
THE METABOLISM OF ORGANIC ACIDS BI FSEUDQMONAS ...
82
THE METABOLISM OF ORGANIC ACIDS BI FSEUDQMONAS AERUGINOSA A.T.C. 9027 JUDITH FRANCES MDORE WHITAKER A THESIS SUBMITTED IN PARTIAL FULFILMENT OF THE REQUIREMENTS FOR THE DEGREE OF MASTER OF ARTS ia the Departments of Biology and Botany and Dairying We accept this thesis as conforming to the standard required for candidates for the degree of M&STER OF ARTS. Members tf£ the Department of Biology and Botany* Hemo<3i7s—or the i/epartment of Dairying. THE UNIVERSITY OF BRITISH COLUMBIA April, 1951.
-
Upload
khangminh22 -
Category
Documents
-
view
2 -
download
0
Transcript of THE METABOLISM OF ORGANIC ACIDS BI FSEUDQMONAS ...

THE METABOLISM OF ORGANIC ACIDS
BI FSEUDQMONAS AERUGINOSA A.T.C. 9027
JUDITH FRANCES MDORE WHITAKER
A THESIS SUBMITTED IN PARTIAL FULFILMENT OF
THE REQUIREMENTS FOR THE DEGREE OF
MASTER OF ARTS
i a the Departments
of
Biology and Botany
and
Dairying
We accept this thesis as conforming to the
standard required for candidates for the
degree of M&STER OF ARTS.
Members tf£ the Department of Biology and Botany*
Hemo<3i7s—or the i/epartment of Dairying.
THE UNIVERSITY OF BRITISH COLUMBIA
Ap r i l , 1951.

ABSTRACT
The present study has been concerned with the oxidation of
the 4-carbon dicarboxylic acids: succinate, fumarate and malate, in the
striot aerobe, Pseudomonas aeruginosa A.T.C. 9027 e -Hesting oells harvested
from a mineral medium in 'which any one of these compounds was the sole
carbon source, metabolized al l three acids immediately and at a constant
and rapid rate© This observation has suggested the operation of the
enzymatic inter-relationship:
Succinate f umarate<r— malate
It is very likely that the oxidation of these substrates follows the
familiar tricarboxylic acid cycle, which has already been shown to occur
in P. aeruginosa 9027*
Questionable quantitative data for the oxidation of the
dicarboxylic acids by dried cell preparations has been obtained. These
preparations did not require either cytochrome c or pyocyanine as a carrier
system for succinate oxidation. The optimum pH range of succinic dehydrogenase
in succinate-grown and acetate-grown dried cells lay between pH 7.0 and 8.0,
an observation which coinoides with values found for other species©
The effect of the two metabolic inhibitors, malonate and uranyl
nitrate, on the respiration of both resting and dried cells of P. aeruginosa
has been investigated. Malonate v/as not inhibitory in moderately high
concentration at pH 7.0, but when the pH was lowered, maximum inhibition
was effected by low concentrations. Apparently, permeability functions as
a governing factor in malonate inhibition, a suggestion which was borne
out in experiments with dried cells, in which the ce l l membrane has been
altered during the drying process. These cells were strongly inhibited
at pH 7.0 by low, concentrations of malonate* An interesting though

complicating feature of the malonata experiments with resting cells'
has "been that P. aeruginosa possesses an adaptive enzyme system for
metabolizing the inhibitor. The inhibitory effect of uranyl nitrate
on the oxidation of Krebs intermediates or of glucose varied with the
substrate - maximum inhibition being obtained with isocitrate oxidation,
and none with malate oxidation, while tho inhibition of other oxidations
f o i l between these two extremes. The use of this non-specific inhibitor
in metabolic studies on whole cells has been discussed and, on the basis
of the present and other studios, its use has been precluded*

AC KNCJl'/LEDG-EMEJHT
Without the unfailing assistance and encouragement
of Dr» J« J. R» Campbell, the enthusiastic interest
of Dean B. A. Eagles, and the financial support of
the Research Council of Ontario, this thesis could
never have been written*

TABLE OF CONTENTS
INTRODUCTION .. . 1
PART I WARBURG STUDIES WITH RESTING CELLS
Introduction 10 Methods
Bacteriological ».... 11 Chemical ...... ««e«.o«.........«.«.«............ 12
Experimental 14 Discussion ........................................» 19
PART II THE METABOLISM OF DRIED CELLS
Introduction •••.••••••.••••••••••..•••«««..•«.•««.. 20 Methods 21 Experimental ....................................... 23 Discussion •••• •.«•••.••...•.•. ••••...« 32
PART III INHIBITOR STUDIES WITH MALONATE AND URANYL NITRATE
Introduction ....................................... 34 Methods 36 Experimental......
A. Malonate Studies ..........«••.............. 38 B. Inhibition by Uranyl Nitrate •• 52
Discussion ......................................... 61
SUMMARY 64
65

- 1 -
DTTRODUCTION
Since carbohydrate is the main source of energy for liv i n g c e l l s ,
the mechanism of i t s degradation has been extensively investigated. In
general, carbohydrate breakdown occurs i n two phases: the anaerobic phase
which ends i n pyruvio acid formation, and the aerobic phase which involves
the u t i l i z a t i o n of pyruvate via tha oxidative pathway. Some aspects of
this second phase have been studied i n the present thesis.
The f i r s t attempts to formulate a scheme of carbohydrate oxidation
are found i n the papers of Thunberg (149),and, of Knoop (69). These workers
suggested that pyruvate was f i r s t converted to acetate, two molecules
of iriiich condensed to give succinate (the Thunberg-WIeland condensation).
The succinate was then cyclically re-converted to acetate, via fumarate,
malate, oxalacetate and pyruvate. The chief weakness of this scheme lay
in the complete lack of evidence in support of the condensation.
A similar scheme was proposed by Toeniessen and Brinkman (151),
except that the condensation involved two molecules of pyruvate, instead of
acetate, an unknown polymerization product being formed, which was then
transformed to succinic acid.
The f i r s t real evidenoe that organic acids function i n respiration
was contained i n the work of Szent-Gyorgyi and his associates (7,8,49), who
found thatsuocinate (or i t s four-oarbon oxidation products)) could stimulate
catalytically the respiration of pigeon breast muscle brei, and that this
oatalytic effect was inhibited by malonate. Szent-^Gyorgyi suggested that

the main, function of succinate, fumarate, malate and oxalacetate i n muscle
"is not to serve as fuel, but to serve as a catalyst j as a catalytic hydrogen
carrier between foodstuff (e.g. pyruvate) ard cytochrome."(147)• The
Szent-Gyorgyi scheme has met with considerable criticism* f i r s t , because i t
rests on the assumption that the succinate systaa is the. only one in animal
tissues which can act directly with the cytochromes, and, second, beoause
the carrier function of oxalacetate~=-^malate merely results i n the accumul
ation of reduced coenzyme I, with no effective transport of hydrogen taking
place.
Shortly after Szent-Gyorgyi had proposed this scheme, Krebs announced
the discovery that, i n addition to succinate, fumarate, malate and oxalacetate,
o6-ketog!utaric and c i t r i c acids also act catalytically on the respiration
of minced muscle, and that their effects are also inhibited by malonate.
On the basis of further investigations, Krebs ( 72) proposed that pyruvate
and oxalacetate might react together to form a 7-carbon compound, from which
citrate, "C-ketoglutarate, and the 4-carbon dicarboxylic acids would then be
re-formed. What had formerly been considered simply as a chain or series of
reactions now became in effect a cycle, the ' c i t r i c acid cycle'. The 7-carbon
condensation product (oxalocitraconio acid) has never been isolat ed, but
Krebs was able to demonstrate the formation of substantial amounts of c i t r i c
acid when pyruvate and oxalaoatata condensed in an anaerobic system (73, 75).
He therefore suggested that pyruvate condensed with oxalacetate to give
citrate, which was oxidized via cis-aconitate, isocitrate, oxalosuccinate,
oC -ketoglutarate and the 4-carbon dicarboxylic acids, with the resulting
oxalacetate becoming available to condense again with pyruvate araid to
repeat the cycle. On the basis of tracer studies, W0od et al.(l60) proposed

- 3 -
that ois-aconitate, and not citrate, i s the i n i t i a l condensation product.
They showed that isotopic carbon, introduced as carbon 'dioxide together
with pyruvate, led to the formation of <<-ketoglutarate which .contained
isotopic carbon only in the carboxyl group next to the ketone group; i f
°c-ketoglutarate was derived from a symmetrical molecule (citrate), the
fixed carbon would be equally distributed in the two carboxyl groups.
Therefore, they concluded that^-ketoglutarate arose from cis-aoonitate,
and that citrate was outside the cycle but in equilibrium with cis-aconitate
through the mediation of aconitase.
Recently, Ogston (103) on purely theoretical grounds, has suggested
that the asymmetrical, occurrence of an isotope in a product cannot be taken
as conclusive evidence against i t s having arisen from a symmetrical preoursor.
He maintains that an asymmetric enzyme (aconitase) can indeed distirg uish
between the identical groups (end carboxyls) of the compound (citrate) which
i t attacks. This conclusion i s based on the fact that enzyme and symmetrical
substrate combine at three points, and that two of the three combination
sites are catalytically different. Stern & Ochoa (l38) have now shown
that citrate is the i n i t i a l product of condensation between oxalacetate and
some "active" 2-carbon compound (derived from pyruvate, and not pyruvate
i t s e l f ) , the condensation being catalysed by condensing enzyme, Coenzyme A,
Hg*" or MET* and adenosine triphosphate. The "active" 2-carbon compound may
be acetic acid or the acetyl radical or possibly acetyl phosphate (55).
By a series of hydrations, dehydrpgenations, and decarboxylations,
citrate i s dissimilated in turn to the following compounds; cis-aconitate,

isocitrata, oxalosuccinate,<<-ketoglutarate, succinate, fumarate, malate
and thence back to oxalacetate. The modern concept of the tricarboxylic
acid cycle is shown in Figure I. Thescheme from oxalacetate back to
succinate corresponds to the catalytic system of Szent-Gyorgyio For each
complete cycle, one molecule of acetate is completely oxidized to carbon
dioxide and wab er. A l l the steps in the cycle may be reversible under
aerobic conditions, but the only hydrogenations which appear to proceed
with any rapidity under these conditions are those connecting the
Szent-GyBrgyi system from oxalacetate to succinate.
The experimental work upon which the Krebs cyole is based was
performed largely with minced pigeon breast muscle (72,73,75). Ochoa
(99, 100,101,140), Wood (158), Barron (126), Vennesland, Peters and
others have demonstrated and discussed the occurrence of the various
reactions in other animal tissue, including kidney, heart and liver.
D.S. Green et a l . (50, 51) have obtained a preparation, the "cyolophorase
system"', which they have defined as a " c omplex of enzymes which catalyses
the oomplete oxidation of pyruvic acid by way of the Krebs oitrio acid
cycle." Microscopic examination of cyolophorase has indicated that i t
is essentially a suspension of intaot mitoohondria, and the enzyme activity
has been shown to be related to the structure of these maoro-granules (141).
- Evidenoe that pyruvate is oxidized through the tricarboxylic acid
cycle in plant tissues comes from three sources: (a) al l the acids which
are intermediates in the Krebs cycle are present inyplantsj (b) a number
of the enzymes catalyzing the reactions of the cycle have been found; (c) the few investigations of pyruvate oxidation in vivo (15,19,21,78) have
presented evidence conforming with the four criteria originally set forth

-2H (CHg.COOH) CH3.CO' (CH3.C0~P04)
COOH.CO.CHg.COOH Oxalacetate
- 2H
COOH.C(OH).CH2.COOH CHg.COOH
Citrate H20
C00H.C.CH2.C00H CH.COOH
+ H20
Cis-aconitate
V COOH.CH.CHg.COOH
CH(OH).COOH
- 2H
Isocitrate
COOH.CH.CHg.COOH CO.COOH
Oxalosucc inate
COOH.CHOH.CH2.COOH
A Malate
COOH.CH:CH.COOH A
Fumarate
-2H
COOH.CHg.CHg.COOH
Succinate -2H -CO2 +H20
CH2.CH2.COOH CO.COOH
od-ketoglutarate
Figure I. The tricarboxylic acid cycle

by Krebs in support of the cycle (72:) • The general picture of the Krebs
cycle in plant tissue has been reviewed by Bonner (20) and by Goddard &
Meeuse(48)»
Scattered information has appeared during the past five years
which indicates that the Krebs cycle is present in microorganisms, Ajl &
Workman (4,5) in studies on the replacement of oarbon dioxide by various
intermediates of the cycle^ suggested that at least parts of the scheme were
operative in Escherichia ooli and Aerobaoter aerogenes, Baskett & Hinshelwood
(16,17) on the baas of growth experiments, have proposed that the cycle oocurs
in Bacterium lactis aerogenes. However, Karlsson & Barker (62) were not
able to demonstrate the presence of this system in Azotobacter agilis, either
by the teohnique of simultaneous adaptation or by radioactive traoer exper
iments, Umbreit (l53),in a review of this data, suggested that part of the
cycle is present, while Stern & Oohoa (139) using extracts of Barker's
organism obtained the condensation of oxalaoetate with "active " aoetate
to citrate. They were able to showithat E, ooli (139) and Streptococcus
faecalis (70) also possess this condensation enzyme. Indirect evidence for
the oxalacetate-"active" acetate condensation in B,coli was given by
Oginsky et a l . (102) Oand by Umbreit (154) in studies on the bactericidal
effects of streptomycin. Further evidenoe for the participation of acetate
in the cycle has been provided by the action of fluoroacetate in inhibiting
the conversion of aoetate to citrate in yeast and baoteria (59)• Hovelli &
Lipmann (96) have found acetyl phosphate to be more effective than acetate
plus adenosine triphosphate in the formation of oitrate by extracts of
E, coli. Using radioactive carbon as a traoer, Saz & Krampitz (128)

- 7 -
demonstrated the same condensation in a lysate of Micrococcus lysodeikticus.
Campbell & Stokes (29) have presented evidence for the tricarboxylic acid
cycle in cells of gseudoraonas aeruginosa, harvested from either a glucose
or an aoetate medium* Several years ago, I»ockwood & Stodola (84) found that
P, fluoresoens produced considerable quantities of<<-ketoglutarate, one of °^
the key intermediates in the cycle, when cells were grown on a glucose
medium under conditions of intense aeration*
In addition to the condensation enzyme, bacterial preparations
have been made of the following enzymes in the Krebs oycle: succinic
dehydrogenase from Corynebaoterium diphtheriae (106), Salmonella aertryoke
(77) and B,ooli (2,57); fumarase from Propionibacterium Shermanii (74);
malio dehydrogenase from a stenothermophilio bacterium (87); oxalaoetio
decarboxylase from M, lysodeiktious (7l), and Azotobaoter vinelandii (108);
pyruvic oxidase from Proteus vulgaris (145)•
Barron et a l , (13) have investigated the occurrence of the Krebs
cycle in yeast, and have given a comprehensive review of previous work* The
cycle may also be operative in the metabolism of molds suoh as Aspergillus
niger (30), Neurospora orassa (82) and Streptomyees species (31), Working
with protozoans, Speck et_ a l , found the Krebs cycle present in the malaria
parasite, Plasmodium gallinaoeum (136), while Harvey obtained no evidence
for its occurrence in Trypanosoma hippicum (56),
Some reoent investigations of acetate oxidation have suggested
that an alternative scheme to the tricarboxylic acid oycle may function in
oertain microorganisms. Entrance into this cycle, the dicarboxylic aoid
cycle (12,42) is made by the Thunberg-lfieland reaction in which two moles
of acetate (possibly phosphorylated acetate) condense to form succinate, or
by the Wood-Werkman reaction of carbon dioxide fixation by pyruvate*

- 8 -
Using radioactive carbon as a tracer, Slade & Werkman (132) and
Kalnitsky et_ al , (60) have obtained evidence for the condensab ion of two
2-carbon molecules to succinic acid, with cell-free preparations of
Aerobacter indologenes ,and B.ooli respectively. Further swpport for this
scheme of acetate oxidation has been given by Ajl (3), working with
A, aerogenes and B.ooli, and by Lenti, al so employing B.ooli (81)• Ten
years ago, Wood & Workman (159) suggested that the Thunberg-Wieiand
condensation might occur in the organism, Propionibaoteriurn pentosaoeum,
but Krebs & Eggleston (74) rejected suoh a possibility in the allied species,
P» Shermanii, On the basis of inhibitor and adaptation experiments,
Barron et a l . (12) have concluded that the dicarboxylic acid oycle, and not
the tricarboxylic acid cycle, is operative in Corynebacterium oreatinovorans.
Foster & Carson (42) in studies on the mechanism of fumaric acid formation
by Rhizopus nigricans observed the four steps fitting the pattern of a
dicarboxylic acid oycle, entrance to the cycle being made by the Thunberg-
Wieiand condensation^
The experiments of Thimann, Bonner, Albaum and others (6,148),
on the action of iodoacetate and organic acids on the growth and proto
plasmic streaming in Avena coleoptiles and whole seedlings, have suggested
that "a respiratory process, which accounts for only a small fraction of
the total respiration, and which involves the 4-carbon acids, is in conb rol
of growth. This particular respiratory process is the one promoted by
auxin," (148). Similarly, Ryan et a l , (127) deduced that a respiratory
process involving the 4-carbon dicarboxylic acids "supplies energy for
growth"' in Heurospora.

r Investigations carried out in this laboratory have shown that
Pseudomonas aeruginosa A.T.C. 9027 oxidizes glucose by way of gluconio
and 2-ketogluconic acids (95,142) and that i t does not possess an Embden-
Meyerhof scheme of metabolism (27)• Both pyruvate and acetate have been
isolated as intermediates in glucose breakdown (28,157). Recently, Campbell
& Stokes (29) presented evidence for the presence cf a tricarboxylic acid
oycle* It is therefore probable that glucose is broken down by the
following pathway:-
Glucose v Gluconate v ^ 2-ketogluconate — — .pyruvate ^acetate
The intermediary steps between 2-ketogluconate and pyruvate may involve a
stepwise degradation of Cg to Cg, C4 and Cg, with alternate phosphorylative
oxidation and decarboxylation); as proposed originally, by Dickens ( 3 6 , 3 . 7 ) .
It is likely that the C3 compound is pyruvate, whioh undergoes oxidative
decarboxylation to "active" acetate, at which point the Krebs cycle is
entered.
The present study has centred about the oxidation of the 4-carbon
dioarboxylio acids, succinate, fumarate and malate, a l l well-established
constituents of the tricarboxylic acid cycle. By using whole resting oells
and dried cell preparations of P. aeruginosa, the interrelationship:
Succinate Fumarate . _ Malate
has been demonstrated. Strong evidence for the occurrence of the enzyme
succinic dehydrogenase, which catalyses the reversible step between succinate
and fumarate, has been obtained in experiments with the competitive inhibitor,
malonate.

- 10 -
PART I
WARBURG STUDIES WITH RESTING CELLS
Numerous studies i n bacterial chemistry have been made with
"resting"' cells* The bacteria are grown in pure culture, centrifuged
and washed, and suspended i n water or saline in a concentration greater
than could be achieved by growth* According to Quastel & Whetham (117),
these "resting" 1 baoteria are f u l l y endowed with metabolic potentialities*
yet unable to multiply because of the lack of essential growth nutrients*
Cells i n this non-proliferating state are well suited for use i n metabolic
studies, as their respiratory mechanisms may be evaluated in the absence
of interfering growth reactions (91).
The pathway of glucose oxidation in P* aeruginosa as now
recognized was arrived at on the basis of resting c e l l studies (91,92)*
Campbell & Stokes (29) used both glucose- and acetate-grown cells to
obtain evidence for a tricarboxylic acid cycle in the same organism*
In the present study, cells have been grown with succinate,
fumarate, malate or acetate as the sole carbon source, end the metabolic
patterns of the c e l l s investigated, with particular reference to the
oxidation of the dicarboxylic acids*

- 11 -
METHODS
Bacteriological: The culture of P. aeruginosa A.T.C. 9027 employed v/as a
typical strongly pigmenting strain. Stocks were maintained in a liver
extract agar of the following composition: 1.0$ tryptone, 0.25$ K2HP04,
0.15& glucose, 0.3$ CaC03, 10.0$ liver extract, 1.5$ agar, pH 7.2. After
growth was initiated at 30° C, cultures were refrigerated. Fresh transfers
of the stock culture were made every month.
Medium for growth of the organism was prepared as follows:
0.3$ (NH4)H2P04, 0.3$ K2HP04, and 0.5 p.p.m. iron (as FeS04.4H20) were
adjusted to pH 6.8, dispensed in 100 ml. quantities in Roux flasks, and
sterilized by autoclaving at 15 lb. pressure for 15 min. Before flasks
were inoculated, the carbon source that had been sterilized through
sintered glass as a 10$ solution was added aseptically to a final conc
entration of 0.3$ to 0.5$, and MgS04.7H20 that had been sterilized by
autoclaving as a 10$ solution was added to a final concentration of 0.1$.
When a dicarboxylic aoid was used as carbon souroe, i t was neutralized to
pH 6.5 with N/l HaOH before being made up to volume.
A 1$ inoculum of a 24 hr. culture, grown in ammonium phosphate
medium with appropriate carbon souroe, was employed. After subculture
from the refrigerated stock into Sullivan's medium (146), the culture was
always transferred at least twice at 24 hr. intervals in ammonium phosphate
medium with appropriate carbon source, before use as inoculum.
The cells for Warburg experiments were harvested after 21 to 24
hours incubation at 30° C , washed with half the growth volume of 0.9$
saline and finally made up to the desired concentration. This concentratio n
of oells gave a light transmission of 65$ to 75$ when 0.2 ml. of the

- 12 -
suspension was mad© upr.to 10.0 ml., and read in the Fisher Electrophotometer.
It is equivalent to 0.30 mg. bacterial nitrogen per ml. (92).
A conventional Warburg apparatus (155) was used to follow the
oxygen uptake, al l experiments being run at 31° C , pH 7.2. Warburg
cups contained 1.5 ml. Sorensen's phosphate buffer (M/l5), pH 7.2, and
0.5 ml. cell suspension in the main compartment. The substrate to be
oxidiged was tipped in from the side-arm after equilibration. The centre
well contained 0.15 ml. of 20$ K0H for the absorption of carbon dioxide.
Total volume of fluid in the cups was 3.15 ml. A l l substrates were
neutralized where necessary and added at suoh a concentration that 0.2 ml.
substrate was equivalent to 18 uM oxygen. The endogenous respiration
was always measured (94). Oxygen uptake due to substrate dissimilation
was determined by subtracting the endogenous oxygen uptake from the
total uptake.
Chemicalt With the exception of sodium pyruvate and cis-aconitic acid,
a l l substrates were commercial preparations. The pyruvate was prepared
in this laboratory by the method of Robertson (122), while cis-aconitate
was prepared from the trans- isomer by the procedure of Malachowski &
Maslowski (86).
Warburg vessels were cleaned by soaking overnight in detergent
solution, then rinsed and treated with hot 10$ M O 3 for an hour. Finally
they were washed with water and dried at 80° C.

- 13 -
EXPERIMENTAL
- Easting oells- of P. aoruginosa harvested from a mineral medium
in which succinate, fumarate or malate was the sole carbon source,
metabolize these dicarboxylic acids immediately and at a constant and
rapid rate (Figures II, III and IV). Other intermediates in the Krebs
cycle are oxidized more slowly or only after a period of adaptation.
This suggests that cells grown on one dicarboxylio acid («.g» succinate)
hava developed the enzymes for metabolizing not only that particular acid
but also fumarate and malate. On the other hand, i f the calls are grown
on acetate (Figure V), or on gluoose or pyruvate (as carried out in tha
present and other investigations, but not shorn), they metabolize the
dicarboxylic acids only after a lag period. According to Karstrom's
terminology (63), cells grown on any one of the 4-carbon dicarboxylio
acids possess "constitutive" enzymes for oxidizing a l l these compounds,
while calls grown on some other carbon source must first develop "adaptive"
enzymes for attacking these substrates.
The sharp division of enzymes into "'constitutive" and "adaptive"
has been questioned during recent years (161, 164). It may well be that it
the lag phase, characteristic of an "adaptive curve, represents the time
necessary for the development of the enzyme systems which will transport
the substrate across the ce l l membrane. It is therefore possible that cells
whioh have been grown on any one of the 4-oarbon dicarboxylio acids may
use the same system for transporting a l l these substrates across the c e l l .
Similarly, cells grown on another carbon souroe, would be expected to
show the same adaptive pattern for each of the 4-carbon acids. Results
obtained (29) with acetate oells or with glucose cells do nob indicate

-.14 -
I S f
ot- XET0GLUTA1Z47E
TYKL
•SULCI MATE
'ALATE
<7b
VATE
MINUTES
Figure I I . O x i d a t i o n of s u c c i n a t e , fumarate, mala te , ° ^ - k e t o g l u t a r a t e , pyruvate and ace ta te by 22 h r . c e l l s ha rves ted from a succ ina t e ammonium phosphate medium. C i t r a t e and i s o c i t r a t e were o x i d i z e d a t a l o w , r a t e , w h i l e c i s - a c o n i t a t e was not me tabo l i zed . ^
Warburg cups con ta ined 0.5 m l . c e l l suspens ion , 1.5 m l . M/15 phosphate b u f f e r pH 7 . 2 , 0 .2 m l . s u b s t r a t e . F i n a l volume 3 .0 m l . T h e o r e t i c a l oxygen uptake f o r complete o x i d a t i o n of any subs t r a t e was 403 u l .

- 15 -
SUCCINATE
PUMARATE
•8
441
3 s
MINUTES Figure III. Oxidation of succinate, fumarate, malate, o^ketoglutarate, pyruvate
and acetate by 23 hr. cells harvested from a fumarate ammonium phosphate medium. Citrate and cis-aconitate were metabolized at a low rate.
Warburg cups contained 0.5 ml. c e l l suspension, 1.5 ml. M/15 phosphate buffer pH 7.2, 0.2 ml. substrate. Final volume was 3.0 ml.
f Theoretical oxygen uptake for complete oxidation of any substrate was 403 u l .

- 16 -
20 o MALATE
ACETATE
VCWTA&.TE
OMLACETATE
6b
MINUTES
Figure IV. Oxidation of malate, succinate, fumarate, oxalacetate, °<-ketoglutarate, pyruvate and acetate by 22 hr. cells harvested from a malate ammonium phosphate medium. Citrate and isocitrate were oxidized at a low rate, while cis-aconitate was not metabolized.
Warburg cups contained 0.5 ml. c e l l suspension, 1.5 ml; M/l5 phosphate buffer pH 7.2, 0.2 ml. substrate. Final volume was 3.0 ml. Theoretical oxygen uptake for complete oxidation of any substrate was 403.ul.

- 17 -
I*
Sao ACETATE
o
MINUTES.
Figure V. Oxidation of acetate, succinate, fumarate, malate and isocitrate by 21.hr. c e l l s harvested from an acetate ammonium phosphate medium. Citrate and cis-acoriitate were oxidized at a low rate.
Warburg cups contained 0.5 ml. c e l l suspension, 1.5 ml. phosphate buffer pH 7.2, 0.2 ml. substrate. Final volume was 3.0 ml. Theoretical oxygen uptate for complete oxidation of any substrate was 403.ul.

- 13 -
the operation of such a transport mechanism. The effect of uranyl
nitrate, vdiich inhibits the transport of substrate across the cell
membrane, has been studied in Part III, and evidence has been obtained
that the same transport mechanism is not used by a l l the 4-carbon
dicarboxylic acids.

- 19 -
' • DISCUSSION
It is suggested that the substrates succinate, fumarate and
malate are metabolized at a constant and rapid rate by cells of P.aeruginosa
A.T.C. 9027 whioh have been grown on any one of these compounds, because of
an enzymatic relationship among themj i.e. :
Succinate v 1 ~ Fumarate ^ Malate succinic fumarafce dehydrogenase
Such a relationship, "vfoioh w s first proposed by Quastel and
coworkers (114,115,116,117jl56) in studies with B. ooli and B.pyooyaneus,
is probably operative in other bacteria, including Azotobacter agilis (62)
and P.saocharophila.(l8)« The enzyme succinio dehydrogenase has been
obtained in cell-free extracts from E.ooli (2,57), S. aertryoke (77),
and C. diphtheriae{106). Conclusive proof of its presence in P.aeruginosa
A.T.C. 9027 is presented in Part III, while suggestive evidence has been
•given for its occurrence in a number of other bacteriaj Azotobacter agilis **• ' n i- r I I --. •
(62), B.laotis aerogenes (16), P. saccharophila (18), Moraxella lweffi
mutant S (85), B. typhosa (52), and certain stenothemophilic bacteria (47).
The enzyme fumarase has been prepared from Propionibacterium Shermanii (74).
In addition to succinate, fumarate and malate,(Ney (91) has shown
that oxalacetate is metabolized at a constant and rapid rate in succinate-
grown cells, and that this substrate is decarboxylated anaerobioally, no
doubt to pyruvate. It is very likely, therefore, that succinate oxidation
follows the familiar tricarboxylic acid cycle, the existence of which in
P. aeruginosa A.T.C. 9027 has already been proposed by Campbell & Stokes (29).

- 20
PART II
THE METABOLISM OF DRIED CELLS
It has been observed in Part I that whole resting oells of
P. aeruginosa metabolize sudcinate, fumarate and malate at approximately
the same rate, regardless of which of these three substrates has been
used for growth. This suggests that there is an enzymatically controlled
relationship between the three compounds. Other intermediates in the
Krebs cycle are metabolized only after a lag phase or period of adaptation,
whioh may represent the time necessary for the elaboration of a system
for transporting the compounds across the cell membrane.
By using dried cell preparations of P. aeruginosa, Campbell &
Stokes (29) were able to increase the range of substrates attacked. Ia
the absence of the complicating factor of permeability, a clear quantit
ative picture was obtained. In the present study, dried cell preparations
of succinate- sad aoetate- grown cells were used in the hope of finding
a quantitative relationship amongst the 4-carbon dicarboxylic acids.
The possible carrier systems and optimum pH of the succinate-
oxidizing enzyme in these dried cells were also investigated.

METHODS
To obtain oells for driad preparations, 20 litres of ammonium
phosphate medium containing succinate or acetate as sole carbon source,
were dispensed in Roux flasks and inooulated with a 24 hour culture of
P. aeruginosa A.T.CV 9027. The cells were collected at the end of 24
hours with a Sharpies centrifuge at 48,000 r.p.m. They were washed once
with 0.9$ saline and re-oentrifuged in a Servall High Speed centrifuge
at 5,000 r.p.m. The resulting pink cell paste was suspended in 50 ml.
distilled water and spread in a thin layer on a large Petri plate. The
cells were dried slowly (3 to 5 days) in vacuo over phosphoric pentoxide
at room temparature. The resulting tanrglassy residue was ground to a
fine powder in a mortar, and stored in air in the freezing compartment of
a refrigerator. The cells were used after a storage period of one week
or more. Two preparations of succinate cells were made, weighing 4.8 and
7.7 grams respectively. One preparation of acetate cells was made, 20
litres of medium yielding 6.2 grams of cells.
For Warburg studies, the dried cells were weighed accurately and
suspended in distilled water so that; each ml. of suspension contained 20
mgm. of cells. An even milky suspension of cells, which could be pipetted
accurately, was obtained after the dried cells had stood for 10 to 15
minutes. Each Warburg vessel contained 1.0 ml. of this suspension. In
addition, a l l Warburg vessels contained 2 uM MgS04.7H20, 2 uM MnCIg
0.4 mMAlCl3, 0.4 mM CaCl 2, 107 ug. DPN, and 5 uM substrate, neutralized,
to pH 7. All runs were made at 31° C , and pH 7.2 or 7.4. In experiments
on possible oarrier systems, 1 and 4 mg. of cytochrome o, and 0.001$ and
0.01$ final concentration of pyocyanine were used. The cytochrome c was

- 22 -
the produot of the Sigma Chemical Company, while the pyocyanine had been
prepared in the laboratory by the method of Elema & Sanders (39).
The optimum pH for succinate oxidation was found by the standard
Thunberg method (155). In the tube were placed 1.0 ml. buffer (phthalate
for pH 4 to 5, phosphate for pH 5.5 to 8, and borate for pH 8 to 10), 1.0
ml. cell suspension (containing 20 mg. dried cells), and 0.3 ml. of 0.01M
malonate. .To the side-bulb were added 0.5 ml. ljlOOO methylene blue, and
0.5 ml. 0.05M sodium succinate. A standard was included which contained
a l l the components of the above system (cells inactivated by boiling for
20 minutes) but with methylene blue at l/lO normal concentration. This
tube represented 90$ reduction of the methylene blue and was used as the
end point of reduction. An endogenous control was also run. Thunberg
tubes were evacuated with a strong water suction pump for 3 minutes before
they were sealed. Following a 10 minute equilibration period at 31° C,
the substrate and methylene blue were tipped in. The end point of the
reaction was recorded as the time required for the colour intensity of
the experimental tube to be reduced to that of the boiled cell standard.
At the end of the reduction period, the seals were checked to confirm the
presence of a vacuum throughout the experiment.

- 23 -
EXPERIMENTAL
Dried oell preparations ,o"£ bacteria have been used for many
years. These preparations have a high endogenous respiration for 4 or
5 days, which drops off to a negligible value when the cells are stored
in the refrigerator. The enzymatic activity remains constant for
several months thereafter. These observations were made forty years
ago by Shattock & Dudgeon (130) and later by Callow (25)• They have been
confirmed by Sleeper et a l . (134) with P. fluoresoens. In this laboratory,
Stokes & Campbell (142) obtained quantitative data on glucose oxidation
by P. aeruginosa, while Campbell & Stokes (29) have presented evidence
for a Krebs cycle in the same organism, both studies being made with dried
oell preparations.
With a view to obtaining a quantitative picture of the oxidation
of the 4-carbon dicarboxylic acids in P.aeruginosa, cells were grown on
succinate and on acetate and dried in vacuo. In Warburg experiments
(Figures VI,VII,VIII), the oxygen consumption was low in comparison to that
obtained by Campbell & Stokes (29) with glucose- or aoetate - grown cells.
In Figure VI, although the values for oxygen consumption are
approximately half the theoretical, a definite relationship is seen, i.e.
24 ul oxygen are required for the step succinate to fumarate, 28 ul for the
step succinate to malate (theoretical in each case is 56 ul), while 56 ul
are used in the oxidation of succinate to oxalacetate (theoretical 112 ul).
The succinate preparation was also able to oxidize citrate, cis-aconitate,
isocitrate, and ^-ketoglutarate, while pyruvate and aoetate were not
metabolized (Figure VII). The enzymes for oxidizing pyruvate and acetate
were probably destroyed during the drying process. ' Similarly, Campbell &
Stokes (29) found that the acetate enzyme was inactivated i f drying took

Bo
MINUTES
Figure VI. Oxidation of succinate, fumarate, malate and oxalacetate by dried cells harvested from a succinate,ammonium phosphate medium.
Warburg cups contained 20 mg. dried cells, 0.5 ml. M/5 phosphate buffer pH 7.2, 2 uM MgSe^HgO, 2 uM MnClg, 0.4 mM A1C13, 0.4 mM CaClg, 107 ug. DPN, and 5 uM substrate. Final volume was 3.0 ml.

- 25 -
Figure VII, Oxidation of succinate, fumarate, isocitrate, 'X'-ketoglutarate, cis-aconitate and citrate by dried cells harvested from a succinate ammonium phosphate medium. Pyruvate and acetate were not metabolized.
Warburg cups contained 20 mg. dried c e l l s , 0.5 ml. M/5 phosphate buffer pH 7.2, 2 uM MgSO^HgO, 2 uM MnClg, 0.4 mM AICI3, 0.4 mM CaCl2, 107 ug DPN, and 5 ui£ substrate. Final volume was 3.0 ml.

- 26 -
longer than 48 hours, an observation whioh was also made by Barron et a l .
(12) with dried cells of C. oraatinovorans.
With aoetate-grown cells, the quantitative picture for the
stepwise oxidation of the 4-carbon dicarboxylic acids more closely approached
the theoretical (Figure VIII). Here the differences in oxygen uptake betweeen
succinate and fumarate and between succinate and malate are 45 and 433 ul
respectively (theoretical in both cases, 56 ul).
With succinate as substrate, dried cells take up approximately
one-third of the oxygen consumed by resting cells. Since the latter have
their oarrier systems and oof actors intact, i t is possible that the low
oxygen uptake charaoteristio of dried cells may arise from the absence of
these necessary constituents. The system for succinate oxidation, known
as the succinoxidase system, has been studied mainly with animal tissue
homogenates (65,106,133) where i t is generally supposed to consist of the
following steps:
Succinate .... Sucoinic dehydrogenase . ... cytochrome b —> cytochrome c
—•^•cytochrome a > cytochrome oxidase (ag) oxygen.
Of the cytochromes, only "c" has been isolated as a soluble pure protein (64),
evidence for the others being based on spectrophotometry observation. In
addition, succinic dehydrogenase and cytochrome oxidase are closely associated
and form an integral part of the large granules or mitochondria in certain
animal cells (58,129). Although the evidence for the existenoe of cytochrome
as a oarrier system in certain species of bacteria is controversial (137,163)
and no large granules are present as sites of succinic dehydrogenase and
cytochrome oxidase activity, i t is probable that P.aeruginosa possesses
the various constituents of the cytochrome system (45,137).
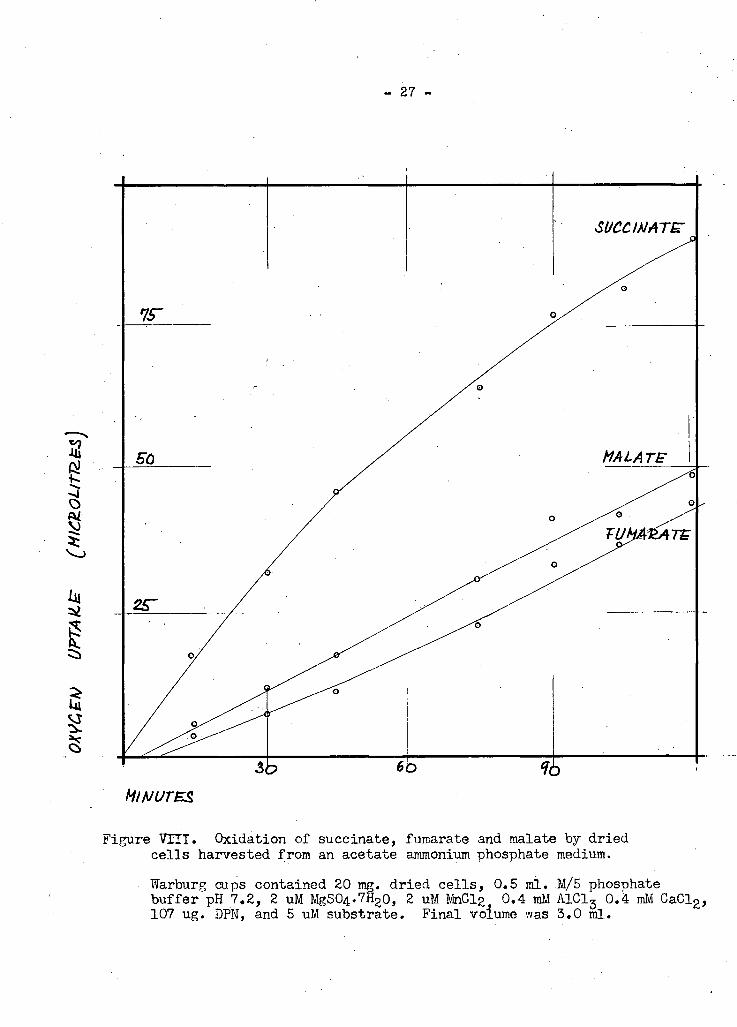
- 27 -
SUCCINATE
1b MINUTES
Figure V X I I . Oxidation o f succinate, fumarate and malate b y dried cells harvested from an acetate ammonium phosphate medium.
Warburg cups contained 20 mg. dried cells, 0.5 ml. M/5 phosphate buffer pH 7.2, 2 uM MgSO^HgO, 2 uM MnClg 0.4 ml A1C13 0.4 ml CaClg, 107 ug. DPN, and 5 uM substrate. Final volume was 3.0 ml.

- 28. -
There is also the possibility of pyocyanine acting as a carrier
sinoe i t is produced copiously by F.aeruginosa on a succinate ammonium
phosphate medium, can be reversibly oxidized and reduced, and has been
found capable of acting as a carrier system with various dehydrogenase
systems in vitro (38).
In addition, a number of metal ions are required as cofactors
for dehydrogenase systems. Ca and Al enhance the activity of animal
succinoxidase (112,113), while Mgt+ and Mn++ are important cofactors for
a large number of enzymes in the glycolytic; and Krebs cycles in both
animal and bacterial tissues (80). These four ions, therefore, were
included in a l l experimental systems.
The effeot of various concentrations of cytochrome c and of
pyocyanine on oxygen uptake are given in Tables I and II. No significant
increase in oxygen consumption is evident. In these particular experiments
there was no endogenous respiration. However, in another experiment where
the oells did oxidize their storage produots, no increase in oxygen uptake
occurred in the presence of added cytochrome o or pyocyanine*
Table I
The effeot of cytochrome o on the oxygen uptake of
dried succinate-grown oells. Substrate: 5VuM succinate.
pH 7.2 .
Oxygen uptake (ul) Time
No 1 mg. 4 mg. (minutes) cytochrome cytochrome cytochrome
60 48 54 43 100 73 75 79 140 91 101 86 180 133 137 131

- 29 -
Table II
The effeot of pyooyanine on the oxygen uptake of dried
suooinate-grown cells* Substrate; 5 uM succinate. pH 7.4 •
T j j n 9 Oxygen uptake (ul) No 0.001$ 0.01$
(minutes) pyocyanine pyooyanine pyocyanine
30 30 37 37 60 ' 70 69 76 90 90 87 97
120 113 105 121 150 131 124 142
The optimum pH of succinic;dehydrogenase has been determined by
the Thunberg method for both succinate and acetate dried cells of
F.aeruginosa,(Figures IX and X). With succinate cells, the optimum
lies between pH 7.0 and 7.5,, while with acetate cells i t occurs at
pH 8.0 .

- 3 0 -
Figure IX. Effect of pH on reduction of M.B. by dried succinate-grown cells, (2 preparations).
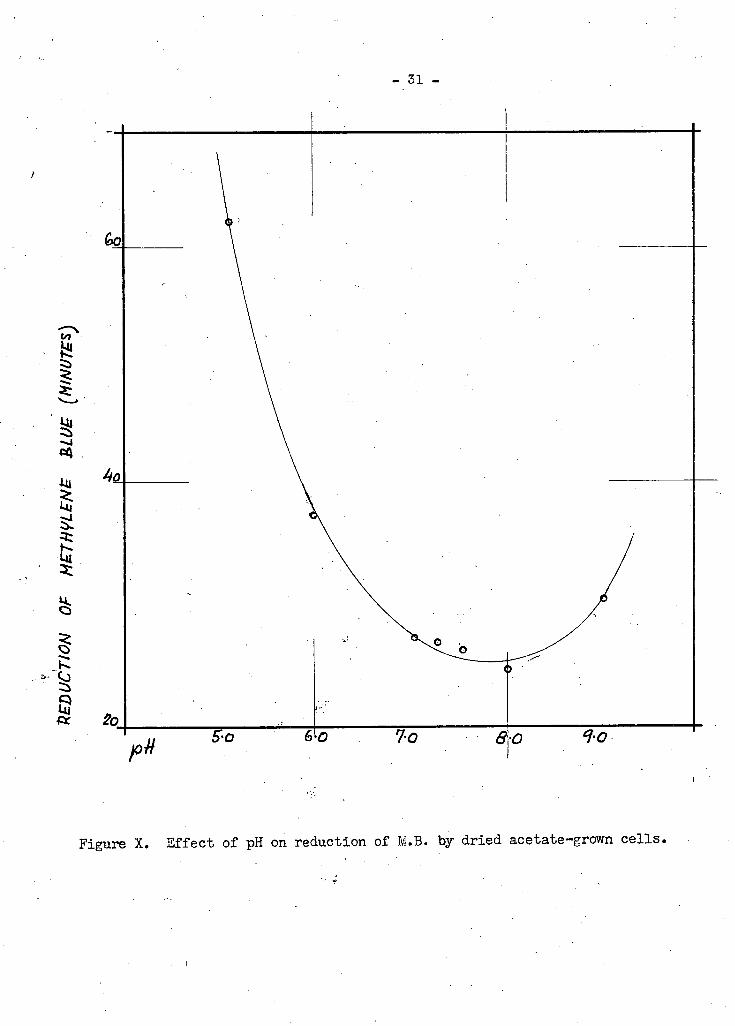
- 3 1 -
F i g u r e X. E f f e c t o f pH o n r e d u c t i o n o f M.B. b y d r i e d a c e t a t e - g r o w n c e l l s .

- 32 -
DISCUSSION
The non-stimulatory effect of cytochrome c on succinate oxidation
by P. aeruginosa may be the result of one or more factors* F i r s t , there is
the possibility that cytochrome o is not a limiting factor- i n this oxidation.
Or, i f we assume that the cytochromes are limiting i n this preparafc ion, the
non-stimulatory effect may result either from the inactivation of cytochrome
oxidase during the drying process, or from the requirement for some cytochrome
component other than "c":. Discussion is restricted to the latter two
p o s s i b i l i t i e s .
"Very l i t t l e work has bean done with bacterial cytochrome oxidase
since Kubowitz & Haas (76) f i r s t obtained evidence for i t s occurrence.-
However, Borei (22) found that the cytochrome oxidase of horse heart became
inactive with increasing age, due to aL terations i n the colloidal struoture.
Such an alteration may also occur i n dried cells of P. aeruginosa. Support
for this possibility i s contained in the work of Militzer et a l . (88) who
found cytochrome oxidase to be the limiting enzyme for the cytochrome chain
in thermophile No. 2184.
The evidenoe for the cytochrome system (a,b and c) i n P.aeruginosa
(45,137) is based solely on spectrophotometric observation. There i s ,
therefore, no guarantee that the bacterial components are structurally
identical with those i n animal tissue(45). Support for this view may be
found i n the fact that neither of the succinoxidase preparations made thus
far from bacterial oells (77,106) require cytochrome c as a carrier.
As noted in the preceding section, pyocyanine does not inorease
succinate oxidation by dried cells of P. aeruginosa. Actually the r . ' .v
evidence for pyocyanine as a carrier system i s fragmentary. Friedheim (43)

_ 33 -
observed an increased oxygen consumption when pyocyanine was added to
suspensions of B. pyooyaneus, an observation which was later confirmed by
Norris (93), Reed & Boyd (121) also comment on the respiratory significance
of the pigment in B. pyocyaneus cultures, while Dickens & Mcllwain (38)
found it capable of acting as a carrier with the hexosemonophosphate system
of yeast juice and red blood cells. However, oontrary results were obtained
by Barron et a l . with C. creatinovorans (12), and by Keilin & Hartree (65)
with a heart muscle preparation. <In these cases, pyocyanine was inhibitory.
The optimum pH for suooinio dehydrogenase in suocinate-grown dried
oells lies between pH 7.0 and 7.5, while with aoetate cells i t occurs at
pH 8.0. These values coincide with others in the literatures Ohlsson (104)
in the original study on bacterial succinic dehydrogenase found. pH 9.0 to
be optimum, while Quastel & Whetham (117), three years later, found a pH
of 7.4 to be the most suitable for the reduction of methylene blue by
resting oells of B.coli. Subsequently, Cook & Aloook (34) found pH 7.6
to 8.0 to be optimum for the aerobic oxidation of succinate by E. coli.
In more recent work, Kun & Abood (77) showed that oell-free preparations
of succinoxidase from S« aertrycke have their optimum activity at pH 7.4,
while Pappenheimer & Hendee (106) found pH 8.0 to be optimum for suocinio
dehydrogenase from C. diphtheriae.
The anaerobic reduction of methylene blue by resting oells of
P. aeruginosa was studied by Randies & Birkeland (120). Guggenheim (52,53)
investigated the same property in a number of pathogens (S. typhosa,.
CI. butylioum, CI. parabotulinumi, and CI. welohii.)

- 34 -
PART III
INHIBITOR STUDIES WITH MALONATE AND URANH, NITRATE
Certain specific information concerning the pathways of cellular
oxidation may be obtained by studying the effects of metabolic inhibitors*
In general, these compounds f a l l into two main groups, depending on the
manner in which they effect inhibition* The first and by far the larger
group act by inhibiting the activity of enzyme systems directly* They
may do so by combining with the enzyme system through one of the following:
the activating protein, the prosthetic group or the carrier system, or by
competing with or otherwise reduoing the effective concentration of substrate*
Malonate, which inhibits the enzyme suecinio dehydrogenase by competing with
the normal substrate for a position on the enzyme surface, has been chosen
from this group*
The second group of inhibitors act indirectly by altering one or
more of the varied meohanisms which regulate the rate and direction of
enzymatic reactions in the living oell. One of these regulating mechanisms
is the state of the oell surface, which is important not only because of
its property of selective permeability, but also because of its more direct
role in the "active" transfer of materials into the cell* That this "active"
transfer is oonnected with phosphorylation has been suggested by Barron (90)
and by Rothstein (123,124) in studies on the metabolism of glucose in yeast
and bacteria, using uranyl nitrate as an inhibitor* An investigation of
the effect of this inhibitor on the metabolism of organic acids and glucose
in P* aeruginosa was undertaken in the present study* In recent years,
ample evidence has appeared for a coupling between the oxidation of organic

acids or glucose and phosphorylation (10,11)• Uranyl nitrate, therefore,
appeared to be a useful tool -whose action might provide more concrete
evidence for this connection between aerobic metabolism and phosphorylation*

- 36 -
METHODS
The methods of maintaining stock cultures were similar to those
described in Part I. Growth medium for the preparation of resting cell
suspensions was similar to that used previously. Resting oell suspensions
of 21 to 24 hours cultures were prepared as in Part I. Dried cell prepar
ations were the same as described in Part II.
Warburg' experiments were conducted as in Part I. In malonate
studies, Sorensen's phosphate buffer was used, while M/7 veronal buffer
was employed xvith uranyl nitrate. 0.3 ml. of inhibitor was added at ten
times the final concentration desired. Thirty to forty minutes contact
between inhibitor and cells was allowed before tipping in the substrate
from the side-arm. In malonate inhibition studies at pH's 5.0 to 5.6,
substrate and mal onate were both neutralized with N/l NaOH to the required
pH using a Beckman pH meter. Uranyl nitrate was also neutralized with
N/i NaOH before being added to the Warburg cups. Percentage inhibition
has been expressed in terms of the % 2 ( u ) values.
Malonate inhibition of dried oell preparations was studied by
both the Warburg method and by the Thunberg technique. In the latter
instance, 0.3 ml. of ten times the desired concentration of malonate was
left in contact with the cell suspension in the main tube for 30 minutes
before adding succinate and methylene blue from the side-bulb.
Chemically-clean glassware was used for determining the influence
of magnesium on growth. This was prepared by soaking overnight in 10$
nitric acid, washing ten times with tap water, three times with distilled
water, three times with re-distilled (glass) water, and finally autoclaving
for 15 minutes at 15 pounds pressure. The basal medium, consisting of

- 37 -
0.11$ KH40H, 0.11$ K2HP04, 0.5 p.p.m. Fe^* (as FeS04.4H20), and 0.33$
sodium succinate, was dispensed in 9 ml. amounts into 125 ml. Erlenmeyer
flasks, and autoclaved. MgS04.7H20 \ms made up in concentrations ranging
from 0.001$ to 10$i and autoolaved separately. 1 ml. of the various Mg++
solutions was added aseptically to the basal medium. Final concentrations
of various constituents were: 0.1$ HH OH, 0.1$ K2HP04, 0.5 p.p.m. Fe ,
0.3$ sodium succinate, and Mg + + ranging from 0.0001$ to 1$.
The inoculum was prepared from a' 21 hour culture of the organism
in basal medium •+• 0.05$ Hg + + . These cells w ere harvested by centrifug-
ation, and washed twice with water re-distilled from glass. They were
re-suspended to give a barely visible turbidity. One drop of this suspension
was employed as inoculum for 10 ml© of medium. Following incubation for
24 hours, the turbidity of the suspension was read in a Fisher Eleotrophoto-
meter using Filter 525 B.

- 38 -
EXPERIMENTAL
A. Malonate Studies
The inhibition of the anzyme succinic dehydrogenase by malonate
was first reported by Quastel & Whet ham (118). Subsequent work (34,49,
116,119) has confirmed their observation and demonstrated a high degree of
specificity for the action of this inhibitor. Szent-Gyorgyi and coworkers
(8,49) observed that malonate inhibited the respiration of pigeon breast
muscle and that succinate was formed oxidatively from fumarate in the presence
of malonate. Further investigations of these effects by Krebs and his assoc
iates (72,73,75) have furnished some of the strongest evidence for the
oxidation of pyruvate by the tricarboxylic acid cycle.
Malonate is assumed to inhibit succinic dehydrogenase by competing
with the enzyme's normal metabolite, sucoinate, for the "hot spots'" or active
centres on the enzyme surface. These centres are thought to be oriented
specifically to pick up succinate as a preliminary to its dehydrogenation,
but may also adsorb closely related molecules such as malonate. This
phenomenon of competitive inhibition, first observed by Qjuastel (115),
results in a more or less complete blocking of the enzymatic reaction© The
effective concentrations of malonate are relative rather than absolite -
i.e. the ratio of suocinate to malonate is the important factor - since the
affinity of the inhibitor for the enzyme is considerably greater than that of
the substrate. Potter & DuBois ( i l l ) obtained 50$ inhibition with, a v
substrate/inhibitor ratio as low as 50/l, while in earlier work Krebs &
Eggleston (73) required a ratio of 9.5/1 •
Initial experiments were conducted with dried cell preparations.
A preliminary study of the effect of pH on malonate inhibition was made
according to the Thunberg teohnique. The tubes contained 20 mg. dried cells
(suspended in 1 ml.), 1.0 ml. phthalate, phosphate or borate buffer

(depending on the desired pH), and 0.3 ml* of either web er or 0*01 M
malonate* In the side-bulb were placed 0*5 ml* of 0.05M suooinate and
0*5 ml* of 1*1000 methylene blue* The ganeral procedure was carried out
as in Part I* Results are given in Tables III and IV.
Table III The effect of pH on malonate inhibition of two
different dried preparations of suocinate-grown cells
Preparation A Preparation B Reduction time Inhibition Reduction t ime Inhibition
(minutes) (per cent) (minutes) (per Succ. Succ. Me i l . Succ. Succ.-+-Mal.
4.1 165 165 0 160 160 0 4.6 108 87 0 115 110 0 5.5 57 129 55 40 67 40 5.9 56 90 39 6*5 40 91 56 35 65 46 7.0 23 55 58 16 43 63 7.5 14 50 72 18 50 64 8*0 17 60 72 20 55 64 9.0 20 40 50 20 44 55
10.0 35 60 42 35 75 53
Note: Endogenous controls showed no reduction at 180 minutes.
Table IV The effect of pH on malonate inhibition of succinic dehydrogenase activity in acetate-grown dried cells
Reduction time (min.) Inhibition pH Succ. Succ . •+• Malon* (per oent)
4.1 180 180 0 5.1 63 57 0 6.0 37 59 37 7.0 27 47 43 7.2 27 42 36 7.5 26 41 37 8.0 24 45 47 9.0 31 46 33
10.0 48 68 29 Note: Endogenous controls showed no reduction at 170 minutes.

- 40 -
It may be concluded that malonate exerts its inhibitory effect
in the pH range of 7.0 to 8.0. This is also the optimum pH range for
succinate dehydrogenation in dried cell preparations, as found in Part II«
Qoiastel & Yfooldridge (119) carried out their experiments at pH 7.4, using
normal resting cells and toluene-treated cells of B. ooli. Since a l l
previous studies with P. aeruginosa have been done at pH 7.2 and 7.4, i t
was decided to use pH 7.4 for further investigation of malonate inhibition.
Warburg studies of the effect of malonate on succinate oxidation
were carried put as in Part II, with both succinate- and acetate-grown dried
cells. The results are summarised in Table V.
Table V. The effect of malonate on the oxygen uptake by dried cells.
Warburg vessels, contained 20 mg, cells, 0.5 ml. M/5 phosphate buffer pH 7.4, 2 uM. MgS04.7H20, 2 uM MnCl2, 0.4 mM AICI3, 0.4 mM CaCl2, 5 o r 10 succinate in side-arm. Malonate was in contact with cells 30 minutes before -.tipping in substrate.
Sxpt. Dried Time Suocin- Malonate Oxygen uptake (ul) Inhibition No. Cells ate.
I Succinate 60 ' 5 uM " 120 « t i 1 8 Q ,1
II Succinate 66 5 uM: " 120 " " 180 "
III Acetate 60 5 uM « 120 11 " 60 " " 120 "
17 Aoetate 60 10 uM " 120 "
60 '» 120 '«
Succ. SuccMale (per c
0.001 M ,51 12 77 11 83 18 78 n 98 34 65
0.005 M 45 0 100 11 59 0 100 n 69 0 loo
0.001 M 62 0 100 tt 89 43 51
0.005 M .62 0 100 11 89 14 84
0.001 M 94 47 50 t i 135 87 36
0.005M 94 9 90 11 135 41 60

It is apparent that malonate in concentrations ranging from
0.001M to 0,005M is able to inhibit the oxidation of suooinate by dried
cell preparations of ff, aeruginosa, harvested from either succinate- or
acetate- ammonium phosphate medium, Table V, Since Krebs (72) considers
that in these concentrations malonate acts exclusively upon the enzyme
succinic dehydrogenase, we may conclude that both types of cells contain
this enzyme. The fact that at a concentration of 0.001 M, malonate
produces a greater inhibitory effect on sucoinate-grown oells than on
acetate-grown cells would tie in with earlier observations that cells which
have been grown on succinate contain a higher relative concentration of
succinic dehydrogenase than those grown on acetate.
In studies with 0.05M malonate at pH 7,4, Ney (91) was unable
to inhibit either the rate or the extent of oxidation of succinate or
glucose by glucose-arhmonium phosphate resting cells of P, aeruginosa.
These experiments were repeated in the present study, using both 0,01M
and 0.05M malonate at pH 7.2 and pH 7.4 • The cells were harvested from a
medium in which sodium succinate constituted the sole oarbon source.
The effect of 0.01M malonate on the oxidation of suocinate,
fumarate and glucose, is given in Table 71. This concentration of malonate
inhibited the rate of oxidation of both succinate and fumarate, but not that
of glucose.
The inhibitory effect of malonate, in concentrations 0.01M and 0.05M,
on the oxidation of suocinate and fumarate has been computed in the form of
^)o2(K) v a l , U 9 S i n ^&^9 VII. Experiment 1 is shown in Figure XI. It is
apparent that 0.01M and 0.05M malonate inhibit the rate of oxidation of
succinate, and to a lesser extent that of fumarate.

- 42 -
Table 71
Oxidations by suooinate-grown resting c e l l s , i a the presence of 0.01M malonate» pH 7>2 . •
Warburg substrate
Time (minutes)
Endog. •+• Malonate
Oxygen uptake (ul)
Substrate Total Endog, + Malonate* Inhibition Substrate (per oent)
Succinate 20 18 174 192 145 25 it 40 38 219 247 230 7 II 60 70 221 291 267 8
Fumarate 20 18i. 164 182 131 29 II 40 38 209 247 192 22 II 60 70 218 288 228 21
Glucose 20 18 29 47 55 0 n 40 38 50 88 . 112 0 it 60 . 70 73 143 190 0
Table 711
Oxidations by auooinate-grown resting c e l l s ,
i n the presence of O.OlM.and 0,05M malonate
Experiment Number pH
7.2 it
Warburg Substrate
Succinate it
Malonate (Molar)
0.01 0.05
9: D 2 ( u )
3220 2040 1190
Inhibition (per cent)
37 63
II
t i
it
t i
7.4: It
Fumarate it
Succinate it
Fumarate tt
0.01 0.05
0.01 0.05
0.01 0.05
2740 2740 1605
2222 1900 1105
2395 1940 1255
0 41
14 50
19 48

- 43 -
12 Q
3
FUMARATE +0OS MAJJOHAU-
\TE_ MALONATE
SUCCINATE HATE
CCINATEU-001H MALtbUATE
SUCCINATE +005~H MALONATE
_MINUTES 4o
Figure X L . Oxidation of succinate and fumarate by 21 hr. cells harvested from succinate-ammonium phosphate medium, i n the presence of 0.O1M and 0.05M malonate.
Warburg cups contained 0.5 ml. c e l l suspension^!.. 5. ml. M/15 phosphate buffer pH 7.2, 0.3 ml. malonate, o.2/substrate. Final volume was 3.0 ml. Theoretical oxygen uptake for complete oxidation of any substrate was 403 u l .

- 44 -
Similar results have been obtained by other workers. Bernstein
(18) observed a decrease in the rate of oxygen uptake by cells of P. sacchar-
ophila when employing 0.01M solutions of malonate. With 0.05M malonate, he
obtained a 71$ inhibition of amount of succinate oxidation and a 59$. inhibition
in the case of fumarate. In studies with a suooinoxidase preparation from
S. aertrycke endotoxin, Kun & Abood (77) obtained 100$ inhibition of succinate
oxidation in the presence of 0.01M malonate. Quastel and his associates
(116,118,119) worked with concentrations of 0.02M malonate and higher to
obtain inhibitory effects in their Thunberg studies with B. coli, B.prodigiosus,
and B. proteus. Quastel & Wheatley (116) reported a 44$ inhibition; of
succinate oxidation in the presence of 0.067M malonate, over a period of 2
hours. In the same study, succinate oxidation by fresh cells of B. coli
was inhibited 24$, while with cells whioh had been stored at 0° C. for
several days a 90$ inhibition was observed* The increase in inhibition
may have been due to an increase of cellular permeability. Most
experiments with animal tissue have been carried out with lower concentrations
of malonate than the above (1,111), although Stoppani (143) required 0.01M
malonate to give a 99$ Inhibition of his succinoxidase preparation from heart
muscle or liver. As to inhibition of glucose oxidation by malonate, Harvey
(56) noted that this compound exerted no effeot at 0.01M in experiments
with Trypanosoma hippicum.
A l l these investigations have been made using malonate concentrations
higher than those recommended by Krebs (72) . Das (35) suggested that at
higher concentrations malonate may interfere with enzymes other than suocinio
dehydrogenase. In this connection, Pardee & Potter (107) claim that malonat<a
inhibits the oxidations of the Krebs cycle by at least two mechanismss

low concentrations (0.004M) completely block suocinate oxidation, -while
higher concentrations (0»02M) effect inhibition of oxalacetate oxidation,
presumably by forming a' complex with free or bound magnesium, a necessary
oofactor for the oxidative decarboxylation of oxalacetate, Evans et al.(41)
also noted that 0,01M malonate completely inhibits the decarboxylation of
oxalacetate. Further evidence that oxalacetate oxidation is the point of
inhibition by malonate in higher concentrations was provided by Potter (110),
He found that malate oxidation is not inhibited when oxalacetate is removed
from the system by transamination, Lwoff & Cailleau (85) working with the
S mutant of Moraxella Lwoffi observed that oxalacetate oxidation was completely
inhibited by malonate at a ratio of malonio acid j oxalacetio aoid of 10,
An interesting though complicating feature of the present studies
on malonate inhibition has been that P, aeruginosa possesses an adaptive
enzyme system for metabolizing the inhibitor, Karlsson & Barker (62) and
Karlsson (61) have found that Azotobacter agilis also is able to adapt to
malonate, .Similarly, Baskett & Hinshelwood (16) observed that cells of
B,lactis aerogenes became adapted to malonate after training on glucose, the
normal growth medium. The original report on the bacterial oxidation of
malonate was made by Butterworth & Walker employing B, pyooyaneus (24)•
To provide conclusive proof for the existence of such an adaptive
enzyme system in P, aeruginosa, the following experiment was conducted.
Cells which had been grown on suocinate and on aoetate mineral media for
21 hours were divided into two parts. One was set aside as a control -
non-adapted cells; the other was incubated for 1 hour with malonate (in the
same concentration as the original growth substrates - via, 0»25$Q added to
the medium - adapted oells. In subsequent Warburg studies, the succinate

" - 46' -
enzyme of both groups of cells was saturated with its substrate in a
concentration of 5 uM, and with a high concentration of malonate (o»011l) . .
The resulting curves (Figures XII and XIII) indicate that there are
indeed two separate enzyme systems present - one for metabolizing succinate,
the other for oxidizing malonate. In the case of the non-adapted succinate-
grown cells, malonate had l i t t l e inhibitory effect, The endogenous curves
indicate that the organism is able to adapt to malonate. With the malonate-
adapted, succinate-grown oells, two distinot curves are apparent. The curve
for oxidation of suocinate (5 uM) by the" adapted cells may be an example of
the "uiauxie" effect of Monod (89), in which a double growth cycle consisting
of two exponential phases is operative. Each curve in this instance would
correspond to the exolusive utilization of one of the constituents in the
growth medium (i.e. either succinate or malonate) due to the inhibitory
effect of one of the compounds on the formation of the enzyme for attacking
the other. The phenomenon and its explanation would appear to be valid only
in the oase of compounds whioh are closely related in structure. Cells whioh
have been grown on acetate show adaptive curves fbrboth suocinate and malonate.
In previous experiments at pH 7 .2 and 7 , 4 , inhibition by malonate in
low concentrations was obtained with dried cell preparations, while high
concentrations -were necessary to inhibit whole resting cells* This suggests
that permeability is involved} i.e. dried oells, in which the cell membrane
has been destroyed or altered, are inhibited, while whole cells, with the
membrane intact, show no inhibition. Turner & Hanly (152) in experiments
with carrot slices found that potassium malonate at pH 4 , 0 caused "inhibition
of that fraction of respiration which is inhibited by cyanide," They conoluded
that although the actual inhibitor is the malonic ion, only undissociated
malonio acid serves as cell penetrant, so that a low pH is essential. Similar

- 47 -
to^H SUCCINATE•_ J
^u/f SUCCINATE /0-0fM MALONATE
MINUT£&
Figure XII. Oxidation of succinate and malonate by non-adapted succinate-grown cells (unbroken lines) and by cells which have been adapted to malonate for 1 hour (broken line s ) . Warburg cups contained 0.5 ml. c e l l suspension, 1.5 ml. M/15 phosphate
buffer pH 7.2, O.g. ml. substrate. Final volume was 3.0 ml.
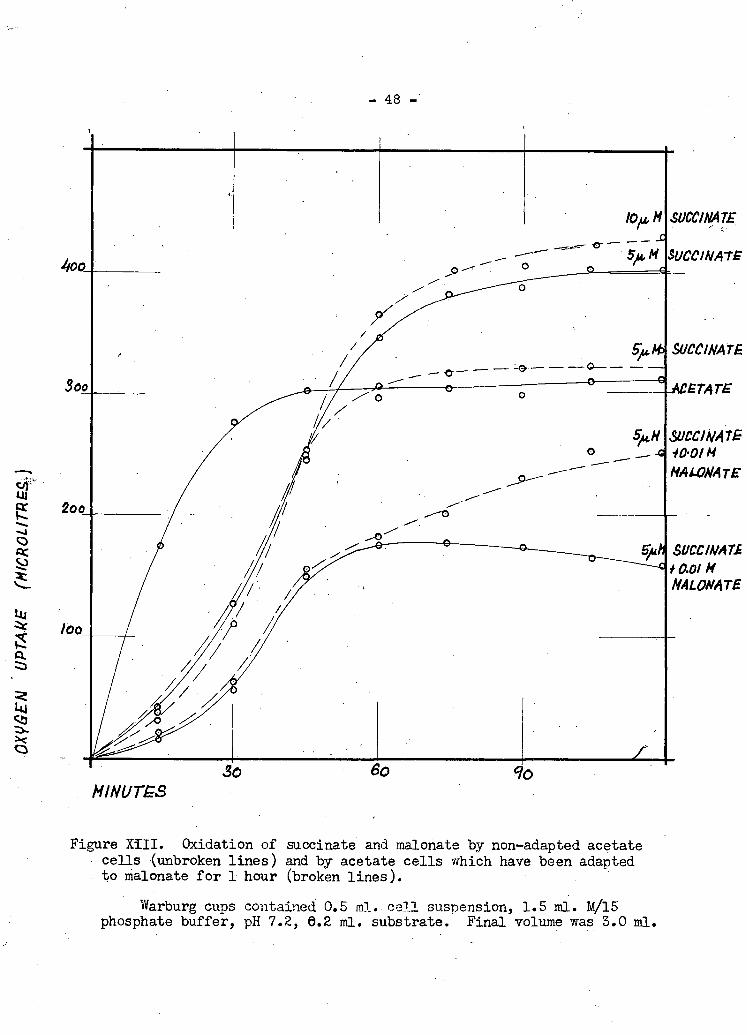
- 48 -
SUCCINATE
SUCCINATE
SUCCINATE:
ACETATE
SUCCWATB •fO-OIH MALONATE
SUCC/NAT1 / 0.01 M MALONATE
MINUTES
Figure XIII. Oxidation of succinate and malonate by non-adapted acetate cells (unbroken lines) and by acetate cells which have been adapted to malonate for 1 hour (broken lines).
Warburg cups contained 0.5 ml. c e l l suspension, 1.5 ml. M/15 phosphate buffer, pH 7.2, 6.2 ml. substrate. Final volume was 3.0 ml.

results were obtained by Bonner & Wildman (21), Bonner (19), and Laties (78), .
with, spinaoh leaves, oat ooleoptiles and barley roots respectively, Gunsalus
(54) working with B, subtilis noted good inhibition at low pH's while none
ocourred at pH 7.0 • The same effect of pH was observed by Barron et a l ,
with C, creatinovorans (12) and with baker's yeast (13),
Results obtained with P, aeruginosa are given in Tables VIII, EC
and X, and in Figure XIV, A l l experiments were conducted at pH 5,0 to
5,6, since at lower values (i.e. pH 3,7 and pH 4,5) no oxidation of substrate
took place. Maximum inhibition of rate and extent of oxygen consumption on
succinate, with succinate-grown cells, was- found at pH 5,0 in the presence
of 0•005M malonate, Malonate affected only the rate of oxidation of other
Krebs intermediates studied, viz. fumarate, malate and V-katoglutarate.
There was no inhibitory effect on,pyruvate oxidation.
Table VIII '
The influence of pH and concentration on malonate inhibition of succinate oxidation,using succinate-grown cells.
Experiment Malonate QQ . Inhibition Final Og Inhibition Number pH (molar) 2 ( N ) of rate % uptake(ul) of amount %
(at 60«) I 5*6 — 895 200
" 0.001 747 16 170 15 M 0.005 494 45 110 45
(at 80') II 5.0 ~ 975 275
III 5.0 n
0.001 812 17 275 0 0.005 283 71 80 71
(at 60') 1845 235
0.002 1 6 4 0 11 225 4 0.003 1 245 33 180 23

- 50 -
Table IX
Inhibition, of succinate and fumarate oxidation by 0*00511 malonate using fumarate-grown oells at pH 5*1
Experimental Conditions °2(N)
Inhibition of 0 2 uptake .."Inhibition rate $ at 90' (ul) of amt. %
Fumarate 880 Fumarate + malonate 706
Succinate 840 Succinate 4-malonate 350
20
58
230 225;
220 130 41
Table X
Inhibition by 0.005M malonate of the oxidation of malate, -ketoglutarate and pyruvate, using oells grown on each of these substrates.
Experimental Conditions
Malate Malate + malonate
PH
5.1 n
-ketoglutarate 5.5 c<-ketoglutarate + "
malonate
Pyruvate' 5*5 Pyruvate 4 malonate "
°2(N)
1310 940
1170 985
1590 1470
Inhibition Og uptake Inhibition of rate % at 90' ul* of amt, %
28
16
215 220
210 195
260 262
Quastel & Wheatley (116) used 0*07M malonate to obtain 30$ inhibition
of fumarate oxidation by B* pyooyaneus over 2 hours, aconcentration which did
not affeot B, coli* These results were typical for organisms which oxidize
fumarate faster than succinate; those with slower fumarate oxidation were
inhibited to the extent of about 90$. Using the same concentration of malonate,
they observed a 6$ inhibition of the amount of oxygen uptake with malate over

- 51
MINUTES Figure XIV. Inhibition of succinate oxidation by various concentrations
of malonate on succinate-grown c e l l s . Warburg cups contained 0.5 ml. c e l l suspension, 1.5 ml. M/l5 phosphate buffer pH 5.0, 0.3 ml. malonate, 0.2 ml. substrate. Final volume was 3.0 ml. Theoretical oxygen uptake for complete oxidation of any substrate was 403 u l .
SUCCINATE
SUCCINATE +0001H MALONATE
SUCCINATE +0-00$ M
MALONATE
SUCCINATE + 0OOS M MALONATE

a period of 2 hours, an amount which is but slightly significant. Although
only one experiment on inhibition was carried out, pyruvate'oxidation by
P, aeruginosa was.found not to be inhibited significantly. This finding
agrees with Lenti's .(81) observation that malonate affects the succinate
but not the pyruvate oxidation of B, co l i . On the other hand, Barron et a l .
(13) found that malonate completely inhibited the oxidation of pyruvic acid ,
by yeast, a finding confirmed by Speck et_ a l , (136) with the malaria parasite.
Similar results were obtained by Szent-Gyorgyi aid his coworkers (8,49) and
by Krebs et a l , (73) using pigeon breast muscle brei, Pardee & Potter (107)
found that 0.004M malonate strongly inhibited pyruvate oxidation in homogen-
ates of rat heart, kidney and brain, but not in liver, Lehninger ( 7 9 ) has
noted that liver homogenates can oxidize pyruvate to acetoacetate in the
presence of malonate,
B, Inhibition by Uranyl Hitrate
Barron (14,90) and Rothstein (123,124) have shown that uranyl
nitrate in low concentrations inhibits metabolism' in whole cells of yeast
and bacteria by combining with the protein of the cell surface. This
uranium-protein complex can be reversed by a number of compounds, including
phosphate, whioh form more strongly associated complexes with the uranyl
ion than does protein (131).
Initial experiments were designed to obviate any complication by
phosphate either in the form of an insoluble precipitate in the culture
medium, or as inorganio phosphate attached to the cell surface. To avoid
precipitation of magnesium ammonium phosphate from the mineral medium, the
concentrations of KH4"*" and P04""" were reduced to 0,1$ while that of Mg*+
was lowered to 0,01$ or 0,005$, depending on the carbon source present.

- 53 -
At this concentration, optimal growth of the bacterium wis maintained, as
shown in Figure XV, which confirms the previous work of Burton et a l . (23).
It was found that by washing the cells three times with 0.9$ saline, the
concentration of phosphate attached to the cell surfaoe could be lowered
to a negligible value. A l l experiments were oonducted with veronal as
buffer© The oxygen consumption of P. aeruginosa at pH 7.2 is the same
whether veronal or phosphate is used as buffer.
x 10 M to 1 x 10_<i M has been investigated. Concentrations of 1 x 10 M
to 4 x 10"^ M reduced the endogenous respiration of P. aeruginosa, as shown
with gluoose oells in Table XI. The higher concentrations, 1 x 10 M and 2
1 x 10 M, caused agglutination of the cells. The inhibition of endogenous
respiration by lower concentrations is probably due to a combination of the
uranyl ion with the proteinaceous storage material of this organism (26),
while at higher concentrations, uranium may be aoting as a general protein
precipitant (131).
The effect of uranyl nitrate in concentrations ranging from 7.5
Table XI The effect of ^ 2 ( ^ 3 ) 2 on the endogenous respiration of cells harvested from a gluoose ammonium phosphate medium.
Duration of experiment was 70 minutes.
Experiment Number.
Experimental Conditions
Oxygen Uptake (ul)
Inhibition (per cent)
I Control 65 35 41 28 81 63 © 0
1 x lO" 4 M UJ 46 37 57
II
2.5 x lO" 4 M U 4 x 10"4 M U Control 1 x lO" 4 M U 1 x 10"3 M U 1 x 10-2 M U
22 100 100

- 54 -
Figure XV. The influence of magnesium on the growth of F. aeruginosa

- .55-
The effect of uranyl nitrate on the oxidation of the Krebs. inter
mediates and of glucose by cells which have been grown on these compounds,
is shown in Tables<XII to XV, and Figure XVI. Uranyl nitrate in concentrat
ions up to 4 x 10"*4 M did not effect a significant inhibition of glucose
oxidation by P. aeruginosa 9027* (Table XII). In contrast to this finding,
Barron (14,90), employing the same species found an inhibition of 25$ with
1 x l O - 4 ! U0 2+ + , and of 69$ with 4 x HT 4 M U0 2
+ + •
Table XII The effect of U02(N03)2 on the oxidation of gluoose by 22 hour cells harvested from a gluoose mineral medium.
Inhibition (per cent)
5 13 17
T h e inhibitory action of uranyl nitrate on the oxidation of the
dicarboxylio acids, succinate, fumarate and malate is shown in Table XIII*
Succinate and fumarate oxidations were decreased only -when a relatively
high concentration of U02 was used, while malate was completely unaffected •
Barron (14) found i t necessary to employ 1 x 10"^ M uranyl nitrate to
inhibit succinate oxidation of E. coli, but at this concentration P.aeruginosa
was agglutinated. The absence of inhibition with malate may be explained
by the fact that this and other hydroxypolycarboxylic acids are able to
reverse uranium inhibition of isolated enzyme systems (131). Barron et a l ,
(14) also observed no inhibition of malate oxidation in yeast, when they
used concentrations of U0 2+ completely toxic to glucose oxidation*
(Molar) * ( »
— 1050 1 x 10"*4 1000
2.5 x 10"4 '925 4 x 10"*4 875

Table XIII
The effect of U02(NQ3)2 on the oxidation of succinate, fumarate
and malate by 21 hour oells harvested from an ammonium phosphate
medium containing one of these compounds as sole carbon souroe.
Experiment Humber®
Experimental Conditions
U0 2(N0 3) 2
(molar) 4 ° (N) Inhibition (per cent)"
I Succinate it it n
1 x 10" 4
2.5 x l O " 4
4 x 10" 4
807 eo4 7853 616
0 3 24
II Fumarate tt it
7.5 x 10~ 5
1 x l O " 4
510 518 480
0 6
III Fumarate ti 4 x l O " 4
522 320 39
IV Malate n tt
1 x 10" 4
4 x 10" 4
1100 1200 1000
0 9
Higher concentrations of uranyl nitrate (2.5 x 10 M and 4 x 10" 4 M)
were required for significant inhibition cf the oxidation of pyruvate and
oc -ketoglutarate (Table XIV), and of acetate (Table XV). The most striking
data were obtained with isooitrate (Figure XVI), i n which even the lowest
concentration (7.5 x 10"^ M) caused greater than 50$ inhibition, and
higher concentrations ( l x 10*"4 M and 2.5 x 10" 4 M) were almost completely
inhibitory. Citrate oxidation, on the .other hand, was but slightly
affected by these conoentrations (Table XV).

- &7; -
Table XIV
The effect of U02(N03)2 on the oxidation of pyruvate and
<-ketoglutarate by 22 hour oells harvested from an ammonium
phosphate medium containing either of these compounds as C souroe
Experiment Number.
Experimental Conditions
UO2(N0?)2
(molar) Q02(N) Inhibition
(per cent)
I Pyruvate 11 tt n
1 x lO**4
2.5 x 10-4 4 x l O - 4
1485 1251 10E8 1034
16 31 30
II <*. -ket oglut ar at e 11 it 7.5 x 10"5
1 x 10"4
538 488 437
9 19
III «* -ketoglutarat e it 11
1 x 10"4
4 x lO""4
1375 1115 920
19 33
Table XV
The effect of 1102(^03)2 °n the oxidation of acetate and
citrate by 24 hour oells harvested from an ammonium phosphate
medium containing either of these compounds as sole C souroe.
Experimental Conditions
Aoetate u ti tt
Citrate tt ti n
U02(NO,)2
(molar)
1 x 10"* 2.5 x l O - 4
4 x 10**4
1 x 10~* 2.5 xlO" 4
4 x lO" 4
°2 Uptake (ul)
(at 120') • 280 248 193 170
(at 105') 158 160 142 112
Inhibition (per cent)
11 31 39
0 10 29

- 58 -
Figure XVI. The inhibitory effect of U0 2(N03) 2 on the oxidation of isocitrate by 22 hour cells harvested from an isocitrate ammonium phosphate medium. Warburg cups contained 0.5 ml. c e l l suspension, 1.5 ml. M/7 veronal buffer pH 7.2, 0.3 ml. U02(N0 3) 2, 0.2 ml. substrate.. Final volume was 3.0 ml.

- 59 -
Attempts were made to reverse the inhibition of uranyl nitrate,
by the addition of phosphate, which forms a strongly associated complex
with the toxio ion. The oxidizable substrate was added .to the main oomparfc-
ment of the Warburg cup (instead of to the side-arm), together with veronal
buffer, cells and water to volume, Uranyl nitrate was added to one side-arm,
and phosphate buffer to the other (0.3 ml. of each). The UOg++ was added
from the first side-arm when the cells had been metabolizing their substrate
at a constant rate for 30 minutes. Fifty minutes later, phosphate was
added from the second sidefearm. In experiments with glucose and with succin
ate oells, no significant decrease in oxidation was observed when the
uranium'was added in this manner, and hence, no reversal of inhibition
was effected by phosphate. Apparently, i t is necessary to have the
uranium and oells in oontact for well over an hour to obtain measurable
inhibition. This finding is in contradiction to the observation^ of
Rothstein (123,124) and Barron (14,90), who obtained an instantaneous
uranium uptake by yeast cells. In addition, Barron (14) always used 38° C.
for bacterial studies, while a l l experiments with P. aeruginosa ATC 9027
have been oonducted at 31° C. At the latter temperature, a much slower
rate of reaction would be operative.
Uranyl nitrate as an inhibitor on dried cell preparations provided
additional evidence that the cell membrane has been altered duringthe drying
process. Concentrations of 4 x 10""4 M and 5 x 10""4 M which had effected a
25$ inhibition with whole resting cells, showed no toxic effect on dried
cells,-in fact, a slight stimulation was observed (Table XVI).

- 6 0 -
Table XVI
The effeot of UOgCNOjOg o n ^he oxidation of succinate by dried
oells harvested from a succinate ammonium phosphate medium.
Experiment UOgCNOg^ Oxygen Uptake % Inhibition
Number (molar) At 60» At 120* At 60* At 120'
17 65 £ x 10-4 21 75.
II — 30 65: S x 10"4 33 70 0 0

DISCUSSION
From studies of malonate inhibition in P. aeruginosa it may be
concluded that this organism possesses the enzyme succinic dehydrogenase*
Additional evidence for a close relationship amongst the compounds succinate,
fumarate and malate was also provided. While malonate strongly inhibits
both the rate and extent of oxidation of suocinate, only rate is affected
in the case of both fumarate and malate. The rate of «W:etoglutarate
oxidation was also inhibitadj an observation which indicates an enzymatio
relationship between this compound and succinates Non-inhibition of
pyruvate oxidation might suggest that this intermediate is not oxidized
via the same meohanism (the Krebs cycle) as those listed above. However,
such a conclusion seems unlikely in the light of other findings (157),
nor can i t be drawn on the basis of a single experiment.
Turning now to uranium, i t is believed to inhibit cellular
metabolism indirectly by combining with the cell surface,, thus causing
decreased permeability to the passage of oxidizable substrates. According
to Knaysi (66,67,68) and others (135), the cell membrane consists of a
lipoid-protein system, with associated ribonucleic acid. Apparently,
uranium complexes with the protein constituents of the membrane, through
certain "active" groups, which Barron (90) and Rothstein (123,124) suggest
may be phosphates, phosphate esters or polyphosphates. Moreover, this
uranium-protein complex can be reversed by the addition of inorganic phosphate
at low concentrations. These facts point to some relationship between
phosphorylation and the "active" transfer of substrates across the cell
membrane. Additional support for this suggestion was found by Gale (46)
in that the passage of. glutamic acid into the cell of Streptococcus ffigpalis
is dependent on an exergonic metabolic reaction involving phosphate.

- 62 -
It also seemed possible that the energy for transport might arise
from a coupling between phosphorylation at the "active" sites in the oell
surface, and the oxidation of substrate - once the reaction had been primed
by energy from some other coupled reaction (44)• That there is a coupling
between phosphorylation and the oxidation of glucose and of certain Krebs
intermediates has been shown by a number of investigators (9,11,32,97,98,
109,144), in studies made mainly with aaimal tissue. Evidence for the
oxidative formation of energy-rich phosphate bonds in bacterial metabolism
has been presented by Lipmann (83),with Lactobaoillus delbruckij by TissiSres
(150) with Aerobaoter aerogenes, and by Hersey & Agl (57) with cell-free
preparations of E,coli.
When the present study was undertaken, i t was hoped that some
association might be uncovered between uranium inhibition and the "active"
(possibly phosphorylative) transfer of substrate across the oell membrane.
However, i t is very difficult to interpret;, the results of the experiments,
since the concentration of uranyl nitrate required for significant inhibition
was not the same in each case. For example, isocitrate oxidation was strongly
inhibited by a l l concentrations used, while malate showed no inhibition what
soever. Inhibition falling between these two extremes was observed with the
other substrates. Similar findings were reported by Rothstein & Meier (125)
in work on molybdate inhibition of surface phosphatases in yeast. It may be
that the protein with which the uranium complexes is not only "active"1 in
the transport of substrate, but is itself the enzyme for metabolizing the
substrate. Thus, different enzymes would show different sensitivities
to uranium. Such an hypothesis i s , however, far beyond the present bounds
of experimental verification.

- 63 -
The greater part of the work on metabolic inhibitors has dealt
with the inhibition of isolated systems. The application of inhibition
studies to the metabolism of whole oells would appear to be open to
question, especially when so l i t t l e i s known of a cell's actual metabolic
pathways, A reduction i n respiration may imply that an inhibitor i s
acting on one or more of the enzymes concerned with carbohydrate metabolism
(162). In addition, an inhibitor may cause an organism to use an alternative
metabolic pathway. Until more i s known of the direct and alternative
pathways i n bacterial metabolism, the use of a non-specific inhibitor such
as uranium is precluded.

- m -
SUMMARY <
Resting cells of P. aeruginosa harvested from a mineral medium,
in which succinate or fumarate or malate was the sole carbon source,
metabolized al l three compounds immediately and at a constant and rapid
rate, thus indicating an enzymatic inter-relationship.
The oxygen consumption of dried oells was considerably lower
than that observed with resting cells. These preparations did not
require cytochrome c or pyocyanine as a carrier system. Optimum pH for
suocinic dehydrogenase in succinate and acetate dried cells lay between
pH 7.0 and 8.0 •
Moderately high concentrations of malonate were neoessary to
inhibit succinate oxidation by resting cells at pH 7.0, but when the pH
was lowered, maximum inhibition v/as effected by low concentrations.
P. aeruginosa possesses an adaptive enzyme system for metabolizing the
inhibitor. Dried cell preparations were strongly inhibited by low
concentrations of malonate at pH.7.0, thus indicating that permeability
is an important factor in malonate inhibition with P. aeruginosa.
Low concentrations of uranyl nitrate caused varying degrees of
inhibition with resting cells in the presence of the Krebs intermediates
or glucose, the most striking effect being observed in the case of
isocitrate oxidation. No reversal of uranium inhibition by phosphate
was obtained.

- 65 -
BIBLIOGRAPHY
7.
3.
9.
10.
11.
12.
13.
Ackermann, W. W. and Potter, V.R. Enzyme inhibitors in relation to chemotherapy. Proc. Soc. Exper. Biol. & Med. 72: 1-9. 1949.
A j l , S.J. Biotin in succinic acid oxidation. Enzymologxa 14: 1.
Ajl, S.J. Acetic acid oxidation by Escherichia coli and Aerobaoter aerogene3, J. Baot* 59: 499-507. 1950.
A j l , S.J. and Workman, C.H. Replacement of carbon dioxide in heterotrophic metabolism. Arch. Biochem. 19: 483-492. 1948.
Ajl, S.J. and Workman, CH. On the mechanism of carbon dioxide replacement in heterotrophic metabolism. J. Bact. 57: 579-593. 1949.
Albaum, H.G. and Commoner, B. The relation between the 4-carbon acids and the growth of oat seedlings. Biol. Bull. 80:314-323. 1941.
and Szent-Gyorgyi, A. uber die Bedeutung der Fumarsaure fur die Tierische Gewebsatmung, Z. physiol.-Chem. 244: 105-152. 1936.
and Szent-Gyorgyi, A. Uber die Bedeutung der Fumarsaure fur die Tierische Gewebsatmung. Z. physiol. Chem. 236: 1-69. 1934.
Ball,. E.G. and Cooper, 0. The activity of succinate oxidase in relation to phosphate and phosphorus compounds. J. Biol, Chem. 180t 113-124. 1949.
Barron, E.S.G. Mechanisms of carbohydrate metabolism. Adv. in Enzymology 3: 149-189. 1943.
Barron, E.S.G. Cellular metabolism and growth* in The Chemistry and physiology of growth. Princeton University press. 1949.
Barron, E.S.G., Ardao, M.I. and Hearon, M, The mechanism of acetate oxidation by Corynebacterium creatinovonans. Arch. Biochem. 29t 130-153. 1950.
Barron, E.S.G., Ardao, M. I., and Hearon, M. Regulatory mechanisms of cellular respiration. III. Enzyme distribution in the c e l l . Its influence on the metabolism of pyruvic acid by baker's yeast. J. Gen. Physiol. 34t 211-224. 1950,
1950.
v.

- 66 -
Barron, E.S.G., Muntz, J.A. and Gasvoda, B. Regulatory mechanisms of cellular-respiration. I. The role of cell membranes: Uranium -inhibition of cellular respiration* J. Genl. Physiol, 32: 163-178, 1948,
15. Barron, E.S.G., Link, G.K.K., Klein, R.M., and Michel, B.E. The metabolism of potato slices, Aroh. Biochem, 28: 377-388, 1950,
16. BSaskett, A.C. and Hinshelwood, Sir C, The utilization of carbon souroes by Bact, lactis aerogenes. I, General survey, Proc, Roy, Soc, B, 136: S20-535. 1949.
17. Baskett, A.C. and Hinshelwood, Sir C, The utilization of carbon sources by Bact, lactis aerogenes, II. The significance of the tricarboxylic acid cycle, Proo, Itoy, Soc, B, 136: 556-543, 1949.
18. Bernstein, D,E, Studies on the assimilation of dicarboxylic acids by Pseudomonas saccharophila. Arch, Biochem, 3* 445-458© 1943.
19. Bonner, J. Biochemical mechanisms in the respiration of the Avena
coleoptile. Arch. Biochem. 17t 311-327. 1948.
20. Bonner, J. Plant biochemistry. Academic Press Ine. New York. 1950.
21. Bonner, J. and Wildman, S.J. Ehzymatio mechanism in the respiration of spinach leaves. Arch. Biochem. 10: 497-518* 1946.
22. Borei, H. Reactivation and protection of cytochrome oxidase preparations. Bioohem. J . 47: 227-233. 1950.
23. Burton, M.Q., Campbell, J.J.R. and Eagles, B.A. The mineral requirements for pyocyanin production. Can. J. Research, C, 26: 15-22.1948.
B4. Butterworth, J. and Walker, T.K. A study of the mechanism of the degradation of -citrio acid by B. pyocyaneus. Part I. Biochem. J. 23: 926-935. 1929.
25. Callow, A.B. The oxygen uptake of bacteria. Biochem. J. 18: 507-518. 1924.
26. Campbell, 0.J.R. (unpublished).
27. Campbell, J.J.R. and Norris, F.C. The intermediate metabolism of Pseudomonas aeruginosa. IV. The absence of an Embden-Meyerhof system as evidenced by phosphorus distribution. Can. J, Researoh, C, 28: 203-212© 1950.

- 67 «
28* Campbell, J.J.R., Norris, F.C. and Norrisj M.E. The intermediate metabolism of Ps eudomonas aeruginos a . • I I , Limitations of simultaneous adaptation as applied to the identification"of acetic aoid, an intermediate in glucose oxidation. Can. J. Research, C, 27: 165-171. 1949.
29. Campbell, J.J.R. and Stokes, F.M. Evidence for the presence of a tricarboxylic acid cycle in Pseudomonas aeruginosa. ' J. Biol. Chem. (in press).
30. Chugtai, I . D . , Pearoe, A.A., "and Walker, T.K. Detection of acetio acid in A. niger growing on-a gluoose medium* Nature 160: 573. 1947. • -
31. Cockrane, V.W. and Dimmick, I . The metabolism of species of Streptomyees. The formation of succinic and other aoids*
J. Bact. 58s 723-731. 1949.
32. Colowick, S.F., Kalokar, H.M. and Cori^ CF. Glucose phosphorylation and oxidation in cell-free tissue extracts. J.Biol.Chem. 137* 343. 1941.
- • ' I
33. Cook,. R.P. A comparison of the dehydrogenations produced by B.ooli communis in the presence of oxygen and methylene blue. ' '»'• [*' Biochem. J. 24: 1538-1550. 1930.
34. Cook, R.P. and Alcock, R.S. • The effect of hydrogen concentration on: some oxidations by B.coli communis. Bioohem. J. 25: 523-533. 1931.
35. Das, N.B. Studies on the inhibition: of the succinic and lactio-malic dehydrogenases. Biochem. J. 31: 1124-1130. 1937.
36. Dickens, F. Mechanisms of carbohydrate oxidation. Nature 1381 "1057. 1936.
37. Diokens, F. and Glock, G.E. Direct oxidation of glucose-6-phosphate by animal tissues. - Nature 166t 33. 1950.
38. Dickens, F. and Mcllwain, H. Phenazine compounds as carriers in the hexose monophosphate system. Biochem. <J. 32: 1615-1625. 1938.
39. Elema, B.. and Sanders, A.C. Studies on the oxidation-reduction of pyocyanine. 1. The biochemical preparation of pyocyanine. Rec. trav. chim. 50t 796-806.-1931.
40. Elli-fssfct, K.A.C. The possible role of intermediate, metabolites-as hydrogen carriers, p.33 in-Symposium on respiratory enzymes* University of Wisconsin Press. 1942*

- 68 -
41. Evans, E.A. Jr., Vennesland, B., and Slotin, L. The mechanism of carbon dioxide fixation in cell-free extracts of pigeon liver. J. Biol. Chem. 147: 771-784. 1943.
42. Foster, J.W. and Carson, S.F. Metabolio exchange of carbon dioxide r with carboxyls and oxidative .synthesis of C* dicarboxylic acids.
Proc.Nat. Acad. Soi. 36: 219-229. 1950.
43. Jfciedheim, E. Pyocyanine, an aocessory respiratory enzyme. J. Exper* Med. 54: 207-221. 1931.
44. Friedkin, M. and Lehninger, A. L. Esterification of inorganic phosphate coupled to electron transport between dihydrodiphosph-pyridine nuoleotide and oxygen I. J. Biol. Chem. 178: 611-623. 1949.
45. Gale, E.F. Chemical activities of bacteria, (p. 39). Academic Press Inc. Nwsr York. 1948.
46. Gale, E. F. The assimilation of amino acids by Bacteria. I. The passage of certain amino acids aoross the cell wall and their concentration in the internal environment of Streptoeooous faeoalis. J. Gen. Microbiol. It 53-76. 1947.
47. Gaughran, B.R.L. Temperature activation of certain respiratory enzymes of stenothermophilic bacteria. J. Gen. Physiol. 32: 313-327. 1949.
48. Goddard, D.R. and Meeuse, B.J.D. Respiration of higher plants. Ann. Rev. Plant Fhysiol. 1: 207-232. 1950. .
49. G&zsy, B. and Szent-Gyorgyjtj A* Uber den Mechanismus der Hauptatmung des Taubenbrustmuskels. Z;. physiol. Chem. 224: 1-11, 1934.
50. Green, D.E. The citr i o a c i d cycle and the cyclophorase.system. pp. 201-205, in Lardy, H.A. Respiratory enzymes. Burgess Publishing Company, Minneapolis, Minn. 1950.
51. Green, D.E.., Loomis, W.F. and ..Auerbach, V.H. Studies on the cyclophorase system. I. The complete oxidation of pyruvio acid and water. J. Biol. Chem. 172: 389-402. 1948.
52. Guggenheim, E. The effeot of various ohemioal and physical agents on the dehydrogenating enzymes of Eberthella typhosa. Biochem. J. 39: 419. 1945.

- 69 -
53. Guggenheim, K. Investigations on the dehydrogenating properties of certain pathogenic obligate anaerobes. J. B act. 47s 313-321. 1944.
54. Gunsalus, I. C. (unpublished).
55. Gurin, S. and Crandall, D. I. The biological oxidation of fatty acids. Cold Spring Hbr. Symp. Quant. Biol. 13: 118-128. 1948.
56. Harvey, S.H. The carbohydrate metabolism of Trypanosoma hippicum. J. Biol.-Chem. 179: 435-463. 1949.
57. Hersey, D.F. and Aj l , S.J. Phosphorylation due to the oxidation of succinic acid by cell-free extracts of Escherichia c o l i . J. Gen. Physiol. 34: 295-304. 1951. ' ' ;
58. Hogeboom, G.H. and Schneider, W.C. Sonic disintegration of isolated liver mitochondria. Nature 166t 302. 1950.
59. Kalnitsky, G. and Barron, E.S.G. The effect of fluoroacetate on the metabolism of yeast and bacteria. J. Biol. Chem. 170: 83-95. 1947
60. Kalnitsky, G., Wood, H.G. and Werkman, CH. Carbon dioxide fixation and succinic acid formation, by a cell-free enzyme preparations of Escherichia c o l i . Arch. Biochem. 2:: 269-281. 1943.
61. Karlsson, J. L. Metabolic studies of Azotobacter agilis by the use of a mutant deficient in pyruvic oxidase. J. Biol. Chem. 183s 549-560. 1950.
62. Karlsson, J.L. and Barker, H. A. Evidenoe against the occurrence of a tricarboxylic acid oycle in Azotobacter agilis. J. Biol. Chem. 175: 913-921. 1948.
63. Karstroiii, H. Enzymatisohe Adaptation bei Mikrborganismen. Ergeb. Bnzymforsch. 7: 350-376. 1937.
64. Keilin, D. and Haftree, E.F. Prepared ion of pure cytochrome o from heart muscle and some of its properties. Proc. Roy. Soc, B, 122: 298. 1937.
65. Keilin, D. and Hartree, B.F. Succinic dehydrogenase-cytochrome system of cells. Intracellular respiratory system oatalyzing aerobic .oxidations of succinio acid. Proc. Roy. Soc, B, 129: 277-306. 1940.
66. Knaysi, G>- On the structure,-morphology, nature and functions of the cytoplasmic membrane in the bacterial oell. J.Bact. 51: 113-121. 1946*

- 70 -
67. Knaysi, G. Cytology of bacteria. II. Bot. Rev. 15s 106-151. 1949.
68. Knaysi, G. Elements of bacterial cytology. Comstook Publishing Co. Inc.,;Ithaca, N.Y. 1950. -
69. Khoop, F. Klin* Wochschr. 2s 60. 1923. As.cited in Krebs, H.A. Adv. in Enzym. 3:- 191-252. 1943.
70. Korkes, S.,, Stern, J.R., Gunsalus, I.C.and Ochoa, S. Enzymatio „ synthesis of citrate from pyruvate and oxalacetate. Nature 166$ 439, 1950.
71. .Kraapitz, L.0. and Workman, C.H. The enzymatic decarboxylation of oxalaoetate. Bioohem. J . 3St 595-602. 1941.
72. Krebs, H.A. ,Biological oxidation of carbohydrate. Adv. in Ehzym. 3s 191-253. 1943. , ,
73. Krebs, H.A. and Sggleston, L.V., Bhe oxidation of pyruvate in pigeon breast muscle. Biochem. J. 34s 442-459. 1940. . ...
74. Krebs, H.A. and Eggleston, L.V. Biological synthesis of oxalacetio acid from pyruvic acid and carbon dioxide. 2. The mechanism of carbon dioxide fixation in. Propionio acid bacteria. Bioohem. J. 35* 676-687. 1941.
75. Krebs, H.A. and Johnson, W.A.. The role of citric aoid in intermediate metabolism in animal tissues. Enzymologia 4's 148-156. 1937.
•76. Kubowitz, F. and Haas, E. Ausbau der phbtochemischen Methoden zur Untersuohung des sauerstoffbetragenden Fermentes. Biochem.Z.-255s 247. 1932.
77. Kun, E. and Abood, L.G., Biochemical properties of suocinoxidase from Salmonella aertrycke. J. Biol. Chem. 180s 813-823. 1949.
78. Laties, G. G. The oxidative formation of succinate in higher plants. Arch. Biochem. 22s 8-15. 1949.
79. Lehninger, A. L. A. quantitative study of the products of fatty acid oxidation in liver suspensions. J. Biol. Chem. 164s 291-306. 1946.
80. Lehninger, A.L. The role of metal ions in enzyme systems. Physiol. Revs. 30s 393. 1950.

- 71 -
81* Lenti, C* • Oxidation of pyruvic acid by Baoterium coli communis* Boll, soo. i t a l * biol* spar* 22: 332-533. 1946. As cited in Chem. Abstr. 41t 7871* 1947.
82. Lewis, R.W. Mutants of Neurospora requiring succinic acid or a biochemically related acid-for •growth* Amer* J* Bot. 35: 292-295. 1948.'
83i: Lipmann, F, An analysis of the pyruvic acid oxidation system* Cold Spring Hbr* Symp. Quant. Biol* 7:"248. 1939. '
84* Lockwood, L.B. and Stodola, F. H. PrJLiminary studies on the production of alpha-ketoglutario aoid by Pseudomonas fluoresoens* J. Biol. Chem. 164: 81-85. 1946. •
854 Ifliroff, A. and Cailleau, R. Production bacteriehe directe d'acid pyruvique aux depens de l'acid malique* Compt. rend. l'acad.Soi*, Paris. 224: 678-679. 1947.
86. Malachowski, • R. and Maslowski,- M. • B 9 r. chem. ges. 61B: 2521. 1928. As cited in Umbreit W.W., Burris, R.H. and Stauffer, J*F, Manometric
• * techniques and tissue metabolism* Burgess Publishing Company* 1949* 87* Militzer, W,, S onderegger,- T*B. and Tuttle, L.C. Thermal enzymes.
Aroh. Biochem. 24s 75-82. 1949.
88* Militzer,' W., Sonderegger,'T.B., Tuttle, L.C. and Georgi, CE. Thermal enzymes. II* Cytochromes* Arch* Biochem* 26: 299-306* 1950*
89* Monod, J* Growth of bacterial cultures* Ann* Rev. Microbiol* 3: 371-394. 1949. — -
90. Muntz, J.A., Singer, T.P. and Barron, E.S.G. Effect of uranium on the'metabolism^of yeast and bacteria. Metallurgical laboratory, Declassified Report CH 3716. 1947* " - '"' - "
91* Ney, P.W. A study of the intermediate metabolism of Pseudomonas aeruginosa. 'Thesis for M.S.A. * Dept. of Dairying, U.B.C-1948*
92* Norris, F* C* "The intermediate metabolism of Pseudomonas aeruginosa: A study of a new pathway of gluoose oxidation* Thesis for"1.1.A.'' Dept. of Dairying, U.B.C. 1949.
93. Norris, Fi C* "(unpublished)* 94. Norris, F.g*, Campbell, J.J.R. and Ney, P*W, The intermediate
metabolism of Pseudomonas aeruginosa* I. The status of the endogenous respiration* Can. J. &esearch, C, 27: 157-164. 1949*

- 72 -
95. Norris, F.C. and Campbell, J.J.R. The intermediate metabolism of Pseudomonas aeruginosa. III. The application of paper chromatography to the identification of gluoonic and 2-ketogluconic acids, intermediates in glucose oxidation. Can. J. Research, C. 27: 253-261. 1949.
96. Novella, G.D. and Lipmann, F. The catalytio function of coenzyme A in citric acid synthesis. J. Biol. Chem. 182: 213-228. 1950.
97.. Ochoa, S. "Coupling" of phosphorylation with, oxidation of pyruvic acid in brain. J. Biol. Chem. 138: 751-773. 1941.
98. .Ochoa, S. Efficiency of aerobic phosphorylation in cell-free heart„ extracts. J . B i o l . Chem..151: 493-505. 1943.
99. Ochoa, S. Biosynthesis of tricarboxylic acids by carbon ;dioxide fixation. III. Enzymatic mechanisms. J. Biol..Chem. .174: 133-'., 157. 1948. . . . . . . .
100. Ochoa, S. Biological mechanism of carboxylation and. decarboxylation. Physiol. Rev. 31: 56-106, 1951. . . " .
101. Ochoa, S. and Weisz-Tabori, E. Biosynthesis of tricarboxylic acids by carbon dioxide fixation., II. Oxalosuccinic,carboxylase. J. B i o l . Chem. 174:.123-132. 1949.
102. Oginsky, E.L.,. Smith, P.H. and Umbreit, W.W. . The action.of streptomycin. I. The nature of the reaction inhibited., J. Bact. 58: 747-758. 1949.
103. .Ogston, A.G. Interpretation of experiments on metabolic prooesses using isotopio tracer elements. Nature 162: 962. 1948.
104. Ohlsson, E. " Skand. "-Arch. Physiol. 41: 77 • 1921. As cited in Lardy, H.A. Respiratory enzymes. Burgess Publishing Company, Minneapolis, Minn. 1949.
105. .Pappenheimer, A.M. Jr.,. and Hendee, E.D. Diphtheria toxin. IV. The iron enzymes of Corynobacterium diphtheriae and their possible relation to diphtheria toxin. J.Biol.Chem. 171: 701-713.. 1947.\
106. .Pappenheimer, A.M. Jr. and Hendee, E.D. Diphtheria toxin. V, A comparison between the diphtherial succinoxidase system and that of beef heart muscle. J, Biol, Chem, 180: 597-609, 1949. . . • • -

- 73 -
107-. .Pardee, A.B. and Potter, V.R. • Malonate inhibition of oxidations in the Krebs tricarboxylic acid cyole. - J• Biol, Chem, 178i 241-250, 1949.
108. .Plaut,; G.W.B. and Lardy, H.A. The oxalacetate decarboxylase of Azotobaoter vinelandii. _ J. Biol. Chem. 180: 13-27. 1949,
109. Potter, V.R. Coupling between phosphorylation and oxidation of- the 4-carbon acids in rat kidney homogenates. Aroh.Biochem. 6: 439-453. 1945.
110. .Potter, V.R. The assay of animal tissues for respiratory enzymes. V, The malic dehydrogenase system. J. Biol. Chem. 165s 311-324, 1946. . i •• » - . . . - -
111. Potter, V.R. And DuBois, K.P. Studies on the mechanism of hydrogen transport in animal tissues. VI, Inhibitor studss- with succinic dehydrogenase, J, Gen, Physiol, 26: 391-405, 1943.
112'. .Potter, V.R. and Schneider, W.C. Studies on the mechanism of hydrogen transport in-animal-tissues• J. Biol. Chem. M 2 t 543-557. 1942.
113. Potter, V.R. and Schneider, W.C. -The assay of animal tissues* for respiratory- enzymes. I-I. Succinic dehydrogenase and cytochrome oxidase. J. Biol. Chem. 149: 217-227. 1943.
114. Quastel, J.H. .On the fermentation of the unsaturated dicarboxylic acids. I. Fumaric acid. Bioohem. J, 18:. 265-280. 1924.
115. Quastel^ J.H. Dehydrogenations produoed by resting bacteria. IV. A theory of the mechanism of oxidations and' reductions in vivo. Bioohem. J, 20: 166-194. 1926.
116. Cvuastel, J.H. and Wheat ley, A.H.M. - Biological oxidations in the succinic acid series. -Biochem. J. 25: 117. 1931.
117*. Quastel, J.H. and Whetham, M.D. The equilibria existing between succinic, fumaric and malic acids in the presence of resting bacteria. Biochem. J. 18: 519. 1924,
118. Quastel, J.H, and Whetham, M.D. Dehydrogenations produoed by resting bacteria. - I. B.ooli communis. Biochem. J. 19P 520.1925.
119. Quastel, J.H. and Wooldridge, ¥f.R. .Some properties of the-dehydrog- . enating enzymes of bacteria, Bioohem, J, 22: 689-703, 1928,

- 74 -
120. Randies, C. I# and Birkaland, J.M. The effects of malic and malonic acids on methylene blue-reduction by bacteria. J. Bact. 54s 275. 1947. - • ,
121« Reed, G.B. and Boyd, E.M. Pyocyanine and growth potential changes of.Psa pyooyaneus. Can. J. Research 8: 173. 1932.
122. Robertson, W.R. The preparation of sodium pyruvate. Science 96s 93-94. 1942.
123. Rothstein,~A., Ffenkel, A. and Larrabee, C. The relationship of the cell surface to metabolism. I l l , Certain characteristics of the uranium complex with cell surface groups of yeast. jT. Cell. & Comp. Physiol. 32t 261-274. 1948.
124. Rothstein, A. and Larrabee, C. The relationship of the cell surface " '" to metabolism, I I , The cell,surface of yeast as the site of
inhibition of glucose metabolism by uranium. J. Cell, & Comp. Physiol. 3.2s 247-260. 1948,
125. Rothstein, A. and Meier, R, The relationship of the cell surface to metabolism, I V , The role of cell surface phosphatases of yeast,' J, Cell. & Comp, Physiol. 34s 97-114, 1949,
126. Rudolph, G.'and BBarron, E.S.G, Biochim, et Biophys. Acta 5s 59. 1950,
127. Ryan, F.J., Tatum, E.L. and Giese, A.G, The 4-carbon respiratory system and growth of tha mold Neurospora, J. Cell. & Comp. Physiol, 23s 83-94. 1944. 1 . . .
128. Saz, H, and Krampitz, L,D, Aoetate oxidation in Micrococcus'lysodeikticus. Bact. Proc; 1950s 126, 1950.
129. Schneider, W.C. and Hogeboom, G.H. Cytochemical studies of mammalian tissuesj the isolation of cell'components by differential centrifug-ation. Cancer Reseafoh lis 1-22. 1951. • ' '
130. Shattock, S.G. and Dudgeon, L.S, Certain results of drying non-spori ng bacteria in a charcoal liquid air vaouum. Proc. Roy. Soc, B, 85s 127-138. 1912.'
131. Singer, T. P., Muntz, J.A., Meyer, J., Gasvoda, B, and Barron, E.S.G. The reversible inhibition of enzymes by uranium. Metallurgical Laboratory, Declassified Report CH-3739. 1947.
132. Slade, H.D. and Werkman, C.H. Assimilation of acetio and suocinioacids containing heavy oarbon by Aerobaoter indologenes. Arch. Biochem, 2s 97-111. 1943. :

- 75 -
133. Slater, E.-C. Arespiratory oatalyst required.for the reduction of cytochrome c by cytochrome b. Biochem. J. 45: 14-32. 1949.
134. Sleeper, B.P., Tauchida, M., and Stanier, R.Y. The bacterial oxidation of aromatio compounds'. II. The preparation of enzymatically active dried cells and the influence thereon of prior patterns of adaptation. J. Bact. 59: 129-133. 1950.
135. Society for General Microbiology. The nature of the bacterial surface. 8th General Meeting of the society. April 1949.
136. Speck, J.F., Moulder, J.W. and Evans, E.A. Jr. The biochemistry o f the malaria parasite. V.« Mechanism of pyruvate oxidation in the malaria parasite. J. Biol. Chem. 164: 119-144. 1946.
137. Stephenson, M. Bacterial metabolism. Longman's, London. 3rd Edition 1949. page 25.
138. Stern, J.R. and Ochoa, S. Enzymatic synthesis of citric acid by condensation of acetate and oxalacetate. J. Biol. Chem. 179: 491-492. 1949.
139. Stern, J.R. and Ochoa, S. Enzymatic synthesis of citric acid. Fed. Proc. 9: 234. 1950.
140. Stern, J.R., Shapiro, B., and Ochoa, S. Synthesis and breakdown of citric acid with crystalline condensing enzyme. Nature 166: 403. 1950. '
141. S t i l l , J. L. and Kaplan, B.H. Localization of oxidases. Exper. Cell! Research. 1: 403-409. 1950.
142. Stokes,'F.N. and Campbell, J.J.R. The oxidation of glucose and gluconic acids by dried cells of Pseudomonas aeruginosa. Arch. Biochem. 30: 121. 1951'. '
143. Stoppani, A.O.M. On the mechanism of suocinoxidase inhibition. Enzymologia 13: 165-176. 1949.
144. Street, H.E. The role of high-energy phosphate bonds in biosynthesis . Soience Progress 38: 43-66. 1950.
145. Stumpf, P.K. pyruvic oxidase of Proteus vulgaris. J. Biol. Chem. 159: 529-544. 1945. '
146. Sullivan, M. Synthetic culture media and the biochemistry of bacterial pigments.- J. Med. Research 14: 109-160. 1905.

- 76 -
147. Szent-Gyorgyi, A. von. On oxidation, fermentation, vitamins, health and disease. Williams & Wilkins, Baltimore. 1939.
148. Thimann, K. V. and Bonner, W. D. Jr. Experiments on the growth and inhibition of isolated plant parts. I. The action of iodoacetate and organic acids on the Avena coleoptile. Amer. J. Bot. 35: 271-281. 1948.
149. Thunberg, T. • Skand. Arch. Physiol. 40s 1. 1920. As cited in Krebs, H.£. Adv. in Bnzymol. 3 : 191-252. 1943.
150. Tissieres, A. Effect of azide on the respiratory system of Aerobaoter aerogenes. Nature 166s 1115. 1950.
151. Toeniessen, E. and Brinkmann, E. Z. physiol. Chem. 187$ 137. 1930. As cited in Krebs,' H. A.' Adv. in Enzymol.3s 191-252. 1943. ' • ;
152. Turner, J.S. and Hanly, V. Malonate and plant respiration. Nature 160$ 296. 1947.
153. Umbreit, W. W. Metabolism of lEoroorganisms. Ann. Rev. Microbiol. 3s 81-96. 1949.
154. Umbreit, W. W. A si-be of action of streptomycin. J. Biol. Chem. 172s 703-714. 1949'.
155. Umbreit, W.W., Burris, R.H. and Stauffer, J.F. Manometric .. techniques and tissue metabolism. Burgess Publishing Company, Minneapolis, Minn. 1949.
156. Walker, T.K., Subramaniam, "V., Stent, H.B. and Butterworth, J. A study of the mechanisms of degradation of citric acid by B. pyooyaneus. (Pseudomonas pyocyanea). II. Action of B. pyooyaneus"on succinic acid. Biochem. <J. 25: 129-137. 1931.
157. Warburton, R.H., Eagles, B.A., and Campbell, J.J.R. The intermediate metabolism of Pseudomonas aeruginosa. V. The identification of pyruvate as an intermediate in gluoose oxidation. Can. J. Research
, (in press).
158. Wood, H. G. The fixation of carbon dioxide and the inter-relationships of thet ricarboxylic acid cycle. Physiol. Rev. 26: 198-247. 1946.
159. Wood, H.G. and Werkman, C.H. The relationship of bacterial u t i l i z ation of carbon dioxide to succinic acid formation. Biochem. J. 34s 129-138. 1940.

77 -
160. Wood, H.G., Workman., C.H., Hemingway, A. and Nier, A.O, Mechanism of fixation of carbon dioxide in the Krebs cycle, J. Biol. Chem, 139: 483-484. 1941.
161. Wooldridge, W.R., Knox, R. and Glass, V. Variability in the activity of bacterial enzymes, I, The effect of the age of oulture. Bioohem, J. 30 : 926-932. 1936.
162. Work, T.S. and Work, -S. The basis of chemotherapy, Oliver & Boyd Limited, Edinburgh, 1948,
163. Yaoi, H.'and Tamiya, H.. On the respiratory pigment, cytochrome, in bacteria, Proc, Imp. Acad, Tokyo. 4: 436. 1928, As cited in Hewitt, L« F, Oxidation-reduction potentials in bacteriology and biochemistry. 5th edition* London -County .Opuacil* . 1948,
164. Yudkin, J. Enzyme variation in mioroorganisms, Biol. Revs, Cambridge Phil, Soc, 13: 93-106. 1938.




















