
Involvement of Ca2+-calmodulin in Cd2+ toxicity during the early phases of radish (Raphanus sativus...
10
Plant, Celt and Environtnent (1997) 20,600-608 Involvement of Ca^^-calmodulin in Cd^^ toxicity during the early phases of radish {Raphanus sativus L.) seed germination A. RIVETTA, N. NEGRINI & M. COCUCCI Dipartimento di Fisiologia delle Piante Coltivate e Chimica Agraria, Universita di Milano, via Celoria 2, 20133 Milano, Italy ABSTRACT The toxicity of Cd^^ in vivo during the early phases of radish {Raphanus sativus L.) seed germination and the in vitro Cd^* effect on radish caimodulin (CaM) were stud- ied. Cd^"^ was taken up in the emhryo axes of radish seeds; the increase in fresh weight of emhryo axes after 24 h of incubation was inhihited significantly in the presence of 10 mmol m~^^ Cd^* in the external medium, when the Cd^* content in the embryo axes was c. 1-1 |imol g~' FW. The reabsorption of K*, which characterizes germination, was inhibited by Cd^*, suggesting that Cd^* affected metabolic reactivation. The slight effect of Cd^^ on the transmem- brane electric potential of the cortical cells of the emhryo axes excluded a generalized toxicity of Cd^* at the plasma membrane level. After 24 h of incubation, Cd^* induced no increase in total acid-soluble thiols and Cd^*-binding pep- tides able to reduce Cd^^ toxicity. Ca^* added to the incu- bation medium partially reversed the Cd^'^-induced inhi- bition of the increase in fresh weight of embryo axes and concomitantly reduced Cd^* uptake. Equilibrium dialysis experiments indicated that Cd^^ bound to CaM and com- peted with Ca^^ in this binding. Cd^* inhibited the activa- tion of Ca^^-CaM-dependent calf-brain phosphodi- esterase, inhibiting the Ca^^-CaM active complex. Cd^^ reduced the binding of CaM to the Ca'^^-CaM binding enzymes present in the soluble fraction of the embryo axes of radish seeds. The possibility that Cd^* toxicity in radish seed germination is mediated by the action of Cd^* on Ca *-CaM is discussed in relation to the in vivo and in vitro effects of Cd^^ Key-words: Raphanus sativus; radish; cadmium; calcium; caimodulin; seed germination. Abbreviations: CaM, caimodulin; £„„ transmembrane electric potential difference; PDE, phosphodiesterase. INTRODUCTION Cadmium is toxic to animals (Hinkle, Kinsella & Osterhoudt 1987) and plants (Woolhouse 1983). The pres- ence of Cd^"^ in the environment is increasing as a result of Correspondence: Matmzio Cocucci, Dipartimento di Fisiologia delle Piante Coltivate e Chimica Agraria, Universita di Milano, via Celoria 2, 20133 Milano, Italy. 600 industrial processes, the presence of the cation as contami- nant in agricultural fertilizers, sludge dispersal and atmo- spheric deposition (Wagner 1993). Cadmium is absorbed and accumulated into plants where it causes severe toxic effects resulting in the inhibition of growth and cell death (Clijster & Van Assche 1985). The mechanism of Cd^"^ toxicity is still unclear, although the responses of plants to Cd^"^ exposure and in particular the role of heavy metal binding peptides (phytochelatins) have been extensively studied (Steffens 1990; Meharg 1994; Rauser 1995) and several toxic effects of Cd^"^ have been investigated. Photosynthesis and mitocbondrial oxidative phosphorylation are inhibited severely by Cd^"^, which appears to increase the passive penneability of the mito- chondrial inner membrane to protons (Kesseler & Brand 1995); Cd^"^, as proposed for other beavy metals, stimulates the production of toxic free radicals (Sandman & Boger 1983) and enhances the level of lipid peroxides (Somashekaraiah, Padmaja & Prasad 1992); Cd""^ can increase the susceptibility of plant cells to oxidative stress through the depletion of glutathione, caused by the synthe- sis of phytochelatins, which are -SH rich peptides induced by Cd^^ (De Vos et al. 1992; Rauser 1995). Cd^^ inhibits the activities of many enzymes (Van Assche & Clijster 1990), and phytochelatins protect these plant enzymes from heavy-metal poisoning (Kneer & Zenk 1992). It has been proposed that Cd'^"^ uptake and toxicity in ani- mals could be caused by an interaction with Ca^"^ (Hinkle et al. 1987). Ca"^"*^ is involved in metabolic control and plant cell signal transduction (Allan & Trewavas 1987; Kaus 1987; Blowers & Trewavas 1988). Many Ca^^- dependent processes are mediated by caimodulin (CaM), a low-molecular-weight soluble Ca^"^-binding protein (Roberts, Lukas & Watterson 1986; Allan & Hepler 1989; Roberts & Harmon 1992). The Cd^^-induced toxicological effect could involve CaM as a primary intracellular target (Klee, Crouch & Richman 1980; Behra 1993), as has been suggested for aluminium toxicity (Siegel & Haug 1983; Rengel 1992). A role of CaM in metabolic reactivation, which charac- terizes the early phases of seed germination, is suggested by the correlation between an increase in the level of CaM and metabolic reactivation in conditions able to inhibit or promote germination (Cocucci & Negrini 1988; Hemandez-Nistal et al. 1989) and by the inhibition of ger- mination by CaM antagonists, suggesting that the © 1997 Blackwell Science Ltd
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Involvement of Ca2+-calmodulin in Cd2+ toxicity during the early phases of radish (Raphanus sativus...

Plant, Celt and Environtnent (1997) 20,600-608
Involvement of Ca^^-calmodulin in Cd^^ toxicity during theearly phases of radish {Raphanus sativus L.) seed germination
A. RIVETTA, N. NEGRINI & M. COCUCCI
Dipartimento di Fisiologia delle Piante Coltivate e Chimica Agraria, Universita di Milano, via Celoria 2, 20133 Milano, Italy
ABSTRACT
The toxicity of Cd^^ in vivo during the early phases ofradish {Raphanus sativus L.) seed germination and the invitro Cd^* effect on radish caimodulin (CaM) were stud-ied. Cd " was taken up in the emhryo axes of radish seeds;the increase in fresh weight of emhryo axes after 24 h ofincubation was inhihited significantly in the presence of10 mmol m~^ Cd^* in the external medium, when the Cd^*content in the embryo axes was c. 1-1 |imol g~' FW. Thereabsorption of K*, which characterizes germination, wasinhibited by Cd^*, suggesting that Cd^* affected metabolicreactivation. The slight effect of Cd ^ on the transmem-brane electric potential of the cortical cells of the emhryoaxes excluded a generalized toxicity of Cd^* at the plasmamembrane level. After 24 h of incubation, Cd^* induced noincrease in total acid-soluble thiols and Cd^*-binding pep-tides able to reduce Cd^^ toxicity. Ca^* added to the incu-bation medium partially reversed the Cd ' -induced inhi-bition of the increase in fresh weight of embryo axes andconcomitantly reduced Cd^* uptake. Equilibrium dialysisexperiments indicated that Cd^^ bound to CaM and com-peted with Ca ^ in this binding. Cd^* inhibited the activa-tion of Ca^^-CaM-dependent calf-brain phosphodi-esterase, inhibiting the Ca^ -CaM active complex. Cd ^reduced the binding of CaM to the Ca' -CaM bindingenzymes present in the soluble fraction of the embryo axesof radish seeds. The possibility that Cd^* toxicity in radishseed germination is mediated by the action of Cd^* onCa *-CaM is discussed in relation to the in vivo and in vitroeffects of Cd^^
Key-words: Raphanus sativus; radish; cadmium; calcium;caimodulin; seed germination.
Abbreviations: CaM, caimodulin; £„„ transmembrane electricpotential difference; PDE, phosphodiesterase.
INTRODUCTION
Cadmium is toxic to animals (Hinkle, Kinsella &Osterhoudt 1987) and plants (Woolhouse 1983). The pres-ence of Cd " in the environment is increasing as a result of
Correspondence: Matmzio Cocucci, Dipartimento di Fisiologiadelle Piante Coltivate e Chimica Agraria, Universita di Milano, viaCeloria 2, 20133 Milano, Italy.
600
industrial processes, the presence of the cation as contami-nant in agricultural fertilizers, sludge dispersal and atmo-spheric deposition (Wagner 1993).
Cadmium is absorbed and accumulated into plants whereit causes severe toxic effects resulting in the inhibition ofgrowth and cell death (Clijster & Van Assche 1985). Themechanism of Cd " toxicity is still unclear, although theresponses of plants to Cd " exposure and in particular therole of heavy metal binding peptides (phytochelatins) havebeen extensively studied (Steffens 1990; Meharg 1994;Rauser 1995) and several toxic effects of Cd " have beeninvestigated. Photosynthesis and mitocbondrial oxidativephosphorylation are inhibited severely by Cd " , whichappears to increase the passive penneability of the mito-chondrial inner membrane to protons (Kesseler & Brand1995); Cd " , as proposed for other beavy metals, stimulatesthe production of toxic free radicals (Sandman & Boger1983) and enhances the level of lipid peroxides(Somashekaraiah, Padmaja & Prasad 1992); Cd"" canincrease the susceptibility of plant cells to oxidative stressthrough the depletion of glutathione, caused by the synthe-sis of phytochelatins, which are -SH rich peptides inducedby Cd^^ (De Vos et al. 1992; Rauser 1995). Cd^^ inhibitsthe activities of many enzymes (Van Assche & Clijster1990), and phytochelatins protect these plant enzymes fromheavy-metal poisoning (Kneer & Zenk 1992).
It has been proposed that Cd' " uptake and toxicity in ani-mals could be caused by an interaction with Ca " (Hinkleet al. 1987). Ca""* is involved in metabolic control andplant cell signal transduction (Allan & Trewavas 1987;Kaus 1987; Blowers & Trewavas 1988). Many Ca^^-dependent processes are mediated by caimodulin (CaM), alow-molecular-weight soluble Ca^"^-binding protein(Roberts, Lukas & Watterson 1986; Allan & Hepler 1989;Roberts & Harmon 1992). The Cd^^-induced toxicologicaleffect could involve CaM as a primary intracellular target(Klee, Crouch & Richman 1980; Behra 1993), as has beensuggested for aluminium toxicity (Siegel & Haug 1983;Rengel 1992).
A role of CaM in metabolic reactivation, which charac-terizes the early phases of seed germination, is suggestedby the correlation between an increase in the level of CaMand metabolic reactivation in conditions able to inhibit orpromote germination (Cocucci & Negrini 1988;Hemandez-Nistal et al. 1989) and by the inhibition of ger-mination by CaM antagonists, suggesting that the
© 1997 Blackwell Science Ltd

in toxicity 601
CaM complex is important in the control of Ca^'^-CaM-dependent activities (Cocucci & Negrini 1989).
In the present paper, the possibility that the toxicity ofcould be explained by an interaction with Ca^^, as
suggested in animal and algal cells (Hinkle et al. 1987;Karez et al. 1990), is considered; thus, an investigation wascarried out on the in vivo Cd " toxicity and the protectiveaction of Ca " on seed germination and the in vitro effectof Cd " on the formation of the Ca~'^-CaM active complexusing plirified radish CaM. The mechanism of Cd " toxic-ity is also discussed in terms of the in vivo and in vitroeffects of Cd " in relation to the action of Ca " .
MATERIALS AND METHODS
Plant material and incubation conditions
Radish seeds (Raphanus sativus L., cv. Tondo RossoQuarantino) from a single year's crop were obtained fromIngegnoli Co, Milan, Italy. Seeds were de-coated and incu-bated (8 seeds cm""*) as described previously (Cocucci &Ballarin-Denti 1981). The incubation media contained0-1 mmol m^^ Mes and CdS04 at the indicated concentra-tions, brought to pH 6 with BTP. After 24 h of incubation,the embryo axes (apical meristems and hypocotyl) wereexcised from the seeds and weighed as described previ-ously (Ballarin-Denti & Cocucci 1979).
Chemicais
CdS04 was purchased from Merck; cAMP, adenosinedeaminase and alkaline phosphatase were purchased fromBoehringer Biochemia Robin, Italy; Fluphenazine 2HC1,Tris, Hepes, Mes, BTP, phenyl methyl sulphonyl fluoride,BSA, cyt c and Ellman's reagent [5,5'-dithio-bis(2-nitrobenzoic acid)] were purchased from Sigma, Italy;Sephadex and Sepharose 6B, epoxy activated, were pur-chased from Pharmacia Fine Chemicals, Sweden.
Measurements of net fluxes andlevels
(4 cm"*) under a continuous flow (about 450 cm"* h ') ofaerated medium, thermoregulated at 26 °C, containing 0-1mol m'^ Mes (adjusted to pH 6 with BTP), 0-25 mol m"^K2SO4 and CdSO4 at the indicated concentrations.Micropipettes were filled with 3 kmol m"" KCl (resistanceof 10-20 MX2) and the E^ recorded using a high-impedance electrometer amplifier (K5-700, WorldPrecision Instruments). After at least 30 min of pre-incuba-tion, the E^^ was measured in the cortical cells of theembryo axes 2—4 mm from the root apex (Ballarin-Denti &Cocucci 1979).
Measurement of total acid-soluble thiols
The embryo axes were homogenized at 2 °C (using a pestleand mortar) in 10 volumes of a solution containing 0-1 molm"- EDTA and 10% (w/v) TCA (10:2, v:v) and the thiolswere measured using Ellman's reagent (Grill & Esterbauer1973).
Equilibrium dialysis
To prevent contamination of the solutions used for equilib-rium dialysis experiments with Ca"" and Mg " , glasswarewas rinsed in EDTA and the dialysis tubing was boiled in asolution containing 5 mol m"" EDTA and then extensivelyrinsed in distilled water (Potter et al. 1983). Ca^'^-freecaimodulin was obtained by passing CaM through aDowex 50 X8 column in H* form (0-7 x 10 cm) eluted with
and
The net fluxes of Cd " and Ca " were measured by thechanges in the concentrations of these ions in the incuba-tion media (Cocucci & Cocucci 1977) and the Cd " andCa " contents determined in the embryo axes excised fromthe seeds (after they were washed twice in ice-cold water),using atomic absorption spectrophotometry (SpectrAA-20,Varian) after complete mineralization of the samplesobtained at 100 °C in the presence of HNO3:H2SO4:HC1O4(5:1:1, v:v:v). The K" level in the incubation medium wasalso measured by atomic absorption spectrophotometry.
Measurements of transmembrane electricpotential difference (Em)
E^ was measured using a conventional proceduredescribed previously (Cocucci et al. 1976) in a lucite cell
© 1997 Blackwell Science Ltd, Plant, Cell and Environment, 20, 600-608
1 mol m""* Tris-HCl at pH 7. Bags made with dialysis tub-ing (1 cm^, 12 000 Mr threshold) were dialysed to equilib-rium (14 h at 25 °C) against 40 cm"* of solution containing10 mol m~ imidazole (pH 7), 100 mol m"" KCl, 48 mmolm""* CaCl2 and CdS04 at the indicated concentrations.
Preparation of cell fractions
Radish embryo axes were homogenized at 2 °C (with amortar and pestle) in 5 volumes of a medium containing 50mol m"- Hepes, 70 mol m " KCl, 10 mol m'^ MgCl2 and250 mol m"^ sucrose at pH 7 2: immediately before use, 15mol m"" /3-mercaptoethanol and 30 fig cvnT^ phenyl-methylsulphonyl fluoride (dissolved in DMSO, 15 mg cm""*) wereadded (Negrini et al. 1995). The homogenate was strainedthrough one layer of muslin, and centrifuged at 500 g for10 min, 13 000 ^ for 15 min and 100 000 g for 60 min at2 °C. The supernatant was analysed on a Sephadex G-75SF column (1 -4 x 75 cm) and the fractions tested for CaMand protein content. The Cd " level in the fractions wasassayed by atomic adsorption spectrophotometry aftercomplete mineralization.
Extraction and purification of CaM
Caimodulin was extracted from radish seeds according toKakiuchi et al. (1981) and purified essentially as describedby Charbonneau & Cormier (1979) for plant caimodulin.CaM was chromatographed on DEAE-cellulose (pH 7-5)

602 A. Rivetta et al.
and then on a fluphenazine-Sepharose-6B affinity column.The CaM obtained was homogeneous when analysed byelectrophoresis on SDS-PAGE (Laemmli 1970).
Assay of CaM
The CaM level was assayed by its capacity to activatephosphodiesterase (PDE) extracted from calf brain andprepared according to Yamagami & Terayama (1981).PDE was assayed spectrophotometrically in 1 cm ^ finalvolume in the presence of alkaline phosphatase (c. 1 unit)and adenosine deaminase (c. 2 units), as described previ-ously (Cocucci 1984).
RESULTS
In wVo experiments
Effect ofCd " on the increase in fresh weight of radishseed embryo axes
When radish seeds were incubated in the presencethe cation was taken up by the seeds and the increase infresh weight of the embryo axes was inhibited. Table 1shows that the fresh weight of the embryo axes increased byc. 87% with respect to the initial weight (time 0) during thefirst 24 h of incubation in the presence of 0-1 mol m""* Mes-BTP (pH 6) as a control. This increase was not affected sig-nificantly when Cd " was supplied at up to 30 mmol m"\At higher concentrations (100 or 300 mmol m"" Cd^^), thefresh weight increased only by c. 65 and 14%, respectively.
Table I (Cd- 'j ) also shows that the Cd"^ content ofthe embryo axes from ungerminated seeds was low anddid not change significantly after 24 h of incubation inthe absence of added Cd""". The Cd"" content after 24 hof incubation in the presence of Cd~^ was much higherand increased as more Cd"" was supplied at the start ofincubation. The level of Cd"" present in the incubationmedia (Table 1, Cd" , ,) after 24 h was much lower thanthat supplied at the beginning of incubation, in agree-ment with the increase of Cd"" in the seeds. Kineticexperiments indicated that the Cd"" contents in theembryo axes reported in Table 1 for each Cd"" concen-tration supplied were attained within the first 7 h of incu-bation (data not shown).
Effect ofCd " on transmembrane electric potential (E^) ofcortical cells of radish seed embryo axes
The increase in the fresh weight of radish embryo axeswas accompanied by the metabolic reactivation of theseeds (Cocucci & Negrini 1988). The effect of Cd " onsome metabolic parameters has been studied to clarifywhether the inhibiting action of Cd " on fresh weightincrease involved metabolic reactivation. The metabolicreactivation of germinating radish seeds is characterizedby a transitory increase in E,.^, which at 24 h of incubationreaches a stable metabolically dependent value of about-110 mV (Ballarin-Denti & Cocucci 1979). Table 2shows that the E^ was not affected significantly, com-pared to the control, in seeds treated with 10, 30 and100 mmol m""* Cd " , but the £,^ decreased by 16% in the300 mmol m"^ Cd " treatment. In germinating radishseeds, K" was released in the medium during the first 12 hof incubation and then reabsorbed (Cocucci & Cocucci1977), reaching very low levels after 24 h of incubation.Cd"^ supplied at up to 30 mmol m""* did not inhibit the
reabsorption of K" , while at higher Cd^^ concentrationsK"*" reabsorption was inhibited (Table 2).
Effect of the presence ofCa^'^ in the incubation mediumon the Cd '^-induced inhibition of fresh weight increase
In other systems, Cd"" uptake and toxicity are reduced byCa-^ (Hinkle et al. 1987; Karez et al. 1990). During thefirst 24 h of incubation of radish seeds, Ca"" was releasedinto the incubation medium and after 24 h of incubationthe Ca " level reached c. 130 mmol m""* (Negrini, Rivetta& Cocucci 1995). The Ca^^ content in the embryo axeswas not significantly affected by Cd"" at the concentra-tions used and was 7-6 ± 0-5 |imol (100 embryos)"'(datum not shown) at 24 h of incubation. The effect ofCd"" on seed germination at high Ca^^ concentration wasexamined; Table 3 shows that the presence of 10 mol m""*Ca"" in the incubation medium did not significantlyaffect the increase in fresh weight of the embryo axes.There was no effect of Ca' ^ on the increase in freshweight of embryo axes at 100 mmol m"" Cd"" , but theinhibition by Cd"" was greatly reduced by Ca " at
^^300 mmol m^ Cd^^; indeed, at this level of Cd^^, theinhibition of the increase in fresh weight was reduced
Incubation conditions(time and treatment)
Fresh weight(g lOOemb."')
Cd^",,,(nmol lOOemb.^')
*n.d.: not detectable.
(mmol
Oh24 h24 h24 h24 h24 h
Control10mmolm~-^Cd^"30 mmol m ' Cd^"
100 mmol m~ Cd^"300 mmol m""* Cd' "
0-230 ± 0-0050-430 ±0-0170-438 ±0-0170-436 ±0-0100-380 ± 0-0080-263 ±0-012
5± 0-7± 1-
44 ± 6-135 ± 5-439 ±15
1167 ±64
5111-3-3
_n.d.*
0-86 ±0-231-56 ±0-148-17 ±0-65
15-00 ±0-41
Table 1. The effect of Cd"" on the increase infresh weight of embryo axes from germinatingradish seeds. De-coated radish seeds wereincubated at 26 °C in an agitated water bath ina medium containing Mes-BTP 0-1 mol m"'pH 6 (Control) and CdS04 at the indicatedconcentrations. At the indicated times, theembryo axes were excised and weighed. Cd^"levels in the embryo axes and in the incubationmedia were assayed by atomic absorptionspectrophotometry. The values are means ±SEof seven experiments run in triplicate
© 1997 Blackwell Science Ltd, Plant, Cell and Environment, 20, 600-608

-caimodulin in toxicity 603
Table 2. Transmembrane electric potential (£•„,) and K* level inthe incubation medium after 24 h of incubation in the presence ofcadmium. De-coated radish seeds were incubated in a mediumcontaining 0-1 mol m"^ Mes-BTP (pH 6) and Cd"* as indicated.After 24 h of germination, the E^ of the embryo axis cells wasmeasured in a perfusion medium containing 0-1 mol m" (pH 6),0-5 mol m " K* (as SO4"") and Cd^* at the concentration attainedfor each treatment after 24 h of incubation (see Table 1). E^, valuesare the mean ±SE of at least 10 single measurements on four seedsrepeated three times. K* was measured in the incubation mediumby atomic absorption spectrophotometry and values are the mean ±SE of four experiments run in triplicate
Incubation conditions(time and treatment)
24 h Control24 h 10mmolm"^Cd^*24 h 30 mmol m"^ Cd^*24 h 100 mmol m"-^Cd-*24 h 300 mmol m" Cd"*
(mV)
-124 ±1-123 ±3-122±3-116±2-104 ± 1
" • out
(/imol 100 seeds"')
6-1 ±0-86-5 ±0-57-0±l-2
16-1 ±3-316-6 ±2-9
in the incubation medium did not significantlyaffect the increase in acid-soluble thiols. This increasewas inhibited by e. 38% when the seeds were incubated inthe presence of 300 mmol m"^ Cd^*. In the presence of100 mmol m"^ Cd^*, c. 75% of the total Cd^* content inthe embryo axes was present in the soluble fraction (cen-trifuged at 100 000 g; data not shown). The separation ofthis fraction on a Sephadex G-75 SE column (Fig. 1)showed that > 70% of the Cd^* was eluted in the zonecorresponding to the total volume of the column, suggest-ing that, after this time of incubation, high-affinity Cd^*-binding peptides were not present. The level of Ca^* pre-sent in the soluble fraction was about 3-fold higher thanCd" * and its separation on the G-75 SF column had a dis-tribution similar to that of Cd^* (data not shown). This
~*suggests that a large proportion of the Cd~* andeluted with the total volume of the column were presentin the cellular free spaces and vacuoles.
from 86% in the absence of Ca2+of Ca" *. The presence of Ca^
^* to 36% in the presencein the incubation medium
2+reduced Cd uptake, so that the Cd""* content in theembryo axes after 24 h of incubation was greatly reducedand the Cd^* level in the medium was concomitantlyhigher (Table 3). Since Mg^* at the same external Ca"*concentration did not reduce Cd"* toxicity (data notshown), the effect of Ca^* appeared to be specific.
Effect on the levels of acid-soluble thiols and onCd" -binding peptides
The exposure of plants to Cd"* was accompanied by anincrease in the level of acid-soluble thiols and high-affin-ity Cd-*-binding peptides (Steffens et al. 1990). Table 4shows that the levels of acid-soluble thiols in the embryoaxes increased approximately 5-fold from their initialvalues after 24 h of incubation; 10, 30 or 100 mmol m"^
In vitro experiments2+Interactions ofCd with the Ca" -CaM complex from
radish seeds
Equilibrium dialysis experiments were set up to study theeffect of Cd^* on the Ca^*-CaM complex. Eigure 2 showsthat, in the absence of added Cd^*, only a small amount ofCd"* was present, dependent on the amount present as acontaminant, and CaM bound about four Ca"* ions. When
2+Cd" was added, less Ca^ was bound to CaM and corre-spondingly more Cd"* bound to CaM. Figure 2 also showsthe Cd^*/Ca"* ratio (Cd"* and Ca^* outside the dialysisbag) at the Cd^* concentrations tested; when the Cd"*/Ca^*ratio was close to 1, the amounts of Ca * and Cd"* boundto CaM were similar.
Calf-brain PDE is activated by CaM in the presence ofcalcium; Fig. 3 shows the effect of Cd"* on PDE activity(reduction in OD260) at different concentrations of CaM.The activity of PDE was dependent on the concentration ofCaM in the assay medium. Figure 3 shows that increasing
Table 3. The effect of the presence of Ca~* in the incubation medium on the Cd"*-induced inhibition of the increase in fresh weight and Cd"*content in embryo axes from radish seeds. De-coated radish seeds were incubated at 26 °C for 24 h in an agitated water bath in the presence of0-1 mol m"" Mes-BTP (pH 6) and 0 or 10 mol m" CaClj, and CdSO4 as indicated. At the end of incubation, the embryo axes were excisedand weighed. Cd"* levels in embryo axes and incubation media were assayed by atomic absorption spectrophotometry. Values are the mean±SE of four experiments run in triplicate
Incubation conditions(time and treatment)
Oh24 h Control24 h 100 mmol nr- Cd'*24 h 300 mmol m"- Cd^*
Fresh weight(g lOOemb."')
Control
0-230 ± 0-0050-398 ±0-0100-365 ±0-0180-253 ±0-012
C a -
0-4010-3700-339
± 0-009± 0-020±0-015
Cd,,,(nmol 100
Control
5 ± 0-58±l-0
390 ±23-2980 ± 59-7
emb. ')
C a -
7±250 ±650 ±
1-210-447-6
(mmol r
Control
n.d.9-0 ±1
14-1 ±1
n" )
-6-0
r^ 2+Ca
n.d10-4 ±98-7 ±
2-08-8
*n.d.: not detectable.
© 1997 Blackwell Science Ltd, Plant, Cell and Environment, 20, 600-608

604 A. Rivetta et al.
Table 4. Acid-soluble thiol levels of embryo axes from radishseeds. De-coated seeds were incubated at 26 °C in an agitatedwater bath in the presence of 0-1 mol m Mes-BTP, pH 6(Control) and Cd " concentrations as indicated. At the indicatedtimes and treatments, the embryo axes were excised from the seedsand the level of acid-soluble thiols was determined using Ellman'sreagent. Values are the mean ± SE of three experiments run intriplicate
Incubation conditions(time and treatment)
Thiols(//mol 100 emb." ')
Oh24 h Control24 h 124 h 30 mmol m"24 h 100 mmol m'^24 h 300 mmol m~
0-23 ±0-01l-26±0171-29 ±0-03l-23±0-151 -27 ± 0-040-87 ±0-06
concentrations progressively inhibited CaM-acti-vated PDE. The Cd" " concentration required to inhibitCaM-activated PDE was higher when PDE activity wasassayed in the presence of higher amounts of CaM; 50%inhibition was obtained in the presence of c. 60, 100 and110 mmol m""* Cd " . At 10 mmol m~ Cd " , when theCd^'^-dependent inhibition of PDE activity becamedetectable, the Cd ' /Ca ' ratio was about 0-05 at all threeCaM concentrations used. The closed symbols in Eig. 3show the sharp increase in the rate of mOD^^o reductionwhen 5'-AMP was added to the assay medium, indicatingthat concentrations of Cd " < 400 mmol m "* did not affectthe enzymatic system coupled to PDE used in the test.
Eigure 4 shows the well-known activation of CaM-depen-dent PDE by Ca^^ (Cocucci & Negrini 1988) and shows thatCd "*" was not able to activate CaM-dependent PDE.
The effect of 10 and 30 mmol m"^ Cd"^ (Eig. 5) on Ca^^activation of CaM-dependent PDE was also examined. Inthe presence of Cd^^, the activation of PDE was similarwhen Ca"" was supplied at concentrations higher thanthose in the absence of Cd" " (control); 50% activation wasobtained at c. 30 and 90 mmol m^^ Ca"" , in the presence of10 and 30 mmol m"" Cd" "* , respectively, as compared toc. 20 mmol m"" Ca^^ in the absence of Cd " (control). As aCa"" concentration > 500 mmol m^^ inhibited CaM-depen-dent PDE activity (data not shown), the Cd^"^-induced inhi-bition could not be reversed.
Effect ofCa^'^ or C(f^ on the distribution of CaM on aSephadex G- 75 SF column
The effect of Cd~^ on Ca~"^-CaM-dependent enzymes ofradish has been studied by analysing the separation ofCaM, present in the soluble fraction of radish embryo axesfrom seeds incubated for 24 h in control conditions, on aSephadex G-75 SE column. As already reported (Negrinietal. 1995), CaM was eluted in the zones corresponding tohigher molecular weight, and very little CaM was eluted inthe zone near 23 500 Mr (Eig. 6a), which corresponds to
the elution zone of 'free' CaM, suggesting that CaM waseluted while bound to CaM-binding proteins. In the pres-ence of Cd " (Eig. 6b), the elution pattern of CaM changed;the amount of CaM eluted in the zone corresponding tohigher molecular weight decreased and that eluted in thezone corresponding to 'free' CaM increased, suggestingthat less CaM was bound to CaM-binding proteins. In thepresence of Cd " , the elution pattern of protein alsochanged, probably as a consequence of the effect ofon CaM and on other Ca^"^-binding proteins.
DISCUSSION
" supplied in the incubation medium at the beginningof the incubation of radish seeds was absorbed and, for
-3external concentrations > c. 10 mmol m (100 mmol mCd " treatment), the increase in fresh weight of the embryoaxes was inhibited (Table 1). This value is similar to exter-nal concentrations inhibiting the growth of other plant tis-sues and organs, ranging between 10 and 40 mmol m~ inCd'^'^-sensitive Silene vulgaris (de Knecht et al. 1994), inmung bean (Somashekaraiah et al. 1992) and in tobaccoseedlings (Vogeli-Lange & Wagner 1996). In this inhibit-ing external Cd " concentration, the Cd " level in the
0 15 20 25 30 35 40 45 50
Fraction number
Figure 1. The distribution of protein and Cd'"" in the solublefraction of embryo axes from radish seeds incubated for 24 h in thepresence of 100 mmol m""' Cd""", after separation on a Sephadex G-75 SF column. De-coated radish seeds were incubated at 26 °C inan agitated water bath in the presence of 0-1 mol m "* Mes-BTP(pH 6) and 100 mmol m - Cd "". The .soluble fraction (100 000 g;0-9 cm" corresponding to about 160 embryos), containing about70% of the total Cd " absorbed by the embryo axes, waschromatographed on a Sephadex G-75 SF column (1-4 x 75 cm),and 2-3 cm fractions were collected and assayed for protein andCd " . The protein concentration ( ) was assayed by measuring/428O and the Cd"* level ( ) by atomic absorptionspectrophotometry after complete mineralization of the fractions.Standard proteins: albumin from bovine serum (68 kDa), albuminfrom hen egg (45 kDa), CaM purified from radish seeds (23 5 kDa)and cytochrome c (12 5 kDa). The elution of low-molecular-weightmolecules was tested using dinitrophenol as a standard.
' 1997 Blackwell Science Ltd, Plant, Cell and Environment, 20, 600-608

-calmodulin in toxicity 605
E•5
Is<0 T3O OO ,—^
T3
COO
.S"cSo
80
60
40
20
0 -
[CaM], =
\
/
o ^ ^ - *
25 mmol m
P
<
0 10 100
] (mmol m )
niO•CMX3O
2
1
0
Figure 2. Equilibrium dialysis experiments of CaM in thepresence of Ca "* and Cd " . Dialysing tubing bags (1 cm' , 12 000Mr threshold) containing 25 mmol m~ CaM were dialysed for 14 hat 25 °C against buffers containing 10 mol m "* imidazole (pH 7),100 mol m" KCI, 48 mmol m" CaCL and CdS04 as indicated.After dialysis, the Ca " and Cd"" bound to CaM were calculated bysubtracting Ca" " and Cd " concentrations outside the dialysis bagfrom the respective concentrations inside the bag. Ca' ' and Cd "contents were assayed using atomic absorption spectrophotometry.(•) Ca^^ bound to CaM; (O) Cd'^ bound to CaM; (•) C d ' ^ratio, calculated outside the bag at the end of dialysis.
was not sufficient to suggest a breakdown of biologicalmembranes dependent on the Cd^'^-induced accumulationof lipid peroxide, in agreement with the lack of effect onplasma membrane observed in dark-grown seedlings ofmung bean (Somashekaraiah et al. 1992).
2-t-Cd ^ is known to interfere with the transport of Ca inanimal (Hinkle et al 1987) and algal cells (Karez et al.1990). Cd "* transport into plant cells may be mediated bycalcium channels in the plant plasma membrane (Harvey,Venis & Trewavas 1989; Schroeder & Thuleau 1991;Thuleau et al. 1994; Malho et al. 1995). In animal cells, theincrease in Ca^^ concentration in the external mediumgreatly decreased Cd"" toxicity, probably through competi-tion with Cd^^ for entry via the calcium channels (Hinkle etal. 1987). In radish seeds, the increase in external Ca"" inthe incubation medium reduced the Cd~"^-induced inhibitionof fresh weight increase in the embryo axes, and this reduc-tion was accompanied by a reduction of Cd " uptake (Table3), indicating an interaction between Cd " and Ca' " .
Cd " has been suggested to inhibit the activities of manyenzymes, for example RuBP carboxylase, nitrate reduc-tase, catalase, superoxide dismutase, malate dehydroge-nase, protease, ATPase and RNAase (Shah & Dubey 1995;and references therein) and mitochondrial oxidative phos-phorylation (Kesseler & Brand 1995), whereas the inter-action between Cd"" and Ca^^ suggests that, as proposedfor aluminium (Siegel & Haug 1983; Rengel 1992), ^^
embryo axes reached 0-439 /imol (100 embryos) ' (Table1), corresponding to c. 1-1 ^mol g"' EW (from Table 1,100 mmol m"'' Cd " treatment) and to c. 3-8 ^umol g ' DW(datum not shown); this value was c. 50% lower than thetoxic value found for other plants (Verkleij et al. 1990;Galli et al. 1996). The higher toxicity of Cd^^ in radishseeds might be a result of the lack of any increase in totalacid-soluble thiols in the presence of Cd^^ compared to thecontrol (Table 4) and the absence of high-affinity Cd ""-binding peptides (Rauser 1995) in the embryo axes after up
2-rto 24 h of incubation (Eig. 1). These Cd^"-binding com-pounds, which sequester Cd " , reduce the level of activeCd^^ in Cd"^-inhibited enzymes (Kneer & Zenk 1992) atidtheir presence has been suggested to be important in Cd "tolerance in plants (Howden et al. 1995).
The reactivation of membrane functions characterizesthe metabolic reactivation of germinating seed, and the F^represents an important parameter of cell plasma mem-brane activity in radish seed embryo axes (Ballarin-Denti& Cocucci 1979). Cd^* reduced F^ by about 6 and 16% inthe inhibiting 100 and 300 mmol m"" Cd^^ treatments(Table 2), suggesting that the inhibition of the increase infresh weight is accompanied by the inhibition of metabolicreactivation. In agreement with these results, the reabsorp-tion of K^ from the incubation medium, which character-izes the metabolic reactivation of seeds (Negrini et al.1995), was inhibited by these treatments (Table 2).However, the reduction in F^ values observed in the pres-ence of inhibiting levels of Cd^^ in the incubation medium
© 1997 Blackwell Science Ltd, Plant, Cell and Environment, 20, 600-608
] (mmol m •')
Figure 3. The effect of Cd"" on PDE activated by three differentconcentrations of CaM. PDE was spectrophotometrieally assayed in1 cm" fmal volume by measuring the reduction in the absorbance at260 nm at 30 °C in the presence of adenosine deaminase andalkaline phosphatase: the reaction mixture contained 50 mol m~Tris-HCI (pH 8), 0-1 mg cm"" BSA, 3 mol m" MgClj, 01 mol m"^cAMP, 0-2 mol m'^ CaCL and CaM and Cd "" as indicated. PDEactivation in the presence of 30 /imol m~ CaM (D), 60 /imol m'^CaM (A) and 180 /fmol m ' CaM (O); Cd^^/Ca^^ ratio (• ) . Closedsymbols indicate the rate of mODjfto reduction when 0-03 mol m~'5'-AMP was added to the assay medium.
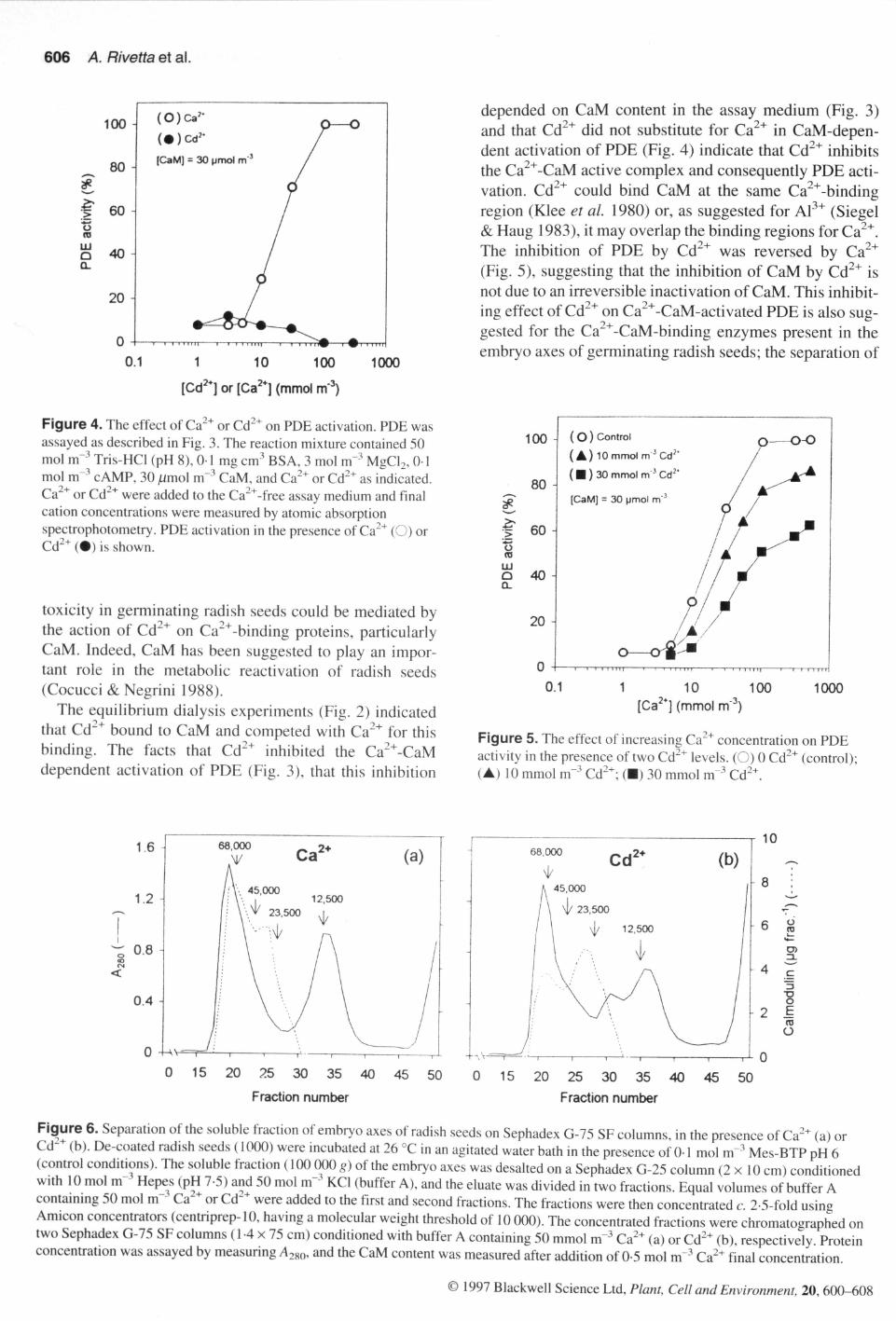
606 A. Rivetta et al.
100
80 -
i 60 -I1g 40Q.
20 -
(O)ca'-
[CaM] = 30 pmol m
0.1 100 1000
*] or [Ca^*] (mmol m' )
depended on CaM content in the assay medium (Fig. 3)and that Cd^* did not substitute for Ca^* in CaM-depen-dent activation of PDE (Eig. 4) indicate that Cd^* inhibitsthe Ca^*-CaM active complex and consequently PDE acti-vation. Cd^* could bind CaM at the same Ca^*-bindingregion (Klee et al. 1980) or, as suggested for Al" * (Siegel& Haug 1983), it may overlap the binding regions for Ca^*.The inhibition of PDE by Cd^* was reversed by ^(Eig. 5), suggesting that the inhibition of CaM by Cd^* isnot due to an irreversible inactivation of CaM. This inhibit-ing effect of Cd^* on Ca^*-CaM-activated PDE is also sug-gested for the Ca^*-CaM-binding enzymes present in theembryo axes of germinating radish seeds; the separation of
Figure 4. The effect of Ca""" or Cd^^ on PDE activation. PDE wasassayed as described in Eig. 3. The reaction mixture contained 50mol m" Tris-HCI (pH 8), 0-1 mg cm^ BSA, 3 mol m ' MgCL, 0" 1mol m "* cAMP, 30 /imol m"'' CaM, and Ca^^ or Cd "" as indicated.
,2+Ca or Cd " were added to the Ca""'-free assay medium and finalcation concentrations were measured by atomic absorptionspectrophotometry. PDE activation in the presence of Ca "" (O) orCd " ( • ) is shown.
toxicity in germinating radish seeds could be mediated bythe action of Cd~* on Ca^*-binding proteins, particularlyCaM. Indeed, CaM has been suggested to play an impor-tant role in the metabolic reactivation of radish seeds(Cocucci & Negrini 1988).
The equilibrium dialysis experiments (Eig. 2) indicatedthat Cd" * bound to CaM and competed with Ca"* for thisbinding. The facts that Cd"* inhibited the Ca^*-CaMdependent activation of PDE (Eig. 3), that this inhibition
20 -
giv
ityP
DE
act
100 -
80 -
60 -
40 -
( O ) Control( A ) 10 mmol m •'
( • ) 30 mmol m '
[CaM] = 30 pmol rr
Cd'-
Cd^-
0.1 10 100] (mmol m )
1000
Figure 5. The effect of increasing Ca " concentration on PDEactivity in the presence of two Cd " levels. (O) 0 Cd "" (control);(A) 10 mmol m"' Cd'^; (•) 30 mmol m"' Cd'^.
L6
1.2
0.8 -
0.4 -
0 -
68,000
/;\\ 45,000
If \ \ V 23,500
i \ \
-V\—r-^: , , r
2*a
12,500
Ar 1
(a)
I1 j-L
0 15 20 25 30 35 40 45 50
Fraction number
68,000 ^
k 45,000
/ \ V 23,500
/ \ i 12,500
(b)-
-
-
10
8
o(0
c
I"TOO
0 15 20 25 30 35 40 45 50Fraction number
Figure 6. Separation of the soluble fraction of embryo axes of radish seeds on Sephadex G-75 SE columns, in the presence of Ca"^ (a) orCd- (b). De-coated radish seeds (1000) were incubated at 26 °C in an agitated water bath in the presence of 0-1 mol m^' Mes-BTP pH 6(control conditions). The soluble fraction (100 000 g) of the embryo axes was desalted on a Sephadex G-25 column (2 x 10 cm) conditionedwith 10 mol m Hepe s (pH 7-5) and 50 mol m~ KCI (buffer A), and the eluate was divided in two fractions. Equal volumes of buffer Acontainmg 50 mol m^ Ca'^ or Cd"^ were added to the first and second fractions. The fractions were then concentrated c 2-5-fold usingAmicon concentrators (centriprep-10, having a molecular weight threshold of 10 000). The concentrated fractions were chromatographed ontwo Sephadex G-75 SF columns (L4 x 75 cm) conditioned with buffer A containing 50 mmol m^' Ca^^ (a) or Cd^^ (b), respectively Proteinconcentration was assayed by measuring A28o, and the CaM content was measured after addition of 0-5 mol m"' Ca"^ final concentration
© 1997 Blackwell Science Ltd, Plant, Cell and Environment, 20, 600-608

-caimodulin in toxicity 607
the soluble fraction on a Sephadex G-75 SF columnshowed that elution in the presence of Cd " greatlyincreased the fraction of free CaM (Fig. 6b), i.e. that notbound to Ca^"^-CaM-binding peptides, suggesting thatCd^^ is able to prevent the CaM binding to Ca^^-CaMbinding enzymes.
These results suggest the involvement of CaM in Cd" *toxicity in germinating radish seed, if Cd^* reaches acytosolic concentration which is able to counteract Ca " inthe activation of CaM. A direct evaluation of Ca " andCd " in the cytosol is very difficult in this material(Negrini et al. 1995), and it is not possible currently todetermine directly the cytosolic Cd " /Ca " ratio. However,the ratio between Cd " and Ca " in the cytosol could besimilar to that calculated from the Cd " and Ca "* contentmeasured in the whole embryo axes. This hypothesis issupported by the facts that Ca " competed with Cd^* foruptake and specifically reduced the toxicity of Cd " (Table3), that the distribution between Cd" " in the externalmedium and in the embryo axes was very similar to that ofCa " (Table 1 and data in the text), that the level of acid-soluble thiols in the embryo axes was low (Table 4) andthat there were no high-affinity Cd~"^-binding peptides pre-sent which could sequester Cd " (Fig. 1). The in vivo inhi-bition of the increase in fresh weight of the embryo axes(Table 1) began at a Cd^^/Ca^+ ratio of c. 0-06, calculatedfrom Cd"" and Ca " contents (Table 1 and datum in thetext), and in vitro the PDE activity began to be inhibited atthis same Cd ' /Ca^"^ ratio value. This supports the hypoth-esis that Cd' ' toxicity in germinating radish seeds involvesCaM-dependent activities.
ACKNOWLEDGMENTS
Research supported by National Research Council of Italy,Special Project RAISA Subproject N. 2, Paper N. 2843,and by Fondazione Lombardia per I'Ambiente.
REFERENCES
Allan E.F. & Hepler P.K. (1989) Caimodulin and calcium-bindingproteins. In The BiochetJiistry of Plants, Vol. 15 (ed. A. Marcus),pp. 455-484. Academic Press, New York.
Allan E.F. & Trewavas A.J. (1987) The role of calcium in metaboliccontrol. In The Biochemistry of Platits, Vol. 12 (eds P. K. Stumpf& E. E. Conn), pp. 117-149. Academic Press, New York.
Ballarin-Denti A. & Cocucci M. (1979) Effects of abscisic acid,gibberellic acid and fusicoccin on the transmembrane potentialduring early phases of germination in radish (Raphanus sativusL.) seeds. Planta 146, 19-23.
Behra R. (1993) In vitro effects of cadmium, zinc and lead oncalmodulin-dependent actions in Oncorhynchus mykiss, Mytilussp. & Chlamydomonas reinhardtii. Archives of EnvironmentalContamituition and Toxicology 24, 21-27.
Blowers D.P. & Trewavas A.J. (1988) Second messengers: theirexistence and relationship to protein kinase. In SecondMessenger in Plant Growth and Development, Vol. 6 (eds W. S.Boss & D. J. Morre), pp. 1-28. Alan R Liss Inc, New York.
Charbonneau H. & Cormier M. (1979) Purification of plant caimodulinby fluphenazine-Sepharose affinity chromatography. Biochemicaland Biophysical Research Communications 90, 1039-1047.
Clijster H. & Van Assche F. (1985) Inhibition of photosynthesis byheavy metals. Photosynthesis Research 7, 31-^0.
Cocucci M. (1984) Increase in caimodulin level in the early phasesof radish seed (Raphanus sativus L.) germination. Plant, Celland Environment 7,215-221.
Cocucci M. & Ballarin-Denti A. (1981) Effects of polar lipids onATPase activity of membrane preparation from germinatingradish seeds. Plant Physiology 68, 377-381.
Cocucci S. & Cocucci M. (1977) Effects of ABA, GA3 and FC onthe development of potassium uptake in germinating radishseeds. Plant Science Eetters 10, 85-95.
Cocucci M. & Negrini N. (1988) Changes in the level of caimodulinand of a caimodulin inhibitor in the early phases of radish(Raphanus sativus L.) seed germination. Plant Physiology 88,910-914.
Cocucci M. & Negrini N. (1989) Caimodulin in metabolic reactiva-tion of germinating seeds. In Plant Membrane Transport: theCurrent Position (eds J. Dainty, M. I. De Michelis, E. Marre & F.Rasi-Caldogno), pp. 355-360. Elsevier Science Publishers,Amsterdam.
Cocucci M., Marre E., Ballarin-Denti A. & Scacchi A. (1979)Characteristics of fusicoccin-induced changes of transmembranepotential and ion uptake in maize root segments. Plant ScienceEetters 6, 143-156.
De Knecht J.A., van Dillen M., Koevoets P.L.M., Schat H., VerkleijJ.A.C. & Ernst W.H.O. (1994) Phytochelatins in cadmium-sensi-tive and cadmium-tolerant Silene vulgaris. Plant Physiology104,255-261.
De Vos C.H.R.. Vonk M.J., Vooijs R. & Schat H. (1992)Glutathione depletion due to copper-induced phytochelatin syn-thesis causes oxidative stress in Silene cucuhalus. PlantPhysiology 98, 853-858.
Galli U., Schuepp H. & Brunold C. (1996) Thiols in cadmium- andcopper-treated maize (Zea mays L. j Planta 198, 139-143.
Grill D. & Esterbauer H. (1973) Quantitative Bestimmung wasser-loslicher Sulfydrylverbindungen in gesunden und geschadigtenNadeln von Picea abies. Phyton 15, 87-101.
Harvey H.J., Venis M.A. & Trewavas A.J. (1989) Partial purifica-tion of a protein from maize (Zea mays) coleoptile membranesbinding the Ca ' -channel antagonist verapamil. BiochetnicalJournal 251, 95-\Q0.
Hemandez-Nistal J., Rodriguez D., Nicolas G. & Alzodaro J (1989)Abscisic acid and temperature modify the level of caimodulin inembryonic axes of Cicerarietinum L. Physiologia Platitaruni 80,891-894.
Hinkle P.M., Kinsella P.A. & Osterhoudt K.C. (1987) Cadmiumuptake and toxicity via voltage-sensitive calcium channels. TheJournal of Biological Chemistry 262, 16333-16337.
Howden R., Goldsbrough P.B., Andersen C.R. & Cobbett CS.(1995) Cadmium-sensitive, cadi mutants of Arabidopsisthaliana are phytochelatin deficient. Plant Physiology 107,1059-1066.
Kakiuchi S., Suboe K., Yamazachi R., Kambayashi J., Sakon &Kosaki G. (1981) Lack of tissue specificity of caimodulin: arapid and high-yield purification method. EEBS Eetters 126,203-207.
Karez C.S., Allemand D., De Renzis G., Gnassia-Barelli M.,Romeo M & Puiseux-Dao S. (1990) Ca-Cd interaction in theprymnesiophyte Cricosphaera elongata. Plant, Cell andEnvironment 13, 483-487.
Kaus H. (1987) Some aspects of calcium-dependent regulation inplant metabolism. Annual Review of Plant Physiology 38,47-72.
Kesseler A. & Brand M.D. (1995) The mechanism of the stimula-tion of state 4 respiration by cadmium in potato tuber (Solanumtuberosum) mitochondria Plant Physiology and Biochemistry33,519-528.
© 1997 Blackwell Science Ltd, Plant, Cell and Environment, 20, 600-608

608 A fl/Veffaetal.
Klee C.B., Crouch T.H. & Richman P.G. (1980) Caimodulin.Annual Review of Biochemistry 49, 489-515.
Kneer R. & Zenk M.H. (1992) Phytochelatins protect plant enzymesfrom heavy metal poisoning. Phytochemistry 31, 2663-2667.
Laemmli H.K. (1970) Cleavage of structural proteins during theassembly of the head of bacteriofage T4. Nature 227, 680-685.
Meharg A.A. (1994) Integrated tolerance mechanisms: constitutiveand adaptative plant responses to elevated metal concentrationsin the environment. Plant, Cell and Environment 17, 989-993.
Malho R., Read N.D., Trewavas A.J. & Salome Pais M. (1995)Calcium channel activity during pollen tube growth and reorien-tation. The Plant Cell 7, 1173-1184.
Negrini N., Rivetta A. & Cocucci M. (1995) Caimodulin levels inradish (Raphanus sativus L.) seeds germinating at low calciumavailability induced by EGTA treatments. Plant, Cell andEnvironment IS, 159-167.
Potter J.D., Strang-Brown P., Walker P.L. & Iida S. (1983) Ca ""binding to Caimodulin. Methods in Enzymology 102, 135-143.
Rauser W.E. (1995) Phytochelatins and related peptides. Structure,biosynthesis and function. Plant Physiology 109, 1141-1149.
Rengel Z. (1992) Disturbance of cell Ca" " homeostasis as a primarytrigger of Al toxicity syndrome. Plant, Cell and Environment 15,931-938.
Roberts D.M. & Harmon A.C. (1992) Calcium-modulated proteins:targets of intercellular calcium signals in higher plants. AnnualReview of Plant Physiology and Plant Molecular Biology 43,375-414.
Roberts D.M., Lukas T.J. & Watterson D.M. (1986) Structure,function and mechanism of action of caimodulin. CRC CriticalReview in Plant Science 4, 311-339.
Sandmann G. & Boger P. (1983) The enzymological function ofheavy metals and their role in electron transfer processes inplants. In Encyclopedia of Plant Physiology, Vol. 15B:Inorganic Pkmt Nutrition (eds A. Lauchli & R. L. Bieleski), pp.563-596. Springer-Verlag, Berlin.
Schroeder I.I. & Thuleau P. (1991) Ca "" channels in higher plantcells. The Pkmt Cell 3, 555-559.
Shah K. & Dubey R.S. (1995) Effect of cadmium on RNA level aswell as activity and molecular forms of ribonuclease in growingrice seedlings. Plant Physiology and Biochemistry 33, 577 584.
Siegel N. & Haug A. (1983) Aluminum interaction with caimod-ulin. Evidence for altered structure and function from opticaland enzymatic studies. Biochimica et Biophysica Acta 744,36^5 .
Somashekaraiah B.V., Padmaja K. & Prasad A.R.K. (1992)Phytotoxicity of cadmium ions on germinating seedlings ofmung bean (Phaseolus vulgaris): involvement of lipid perox-ides in chlorophyll degradation. Physiologia Plantarum 85,85-89.
Steffens J.C. (1990) The heavy metal-binding peptides of plants.Annual Review of Plant Physiology and Plant Molecular Biology41, 553-575.
Thuleau P., Ward J.M., Ranjeva R. & Schroeder J.I. (1994)Voltage-dependent calcium permeable channels in the plasmamembrane of higher plant cell. The EMBO Journal 13,2970-2975.
Van Assche F. & Clijsters H. (1990) Effects of metals on enzymeactivity in plants. Plant, Cell and Environment 13, 195-206.
Verkleij J.A.C, Koevoets P., Van't Riet J., Bank R., Nijdam Y. &Ernst W.H.O. (1990) Poly ((-glutamylcysteinyl)glycines or phy-tochelatins and their role in cadmium tolerance of Silene vul-garis. Plant, Cell and Environment 13, 913-921.
Vogeli-Lange R. & Wagner G.J. (1996) Relationship between cad-mium, glutathione and cadmium-binding peptides (phy-tochelatins) in leaves of intact tobacco seedlings. Plant Science114, 11-18.
Wagner G.J. (1993) Accumulation of cadmium in crop plants andits consequences to human health. In Advances in Agronomy (ed.D. L. Sparks), pp. 173-212. Academic Press, Inc., California.
Woolhouse H.W. (1983) Toxicity and tolerance in the responses ofplants to metals. In Encyclopedia of Plant Physiology, Vol. 12C:Responses to the Chemical and Biological Environment (eds O.L. Lange, P. S. Nobel, C B. Osmond & H. Ziegler), pp. 245-300.Springer-Verlag, Berlin.
Yamagami K. & Tarayama H. (1981) Dynamics of caimodulin andcyclic AMP phosphodiesterase in plasma membranes of rat liv-ers and ascites hepatomas. Biochimica et Biophysica Acta 677,174-183.
Received 7 August 1996: received in revised form 9 October 1996;accepted for publication 11 October 1996
© 1997 Blackwell Science Ltd, Plant, Cell and Environment, 20, 600-608





















