
Increases in flood magnitudes in California under warming climates
Upload
independentCategory
view
1download
0
INVITED REVIEW
The hydroclimatic and ecophysiological basis of cloud forest distributions undercurrent and projected climates
Rafael S. Oliveira1,2,*, Cleiton B. Eller1, Paulo R. L. Bittencourt1 and Mark Mulligan3
1Departamento de Biologia Vegetal, Instituto de Biologia, Universidade Estadual de Campinas (UNICAMP), Campinas, SP, Brazil,2School of Plant Biology, Faculty of Natural and Agricultural Sciences, The University of Western Australia, 35 Stirling Highway,
WA 6009, Australia and 3Department of Geography, King’s College London, Strand, London WC2R 2LS, UK* For correspondence. E-mail [email protected]
Received: 8 January 2014 Returned for revision: 28 January 2014 Accepted: 4 March 2014
† Background Tropical montane cloud forests (TMCFs) are characterized by a unique set of biological and hydrocli-matic features, including frequent and/or persistent fog, cool temperatures, and high biodiversity and endemism.These forests are one of the most vulnerable ecosystems to climate change given their small geographic range,high endemism and dependence on a rare microclimatic envelope. The frequency of atmospheric water deficitsfor some TMCFs is likely to increase in the future, but the consequences for the integrity and distribution of theseecosystems are uncertain. In order to investigate plant and ecosystem responses to climate change, we need toknow how TMCF species function in response to current climate, which factors shape function and ecology mostand how these will change into the future.† Scope This review focuses on recent advances in ecophysiological research of TMCF plants to establish a linkbetween TMCF hydrometeorological conditions and vegetation distribution, functioning and survival. The hydrauliccharacteristics of TMCF trees are discussed, together with the prevalence and ecological consequences of foliaruptake of fog water (FWU) in TMCFs, a key process that allows efficient acquisition of water during cloud immersionperiods, minimizing water deficits and favouring survival of species prone to drought-induced hydraulic failure.† Conclusions Fog occurrence is the single most important microclimatic feature affecting the distribution and func-tion of TMCF plants. Plants in TMCFs are very vulnerable to drought (possessing a small hydraulic safety margin),and the presence of fog and FWU minimizes the occurrence of tree water deficits and thus favours the survival ofTMCF trees where such deficits may occur. Characterizing the interplay between microclimatic dynamics andplant water relations is key to foster more realistic projections about climate change effects on TMCF functioningand distribution.
Key words: Tropical montane cloud forest, plant water relations, drought, cavitation, fog, hydraulic failure,ecohydrology, foliar water uptake, Drimys brasiliensis, climate change, neotropics.
INTRODUCTION
The climatic conditions associated with elevation in tropicallandscapes favour the occurrence of a unique and endangeredecosystem known as tropical montane cloud forests (TMCFs).Despite the occurrence of TMCFs in a wide range of climaticenvelopes (Jarvis and Mulligan, 2010), the main common cli-matic attribute for every TMCF is frequent and persistentcloud immersion (i.e. fog; Scholl et al., 2010; Bruijnzeel et al.,2011). Fog frequency and intensity is an important factor deter-mining several structural features of TMCFs (Grubb andWhitmore, 1966; Bruijnzeel and Veneklaas, 1998; Bruijnzeeland Hamilton, 2000; Bruijnzeel, 2001). As a general rule, thereis an increase in epiphyte cover and decrease in tree height,canopy stratification and leaf area index in high-altitudeTMCFs (also called upper montane cloud forests). TMCFslocated at lower altitudes (lower montane cloud forests) arecloser structurally to lowland tropical forests (Bruijnzeel andHamilton, 2000; Bruijnzeel, 2001; Bruijnzeel et al., 2011).
The climatic and structural characteristics of TMCFs arewidely assumed to be responsible for some of the ecosystem ser-vices provided by TMCFs. The environments of TMCFs are
thought to increase streamflow volume, not only because of theadditional inputs of cloud water interception (CWI), but alsobecause of the low average atmospheric demand and thus lowevapotranspiration, caused by the frequent cloud immersion(Bruijnzeel et al., 2011). Water quality may also be improvedby the role of the TMCF cover in reducing soil erosion and land-slides compared with other land uses (Sidle et al., 2006). Theseecosystem services might be extremely valuable in some regionsin which significant populations occur downslope and down-stream of cloud forests, and are sometimes considered as abasis for TMCF conservation programmes through ‘paymentfor ecosystem services’ schemes (Bruijnzeel et al., 2011).
Cloudforests are also extremely valuable froma biological con-servation point of view; the uniqueness of TMCF environments isalso reflected in their high biodiversity and endemism levels(Bruijnzeel et al., 2010a, b). These ecosystems have a unique flor-istic composition, significantly distinct from that of lowland trop-ical forest (Grubb and Whitmore, 1966; Bertoncello et al., 2011).Neotropical TMCFs present an abundance of temperate-climatetaxa, such as Podocarpus, Alnus, Drimys, Weinmannia andMagnoliaceae (Webster, 1995; Bertoncello et al., 2011). Basedon the disjunct distribution of these taxa in tropical landscapes
# The Author 2014. Published by Oxford University Press on behalf of the Annals of Botany Company. All rights reserved.
For Permissions, please email: [email protected]
Annals of Botany 113: 909–920, 2014
doi:10.1093/aob/mcu060, available online at www.aob.oxfordjournals.org
at BIB
LIO
TE
CA
CE
NT
RA
L U
NIV
EST
AD
UA
L C
AM
PINA
S on April 24, 2014
http://aob.oxfordjournals.org/D
ownloaded from

and palinological records, some authors suggest that the modernfloristic composition and distribution of Neotropical TMCFscould be explained by Pleistocene climatic fluctuations, causingexpansions and retractions in vegetation (Webster, 1995;Meireles, 2003; Bertoncello et al., 2011) and the reconnectionbetween North and South America during the Pliocene, whichallowed the migration of Andean and cordilleran taxa betweennorth and south (Webster, 1995). The relatively low endemismat species level, despite high generic endemism, suggests recentand rapid speciation in TMCFs (Webster, 1995).
Various assessments of the distribution of TMCFs exist. Themost comprehensive assessment of the distribution of cloudforests throughout the tropics is that compiled under the auspicesof UNEP–WCMC by Aldrich et al. (1997). This is a databasecomprising .560 point observations distributed throughout thetropics and representing areas that have been defined as cloudforests in the literature or by local experts. These point observa-tions have been used to help develop spatial assessments ofcloud forest distribution on the basis of nationally or regionallydefined elevational bands and remotelysensed forest cover assess-ments (Bubb et al., 2004; Scatena et al., 2010). The derived totalcover of TMCFs was estimated to be in the order of 215 000 km2
(1.4 % of the total area of all tropical forests). However, TMCFsare defined by the frequency and persistence of cloud cover, notby elevation, and Jarvis and Mulligan (2011) stress the verywide range ofclimatic and landscape situations (temperature, rain-fall, altitude, distance to sea and mountain size) represented by the.560 observed UNEP–WCMC cloud forest sites. Because thisclimatic variability is not just controlled by elevation, elevation-based approaches to estimate cloud forest distribution will beable to indicate the major cloud forest areas but they are notlikely to identify all cloud-affected forests and may thus representan underestimate of the true cloud forest distribution and extent.
The cloud frequency-based pan-tropical assessment ofMulligan (2010) models the distributions of cloud forest hydrocli-matically todefine the distribution of hydroclimaticcloud-affectedforests (CAFs) rather than elevationally or ecologically definedTMCFs (Fig. 1). The most affected CAFs will have ecologicaladaptations that are characteristic of TMCFs (ecologically or ele-vationally defined), but lesser CAFs may still be hydrologicallyand ecologically distinct from forests that are not cloud affectedbut might not be considered as cloud forest structurallyorecologic-ally. CAFs represent some 14.2 % of all tropical forests and coveran area of 2.21 Mkm2 between 23.58N and 358S (Mulligan, 2010).
The archipelagic distribution of TMCFs (Luna-Vega et al.,2001) and the relationship between altitude and TMCF structureand composition (Grubb and Whitmore, 1966; Bruijnzeel andHamilton, 2000; Bruijnzeel, 2001; Bertoncello et al., 2011;Bruijnzeel et al., 2011) raises the question of which ecophysiolo-gical traits TMCF plants possess that allow and restrict their dis-tributions to these specific hydroclimatic conditions. Addressingthis question will help provide a mechanistic basis to investigatehow these ecosystems will respond to the climatic changes pro-jected to affect tropical montane regions.
Temperature projections of general circulation models (GCMs)agree reasonably well that tropical mountains will see warmingover the next decades. Some models project an increase in theheight of cloud formation (‘cloud uplift’) and higher evapotrans-piration in tropical montane regions as a consequence of increas-ing earth surface temperatures (Still et al., 1999). These changes
may affect TMCF structure and functioning in a number ofways, from drought-induced mortality of some tree species(Lowry et al., 1973; Werner, 1988) to an upward shift inlowland fauna and flora and invasion of pre-montane andlowland tropical species (Pounds et al., 1999). There is muchless agreement between GCMs concerning the projected distribu-tion of rainfall in tropical mountains (Mulligan et al., 2011), anddifferent GCMs disagree in both the magnitude and direction ofchange of rainfall at the regional scale (Bruijnzeel et al., 2011).Given the spatial complexity of climate in general and rainfall inparticular in tropical mountains, the local scale impacts of theserainfall changes are impossible to project (Oliveira et al., 2014).Given their limitedgeographicextent, island isolationbyelevationand surrounding land use change, and strong dependence on aunique set of climate characteristics, it is clear that changes in rain-fall and temperaturewill lead to significant stress on these systems.
In this review, we intend to link TMCF unique hydrometeoro-logical conditions with TMCF vegetation distribution, function-ing and survival in current and future climates. We will do thatby coupling published and new data about TMCF plant waterrelations, including recent advances regarding foliar wateruptake (FWU; Eller et al., 2013; Goldsmith et al., 2013) and thehydraulic safety margin (Choat et al., 2012), with published andnew data about current and projected TMCF microclimate.
HYDROCLIMATIC CONDITIONS ANDHYDRAULIC FUNCTIONING OF TMCF TREES
Temporal and spatial patterns of fog occurrence in TMCFs
Mulligan (2010) calculates the lifting condensation level (LCL)for four periods of the day for each month on the basis of pan-tropical climatological data and finds very high frequencies atwhich LCL is at ground level (i.e. fog is possible) in the Andesand Central America, but also in Africa and to a lesser extentparts of South-east Asia. However, elevation was not a good sur-rogate for satellite-observed cloud frequency across the tropics.Although the minimum observed cloud frequency does increaselinearly with altitude (areas close to sea level having cloud fre-quencies of around 30 % in the tropics), sites at a particular alti-tude can show a range of cloud frequencies, depending on otherfactors. Nevertheless, at altitudes .1400 m a.s.l., cloud frequen-cies are generally .65 % (Mulligan, 2010).
Jarvis and Mulligan (2011) found the climate of the UNEP–WCMC TMCF sites to be highly variable, with an average rainfallof 2000mm year–1 and an average temperature of 17.7 8C. Theyalso found TMCFs to be wetter (rainfall being 184 mm year–1
higher on average), cooler (by 4.2 8C on average) and less season-ally variable than the average forall montane forests (defined as alltropical forests at .500 m elevation). These global generaliza-tions hide significant variability within and between sites.
Fog tends to occur much more frequently in the afternoon andnight (Mulligan, 2010) and may persist through the dryseason when rainfall is low or zero. This may be important hydro-logically and ecophysiologically in seasonally dry environments(Bruijnzeel et al., 2011). Observations of cloud frequency(2001–2006) based on the MODIS cloud climatology developedby Mulligan (2010) for CAF areas in Colombia (forest cover.40 %) show an area-average frequency of 0.66. Rainfallextracted from WorldClim for the same CAF areas and months
Oliveira et al. — Hydroclimatic conditions and ecophysiology of cloud forests910
at BIB
LIO
TE
CA
CE
NT
RA
L U
NIV
EST
AD
UA
L C
AM
PINA
S on April 24, 2014
http://aob.oxfordjournals.org/D
ownloaded from

FI G. 1. Global distribution of cloud-affected forests (CAFs) defined hydroclimatically (Mulligan, 2010) in South-east Asia and Oceania (A), Paleotropics (B) andNeotropics (C). Areas with .40 % tree cover are shown; the darkest shades are 100 % tree cover.
Oliveira et al. — Hydroclimatic conditions and ecophysiology of cloud forests 911
at BIB
LIO
TE
CA
CE
NT
RA
L U
NIV
EST
AD
UA
L C
AM
PINA
S on April 24, 2014
http://aob.oxfordjournals.org/D
ownloaded from

show 179 mm month–1 (Table 1). Diurnality of fog frequency forColombian TMCFs defined using the elevational limits of Bubbet al. (2004), CAFs defined by Mulligan (2010) and all land inColombia is shown in Table 2. Clearly CAFs do not have signifi-cantly greater cloud frequency than all land in Colombia except inthe evening – whereas the elevationally defined TMCFs have verylow observed cloud frequency at this time. High cloud frequencyduring the day will lead to a lower incident solar radiation andphotosyntheticallyactive radiation (PAR) loads, with an increaseddiffuse fraction of light radiation (Letts and Mulligan, 2005;Mercado et al., 2009), whereas high night-time cloud frequencywill tend to reduce outgoing long-wave radiation and thus dailytemperature range. In contrast to the pan-tropical mean, forColombia the mean annual rainfall for CAFs and TMCFs islower than for all land, though the monthly minimum for CAFs(97 mm) is higher than the minimum for all land (75 mm).Mean annual temperature for CAFs in Colombia is 18 8C (lowerthan all land at 24 8C) but not as low as for TMCFs at 13.25 8C.Jarvis and Mulligan (2011) show that rainfall seasonality ishighly variable between TMCF sites, with most showing low sea-sonality but some having a strong seasonality of rainfall.
Fog impacts the solar radiation, temperature and precipitationmean and seasonal behaviourof TMCFs, but perhaps the key com-ponent of TMCF climate of relevance to climate change studies isthe altitudinally and topographically controlled spatial variabilityof climate, which means that cloud forests occur over highlyrestricted ranges with sharp climatic gradients. Table 3 showschange in temperature, precipitation and cloud frequency for allland, TMCFsand CAFs (.40 % tree cover) in Colombia and indi-cates that, though gradients of cloud frequency are only slightlysteeper in TMCFs and CAFs compared with all land, gradientsof rainfall and temperature are much steeper and it is these
gradients that create sensitivity to climate change in cloud forestecosystems since these gradients create barriers to dispersal andmigration as cloud forest climates change.
Water use patterns of TMCF trees
The linkages between the highly variable hydrometeorologicalconditions in TMCFs and vegetation water use remain poorlyexplored. Though there are a paucity of studies quantifying treetranspiration in TMCFs compared with other systems,Bruijnzeel et al. (2011) are able to show a general negative rela-tionship between TMCF vegetation water use and altitude(Bruijnzeel et al., 2011). Forests located at higher altitudes aremore affected by fog (upper montane cloud forests and elfincloud forests) and transpire less (380.4+31.8 mm year–1) thanlower montane cloud forests (646+38.8 mm year–1) andlowland evergreen rain forests (1004+81.6 mm year–1). Thisnegative relationship between vegetation water use and altitudecould be attributed mostly to increased cloud cover (and thusreduced evaporative demand) at higher altitudes (Zotz et al.,1998) as well as reduced leaf area index (Bruijnzeel et al.,2011). Cavelier (1996) proposed that hydraulic inefficiencycould constrain TMCF tree transpiration, but several studiesshowed thatpeak transpiration ratesofTMCFtreesarecomparablewith those of lowland forests (Zotz et al., 1998; Feild andHolbrook, 2000; Santiago et al., 2010). Santiago et al. (2010)even showed that xylem area per unit of leaf area increased withaltitude in the Hawaiian tree species Metrosideros polymorpha.
TABLE 1. Climatic characteristics of CAFs, TMCFs and all land in Colombia
Variable Area Jan Feb Mar Apr May Jun Jul Aug Sep Oct Nov Dec Mean
Cloud frequency(fraction)
TMCFs* 0.36 0.51 0.63 0.76 0.79 0.74 0.72 0.66 0.77 0.74 0.62 0.57 0.66
CAFs 0.39 0.53 0.63 0.78 0.79 0.74 0.74 0.66 0.76 0.76 0.63 0.58 0.67All land 0.36 0.53 0.65 0.78 0.79 0.75 0.74 0.69 0.76 0.75 0.63 0.58 0.67
Rainfall (mm h– 1) TMCFs* 75 86 110 180 180 140 130 120 130 190 160 110 134.25CAFs 97 110 140 230 240 200 170 170 190 250 210 140 178.92All land 91 110 140 250 300 280 260 250 240 280 220 140 213.42
Temperature (8C) TMCF* 13 13 14 14 14 13 13 13 13 13 13 13 13.25CAFs 18 19 19 19 19 18 18 18 18 18 18 18 18.33All land 24 24 25 24 24 24 23 24 24 24 24 24 24.00
*Bubb et al. (2004); 2000–3500 m a.s.l.
TABLE 2. Diurnality of satellite observed cloud frequency (2000–2006) for CAFs, TMCF and all land in Colombia
Local time TMCFs* CAFs All land
0600–1200 0.62 0.66 0.671200–1800 0.78 0.75 0.751800–2400 0.56 0.8 0.762400–0600 0.62 0.6 0.62
*Bubb et al. (2004); 2000–3500 m a.s.l.
TABLE 3. Spatial variability in the climate characteristics ofTMCFs, CAFs and all land in Colombia expressed as the mean
gradient for each variable in each zone
Variable Area Mean gradient (units per km)
Cloud frequency (fraction) TMCFs* 0.012CAFs 0.012All land 0.011
Rainfall (mm h– 1) TMCFs* 90CAFs 110All land 46
Temperature (8C) TMCFs* 0.9CAFs 0.8All land 0.3
*Bubb et al. (2004); 2000–3500 m a.s.l.
Oliveira et al. — Hydroclimatic conditions and ecophysiology of cloud forests912
at BIB
LIO
TE
CA
CE
NT
RA
L U
NIV
EST
AD
UA
L C
AM
PINA
S on April 24, 2014
http://aob.oxfordjournals.org/D
ownloaded from

The TMCFs located at higher altitudes are usually exposed tomore persistent fog events (Grubb and Whitmore, 1966; Jarvisand Mulligan, 2011), a microclimatic condition that affects treewater relations through transpiration suppression and by the add-ition of a water subsidy to the ecosystem (Fig. 2). Transpirationsuppression caused by fog has been described in severalfog-affected ecosystems (Burgess and Dawson, 2004; Reinhardtand Smith, 2008; Limm et al., 2009), including TMCFs (Gotschet al., 2014; C. B. Eller et al., unpubl. data). The mechanismbehind this suppression is probably the decrease in atmosphericvapour pressure deficit (VPD) and PAR associated with fogevents (Reinhardt and Smith, 2008), which decreases the drivinggradient for water loss by the vegetation. The formation of awater film on leaves also limits gas exchange and contributes totranspiration suppression (Smith and McClean, 1989; Brewerand Smith, 1997; Letts and Mulligan, 2005). Moreover, high-altitude TMCFs are subjected to lower mean air temperatureswhen compared with lowland forests (Bruijnzeel et al., 2011;Jarvis and Mulligan, 2011), which leads to lower VPD and, conse-quently, lower plant transpiration rates.
Night-time transpiration is another common and importantcomponent of tree and ecosystem water balance in TMCFs(Dawson et al., 2007). The few studies of night-time transpir-ation in TMCFs show moderate to very high water losses atnight (Feild and Holbrook, 2000; Rosado et al., 2012; Gotschet al., 2014). The functional meaning of night-time transpirationis not completely clear, but it is often suggested that it can con-tribute to nutrient acquisition (Scholz et al., 2007; Snyderet al., 2008). Nocturnal sap flow in TMCF trees during drier
nights could compensate for the lack of nutrient acquisitionduring periods in which transpiration is suppressed by leaf-wetting events.
Soil water conditions might pose additional constraints on thewater use of TMCF vegetation. Extreme conditions, such aswater logging, constrain plant transpiration in some TMCFsbecause of poorly developed root systems or lower leaf area oftrees inhabiting anoxic soils (Jane and Green, 1985; Santiagoet al., 2000). Soil water deficits, documented in seasonally dryTMCF areas (Jarvis and Mulligan, 2011), can cause a decreasein tree crown conductance and constrain plant transpiration(Kumagai et al., 2004; 2005; Chu et al., 2014).
Stomatal behaviour of TMCF trees might be quite conserva-tive, closing in response to relatively low VPD (Jane andGreen, 1985), leading to the inhibitory effects of high VPD(.1–1.2 kPa) on tree transpiration even under non-limitingsoil water conditions (Motzer, 2005). This type of stomatalbehaviour is usually associated with plants vulnerable tohydraulic failure (McDowell et al., 2008). Despite the paucityof tree hydraulic data for TMCFs, Santiago et al. (2000) demon-strated that M. polymorpha trees from TMCFs are more suscep-tible to xylem cavitation than lowland forest trees. Drimysbrasiliensis, one of the most abundant and ubiquitous treespecies in Brazilian TMCFs (Bertoncello et al., 2011), also hasa hydraulic system that is a very vulnerable to drought, losing50 % of its hydraulic conductivity at –1.56 MPa (Fig. 3), avery high value when compared with the average –2.6 MPa fortropical forests (Choat et al., 2012). In addition, this specieshas a very narrow xylem hydraulic safety margin, indicating
A Clear day Long duration/high magnitudeleaf-wetting events
Short duration/low magnitudeleaf-wetting events
B C
FWU FWU
CWICWI
E
ysoil = –0·48 MPa ysoil = –0·19 MPa ysoil = –0·77 MPa
FI G. 2. Scenarios illustrating the direction and magnitude of water fluxes in tropical montane cloud forests (TMCFs) under contrasting micrometeorological condi-tions. In scenario (A), clear days and nights, TMCF trees lose water to the atmosphere by transpiration (E). In scenarios (B) and (C), leaf-wetting events suppress tran-spiration of TMCFtrees and provide additional water supply to the vegetation by cloudwater interception (CWI), which is thewater intercepted by the plant aerial tissuethat then drips to the soil, and by foliar water uptake (FWU) that is the water directly intercepted and absorbed by plant leaves which may be redistributed downwardsthrough the plant xylem to the soil (see Eller et al., 2013). The magnitude of FWU, CWI and water drip to the soil will depend on: (1) the duration and magnitude of fogevents; (2) canopy water storage capacity; and (3) atmosphere–soil water potential gradient (WPG). In scenario (B), fog events of high magnitude and long durationsaturate canopy water storage capacity and increase CWI, causing an increase in soil water potential and a decrease in FWU. In scenario (C), hydrological inputs of lowmagnitude and/or duration wet the canopy but not the soil, increasing the WPG and the magnitude of FWU. However, during the wet season or in very humid TMCFs,when the soil has high water potential, the FWU should be minor regardless of fog event intensity, because of the small WPG. Soil water potential values are monthly
means of the wettest month (–0.19 MPa) and driest month (–0.77 MPa) in a Brazilian cloud forest stand.
Oliveira et al. — Hydroclimatic conditions and ecophysiology of cloud forests 913
at BIB
LIO
TE
CA
CE
NT
RA
L U
NIV
EST
AD
UA
L C
AM
PINA
S on April 24, 2014
http://aob.oxfordjournals.org/D
ownloaded from

that this species operates close to the steepest point of its xylemvulnerability curve and is therefore very prone to catastrophicembolism (Fig. 3). These results support the view that TMCFtrees are particularly vulnerable to droughts and might dependon alternative water sources, such as cloud water, to avoid hy-draulic failure.
CLOUD WATER INPUTS IN TMCFs
Cloud water interception
Cloud water interception (CWI) and its subsequent precipitationas fog drip may represent a major hydrological input to TMCFs.There is a general trend of higher altitude TMCFs presentinghigher CWI values (Giambelluca and Gerold, 2011); however,the relative importance of this hydrological input varies consid-erably between sites because of the importance of vegetationstructure and epiphytism, fog frequency, fog water content, topo-graphic exposure, wind direction and wind speed (Bruijnzeelet al., 2011). Holwerda (2010) found CWI values as low as0.15 mm d– 1 (1.7 % of the rainfall at the site) in a Mexicanlower montane cloud forest, while Takahashi et al. (2010)found values as high as 3.3 mm d– 1 (37 % of the rainfall at thesite) in a Hawaiian lower montane cloud forest.
The hydrological relevance of CWI might vary seasonally andpeak during dry seasons when rainfall inputs are lowest. Brown(1996) used throughfall data (water captured below the canopyduring fog or rainfall) to investigate seasonal variation in CWIin a TMCF in Guatemala. He found that throughfall in anupper montane cloud forest, despite being relatively highduring the entire year, can exceed rainfall by 147 mm duringthe dry season. Holder (2004) estimates that the contribution offog precipitation to the hydrological budget in GuatemalanTMCF is 1 mm d– 1 during the dry season and 0.5 mm d– 1
during the rainy season. The impact of the seasonality of this
water input on vegetation water use has yet to be demonstrateddirectly in TMCFs. Plants from redwood forests, a non-montanefog-affected ecosystem, use significantly more fog water duringthe dryseason, when fog incidence is higher (Dawson, 1998). It islikely that fog inputs have the greatest impacts hydrologically inlow rainfall, seasonally dry but frequently foggy and highlyexposed forests (Bruijnzeel et al., 2011).
Foliar water uptake
Recent studies have suggested that direct foliar water uptake isan ecophysiologically important input in TMCFs (Eller et al.,2013; Goldsmith et al., 2013). Unlike CWI, FWU is a waterflux within the plant, driven by water potential gradientsbetween sources and sinks along the soil–plant–atmospherecontinuum (SPAC) and the hydraulic conductivity betweenSPAC compartments (Fig. 2). Simonin et al. (2009) suggestedthat FWU can be described using a simple equation based onDarcy’s law:
FWU = kAtm−LDcAtm−L
where kAtm2L is the efficiency of leaf water uptake, which is ba-sically the leaf surface total conductivity to water entry, andDcAtm2L is the water potential (cH2O
) gradient between theinside and the outside of the leaf. During fog events, the atmos-pheric boundary layer surrounding leaves is saturated with mois-ture and the cH2O
outside the leaf should be close to zero. If leafcH2O
is negative, FWU should be higher than 0 during most leaf-wetting events provided that the leaf surface is hydrophilicenough to allow water film formation.
With constant kAtm2L, we should expect higher FWU rates inleaves experiencing water deficits, which should be morecommon during periods of low soil water availability (Fig. 2B, C).
–3
Safety margin = 0·01 MPa
Drought alleviation after fog = 0·4 MPa
–2
x (MPa)
min f
50
88
–1 0
0
20
40
60P
LC (
%)
80
100
–4
FI G. 3. Embolism vulnerability curve showing loss of hydraulic conductivity (PLC, %) as a function of xylem water potential (Cx, MPa) for branches of Drimysbrasiliensis (Winteraceae), a dominant species in Brazilian tropical montane cloud forests. C50 (–1.55 MPa) and C88 are the xylem water potential inducing 50and 88 % embolism, respectively. Cmin (–1.54 MPa) is the minimum xylem water potential measured in the field during 24 months. Cf is the increase in Cmin dueto fog occurrence. The difference between C50 and Cmin (vertical red bar) represents the ‘safety margin’ that the plant operates in the driest conditions, which is0.01 MPa. The blue arrow represents the increase in leaf water potential and hydraulic safety margin after fog exposure and foliar water uptake (FWU) (from Elleret al., 2013). The curve was fitted using an exponential sigmoidal equation: PLC ¼ 100/{1 – exp[a(Cx – C50)]}, where a is the slope of the curve. The R2 value
for the fit (–0.72) was obtained with linear regression of the transformed data (Pammenter and Van der Willigen, 1998).
Oliveira et al. — Hydroclimatic conditions and ecophysiology of cloud forests914
at BIB
LIO
TE
CA
CE
NT
RA
L U
NIV
EST
AD
UA
L C
AM
PINA
S on April 24, 2014
http://aob.oxfordjournals.org/D
ownloaded from

Supporting this prediction, Breshears et al. (2008) shows thatthe effect of FWU on leaf water potential is greater when theplant is subjected to water stress. Also, sap flow reversals ofhigher magnitude have been observed during the dry season inD. brasiliensis at a Brazilian TMCF (C. B. Eller et al., unpubl.data). However, Burgess and Dawson (2004) observed that well-watered leaves of Sequoia sempervirens absorbed more fog waterthan water-stressed leaves, implying that kAtm2L is moredynamic in some species than others. Therefore, FWU couldbe more controlled by DcAtm2L in some species, while inothers the kAtm2L should play a larger role.
The kAtm2L should be largely determined by leaf cuticle perme-ability towaterand the occurrence of structures that facilitatewateruptake. Despite their role in restricting molecular diffusion andthus water loss, the cuticles of leaves are known to be permeableto various molecules (Schonherr and Riederer, 1989; Schreiberand Riederer, 1996; Niederl et al., 1998). Water might diffusethrough a lipophilic pathway in the cuticle, with lipophilic cutinand wax domains forming its transport path (Schreiber, 2005),or aqueous pores, which are formed by the hydration of dipolesand ionic functional groups (Schonherr, 2006). It is important tonote that cuticle permeability to water might vary by severalorders of magnitude between species (Kerstiens, 1996), andmight be quite sensitive to changes in environmental conditions,increasing under high temperature (Schreiber, 2001) and high at-mospheric humidity (Schreiber et al., 2001; Eller et al., 2013).
The occurrence of structures on leaf epidermis that facilitatewater uptake can increase kAtm2L even further. Trichomes(Schreiber et al., 2001; Schonherr, 2006), hydathodes (Martinand von Willert, 2000), guard cells (Schlegel et al., 2005) and sto-matal plugs (Westhoff et al., 2009) are examples of epidermalstructures that might be preferential paths to FWU in somespecies because of differential properties of the cuticle on thesestructures. For example, Schonherr (2006) shows that aqueouspores are more likely to occur at the base of trichomes(Schonherr, 2006). Spatial heterogeneity on the wax content ofthe cuticle might also strongly affect water permeability(Schonherr and Lendzian, 1981; Becker et al., 1986).
There is also substantial empirical evidence that water mightenter into leaves through stomatal apertures (Eichert et al.,2008; Burkhardt et al., 2012). Until recently, direct water entrythrough stomata was considered physically impossible becauseof high water surface tension and the morphology of stomata(Schonherrand Bukovac, 1972), but the recent hypothesis of ‘hy-draulic activation of stomata’ by Burkhardt (2010) provides apossible explanation for this process. Burkhardt (2010) sug-gested that the deposition of hygroscopic particles around theguard cells and sub-stomatal cavity might break water surfacetension and allow the formation of thin water films along thestomata, establishing a hydraulic connection between theoutside surfaces of the leaf and the apoplast.
Considering the multiple water entry pathways on the leaf, it isnot surprising that the occurrence of FWU in plants is a verywidespread phenomenon, confirmed in .70 species (.85 %of all the studied species; Goldsmith et al. 2013). To our knowl-edge, all the studies investigating FWU in TMCFs found that thismechanism was present at least to some extent in the studied trees(Lima, 2010; Cassana and Dillenburg, 2012; Eller et al., 2013;Goldsmith et al., 2013). The prevalence of this mechanism inTMCFs enhances vegetation survival during seasonal droughts
(Eller et al., 2013), and might affect biotic interactions, foliartraits associated with fog interception efficiency (Martorell andEzcurra, 2007) and perhaps even hydraulic niche differentiation(Silvertown et al., 1999) and community assembly patterns. Weshould also note that this process adds a potentially importantbiotic component to TMCF water fluxes that has been ignoredin hydrometeorological models until now.
Ecological consequences of cloud immersion
Cloud immersion generally has a positive effect on leaf, plantand forest water balance (Bruijnzeel et al., 2011; Eller et al.,2013; Goldsmith et al., 2013). Even if a certain tree species isnot capable of significant FWU, the suppressive effect on planttranspiration (Limm et al., 2009; Gotsch et al., 2014) and add-itional soil water input by fog drip can provide an importantwater subsidy for plants (Dawson, 1998; Liu et al., 2004).However, there are important ecological differences in thewater subsidy provided by FWU and fog drip. First, part of thewater of some leaf-wetting events might not even reach the soilbecause of the canopy storage and subsequent evaporation.Thus, species capable of FWU could benefit from the waterinput even of a weak leaf-wetting event. Also, the water absorbedby FWU might be redistributed inside the plant and even reachthe plant rhizosphere (Eller et al., 2013). The increase in rootmoisture associated with this transport should cause ecologicalconsequences to plants similar to those when water is redistribu-ted between roots in different soil layers (hydraulic redistribu-tion; Burgess et al., 1998; Oliveira et al., 2005a). Theseconsequences include a decrease in branch and root embolism,an increase on root life span (Domec et al., 2004, 2006;Bauerle et al., 2008), benefits to mycorrhizal development(Querejeta et al., 2007) and even increased nutrient availabilityin the soil close to the roots (Dawson, 1997; Pang et al., 2013).The effects of this water transport on biotic interactions andbelow-ground resource competition could also be significant(Dawson, 1993; Zou et al., 2005; Prieto et al., 2011). However,there are also a number of possibly negative ecological conse-quences associated with FWU. If FWU occurs directly throughthe cuticle, as seems to be the case in some species (Schonherr,1976, 2006; Kerstiens, 2006; Eller et al., 2013), this additionalwater permeability could work both ways, leading to higher cu-ticular conductance, which can be detrimental to plant droughtresistance, mostly because of the reduced capacity to controlleaf water loss during droughts (Burkhardt and Riederer,2003). Another possible cost associated with FWU comes fromthe potential negative relationship between FWU and leafwater repellency (LWR) (Fig. 4; Grammatikopoulos andManetas, 1994; Rosado et al., 2010; Rosado and Holder,2013). FWU is thought to be favoured in plants with lowerLWR (i.e. plants that stay wet for longer). Comparative studiesshow that LWR in cloud forests is lower than in lowland forests(Holder, 2007a), which indirectly reinforces the proposedLWR–FWU relationship (Fig. 4), now that we have evidencethat FWU occurs in TMCFs (Lima, 2010; Eller et al., 2013;Goldsmith et al., 2013). Low LWR might have severalnegative consequences for the leaf, such as facilitation of patho-gen infection (Reynolds et al., 1989; Evans et al., 1992), foliarnutrient leaching (Cape, 1996), epiphyll growth (Holder,2007b), decrease in leaf self-cleaning properties (Barthlott and
Oliveira et al. — Hydroclimatic conditions and ecophysiology of cloud forests 915
at BIB
LIO
TE
CA
CE
NT
RA
L U
NIV
EST
AD
UA
L C
AM
PINA
S on April 24, 2014
http://aob.oxfordjournals.org/D
ownloaded from
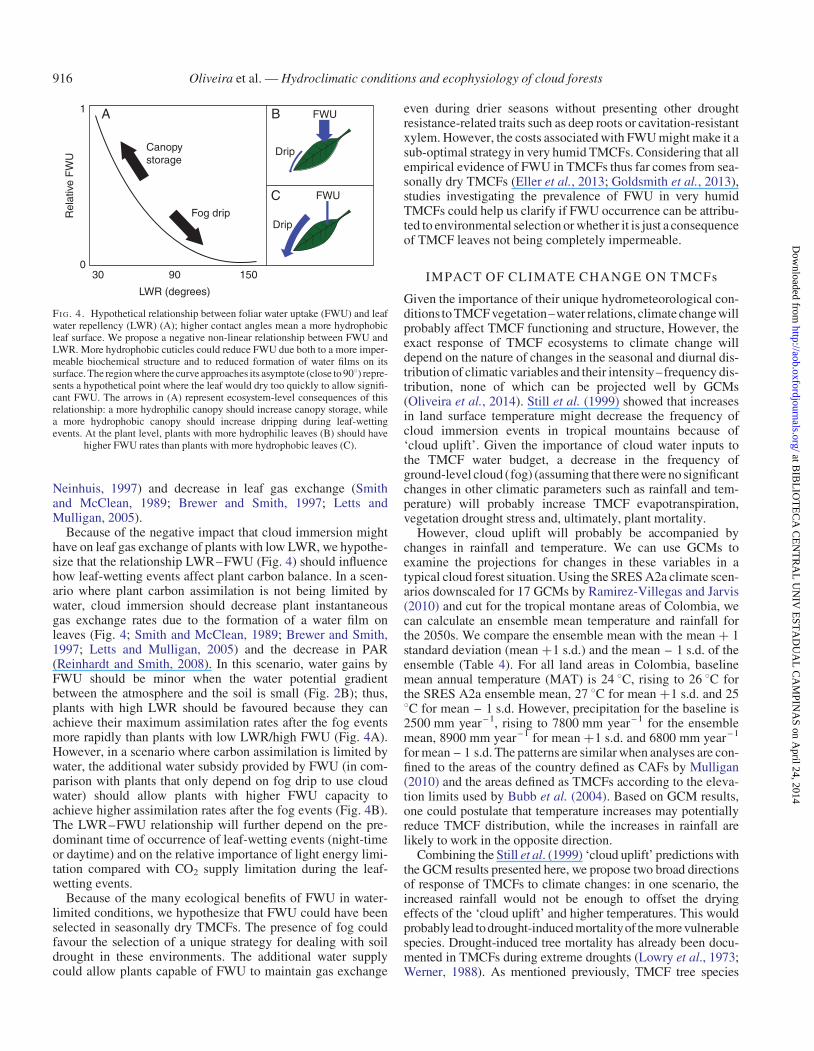
Neinhuis, 1997) and decrease in leaf gas exchange (Smithand McClean, 1989; Brewer and Smith, 1997; Letts andMulligan, 2005).
Because of the negative impact that cloud immersion mighthave on leaf gas exchange of plants with low LWR, we hypothe-size that the relationship LWR–FWU (Fig. 4) should influencehow leaf-wetting events affect plant carbon balance. In a scen-ario where plant carbon assimilation is not being limited bywater, cloud immersion should decrease plant instantaneousgas exchange rates due to the formation of a water film onleaves (Fig. 4; Smith and McClean, 1989; Brewer and Smith,1997; Letts and Mulligan, 2005) and the decrease in PAR(Reinhardt and Smith, 2008). In this scenario, water gains byFWU should be minor when the water potential gradientbetween the atmosphere and the soil is small (Fig. 2B); thus,plants with high LWR should be favoured because they canachieve their maximum assimilation rates after the fog eventsmore rapidly than plants with low LWR/high FWU (Fig. 4A).However, in a scenario where carbon assimilation is limited bywater, the additional water subsidy provided by FWU (in com-parison with plants that only depend on fog drip to use cloudwater) should allow plants with higher FWU capacity toachieve higher assimilation rates after the fog events (Fig. 4B).The LWR–FWU relationship will further depend on the pre-dominant time of occurrence of leaf-wetting events (night-timeor daytime) and on the relative importance of light energy limi-tation compared with CO2 supply limitation during the leaf-wetting events.
Because of the many ecological benefits of FWU in water-limited conditions, we hypothesize that FWU could have beenselected in seasonally dry TMCFs. The presence of fog couldfavour the selection of a unique strategy for dealing with soildrought in these environments. The additional water supplycould allow plants capable of FWU to maintain gas exchange
even during drier seasons without presenting other droughtresistance-related traits such as deep roots or cavitation-resistantxylem. However, the costs associated with FWU might make it asub-optimal strategy in very humid TMCFs. Considering that allempirical evidence of FWU in TMCFs thus far comes from sea-sonally dry TMCFs (Eller et al., 2013; Goldsmith et al., 2013),studies investigating the prevalence of FWU in very humidTMCFs could help us clarify if FWU occurrence can be attribu-ted to environmental selection or whether it is just a consequenceof TMCF leaves not being completely impermeable.
IMPACT OF CLIMATE CHANGE ON TMCFs
Given the importance of their unique hydrometeorological con-ditions to TMCF vegetation–water relations, climate changewillprobably affect TMCF functioning and structure, However, theexact response of TMCF ecosystems to climate change willdepend on the nature of changes in the seasonal and diurnal dis-tribution of climatic variables and their intensity–frequency dis-tribution, none of which can be projected well by GCMs(Oliveira et al., 2014). Still et al. (1999) showed that increasesin land surface temperature might decrease the frequency ofcloud immersion events in tropical mountains because of‘cloud uplift’. Given the importance of cloud water inputs tothe TMCF water budget, a decrease in the frequency ofground-level cloud (fog) (assuming that therewere no significantchanges in other climatic parameters such as rainfall and tem-perature) will probably increase TMCF evapotranspiration,vegetation drought stress and, ultimately, plant mortality.
However, cloud uplift will probably be accompanied bychanges in rainfall and temperature. We can use GCMs toexamine the projections for changes in these variables in atypical cloud forest situation. Using the SRES A2a climate scen-arios downscaled for 17 GCMs by Ramirez-Villegas and Jarvis(2010) and cut for the tropical montane areas of Colombia, wecan calculate an ensemble mean temperature and rainfall forthe 2050s. We compare the ensemble mean with the mean + 1standard deviation (mean +1 s.d.) and the mean – 1 s.d. of theensemble (Table 4). For all land areas in Colombia, baselinemean annual temperature (MAT) is 24 8C, rising to 26 8C forthe SRES A2a ensemble mean, 27 8C for mean +1 s.d. and 258C for mean – 1 s.d. However, precipitation for the baseline is2500 mm year– 1, rising to 7800 mm year– 1 for the ensemblemean, 8900 mm year– 1 for mean +1 s.d. and 6800 mm year– 1
for mean – 1 s.d. The patterns are similar when analyses are con-fined to the areas of the country defined as CAFs by Mulligan(2010) and the areas defined as TMCFs according to the eleva-tion limits used by Bubb et al. (2004). Based on GCM results,one could postulate that temperature increases may potentiallyreduce TMCF distribution, while the increases in rainfall arelikely to work in the opposite direction.
Combining the Still et al. (1999) ‘cloud uplift’ predictions withthe GCM results presented here, we propose two broad directionsof response of TMCFs to climate changes: in one scenario, theincreased rainfall would not be enough to offset the dryingeffects of the ‘cloud uplift’ and higher temperatures. This wouldprobably lead todrought-induced mortalityof the more vulnerablespecies. Drought-induced tree mortality has already been docu-mented in TMCFs during extreme droughts (Lowry et al., 1973;Werner, 1988). As mentioned previously, TMCF tree species
Canopystorage
Fog drip
0
1 A B
C
30 90
LWR (degrees)
150
Drip
Drip
FWU
FWU
Rel
ativ
e F
WU
FI G. 4. Hypothetical relationship between foliar water uptake (FWU) and leafwater repellency (LWR) (A); higher contact angles mean a more hydrophobicleaf surface. We propose a negative non-linear relationship between FWU andLWR. More hydrophobic cuticles could reduce FWU due both to a more imper-meable biochemical structure and to reduced formation of water films on itssurface. The region where the curve approaches its asymptote (close to 908) repre-sents a hypothetical point where the leaf would dry too quickly to allow signifi-cant FWU. The arrows in (A) represent ecosystem-level consequences of thisrelationship: a more hydrophilic canopy should increase canopy storage, whilea more hydrophobic canopy should increase dripping during leaf-wettingevents. At the plant level, plants with more hydrophilic leaves (B) should have
higher FWU rates than plants with more hydrophobic leaves (C).
Oliveira et al. — Hydroclimatic conditions and ecophysiology of cloud forests916
at BIB
LIO
TE
CA
CE
NT
RA
L U
NIV
EST
AD
UA
L C
AM
PINA
S on April 24, 2014
http://aob.oxfordjournals.org/D
ownloaded from

operate close to their limit of hydraulic failure (Fig. 3). This meansthat the dry-season changes in soil water availability and atmos-pheric demand expected in this drier scenario might seriouslydamage the species hydraulic system and increase the chance oflarge-scale vegetation mortality. Plants with high FWU capacitycould be particularly vulnerable to the decrease in leaf-wettingevents, not only because of the key role of FWU in the mainten-ance of ecophysiological performance during drought (Simoninet al., 2009; Eller et al., 2013), but also because of the role thatFWU might play in hydraulic failure avoidance. The increase inleaf water potential associated with FWU (average of 0.4 MPain D. brasiliensis; Eller et al., 2013) might decrease xylemtension (Brodersen and McElrone, 2013) and increase the planthydraulic safety margin (Fig. 3). Foliar water uptake can also bean important mechanism responsible for successful embolismrepair in leaves and stems of TMCF plants (Limm et al., 2009;Simonin et al., 2009; Eller et al., 2013). Cuticular absorptioncould reduce the tension on the xylem enough to allow forrefilling (Burgess and Dawson, 2007; Limm et al., 2009,Oliveira et al., 2005b).
In another possible scenario, the increased rainfall completelyoffsets the reduced cloud water contribution to TMCF waterbudget and increased atmospheric demand caused by higher tem-peratures. This would expose TMCF vegetation to a warmer, lessfoggy but rainier climate, similar to the climatic envelope oflowland tropical forests. This kind of change could favour the in-vasion of TMCFs by lower elevation species (Loope andGiambelluca, 1998; Pauchard et al., 2009). Lowland species arelikely to be better competitors in this climatic scenario, due totheir higher leaf area index (Bruijnzeel et al., 2011) and higheroptimum temperature for photosynthesis (Allen and Ort, 2001),which leads to faster growing rates and potentially higher seedoutput. The invasion of TMCFs by lowland animals observedby Pounds et al. (1999) and associated with climate changecould also increase dispersion rates of lowland tree speciesupwards into the mountains.
In both of the hypothetical scenarios, TMCF functioning andstructure would be altered. In the drier scenario, drought shouldinduce widespread mortality of less drought-resistant species,while in the warmer scenario, TMCF species could be competi-tively displaced by lower elevation species. More knowledgeabout TMCF vegetation and ecosystem functioning is necessaryin order to understand more precisely towhat extent each particu-lar scenario could affect TMCFs. It is possible that differentTMCFs would be more or less vulnerable to a particular scenario
depending on their current climate characteristics. For example,current seasonally dry TMCFs could be less vulnerable to a drierscenario, because one could assume that species from theseTMCFs are already more drought resistant.
CONCLUDING REMARKS AND PERSPECTIVES
In this review, we propose that TMCF distribution dependsstrongly on the relationship between particular plant ecophysio-logical traits, such as FWU (increasing the hydraulic safetymargin), and unique hydrometeorological conditions ofTMCFs. Changes in these conditions, especially related tocloud immersion events, could drastically change the costs andbenefits associated with FWU and, consequently, TMCF struc-ture and functioning. More information about the mechanismsbehind drought-induced mortality in TMCF plants is needed toclarify how drought events might affect population dynamicsand community structure of TMCFs under drier climates.Despite knowing that leaf-wetting events and FWU might be im-portant to some TMCF species during droughts (Eller et al.,2013), we do not know what proportion of TMCF speciesdepend on FWU for survival during drought. We also do notknow if a small hydraulic safety margin (Fig. 3) is a widespreadtrait in different TMCF species.
Potential effects of increased precipitation – which will varyhighly between cloud forests in different topographic and contin-ental settings – could either compensate for the reduction of leaf-wetting events or combine with warming to create a microclimaticenvelope that could facilitate the invasion of TCMFs by lowlandspecies. Competitive interactions between lowland forestspecies under different environmental conditions are alsoneeded to illustrate TMCF vulnerability to lowland species inva-sion and the consequences for TMCF community structure andecosystem-level processes.
We believe that the inclusion of non-standard climate vari-ables (fog frequency and terrain exposure) and species function-al attributes is essential for an accurate niche-based modelling ofspecies distribution and also for more accurate predictions ofecophysiological models, especially under climate change.The FWU phenomenon in TMCFs, for example, adds an import-ant component that needs to be taken into consideration in TMCFecophysiological models, since it could increase the predictedcontribution of fog to the ecophysiology of these ecosystemsand also affect canopy water storage and re-evaporation to the at-mosphere. The water subsidy provided by fog could also allow
TABLE 4. Climate change uncertainty for TMCFs, CAFS and all land in Colombia based on a 17 GCM ensemble for a SRES A2ascenario
TMCFs* CAFs All land
MAT (8C)Precipitation(mm year– 1) MAT (8C)
Precipitation(mm year– 1) MAT (8C)
Precipitation(mm year– 1)
Baseline (1950–2000) 13 1600 18 2200 24 2500Mean of 17 GCMs A2a 2050s 16 5100 21 6700 26 7800Mean of 17 GCMs +1 s.d. A2a 2050s 17 6000 22 7500 27 8900Mean of 17 GCMs – 1 s.d. A2a 2050s 15 4300 20 5800 25 6800
*Bubb et al. (2004); 2000–3500 m a.s.l.MAT, mean annual temperature.
Oliveira et al. — Hydroclimatic conditions and ecophysiology of cloud forests 917
at BIB
LIO
TE
CA
CE
NT
RA
L U
NIV
EST
AD
UA
L C
AM
PINA
S on April 24, 2014
http://aob.oxfordjournals.org/D
ownloaded from

species capable of FWU to occur in places where they could nototherwise occur, if they depended only on soil water.
ACKNOWLEDGEMENTS
This review was based, in part, ona plenary lecture presented at theComBio2013, Perth, Australia, sponsored by the Annals ofBotany. The authors would like to express their thanks to theGraduate Program in Ecology from the University of Campinas(UNICAMP), and to Hans Lambers and Tim Colmer(University of Western Autralia) for the invitation to present alecture at ComBio2013. This work was supported by the SaoPaulo Research Foundation (FAPESP) (grant no. 10/17204-0),FAPESP/Microsoft Research (grant no. 11/52072-0) awarded toR.S.O., and the Higher Education Co-ordination Agency(CAPES) (scholarship to C.B.E. and P.L.B.). The cloud forestmapping was supported by the UK Department for InternationalDevelopment Forestry Research Programme (ZF0216).
LITERATURE CITED
Aldrich M, Billington C, Edwards M, Laidlaw R. 1997. A global directory oftropical montane cloud forests. Cambridge, UK: UNEP–WCMC.
Allen DJ, Ort DR. 2001. Impacts of chilling temperatures on photosynthesis inwarm-climate plants. Trends in Plant Science 6: 36–42.
Barthlott W, Neinhuis C. 1997. Purity of sacred lotus, or escape from contam-ination in biological surfaces. Planta 202: 1–8.
Bauerle TL, Richards JH, Smart DR, Eissenstat DM. 2008. Importance of in-ternal hydraulic redistribution for prolonging the lifespan of roots in dry soil.Plant, Cell and Environment 31: 177–186.
Becker M, Kerstiens G, Schonherr J. 1986. Water permeability of plant cuti-cles: permeance, diffusion and partition coefficients. Trees 1: 54–60.
Bertoncello R, Yamamoto K, Meireles LD, Shepherd GJ. 2011. A phytogeo-graphic analysis of cloud forests and other forest subtypes amidst theAtlantic forests in south and southeast Brazil. Biodiversity and Conservation20: 3413–3433.
Breshears DD, McDowell NG, Goddard KL, et al. 2008. Foliar absorption ofintercepted rainfall improves woody plant water status most duringdrought. Ecology 89: 41–47.
Brewer CA, Smith WK. 1997. Patterns of leaf surface wetness for montane andsubalpine plants. Plant, Cell and Environment 20: 1–11.
Brodersen CR, McElrone AJ. 2013. Maintenance of xylem network transportcapacity: a review of embolism repair in vascular plants. Frontiers inPlant Science 4: 1–11.
Brown MB, de la Roca I, Vallejo A, et al. 1996. Avaluation analysis of the role ofcloud forests in watershed protection. Sierra de las Minas BiosphereReserve, Guatemala and Cusuco N.P., Honduras. Philadelphia, PA:RARE Center for Tropical Conservation.
Bruijnzeel LA. 2001. Hydrology of tropical montane cloud forest: a reassess-ment. Land Use and Water Resources Research 1: 1–18.
Bruijnzeel LA, Hamilton LS. 2000. Decision time for cloud forests. IHP HumidTropics Programme Series no. 13. Paris: IHP-UNESCO, Amsterdam:IUCN-NL, and Gland: WWF International.
Bruijnzeel LA, Veneklas E. 1998. Climatic conditions and tropical montaneforest productivity: the fog has not lifted yet. Ecology 79: 3–9.
Bruijnzeel LA, Scatena FN, Hamilton LS, eds. 2010a. Tropical montane cloudforests. Science for conservation and management. Cambridge: CambridgeUniversity Press.
Bruijnzeel LA, Kappelle M, Mulligan M, Scatena FN. 2010b. Tropicalmontane cloud forests: state of knowledge and sustainability perspectivesin a changing world. In: Bruijnzeel LA, Scatena FN, Hamilton LS, eds.Tropical montane cloud forests. Science for conservation and management.Cambridge: Cambridge University Press, 691–740.
Bruijnzeel LA, Mulligan M, Scatena FN. 2011. Hydrometeorology of tropicalmontane cloud forests: emerging patterns. Hydrological Processes 25:465–498.
Bubb P, May I, Miles L, Sayer J. 2004. Cloud forest agenda. Cambridge:UNEP–WCMC.
Burgess SSO, Dawson TE. 2004. The contribution of fog to thewater relations ofSequoia sempervirens (D. Don): foliar uptake and prevention of dehydra-tion. Plant, Cell and Environment 27: 1023–1034.
Burgess SSO, Dawson TE. 2007. Predicting the limits to tree height using stat-istical regressions of leaf traits. New Phytologist 174: 626–636.
Burgess SSO, Adams MA, Turner NC, Ong CK. 1998. The redistribution ofsoil water by tree root systems. Oecologia 115: 306–311.
Burkhardt M. 2010. Hygroscopic particles on leaves: nutrients or desiccants?Ecological Monographs 80: 369–399.
Burkhardt M, Riederer M. 2003. Ecophysiological relevance of cuticular tran-spiration of deciduous and evergreen plants in relation to stomatal closureand leaf water potential. Journal of Experimental Botany 54: 1941–1949.
Burkhardt J, Basi S, Pariyar S, Hunsche M. 2012. Stomatal penetration byaqueous solutions – an update involving leaf surface particles. NewPhytologist 196: 774–787.
Cape JN. 1996. Surface wetness and pollutant deposition. In: Kerstiens G, ed.Plant cuticles: an integrated approach. Oxford: BIOS ScientificPublishers, 283–300.
Cavelier J. 1996. Tissue water relations in elfin cloud forest tree species ofSerrania de Macuira, Guajira, Colombia. Trees 4: 155–163.
Cassana FF, Dillenburg LR. 2012. The periodic wetting of leaves enhanceswater relations and growth of the long-lived conifer Araucaria angustifolia.Plant Biology 15: 75–83.
Choat B, Jansen S, Brodribb TJ, et al. 2012. Global convergence in the vulner-ability of forests to drought. Nature 491: 752–755.
Chu H-S, Chang S-C, Klemm O, et al. 2014. Does canopy wetness matter?Evapotranspiration from a subtropical montane cloud forest in Taiwan.Hydrological Processes 28: 1190–1214.
Dawson TE. 1993. Hydraulic lift and the water use by plants: implications forwater balance, perfomance and plant–plant interactions. Oecologia 95:565–574.
Dawson TE. 1997. Water loss from tree roots influences soil water and nutrientstatus and plant performance. In: Flore HE, Lynch JP, Eissenstat DM, eds.Radical biology: advances and perspectives on the function of plant roots.Rockville, MD: American Society of Plant Physiologists, 235–250.
Dawson TE. 1998. Fog in the California redwood forest: ecosystem inputs anduse by plants. Oecologia 117: 476–485.
Dawson TE, Burgess SS, Tu KP, et al. 2007. Nighttime transpiration in woodyplants from contrasting ecosystems. Tree Physiology 27: 561–575.
Domec JC, Warren JM, Meinzer FC. 2004. Native root xylem embolism andstomatal closure in stands of Douglas-fir and ponderosa pine: mitigationby hydraulic redistribution. Oecologia 141: 7–16.
Domec JC, Scholz FG, Bucci SJ, Meinzer FC, Goldstein G, Villalobos-VegaR. 2006. Diurnal and seasonal changes in root xylem embolism inNeotropical savanna woody species: impact on stomatal control of plantwater status. Plant, Cell and Environment 29: 26–35.
Eichert T, Kurtz A, Steinerb U, Goldbach H. 2008. Size exclusion limits andlateral heterogeneity of the stomatal foliar uptake pathway for aqueoussolutes and water suspended nanoparticles. Physiologia Plantarum 134:151–160.
Eller CB, Lima AL, Oliveira RS. 2013. Foliar uptake of fog water and transportbelowground alleviates drought effects in the cloud forest tree species,Drimys brasiliensis (Winteraceae). New Phytologist 199: 151–162.
Evans KJ, Nyquist WE, Latin RX. 1992. A model based on temperature and leafwetness duration for establishment of Alternaria leaf blight of muskmelon.Phytopathology 82: 890–895.
Feild TS, Holbrook NM. 2000. Xylem sap flow and stem hydraulics of the ves-selless angiosperm Drimys granadensis (Winteraceae) in a Costa Rican elfinforest. Plant, Cell and Environment 23: 1067–1077.
Giambelluca TW, Gerold G. 2011. Hydrology and biogeochemistry of tropicalmontane cloud forests. In: Levia DF, Carlyle-Moses D, Tanaka T, eds.Hydrology and biogeochemistry of forest ecosystems. New York: SpringerVerlag, 221–259.
Goldsmith GR, Matzke NJ, Dawson TE. 2013. The incidence and implicationsof clouds for cloud forest plant water relations. Ecology Letters 16:307–314.
Gotsch SG, Asbjornsen H, Holwerda F, Goldsmith GR, Weintraub AE,Dawson TE. 2014. Foggy days and dry nights determine crown-levelwater balance in a seasonal tropical montane cloud forest. Plant, Cell andEnvironment 37: 261–272.
Grammatikopoulos G, Manetas Y. 1994. Direct absorption of water by hairyleaves of Phlomis fruticosa and its contribution to drought avoidance.Canadian Journal of Botany 72: 1805–1811.
Oliveira et al. — Hydroclimatic conditions and ecophysiology of cloud forests918
at BIB
LIO
TE
CA
CE
NT
RA
L U
NIV
EST
AD
UA
L C
AM
PINA
S on April 24, 2014
http://aob.oxfordjournals.org/D
ownloaded from

Grubb PJ, Whitmore TC. 1966. A comparison of montane and lowland rainforest in Ecuador. II. The climate and its effects on the distributionand physi-ognomy of the forests. Journal of Ecology 54: 303–333.
Holder CD. 2004. Rainfall interception and fog precipitation in a tropicalmontane cloud forest of Guatemala. Forest Ecology and Management190: 373–384.
Holder CD. 2007a. Leaf water repellency of species in Guatemala and Colorado(USA) and its significance to forest hydrologystudies. Journal of Hydrology336: 147–154.
Holder CD. 2007b. Leaf water repellency as an adaptation to tropical montanecloud forest environments. Biotropica 39: 767–770.
Holwerda F, Bruijnzeel LA, Oord AL, et al. 2010. Fog interception in a PuertoRican elfin cloud forest: a wet-canopy water budget approach. In: BruijnzeelLA, Scatena FN, Hamilton LS, eds. Tropical montane cloud forests. Sciencefor conservation and management. Cambridge: Cambridge UniversityPress, 282–292.
Jane GT, Green TGA. 1985. Patterns of stomatal conductance in six evergreentree species from a New Zealand cloud forest. Botanical Gazette 146:413–420.
Jarvis A, Mulligan M. 2011. The climate of cloud forests. HydrologicalProcesses 25: 327–343.
Kerstiens G. 1996. Cuticular water permeability and its physiological signifi-cance. Journal of Experimental Botany 47: 1813–1832.
Kerstiens G. 2006. Water transport in plant cuticles: an update. Journal ofExperimental Botany 57: 2493–2499.
Kumagai T, Saitoh TM, Sato Y, et al. 2004. Transpiration, canopy conductanceand the decoupling coefficient of a lowland mixed dipterocarp forest inSarawak, Borneo: dry spell effects. Journal of Hydrology 287: 237–251.
Kumagai T, Saitoh TM, Sato Y, et al. 2005. Annual water balance and season-ality of evapotranspiration in a Bornean tropical rainforest. Agricultural andForest Meteorology 128: 81–92.
Letts MG, Mulligan M. 2005. The impact of light quality and leaf wetness onphotosynthesis in north-west Andean tropical montane cloud forest.Journal of Tropical Ecology 21: 549–557
Lima AL. 2010. The ecological role of fog and foliar water uptake in three woodyspecies from Southeastern Brazilian Cloud Forest. Masters thesis,University of Campinas (UNICAMP), Brazil.
Limm E, Simonin K, Bothman A, Dawson T. 2009. Foliar water uptake: acommon water acquisition strategy for plants of the redwood forest.Oecologia 161: 449–459.
Liu W, Meng F, Zhang Y, Liu Y, Li H. 2004. Water input from fog drip in thetropical seasonal rain forest of Xishuangbanna, South-West China.Journal of Tropical Ecology 20: 517–524.
Loope LL, Giambelluca TW. 1998. Vulnerability of island tropical montanecloud forests to climate change, with special reference to east Maui,Hawaii. Climatic Change 39: 503–517.
Lowry JB, Lee DW, Stone BC. 1973. Effects of drought on Mount Kinabalu.Malayan Nature Journal 26: 178–179.
Luna-Veja I, Morrone JJ, Ayala OAA, Organista DE. 2001. Biogeographicalaffinities among Neotropical cloud forests. Plant Systematics and Evolution228: 229–239.
Martin CE, von Willert DJ. 2000. Leaf epidermal hydathodes and the ecophy-siological consequences of foliar water uptake in species of Crassula fromthe Namib Desert in southern Africa. Plant Biology 2: 229–242.
Martorell C, Ezcurra E. 2007. The narrow-leaf syndrome: a functional and evo-lutionary approach to the form of fog-harvesting rosette plants. Oecologia151: 561–73.
McDowell N, Pockman WT, Allen CD, et al. 2008. Mechanisms of plant sur-vival and mortality during drought: why do some plants survive whileothers succumb to drought? New Phytologist 178: 719–739.
Meireles LD. 2003. Florıstica das fisionomias vegetacionais e estrutura da flor-esta alto-montana de Monte Verde, Serra da Mantiqueira, MG. Mastersthesis, University of Campinas (UNICAMP), Brazil.
Mercado LM, Bellouin N, Sitch S, et al. 2009. Impact of changes in diffuse ra-diation on the global land carbon sink. Nature 458: 1014–1017.
Motzer T. 2005. Micrometeorological aspects of a tropical mountain forest.Agricultural and Forest Meteorology 135: 230–240.
Mulligan M. 2010. Modeling the tropics-wide extent and distribution of cloudforest and cloud forest loss, with implications for conservation priority.In: Bruijnzeel LA, Scatena FN, Hamilton LS, eds. Tropical montanecloud forests. Science for conservation and management. Cambridge:Cambridge University Press, 14–39.
Mulligan M, Fisher M, Sharma B, et al. 2011. The nature and impact of climatechange in the Challenge Program on Water and Food (CPWF) basins. WaterInternational 36: 96–124.
Niederl S, Kirsch T, Riederer M, Schreiber L. 1998. Co-permeability of3H-labelled water and 14C-labelled organic acids across isolated plant cuti-cles: investigating cuticular paths of diffusion and predicting cuticular tran-spiration. Plant Physiology 116: 117–123.
Oliveira RS, Dawson TE, Burgess SSO, Nepstad DC. 2005a. Hydraulic redis-tribution in three Amazonian trees. Oecologia 145: 354–363.
Oliveira RS, Dawson TE, Burgess SSO. 2005b. Evidence for direct water ab-sorption by the shoot of the desiccation-tolerant plant Vellozia flavicans inthe savannas of central Brazil. Journal of Tropical Ecology 21: 585–588.
Oliveira RS, ChristoffersenBO, Barros FV, et al. 2014. Changing precipitationregimes and the water and carbon economies of trees. Theoretical andExperimental Plant Physiology. doi:10.1007/s40626-014-0007-1.
Pang J, Wang Y, Lambers H, Tibbet M, Siddique KHM, Ryan MH. 2013.Commensalism in an agroecosystem: hydraulic redistribution by deep-rooted legumes improves survival of a droughted shallow-rooted legumecompanion. Physiologia Plantarum 149: 79–90.
Pammenter NW, Van der Willigen C. 1998. A mathematical and statistical ana-lysis of the curves illustrating vulnerability of xylem to cavitation. TreePhysiology 18: 589–593.
Pauchard A, Kueffer C, Dietz H, et al. 2009. Ain’t no mountain high enough:plant invasions reaching new elevations. Frontiers in Ecology and theEnvironment 7: 479–486.
Pounds JA, Fogden MP, Campbell JH. 1999. Biological response to climatechange on a tropical mountain. Nature 398: 611–615.
Prieto I, Padilla FM, Armas C, Pugnaire FI. 2011. The role of hydraulic lift onseedlingestablishmentunderanurseplant species inasemi-aridenvironment.Perspectives in Plant Ecology, Evolution and Systematics 13: 181–187.
Querejeta JI, Egerton-Warburton LM, Allen MF. 2007. Hydraulic lift maybuffer rhizosphere hyphae against the negative effects of severe soildrying in a California oak savanna. Soil Biology and Biochemistry 39:409–417.
Ramirez-Villegas J, Jarvis A. 2010. Downscaling global circulation modeloutputs: the delta method decision and policy analysis. Working PaperNo. 1. Colombia: Decision and Policy Analysis, CIAT.
Reinhardt K, Smith WK. 2008. Impacts of cloud immersion on microclimate,photosynthesis and water relations of Abies fraseri (Pursh.) Poiret in a tem-perate mountain cloud forest. Oecologia 158: 229–238.
Reynolds KM, Madden LV, Richard DL, Ellis MA. 1989. Splash dispersal ofPhytophthora cactorum from infected strawberry fruit by simulated canopydrip. Phytopathology 79: 425–432.
Rosado BHP, Holder CD. 2013. The significance of leaf water repellency in eco-hydrological research: a review. Ecohydrology 6: 150–161.
Rosado BHP, Oliveira RS, Aidar MPM. 2010. Is leaf water repellency related tovapor pressure deficit and crown exposure in tropical forests? ActaOecologica 36: 645–649.
Rosado BHP, Oliveira RS, Joly CA, Aidar MPM, Burgess SSO. 2012.Diversity in nighttime transpiration behavior of woody species of theAtlantic Rain Forest, Brazil. Agricultural Forest Meteorology 158–159:13–20.
Santiago LS, Jones T, Goldstein G. 2010. Physiological variation in HawaiianMetrosideros polymorpha across a range of habitats: from dry forests tocloud forests. In: Bruijnzeel LA, Scatena FN, Hamilton JG, Juvik JO,Bubb P, eds. Mountains in the mist: science for conserving and managingtropical montane cloud forests. Honolulu: University of Hawaii Press,456–464.
Santiago LS, Goldstein G, Meinzer FC, Fownes J, Mueller-Dombois D. 2000.Transpiration and forest structure in relation to soil waterlogging in aHawaiian montane cloud forest. Tree Physiology 20: 673–681.
ScatenaFN, BruijnzeelLA, Bubb P, Das S.2010. Setting the stage. In: BruijnzeelLA, Scatena FN, Hamilton LS, eds. Tropical montane cloud forests. Sciencefor conservation and management. Cambridge: Cambridge University Press,3–13.
Schlegel TK, Schonherr J, Schreiber L. 2005. Size selectivity of aqueous poresin stomatous cuticles of Vicia faba leaves. Planta 221: 648–655.
Scholl M, Eugster W, Burkard R. 2010. Understanding the role of fog in foresthydrology: stable isotopes as tools for determining input and partitioning ofcloud water in montane forests. Hydrological Processes 25: 353–366.
Scholz FG, Bucci SJ, Goldstein G, Meinzer FC, Franco AC, Miralles-Wilhelm F. 2007. Removal of nutrient limitations by long-term fertilization
Oliveira et al. — Hydroclimatic conditions and ecophysiology of cloud forests 919
at BIB
LIO
TE
CA
CE
NT
RA
L U
NIV
EST
AD
UA
L C
AM
PINA
S on April 24, 2014
http://aob.oxfordjournals.org/D
ownloaded from

decreases nocturnal water loss in savanna trees. Tree Physiology 27:551–559.
Schonherr J. 1976. Water permeability of isolated cuticular membranes: theeffect of pH and cations on diffusion, hydrodynamic permeability andsize of polar pores. Planta 128: 113–126.
Schonherr J. 2006. Characterization of aqueous pores in plant cuticles and per-meation of ionic solutes. Journal of Experimental Botany 57: 2471–2491.
Schonherr J, Bukovac MJ. 1972. Penetration of stomata by liquids: dependenceon surface tension, wettability, and stomatal morphology. Plant Physiology49: 813–819.
Schonherr J, LendzianK. 1981.A simpleand inexpensive methodof measuringwater permeability of isolated plant cuticular membranes. Zeitschrift furPflanzenphysiologie 102: 321–327.
Schonherr J, Riederer M. 1989. Foliar penetration and accumulation of organicchemicals in plant cuticles. Reviews of Enviromental Contamination andToxicology 108: 1–70.
Schreiber L. 2001. Effect of temperature on cuticular transpiration of isolated cu-ticular membranes and intact leaf disks. Journal of Experimental Botany 52:1893–1900.
Schreiber L. 2005. Polar paths of diffusion across plant cuticles: new evidencefor an old hypothesis. Annals of Botany 95: 1069–1073.
Schreiber L, Riederer M. 1996. Ecophysiology of cuticular transpiration: com-parative investigation of cuticular water permeability of plant species fromdifferent habitats. Oecologia 107: 426–432
Schreiber L, Skrabs M, Hartmann KD, Diamantopoulos P, Simanova E,Santrucek J. 2001. Effect of humidity on cuticular water permeability ofisolated cuticular membranes and leaf disks. Planta 214: 274–282.
Sidle RC, Ziegler AD, Negishi JN, Nik AR, Siew R, Turkelboom F. 2006.Erosion processes in steep terrain: truths, myths, and uncertainties relatedto forest management in Southeast Asia. Forest Ecology and Management224: 199–225.
Silvertown J, Dodd ME, Gowing D, Mountford O. 1999. Hydrologically-defined niches reveal a basis for species-richness in plant communities.Nature 400: 61–63.
Simonin KA, Santiago LS, Dawson TE. 2009. Fog interception by Sequoiasempervirens (D.Don) crowns decouples physiology from soil waterdeficit. Plant, Cell and Environment 32: 882–892.
Smith WK,McClean TM. 1989.Adaptive relationship between leaf water repel-lency, stomatal distribution, and gas exchange. American Journal of Botany76: 465–469.
Snyder KA, James JJ, Richards JH, Donovan LA. 2008. Does hydraulic lift ornighttime transpiration facilitate nitrogen acquisition? Plant and Soil 306:159–166.
Still CJ, Foster PN, Schneider SH. 1999. Simulating the effects of climatechange on tropical montane cloud forests. Nature 398: 608–610.
Takahashi M, Giambelluca TW, Mudd RG, et al. 2010. Rainfallpartitioning and cloud water interception in native forest and invadedforest in Hawai’i Volcanoes National Park. Hydrological Processes 25:448–464.
Westhoff M, Zimmermann D, Zimmermann G, et al. 2009. Distribution andfunction of epistomatal mucilage plugs. Protoplasma 235: 101–105.
Webster GL. 1995. The panorama of Neotropical cloud forests. In: Churchill SP,Balslev H, Forero E, Luteyn JL, eds. Biodiversity and conservation ofNeotropical montane forests. Neotropical Montane Forest Biodiversityand Conservation Symposium 1. New York Botanical Garden, 53–77.
Werner WL. 1988. Canopy dieback in the upper montane rain forests of SriLanka. GeoJournal 17: 245–248.
Zotz G, Tyree MT, Patino S, Carlton MR. 1998. Hydraulic architecture andwater use of selected species from a lower montane cloud forest inPanama. Trees 12: 302–309.
Zou C, Barnes P, Archer S, McMurtry C. 2005. Soil moisture redistribution asa mechanism of facilitation in savanna tree–shrub clusters. Oecologia 145:32–40.
Oliveira et al. — Hydroclimatic conditions and ecophysiology of cloud forests920
at BIB
LIO
TE
CA
CE
NT
RA
L U
NIV
EST
AD
UA
L C
AM
PINA
S on April 24, 2014
http://aob.oxfordjournals.org/D
ownloaded from
Copyright © 2022 FDOKUMEN




















