taxodioid pits observed in the cross-fields (scale bar D 19 µm).5. ‘Palmae’. Reticulate vessel element from the metaxylem (scale bar D 10 µm).6. ‘Vaccinium’, reticulate perforation plate observed in radial section (scale bar D 8 µm).
in the Miocene lignites of this area (Van der Burgh,1987)
Family Ericaceae
‘Vaccinium’ (Plate II, 6)
TS — diffuse porous wood; pores very small (lessthan 30 µm) and numerous, isolated or in smallgroups; diffuse parenchyma; rays quite indistinct anduniseriate.
LTS — not observed as fragment of charcoal toosmall.
LRS — scalariform perforation plates with up to10 bars; a single reticulate perforation plate was alsonoted in our specimen; ray cell walls densely pitted;rays heterogeneous.
Anatomical features observed in this Ericaceaeare very similar to those characterizing the ex-tant genus Vaccinium. The two fragments of char-coal assigned to this genus have been found in theGarzweiler II Seam. Fruits of Vaccinium miocenicumVan der Burgh have been identified by Van der Burgh(1987) from lignite and sand deposits.
Family: Ericaceae (?)
Anatomical features are similar to those describedabove, except for the size of the pores, which areeven smaller (less than 20 µm) and very abundant.Characters observed in transverse section are verysimilar to those characterizing modern Rhododen-dron. However, ray features are different.
Family Fagaceae
‘Quercus (deciduous)’ (Plate III, 1)
TS — pores of two sizes: early wood with largepores (more than 150 µm diameter) in sparse rows,

250 I. Figueiral et al. / Review of Palaeobotany and Palynology 104 (1999) 239–266
PLATE III

I. Figueiral et al. / Review of Palaeobotany and Palynology 104 (1999) 239–266 251
late wood with an oblique arrangement of muchsmaller pores; tyloses abundant in large vessels;parenchyma apotracheal and in tangential bands.
LTS — rays uniseriate, at least up to 8 cells high;multiseriate rays not observed.
LRS — ray tissue homogeneous; vessels withsimple perforation plates; large apertures of vessel-ray pits, often oval.
Anatomical characters closely resemble those ofa modern Quercus (deciduous). The fragments iden-tified come from the Garzweiler I seam. Cupules ofQuercus robur L., from the Garzweiler seam, havebeen identified by Van der Burgh (1987). However,multiseriate rays characteristic of modern oaks werenot observed in these samples which may be duemerely to the small size of the specimens. Otherpossibilities include Castanea with characters sim-ilar to those of deciduous oak, except for the rayswhich are never multiseriate. Confirmation will bepossible only when other similar fragments becomeavailable. It is interesting to note that Miocene andPliocene leaf remains of Quercus have also beenrecovered in the Lower Rhine Basin from clastichorizons (Belz, 1992; Belz and Mosbrugger, 1994).According to Van der Burgh (1993), leaves formerlyidentified as Quercus pseudocastanea and Quercusroburoides should be considered as the fossil coun-terparts of extant Quercus petraea. Fossil leaves ofthis species, recovered at Hambach, have been exam-ined for palaeoatmospheric signatures (Kurschner,1996).
Family Hamamelidaceae (Plate III, 2 and 3)
TS — not observed due to compression.LTS — rays high.LRS — ray tissue heterogeneous; large gash-bor-
PLATE III
1. ‘Quercus (deciduous)’, large pits in vessel to ray intersection, observed in radial section (scale bar D 10 µm).2. ‘Hamamelidaceae’, heterogenous ray tissue with large pits (scale bar D 48 µm).3. ‘Hamamelidaceae’, radial section — detail of rays showing large gash-like pits from ray to vessel and a scalariform perforation plate
(scale bar D 19 µm).4. ‘Lauraceae’, transverse section showing vessel distribution (solitary and in small multiples) (scale bar D 29 µm).5. ‘Lauraceae’, transverse section: detail of isolated pores with vasicentric parenchyma and sheath cells (oil or mucilage cells) in rays
(scale bar D 71 µm).6. ‘Lauraceae’ (?), ray pits in radial section (scale bar D 19 µm)7. ‘Lauraceae’ (?), detail of vessel to ray pits in radial section (scale barD 5 µm).
dered pits between rays and vessels; vessels withscalariform perforation plates.
The few characters observed are usually presentin modern species belonging to the family Hamamel-idaceae; this taxon has been identified in GarzweilerII.
The fossil record from the Lower Rhine Basincomprises several species belonging to this family.Fruits of Proserpinaca reticulata Reid et Reid havebeen distinguished, as well as seeds of Distyliumuralense Kolesnikova, Liquidambar magniloculataCzeczott et Skirgiello, and Corylopsis urselensisReid et Reid. Material assigned to these specieswas recovered from lignite and sand deposits.
Fossil wood identified as Liquidambaroxylonspeciosum Felix and Hamamelidoxylon rhenanumVan der Burgh has also been recorded (Van derBurgh, 1973).
Family Lauraceae (Plate III, 4 and 5)
TS — vessels solitary or in small multiples upto 70–80 µm diameter; parenchyma paratracheal,tending to be aliform.
LTS — rays biseriate, rarely triseriate, up to 20cells high; sheath cells (oil or mucilage cells) in rays.
LRS — ray tissue slightly heterogeneous; vesselswith simple perforation plates; ray-vessel pits withenlarged apertures.
Fragments of Lauracean wood have been identi-fied in Garzweiler I and II.
Fossil leaves and fruits of Lauraceae have beenidentified from the Tertiary of the Lower RhineEmbayment. Endocarps and fruits of Laurocarpumrecovered from sand and silt deposits from the topof the Indener Schichten have been identified by Vander Burgh (1987).

252 I. Figueiral et al. / Review of Palaeobotany and Palynology 104 (1999) 239–266
PLATE IV

I. Figueiral et al. / Review of Palaeobotany and Palynology 104 (1999) 239–266 253
Fossil wood identified as Laurinoxylon nectandri-oides Krausel et Schonfeld and Laurinoxylon see-mannianum Madel has also been recorded (Van derBurgh, 1973 and references therein). The familyLauraceae is not recorded in the pollen diagrams,due to poor preservation.
Our fragments present characters similar (in thetransverse section) to those of some species of mod-ern Cinnamomum.
Family Myricaceae
‘Myrica’ (Plate IV, 1 and 2)
TS — diffuse porous. Pores mostly isolated, ca.30–50 µm diameter
LTS — rays 1–3 cells wide and up to 15 cellshigh.
LRS — ray tissue heterogeneous.Vessels withscalariform perforation plates with few bars (up to 9in this case).
Characters observed in the single fragment anal-ysed (from Garzweiler II) are very similar to thosecharacterizing modern Myrica faya.
Several species of Myrica have been identifiedfrom the Rhine Embayment based on the morphol-ogy of endocarps:M. boveyana (Heer) Chandler,M.ceriferiformis Kownas, M. minima Negru and M.suppanii Kirchheimer (Van der Burgh, 1987). Sam-ples studied were collected in lignite layers, and insand and silt deposits.
Family Oleaceae
‘Fraxinus’
TS — ring porous wood; earlywood pores looselypacked; latewood pores solitary or in groups oftwo. Paratracheal parenchyma, vasicentric and con-fluent.
PLATE IV
1. ‘Myrica’, transverse section showing solitary pores (scale bar D 47 µm).2. ‘Myrica’, scalariform perforation plate with eight, thin bars (radial section) (scale bar D 8 µm).3. ‘Linociera (D Chionanthus)=Olea’, transverse section showing pores in short radial groups and paratracheal parenchyma (scale barD 74 µm).
4. ‘Linociera (D Chionanthus)=Olea’, transverse section — detail of group of pores and parenchyma (scale bar D 47 µm).5. ‘Linociera (D Chionanthus)=Olea’, tangential section showing biseriate ray (scale bar D 40 µm).6. ‘Linociera (D Chionanthus)= Olea’, heterogeneous ray tissue is observed in radial section (scale bar D 95 µm).
LTS — rays 1–2 cells wide, up to 30 cells high.LRS — rays homogeneous.Anatomical features observed in transverse sec-
tion are very similar to those observed in Fraxinusornus, i.e. early wood pores do not form a continuousring.
Fragments from this taxon have been identified inGarzweiler I.
The possible presence of Fraxinus at Hambachhas been suggested by pollen analysis (Ashraf andMosbrugger, 1996) as well as by the study of fruitsand seeds (M. Pingen, pers. commun., 1997).
‘Linociera (D Chionanthus)–Olea ’ (Plate IV, 3–6)
TS — diffuse porous. Pores isolated or in shortradial groups (mostly 2). Paratracheal parenchyma.Rays with short cells (square-shaped).
LTS — rays mostly biseriate.LRS — rays heterogeneous. Vessels with simple
perforation plates.Features observed are identical to those of modern
Olea europaea although the density of vessels islower, and also very similar to those of modernLinociera (D tropical Chionanthus). The presence oftaxa with anatomical characteristics similar to thosefound in our fragments has never been reported forthe Lower Rhine Embayment.
Features observed (such as size of vessels andray-width) might be closer to those characterizingthe genus Olea. The low density of vessels noticed inthe fossil material has also been observed in charcoalfragments from Spanish and French archaeologicalsites (Neolithic–Roman period) situated in the Me-somediterranean Bioclimatic Stage (Terral, 1997).At present, this bioclimatic stage is characterized byan average annual temperature of 13–17ºC, averagemaximum temperature of coldest months D 9–14ºC,average minimum temperature of coldest months D

254 I. Figueiral et al. / Review of Palaeobotany and Palynology 104 (1999) 239–266
�1 to 4ºC (Rivas-Martinez and Armaiz, 1984). How-ever, assignment of our specimen to one of the twogenera will need further confirmation. This taxon hasbeen identified only in Garzweiler II.
Endocarps of Chionanthus ruehii Gregor havebeen identified in samples from the lignites of theGarzweiler seam at Eschweiler by Van der Burgh(1987).
Family Symplocaceae
‘Symplocos’
TS — not observed due to compression.LTS — rays high and up to 5 cells wide.LRS — rays heterogeneous. Vessels with scalari-
form perforation plates, with at least up to 50 bars;bars ca. 2 µm thick and closely spaced.
Wood fragments identified as Symplocos arepresent in both Garzweiler I and II.
Several species of Symplocaceae have been iden-tified based on the morphology of endocarps: Sym-plocos germanica Mai, S. gothanii Kirchheimer,S. lignitarum (Quenstedt) Kirchheimer, S. minu-tula (Von Sternberg) Kirchheimer, S. pseudogregariaKirchheimer, S. salzhausenensis (Ludwig) Kirch-heimer and S. schereri (Kirchheimer). Material iden-tified comes from lignite, clay and sandy levels,formed during the Miocene and Pliocene.
Species identified on the basis of wood anatomyinclude Symplocoxylon latiporosum Van der Burgh,Symplocoxylon krauselii Van der Burgh and Sym-plocoxylon eschweilerense Van der Burgh. Featuresobserved in the longitudinal sections of our speci-men are similar to those described by Van der Burgh(1964, 1973) as Symplocoxylon latiporosum.
Family Styracaceae
‘Styrax’
TS — not observed.LTS — rays 2–4 cells wide.LRS — Slit-like vessel pits, vessels with rare
scalariform perforation plates, with few bars. Pres-ence or absence of other type of perforation platesmay not be verified due to bad preservation.
Characters observed in our specimen closely re-semble those described by Van der Burgh (1978b)
as genus Styrax. This taxon which is present in bothGarzweiler I and II, has not been recorded by pollenanalysis.
Endocarps, fruits and seeds of three species offamily Styracaceae have been identified from lig-nites, clay and sandy deposits: Halesia crassa (Reidet Reid) Kirchheimer, Rehderodendron ehrenbergii(Kirchheimer) Mai, and Styrax maximus (Weber)Kirchheimer (Van der Burgh, 1987).
Family Lauraceae (?) (Plate III, 6, 7)
TS — not observed due to compression.LTS — not observed due to compression.LRS — pits in vessel-ray intersections large; ves-
sels with scalariform perforation plates with thickbars.
Features observed in the radial section closelyresemble those of some Lauraceae. Further confir-mation is needed. The single fragment assigned tothis taxon has been identified in Garzweiler I.
Some final comments on the anatomy of the taxadistinguished, concern features which may provideinformation on the climatic conditions under whichthese plants were growing (Chaloner and Creber,1973; Creber and Chaloner, 1984; Baas and Schwe-ingruber, 1987; Wheeler and Baas, 1991). Althoughthe material is scarce and mostly compressed wehave been able to observe that:
(1) Conifers seem better preserved than an-giosperms.
(2) Conifer wood shows distinct growth rings(growth rings in Sciadopitys are less conspicuous butthis is characteristic of the genus) indicating seasonalvariations. Most Cupressaceae seem to present anabrupt transition from early- to latewood, with thelatter being less than 1=4 of the growth ring width;this seems to indicate a long growing season (PlateI, 3). This contrasts with the pine specimen whichshows a conspicuous band of late wood (Plate II, 1).Many of the specimens observed present a sequenceof analogous growth rings. However, other fragmentsshow clear differences in growth ring width, whichmight indicate punctuated or variable environmentalconditions.
(3) Eight out of the twelve taxa of angiospermwood distinguished possess scalariform perfora-tion plates, supposedly useful to trap embolisms

I. Figueiral et al. / Review of Palaeobotany and Palynology 104 (1999) 239–266 255
in thawing xylem sap (Zimmermann, 1983; Car-lquist, 1988). In the modern flora, the incidence ofscalariform perforation plates is believed to showecological trends. Abundance of scalariform perfo-ration plates are usually a sign of mesic environ-ments (Baas and Schweingruber, 1987) which seemsto agree with the palaeoenvironment data obtainedfor this area. However, this anatomical feature isalso particularly frequent in cool temperate to arctic,and tropical high montane floras (Baas, 1976, 1986;Wheeler and Baas, 1991) which is apparently notconsistent with the palaeoenvironment and palaeo-geographic data already obtained for this area.
4. Quantitative results
The quantitative results obtained so far are givenin Table 2.
The majority of wood remains consists ofgymnosperms (77%), mostly Taxodiaceae andCupressaceae. Amongst these ‘Taxodium’ and‘Chamaecyparis–Cupressus’ are the most frequentand abundant taxa. Frequencies of ‘Taxodium’ in-crease clearly at the top of Garzweiler I.
The presence of Sciadopitys is noted in both unitsof the seam.
Angiosperm wood is much less frequent; of the708 fragments identified, only 140 belong to An-giosperms (19.5%). Among these, Symplocos seemsto be the most frequent and abundant taxon, followedby Lauraceae and Styrax.
We would also like to draw attention to:(1) The high frequencies of fragments identified
only as ‘Conifer’, due to advanced compression.(2) The total absence of woody material at certain
levels of the seams, especially in Garzweiler II. Thisabsence is particularly noticeable from 5.60 to 6.60m.
(3) The presence of charcoalified fragments, re-covered from the following horizons: GarzweilerI — 0.20–0.40 m, 1.40–1.60 m, 2.80–3.00 m,5.80–6.00 m; Garzweiler II — 4.40–4.60 m, 9.60–9.80 m, 10.00–10.20 m, 10.20–10.40 m, 12.60–12.80 m, 13.20–13.50 m. Taxa affected by wild-fire include Chamaecyparis–Cupressus, Pinus, Glyp-tostrobus, Palmae, Quercus, Cornaceae, Vacciniumand Ericaceae (?).
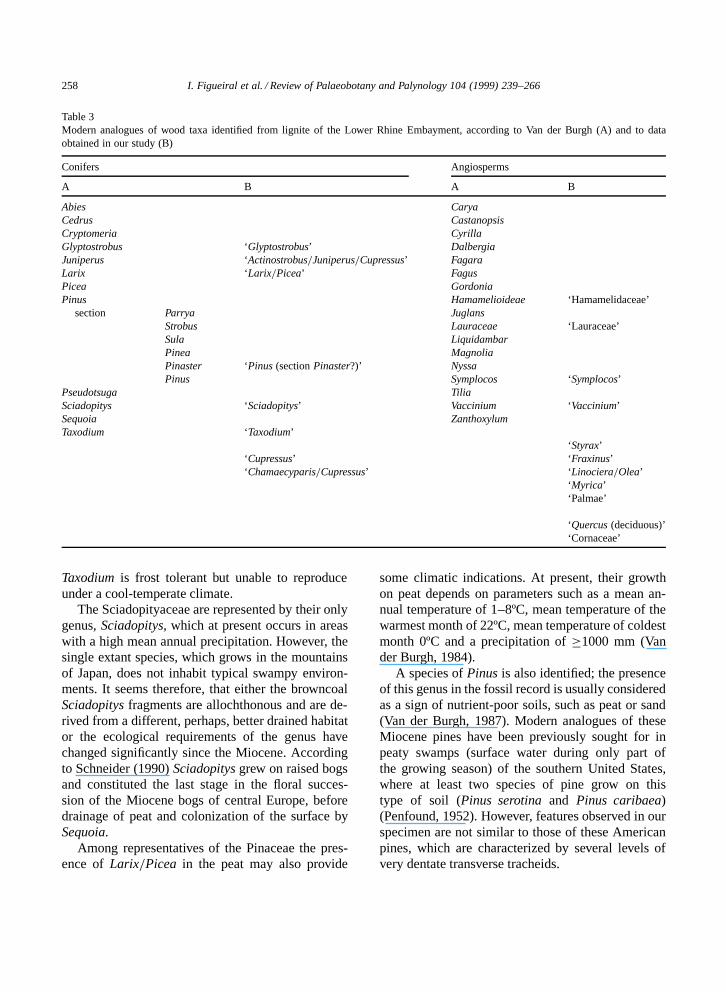
The floral assemblage is composed of a mixtureof evergreen coniferous and broadleaved (evergreenand deciduous) taxa. Some of which have not beenpreviously recorded as wood remains from the LowerRhine Embayment (Table 3).
It is interesting to note that Garzweiler II ap-pears to show a greater variety of taxa, particularlyAngiosperms, and this is observed from a smallersample set. Eighteen different taxa are identifiedin Garzweiler II for 297 fragments, while only 10taxa are recognized amongst the 405 fragments fromGarzweiler I (Table 1). This might reflect an increas-ingly lower plant diversity of the area towards the topof the seam. However, until further large sampling iscarried out this hypothesis cannot be confirmed.
5. Climatic implications of identified woody taxa
Some of the taxa identified may provide climaticinformation based on the climatic tolerances of mod-ern relatives of fossil plants (nearest living relativemethod) (Mosbrugger and Utescher, 1997). How-ever, this information must be considered approxi-mate as the ecological requirements=tolerances of cer-tain genera=species may have changed through time.Palaeoecological information presented here followMai (1964, 1995), Van der Burgh (1973, 1984, 1987),Schneider (1990) and Mosbrugger et al. (1994).
Among the conifer woods members of the familyTaxodiaceae generally indicate high and continu-ous levels of humidity. Modern Taxodium grows inmarshy environments, mostly in southeastern UnitedStates (coastal plains from Virginia southwards toLouisiana) (Penfound, 1952). Both Taxodium dis-tichum (southern cypress) and Taxodium ascendens(pond cypress) extend into the fresh water swamps(deep swamps), which are covered in water through-out most of the growing season (Penfound, 1952).
Some authors consider that Glyptostrobus growsexclusively in marshy habitats (Michaux et al.,1979). Schneider (1990) identifies Glyptostrobusswamp as an early stage in the floral succession ofMiocene bogs in central Europe. However, the authoralso notes that modern Glyptostrobus requires dryperiods for seed germination. During periods witha high water level, the plant forms pneumatophoresfor respiration. Other authors (Li, 1953; Tanai et al.,

256 I. Figueiral et al. / Review of Palaeobotany and Palynology 104 (1999) 239–266
Table 2Frequencies of taxa from Garzweiler Seam I (uppermost part), II (middle) and III (lower part). Unit II presents greater taxonomic varietythan unit I. High frequencies of ‘Taxodium’ and ‘Chamaecyparis–Cupressus ’ are noted. The number of specimens identified only to thelevel of ‘Conifer’ is high. Taxa are ordered according to their appearance during identification
1963) note that, at present, this tree grows in moistriver margins rather than in swampy areas. It is dis-tributed in the lowlands of the southern China (Tanai
et al., 1963; Ying et al., 1993) which are regionscharacterized by a warm-temperate climate. ModernGlyptostrobus is also frost sensitive, whereas extant

I. Figueiral et al. / Review of Palaeobotany and Palynology 104 (1999) 239–266 257
Table 2 (continued)
258 I. Figueiral et al. / Review of Palaeobotany and Palynology 104 (1999) 239–266
Table 3Modern analogues of wood taxa identified from lignite of the Lower Rhine Embayment, according to Van der Burgh (A) and to dataobtained in our study (B)
Conifers Angiosperms
A B A B
Abies CaryaCedrus CastanopsisCryptomeria CyrillaGlyptostrobus ‘Glyptostrobus’ DalbergiaJuniperus ‘Actinostrobus=Juniperus=Cupressus’ FagaraLarix ‘Larix=Picea’ FagusPicea GordoniaPinus Hamamelioideae ‘Hamamelidaceae’
section Parrya JuglansStrobus Lauraceae ‘Lauraceae’Sula LiquidambarPinea MagnoliaPinaster ‘Pinus (section Pinaster?)’ NyssaPinus Symplocos ‘Symplocos’
Pseudotsuga TiliaSciadopitys ‘Sciadopitys’ Vaccinium ‘Vaccinium’Sequoia ZanthoxylumTaxodium ‘Taxodium’
‘Styrax’‘Cupressus’ ‘Fraxinus’‘Chamaecyparis=Cupressus’ ‘Linociera=Olea’
‘Myrica’‘Palmae’
‘Quercus (deciduous)’‘Cornaceae’
Taxodium is frost tolerant but unable to reproduceunder a cool-temperate climate.
The Sciadopityaceae are represented by their onlygenus, Sciadopitys, which at present occurs in areaswith a high mean annual precipitation. However, thesingle extant species, which grows in the mountainsof Japan, does not inhabit typical swampy environ-ments. It seems therefore, that either the browncoalSciadopitys fragments are allochthonous and are de-rived from a different, perhaps, better drained habitator the ecological requirements of the genus havechanged significantly since the Miocene. Accordingto Schneider (1990) Sciadopitys grew on raised bogsand constituted the last stage in the floral succes-sion of the Miocene bogs of central Europe, beforedrainage of peat and colonization of the surface bySequoia.
Among representatives of the Pinaceae the pres-ence of Larix=Picea in the peat may also provide
some climatic indications. At present, their growthon peat depends on parameters such as a mean an-nual temperature of 1–8ºC, mean temperature of thewarmest month of 22ºC, mean temperature of coldestmonth 0ºC and a precipitation of ½1000 mm (Vander Burgh, 1984).
A species of Pinus is also identified; the presenceof this genus in the fossil record is usually consideredas a sign of nutrient-poor soils, such as peat or sand(Van der Burgh, 1987). Modern analogues of theseMiocene pines have been previously sought for inpeaty swamps (surface water during only part ofthe growing season) of the southern United States,where at least two species of pine grow on thistype of soil (Pinus serotina and Pinus caribaea)(Penfound, 1952). However, features observed in ourspecimen are not similar to those of these Americanpines, which are characterized by several levels ofvery dentate transverse tracheids.

I. Figueiral et al. / Review of Palaeobotany and Palynology 104 (1999) 239–266 259
Species belonging to the Cupressaceae, such asChamaecyparis thyoides (southern white cedar), alsogrow in these American peat swamps. Its presenceis noted both along the edges and in the shallowerareas of the deep fresh-water swamps. The genusChamaecyparis grows only in eastern Asia (threespecies in Taiwan and Japan) and North America(three species, one growing in the Atlantic areas,while the other two are distributed along the Pacificcoast). It might also be significant that, in Japan,Chamaecyparis often grows in association with Sci-adopitys (Li, 1953).
The presence of Cupressus usually suggests a dryand well drained habitat and does not seem to in-dicate the wet environment of a peat bog. However,some species of this genus also grow in moist habi-tats, along river banks or close to bays and coastlines (Li, 1953).
Modern analogues to the angiosperm woodsfound in this study may also be found. The singlemonocotyledonous stem fragment identified presentscharacters close to those of extant Palmae which de-velop under a climate with mean annual temperatureof ½12ºC, mean temperature coldest month of ½8ºC(Van der Burgh, 1984).
Among dicotyledonous taxa Symplocos is themost frequent; this is a large genus comprising ca.250 species growing in Asia, Oceania and Amer-ica (Nooteboom, 1975) which, as a whole, show acertain resistance to cold weather. Some species indi-cate permanent high groundwater table, while othersmay grow in upland forest.
Lauraceae are also relatively frequent; the fam-ily is composed of ca. 2000 species, which occurmostly in tropical or warm-temperate regions, in ar-eas with a moist and equable climate, and accordingto Cronquist (1981, p. 59) “...occupy no particularecological niche...”
Different species of Myrica may also representdifferent habitats such as marshy environments, peatbogs, upland forests and coniferous forests. In thefossil record its presence is usually considered asrepresenting acid, oligotrophic conditions, and asgrowing in the midst of the Taxodium dominatedvegetation of swampy areas (Van der Burgh, 1987).
Vaccinium is also regarded as a genus preferringwet conditions. In the Lower Rhine Embayment itspresence has been associated with acidic conifer-
ous forest and peat bog vegetation (Van der Burgh,1987).
The identification of woody taxa similar, in termsof anatomical characters, to modern Olea is some-what unexpected as this genus has not been reportedbefore in this area. However, it is also possiblethat the specimen might belong to genus Linociera(D Chionanthus). Although the genera Linocieraand Chionanthus were combined by Stearn (1976),their respective wood anatomies are not very similar.Genus Chionanthus is very diverse and two groupshave been distinguished (Baas et al., 1988): GroupA — tropical Chionanthus (from which most specieswere previously named Linociera) and Group B —temperate Chionanthus. According to these authors,Group A (the one that might interest us) is mostlycomposed of evergreen trees or shrubs growing inwet or dry forests at low to medium altitude.
Olea is a subtropical genus, consisting of 20species (Cronquist, 1981), growing normally in thesubtropical and mediterranean regions. In the north-ern hemisphere, it is present between 22 and 45ºN,while in the southern hemisphere it grows from theEquator to 37ºS (Baldy, 1990). Olea may grow nat-urally in regions characterized by very different pre-cipitation levels, from 200 to 1000 mm (Le Houerou,1959).
The Coexistence approach (Mosbrugger andUtescher, 1997) was also applied to the woodyfloras. The results obtained from the fossil woodassemblage from Garzweiler Seam II suggests amean annual temperature of 14.6–16.6ºC and thatfrom Garzweiler Seam I of 14.8–16.6ºC. Thesepalaeotemperatures are consistent with those derivedfrom fruits and seeds from the same horizon. Theyalso provide a reasonably narrow palaeotemperaturerange when considered in terms of the comparativelylow plant diversity combined with a high statisticalsignificance, as all the nearest living relatives of theidentified taxa may coexist under the same climaticconditions.
At least some of the taxa identified are able togrow in wet habitats, such as a bog, which arecurrently interpreted for the browncoal depositionenvironment. However, and based on the data avail-able, we believe that the possibility of punctual ortemporary drainage of the peat bog should also betaken into consideration. A partial and temporary

260 I. Figueiral et al. / Review of Palaeobotany and Palynology 104 (1999) 239–266
desiccation of the surface (or part of the surface)would allow (i) possibility of gaseous diffusion andthe presence of oxygen in the soil for the ger-mination of seeds of Glyptostrobus and Taxodium,(ii) colonization of poor soils by Pinus and Cu-pressus, (iii) development of herbaceous vegetation,as shown by palynology (Ashraf and Mosbrugger,1995, 1996), and (iv) occurrence of natural wildfiresas indicated by the presence of charcoal and transi-tion fossils lignite=charcoal in some horizons. Taxaaffected by fire include Chamaecyparis–Cupressus,Pinus, Glyptostrobus, Palmae, Quercus, Cornaceae,Vaccinium and Ericaceae (?). At present, little dam-age is caused by fire in fresh-water marshes whensurface water is present. However, during periodsof drought, fire may significantly affect the wholemarshy environment (Penfound, 1952; Cypert, 1972;Cohen, 1974).
6. Comparison with pollen analysis
Over the last 50 years the Tertiary pollen andspores of the Lower Rhine Embayment have beenstudied in some detail (see Ashraf and Mosbrugger,1995, for a summary). It is therefore necessary tocompare the results from our analysis of macrofossilremains with the interpretations derived from paly-nological investigations. We have concentrated onthe most recent palynological studies in which ourworking group was involved. Ashraf and Mosbrug-ger (1995, 1996) analysed and described the MiddleMiocene to Pliocene palynoflora based on a 270 mprofile from the open-cast mine Hambach and on a570 m core, about 7 km to the west of Hambach.Using multivariate statistical analysis of the paly-nological data from the Hambach profile, Huhn etal. (1997) provided a detailed reconstruction of thepalaeovegetation change from the Garzweiler Seam(Fig. 2). It is important to note that the profile sam-pled for palynology was situated approximately 1 kmto the west of the section from where our wood andcharcoal fragments were collected; in addition, thestandard sampling intervals used in the palynologicalstudies were also at 10 to 20 cm intervals, whichjustifies a comparison of macro remains with thepalynological data of Ashraf and Mosbrugger (1995,1996) and Huhn et al. (1997).
The palynological samples were rich in fossilmaterial. Families identified include:
Spores: Gleicheniaceae, Lycopodiaceae, Osmun-daceae, Polypodiaceae, Pteridaceae, Schizaeaceae,Selaginellaceae, Sphagnaceae.
Pollen: Gymnosperms — Cupressaceae, Pinaceae,Taxodiaceae, Sciadopityaceae, Ephedraceae;
Angiosperms (monocotyledons) — Cyperaceae,Gramineae=Poaceae, Palmae, Sparganiaceae;
Angiosperms (dicotyledons) — Aceraceae, Anac-ardiaceae, Aquifoliaceae, Araliaceae, Betulaceae,Clethraceae, Cyrillaceae, Compositae, Empetraceae,Ericaceae, Fagaceae, Hamamelidaceae, Juglan-daceae, Leguminosae, Magnoliaceae, Mastixiaceae,Myricaceae, Nymphaeaceae, Nyssaceae, Oleaceae,Salicaceae, Sapotaceae, Symplocaceae, Theligo-naceae, Tiliaceae, Ulmaceae, Vitaceae.
With 44 families (8 families of pteridophytes,5 families of gymnosperms, 31 families of an-giosperms) the palynoflora of Garzweiler Seam isclearly more diverse than the wood flora. There area number of reasons for the diversity difference be-tween the macro-and micro-fossil records. The ashcontent of the Garzweiler Seam is low, therefore onemight expect that transport of wood fragments insidethe peat area was minimal. In contrast, pollen andspores will readily be dispersed over large distancesby wind, insects, or water transport. Hence, the woodfragments of each sample will only reflect the verylocal vegetation, whereas a palynological sample ofthe same size will represent a larger catchment area.In addition, it comes as no surprise that herbaceousplants such as pteridophytes, Cyperaceae, Poaceae,Compositae or Nymphaeaceae, which are well doc-umented in the palynoflora, are missing from themacrofossil-lignite flora. Natural charcoal producedby wildfires probably represents the only means bywhich herbaceous plants can be preserved. Still,these two reasons are not sufficient to explain thedegree of diversity between the wood flora and thepalynoflora, in particular between the abundance ofgymnosperms and angiosperms. Further differencesin preservation potential might play a considerablerole.
The pollen record identified five families ofgymnosperms, out of which, four (Cupressaceae,Pinaceae, Taxodiaceae and Sciadopityaceae) are alsofound from wood remains (Table 4); it is only the

I.Figueiral
etal./R
eviewof
Palaeobotanyand
Palynology104
(1999)239–266
261
Fig. 2. Distribution of vegetation groups in the Garzweiler Seam, as shown by cluster analysis of pollen data (Huhn et al., 1997).

262 I. Figueiral et al. / Review of Palaeobotany and Palynology 104 (1999) 239–266
Table 4Taxa identified in Garzweiler Seam: comparison between wood remains and palynology (G D Garzweiler)
Wood Palynomorphs Wood Palynomorphs
Gymnosperms Angiosperms (dicotyledons)Cupressaceae Empetraceae GI
Cupressus sempervirens GI EricaceaeChamaecyparis=Cupressus GI, GII GI Erica GI, GII, GIIIActinost.=Juniper.=Cupress. GI Vaccinium GII GI, GII, GIII
Ephedraceae FagaceaeEphedra GI, GII Castanea GI, GII, GIII
Pinaceae Fagus GI, GIIAbies GII, GIII Quercus GII GI, GII, GIIILarix=Picea GII Trigonobalanus GI, GII, GIIIPicea GII Hamamelidaceae GIIPinus GII Liquidambar GI, GII, GIII
Taxodiaceae JuglandaceaeGlyptostrobus GI, GII, GIII(?) Carya GI, GII, GIIIGlyptostrobus lineatus GI Juglans GISequoia polyformosus GI, GII, GIII Platycarya GIISequoia sp., Cryptomeria GI Lauraceae GI(?), GII
Sciadopityaceae Leguminosae GI, GII, GIIISciadopitys verticillata GI, GII MagnoliaceaeSciadopitys GI,GII Liriodendron GII
Magnolia GI, GIIAngiosperms (monocotyledons) MastixiaceaeCyperaceae Mastixia GI, GII, GIII
Carex GI MyricaceaeGraminea–Poaceae GI Myrica GII GI, GII, GIIIPalmae GII GI Nymphaeaceae
Sabal, Pseudophoenix GI, GII Nuphar GIIPhoenix GI, GII Nyssaceae
Sparganiaceae Nyssa GI, GII, GIIITypha GI, GII Oleaceae
Fraxinus GI GI, GIIAngiosperms (dicotyledons) Linociera=Olea GIIAceraceae Salicaceae
Acer GI, GII Salix GIAnacardiaceae Sapotaceae GII
Rhus GI, GII, GIII StyracaceaeAquifoliaceae Styrax GI, GII
Ilex GI, GII, GIII SymplocaceaeAraliaceae Symplocos GI, GII GI, GII, GIII
Aralia GI, GII TheligonaceaeBetulaceae Theligonum GI
Betula GI, GII, GIII TiliaceaeCarpinus GI, GII, GIII Brownlowia GI, GIIAlnus GI, GII Ulmaceae
Clethraceae Celtis GI, GII, GIIIClethra GI, GII, GIII Ulmus GI, GII, GIII
Compositae Zelkova GI, GIICirsium, Asteroideae GI Vitaceae
Cornaceae GII Cissus GI, GIICyrillaceae, Clethrac. Rosaceae GI, GII, GIII Vitis GI, GII

I. Figueiral et al. / Review of Palaeobotany and Palynology 104 (1999) 239–266 263
family Ephedraceae which has not yet been foundin the fossil wood record. In contrast, out of thethirty-one angiosperm families identified in the pa-lynoflora, only seven are also represented by woodfragments (Table 4). Even, without considering typ-ical herbaceous families (e.g. Poaceae, Compositae)the angiosperms are clearly under-represented in thewood flora compared with the palynoflora (Table 4).This observation is not new and has been explainedin two ways (Van der Burgh, 1973; Von der Brelieand Wolf, 1981). One hypothesis stresses that thegeneral preservation potential of angiosperm woodis lower that of gymnosperms. Another hypothesisrelates to differences in growth habits and has beenput forward for all the previous studies of Tertiarywood floras, in which only larger pieces of woodwere collected; indeed many studies have concen-trated on large preserved trunks, and ignored thesmall fossil wood component entirely. In these casesthe angiosperms may be underrepresented becausethe gymnosperms predominantly formed larger treeswhereas the angiosperms were mostly restricted tomore shrubby growth forms (Van der Burgh, 1973)and therefore were impoverished in the wood macro-fossil record. With our sampling technique for themacro remains, however, we can rule out this latterhypothesis because both trees and shrubs have anequal potential of being sampled.
This study clearly supports the possibility that onaverage, angiosperm wood decays more rapidly thangymnosperm wood. Recently, a biochemical expla-nation for this diagenetic bias towards gymnospermwood has become available. As Hatcher and Clifford(1997) pointed out, lignin from angiosperms typi-cally decays into water soluble parts and hence israpidly destroyed whereas the decay of gymnospermlignin includes water-insoluble products.
On the other hand, it is interesting to note thatthe wood flora also contains some families, suchas Cornaceae, Lauraceae and Styracaceae, whichare not documented in the palynoflora. Also, withinOleaceae, neither Olea nor Linociera (D Chionan-thus) have been identified from pollen. Similarly,Glyptostrobus and Pinus are not unequivocally re-ported in the pollen record. Again, several reasonsmay be responsible for the absence of some taxa inthe palynoflora. Taphonomy presumably accountsfor the absence of Lauraceae pollen, which are
well known to be thin-walled and to decay eas-ily (Christophel, 1989). Certainly, problems of tax-onomy and identification also play an importantrole. For instance, the pollen type ‘Inaperturopol-lenites’, which is abundant in the Garzweiler Seam,may encompass pollen from Taxodium and Glyp-tostrobus, although there are good reasons to assumethat the pollen mostly belongs to Taxodium (Ashrafand Mosbrugger, 1996, p. 10). Similarly, the distinc-tion of Taxodium, Sequoia and Glyptostrobus basedon wood anatomy alone is difficult and differentopinions about the diagnostic characters still exist.
In summary, there are pronounced differences intaxonomic composition between the palynoflora andthe wood flora which may be largely explained by(1) differences in the catchment area, (2) differencesin the taphonomy and preservational potential and(3) by problems of taxonomic identification. Thereare also differences in the ecological interpretationof the wood and the palynoflora.
Based on their multivariate statistical analysis ofthe palynological data, Huhn et al. (1997) identified,for the Garzweiler Seam, six vegetation units with acharacteristic succession in time (Fig. 2). The domi-nance of Pinaceae–Ericaceae is noted at the base ofthe sequence, followed by an increase of Myrica andCyrilla. The spread of pollen interpreted as repre-senting Taxodium and Nyssa swamps occurs in theuppermost part of the sequence. According to theauthors, this sequence could reflect a rising ground-water table caused by a transgression phase. Thisinterpretation is sustained by a recent coal petrogra-phy analysis of the Garzweiler Seam (Weissenberger,1995). In our analysis of the wood flora (Table 2)a concentration of Taxodium towards the top of theseam is also recognizable and supports the palyno-logical interpretation. However, there also exist cleardifferences between the quantitative and ecologicaldata obtained from wood and pollen. From the abovediscussion it is already evident that an angiospermdominated peat vegetation, as is suggested from thepalynoflora, cannot be derived from wood macro-fossils. Moreover, the relatively high abundance of‘Pityosporites’ (Pinaceae) in the palynological dia-gram contrasts with the low frequencies of Pinaceaewoods. However, pollen of genera from this family iscommonly over represented in Tertiary and Quater-nary pollen spectra due to its high production and=or

264 I. Figueiral et al. / Review of Palaeobotany and Palynology 104 (1999) 239–266
long-distance dispersal. Finally the high frequenciesof ‘Sequoiapollenites’ (assigned to Sequoia) in thepalynological record contrast with its absence fromthe woody remains. Here again taxonomic problemsmay play a major role. The absence of Sequoiafrom the wood flora may simply reflect the fact thatwe failed to recognize features characterizing mod-ern Sequoia sempervirens. It is also possible, thatthe pollen type ‘Sequoiapollenites’ does not (or notonly) belong to Sequoia but (also) to other Taxo-diaceae (cf. Ashraf and Mosbrugger, 1996, pp. 11–12).
7. General conclusions
The data obtained show the potential of (1) thebrowncoal deposits as a source of large amounts offossil wood material in which anatomical charactersmay still be recognized and of (2) the new methodsused to collect and study material of this age. Theimportance of exhaustive sampling is made clearas some of the taxa (such as Myrica, Vaccinium,Cornaceae, Palmae, Ericaceae?) were only identifiedfrom very small pieces (ca. 2 mm), while others areidentified for the first time in this area.
Based on the observation of the available remains,the preservation of conifer woods is often signifi-cantly better than that of angiosperms, where severecompaction is observed in transverse and tangentialsections. In fact, angiosperm wood apparently decaysmore rapidly than gymnosperm wood. It appears thatlignin from angiosperms typically decays into watersoluble parts and is rapidly destroyed while the de-cay of gymnosperm lignin includes water-insolubleproducts (Hatcher and Clifford, 1997).
Charcoal fragments are recorded but, in somecases, are too small for unequivocal identification;their presence reflects the existence of wildfires andappears to indicate that fire affects certain taxa.
Palaeoenvironmental information may eventuallybe obtained based on anatomical features (such asgrowth rings, type of perforation plates) but a largeamount of material is needed in order to have statis-tically reliable results.
Comparison with palynology seems particu-larly interesting despite differences concerning theamount of material available. Similarities allow us
to confirm results, such as the peak in Taxodium,while differences need to be investigated. Althoughthe taxonomic list obtained based on the wood re-mains will always be very short when comparedwith palynological spectra, the results may prove tobe complementary as it includes taxa that are diffi-cult to trace by palynology due to low fossilizationpotential, such as thin exine, and low levels of pollenproduction.
Acknowledgements
This study is part of a research program supportedby the Deutsche Forschungsgemeinschaft (DFG) onthe distribution, palaeobotany, palaeoecology andisotopic composition of Miocene fossil wood andcharcoal from the Lower Rhine Embayment.
We would like to thank the board of Rheinbraunfor permission to work in Hambach and in particular,Dr. Von der Hocht for help and advice during fieldsampling. We also thank Dr. J. Hilton (Universityof Wales, Cardiff) for help in field collecting, Dr.P. Detienne (Cirad-Foret, Montpellier) for help con-cerning taxonomic identifications and Herr Hutte-mann (Universitat Tubingen) for help with the SEM.We also thank Prof. D. Ferguson and an anonymousreferee for their comments on the manuscript. This ispublication no. ISEM 98-104 UMR 5554 CNRS.
References
Anagnost, S.E., Meyer, R.W., De Zeeuw, C., 1994. Confirmationand significance of Bartholin’s method for the identification ofthe wood of Picea and Larix. IAWA J. 15 (2), 171–184.
Ashraf, A.R., Mosbrugger, V., 1995. Palynologie und Palynos-tratigraphie des Neogens der Niederrheinischen Bucht. Teil 1:Sporen. Palaeontographica B 235, 61–173.
Ashraf, A.R., Mosbrugger, V., 1996. Palynologie und Palynos-tratigraphie des Neogens der Niederrheinischen Bucht. Teil 2:Pollen. Palaeontographica B 241, 1–98.
Baas, P., 1976. Some functional and adaptive aspects of ves-sel member morphology. In: Baas, P., Bolton, A.J., Catling,D.M. (Eds.), Wood Structure in Biological and TechnologicalResearch. Leiden Bot. Ser. 3, 157–181.
Baas, P., 1986. Ecological patterns of xylem anatomy. In:Givnish, J. (Ed.), On the Economy of Plant Form and Func-tion. Cambridge Univ. Press, Cambridge, pp. 327–352.
Baas, P., Schweingruber, F.H., 1987. Ecological trends in thewood anatomy of trees, shrubs and climbers from Europe.

I. Figueiral et al. / Review of Palaeobotany and Palynology 104 (1999) 239–266 265
IAWA Bull. N. S. 8, 245–274.Baas, P., Esser, P.M., Van der Westen, M.E.T., Zandee, M., 1988.
Wood anatomy of the Oleaceae. IAWA Bull. N. S. 9, 103–182.Baldy, C., 1990. Le Climat de l’olivier (Olea europea L.) Ecol.
Mediterr. 16, 113–121.Bartholin, T., 1979. The Picea–Larix problem. IAWA Bull. N. S.
1, 68–70.Belz, G., 1992. Systematisch-palaookologische und palaok-
limatologische Analyse von Blattformen in Mio-=Pliozander Niederrheinischen Bucht (NW-Deutschland). Diss., Univ.Tubingen, 179 pp.
Belz, G., Mosbrugger, V., 1994. Systematisch-palaookologis-che und palaoklimatologische analyse von Blattformen inMio-=Pliozan der Niederrheinischen Bucht (NW-Deutschland).Palaeontographica B 233, 19–156.
Carlquist, S., 1988. Comparative Wood Anatomy. Systematic,Ecological, and Evolutionary Aspects of Dicotyledon Wood.Springer, Berlin, 436 pp.
Chaloner, W.G., Creber, C.T., 1973. Growth rings in fossil woodsas evidence of past climates. In: Tarling, D.H., Runcorn,S.K. (Eds.), Implications of Contimental Drift to the EarthSciences. Academic Press, London, pp. 425–437.
Christophel, D.C., 1989. Evolution of the Australian florathrough the Tertiary. In: Ehrendorfer, F. (Ed.), Woody plants— Evolution and Distribution since the Tertiary. Springer,Vienna, pp. 63–78.
Cohen, A.D., 1974. Evidence of fires in the ancient Evergladesand coastal swamps of Southern Florida. Miami Geol. Soc.Mem. 2, 213–218.
Creber, C.T., Chaloner, W.G., 1984. Climatic implications fromgrowth rings in fossil woods. In: Brenchley, P. (Ed.), Fossilsand Climate. Wiley & Sons, London, pp. 49–74.
Cronquist, A., 1981. An integrated System of Classification ofFlowering Plants. Columbia Univ. Press, New York, NY, 1261pp.
Cypert, E., 1972. Plant succession on burned areas in Okefeno-kee Swamp following the fires of 1954 and 1955. Proc. TallTimbers Fire Ecol. Conf. 12, 199–217.
Detienne, P., Jacquet, P., 1983. Atlas d’identification des boisde l’Amazonie et des regions voisines. Centre TechniqueForestier Tropical, Noguent-sur-Marne, 640 pp.
Figueiral, I., in press. Lignified and charcoalified fossil wood.In: Jones, T.P., Rowe, N.P. (Eds.), Fossil Plants and Spores,Modern Techniques. Spec. Publ. Geol. Soc. London.
Greguss, P., 1955. Identification of Living Gymnosperms on theBasis of Xylotomy. Akad. Kiado, Budapest, 263 pp.
Greguss, P., 1959. Holzanatomie der Europaischen Laubholzerund Straucher. Akad. Kiado, Budapest, 330 pp.
Grosser, D., 1977. Die Holzer Mitteleuropas. Ein mikropho-tographischer Lehratlas. Springer, Berlin, 208 pp.
Hatcher, P.G., Clifford, D.J., 1997. The organic geochemistry ofcoal: from plant materials to coal. Org. Geochem. 5–6, 251–274.
Huhn, B., Utescher, T., Ashraf, A.R., Mosbrugger, V., 1997. Thepeat-forming vegetation in the Middle Miocene Lower RhineEmbayment, an analysis based on palynological data. SFB350, Univ. Bonn, 12 pp.
Jacquiot, C., 1955. Atlas d’Anatomie des bois des Coniferes.Centre Technique du Bois, Paris, 2 vol.
Jacquiot, C., Trenard, Y., Dirol, D., 1973. Atlas d’anatomie desbois des Angiosperms. Centre Technique du Bois, Paris, 175pp.
Jones, T., Fortier, S.M., Mosbrugger, V., Roesler, J., Utescher, T.,1996. 13C=12C ratio double cyclicity in a Miocene browncoal:isotopic signals and orbital forcing. Terra Nova 9, 19–23.
Kilpper, K., 1968a. Koniferen aus den tertiaren Deckschichtendes Niederreinischen Hauptflozes, 2. Genus Pinus L. Palaeon-tographica B 123, 213–227.
Kilpper, K., 1968b. Koniferen aus den tertiaren Deckschichtendes Niederrheinischen Hauptflozes, 3: Taxodiaceae und Cu-pressaceae. Palaeontographica B 124, 102–111.
Kurschner, W.M., 1996. Leaf stomata as biosensors of palaeoat-mospheric CO2 levels. LPP Found. Utrecht LPP Contrib. Ser.5, 153 pp.
Le Houerou, H.N., 1959. Ecologie, Phytosociologie et Productiv-ite de l’olivier en Tunisie meridionale. CNRS Extr. Bull. Serv.Carte Phytogeogr. B IV (1), 7–72.
Li, H.-L., 1953. Present distribution and habitats of the Conifersand Taxads. Evolution 1952, 245–261.
Mai, D.H., 1964. Die Mastixioideen-Floren im Tertiar der Ober-lausitz. Palaontol. Abh. B II, 1–92.
Mai, D.H., 1995. Tertiare Vegetationsgeschichte Europas. Fis-cher, Jena, 691 pp.
Metcalfe, C.R., Chalk, L., 1950. Anatomy of the Dicotyledons.Clarendon Press, Oxford, 2 vol., 1500 pp.
Metcalfe, C.R., Chalk, L., 1983. Anatomy of the Dicotyledons,II. Clarendon Press, Oxford, 297 pp.
Michaux, J., Suc, J.-P., Vernet, J.-L., 1979. Climatic inferencefrom the history of the Taxodiaceae during the Pliocene andthe Early Pleistocene in Western Europe. Rev. Palaeobot. Pa-lynol. 27, 185–191.
Minnigerode, C., Richter, H.G., 1986. Holzanatomische Unter-suchungen fossiler wurzelholzer aus dem Miozan der Nieder-rheinische Bucht. Rev. Palaeobot. Palynol. 48, 253–268.
Mosbrugger, V., Utescher, T., 1997. The coexistence approach— a method for quantitative reconstructions of Tertiary ter-restrial palaeoclimate data using plant fossils. Palaeogeogr.,Palaeoclimatol., Palaeoecol. 134, 61–86.
Mosbrugger, V., Gee, C.T., Belz, G., Ashraf, A.R., 1994. Three-dimensional reconstruction of an in-situ Miocene peat forestfrom the Lower Rhine Embayment, northwestern Germany— new methods in palaeovegetation analysis. Palaeogeogr.,Palaeoclimatol., Palaeoecol. 110, 295–317.
Nooteboom, H.P., 1975. Revision of the Symplocaceae of theOld World, New Caledonia excepted. Univ. Press, Leiden, 335pp.
Page, C.N., 1990. Pinatae. In: Kubitzki, K. (Ed.), Pteridophytesand Gymnosperms. Springer, Berlin, vol. I, pp. 290–361.
Penfound, W.T., 1952. Southern swamps and marshes. Bot. Rev.18, 413–446.
Phillips, E.W.J., 1949. Identification of softwoods by their micro-scopic structure, 3rd ed. Forest Products Res. Bull. 22, H.M.Stationary Office, London, 1968.
Pingen, M., 1990. The fruit and seed flora of Kreuzau, Western

266 I. Figueiral et al. / Review of Palaeobotany and Palynology 104 (1999) 239–266
Germany (Miocene). In: Knobloch, E., Kvacek, Z. (Eds.),Proc. Symp. Paleofloristic and Paleoclimatic Changes in theCretaceous and Tertiary, 1989, Prague, pp. 213–214.
Price, R.A., 1995. Familial and generic classification of theconifers. Am. J. Bot. 82 (6 Suppl.), 110 pp.
Price, R.A., Thomas J., Strauss, H., Gadek, P.A., Quinn, C.J.,Palmer, J.D., 1993. Familial relationships of the conifers fromrbcL sequence data. Am. J. Bot. 80, 172 pp.
Rivas-Martinez, S., Armaiz, C., 1984. Bioclimatologia y Vegeta-cion en la Peninsula Iberica. Bull. Soc. Bot. Fr. 131, Actual.Bot. (2–4), 111–120.
Schafer, A., Hilger, D., Gross, G., von der Hocht, F., 1996.Cyclic sedimentation in Tertiary Lower-Rhine Basin (Ger-many) — the ‘Liegendrucken’ of the brown-coal open-castFortuna mine. Sediment. Geol. 103, 229–247.
Schneider, W., 1990. Floral successions in Miocene bogs of Cen-tral Europe. In: Knobloch, E., Kvacek, Z. (Eds.), Proc. Symp.Paleofloristc and Paleoclimatic changes in the Cretaceous andTertiary, Prague, 1989, pp. 205–212.
Schweingruber, F.H., 1978. Mikroskopische Holzanatomie.Zurcher AG, Zug, 226 pp.
Schweingruber, F.H., 1990. Anatomie europaischer Holzer. EinAtlas zur Bestimmung europaischer Baum-, Strauch- und Zw-ergstrauchholzer. Haupt, Stuttgart, 800 pp.
Slocum, D.H., McGuinnes Jr., E.A., Beall, F.C., 1978. Charcoalyield, shrinkage and density changes during carbonization ofOak and Hickory woods. Wood Sci. 11 (1), 42–47.
Stearn, W.T., 1976. Union of Chionanthus and Linociera(Oleaceae). Ann. Mo. Bot. Gard. 63, 355–357.
Stefanovic, S., Jager, M., Deutsch, J., Broutin, J., Masselot,M., 1998. Phylogenetic relationships of conifers inferred frompartial 28S rRNA gene sequences. Am. J. Bot. 85 (5), 688–697.
Tanai, T., Suzuki N., Huzioka, K., Matsuo, H., 1963. Tertiaryfloras of Japan. Miocene floras. Coll. Assoc. Commem. 80thAnniv. Geol. Survey Jpn., Tokyo, 262 pp.
Terral, J.F., 1997. La domestication de l’olivier (Olea eu-ropaea L.) en Mediterranee nord-occidentale: approche mor-phometrique et implications paleoclimatiques. These, Univ.Montpellier II, 2 vol.
Utescher, T., Ashraf, A.R., Mosbrugger, V., 1992. Zur Faziesen-twicklung im Neogen der Niederrheinischen Bucht. In: Ko-var-Eder, J. (Ed.), Palaeovegetational Development in Europe.
Mus. Nat. History, Vienna, pp. 235–243.Van der Burgh, J., 1964. Holzer der Niederrheinischen Braun-
kohlenformation. I. Acta Bot. Neerl. 13, 250–301.Van der Burgh, J., 1973. Holzer der Niederrheinischen Braun-
kohlenformation. II. Rev. Palaeobot. Palynol. 15, 73–275.Van der Burgh, J., 1978a. The Pliocene flora of Fortuna Garsdorf,
I Fruits and seeds of Angiosperms. Rev. Palaeobot. Palynol.26, 173–211.
Van der Burgh, J., 1978b. Holzer aus dem Pliozan der Nieder-rheinischen Bucht. Fortschr. Geol. Rheinl. Westfalen 28, 213–275.
Van der Burgh, J., 1984. Some palms in the Miocene of theLower Rhenish plain. Rev. Palaeobot. Palynol. 40, 359–374.
Van der Burgh, J., 1987. Miocene floras in the Lower Rhen-ish Basin and their ecological interpretation. Rev. Palaeobot.Palynol. 52, 299–366.
Van der Burgh, J., 1993. Oaks related to Quercus petraea fromthe Upper Tertiary of the Lower Rhenish Basin. Paleonto-graphica B 230, 195–201.
Vernet, J.L., 1973. Etude sur l’histoire de la vegetation dusud-est de la France au Quaternaire d’apres les charbons debois principalement. Paleobiol. Cont. 4 (1), 90 pp.
Von der Brelie, G., Wolf, M., 1981. Zur Petrographie hellerund dunkler Schichten im rheinischen Hauptbraunkohlenfloz.Fortschr. Geol. Rheinl. Westfalen 29, 95–163.
Weissenberger, R., 1995. Kohlenpetrographische Untersuchun-gen zur Faziesentwicklung in den Flozen Frimmersdorf undGarzweiler (Tagebau Hambach, Rheinisches Braunkohlenre-vier, Miozan). Dipl. Thesis, Univ. Tubingen, 108 pp.
Wheeler, E.A., Baas, P., 1991. A survey of the fossil record fordicotyledonous wood and its significance for evolutionary andecological wood anatomy. IAWA Bull. N. S. 12 (3), 275–332.
Ying, T.-S., Zhang, Y.-L., Boufford, D.E., 1993. The EndemicGenera of Seed Plants of China. Science Press, Beijing, 824pp.
Zagwijn, W.H., Hager, H., 1987. Correlation of continental andmarine deposits in the southeastern Netherlands and the LowerRhine district. Meded. Werkgroep Tertiair Kwartair Geol. 24,59–78.
Zimmermann, M.H., 1983. Xylem Structure and the Ascent ofSap. (Springer Series in Wood Science, 1.) Springer, Berlin,143 pp.




















