
Paleoclimatic history and vicariant speciation in the "sand goby" group (Gobiidae, Teleostei)
Upload
independentCategory
view
1download
0
ORIGINAL PAPER
The Gyrodactylus (Monogenea, Gyrodactylidae) parasite faunaof freshwater sand gobies (Teleostei, Gobioidei) in their centreof endemism, with description of seven new species
Maarten P. M. Vanhove & Alcibiades N. Economou &
Stamatis Zogaris & Sofia Giakoumi & Davor Zanella &
Filip A. M. Volckaert & Tine Huyse
Received: 26 August 2013 /Accepted: 7 November 2013# Springer-Verlag Berlin Heidelberg 2013
Abstract While Gobioidei comprises showcases of(adaptive) radiation, the scientific interest they yielded didnot ensure full understanding of goby biodiversity. Even in awell-studied region like Europe, wide knowledge gaps re-main. Sand gobies represent one of the few clades whosemonogenean parasites have been thoroughly studied.However, in the Balkans, part of the sand gobies' centre ofendemism, these parasites were unstudied.We focus on Greekand Croatian freshwater gobies. From five sand goby species,the first parasites are reported, describing seven newGyrodactylus species. Economidichthys pygmaeus harboursGyrodactylus benedeni sp. n. and Gyrodactylus dorlodoti sp.n. Its congener E. trichonis hosts G. meelkopae sp. n.Knipowitschia milleri was found to host G. charon sp. n.,
K. thessala is infected byG. bios sp. n., andK. croatica byG.douglasadamsi sp. n. and G. hellemansi sp. n. Gyrodactylusbubyri was found on its type host K. caucasica . A diverseparasite fauna is expected for a region known for its biodiver-sity and endemism. The contribution of parasites to speciesrichness in such hotspots is overlooked. The observed speciesrichness per host is rather low compared to the better-studiedeastern Atlantic sand gobies. Host vicariance is considered tomediate parasite specificity in this fauna. Some new flatwormspecies display unique morphological features, such as theremarkable size of the marginal hook sickle propercompared to its foot in the Economidichthys parasites,or a characteristically kinked marginal hook sickle in G.douglasadamsi sp. n. These features reflect their hosts'endemism in the Balkans.
Introduction
Parasites account for more than 50 % of the total speciesrichness on this planet (Timm and Clauson 1987; Windsor1998). Their common tendency of exploiting a single hostspecies or infection site is a powerful driver of diversification(de Meeûs et al. 1998). Moreover, parasites are of consider-able importance in terms of biomass (Kuris et al. 2008),ecosystem function (Marcogliese 2004) and indicator species(Sures 2004; Palm and Rückert 2009). Studying the diversityand systematics of parasites helps to unravel the mechanismsunderlying species richness, and will enrich our understandingof this diverse and highly important group of organisms(Brooks and McLennan 1993; Thompson 1999; Brooks andHoberg 2001; Littlewood 2011). A promising approach tounderstand parasite biodiversity is the use of monogeneanflatworms infecting closely related host fishes, in view of their
M. P. M. Vanhove : F. A. M. Volckaert : T. HuyseLaboratory of Biodiversity and Evolutionary Genomics, Departmentof Biology, University of Leuven, Ch. Deberiotstraat 32,3000 Leuven, Belgium
M. P. M. Vanhove :A. N. Economou : S. Zogaris : S. GiakoumiInstitute of Marine Biological Resources and InlandWaters, HellenicCentre for Marine Research, 46.7 km Athens-Sounio Avenue,PO Box 712, Anavyssos 190 13, Greece
D. ZanellaDepartment of Zoology, Faculty of Science, University of Zagreb,Rooseveltov trg 6, HR-10000 Zagreb, Croatia
T. HuyseDepartment of Biology, Royal Museum for Central Africa,Leuvensesteenweg 13, 3080 Tervuren, Belgium
Present Address:M. P. M. Vanhove (*)Department of Botany and Zoology, Faculty of Science, MasarykUniversity, Kotlářská 2, 611 37 Brno, Czech Republice-mail: [email protected]
Parasitol ResDOI 10.1007/s00436-013-3693-8

direct (one-host) life cycle, relative harmless nature (cf. "neu-tral" markers) and close relationship with one or some hostspecies (Pariselle et al. 2003). One of the few cases in which afish lineage has been intensively scrutinized for monogeneanspecies throughout a substantial part of its distribution in-volves the sand gobies and their Gyrodactylus vonNordmann, 1832 parasites (Bakke et al. 2007; see Table 1for an overview of Gyrodactylus species from gobies).
The scientific literature on sand gobies is relatively prolificand recent. This ecologically and behaviorally diverse cladeincludes Pomatoschistus Gill, 1863,Knipowitschia Iljin, 1927,Economidichthys Bianco, Bullock, Miller and Roubal, 1987andGobiusculus Duncker, 1928 (although other closely relatedgenera are confirmed to form part of an "expanded sand gobygroup" by Agorreta et al. 2013). One of its representatives, E.trichonis , is noted as the smallest European freshwater fish(Kottelat and Freyhof 2007).Most of the diversification of sandgobies was allopatric, fuelled by historical vicariance and dis-persal events (Huyse et al. 2004b; Vanhove et al. 2012).However, adaptive processes (e.g., linked to the sensory sys-tem) are thought to contribute to their potential for radiation(Larmuseau et al. 2010). This has also been shown in other(marine) gobies, namely, Elacatinus Jordan, 1904 s.l.gobiosomatines (Larmuseau et al. 2011) and illustrates thecombined importance of biogeographical and adaptive process-es in goby speciation in particular and radiation in general.Indeed, gobies represent some of the prime examples of radi-ations (Miller 1979; Fishelson 1989; Rüber et al. 2003; Taylorand Hellberg 2005). Their highly adaptive nature allowed themto exploit niches that remained fallow ground for most fishtaxa, such as an amphibious lifestyle (Ishimatsu and Gonzales2011) or freshwater habitats of oceanic islands (Ryan 1991).Hence, also these partners in the host–parasite system areideally suited to study speciation mechanisms.
While neutral and protein-coding genes, as well as othercharacters (e.g., drawn from behavior and life history;Malavasi et al. 2012) clearly contributed to our understandingof sand goby evolution, biogeography and taxonomy, consid-erable knowledge gaps remain. A lot of baseline informationon gobies is lacking, including an accurate species delineationand a full grasp of their ecological and distributional ranges.Kovačić and Patzner (2011) mention a discrepancy betweenspecies richness and available data in Mediterranean marinegobies. Even in European freshwaters, where the ichthyofaunain general is relatively depauperate (Lundberg et al. 2000), thisknowledge gap is obvious (Economou et al. 2007; Kottelatand Freyhof 2007; Vanhove et al. 2011a), not to mention ourscant data on their parasite fauna. Regarding sand gobies,Vanhove et al. (2012) recently complemented the phylogenet-ic study by Huyse et al. (2004b) with species from the sandgobies' centre of endemism, the eastern Mediterranean, wherethey are the most important freshwater gobies in terms ofspecies numbers. Likewise, Malavasi et al. (2012) included a
limited number of those sand gobies in a behavior and lifehistory based phylogeny. However, knowledge of taxaparasitising sand gobies is currently limited to the north-eastern Atlantic region and the Western MediterraneanSea, and mostly restricted to marine Pomatoschistus andGobiusculus species (Table 1).
Parasites have proven useful in host identification, biogeog-raphy and evolutionary history, both at the interspecific (Paugyet al. 1990;McDowall 2000; Choudhury andDick 2001; Boegerand Kritsky 2003; Barson et al. 2010; Pariselle et al. 2011;Gillardin et al. 2012; Muterezi Bukinga et al. 2012; Vanhove2012; Vanhove et al. 2013) and intraspecific (MacKenzie 2002;Nieberding et al. 2004; Criscione et al. 2006; Blakeslee et al.2008; Vanhove 2012) level. On a more applied level, a betterunderstanding of goby parasites in South-east Europe is useful inview of the potential effect of introduced Ponto-Caspian gobiesthroughout Europe (Kvach and Skóra 2007; Ondračková et al.2009; Kvach and Winkler 2011). Also, in view of the conserva-tion risks that many sand gobies are facing (Miller 1990;Economidis 1995), potential co-extinction should be taken intoaccount. Indeed, if a host species dies out, many (specific)parasites follow suit (Stork and Lyal 1993; Koh et al. 2004;Dunn et al. 2009; Justine et al. 2012). First and foremost, moreknowledge is needed on the diversity of monogenean parasitesinfecting sand gobies in their centre of endemism, the easternMediterranean. It is the aim of this study to cover that aspect.
Material and methods
Gobies were collected by hand and seine netting, and back-pack electrofishing. They were preserved in 99 % ethanolawaiting further processing. Localities from which parasiteswere retrieved are shown in Fig. 1; they have been describedby Vanhove et al. (2012). In view of sympatry withKnipowitschia caucasica , Proterorhinus semilunaris fromthe Evros river was also inspected for parasites. This speciesbelongs to a lineage clearly distinct from the sand gobies(Thacker and Roje 2011); in fact, it should be noted that sandgobies may even belong to Gobionellidae rather than toGobiidae (Thacker 2013), though Agorreta et al. (2013) con-sider them gobionelline-like gobiids.
Gills, body, fins and recipient were inspected for monoge-neans under an Olympus SZX12 stereomicroscope.Gyrodactylids were removed with a dissection needle.Specimens were treated with 10 % sodium dodecyl sulphatefor a couple of minutes, subsequently fixed with ammoniumpicrate-glycerine (Malmberg 1957) and mounted on a slideunder a cover-slip. Morphological study of the retrieved par-asites focused on the haptoral hard parts, in view of theirtaxonomical importance (García-Vásquez et al. 2012), nextto other features such as the arrangement of the male copula-tory organ (MCO). Measurements, based on Shinn et al.
Parasitol Res

Table 1 Summary of the cur-rently known formally describedGyrodactylus species parasitisingEuropean gobies, with host spe-cies and infection sites, includingthe new species described
Host range data are based onHuyse and Volckaert (2002,2005), Huyse et al. (2003, 2004,2006), Longshaw et al. (2003),Harris et al. (2004), Huyse andMalmberg (2004), Dmitrieva andSkidan (2005), Pugachev et al.(2009), Mierzejewska et al.(2011) and references therein,supplemented with our ownfindings
Parasite species Goby host species
Gyrodactylus alviga Dmitrieva and Gerasev, 2000 Neogobius melanostomus (Pallas, 1814)
Gyrodactylus arcuatus Bychowsky, 1933 Gobiusculus flavescens (Fabricius, 1779)
Knipowitschia panizzae (Verga, 1841)
Pomatoschistus microps (Krøyer, 1838)(Baltic Sea)
Proterorhinus marmoratus (Pallas, 1814)
Gyrodactylus arcuatoides Huyse, Malmberg andVolckaert, 2004
Po. pictus (Malm, 1865)
G. benedeni sp. n. Economidichthys pygmaeus (Holly, 1929)
G. bios sp. n. K. thessala (Vinciguerra, 1921)
G. branchialis Huyse, Malmberg and Volckaert, 2004 Po. marmoratus (Risso, 1810) (Mediterranean)
Po. microps (Krøyer, 1838) (North Sea,Mediterranean)
Po. minutus (Pallas, 1770) (Mediterranean)
G. bubyri Osmanov, 1965 K. caucasica (Kawrajsky in Berg 1916)
G. charon sp. n. K. milleri (Ahnelt and Bianco, 1990); K. sp.
G. dorlodoti sp. n. E. pygmaeus
G. douglasadamsi sp. n. K. croatica
Mrakovčić, Kerovec, Mišetić and Schneider, 1996G. flavescensis Huyse, Malmberg and Volckaert, 2004 Gobiusculus flavescens
G. flesi Malmberg, 1957 Gobius niger Linnaeus, 1758
Neogobius melanostomus
G. gondae Huyse, Malmberg and Volckaert, 2004 Po. lozanoi (de Buen, 1923)
Po. minutus (North Sea, Mediterranean)
G. hellemansi sp. n. K. croatica
Mrakovčić, Kerovec, Mišetić and Schneider, 1996G. leopardinus Dmitrieva and Skidan, 2005 Po. marmoratus
G. longidactylus Geets, Malmberg and Ollevier, 1998 Po. lozanoi
G. meelkopae sp. n. E. trichonis Economidis and Miller, 1990
G. micropsi Gläser, 1974 Po. microps
G. ostendicus Huyse and Malmberg, 2004 K. panizzae
Po. marmoratus (Mediterranean)
Po. microps (North Sea, Mediterranean)
G. proterorhini Ergens, 1967 Babka gymnotrachelus (Kessler, 1857)
Gobius cobitis Pallas, 1814
Gobius niger
Mesogobius batrachocephalus (Pallas, 1814)
Neogobius fluviatilis (Pallas, 1814)
N. melanostomus
Ponticola kessleri (Günther, 1861)
Ponticola platyrostris (Pallas, 1814)
Pr. marmoratus
Pr. semilunaris (Heckel, 1837)
Zosterisessor ophiocephalus (Pallas, 1814)
G. quadratidigitus Longshaw, Pursglove and Shinn, 2003 Thorogobius ephippiatus (Lowe, 1839)
G. rugiensis Gläser, 1974 Po. microps (North Sea, Mediterranean)
Po. minutus (Mediterranean)
G. rugiensoides Huyse and Volckaert, 2002 Po. lozanoi
Po. minutus (North Sea, Mediterranean)
Po. pictus
Parasitol Res

(2004), were taken using a Zeiss Axio Imager Z1 microscopeat a magnification of 100x (oil immersion, 10× ocular) underdifferential interference contrast, with an AxioCamMR3 cam-era and AxioVision v.4.2.8 software.
Identification and comparison with known congeners wasbased on Osmanov (1971), Ergens (1985), Geets et al. (1998),Huyse and Volckaert (2002), Huyse et al. (2003, 2004a),Longshaw et al. (2003), Huyse and Malmberg (2004),Dmitrieva and Skidan (2005), GyroDb (Harris et al. 2008;Shinn et al. 2010), Pugachev et al. (2009) and Mierzejewskaet al. (2011). Taxon names and authorities follow Eschmeyer(2013) for hosts and MonoDb (http://www.monodb.org) forparasites.
Type material is deposited in the Natural History Museum(London, United Kingdom) (NHMUK), the MuséumNational d'Histoire Naturelle (Paris, France) (MNHN) andthe invertebrate type collection of the Department of
Zoology, Swedish Museum of Natural History (Stockholm,Sweden) (SMNH). Vouchers of previously described speciesare in the general invertebrate collection of the Department ofZoology, SMNH. Symbiotypes and host vouchers are kept inthe collection of the research group Zoology: Biodiversity andToxicology of Hasselt University (Diepenbeek, Belgium)(HU) or in the reference collection of the Institute of MarineBiological Resources and Inland Waters, Hellenic Centre forMarine Research (Anavyssos, Greece) (HCMR).
Results
At least nineGyrodactylus species were collected, amongwhichthere are two known species. The specimens infectingCaucasian dwarf goby Knipowitschia caucasica (HUSYMBIOTYPE 004) sampled in the Strymon River on 8 July
Fig. 1 Map showing sampling localities where Gyrodactylus was re-trieved from sand gobies. (1) Ferres, Evros River (40°54′ N, 26°12′ E),(2 ) Strymoniko Bridge, Strymon River (41°3′ N, 23°21′ E), (3 )Farkadona Bridge, Pinios River (39°34′ N, 22°5′ E), (4) Kalamas Delta,Kalamas River (39°32′ N, 20°12′ E), (5) Acheron Delta, Acheron River(39°15′ N, 20°29′ E), (6) Skala Springs, Louros River (39°10′ N, 20°46′E), (7) Agios Georgios Springs, Arachthos River (39°6′ N, 21°1′ E), (8)Vlychos Spring, Vonitsa Lagoon (38°55′ N, 20°52′ E), (9) AstakosSprings (38°33′ N, 21°6′ E), (10) Panaitolio Canal, Lake Trichonis
(38°35′ N, 21°28′ E), (11) Myrtia Stream, Lake Trichonis (38°36′ N,21°33′ E), (12) Lake Trichonis (38°33′ N, 21°35′ E), (13) AitolikoStream, Acheloos Delta (38°27′ N, 21°19′ E), (14) Kryoneri Springs,Evinos Delta (38°21′N, 21°36′ E), (15) Chiliadou Springs, Mornos Delta(38°24′N, 21°55′E), (16) Kalogria Laspoloutra, Prokopos Lagoon (38°9′N, 21°24′ E), (17) Kalogria Channel, Prokopos Lagoon (38°10′N, 21°23′E), (18) Ponari, Morača River, (19) just under the Vučija Spring, VrgoracField, Matica River, Neretva basin (43°10′ N, 17°22′ E)
Parasitol Res

2009 were identified as G. bubyri , the type host of which isindeed K. caucasica . Specimens (SMNH 131463–131467)were recovered from fins (pelvic and second dorsal), skin (head,flank, operculum) and gills. Evros River Proterorhinussemilunaris (HU SYMBIOTYPE 041, collected 13 July 2009)yieldedG. proterorhini (SMNH131468–131469) as fin parasite(pectoral and pelvic). One specimen on the gills of Pr.semilunaris is similar to G. bubyri , but cannot be assigned withcertainty, lacking additional specimens. The fact that both para-sites had not been formally recorded from Pr. semilunaris in theavailable literature (Longshaw et al. 2003; Mierzejewska et al.2011) is due to recent taxonomic changes in this fish genus(Kottelat and Freyhof 2007). Indeed, earlier records of G.proterorhini (ascribed to Pr. marmoratus) very likely concernthe same host. It is easy to recognizeG. proterorhini by virtue ofthe additional ear-shaped haptoral elements, just anterior to theanchors. The case of G. thysi Vanhove, Snoeks, Volckaert andHuyse, 2011 might suggest a link between such structures and aproportionally short anchor root (Vanhove et al. 2011b). A singleGyrodactylus specimen was found on the gills of Montenegrodwarf gobyK.montenegrina Kovačić and Šanda, 2007 from theMorača River (HU SYMBIOTYPE 027, collected 4 September2007), unfortunately damaged — only haptoral parts in uterowere available (Fig. 3l). Therefore, it cannot serve for a formalspecies description. Gyrodactylus species were found that arenew to science on Economidichthys pygmaeus , E. trichonis , K.croatica , K. milleri , K. thessala and K. sp. They are describedbelow. Data on the number of host specimens investigated andon prevalence and infection intensity are provided in Table 2.Morphometric data of the proposed new species are shown inTable 3.
Gyrodactylus benedeni sp. n. Vanhove and Huyse(Figs. 2a–e and 4a; Tables 2 and 3)Type host: Economidichthys pygmaeus (Holly, 1929)
(Economidis' goby).Type locality: Kryoneri Springs (Evinos Delta), Greece
(7 June 2008, on HU SYMBIOTYPE 018).Other localities: Kalamas Delta (Kalamas River) (8 June
2008, on HU SYMBIOTYPE 011–016), Lake Trichonis(9 June 2008, on HU SYMBIOTYPE 019–023), AitolikoStream (Acheloos Delta) (28 September 2006, on HCMREpy - Alc 017 and 022), Skala Springs (Louros River) (6 July2006, on HU SYMBIOTYPE 010), Agios Georgios Springs(Arachthos River) (9 May 2007, on HU SYMBIOTYPE 005),Chiliadou Springs (Mornos Delta) (15 January 2007, on HUSYMBIOTYPE 008-009), Astakos Springs (4 July 2006, onHU SYMBIOTYPE 006–007), Vlychos Spring (VonitsaLagoon) (4 July 2006, on HU SYMBIOTYPE 024) (all inGreece), on type host.
Material studied: 58 specimens.Type material: holotype: NHMUK 2013.7.24.1; paratypes:
NHMUK 2013.7.24.2-11, MNHN HEL324-331, SMNHType-8501-8512.
Infection site: fins (pectoral, pelvic, both dorsals, caudal),skin (eye, flank, mouth, anus).
Etymology: the species epithet honors Prof. Pierre-JosephVan Beneden (1809–1894), a Belgian zoologist active infields as diverse as fossil cetaceans and aquatic parasites; hewas the first ever to found a marine biological station (inOstend, Belgium) and to disentangle the tapeworm's life cy-cle. He also studiedGyrodactylus and other monogeneans andwas professor at the University of Leuven, where the lab workfor this study was carried out.
Diagnosis: anchor slender, sharply curved. Anchor rootslightly curved inwards. Ventral bar slender, with laterallypointing, well-developed but blunt processes (in contrast tothe larger, more ear-shaped processes pointing anteriorly in,e.g., G. arcuatus or G. proterorhini). Ventral bar shield trian-gular but rounded, without clear striation, similar to other sandgoby parasites like G. rugiensis and G. rugiensoides . Theshape of the marginal hook is however very distinctive: with asubstantial difference in proximal and distal width of thesickle, the sickle foot is minute in comparison to the sickleproper, which is about as thick as the foot. Sickle point onlyweekly curved or sharpened. MCO with seven to ninespinelets in a single row and one apical spine.
Gyrodactylus bios sp. n. Vanhove and Huyse(Figs. 3d–f and 4b; Tables 2 and 3)Type host: Knipowitschia thessala (Vinciguerra, 1921)
(Thessaly goby).Type locality: near Farkadona Bridge, main course of the
Pinios River (Greece) (July 2009, on HU SYMBIOTYPE039–040).
Material studied: eight specimens.Type material: holotype: NHMUK 2013.7.24.21;
paratypes: NHMUK 2013.7.24.20, MNHN HEL339, SMNHType-8522.
Infection site: skin (eye, flank), fins (caudal, dorsal).Etymology: the species epithet, as a noun in apposition, is
derived from BIOS, the biology student union of the Universityof Leuven, on the occasion of its 60th anniversary in 2012 and ingratitude of many joyful memories of the first author, formerly aquaestor, praeses and board member, now an honorary member.
Diagnosis: similar to several congeners infecting sand gobies(G. bubyri ,G. micropsi ,G. rugiensis ,G. rugiensoides),G. biossp. n. has slender anchors and a simple ventral bar with short,blunt processes and a modest, triangular, rounded shield.However, it differs from these species by its marginal hook sicklepoint, extending beyond its toe. This distinguishes it also fromG.longidactylus . Other species with such far-reaching marginalhook point include G. quadratidigitatus and G. leopardinus ,whose ventral bar processes are however larger. In addition, G.quadratidigitatus has a firmer anchorwith short root. One row ofseven to nine spinelets and an apical spine in the MCO.
Gyrodactylus charon sp. n. Vanhove and Huyse(Figs. 3a–c and 4c; Tables 2 and 3)
Parasitol Res

Type host: Knipowitschia milleri (Ahnelt and Bianco,1990) (Miller's dwarf goby).
Additional host: Knipowitschia sp.Type locality: Acheron Delta (Acheron River)
(Greece), on type host (5 July 2006, on HCMR Kn -Alc 025; 17 January 2007, on HU SYMBIOTYPE 031–033; 8 June 2008, on HU SYMBIOTYPE 028–030 and034).
Additional localities: Kalogria Laspoloutra (ProkoposLagoon) (5 June 2008, on HU SYMBIOTYPE 036),Kalogria Channel (Prokopos Lagoon) (5 June 2008, on HUSYMBIOTYPE 035 and 037–038), Greece, on K. sp.
Material studied: 29 specimens.Type material: holotype: NHMUK 2013.7.24.14;
paratypes: NHMUK 2013.7.24.15-19, MNHN HEL334-338,SMNH Type-8515-8521.
Infection site: fins (both dorsals, anal, pectoral, pelvic),skin (head, flank), gill filaments.
Etymology: the species epithet is used as a noun in appo-sition. Charon is the mythological ferryman ferrying souls tothe underworld; one of the entrances can be reached bycrossing the river Acheron, the type locality of this species.
Diagnosis: like inG. bios sp. n., marginal hooks clearly notbelonging to the same type as in Economidichthys parasites.Similarly, marginal hook sickle point extending beyond its toe. Itis distinct from the sickle of G. bios sp. n. by the point directedalong a more downwards angle, and by its toe with a gentlerslope (marginal hook foot ends more abruptly in a straightdownwards pointing toe in G. bios sp. n., though not to theextent of G. quadratidigitatus where the toe is blunt and"square"). Conversely, the marginal hook of G. bubyri has asomewhat sturdier sickle proper and a toe pointing further
Table 2 Number of host specimens investigated, number of host specimens infected, and number of parasite specimens retrieved per infected hostspecimen, for Gyrodactylus recovered from freshwater sand gobies
Parasite species Sampling localityand date
# host specimensstudied
# host specimensinfected
# parasite specimenson infected specimens(minimum–maximum)
G. benedeni sp. n. Agios Georgios Springs, May 2007 2 1 1
Aitoliko Stream, September 2006 8 6 1–11
Astakos Springs, July 2006 2 2 1–2
Chiliadou Springs, January 2007 2 2 1–>25
Kalamas Delta, June 2008 16 8 1–4
Kryoneri Springs, June 2008 4 2 4–6
Lake Trichonis, June 2008 4 3 2–4
Vlychos Spring, July 2006 2 1 1
Skala Springs, July 2006 2 1 3
G. bios sp. n. Pinios River, July 2009 4 3 2–8
G. bubyri Strymon River 4 1 6 (fins or skin)
1 (gills)
G. charon sp. n. Acheron Delta, July 2006 8 1 13 (fins or skin)
2 (gills)
Acheron Delta, January 2007 6 3 (fins or skin) 2–4 (fins or skin)
Acheron Delta, June 2008 4 4 (fins or skin) 1–8 (fins or skin)
1 (gills) 1 (gills)
Kalogria Channel, June 2008 3 1 (gills) 1
Kalogria Laspoloutra, June 2008 4 1 (gills) 1
G. dorlodoti sp. n. Aitoliko Stream, September 2006 8 4 1–4
Lake Trichonis, June 2008 4 1 4
G. douglasadamsi sp. n. Matica River, May 2012 3 3 (fins or skin) 6–15 (fins or skin)
2 (gills) 1–2 (gills)
G. hellemansi sp. n. 3 2 1–7
G. meelkopae sp. n. Myrtia Stream, May 2009 42 1 1(+ 1 in recipient)
Panaitolio Canal, May 2009 7 1 (gills) 1
G. sp. on Knipowitschia montenegrina Morača River, September 2007 6 1 (gills) 1
See Fig. 1 and species descriptions for more detailed sampling data
Parasitol Res

Tab
le3
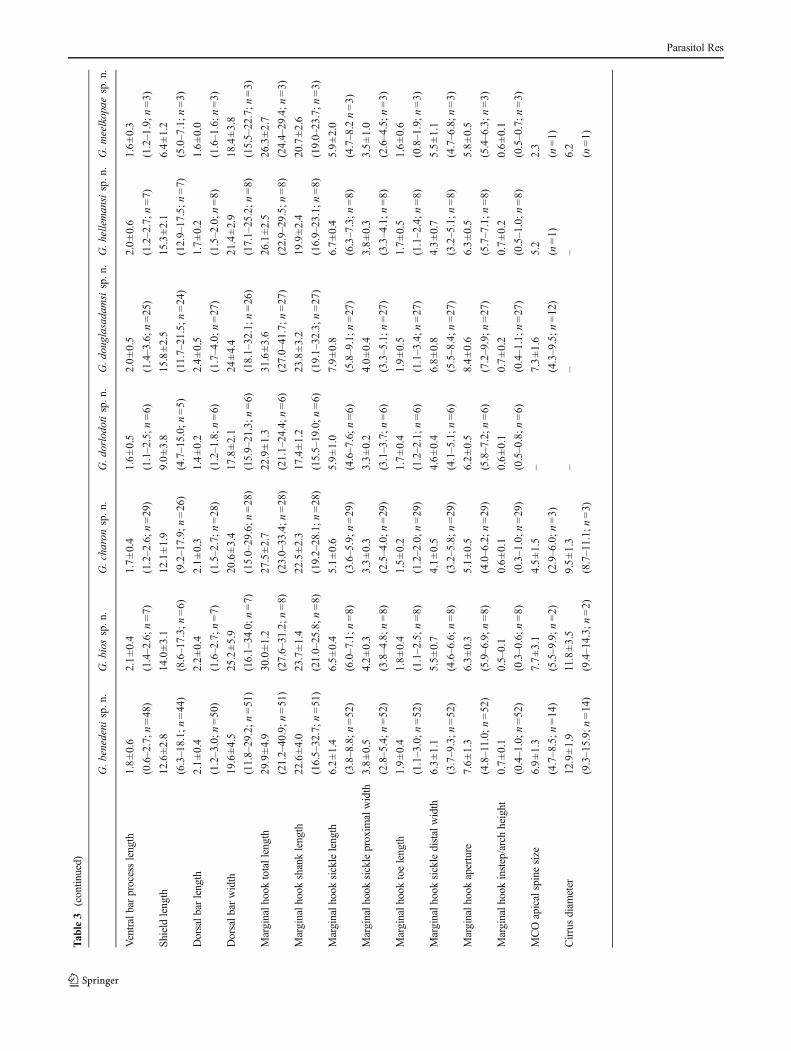
Morphom
etricdataof
body,haptoralhardpartsandmalecopulatory
organ(M
CO)o
fthe
newlydescribedGyrodactylusspecies,allinμm(distances)o
r°(angles),depictedas
average±standard
deviation,with
rangeandnumberof
measurementsin
parentheses
G.benedenisp.n.
G.biossp.n.
G.charonsp.n.
G.dorlodotisp.n.
G.douglasadam
sisp.n.
G.hellemansisp.n.
G.m
eelkopae
sp.n.
Bodywidth
84.3±18.6
152.1±5.5
100.9±22.2
73.6±13.6
111.3±22.1
116.5±24.1
77.2
(46.6–122.7;
n=39)
(145.6–158.1;n
=4)
(73.8–152.5;
n=12)
(53.6–84.4;n
=4)
(80.6–180.5;
n=20)
(100.3–151.7;n
=4)
(n=1)
Bodylength
248.3±40.9
337.4±21.0
253.8±42.1
230.3±30.3
324.9±66.5
323.3±105.4
196.4
(168.2–382.3;n
=38)
(314.0–358.4;n
=4)
(202.3–316.2;n
=12)
(207.1–274.8;n
=4)
(213.0–464.4;n
=20)
(242.2–478.2;n
=4)
(n=1)
Anchoraperture
15.7±1.8
17.1±2.6
15.4±2.1
16.4±3.5
17.6±2.4
16.1±1.6
18.8±11.4
(12.7–22.0;n
=51)
(14.0–20.2;n
=4)
(11.1–19.5;n
=25)
(11.3–19.6;n
=6)
(13.7–22.0;n
=28)
(14.3–18.9;n
=8)
(9.1–31.3;
n=3)
Anchorproxim
alshaftw
idth
6.4±0.6
7.1±0.4
6.5±0.6
4.8±0.7
6.7±0.5
5.8±0.3
6.1±0.7
(4.6–8.2;n
=51)
(6.6–7.7;n
=7)
(5.2–7.9;n
=29)
(4.2–6.0;n
=6)
(5.7–8.0;n
=29)
(5.4–6.2;n
=8)
(5.6–6.9;n
=3)
Anchorpointlength
25.1±2.2
28.2±0.6
24.2±1.3
16.1±1.4
28.6±1.2
19.4±0.8
22.4±2.6
(20.8–28.8;n
=51)
(27.6–28.9;n
=4)
(21.2–26.7;n
=25)
(13.9–17.9;n
=6)
(26.1–30.7;n
=28)
(18.3–20.6;n
=8)
(20.1–25.3;n
=3)
Anchordistalshaftw
idth
3.8±0.3
4.4±0.3
4.1±0.5
2.9±0.4
4.2±0.4
3.6±0.5
4.2±0.6
(3.2–4.7;n
=51)
(4.0–4.8;n
=4)
(3.3–5.2;n
=25)
(2.4–3.6;n
=6)
(3.6–5.0;n
=27)
(2.8–4.2;n
=8)
(3.8– 5.0;n
=3)
Anchorshaftlength
30.3±2.3
32.2±1.0
28.6±1.8
22.7±2.1
33.0±1.5
26.1±1.1
26.3±5.8
(26.0–34.2;n
=51)
(31.0–33.4;n
=4)
(24.4–31.2;n
=25)
(19.9–24.8;n
=6)
(28.3–35.5;n
=26)
(23.9–27.4;n
=8)
(21.6–32.7;n
=3)
Anchorinnercurvelength
2.4±1.2
1.9±0.4
1.4±0.6
2.1±1.1
1.9±0.6
1.3±0.7
1.6±1.1
(0.7–6.6;n
=51)
(1.5–2.3;n
=4)
(0.6–3.1;n
=25)
(1.0–3.5;n
=6)
(0.8–2.9;n
=27)
(0.6–2.6;n
=8)
(0.9–2.9;n
=3)
Anchoraperture
angle
32.55±3.6
33.7±4.7
34.0±5.6
48.1±7.8
33.4±4.9
39.5±5.5
39.2±14.2
(26.4–48.8;n
=51)
(28.7–40.1;n
=4)
(24.1–48.8;n
=24)
(38.7–58.3;n
=6)
(26.2–44.6;n
=27)
(30.1–45.7;n
=8)
(24.1–52.4;n
=3)
Anchorpointcurve
angle
7.5±4.0
5.0±1.1
4.8±1.9
11.4±6.7
5.0±1.8
6.4±2.4
7.0±4.3
(2.2–21.0;
n=51)
(4.0–6.5;n
=4)
(2.0–9.2;n
=24)
(5.4–19.9;
n=6)
(2.2–8.2;n
=27)
(3.6–10.6;
n=8)
(3.6–11.8;
n=3)
Inneranchor
aperture
angle
37.1±4.0
39.2±5.5
39.5±6.6
54.4±8.3
38.1±5.5
47.0±5.1
45.3±15.8
(31.0–55.8;n
=51)
(33.1–46.4;n
=4)
(28.1–56.7;n
=24)
(44.3–65.3;n
=6)
(30.4–50.2;n
=27)
(41.2–53.0;n
=8)
(28.3–59.4;n
=3)
Anchorroot
length
15.3±1.6
18.2±1.3
16.1±1.4
13.2±1.3
17.7±1.8
13.9±1.0
12.3±2.7
(11.2–18.5;n
=51)
(16.3–20.1;n
=7)
(13.9–19.2;n
=29)
(11.7–15.2;n
=6)
(14.5–22.6;n
=27)
(12.3–15.2;n
=8)
(10.0–15.2;n
=3)
Anchortotallength
48.2±3.5
52.2±1.9
47.4±2.4
37.9±1.7
53.9±1.7
41.9±1.3
40.8±6.4
(40.3–53.7;n
=50)
(50.6–54.9;n
=4)
(41.7–52.6;n
=25)
(35.7–40.1;n
=6)
(49.6–57.8;n
=26)
(40.2–43.4;n
=8)
(35.4–47.8;n
=3)
Ventralbartotalw
idth
22.3±3.3
27.9±2.5
22.7±2.9
17.2±1.7
28.2±5.3
22.4±3.4
17.3±1.6
(13.4–29.4;n
=46)
(23.4–30.8;n
=7)
(17.3–28.1;n
=28)
(15.3–19.7;n
=6)
(20.3–39.0;n
=20)
(19.5–29.5;n
=7)
(15.6–18.6;n
=3)
Ventralbartotallength
20.3±3.0
21.8±2.7
20.1±2.2
13.8±3.8
23.7±2.6
21.3±2.1
11.7±1.0
(13.9–25.9;n
=44)
(17.0–25.0;n
=6)
(15.8–25.9;n
=26)
(9.1–19.6;
n=5)
(19.3–29.1;n
=24)
(18.7–24.1;n
=7)
(10.7–12.6;n
=3)
Ventralbarprocess-to-m
id-length
2.9±0.9
1.7±1.0
2.6±1.1
1.6±0.5
1.9±0.5
1.6±0.5
1.5±0.7
(0.9–5.2;n
=47)
(0.7–3.2;n
=7)
(0.8–4.7;n
=28)
(1.1–2.2;n
=6)
(0.6–2.5;n
=18)
(1.0–2.1;n
=6)
(0.8–2.3;n
=3)
Ventralbarmedianlength
5.2±0.7
6.5±0.8
5.5±1.1
3.2±0.4
6.5±0.9
4.4±0.3
4.1±0.2
(3.9–7.5;n
=50)
(5.8–8.1;n
=7)
(3.8–8.7;n
=28)
(2.8–3.8;n
=6)
(5.3–8.1;n
=25)
(3.9–4.7;n
=6)
(4.0–4.3;n
=3)
Parasitol Res
Tab
le3
(contin
ued)
G.benedenisp.n.
G.biossp.n.
G.charonsp.n.
G.dorlodotisp.n.
G.douglasadam
sisp.n.
G.hellemansisp.n.
G.m
eelkopae
sp.n.
Ventralbarprocesslength
1.8±0.6
2.1±0.4
1.7±0.4
1.6±0.5
2.0±0.5
2.0±0.6
1.6±0.3
(0.6–2.7;n
=48)
(1.4–2.6;n
=7)
(1.2–2.6;n
=29)
(1.1–2.5;n
=6)
(1.4–3.6;n
=25)
(1.2–2.7;n
=7)
(1.2–1.9;n
=3)
Shield
length
12.6±2.8
14.0±3.1
12.1±1.9
9.0±3.8
15.8±2.5
15.3±2.1
6.4±1.2
(6.3–18.1;
n=44)
(8.6–17.3;
n=6)
(9.2–17.9;
n=26)
(4.7–15.0;
n=5)
(11.7–21.5;n
=24)
(12.9–17.5;n
=7)
(5.0–7.1;n
=3)
Dorsalb
arlength
2.1±0.4
2.2±0.4
2.1±0.3
1.4±0.2
2.4±0.5
1.7±0.2
1.6±0.0
(1.2–3.0;n
=50)
(1.6–2.7;n
=7)
(1.5–2.7;n
=28)
(1.2–1.8;n
=6)
(1.7–4.0;n
=27)
(1.5– 2.0;n
=8)
(1.6–1.6;n
=3)
Dorsalb
arwidth
19.6±4.5
25.2±5.9
20.6±3.4
17.8±2.1
24±4.4
21.4±2.9
18.4±3.8
(11.8–29.2;n
=51)
(16.1–34.0;n
=7)
(15.0–29.6;n
=28)
(15.9–21.3;n
=6)
(18.1–32.1;n
=26)
(17.1–25.2;n
=8)
(15.5–22.7;n
=3)
Marginalh
ooktotallength
29.9±4.9
30.0±1.2
27.5±2.7
22.9±1.3
31.6±3.6
26.1±2.5
26.3±2.7
(21.2–40.9;n
=51)
(27.6–31.2;n
=8)
(23.0–33.4;n
=28)
(21.1–24.4;n
=6)
(27.0–41.7;n
=27)
(22.9–29.5;n
=8)
(24.4–29.4;n
=3)
Marginalh
ookshanklength
22.6±4.0
23.7±1.4
22.5±2.3
17.4±1.2
23.8±3.2
19.9±2.4
20.7±2.6
(16.5–32.7;n
=51)
(21.0–25.8;n
=8)
(19.2–28.1;n
=28)
(15.5–19.0;n
=6)
(19.1–32.3;n
=27)
(16.9–23.1;n
=8)
(19.0–23.7;n
=3)
Marginalh
ooksicklelength
6.2±1.4
6.5±0.4
5.1±0.6
5.9±1.0
7.9±0.8
6.7±0.4
5.9±2.0
(3.8–8.8;n
=52)
(6.0–7.1;n
=8)
(3.6–5.9;n
=29)
(4.6–7.6;n
=6)
(5.8–9.1;n
=27)
(6.3–7.3;n
=8)
(4.7–8.2n=3)
Marginalh
ooksickleproxim
alwidth
3.8±0.5
4.2±0.3
3.3±0.3
3.3±0.2
4.0±0.4
3.8±0.3
3.5±1.0
(2.8–5.4;n
=52)
(3.8–4.8;n
=8)
(2.5–4.0;n
=29)
(3.1–3.7;n
=6)
(3.3–5.1;n
=27)
(3.3–4.1;n
=8)
(2.6–4.5;n
=3)
Marginalh
ooktoelength
1.9±0.4
1.8±0.4
1.5±0.2
1.7±0.4
1.9±0.5
1.7±0.5
1.6±0.6
(1.1–3.0;n
=52)
(1.1–2.5;n
=8)
(1.2–2.0;n
=29)
(1.2–2.1;n
=6)
(1.1–3.4;n
=27)
(1.1–2.4;n
=8)
(0.8–1.9;n
=3)
Marginalh
ooksickledistalwidth
6.3±1.1
5.5±0.7
4.1±0.5
4.6±0.4
6.8±0.8
4.3±0.7
5.5±1.1
(3.7–9.3;n
=52)
(4.6–6.6;n
=8)
(3.2– 5.8;n
=29)
(4.1–5.1;n
=6)
(5.5–8.4;n
=27)
(3.2–5.1;n
=8)
(4.7–6.8;n
=3)
Marginalh
ookaperture
7.6±1.3
6.3±0.3
5.1±0.5
6.2±0.5
8.4±0.6
6.3±0.5
5.8±0.5
(4.8–11.0;
n=52)
(5.9–6.9;n
=8)
(4.0–6.2;n
=29)
(5.8–7.2;n
=6)
(7.2–9.9;n
=27)
(5.7–7.1;n
=8)
(5.4–6.3;n
=3)
Marginalh
ookinstep/archheight
0.7±0.1
0.5–0.1
0.6±0.1
0.6±0.1
0.7±0.2
0.7±0.2
0.6±0.1
(0.4–1.0;n
=52)
(0.3–0.6;n
=8)
(0.3–1.0;n
=29)
(0.5–0.8;n
=6)
(0.4–1.1;n
=27)
(0.5–1.0;n
=8)
(0.5–0.7;n
=3)
MCOapicalspinesize
6.9±1.3
7.7±3.1
4.5±1.5
–7.3±1.6
5.2
2.3
(4.7–8.5;n
=14)
(5.5–9.9;n
=2)
(2.9–6.0;n
=3)
(4.3–9.5;n
=12)
(n=1)
(n=1)
Cirrusdiam
eter
12.9±1.9
11.8±3.5
9.5±1.3
––
–6.2
(9.3–15.9;
n=14)
(9.4–14.3;
n=2)
(8.7–11.1;
n=3)
(n=1)
Parasitol Res

downwards compared to G. charon sp. n. One row of seven tonine spinelets and an apical spine in the MCO.
Gyrodactylus dorlodoti sp. n. Vanhove and Huyse(Figs. 2f–h and 4d; Tables 2 and 3)
Fig. 2 Micrographs of the hardparts of the new Gyrodactylusspecies living on Economidichthyssand gobies. a G. benedeni sp. n.(type locality): haptor. b G.benedeni sp. n. (type locality):marginal hooks. c G. benedeni sp.n. (Mornos): haptor. d G. benedenisp. n. (Mornos): marginal hooks. eG. benedeni sp. n. (Mornos): malecopulatory organ. f G. dorlodotisp. n. (type locality): anchor. g, hG. dorlodoti sp. n. (type locality):marginal hook sickles. i G.meelkopae sp. n. (type locality):anchors. j G. meelkopae sp. n.(type locality): marginal hooksickles. k G. meelkopae sp. n.(type locality): male copulatoryorgan. l G. meelkopae sp. n.(Panaitolio Canal): marginal hooksickles. All scale bars=20 μmexcept for e, j=10 μm andk=5 μm
Parasitol Res
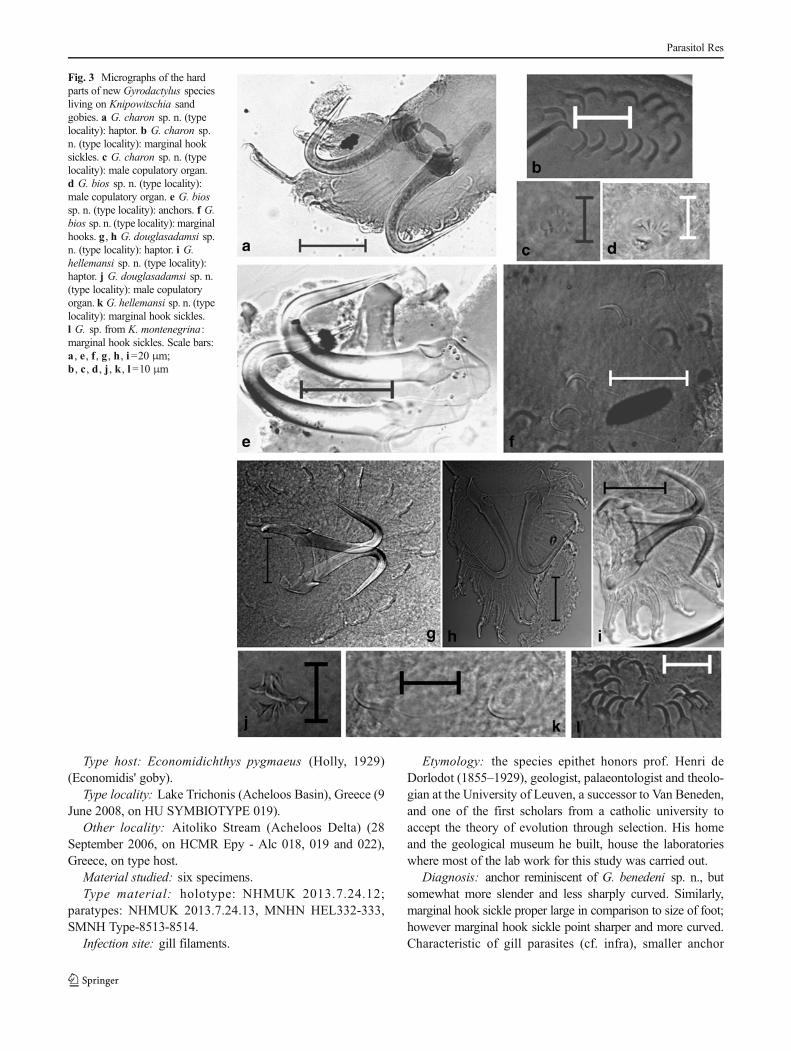
Type host: Economidichthys pygmaeus (Holly, 1929)(Economidis' goby).
Type locality: Lake Trichonis (Acheloos Basin), Greece (9June 2008, on HU SYMBIOTYPE 019).
Other locality: Aitoliko Stream (Acheloos Delta) (28September 2006, on HCMR Epy - Alc 018, 019 and 022),Greece, on type host.
Material studied: six specimens.Type material: holotype: NHMUK 2013.7.24.12;
paratypes: NHMUK 2013.7.24.13, MNHN HEL332-333,SMNH Type-8513-8514.
Infection site: gill filaments.
Etymology: the species epithet honors prof. Henri deDorlodot (1855–1929), geologist, palaeontologist and theolo-gian at the University of Leuven, a successor to Van Beneden,and one of the first scholars from a catholic university toaccept the theory of evolution through selection. His homeand the geological museum he built, house the laboratorieswhere most of the lab work for this study was carried out.
Diagnosis: anchor reminiscent of G. benedeni sp. n., butsomewhat more slender and less sharply curved. Similarly,marginal hook sickle proper large in comparison to size of foot;however marginal hook sickle point sharper and more curved.Characteristic of gill parasites (cf. infra), smaller anchor
Fig. 3 Micrographs of the hardparts of new Gyrodactylus speciesliving on Knipowitschia sandgobies. a G. charon sp. n. (typelocality): haptor. b G. charon sp.n. (type locality): marginal hooksickles. c G. charon sp. n. (typelocality): male copulatory organ.d G. bios sp. n. (type locality):male copulatory organ. e G. biossp. n. (type locality): anchors. f G.bios sp. n. (type locality): marginalhooks. g , h G. douglasadamsi sp.n. (type locality): haptor. i G.hellemansi sp. n. (type locality):haptor. j G. douglasadamsi sp. n.(type locality): male copulatoryorgan. k G. hellemansi sp. n. (typelocality): marginal hook sickles.l G. sp. from K. montenegrina:marginal hook sickles. Scale bars:a , e , f , g , h, i=20 μm;b , c , d , j , k , l=10 μm
Parasitol Res

compared to G. benedeni sp. n. infesting skin and fins of thesame fish species. Slender appearance of ventral bar because ofthin median portion. MCO not observed.
Gyrodactylus douglasadamsi sp. n. Vanhove and Huyse(Figs. 3g,h,j and 4f; Tables 2 and 3)Type host: Knipowitschia croatica Mrakovčić, Kerovec,
Mišetić and Schneider, 1996 (Vrgorac goby).Type locality: just under the Vučija Spring, Vrgorac Field,
Matica River, Neretva Basin, Croatia (11 May 2012, on HUSYMBIOTYPE 001–003).
Material studied: 29 specimens.Type material: holotype: NHMUK 2013.7.24.23;
paratypes: NHMUK 2013.7.24.24-27, MNHN HEL341-344,SMNH Type-8524-8526.
Infection site: fins (pectoral, pelvic, both dorsals, base offirst dorsal fin), gill filaments.
Etymology: the species epithet honors the English writerDouglas Adams (1952–2001). Not only did he create thefascinating universes of The Hitchhiker's Guide to theGalaxy and Dirk Gently, he was also an avidenvironmentalist/conservationist. This is shown exquisitelyin Last Chance to See , a telling account on endangeredspecies and people's perception of them.
Diagnosis: anchor slender and sharply curved. Large ven-tral bar with broad triangular shield and well-developed, bluntprocesses. Marginal hook sickle with relatively small andclearly arched foot. Sickle proper of marginal hook character-istically kinked. This feature seems new to the genus and
Fig. 4 Drawings of anchor andmarginal hook sickle of aGyrodactylus benedeni sp. n. b G.bios sp. n. c G. charon sp. n. d G.dorlodoti sp. n. e G. meelkopaesp. n. f G. douglasadamsi sp. n.g G. hellemansi sp. n. All scalebars=8 μm
Parasitol Res

suffices to distinguish G. douglasadamsi sp. n. from its con-geners. As it was observed in the haptors and in utero in allstudied specimens, we are confident this is not an artefact fromstraining or flattening during the mounting process. MCOwith apical spine and a single row of five to six spinelets.
Gyrodactylus hellemansi sp. n. Vanhove and Huyse(Figs. 3i,k and Fig. 4g; Tables 2 and 3)Type host: Knipowitschia croatica Mrakovčić, Kerovec,
Mišetić and Schneider, 1996 (Vrgorac goby).Type locality: just under the Vučija Spring, Vrgorac Field,
Matica River, Neretva Basin, Croatia (11 May 2012, on HUSYMBIOTYPE 002–003).
Material studied: eight specimens.Type material: holotype: NHMUK 2013.7.24.28;
paratypes: NHMUK 2013.7.24.29-31, MNHN HEL345,SMNH Type-8527.
Infection site: gill filaments.Etymology: the species epithet honors senior research tech-
nician ing. Bart Hellemans (Belgium), in gratitude for theskillful and patient way in which he taught the first author,and many other students and scholars, the ropes of moleculargenetic laboratory work, and for his many other contributionsto our research.
Diagnosis: this parasite, only found on gills, has a smallerand more delicate anchor than G. douglasadamsi sp. n., itscongener on both fins and gills of the same host species.Marginal hook sickle with a distinctly curved foot; the slendersickle point extends far beyond the toe. This latter feature isreminiscent ofG. bios sp. n.; inG. hellemansi sp. n., however,the heel of the marginal hook sickle is larger in proportion tothe sharply downward pointing toe. MCO with apical spineand six spinelets arranged in one row.
Gyrodactylus meelkopae sp. n. Vanhove and Huyse(Figs. 2i–l and 4e; Tables 2 and 3)Type host: Economidichthys trichonis Economidis and
Miller, 1990 (Trichonis goby).Type locality: Myrtia Stream (Lake Trichonis), Greece
(5 May 2009, on HU SYMBIOTYPE 025).Additional locality: Panaitolio Canal (Lake Trichonis),
Greece (5 May 2009, on HU SYMBIOTYPE 026).Material studied: three specimens.Type material: holotype: NHMUK 2013.7.24.22;
paratypes: MNHN HEL340, SMNH Type-8523.Infection site: head, body, gill rakers.Etymology: the species epithet honors biologist dr. Ellen
Meelkop (The Netherlands/Belgium/Australia), a close friendof the first author. Amongst her many good influences, shealso taught him the use of the microscope with which themeasurements were performed.
Diagnosis: the long and thick marginal hook sickle proper(compared to its foot) is indicative of an affinity withEconomidichthys parasites, despite some intra-individual dif-ferences in shape. Marginal hook toe and heel point
downwards, hence the foot appears curved. Anchor moredistinct, especially in the root, which is firmer and blunterthan in G. benedeni sp. n. or G. dorlodoti sp. n. One circle ofeight spinelets in MCO; MCO seems minute both in terms ofcirrus diameter and length of the apical spine (though cautionis needed since theMCOwas observed in one specimen only).
Discussion
TheGyrodactylus fauna of sand gobies has fostered numerousstudies on the diversity and evolution of fishes and theirparasites (Huyse and Volckaert 2002, 2005; Huyse andMalmberg 2004; Huyse et al. 2003, 2004a, b, 2006). It istherefore considered one of the best-studied host systems withregard to (gyrodactylid) monogeneans (Bakke et al. 2007).Unfortunately, the centre of endemism of the sand gobies wasmissing in both fish and parasite studies until recently(Vanhove et al. 2012). Here, we include at least seven nominalsand goby species fromGreece and Croatia, and describe theirhitherto unknown parasite fauna. This is the first report of anyparasite living on Economichthys pygmaeus (Economou et al.2004a), E. trichonis (Economou et al. 2004b), K. croatica(Miller 2004), K. milleri (Miller et al. 2004), and K. thessala(Miller and Economidis 2004). The same goes for the recentlydescribed K. montenegrina . The only earlier study on mono-geneans of endemic Greek freshwater fishes that we found inthe literature yielded three new Dactylogyrus Diesing, 1850species on the endemic barb Barbus prespensis Karaman,1924 from the Balkans (Dupont and Lambert 1986). In viewof the high aquatic endemicity of the Balkans in general(Kryštufek and Reed 2004) and of the Greek ichthyofaunain particular (Economou et al. 2007), the discovery of newspecies, certainly in an understudied group like monogeneans,can be expected in the region. In the context of endemism, it isnot surprising that this parasitological survey of an endemicgoby radiation yields a parasite fauna of similar endemicity,which is morphologically deviant from its congeners onPomatoschistus . For example, the kinked marginal hook sick-le proper of G. douglasadamsi sp. n. seems a new trait to thegenus. Similarly, it is noteworthy that Gyrodactylus speciesliving on Economidichthys display a hitherto unique marginalhook sickle shape, with a proportionally quite small foot andthick sickle proper. Further surveys should assess whether thischaracterizes an endemic lineage to the Acheloos Basin or theIonian Ecoregion, as is the case for this host genus.Knipowitschia also occurs in Lake Trichonis, in sympatry withboth Economidichthys species (e.g., Daoulas et al. 1993). It ishence worthwhile to scrutinize the Lake Trichonis representa-tives of the former genus for Gyrodactylus .
Some of the newly described species shed additional lighton the taxonomy of their host. Vanhove et al. (2012) suggestedthat the various Ionian Ecoregion Knipowitschia sp.
Parasitol Res

populations are conspecific to K. milleri , the alleged endemicof the Acheron River. Knipowitschia milleri as well asProkopos Lagoon Knipowitschia sp. share G. charon sp. n.as their parasite. As no other Balkan freshwater sand gobyspecies share Gyrodactylus species, we propose this fact is anadditional argument for the conspecificity of host populations.The same reasoning goes for the genetically highly divergentMornos population of E. pygmaeus (Vanhove et al. 2012),which is infected by G. benedeni sp. n., like all other E.pygmaeus populations.
Conversely, all parasite species observed seem to be con-fined to a single sand goby host species. This observationcombined with the high level of endemism in the Balkanichthyofauna, suggests that the species richness of fish para-sites and other symbionts is considerable and at least mirrorsthat of the hosts. However, host diversity cannot be the onlycause of parasite species richness. Indeed, E. pygmaeus andK. croatica both harbour twoGyrodactylus species. Althoughsampling bias is hard to rule out, the minute size of E.trichonis and the other Knipowitschia species might relateto their less speciose Gyrodactylus fauna (see Poulin 2002).
It is well known that Gyrodactylus species can be site-specific. Preferential infection sites correlate with consistentmorphological differences (Huyse and Malmberg 2004). Inthe case of G. bubyri , G. charon sp. n. and G. meelkopae sp.n., single species infect different sites (fins/body vs. gills).Conversely, for G. benedeni sp. n. and G. dorlodoti sp. n., aswell as for G. douglasadamsi sp. n. and G. hellemansi sp. n.,haptor morphology (smaller anchors ofG. dorlodoti sp. n. andG. hellemansi sp. n.) may reflect the characteristic morpho-logical differences between fin and gill parasites, respectively(although site preference is less strict inG. douglasadamsi sp.n.). Sometimes species exploiting different infection sites ofone host species have a sister-group relation (see Přikrylováet al. 2012 for an example from mochokid catfishes). Thisphenomenon of within-host speciation also took place in sandgobies, albeit infrequently (Huyse and Volckaert 2005).However, infection site may change rapidly. Several casesare known of site-switching within conspecifics within a hostspecies (Raeymaekers et al. 2008), within conspecifics onseveral host species (Vanhove et al. 2011b) and within line-ages (Huyse et al. 2003). Related to this low phylogeneticconstraint on infection site, the presence of both a gill and a finparasite on Pomatoschistus sand gobies is mostly a result ofthe fact that several lineages colonized this genus at somepoint in time (Huyse and Volckaert 2005). Co-phylogeneticanalyses on the basis of molecular data are ongoing, to assessto what extent host-switching and within-host duplicationevents contributed to the sand goby gyrodactylid fauna inthe eastern Mediterranean, and to verify whether co-speciation events occurred. By establishing the sequence ofdivergence events and the direction of potential host-switches,genetic data will also answer the question whether the eastern
Mediterranean is, just like for the hosts, the centre of ende-mism of these parasites.
One might a fortiori expect numerous unknown speciesliving on an ichthyofauna of which our understanding, even inEurope, is relatively limited. This study expands the work ongyrodactylid parasitic flatworms living on sand gobies to thecentre of endemism of the hosts, the eastern Mediterranean.Gyrodactylus infecting gobies represents one of the few casesin which taxonomic understanding of monogenean parasiteson a given host clade offers a firm basis. This study leads us toconclude that the endemic character of the Balkans is reflectedin its Gyrodactylus fauna. For many of the host goby speciesstudied here, we report only a singleGyrodactylus species perhost species. It is recommended that more specimens of theBalkan gobies be sampled, and in other seasons, to confirmthis observation. In any case, the present study does notindicate a higher species richness than in their more northerncounterparts, often infected by several species (Table 1). Theabove-mentioned host-switching events, important to parasitediversification in Pomatoschistus , may be less likely in fresh-water sand gobies. Hence, the distribution of the parasitefauna seems driven by host vicariant isolation rather than byadditional environmental factors fuelling diversification in thehighly biodiverse Balkans. Conversely, unique haptoral struc-tures are reported, such as the large size of the marginal hooksickle proper, as compared to its foot, inG. benedeni sp. n.,G.dorlodoti sp. n. and G. meelkopae sp. n., and the kink in themarginal hook sickle of G. douglasadamsi sp. n. This obser-vation, rather than considerations on species richness, rendersthis parasite fauna interesting for subsequent studies on theevolution and biogeography of the sand goby– Gyrodactylussystem. Specifically, molecular analyses are needed toascertain which speciation mechanisms contributed to thebiodiversity of these flatworms in the Balkans, assessingthe occurrence of within-host duplication (possibly fuelledby site preference), host-switching or co-speciationevents.
Acknowledgements We thank D. Mrdak, V. Pešić (University ofMon-tenegro), E. Kalogianni, L. Vardakas, Y. Chatzinikolaou, V. Tachos, D.Kommatas, N. Koutsikos and Y. Kapakos (HCMR) for help with sam-pling; L. Schoofs, L. Arckens, E. Meelkop, J. Nys, J. Aerts and L.Minerva (KU Leuven) for (help with) the use of a ZEISS microscope;B. Geens for help with collecting part of theGyrodactylus data; A. Vidalis(Biodiversity East) for providing Fig. 1; and W.A. Boeger (UniversidadeFederal do Paraná, Brazil), J. Cable (Cardiff University, Wales, UnitedKingdom), A. Pariselle (Institut de Recherche pour le Développement,France), M. Ondračková (Institute of Vertebrate Biology, Academy ofSciences of the Czech Republic) and I. Přikrylová (Masaryk University,Czech Republic) for useful suggestions on monogenean work. M.P.M.Vand T.H. benefited from a PhD and a postdoctoral fellowship, respective-ly, of the Research Council – Flanders (FWO – Vlaanderen). Fieldworkwas partly financed by a travel grant to M.P.M.V. from BeNCoRe(Belgian Network for Coastal Research; ENCORA coordination actioncontract number FP6-2004-Global-3–518120). M.P.M.V. is currentlysupported by ECIP - Centre of excellence, GAČRNo. GBP505/12/G112.
Parasitol Res

References
Agorreta A, San Mauro D, Schliewen U, Van Tassell JL, Kovačić M,Zardoya R, Rüber L (2013) Molecular phylogenetics of Gobioideiand phylogenetic placement of European gobies. Mol PhylogenetEvol 69(3):619–633. doi:10.1016/j.ympev.2013.07.017
Bakke TA, Cable J, Harris PD (2007) The biology of gyrodactylidmonogeneans: the “Russian-doll Killers”. Adv Parasit 64:161–276
BarsonM, Přikrylová I, Vanhove MPM, Huyse T (2010) Parasite hybrid-ization in African Macrogyrodactylus spp. (Monogenea,Platyhelminthes) signals historical host distribution. Parasitology137:1585–1595
Blakeslee AMH, Byers JE, Lesser MP (2008) Solving cryptogenic his-tories using host and parasite molecular genetics: the resolution ofLittorina littorea’s North American origin. Mol Ecol 17:3684–3696
Boeger WA, Kritsky DC (2003) Parasites, fossils and geologic history:historical biogeography of the South American freshwater croakers,Plagioscion spp. (Teleostei, Sciaenidae). Zool Scr 32(1):3–11
Brooks DR, Hoberg EP (2001) Parasite systematics in the 21st century:opportunities and obstacles. Trends Parasitol 17(6):273–275
Brooks DR, McLennan DA (1993) Parascript. Parasites and the languageof evolution. Smithsonian Institution Press, Washington, DC
Choudhury A, Dick TA (2001) Sturgeons (Chondrostei: Acipenseridae)and their metazoan parasites: patterns and processes in historicalbiogeography. J Biogeogr 28(11–12):1411–1439
Criscione CD, Cooper B, Blouin MS (2006) Parasite genotypes identifysource populations of migratory fish more accurately than fishgenotypes. Ecology 87(4):823–828
Daoulas C, Economou AN, Psarras T, Barbieri-Tseliki R (1993)Reproductive strategies and early development of three freshwatergobies. J Fish Biol 42:749–776
de Meeûs T, Michalakis Y, Renaud F (1998) Santa Rosalia revisited: orwhy are there so many kinds of parasites in ‘the garden of earlydelights’? Parasitol Today 14:10–13
Dmitrieva EV, Skidan N (2005) Gyrodactylus leopardinus sp. n.(Monogenea) – a parasite of Pomatoschistus marmoratus(Gobiidae) from the Sea of Azov. Vestn Zool 39(3):17–22
Dunn RR, Harris NC, Colwell RK, Koh LP, Sodhi NS (2009) The sixthmass coextinction: are most endangered species parasites and mu-tualists? P Roy Soc B-Biol Sci 276:3037–3045
Dupont F, Lambert A (1986) Study of communities of MonogeneaDactylogyridae parasites of the Cyprinidae in Lake Mikri Prespa(northern Greece). Description of 3 new species from an endemicBarbus: Barbus cyclolepis prespensis Karaman, 1924. AnnParasitol Hum Comp 61(6):597–616 [in French]
Economidis PS (1995) Endangered freshwater fishes of Greece. BiolConserv 72:201–211
Economou AN, Miller PJ, Economidis PS (2004a) Economidichthyspygmaeus . In: Miller PJ (ed) The freshwater fishes of Europe –Gobiidae 2. AULA-Verlag GmbH, Wiebelsheim, pp 443–458
Economou AN, Miller PJ, Economidis PS (2004b) Economidichthystrichonis . In: Miller PJ (ed) The freshwater fishes of Europe –Gobiidae 2. AULA-Verlag GmbH, Wiebelsheim, pp 459–468
Economou AN, Giakoumi S, Vardakas L, Barbieri R, Stoumboudi M,Zogaris S (2007) The freshwater ichthyofauna of Greece – an updatebased on a hydrographic basin survey. Mediterr Mar Sci 8:91–166
Ergens R (1985) Order Gyrodactylidea Bychowsky, 1937. In: Gusev AV(ed) Key to the parasites of the freshwater fish fauna of the USSR,vol 2. Nauka, Leningrad, pp 269–347
Eschmeyer WN (ed) (2013) Genera, species, references. Catalog ofFishes. http://research.calacademy.org/research/ichthyology/catalog/fishcatmain.asp. Accessed 27 July 2013
Fishelson L (1989) Bisexuality and pedogenesis in gobies (Gobiidae,Teleostei) and other fish, or, why so many little fish in tropical seas.Senck Marit 20(3–4):147–169
García-Vásquez A, Shinn AP, Bron JE (2012) Development of a lightmicroscopy stain for the sclerites of Gyrodactylus von Nordmann,1832 (Monogenea) and related genera. Parasitol Res 110:1639–1648
Geets A, Malmberg G, Ollevier F (1998) Gyrodactylus longidactylus sp.nov., a monogenean from Pomatoschistus lozanoi (de Buen)(Gobiidae) from the North Sea. Syst Parasitol 41:63–70
Gillardin C, VanhoveMPM, Pariselle A, Huyse T, Volckaert FAM (2012)Ancyrocephalidae (Monogenea) of Lake Tanganyika: II.Description of the first Cichlidogyrus spp. parasites fromTropheini fish hosts (Teleostei, Cichlidae). Parasitol Res 110(1):305–313
Harris PD, Shinn AP, Cable J, Bakke TA (2004) Nominal species of thegenus Gyrodactylus von Nordmann 1832 (Monogenea:Gyrodactylidae), with a list of principal host species. Syst Parasitol59:1–27
Harris PD, Shinn AP, Cable J, Bakke TA, Bron JE (2008) GyroDb:gyrodactylid monogeneans on the web. Trends Parasitol 24:109–111
Huyse T, Malmberg G (2004) Molecular and morphological comparisonsbetween Gyrodactylus ostendicus n. sp. (Monogenea:Gyrodactylidae) on Pomatoschistus microps (Krøyer) and G.harengi Malmberg, 1957 on Clupea harengus membras L. SystParasitol 58:105–113
Huyse T, Volckaert FAM (2002) Identification of a host-associated spe-cies complex using molecular and morphometric analyses, with thedescription of Gyrodactylus rugiensoides n. sp. (Gyrodactylidae,Monogenea). Int J Parasitol 32:907–919
Huyse T, Volckaert FAM (2005) Comparing host and parasite phyloge-nies:Gyrodactylus flatworms jumping from goby to goby. Syst Biol54(5):710–718
Huyse T, Audenaert V, Volckaert FAM (2003) Speciation and host–parasite relationships in the parasite genus Gyrodactylus(Monogenea, Platyhelminthes) infecting gobies of the genusPomatoschistus (Gobiidae, Teleostei). Int J Parasitol 33(14):1679–1689
Huyse T, Malmberg G, Volckaert FAM (2004a) Four new species ofGyrodactylus von Nordmann, 1832 (Monogenea, Gyrodactylidae)on gobiid fishes: combined DNA and morphological analyses. SystParasitol 59:103–120
Huyse T, Van Houdt J, Volckaert FAM (2004b) Paleoclimatic history andvicariant speciation in the “sand goby” group (Gobiidae, Teleostei).Mol Phylogenet Evol 32:324–336
Huyse T, Pampoulie C, Audenaert V, Volckaert FAM (2006) First reportof Gyrodactylus spp. (Platyhelminthes: Monogenea) in the westernMediterranean sea: molecular and morphological descriptions. JParasitol 92(4):682–690
Ishimatsu A, Gonzales TT (2011) Mudskippers: front runners in themodern invasion of land. In: Patzner RA, Van Tassell JL, KovačićM, Kapoor BG (eds) The biology of Gobies. Science Publishers,Enfield, pp 609–638
Justine J-L, Beveridge I, Boxshall GA, Bray RA, Miller TL, Moravec F,Trilles J-P, Whittington ID (2012) An annotated list of fish parasites(Isopoda, Copepoda, Monogenea, Digenea, Cestoda, Nematoda)collected from Snappers and Bream (Lutjanidae, Nemipteridae,Caesionidae) in New Caledonia confirms high parasite biodiversityon coral reef fish. Aquat Biosyst 8:22
Koh LP, Dunn RR, Sodhi NS, Colwell RK, Proctor HC, Smith VS (2004)Species coextinctions and the biodiversity crisis. Science 305:1632–1634
Kottelat M, Freyhof J (2007) Handbook of European freshwater fishes.Kottelat, Cornol and Freyhof, Berlin
Kovačić M, Patzner RA (2011) North-Eastern Atlantic andMediterranean gobies. In: Patzner RA, Van Tassell JL, Kovačić M,Kapoor BG (eds) The biology of gobies. Science Publishers,Enfield, pp 177–206
Parasitol Res

Kryštufek B, Reed JM (2004) Pattern and process in Balkan biodiversity– an overview. In: Griffiths HI, Kryštufek B, Reed JM (eds) Balkanbiodiversity: pattern and process in the European hotspot. KluwerAcademic Publishers, Dordrecht, pp 1–8
Kuris AM, Hechinger RF, Shaw JC, Whitney KL, Aguirre-Macedo L,Boch CA, Dobson AP, Dunham EJ, Fredensborg BL, Huspeni TC,Lorda J, Mababa L, Mancini FT, Mora AB, Pickering M, TalhoukNL, Torchin ME, Lafferty KD (2008) Ecosystem energetic implica-tions of parasite and free-living biomass in three estuaries. Nature454:515–518
Kvach Y, Skóra KE (2007) Metazoa parasites of the invasive round gobyApollonia melanostoma (Neogobius melanostomus) (Pallas)(Gobiidae: Osteichthyes) in the Gulf of Gdańsk, Baltic Sea,Poland: a comparison with the Black Sea. Parasitol Res 100:767–774
Kvach Y, Winkler HM (2011) The colonization of the invasive roundgoby Neogobius melanostomus by parasites in new localities in thesouthwestern Baltic Sea. Parasitol Res 109:769–780
Larmuseau MHD, Huyse T, Vancampenhout K, Van Houdt JKJ,Volckaert FAM (2010) High molecular diversity in the rhodopsingene in closely related goby fishes: a role for visual pigments inadaptive speciation? Mol Phylogenet Evol 55:689–698
Larmuseau MHD, Vanhove MPM, Huyse T, Volckaert FAM, Decorte R(2011) Signature of selection on the rhodopsin gene in the marineradiation of American seven-spined gobies (Gobiidae,Gobiosomatini). J Evol Biol 24(7):1618–1625
Littlewood DTJ (2011) Systematics as a cornerstone of parasitology:overview and preface. Parasitology 138:1633–1637
Longshaw M, Pursglove M, Shinn AP (2003) Gyrodactylusquadratidigitus n. sp. (Monogenea: Gyrodactylidae), a parasite ofthe leopard-spotted goby Thorogobius ephippiatus (Lowe) from thesouth-western coast of the UK. Syst Parasitol 55:151–157
Lundberg JG, Kottelat M, Smith GR, Stiassny M, Gill T (2000) So manyfishes, so little time: an overview of recent ichthyological discover-ies in fresh waters. Ann Missouri Bot Gard 87:26–62
MacKenzie K (2002) Parasites as biological tags in population studies ofmarine organisms: an update. Parasitology 124:S153–S163
Malavasi S, Gkenas C, Leonardos I, Torricelli P, McLennan DA (2012)The phylogeny of a reduced ‘sand goby’ group based on behaviour-al and life history characters. Zool J Linn Soc-London 165:916–924
Malmberg G (1957) On the occurrence of Gyrodactylus on Swedishfishes. Skr Söd Sver Fiskför Asskr 1956:19–76 [in Swedish]
Marcogliese DJ (2004) Parasites: small players with crucial roles in theecological theater. EcoHealth 1:151–164
McDowall RM (2000) Biogeography of the southern cool-temperategalaxioid fishes: evidence from metazoan macroparasite faunas. JBiogeogr 27(5):1221–1229
Mierzejewska K, Martyniak A, Kakareko T, Dzika E, Stańczak K, HliwaP (2011) Gyrodactylus proterorhini Ergens, 1967 (Monogenoidea,Gyrodactylidae) in gobiids from the Vistula River – the first recordof the parasite in Poland. Parasitol Res 108:1147–1151
Miller PJ (1979) Adaptiveness and implications of small size in teleosts.Sym Zool S 44:263–306
Miller PJ (1990) The endurance of endemism – the Mediterranean fresh-water gobies and their prospects for survival. J Fish Biol 37:145–156
Miller PJ (2004) Knipowitschia croatica. In: Miller PJ (ed) The freshwa-ter fishes of Europe – Gobiidae 2. AULA-Verlag GmbH,Wiebelsheim, pp 365–369
Miller PJ, Economidis PS (2004) Knipowitschia thessala . In: Miller PJ(ed) The freshwater fishes of Europe – Gobiidae 2. AULA-VerlagGmbH, Wiebelsheim, pp 423–429
Miller PJ, Madurell T, Economou AN (2004) Knipowitschia milleri. In:Miller PJ (ed) The freshwater fishes of Europe –Gobiidae 2. AULA-Verlag GmbH, Wiebelsheim, pp 399–404
Muterezi Bukinga F, Vanhove MPM, Van Steenberge M, Pariselle A
(2012) Ancyrocephalidae (Monogenea) of Lake Tanganyika: III.Cichlidogyrus infecting the world’s biggest cichlid and the non-endemic tribes Haplochromini, Oreochromini and Tylochromini(Teleostei, Cichlidae). Parasitol Res 111(5):2049–2061
Nieberding C,Morand S, Libois R,Michaux JR (2004) A parasite revealscryptic phylogeographic history of its host. P Roy Soc B-Biol Sci271(1557):2559–2568
Ondračková M, Dávidová M, Blažek R, Gelnar M, Jurajda P (2009) Theinteraction between an introduced fish host and local parasite fauna:Neogobius kessleri in the middle Danube River. Parasitol Res 105:201–208
Osmanov SO (1971) Parasites of Uzbekistan’s fishes. FAN, Tashkent [InRussian]
Palm HW, Rückert S (2009) A new approach to visualize ecosystemhealth by using parasites. Parasitol Res 105:539–553
Pariselle A, Morand S, Deveney M, Pouyaud L (2003) Parasite speciesrichness of closely related hosts: historical scenario and “genetic”hypothesis. In: Combes C, Jourdan J (eds) Hommage à LouisEuzet—Taxonomie, écologie et évolution des métazoaires parasites.Taxonomy, ecology and evolution of metazoan parasites. PressesUniversitaires de Perpignan, Perpignan, pp 147–166
Pariselle A, Boeger WA, Snoeks J, Bilong Bilong CF, Morand S,Vanhove MPM (2011) The monogenean parasite fauna of cichlids:a potential tool for host biogeography. Int J Evol Biol 2011:471480
Paugy D, Guégan JF, Agnèse JF (1990) Three simultaneous and inde-pendent approaches to the characterization of a new species ofLabeo (Teleostei, Cyprinidae) from West Africa. Can J Zool 68:1124–1131
Poulin R (2002) The evolution of monogenean diversity. Int J Parasitol32:245–254
Přikrylová I, Blažek R, Vanhove MPM (2012) An overview of theGyrodactylus (Monogenea: Gyrodactylidae) species parasitizingAfrican catfishes, and their morphological and molecular diversity.Parasitol Res 110:1185–1200
Pugachev ON, Gerasev PI, Gussev AV, Ergens R, Khotenowsky I (2009)Guide to Monogenoidea of freshwater fish of Palaearctic and AmurRegions. Ledizione-Ledi Publishing, Milan
Raeymaekers JAM, Huyse T, Maelfait H, Hellemans B, Volckaert FAM(2008) Community structure, population structure and topographi-cal specialisation ofGyrodactylus (Monogenea) ectoparasites livingon sympatric stickleback species. Folia Parasit 55:187–196
Rüber L, Van Tassell JL, Zardoya R (2003) Rapid speciation and ecolog-ical divergence in the American seven-spined gobies (Gobiidae,Gobiosomatini) inferred from a molecular phylogeny. Evolution57(7):1584–1598
Ryan PA (1991) The success of the Gobiidae in tropical Pacific insularstreams. New Zeal J Zool 18:25–30
Shinn AP, Hansen H, Olstad K, Bachmann L, Bakke TA (2004) The useof morphometric characters to discriminate specimens of laboratory-reared andwild populations ofGyrodactylus salaris andG. thymalli(Monogenea). Folia Parasit 51:239–252
Shinn AP, Harris PD, Cable J, Bakke TA, Paladini G, Bron JE (eds)(2010) GyroDb. http://www.gyrodb.net, version (06/2010)
Stork NE, Lyal CHC (1993) Extinction or ‘co-extinction’ rates? Nature366:307
Sures B (2004) Environmental parasitology: relevancy of parasites inmonitoring environmental pollution. Trends Parasitol 20(4):170–177
Taylor MS, Hellberg ME (2005) Marine radiations at small geographicscales: speciation in neotropical reef gobies (Elacatinus) . Evolution59:374–385
Thacker CE (2013) Phylogenetic placement of the European sand gobiesin Gobionellidae and characterization of gobionellid lineages(Gobiiformes: Gobioidei). Zootaxa 3619(3):369–382
Thacker CE, Roje DM (2011) Phylogeny of Gobiidae and identificationof gobiid lineages. Syst Biodivers 9(4):329–347
Parasitol Res

Thompson JN (1999) The evolution of species interactions. Science 284:2116–2118
Timm RM, Clauson BL (1987) Coevolution: Mammalia. In: Parker SP(ed) (1988) McGraw-Hill yearbook of science & technology.McGraw-Hill, New York, pp 212–214
Vanhove MPM (2012) Species flocks and parasite evolution. Towards aco-phylogenetic analysis of monogenean flatworms of cichlids andgobies. PhD thesis, University of Leuven
VanhoveMPM, KovačićM, Koutsikos NE, Zogaris S, Vardakas LE, HuyseT, Economou AN (2011a) First record of a landlocked population ofmarine Millerigobius macrocephalus (Perciformes: Gobiidae): obser-vations from a unique spring-fed karstic lake (Lake Vouliagmeni,Greece) and phylogenetic positioning. Zool Anz 250(3):195–204
Vanhove MPM, Snoeks J, Volckaert FAM, Huyse T (2011b) First de-scription of monogenean parasites in Lake Tanganyika: the cichlid
Simochromis diagramma (Teleostei, Cichlidae) harbours a highdiversity of Gyrodactylus species (Platyhelminthes, Monogenea).Parasitology 138(3):364–380
Vanhove MPM, Economou AN, Zogaris S, Larmuseau MHD, GiakoumiS, Kalogianni E, Volckaert FAM, Huyse T (2012) Phylogenetics andbiogeography of the Balkan "sand gobies" (Teleostei, Gobiidae):vulnerable species in need of taxonomic revision. Biol J Linn Soc105(1):73–91
VanhoveMPM, Van SteenbergeM, Dessein S, Volckaert FAM, Snoeks J,Huyse T, Pariselle A (2013) Biogeographical implications ofZambezian Cichlidogyrus species (Platyhelminthes: Monogenea:Ancyrocephalidae) parasitizing Congolian cichlids. Zootaxa3608(5):398–400
Windsor DA (1998) Most of the species on Earth are parasites. Int JParasitol 28:1939–1941
Parasitol Res
Copyright © 2022 FDOKUMEN




















