
Infection level of monogenean gill parasite, Diplozoon kashmirensis (Monogenea, Polyopisthocotylea)...
10
This article appeared in a journal published by Elsevier. The attached copy is furnished to the author for internal non-commercial research and education use, including for instruction at the authors institution and sharing with colleagues. Other uses, including reproduction and distribution, or selling or licensing copies, or posting to personal, institutional or third party websites are prohibited. In most cases authors are permitted to post their version of the article (e.g. in Word or Tex form) to their personal website or institutional repository. Authors requiring further information regarding Elsevier’s archiving and manuscript policies are encouraged to visit: http://www.elsevier.com/copyright
-
Upload
independent -
Category
Documents
-
view
5 -
download
0
Transcript of Infection level of monogenean gill parasite, Diplozoon kashmirensis (Monogenea, Polyopisthocotylea)...

This article appeared in a journal published by Elsevier. The attachedcopy is furnished to the author for internal non-commercial researchand education use, including for instruction at the authors institution
and sharing with colleagues.
Other uses, including reproduction and distribution, or selling orlicensing copies, or posting to personal, institutional or third party
websites are prohibited.
In most cases authors are permitted to post their version of thearticle (e.g. in Word or Tex form) to their personal website orinstitutional repository. Authors requiring further information
regarding Elsevier’s archiving and manuscript policies areencouraged to visit:
http://www.elsevier.com/copyright

Author's personal copy
Veterinary Parasitology 189 (2012) 218– 226
Contents lists available at SciVerse ScienceDirect
Veterinary Parasitology
jou rn al h om epa ge: www.elsev ier .com/ locate /vetpar
Infection level of monogenean gill parasite, Diplozoon kashmirensis(Monogenea, Polyopisthocotylea) in the Crucian Carp, Carassiuscarassius from lake ecosystems of an altered water quality: Whatfactors do have an impact on the Diplozoon infection?
U.R. Zargara,∗, M.Z. Chishti a, A.R. Yousufa, A. Fayazb
a Centre of Research for Development, University of Kashmir, Indiab Department of Zoology, University of Kashmir, India
a r t i c l e i n f o
Article history:Received 16 February 2012Received in revised form 20 April 2012Accepted 21 April 2012
Keywords:MonogeneanDiplozoon kashmirensisCarassius carassiusKashmir HimalayasSynergistic effectMicrohabitatCondition factorHypertrophic habitats
a b s t r a c t
Monogeneans are of great interest to the ecologists because of their simple life cycle andare considered as one of the important and sensitive parasites to any changes in waterquality. The monogenean gill parasite, Diplozoon kashmirensis of the Carassius carassius wasexamined with the aim to evaluate the infection level and the factors influencing the infec-tion. Our results showed that highest prevalence (34.22%) of D. kashmirensis was in the lakehaving high trophic status and least prevalence (10.90%) in the lake having least trophicstatus. Infection levels were significantly higher at the basins/sites having deterioratedwater quality in comparison to the basins/sites having better water quality. The combinedeffect of pollutants and eutrophication showed decrease in intensity of D. kashmirensis inone of the polluted/hypertrophied site in the hypertrophic lake (Anchar Lake), while assynergistic effect (i.e. increase) on prevalence and intensity was observed in one of the pol-luted/hypertrophied site in the eutrophic lake (Dal Lake). Sex was not an important factorinfluencing the D. kashmirensis in three lakes. The study showed that condition factor wassignificantly higher in the hypertrophic lake and in an uninfected fish. In addition micro-habitat influenced the Diplozoon infection. Most preferable site for the D. kashmirensiswas middle sector of the gill. Furthermore, the gill parasite showed significantly posi-tive correlation (rp = 0.6, P < 0.05) with water temperature in the hypertrophic lake, thusshowing the impact of water temperature on this parasite. It was concluded that intra lakedifferences, season, condition factor, microhabitat and water temperature influenced theinfection of D. kashmirensis. It seems from the data that eutrophic and hypertrophic habitatswere favourable for the D. kashmirensis. The D. kashmirensis showed both antagonistic andsynergistic response to the combined effect of pollution and eutrophication.
© 2012 Elsevier B.V. All rights reserved.
∗ Corresponding author at: Centre of Research for Development(CORD), University of Kashmir (NAAC Accredited Grade ‘A’ University),Srinagar 190006, Kashmir, India. Tel.: +91 9797283587.
E-mail address: [email protected] (U.R. Zargar).
1. Introduction
Monogenean gill parasites are an important class oftrematoda of Phylum Platyhelminths. Majority of themare ectoparasites and have direct lifecycle. Monogeneanparasites are seen both in fresh water and marine fishes.In most of cases, monogenean parasites are host-specific,although some may infect several hosts from different fam-ilies (Poulin, 1992). Monogeneans mostly infect gills of fish
0304-4017/$ – see front matter © 2012 Elsevier B.V. All rights reserved.http://dx.doi.org/10.1016/j.vetpar.2012.04.029

Author's personal copy
U.R. Zargar et al. / Veterinary Parasitology 189 (2012) 218– 226 219
and are found on gill filaments, gill rakers or the lateralsurfaces of gill arches (Bychowsky, 1961).
Three species of the Genus Diplozoon have beenreported from different fish species in the KashmirHimalayas and taxonomic status of these parasites hasbeen confirmed (Ahmed and Chishti, 2002). The speciesinclude Diplozoon kashmirensis, Diplozoon guptai and Diplo-zoon aegeptensis. In the past, researchers have studied someof the ecological aspects of helminth parasites includingDiplozoon spp. in this region and have stated that temper-ature, pH and O2 are the important factors influencing thehelminth parasites (Chishti et al., 2000; Zargar et al., 2012).However, previous studies have not used holistic approachto ascertain the effect of various factors which influenceinfection levels of Diplozoon spp. in the Kashmir Himalayas.In order to unravel the effect of various factors like waterquality, seasonality, sex, condition factor, microhabitat andmacrohabitat on the Diplozoon infection, we carried out aninvestigation in three valley lakes on exotic fish, Carassiuscarassius.
It was predicted that D. kashmirensis may show inter andintra lake variation due to altered water quality. It was alsopredicted that season, sex and microhabitat could affect themonogenean infection. Furthermore, it was predicted thatthe combined effect of pollutants and nutrient enrichmentmay affect Diplozoon infection either antagonistically orsynergistically depending upon the type of environmentand adaptability of the parasite.
2. Materials and methods
2.1. Study sites
The valley of Kashmir is situated in the mid-Himalayasin the North West and South-East direction within thecoordinates 33◦01′–35◦00′N latitude and 73◦48′–75◦30′Elongitude at an altitude of ≥1500 m above sea level. Thestudy was carried out in three valley lakes viz., the AncharLake (34◦01′N–74◦02′E), the Dal Lake (34◦07′N–74◦52′E)and the Manasbal Lake (34◦15′N–74◦40′E). The Anchar Lakeis situated in the North West of Srinagar city at an eleva-tion of 1583 m. The lake is dominated throughout its basinwith submerged as well as free floating macrophytes. Inthe present study three sites were selected (Anchar Ghat,Anchar Centre and Anchar Inlet). The Dal Lake is an urbanlake which lies in the east of Srinagar city, at the foot ofZabarwan Hills, situated at an elevation of 1584 m abovethe sea level. Four sites were selected in Dal Lake, one eachin Hazratbal Basin, Gagribal Basin, Nagin and Bud Dal. Man-asbal Lake is a rural lake situated at a distance of 32 km fromthe Srinagar city. Its length and breadth is about 3.2 kmand 1 km, respectively. The lake is situated at an altitudeof 1584 m above sea level. Four sites were selected in thislake, one at the centre of the lake and other three sites werein the peripheral zone of the lake.
2.2. Physico-chemical characteristics of the water
The physico-chemical characteristics of water wereanalyzed as per the methods described by the Council forScientific and Industrial Research (CSIR, 1974), Mackereth
et al. (1978) and the American Public Health Association(APHA, 1998).
2.3. Host and parasite collection
The present study was carried out between September2010 and August 2011. In total, 321 (122 specimens inthe Anchar Lake, 135 specimens in the Dal Lake and 64 inthe Manasbal Lake) individuals of C. carassius were exam-ined in three lakes. Fish were caught by local fishermenusing nets and were killed by pithing. Further investiga-tion was carried out in the laboratory, where total length(TL), standard length (SL), and body weight (BW) andsex for each fish was determined. Biometrical indices likelength–weight relationship was calculated by Le Cren’s(1951) method (W = aLb) and condition factor by Fulton’sIndex (K = W/L3 × 105).
Fish collected from nearby lakes were immediately dis-sected and some fishes were kept alive in aquarium forshort period of time (up to half an hour) so that they remainfresh for parasitological investigation. Gills were carefullyrecovered soon after the death of fish and kept in a Petri dishcontaining normal saline (0.85% NaCl). In case of distantlakes, live fish samples were carried in buckets with waterand death fish were transported in containers containingice packs and were immediately analyzed after reachingthe laboratory (within 12 h). With the aid of magnifyingglasses the gills were searched for parasites. The attachedparasites were detached from the gills by carefully teasingthe filament with the help of brush. All the informationcollected was maintained in a proper way. For studyingmicrodistribution of Diplozoon parasites, the methodologyof Turgut et al. (2006) and Geets et al. (1997) were adopted.Gill arches were numbered 1–4 from the anterior gill archbelow the operculum to the posterior. The sites of parasiteson left and right side were recorded. In addition we dividedgill into 6 sectors (following Turgut et al., 2006).
2.4. Experimental design
We selected three different lakes viz., Anchar, Dal, andManasbal for inter lake variation of Diplozoon infection.Anchar and Dal lakes were selected for intra lake varia-tion in Diplozoon infection. We did not included the intralake differences in the Manasbal Lake because differentstudy sites showed very small differences and also becausefish samples were not available at all sites for comparativeparasitological analysis. Two sites, one each in the Anchar(Anchar Ghat) and the Dal (Gagribal) were also consideredas polluted sites because of high level of contamination inaddition to eutrophic condition.
2.5. Data analysis
The level of Diplozoon infection was quantified usingprevalence, mean intensity and mean abundance as perMargolis et al. (1982) and Bush et al. (1997).
Pearson’s correlation was used to assess associationsbetween different physico-chemical features and Diplo-zoon infection. Correlation and other statistical analysis ofdata were carried out with SPSS 11.5 (SPSS Inc., Chicago, IL,
Author's personal copy
220 U.R. Zargar et al. / Veterinary Parasitology 189 (2012) 218– 226
Table 1The range of physico-chemical characteristics of water in three Lakes.
Parameter Anchar Lake Dal Lake Manasbal Lake
Max Min Max Min Max Min
Water temp. (◦C) 30.0 6.0 30.0 6.5 29.3 6.0Air temp. (◦C) 34.0 9.0 31.2 6.8 31.6 6.9Depth (m) 1.7 0.6 2.6 2.0 7.4 6.1Transparency (m) 0.94 0.4 2.0 1.5 3.6 2.9pH 8.4 6.5 8.4 7.4 8.5 7.2Conductivity (�S cm−1) 265.0 212.2 280.0 243.0 200.0 111.0Dissolved oxygen (mg l−1) 4.4 1.15 6.3 4.9 6.9 5.6Free CO2 (mg l−1) 34.2 18.9 3.5 1.5 1.2 0.5Alkalinity (mg l−1) 272.3 207.5 164.2 129.3 192.0 131.6Chloride (mg l−1) 42.4 17.6 20.3 9.9 24.3 10.5Ammonia-N (�g l−1) 560.0 374.0 190.3 149.2 194.0 169.0Nitrate-N (�g l−1) 253.7 215.4 330.1 307.3 197.2 140.2Total phosphate (�g l−1) 262.5 160.3 202.0 159.4 70.2 45.2Total hardness (mg l−1) 255.2 125.2 170.2 140.0 130.2 97.2
Table 2Intra lake variation of physico-chemical (±SD) features in Dal and Anchar lakes.
Lake parameter Dal Lake Anchar Lake
Gagribal Hazratbal Nishat Nigeen Anchar Ghat Anchar Centre Anchar Inlet
Water temp. (◦C) 17.6 ± 8.6 17.4 ± 8.4 17.0 ± 7.6 16.9 ± 8.4 17.9 ± 3.3 17.5 ± 7.49 16.9 ± 5.7Depth (m) 1.16 ± 0.84 2.23 ± 0.21 2.43 ± 0.15 2.45 ± 0.05 0.75 ± 0.13 1.59 ± 0.19 1.3 ± 0.36Transparency (m) 0.649 ± 0.35 1.5 ± 0.0 1.66 ± 0.15 1.93 ± 0.05 0.50 ± 0.0 0.6 ± 0.1 0.76 ± 0.24pH 5.52 ± 4.23 8.16 ± 0.25 7.66 ± 0.21 7.53 ± 0.11 7.02 ± 0.55 7.06 ± 0.35 7.52 ± 0.66Conductivity (�S) 162.44 ± 12.4 270 ± 10.0 259 ± 3.60 246.33 ± 3.05 254.5 ± 9.53 213.63 ± 2.06 236.25 ± 11.08Dissolved oxygen (mg l−1) 2.81 ± 2.31 5.1 ± 0.20 5.8 ± 0.45 6.2 ± 0.1 2.57 ± 0.63 2.05 ± 0.83 4.2 ± 0.14Free CO2 (mg l−1) 15.81 ± 12.20 3.33 ± 0.15 2.9 ± 0.43 2.13 ± 0.15 29.82 ± 4.08 20 ± 1.57 22.52 ± 1.73Alkalinity (mg l−1) 167.18 ± 8.9 158.2 ± 7.21 160.5 ± 3.97 143.1 ± 12.21 267.05 ± 5.56 233.45 ± 14.6 218.56 ± 11.36Chloride (mg l−1) 16.15 ± 11.11 19.03 ± 1.35 16.6 ± 0.81 15.26 ± 1.30 33.42 ± 9.09 22.55 ± 3.32 19.2 ± 1.65Ammonia-N (�g l−1) 326.86 ± 7.2 184.0 ± 6.97 169.2 ± 7.96 151.63 ± 3.38 540.0 ± 27.08 492.77 ± 27.8 424.6 ± 82.16Nitrate-N (�g l−1) 320.93 ± 11.88 152.54 ± 12.8 130.3 ± 5.89 120.1 ± 0.80 244.62 ± 9.03 240.83 ± 10.10 221.52 ± 6.11Total phosphate (�g l−1) 190.83 ± 10.77 170.90 ± 13.4 175.93 ± 4.28 150.13 ± 5.61 244.0 ± 19.68 257.12 ± 5.49 216.83 ± 49.22Total hardness (mg l−1) 165.63 ± 4.11 156.96 ± 8.4 159.66 ± 4.53 148.83 ± 7.68 188.72 ± 51.73 166.12 ± 55.4 130.43 ± 49.22
USA). One-way analysis of variance (ANOVA) was used tosee the significant differences in intensity and abundancewithin different sites in two lakes and between three lakes.In addition, One-way analysis of variance (ANOVA) wasused to evaluate seasonal differences in mean intensity intwo lakes and multiple comparisons (Tukey test, LSD) werealso carried out. Chi Square analysis was used to demon-strate the effect of month and season on the prevalence.Furthermore, Chi Square analysis was also used for intralake variation in prevalence. The effect of microhabitat wasanalyzed by using ANOVA and Kruskal–Wallis and to testthe differences at different levels.
3. Results
The range of various physico-chemical features, calcu-lated from the three lakes is presented in Table 1. These
lakes have been studies from time to time by different lim-nologists with the aim to categorize these lakes accordingto approved international standards (OECD, 1982; APHA,1998). The present study showed that all the three lakeshave reached to the level of eutrophic condition, but thelevel of trophic status varies. The Anchar Lake was the mosteutrophic whereas, the Manasbal Lake was the least nutri-ent enriched. This study is also supported by the findingsof Pandit and Yousuf (2002). The marked differences inwater quality of three lakes clearly depict the influence ofanthropogenic stress on lakes.
Intra lake variation was observed in physico-chemicalfeatures in different basins/study sites in Dal and Ancharlakes (Table 2). The intra lake variation in physicochemical features in the Dal Lake was in the orderof: Gagribal > Hazratbal Basin > Nishat > Nigeen and in theAnchar Lake it was in the order of: Anchar Ghat > Anchar
Table 3Intra lake variation of Diplozoon infection (±SD) in two lakes.
Dal Lake (N = 135) Anchar Lake (N = 122)
Gagribal (N = 30) Hazratbal basin (N = 35) Bud Dal (N = 40) Nigeen (N = 30) Anchar Ghat(N = 50)
Anchar Centre(N = 40)
Anchar Inlet(N = 32)
Prevalence 38.46 21.9 12.73 12.16 50.2 36.4 10.03MI 3.38 ± 1.46 3.53 ± 1.11 1.87 ± 0.71 1.06 ± 0.11 2.9 ± 1.63 4.18 ± 1.84 1.17 ± 0.15MA 2.29 ± 0.83 1.54 ± 0.83 0.32 ± 0.25 0.19 ± 0.18 1.36 ± 0.48 1.43 ± 0.61 0.09 ± 0.02

Author's personal copy
U.R. Zargar et al. / Veterinary Parasitology 189 (2012) 218– 226 221
Table 4Multiple comparison tests (Tukey HSD and LSD) in mean intensity (MI) and mean abundance (MA) between different study sites in Anchar and Dal lakes.
Tukey HSD P-value LSD P-value
Anchar LakeMI
Anchar Ghat vs. Anchar Centre 0.52 Anchar Ghat vs. Anchar Centre 0.28Anchar Ghat vs. Anchar Inlet 0.18 Anchar Ghat vs. Anchar Inlet 0.08Anchar Centre vs. Anchar Inlet 0.04* Anchar Centre vs. Anchar Inlet 0.17
MAAnchar Ghat vs. Anchar Centre 0.97 Anchar Ghat vs. Anchar Centre 0.82Anchar Ghat vs. Anchar Inlet 0.02* Anchar Ghat vs. Anchar Inlet 0.009*
Anchar Centre vs. Anchar Inlet 0.01* Anchar Centre vs. Anchar Inlet 0.006*
Dal LakeMI
Hazratbal Basin vs. Gagribal 0.99 Hazratbal Basin vs. Gagribal 0.85Hazratbal Basin vs. Bud Dal 0.24 Hazratbal Basin vs. Bud Dal 0.73Hazratbal Basin vs. Nigeen 0.06a Hazratbal Basin vs. Nigeen 0.02*
Bud Dal vs. Gagribal 0.31 Bud Dal vs. Gagribal 0.09Nigeen vs. Gagribal 0.08 Nigeen vs. Gagribal 0.02*
Nigeen vs. Bud Dal 0.75 Nigeen vs. Bud Dal 0.35MA
Hazratbal Basin vs. Gagribal 0.47 Hazratbal Basin vs. Gagribal 0.17Hazratbal Basin vs. Bud Dal 0.147 Hazratbal Basin vs. Bud Dal 0.04*
Hazratbal Basin vs. Nigeen 0.101 Hazratbal Basin vs. Nigeen 0.03*
Bud Dal vs. Gagribal 0.02* Bud Dal vs. Gagribal 0.004*
Nigeen vs. Gagribal 0.01* Nigeen vs. Gagribal 0.003*
Nigeen vs. Bud Dal 0.99 Nigeen vs. Bud Dal 0.8
a Tendency to differ.* P ≤ 0.05.
Centre > Anchar Inlet. The water quality of Intra Lake siteswas categorized as per APHA (1998) and OECD (1982).
3.1. Intra lake variation of D. kashmirensis infection
The results showed no significant intra lake variationin prevalence in the Dal Lake (Chi Square = 5.6; P = 0.22),while as significant differences were observed betweenstudy sites in Anchar Lake (Chi Square = 14.7; P = 0.002).The prevalence was significantly higher at highly pol-luted/hypereutrophied site (Prevalence = 50.2; P < 0.05) incomparison to eutrophic site (Prevalence = 10.03; P < 0.05)in the Anchar Lake (Table 3).
Mean intensity and mean abundance showed signifi-cant differences between different study sites in the DalLake (MI: F = 8.16; P = 0.008, MA: F = 4.38; P = 0.04) and inthe Anchar Lake (MI: F = 4.5; P = 0.04, MA: F = 8.04; P = 0.01).Interestingly, MI and MA were significantly (P < 0.05) lessat polluted site (Anchar Ghat) in comparison to eutrophicsite (Anchar Centre) in the Anchar Lake. The multiple com-parison tests also showed significant differences betweendifferent basins in the Dal and Anchar lakes (Table 4).In comparison to the Anchar Lake, polluted/hypertrophicsite in Dal Lake showed significantly high prevalenceand mean intensity of the D. kashmirensis (P = 38.46%,MI = 3.38 ± 1.46).
3.2. Inter lake variation of D. kashmirensis infection
Mean prevalence of monogenean gill infection inthree lakes was 22.24%. The comparative analysis showedhighest Diplozoon infestation in the water body whichhas reached hypertrophic level (Anchar Lake: Preva-lence = 34.22%), while as it was least in the lake with least
eutrophic condition (Manasbal Lake: Prevalence = 10.9%).Similarly, mean intensity and mean abundance showed thesame trend (Table 5).
A comparison of mean intensity and mean abundance inthree lakes showed insignificant relationship (MI: F = 0.034,P > 0.05; MA: F = 0.034, P > 0.05).
3.3. Effect of month and season on the infection
The monthly prevalence showed insignificant dif-ferences (P > 0.05), whereas seasons showed significantdifferences in prevalence (Table 6; P < 0.05). Clear seasonaltrend was observed in Dal and Anchar lakes with maxi-mum infection level during summer months and least inwinter months (Table 6). In Anchar Lake, however, infec-tion level showed abrupt increase from winter onwards.The prevalence among different seasons when comparedstatistically revealed significant differences: spring vs.summer (P < 0.05), autumn vs. summer (P < 0.05), wintervs. summer (P < 0.05). The seasons showed significant dif-ferences in mean intensity (F = 4.3; P = 0.05) and meanabundance (F = 11.7; P = 0.04) in Dal Lake while as insignif-icant difference was observed in mean intensity (F = 3.6;P = 0.07) and significant differences in mean abundance(F = 4.8; P = 0.04) in Anchar Lake. Multiple comparisons inMI and MA showed significant differences among differentseasons in the Dal Lake (Table 7).
3.4. Influence of sex and condition factor on the level ofinfection
In all three lakes, females showed insignificantly highDiplozoon infection in comparison to males (P > 0.05;Table 8). Condition factor showed significant differences

Author's personal copy
222 U.R. Zargar et al. / Veterinary Parasitology 189 (2012) 218– 226
Table 5Inter lake variation of Diplozoon infection (±SD).
Name of the parasite Parameter Anchar Dal Manasbal
D. kashmirensis Sample size 122 135 64Prevalence 34.22 21.6 10.9MI 2.86 ± 1.8 2.70 ± 1.6 2.7 ± 0.88MA 1.04 ± 0.74 0.66 ± 0.8 0.29 ± 0.16
Table 6Seasonal variation of infection indices in two lakes.
Infection parameter Anchar Lake Dal Lake
Autumn Winter Spring Summer Autumn Winter Spring Summer
Prevalence 10.03 28.53333 45.36667 62.35 20.5 12.76667 16.4 44.35MI 1 ± 0 4.76 ± 1.56 3.08 ± 1.56 2.5 ± 0.14 1.16 ± 0.28 2.6 ± 1.15 2.83 ± 1.75 4.9 ± 0.14MA 0.09 ± 0.09 1.35 ± 0.52 1.34 ± 0.79 1.55 ± 0.07 0.26 ± 0.06 0.31 ± 0.31 0.42 ± 0.32 2.15 ± 0.91
Table 7Multiple comparison tests (Tukey HSD and LSD) in mean intensity (MI)and mean abundance (MA) between different seasons in Anchar and Dallakes.
Anchar Lake Dal Lake
Tukey HSD P-value Tukey HSD P-value
MIAutumn vs. Winter 0.053 Autumn vs. Summer 0.03*
Winter vs. Spring 0.34 Winter vs. Summer 0.22Summer vs. Summer 0.66 Summer vs. Spring 0.27
MIAutumn vs. Winter 0.07 Autumn vs. Summer 0.005*
Winter vs. Spring 0.07 Winter vs. Summer 0.006*
Summer vs. Summer 0.06 Summer vs. Spring 0.008*
* P ≤ 0.05.
between three lakes (P = 0.00) with higher condition fac-tor in the hypertrophic lake (Fig. 1). The condition factorwas significantly higher in uninfected fish in comparisonto infected fish (P = 0.015, Fig. 2).
3.5. Effect of microhabitat on the infection level
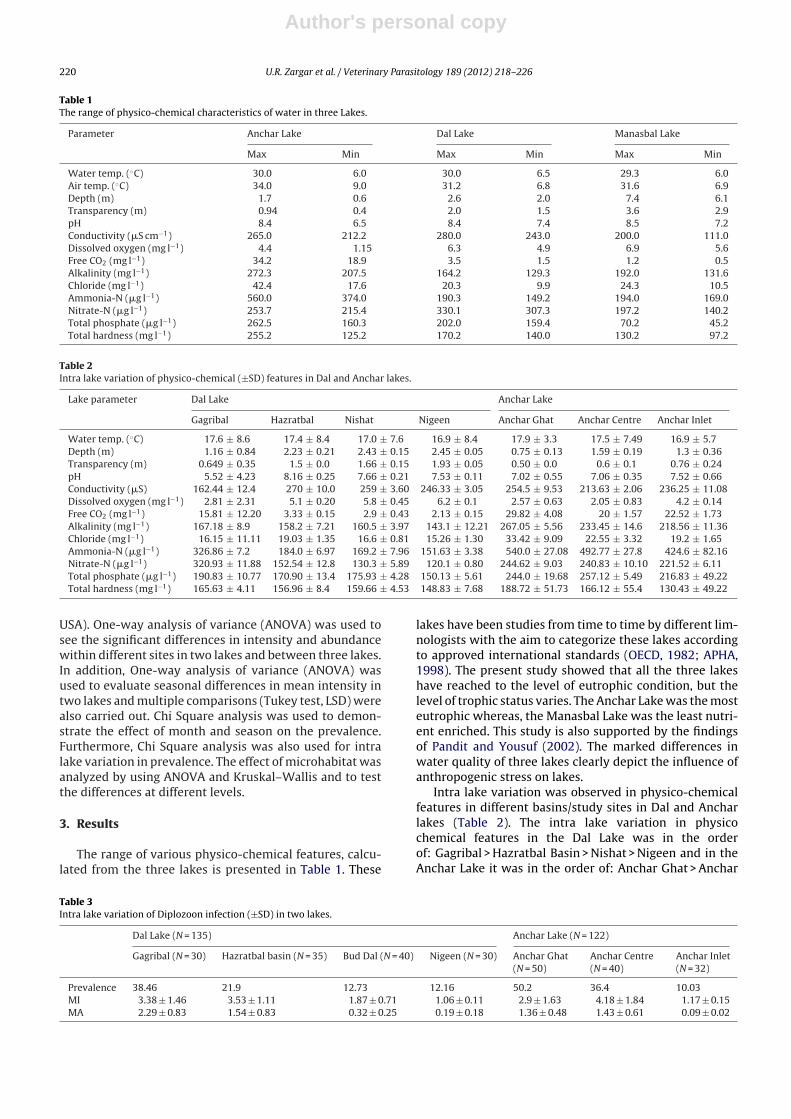
The results showed significant differences betweenright and left sides of gill (Kruskal–Wallis: X2 = 10.9,P = 0.001; Fig. 3). The number of Diplozoon parasites
Table 8Infection level in male and female fish in three lakes.
Infection parameter Male Female P-value
Anchar LakeSample size 51 53Prevalence 37.25 45.28 >0.05Mean intensity 3.05 2.54Mean abundance 1.14 1.15
Dal LakeSample size 58 61Prevalence 22.4 24.6 >0.05Mean intensity 3.15 5.13Mean abundance 0.75 1.24
Manasbal LakeSample size 33 31 >0.05Prevalence 9.09 12.9Mean intensity 2.0 3.25Mean abundance 0.18 0.42
Note: P-value is for prevalence between male and female.
Fig. 1. Overall comparative analysis of condition factor of fish in threelakes with different water quality.
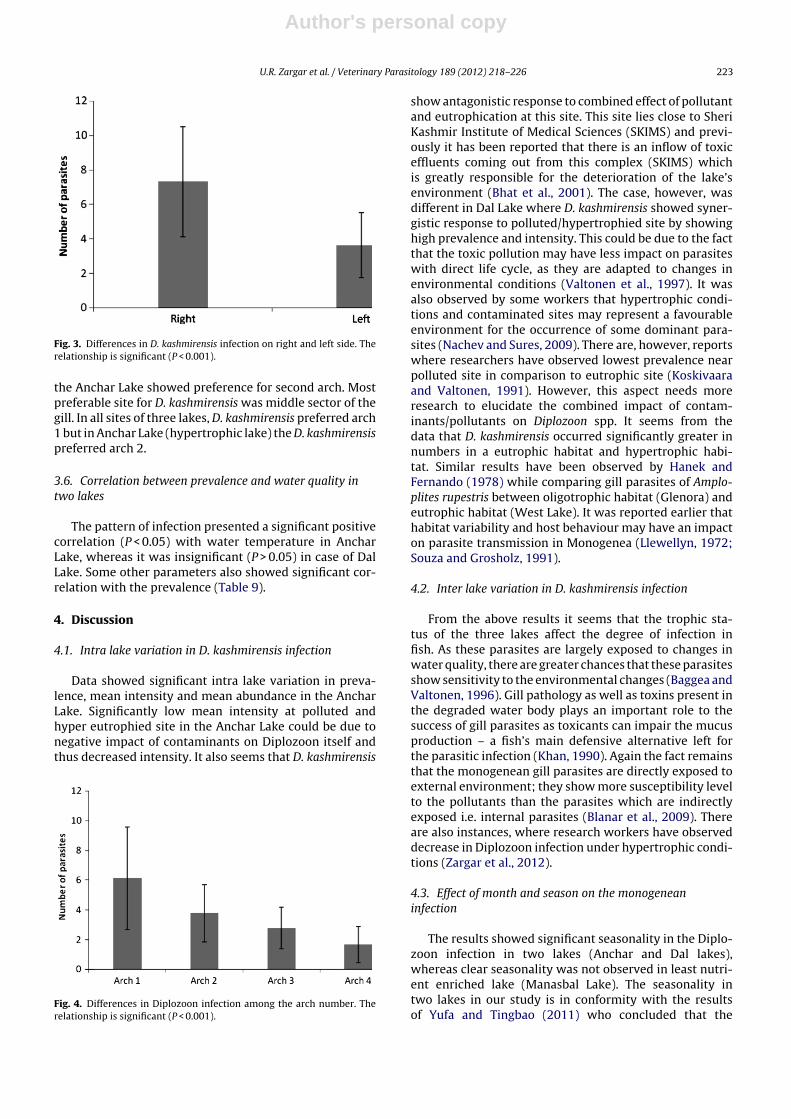
showed significant differences among arch number(Kruskal–Wallis: X2 = 10.90, P < 0.001). Arch 1 showedhighest average infestation of Diplozoon infection incomparison to other three arches (average no. of para-sites = 6.1 ± 3.4) and least was found on arch 4 (averageno. of parasites = 1.6 ± 1.2; Fig. 4). However, specimens in
uninfectedinfected
2.2
2.0
1.8
1.6
1.4
1.2
1.0
Fig. 2. Condition factor of infected and uninfected fish. The condition fac-tor (k) is significantly higher in uninfected fish (ANOVA: F = 16.5; P = 0.015).
Author's personal copy
U.R. Zargar et al. / Veterinary Parasitology 189 (2012) 218– 226 223
Fig. 3. Differences in D. kashmirensis infection on right and left side. Therelationship is significant (P < 0.001).
the Anchar Lake showed preference for second arch. Mostpreferable site for D. kashmirensis was middle sector of thegill. In all sites of three lakes, D. kashmirensis preferred arch1 but in Anchar Lake (hypertrophic lake) the D. kashmirensispreferred arch 2.
3.6. Correlation between prevalence and water quality intwo lakes
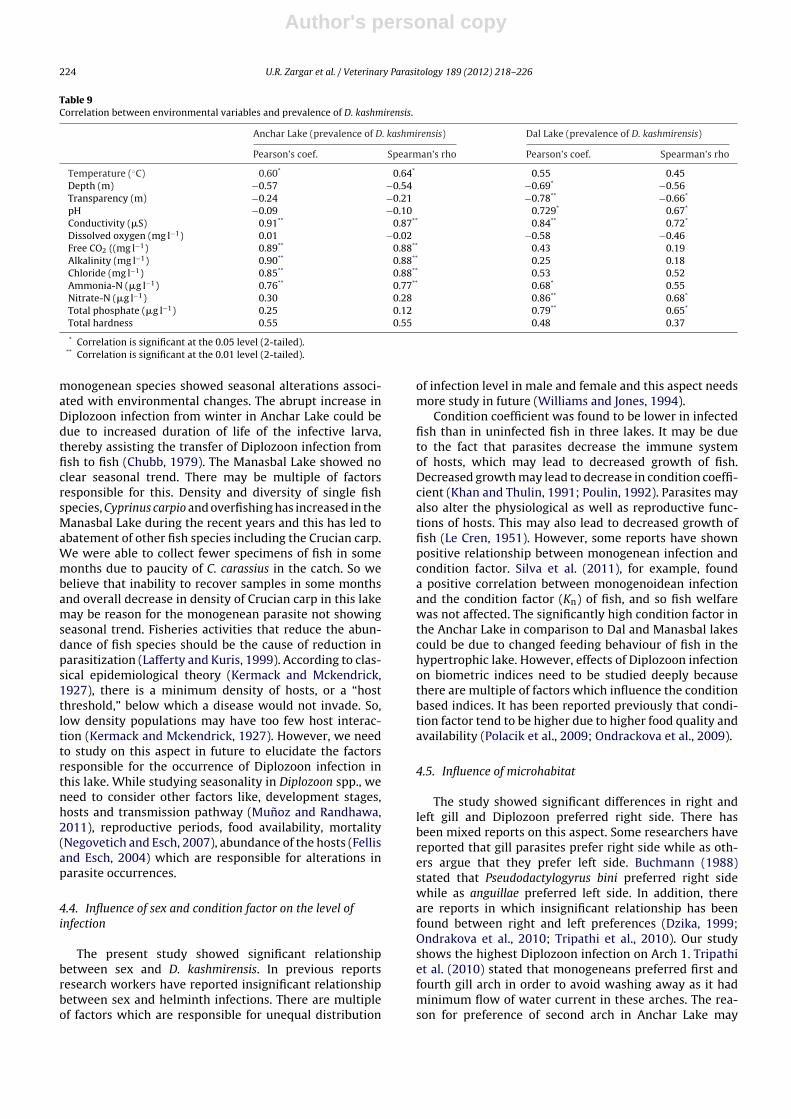
The pattern of infection presented a significant positivecorrelation (P < 0.05) with water temperature in AncharLake, whereas it was insignificant (P > 0.05) in case of DalLake. Some other parameters also showed significant cor-relation with the prevalence (Table 9).
4. Discussion
4.1. Intra lake variation in D. kashmirensis infection
Data showed significant intra lake variation in preva-lence, mean intensity and mean abundance in the AncharLake. Significantly low mean intensity at polluted andhyper eutrophied site in the Anchar Lake could be due tonegative impact of contaminants on Diplozoon itself andthus decreased intensity. It also seems that D. kashmirensis
Fig. 4. Differences in Diplozoon infection among the arch number. Therelationship is significant (P < 0.001).
show antagonistic response to combined effect of pollutantand eutrophication at this site. This site lies close to SheriKashmir Institute of Medical Sciences (SKIMS) and previ-ously it has been reported that there is an inflow of toxiceffluents coming out from this complex (SKIMS) whichis greatly responsible for the deterioration of the lake’senvironment (Bhat et al., 2001). The case, however, wasdifferent in Dal Lake where D. kashmirensis showed syner-gistic response to polluted/hypertrophied site by showinghigh prevalence and intensity. This could be due to the factthat the toxic pollution may have less impact on parasiteswith direct life cycle, as they are adapted to changes inenvironmental conditions (Valtonen et al., 1997). It wasalso observed by some workers that hypertrophic condi-tions and contaminated sites may represent a favourableenvironment for the occurrence of some dominant para-sites (Nachev and Sures, 2009). There are, however, reportswhere researchers have observed lowest prevalence nearpolluted site in comparison to eutrophic site (Koskivaaraand Valtonen, 1991). However, this aspect needs moreresearch to elucidate the combined impact of contam-inants/pollutants on Diplozoon spp. It seems from thedata that D. kashmirensis occurred significantly greater innumbers in a eutrophic habitat and hypertrophic habi-tat. Similar results have been observed by Hanek andFernando (1978) while comparing gill parasites of Amplo-plites rupestris between oligotrophic habitat (Glenora) andeutrophic habitat (West Lake). It was reported earlier thathabitat variability and host behaviour may have an impacton parasite transmission in Monogenea (Llewellyn, 1972;Souza and Grosholz, 1991).
4.2. Inter lake variation in D. kashmirensis infection
From the above results it seems that the trophic sta-tus of the three lakes affect the degree of infection infish. As these parasites are largely exposed to changes inwater quality, there are greater chances that these parasitesshow sensitivity to the environmental changes (Baggea andValtonen, 1996). Gill pathology as well as toxins present inthe degraded water body plays an important role to thesuccess of gill parasites as toxicants can impair the mucusproduction – a fish’s main defensive alternative left forthe parasitic infection (Khan, 1990). Again the fact remainsthat the monogenean gill parasites are directly exposed toexternal environment; they show more susceptibility levelto the pollutants than the parasites which are indirectlyexposed i.e. internal parasites (Blanar et al., 2009). Thereare also instances, where research workers have observeddecrease in Diplozoon infection under hypertrophic condi-tions (Zargar et al., 2012).
4.3. Effect of month and season on the monogeneaninfection
The results showed significant seasonality in the Diplo-zoon infection in two lakes (Anchar and Dal lakes),whereas clear seasonality was not observed in least nutri-ent enriched lake (Manasbal Lake). The seasonality intwo lakes in our study is in conformity with the resultsof Yufa and Tingbao (2011) who concluded that the
Author's personal copy
224 U.R. Zargar et al. / Veterinary Parasitology 189 (2012) 218– 226
Table 9Correlation between environmental variables and prevalence of D. kashmirensis.
Anchar Lake (prevalence of D. kashmirensis) Dal Lake (prevalence of D. kashmirensis)
Pearson’s coef. Spearman’s rho Pearson’s coef. Spearman’s rho
Temperature (◦C) 0.60* 0.64* 0.55 0.45Depth (m) −0.57 −0.54 −0.69* −0.56Transparency (m) −0.24 −0.21 −0.78** −0.66*
pH −0.09 −0.10 0.729* 0.67*
Conductivity (�S) 0.91** 0.87** 0.84** 0.72*
Dissolved oxygen (mg l−1) 0.01 −0.02 −0.58 −0.46Free CO2 ((mg l−1) 0.89** 0.88** 0.43 0.19Alkalinity (mg l−1) 0.90** 0.88** 0.25 0.18Chloride (mg l−1) 0.85** 0.88** 0.53 0.52Ammonia-N (�g l−1) 0.76** 0.77** 0.68* 0.55Nitrate-N (�g l−1) 0.30 0.28 0.86** 0.68*
Total phosphate (�g l−1) 0.25 0.12 0.79** 0.65*
Total hardness 0.55 0.55 0.48 0.37
* Correlation is significant at the 0.05 level (2-tailed).** Correlation is significant at the 0.01 level (2-tailed).
monogenean species showed seasonal alterations associ-ated with environmental changes. The abrupt increase inDiplozoon infection from winter in Anchar Lake could bedue to increased duration of life of the infective larva,thereby assisting the transfer of Diplozoon infection fromfish to fish (Chubb, 1979). The Manasbal Lake showed noclear seasonal trend. There may be multiple of factorsresponsible for this. Density and diversity of single fishspecies, Cyprinus carpio and overfishing has increased in theManasbal Lake during the recent years and this has led toabatement of other fish species including the Crucian carp.We were able to collect fewer specimens of fish in somemonths due to paucity of C. carassius in the catch. So webelieve that inability to recover samples in some monthsand overall decrease in density of Crucian carp in this lakemay be reason for the monogenean parasite not showingseasonal trend. Fisheries activities that reduce the abun-dance of fish species should be the cause of reduction inparasitization (Lafferty and Kuris, 1999). According to clas-sical epidemiological theory (Kermack and Mckendrick,1927), there is a minimum density of hosts, or a “hostthreshold,” below which a disease would not invade. So,low density populations may have too few host interac-tion (Kermack and Mckendrick, 1927). However, we needto study on this aspect in future to elucidate the factorsresponsible for the occurrence of Diplozoon infection inthis lake. While studying seasonality in Diplozoon spp., weneed to consider other factors like, development stages,hosts and transmission pathway (Munoz and Randhawa,2011), reproductive periods, food availability, mortality(Negovetich and Esch, 2007), abundance of the hosts (Fellisand Esch, 2004) which are responsible for alterations inparasite occurrences.
4.4. Influence of sex and condition factor on the level ofinfection
The present study showed significant relationshipbetween sex and D. kashmirensis. In previous reportsresearch workers have reported insignificant relationshipbetween sex and helminth infections. There are multipleof factors which are responsible for unequal distribution
of infection level in male and female and this aspect needsmore study in future (Williams and Jones, 1994).
Condition coefficient was found to be lower in infectedfish than in uninfected fish in three lakes. It may be dueto the fact that parasites decrease the immune systemof hosts, which may lead to decreased growth of fish.Decreased growth may lead to decrease in condition coeffi-cient (Khan and Thulin, 1991; Poulin, 1992). Parasites mayalso alter the physiological as well as reproductive func-tions of hosts. This may also lead to decreased growth offish (Le Cren, 1951). However, some reports have shownpositive relationship between monogenean infection andcondition factor. Silva et al. (2011), for example, founda positive correlation between monogenoidean infectionand the condition factor (Kn) of fish, and so fish welfarewas not affected. The significantly high condition factor inthe Anchar Lake in comparison to Dal and Manasbal lakescould be due to changed feeding behaviour of fish in thehypertrophic lake. However, effects of Diplozoon infectionon biometric indices need to be studied deeply becausethere are multiple of factors which influence the conditionbased indices. It has been reported previously that condi-tion factor tend to be higher due to higher food quality andavailability (Polacik et al., 2009; Ondrackova et al., 2009).
4.5. Influence of microhabitat
The study showed significant differences in right andleft gill and Diplozoon preferred right side. There hasbeen mixed reports on this aspect. Some researchers havereported that gill parasites prefer right side while as oth-ers argue that they prefer left side. Buchmann (1988)stated that Pseudodactylogyrus bini preferred right sidewhile as anguillae preferred left side. In addition, thereare reports in which insignificant relationship has beenfound between right and left preferences (Dzika, 1999;Ondrakova et al., 2010; Tripathi et al., 2010). Our studyshows the highest Diplozoon infection on Arch 1. Tripathiet al. (2010) stated that monogeneans preferred first andfourth gill arch in order to avoid washing away as it hadminimum flow of water current in these arches. The rea-son for preference of second arch in Anchar Lake may

Author's personal copy
U.R. Zargar et al. / Veterinary Parasitology 189 (2012) 218– 226 225
be due to low dissolved O2 conditions which change thesite specificity of D. kashmirensis (Chapman et al., 2000).Ondrakova et al. (2010) and Hanek and Fernando (1978)observed a preference of monogenean parasite for sec-ond gill arch. Turgut et al. (2006) observed a significantlygreater number of Dactylogyrus auriculatus on the secondand third gill arches (Koskivaara, 1992). There have beenalso reports that monogeneans often have over lappingniches (Rohde, 1981). Most monogenean species infect spe-cific host species and varies with host habitat, behaviourand other features (Rohde, 1979). Wiles (1967) reportedthat D. paradoxum showed random microdistribution ongills and pointed out that D. paradoxum mostly prevails onmedian sectors of gills.
Direct competition or interactive segregation maybe reason for microhabitat selection, in helminth para-sites (Holmes, 1972) while as internal factors are moreimportant for monogenean site selection (Rohde, 1979).However, effects of environmental factors cannot be ruledout as monogeneans are in direct contact with externalenvironment and may show response to physico chemicalfeatures (Bychowsky, 1961).
4.6. Effect of water quality on D. kashmirensis
The present study showed that some of the physicochemical features showed a significant positive correla-tion with the prevalence. Modu et al. (2011) showed thatthere exists a significant correlation between monogeneaninfection and water quality parameters in a pond. A num-ber of workers (Beer and German, 1993; Kennedy andWatt, 1994; Marcogliese, 2001; Lafferty and Kuris, 2005)have suggested that natural abiotic factors such as tem-perature, oxygen, salinity, hydrogen ion concentration andeutrophication have a positive influence on the occurrenceof parasite populations. Evidence from the present studysuggests that the water temperature plays an importantrole in the progression of D. kashmirensis. Dissolved O2 hasbeen found to be the predictor of habitat selection in mono-geneans (Ernst et al., 2005). Our results, however, showedinsignificant relationship between O2 level and D. kash-mirensis infection.
Positive relationship between the infection indices ofgill parasite and water quality suggest that the monoge-nean gill parasites can be used for stress indicator/effectindicator. There has been also debate on whether to usemonogenean parasites as pollution indicators as they arein direct contact with water. Nevertheless, there are someparasite/contamination studies, which are still unclear andneed more research. But monogeneans including Diplo-zoon parasites can act as an excellent non-specific earlywarning indicators. Recent meta-analysis by Blanar et al.(2009) showed that fresh water monogeneans are goodcandidates as potential biological indicators of pollution inaquatic ecosystems.
5. Conclusions and suggestions
In summary, intra lake variation, season, conditionfactor, microhabitat and water quality seem to have a sig-nificant impact on the D. kashmirensis infection. It also
seems from the data that eutrophic and hypertrophic habi-tats were favourable for D. kashmirensis. D. kashmirensisshowed both antagonistic and synergistic response to thecombined effect of pollution and eutrophication. It is sug-gested that more field and in vitro studies need to be carriedout to know the joint impact of pollution and eutrophica-tion on the Diplozoon infection. Present study will pavethe way for future studies on different ecological aspectsof Diplozoon parasites in this region as well as in simi-lar climatic regions of the world. Furthermore we need tounravel the specific factors which have major impact on theoccurrence of Monogenea in this region. Monogeneans inparticular should be integrated with bio monitoring pro-grammes, as these parasites can provide supplementaryinformation on pollution. It is also suggested that monoge-neans in general and Diplozoon spp. in particular should beanalyzed for accumulation capability of different contam-inants under experimental conditions in order to validatethe role of Diplozoon parasites in environmental monitor-ing.
Acknowledgements
We acknowledge and appreciate the kind help fromDirector Prof. A. Nahid Kamilli of Centre of Research forDevelopment (CORD), University of Kashmir for providingdepartmental vehicle for carrying out sampling trips to dif-ferent lakes. In addition financial support from UniversityGrants Commission (UGC) for providing fellowship to 1stauthor, Ummer Rashid Zargar is also acknowledged. Wealso acknowledge Mr. Sajad Hussain from Department ofLiguistics, University of Kashmir for his help during proofreading of the manuscript.
References
Ahmed, F., Chishti, M.Z., 2002. Fish trematode parasites of Kashmir.Part I. Genus Diplozoon Nordmann 1832 (Monogenea, Polyophistho-cotylea). Orient. Sci. 4, 79–91.
American Public Health Association, Washington, DC, 1998. Standardmethods for the examination of water and wastewater, 20th ed. Amer-ican Public Health Association, the American Water Works Associationand the Water Environment Federation, 1220 pp.
Baggea, A.M., Valtonen, E.T., 1996. Experimental study on the influence ofpaper and pulp mill effluent on the gill parasite communities of roach(Rutilus rutilus). Parasitology 112, 499–508.
Beer, S.A., German, S.M., 1993. Ecological prerequisites of worsening of thecercariosis situation in cities of Russia (Moscow region as an example).Parazitologiya 27, 441–449.
Bhat, S.A., Rather, S.A., Pandit, A.K., 2001. Impact of effluents from Sheri-Kashmir Institute of Medical Sciences (SKIMS), Soura on Anchar Lake.J. Res. Dev. 1, 30–37.
Blanar, C.A., Munkittrick, K.R., Houlahan, J., Mac Latchy, D.L., Marcogliese,D.J., 2009. Pollution and parasitism in aquatic animals: a meta-analysisof effect size. Aquat. Toxicol. 93, 1–80.
Buchmann, K., 1988. Interactions between the gill-parasitic monogeneansPseudodactylogyrus anguillae and P. bini and the fish host Anguillaanguilla B. Eur. Assoc. Fish Pat. 8, 98–99.
Bush, A.O., Lafferty, K.D., Lotz, J.M., Shostak, A.W., 1997. Parasitology meetsecology on its own terms: Margolis et al. revisited. J. Parasitol. 83,575–583.
Bychowsky, B.E., 1961. Monogenetic Trematodes: Their Systematics andPhylogeny. Transcript of a Russian Monograph. American Institute ofBiological Sciences, Washington, DC.
Chapman, L.J., Carmine, A., Lanciani Colin Chapman, A., 2000. Ecology of aDiplozoon parasite on the gills of the African cyprinid Barbus neumay-eri. East. Afr. Wildl. J. 38, 312–320.

Author's personal copy
226 U.R. Zargar et al. / Veterinary Parasitology 189 (2012) 218– 226
Chishti, M.A., Hassan, M., Feroz, S., Jabeen, U., 2000. Effects of tempera-ture, pH, and O2 on cestode and trematode infection in some fishesof Wular Lake, Kashmir. In: Khan, M.A., Zargar, M.Y. (Eds.), Agricul-ture and Environment. A. P. H. Publication Corporation, New Delhi,pp. 323–333.
Chubb, J.C., 1979. Seasonal occurrence of helminths in freshwater fishes.Part II. Trematoda. Adv. Parasitol. 17, 141–313.
CSIR, 1974. An Analytical Guide. Part 1. National Institute for WaterResearch, Pretoria, South Africa.
Dzika, E., 1999. Microhabitat of Pseudodactylogyrus anguillae and P. bini(Monogenea: dactylogyridae) on the gills of large-size European ell,Anguilla anguilla from Lake Gaj, Poland. Folia Parasitol. 46, 33–367.
Ernst, I., Whittington, I.D., Corneillie, S., Talbot, C., 2005. Effects of tem-perature, salinity, desiccation, hatchery & chemical treatment on eggembryonation & hatchery success of Benedenia sericola (Monogenea,Capsalidae a parasite of farmed Sericola spp.). J. Fish Dis. 28, 157–164.
Fellis, K.J., Esch, G.W., 2004. Community structure and seasonal dynamicsof helminth parasites in Lepomis cyanellus and L. macrochirus fromCharlie’s Pond, North Carolina, host size and species as determinantsof community structure. J. Parasitol. 90, 41–49.
Geets, A., Coene, H., Ollevier, F., 1997. Ectoparasites of the whitespottedrabbitfish, Siganus sutor (Valenciennes, 1835) of the Kenyan coast: dis-tribution within the host population and site selection on the gills.Parasitology 115, 69–79.
Hanek, G., Fernando, C.H., 1978. Spatial distribution of gill parasites ofLepomis gibbosus (L.) and Ambloplites surestris (Raf.). Can. J. Zool. 56,1235–1240.
Holmes, J.C., 1972. Site selection by parasitic helminths: interspecificinteractions, site segregation, and their importance to the develop-ment of helminth communities. Can. J. Zool. 51, 333–347.
Kennedy, C.R., Watt, R.J., 1994. The decline and natural recovery of anunmanaged coarse fishery in relation to changes in land use and atten-dant eutrophication. In: Cowx, I.G. (Ed.), Rehabilitation of FreshwaterFisheries. Blackwell Scientific, Oxford, pp. 366–375.
Kermack, W.O., Mckendrick, A.G., 1927. A contribution to the mathemat-ical theory of epidemics. Part 3. In further studies of the problem ofendemicity. Proc. Roy. Soc. Lond. A: Mat. 141, 92–122.
Khan, R.A., 1990. Parasitism in marine fish after chronic exposure topetroleum hydrocarbons in the laboratory and to the Exxon Valdezoil spill. Bull. Environ. Contam. Toxicol. 44, 759–763.
Khan, R.A., Thulin, J., 1991. Influence of pollution on parasites of aquaticanimals. Adv. Parasitol. 30, 201–238.
Koskivaara, M., 1992. Environmental factors affecting monogeneans par-asitic on freshwater fish. Parasitol. Today 8, 339–342.
Koskivaara, M., Valtonen, E.T., 1991. Paradiplozoon homoin (Monogenea)and other gill parasites of Roach, Rutilus tutilus in Finland. Aqua Fen-nica 21, 137–143.
Lafferty, K.D., Kuris, A.M., 1999. How environmental stress affects theimpacts of parasites. Limnol. Oceanogr. 44, 925–931.
Lafferty, K.D., Kuris, A.M., 2005. Parasitism and environmental distur-bances. In: Thomas, F., Guégan, J.F., Renaud, F. (Eds.), Parasitism andEcosystems. , pp. 113–123.
Le Cren, E.D., 1951. The length–weight relationship and seasonal cycle ingonadal weight and condition in the perch. Perca fluviatilus. J. Anim.Ecol. 20, 201–219.
Llewellyn, J., 1972. Behaviour of monogeneans. In: Canning, E.U., Wright,C.A. (Eds.), Behavioural Aspects of Parasite Transmission. AcademicPress, London, pp. 19–30.
Mackereth, F.J.H., Heron, J., Talling, J.F., 1978. Water Analysis: SomeRevised Methods for Limnologists. Freshwater Biological Association,Scientific Publication, 120 pp.
Marcogliese, D.J., 2001. Implications of climate change for parasitism ofanimals in the aquatic environment. Can. J. Zool. 79, 1331–1352.
Margolis, L., Esch, G.W., Holmes, J.C., Kuris, A.M., Schad, G.A., 1982. Theuse of ecological terms in parasitology (report Bush et al.—ParasiteEcology and Terminology 583 of an adhoc committee of the AmericanSociety of Parasitologists). J. Parasitol. 68, 131–133.
Modu, B.M., Saiful, M., Kartini, M., Kassim, Z., Hassan, M., Shaharom-Harrison, F.M., 2011. Impact of Monogenean Parasite in Relation to
Water quality Effects on the Structural changes in the Gills of Fresh-water Cat Fish, Hemibagrus nemurus Valenciennes 1840. EmpoweringScience, Technology and Innovation Towards a Better TomorrowUMTAS 2011 LSO17.
Munoz, G., Randhawa, H.S., 2011. Monthly variation in the parasitecommunities of the intertidal fish Scartichthys viridis (Blenniidae)from central Chile: are there seasonal patterns? Parasitol. Res. 109,53–62.
Nachev, M., Sures, B., 2009. The endohelminth fauna of barbel (Barbusbarbus) correlates with water quality of the Danube River in Bulgaria.Parasitology 136, 545–552.
Negovetich, N.J., Esch, G.W., 2007. Long-term analysis of Charlie’s pond:fecundity and trematode communities of Helisoma anceps. J. Parasitol.93, 1311–1318.
Organisation for Economic Co-operation & Development, 1982. Eutroph-ication of Waters. Monitoring, Assessment and Control. Organisationfor Economic Co-operation & Development, Paris, 154 pp.
Ondrackova, M., Davidova, M., Blazek, R., Gelnar, M., Jurajda, P., 2009.The interaction between an introduced fish host and local parasitefauna, Neogobius kessleri in the middle Danube River. Parasitol. Res.105, 201–208.
Ondrakova, M., Davidova, M., Prikrylova, I., Pecınkova, M., 2010. Mono-genean parasites of introduced pumpkinseed Lepomis gibbosus(Centrarchidae) in the Danube River Basin. J. Helminthol. 85, 435–441.
Pandit, A.K., Yousuf, A.R., 2002. Trophic status of Kashmir Himalayan lakesas depicted by water chemistry. J. Res. Dev. 2, 0972–5407.
Polacik, M., Janac, M., Jurajda, P., Adamek, Z., Ondrackova, M., Trichkova,T., Vassilev, M., 2009. Invasive gobies in the Danube: invasion suc-cess facilitated by availability and selection of superior food resources.Ecol. Freshw. Fish 8, 640–649.
Poulin, R., 1992. Toxic pollution and parasitism in freshwater fish. Para-sitol. Today 8, 58–60.
Rohde, K., 1979. A critical evaluation of intrinsic and extrinsic factorsresponsible for niche restriction in parasites. Am. Nat. 114, 648–671.
Rohde, K., 1981. Niche width of parasites in species rich and species-poorcommunities. Experientia 37, 359–361.
Silva, A., Tavares-Dias, M., Gt Jerônimo, Martins, M., 2011. Parasite diver-sity in Oxydoras niger (Osteichthyes: Doradidae) from the basin ofSolimões River, Amazonas state, Brazil, and the relationship betweenmonogenoidean and condition factor. Braz. J. Biol. 71, 791–796.
Souza, W.P., Grosholz, E.D., 1991. The influence of habitat structure onthe transmission of parasites. In: Bell, S.S., McCoy, E.D., Mushinsky,H.R. (Eds.), Habitat Structure: The Physical Arrangement of Objects inSpace. , pp. 300–324.
Tripathi, P., Agarwal, N., Pant, R., Agarwal, G.G., 2010. Microhabitat dis-tribution of some monogeneoideans, parasitizing the gills of Wallagoattu (Bl. And Sch., 1801) and their seasonal variation. J. Parasitic Dis.34, 24–28.
Turgut, E., Shinn, A., Wootten, R., 2006. Spatial distribution of Dactylogyrus(Monogenan) on the Gills of the Host. Turk. J. Fish Aquat. Sci. 6, 93–98.
Valtonen, E.T., Holmes, J.C., Koskivaara, M., 1997. Eutrophication, pollu-tion, and fragmentation: effects on parasite communities in roach(Rutilus rutilus) and perch (Perca fluviatilis) in four lakes in centralFinland. Can. J. Fish. Aquat. Sci. 54, 572–585.
Wiles, W., 1967. The occurrence of Diplozoon paradoxum Nordmann, 1832(Trematoda: Monogenea) in certain waters of northern England andits distribution on the gills of certain Cyprinidae. Parasitology 58,61–70.
Williams, H., Jones, A., 1994. Parasitic Worms of Fish. Taylor & Francis Ltd,p. 593.
Yufa, L., Tingbao, Y., 2011. Seasonal patterns in the community ofgill monogeneans on wild versus cultured orange-spotted grouper,Epinephelus coioides Hamilton, 1822 in Daya Bay, South China Sea.Aquac. Res., 1–11.
Zargar, U.R., Yousuf, A.R., Chishti, M.Z., Ahmed, F., Bashir, H., Ahmed, F.,2012. Effects of water quality and trophic status on helminth infec-tions in the cyprinid fish, Schizothorax niger Heckel, 1838 from threelakes in the Kashmir Himalayas. J. Helminthol. 86, 70–76.




















