
The Indian Erysiphaceae revisited
26
371 Nova Hedwigia 89 3—4 371—395 Stuttgart, November 2009 DOI: 10.1127/0029-5035/2009/0089-0371 0029-5035/09/0089-0371 $ 6.25 © 2009 J. Cramer in der Gebrüder Borntraeger Verlagsbuchhandlung, D-14129 Berlin · D-70176 Stuttgart The Indian Erysiphaceae revisited by Uwe Braun 1* and Yudhbir Singh Paul 2 1 Martin-Luther-Universität, Institut für Biologie, Bereich Geobotanik und Botanischer Garten, Herbarium, Neuwerk 21, D-06099 Halle (Saale), Germany e-mail: [email protected] 2 Department of Plant Pathology, C.S.K. Himachal Pradesh Krishi Vishvavidyalaya, Palampur - 176062 (H.P.), India e-mail: [email protected] With 11 figures Braun, U. & Y.S. Paul (2009): The Indian Erysiphaceae revisited. - Nova Hedwigia 89: 371–395. Abstract: Taxonomy and nomenclature of new Indian powdery mildew taxa described by Paul & Thakur (2006) have been revised and reassessed on the base of the current phylogenetic generic concept of the Erysiphaceae. The following species are validated, redescribed and illustrated: Erysiphe farmanii sp. nov., E. ficicola sp. nov., E. machiliana sp. nov., E. malloticola sp. nov., E. quercifolia sp. nov., Phyllactinia bauhiniae sp. nov. and P. cassiae-fistulae sp. nov. Furthermore, the following new combinations are introduced: Erysiphe religiosa comb. nov. (= Uncinula aspera var. solani), E. cyclobalanopsidis comb. nov. and E. kumaonensis comb. nov. Key words: Erysiphales, Erysiphe, Himachal Pradesh, Phyllactinia, powdery mildew fungi. Introduction Paul & Thakur (2006) published a monographic treatment of the powdery mildew fungi (Erysiphaceae) of Himachal Pradesh, India, and provided a comprehensive general survey (check list) of this fungal group in India. Numerous new species and varieties were introduced. Type material of all new taxa has recently been re-examined by the first author in connection with attempts to prepare a new, updated version of his old powdery mildew monograph (Braun 1987). However, all new taxa are unfortunately invalid according to ICBN, Art. 37.6, which demands that since 1990 indication of type material of new taxa must include one of the words “holotypus” or “typus” or its equivalent in a modern language in order to be validly published. The generic taxonomy of the new taxa in Paul & Thakur (2006) was largely based *corresponding author
Transcript of The Indian Erysiphaceae revisited

371
Nova Hedwigia 89 3—4 371—395 Stuttgart, November 2009
DOI: 10.1127/0029-5035/2009/0089-0371 0029-5035/09/0089-0371 $ 6.25© 2009 J. Cramer in der Gebrüder Borntraeger
Verlagsbuchhandlung, D-14129 Berlin · D-70176 Stuttgart
The Indian Erysiphaceae revisited
by
Uwe Braun1* and Yudhbir Singh Paul2
1Martin-Luther-Universität, Institut für Biologie, Bereich Geobotanik und Botanischer Garten,Herbarium, Neuwerk 21, D-06099 Halle (Saale), Germany
e-mail: [email protected] of Plant Pathology, C.S.K. Himachal Pradesh Krishi Vishvavidyalaya,
Palampur - 176062 (H.P.), Indiae-mail: [email protected]
With 11 figures
Braun, U. & Y.S. Paul (2009): The Indian Erysiphaceae revisited. - Nova Hedwigia 89: 371–395.
Abstract: Taxonomy and nomenclature of new Indian powdery mildew taxa described by Paul &Thakur (2006) have been revised and reassessed on the base of the current phylogenetic genericconcept of the Erysiphaceae. The following species are validated, redescribed and illustrated: Erysiphefarmanii sp. nov., E. ficicola sp. nov., E. machiliana sp. nov., E. malloticola sp. nov., E. quercifoliasp. nov., Phyllactinia bauhiniae sp. nov. and P. cassiae-fistulae sp. nov. Furthermore, the followingnew combinations are introduced: Erysiphe religiosa comb. nov. (= Uncinula aspera var. solani),E. cyclobalanopsidis comb. nov. and E. kumaonensis comb. nov.
Key words: Erysiphales, Erysiphe, Himachal Pradesh, Phyllactinia, powdery mildew fungi.
Introduction
Paul & Thakur (2006) published a monographic treatment of the powdery mildewfungi (Erysiphaceae) of Himachal Pradesh, India, and provided a comprehensivegeneral survey (check list) of this fungal group in India. Numerous new species andvarieties were introduced. Type material of all new taxa has recently been re-examinedby the first author in connection with attempts to prepare a new, updated version ofhis old powdery mildew monograph (Braun 1987). However, all new taxa areunfortunately invalid according to ICBN, Art. 37.6, which demands that since 1990indication of type material of new taxa must include one of the words “holotypus”or “typus” or its equivalent in a modern language in order to be validly published.The generic taxonomy of the new taxa in Paul & Thakur (2006) was largely based
*corresponding author
eschweizerbartxxx author

372
on the traditional concept as circumscribed in Braun (1987). However, comprehensivephylogenetic analyses of powdery mildew fungi have been published since 1999 andinfluenced the generic taxonomy of the Erysiphales strongly (Takamatsu et al. 1999,2000, 2008 a, b; Saenz and Taylor 1999; Mori et al. 2000; Hirata et al. 2000;Khodaparast et al. 2001; Matsuda & Takamatsu 2003; Hirose et al. 2005). Braun(1999) and Braun & Takamatsu (2000) discussed taxonomic consequences of theresults of molecular sequence analyses and proposed a new phylogenetic genericconcept. Cook et al. (1997) examined powdery mildew anamorphs in detail using ascanning electron microscopic approach, and introduced a new concept of theanamorph genus Oidium Link. Phylogeny and current taxonomy of the Erysiphaceaewere summarised by Braun et al. (2002). The re-examination of the new Indianpowdery mildew taxa was necessary in order to compare them within a worldwidecontext of the Erysiphaceae, the new phylogenetic generic taxonomy and therequirements of the ICBN.
Material and methods
The collections examined were described, mounted in distilled water, using oil immersion (brightfield and phase contrast), but without any staining, by means of standard light microscopy (OlympusBX 50, Hamburg, Germany). 30 measurements (× 1000 magnification) of chasmothecia, conidiaand other structures were made, with the extremes given in parentheses. The collections examinedare deposited at the herbarium DPDH (Herbarium, Department of Plant Pathology, C.S.K. HimachalPradesh Krishi Vishvavidyalaya, Palampur, H.P., India) and partly at HAL (Herbarium, Martin-Luther-University, Institute of Biology, Geobotany and Botanical Garden, Halle, Germany).
Results and discussion
New taxa of powdery mildew fungi described by Paul & Thakur (2006) fromHimachal Pradesh, India, are listed in alphabetical order, followed by the taxonomicand nomenclatural status of the taxa concerned, based on the current generic taxonomy(Braun et al. 2002), brief discussions and illustrations of recognised, validated species(accepted names in bold).
(1) Arthrocladiella althaeae Y.S.Paul & V.K.Thakur, Indian Erysiphaceae: 57,Jodhpur 2006, nom. inval. (ICBN, Art. 37.6).
= Leveillula taurica (Lév.) G.Arnaud.
ILLUSTRATION: Paul & Thakur (2006: 58, Fig. 14).
MATERIAL EXAMINED: On Althaea rosea (L.) Cav. (Malvaceae), India, Himachal Pradesh, Sharboo(Kinnaur), 21 Sep. 2002, Y.S.Paul & V.K.Thakur (DPDH-407).
NOTES: The collection on Althaea rosea proved to be conspecific with L. taurica.The mycelium is partly internal and the anamorph pertains to the genus OidiopsisScalia. Relatively small catenate conidia mentioned in the original description belongpossibly to another anamorphic powdery mildew on the host leaves. The chasmotheciacoincide with those of L. taurica as well (170–225 μm diam., appendages numerous,irregularly branched, often coralloid; asci numerous, up to 30, stipitate, 80–110 ×
eschweizerbartxxx author

373
25–40 μm; with two large ascospores, 30–40 × 15–23 μm). Appendages ofArthrocladiella Vassilkov are different by being strictly dichotomously branched.
(2) Erysiphe aquilegiae var. clematidis Y.S.Paul & V.K.Thakur, Indian Erysiphaceae:38, Jodhpur 2006, nom. inval. (ICBN, Art. 37.6).
= Erysiphe aquilegiae var. ranunculi (Grev.) R.Y.Zheng & G.Q.Chen.
ILLUSTRATION: Paul & Thakur (2006: 43, Fig. 8a).
MATERIAL EXAMINED: On Clematis gouriana Roxb. (Ranunculaceae), India, Himachal Pradesh,Kamrunag (Lahaul & Spiti), 21 Oct. 2001, Y.S.Paul (DPDH-322).
NOTES: Collections on Clematis spp. are known to belong to E. aquilegiae var.ranunculi (Braun 1987). The specimen on C. gouriana is not sufficiently distinctfrom var. ranunculi to warrant a separate variety.
(3) Erysiphe betae var. spinacearum Y.S.Paul & V.K.Thakur, Indian Erysiphaceae:29, Jodhpur 2006, nom. inval. (ICBN, Art. 37.6).
= Erysiphe betae (Vaňha) Weltzien.
MATERIAL EXAMINED: On Spinacia oleracea L. (Chenopodiaceae), India, Himachal Pradesh, Bajaura,25 May 2003, Kumar (DPDH-425).
NOTES: Collections on Spinacea species are morphologically very close to andindistinguishable from E. betae on Beta spp. The appendages are occasionallybranched, and the conidia are cylindrical, 30–45 × 15–20 μm.
(4) Erysiphe cichoracearum var. saussureae Y.S.Paul & V.K.Thakur, IndianErysiphaceae: 27, Jodhpur 2006, nom. inval. (ICBN, Art. 37.1 and 37.6).
= Leveillula taurica (Lév.) G.Arnaud and Golovinomyces cichoracearum (DC.)Heluta.
ILLUSTRATION: Paul & Thakur (2006: 29, Fig. 4a,c).
MATERIAL EXAMINED: On Saussurea costus (Falc.) Lipsch. and Matricaria recutita L. (Asteraceae),India, Himachal Pradesh, Sharboo (Kinnaur), 21 Sep. 2002, Y.S.Paul & V.K.Thakur (DPDH-404,406).
NOTES: This variety was introduced on the base of two collections, one on Saussureacostus and another one on Matricaria recutita, but without designation of a type.The material on the two hosts is heterogeneous. The fungus on Saussurea pertains toLeveillula taurica (chasmothecia 150–190 μm diam.; anamorph belonging to Oidiopsis,primary conidia lanceolate, attenuated towards the apex, 50–60 × 15–20 μm,secondary conidia more or less cylindrical, 40–50 × 15–20 μm). The powdery mildewon Matricaria is identical with Golovinomyces cichoracearum (chasmothecia100–130 μm diam.; conidia in chains, 30–40 × 14–20 μm).
(5) Erysiphe cruciferarum var. indica Y.S.Paul & V.K.Thakur, Indian Erysiphaceae:29, Jodhpur 2006, nom. inval. (ICBN, Art. 37.6), var. chandra Y.S.Paul &V.K.Thakur (l.c.: 31), nom. inval. (ICBN, Art. 37.6) and var. cleomes (‘cleomae’)Y.S.Paul & V.K.Thakur (l.c.: 31), nom. inval. (ICBN, Art. 37.6).
eschweizerbartxxx author

374
= Erysiphe cruciferarum Opiz ex L.Junell.
ILLUSTRATION: Paul & Thakur (2006: 45, Fig. 9d,e).
MATERIAL EXAMINED: On Lepidium latifolium L. (Brassicaceae), India, Himachal Pradesh, Tabo(Lahaul & Spiti), 10 Aug. 2002, Y.S.Paul (DPDH-385); on Capsella bursa-pastoris (L.) Med.(Brassicaceae), Palampur, 16 Jul. 2002, Y.S.Paul (DPDH-375); on Cleome spinosa L. (Capparaceae),Maranda (Kangra), 6 Apr. 2003, Y.S.Paul (DPDH-422).
NOTES: The three varieties coincide with the morphological variability of this specieson a worldwide basis. Powdery mildew on Cleome spp. was described as Erysipheradulescui Docea. Braun (1987) reduced this species to synonym with E. cruciferarum.Powdery mildew fungi on hosts of the allied families Brassicaceae and Capparaceaeare morphologically indistinguishable, but it remains to be proven if they arebiologically and phylogenetically distinct.
(6) Erysiphe heraclei var. himalayensis Y.S.Paul & V.K.Thakur, Indian Erysiphaceae:25, Jodhpur 2006, nom. inval. (ICBN, Art. 37.6).
= Erysiphe heraclei DC.
ILLUSTRATION: Paul & Thakur (2006: 43, Fig. 8c).
MATERIAL EXAMINED: On Coriandrum sativum L. (Apiaceae), India, Himachal Pradesh, Palampur,23 Apr. 2004, V.K.Thakur (DPDH-364).
NOTES: Coriandrum sativum is known as host of E. heraclei (Amano 1986; Braun1987). The chasmothecial appendages are partly branched, and the structure and sizeof the conidiophores and the cylindrical conidia, 30–40 × 12–16 μm, agree wellwith those of E. heraclei.
(7) Erysiphe karpoorii Y.S.Paul & V.K.Thakur, Indian Erysiphaceae: 25, Jodhpur2006, nom. inval. (ICBN, Art. 37.6).
= Neoerysiphe geranii (Y.Nomura) U.Braun.
ILLUSTRATION: Paul & Thakur (2006: 47, Fig. 10d).
MATERIAL EXAMINED: On Geranium wallichianum D.Don (Geraniaceae), India, Himachal Pradesh,Karsog (Mandi), 22 Oct. 2001, Y.S.Paul & V.K.Thakur (DPDH-307).
NOTES: Due to a combination of lobed appressoria, conidia formed in chains andchasmothecia with immature asci before overwintering, this species belongs in thegenus Neoerysiphe. It is conspecific with N. geranii described by Nomura (1997)from Japan. The chasmothecia are 100–145 μm diam.; peridial cells irregularly shaped,5–20 μm diam.; appendages mycelioid, septate, thin-walled, 0.5–1.5 times as longas the chasmothecial diam., at first hyaline, later brown below and paler towards theapex; asci numerous, 40–60 × 25–35 μm, immature, without any spores beforeoverwintering. The appressoria are usually solitary, almost nipple-shaped to usuallylobed, 2–5 μm diam. The foot-cells of the conidiophores are cylindrical, 30–40 ×9–11 μm, followed by 1–3 short cells. The conidia are formed in chains, ellipsoid-doliiform, 25–35 × 12–16 μm.
eschweizerbartxxx author

375
(8) Erysiphe lindelofiae (‘lindelophiae’) Y.S.Paul & V.K.Thakur, Indian Erysi-phaceae: 48, Jodhpur 2006, nom. inval. (ICBN, Art. 37.6).
= Leveillula taurica (Lév.) G.Arnaud.
ILLUSTRATION: Paul & Thakur (2006: 47, Fig. 10c).
MATERIAL EXAMINED: On Lindelofia stylosa Brand (Boraginaceae), India, Himachal Pradesh, Moolang(Lahaul & Spiti), 10 Aug. 2002, Y.S.Paul (DPDH-391).
NOTES: E. lindelofiae proved to be identical with L. taurica. The anamorph pertainsto the genus Oidiopsis (mycelium partly internal, external mycelium forming dense,pannose, white patches; conidia solitary, primary conidia lanceolate, attenuated towardsthe apex, 40–70 × 15–24 μm; secondary conidia subcylindrical to ellipsoid-doliiform)and the chasmothecia agree well with those of L. taurica (mature chasmothecialarge, 150–190 μm diam.; appendages coralloid, often irregularly branched, 0.5–1(–2)times as long as the chasmothecial diam.; asci numerous, up to about 20, 65–100 ×30–50 μm, stipitate; with two large ascospores, 23–40 × 14–24 μm).
(9) Erysiphe orontii var. papaveris (‘papaverae’) Y.S.Paul & V.K.Thakur, IndianErysiphaceae: 47, Jodhpur 2006, nom. inval. (ICBN, Art. 37.6).
= Golovinomyces orontii (Castagne) Heluta.
ILLUSTRATION: Paul & Thakur (2006: 45, Fig. 9c).
MATERIAL EXAMINED: On Papaver somniferum L. (Papaveraceae), India, Himachal Pradesh, Kukumseri(Lahaul & Spiti), 12 Aug. 2002, Y.S.Paul (DPDH-378).
NOTES: Papaver spp. are known to be host species of G. orontii (Braun 1987). TheIndian specimen on P. somniferum agrees morphologically well with the latter species.Although Matsuda & Takamatsu (2003) published comprehensive examinations ofthe phylogenetic structure of the Golovinomyces cichoracearum complex, thetaxonomy of G. oronii is still insufficiently known, i.e. the latter species remains acompound species.
(10) Erysiphe pisi var. kukumi Y.S.Paul & V.K.Thakur, Indian Erysiphaceae: 25,Jodhpur 2006, nom. inval. (ICBN, Art. 37.6) and var. medicaginis Y.S.Paul &V.K.Thakur (l.c.: 26), nom. inval. (ICBN, Art. 37.6).
= Ersiphe pisi DC.
ILLUSTRATION: Paul & Thakur (2006: 30, Fig. 5c,d).
MATERIAL EXAMINED: On Medicago falcata L. and M. sativa L. (Fabaceae), India, Himachal Pradesh,Kukumseri (Lahaul & Spiti), 5 Aug. 2001, Y.S.Paul (DPDH-280, 281).
NOTES: The two varieties coincide with the morphological variability of this specieson a worldwide basis.
(11) Erysiphe polygoni var. rumicis Y.S.Paul & V.K.Thakur, Indian Erysiphaceae:35, Jodhpur 2006, nom. inval. (ICBN, Art. 37.6), var. kailashi Y.S.Paul &V.K.Thakur (l.c.: 35), nom. inval. (ICBN, Art. 37.6) and var. fagopyri Y.S.Paul &V.K.Thakur (l.c.: 36), nom. inval. (ICBN, Art. 37.6).
eschweizerbartxxx author

376
= Erysiphe polygoni DC.
ILLUSTRATION: Paul & Thakur (2006: 36, Fig. 6b–d).
MATERIAL EXAMINED: On Rumex patientia L. (Polygonaceae), India, Himachal Pradesh, Tabo (Lahaul& Spiti), 10 Aug. 2002, Y.S.Paul (DPDH-386); on Rheum australe D.Don, Sharboo (Kinnaur),21 Sep. 2002, Y.S.Paul & V.K.Thakur (DPDH-403); on Fagopyrum esculentum Moench, Kukumseri(Lahaul & Spiti), 10 Sep. 2001, Singh (DPDH-376).
NOTES: Braun (1987) assigned collections on Fagopyrum, Rheum and Rumex speciesto E. polygoni. They are morphologically indistinguishable from specimens onPolygonum species. Detailed molecular sequence analyses of E. polygoni on differenthosts are not yet available.
(12) Erysiphe rhododendri var. barotii Y.S.Paul & V.K.Thakur, Indian Erysiphaceae:46, Jodhpur 2006, nom. inval. (ICBN, Art. 37.6).
= Erysiphe rhododendri J.N.Kapoor.
ILLUSTRATION: Paul & Thakur (2006: 47, Fig. 10a).
MATERIAL EXAMINED: On Rhododendron sp. (Ericaceae), India, Himachal Pradesh, Tikkan, 2 Feb.2002, Y.S.Paul (DPDH-327).
NOTES: The differences mentioned in the original description are not sufficient for aseparate variety. The variability of E. rhododendri is insufficiently known.
(13) Erysiphe sordida var. similensis Y.S.Paul & V.K.Thakur, Indian Erysiphaceae:42, Jodhpur 2006, nom. inval. (ICBN, Art. 37.6) and var. spitiana Y.S.Paul &V.K.Thakur (l.c.: 44), nom. inval. (ICBN, Art. 37.6).
= Golovinomyces sordidus (L.Junell) Heluta.
ILLUSTRATION: Paul & Thakur (2006: 45, Fig. 9a,b).
MATERIAL EXAMINED: On Plantago major L. (Plantaginaceae), India, Himachal Pradesh, Kukumseri(Lahaul & Spiti), 5 Aug. 2001, Y.S.Paul (DPDH-284); on Plantago tibetica Hook. f., Tabo (Lahaul& Spiti), 10 Aug. 2002, Y.S.Paul (DPDH-410).
NOTES: The two varieties coincide with the morphological variability of this specieson a worldwide basis.
(14) Farmanomyces sphaerosporus (‘sphaerospora’) Y.S.Paul & V.K.Thakur, IndianErysiphaceae: 79, Jodhpur 2006, nom. inval. (ICBN, Art. 37.6).
≡ Erysiphe farmanii U.Braun & Y.S.Paul, sp. nov. Fig. 1
MycoBank, MB 512648
E. sikkimensis valde similis, sed appendicibus chasmotheciorum longioribus (ad 150 μm longis),non ramosis, et ascis 2–4-sporis.
ETYMOLOGY: Derived from Farman, in memory of the beloved father of Y.S.Paul.
ILLUSTRATION: Paul & Thakur (2006: 79, Fig. 22).
HOLOTYPE: On Quercus glauca Thunb. [≡≡≡≡≡ Cyclobalanopsis glauca (Thunb.) Oerst.] (Fagaceae),India, Himachal Pradesh, Salogra (Solan), 17 May 2003, Y.S.Paul & V.K.Thakur (Herbarium of theDepartment of Plant Pathology, C.S.K. Himachal Pradesh Krishi Vishvavidyalaya, Palampur, India,DPDH-424). ISOTYPES: HAL 2292 F, HCIO 45054, IMI 390879.
eschweizerbartxxx author

377
Mycelium mainly epiphyllous, forming white patches or effuse; conidiophores arisingfrom superficial hyphae, erect, cylindrical-filiform, 65–110 × 7–10 μm, a long foot-cell followed by 1–2 short cells; conidia solitary, subcylindrical, 24–32 × 10–13 μm.Chasmothecia mainly epiphyllous, scattered to gregarious, 60–85(–90) μm diam.;peridium thin, only composed of 2–3 cell layers, light brown, occasionally semitrans-parent, peridial cells irregularly polygonal, (5–)10–15(–20) μm diam.; appendagesfew, sparingly developed, to relatively numerous and well-developed, 0.3–1.5 timesas long as the chasmothecial diam. (25–150 × 3–8 μm), unbranched, mycelioid,colourless, wall somewhat thickened, aseptate, rough-walled; asci few, broadly ovoidto subglobose, sessile or subsessile, 40–50 × 40–45 μm when mature, wall up to1.5(–2) μm wide, 2–4-spored; ascospores subglobose to broadly ellipsoid-ovoid,12–20 × 8–15 μm, colourless.
NOTES: Erysiphe farmanii is morphologically close to E. sikkimensis Chona,J.N.Kapoor & H.S.Gill (Braun 1987; Chen et al. 1987), known from China andIndia on Quercus spp., including Q. glauca. The latter species is, however,distinguished by having very short, often branched appendages and (3–)4–7(–8)-spored asci. E. gracilis R.Y.Zheng & G.Q.Chen (Braun, 1987; Chen et al. 1987) isanother similar species on Quercus spp. in Asia, but differs in having largerchasmothecia, 80–110(–120), with long, often branched appendages, 0.5–4 times aslong as the chasmothecia, and (3–)5–7-spored asci. The new genus FarmanomycesY.S.Paul & V.K.Thakur (Paul & Thakur 2006: 79) was compared with ErysipheDC. and Typhulochaeta S.Ito & Hara and distinguished by its thin peridial wall. Inthis respect, Farmanomyces is similar to the genus Brasiliomyces Viégas (Zheng1984; Braun 1987). However, the structure of the peridium in E. farmanii isintermediate between Erysiphe and Brasiliomyces. The peridia are rather thin,
Fig. 1. Erysiphe farmanii. A. conidia, B. chasmothecium, C. appendage, D. asci. Bars: Chasmothecium= 50 μm, conidia and asci = 10 μm. U.Braun del.
eschweizerbartxxx author

378
composed of two to three cell layers, light brown and only slightly transparent. InBrasiliomyces, the peridium is thinner, paler and semitransparent. Due to its2–4-spored asci, E. farmanii resembles Brasiliomyces trina (Harkn.) R.Y.Zheng(= Erysiphe trina Harkn.), known from the southern USA on Lithocarpus sp. andQuercus agrifolia. Mori et al. (2000) carried out comprehensive molecular sequenceanalyses. B. trina was included in these examinations and clustered among otherspecies of the Erysiphe emend. U.Braun & S.Takam. (incl. Microsphaera Lév. andUncinula Lév.) clade. Therefore, the original status of this species as member of thegenus Erysiphe has to be reinstated, and Brasiliomyces should be reduced to synonymywith the latter genus. Braun & Takamatsu (2000) and Braun et al. (2002) tentativelymaintained Brasiliomyces, above all since the anamorphs of Brasiliomyces specieswere still unknown. Although conidiophores and conidia of E. malachae Seaver(= Brasiliomyces malvastri Viégas), the type species of Brasiliomyces, are stillunknown, the hyphae of this species are provided with lobed appressoria as in Erysiphespp. A second Brasiliomyces-like species of Erysiphe has been described from Taiwanon Quercus glauca: Erysiphe cyclobalanopsidis (K.C.Kuo, W.H.Hsieh & L.S.Leu)U.Braun, comb. nov. (Basionym: Brasiliomyces cyclobalanopsidis K.C.Kuo,W.H.Hsieh & L.S.Leu, Mycol. Res. 96: 702, 1992; MycoBank, MB 512650). Thisspecies is, however, quite distinct from E. farmanii by having 8-spored asci andlacking chasmothecial appendages (Kuo et al. 1992). Erysiphe kumaonensis(N.Ahmad, A.K.Sarbhoy & Kamal) U.Braun, comb. nov. (Basionym: Brasiliomyceskumaonensis N.Ahmad, A.K.Sarbhoy & Kamal, Mycol. Res. 102: 31, 1998;MycoBank, MB 512653), an Indian species on Quercus sp., is also easily distin-guishable from E. farmanii by its larger chasmothecia, 80–95 μm diam., with4–6-spored asci (Ahmad et al. 1998). To-anun et al. (2003) described the new speciesBrasiliomyces chiangmaiensis To-anun & S.Takam. (on Dalbergia cultrata, Thailand)and provided a key to Brasiliomyces species. The following key contains species ofthe genus Erysiphe with lacking or mycelioid appendages (Erysiphe sect. Erysiphe)on Quercus spp.:
1. Chasmothecia large, 75–110(–120) μm diam., average above 80 μm............................................. 21. Chasmothecia smaller, average below 80 μm................................................................................ 3
2. Chasmothecia 80–110(–120) μm diam.; appendages long, 0.5–4 times as long as the chasmothecialdiam., often branched; asci (3–)5–7-spored; on Quercus spp., Asia............................... E. gracilis
2. Chasmothecia (75–)80–90(–95) μm diam.; peridial wall thin, one-layered; appendages lackingor almost so, few and rudimentary; asci 4–6-spored; on Quercus sp., India............. E. kumaonensis
3. Chasmothecia very small, diameter below 50 μm; appendages lacking; asci two, 8-spored; onQuercus glauca, Taiwan................................................................................ E. cyclobalanopsidis
3. Chasmothecia larger, 50–85 μm diam., appendages mostly present.............................................. 4
4. Chasmothecia 50–70 μm diam.; peridium thin, one-layered, semitransparent; asci 2-spored; asco-spores large, 25–40 × 13–18 μm; on Quercus agrifolia, USA........................................ E. trina
4. Chasmothecia 60–85 μm diam.; peridium thicker, at least two cell layers, not or only slightlytransparent; asci 2–8-spored; ascospores smaller, up to 25 × 15 μm.............................................. 5
5. Appendages very short, 0.1–0.5 times as long as the chasmothecial diam., often branched; asci(3–)4–7(–8)-spored; on Castanopsis and Quercus spp., Asia.................................. E. sikkimensis
5. Appendages longer, 0.3–1.5(–2) times as long as the chasmothecial diam., unbranched; asci2–4-spored; on Quercus glauca, India.......................................................................... E. farmanii
eschweizerbartxxx author

379
(15) Phyllactinia bauhiniae Y.S.Paul & V.K.Thakur, Indian Erysiphaceae: 72,Jodhpur 2006, nom. inval. (ICBN, Art. 37.6).
= Phyllactinia bauhiniae Y.S.Paul, sp. nov. Fig. 2
MycoBank, MB 512654
Phyllactiniae cassiae similis, sed conidiis secundariis lageniformibus, apice inflato, et chasmotheciismajoribus, 180–220 μm diam., appendicibus brevioribus, 100–160 μm longis. Differt a Phyllactiniacaesalpiniae mycelio persistenti et appendicibus brevioribus, 100–160 μm longis.
ETYMOLOGY: Derived from the host genus.
ILLUSTRATION: Paul & Thakur (2006: 75, Fig. 20d).
HOLOTYPE: On Bauhinia sp. (Fabaceae), India, Himachal Pradesh, Samela (Kangra), 8 Mar. 2002,Y.S.Paul & V.K.Thakur (Herbarium of the Department of Plant Pathology, C.S.K. Himachal PradeshKrishi Vishvavidyalaya, Palampur, India, DPDH-345). ISOTYPE: HAL 2283 F.
Mycelium hypophyllous, effuse, thin, white, persistent, internal and external;conidiophores arising from superficial hyphae, erect, straight, cylindrical-filiform,80–95 μm long and (4–)5–7(–8) μm wide, composed of a long foot-cell, followedby short cells; conidia solitary, dimorphic, primary conidia lanceolate, secondaryconidia pin-like (narrowly lageniform with enlarged apex), apically rounded tosubtruncate, 50–80 × 10–17 μm, germ tubes lateral near the base, short to moderatelylong, apex unswollen to somewhat swollen. Chasmothecia hypophyllous, scattered
Fig. 2. Phyllactinia bauhiniae. A. conidiophore with conidium, B. primary conidium, C. primaryconidia with germ tube, D. secondary conidia. Bar = 10 μm. U.Braun del.
eschweizerbartxxx author

380
to gregarious, 180–220 μm diam., peridial cells irregularly polygonal, 10–20 μmdiam.; penicillate cells terminal; appendages 6–8, equatorial, acicular with bulbosebase, shorter than the chasmothecial diam. (120–160 μm), hyaline, bulbose base30–35 μm diam., attenuated towards the apex, hyaline, wall thickened below,thin above, smooth; asci numerous, more than 15, ellipsoid-ovoid, saccate,stipitate, 70–85 × 25–40 μm, 2-spored; ascospores broadly ellipsoid-ovoid,25–32 × 14–18 μm.
NOTES: The new species is morphologically very similar to Phyllactinia cassiaeG.J.M.Gorter & Eicker, described from South Africa on Cassia beareana, Caesal-piniaceae (Gorter & Eicker 1987), a species which is also characterised by havinglanceolate-ventricose conidia, resembling those of Leveillula taurica. However,P. cassiae is easily distinguishable from P. bauhiniae by its smaller chasmothecia,(125–)160–165(–190) μm diam., with longer appendages, about as long as thechasmothecial diam. The conidia may become somewhat lageniform, but they arenot pin-shaped, i.e. the apex remains unswollen. P. caesalpiniae Y.N.Yu (Chen etal. 1987), described from China on Caesalpinia spp. and Gleditsia macracantha,differs from P. bauhiniae in having evanescent mycelium and much longerchasmothecial appendages (up to twice as long as the chasmothecial diam.). Theanamorph of P. caesalpiniae is still unknown.
(16) Phyllactinia bauhiniae var. cassiae Y.S.Paul & V.K.Thakur, IndianErysiphaceae: 72, Jodhpur 2006, nom. inval. (ICBN, Art. 37.6).
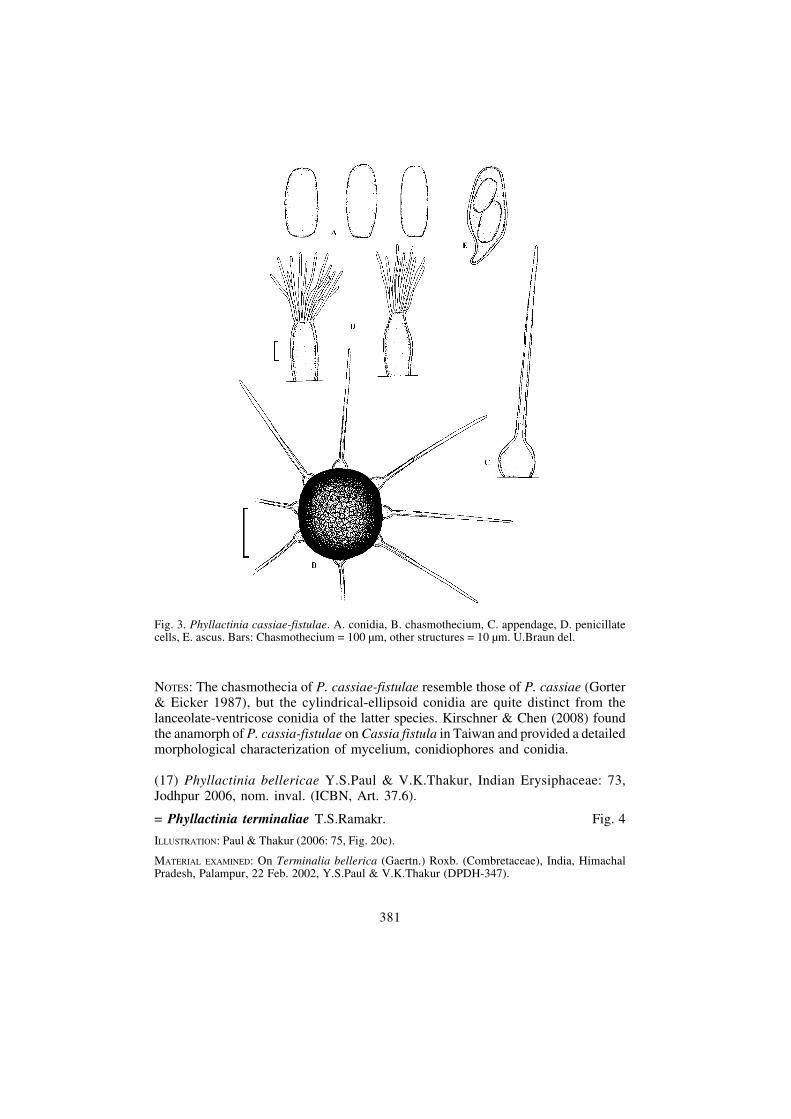
≡ Phyllactinia cassiae-fistulae U.Braun & Y.S.Paul, sp. nov. Fig. 3
MycoBank, MB 512655
Phyllactiniae cassiae similis, sed conidiis cylindraceis-ellipsoidibus.
ETYMOLOGY: Derived from the host genus.
DESCRIPTION (anamorph): Kirschner & Chen (2008).
ILLUSTRATION: Paul & Thakur (2006: 75, Fig. 20e), Kirschner & Chen (2008: Figs 1, 2 A-F).
HOLOTYPE: On Cassia fistula L. (Fabaceae), India, Himachal Pradesh, Panjgrain (Barmana), 12 Dec.2001, Y.S.Paul & V.K.Thakur (Herbarium of the Department of Plant Pathology, C.S.K. HimachalPradesh Krishi Vishvavidyalaya, Palampur, India, DPDH-325). ISOTYPE: HAL 2285 F.
Mycelium amphigenous, mainly hypophyllous, effuse, thin to dense, white, internaland external; conidiophores arising from external hyphae, erect, straight, cylindrical-filiform, up to about 130 × 6 μm; conidia solitary, cylindrical-ellipsoid, 40–50 ×10–20 μm, base rounded to truncate, apex broadly rounded, germ tubes at the ends.Chasmothecia mainly epiphyllous, scattered, (150–)160–190 μm diam.; peridial cellsirregularly polygonal, 10–20 μm diam.; penicillate cells terminal, 40–60 × 10–15 μm,stem subcylindrical to ampulliform, unbranched; appendages 8–12, equatorial,acicular with bulbose base, 0.75–1.25 times as long as the chasmothecial diam.(80–185 μm), hyaline, bulbose base 25–35 μm diam., attenuated towards the tip,wall thickened below, thin above, smooth; asci numerous, more than 20, saccate,stipitate, 50–75 × 25–30 μm, wall up to 2 μm wide, 2-spored; ascospores broadlyellipsoid-ovoid, 20–32 × 9–19 μm.
eschweizerbartxxx author
381
NOTES: The chasmothecia of P. cassiae-fistulae resemble those of P. cassiae (Gorter& Eicker 1987), but the cylindrical-ellipsoid conidia are quite distinct from thelanceolate-ventricose conidia of the latter species. Kirschner & Chen (2008) foundthe anamorph of P. cassia-fistulae on Cassia fistula in Taiwan and provided a detailedmorphological characterization of mycelium, conidiophores and conidia.
(17) Phyllactinia bellericae Y.S.Paul & V.K.Thakur, Indian Erysiphaceae: 73,Jodhpur 2006, nom. inval. (ICBN, Art. 37.6).
= Phyllactinia terminaliae T.S.Ramakr. Fig. 4
ILLUSTRATION: Paul & Thakur (2006: 75, Fig. 20c).
MATERIAL EXAMINED: On Terminalia bellerica (Gaertn.) Roxb. (Combretaceae), India, HimachalPradesh, Palampur, 22 Feb. 2002, Y.S.Paul & V.K.Thakur (DPDH-347).
Fig. 3. Phyllactinia cassiae-fistulae. A. conidia, B. chasmothecium, C. appendage, D. penicillatecells, E. ascus. Bars: Chasmothecium = 100 μm, other structures = 10 μm. U.Braun del.
eschweizerbartxxx author

382
NOTES: The material on T. bellerica agrees well with P. terminaliae, described fromBihar (India) on T. chebula Retz. (Ramakrishnan 1957). Patil (1962) recorded anddescribed this species from Maharashtra, India, on T. bellerica, T. chebula andT. tomentosa Wight & Arn.. The collection on T. bellerica from Himachal Pradeshis characterised as follows: Chasmothecia 200–250 μm diam.; penicillate cells up to100 μm long, stem subcylindrical, 8–15 μm wide, unbranched or with 1–3 short,peg-like terminal branchlets; appendages 5–12, (0.5–)1–2.5 times as long as thechasmothecial diam.; asci 6–12, 70–85 × 25–45 μm, 2-spored; ascospores 20–45 ×15–27 μm. Braun (1987) considered P. terminaliae a synonym of P. guttata(Wallr. : Fr.) Lév. However, based on molecular sequence analyses (Takamatsu etal. 2008a) and morphological differences in the shape and size of penicillate cells(Shin & Li 2002 ), it could be shown that a narrower species concept has to beapplied within Phyllactinia, i.e. the compound species P. guttata s. lat. has to besplit into several species.
(18) Phyllactinia guttata var. cedrelae Y.S.Paul & V.K.Thakur, Indian Erysiphaceae:78, Jodhpur 2006, nom. inval. (ICBN, Art. 37.6).
= Phyllactinia toonae Y.X.Yu & Y.Q.Lai. Fig. 5
ILLUSTRATION: Paul & Thakur (2006: 76, Fig. 21e).
MATERIAL EXAMINED: On Toona ciliata M.Roem. [≡≡≡≡≡ Cedrela toona Roxb.] (Meliaceae), India, HimachalPradesh, Palampur, 19 Feb. 2002, Y.S.Paul & V.K.Thakur (DPDH-342).
NOTES: The Indian Phyllactinia collection on Toona ciliata (≡≡≡≡≡ Cedrela toona) agreesperfectly with P. toonae, known from China on Melia azedarach L., Toona sinensis(A.Juss.) M.Roem. (= Cedrela sinensis A.Juss.) and T. microcarpa (DC.) Harms(Chen et al. 1987). The conidia are broadly clavate-ovoid to soleiform-cymbiformto somewhat rhomboidal, sometimes with an apiculus-like apex, 40–80 × 18–32 μm.The Phyllactinia on Toona spp. is considered to be a separate species, easilydistinguishable from P. guttata s. str. by its conidial shape and penicillate cells with
Fig. 4. Phyllactinia terminaliae. A. penicillate cells, B. ascus. Bar = 10 μm. U.Braun del.
eschweizerbartxxx author

383
a subcylindrical, unbranched stem. P. guttata s. str. on Corylus spp. is distinguishedby a unique type of penicillate cells characterised by having a subcylindrical stem,divided into 2–10(–16) short, bulbose branchlets.
(19) Phyllactinia guttata var. populina Y.S.Paul & V.K.Thakur, Indian Erysiphaceae:77, Jodhpur 2006, nom. inval. (ICBN, Art. 37.6).
= Phyllactinia populi (Jacz.) Y.N.Yu.
ILLUSTRATION: Paul & Thakur (2006: 76, Fig. 21d).
MATERIAL EXAMINED: On Populus sp. (Salicaceae), India, Himachal Pradesh, Karsog (Mandi),20 Oct. 2001, Y.S.Paul & V.K.Thakur (DPDH-333).
NOTES: Yu (in Yu & Lai 1979) introduced the combination Phyllactinia populi forAsian collections on Populus spp. This name was based on P. suffulta f. populi Jacz.(Jaczewski 1927), which is with regard to morphological characters a nomen nudum,solely based on host specificity, i.e. on its occurrence on Populus spp. However, inplant pathogenic taxa such names are not considered to be invalid. Thus, P. populi isavailable as valid name. Braun (1987) reduced P. populi to synonymy with P. guttatas. lat., but Phyllactinia collections on Populus spp. differ from P. guttata s. str. (onCorylus spp.) in having quite distinct penicillate cells, usually with a subcylindrical,unbranched stem.
The Indian material on Populus sp. is characterised as follows: Mycelium hypo-phyllous, effuse, white, evanescent to subpersistent; conidia solitary, clavate-cymbiform, 60–80 × 20–25 μm. Chasmothecia amphigenous, scattered to gregarious,165–250 μm diam.; peridial cells irregularly polygonal, 10–20 μm diam.; penicillatecells terminal, up to 80 μm long and 10–15 μm wide, stem subcylindrical, unbranched;appendages 5–8, equatorial, acicular with bulbose base, 25–35(–40) μm diam.,attenuated towards the apex, about 1–1.5 times as long as the chasmothecial diam.(150–300 μm), hyaline, smooth, wall somewhat thickened below, thin-walled above;asci numerous, usually more than 15, saccate, short-stalked, 60–85 × 30–40 μm,wall up to 2 μm wide, 2-spored; ascospores broadly ellipsoid-ovoid, hyaline,25–30(–35) × 15–25 μm.
Fig. 5. Phyllactinia toonae. A. conidia, B. conidium with germ tube. Bar = 10 μm. U.Braun del.
eschweizerbartxxx author

384
(20) Phyllactinia guttata var. pyricola Y.S.Paul & V.K.Thakur, Indian Erysiphaceae:77, Jodhpur 2006, nom. inval. (ICBN, Art. 37.6).
= Phyllactinia pyri-serotinae Sawada Fig. 6
ILLUSTRATION: Paul & Thakur (2006: 76, Fig. 21f).
MATERIAL EXAMINED: On Pyrus sp. (Rosaceae), India, Himachal Pradesh, Palampur, 22 Nov. 2002,Y.S.Paul & V.K.Thakur (DPDH-419).
NOTES: The original material of var. pyricola is characterised as follows: Chasmothecia185–225 μm diam.; appendages 5–8, about as long as the chasmothecial diam., basalbulbose swelling 30–40 μm diam.; penicillate cells up to 100 μm long and 10–20 μmbroad, stem subcylindrical to ampulliform, with up to three short terminal branchlets;asci more than 20, saccate, stipitate, 60–80 × 30–40 μm, 2-spored; ascospores 30–38 ×18–25 μm. Collections of Phyllactinia on hosts of the Rosaceae are usually placed inPhyllactinia mali (Duby) U.Braun (Blumer 1967; Braun 1987; Shin 2000; Shin & Li2002). However, this name was based on European samples on species of Malus, Pyrusand other rosaceous hosts, characterised by having rather small chasmothecia, 140-185μm diam. (average below 170 μm), and clavate conidia. Asian collections on Pyrus spp.are distinguished by having larger chasmothecia and clavate-soleiform conidia. The nameP. pyri-serotinae is the oldest name available for this species. P. pyri (Castagne) Hommawas based on European material and is conspecific with P. mali.
(21) Phyllactinia guttata var. rosacearum Y.S.Paul & V.K.Thakur, IndianErysiphaceae: 74, Jodhpur 2006, nom. inval. (ICBN, Art. 37.6).
= Phyllactinia pyri-serotinae Sawada
ILLUSTRATION: Paul & Thakur (2006: 75, Fig. 20c).
MATERIAL EXAMINED: On Cotoneaster bacillaris Wall. ex Lindl. (Rosaceae), India, Himachal Pradesh,Shimla, 21 Oct. 2001, Y.S.Paul & V.K.Thakur (DPDH-309).
NOTES: The chasmothecia are 170–265 μm diam.; with 5–10 appendages, 0.75–2times as long as the chasmothecial diam.; penicillate cells up to 100 μm long and 10–20 μm wide, stem subcylindrical to lageniform, terminally bifurcate or with up tofour branchlets; asci 20–30, 70–90 × 30–45 μm, 2-spored; ascospores 20–35 ×(10–)15–20 μm. The material on Cotoneaster from India coincides well withcollections of P. pyri-serotinae on Pyrus spp.
(22) Podosphaera tridactyla var. prunicola Y.S.Paul & V.K.Thakur, IndianErysiphaceae: 51, Jodhpur 2006, nom. inval. (ICBN, Art. 37.6).
= Podosphaera tridactyla (Wallr.) de Bary.
ILLUSTRATION: Paul & Thakur (2006: 50, Fig. 11c).
MATERIAL EXAMINED: on Prunus cornuta (Wall. ex Royle) Steud. (Rosaceae), India, Himachal Pradesh,Madgran (Lahaul & Spiti), 12 Aug. 2002, Y.S.Paul & V.K.Thakur (DPDH-281).
NOTES: The specimen on P. cornuta is morphologically not distinct from othercollections on Prunus spp.
eschweizerbartxxx author

385
(23) Sphaerotheca ainsliaeae (‘ainsliae’) Y.S.Paul & V.K.Thakur, IndianErysiphaceae: 21, Jodhpur 2006, nom. inval. (ICBN, Art. 37.6).
= Podosphaera fusca (Fr.) U.Braun & Shishkoff.
ILLUSTRATION: Paul & Thakur (2006: 20, Fig. 2c).
MATERIAL EXAMINED: On Ainsliaea aptera DC. (Asteraceae), India, Himachal Pradesh, Karsog (Mandi),22 Oct. 2001, Y.S.Paul (DPDH-305).
NOTES: Small chasmothecia, 60–90 μm diam., and relatively small terminal oculi ofthe asci render this species a synonym of P. fusca s. str. (Braun & Takamatsu 2000).
(24) Sphaerotheca aphanis var. potentillae Y.S.Paul & V.K.Thakur, IndianErysiphaceae: 21, Jodhpur 2006, nom. inval. (ICBN, Art. 37.6).
= Podosphaera aphanis (Wallr.) U.Braun & S.Takam. var. aphanis.
ILLUSTRATION: Paul & Thakur (2006: 20, Fig. 2b).
MATERIAL EXAMINED: On Potentilla sp. (Rosaceae), India, Himachal Pradesh, Hansa (Lahaul & Spiti),9 Aug. 2002, Y.S.Paul (DPDH-281).
NOTES: The characters of var. potentillae coincide with the morphological variabilityof P. aphanis var. aphanis. The appendages are (0.5–)1–1.5 times as long as thechasmothecial diam., septate and pigmented. Potentilla spp. are infected byP. aphanis var. aphanis as well as P. aphanis var. hyalina (U.Braun) U.Braun &S.Takam. The latter variety differs from var. aphanis in having uniformly short,narrow, more or less hyaline chasmothecial appendages (Braun 1987).
Fig. 6. Phyllactinia pyri-serotinae. A. penicillate cells, B. ascus. Bar = 10 μm. U.Braun del.
eschweizerbartxxx author

386
(25) Sphaerotheca fusca var. compositarum Y.S.Paul & V.K.Thakur, Indian Erysi-phaceae: 20, Jodhpur 2006, nom. inval. (ICBN, Art. 37.6).
= Podosphaera fusca (Fr.) U.Braun & Shishkoff.
ILLUSTRATION: Paul & Thakur (2006: 19, Fig. 1e).
MATERIAL EXAMINED: On Senecio chrysanthemoides DC. (Asteraceae), India, Himachal Pradesh,Kukumseri (Lahaul & Spiti), 10 Aug. 2001, Y.S.Paul & V.K.Thakur (DPDH-281).
NOTES: The size of the chasmothecia and the characters of the asci agree with theconcept of P. fusca s. str. (Braun & Takamatsu 2000).
(26) Sphaerotheca fusca var. cucumis (‘cucumae’) Y.S.Paul & V.K.Thakur, IndianErysiphaceae: 18, Jodhpur 2006, nom. inval. (ICBN, Art. 37.6).
= Podosphaera xanthii (Castagne) U.Braun & Shishkoff.
ILLUSTRATION: Paul & Thakur (2006: 19, Fig. 1f).
MATERIAL EXAMINED: On Cucumis sativus L. (Cucurbitaceae), India, Himachal Pradesh, Sharboo(Kinnaur), 21 Sep. 2002, Y.S.Paul & V.K.Thakur (DPDH-402).
NOTES: Based on phylogenetic results (Hirata et al. 2000) and morphology, Braun etal. (2001) assigned previous collections of Sphaerotheca fusca (incl. S. cucurbitae(Jacz.) Z.Y.Zhao) on hosts of the Cucurbitaceae to Podosphaera xanthii. Thechasmothecia are relative large, 70–125 μm diam., with asci having large terminaloculi, 15–25 μm diam.
(27) Sphaerotheca heteropogonis (‘heteropogoni’) Y.S.Paul & V.K.Thakur, IndianErysiphaceae: 23, Jodhpur 2006, nom. inval. (ICBN, Art. 37.6).
= Podosphaera xanthii (Castagne) U.Braun & Shishkoff.
ILLUSTRATION: Paul & Thakur (2006: 20, Fig. 2d).
MATERIAL EXAMINED: On ‘Heteropogon contortus Roem. & Schult.’ (Poaceae), India, HimachalPradesh, Padar (Madi), 21 Oct. 2001, Y.S.Paul & V.K.Thakur (DPDH-341).
NOTES: Morphologically this species fully agrees with the feature of P. xanthii. Thechasmothecia are large, 70–130 μm diam., and the asci possess large terminal oculi,15–30 μm diam. The identity of the host plant is unclear, but in any case it does notbelong to Heteropogon contortus, and it is no member of the Poaceae at all. Theleaves, which are not grass-like, are lobed, possess a pinnate leaf venation, andbelong possibly to a composite.
(28) Uncinula aspera var. solani Y.S.Paul & V.K.Thakur, Indian Erysiphaceae: 60,Jodhpur 2006, nom. inval. (ICBN, Art. 37.6).
= Erysiphe religiosa (T.S.Ramakr.) U.Braun & Y.S.Paul, comb. nov. Fig. 7
MycoBank, MB 512656
Basionym: Uncinula religiosa T.S.Ramakr., Proc. Indian Acad. Sci., B, 48: 124, 1959.
ILLUSTRATION: Bappammal et al. (1995: 168, Fig. 105), Paul & Thakur (2006: 63, Fig. 16b).
DESCRIPTION: Bappammal et al. (1995: 169).
eschweizerbartxxx author

387
MATERIAL EXAMINED: On Ficus religiosa L. (Moraceae), India, Himachal Pradesh, Subathu (Solan),18 Jan. 2002, Y.S.Paul & V.K.Thakur (DPDH-332).
Mycelium amphigenous, persistent white patches or covers; hyphae branched, septate,thin-walled, smooth or almost so, 3–6.5 μm wide; appressoria multilobed; conidio-phores erect, straight, 50–90 μm long, foot-celles subcylindrical, straight to somewhatflexuous, 15–30 × 7.5–10 μm; conidia solitary, ellipsoid-cylindrical, 30–50 ×10–16 μm. Chasmothecia hypophyllous, scattered to subgregarious, 80–105 μm diam.;peridial cells irregularly polygonal, 10–20 μm diam.; appendages 14–25, more orless equatorial, (1–)1.5–2 times as long as the chasmothecial diam. (100–175 μm),4–6(–8) μm wide below, width mostly gradually increasing towards the apex, up to10 μm wide above, wall almost smooth to verruculose, above all below, wall thickenedbelow, becoming gradually thinner towards the tip, apex tightly circinate (-subheli-coid), coiled part not enlarged; asci 2–5, saccate, short-stalked, 45–60 × 30–45 μm,3–5-spored; ascospores ellipsoid-ovoid, colourless, 20–30 × 8–17 μm.
NOTES: Indian collections on Ficus religiosa, described as Uncinula religiosa, agreemorphologically well with Erysiphe aspera (Doidge) U.Braun & S.Takam.(= Uncinula aspera Doidge), described from South Africa on Ficus petersii, andwere included in the latter species by Braun (1987). However, Hosagoudar et al.
Fig. 7. Erysiphe religiosa. A. chasmothecium, B. appendage, C. ascus. Bars: Chasmothecium =50 μm, appendage and ascus = 10 μm. U.Braun del.
eschweizerbartxxx author

388
(1992) and Bappammal et al. (1995) considered U. religiosa a separate speciesmorphologically distinct from U. aspera. E. religiosa is distinguished from E. asperaby having smaller chasmothecia and appendages with a width gradually increasingfrom base to apex. In E. aspera, the width of the appendages is gradually increasingfrom base to apex or only increasing up to 2/3 to 3/4, and then again decreasing inwidth. Based on the original collection of U. aspera var. solani, these differencesand the status of the Ficus religiosa powdery mildew as separate species could beconfirmed. However, U. religiosa has to be reallocated to the genus Erysiphe sinceUncinula has been reduced to synonym with the latter genus due to a phylogeneticand morphological reassessment of the whole generic concept within the Erysipheae(Braun & Takamatsu 2000; Braun et al. 2002).
(29) Uncinula malloti (‘mallotii’) Y.S.Paul & V.K.Thakur, Indian Erysiphaceae:64, Jodhpur 2006, nom. inval. (ICBN, Art. 37.6).
≡ Erysiphe malloticola U.Braun & Y.S.Paul, sp. nov. Fig. 8
MycoBank, MB 512657
E. alchorneae similis, sed appendicibus basim versus crassitunicatis et latitude apicem versus saepegradatim crescenti.
ETYMOLOGY: Derived from the host genus (dwelling on Mallotus).
ILLUSTRATION: Paul & Thakur (2006: 65, Fig. 17b).
HOLOTYPE: On Mallotus philippinensis Müll.Arg. (Euphorbiaceae), India, Himachal Pradesh, Kangar(Nahan), 17 Jan. 2002, Y.S.Paul & V.K.Thakur (Herbarium of the Department of Plant Pathology,C.S.K. Himachal Pradesh Krishi Vishvavidyalaya, Palampur, India, DPDH-329). ISOTYPE: HAL2281 F.
Mycelium epiphyllous, forming thin, white, persistent patches. Chasmothecia amphi-genous, gregarious, (75–)90–140(–150) μm diam., peridium at first rather pale,yellowish to olivaceous-brown, later darker brown, cells irregularly polygonal,(5–)10–25(–30) μm diam., wall of the cells up to 1.5 μm wide; appendages (5–)10–40,more or less equatorial or occasionally somewhat in the upper half, straight tosomewhat curved, stiff to somewhat flexuous, 1–2 times as long as the chasmothecialdiam. (100–200 μm), 4–7 μm wide below, occasionally somewhat wider at the verybase, width subequal throughout or somewhat increasing towards the apex, but upperpart just below the coiled tip usually distinctly enlarged, up to 10 μm wide above,aseptate or with a single basal septum, up to 30 μm distant from the point of attachment,thick-walled below (up to 2 μm wide), becoming thin-walled towards the apex,smooth or verruculose below, apex tightly uncinate, circinate to helicoid, oftensomewhat enlarged; asci 8–15, broadly obovoid-saccate, short-stalked, 50–80 ×30–50 μm, wall up to 2 μm wide, 3–6-spored; ascospores broadly ellipsoid-ovoid,colourless, 20–35 × 12–18 μm.
NOTES: Among Uncinula-like species of the genus Erysiphe, E. malloticola resemblesE. alchorneae (R.Y.Zheng & G.Q.Chen) U.Braun & S.Takam. with regard tochasmothecial size, number of appendages, asci and ascospores (Braun 1987), but itdiffers in having appendages which are thick-walled towards the base and usuallyincreasing in width from base to apex. The epithet ‘malloti’ could not be used withinErysiphe since already preoccupied by E. malloti Zhi X.Chen & R.X.Gao.
eschweizerbartxxx author

389
(30) Uncinula miyabei var. alnicola Y.S.Paul & V.K.Thakur, Indian Erysiphaceae:61, Jodhpur 2006, nom. inval. (ICBN, Art. 37.6).
= Erysiphe miyabei (S.Salmon) U.Braun & S.Takam.
ILLUSTRATION: Paul & Thakur (2006: 63, Fig. 16d).
MATERIAL EXAMINED: On Alnus nitida Endl. (Betulaceae), India, Himachal Pradesh, Jhatingri, 2 Feb.2002, Y.S.Paul (DPDH-328).
NOTES: The Indian collection on Alnus nitida agrees well with the concept ofE. miyabei. 5–7-spored asci have been observed, although the asci were said to be2–3-spored.
(31) Uncinula mori var. moricola Y.S.Paul & V.K.Thakur, Indian Erysiphaceae:59, Jodhpur 2006, nom. inval. (ICBN, Art. 37.6).
= Erysiphe mori (Miyake) U.Braun & S.Takam.
ILLUSTRATION: Paul & Thakur (2006: 62, Fig. 15c).
MATERIAL EXAMINED: On Morus serrata Roxb. (Moraceae), India, Himachal Pradesh, Shimla,21 Oct. 2001, Y.S.Paul (DPDH-318).
NOTES: The features of the collection on M. serrata coincide with the variability ofE. mori.
Fig. 8. Erysiphe malloticola. A. chasmothecium, B. appendage, C. tips of appendages, D. ascus.Bars: Chasmothecium = 50 μm, appendage, tips and ascus = 10 μm. U.Braun del.
eschweizerbartxxx author

390
(32) Uncinula quercifolia Y.S.Paul & V.K.Thakur, Indian Erysiphaceae: 62, Jodhpur2006, nom. inval. (ICBN, Art. 37.6).
≡ Erysiphe quercifolia U.Braun & Y.S.Paul, sp. nov. Fig. 9
MycoBank, MB 512658
E. pyrenaicae similis, sed chasmotheciis 100–135 μm diam., appendicibus pluribus, (15–)25–40, etascis pluribus, 8–12.
ETYMOLOGY: Referring to the host genus.
ILLUSTRATION: Paul & Thakur (2006: 65, Fig. 17a).
HOLOTYPE: On Quercus incana Roxb. (Fagaceae), India, Himachal Pradesh, Subathu (Solan),18 Jan. 2002, Y.S.Paul & V.K.Thakur (Herbarium of the Department of Plant Pathology, C.S.K.Himachal Pradesh Krishi Vishvavidyalaya, Palampur, India, DPDH-330). ISOTYPE: HAL 2282 F.
Mycelium usually epiphyllous, forming white patches or effuse; conidia formedsingly, ellipsoid-cylindrical, 25–35 × 12–15 μm. Chasmothecia scattered to gregarious,100–135 μm diam.; peridial cells irregularly polygonal, (5–)10–20(–25) μm diam.;appendages (15–)25–40, more or less equatorial, straight to flexuous, 0.75–2 timesas long as the chasmothecial diam. (75–230 μm), 4–6 μm wide below, width almostequal throughout or width mostly gradually increasing towards the apex, up to
Fig. 9. Erysiphe quercifolia. A. conidia, B. chasmothecium, C. appendage, D. tips of appendages,E. ascus. Bars: Chasmothecium = 50 μm, appendage, tips and ascus = 10 μm. U.Braun del.
eschweizerbartxxx author

391
10 μm just below the coiled part, thick-walled below, up to 2 μm wide, thin-walledabove, rough-walled, aseptate or with a single septum at the very base, colourless,apex tightly circinate to subhelicoid, often slightly enlarged; asci 8–12, obovoid-saccate, short-stalked, thin-walled (wall up to 1.5 μm wide), 50–70 × 30–40 μm,(2–)3–5-spored; ascospores 20–35 × 10–18 μm.
NOTES: The new species on Quercus incana resembles Erysiphe pyrenaica (Vienn.-Bourg.) U.Braun & S.Takam., known from France on Quercus pyrenaica (Braun1987), but the latter species differs from E. quercifolia in having smaller chasmothecia,95–115 μm diam., with a smaller number of appendages (9–20), and few asci (3–5).
(33) Uncinuliella ficicola Y.S.Paul & V.K.Thakur, Indian Erysiphaceae: 68, Jodhpur2006, nom. inval. (ICBN, Art. 37.6).
≡ Erysiphe ficicola U.Braun & Y.S.Paul, sp. nov. Fig. 10
MycoBank, MB 512659
LATIN DESCRIPTION: Paul & Thakur (2006: 68, sub Uncinuliella ficicola).
ETYMOLOGY: Derived from the host genus (dwelling on Ficus).
ILLUSTRATION: Paul & Thakur (2006: 68, Fig. 18c).
HOLOTYPE: On Ficus roxburghii Wall. (Moraceae), India, Himachal Pradesh, Subathu (Solan),18 Jan. 2002, Y.S.Paul & V.K.Thakur (Herbarium of the Department of Plant Pathology, C.S.K.Himachal Pradesh Krishi Vishvavidyalaya, Palampur, India, DPDH-344). ISOTYPE: HAL 2279 F.
Mycelium amphigenous, forming white patches, persistent or almost so. Chasmotheciamainly epiphyllous, scattered to gregarious, 80–100 μm diam.; peridial cellsirregularly polygonal, 10–25 μm diam.; appendages dimorphic, with 15–35 long,Uncinula-like appendages, straight to slightly curved, not flexuous, 1.5–2.5 times aslong as the chasmothecial diam. (120–240 μm), 4–6 μm wide below, width increasingtowards the apex, 7–15 μm wide just below the coiled part, which is, however, notenlarged (rather slightly attenuated towards the ultimate tip), hyaline, occasionallybrown at the very base, aseptate, thick-walled below, becoming gradually thinnertowards the apex, smooth to verruculose below, apex tightly circinate; in additionwith numerous short, bristle-like appendages arising above the long Uncinula-likeappendages, subcylindrical to mostly attenuated towards the apex, unbranched, upto 30 × 6 μm, hyaline, aseptate, wall thin to slightly thickened, smooth or almost so;asci (2–)4–6, saccate, subsessile to short-stalked, 50–65 × 30–50 μm, wall up to2.5 μm wide, (2–)4–5-spored; ascospores broadly ellipsoid-ovoid, hyaline, 18–35 ×11–18 μm.
NOTES: This species belongs to Erysiphe species with dimorphic chasmothecialappendages, which have previously been placed in the genus Uncinuliella R.Y.Zheng& G.Q.Chen. In molecular sequence analyses, it could be demonstrated that formerUncinuliella species cluster together with other species of Erysiphe, incl. Microsphaeraand Uncinula (Mori et al. 2000). Hence, this genus was reduced to synonym withErysiphe emend. (Braun & Takamatsu 2000; Braun et al. 2002). E. ficicola is thefirst species of this morphological group, i.e. the former genus Uncinuliella, on amember of the Moraceae. There is no comparable species.
eschweizerbartxxx author

392
(34) Uncinuliella machiliana (‘machilliana’) Y.S.Paul & V.K.Thakur, IndianErysiphaceae: 67, Jodhpur 2006, nom. inval. (ICBN, Art. 37.6).
≡ Erysiphe machiliana U.Braun & Y.S.Paul, sp. nov. Fig. 11
MycoBank, MB 512660
LATIN DESCRIPTION: Paul & Thakur (2006: 67, sub Uncinuliella machiliana).
ETYMOLOGY: Referring to the host genus.
ILLUSTRATION: Paul & Thakur (2006: 68, Fig. 18b).
HOLOTYPE: On Machilus odoratissima Nees [= Persea odoratissima (Nees) Kosterm.] (Lauraceae),India, Himachal Pradesh, Saproon (Solan), 18 Jan. 2002, Y.S.Paul & V.K.Thakur (Herbarium ofthe Department of Plant Pathology, C.S.K. Himachal Pradesh Krishi Vishvavidyalaya, Palampur,India, DPDH-329). ISOTYPE: HAL 2280 F.
Mycelium amphigenous, mainly epiphyllous, in white patches or effuse, thin; conidiaformed singly, subcylindrical, 25–35 × 12–15 μm. Chasmothecia (80–)100–140 μmdiam., peridial cells irregularly polygonal, (5–)10–20(–25) μm diam.; appendagesdimorphic, with 20–35 long, Uncinula-like appendages, more or less equatorial,0.75–1.5(–2) times as long as the chasmothecial diam., 4–6 μm wide below,occasionally somewhat wider at the very base, width increasing towards the apex, up
Fig. 10. Erysiphe ficicola. A. chasmothecium, B. long, Uncinula-like appendage, C. tip of anappendage, D. short, bristle-like appendages, E. ascus. Bars: Chasmothecium = 50 μm, appendage,tips and ascus = 10 μm. U.Braun del.
eschweizerbartxxx author

393
to 13 μm in the coiled part and just below, hyaline, occasionally brown at the verybase, aseptate or with a single basal septum, wall thickened below, becoming graduallythinner towards the apex, completely smooth or verruculose below and smooth above,apex tightly circinate; in addition numerous short, bristle-like appendages arisingabove the long Uncinula-like appendages, subcylindrical to mostly attenuated towardsthe apex, unbranched, up to 30 × 7 μm, hyaline, aseptate, wall thin to slightlythickened, verruculose; asci 7–15, saccate, subsessile to short-stalked, 45–65 ×30–40 μm, wall thin, up to 1.5 μm wide, (2–)3–5-spored; ascospores broadly ellipsoid-ovoid, hyaline, 20–35(–40) × 9–15 μm.
NOTES: This species belongs to Erysiphe species with dimorphic chasmothecialappendages, which have previously been placed in the genus Uncinuliella.E. machiliana is the first species of this morphological group on a member of theLauraceae. There is no comparable species.
References
AHMAD, N., A.K. SARBHOY & KAMAL (1998): A new variety and two new species ofpowdery mildew from India. - Mycol. Res. 102: 30–31.
Fig. 11. Erysiphe machiliana. A. conidia, B. chasmothecium, C. long, Uncinula-like appendage,D. tip of an appendage, E. short, bristle-like appendages, F. ascus. Bars: Chasmothecium = 50 μm,appendage, tips and ascus = 10 μm. U.Braun del.
eschweizerbartxxx author

394
AMANO, K. (1986): Host range and geographical distribution of the powdery mildew fungi. -Tokyo.
BAPPAMMAL, M., V.B. HOSAGOUDAR & K. UDAIYAN (1995): Powdery mildews of TamilNadu, India. - New Botanist 22: 81–175.
BLUMER, S. (1967): Echte Mehltaupilze (Erysiphaceae). - Gustav Fischer, Jena.
BRAUN, U. (1987): A monograph of the Erysiphales (powdery mildews). - Beih. Nova Hedwigia89: 1–700.
BRAUN, U. (1999): Some critical notes on the classification and generic concept of the Erysiphaceae.- Schlechtendalia 3: 49–55.
BRAUN, U. & S. TAKAMATSU (2000): Phylogeny of Erysiphe, Microsphaera, Uncinula(Erysipheae) and Cystotheca, Podosphaera, Sphaerotheca (Cystotheceae) inferred from rDNA ITSsequences - some taxonomic consequences. - Schlechtendalia 4: 1–33.
BRAUN, U., N. SHISHKOFF & S. TAKAMATSU (2001): Phylogeny of Podosphaera sect.Sphaerotheca subsect. Magnicellulatae (Sphaerotheca fuliginea s.lat.) inferred from rDNA ITSsequences - a taxonomic interpretation. - Schlechtendalia 7: 45–52.
BRAUN, U., R.T.A. COOK, A.J. INMAN & H.D. SHIN (2002): The taxonomy of the powderymildew fungi. In BÉLANGER R.R. et al. (eds): The powdery mildews: a comprehensive treatise:13–54. APS Press, St. Paul.
CHEN, G.-Q., S.-J. HAN, Y.-Q. LAI, Y.-N. YU, R.-Y. ZHENG & Z.-Y. Zhao (1987): FloraFungorum Sinicorum, Vol. 1, Erysiphales. - Science Press, Beijing.
COOK, R.T.A., A.J. INMAN & C. BILLINGS (1997): Identification of anamorphs of powderymildews using morphological and host range data. - Mycol. Res. 101: 975–1002.
GORTER, G.J.M.A. & A. EICKER (1987): Additional first records of some powdery mildew fungiin South Africa, including a new species. - S. African J. Bot. 53: 93–97.
HIRATA, T., J.H. CUNNINGTON, U. PAKSIRI, S. LIMKAISANG, N. SHISHKOFF,B. GRIGALIUNAITE, Y. SATO & S. TAKAMATSU (2000): Evolutionary analysis of subsectionMagnicellulatae of Podosphaera section Sphaerotheca (Erysiphales) based on the rDNA internaltranscribed spacer sequences with special reference to host plants. - Canad. J. Bot. 78: 1521–1530.
HIROSE, S., S. TANDA, L. KISS, B. GRIGALIUNAITE, M. HAVRYLENKO & S. TAKA-MATSU (2005): Molecular phylogeny and evolution of the maple powdery mildew (Sawadaea,Erysiphaceae) inferred from nuclear rDNA sequences. - Mycol. Res. 109: 912–922.
HOSAGOUDAR, V.B., SIDDAPPA, V. VIJAYANTHI & K. UDAIYAN (1992): New ornoteworthy species of powdery mildews from Tamil Nadu, India. - Cryptog. Bot. 2: 351–345.
JACZEWSKI, A.A. (1927): Karmannyj opredelitel’ gribov. Muchnisto-rosjanye griby. - Leningrad.
KHODAPARAST, S.A., S. TAKAMATSU & G.-A. HEDJAROUDE (2001): Phylogeneticstructure of the genus Leveillula (Erysiphales: Erysiphaceae) inferred from the nucleotide sequencesof the rDNA ITS region with special reference to the L. taurica species compex. - Mycol. Res. 105:909–918.
KIRSCHNER, R. & C.-J. CHEN (2008): The Ovulariopsis anamorph of Phyllactinia bauhiniaevar. cassiae: first record outside India and morphological characterization. - Sydowia 60: 57–67.
KUO, K.-C., W.-H. HSIEH & C.-S. LEU (1992): Brasiliomyces cyclobalanopsidis sp. nov., a newpowdery mildew on Cyclobalanopsis glauca. - Mycol. Res. 96: 702–703.
MATSUDA, S. & S. TAKAMATSU (2003): Evolution of host-parasite relationships ofGolovinomyces (Ascomycetes: Erysiphaceae) inferred from nuclear rDNA sequences. - Molec.Phylogen. Evol. 27: 314–327.
eschweizerbartxxx author

395
MORI, Y., Y. SATO & S. TAKAMATSU (2000): Evolutionary analysis of the powdery mildewfungi using nucleotide sequences of the nuclear ribosomal DNA. - Mycologia 92: 74–93.
NOMURA, Y. (1997): Taxonomical study of the Erysiphaceae of Japan. - Yokendo LTD., Tokyo.
PATIL, S.D. (1962): The genus Phyllactinia Lév. in Maharashtra State. - J. Univ. Poona, Sci.Techol. Sect., 22: 119–125.
PAUL, Y.S. & V.K. THAKUR (2006): Indian Erysiphaceae. - Scientific Publishers, Jodhpur.
RAMAKRISHNAN, T.S. (1957): Notes on some fungi. - Proc. Indian Acad. Sci., B, 45: 176–180.
SAENZ, G.S. & J.W. TAYLOR (1999): Phylogeny of the Erysiphales (powdery mildews) inferredfrom internal transcribed spacer ribosomal DNA sequences. - Canad. J. Bot. 77: 150–168.
SHIN, H.-D. (2000): Erysiphaceae of Korea. - National Institute of Agricultural Science andTechnology, Suwon.
SHIN, H.-D. & H.-J. LI (2002): Morphology of penicillate cells in the genus Phyllactinia and itstaxonomic application. - Mycotaxon 83: 301–325.
TAKAMATSU, S., T. HIRATA, Y. SATO & Y. Nomura (1999): Phylogenetic relationships ofMicrosphaera and Erysiphe sect. Erysiphe (powdery mildews) inferred from the rDNA ITS sequences.- Mycoscience 40: 259–268.
TAKAMATSU, S., T. HIRATA & Y. Sato (2000): A parasitic transition from trees to herbsoccurred at least two times in tribus Cystotheceae (Erysiphaceae): Evidence from nuclear ribosomalDNA. - Mycol. Res. 104: 1304–1311.
TAKAMATSU, S., M. INAGAKI, S. NIINOMI, S.A. KHODAPARAST, H.D. SHIN,B. GRIGALIUNAITE & M. HAVRYLENKO (2008a): Comprehensive molecular phylogeneticanalysis and evolution of the genus Phyllactinia (Ascomycota: Erysiphales) and its allied genera. -Mycol. Res. 112: 299–315.
TAKAMATSU, S., M. HAVRYLENKO, S.M. WOLCAN, S. MATSUDA & S. NIINOMI(2008b): Molecular phylogeny and evolution of the genus Neoerysiphe (Erysiphaceae, Ascomycota).- Mycol. Res. 112: 639–649.
TO-ANUN, C., S. LIMKAISANG, W. FANGFUK, Y. SATO, U. BRAUN & S. TAKAMATSU(2003): A new species of Brasiliomyces (Erysiphaceae) on Dalbergia cultrata var. cultrata fromThailand. - Mycoscience 44: 447–451.
YU, Y.N. & Y.Q. LAI (1979): Taxonomic studies on the genus Phyllactinia of China II. Phyllactiniawith short perithecial appendages. - Acta Microbiol. Sin. 19: 11–23.
ZHENG, R.-Y. (1984): The genus Brasiliomyces (Erysiphaceae). - Mycotaxon 19: 281–289.
Received 11 December 2008, accepted in revised form 18 December 2008.
eschweizerbartxxx author

396
eschweizerbartxxx author




















