
The Involvement of Ca 2+/Calmodulin-Dependent Protein Kinase in Memory Formation in Day-Old Chicks
12
NEUROBIOLOGY OF LEARNING AND MEMORY 66, 24–35 (1996) ARTICLE NO. 0040 The Involvement of Ca 2/ /Calmodulin-Dependent Protein Kinase in Memory Formation in Day-Old Chicks W. Q. ZHAO, P. BENNETT, N. RICKARD, G. L. SEDMAN, M. E. GIBBS AND K. T. NG 1 La Trobe University, Bundoora, Victoria, Australia 3083 (Lynch, Kessler, Arai, & Larson, 1990; Denny, Po- Day-old chicks trained on a single trial passive avoid- lan, Ghuman, Wayner, & Armstrong, 1990; Fazeli, ance learning task showed a significant increase, relative Errington, Dolphin, & Bliss, 1990), through an in- to untrained controls, in activity of the Ca 2/ /calmodulin- crease in intracellular Ca 2/ levels. Among these dependent protein kinase (CaMK) in the particulate frac- Ca 2/ -dependent enzymes, the Ca 2/ /calmodulin-de- tion from tissues from the intermediate medial hyperstria- pendent protein kinase II (CaMK II) is a major class tum ventrale region of the forebrain. The increased kinase of Ca 2/ /calmodulin-dependent protein kinases in the activity was observed within 10 min following training brain that contributes to various neuronal processes, and persisted for at least 70 min posttraining. Amnesia including a number associated with learning and for the task was induced by micromolar concentrations of memory formation. Other kinases, such as protein the specific CAMK II antagonist, KN-62, administered kinase C (PKC) (Nishizuka, 1986; Suzuki, 1994; into the neostriatal/hyperstriatal region of the forebrain. The effect of KN-62 was lateralized. In the right hemi- Zhao, Gibbs, Sedman, & Ng, 1994), cAMP-dependent sphere, KN-62 induced amnesia only when injected within protein kinase (PKA) (Drain, Folders, & Quinn, 2.5 min following training, with memory loss evident by 1991; Frey, Huang, & Kandel, 1993; Huang & Kan- 5 min posttraining. In contrast, in the left hemisphere del, 1994; Suzuki, 1994; Zhao, Polya, Wang, Gibbs, amnesia was induced by KN-62 administered as late as 5 Sedman, & Ng, 1995a; (Zhao, Polya, Wang, Bennett, min posttraining, with onset of amnesia occurring after Gibbs, Sedman, & Ng, 1995b), and tyrosine kinase 10 min posttraining. The findings were interpreted within (Grant, O’Dell, Karl, Stein, Soriano, & Kandel, 1992; the context of a three-stage model of memory formation. Grant & Silva, 1994) are also implicated in learning q 1996 Academic Press, Inc. and memory. CaMK II is abundantly distributed in mammalian INTRODUCTION brain, with a particularly high concentration in areas shown to be involved in memory formation Protein phosphorylation underlies a variety of cel- and in the possibly memory-related phenomenon of lular processes in the central nervous system long-term potentiation (LTP), as well as in areas (Nairn, Hemmings, & Greengard, 1985; Hemmings, rich in glutamate-binding activity (Soderling, Tan, Nairn, McGuinness, Huganir, & Greengard, 1989). McGlade-McCuulloh, Yamamoto, & Fukunaga, Many extracellular messengers trigger intracell- 1994; Erondu & Kennedy, 1985; Quimet, McGuin- ular events by activating Ca 2/ -dependent enzymes, ness, & Greengard, 1984). The finding that the 50- such as protein kinases (Nishizuka, 1986; Suzuki, kDa subunit of CaMK II is identical to the major 1994; Hemmings et al., 1989; Rodnight & Wofchuk, protein found in the postsynaptic density (PSD) 1992), protein phosphatases (Halpain, Girault, & (Kennedy, Bennett, & Erondy, 1983; Kelly, McGuin- Greengard, 1990; Mulkey, Heron, & Malenka, 1993; ness, & Greengard, 1984; Goldenring et al., 1984) Nichols, Suplick, & Brown, 1994), and proteases and is metabolized concomitantly with the other ma- jor proteins of the PSD (Sedman, Jeffrey, Austin, & Rostas, 1986) has led to the suggestion that this ki- 1 This research was supported by a grant from the Australian nase may play an important role in synaptic plastic- Research Council. Address reprint requests to Dr. W. Q. Zhao at ity. CaMK II appears to be involved in the regulation present address: Department of Psychology, Monash University, Clayton, Victoria, Australia 3168. of neurotransmitter release by phosphorylating a 24 1074-7427/96 $18.00 Copyright q 1996 by Academic Press, Inc. All rights of reproduction in any form reserved.
Transcript of The Involvement of Ca 2+/Calmodulin-Dependent Protein Kinase in Memory Formation in Day-Old Chicks

NEUROBIOLOGY OF LEARNING AND MEMORY 66, 24–35 (1996)ARTICLE NO. 0040
The Involvement of Ca2//Calmodulin-Dependent Protein Kinase inMemory Formation in Day-Old Chicks
W. Q. ZHAO, P. BENNETT, N. RICKARD, G. L. SEDMAN, M. E. GIBBS AND K. T. NG1
La Trobe University, Bundoora, Victoria, Australia 3083
(Lynch, Kessler, Arai, & Larson, 1990; Denny, Po-Day-old chicks trained on a single trial passive avoid- lan, Ghuman, Wayner, & Armstrong, 1990; Fazeli,
ance learning task showed a significant increase, relative Errington, Dolphin, & Bliss, 1990), through an in-to untrained controls, in activity of the Ca2//calmodulin- crease in intracellular Ca2/ levels. Among thesedependent protein kinase (CaMK) in the particulate frac- Ca2/-dependent enzymes, the Ca2//calmodulin-de-tion from tissues from the intermediate medial hyperstria-
pendent protein kinase II (CaMK II) is a major classtum ventrale region of the forebrain. The increased kinaseof Ca2//calmodulin-dependent protein kinases in theactivity was observed within 10 min following trainingbrain that contributes to various neuronal processes,and persisted for at least 70 min posttraining. Amnesiaincluding a number associated with learning andfor the task was induced by micromolar concentrations ofmemory formation. Other kinases, such as proteinthe specific CAMK II antagonist, KN-62, administeredkinase C (PKC) (Nishizuka, 1986; Suzuki, 1994;into the neostriatal/hyperstriatal region of the forebrain.
The effect of KN-62 was lateralized. In the right hemi- Zhao, Gibbs, Sedman, & Ng, 1994), cAMP-dependentsphere, KN-62 induced amnesia only when injected within protein kinase (PKA) (Drain, Folders, & Quinn,2.5 min following training, with memory loss evident by 1991; Frey, Huang, & Kandel, 1993; Huang & Kan-5 min posttraining. In contrast, in the left hemisphere del, 1994; Suzuki, 1994; Zhao, Polya, Wang, Gibbs,amnesia was induced by KN-62 administered as late as 5 Sedman, & Ng, 1995a; (Zhao, Polya, Wang, Bennett,min posttraining, with onset of amnesia occurring after Gibbs, Sedman, & Ng, 1995b), and tyrosine kinase10 min posttraining. The findings were interpreted within
(Grant, O’Dell, Karl, Stein, Soriano, & Kandel, 1992;the context of a three-stage model of memory formation.Grant & Silva, 1994) are also implicated in learning
q 1996 Academic Press, Inc.and memory.
CaMK II is abundantly distributed in mammalianINTRODUCTION brain, with a particularly high concentration in
areas shown to be involved in memory formationProtein phosphorylation underlies a variety of cel- and in the possibly memory-related phenomenon of
lular processes in the central nervous system long-term potentiation (LTP), as well as in areas(Nairn, Hemmings, & Greengard, 1985; Hemmings, rich in glutamate-binding activity (Soderling, Tan,Nairn, McGuinness, Huganir, & Greengard, 1989). McGlade-McCuulloh, Yamamoto, & Fukunaga,Many extracellular messengers trigger intracell- 1994; Erondu & Kennedy, 1985; Quimet, McGuin-ular events by activating Ca2/-dependent enzymes, ness, & Greengard, 1984). The finding that the 50-such as protein kinases (Nishizuka, 1986; Suzuki, kDa subunit of CaMK II is identical to the major1994; Hemmings et al., 1989; Rodnight & Wofchuk, protein found in the postsynaptic density (PSD)1992), protein phosphatases (Halpain, Girault, & (Kennedy, Bennett, & Erondy, 1983; Kelly, McGuin-Greengard, 1990; Mulkey, Heron, & Malenka, 1993; ness, & Greengard, 1984; Goldenring et al., 1984)Nichols, Suplick, & Brown, 1994), and proteases and is metabolized concomitantly with the other ma-
jor proteins of the PSD (Sedman, Jeffrey, Austin, &Rostas, 1986) has led to the suggestion that this ki-1 This research was supported by a grant from the Australiannase may play an important role in synaptic plastic-Research Council. Address reprint requests to Dr. W. Q. Zhao atity. CaMK II appears to be involved in the regulationpresent address: Department of Psychology, Monash University,
Clayton, Victoria, Australia 3168. of neurotransmitter release by phosphorylating a
241074-7427/96 $18.00Copyright q 1996 by Academic Press, Inc.All rights of reproduction in any form reserved.
AID NLM 3700 / 6v06$$$141 05-20-96 10:36:10 nlma AP: NLM

25CaMK AND MEMORY FORMATION IN CHICK
presynaptic substrate protein, synapsin I (Llinas, modulin-dependent cellular processes, such as acti-vation of nitric oxide synthase, which also has beenMcGuinness, Leonard, Sugimori, & Greengard,
1985; Llinas, Gruner, Sugimori, McGuinness, & implicated in memory formation using the same be-havioral paradigm (Holscher & Rose, 1993; Rickard,Greengard, 1991; Greengard, Valtorta, Czernik, &
Benfenati, 1993). Injection of CaMK II into presyn- Ng, & Gibbs, 1994).We have shown previously that PKC (Zhao et al.,aptic terminals increased evoked neurotransmitter
release at the squid giant synapse, whereas injection 1994) and PKA (Zhao et al., 1995a,b), as well astyrosine kinase (unpublished results), are involvedof dephosphorylated synapsin I reduced both sponta-
neous and evoked neurotransmitter release at the in different stages of memory formation followingsingle trial passive discriminated avoidance trainingsame synapse (Llinas et al., 1985; Lin, Sugimor, Lli-
nas, McGuinness, & Greengard, 1990). Similar re- in the neonate chick. The role of CaMK is of interestin this context. In this paper we report results fromsults have been reported in vertebrates. Activated
CaMK II in rat brain increased glutamate and nor- studies on the effects of KN62, which has been re-ported to specifically inhibit CaMK II by preventingadrenaline release from nerve terminals, while inhi-
bition of CaMK II depresses the release of these two Ca2/-dependent autophosphorylation of the kinase(Tokumitsu et al., 1990).transmitters (Nichols, Sihra, Czernik, Nairn, &
Greengard, 1990).CaMK II also has been found to be activated dur- METHODS
ing induction of long-term potentiation (for reviews,Animalssee Colbran, 1992; Kelly, 1992; Suzuki, 1994), a pu-
tative cellular mechanism for long-term memoryDay-old black Australorp white Leghorn cockerels(Bliss & Collingridge, 1993). Induction of LTP in the
were obtained from a local hatchery on the morningCA1 region of rat hippocampal slices resulted in anof each experimental day. In the laboratory, theyincrease in CaMK II activity and in the autophos-were housed in pairs in wooden boxes maintainedphorylation of this kinase (Fukunaga, Stoppini,at a temperature of 26–297C, with ad lib. food. AllMiyamoto, & Muller, 1993; Fukunaga, Muller, & Mi-experimental procedures were carried out on pairedyamoto, 1995), while CaMK II inhibitors preventedchicks to avoid the effects of isolation stress (see deLTP formation in rat hippocampal and guinea pigVaus, Gibbs, & Ng, 1980). Twenty different chickssuperior colliculus slices (Malinow, Schulman, &were used for each data point on each day of experi-Tsien, 1989; Malenka, Kauer, Perkel, Mauk, Kelly,mentation. Where a given experiment extended be-Nicoll, & Waxham, 1989; Tokumitsu, Chijiwa, Hagi-yond 1 day, selected data points were replicated towara, Mizutani, Terasawa, & Kidaka, 1990; Ito, Hi-check on possible batch differences. Consequently,daka, & Sugiyama, 1991; Malgaroli, Malinow, Schul-some data points may have sample sizes exceedingman, & Tsien, 1992; Miyamoto & Okada, 1993).20 chicks.Less is known about the involvement of CaMK in
learning and memory. The specific CaMK II inhibi- Behavioral Experimentstor KN-62 administered into the CA1 region of therat hippocampus immediately after step-down pas- Procedures. The training procedures have been
described in detail elsewhere (Gibbs & Ng, 1977).sive avoidance training in rats induced retrogradeamnesia for the task (Wolfman, Fin, Dias, Bianchin, Briefly, chicks were pretrained in pairs to peck at a
chrome bead (2 mm diameter) coated with water andDa Silva, Schmits, Medina, & Izquierdo, 1994). Mu-tant mice missing CaMK II genes (Silva, Stevens, presented twice for as long as necessary to encourage
the chicks to peck freely. These were followed by twoTonegawa, & Wang, 1992a) were shown to be bothunable to establish LTP and deficient in spatial further pretraining trials, the first with a red glass
bead (4 mm diameter) coated with water and pre-learning (Silva, Paylor, Wehner, & Tonegawa,1992b). The calmodulin inhibitor trifluoperazine sented for 10 s and the second a blue glass bead (4
mm diameter), also coated with water and presented(TFP) was reported to induce amnesia following sin-gle trial passive avoidance training in the chick (Ser- for 10 s. The number of pecks and the latency to the
first peck at these beads for each chick were recordedrano, Beniston, Oxonian, Rodriguez, Rosenzweig, &Bennett, 1994). However, this effect cannot be at- on an on-line computer. In a single 10-s training
trial, the chicks were presented with a red bead simi-tributed unequivocally to inhibition of CaMK activ-ity since TFP also depresses PKC activity (Mehta & lar to that used in pretraining, but coated with the
chemical aversant, methylanthranilate (MeA). TheKaul, 1990). Furthermore, interference with theubiquitous calmodulin regulator can affect other cal- number of pecks and latency to first peck were re-
AID NLM 3700 / 6v06$$$142 05-20-96 10:36:10 nlma AP: NLM

26 ZHAO ET AL.
corded on the on-line computer. Chicks that failed ministration experiment, the drug and the vehiclewere given at various times before or after training.to peck this bead or to show typical disgust reactions,
such as head shaking and beak wiping, were ex-cluded from subsequent data analyses. Typically, no Biochemical Experimentmore than 10% of chicks were excluded for this rea-
Behavioral procedures. To measure CaMK II ki-son. In 10-s retention tests, carried out at variousnase activity at different stages of memory forma-times after the training trial, the chicks were pre-tion, 1-day-old chicks were divided into four groupssented with a dry red bead, followed by a dry bluedesigned for different training–sacrifice intervalsbead, similar to those used in the pretraining trials.(immediately and 10, 30, and 70 min after training)Again, the number of pecks and latency to first peckand trained as described above. At each of four train-were recorded. All testing procedures were carrieding–sacrifice interval points six chicks were killedout by an investigator blind to the drug conditionby decapitation and the heads immersed immedi-(see below) for each data point.ately in liquid nitrogen. In order to distinguishMemory was indexed by a discrimination ratiotraining-specific from non-training-related changes(DR), defined as the ratio of the number of pecks ofbrought about by either the pretraining proceduresthe blue bead to the total number of pecks of the redor by stress associated with the aversive taste ofand the blue bead for any chick which pecked at theMeA, four groups of six chicks each were subjected toblue bead on the retention test (Ng & Gibbs, 1991).the pretraining procedures only and killed at timesAll statistical tests were carried out with Type I er-yoked to those of the experimental groups, and threeror rate set at a Å 0.05 and statistical values re-groups of six otherwise naive chicks each were givenported to two decimal place accuracy. Where un-a drop of MeA on the beak and sacrificed immedi-equal sample sizes were deemed to be the result ofately, 30 min, or 70 min after exposure to MeA. Afactors not associated with the treatments, un-group of six naive chicks not exposed to any behav-weighted means analyses were used.ioral manipulation was used to determine restinglevels of kinase activity. The above behavioral exper-Drug administration. 1-(N,O-bis-[5-isoquinol-iment was repeated three times.inesulfonyl]-N-methyl-L-tyrosyl)-4-phenylpipera-
zine (KN-62) was purchased from Calbiochem. MeA Preparation of brain tissue. Since the intermedi-and calmodulin were purchased from Sigma. The ate medial hyperstriatum ventrale (IMHV) isexogenous CaMK II substrate [Arg3]GS1-10 peptide thought to be critical to learning and memory in(Pearson, Woodgett, Cohen, & Kemp, 1985; Klann, the chick (Horn, 1985, 1991), tissue from the IMHVChen, & Sweatt 1991), was purchased from Auspep. rather than from the whole forebrain was used to[g-32P]ATP (3000 Ci/mmol) was purchased from Am- investigate changes in CaMK II activity followingersham. KN-62 was dissolved in demethyl sulfoxide learning. Brain areas were defined according to(Me2SO) first and then diluted with 0.9% NaCl to Youngren and Phillips (1978), by preparing a setvarious concentrations, as required. In each experi- of forebrain slices stained with cresyl violet. Brainment the Me2SO vehicle was used as the control dissection was carried out in a Frigocut cryostat setdrug. KN-62 or the Me2SO vehicle was injected free- at 0207C. Forebrains of chicks were removed andhand into the center of the left, the right, or both sliced from the anterior brain until 110 1 50 mmforebrain hemispheres of chicks in volumes of 10 ml slices were obtained. IMHV tissues from both hemi-per hemisphere using a Hamilton repeating dis- spheres (Fig. 1) of six brains in each instance werepenser syringe, fitted with a 27-gauge needle. A stop pooled for tissue fraction preparation.on the needle regulated injection depth to 3.5 mm,to ensure that the drug was delivered into the neo- Preparation of tissue fractions. Subcellular frac-striatal/hyperstriatal complex, an area of the fore- tions were prepared according to the procedures ofbrain shown to be metabolically active following Kraft and Anderson (1983) and Akers, Lovinger, Col-imprinting (Horn, 1991) and passive avoidance ley, Linden, and Routtenberg (1986). IMHV tissuelearning (Rose & Csillag, 1985; Sedman, O’Dowd, from each group was homogenised in a medium con-Rickard, Gibbs, & Ng, 1991) in the neonate chick. taining 50 mM Tris (pH 7.1), 10 mM EGTA, 2 mMAccuracy of needle placements was checked by ran- EDTA, 0.2 mM phenymethylsulfonyl fluoridedom histological examinations. For the dose re- (PMSF), 1 mg% leupeptin, 0.3% b-mercaptoethanol,sponse and retention function experiments, KN-62 and 0.05% sodium azide. The homogenates were cen-and Me2SO vehicle were administered into chicks trifuged at 100,000g for 30 min, and the supernatant
collected as the soluble (cytosolic) fraction (Kraft etimmediately after training, while, in the time of ad-
AID NLM 3700 / 6v06$$$142 05-20-96 10:36:10 nlma AP: NLM

27CaMK AND MEMORY FORMATION IN CHICK
liability and the resulting counts averaged acrossreplicates.
Data analyses. Possible batch differences in iso-topes and substrates, and random factors affectingexperiments carried out on different days, makecomparisons of raw phosphorylation data not partic-ularly meaningful. Thus, scintillation counting val-ues for CaMK II activity in soluble and particulatefractions from each pooled sample from the experi-mental and pretraining-yoked groups were aver-aged, and converted to percentage of the meanCaMK II activity of the corresponding fraction fromthe sample from the naive control group, an ap-proach usually used in most correlational studies(see for example, Klann et al., 1991; Fukunaga etal., 1993). Data from four independently repeatedFIG. 1. A outline diagram of coronal sections of the forebrainbehavioral experiments for each group were sub-of a day-old chick. The position of the IMHV region was definedjected to statistical analyses. Data from trained ani-according to Youngren et al. (1978). HV, hyperstriarum accessor-
ium; IMHV, intermediate medial hyperstriatum ventrale; N, neo- mals were compared with those of pretrained onlystriatum. animals using two-way ANOVAs. Data from the
chicks exposed only to MeA were subjected to one-way ANOVAs.
al., 1983; Akers et al., 1986). The pellets werewashed with and resuspended in the same medium RESULTScontaining 0.5% (v/v) Triton X-100. After 20 min onice, the samples were centrifuged at 100,000g for 30 Effects of Training on CaMK II Activitymin. The supernatant from this step was collected
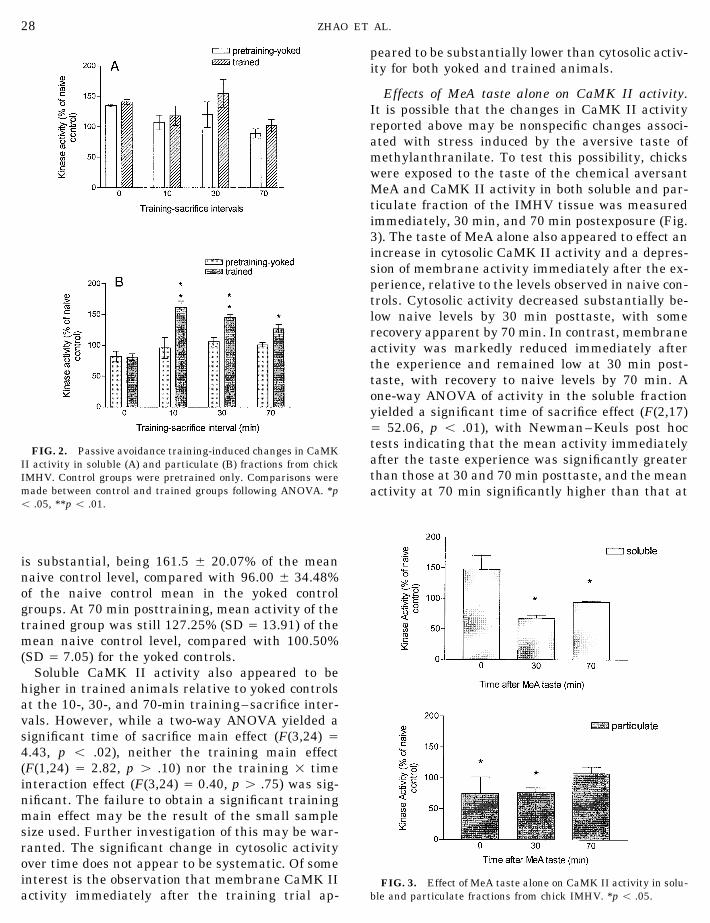
Training-induced changes in CaMK II activity.as the particulate (membrane) fraction (Kraft et al.,When CaMK II activity in IMHV tissue was mea-1983; Akers et al., 1986). Protein concentrationssured using the Pro-Leu-Ser-Arg-Thr-Leu-Ser-Val-were measured by a modified Lowry method (1951)Ser-Ser-amide peptide in the absence of Ca2//cal-using Bio-Rad DC protein assay reagents.modulin, there were no apparent differences be-tween trained animals and naive controls (data notCaM kinase II assays. CaMK II activity was as-
sayed using the exogenous CaMK II substrate, Pro- shown). However, changes in CaMK II activity fol-lowing training were detected when this activity wasLeu-Ser-Arg-Thr-Leu-Ser-Val-Ser-Ser-amide peptide
(Pearson et al., 1985; Klann et al., 1991). The total measured in the presence of Ca2//calmodulin (Fig.2). Activity in the particulate fraction of yoked con-reaction medium of 100 ml vol contained 50 mM
Tris–HCl buffer, 0.5 mM EGTA, 0.5 mM EDTA, 1 trols was maintained at the level of naive controlsat all training–sacrifice intervals. In contrast, activ-mM Mg2/, 40 mM ATP, 50 mM Pro-Leu-ser-Arg-Thr-
Leu-ser-Val-Ser-Ser-amide peptide, and 2.5 mCi ity for trained animals showed a substantial rise by10 min following training, with the increased activ-[g-32P]ATP, with or without 1.5 mM Ca2/ and 1 mg/
ml calmodulin. Reactions were initiated by adding ity maintained up to 70 min posttraining, the longesttraining–sacrifice interval investigated. A two-way3.2 mg (10 ml) protein of tissue fractions to yield a
final concentration of 32 mg/ml protein. Companion ANOVA yielded significant training (F(1,24) Å26.50, p õ .01) and time (F(3,24) Å 11.59, p õ .01)reactions for each sample were carried out in the
absence of Ca2//calmodulin. Reaction mixtures were main effects, and a significant training 1 time inter-action effect (F(3,24) Å 4.79, p õ .01). Simple mainincubated at 307C for 15 min and were terminated
by spotting duplicates of 30 ml of the mixture on 1.5 effects analyses showed a significant variation of ac-tivity over time for the trained (p õ .01) but not1 1.5 cm phosphocellulose filter squares (Whatman,
P-81), which were subsequently washed three times the yoked control groups, and significant differencesbetween trained and yoked control groups at thein 75 mM H3PO4, followed by two washes in 100%
ethanol. Squares were dried with a hair drier and 10-, 30-, and 70-min training–sacrifice intervals (allp õ .05), but not immediately after training. Thecounted in a scintillation counter. All assays were
carried out in duplicate or triplicate to check for re- increase in membrane activity 10 min posttraining
AID NLM 3700 / 6v06$$$142 05-20-96 10:36:10 nlma AP: NLM
28 ZHAO ET AL.
peared to be substantially lower than cytosolic activ-ity for both yoked and trained animals.
Effects of MeA taste alone on CaMK II activity.It is possible that the changes in CaMK II activityreported above may be nonspecific changes associ-ated with stress induced by the aversive taste ofmethylanthranilate. To test this possibility, chickswere exposed to the taste of the chemical aversantMeA and CaMK II activity in both soluble and par-ticulate fraction of the IMHV tissue was measuredimmediately, 30 min, and 70 min postexposure (Fig.3). The taste of MeA alone also appeared to effect anincrease in cytosolic CaMK II activity and a depres-sion of membrane activity immediately after the ex-perience, relative to the levels observed in naive con-trols. Cytosolic activity decreased substantially be-low naive levels by 30 min posttaste, with somerecovery apparent by 70 min. In contrast, membraneactivity was markedly reduced immediately afterthe experience and remained low at 30 min post-taste, with recovery to naive levels by 70 min. Aone-way ANOVA of activity in the soluble fractionyielded a significant time of sacrifice effect (F(2,17)Å 52.06, p õ .01), with Newman–Keuls post hoctests indicating that the mean activity immediately
FIG. 2. Passive avoidance training-induced changes in CaMKafter the taste experience was significantly greaterII activity in soluble (A) and particulate (B) fractions from chickthan those at 30 and 70 min posttaste, and the meanIMHV. Control groups were pretrained only. Comparisons were
made between control and trained groups following ANOVA. *p activity at 70 min significantly higher than that atõ .05, **p õ .01.
is substantial, being 161.5 { 20.07% of the meannaive control level, compared with 96.00 { 34.48%of the naive control mean in the yoked controlgroups. At 70 min posttraining, mean activity of thetrained group was still 127.25% (SD Å 13.91) of themean naive control level, compared with 100.50%(SD Å 7.05) for the yoked controls.
Soluble CaMK II activity also appeared to behigher in trained animals relative to yoked controlsat the 10-, 30-, and 70-min training–sacrifice inter-vals. However, while a two-way ANOVA yielded asignificant time of sacrifice main effect (F(3,24) Å4.43, p õ .02), neither the training main effect(F(1,24) Å 2.82, p ú .10) nor the training 1 timeinteraction effect (F(3,24) Å 0.40, p ú .75) was sig-nificant. The failure to obtain a significant trainingmain effect may be the result of the small samplesize used. Further investigation of this may be war-ranted. The significant change in cytosolic activityover time does not appear to be systematic. Of someinterest is the observation that membrane CaMK II FIG. 3. Effect of MeA taste alone on CaMK II activity in solu-
ble and particulate fractions from chick IMHV. *p õ .05.activity immediately after the training trial ap-
AID NLM 3700 / 6v06$$$143 05-20-96 10:36:10 nlma AP: NLM
29CaMK AND MEMORY FORMATION IN CHICK
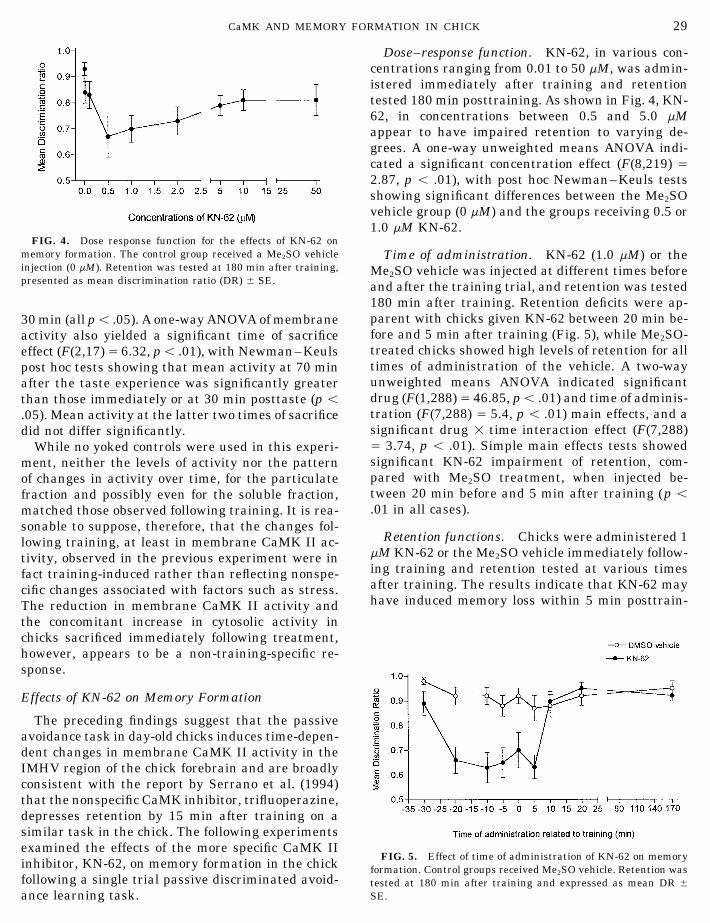
Dose–response function. KN-62, in various con-centrations ranging from 0.01 to 50 mM, was admin-istered immediately after training and retentiontested 180 min posttraining. As shown in Fig. 4, KN-62, in concentrations between 0.5 and 5.0 mMappear to have impaired retention to varying de-grees. A one-way unweighted means ANOVA indi-cated a significant concentration effect (F(8,219) Å2.87, p õ .01), with post hoc Newman–Keuls testsshowing significant differences between the Me2SOvehicle group (0 mM) and the groups receiving 0.5 or1.0 mM KN-62.
FIG. 4. Dose response function for the effects of KN-62 onmemory formation. The control group received a Me2SO vehicle Time of administration. KN-62 (1.0 mM) or theinjection (0 mM). Retention was tested at 180 min after training, Me2SO vehicle was injected at different times beforepresented as mean discrimination ratio (DR) { SE. and after the training trial, and retention was tested
180 min after training. Retention deficits were ap-parent with chicks given KN-62 between 20 min be-30 min (all põ .05). A one-way ANOVA of membranefore and 5 min after training (Fig. 5), while Me2SO-activity also yielded a significant time of sacrificetreated chicks showed high levels of retention for alleffect (F(2,17)Å 6.32, põ .01), with Newman–Keulstimes of administration of the vehicle. A two-waypost hoc tests showing that mean activity at 70 minunweighted means ANOVA indicated significantafter the taste experience was significantly greaterdrug (F(1,288)Å 46.85, põ .01) and time of adminis-than those immediately or at 30 min posttaste (p õtration (F(7,288) Å 5.4, p õ .01) main effects, and a.05). Mean activity at the latter two times of sacrificesignificant drug 1 time interaction effect (F(7,288)did not differ significantly.Å 3.74, p õ .01). Simple main effects tests showedWhile no yoked controls were used in this experi-significant KN-62 impairment of retention, com-ment, neither the levels of activity nor the patternpared with Me2SO treatment, when injected be-of changes in activity over time, for the particulatetween 20 min before and 5 min after training (p õfraction and possibly even for the soluble fraction,.01 in all cases).matched those observed following training. It is rea-
sonable to suppose, therefore, that the changes fol-Retention functions. Chicks were administered 1lowing training, at least in membrane CaMK II ac-
mM KN-62 or the Me2SO vehicle immediately follow-tivity, observed in the previous experiment were ining training and retention tested at various timesfact training-induced rather than reflecting nonspe-after training. The results indicate that KN-62 maycific changes associated with factors such as stress.have induced memory loss within 5 min posttrain-The reduction in membrane CaMK II activity and
the concomitant increase in cytosolic activity inchicks sacrificed immediately following treatment,however, appears to be a non-training-specific re-sponse.
Effects of KN-62 on Memory Formation
The preceding findings suggest that the passiveavoidance task in day-old chicks induces time-depen-dent changes in membrane CaMK II activity in theIMHV region of the chick forebrain and are broadlyconsistent with the report by Serrano et al. (1994)that the nonspecific CaMK inhibitor, trifluoperazine,depresses retention by 15 min after training on asimilar task in the chick. The following experimentsexamined the effects of the more specific CaMK II
FIG. 5. Effect of time of administration of KN-62 on memoryinhibitor, KN-62, on memory formation in the chick formation. Control groups received Me2SO vehicle. Retention wasfollowing a single trial passive discriminated avoid- tested at 180 min after training and expressed as mean DR {
SE.ance learning task.
AID NLM 3700 / 6v06$$$143 05-20-96 10:36:10 nlma AP: NLM

30 ZHAO ET AL.
FIG. 7. Lateralization of the effects of KN-62 on memorystages. KN-62 (1 mM) was unilaterally injected into either the leftor the right hemisphere immediately after training. RetentionFIG. 6. Effect of KN-62 on retention functions. Control groupswas tested at various times following training and expressed aswere given the Me2SO vehicle. Retention was expressed as meanmean DR { SE.DR { SE.
sphere 1 time interaction effect (F(6,323) Å 2.69, ping, with the loss persisting at least until 180 minõ .02). Simple main effects analyses confirmed thatposttraining (Fig. 6). Consistent with this, a two-only the left hemisphere showed significant time ofway unweighted means ANOVA yielded a significanttest variations (p õ .01) and that significant differ-drug main effect (F(1,427) Å 98.15, p õ .01), but noences between hemispheres were present only at 5significant time main effect (F(8,427) Å 1.19, p úand 10 min posttraining (both p õ .01). Two control.30) or drug 1 time interaction effect (F(8,427) Ågroups injected with the Me2SO vehicle immediately0.71, pÅ .68). Thus, KN-62 appears to have impairedafter training, into the left or the right hemisphere,memory processing within 5 min following learning.yielded mean DRs of 0.92 (SD Å 0.13) and 0.94 (SDÅ .10), respectively, at 180 min posttraining, con-Lateralization of Effects of KN-62firming that KN-62 injected into either hemisphereyielded substantial memory loss at this time.Protein kinase C inhibitors, such as melittin, have
been reported to bring about amnesia only when ad-Time of administration functions for unihemi-ministered in the left hemisphere of the chick fore-
spheric application of KN-62. KN-62 (1 mM) wasbrain (Burchuladze, Potter, & Rose, 1990; Zhao etadministered at various times within 10 min afteral., 1994). Rose (1991) has suggested that memorytraining into the left or the right hemisphere, andtraces may not be stably located in a single neuronalretention was tested 180 min posttraining (Fig. 8).ensemble, but may be dynamic and fluid, movingThe results suggest that KN-62 was effective in in-from site to site within the brain and possibly acrossducing amnesia when administered into left hemi-hemispheres. The following experiments examined
possible lateralization of the effects of KN-62.
Retention functions following unihemispheric ap-plication of KN-62. KN-62 (1 mM) was adminis-tered into the left or the right hemisphere immedi-ately after training and retention was tested at vari-ous times following training (Fig. 7). The resultssuggest that KN-62 is capable of inducing amnesiawhen administered into either chick forebrain hemi-sphere. However, retention loss occurred within 5min following training when KN-62 was applied tothe right hemisphere, but only by 20 min followingtraining when the drug was administered into theleft hemisphere. A two-way unweighted means AN-OVA indicated significant hemisphere (F(1,323) Å FIG. 8. Time of administration function for KN-62 given to25.75, p õ .01) and time of test (F(6,323) Å 2.89, p the left or the right hemisphere. Retention was tested at 180 min
after training and presented as mean DR { SE.õ .01) main effects, as well as a significant hemi-
AID NLM 3700 / 6v06$$$143 05-20-96 10:36:10 nlma AP: NLM

31CaMK AND MEMORY FORMATION IN CHICK
sphere up to at least 5 min posttraining, but not mixture. It may be possible also that translocationmechanism of CaMK II is not involved in memorywhen given at 10 min posttraining. In sharp con-
trast, right hemispheric application of KN-62 was formation.In contrast to the marked increase in CaMK IIeffective only when carried out immediately after
training, with no apparent retention loss when the in particulate fractions following training, in chicksexposed to the taste of MeA alone, CaMK II activitydrug was given 2.5 min or later following training.
An unweighted means two-way ANOVA yielded sig- was suppressed immediately and 30 min after expo-sure to the aversant. The explanation for this is notnificant hemisphere (F(1,142) Å 9.91, p õ .01) and
time of administration (F(3,142) Å 5.61, p õ .01) obvious, although it is interesting to note that thetaste of MeA alone has been shown also to depressmain effects and a significant hemisphere 1 time
interaction effect (F(3,142) Å 4.14, p õ .01). Simple Na//K/ ATPase activity in neonate chicks (Hajek,Sykova, Sedman, & Ng, 1994). In any event, it ismain effects analyses confirmed significant changes
in mean retention levels across times of administra- clear that the increase in CaMK II activity observedin the particulate fraction following the trainingtion for both hemispheres (p õ .02) and significant
differences between hemispheres only for the 2.5- trial was learning-induced.Although Ca2/-independent CaMK II activity hasand 5-min times of administration.
been shown previously to be increased in severaltypes of in vivo systems following depolarization (seeDISCUSSIONColbran, 1992) and LTP (Fukunaga et al., 1993),none was detected in the present study. Klann et al.In the present study, phosphorylation of the Pro-
Leu-Ser-Arg-Thr-Leu-Ser-Val-Ser-Ser-amide pep- (1991) also have reported a marked elevation of PKCactivity following LTP, but no significant Ca2/-inde-tide was significantly elevated in tissue from the
IMHV region of the chick forebrain following the pendent CaMK II activity. It has been found thatCa2/-independent CaMK II activity observed in dif-training experience. Relative to naive controls, and
in the presence of Ca2//calmodulin, CaMK II activity ferent cell types does not persist for more than a fewminutes due to high cellular phosphatase activityin the particulate fraction was significantly lower
than that in the soluble fraction immediately (0 min) (Colbran, 1992). It is possible that any autophos-phorylated CaMK II resulting from the learning ex-after training, with cytosolic levels substantially ele-
vated in trained animals over naive controls. These perience in the present study may have been dephos-phorylated during tissue preparation since a phos-changes may be attributed to nonspecific factors as-
sociated with the behavioral procedures, since they phatase inhibitor was not included in the buffers.It has been suggested also that Ca2/-independentwere detected also in chicks exposed only to the pre-
training trials or to the taste alone of the chemical CaMK II activity represents only a small fractionof CaMK activity in the brain (Richard Rodnight,aversant, MeA. On the other hand, in the particulate
fraction, CaMK II activity in trained chicks in- personal communication) and may not be detectablein our assay system. Only crude brain cytosolic orcreased sharply over the levels observed in pre-
trained control chicks by 10 min posttraining, with membrane fractions, rather than purified CaMK II,were added to the reactions in our assays. These willthis increase more or less maintained through to at
least 70 min posttraining. Although translocation of contain various CaMKs in addition to CaMK II, aswell as various proteins which may be phosphory-CaMK II across subcellular compartments has been
reported previously in brain tissues of animals sub- lated by these CaMKs in the presence of Ca2//cal-modulin. The effect would be to mask the smalljected to non-learning-related conditions such as
ischemia (Aronowski, Grotta, & Waxham, 1992; amount of Ca2/-independent CaMK II activity. Pos-sible changes in autophosphorylation of CaMK II asKolb, Hudman, & Waxham, 1995; Hu & Wieloch,
1995; Morioka, Fukunaga, Nagahiro, Kurino, Us- a consequence of learning may nonetheless be im-portant in memory processing. We are currently in-hio, & Miyamoto, 1995), hypoglycemia (Hu, Kuri-
hara, & Wieloch, 1995), and status epilepticus vestigating these. For the present, the fact that,when assayed in the presence of Ca2//calmodulin,(Wasterlain, Bronstern, Morin, Dwyer, & Sankar,
1992), there was no evidence of such a translocation persistently high CaMK II activity was observed inthe IMHV tissue of trained animals within 10 minin the present study, cytosolic CaMK II activity re-
maining essentially unchanged. It may be possible of training indicates that a certain form of activatedCaMK II was generated as a result of training, andthat decreased CaMK II activity in the cytosol is
masked by Ca2//calmodulin-dependent phosphoryla- the fact that amnesia was induced shortly aftertraining by KN-62 confirms a role for CaMK II intion of other brain proteins present in the reaction
AID NLM 3700 / 6v06$$$143 05-20-96 10:36:10 nlma AP: NLM

32 ZHAO ET AL.
early memory processes (Silva et al., 1992b; Wolf- While the findings do not rule out a role for CaMKII in LTP-related memory processes (Bliss & Col-man et al., 1994; Serrano et al., 1994).
The behavioral studies reported here clearly show lingridge, 1993; Silva et al., 1992a,b; Malinow et al.,1989; Malenka et al., 1989; Ito et al., 1991), the am-that bihemispheric administration of KN-62 into the
hyperstriatal/neostriatal region of the chick fore- nestic effects of KN-62 cannot be attributed solelyto interference with LTP induction since onset ofbrain induced substantial retention deficits within 5
min following learning, provided the injections were amnesia occurs considerably earlier than has beenobserved in an identical behavioral paradigm withgiven shortly after training. The time of onset of
amnesia achieved with KN-62 differs from that re- bihemispheric blockade of metabotropic, NMDA, orAMPA glutamate receptors. These latter treatmentsported for trifluoperazine (TFP; Serrano et al., 1994),
for which onset occurred after 15 min posttraining. resulted in amnesia around 90 min posttraining, in-dependent of the time of administration of the drugsWe have unpublished results confirming Serrano et
al.’s finding. The reason for the discrepancy between (Rickard et al., 1994; Rickard & Ng, 1995).While the results from bihemispheric injections ofKN-62 and TFP in the time of amnesia onset is un-
clear. However, TFP is not a specific CaM kinase KN-62 suggest that CaMK II activity is implicatedin memory processing from as early as 5 min afterinhibitor with inhibitory effect also on PKC activity
as well as numerous other calmodulin-dependent training, this appears to be due entirely to the effectsof inhibiting CaMK II activity in the right hemi-proteins and enzymes in both the brain and non-
nervous system (Weiss, Prozialeck, Cimino, Bar- sphere. KN-62 in the left hemisphere resulted in am-nesia onset only after 10 min following learning.nette, & Wallace, 1980; VanBelle, 1984; Motohashi,
1991; Mittal & Jadhav, 1994; Mehta & Kaul, 1990). This hemispheric dissociation of the effects of KN-62 is reflected also in the time of administrationTherefore, the amnesia induced by TFP (Serrano et
al., 1994) may not be solely attributed to the inhibi- functions, with right hemisphere injections being ef-fective only when given before 2.5 min posttrainingtion of CaMK II. We now have evidence that TFP
interferes with different stages of memory formation and left hemisphere injections inducing amnesiawhen administered as late as 5 min posttraining.depending on the time of administration (unpub-
lished data). In contrast, KN-62 has been reported to Although there is no direct evidence indicatingwhether KN-62 diffuses across hemispheres, previ-specifically inhibit CaMK II activity and to prevent
Ca2/-dependent autophosphorylation of CaMK II, ous reports with a different drug has shown littleinterhemispheric diffusion following intracranial ad-without affecting other kinases such as PKA and
PKC (Tokumitsu et al., 1990). In the present study, ministration (Gibbs, Richdale, & Ng, 1979). In addi-tion, that inhibitors of PKC (Burchuladze et al.,KN-62 significantly inhibited memory formation at
micromolar concentrations. 1990; Zhao et al., 1994) and PKA (Zhao et al., 1995b)induce amnesia only when administered into the leftThe fact that KN-62 had to be administered no
later than 5 min posttraining to be effective is consis- hemisphere of the chick brain also suggests thatthese drugs do not diffuse across hemispheres. It hastent with an earlier report that KN-62 was most effec-
tive in inducing retrograde amnesia in the rat when been suggested that the left and right hemispheresof the chick may have differentiated roles in memorygiven immediately after training (Wolfman et al.,
1994). KN-62 has been demonstrated to have no in- processing (Rose, 1991; McCabe, 1991; Andrew,1991). The present evidence with KN62, and thosehibitory effects on already autophosphorylated and
Ca2//calmodulin-independent CaMK II (Tokumitsu reported previously for other kinases and other am-nestic agents (Serrano, Ramus, Bennett, & Rosen-et al., 1990). It has been shown that once autophos-
phorylated in the presence of Ca2//calmodulin, CaMK zweig, 1992), is consistent with this view and under-scores Rose’s (1991) caution against assuming a lin-II becomes persistently and constitutively active and
is no longer dependent on Ca2//calmodulin (Colbran, ear sequence of memory processes in a singleneuronal assembly.1992; Miller & Kennedy, 1986; Lai, Nairn, &
Greengard, 1986). This switch-like phenomenon hasencouraged speculation that CaMK II may be an ideal REFERENCESmolecular mechanism underlying memory storage in
Akers, R. F., Lovinger, D. C., Colley, P. E., Linden, D. J., & Rout-the brain (Lisman, 1994; Lisman & Goldering, 1988).tenberg, A. (1986). Translocation of protein kinase C activity
It is plausible to suggest that memory mechanisms may mediate hippocampal long-term potentiation. Science,early after training may be dependent on an initial 231, 587–589.Ca2/-dependent autophosphorylation of CaMK II sus- Andrew, L. J. (1991). The nature of behavioural lateralization in
the chick. In R. J. Andrew (Ed.), Neural and behaviouralceptible to blockade by KN-62.
AID NLM 3700 / 6v06$$$144 05-20-96 10:36:10 nlma AP: NLM

33CaMK AND MEMORY FORMATION IN CHICK
plasticity: The use of the domestic chicken as a model & Kandel, E. R. (1992). Impaired long-term potentiation, spa-tial learning, and hippocampal development in fyn mutant(pp. 536–554). Oxford: Oxford Univ. Press.mice. Science, 258, 1903–1910.Aronowski, J., Grotta, J. C., & Waxham, M. N. (1992). Ischaemia-
induced translocation of Ca2//calmodulin-dependent protein Grant, S. G., & Silva, A. J. (1994). Targeting learning. Trends inkinase II: potential role in neuronal damage. Journal of Neu- Neuroscience, 17, 71–75.rochemistry, 58, 1743–1753. Greengard, P., Valtorta, F., Czernik, A. J., & Benfenati, F. (1993).
Bliss, T. V. P., & Collingridge, G. C. (1993). A synaptic model of Synaptic vesicle phosphoproteins and regulation of synapticmemory: Long-term potentiation in the hippocampus. Na- function. Science, 259, 780–785.ture, 361, 31–39. Hajek, I., Sykova, E., Sedman, G., & Ng, K. T. (1994). Na/,
Burchuladze, R., Potter, J., & Rose, S. P. R. (1990). Memory for- K/-ATPase activity in young chicks after taste stimulation.mation in the chick depends on membrane-bound protein ki- Brain Research Bulletin, 33, 87–91.nase C. Brain Research, 535, 131–138. Halpain, S., Girault, J. A., & Greengard, P. (1990). Activation of
Colbran, R. J. (1992). Regulation and role of brain calcium/cal- NMDA receptors induces dephosphorylation of DARPP-32 inmodulin-dependent protein kinase II. Neurochemistry Inter- rat striatal slices. Nature, 343, 369–372.national, 21, 469–497. Hemmings, H. C., Jr., Nairn, A. C., McGuinness, T. L., Huganir,
Denny, J. B., Polan, C. J., Ghuman, A., Wayner, M. J., & Arm- R. L., & Greengard, P. (1989). Role of protein phosphorylationstrong, D. L. (1990). Calpain inhibitors block long-term poten- in neuronal signal transduction. FASEB Journal, 3, 1538–tiation. Brain Research, 534, 317–320. 1592.
de Vaus, J. E., Gibbs, M. E., & Ng, K. T. (1980). Effect of social Holscher, C., & Rose, S. P. (1993). Inhibiting synthesis of theisolation on memory formation. Behavioral and Neural Biol- putative retrograde messenger nitric oxide results in amne-ogy, 29, 473–480. sia in a passive avoidance task in the chick. Brain Research,
619, 89–94.Drain, P., Folders, E., & Quin, W. G. (1991). cAMP-dependentprotein kinase and the disruption of learning in transgenic Horn, G. (1985). Memory, imprinting, and the brain. Oxford:flies. Neuron, 6, 71–82. Clarendon Press.
Erondu, N. E., & Kennedy, M. B. (1985). Regional distribution of Horn, G. (1991). Imprinting and recognition memory: A reviewtype II Ca2/ calmodulin-dependent protein kinase in rat of neural mechanisms. In R. J. Andrew (Ed.), Neural andbrain. Journal of Neuroscience, 5, 3270–3277. behavioural plasticity: The use of the domestic chick as a
model (pp. 219–261). Oxford: Oxford Univ. Press.Fazeli, M. S., Errington, M. L., Dolphin, A. C., & Bliss, T. V.(1990). Increase efflux of a haemoglobin-like protein and an Hu, B. R., Kurihara, J., & Wieloch, T. (1995). Persistent transloca-80 Kda protease into push-pull perfusates following the in- tion and inhibition of Ca2//calmodulin-dependent protein ki-duction of long-term potentiation in the dentate gyrus. Brain nase II in the crude synaptosomal fraction of the vulnerableResearch, 521, 247–253. hippocampus following hypoglycemia. Journal of Neurochem-
istry, 64, 1363–1369.Frey, U., Huang, Y. Y., & Kandel, E. R. (1993). Effects of cAMPsimulate a late stage of LTP in hippocampal CA1 neurones. Hu, B. R., & Wieloch, T. (1995). Persistent translocation ofScience, 260, 1661–1664. Ca2//calmodulin-dependent protein kinase II to synaptic
junctions in the vulnerable hippocampal CA1 region follow-Fukunaga, K., Rich, D. P., & Soderling, T. R. (1989). Generationing transient ischaemia. Journal of Neurochemistry, 64, 277–of the Ca2/-independent from of Ca2//calmodulin-dependent284.protein kinase II in cerebellar granule cells. Journal of Bio-
logical Chemistry, 264, 830–836. Huang, Y. Y., & Kandel, E. R. (1994). Recruitment of long-lastingand protein kinase A-dependent long-term potentiation in theFukunaga, K., Stoppini, L., Miyamoto, E., & Muller, D. (1993).CA1 region of hippocampus requires repeated tetanisation.Long-term potentiation is associated with an increased activ-Learning and Memory, 1, 74–82.ity of Ca2//calmodulin-dependent protein kinase II. Journal
of Biochemistry, 268, 7863–7867. Ito, I., Hidaka, H., & Sugiyama, H. (1991). Effects of KN-62, aspecific inhibitor of calcium/calmodulin-dependent protein ki-Fukunaga, K., Muller, D., & Miyamoto, E. (1995). Increased phos-nase II, on long-term potentiation in the rat hippocampus.phorylation of Ca2//calmodulin-dependent protein kinase IINeuroscience Letters, 121, 119–121.and its endogenous substrates in the induction of long-term
potentiation. Journal of Biological Chemistry, 270, 6119– Kelly, P. T. (1992). Calmodulin-dependent protein kinase II. Mo-6124. lecular Neurobiology, 5, 153–177.
Gibbs, M. E., & Ng, K. T. (1977). Psychology of memory: Towards Kelly, P. T., McGuinness, T. L., & Greengard, P. (1984). Evidencea model of memory formation. Biobehavioral Reviews, 1, 113– that the major postsynaptic density protein is a component136. of a Ca2//calmodulin-dependent protein kinase. Proceedings
of the National Academy of Science, USA, 81, 945–949.Gibbs, M. E., Richdale, A. L., & Ng, K. T. (1979). Biochemicalaspects of protein synthesis inhibition by cycloheximide in Kennedy, M. B., Bennett, M. K., & Erondu, N. E. (1983). Biochem-one or both hemispheres of the chick brain. Pharmacology, ical and immunochemical evidence that the major postsynap-Biochemistry and Behavior, 10, 929–931. tic density protein is a subunit of a calmodulin-dependent
kinase. Proceedings of the National Academy of Science, USA,Goldenring, J. R., McGuire, J. S., & Delorenzo, R. J. (1984). Identi-80, 7357–7361.fication of the major postsynaptic density protein as homolo-
gous with the major calmodulin-binding subunit of a calmod- Klann, E., Chen, S. J., & Sweatt, J. D. (1991). Persistent proteinulin-dependent protein kinase. Journal of Neurochemistry, kinase activation in the maintenance phase of long-term po-42, 1077–1084. tentiation. Journal of Biological Chemistry, 266, 24253–
24256.Grant, S. G., O’Dell, T. J., Karl, K. A., Stein, P. L., Soriano, P.,
AID NLM 3700 / 6v06$$$144 05-20-96 10:36:10 nlma AP: NLM

34 ZHAO ET AL.
Kolb, S. J., Hudmon, A., & Waxham, M. N. (1995). Ca2//calmodu- Mittal, C. K., & Jadhav, A. L. (1994). Calcium-dependent inhibi-tion of constitutive nitric oxide synthase. Biochemical andlin kinase II translocates in a hippocampal slice model of
ischaemia. Journal of Neurochemistry, 64, 2147–2156. Biophysical Research Communications, 203, 8–15.
Kraft, A. S., & Anderson, W. B. (1983). Phorbol esters increase Miyamoto, T., & Okada, Y. (1993). NMDA receptor, protein kinasethe amount of Ca2/, phospholipid-dependent protein kinase C and calmodulin system participate in the long-term poten-associated with plasma membrane. Nature, 301, 621–623. tiation in guinea pig superior colliculaus slices. Brain Re-
search, 605, 287–292.Lai, Y., Nairn, A. C., & Greengard, P. (1986). Autophosphoryla-tion reversibly regulates the Ca2//calmodulin-dependence of Morioka, M., Fukunaga, K., Nagahiro, S., Kurino, M., Ushio, Y.,Ca2//calmodulin-dependent protein kinase II. Proceedings of & Miyamoto, E. (1995). Glutamate-induced loss of Ca2//cal-the National Academy of Science, USA, 83, 4253–4257. modulin-dependent protein kinase II activity in cultured rat
hippocampal neurones. Journal of Neurochemistry, 64, 2132–Lin, J. W., Sugimori, M., Llinas, R. R., McGuinness, T. L.,2139.& Greengard, P. (1990). Effects of synapsin I and calcium/
calmodulin-dependent protein kinase II on spontaneous neu- Motohashi, N. (1991). Phenothiazines and calmodulin. Anticancerrotransmitter release in the squid giant synapse. Proceedings Research, 11, 1125–1164.of the National Academy of Science, USA, 87, 8257–8261. Mulkey, R. M., Heron, C. E., & Malenka, R. C. (1993). An essential
Lisman, J. E. (1994). The CaM kinase II hypothesis for the storage role for protein phosphatase in hippocampal long-term de-of synaptic memory. Trends in Neuroscience, 17, 406–412. pression. Science, 261, 1051.
Lisman, J. E., & Goldering, M. (1988). Evaluation of a model of Nairn, A. C., Hemmings, H. C., Jr., & Greengard, P. (1985). Pro-long-term memory based on the properties of the tein kinase in the brain. Annual Review of Biochemistry, 54,Ca2//calmodulin-dependent protein kinase. Journal of Physi- 931–976.ology, 83, 187–189. Ng, K. T., & Gibbs, M. E. (1988). A biological model for memory
Llinas, R. R., McGuinness, T. L., Leonard, C. S., Sugimori, M., formation. In H. J. Markowitsch (Ed.), Information pro-& Greengard, P. (1985). Intraterminal injection of synapsin cessing by the brain (pp. 151–178). Toronto: Hans Huber.I or calcium/calmodulin-dependent protein kinase II alters Ng, K. T., & Gibbs, M. E. (1991). Stages in memory formation: aneurotransmitter release at the squid giant synapse. Proceed- review. In R. J. Andrew (Ed.), Neural and behavioural plas-ings of the National Academy of Science, USA, 82, 3035– ticity: The use of the domestic chick as a model (pp. 351–369).3039. Oxford: Oxford Univ. Press.
Llinas, R. R., Gruner, J. A., Sugimori, M., McGuinness, T. L.,Nichols, R. A., Sihra, T. S., Czernik, A. J., Nairn, A. C.,
& Greengard, P. (1991). Regulation by synapsin I and Ca2/-& Greengard, P. (1990). Calcium/calmodulin-dependent pro-
calmodulin-dependent protein kinase II of the transmittertein kinase II increases glutamate and noradrenaline release
release in squid giant synapse. Journal of Physiology, 436,from synaptosomes. Nature, 343, 647–651.
257–282.Nichols, R. A., Suplick, G. R., & Brown, J. M. (1994). Calcineurin-
Lowry, O. H., Rosebrough, N. J., Farr, A. L., & Randall, R. J.mediated protein dephosphorylation in brain nerve terminals
(1951). Protein measurement with the folin phenol reagent.regulates the release of glutamate. Journal of Biological
Journal of Biological Chemistry, 193, 265–275.Chemistry, 269, 23817–23823.
Lynch, G., Kessler, M., Arai, A., & Larson, J. (1990). The natureNishizuka, Y. (1986). Studies and perspectives of protein kinaseand causes of hippocampal long-term potentiation. Progress
C. Science, 233, 305–312.in Brain Research, 83, 233–250.Pearson, R. B., Woodgett, J. R., Cohen, P., & Kemp, B. E. (1985).McCabe, B. J. (1991). Hemispheric asymmetry of learning-in-
Substrate specificity of a multifunctional calmodulin-depen-duced changes. In R. J. Andrew (Ed.), Neural and behav-dent protein kinase. Journal of Biological Chemistry, 260,ioural plasticity—The use of the domestic chick as a model14471–14476.(pp. 262–276). Oxford: Oxford Univ. Press.
Ouimet, C. C., McGuinness, T. L., & Greengard, P. (1984). Immu-Malenka, R. C., Kauer, J. A., Perkel, D. J., Mauk, M. D., Kelly,nocytochemical localization of calcium/calmodulin-dependentP. T., Nicoll, R. A., & Waxham, M. N. (1989). An essentialprotein kinase II in rat brain. Proceedings of the Nationalrole for postsynaptic calmodulin and protein kinase activityAcademy of Science, USA, 81, 5604–5608.in long-term potentiation. Nature, 340, 554–557.
Rickard, N. S., Poot, A. C., Gibbs, M. E., & Ng, K. T. (1994).Malgaroli, A., Malinow, R., Schulman, H., & Tsien, R. W. (1992).Antagonism of either NMDA or non-NMDA glutamate recep-Persistent signalling and changes in presynaptic function intors impairs long-term memory consolidation in the day-oldlong-term potentiation. Ciba Foundation Symposium, 164,chick. Behavioral and Neural Biology, 62, 33–40.176–191.
Rickard, N. S., Ng, K. T., & Gibbs, M. E. (1994). A nitric oxideMalinow, R., Schulman, H., & Tsien, R. W. (1989). Inhibitionagonist stimulates consolidation of long-term memory in theof postsynaptic PKC or CaMK II blocks induction but not1-day-old chick. Behavioral Neroscience, 108, 640–644.expression of LTP. Science, 245, 862–866.
Rickard, N. S., & Ng, K. T. (1995). Blockade of metabotropic gluta-Mehta, U., & Kaul, D. (1990). Effect of trifluoperazine on choles-mate receptors prevents long-term memory consolidation.terol metabolism of smooth muscle cells exposed to hypercho-Brain Research Bulletin, 36, 355–359.lesterolemic medium in vitro. Biochemical Medicine Meta-
Rodnight, R., & Wofchuk, S. T. (1992). Roles for protein phosphor-bolic Biology, 44, 151–158.ylation in synaptic transmission. Essays in Biochemistry, 27,Miller, S. G., & Kennedy, M. B. (1986). Regulation of brain type91–102.II Ca2//calmodulin-dependent protein kinase by autophos-
phorylation: a Ca2/-triggered molecular switch. Cell, 44, Rose, S. P. R., & Csillag, A. (1985). Passive avoidance trainingresults in lasting changes in deoxyglucose metabolism in left861–870.
AID NLM 3700 / 6v06$$$144 05-20-96 10:36:10 nlma AP: NLM

35CaMK AND MEMORY FORMATION IN CHICK
hemisphere regions of chick brain. Behavioral and Neural sawa, M., & Kidaka, H. (1990). KN-62, 1-[N, O-bis(5-isoqui-nolinesulfonyl)-N-methyl-L-tyrosyl]-4-phenylpiperazine, aBiology, 44, 315–325.specific inhibitor of Ca2//calmodulin-dependent protein ki-Rose, S. P. R. (1991). How chicks make memories: the cellularnase II. Journal of Biological Chemistry, 265, 4315–4320.cascade from c-fos to dendritic remodelling. Trends in Neuro-
Van Belle, H. (1984). The effect of drugs on calmodulin and itsscience, 14, 390–397.interaction with phosphodiesterase. Advances in Cyclic Nu-Sedman, G. L., Jeffrey, P. L., Austin, L., & Rostas, J. A. P. (1986).cleotide and Protein Phosphorylation Research, 17, 557–567.The metabolic turnover of the major proteins of the postsyn-
Wasterlain, C. G., Bronstern, J. M., Morin, A. M., Dwyer, B. E., &aptic density. Molecular Brain Research, 1, 221–230.Sankar, R. (1992). Translocation and autophosphorylation ofSedman, G., O’Dowd, B., Rickard, N., Gibbs, M. E., & Ng, K. T.brain calmodulin kinase II in status epilepticus. Epilepsy Re-(1991). Brain metabolic activity associated with long-termsearch Supplement, 9, 231–238.memory consolidation. Molecular Neurobiology, 5, 351–354.
Weiss, B., Prozialeck, W., Cimino, M., Barnette, M. S., & Wallace,Serrano, P. E., Ramus, S. J., Bennett, E. L., & Rosenzweig,T. L. (1980). Pharmacological regulations of calmodulin. An-M. R. (1992). Comparative study of roles of the lobus parolfac-nals of the New York Academy of Science, 356, 319–345.torius and intermediate medial hyperstriatum ventrale in
Wolfman, C., Fin, C., Dias, M., Bianchin, M., Da Silva, R. C.,memory formation in the chick brain. Pharmacology, Bio-Schmits, P. K., Medina, J. H., & Izquierdo, I. (1994). Intrahip-chemistry and Behavior, 41, 761–766.pocampal or intraamygdala infusion of KN-62, a specific in-
Serrano, P. E., Beniston, D. S., Oxonian, M. G., Rodriguez, W. A.,hibitor of Calcium/calmodulin-dependent protein kinase II,
Rosenzweig, M. R., & Bennett, E. L. (1994). Differential ef-causes retrograde amnesia in the rat. Behavioral and Neural
fects of protein kinase inhibitors and activators on memoryBiology, 61, 203–205.
formation in the 2-day-old chick. Behavioral and Neural Biol-Youngren, O. M., & Phillips, R. S. (1978). A stereotaxis atlasogy, 61, 60–72.
of the brain of the three-day-old domestic chick. Journal ofSilva, A. J., Stevens, C. F., Tonegawa, S., & Wang, Y. (1992a). Comparative Neurology, 181, 567–600.
Deficient hippocampal long-term potentiation in a-calcium-Zhao, W. Q., Gibbs, M. E., Sedman, G. L., & Ng, K. T. (1994).calmodulin kinase II mutant mice. Science, 257, 201–206.
Effects of PKC inhibitors and activators on memory. Behav-Silva, A. J., Paylor, R., Wehner, J. M., & Tonegawa, S. (1992b). ioral and Brain Research, 60, 151–160.
Impaired spatial learning in a-calcium-calmodulin kinase IIZhao, W. Q., Polya, G. M., Wang, B. H., Gibbs, M. E., Sedman,mutant mice. Science, 257, 206–211.
G. L., & Ng, K. T. (1995a). Inhibitors of cAMP-dependentSoderling, T. R., Tan, S. E., McGlade-McCuulloh, E., Yamamoto, protein kinase impair long-term memory formation in day-
H., & Fukunaga, K. (1994). Excitatory interactions between old chicks. Neurobiology of Learning and Memory, 64, 106–glutamate receptors and protein kinases. Journal of Neurobi- 118.ology, 25, 304–311. Zhao, W. Q., Polya, G. M., Wang, B. H., Bennett, P., Gibbs,
Suzuki, T. (1994). Protein kinases involved in the expression of M. E., Sedman, G. L., & Ng, K. T. (1995b). The involvementlong-term potentiation. International Journal of Biochemis- of cAMP-dependent protein kinase in long-term memory for-try, 26, 735–744. mation. Proceedings of the Australian Neuroscience Society,
6, 119.Tokumitsu, H., Chijiwa, T., Hagiwara, M., Mizutani, A., Tera-
AID NLM 3700 / 6v06$$$144 05-20-96 10:36:10 nlma AP: NLM




















