
The mangrove communities in the Arroyo Seco deltaic fan, Jalisco, Mexico, and their relation with...
16
The mangrove communities in the Arroyo Seco deltaic fan, Jalisco, Mexico, and their relation with the geomorphic and physical–geographic zonation A.P. Méndez Linares a, ⁎ , J. López-Portillo b , J.R. Hernández-Santana a , M.A. Ortiz Pérez a , O. Oropeza Orozco a a Instituto de Geografía, Universidad Nacional Autónoma de México, Apdo. Postal 01000, México 04510 D. F., México b Departamento de Ecología Funcional, Instituto de Ecología A. C., Apdo. Postal 63, Xalapa 91070 Veracruz, México Received 26 August 2004; received in revised form 17 March 2006; accepted 5 May 2006 Abstract This work investigates mangrove tree zonation dynamics in a deltaic fan in the southwestern coast of Mexico. A hypothetical evolution that led to the formation and consolidation of the Arroyo Seco deltaic fan based on the environmental components from the hydrographic basin is proposed. Also, the relief morphogenetic types, along with their sedimentary composition, are derived from geomorphologic and edaphic diagnose methods. Finally, mangrove tree zonation, basal area and height along transects traced parallel to the relief are related to morphogenetic types and to the water table depth during the dry period. There was a clear sedimentological evidence of the accretion process in the deltaic fan. The accretion horizons conditioned the water table, which increased in depth from the margin of the lagoon to the hinterland, thus changing among geoforms. It was much greater at the middle part of the transects, where continental or lagoon water did not infiltrate due to high resistance of clayey soils. The geomorphic zonation of mangrove communities in the Arroyo Seco deltaic fan is: (1) a low fluvial distributary accumulative plain, with coconut palm plantations; (2) a bank fluvial accumulative plain between distributary channels, with coconut palm plantations and patches of low tropical forest; (3) a relatively high swamp accumulative plain, flooded during high water levels, with Avicennia germinans; (4) a low swamp, temporarily flooded accumulative plain, with Rhizophora mangle, La- guncularia racemosa and A. germinans; (5) a very low swamp, permanently flooded accumulative plain, with distributary channels, lateral microbars and tide microdepressions, where Rhizophora and Laguncularia codominate; and (6), a Rhizophora fringe and colonization area along the fan's frontal edge on the same plain. Along this gradient composed of six mangrove communities, basal area and height tended to increase as the water table reached soil level. The relevance of this study rests on the use of both geomorphological and ecological methods to explain the development and distribution of mangrove communities in an actively accreting area. It also highlights the importance of the water table on the survival, structure and zonation of mangrove communities. © 2006 Elsevier B.V. All rights reserved. Keywords: Deltaic fan; Mangrove zonation; Sedimentary accretion 1. Introduction Coastal ecosystems, including mangroves, have a signif- icant ecological and economic importance at global, regional and local scales. They reduce erosion in coastal areas by ameliorating the effects of waves and currents, provide protection from extreme hydrodynamic and meteorological phenomena and changes, contribute to soil formation, shelter many marine animal species, contribute to coastal produc- tivity by functioning as nursing areas for fish and crustaceans, favor tourism development in coasts as leisure areas, and constitute an important source of forest resources, Catena 70 (2007) 127 – 142 www.elsevier.com/locate/catena ⁎ Corresponding author. Tel.: +52 55 56224356; fax: +52 55 56162145. E-mail address: [email protected] (A.P. Méndez Linares). 0341-8162/$ - see front matter © 2006 Elsevier B.V. All rights reserved. doi:10.1016/j.catena.2006.05.010
Transcript of The mangrove communities in the Arroyo Seco deltaic fan, Jalisco, Mexico, and their relation with...

) 127–142www.elsevier.com/locate/catena
Catena 70 (2007
The mangrove communities in the Arroyo Seco deltaic fan, Jalisco,Mexico, and their relation with the geomorphic
and physical–geographic zonation
A.P. Méndez Linares a,⁎, J. López-Portillo b, J.R. Hernández-Santana a, M.A. Ortiz Pérez a,O. Oropeza Orozco a
a Instituto de Geografía, Universidad Nacional Autónoma de México, Apdo. Postal 01000, México 04510 D. F., Méxicob Departamento de Ecología Funcional, Instituto de Ecología A. C., Apdo. Postal 63, Xalapa 91070 Veracruz, México
Received 26 August 2004; received in revised form 17 March 2006; accepted 5 May 2006
Abstract
This work investigates mangrove tree zonation dynamics in a deltaic fan in the southwestern coast of Mexico. A hypothetical evolutionthat led to the formation and consolidation of the Arroyo Seco deltaic fan based on the environmental components from the hydrographicbasin is proposed. Also, the relief morphogenetic types, along with their sedimentary composition, are derived from geomorphologic andedaphic diagnose methods. Finally, mangrove tree zonation, basal area and height along transects traced parallel to the relief are related tomorphogenetic types and to the water table depth during the dry period. There was a clear sedimentological evidence of the accretion processin the deltaic fan. The accretion horizons conditioned the water table, which increased in depth from the margin of the lagoon to thehinterland, thus changing among geoforms. It was much greater at the middle part of the transects, where continental or lagoon water did notinfiltrate due to high resistance of clayey soils. The geomorphic zonation of mangrove communities in the Arroyo Seco deltaic fan is: (1) alow fluvial distributary accumulative plain, with coconut palm plantations; (2) a bank fluvial accumulative plain between distributarychannels, with coconut palm plantations and patches of low tropical forest; (3) a relatively high swamp accumulative plain, flooded duringhigh water levels, with Avicennia germinans; (4) a low swamp, temporarily flooded accumulative plain, with Rhizophora mangle, La-guncularia racemosa and A. germinans; (5) a very low swamp, permanently flooded accumulative plain, with distributary channels, lateralmicrobars and tide microdepressions, where Rhizophora and Laguncularia codominate; and (6), a Rhizophora fringe and colonization areaalong the fan's frontal edge on the same plain. Along this gradient composed of six mangrove communities, basal area and height tended toincrease as the water table reached soil level.
The relevance of this study rests on the use of both geomorphological and ecological methods to explain the development and distributionof mangrove communities in an actively accreting area. It also highlights the importance of the water table on the survival, structure andzonation of mangrove communities.© 2006 Elsevier B.V. All rights reserved.
Keywords: Deltaic fan; Mangrove zonation; Sedimentary accretion
1. Introduction
Coastal ecosystems, including mangroves, have a signif-icant ecological and economic importance at global, regional
⁎ Corresponding author. Tel.: +52 55 56224356; fax: +52 55 56162145.E-mail address: [email protected]
(A.P. Méndez Linares).
0341-8162/$ - see front matter © 2006 Elsevier B.V. All rights reserved.doi:10.1016/j.catena.2006.05.010
and local scales. They reduce erosion in coastal areas byameliorating the effects of waves and currents, provideprotection from extreme hydrodynamic and meteorologicalphenomena and changes, contribute to soil formation, sheltermany marine animal species, contribute to coastal produc-tivity by functioning as nursing areas for fish andcrustaceans, favor tourism development in coasts as leisureareas, and constitute an important source of forest resources,

128 A.P. Méndez Linares et al. / Catena 70 (2007) 127–142
mostly at a local level (Dawes, 1986; Gallegos, 1986; Valdéz,1991; Blasco et al., 1996). For these reasons, the scientificcommunity has led efforts to investigate the structural,functional and dynamic regularities that characterize thesefragile ecosystems, which may be aimed at establishing thebases for the sustainable management of littoral resources.
Coastal zones represent areas that include high-energyphysical stages, where geophysical, terrestrial and oceanicprocesses interact. These processes permanently model thecontinental edge and influence the structural features andspatial distribution of the ecosystems associated to it(Pannier, 1992). In turn, these territories are influenced bychanges in mean sea level, as well as by extreme naturalphenomena such as earthquakes, hurricanes and tsunamis,which may give rise to radical physical modifications overgeological or historical periods. Since plant distributionresponds to changes in habitat, the modifications induced bygeomorphic processes can occur regardless of biotic factors.This concept has been applied to low tropical coasts,particularly where mangrove communities develop, sincethese communities frequently grow in habitats that varysubstantially in their geomorphological characteristics(Thom, 1967; Semeniuk and Semeniuk, 1995).
Mangroves display distinctive morphological, structural,physiological and floristic characteristics that differentiatethem from other plant communities. The frequent distributionin stripes or zonation of the different species is worth noting,since zones can be dominated by a given species, orcodominated by two or more (Valdéz, 1991; Young andHarvey, 1996; Blasco et al., 1996; Duke and Ball, 1998; Ewelet al., 1998a,b; Ball, 1998). Zonation is likely the mangroveecosystem's response to external factors rather than a temporalsequence induced by the plants themselves. It is generallyconsidered that the spatial distributionmight also be associatedto changes in water level and salinity (Rzedowski, 1978; Rico-Gray and Palacios-Ríos, 1996). For instance, in a coastallagoon in the Gulf of Mexico, the highest plant diversity andlargest trunk diameter in three mangrove species (Rhizophoramangle, Laguncularia racemosa and Avicennia germinans)were correlated to relatively low salinities found in theinterfluvial basin. On the other hand, A. germinans cover wasgreater in the mudflat, a geoform with a higher salinity. It alsoseems clear that the changes in microtopography due toaccretion or subsidence exert a controlling role on mangrovedistribution, this being a most important factor, along withsubstrate type and extreme hydrological and oceanographicregimes (Thom, 1967; López-Portillo and Ezcurra, 1989;López-Portillo et al., 1989; Smith, 1992).
A number of classifications of mangrove communitytypes exist, but the main criteria upon which these are basedinclude two types: physiognomic and geomorphological.Because of its physiognomic approach, the paper mostfrequently cited in the literature is that of Lugo and Snedaker(1974), who classified mangrove communities into fiveforest groups: fringe, riverine, overwash, basin and dwarf.This classification has been amply used, even to suggest the
provision of different goods and services by mangroves(Ewel et al., 1998a). In Mexico, for example, it has beenapplied to describe riverine, and basin mangrove habitatswhere R. mangle, L. racemosa, and A. germinans coexist,withR. mangle as the dominant species in the state of Chiapas(Gallegos, 1986). Also, it has been applied in the extensivefringe mangrove forests along marine and lagoon margins,and in the high basin, low swamp known as Petén (hammock)in the Celestún area within the calcareous Yucatan Peninsula,where four major mangrove types were identified, and furthersubdivided into 10 variants (Trejo-Torres et al., 1993).
However, a geomorphological perspective is essential tounderstand the history, distribution and dynamics ofmangrovecommunities in deltaic zones. Nichols et al. (1998), O'Neillet al. (1988), Müller et al. (2004), Coblentz and Ritters (2004),and Priego-Santander (2004) have reported a significantcorrelation between geomorphologic heterogeneity and bio-diversity, which in one case (Priego-Santander, 2004)accounted for as much as 82% of the variability in plantspecies distribution. These papers show that slope, aspect anddrainage are of great relevance to explain plant communitydistribution. Such approaches, suitable for large territories andmarked topographical gradients, are less effective in smallerareas where the slope may be lower than 0.1% andwhere plantcommunities and species are fewer, as in our study site, thealluvial fan in Arroyo Seco.
The geomorphological approach deals with the morphom-etry, genesis, structure, dynamics and age of the relief, some ofwhich are not considered in the previously cited reports. In thiscontext, Thom (1967) made significant contributions tomangrove forest geomorphology. Based on the stratigraphyfrom profiles up to 4m deep in the Grijalva–Usumacinta delta,Tabasco, Thom (1967) inferred the evolution of mangrovehabitats within the context of an active delta, and showed thatsedimentation rates and subsidence model landscape mor-phology. More evidences have been published concerning theclose correlation between vegetation patterns, sedimentationand coastal development (Thom, 1984; Woodroffe, 1992).This approach is used in the present work, by studying thegeomorphological structure of the Arroyo Seco basin,particularly its deltaic fan, in the state of Jalisco.
In this paper, a morphogenetic and chronologic classifi-cation of the relief is enriched with a physico–geographicalapproach that includes soil descriptions, the depth of thewater table and substrate salinity. In this sense, the mainobjective of the present work was to deduce the developmentof the Arroyo Seco deltaic fan, an actively accretingenvironment, and to relate it to the sedimentary compositionand the distribution in time and space of mangrove species.
2. Materials and methods
2.1. Study area
The Arroyo Seco deltaic fan is located to the southeast ofthe Jalisco coast in the Pacific coast of Mexico (19°10′50″–
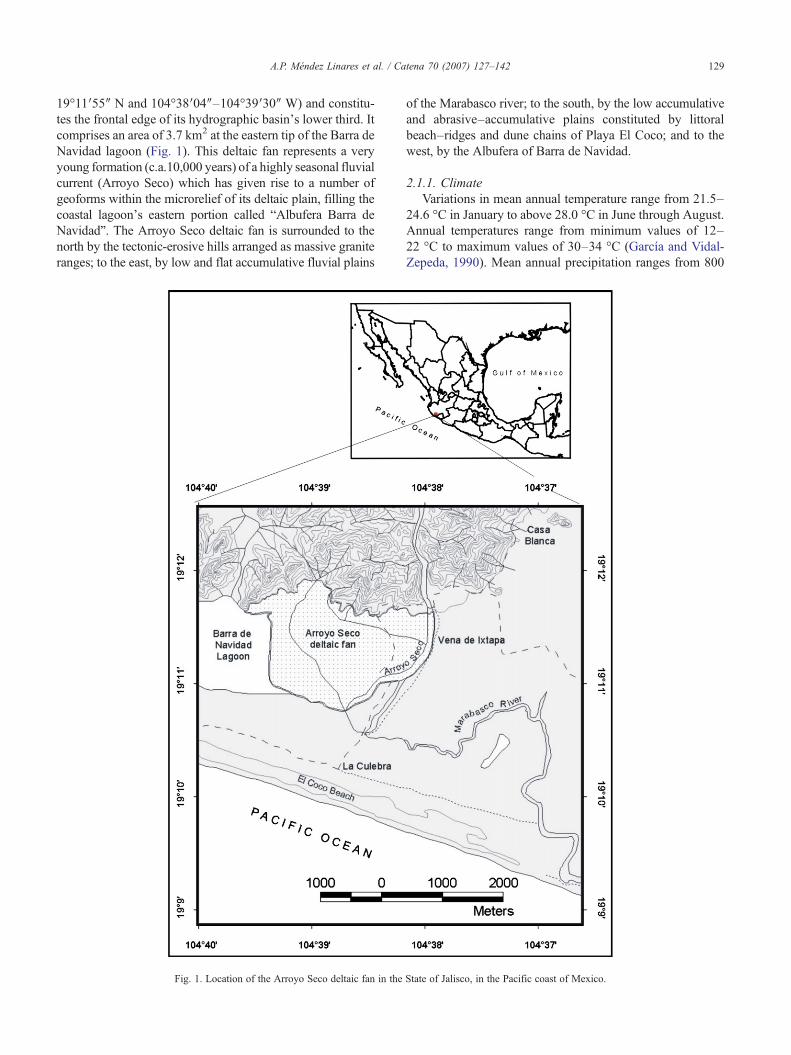
129A.P. Méndez Linares et al. / Catena 70 (2007) 127–142
19°11′55″ N and 104°38′04″–104°39′30″ W) and constitu-tes the frontal edge of its hydrographic basin's lower third. Itcomprises an area of 3.7 km2 at the eastern tip of the Barra deNavidad lagoon (Fig. 1). This deltaic fan represents a veryyoung formation (c.a.10,000 years) of a highly seasonal fluvialcurrent (Arroyo Seco) which has given rise to a number ofgeoforms within the microrelief of its deltaic plain, filling thecoastal lagoon's eastern portion called “Albufera Barra deNavidad”. The Arroyo Seco deltaic fan is surrounded to thenorth by the tectonic-erosive hills arranged as massive graniteranges; to the east, by low and flat accumulative fluvial plains
Fig. 1. Location of the Arroyo Seco deltaic fan in the
of the Marabasco river; to the south, by the low accumulativeand abrasive–accumulative plains constituted by littoralbeach–ridges and dune chains of Playa El Coco; and to thewest, by the Albufera of Barra de Navidad.
2.1.1. ClimateVariations in mean annual temperature range from 21.5–
24.6 °C in January to above 28.0 °C in June through August.Annual temperatures range from minimum values of 12–22 °C to maximum values of 30–34 °C (García and Vidal-Zepeda, 1990). Mean annual precipitation ranges from 800
State of Jalisco, in the Pacific coast of Mexico.

130 A.P. Méndez Linares et al. / Catena 70 (2007) 127–142
to 1200 mm at the basin's lower third, and from 1200 to1500 mm at its upper half, concentrated in summer and earlyautumn showers with occasional intensities of 200–400 mmin 24 h (Vidal-Zepeda, 1990a,b).
2.1.2. GeologyThe Arroyo Seco hydrographic basin is located in the
coastal portion of a large Sierra Madre del Sur Geomorphic–Tectonic Unit, formed between the upper Mesozoic and theTertiary (López-Ramos, 1983). The region's relief isconditioned by tectonic activity dating from the Neogene(Miocene–Pliocene) and still intense at present (Lugo Hubp,1990). The basin substrate is composed of igneous rocks(Jurassic and Cretaceous granites and granodiorites, tuffs andacid and basic extrusive components), marine carbonatedsedimentary complexes, and Quaternary terrigenous conti-nental elements (Fig. 2). Quaternary deposits consist of
Fig. 2. Geological constitution of the Arroyo Seco hydrographic basin, State of Jalis(5) sandstone and conglomerates; (6) residual sediments; (7) alluvial sediments; (8)E13B31 (1975), E13B32 (1975), E13B41 (1974) and E13B42 (1974).
alluvial sequences of clays, sands and non-consolidatedpebbles in intramountain valleys, alluvial plains, and thebottom of fluvial valleys. Marine accumulative ancient spitgeoforms occur along the littoral, occasionally consolidatedin tombolo, or formed by non-consolidated littoral sediments(mainly sands) deposited by wave action and littoral driftcurrents (López-Ramos, 1983; Lugo Hubp, 1990).
2.1.3. GeomorphologyThe hydrographic basin is within the Jalisco morphos-
tructural macroblock, where denudation, erosion and abrasionled to the development of extensive relief planation summitand watershed surfaces and highland fragments, originallymarine and subsequently remodeled by weathering anderosive–denudation processes. Standing out in the ArroyoSeco hydrographic basin spectrum of geomorphic steps are theyounger plain relief types (H, heightsb40 masl), coastal hills
co, Mexico. (1) Granite; (2) basic and acid extrusives; (3) tuffs; (4) limestone;swamp sediments. Sources: CETENAL 1:50,000 geological charts numbers

131A.P. Méndez Linares et al. / Catena 70 (2007) 127–142
or elevations (Hb220m), premountains (400bHb800m) andlow mountains (800bHb1200 m) (Hernández-Santana et al.,1996).
2.1.4. HydrologyMean annual runoff values oscillate between 500 and
1000 mm, mostly between July and October (Jiménez andMaderey-Rascón, 1990). By pluvial period, runoff valuesconcentrate 500 mm in the rainy season (June–October) andbetween 100 and 500 mm in the dry season (November–May).High runoff in summer and early autumn result in the transport oflarge volumes of sediments, mostly sandy, among the highest inMexico, in the range of 500 to 10,000 Mg km−2 year−1
(Maderey-Rascón, 1990). This hydro-meteorological phenome-non constitutes the main mechanism among the alluvial pro-cesses of the deltaic fan's formation, growth and modification.
2.1.5. SoilsThe soil types in the study area correspond to Regosols,
Lithosols and Faeozem, all of which are characterized by theabsence of diagnose horizons (FAO-UNESCO, 1990). Thesesoil types present a rocky cover that contributes further to theheavy denudation of the relief, and the growth of fluvialdeposition areas located along the whole valley and endingwith the deltaic fan.
2.1.6. Vegetation and land useOak forests dominate at the mountainous portion to the
east of the basin, where the highest precipitation occurs;small patches of tropical deciduous forests alternate withinduced pastureland to the west. There are large extensionsof tropical deciduous forests interspersed with sparse patchesof oak forests and grasslands in premountains and hills.Large irrigated agricultural lands occur on coastal alluvialplains. At the deltaic fan, mangrove forests develop along thetransition zone with the Barra de Navidad lagoon, which isbordered by dune vegetation at the littoral zone.
2.2. Evolution of the deltaic fan
The hypothetical evolution of the deltaic fan is based onthe morphodynamic and sedimentary analysis of the coastalzone (Rice, 1977; Tarbuck and Lutgens, 1999) and on theevaluation of comparative scenarios on the formation of spitsand tombolo in the Caribbean (Hernández-Santana et al.,2002). The geoforms, which are made up of littoral beach–ridges parallel to the coast and former islands merged tothem, form a complex spit–tombolo geoform that constitutesthe structural conditions for the local deposition of theArroyo Seco deltaic fan.
The proposed sequence of development is based on theestimated age of the littoral beach–ridges from the near statesof Nayarit and Sinaloa in Mexico (Curray et al., 1969). Theestimated age is 3600–4750 years BP (Early to Late Holo-cene). Judging by the time at which transgression began on theLate Pleistocene to Early Holocene (Curray andMoore, 1964).
The deposition from the littoral beach–ridges to the low tidalterraces and the advancement of the spit that consolidated thetombolo probably corresponds to Late Holocene.
2.3. Cartography and fieldwork
The thematic cartography was developed using the Arc/View 3.2 GIS. To characterize the relief of the Arroyo Secodeltaic fan the methods from general geomorphologicalanalyses were followed (Spiridonov, 1975; Díaz Díaz, 1986;Hernández-Santana and Reyes-González, 2002). Two1:50000 topographic charts (E13B41 and E13B42; INEGI,1992) were used as base and the delimitation of the reliefgenetic types were interpreted from a 1:20000 digital ortho-photo (E13B42a; INEGI, 1994) (Fig. 3). The basin'sgeological characterization was derived from thematic1:50000 cartography (E13B31, E13B32, E13B41 andE13B42; CETENAL, 1974, 1975, 1981). The geomorpho-logical classification of relief types was also derived fromtopographic interpretation with the aid of the previously citedorthophoto, the correlation between relief and geologicalstructure, and field verification.
Sampling points for seven representative edaphic profileswere selected within and in the vicinity of the deltaic fan,recording the existing geoforms in each of them. The horizonsfrom edaphic profiles were describedwith respect to thickness,depth, color, texture, structure and other properties related toorganic matter, carbonate and moisture content and defined inaccordance to the soil world classification (FAO-UNESCO,1990) modified by INEGI (1991). Two profiles were made onthe granitic basement on the coastal hills that limit the deltaicfan to the north. Four of the seven profiles (3–6) were used inthe elaboration of the fan's stratigraphy and of the generaledaphic profiles. The last profile was made on the actual outletof the Arroyo Seco in the Albufera Barra de Navidad, toexplore differences in sedimentary environments between theoutlet and the mangrove front.
Once the geoform typological units were interpreted,fieldwork was undertaken in July 2001 on two diagnoseprofiles (A–A′ and B–B′), both with an east–westorientation and perpendicular to the mangrove communityfront. Three analytical transects (I–I′, II–II′ and III–III′)were traced on December 2001 after photo interpretation(Fig. 3). The microrelief along these transects was measuredin the field with the help of a transit level at 20 m intervals.Also, following the methodology described by Matteucciand Colma (1982) and López-Portillo and Ezcurra (1989),plant sampling stations were placed every 40 m alongtransects I to III, making a total of 14, 17 and 11 samplingstations, respectively (the number of stations depending onthe length of each transect). In each sampling station, theheight and perimeter at breast height (which was laterconverted to basal area) of the four tallest trees of eachspecies were measured with a clinometer and a measuringtape, respectively. If the mangal consisted of two species,five individuals per species were measured, and ten

Fig. 3. Digital orthophoto including the Arroyo Seco deltaic fan in the Pacific coast of Mexico. White lines indicate field transects I, II and III, and profiles A andB. Dots mark the location of edaphic wells 1–7. Original source: INEGI, (1994) E13B42(1) 1: 20,000 digital orthophoto.
132 A.P. Méndez Linares et al. / Catena 70 (2007) 127–142
individuals if it was a monospecific mangal. We alsomeasured the cover of living and dead plants with a ConcaveSpherical Crown densiometer (Forestry Suppliers, Inc.,Jackson, Mississippi) on four measurement points alongthe main cardinal directions, and at 8-m distance from thecenter of the sampling station, which were later averaged togive one value per sampling station. Finally, we counted thenumber of seedlings and saplings in two 1 m2 quadrants at adistance of 2 m from the sampling point and a generaldescription of the plant community was done.
In each sampling point, we measured water table depth andsalinity during the dry season (onDecember 2001) with the aidof a hand auger with extensions to reach down to 1.8 m. Oncethe water table was reached, all water was extracted and thewater level allowed recovering for 30 min at least in order tomeasure its depth, after which a water sample was taken tomeasure salinity with the aid of an YSI 85 Water QualitySystem (YSI Environmental, Yellow Springs, Ohio).
2.4. Multivariate analysis of geomorphic, environmentaland mangrove vegetation properties
We used Canonical Correspondence Analysis (CCA) torelate an environmental data matrix (including the geoform,soil texture and organic matter, interstitial water salinity andwater table depth) with a biotic data matrix (including thebasal area of the three mangrove species and mangrove forest
height). Data under consideration corresponded to 31sampling stations (14 from transect I and 17 from transectII). We used CANOCO (Ter Braak, 1998; Lepš andŠmilauer, 2003) as an extension of the Cornell Ecologyprogram DECORANA (Hill, 1979). For ease of processing,we coded some of the variables as follows:
Geoforms: 1. Fluvial accumulative plain, of bank; 2.Swamp accumulative plain, low; 3. Swamp accumulativeplain, transitional; 4. Swamp accumulative plain, verylow; 5. Swamp plain, frontal;Soil texture: 1. Sandy, 2. Clayey–sandy, 3. Sandy–silty,4. Clayey–silty, 5. Silty, 6. Clayey;Carbonates (1.High, 2. Middle, and 3. Low reaction toHCl);Organic Matter (1.High, N50%; 2. Medium 25–50%; 3.Low, b25%; 4. Absent).
The remaining environmental variables, and all the bioticvariables were considered as continuous.
3. Results
3.1. Evolution of the deltaic fan
The basin's igneous nature (mostly granitic intrusiveelements) and the influence of the tropical hydrothermal

Fig. 4. Hypothetical evolution of relief in the Arroyo Seco deltaic fan and its surrounding areas in the Pacific coast of Mexico. Tones and symbols orderedaccording to the following numbers: Relief types: (1) tectonic-erosive hills; (2) abrasive and abrasive–accumulative plains; (3) accumulative fluvial plains; (4)deltaic fan accumulative fluvial plains; (5) swamp plains; (6) abrasive planation surfaces; (7) beaches. Relief forms: (8) dune chains and littoral beach–ridge axes;(9) abrasive coasts; (10) direction of littoral drift currents; (11) faults; (12) morphoalignments; (13) fluvial streams; (14) deltaic fan fluvial distributaries.
133A.P. Méndez Linares et al. / Catena 70 (2007) 127–142
regime determines a heavy weathering that facilitates theerosion and transport of large volumes of sediments whichconstitute the source for the littoral accumulative formsincluding the Arroyo Seco deltaic fan and Playa El Cocolittoral beach–ridges. Another sediment supply for theselandforms is transported by the Marabasco River, the coastalaccumulative plain which alternates with that of Arroyo Secoand that originally represented the major source for thedevelopment of littoral beach–ridges linking the mainland tothe ancient Cerro San Francisco (Navidad) island. Thisformer island, a littoral geoform, acted as a “sediment trap”during the development and extension of the Arroyo Secodeltaic fan. The linear direction of the beach–ridgesconnecting the former island indicates the transport of greatvolumes of sand by strong littoral currents.
The formation and consolidation of today's coastal edgetook place during the Quaternary period, as a response to theglobal scale variations in sea level along with the regional andlocal earth-crust neotectonic activity. Under these conditionsand from the evidence found during the geomorphologicalcharacterization and classification, the following hypotheticalevolutionary stages of the Arroyo Seco deltaic fan's relief andsurrounding area are proposed (Fig. 4).
• Mid-upper Pleistocene (QII–III): Morphostructural regu-larities resembling those of today's coastal geomorphicscenario become evident along the coastline, with theextension and growth of premountain ranges andfragments uplifted from the continental shelf with aseaward direction. In this development stage, the currentSan Francisco hills constituted small elevations (tens ofmeters high) located very close to today's coastal hillsthat were derived from the uplifted continental fringe,initially representing small islands with fragments ofmarine plains.
• Late upper Pleistocene–Early Holocene (QIII2 –IV
1 ): In thisstage the tectonic uplifting determined the territory'sgeneral elevation, contributing to an increase in erosionprocesses at mountain and premountain zones, with thesubsequent transport and accumulation in lower coastalareas. Plains fragments from the previous stage were remo-deled in uplifting events and shaped as hills surrounded by awider abrasive-accumulative plain, while at the same timethe Marabasco River alluvial plain was formed. In this waythe paragenetic development of a number of plains starts. Inturn, the littoral sea current regime gave rise to sedimentarytransport, constituting the remains of a small accumulative

134 A.P. Méndez Linares et al. / Catena 70 (2007) 127–142
spit. At this point, the first frontal sediments of the ArroyoSeco deltaic fan started to build up.
• Late Holocene (QIV2 ): The hills' coastal domain widens,
hence increasing the relief energy and fluvial accumula-tion in the Marabasco and Arroyo Seco alluvial plains.Under these conditions the spit got closer to the SanFrancisco Island, linking it with the mainland. At thismoment the basic geoforms of both the littoral relief anddeltaic fan were fully established.
• Late Holocene–Recent: In this final stage the swampplains surrounding the inner border of the Albufera deNavidad were consolidated, and different geoform typesbecame established and so did mangrove habitats, fol-lowing the micro-conditions created by relief. The fan is
Fig. 5. Geomorphological map of the Arroyo Seco deltaic fan and its surroundingnumbers. H in legend denotes altitude and h denotes relief relative height or dissectito Quaternary: (1) tectonic-erosive, block (20bHb220 m; h=60–100 m). Plains,accumulative, flat, slightly inclined (H=5–10 m); (3) fluvial accumulative, low, fladeltaic fan facies and littoral sea forms (Hb2–3 m); (5) fluvial accumulative, debetween deltaic fan distributaries, low (Hb2–3 m). Marine: (7) marine accumul(H=3–4 m; h=1–2 m), dune chains (H=6–8 m; h=4–5 m) and by beach sepermanently flooded, with a coastal alluvial fan microrelief (high water flow distribuswamp accumulative, low, flooded during high water flows, with distributing chanfluvial–marine, low and very low, resulting from the contact between alluvial andwhere lagoons occur today, consolidated during the Holocene (Hb2 m). Anthropi4 m). Submarine relief types: continental shelf plains: (12) abrasive–accumuundifferentiated; (13) accumulative, poorly consolidated, of deltaic fan submarinesubmarine stream, shallow (H=−2 to −4 m). Relief forms of different genesis: (15and filled seasonal intermittent stream beds and high water flows; (17) lagoons andlittoral dune ridges. Relief forms: (19) intermittent streams; (20) main interfluvial
still advancing towards the west, filling the eastern sectorof the lagoon.
The present dynamics of the deltaic fan geoforms revealssectors with noticeable accumulation levels, evident also bythe buildup of sediments and detritus over the mangroveforest surface, including the base trees. There are also R.mangle dominated islands trapped by large accumulativevolumes that determined changes in habitat, upstreampenetration of R. mangle trees along distributary channelsenlarged by local tides, R. mangle seedlings to the front ofrecent accretion, and other mangrove distribution patternsoriginated by the variations in the structure and functioningof microrelief dynamics.
areas in the Pacific coast of Mexico. Tones and symbols are according toon depth: Emerged relief types: Hills consolidated during the upper Plioceneconsolidated during the upper Pleistocene to Holocene: (2) fluvial erosive–t (H=5–10 m); (4) fluvial accumulative, low, flood–plain in transition withltaic fan distributors (Hb2–3 m); Fluvial: (6) fluvial accumulative, banksative and abrasive–accumulative, constituted by low littoral beach–ridgesctors (Hb2–2.5 m). Swamp: (8) swamp accumulative, very low, muddy,tion channels, lateral microbars, tidal microdepressions) (Hb0.5–1.0 m); (9)nels and laminar accumulation sectors (Hb2 m); (10) swamp accumulative,coastal marine facies, with remains of ancient littoral beach–ridge networksc: (11) accumulative, remodelled and transformed by human action (Hb3–lative, stepped and inclined, with remains of quaternary sea terraces,sectors in inner lagoon structures (H=−2 to −3 m); (14) accumulative, of) beach strips (sun strip, berm and embryonic dune strips); (16) flood–plainother water bodies of fluvial or fluvial–marine origin, undifferentiated; (18)divisions; (21) microrelief of fluvial distributaries in the deltaic fan.
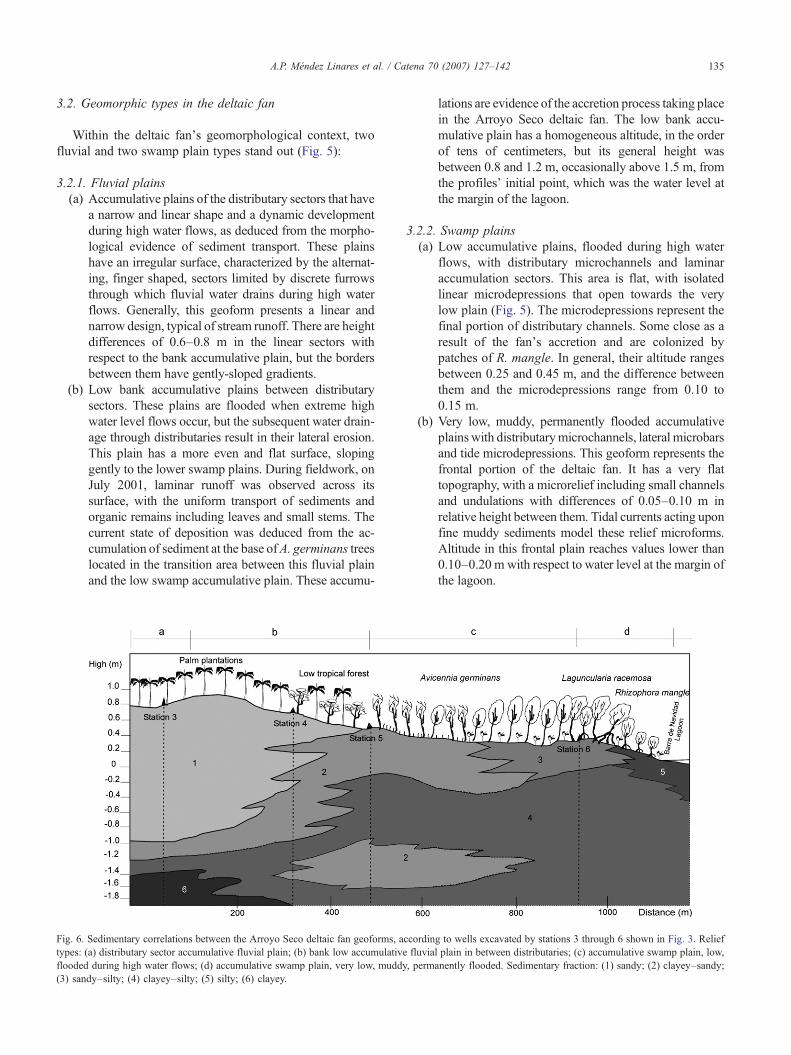
135A.P. Méndez Linares et al. / Catena 70 (2007) 127–142
3.2. Geomorphic types in the deltaic fan
Within the deltaic fan's geomorphological context, twofluvial and two swamp plain types stand out (Fig. 5):
3.2.1. Fluvial plains(a) Accumulative plains of the distributary sectors that have
a narrow and linear shape and a dynamic developmentduring high water flows, as deduced from the morpho-logical evidence of sediment transport. These plainshave an irregular surface, characterized by the alternat-ing, finger shaped, sectors limited by discrete furrowsthrough which fluvial water drains during high waterflows. Generally, this geoform presents a linear andnarrow design, typical of stream runoff. There are heightdifferences of 0.6–0.8 m in the linear sectors withrespect to the bank accumulative plain, but the bordersbetween them have gently-sloped gradients.
(b) Low bank accumulative plains between distributarysectors. These plains are flooded when extreme highwater level flows occur, but the subsequent water drain-age through distributaries result in their lateral erosion.This plain has a more even and flat surface, slopinggently to the lower swamp plains. During fieldwork, onJuly 2001, laminar runoff was observed across itssurface, with the uniform transport of sediments andorganic remains including leaves and small stems. Thecurrent state of deposition was deduced from the ac-cumulation of sediment at the base ofA. germinans treeslocated in the transition area between this fluvial plainand the low swamp accumulative plain. These accumu-
Fig. 6. Sedimentary correlations between the Arroyo Seco deltaic fan geoforms, atypes: (a) distributary sector accumulative fluvial plain; (b) bank low accumulativeflooded during high water flows; (d) accumulative swamp plain, very low, muddy(3) sandy–silty; (4) clayey–silty; (5) silty; (6) clayey.
lations are evidence of the accretion process taking placein the Arroyo Seco deltaic fan. The low bank accu-mulative plain has a homogeneous altitude, in the orderof tens of centimeters, but its general height wasbetween 0.8 and 1.2 m, occasionally above 1.5 m, fromthe profiles' initial point, which was the water level atthe margin of the lagoon.
3.2.2. Swamp plains(a) Low accumulative plains, flooded during high water
flows, with distributary microchannels and laminaraccumulation sectors. This area is flat, with isolatedlinear microdepressions that open towards the verylow plain (Fig. 5). The microdepressions represent thefinal portion of distributary channels. Some close as aresult of the fan's accretion and are colonized bypatches of R. mangle. In general, their altitude rangesbetween 0.25 and 0.45 m, and the difference betweenthem and the microdepressions range from 0.10 to0.15 m.
(b) Very low, muddy, permanently flooded accumulativeplainswith distributarymicrochannels, lateral microbarsand tide microdepressions. This geoform represents thefrontal portion of the deltaic fan. It has a very flattopography, with a microrelief including small channelsand undulations with differences of 0.05–0.10 m inrelative height between them. Tidal currents acting uponfine muddy sediments model these relief microforms.Altitude in this frontal plain reaches values lower than0.10–0.20 m with respect to water level at the margin ofthe lagoon.
ccording to wells excavated by stations 3 through 6 shown in Fig. 3. Relieffluvial plain in between distributaries; (c) accumulative swamp plain, low,, permanently flooded. Sedimentary fraction: (1) sandy; (2) clayey–sandy;

136 A.P. Méndez Linares et al. / Catena 70 (2007) 127–142
3.3. Accretion processes in the deltaic fan
The Arroyo Seco deltaic fan has a transitional nature in itsvarious sedimentary components, both horizontally andvertically. Sandy horizons are appreciated in the accumula-tive fluvial plains, alternating with a clayey–sandy layertowards the lowest swamp plains. This clayey–sandyhorizon lies below the sandy horizon in accumulative fluvialplains, advancing to the fan's front. At the lowest swampplain, the clayey–sandy horizon alternates with a sandy–silty one, mostly at the surface of the very low swamp plain.This marked accumulation of the sandy fraction, bothhomogeneous and mixed with the clay and loam fractionsat the fan's lower portions results from the recent advancingaccretion process, reflected in the distribution of the coarsersediments upon the finer horizons (Fig. 6).
3.4. Soil types
On higher coastal ground, over granitic substrate, soils arealbic and haplic luvisols developed over granitic complexes.There are poorly differentiated horizons, 10 to 30 cm deep,with gravelly to sandy texture, with blocks of angular orround structure, with no carbonates and organic matter.These soils reflect an intense physical intemperism and resultfrom the destruction and accumulation in situ.
Soils in the fluvial plains are eutric fluvisols, and a 1.75 mhorizon could be appreciated in the profile near the upland limitof this geoform, with no structure and no carbonates, whichreflected aluvial deposition conditions. In the three subjacenthorizons, 20 to 40 cmwide, texture was clayey–silty, structurelaminar and massive, and the horizons were humid andcompact. This is characteristic of the actual surface in the frontof the aluvial fan, and therefore indicates its previous location.
Soils in the swamp plains, distributed in low bankaccumulative plains located between distributary sectors,were characterized as umbric fluvial. Superficial horizons are20–30 cm deep, and texture was clayey–sandy structured inblocks. Lower horizons had a clayey–limey texture,unstructured, and were humid and mildly carbonatedreflecting a lagoon habitat in a very fine sedimentary facie,which, along with traces of embedded stem and root wood,indicated the former mangrove front. The contact betweenfluvial and lagoon habitats was found at station 5 (Fig. 3),where the amount of carbonates increased with the humidityof the horizons. There were also seashells in these horizons,showing the “paleofront” of the mangrove community.Finally, lagoon habitat soils were Gleysols, characterizedby deep horizons (greater than 2.8 m), with a clayey tosandy–limey texture, without structure, very humid, with ahigh content of organic matter (humus and peat) and mildlycarbonated, all properties presently found inmangrove fronts.
South of the aluvial fan and on the very low swamp plain(Station 7, Fig. 3), the superficial horizons were 0.1–0.3 mwide and of sandy texture. The underlying horizon was 0.3–1 m wide and texture was sandy–clayey, which changed to
clayey–limey on the lower horizon. From these deephorizons it can be inferred that the sandy sedimentstransported during extreme high water runoff from the actualArroyo Seco River buried the low, permanently flooded,lagoon plain of this site.
3.5. Water table depth
Maximum depth of the water table in transect I–I′ reached−0.81 m in the higher part of the low cumulative swampplain (Fig. 7a) and the level increased to −0.5 m at thecontact with the mangrove front. This pattern is repeated intransect II–II′ (not shown), which was traced on the samedirection and over the same two geoform types. In thistransect, the water table was found at −1.71 m at the highpart of the fluvial plain and it increased to the surface level atthe lower swamp plain.
Transect III–III′, traced on the low temporally floodedcumulative swamp plain, was parallel to the mangrove front.The water table varied from −1.27 to −0.23 m, depending onrelief. There were sand mounds where the water table wasdeeper, a consequence of the sediments transported duringhigh energy flow events by the Arroyo Seco. After this zone,the water table was somewhat similar along the swampwhere there was less lateral sand accretion.
3.6. Salinity
Salinity in the water table depended not only on seawaterbut on freshwater infiltration and evapotranspiration. At theinland initial point of the I–I′ transect (Fig. 7b) the salinitywas greater than that of seawater (78 vs. 36‰). In the swampplains, salinity decreased in a downslope direction from 64.5to 29‰. This pattern is similar for transect II–II′ (Fig. 7f):19.6‰ in the low bank accumulative fluvial plains, 45.5 to91.6‰ in the low accumulative swamp plains, and 40 to15.2‰ in the very low, muddy, permanently floodedaccumulative swamp plains. In transect III–III′, salinity isvery low (0.7‰) near the Arroyo Seco river, but it increasesdramatically from 50 to 71‰ into the low accumulativeswamp plain. Salinity is greater in where there is slowinfiltration of rain or tidal water and high evapotranspiration.
3.7. Physiognomy and structure of mangrove communities
3.7.1. Basal areaThere is a marked zonation among species and along
transects I–I′ and II–II′ (Fig. 7.d–h). Over the first 80 malong transect I–I′, on the cumulative swamp plain, the basalarea of Avicennia is around at 0.024 m2. This value increasesto 0.074 m2 on the 120–240 m transect segment and peaks at0.18 m2 on the next 280 to 360 m interval. Finally, in thepermanently flooded swamp accumulative plain, the basalarea of Avicennia decreases to 0.003 m2 and the maximumdimensions for Rhizophora and Laguncularia (0.22 and0.09 m2, respectively) are found.

Fig. 7. Soil surface relief and water table depth (a, e), water table salinity (b, f), mangrove tree community cover (c, g: hatched bars, dead tree cover; filled bars,living tree cover) and basal area (d, h) along transects I–I′ (a–d) and II–II′ (e–h). See Fig. 3 for transect location.
137A.P. Méndez Linares et al. / Catena 70 (2007) 127–142
There was a low tropical forest with Pithecellobium dulce,Senna racemosa and Mimosa sp in the highest section oftransect II–II (Fig. 7h) with BA between 0.007 and 0.012m2.Batis maritima is along the first 40 m of the transect (100%cover, 0.7 m height). It is interspersed with patches of Avi-cennia where BA is between 0.1 and 0.007 m2 in a 120 msegment. The minimum values correspond to two distributivebeds within the plain. BA distribution of Avicennia is bi-modal with peak values (0.16–0.17 m2) at 240 m and 520 malong the transect. Further along the transect, the BA ofAvicennia decreases to 0.02 m2 while that of Lagunculariaand Rhizophora is as high as 0.096 m2 and 0.114 m2,respectively, in the flooded swamp plain. The basal area ofRhizophora is consistently higher than that of Laguncularia(0.15 m2 vs. 0.077 m2, but variability around the average isvery high. Rhizophora is the sole species in the final part ofthe transect (720 m), with a basal area of 0.132 m2.
Transect III–III′, perpendicular to the other transects,began at the margin of the Arroyo Seco, starting at aninduced grassland. The intrusion of sand from the river
created mounds in the 40 to 80 m marks along the transectthat buried the trunk bases of Laguncularia and Rhizophoratrees with 0.11 and 0.16 m2 BA, respectively, causing thedeath of branches and the leaf cover. As surface sanddecreases along this plain, at 120 m, there is a depressionindicating a former distributive bed where the BA of Avi-cennia (0.02 m2) is lower than that of Laguncularia andRhizophora (both with 0.11 m2). The BA of Avicenniaincreases again along the transect up to 0.15 m2, while that ofLaguncularia decreases until it is not and is not found in therest of the transect. There is a bimodal distribution of BA ofAvicennia on the 200 to 400 m transect segment whosevalley corresponds with a zone of high mortality.
3.7.2. HeightChanges in tree height are similar to those of BA. In the
high part of the low swamp accumulative plain temporallyflooded, at the first 80 m of transect I–I′ Avicennia heightreaches up to 4.5 m and increases up to 7 m in the 120–240 minterval and up to 11 m in the 280–360 m interval. The height

Fig. 8. Tree height as a function of the depth of the water table level depth.Height tends to increase as the water table reaches the soil surface. Circles,Avicennia germinans; triangles, Laguncularia racemosa; squares, Rhizo-phora mangle. The linear regression line and statistics includes only A.germinans.
138 A.P. Méndez Linares et al. / Catena 70 (2007) 127–142
of this species decreases again to 6.4 m when in contact withthe very low swamp accumulative plain, permanently flooded(400 m). Rhizophora and Laguncularia reach up to 11 m and9.5 m respectively near the lagoon margin (400 m).
The tropical lowland forest trees reached heights between2.5 and 3.4 m at the beginning of transect II–II′. At the 80 mmark, within the transect, the average height of Avicennia is7.2 m, but decreases to 1.5–2.8 m in the 120–200 m interval.There is a bimodal distribution in height of Avicennia withpeaks at the 240 (6.30 m) and 520 m (9.30 m) transect marks.The height of Avicennia decreased to 3.40 m as the transectpasses over the low swamp where Laguncularia and Rhi-zophora reached heights of 8 to 9 m. At the 40–80 mmark of
Fig. 9. Canonical Correspondence Analysis triplot with species, sites (1–31) and enMexico. Eigenvalues: Axis 1=0.898; Axis 2=0.064; Species–environment correlaccounted by axes 1 and 2, respectively; finally, 93.3 and 6.7% of variance in the sanalysis considers geomorphic, edaphic and water table properties, and basal aremangle). Four zones are clearly defined: the monospecific Avicennia forests (A)(ALR), the mixed forest composed of Laguncularia and Rhizophora (LR), and th
transect III–III′, the sector codominated by Lagunculariaand Rhizophora has maximum average heights of 11.5 m. Atthe 240–400 m sector, there is a bimodal pattern in Avicen-nia height with peaks at 8 m.
3.7.3. Geomorphological and physico–geographical zona-tion of mangrove species associations
The correlation of mangrove associations and othervegetation types with geoforms led to the following zonation:a first zone of alluvial plain distributor sectors in the accu-mulative fluvial plain, with palm plantations; a second zone inthe bank accumulative fluvial plain, with low tropical forestand patches of palm plantations; a third zone, in the lowaccumulative swamp plain with A. germinans mangal for-mations, which towards the lowest swamp plain wereassociated with L. racemosa and R. mangle, constituting arather narrow fourth zone; a fifth zone in the very lowaccumulative swamp plain, where the L. racemosa–R. mangleassociation developed; and finally a sixth narrow zone with R.mangle located at the fan's frontal edge.
3.7.4. Plant coverIn transect I–I′, dead Avicennia tree cover (Fig. 7c,
hatched bars) was greater at the interval between 0 and240 m and there was practically no live trees. Starting at280 m, dead tree cover decreased abruptly and there was a 70to 90% cover of living Avicennia trees (Fig. 7c, filled bars).In transect II–II′, the greatest cover of dead Avicennia trees(Fig. 7g, hatched bars) was at the central part (240–440 m)with a maximum of 60%, and the cover of live trees (Fig. 7g,
vironmental variables of mangrove forests from the Arroyo Seco deltaic fan,ation: Axis 1=0.969; Axis 2=0.719; 83% and 5.9% of variance in speciespecies–environmental relation accounted by axes 1 and 2, respectively. Thisa and height of mangrove tree species (A. germinans, L. racemosa and R., the mixed forests composed of Avicennia, Laguncularia and Rhizophorae monospecific Rhizophora fringe (R).

139A.P. Méndez Linares et al. / Catena 70 (2007) 127–142
filled bars) was highest towards the final part of the transectwith values up to 90%. Finally, in transect III–III′ (notshown), live tree cover was between 30% and 50%, and thatof dead trees was around 15%, but total cover is not higherthan 60%.
3.7.5. Seedling/sapling densityIn transect I–I′, Avicennia seedling density was between
3.4–14.5/m2 in the sector between 280 to 400 m, whereheight and basal area and cover of the healthy adults wasgreater. Starting along the transect at 440 m and until 520 m,seedling of Laguncularia and Rhizophora were registered,but with low density (0.03–0.10 m−2). In the 320 to 480 msector of transect II–II′, there were up to 32 Avicennia seed-lings/m2, with heights up to 13 cm. Seedlings/saplings ofLaguncularia and Rhizophora which were 0.5 to 1.2 m tallappeared at the 600 m mark, but their density was relativelylow (1–2.2/m2). At the Rhizophora margin, seedling/sapling(0.7 to 1 m tall) density was 1.1/m2.. Finally, in transect III–III′, there were 0.4 and 0.8 Avicennia seedlings/saplings/m2
with heights between 0.08 and 0.4 m., and no Lagunculariaor Rhizophora seedlings/saplings were found.
3.7.6. Tree height and water table salinity and depthThere was no significant correlation between tree height
and water table salinity (r=0.31, P=0.09); however, thelinear regression between tree height and water table depthwas significant (r=0.74, Pb0.001, Fig. 8), which suggeststhat most of the individuals that grow under conditions ofdeeper water table are deteriorated or dead.
3.7.7. Tree cover and water table salinity and depthThere was no significant correlation between the cover of
living trees and water table salinity (r=0.32, P=0.91, butthis correlation was significant when considering dead treecover (r=0.54, P=0.003, indicating that tree mortality isassociated with higher salinity. On the other hand, tree coverwas significantly correlated to water table depth (r=0.49,P=0.008), indicating greater cover as the water table reachesthe soil surface. This correlation was not significant whenusing the cover of dead trees (r=0.098, P=0.61). However,the cover dead trees increase when water table was deeperthan 40 cm.
3.7.8. Seedling/sapling density and cover vs. water tablesalinity and depth
There was a significant correlation between saplingdensity and water table depth (P=0.029) and data areconcentrated at relatively superficial depths (0 to 50 cm).Sapling height seemed to increase as water table approachesthe soil surface, but this trend was not statistically significant(F=0.13; P=2.5).
3.7.9. Multivariate analysisThe CCA ordination triplot for sampling sites, environ-
mental variables and species is shown in Fig. 9. The first axis
accounts for 83% of data variability and the second for 7% ofthe species data set. Four distinctive mangrove forests can beclearly distinguished from left to right (Fig. 9): themonospecific Avicennia mangals, mixed Rhizophora–La-guncularia–Avicennia forests, mixed Rhizophora–Lagun-cularia forests, and the Rhizophora fringe forest at lagoonmargin. There is a strong correlation between environmentalfactors and mangrove community distribution, since the thefirst axis accounts for 93% of data variability and the secondfor the remaining 7%. As arrows in Fig. 9 indicate, whencompared to the other mangrove forests, the monospecificAvicennia mangal is correlated with greater water tablesalinities, higher soil microrelief and temperature, and lowersoil organic content. On the other hand, the remainingmangrove forests are related to a more superficial water tablelevel, finer soil texture, and higher carbonate content. Ascoded in the CCA, the geoforms occupied by the Avicenniamangals have the highest code values, corresponding to thetransitional, frontal and very low swamp plains in contrast tothe fluvial accumulative plain and the low swamp accumu-lative plain, which are occupied by the other three mangrovecommunities. The results from Fig. 9 are characteristic ofstrong environmental gradients with a marked zonation (e.g.Lachance and Lavoie, 2004), as described in Fig. 7.
4. Discussion
From a geomorphologic viewpoint the morphogenetictypes are determined by sediment transport along the deltaicfan, predominantly sand sediments in fluvial geoforms andclay–lime sediments in swamp geoforms. There are cleartransitory stratifications that interdigitate at contact zonesbetween geoforms as can be deduced by changes ingranulometry that result from the selective transport anddeposition processes characteristic of deltaic zones. Thedeltaic fan of Arroyo Seco shows, both horizontally andvertically, differential particle size from coarser to finersediments. This accretion is currently active and has con-sequences on mangal distribution and mortality: at the lowaccumulative swamp plain, there is a high proportion of deadtrees since sediment accretion increased the distance be-tween roots and the water table, especially during the dryperiod. Also, sediment accretion can be observed in thetransition zones between mangal associations and in thecolonization of the lagoon fringe. Field data indicated thatthis process of horizontal succession could also be deducedfrom stratigraphy as a “vertical succession”.
There are evidences of the actual accretion processin the advance of the Rhizophora fringe, the overlap ofhorizons with soils having a sandy texture over soils domi-nated by clays with mangrove wood debris, and of sandysediments that bury Avicennia pneumatophores. Theseresults support the hypothesis that in deltaic zones man-grove zonation is related with the geomorphic mosaic andits spatio-temporal dynamics (Thom, 1967, 1984; Thomet al., 1975).

140 A.P. Méndez Linares et al. / Catena 70 (2007) 127–142
As in other areas (Clarke andHannon, 1970; López-Portilloand Ezcurra, 1989; López-Portillo et al., 1989), there is agradient that depends on microrelief, soil texture and waterlevel (in our case, the lowest water level during the dry seasonas indicated by the depth of the water table) that is reflected inthe composition and structure of the mangrove communities.This gradient is clear in the CCA triplot shown in Fig. 9.
The geomorphic viewpoint allows us to describe zonationand the acknowledgement of its genesis allows us to predictits future evolution. In this zone, as in others along thesoutheast Pacific coast of Mexico, there are short and intenseperiods of rainfall that result in very erosive fluvial outflows(Vidal-Zepeda, 1990a). Great volumes of sediments modifythe microrelief of the zone (Hernández-Santana, unpubl.data). The differential accumulation of upland, lagoon andmarine of sediments changes the patterns of superficial flowand soil texture, which impacts the degree of continental andtidal water influence (Semeniuk, 1980). In our study site,there are evidences that, as a consequence of the accretion,each mangal association has a “front of advance” indicatedby seedling and sapling presence, and a back zone of mor-tality as patches of dead trees are left behind.
Water level has been traditionally mentioned as a factorinfluencing the physiognomy and structure of mangrovecommunities (Chapman, 1976; Tomlinson, 1986; Ball,1996). However, the water table can be very low especiallyin the dry season and therefore decrease water availability toroot systems. The fact that the water table in our study site isnot horizontal along the transects, which were made parallelto the slope, suggests a strong resistance to subsurface flowprobably due to the fine texture from the sediments as thelagoon margin is approached. Since dead trees of Avicenniaare found in the areas where there is a deep water table, suchdeath may be a consequence of a double stress caused bysubstrate salinity and drought. Both types of stress areconsidered as similar from the physiological viewpoint (Lugoet al., 1981; Tomlinson, 1986; Ball, 1996). It can be predictedthat salinity concentration would be less limiting for mangaldevelopment if soils were saturated with water.
This geomorphologic viewpoint allows us to explain azonation that would be misleading under the usual clas-sification by Lugo and Snedaker (1974). According to it, thesequence of associations at the Arroyo Seco deltaic fan wouldcorrespond to a basin and a fringe mangal type, but thephysiognomy we observed is much more complex. It isconvenient to define mangrove association patterns as afunction of the dynamics within their habitats, consideringthat they frequently develop in areas where there is a highinterchange of continental–marine energy, and where geo-morphic processes play an important role (Woodroffe, 1992;Ellison et al., 2000).
5. Conclusions
(1) The Arroyo Seco hydrographic basin, a source of runoffsupply to the deltaic fan, is located under geological–
geomorphological and climatic–hydrological condi-tions that favor intense physical weathering and erosionprocesses that facilitate sediment transport and deposi-tion, contributing to continued superficial accretion.
(2) This alluvial fan's geomorphic system is divided intotwo subsystems: fluvial geoforms and swamp geo-forms, which determine the vegetation successionalstages. The system contemplates two types of fluvialplains plus two types of swamp plains, located in stepsfrom the hinterland towards the front of the deltaic fan:bank and distributary accumulative fluvial plains, andlow and very low swamp accumulative plains.
(3) Sediment analyses, both horizontal and vertical,revealed a transitional character of the sedimentarycomponents in the deltaic fan that corresponded toaccumulative geoforms whose structure and composi-tion reflect an active accretion process. To this respect,the R. mangle or L. racemosa remains found in deephorizons of edaphic wells over low tropical forest andA. germinans communities evince the ancient fanfront, buried by accretion. The lower portion of thedeltaic fan's general profile, shows, under the clayey–sandy horizon, an alternating transit of fine fractions,including mud and clay, and a clayey–muddy transi-tion between both horizons, a typical granulometricsequence for an actively growing deltaic fan.
(4) The zonation of mangrove communities a result of thecontinued sedimentary accretion process in the fan.Additionally, high water flows occur in the areaand the transport of enormous amounts of sedi-ments modify the microrelief and influence superfi-cial and intertidal runoff, giving rise to the changesin sediment texture and to water table depth anddistribution.
(5) Water table depth seems to be an important factor inthe physiognomic-structural development of mangals.In the areas where the water table is deep (N40 cm)mangrove communities have a high proportion of deadtrees or poorly developed individuals. This suggeststhat even if salinity is high (up to 95‰ in the dryseason), mangroves can develop if soils are water-saturated.
(6) There is a clear geomorphic zonation pattern ofmangrovecommunities in the deltaic fan, with six speciesassociation zones. Since the succession of these commu-nities is a consequence of the fan's accretion dynamics,the changes in these ecosystems are predictable andtherefore able to be managed in an adequate manner.
Acknowledgements
The authors appreciate the assistance of A. Ramírez, C.Enríquez, M. Vilchis, M.P. Salazar and J.Valdiri duringfieldwork and I. Trejo-Vázquez and G. Vázquez in the CCAanalysis. Two anonymous reviewers provided constructivecriticism and suggestions.

141A.P. Méndez Linares et al. / Catena 70 (2007) 127–142
References
Ball, M.C., 1996. Comparative ecophysiology of mangrove forest andtropical lowland moist rainforest. In: Mulkey, S.S., Chazdon, R.L.,Smith, A.P. (Eds.), Tropical Forest Plant Ecophysiology. Chapman &Hall, New York, pp. 461–496.
Ball, M.C., 1998. Mangrove species richness in relation to salinity andwaterlogging: a case study along the Adelaide River floodplain, NorthernAustralia. Global Ecology and Biogeography Letters 7, 73–82.
Blasco, F., Saenger, P., Janodet, E., 1996. Mangroves as indicators of coastalchange. Catena 27, 167–178.
Chapman, V.J., 1976. Mangrove Vegetation. J. Cramer, Vaduz.Clarke, L.D., Hannon, H.J., 1970. The mangrove swamp and salt marsh
communities of the Sidney district III. Plant growth in relation to salinityand waterlogging. Journal of Ecology 58, 351–369.
Coblentz, D.D., Ritters, K.H., 2004. Topographic controls on the regional-scale biodiversity of the south-western USA. Journal of Biogeography31, 1125–1138.
Curray, J.R., Moore, D.G., 1964. Pleistocene deltaic progradation of con-tinental terrace, Costa de Nayarit, Mexico. In: Van Andel, T.H., Shor Jr.,G.G. (Eds.), Marine Geology of the Gulf of California. American Assoc.Petrol. Geol., Tulsa, Oklahoma, pp. 193–215.
Curray, J.R., Emmel, F.J., Crampton, J.S., 1969. Holocene history of a strandplain, lagoonal coast, Nayarit,Mexico. In: Ayala-Castañares, A., Phleger,F.B. (Eds.), Coastal Lagoons, a Symposium. UNAM, Mexico City,pp. 63–100.
Dawes, C., 1986. Botánica Marina. LIMUSA, Mexico City.Díaz Díaz, J.L., 1986. Morfoestructuras de Cuba Occidental y su geodinámica
reciente. Ph.D. Thesis, Geography Institute, Moscow, Russia, 232 pp.Duke, N.C., Ball, M.C., Ellison, J.C., 1998. Factors influencing biodiversity
and distributional gradients in mangrove. Global Ecology andBiogeography Letters 7, 27–47.
Ellison, A.M., Mukherjee, B.B., Karim, A., 2000. Testing patterns ofzonation in mangroves: scale dependence and environmental correlatesin the Sundarbans of Bangladesh. Journal of Ecology 88, 813–824.
Ewel, K.C., Twilley, R., Ong, J.E., 1998a. Different kinds mangrove forestprovides different goods and services. Global Ecology and Biogeogra-phy Letters 7, 83–94.
Ewel, K.C., Bouergeois, J.A., Cole, T.G., Zheng, S., 1998b. Variation inenvironmental characteristics and vegetation in high-rainfall mangroveforest, Kosrae, Micronesia. Global Ecology and Biogeography Letters 7,49–56.
FAO-UNESCO, 1990. Mapa mundial de suelos a escala 1: 50 000 000 einforme sobre recursos mundiales de suelos. Organización de lasNaciones Unidas para la Agricultura y la Alimentación, Roma.
Gallegos, M., 1986. Petróleo y manglar. Centro de Ecodesarrollo, MexicoCity. 101 pp.
García, E. Vidal-Zepeda, R., 1990. Temperaturas extremas. In: García deFuentes, A. (Ed.), Atlas Nacional de México. Instituto de Geografía,UNAM, México. Vol. III. p. IV.4.5.
Hernández-Santana, J.R., Reyes-González, R., 2002. Regeneración artificialde playas y desarrollo sostenible: experiencias ambientales en la Playade Varadero, Península de Hicacos, Cuba. Investigaciones Geográficas49, 43–56.
Hernández-Santana, J.R., Ortiz-Pérez, M.A., Zamorano-Orozco, J.J., 1996.Regionalización morfoestructural de la Sierra Madre del Sur, México.Investigaciones Geográficas 31, 45–67.
Hernández-Santana, J.R., Seguinot-Barbosa, J., Reyes-González, R., 2002.Islas de Puerto Rico y Culebra: regularidades geomorfológicas de surelieve costero y submarino. Investigaciones Geográficas 47, 7–19.
Hill, M.O., 1979. TWINSPAN — a FORTRAN Program for ArrangingMultivariate Data in an Ordered Two-way Table by Classification of theIndividuals and Attributes. Section of Ecology and Systematics, CornellUniversity, Ithaca.
Jiménez, R.A. Maderey-Rascón, L.E., 1990. Escurrimiento medio anual. In:García de Fuentes, A. (Ed.), Atlas Nacional de México. Instituto deGeografía, UNAM, México. V. 3, p.IV.6.4.
Lachance, D., Lavoie, C., 2004. Vegetation of Sphagnum bogs in highlydisturbed landscapes: relative influence of abiotic and anthropogenicfactors. Applied Vegetation Science 7, 183–192.
Lepš, J., Šmilauer, P., 2003. Multivariate Analysis of Ecological Data usingCANOCO. Cambridge University Press, U.K. 269 pp.
López-Portillo, J., Ezcurra, E., 1989. Response of three mangroves tosalinity in two geoforms. Functional Ecology 3, 355–361.
López-Portillo, J., Ezcurra, E., Maass, J.M., 1989. Los petenes de SianKa'an, Quintana Roo y su relación con gradientes de presión hídrica.Acta Botánica Mexicana 5, 19–29.
López-Ramos, E., 1983. Geología de México. Published by the author.Vol. 3, 3rd ed. Mexico City.
Lugo, A.E., Snedaker, S.C., 1974. The ecology of mangroves. AnnualReview of Ecology and Systematics 5, 39–64.
Lugo, A.E., Cintrón, G., Goenaga, C., 1981. Mangrove ecosystems understress. In: Barrett, G.W., Rosenberg, R. (Eds.), Stress Effects on NaturalEcosystems. John Wiley & Sons, Ltd., Chichester, pp. 129–153.
Lugo Hubp, J., 1990. El Relieve de la República Mexicana. Revista delInstituto de Geología 9, 82–111.
Maderey-Rascón, L.E., 1990. Volumen medio anual de sedimentos ensuspensión. In: García de Fuentes,A. (Ed.) Atlas Nacional de México,Sección Naturaleza, Instituto de Geografía, UNAM, México. vol. III.pp. IV.6.4.
Matteucci, K.L., Colma, A., 1982. Metodología para el estudio de lavegetación. Secretaría General de la Organización de los EstadosAmericanos, Washington, D. C. 168 pp.
Müller, C., Berger, G., Glemnitz, M., 2004. Quantifying geomorphologicalheterogeneity to assess species diversity of set-aside arable land.Agricultural Ecosystem & Environment 104, 587–594.
Nichols, W.F., Killingbeck, K.T., August, P.V., 1998. The influence ofgeomorphology heterogeneity on biodiversity. A landscape perspective.Conservation Biology 2, 371–379.
O'Neill, R.V., Krummel, J.R., Gardner, R.H., Sugihara, G., Jackson, B.,DeAngelis, D.L., Milne, B.T., Turner, M.G., Zygmunt, B., Christensen, S.,Dale, V.H., Graham, R.L., 1988. Indices of landscape pattern. LandscapeEcology 1, 153–162.
Pannier, F., 1992. El ecosistema de manglar como indicador de cambiosglobales en la zona costera tropical. Ciencia 43, 111–113.
Priego-Santander, A.G., 2004. Relación entre la heterogeneidad geoecoló-gica y la biodiversidad en ecosistemas costeros tropicales. Ph D Thesis,Instituto de Ecología, A. C., Xalapa, Veracruz, 117 pp.
Rice, R.J., 1977. Fundamentals of Geomorphology. Longman, New York.Rico-Gray, V., Palacios-Ríos, M., 1996. Salinidad y nivel del agua como
factores en la distribución de la vegetación en la ciénaga del NW deCampeche, México. Acta Botánica Mexicana 34, 53–61.
Rzedowski, J., 1978. Vegetación de México. LIMUSA, Mexico City.Semeniuk, V., 1980. Mangrove zonation along an eroding coastline in King
Sound, North-Western Australia. Journal of Ecology 68, 789–812.Semeniuk, C., Semeniuk, V., 1995. A geomorphic approach to global
classification for inland wetlands. Vegetatio 118, 103–124.Smith III, T.J., 1992. Forest structure. In: Robertson, A.I., Alongi, D.M.
(Eds.), Tropical Mangrove Ecosystems. Coastal and Estuarine Studies,vol. 41. American Geophysical Union, Washington, pp. 101–136.
Spiridonov, A.I., 1975. Geomorfologuicheskoe kartografiravanie. Nedra,Moscow (in Russian), 182 pp.
Tarbuck, E.J., Lutgens, F.K., 1999. Earth. An Introduction to PhysicalGeology, 6th ed. Prentice Hall, Madrid.
Ter Braak, C.J.F., 1998. Program CANOCO, Version 4.0. Centre forBiometry Wageningen, The Netherlands.
Thom, B.G., 1967. Mangrove ecology and deltaic geomorphology: Tabasco,Mexico. Journal of Ecology 55, 301–343.
Thom, B.G., 1984. Coastal landforms and geomorphic processes. In:Snedaker, S.C., Snedaker, J.C. (Eds.), The Mangrove Ecosystems:Research Methods. UNESCO, U. K., pp. 3–17.
Thom, B.G., Wright, L.D, Coleman, J.M., 1975. Mangrove ecology anddeltaic–estuarine geomorphology — Cambridge Gulf–Ord RiverWestern-Australia. Journal of Ecology 63 (1), 203–232.

142 A.P. Méndez Linares et al. / Catena 70 (2007) 127–142
Tomlinson, P.B., 1986. The Botany of Mangroves. Cambridge UniversityPress, Great Britain.
Trejo-Torres, J., Durán, R., Olmsted, I., 1993. Manglares de la Penínsulade Yucatán. In: Salazar-Vallejo, S.I., González, N.E. (Eds.), Biodiver-sidad Marina y Costera de México. CONABIO-CIQRO, Mexico City,pp. 661–672.
Valdéz, H.J., 1991. Estructura fisonómica del bosque de mangles de lalaguna de Agua Brava, Nayarit. Ph.D. Thesis, Chapingo, Mexico.
Vidal-Zepeda, R., 1990a. Precipitación. In: García de Fuentes, A. (Ed.),Atlas Nacional de México. SIGSA, Mexico City, pp. IV.4.6.
Vidal-Zepeda, R., 1990b. Precipitación máxima en 24 horas. In: García deFuentes,A. (Ed.), Atlas Nacional de México, Ed. SIGSA , Mexico City,pp. IV.4.5.
Woodroffe, C., 1992. Mangrove sediments and geomorphology. In:Robertson, A.I., Alongi, D.M. (Eds.), Tropical Mangrove Ecosystems.Coastal and Estuarine Studies, vol. 41. American Geophysical Union,Washington, pp. 7–41.
Young, B.M., Harvey, L.E., 1996. A spatial analysis of the relationshipbetween mangrove (Avicennia marina var. australasica) physiognomyand sediment accretion in the Hauraki Plains, New Zealand. Estuarine,Coastal and Shelf Science 42, 231–246.




















