
The Limits and Intensity of Plasmodium falciparum Transmission: Implications for Malaria Control and...
25
Table S1. National level estimates of the area occupied by and number of people at risk of Plasmodium falciparum malaria transmission in 2007. The data are presented for World Health Organization (WHO) region: AFRO, African Regional Office of the WHO, AMRO, American Regional Office of the WHO, EMRO, Eastern Mediterranean Regional Office of the WHO, EURO, European Regional Office of the WHO, SEARO, South East Asian Regional Office of the WHO and WPRO, Western Pacific Regional Office of the WHO. Areas are in millions km 2 and populations at risk in millions. The populations are further stratified as those at stable (P. falciparum annual parasite incidence (PfAPI) of more than 0.1 per 1000 population per annum (pa)) and unstable (PfAPI < 0.1‰ pa). Region/Country Area at risk (km 2 ) Population at risk (millions) Stable Unstable All Stable Unstable All AFRO Angola 1.26 0.02 1.28 15.41 0.28 15.70 Benin 0.12 0.00 0.12 7.66 0.00 7.66 Botswana 0.43 0.00 0.43 0.89 0.00 0.89 Burkina Faso 0.28 0.00 0.28 14.23 0.00 14.23 Burundi 0.02 0.00 0.02 5.66 0.00 5.66 Cameroon 0.47 0.00 0.47 16.95 0.00 16.95 Cape Verde 0.00 0.00 0.00 0.00 0.25 0.25 Central African Republic 0.63 0.00 0.63 4.15 0.00 4.15 Chad 0.69 0.63 1.32 9.76 0.16 9.92 Comoros 0.00 0.00 0.00 0.59 0.00 0.59 Congo 0.34 0.00 0.34 3.33 0.00 3.33 Côte d'Ivoire 0.33 0.00 0.33 17.80 0.00 17.80 Dem. Republic of the Congo 2.32 0.00 2.32 57.97 0.00 57.97 Equatorial Guinea 0.03 0.00 0.03 0.52 0.00 0.52 Eritrea 0.07 0.06 0.13 3.33 0.95 4.28 Ethiopia 0.93 0.07 0.99 46.08 1.50 47.59 Gabon 0.27 0.00 0.27 1.34 0.00 1.34 Gambia 0.01 0.00 0.01 1.51 0.00 1.51 Ghana 0.24 0.00 0.24 22.21 0.00 22.21 Guinea 0.25 0.00 0.25 9.23 0.00 9.23 Guinea-Bissau 0.03 0.00 0.03 1.43 0.00 1.43 Kenya 0.00 0.03 0.03 25.61 0.18 25.79 Liberia 0.10 0.00 0.10 3.43 0.00 3.43 Madagascar 0.61 0.09 0.70 17.28 0.00 17.28 Malawi 0.12 0.00 0.12 13.45 0.00 13.45 Mali 0.65 0.67 1.32 13.46 0.47 13.93 Mauritania 0.28 0.53 0.81 0.93 0.40 1.33 Mayotte 0.00 0.00 0.00 0.00 0.27 0.27 Mozambique 0.82 0.00 0.82 21.06 0.00 21.06 Namibia 0.35 0.19 0.54 1.25 0.38 1.64 Niger 0.45 0.79 1.25 13.19 0.62 13.81 Nigeria 0.93 0.00 0.93 134.60 0.00 134.60 Rwanda 0.02 0.00 0.02 5.03 0.00 5.03 Sao Tome and Principe 0.00 0.00 0.00 0.13 0.00 0.13 Senegal 0.20 0.00 0.20 10.82 0.00 10.82
Transcript of The Limits and Intensity of Plasmodium falciparum Transmission: Implications for Malaria Control and...

Table S1. National level estimates of the area occupied by and number of people at risk of
Plasmodium falciparum malaria transmission in 2007. The data are presented for World
Health Organization (WHO) region: AFRO, African Regional Office of the WHO, AMRO,
American Regional Office of the WHO, EMRO, Eastern Mediterranean Regional Office of
the WHO, EURO, European Regional Office of the WHO, SEARO, South East Asian
Regional Office of the WHO and WPRO, Western Pacific Regional Office of the WHO. Areas
are in millions km2 and populations at risk in millions. The populations are further stratified as
those at stable (P. falciparum annual parasite incidence (PfAPI) of more than 0.1 per 1000
population per annum (pa)) and unstable (PfAPI < 0.1‰ pa).
Region/Country Area at risk (km2) Population at risk (millions)
Stable Unstable All Stable Unstable All AFRO Angola 1.26 0.02 1.28 15.41 0.28 15.70Benin 0.12 0.00 0.12 7.66 0.00 7.66Botswana 0.43 0.00 0.43 0.89 0.00 0.89Burkina Faso 0.28 0.00 0.28 14.23 0.00 14.23Burundi 0.02 0.00 0.02 5.66 0.00 5.66Cameroon 0.47 0.00 0.47 16.95 0.00 16.95Cape Verde 0.00 0.00 0.00 0.00 0.25 0.25Central African Republic 0.63 0.00 0.63 4.15 0.00 4.15Chad 0.69 0.63 1.32 9.76 0.16 9.92Comoros 0.00 0.00 0.00 0.59 0.00 0.59Congo 0.34 0.00 0.34 3.33 0.00 3.33Côte d'Ivoire 0.33 0.00 0.33 17.80 0.00 17.80Dem. Republic of the Congo 2.32 0.00 2.32 57.97 0.00 57.97Equatorial Guinea 0.03 0.00 0.03 0.52 0.00 0.52Eritrea 0.07 0.06 0.13 3.33 0.95 4.28Ethiopia 0.93 0.07 0.99 46.08 1.50 47.59Gabon 0.27 0.00 0.27 1.34 0.00 1.34Gambia 0.01 0.00 0.01 1.51 0.00 1.51Ghana 0.24 0.00 0.24 22.21 0.00 22.21Guinea 0.25 0.00 0.25 9.23 0.00 9.23Guinea-Bissau 0.03 0.00 0.03 1.43 0.00 1.43Kenya 0.00 0.03 0.03 25.61 0.18 25.79Liberia 0.10 0.00 0.10 3.43 0.00 3.43Madagascar 0.61 0.09 0.70 17.28 0.00 17.28Malawi 0.12 0.00 0.12 13.45 0.00 13.45Mali 0.65 0.67 1.32 13.46 0.47 13.93Mauritania 0.28 0.53 0.81 0.93 0.40 1.33Mayotte 0.00 0.00 0.00 0.00 0.27 0.27Mozambique 0.82 0.00 0.82 21.06 0.00 21.06Namibia 0.35 0.19 0.54 1.25 0.38 1.64Niger 0.45 0.79 1.25 13.19 0.62 13.81Nigeria 0.93 0.00 0.93 134.60 0.00 134.60Rwanda 0.02 0.00 0.02 5.03 0.00 5.03Sao Tome and Principe 0.00 0.00 0.00 0.13 0.00 0.13Senegal 0.20 0.00 0.20 10.82 0.00 10.82

Region/Country Area at risk (km2) Population at risk (millions) Stable Unstable All Stable Unstable All
Sierra Leone 0.07 0.00 0.07 5.50 0.00 5.50South Africa 0.07 0.05 0.12 3.44 2.95 6.39Swaziland 0.01 0.00 0.01 0.23 0.00 0.23Togo 0.06 0.00 0.06 5.45 0.00 5.45Uganda 0.24 0.00 0.24 27.03 0.00 27.03United Republic of Tanzania 0.92 0.00 0.92 39.84 0.00 39.84Zambia 0.78 0.00 0.78 11.84 0.00 11.84Zimbabwe 0.31 0.00 0.31 7.44 0.00 7.44Total 15.70 3.11 18.81 601.06 8.43 609.49 AMRO Belize 0.00 0.01 0.01 0.00 0.17 0.17Bolivia 0.12 0.56 0.68 0.22 2.61 2.83Brazil 4.15 0.41 4.56 12.79 16.69 29.47Colombia 0.52 0.31 0.83 5.26 7.74 13.00Dominican Republic 0.02 0.01 0.03 1.41 2.83 4.24Ecuador 0.11 0.07 0.17 4.12 1.65 5.77French Guiana 0.08 0.00 0.08 0.14 0.00 0.14Guatemala 0.05 0.03 0.08 1.02 5.27 6.30Guyana 0.17 0.04 0.20 0.14 0.52 0.66Haiti 0.03 0.00 0.03 8.58 0.02 8.60Honduras 0.05 0.03 0.08 0.88 2.63 3.51Nicaragua 0.09 0.03 0.11 1.57 2.12 3.70Panama 0.02 0.00 0.02 0.90 0.00 0.90Peru 0.50 0.06 0.57 3.87 1.70 5.57Suriname 0.02 0.11 0.13 0.01 0.05 0.06Venezuela 0.15 0.49 0.64 0.22 6.24 6.46Total 6.06 2.17 8.23 41.13 50.23 91.37 EMRO Afghanistan 0.05 0.21 0.26 4.56 12.53 17.10Djibouti 0.00 0.02 0.02 0.02 0.41 0.43Iran 0.01 0.26 0.27 0.15 2.72 2.87Pakistan 0.21 0.55 0.76 30.74 68.30 99.04Saudi Arabia 0.01 0.03 0.03 0.72 1.22 1.94Somalia 0.58 0.06 0.64 10.04 0.55 10.59Sudan 1.52 1.06 2.57 28.99 6.84 35.83Yemen 0.21 0.25 0.47 15.93 5.72 21.65Total 2.60 2.43 5.02 91.14 98.30 189.44 EURO Kyrgyzstan 0.00 0.02 0.02 0.00 1.20 1.20Tajikistan 0.00 0.02 0.02 0.00 2.16 2.16Total 0.00 0.03 0.03 0.00 3.36 3.36 SEARO Bangladesh 0.02 0.05 0.07 15.12 47.99 63.11Bhutan 0.01 0.00 0.01 0.79 0.47 1.26India 1.63 1.32 2.95 414.53 535.59 950.12Indonesia 1.31 0.43 1.74 68.59 81.93 150.52Myanmar 0.68 0.01 0.69 42.88 1.91 44.79Nepal 0.01 0.02 0.04 3.40 6.15 9.54

Region/Country Area at risk (km2) Population at risk (millions) Stable Unstable All Stable Unstable All
Sri Lanka 0.01 0.03 0.05 1.75 7.53 9.28Thailand 0.20 0.27 0.47 16.53 30.53 47.06Timor-Leste 0.01 0.00 0.01 0.96 0.00 0.96Total 3.89 2.14 6.03 564.54 712.08 1276.62 WPRO Cambodia 0.18 0.01 0.19 10.77 2.55 13.33China 0.20 0.16 0.37 17.13 20.32 37.45Lao People's Dem. Republic 0.24 0.00 0.24 5.30 0.01 5.31Malaysia 0.23 0.00 0.23 6.29 16.17 22.46Papua New Guinea 0.40 0.00 0.40 4.11 0.00 4.11Philippines 0.16 0.06 0.22 26.95 20.41 47.35Solomon Islands 0.02 0.00 0.02 0.43 0.00 0.43Vanuatu 0.01 0.00 0.01 0.22 0.00 0.22Viet Nam 0.16 0.18 0.33 19.31 53.31 72.62Total 1.60 0.41 2.02 90.51 112.77 203.28 Global Totals 29.85 10.29 40.14 1388.39 985.17 2373.56

Protocol S1. Methods and sources of medical intelligence used to describe Plasmodium falciparum annual parasite incidence (PfAPI) globally
We defined areas of extremely low, unstable transmission risk as spatial units reporting a
PfAPI of less than 0.1 per 1000 population per annum (pa). This is because this criterion
was found to be a reliable indicator for the cessation of indoor residual spraying during the
consolidation phase of the Global Malaria Eradication Programme [1-4]. During this period,
the limit was reduced from 0.5‰ as it became recognized that surveillance, including
passive and active case detection of incident cases, was often less accurate and reliable
than nations thought: malaria often resumed after the cessation of spraying from 0.5‰, but
rarely from 0.1‰. This more conservative categorization of unstable transmission around
one clinical P. falciparum case 10,000 population (0.1‰ pa), thus, helps compensate for the
vagaries of district or provincial level reporting of parasitologically confirmed cases [5-7].
Using a higher threshold between ‘stable’ and ‘unstable’ transmission would have required a
greater confidence in the fidelity of the source information and the underlying surveillance.
PfAPI data sourcing
Table SI A1 summarizes the PfAPI data collection process and availability for all P.
falciparum malaria endemic countries. Complete PfAPI data were not available for Ecuador,
Bolivia, Haiti, Suriname and Venezuela. Risk maps presented at a regional malaria meeting
in Cartagena, Colombia [8], were used, showing high, medium, low and no risk areas for P.
falciparum and P. vivax. These maps were digitized and then combined with first-level
administrative unit species-specific API risk data reported in 2002 [9] to constrain the Pf-
limits within each country and map areas of either <0.1 or ≥0.1 cases per 1000 people pa.
When these first-level administrative unit case reports were less than 10 in 2002 [9], it was
assumed that contemporary risks in these areas were <0.1 ‰ pa. In Venezuela, the high risk
areas reported in the Cartagena meeting [8] corresponded with descriptions of international
travel advisories which state that risk in Amazonas and Bolivar is mainly restricted to the
Orinoco River basin and its tributaries in the former, and along the same river in the latter but
also in areas bordering the states of Apure and Guarico [10]. These descriptions were used
to create a 60 km distance buffer of high risk along the Orinoco and its tributaries. The map
of Venezuela in the Cartagena report shows high risk areas towards the north mainly in the
state of Sucre [8]. This disagrees with published reports [9,11] of API in Sucre being
attributed only to P. vivax, however, and probably refers to the local high risk of P. vivax
malaria. Sucre was, therefore, classified at no risk for P. falciparum. For Suriname, similar
descriptions of high risk restricted to the Maroni river are available [8] and, therefore, a

similar 60 km buffer was created along this river and classified as high risk. The latter buffer
did not capture two small high risk areas west of the river shown in the Cartagena map [8]
and these were left as unstable risk due to lack of geographic information to digitize them
with precision.
Plasmodium falciparum API data were not available for most countries within the WHO
Africa region, largely because these are not regarded as priority malaria metrics for this
region and reporting systems are probably the least reliable when compared to other regions
of the world [5-7,12,13]. Southern Africa defined as Swaziland, South Africa, Zimbabwe,
Botswana and Namibia, represents an exception with a combination of published risk data
[14,15] and expert opinion from national malaria control programmes [16] used to estimate
the limits of risk through matched administrative boundaries or digitized maps in these
countries. For island populations risks were reviewed from published sources to confirm
current P. falciparum transmission status including Cape Verde [17], Comoros [18], Mayotte
[19], and Mauritius [19]. The islands of Reunion and Seychelles were excluded as malaria
endemic as confirmed through published literature [19].
Malaria programme managers in EMRO confirmed that the small foci of malaria transmission
in Ihrir, Illizi Department, Algeria and the Fayoum Governate, Egypt, are entirely due to P.
vivax. Similarly, the limited cases reported in Syria were all due to P. vivax. These countries
were, therefore, confirmed as P. falciparum free. Tajikistan and Kyrgyzstan were regarded
as the only P. falciparum endemic countries in the WHO’s European region. Both countries
have reported P. falciparum malaria cases since 2003 [20], with very few cases reported in
2006, however, and with elimination scheduled by 2010 [21].
Medical intelligence confirmed the P. vivax only status of the Korean Peninsula [22,23],
Argentina [24], Paraguay [24], Iraq [25], Uzbekistan [26], Turkey [25], Azerbaijan [25] and
Mauritius [27]. The United Arab Emirates have not reported any autochthonous cases of P.
falciparum malaria for over five years [28] and satisfied the criteria for the certification of the
eradication of malaria in 2007 [29]. The Chiapas region of Mexico, Canton Matua, in Costa
Rica and Ahuachapa in El Salvador have all had only 1 to12 P. falciparum cases reported
over the last five years, and these countries are regarded as largely P. falciparum free [24].
Mapping PfAPI data
In order to map PfAPI data consistently, digital boundaries of first and second level units
were obtained from the United Nations' Second Administrative Level Boundaries (SALB)

dataset (n=24) [30] and the Global Administrative Unit Layers (GAUL) developed by the
Food and Agriculture Organization (FAO; n=82) [31]. In addition, PfAPI classifications were
reconciled at third-level administrative units for Nepal (Global Administrative Areas database
of the University of California at Berkeley, http://biogeo.berkeley.edu/gadm) and restricted
areas in Peru through the Peruvian Dirección General de Epidemiología
(http://www.oge.sld.pe). In South Africa, third-level administrative data [16] were digitized
against boundary data provided by FAO Geonetwork [32].
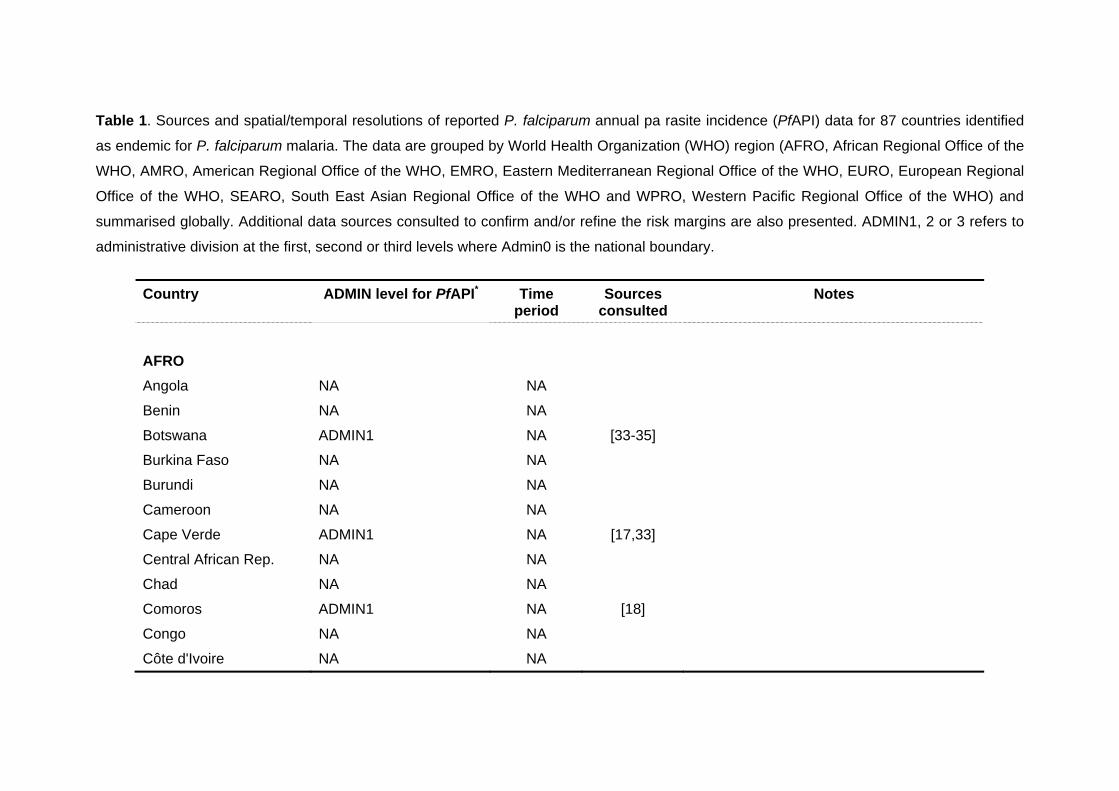
Table 1. Sources and spatial/temporal resolutions of reported P. falciparum annual pa rasite incidence (PfAPI) data for 87 countries identified
as endemic for P. falciparum malaria. The data are grouped by World Health Organization (WHO) region (AFRO, African Regional Office of the
WHO, AMRO, American Regional Office of the WHO, EMRO, Eastern Mediterranean Regional Office of the WHO, EURO, European Regional
Office of the WHO, SEARO, South East Asian Regional Office of the WHO and WPRO, Western Pacific Regional Office of the WHO) and
summarised globally. Additional data sources consulted to confirm and/or refine the risk margins are also presented. ADMIN1, 2 or 3 refers to
administrative division at the first, second or third levels where Admin0 is the national boundary.
Country ADMIN level for PfAPI* Time period
Sources consulted
Notes
AFRO
Angola NA NA
Benin NA NA
Botswana ADMIN1 NA [33-35]
Burkina Faso NA NA
Burundi NA NA
Cameroon NA NA
Cape Verde ADMIN1 NA [17,33]
Central African Rep. NA NA
Chad NA NA
Comoros ADMIN1 NA [18]
Congo NA NA
Côte d'Ivoire NA NA
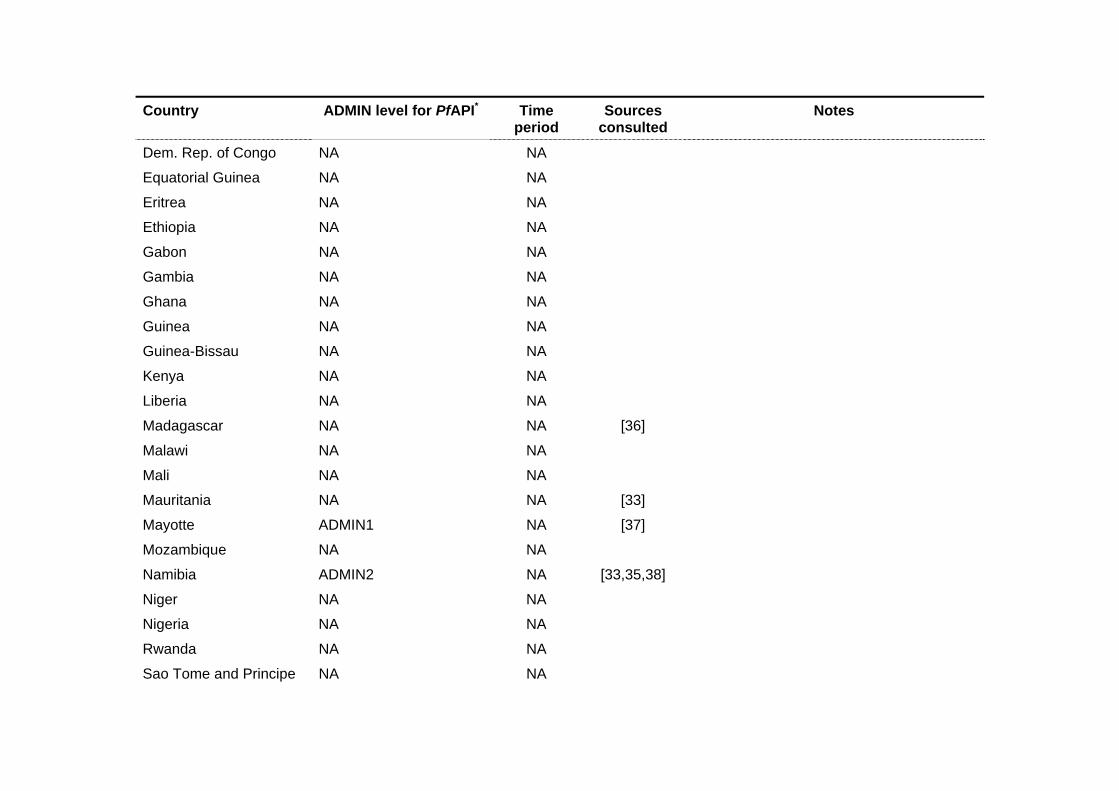
Country ADMIN level for PfAPI* Time period
Sources consulted
Notes
Dem. Rep. of Congo NA NA
Equatorial Guinea NA NA
Eritrea NA NA
Ethiopia NA NA
Gabon NA NA
Gambia NA NA
Ghana NA NA
Guinea NA NA
Guinea-Bissau NA NA
Kenya NA NA
Liberia NA NA
Madagascar NA NA [36]
Malawi NA NA
Mali NA NA
Mauritania NA NA [33]
Mayotte ADMIN1 NA [37]
Mozambique NA NA
Namibia ADMIN2 NA [33,35,38]
Niger NA NA
Nigeria NA NA
Rwanda NA NA
Sao Tome and Principe NA NA
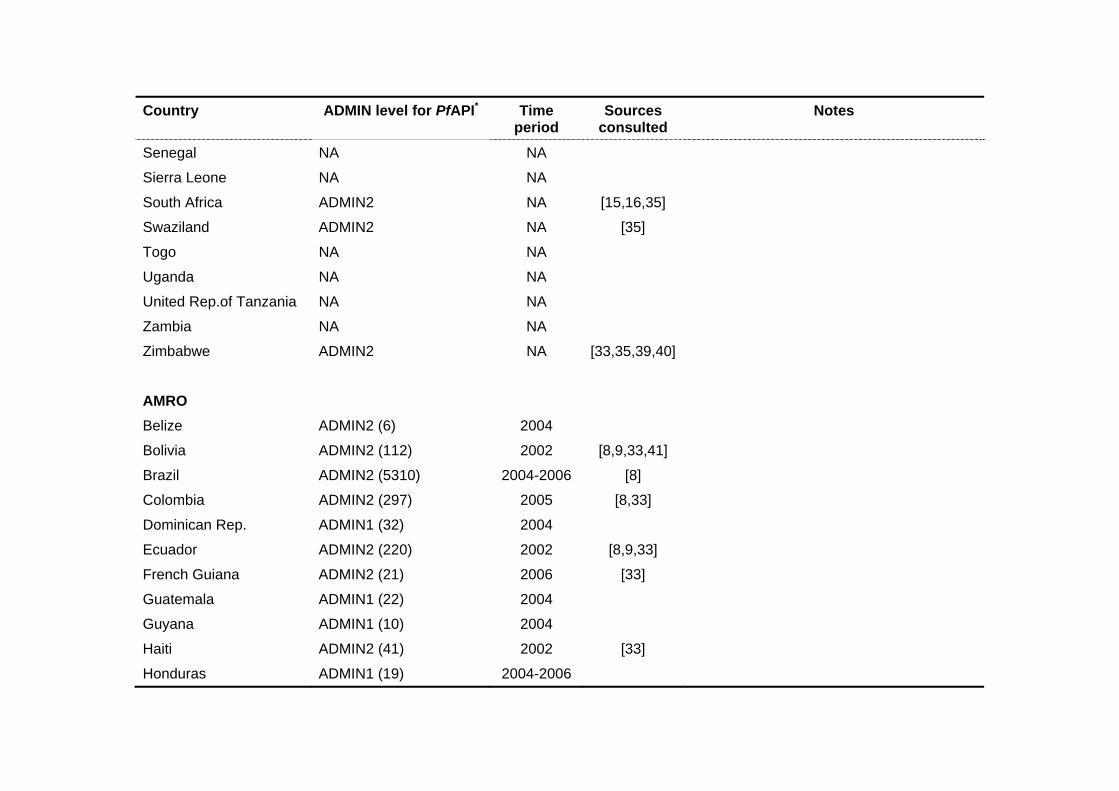
Country ADMIN level for PfAPI* Time period
Sources consulted
Notes
Senegal NA NA
Sierra Leone NA NA
South Africa ADMIN2 NA [15,16,35]
Swaziland ADMIN2 NA [35]
Togo NA NA
Uganda NA NA
United Rep.of Tanzania NA NA
Zambia NA NA
Zimbabwe ADMIN2 NA [33,35,39,40]
AMRO
Belize ADMIN2 (6) 2004
Bolivia ADMIN2 (112) 2002 [8,9,33,41]
Brazil ADMIN2 (5310) 2004-2006 [8]
Colombia ADMIN2 (297) 2005 [8,33]
Dominican Rep. ADMIN1 (32) 2004
Ecuador ADMIN2 (220) 2002 [8,9,33]
French Guiana ADMIN2 (21) 2006 [33]
Guatemala ADMIN1 (22) 2004
Guyana ADMIN1 (10) 2004
Haiti ADMIN2 (41) 2002 [33]
Honduras ADMIN1 (19) 2004-2006

Country ADMIN level for PfAPI* Time period
Sources consulted
Notes
Nicaragua ADMIN1 (18) 2004
Panama ADMIN2 (68) 2006
Peru ADMIN2 (191) and ADMIN3 (14)
2004 [8,33]
Suriname ADMIN2 (62) 2002 [9,33] A 60km buffer around the Maroni river was assumed to correspond to high risk
Venezuela ADMIN2 (318) 2002 [9,33,41] Assumptions of risk confined mainly near the Orinoco and its main tributaries (60km buffer) were deemed sensible to avoid overestimating risk in Bolivar and Amazonas
EMRO
Afghanistan ADMIN1 (32) 2005 [42]
Djibouti ADMIN1 NA [43]
Iran ADMIN2 (251) 2004-2006 [44,45]
Pakistan ADMIN2 (119) 2004-2006 [46] No information available for six ‘tribal areas’ in the Fata region of Pakistan and 15 ADMIN1 units in the disputed territory of Jammu Kashmir.
Saudi Arabia ADMIN1 (13) 2005-2006 [47]
Somalia ADMIN2 NA
Sudan ADMIN1 NA
Yemen ADMIN1 (19) 2002, 2005-2006

Country ADMIN level for PfAPI* Time period
Sources consulted
Notes
EURO
Kyrgyzstan ADMIN2 (40) 2002-2005 [20,48]
Tajikistan ADMIN2 (56) 2005-2006 [20,48]
SEARO
Bangladesh ADMIN2 (64) 2003-2004 [49]
Bhutan ADMIN1 (20) 2002-2004 [33,49]
India ADMIN2 (538) 2002-2004 Twenty-two ADMIN1 units could not be reconciled or matched to any reported PfAPI data in India and were left as missing data.
Indonesia ADMIN1/2 (281) 2005 [49] The following rules were used to interpolate ADMIN1 data to some ADMIN2 polygons: i) in 5 cases, no ADMIN2 data were available for the whole ADMIN1 and this was defaulted to 2005 ADMIN1 data; ii) there were 50 ADMIN2s for which no data were available but other ADMIN2 units within the same ADMIN1 had data; these missing ones were assigned the overall ADMIN1 value.
Myanmar ADMIN1 (14) and ADMIN2 (11)
2003-2004 [33,49]
Nepal ADMIN3 (175) 2002-2003 [33,49]
Sri Lanka ADMIN2 (25) 2004 [33,50]
Thailand ADMIN1 (76) 2003-2004 [33,51]

Country ADMIN level for PfAPI* Time period
Sources consulted
Notes
Timor Leste ADMIN1 (13) 2004-2005
WPRO
Cambodia ADMIN1 (24) 2003-2005
China ADMIN1 (32) ADMIN2 (16)i
2003-2005 [33,52-54]
Lao Dem. Peoples Rep. ADMIN1 (18) 2003-2005 [33,55]
Malaysia ADMIN1 (15) 2003-2005 [56]
Papua New Guinea ADMIN1 (20) 2003-2005 [57]
Philippines ADMIN2 (79) 2003-2005 [33]
Solomons ADMIN1 (10) 2003-2005
Vanuatu ADMIN1 (6) 2003-2005
Vietnam ADMIN1 (61)i 2003-2005 [33,55]
*Missing data: Apart from data missing in Pakistan and India (specified in the table), data were not available for the following territories: The
Santanilla islands of Honduras, D.I. Yogyakarta of Indonesia, the district of Kilinochchi in Sri Lanka and the province of Bac Nihn in Vietnam.

References
1. Black R (1968) Manual of epidemiology and epidemiological services in malaria programmes. Geneva: World Health Organization. 223 p.
2. Hay SI, Smith DL, Snow RW (2007) Measuring malaria endemicity from intense to interrupted transmission. Lancet Infect Dis: in press.
3. Pampana E (1969) A textbook of malaria eradication. London: Oxford University Press. 593 p.
4. Yekutiel P (1980) Eradication of infectious diseases: a critical study. 2 Contributions to epidemiology and biostatistics. Basel, Switzerland: Karger. pp. 164.
5. Erhart A, Thang ND, Xa NX, Thieu NQ, Hung LX, et al. (2007) Accuracy of the health information system on malaria surveillance in Vietnam. Trans R Soc Trop Med Hyg 101: 216-225.
6. Snow RW, Guerra CA, Noor AM, Myint HY, Hay SI (2005) The global distribution of clinical episodes of Plasmodium falciparum malaria. Nature 434: 214-217.
7. Sharma VP (2007) Battling the malaria iceberg with chloroquine in India. Malar J 6: 105. 8. PAHO (2000) The Roll Back Malaria Initiative in the Rainforest Region of South America.
Cartagena: Pan American Health Organization. 9. PAHO (2003) Report on the Status of Malaria Programs in the Americas (Based on 2002
data) CD44/INF/3. Washington, D. C.: Pan American Health Organization / World Health Organization - 44th Directing Council/55th Session of the Regional Committee.
10. IAMAT (2004) World Malaria Risk Chart. Available: http://www.iamat.org/pdf/WorldMalariaRisk.pdf. Accessed: July 2007.
11. Caceres JL, Pizzo N, Vela FA, Perez W, Rojas JG, et al. (2005) Impacto de la cura radical masiva sobre la incidencia malaria del estado de Sucre, Venezuela. Bol Malar Sal Amb 45: 27-36.
12. Chilundo B, Sundby J, Aanestad M (2004) Analysing the quality of routine malaria data in Mozambique. Malar J 3: 3.
13. Gething PW, Noor AM, Gikandi PW, Ogara EA, Hay SI, et al. (2006) Improving imperfect data from health management information systems in Africa using space-time geostatistics. PLoS Med 3: e271.
14. Kleinschmidt I, Sharp B, Mueller I, Vounatsou P (2002) Rise in malaria incidence rates in South Africa: a small-area spatial analysis of variation in time trends. Am J Epidemiol 155: 257-264.
15. Kleinschmidt I, Sharp BL, Clarke GP, Curtis B, Fraser C (2001) Use of generalized linear mixed models in the spatial analysis of small-area malaria incidence rates in Kwazulu Natal, South Africa. Am J Epidemiol 153: 1213-1221.
16. Malaria in Southern Africa (2005) Malaria risk update 2005. http://www.malaria.org.za/malaria_risk/update2005.
17. Alves J, Roque AL, Cravo P, Valdez T, Jelinek T, et al. (2006) Epidemiological characterization of Plasmodium falciparum in the Republic of Cabo Verde: implications for potential large-scale re-emergence of malaria. Malar J 5: 32.
18. Blanchy S, Julvez J, Mouchet J (1999) Stratification epidemiologique du paludisme dans l'archipel des Comores. Bull Soc Pathol Exot 92: 177-184.
19. Tchen J, Ouledi A, Lepere JF, Ferrandiz D, Yvin JL (2006) Epidemiologie et prevention du paludisme dans les iles du sud-ouest de l'Ocean Indien. Med Trop (Mars) 66: 295-301.
20. Ejov M (2005) Scaling up the response to malaria in the WHO European Region: Progress toward curbing an epidemic 2000-2004. Copenhagen: World Health Organization Regional Office for Europe.
21. WHO Regional Office for Europe (2006) Regional strategy: from malaria control to elimination in the WHO European Region 2006-2015. WHO-EUR/06/5061322. Copenhagen: World Health Organization Regional Office for Europe. 41 p.

22. Yeom JS, Kim TS, Oh S, Sim JB, Barn JS, et al. (2007) Plasmodium vivax malaria in the Republic of Korea during 2004-2005: changing patterns of infection. Am J Trop Med Hyg 76: 865-868.
23. Yeom JS, Ryu SH, Oh S, Lee WJ, Kim TS, et al. (2005) Status of Plasmodium vivax malaria in the Republic of Korea during 2001-2003. Am J Trop Med Hyg 73: 604-608.
24. PAHO (2006) Regional Strategic Plan for Malaria in the Americas 2006-2010. Washington DC: Pan American Health Organization.
25. Korenromp E (2005) Malaria incidence estimates at country level for the year 2004 - Proposed estimates and draft report. Geneva, Switzerland.
26. Severini C, Menegon M, Di Luca M, Abdullaev I, Majori G, et al. (2004) Risk of Plasmodium vivax malaria reintroduction in Uzbekistan: genetic characterization of parasites and status of potential malaria vectors in the Surkhandarya region. Trans R Soc Trop Med Hyg 98: 585-592.
27. WHO (2005) International Travel and Health: Situation as on 1 January 2005. Geneva: World Health Organization.
28. WHO Regional Office for the Eastern Mediterranean (2007) Strategic plan for malaria control and elimination in the WHO Eastern Mediterranean Region 2006–2010. WHO-EM/MAL/340/E. Cairo: World Health Organization Regional Office for the Eastern Mediterranean. 41 p.
29. WHO (2007) United Arab Emirates certified malaria-free. Wkly Epidemiol Rec 82: 25-32. 30. SALB (2007) Second Administrative Level Boundaries.
http://www3.who.int/whosis/gis/salb/salb_home.htm. 31. FAO (2005) The Global Administrative Unit Layers (GAUL). Rome: Food and Agriculture
Organization of the United Nations. 32. FAO (2007) FAO GeoNetwork. http://www.fao.org/geonetwork/srv/en/about. Accessed:
June 2007. 33. CDC (2007) Travelers' Health. http://www.cdc.gov/travel/regionalmalaria/. Accessed:
July 2007. 34. Ministry of Health of the Republic of Botswana (2001) Botswana Roll Back Malaria
Strategic Plan 2002-2005. National Malaria Control Programme. 7 p. 35. SAMC Malaria risk maps in Southern Africa. http:/www.malaria.org.zw/countries.
Accessed: July 2007. 36. Pasteur Institute of Madagascar, Roll Back Malaria (2004) Madagascar Malaria Map.
Malaria Reserach Unit. 37. Julvez J, Galtier J, Halidi MA, Henry M, Mouchet J (1987) Actualites epidemiologiques
de paludisme. Epidemiologie du paludisme et lute antipaludique a Mayotte (Archipel des Comores, Ocean Indien). Bull Soc Pathol Exot Filiales 80: 505-519.
38. Ministry of Health & Social Sciences of the Republic of Namibia (1995 ) National policy and strategy for malaria control. Ministry of Health & Social Sciences, Republic of Namibia. 95 p.
39. Root G (2006) Malaria prevalence and community-based survey. Ministry of Health and Child Welfare of Zimbabwe and World Health Organization.
40. Shiff C (2002) Integrated approach to malaria control. Clin Microbiol Rev 15: 278-293. 41. PAHO. Meeting of national directors of epidemiology and malaria programs; 2005 7–10
November 2005; San José. Pan American Health Organization. 42. Asha AW (2004) Afghanistan country report at 4th RBM-EMRO meeting at Isfahan, Iran
25-28 May 2004. Isfahan, Iran. 43. National Malaria Control Programme Manager (2007). Djibouti. 44. Raesi A (2007) National Malaria Control Programme Manager. Iran. 45. Zamani G (2007) RBM Monitoring and Evaluation coordinator for EMRO. 46. Mansoor F (2007) National Malaria Control Programme Manager. Pakistan. 47. Saudi Arabia National Malaria Control Programme Manager (2007). 48. WHO European Region (2006) Centralized Information System for Infectious Diseases
(CISID). http://data.euro.who.int/cisid/. 49. SEARO (2007). www.searo.who.int/EN/section10.

50. Briet OJ, Gunawardena DM, van der Hoek W, Amerasinghe FP (2003) Sri Lanka malaria maps. Malar J 2: 22.
51. Zhou G, Sirichaisinthop J, Sattabongkot J, Jones J, Bjornstad ON, et al. (2005) Spatio-temporal distribution of Plasmodium falciparum and P. vivax malaria in Thailand. Am J Trop Med Hyg 72: 256-262.
52. Zhou SS, Tang LH, Sheng HF (2005) Malaria situation in the People's Republic of China in 2003. Zhongguo Ji Sheng Chong Xue Yu Ji Sheng Chong Bing Za Zhi 23: 385-387.
53. Zhou SS, Tang LH, Sheng HF, Wang Y (2006) Malaria situation in the People' s Republic of China in 2004. Zhongguo Ji Sheng Chong Xue Yu Ji Sheng Chong Bing Za Zhi 24: 1-3.
54. Zhou SS, Wang Y, Tang LH (2006) Malaria situation in the People's Republic of China in 2005. Zhongguo Ji Sheng Chong Xue Yu Ji Sheng Chong Bing Za Zhi 24: 401-403.
55. Pratap S (2003) Update on malaria, multidrug resistance and economic development in the Mekong region of South East Asia. Southeast Asian J Trop Med Public Health 34: 105 pages.
56. WPRO (2005) Malaria Epidemiology, Cambodia. www.wpro.who.int/sites/mvp/epidemiology/malaria/.
57. Muller I, Bockarie M, Alpers M, Smith T (2003) The epidemiology of malaria in Papua New Guinea. Trends Parasitol 19: 253-259.

Protocol S2: Developing global biological limits for Plasmodium falciparum transmission
The temperature mask
Temperature affects many aspects of mosquito physiology [1]. One aspect critical for malaria
transmission is the temperature dependence of sporogony: the time it takes for sporozoites
to develop and become infectious in Anopheles. A method for estimating the duration of
sporogony has been proposed, based on the number of degree-days required by the
parasite to complete development [2,3], for malaria the sum of the number of degrees in a
day by which the mean temperature exceeds the minimum required for the development of
sporozoites. Nikolaev [4] showed that the degree-days required for the maturation of
sporozoites in an An. maculipennis population from Russia were 105 for P. vivax, 111 for P.
falciparum and 144 for P. malariae and that parasite development ceased below 16oC for P.
falciparum and P. malariae and below 14.5oC for P. vivax. The duration of sporogony (DS) in
days can thus be calculated as:
MINMEAN TTDS
−=
DD [1]
where DD are the parasite species-specific degree-days, TMEAN is the mean ambient
temperature and TMIN is the minimum temperature required for parasite development.
Figure 1 plots the results of the above equation for P. falciparum, P. vivax and P. malariae
development in An. maculipennis. It shows that P. vivax is able to develop at the lowest
temperatures, followed by P. falciparum and P. malariae, thus helping explain the species
specific latitudinal limits of the parasites globally [5,6]. The figure also shows that the length
of sporogony of P. falciparum increases rapidly when temperatures drop below 22oC. The
curve never reaches a true asymptote on the y-axis; but the duration of sporogony becomes
so extended that few anophelines will survive long enough to inoculate humans and at 16oC
parasite development ceases entirely. Conversely, as temperature increases the duration of
sporogony decreases, so that at 30oC it can take less than ten days. Obviously, the duration
of sporogony then becomes limited by parasite and vector survival, which plummet as
temperatures rise above 32oC [7]. The duration of sporogony is dependent fundamentally on
enzyme kinetics [8] and thus widely assumed to be relatively independent of vector species.
It is the interplay between the duration of sporogony and the species-specific longevity of the
Anopheles vector that forms the basis of the temperature mask.
1
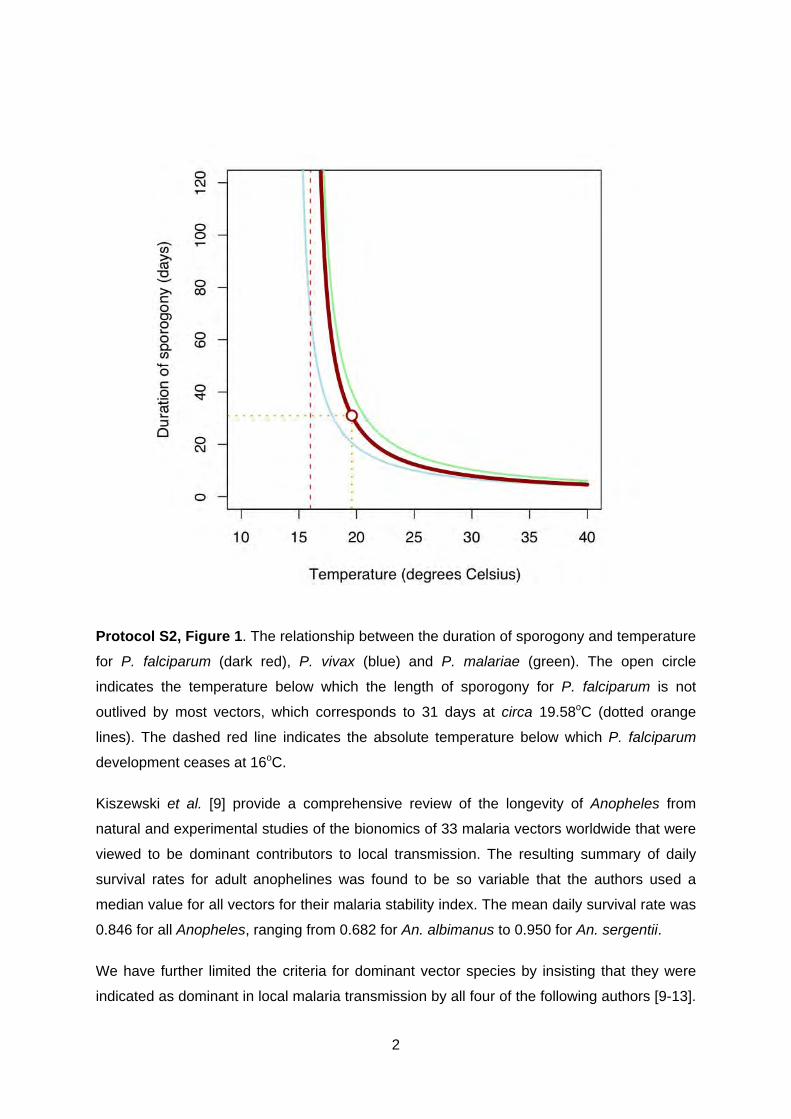
Protocol S2, Figure 1. The relationship between the duration of sporogony and temperature
for P. falciparum (dark red), P. vivax (blue) and P. malariae (green). The open circle
indicates the temperature below which the length of sporogony for P. falciparum is not
outlived by most vectors, which corresponds to 31 days at circa 19.58oC (dotted orange
lines). The dashed red line indicates the absolute temperature below which P. falciparum
development ceases at 16oC.
Kiszewski et al. [9] provide a comprehensive review of the longevity of Anopheles from
natural and experimental studies of the bionomics of 33 malaria vectors worldwide that were
viewed to be dominant contributors to local transmission. The resulting summary of daily
survival rates for adult anophelines was found to be so variable that the authors used a
median value for all vectors for their malaria stability index. The mean daily survival rate was
0.846 for all Anopheles, ranging from 0.682 for An. albimanus to 0.950 for An. sergentii.
We have further limited the criteria for dominant vector species by insisting that they were
indicated as dominant in local malaria transmission by all four of the following authors [9-13].
2

The daily survival rate for the 19 main/dominant vectors by region was used to determine the
fraction of the population surviving over successive days (Figure 2). Although temperature
will affect other parameters of the basic reproduction rate of infection [14], including biting
and resting vector habits, it seems reasonable to consider the proportion of the population
surviving 31 days as the critical point of interruption of P. falciparum transmission. To a close
approximation, most vector populations would have been reduced to 99% of their original
population size within 31 days. The regional variation encountered in this approximation was
then explored further.
In AMRO, the mean daily survival probability for the five regionally dominant vectors is
0.782. This means that 99.95% of an initial population would not survive 31 days (Figure 2,
top left). By far the longest-lived vector in the region is An. pseudopunctipennis for which
98.09% of the population will have perished in 31 days; this makes this vector resistant to
higher altitudes across its distribution in the Andean slopes [15]. Similarly, the mean daily
survival probability for the three regionally dominant vectors in AFRO is 0.780. This means
that 99.95% of an initial population would not survive 31 days (Figure 2, top right). In
SEARO/WPRO the mean daily survival probability for the eight regionally dominant vectors
is 0.836. This predicts that 99.62% of an initial population would not survive 31 days (Figure
2, bottom right). The longest lived vector in the region is An. dirus, for which 93.41% of the
population will have perished in 31 days. This species is mainly a forest dweller [16,17] and
therefore is restricted geographically and ecologically to a niche that does not suffer extreme
temperature limits. Excluding areas where average temperatures were such that sporogony
would not complete in 31 days was thus considered a biologically plausible and conservative
limit of P. falciparum malaria transmission in these regions.
In EMRO the mean daily survival probability for the five regionally dominant vectors is 0.846.
This indicates that 99.45% of an initial population would not survive 31 days (Figure 2,
bottom left). This mean disguises two relatively long-lived vectors in the region: An. sergentii,
which is an oasis breeder, and An. superpictus, which is another mountain foothill breeder
[11,12,18]. These species have longevities that predict that only 79.61% and 82.69% of their
populations would have died after a 31-day period. The 31 days time limit was applied for
this region, except where these species are dominant, where it was doubled to 62 days. This
duration would result in a population mortality of 95.84% and 97.00% for An. sergentii and
An. superpictus, respectively. The distributions of these vectors described by White [13]
were digitized and the 62-day rule applied within these species ranges.
3

Protocol S2, Figure 2. The fraction of the original population of Anopheles surviving by day
for each dominant vector species of the World Health Organization (WHO) geographical
regions. (AFRO, African Regional Office of the WHO, AMRO, American Regional Office of
the WHO, EMRO, Eastern Mediterranean Regional Office of the WHO, EURO, European
Regional Office of the WHO, SEARO, South East Asian Regional Office of the WHO and
WPRO, Western Pacific Regional Office of the WHO). In all panels, the red line is the
average survival fraction for the region and each species is identified by a unique colour
shown in the top right of the panel. The black dotted lines mark the 31 days duration of
sporogony limit. The green dotted line in the EMRO/EURO panel marks the 62 days criterion
applied for An. sergentii and An. superpictus.
4

In summary, with the exception of An. sergentii and An. superpictus, it is rare for adult
dominant vectors of malaria to survive longer than a month, with more than 99% of the
average population dying after 31 days. The longer-lived vectors are generally those
adapted to survive at higher altitudes or harsher conditions, such as is the case of An.
superpictus and An. sergentii. Despite the fact that a relatively small proportion of the
populations of these vectors are normally able to survive longer than one month, the
numbers of individuals surviving might still pose a significant risk for malaria transmission by
being able to support parasite development at lower temperatures. After two months,
however, most individuals of both species (>95%) would also have succumbed.
Using average monthly temperature records estimated from a global climate surface [19],
the duration of P. falciparum sporogony was estimated for each month. Those pixels where
the duration of sporogony was 31 days or less were identified in each month. This provided
12 images with a binary outcome of whether P. falciparum sporogony could be completed in
more or less than 31 days. The images were combined to identify the number of temperature
suitable months available in a synoptic year (Figure 3). All pixels where the 31 days limit was
not achieved for any single month (i.e. grey pixels in Figure 3), or two consecutive months in
the geographic range of An. sergentii and An. superpictus, were used as a conservative
mask to exclude areas where transmission is highly unlikely to occur.
Protocol S2, Figure 3. An overlay of 12 monthly images of where the duration of sporogony
exceeds 31 days, restricted to P. falciparum Malaria Endemic Countries (PfMECs). Pixels
where the temperature did not reach 19.58oC in any single month of a synoptic year (here
5

shown as grey areas within the PfMECs) were used to mask in all areas (except within the
range of An. sergentii and An. superpictus were two consecutive months were required).
The aridity mask
Upper temperature limits were not defined on the basis of physiological tolerances of vectors
measured in laboratories [20-23] as these were so high as to be rarely achieved in nature
and often subject to behavioural avoidance [20]. We preferred a partial surrogate for extreme
aridity: a hybrid measure encompassing both high temperature and low water availability.
The ability of adult vectors to survive long enough to contribute to parasite transmission and
their eggs and larvae to survive in sufficient numbers to sustain transmission is dependent
on the level of humidity and the species-specific ability to withstand arid conditions [24-26].
Hyper-aridity is the main criterion used to define a desert biome [27] and, therefore,
identifying desert extents was assumed to be an accurate proxy for the extreme mask to limit
the risk of malaria transmission.
These potentially limiting conditions prevail in deserts and their fringes found in malaria
endemic countries, notably the Sahara (and the Sahel), the Namib, the Arabian and the Thar
deserts, as well as the northern arid areas of East Africa and Peru (Figure SI B4). Since in
these areas optimum growth of most plants is hindered, a proxy for vegetation cover can be
used to classify arid areas [28]. Such a proxy can be derived from satellite sensors by
combining the information of different channels of the electromagnetic spectrum to derive
vegetation indices [29]. One of the most commonly used is the normalized difference
vegetation index (NDVI) [30], available as a multitemporal series from the Advanced Very
High Resolution Radiometer sensor [31] and, more recently, from the MODerate-resolution
Imaging Spectroradiometer (MODIS) sensor on board the Terra and Aqua satellites [32-34].
In addition to NDVI, MODIS products include the enhanced vegetation index (EVI) [34]. EVI
is calculated similarly to NDVI, which is derived from two channels of the electromagnetic
spectrum (red and near-infrared). EVI incorporates a third channel (blue) and corrects for
some of the distortion caused by atmospheric particles and ground cover beneath the
vegetation. This makes EVI a more robust index by offering improved sensitivity, particularly
in areas with high biomass content where it saturates less than NDVI, but also reduced
contamination throughout by particles in the air [32-34].
Temporal Fourier processed, monthly, bi-directional reflectance distribution function
corrected EVI images [32,35,36] were reclassified using ArcView GIS 3.2 (ESRI 1999) to
give a binary output of areas where EVI ≤0.1 and EVI >1. These reclassified images were
6

then overlaid in pairs to produce 12 new images. The 12 pairs were then combined to
identify pixels where conditions were suitable for transmission (i.e. where EVI pixel values in
a synoptic year were higher than 0.1 for at least 2 consecutive months).
Despite strict quality control, these data are affected by atmospheric contamination in the
form of clouds and aerosols, although these effects are less frequent in arid low rainfall
areas with infrequent cloud cover. To avoid the introduction of these errors in the final mask,
a sub-mask was used of only those territories within P. falciparum endemic countries that
were defined as being at some level of risk according to PfAPI data and that overlapped with
the arid areas defined by the EVI threshold. This included whole or partial territories of 20
PfMECs as follows: Afghanistan, Angola, Chad, Djibouti, Eritrea, Ethiopia, India (northwest),
Iran, Kenya, Kyrgyzstan, Mali, Mauritania, Niger, Pakistan, Peru (northwest), Saudi Arabia
(southwest), Somalia, Sudan, Tajikistan and Yemen.
The aridity sub-mask was applied in a step-wise fashion by which risk was down-regulated
one class (i.e. stable to unstable and unstable to no risk). Therefore, the only areas where
risk was excluded were those where PfAPI had already defined limited risk of malaria. The
sub-mask was then applied on top of the PfAPI and temperature masks (Figure 1, bottom
panel, main paper).
Protocol S2, Figure 4. Overlay of reclassified monthly EVI images (≤0.1 and >0.1). The
scale shows the number of ‘arid’ months occurring in each pixel in a synoptic year. Despite
quality control, cloud contamination was still evident in some humid areas (e.g. Gulf of
7

8
Guinea) as fine speckle. The red outline indicates P. falciparum risk areas where the aridity
mask was applied to avoid introducing cloud-contaminated pixels in the final image.

References 1. Clements AN (1999) The Biology of Mosquitoes. Wallingford, UK: CABI Publishing. 740 p. 2. Detinova TS (1962) Age grouping methods in Diptera of medical importance with special
reference to some vectors of malaria. Geneva: World Health Organization. 3. Moshkovsky SD (1946) [The dependence upon temperature of the speed of development
of malaria plasmodia in the mosquito]. Med Parazitol (Mosk) 15: 19. 4. Nikolaev BP (1935) The influence of temperature on the development of malaria
plasmodia in the mosquito. Tr Pasteur Inst Epidem Bakt (Leningr) 2: 108-109. 5. Lysenko AY, Semashko IN (1968) Geography of Malaria: a medico-geographic profile of
an ancient disease. In: Lebedew AW, editor. Medicinskaja Geografija. Moscow: Academy of Sciences. pp. 25-146.
6. Lysenko AJ, Beljaev AE (1969) An analysis of the geographical distribution of Plasmodium ovale. Bull World Health Organ 40: 383-394.
7. Muirhead-Thompson RC (1951) Mosquito behaviour in relation to malaria transmission and control in the tropics. London: Edward Arnold & Co.
8. Sharpe PJ, DeMichele DW (1977) Reaction kinetics of poikilotherm development. J Theor Biol 64: 649-670.
9. Kiszewski A, Mellinger A, Spielman A, Malaney P, Sachs SE, et al. (2004) A global index representing the stability of malaria transmission. Am J Trop Med Hyg 70: 486-498.
10. Mouchet J, Carnevale P, Coosemans M, Julvez J, Manguin S, et al. (2004) Biodiversité du paludisme dans le monde. Paris: John Libbey Eurotext. 428 p.
11. Service MW (1993) Mosquitoes (Culicidae). In: Lane RP, Crosskey RW, editors. Medical Insects and Arachnids. London: Chapman & Hall. pp. 120-240.
12. Service MW (1993) The Anopheles vector. Bruce-Chwatt's Essential Malariology. London: Edward Arnold. pp. 96-123.
13. White GB (1989) Malaria. In: Sloof R, editor. Geographical distribution of arthropod-borne diseases and their principal vectors WHO/VBC/89967. Geneva: World Health Organization, Division of Vector Biology and Control. pp. 7-22.
14. Rogers DJ, Randolph SE (2006) Climate change and vector-borne diseases. Adv Parasitol 62: 345-381.
15. Rubio-Palis Y, Zimmerman RH (1997) Ecoregional classification of malaria vectors in the neotropics. J Med Entomol 34: 499-510.
16. Guerra CA, Snow RW, Hay SI (2006) A global assessment of closed forests, deforestation and malaria risk. Ann Trop Med Parasitol 100: 189-204.
17. Obsomer V, Defourny P, Coosemans M (2007) The Anopheles dirus complex: spatial distribution and environmental drivers. Malar J 6: 26.
18. Zahar AR (1984) Vector control operations in the African context. Bull World Health Organ 62 Suppl: 89-100.
19. Hijmans RJ, Cameron SE, Parra JL, Jones PG, Jarvis A (2005) Very high resolution interpolated climate surfaces for global land areas. Int J Climatol 25: 1965-1978.
20. Kirby MJ, Lindsay SW (2004) Responses of adult mosquitoes of two sibling species, Anopheles arabiensis and A. gambiae s.s. (Diptera: Culicidae), to high temperatures. Bull Entomol Res 94: 441-448.
21. Bayoh MN, Lindsay SW (2003) Effect of temperature on the development of the aquatic stages of Anopheles gambiae sensu stricto (Diptera: Culicidae). Bull Entomol Res 93: 375-381.
22. Bayoh MN, Lindsay SW (2004) Temperature-related duration of aquatic stages of the Afrotropical malaria vector mosquito Anopheles gambiae in the laboratory. Med Vet Entomol 18: 174-179.
23. Jepson WF, Moutia A, Courtois C (1947) The malaria problem in Mauritius: the bionomics of Mauritian anophelines. Bull Entomol Res 38: 177-208.
9

10
24. Shililu JI, Grueber WB, Mbogo CM, Githure JI, Riddiford LM, et al. (2004) Development and survival of Anopheles gambiae eggs in drying soil: influence of the rate of drying, egg age, and soil type. J Am Mosq Control Assoc 20: 243-247.
25. Omer SM, Cloudsley-Thompson JL (1970) Survival of female Anopheles gambiae Giles through a 9-month dry season in Sudan. Bull World Health Organ 42: 319-330.
26. Omer SM, Cloudsley-Thomson JL (1968) Dry season biology of Anopheles gambiae Giles in the Sudan. Nature 217: 879-880.
27. UNEP (2006) Global Deserts Outlook. Nairobi: Division of Early Warning and Assessment (DEWA), United Nations Environment Programme. 184 p.
28. Suzuki R, Xu JQ, Motoya K (2006) Global analyses of satellite-derived vegetation index related to climatological wetness and warmth. Int J Climatol 26: 425-438.
29. Myneni RB, Maggion S, Iaquinto J, Privette JL, Gobron N, et al. (1995) Optical remote-sensing of vegetation - modeling, caveats, and algorithms. Remote Sens Environ 51: 169-188.
30. Tucker CJ (1979) Red and photographic infrared linear contributions for monitoring vegetation. Remote Sens Environ 8: 127-150.
31. Hay SI (2000) An overview of remote sensing and geodesy for epidemiology and public health application. Adv Parasitol 47: 1-35.
32. Hay SI, Tatem AJ, Graham AJ, Goetz SJ, Rogers DJ (2006) Global environmental data for mapping infectious disease distribution. Adv Parasitol 62: 37-77.
33. Guerra CA, Hay SI (2005) Remote Sensing: Generalities and Data Products for Malaria Risk Mapping in the Americas. In: Confalonieri UEC, Marinho DP, editors. Remote Sensing and the Control of Infectious Diseases: Proceedings from an Interamerican Workshop. Rio de Janeiro: ENSP/FIOCRUZ. pp. 71-89.
34. Tatem AJ, Goetz SJ, Hay SI (2004) Terra and Aqua: new data for epidemiology and public health. Int J Appl Earth Obs 6: 33-46.
35. Scharlemann J, Benz D, Hay SI, Purse B, Tatem AJ, et al. (2008) Global data for ecology and epidemiology: a novel algorithm for temporal Fourier processing MODIS data. PLoS One: in press.
36. Rogers DJ (2000) Satellites, space, time and the African Trypanosomiases. Adv Parasitol 47: 128-171.



















