
The melanin-concentrating hormone gene in human: flanking region analysis, fine chromosome mapping,...
13
Ž . Molecular Brain Research 46 1997 243–255 Research report The melanin-concentrating hormone gene in human: flanking region analysis, fine chromosome mapping, and tissue-specific expression Agnes Viale a , Yao Zhixing a,1 , Christophe Breton a,2 , Florence Pedeutour b , ` Antoine Coquerel c , Daniel Jordan d , Jean-Louis Nahon a, ) a Institut de Pharmacologie Moleculaire et Cellulaire, CNRS UPR 411, 660 route des Lucioles, Sophia-Antipolis, 06560 Valbonne, France ´ b LGMCH, CNRS URA 1462, Faculte de Medecine, aÕenue de Valombrose, 06107 Nice Cedex 2, France ´ ´ ´ c Laboratoire de Radioanalyse, CHU de Rouen, 76031 Rouen Cedex, France ´ d Anatomie Pathologique, Faculte de Medecine Alexis-Carrel, rue Guillaume Paradin, 69372 Lyon Cedex 08, France ´ ´ ´ Accepted 29 October 1996 Abstract Ž . Genomic sequences encoding the human melanin-concentrating hormone MCH were isolated from a YAC library and subcloned in pUC vector using a novel E. coli transformation method. A 4.1-kb fragment encompassing f1.0 kb of the 5 X -end-flanking region, the three exons–two introns of the coding region and f1.7 kb of the 3 X -end-flanking region, was sequenced. Comparison with the rat MCH gene indicated strong conservation in the 5 X -flanking region, in particular over the putative TATA box, CAAT box, GRE and AP-1 elements that could potentially regulate MCH gene expression. FISH with a fluorescent MCH genomic probe on human chromosomes and PCR analysis of a YAC panel mapped MCH to chromosome 12q23.1 in a region flanked by D12S1074 and D12S1030 markers. Ž . Expression of the MCH RNA species and pro-MCH-derived peptides MCH and NEI was investigated in human tissues by combining Northern blotting, RT-PCR, in situ hybridization, immunohistochemistry and RIA. In the human brain, MCH mRNA and MCHrNEI peptides were predominantely expressed in the lateral hypothalamus in agreement with the known distribution of MCH expression in rat. In addition, MCH gene products were detected in extra-hypothalamic sites, such as the pallidum, neocortex and cerebellum. In peripheral tissues, MCH mRNA was identified in several organs, including the thymus, brown adipose tissue, duodenum and testis. An additional shorter MCH gene transcript, likely the result of alternate splicing, was revealed in several brain areas and peripheral tissues. While only fully processed MCH and NEI were found in hypothalamus, a different peptide form, bearing MCH and NEI epitopes, was detected in peripheral organs. This represents the first evidence for differential processing of pro-MCH in mammals. q 1997 Elsevier Science B.V. All rights reserved. Keywords: MCH; NEI; Chromosome 12q23; Neuropeptide gene expression; Hypothalamus; Thymus; Brown adipose tissue; Differential processing 1. Introduction Ž . Melanin-concentrating hormone MCH is a cyclic pep- w x tide originally isolated from salmon pituitary gland 25 w x and later from rat and human hypothalami 32,52 . Mam- malian MCH is encoded within a precursor of 165 amino acids which also gives rise to two other peptides, neu- Ž . ropeptide-glutamic acid-isoleucine NEI and neuropep- ) Ž. Corresponding author. Fax: q33 4 9395-7708; E-mail: [email protected] 1 Present address: Department of Biology, Beijing Normal University, Beijing 100875, P.R. China. 2 Present address: Centre CNRS-INSERM de Pharmacologie-Endo- crinologie, 141 rue de la Cardonille, 34094 Montpellier, France. Ž . w x tide-glycine-glutamic acid NGE 33,34 . Of these two peptides, only NEI has so far been isolated and character- w x ized from rat hypothalamic cells 36 and peripheral tissues w x 22,23 . The main sites of synthesis of MCH mRNA and derived peptides in rat brain are the lateral hypothalamus area Ž . Ž . wx LHA and neighboring zona incerta ZI 4 . The same locations for MCH perikarya were found in other mam- w x malian species, including man 1,33 . The restricted local- ization of neurons expressing pro-MCH-derived peptides in the hypothalamus together with a widespread distribu- tion of fibers throughout the brain strongly suggests that the MCH networks represents an interneuronal system possibly involved in many neuronal functions. I.c.v. injec- tions of MCH inhibit or activate food-intake behavior 0169-328Xr97r$17.00 Copyright q 1997 Elsevier Science B.V. All rights reserved. Ž . PII S0006-8993 97 00015-2
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of The melanin-concentrating hormone gene in human: flanking region analysis, fine chromosome mapping,...

Ž .Molecular Brain Research 46 1997 243–255
Research report
The melanin-concentrating hormone gene in human: flanking regionanalysis, fine chromosome mapping, and tissue-specific expression
Agnes Viale a, Yao Zhixing a,1, Christophe Breton a,2, Florence Pedeutour b,`Antoine Coquerel c, Daniel Jordan d, Jean-Louis Nahon a,)
a Institut de Pharmacologie Moleculaire et Cellulaire, CNRS UPR 411, 660 route des Lucioles, Sophia-Antipolis, 06560 Valbonne, France´b LGMCH, CNRS URA 1462, Faculte de Medecine, aÕenue de Valombrose, 06107 Nice Cedex 2, France´ ´ ´
c Laboratoire de Radioanalyse, CHU de Rouen, 76031 Rouen Cedex, France´d Anatomie Pathologique, Faculte de Medecine Alexis-Carrel, rue Guillaume Paradin, 69372 Lyon Cedex 08, France´ ´ ´
Accepted 29 October 1996
Abstract
Ž .Genomic sequences encoding the human melanin-concentrating hormone MCH were isolated from a YAC library and subcloned inpUC vector using a novel E. coli transformation method. A 4.1-kb fragment encompassing f1.0 kb of the 5X-end-flanking region, thethree exons–two introns of the coding region and f1.7 kb of the 3X-end-flanking region, was sequenced. Comparison with the rat MCHgene indicated strong conservation in the 5X-flanking region, in particular over the putative TATA box, CAAT box, GRE and AP-1elements that could potentially regulate MCH gene expression. FISH with a fluorescent MCH genomic probe on human chromosomesand PCR analysis of a YAC panel mapped MCH to chromosome 12q23.1 in a region flanked by D12S1074 and D12S1030 markers.
Ž .Expression of the MCH RNA species and pro-MCH-derived peptides MCH and NEI was investigated in human tissues by combiningNorthern blotting, RT-PCR, in situ hybridization, immunohistochemistry and RIA. In the human brain, MCH mRNA and MCHrNEIpeptides were predominantely expressed in the lateral hypothalamus in agreement with the known distribution of MCH expression in rat.In addition, MCH gene products were detected in extra-hypothalamic sites, such as the pallidum, neocortex and cerebellum. In peripheraltissues, MCH mRNA was identified in several organs, including the thymus, brown adipose tissue, duodenum and testis. An additionalshorter MCH gene transcript, likely the result of alternate splicing, was revealed in several brain areas and peripheral tissues. While onlyfully processed MCH and NEI were found in hypothalamus, a different peptide form, bearing MCH and NEI epitopes, was detected inperipheral organs. This represents the first evidence for differential processing of pro-MCH in mammals. q 1997 Elsevier Science B.V.All rights reserved.
Keywords: MCH; NEI; Chromosome 12q23; Neuropeptide gene expression; Hypothalamus; Thymus; Brown adipose tissue; Differential processing
1. Introduction
Ž .Melanin-concentrating hormone MCH is a cyclic pep-w xtide originally isolated from salmon pituitary gland 25
w xand later from rat and human hypothalami 32,52 . Mam-malian MCH is encoded within a precursor of 165 aminoacids which also gives rise to two other peptides, neu-
Ž .ropeptide-glutamic acid-isoleucine NEI and neuropep-
) Ž .Corresponding author. Fax: q33 4 9395-7708; E-mail:[email protected]
1 Present address: Department of Biology, Beijing Normal University,Beijing 100875, P.R. China.
2 Present address: Centre CNRS-INSERM de Pharmacologie-Endo-crinologie, 141 rue de la Cardonille, 34094 Montpellier, France.
Ž . w xtide-glycine-glutamic acid NGE 33,34 . Of these twopeptides, only NEI has so far been isolated and character-
w xized from rat hypothalamic cells 36 and peripheral tissuesw x22,23 .
The main sites of synthesis of MCH mRNA and derivedpeptides in rat brain are the lateral hypothalamus areaŽ . Ž . w xLHA and neighboring zona incerta ZI 4 . The samelocations for MCH perikarya were found in other mam-
w xmalian species, including man 1,33 . The restricted local-ization of neurons expressing pro-MCH-derived peptidesin the hypothalamus together with a widespread distribu-tion of fibers throughout the brain strongly suggests thatthe MCH networks represents an interneuronal systempossibly involved in many neuronal functions. I.c.v. injec-tions of MCH inhibit or activate food-intake behavior
0169-328Xr97r$17.00 Copyright q 1997 Elsevier Science B.V. All rights reserved.Ž .PII S0006-8993 97 00015-2

( )A. Viale et al.rMolecular Brain Research 46 1997 243–255244
depending on the concentration of peptide used andror ratw xstrains studied 41,42 . In addition, MCH and NEI act as
functional antagonists in the activation of the hypothala-w xmus-pituitary-adrenal gland axis after an acute stress 5 .
Both MCH and NEI induce secretion of oxytocin in thew xneurohypophysis 36 . Centrally, MCH was shown to an-
w xtagonize a-MSH on grooming behavior 14 , on sensori-w x w xmotor integration 30 , on learning acquisition 29 or on
w xaggression and anxiety 17 . All of these data are consis-tent with the down-regulation of MCH gene activity ob-
w x w xserved after chronic stress 38 or osmotic stress 39 .Peripherally, MCH and associated peptides predominate inthe gastrointestinal and genital tracts where they appear to
w xact as paracrine or autocrine factors 21–23 .The exon–intron structure of the MCH genes was estab-
lished after PCR amplification of genomic DNA from thew xrat, mouse and human 7 . The rat MCH gene has been
also cloned following screening of a genomic library butonly 380 nucleotides upstream of the transcriptional initia-tion site were published and no regulatory element was
w xmapped 50 . Strikingly, a single MCH gene was identifiedin rodents while two distinct gene systems were discoveredin human, one corresponding to the authentic gene and
w xhighly homologous to its rat and mouse counterparts 7and the other defined as the variant gene on the basis of
w xpartial sequence homology with the authentic gene 8 . TheŽ .human authentic gene PMCH locus was mapped to
12q23–q24 and two copies of the variant gene, designedPMCHL1 and PMCHL2, were located on chromosome
w x5p14 and 5q12–q13, respectively 37 . The authentic MCHgene might be involved in human neural disorders, such as
w xDarier’s disease 12 or the spinocerebellar ataxia type 2Ž . w xlocus SCA 2 16 .
In order to initiate genetic linkage studies and to furtherinvestigate the role and regulation of MCH and associatedpeptides in human complex behaviors, such as food-intakebehavior, it is necessary to isolate the 5X- and 3X-flankingregions of the MCH gene and to identify the products ofMCH gene transcription and MCH mRNA translation. Inthis article, we have established the genomic organizationof the human authentic MCH gene and identified severalregulatory elements which might be involved in its regula-tion. In addition, MCH RNA species were identified inunexpected sites of the human brain and in several periph-eral organs as well. Finally, a particular processing ofpro-MCH was found in peripheral tissues of humanneonates and adults.
2. Materials and methods
2.1. YAC library screening
Ž .YAC libraries from the CEPH Paris, France werescreened by PCR using 30 pmol of two MCH-specific
Ž Xprimers: HMCH2 5 CCA GCA AGA TTG GTA GAC
X. Ž XTCT TCC CAG CAT 3 and HMCH6 5 TTC TTA AATX.CAT GGT CTG CCA CTG AAT CTG 3 in a final
volume of 30 ml containing 0.2 U of Taq polymeraseŽ .Appligene and 0.8 mM dNTPs. Amplification conditionswere as follows: denaturation at 948C, annealing at 528Cand extention at 728C for 1 min each during 30 cycles. ThePCR products were separated on a 0.8% agarose gel,stained in presence of ethidium bromide, visualised underUV, transferred to nylon membrane and hybridized with a
Ž .human MCH cDNA probe see below as described else-w xwhere 21 . First screening yielded 8 positive pools. Each
pool was then screened following the same strategy andtwo YAC clones were finally analyzed.
2.2. Subcloning and sequencing of a MCH gene fragment
Genomic YAC DNA was purified according to standardw xmethods 46 and fragments were subcloned into the PUC
19 plasmid vector following a novel transformation proto-col. Briefly, YAC DNA were cut either with XhoI andBamHI or XbaI to yield fragments of 8.9 and 3.36 kb,respectively. These fragments were ligated into pUC19
Ž .vector Boehringer-Mannheim, Germany after which 2 mlof 1.0 mgrml ethidium bromide were added to 6 ml of theligation mixture and the solution incubated at 808C for 5min and chilled at 08C. The 8 ml reaction mixture wasused to transform 200 ml of competent XLI Blue cellsŽ .Stratagene, USA according to conventional calcium chlo-
w xride method or by electroporation 46 . All details concern-ing the ethidium bromide incubation conditions have been
Ž .defined elsewhere Zhixing and Nahon, submitted . E. colicells were plated on LB agar-containing ampicillin, IPTG
w xand Xgal 46 and the white colonies were subsequentlyused for detailed analysis.
The sequence of MCH recombinant clones was deter-mined from double-stranded DNA using Taq dye deoxy
Ž .terminator cycle sequencing kit Applied Biosystems onan Applied biosystems model 373A sequencer. Computeranalysis of nucleotide and deduced protein sequences uti-lized the DNA strider program and programs from Bisance
w x Ž .service 13 SIGNUC, FASTA and BLAST .
2.3. Chromosomal mapping
Ž .Fluorescence in situ hybridization FISH was per-formed on human metaphase chromosomes. Biotinylated
Ž Ž . .probe phMCH I ; Fig. 1A was co-precipitated with Cot.Ž .1 DNA Gibco-BRL and hybridized on slides and the
w xstaining revealed as described elsewhere 37 .
2.4. RNA preparation and Northern blot analysis
Total RNA was extracted from different human tissuesŽ .neonates or adults by the acid guanidinium-thiocyanate-
w xphenol-chloroform method 11 .

( )A. Viale et al.rMolecular Brain Research 46 1997 243–255 245
Ž .qFor Northern blot analyses, f5 mg of total or poly ARNA were denaturated in 2.2 M formaldehyder50% for-mamide solution and fractioned electrophoretically on a1.2% agaroserformaldehyde gel in morpholino-propan-
Ž .esulfonic MOPS buffer. RNAs were transferred to nylonmembrane, hybridized with specific human MCH or b-actin cDNA probes and blots were exposed to X-ray films.Full experimental details have been described elsewherew x38 . The relative abundance of MCH mRNA on theautoradiographs was determined by scanning densitometryusing an Hagfa arcus scanner and quantified using a
Žcomputerized image analysis system NIH image, version.1.51 .
2.5. RT-PCR analysis
Ž .First-strand synthesis of cDNA from poly A q or totalRNA was carried out using moloney murine leukaemia
Ž .virus reverse transcriptase Gibco-BRL, France . The sin-gle-stranded cDNA was subsequently amplified by PCRunder the same conditions as described for YAC screening
Ž .using HMCH2, HMCH6 see above , HPCR3 and HPCR4w xprimers 7 . Detailed protocols of the reverse transcription
w xand amplification methods were submitted previously 21 .The PCR products were separated onto a 0.8 or 1.5%agarose gel, visualized under UV after staining with ethid-ium bromide and Southern blotted with a 32 P-labeled
w xhMCH cDNA probe 40 .
2.6. In situ hybridization
Four post-mortem human hypothalami were used for insitu hybridization. The results presented in Fig. 6A arefrom the brain of a 52-year-old man deceased by accident;post-mortem delay was f20 h.
Ž .Frozen sections 12 mm-thick were cut on a cryostat aty208C and thaw-mounted on silane-coated slides. Tissuessections were fixed in 4% paraformaldehyder1=PBSŽ0.14 M NaCl, 4 mM Na HPO , 1.5 mM KH PO , pH2 4 2 4
.7.4 for 10 min at room temperature, washed in 4=SSCfor 5 min 2= and pre-hybridized for at least 1 h in a4=SSCr1=Denhardt’s solution. Adequate volume ofhybridization mixture containing 33 P-labeled RMCH2oligoprobe was spotted on each slide, sealed under cover-slip, placed in a humidified chamber and incubated
Ž .overnight at 258C below the temperature of fusion Tmw x21 . The slides were rinsed 3= for 20 min in 1 SSC at428C under agitation and deshydrated in ethanolr0.3 Mammonium acetate. Finally, sections were exposed to
Ž .Biomax MR film Kodak for 2 days.
2.7. Immunohistochemistry
After a post-fixation step of 10 min in PFA 4%r1=
Ž .KPBS same as 1=PBSq2.7 mM KCl , tissue sectionswere washed 2= in KPBS solution and incubated for 48 h
Žat 48C with primary antiserum MCH and NEI antisera
Ž . Ž .Fig. 1. Structure of the human MCH gene. A: partial restriction site map of the phMCH I and phMCH II clones are shown. The exons are boxed. B:exon–intron organization of the MCH gene and locations of primers used for RT-PCR. The expected lengths of the PCR products are indicated. Thesecorresponding to amplification of mature MCH mRNA are noted in brackets.
( )A. Viale et al.rMolecular Brain Research 46 1997 243–255246

( )A. Viale et al.rMolecular Brain Research 46 1997 243–255 247
were kindly provided by Dr. J. Vaughan, The Salk Insti-.tute, La Jolla, USA at a dilution of 1 : 1000 in KPBS
buffer containing 0.3% Triton-X-100r0.02% normal goatŽserum. Sections were then washed in KPBS solution 2=
.for 10 min and incubated for 1 h with fluoresceinylatedŽgoat anti rabbit IgG 1 : 100 dilution; HqL Affinity puri-
.fied, Vector flow . After washing, the sections weremounted in glycerolrmoviol and photographed under fluo-rescence illumination.
2.8. Peptide extraction
After homogenization by polytron, tissues were boiledfor 10 min in 0.1 M HCl and centrifuged 2= at 10 000=gat room temperature for 20 min. The supernatants wereneutralized with NAOH and the peptide solutionslyophilized. The dried extracts were re-suspended in 10mM phosphate buffer and used for either a direct RIA oran HPLC purification.
2.9. HPLC condition
Ž .Reverse-phase HPLC RP-HPLC was performed with aŽ .C158 Merck Lithosorb column 100-RP 5 mm . The mo-
bile phase was 0.1% TFA solution. The peptides wereeluted with an acetonitrile-TFA 0.1% gradient from 20 to60% in 40 min with a 1 mlrmin flow rate after anisocratic step at 20% for 10 min. The retention time of thesynthetic ratrhuman MCH and NEI were 32 and 15 min,respectively. Synthetic MCH and NEI were gifts from Drs.
ŽCarl M. Hoeger and Jean Rivier Salk Institute, La Jolla,. w xCA . Synthetic NEI-GRR 45 were custom synthesized by
Ž .Neosystem Belgium .
2.10. Radioimmunoassay
w xRIA were performed as described 23 . Briefly, 50 mlŽ .of rat MCH or NEI antisera 1 : 60 000 , 50 ml of the
standard peptide or biological samples, 50 ml of theŽ 5 .iodinated peptide 10 cpmrml and 100 ml of phosphate
buffer were mixed. After 24 or 48 h of incubation at 48C,bound radioactivity was precipitated by adding 50 ml ofbovine serum, 50 ml of 0.6% Tween-20 and 350 ml ofpolyethyleneglycol 6000 25%. Tubes were then cen-trifuged for 10 min at 4000 rpm at 48C and the supernatantwas discarded. The pellets were counted in a Crystal
Ž .Packard for automatic calculation of the boundrtotal BrTŽratio. Blank values i.e. radioactivity precipitated without
.antibodies were always -6%. The routine sensitivity ofthe assays was 250 and 150 fmolrsample for the MCH
and NEI RIA, respectively. The NEI antiserum recognizedequally well the amidated NEI peptide and NEI-GR but
w xNEI-GRR cross-reacted -0.1% with this antiserum 45 .
2.11. Citraconic anhydride actionr trypsinrunblocking( )CTU method
w xThe CTU procedure described by Bidard et al. 3 wasused to characterize the 44-min HPLC peak containingMCH immunoreactivity. Dried samples of crude peptidicextracts or after RP-HPLC purification were dissolved in
Ž .0.05% wrv Triton-X-100, HepesrNaOH buffer and cit-Ž .raconic anhydre 5 mM final concentration , tryptic reac-
w xtion was carried out, then unblocking was performed 3 .The samples were finally analyzed on RP-HPLC as de-scribed above.
3. Results
3.1. Cloning of the coding and flanking parts of the humanMCH gene
Isolation of the MCH gene was achieved by screening aYAC library. Two YAC clones, 908f7 and 886a6, wereclearly positive after DNA amplification with MCH gene-
Ž .specific primers, viz. HPCR3 and HPCR4 Fig. 1B . ABamHIrXhoI fragment of f8.9 kb in length and a XbaIfragment of f3.3 kb in length were then subcloned inpUC19 vector by using a modified procedure, including
Žethidium bromide treatment before transformation Zhix-.ing and Nahon, submitted . Several bacterial colonies were
selected and maxi-preparation of DNA was performedŽ .with two colonies representative of the phMCH I clone
Ž .bearing the 8.9-kb XhoIrBamHI fragment or theŽ . Ž .phMCH II clone bearing the 3.3-kb XbaI fragment .
Digestion of these cloned DNA and whole cell DNA fromhuman brain with several restriction enzymes and probingwith MCH cDNA demonstrated overlapping patterns of
Ž .hybridization Fig. 1A, and not shown .
3.2. Nucleotide sequence analysis of the cloned humanMCH gene
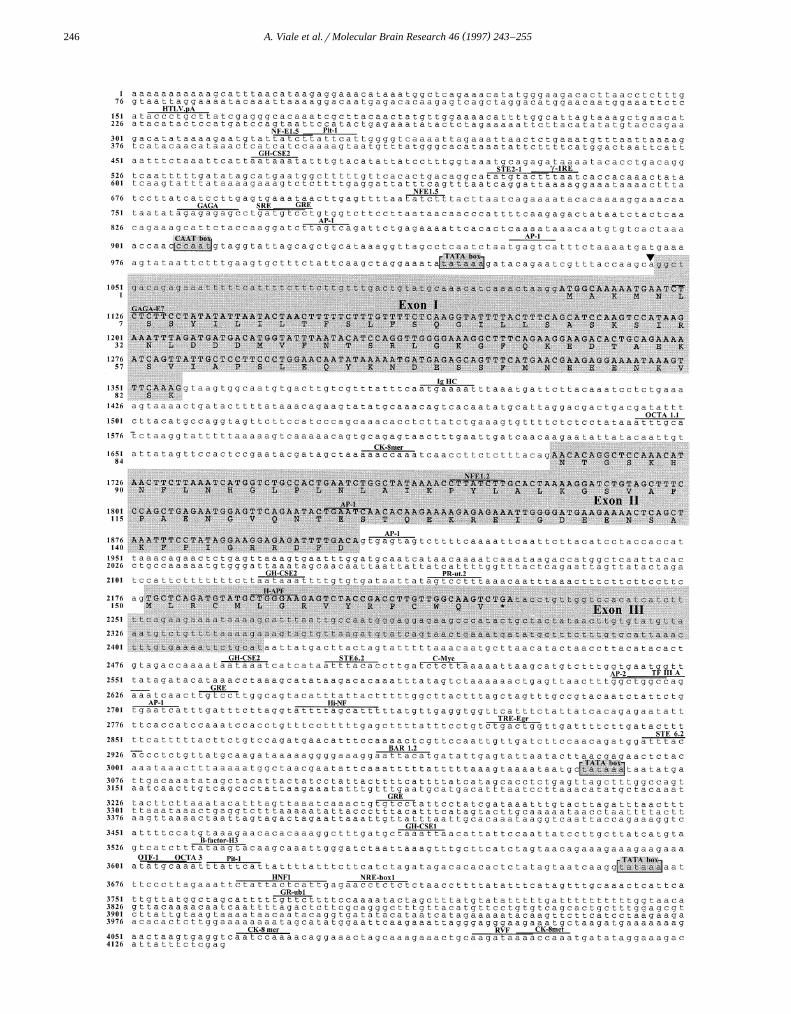
The sequence of the human MCH gene is shown in Fig.Ž .2. The human MCH gene contains 3 exons I–III of 279,
199 and 240 bp, respectively, as previously found bycloning and sequencing a PCR fragment named phMCH-
w xG34 8 . Intron–exon boundaries were determined by com-Ž .parison of phMCH I clone and phMCH cDNA sequences
Fig. 2. Partial DNA sequence of human MCH gene and deduced amino-acid sequence. The genomic structure and deduced protein structure wasŽ . Ž .determined by sequencing phMCH I and phMCH II clones. DNA sequence begins at nucleotide 1 of the sequenced fragment and amino-acid sequence
begins at the initiation codon ATG. Exons are boxed. The translated regions of MCH gene are noted in capital letters. Upperlined and named are theconsensus sequences for transcriptional regulatory elements identified by computer analysis. Transcription start site is indicated with an arrowhead.

( )A. Viale et al.rMolecular Brain Research 46 1997 243–255248
Fig. 3. Comparison of the promoter region of MCH gene in rat and human. Identical nucleic acids are shaded. TATA box and CAAT box are boxed withsolid and broken lines, respectively. Upperlined are the consensus sequence for controlling elements conserved between rat and human MCH gene. Base
w xnumbering is the same as in Fig. 2 for the human MCH gene and as reported previously for the rat MCH gene 50 .
and were found to be identical to those identified in thephMCH-G34 clone.
The transcriptional initiation site was tentatively local-ized to 30 bases 3X of the first thymidine of a consensusTATA box by comparison with that found in the rat MCH
w x X Xgene 38,50 . Analysis of the 5 - and 3 -end-flanking se-quences as well as of the central part of the human MCHgene revealed the presence of consensus sequences corre-sponding to potential regulatory factor-binding sites. Thepossible functional relevance of these regulatory elementswill be discussed below. Comparison of the 5X-flankingregion of the human MCH with the available sequencecounterpart of the rat MCH gene revealed strong sequenceidentities within some of these putative transcriptional
w xregulatory elements, such as a partial GRE 24 , a GAGAw x w x w xbox 18 , a NF-E1-binding site 53 and two AP-1 sites 27
Ž .Figs. 2 and 3 .
3.3. Chromosomal mapping
Using a FISH technique on human metaphase chromo-somes, the MCH gene was mapped with the genomic
Ž . Ž .MCH probe phMCH I on chromosome 12q23 Fig. 4A .No signal was identified outside the chromosome 12 label-ing, even on chromosome 5p and 5q where the variant
w xMCH gene loci were mapped 37 . The two MCH-positiveYACs described above, overlapped on the physical map of
w xhuman chromosome 12 established by Krauter et al. 26 .Hence, PMCH was located to the region, including theIGF1 gene, and flanked by D12S1074 on the centromic
Ž .side and D12S1030 on the telomeric side Fig. 4B .
3.4. MCH gene expression and cellular localization inhuman brain
RNA samples from rat, mouse and human hypothalamiwere examined first by Northern blot to determine the size
and relative abundance of MCH gene transcripts. MajorRNA species of 1.0, 1.1 and 0.95 kb were identified in rat,
Ž .mouse and human, respectively Fig. 5A , consistent withw xthe length of MCH cDNA in human 40 and the size of
w xthe rodent MCH mRNA reported previously 7,38 . Quan-
Fig. 4. Chromosomal mapping of PMCH in human. A: in situ hybridiza-Ž .tion of the phMCH I clone to human chromosomes. The bright spots are
located on sister chromatids of chromosomes identified as 12 by DAPI-Ž .staining analysis arrowheads . Note the absence of fluorescence over
other chromosomes. B: partial physical map of human chromosome 12.Two overlapping YACs bearing MCH gene sequences, named 908f7 and889a6, are noted with double arrows. The location of PMCH locus isindicated within the hatched lines. The names and positions of the
w xmarkers are as reported by Krauter et al. 26 .
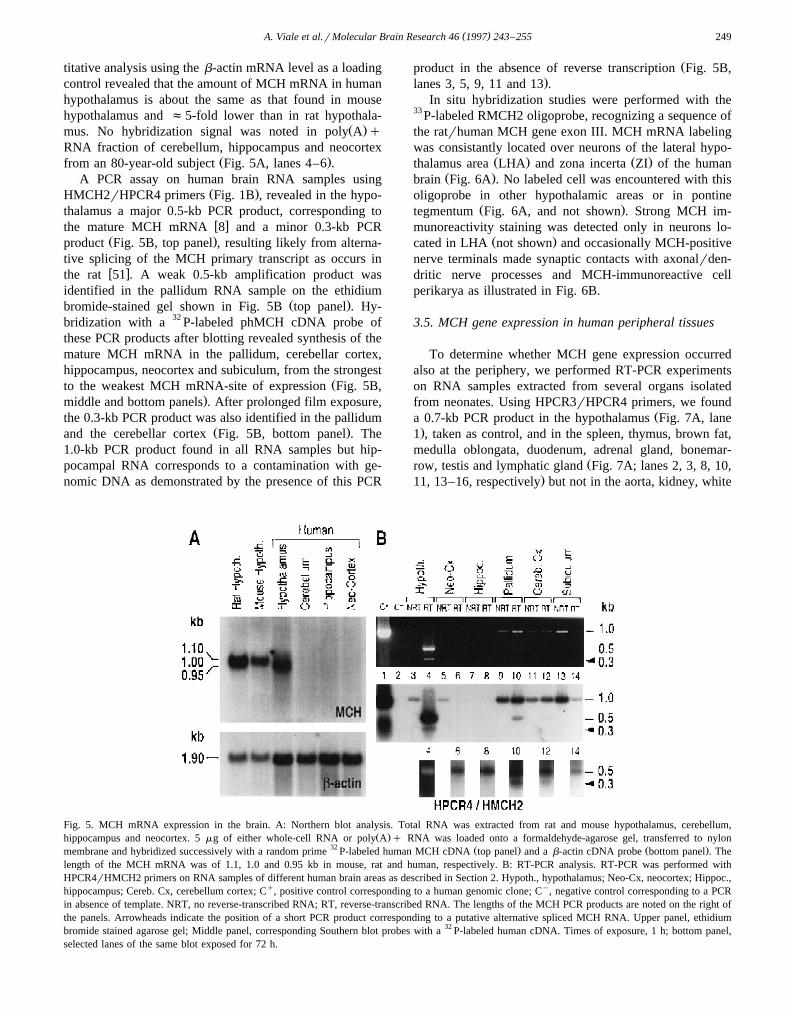
( )A. Viale et al.rMolecular Brain Research 46 1997 243–255 249
titative analysis using the b-actin mRNA level as a loadingcontrol revealed that the amount of MCH mRNA in humanhypothalamus is about the same as that found in mousehypothalamus and f5-fold lower than in rat hypothala-
Ž .mus. No hybridization signal was noted in poly A qRNA fraction of cerebellum, hippocampus and neocortex
Ž .from an 80-year-old subject Fig. 5A, lanes 4–6 .A PCR assay on human brain RNA samples using
Ž .HMCH2rHPCR4 primers Fig. 1B , revealed in the hypo-thalamus a major 0.5-kb PCR product, corresponding to
w xthe mature MCH mRNA 8 and a minor 0.3-kb PCRŽ .product Fig. 5B, top panel , resulting likely from alterna-
tive splicing of the MCH primary transcript as occurs inw xthe rat 51 . A weak 0.5-kb amplification product was
identified in the pallidum RNA sample on the ethidiumŽ .bromide-stained gel shown in Fig. 5B top panel . Hy-
bridization with a 32 P-labeled phMCH cDNA probe ofthese PCR products after blotting revealed synthesis of themature MCH mRNA in the pallidum, cerebellar cortex,hippocampus, neocortex and subiculum, from the strongest
Žto the weakest MCH mRNA-site of expression Fig. 5B,.middle and bottom panels . After prolonged film exposure,
the 0.3-kb PCR product was also identified in the pallidumŽ .and the cerebellar cortex Fig. 5B, bottom panel . The
1.0-kb PCR product found in all RNA samples but hip-pocampal RNA corresponds to a contamination with ge-nomic DNA as demonstrated by the presence of this PCR
Žproduct in the absence of reverse transcription Fig. 5B,.lanes 3, 5, 9, 11 and 13 .
In situ hybridization studies were performed with the33 P-labeled RMCH2 oligoprobe, recognizing a sequence ofthe ratrhuman MCH gene exon III. MCH mRNA labelingwas consistantly located over neurons of the lateral hypo-
Ž . Ž .thalamus area LHA and zona incerta ZI of the humanŽ .brain Fig. 6A . No labeled cell was encountered with this
oligoprobe in other hypothalamic areas or in pontineŽ .tegmentum Fig. 6A, and not shown . Strong MCH im-
munoreactivity staining was detected only in neurons lo-Ž .cated in LHA not shown and occasionally MCH-positive
nerve terminals made synaptic contacts with axonalrden-dritic nerve processes and MCH-immunoreactive cellperikarya as illustrated in Fig. 6B.
3.5. MCH gene expression in human peripheral tissues
To determine whether MCH gene expression occurredalso at the periphery, we performed RT-PCR experimentson RNA samples extracted from several organs isolatedfrom neonates. Using HPCR3rHPCR4 primers, we found
Ža 0.7-kb PCR product in the hypothalamus Fig. 7A, lane.1 , taken as control, and in the spleen, thymus, brown fat,
medulla oblongata, duodenum, adrenal gland, bonemar-Žrow, testis and lymphatic gland Fig. 7A; lanes 2, 3, 8, 10,
.11, 13–16, respectively but not in the aorta, kidney, white
Fig. 5. MCH mRNA expression in the brain. A: Northern blot analysis. Total RNA was extracted from rat and mouse hypothalamus, cerebellum,Ž .hippocampus and neocortex. 5 mg of either whole-cell RNA or poly A q RNA was loaded onto a formaldehyde-agarose gel, transferred to nylon
32 Ž . Ž .membrane and hybridized successively with a random prime P-labeled human MCH cDNA top panel and a b-actin cDNA probe bottom panel . Thelength of the MCH mRNA was of 1.1, 1.0 and 0.95 kb in mouse, rat and human, respectively. B: RT-PCR analysis. RT-PCR was performed withHPCR4rHMCH2 primers on RNA samples of different human brain areas as described in Section 2. Hypoth., hypothalamus; Neo-Cx, neocortex; Hippoc.,hippocampus; Cereb. Cx, cerebellum cortex; Cq, positive control corresponding to a human genomic clone; Cy, negative control corresponding to a PCRin absence of template. NRT, no reverse-transcribed RNA; RT, reverse-transcribed RNA. The lengths of the MCH PCR products are noted on the right ofthe panels. Arrowheads indicate the position of a short PCR product corresponding to a putative alternative spliced MCH RNA. Upper panel, ethidiumbromide stained agarose gel; Middle panel, corresponding Southern blot probes with a 32 P-labeled human cDNA. Times of exposure, 1 h; bottom panel,selected lanes of the same blot exposed for 72 h.

( )A. Viale et al.rMolecular Brain Research 46 1997 243–255250
Fig. 6. Distribution of MCH-expressing cells in human hypothalamus. A:in situ hybridization with 33 P-labeled RMCH2 oligoprobe of two repre-sentative horizontal sections of a 52-year-old human hemi-hypothalamus.LHA: lateral hypothalamus area; O.C., Optic chiasma; 3v, third ventricle.B: an isolated MCH-expressing neuron identified with a ratrhuman MCHantiserum. Note the presence of immunoreactive MCH nerve terminalssurrounding the perikarion and dendrites of the MCH-expressing cell.Magnification: =1 in A; =250 in B.
Žfat, bladder, stomach and pyllorus Fig. 6A, lanes 4–7, 9.and 12, respectively . The tissue-specific pattern of MCH
mRNA expression was identical when using theHMCH6rHMCH2 primers which amplified a 0.2-kbMCH-specific fragment and a 0.5-kb fragment correspond-
Ž .ing to an intron B-containing PCR product Fig. 7B .Contamination with genomic DNA was found in someRNA samples.
Marked variations in the levels of MCH mRNA werenoted among the tissues.When compared to the hypothala-
Ž . Ž .mus Fig. 7, lane 1 , the thymus lane 3 and brownŽ .adipose tissues lane 8 expressed f2- and 8-fold lower
MCH mRNA levels, respectively. All other positive tissueexhibited f20-fold lower levels. These differences inMCH mRNA content may reflect true physiological varia-tions but differential degradation of RNA species in humantissues andror difficulties in amplification efficiencies maynot be excluded.
Ž .Using HPCR3rHPCR4 primers Fig. 1B , a 0.5-kbPCR product was found in the thymus and testis but not in
Ž .other peripheral organs arrowhead in Fig. 7A . This corre-sponds likely to the 0.3-kb RNA product already identifiedin the hypothalamus of adult human using
Ž .HMCH2rHPCR4 primers arrowhead in Fig. 5B . Thisshort PCR product was lacking in the hypothalamus, thy-mus and testis when HMCH6 and HMCH2 primers were
Ž .used Fig. 7B , suggesting that part or the whole of exon IIwas missing in the corresponding RNA.
3.6. Differential processing of the MCH precursor in thebrain Õs. peripheral tissues
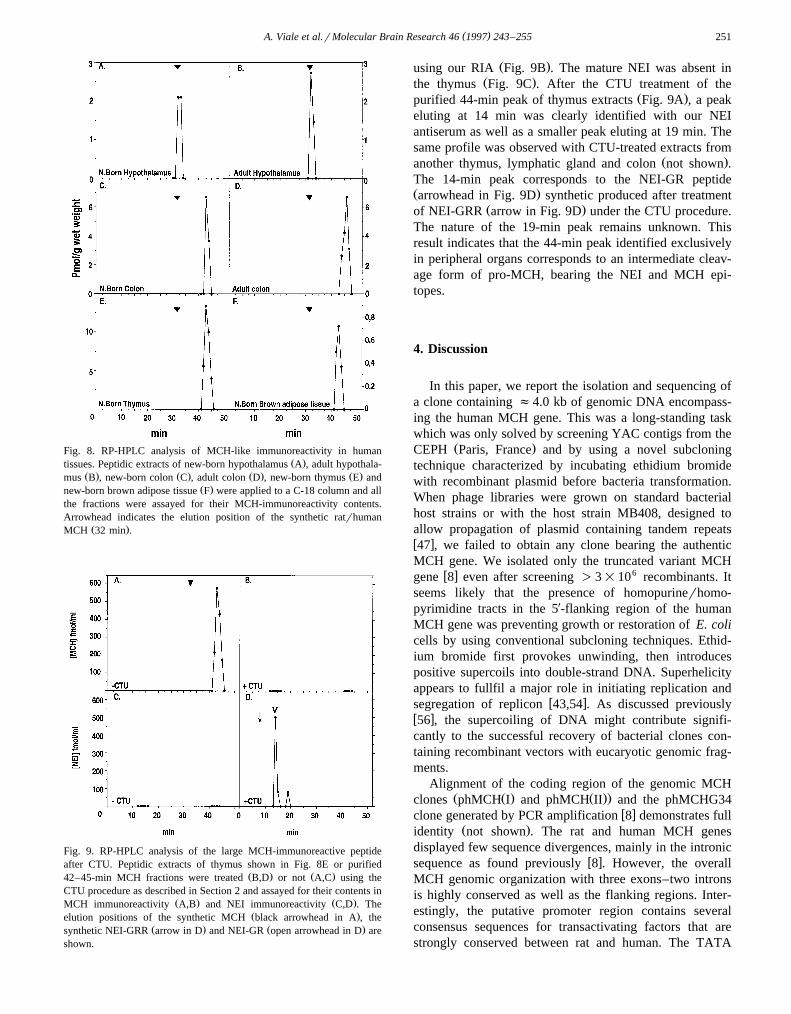
Hypothalamic peptide extracts from neonate or adulthuman were chromatographed on RP-HPLC and MCHcontent was determined with a RIA. As shown in Fig.8A,B, the totality of the MCH immunoreactivity co-elutedwith the synthetic peptide with a peak at 32 min in bothneonatal and adult hypothalami. In contrast, for all periph-
Ž .eral organs tested from neonates Fig. 8C,E,F or adultsŽ .Fig. 8D , the elution profile from RP-HPLC showed asingle peak at 44 min and no MCH immunoreactivity wasfound co-eluting with synthetic MCH. The NEI antiserafailed to recognize this as well as other peaks in peripheraltissue extracts while a peak at 15 min, characteristic of theNEI-amidated peptide, was detected in the hypothalamic
Ž .extracts not shown .To test the possibility that the MCH-immunoreactive
material found in peripheral tissues could be a processingintermediate joining NEI and MCH, we tried to identify apeptide recognized by the NEI antiserum in the materialcorresponding to the 44-min peak by using the citraconyla-
Ž . w xtionrtrypsinrunblocking processing CTU 3 . The aminefunctions of the lysine side chains of the putative productwere protected by treatment with citraconic anhydride,then trypsin was added to cleave the citraconylated pep-tides at the C-terminal sides of arginine residues. Underthese conditions, a putative NEI-MCH intermediate wouldgenerate a NEI-GR form and several short fragmentscorresponding to MCH digestion products undetectable
Fig. 7. Expression of MCH RNA species in peripheral organs. RT-PCRŽ .experiments were performed with HPCR3rHPCR4 primers A or
Ž .HMCH6rHMCH2 primers B as described in Section 2. RepresentativeSouthern blots of RT-PCR products probed with 32 P-labeled humanMCH cDNA probe are shown. The PCR products are indicated as in Fig.5B. Hypoth., hypothalamus; Medulla. ob., medulla oblongata; AdrenalGl., adrenal gland; Lymphatic Gl., lymphatic gland.
( )A. Viale et al.rMolecular Brain Research 46 1997 243–255 251
Fig. 8. RP-HPLC analysis of MCH-like immunoreactivity in humanŽ .tissues. Peptidic extracts of new-born hypothalamus A , adult hypothala-
Ž . Ž . Ž . Ž .mus B , new-born colon C , adult colon D , new-born thymus E andŽ .new-born brown adipose tissue F were applied to a C-18 column and all
the fractions were assayed for their MCH-immunoreactivity contents.Arrowhead indicates the elution position of the synthetic ratrhuman
Ž .MCH 32 min .
Fig. 9. RP-HPLC analysis of the large MCH-immunoreactive peptideafter CTU. Peptidic extracts of thymus shown in Fig. 8E or purified
Ž . Ž .42–45-min MCH fractions were treated B,D or not A,C using theCTU procedure as described in Section 2 and assayed for their contents in
Ž . Ž .MCH immunoreactivity A,B and NEI immunoreactivity C,D . TheŽ .elution positions of the synthetic MCH black arrowhead in A , the
Ž . Ž .synthetic NEI-GRR arrow in D and NEI-GR open arrowhead in D areshown.
Ž .using our RIA Fig. 9B . The mature NEI was absent inŽ .the thymus Fig. 9C . After the CTU treatment of the
Ž .purified 44-min peak of thymus extracts Fig. 9A , a peakeluting at 14 min was clearly identified with our NEIantiserum as well as a smaller peak eluting at 19 min. Thesame profile was observed with CTU-treated extracts from
Ž .another thymus, lymphatic gland and colon not shown .The 14-min peak corresponds to the NEI-GR peptideŽ .arrowhead in Fig. 9D synthetic produced after treatment
Ž .of NEI-GRR arrow in Fig. 9D under the CTU procedure.The nature of the 19-min peak remains unknown. Thisresult indicates that the 44-min peak identified exclusivelyin peripheral organs corresponds to an intermediate cleav-age form of pro-MCH, bearing the NEI and MCH epi-topes.
4. Discussion
In this paper, we report the isolation and sequencing ofa clone containing f4.0 kb of genomic DNA encompass-ing the human MCH gene. This was a long-standing taskwhich was only solved by screening YAC contigs from the
Ž .CEPH Paris, France and by using a novel subcloningtechnique characterized by incubating ethidium bromidewith recombinant plasmid before bacteria transformation.When phage libraries were grown on standard bacterialhost strains or with the host strain MB408, designed toallow propagation of plasmid containing tandem repeatsw x47 , we failed to obtain any clone bearing the authenticMCH gene. We isolated only the truncated variant MCH
w x 6gene 8 even after screening )3=10 recombinants. Itseems likely that the presence of homopurinerhomo-pyrimidine tracts in the 5X-flanking region of the humanMCH gene was preventing growth or restoration of E. colicells by using conventional subcloning techniques. Ethid-ium bromide first provokes unwinding, then introducespositive supercoils into double-strand DNA. Superhelicityappears to fullfil a major role in initiating replication and
w xsegregation of replicon 43,54 . As discussed previouslyw x56 , the supercoiling of DNA might contribute signifi-cantly to the successful recovery of bacterial clones con-taining recombinant vectors with eucaryotic genomic frag-ments.
Alignment of the coding region of the genomic MCHŽ Ž . Ž ..clones phMCH I and phMCH II and the phMCHG34
w xclone generated by PCR amplification 8 demonstrates fullŽ .identity not shown . The rat and human MCH genes
displayed few sequence divergences, mainly in the intronicw xsequence as found previously 8 . However, the overall
MCH genomic organization with three exons–two intronsis highly conserved as well as the flanking regions. Inter-estingly, the putative promoter region contains severalconsensus sequences for transactivating factors that arestrongly conserved between rat and human. The TATA

( )A. Viale et al.rMolecular Brain Research 46 1997 243–255252
box which is required for adequate initiation of transcrip-tion by RNA polymerase II is located 30 bp upstream of
Ž .the cap site boxed in Fig. 1B . A putative CAAT box asw xdefined by Thompson and Watson 50 is found at the
same location upstream from the TATA box of the humanand rat MCH gene. Besides these two sequences, otherregulatory elements that we identified in the promoterregion of the human MCH were overlooked in the analysis
w xof the rat MCH gene 50 . Consensus AP-1-binding sitesŽ .TGANTCA are located y94ry89 upstream from the
Žcap site of the humanrrat MCH gene and in exon II Fig..2BFig. 3 . They may be involved in regulation of MCH
gene expression by Fosrjun regulatory factor family asw xfound for many neuronal systems 49 . Derived AP-1-bind-
ing sites are also present in the promoter regionŽ . Ž .TTAGTCA or in intron B TGAGTGA of the human
Ž .MCH gene Fig. 2B and could possibly be recognized byŽ .Fos complexes. Similarly, an interferon g IFN-g re-
Ž . w xsponse element IRE 55 and partial sequences of aŽ . w xglucocorticoid response element GRE 24 are mapped
within the 5X-flanking region of the ratrhuman MCH gene.The IRE can play a role in the transcriptional regulation of
w xthe MCH gene in macrophages 23 and the GRE can beresponsible for the induction of the MCH gene by gluco-
w xcorticoids in vivo 38 . Other cis-regulatory elements havebeen mapped in the 5X-end-flanking, central coding and3X-end flanking regions of the human MCH gene but theovert functions of these sequences need to be investigatednow by using reporter-gene assay, gel-shift and DNA-footprinting analyses.
Previous chromosomal mapping using FISH has indi-cated that PMCH was located on human chromosome
w x12q23–24 37 . We confirmed this result by using a largegenomic fragment bearing flanking regions of the authen-tic MCH gene and we demonstrated the absence of largeportions of this gene on other chromosomes. This dataindicated that the region of homology between the authen-tic and the variant gene is actually restricted to part of the
w xcoding sequence 8 . Based upon screening of YAC con-tigs by PCR we have placed the authentic human MCHgene to a 5-cM genetic interval bounded by D12S1074 andD12S1030. This region contains the insulin-like growth
Ž .factor 1 IGF 1 locus as well as two additional polymor-phic markers that might be useful in more precisely locat-
w xing the MCH gene 26 . The MCH gene was a possiblecandidate gene for Darier’s disease and spinocerebellar
w xataxia type 2 37 . However, these loci have been assignedto regions flanked, respectively, by D12S105 and D12S129
w xfor the Darier’s disease 12 and D12S84 and D12S79 andw xfor the SCA2 2,20 , i.e. on the telomeric side of the
PMCH locus and excluding, therefore, the MCH gene asinvolved in either of these diseases.
In the present study, we demonstrate that both MCHmRNA and MCH peptide are synthesized in neurons of thedorso-lateral hypothalamus of adult human brain, extend-
w xing earlier immunohistochemical studies 6,32 . In addi-
tion, expression of MCH gene was also revealed by usingRT-PCR in other brain areas, such as the pallidum, thesubiculum, the cerebellar cortex and the neocortex. Noneof these brain areas was found to express MCH mRNA in
w xthe rat 4,38 , suggesting that synthesis of MCH RNA inthese novel sites may be restricted to the human brain. Atthe present time, however, direct evidence for intrinsicsynthesis of MCH gene product in extra-hypothalamicareas of the human brain using in situ hybridization islacking. The low amount of MCH mRNA in these areas
w xcould reflect transcriptional leakage 10 or an axonalorigin as demonstrated for the hypothalamic vasopressin
w xand oxytocin gene transcripts 31 .Expression of the MCH mRNA and a MCH-like pep-
tide was also found in various peripheral human organs.The highest levels of MCH-specific PCR products andMCH-like peptides were found in the thymus of neonatesand low but significant amounts of MCH mRNA wereidentified in spleen, bonemarrow and lymphatic gland. Thepresence of MCH mRNA and a pro-MCH-derived peptidein organs forming parts of the immune system may reflectan involvement of MCH-like peptides in the control ofimmune responses. It is worth noting in this context thatMCH and NEI are co-localized in resident macrophages of
w x Žthe intestine 23 and spleen Hervieu and Nahon, unpub-.lished data in the rat. In addition, MCH mRNA was also
visualized in the duodenum and testis, two organs wherew xMCH-positive cells were localized in rat 21,22 .
One intringuing site of MCH mRNA and MCH-likepeptide synthesis in peripheral tissues of human neonates
Ž .is brown adipose tissue Fig. 7, lane 8; and Fig. 8F . Thebrown fat is designed to produce heat through uncouplingfuel oxidation in mitochondria and represents, therefore,
w xthe physiological opposite of white fat 15 . The absence ofMCH expression in white adipose tissue may indicate arestricted function of MCH in the release of energy. Ofpotentially greater importance is the fact that MCH is
w xover-expressed in obrob mice 42 and participate as amajor component in the control of food-intake behavior in
w xthe rat 41,42 . Whether MCH in adipose tissue is alsoinvolved in the physiological regulation of energy expendi-ture in human will need further investigation.
An additional MCH RNA species of shorter length thanmature MCH mRNA was identified in hypothalamic andperipheral RNA samples of human neonates and adults.RT-PCR analysis with several primers revealed that exonII of the MCH gene was lacking in the resulting processed
Ž .RNA Fig. 7, and not shown . Very recently, we haveestablished that a short form of MCH RNA resulting froman alternative splicing may be produced in the rat hypo-
w xthalamus 51 . This alternative RNA splicing would allowsynthesis of a chimeric protein with a N-terminus identicalto that of pro-MCH encoded in exon I and a C-terminalregion overlapping in a different reading frame the codingsequence and non-coding part of the exon III MCH gene.Cloning and sequencing of the short RT-PCR product from

( )A. Viale et al.rMolecular Brain Research 46 1997 243–255 253
human tissues should clarify the nature of the correspond-ing RNA species.
Interestingly, both the mature MCH mRNA and theshort RNA species were identified in some brain areas and
Ž .peripheral organs Fig. 5BFig. 7 whereas a single full-length MCH mRNA was found in other non-hypothalamic
Ž .brain areas neocortex, hippocampus and subiculum or inŽperipheral neonatal tissues e.g. brown adipose tissue and
.bonemarrow , suggesting a tissue-specific regulation ofMCH RNA splicing with a possible functional relevance.
A striking result in analyzing MCH expression in hu-man tissues is the occurrence of a large MCH-im-munoreactive peptide in peripheral tissues isolated fromneonates or adults. In contrast, only the small biologicallyactive peptides MCH and NEI were identified in thehuman hypothalamus. The identification of NEI-GR as aproduct of the CTU procedure suggested that this largepeptide found in peripheral organs is composed, at least, ofa NEI sequence in its N-terminus and ends with a C-termi-nal MCH sequence. The pro-MCH processing intermediatewas found in large amount in many peripheral tissues,including thymus, colon, duodenum and lymphatic gangliaŽ .Fig. 8C–F, and not shown , and it may represent either aninactive processing intermediate or a peptide with hithertounknown biologically activities. Tissue-specific and differ-ential processing of a peptidic precursor is well-illustrated
w x w xwith pro-opiomelanocortin 35 , proglucagon 28,44 andw xproneurotensin 9 . The factors that control the processing
of the mammalian pro-MCH have not yet been identified.ŽIn particular, the role of the prohormone convertases re-
w x.viewed in 19,48 in the cell-specific processing of thehuman pro-MCH should be investigated.
In summary, we have cloned and sequenced the humangene for MCH and have localized it to a restricted area ofthe chromosome 12q23.1. Strong MCH gene expressionwas found in hypothalamus and in some peripheral tissues.A shorter MCH RNA product, likely the result of alternatesplicing, was identified in hypothalamus but also in pal-lidum, cerebellar cortex as well as in thymus and testis.Unexpectedly, whereas in the hypothalamus expression ofthe full MCH mRNA gave rise to mature MCH and NEIpeptides, in the periphery, it led to the synthesis of aC-terminal intermediate encompassing NEI and MCH.These results provide strong evidence that tissue-specificproduction of MCH and associated peptides is regulatedboth at the level of transcript synthesis and at that ofpro-MCH processing in human brain and peripheral tis-sues.
Acknowledgements
ŽWe are grateful to Drs. A. Beaudet McGill University,.Montreal, Quebec, Canada , P. Kitabgi and F. Presse
Ž .IPMC, Valbonne, France for critical reading of theŽmanuscript and to Dr. J. Grosgeorge LGMCH, Nice,
.France for technical assistance. We thank very much Drs.Ž . ŽD. Le Paslier CEPH, Paris, France , P. Vernier Institut A.
. ŽFessard, Gif-sur-Yvette, France and F. Agid Hopitalˆ.Pitie-Salpetriere, Paris, France for the generous gift of the´ ´ `
YAC library, human RNA and tissues samples, respec-tively. We thank J. Kervella for typing the manuscript andF. Aguila for preparation of artworks. This work wassupported by the Association pour la Recherche sur le
Ž .Cancer ARC 6987 , the Groupement de Recherches etŽ .d’Etudes sur les Genomes GREG 100r94 and the Min-´
Ž .istere de l’Education Nationale 95G0099 . A. Viale and`Y. Zhixing are recipients of fellowships from the ADER-
Ž .PACA CAR 9312r2679 and the ARC, respectively.
References
w x1 Baker, B.I., Melanin-concentrating hormone: a general vertebrateŽ .neuropeptide, Int. ReÕ. Cytol., 126 1991 1–47.
w x2 Belal, S., Cancel, G., Stenavin, G., Hentati, F., Khati, C., BenHamida, C., Auberger, G., Agid, Y., Ben Hamida, M. and Brice, A.,Clinical and genetic analysis of a Tunisian family with autosomaldominant cerebellar ataxia type 1 linked to the SCA2 locus, Neurol-
Ž .ogy, 44 1994 1423–1426.w x3 Bidard, J.N., De Nadai, F., Rovere, C., Moinier, D., Laur, J.,`
Martinez, J., Cuber, J.C. and Kitabgi, P., Immunological and bio-chemical characterization of processing products from the neu-rotensinrneuromedin N precursor in the rat medullary thyroid carci-
Ž .noma 6-23 cell line, Biochem. J., 291 1993 225–233.w x4 Bittencourt, J.C., Presse, F., Arias, C., Peto, C., Vaughan, J., Nahon,
J.L., Vale, W. and Sawchenko, P.E., The melanin-concentratinghormone system of the rat brain: an immuno- and hybridization
Ž .histochemical characterization, J. Comp. Neurol., 319 1992 218–245.
w x5 Bluet-Pajot, M.T., Presse, F., Voko, Z., Hoeger, C., Mounier, F.,¨Epelbaum, J. and Nahon, J.L., Neuropeptide-E-I antagonizes theaction of melanin-concentrating hormone on stress-induced release
Ž .of adrenocorticotropin in the rat, J. Neuroendocrinol., 7 1995297–303.
w x6 Bresson, J.L., Clavequin, M.C., Fellman, D. and Bugnon, C., Hu-man hypothalamic neuronal system revealed with a salmon
Ž .melanin-concentrating hormone MCH antiserum, Neurosci. Lett.,Ž .102 1989 39–43.
w x7 Breton, C., Presse, F., Hervieu, G. and Nahon, J.L., Structure andregulation of the mouse melanin-concentrating hormone mRNA and
Ž .gene, Mol. Cell. Neurosci., 4 1993 271–284.w x8 Breton, C., Schorpp, M. and Nahon, J.L., Isolation and characteriza-
tion of the human melanin-concentrating hormone gene and a vari-Ž .ant gene, Mol. Brain. Res., 18 1993 297–310.
w x9 Carraway, R.E. and Mitra, S.P., Posttranslational proccessing of theneurotensinrneuromedin-N precursor. In P. Kitabgi and C.B. Ne-
Ž .meroff Eds. , The Neurobiology of Neurotensin, Vol. 668, NewYork Academy Science, New York, NY, 1992, pp. 1–16.
w x10 Chelly, J., Concordet, J.-P., Kaplan, J.-C. and Kahn, A., Illegitimatetranscription: transcription of any gene in any cell type, Proc. Natl.
Ž .Acad. Sci. USA, 86 1989 2617–2621.w x11 Chomczynski, P. and Sacchi, N., Single-step method of RNA isola-
tion by acid guanidinium thiocyanate-phenol-chloroform extraction,Ž .Anal. Biochem., 162 1987 156–159.
w x12 Craddock, N., Dawson, E., Burge, S., Parfitt, L., Mant, B., Roberts,Q., Daniels, J., Gill, M., McGuffin, P., Powell, J. and Owen, M.,The gene for Darier’s disease maps to chromosome 12q23–q24.1,
Ž .Hum. Mol. Genet., 2 1993 1941-1943.

( )A. Viale et al.rMolecular Brain Research 46 1997 243–255254
w x13 Dessen, P., Fondrat, C., Valencien, C. and Mugnier, C., Bisance: aFrench service for access to biomolecular sequence databases,
Ž .Cabios, 6 1990 355–356.w x Ž .14 Eberle, A.N., Melanin-concentrating hormone. In A.N. Eberle Ed. ,
The Melanotropins, Karger, Basel, Switzerland, 1988, pp. 321–332.w x15 Flier, J.S., The adipocyte: storage depot or node on the energy
Ž .information superhighway? Cell, 80 1995 15–18.w x16 Gispert, S., Twells, R., Orozco, G., Brice, A., Weber, J., Heredero,
L., Scheufler, K., Riley, B., Allotey, R., Nothers, C., Hillermann, R.,Lunkes, A., Khati, C., Stevanin, G., Hernandez, A., Magarino, C.,Klockgether, T., Durr, A., Chneiweiss, H., Enczmann, J., Farrall,M., Beckmann, J., Mullan, M., Wernet, P., Agid, Y., Freund, H.-J.,Williamson, R., Auburger, G. and Chamberlain, S., Chromosomalassignment of the second locus for autosomal dominant cerebellar
Ž . Ž .ataxia SCA2 to chromosome 12q23–24.1, Nature Genet., 4 1993295–299.
w x17 Gonzalez, M.I., Vaziri, S. and Wilson, C.A., Behavioral effects ofa-MSH and MCH after central administration in the female rat,
Ž .Peptides, 17 1996 171–177.w x18 Granok, H., Leibovitch, B.A., Shaffer, C.D. and Elgin, S.C.R.,
Ž .Ga-ga over GAGA factor, Curr. Biol., 5 1995 238–241.w x19 Halban, P.A. and Irminger, J.-C., Sorting and processing of secre-
Ž .tory proteins, Biochem. J., 299 1994 1–18.w x20 Hernandez, A., Magarino, C., Gispert, S., Santos, N., Lunkes, A.,
Orozco, G., Heredero, L., Beckmann, J. and Auberger, G., GeneticŽ .mapping of the spinocerebellar ataxia 2 SCA2 locus on chromo-
Ž .some 12q23–q24.1, Genomics, 25 1995 433–435.w x21 Hervieu, G. and Nahon, J.L., Pro-melanin concentrating hormone
messenger ribonucleic acid and peptides expression in peripheralŽ .tissues of the rat, Neuroendocrinology, 61 1995 348–364.
w x22 Hervieu, G., Segretain, D. and Nahon, J.L., Developmental andstage-dependent expression of melanin-concentrating hormone in
Ž .mammalian germ cells, Biol. Reprod., 54 1996 1161–1172.w x23 Hervieu, G., Volant, K., Grishina, O., Descroix-Vagne, M. and
Nahon, J.L., Similarities in cellular expression and functions ofmelanin-concentrating hormone and atrial natriuretic factor in the rat
Ž .digestive tract, Endocrinology, 137 1996 561–571.w x24 Karin, M., Haslinger, A., Holtgreve, H., Richards, R.I., Krauter, P.,
Westphal, H.M. and Beato, M., Characterization of DNA sequencesthrough which cadmium and glucocorticoid hormones induce human
Ž .metallothionein-II gene, Nature, 308 1984 513–519.Aw x25 Kawauchi, H., Kawazoe, I., Tsubokawa, M., Kishida, M. and Baker,
B.I., Characterization of melanin-concentrating hormone in chumŽ .salmon pituitaries, Nature, 305 1983 321–323.
w x26 Krauter, K., Montgomery, K., Yoon, S.-J., LeBlanc-Straceski, J.,Renault, B., Marondel, I., Herdman, V., Cupelli, L., Banks, A.,Lieman, J., Menninger, J., Bray-Ward, P., Nadkarni, P., Weis-senbach, J., Le Paslier, D., Rigault, P., Chumakov, I., Cohe, D.,Miller, P., Ward, D. and Kucherlapati, R., A second-generation
Ž .YAC contig map of human chromosome 12, Nature, 377 1995321–333.
w x27 Lee, W., Mitchell, P. and Tjian, R., Purified transcription factorAP-1 interacts with TPA-inducible enhancer elements, Cell, 49Ž .1987 741–752.
w x28 Lui, E.Y., Asa, S.L., Drucker, D.J., Lee, Y.C. and Brubaker, P.L.,Glucagon and related peptides in fetal rat hypothalamus in vivo and
Ž .in vitro, Endocrinology, 126 1990 110–117.w x29 McBride, R.B., Beckwith, B.E., Swenson, R.R., Sawyer, T.K.,
Hadley, M.E., Matsunaga, T.O. and Hruby, V.J., The actions ofŽ .melanin-concentrating hormone MCH on passive avoidance in
Ž .rats: a preliminary study, Peptides, 15 1994 757–759.w x30 Miller, C.L., Hruby, V.J., Matsunaga, T.O. and Bickford, P.C.,
Alpha-MSH and MCH are functional antagonists in a CNS auditoryŽ .gating paradigm, Peptides, 14 1993 431–440.
w x31 Mohr, E., Fehr, S. and Richter, D., Axonal transport of neuropeptideencoding mRNAs within the hypothalamo-hypophyseal tract of rats,
Ž .EMBO J., 10 1991 2419–2424.
w x32 Mouri, T., Takahashi, K., Kawauchi, H., Sone, M., Totsune, K.,Murakami, O., Itoi, K., Ohneda, M., Sasano, H. and Sasano, N.,Melanin-concentrating hormone in the human brain, Peptides, 14Ž .1993 643–646.
w x33 Nahon, J.L., The melanin-concentrating hormone: from the peptideŽ .to the gene, Crit. ReÕ. Neurobiol., 8 1994 221–262.
w x34 Nahon, J.L., Presse, F., Bittencourt, J.C., Sawchenko, P. and Vale,W., The rat melanin-concentrating hormone mRNA encodes multi-ple putative neuropeptides coexpressed in the dorsolateral hypothala-
Ž .mus, Endocrinology, 125 1989 2056–2065.w x35 O’Donohue, T.L. and Dorsa, D., The opiomelanotropinergic neu-
Ž .ronal and endocrine system, Peptides, 3 1982 353–395.w x36 Parkes, D.G. and Vale, W., Contrasting actions of melanin-con-
centrating hormone and neuropeptide-E-I on posterior pituitary func-Ž .tion. In A. Eberle and H. Vaudry Eds. , The Melanotropic Peptides,
Vol. 680, New York Academy of Sciences Series, 1993, pp. 588–590.w x37 Pedeutour, F., Szpirer, C. and Nahon, J.L., Assignment of the human
Ž .pro-melanin-concentrating hormone gene PMCH to chromosomeŽ .12q23–q24 and two variant genes PMCHL1 and PMCHL2 to
Ž .chromosome 5p14 and 5q12–q13, Genomics, 19 1994 31–37.w x38 Presse, F., Hervieu, G., Imaki, T., Sawchenko, P.E., Vale, W. and
Nahon, J.L., Rat melanin-concentrating hormone messenger ribonu-cleic acid expression: marked changes during development and after
Ž .stress and glucocorticoid stimuli, Endocrinology, 131 1992 1241–1250.
w x39 Presse, F. and Nahon, J.L., Differential regulation of melanin-con-centrating hormone gene expression in distinct hypothalamic areas
Ž .under osmotic stimulation in rat, Neuroscience, 55 1993 709–720.w x40 Presse, F., Nahon, J.L., Fischer, W.H. and Vale, W., Structure of the
human melanin-concentrating hormone mRNA, Mol. Endocrinol., 4Ž .1990 632–637.
w x41 Presse, F., Sorokovsky, I., Max, J.P., Nicolaidis, S. and Nahon, J.L.,Melanin-concentrating hormone is a potent anorectic peptide regu-lated by food-deprivation and glucopenia in the rat, Neuroscience,
Ž .71 1996 735–745.w x42 Qu, D., Ludwig, D.S., Gammeltoft, S., Piper, M., Pelleymounter,
M.A., Cullen, M.J., Mathes, W.F., Przupek, J., Kanarek, R. andMaratos-Flier, E., A role for melanin-concentrating hormone in the
Ž .central regulation of feeding behaviour, Nature, 380 1996 243–247.w x43 Reece, R.J. and Maxwell, A., DNA gyrase: structure and function,
Ž .Crit. ReÕ. Biochem. Mol. Biol., 26 1991 335-375.w x44 Rouille, Y., Martin, S. and Steiner, D.F., Differential processing of´
proglucagon by the subtilisin-like prohormone convertases PC2 andPC3 to generate either glucagon of glucagon-like peptides, J. Biol.
Ž .Chem., 270 1995 26488–26496.w x45 Rovere, C., Viale, A., Nahon, J.L. and Kitabgi, P., Impaired process-`
ing of brain pro-neurotensin and pro-melanin-concentrating hormoneŽ .in obese fatr fat mice, Endocrinology, 137 1996 2954–2958.
w x46 Sambrook, J., Fritsch, E.F. and Maniatis, T., Molecular Cloning. ALaboratory Manual, 2nd edn., Cold Spring Harbor Laboratory Press,Cold Spring Harbor, USA, 1989.
w x Ž .47 Schatz, D.G., Oettinger, M.A. and Baltimore, D., The V D J recom-Ž .bination activating gene, RAG-1, Cell, 59 1989 1035–1048.
w x48 Seidah, N.G. and Chretien, M., Proprotein and prohormone conver-´tases of the subtilisin family. Recent developements and future
Ž .perspectives, Trends Endocrinol. Metab., 3 1992 133–140.w x49 Sheng, M. and Greenberg, M.E., The regulation and function of
c-fos and other immediate early genes in the nervous system,Ž .Neuron, 4 1990 477–485.
w x50 Thompson, R.C. and Watson, S.J., Nucleotide sequence and tissue-specific expression of the rat melanin concentrating hormone gene,
Ž .DNA Cell Biol., 9 1990 637–645.w x51 Toumaniantz, G., Bittencourt, J.C. and Nahon, J.L., The rat
melanin-concentrating hormone gene encodes an additional putativeŽ .protein in a different reading frame, Endocrinology, 137 1996
4518–4521.w x52 Vaughan, J.M., Fischer, W.H., Hoeger, C., Rivier, J. and Vale, W.,

( )A. Viale et al.rMolecular Brain Research 46 1997 243–255 255
Characterization of melanin-concentrating hormone from rat hypo-Ž .thalamus, Endocrinology, 125 1989 1660–1665.
w x X53 Wall, L., De Boer, E. and Grosveld, F., The human b-globin gene 3enhancer contains multiple binding sites for an erythroid-specific
Ž .protein, Genes DeÕ., 1988 1089–1100.w x54 Wang, J.C., Interactions between twisted DNAs and enzymes: the
Ž .effects of superhelical turns, J. Biol. Chem., 87 1974 797-816.
w x55 Yang, Z., Sugawara, M., Ponath, P.D., Wessendorf, L., Banerji, J.,Li, Y. and Strominger, J.L., Interferon g response region in thepromoter of the human DPA gene, Proc. Natl. Acad. Sci. USA, 87Ž .1990 9226–9230.
w x56 Zhixing, Y. and Nahon, J.L., DNA gyrase improves DNA transfor-mation of E. coli cells with large recombinant plasmids, Nucleic
Ž .Acids Res., 23 1995 3353–3354.




















