
numa romaria poiitica , intormou o mmistro do fas telefonicas ...
Upload
independentCategory
view
0download
0
The Fas-Fas Ligand System and Other Modulators ofApoptosis in the Cornea
Steven E. Wilson *"\ Qian Li,\ Jian Weng,\ Patricia A. Barry-Lane,^ James V. Jester,fQianwa Liang,* and Robert J. WordingerX
Purpose. Previous studies have suggested that the disappearance of anterior keratocytes afterinjury to the overlying epithelium is mediated by apoptosis. The authors examined the expres-sion of the apoptosis-related modulators, Fas (receptor), Fas ligand, Bax, Bcl-2, Bcl-XL, andinterleukin-1 beta converting enzyme (ICE) in corneal cells as candidate mediators of thisresponse and tested the effect of Fas receptor-stimulating antibody on corneal stromal fibro-blast cells in vitro.Methods. Reverse-transcription-polymerase chain reaction was used to detect FAS, FAS ligand,Bax, Bcl-2, Bcl-XL, and ICE mRNA expression in primary cultures of human corneal epithelial,stromal fibroblast, and endothelial cells. Immunohistochemistry was applied to detect Fas andFas ligand proteins in fresh-frozen sections of normal human cornea. The effect of FAS-stimulating monoclonal antibody on first-passage stromal fibroblasts was studied using a DNAfragmentation assay, the live-dead assay with fluorescent microscopy, toluidene blue stainingwith light microscopy, and electron microscopy.Results. FAS, Fas ligand, Bax, Bcl-2, Bcl-XL, and ICE mRNAs are expressed in all three majorcell types of the cornea. Fas protein is expressed in corneal epithelial, keratocyte, and endothe-lial cells in fresh-frozen human cornea. Fas ligand protein, however, was detected in cornealepithelial and endothelial, but not keratocyte, cells. Fas-stimulating antibody induced first-passage stromal fibroblast cell death with morphologic changes and DNA fragmentationconsistent with apoptosis.Conclusions. The Fas system (Fas and Fas ligand) modulators and final common pathwaymediators of apoptosis are expressed in corneal cells. The distribution of Fas (epithelial,keratocyte, and endothelial cells) and Fas ligand (epithelial and endothelial cells) proteinexpression in fresh-frozen corneal tissue suggests that Fas ligand expressed in corneal epithe-lial and endothelial cells modulates functions in keratocyte cells and, possibly, autocrine-juxtacrine functions in epithelium and endothelium. The Fas-Fas ligand system is expressedin the cornea and could have important functions in normal corneal physiology and inthe pathophysiology of corneal disease, including modulation of keratocyte apoptosis afterepithelial injury. Invest Ophthalmol Vis Sci. 1996;37:1582-1592.
Apoptosis (programmed cell death) is a fundamentalprocess that occurs during development, homeostasis,and wound healing in the tissues of essentially allmulticellular organisms.1'2 We recently demonstrated
From the *Eye Institute and the Department of Cell Biology, The Cleveland ClinicFoundation, Cleveland, Ohio; the f Department of Ophthalmology, University ofTexas Southiuestern Medical Center at DalUis; and the %North Texas Eye ResearchInstitute, University of North Texas Health Sciences Center, Fort Worth, Texas.Presented in part at the Ocular Cell and Molecular Biology Symposium, San Diego,California, August 1995, and the Association for Research in Vision andOphthalmology, Fort Lauderdale, Florida, April 1996.Supported by US Public Health Service grant EY10056 from the National Institutesof Health.Submitted for publication January 17, 1996; revised March 21, 1996; acceptedMarch 22, 1996.Proprietary interest calegoiy: N.Reprint requests: Steven E. Wilson, Eye Institute and Department of Cell Biology/A31, The Cleveland Clinic Foundation, 9500 Euclid Avenue, Cleveland, OH44195.
apoptosis of keratocytes mediated through epithelial -stromal interactions after wounding of the cornealepithelium,3 providing a mechanism for the disap-pearance of keratocytes in the anterior stroma aftercorneal wounding initially described by Nakayasu4 inthe rat and Crosson5 in the rabbit and recently con-firmed in primates.6 We hypothesized a fundamentalrole for this system in the maintenance of cornealtissue organization, the response to injury, and thepathophysiology of corneal diseases.3 For example,apoptosis of the anterior stromal keratocytes that oc-curs after excimer laser photorefractive keratectomyprobably is an initiating event in the subsequentwound healing response.3
The molecular mechanisms underlying the initia-
1582Investigative Ophthalmology & Visual Science, July 1996, Vol. 37, No. 8Copyright © Association for Research in Vision and Ophthalmology

Apoptosis-Related Modulators 1583
tion and regulation of apoptosis are topics of intenseinvestigation throughout the scientific community.Numerous mutations affecting mediators of specificstages of apoptosis have been identified in the nema-tode Caenorhabditis elegans7 and analogous modulatorshave been identified in higher organisms (Fig. I).8
For example, the C. elegans death gene, ced-S, is analo-gous to interleukin-1 beta converting enzyme (ICE)-like protease, and the ced-9 apoptosis inhibitory geneis analogous to Bcl-2.8 Specific cell types have beenshown to have multiple, alternative, extracellular andintracellular apoptosis signaling pathways that con-verge on a single common death pathway (Fig. I).8
For example, our recent studies have demonstratedthat interleukin (IL)-l alpha and ILrl beta can induceapoptosis in corneal stromal fibroblasts in vitro andkeratocytes in vivo.3 Because IL-1 alpha is expressedby corneal epithelial cells14'15 and ILrl receptor is ex-pressed by keratocytes,1617 we hypothesized that in-jury- or death-induced release of IL-1 alpha from cor-neal epithelial cells activates the final common apo-ptotic pathway in keratocytes, inducing cell death. Wecould not inhibit the in vivo apoptotic response ofkeratocytes to epithelial wounding, however, by priorinjection of IL-1 receptor antagonist into the stroma.3
One possible explanation for the ineffectiveness of IL-1 receptor antagonist at inhibiting this response isexpression of multiple systems activating the death ofthe keratocyte cells in response to epithelial cell in-jury. An important lesson learned from transgenic ani-mal models is that such duplication of regulatory sys-tems is the norm for many cytokine- and growth factor-mediated processes.18 In the current study, we haveexamined corneal cell expression of several modula-tors known to be involved in apoptosis. These includemodulators that are likely to regulate intercellularcommunication leading to apoptosis, such as Fas(APO-1) and Fas ligand, as well as members of thefinal common apoptotic pathway (ICE, Bcl-2, Bcl-XL,and Bax). We also examined the effect of a Fas-stimu-lating antibody on corneal stromal fibroblasts.
METHODS AND MATERIALS
,,. CORTICO-I STEROIDS
;>*0 i, x
CENTRALDEATH SIGNAL MODULATOR
"REAPER-DROSPHILA"
ICE-LIKE IPROTEASE;
CELLDEATH
FIGURE l. Cell-specific and common pathways of apoptosis.Examples of cell-specific signaling pathways that produceapoptosis in cells are diagrammed above the broken hori-zontal line. A particular cell type may undergo apoptosisin response to several different signals. All the cell-specificpathways shown in the diagram are dependent on extracellu-lar signals. Cell-specific signals also may be intracellular.Each of the cell-specific pathways is thought to trigger acommon apoptotic pathway (diagrammed schematically be-low the broken line) by a central death signal. The macro-molecule serving this function in higher organisms has notbeen identified. A gene called reaper that appears to servesuch a function has been identified in Drosophila melanogas-ter.9 The reaper gene product has been shown to integrateinformation from alternative signaling pathways. Deletionsof reaperhave been found to suppress the apoptotic responseto every stimulus evaluated to date. The subsequent steps(arrows) leading to apoptotic cell death are likely orderedin a pathway. Many participants in the pathway have yet tobe identified in higher organisms. Interleukin-1 beta con-verting enzyme (ICE) or an ICE-like protein that is a mam-malian homologue of the Caenorhabditis elegans cell deathgene ced-S is one of the modulators in the common path-
Immunohistochemistry for Fas and Fas Ligand wav- T h e Bcl-2 protein is able to suppress many apoptotic
Corneoscleral rims were excised from eyes of patientswith conjunctival melanoma not involving the corneaor choroidal melanoma within 5 minutes of eviscera-tion or enucleation, respectively, embedded in HistoPrep (Fisher, Fairlong, NJ), snap frozen in liquid ni-trogen, and stored at — 85°C. The research followedthe tenets of the Declaration of Helsinki, and in-formed consent was obtained from each patient be-fore surgery after the nature and the possible conse-quences of the study were explained. This study wasapproved by the Institutional Review Boards at the
death programs in higher organisms (dash indicates inhibi-tion)." Bcl-2 is the mammalian homologue to ced-9, thegene product that suppresses apoptosis in cells of C. elegans.Another member of the Bcl family that appears to suppressapoptosis Bcl-XL.
12 Both Bcl-2 and Bcl-XL heterodimerizewith a product of another gene called Bax.13 Overexpressionof Bax accelerates apoptosis. The exact relationship of Bcl-2, Bcl-X ,̂ and Bax to cell survival versus apoptosis is un-clear.""13 It appears, however, that the equilibrium betweenhomodimers and heterodimers of these proteins may havea role in regulating the common apoptotic pathway. Expres-sion in corneal cells of modulators within hatched boxes isinvestigated in this study.

1584 Investigative Ophthalmology & Visual Science, July 1996, Vol. 37, No. 8
TABLE l. Polymerase Chain Reaction Primers
Modulator Reference Size Upstream Primer Downstream Primer
BCL^2 alpha 11 332/>332 TTGTGGCCTTCTTTGAGTTCG (exon 1)BCL^X,. 12 379/unknown GGAGCTGGTGGTTGACTTTCTBAX alpha 13 482/=>482 AGACAGGGGCCCTTTTGCTTC (exon 2)Beta actin 20 321/772 AGGCCAACCGCGAGAAGATGACC (exon 3)ICE 21 424/=>424 AGCTTTGATTGACTCCGTTAT (exon 2)FAS 22, 23 413/>4000 AGACTGCGTGCCCTGCCAAGA (exon 3)FAS LIGAND 24, 25 177/177 TTCTTCCCTGTCCAACCTCTG (exon 1)
750/^750
TACTGCTTTAGTGAACCTTTT (exon 2)CCGGAAGAGTTCATTCACTACGAGCACTCCCGCCACAAAGAT (exon 6)GAAGTCCAGGGCGACGTAGCAC (exon 4)CAGATTTTGTAGCAGCATTGT (exon 5)CAGGATTTAAGGTTGGAGATT (exon 7)AAAACATCACAAGGAGACACA (exon 1)TCTTCCCCTCCATCATCACCA (exon 4)
Cleveland Clinic Foundation, (Cleveland, OH) andthe University of Texas Southwestern Medical Center(Dallas, TX).
Seven-micrometer sections were prepared with aReichert-Jung (Leica, Deerfield, IL) cryostat. Tissuesections were fixed in acetone at — 20°C for 10 min-utes. Immunohistochemistry for Fas or Fas ligand wasperformed using standard methods involving biotinyl-ated secondary antibodies and streptavidin-conju-gated peroxidase (Universal LSAB + Kit, peroxidase;DAKO, Carpinteria, CA) according to the manufactur-er's instructions, except that sections were incubatedovernight at 37°C with Fas, Fas ligand, or control pri-mary antibody. Fas immunohistochemistry was per-formed with an anti-human Fas mouse IgM mono-clonal antibody (Upstate Biotechnology, Lake Placid,NY). Control immunohistochemistry was performedwith a nonimmune mouse IgM (RD Systems, Minneap-olis, MN). Both Fas and control antibodies were usedat a concentration of 10 /ig/ml. An anti-Fas ligandrabbit polyclonal antibody (Fas-L N-20; Santa CruzBiotechnology, Santa Cruz, CA) was used at a concen-tration of 2 /Ltg/ml. Control preabsorption was per-formed for Fas ligand by preincubating the antibodywith sc-834P control Fas ligand peptide (Santa CruzBiotechnology) at a concentration of 20 fxg/ml for 30minutes before incubating with tissue sections over-night at 37°C.
Reverse-Transcription-Polymerase ChainReaction Method for the Detection ofMessenger RNA
Total cellular ribonucleic acid (RNA) was isolated,and cDNA was synthesized from human primary cul-tures of corneal epithelial, stromal fibroblast, and en-dothelial cells, as previously described.19 The qualityof cDNA synthesis was monitored through the ampli-fication of beta actin. Only cDNA yielding beta actinamplifications of the expected size for beta actinmRNA, without contamination with genomic beta ac-tin amplification product of a larger size, was used forexperimental amplification.19 Polymerase chain reac-tion primers for ICE, Fas, Fas ligand, Bcl-2, Bcl-XL,and Bax were designed from the previously reportedsequences (Table 1)-"-13-iJ()-25 Polymerase chain reac-
tion reactions were performed with a temperature cy-cler (MJ Research, Watertown, MA) according to apreviously described method.19 A modified hot-startmethod was used in which anti-TAQ polymerase anti-body (Clontech, Palo Alto, CA) was added to the reac-tions, according to the manufacturer's instructions,to prevent initiation of amplification until after thereactions initially were raised to the antibody denatur-ing temperature of 70°C. Polymerase chain reactionamplification products were run on agarose gels aspreviously described.19 Polymerase chain reactionproducts were cut from agarose gels, cloned into thePCR II Cloning Vector (Invitrogen, San Diego, CA),and sequenced (Sequenase 2.0, United States Bio-chemical, Cleveland, OH) according to the manufac-turer's protocols.
Effect of Fas-Stimulating Antibody on StromalFibroblasts
Primary human stromal fibroblast cells were culturedas previously described.19 First-passage stromal fibro-blast cells were plated at densities from 5 X 102 to 1X 105 cells/cm2 in standard six-well plates (Corning,Corning, NY) in Eagle's modified essential mediumwith 10% fetal bovine serum. Twenty-four hours afterplating, the medium was changed to Eagle's modifiedessential medium with 0.5% fetal bovine serum beforeeither 100 ng/ml of anti-human Fas mouse mono-clonal IgM antibody (Upstate Biotechnology) or 100ng/ml control mouse IgM (RD Systems) was added.
After 24 hours, the percent of dead cells per fieldwas determined in 10 randomly selected 100X in-verted microscope fields (model TMF; Nikon, Mel-ville, NY) for both the anti-Fas and control antibodygroups. Statistical comparisons were made using theMann-Whitney Test. P < 0.05 was considered statisti-cally significant. Photographs also were obtained withthe inverted microscope.
First-passage human stromal fibroblasts exposedto anti-Fas or control antibody for 24 hours were testedfor viability with the Live/Dead Eukolight Viability/Cytotoxicity Assay (Molecular Probes, Eugene, OR)according to the manufacturer's instructions. Livecells have ubiquitous intracellular esterase activity thatconverts nonfluorescent, cell-permeant, calcein acet-

Apoptosis-Relatcd Modulators
EPI SFBP M 1 2 1 2600 • • • • • • • »4 0 0 - ^ ^ ^ ^ ^ ^ E l
3— y
HCN1 2
—a a
C
413
FIGURE 2. Fas mRNA expression in human corneal cells. Pri-mary cultures of human corneal epithelial {EPI), stromalfibroblast (SF), and endothelial (HCN) cells were evaluatedfor the production of Fas mRNA by reverse-transcription -polymerase chain reaction. Each corneal cell type yieldedamplification bands of the expected size of 413 bp for FasmRNA. M = 100 bp marker with sizes in bp (BP) indicatedto the left. C = simultaneous control reaction without addedcDNA target.
oxymethyl ester to calcein resulting in green fluores-cence with a fluorescein isothiocyanate filter. Thelevel of green fluorescence diminishes as cells die.Concurrently, ethidium homodimer enters dead cellsbecause of increased permeability from membranedamage and binds nucleic acids. Chromatin withindead cells fluoresces red with a rhodamine filter.Chromatin condensation frequently can be detectedin cells undergoing apoptosis. Computer-generatedcomposites that allow both colors to be displayed si-multaneously were obtained using cells plated onto60 mm Petriperm tissue culture dishes (Bachofer,Reutlingen, Germany) and were viewed using a LeitzFluovert FU microscope (Leica, Deerfield, IL)equipped with a Thermal Liquid Coupled Micro Incu-bator (Adams and List Associates, Westbury, NY) andconstant 5% CO^ in an air perfusion system. Fluores-cent images were captured digitally using a high-per-formance CCD camera (COHU, San Diego, CA) andintegrator-frame storer (Colorado Video, Boulder,CO). Images were digitized using a 486 personal com-puter with a Data Translation DT3852 image acquisi-tion card (Marlboro, MA) and 8 Mbytes of on-boardmemory. Individual images were transferred to a Sili-con Graphics (Mountain View, CA) workstation (Per-sonal Iris 4D-35G) and processed using the ANA-LYZE image processing software program (Mayo Med-ical Ventures, Rochester, MN). Final images werephotographed using an AGFA-Matrix film recorder(model 6564; Orangeburg, NY) and 4X5 Ektachrome64T (Eastman Kodak, Rochester, NY).
First-passage human stromal fibroblasts were ex-posed to anti-Fas or control antibody for 24 hours.Dead cells were combined with living cells that hadbeen removed from the culture flask by trypsinization.DNA was isolated, and ethidium bromide-stained gelswere used to detect internucleosomal DNA fragmenta-tion as previously described.20 The experiment wasrepeated three times.
First-passage stromal fibroblasts exposed to anti-Fas monoclonal or control antibody were trypsinized,
1585
washed with medium containing 10% fetal bovine se-rum, pelleted in a 1.5 ml Eppendorf tube, and fixed in3% gluteraldehyde and 1% paraformaldehyde. One-micron sections were stained with toluidine blue 1%in borate buffer and photographed with a light micro-scope (Optiphot-2, Nikon). Electron microscopy wasperformed as previously described27 on the fixed cellpellets. Electron microscopy sections were cut at 70nm and stained with 3% uranyl acetate for 15 minutes,followed by 3 minutes in Reynold's lead citrate.
RESULTS
Fas mRNA was detected by reverse-transcription-poly-merase chain reaction (RT-PCR) in corneal epithe-lial, stromal fibroblast, and endothelial cells (Fig. 2)in primary culture. Nucleic acid sequencing demon-strated that the amplification product of the expectedsize was identical to the known sequence for Fas. Fasprotein was detected by immunohistochemistry in cor-neal epithelial, keratocyte, and endothelial cells (Fig.3). Although each cell type stained diffusely, perinu-clear staining was prominent.
Fas ligand mRNA was detected by RT-PCR in all
o
c>
A
I
00
\i B
0
C0D
FIGURE 3. Immunohistochemical detection of Fas in humancorneal cells. Fas protein was detected in human cornealepithelial (A, between hollow arroios), keratocyte (A,C, arrows),and endothelial (C, holloxv arrow) cells. The morphology ofthe human endothelial cells was distorted by cryostat sec-tioning, but this did not influence immunohistologic stain-ing. Each cell type appeared to stain diffusely, althoughperinuclear staining seemed most prominent. No stainingof cells was noted in adjacent sections when a nonimmuneIgG was used as a control (B,D). Magnification, X200.

1586 Investigative Ophthalmology & Visual Science, July 1996, Vol. 37, No. 8
FIGURE 4. Fas ligand mRNA expression in human cornealcells. Primary cultures of human corneal epithelial (EPI),stromal fibroblast (SF), and endothelial (HCN) cells wereevaluated for the production of Fas mRNA by reverse-tran-scription-polymerase chain reaction. Each corneal cell typeyielded amplification bands of the expected size of 177 bpfor Fas ligand mRNA. M = a 100 bp marker with sizes inbp (BP) indicated to the left. C = a simultaneous controlreaction without added cDNA target.
three major cell types of the cornea in primary culture(Fig. 4). Nucleic acid sequencing demonstrated thatthe amplification product of the expected size wasidentical to the known sequence for Fas ligand. Afterthe genomic organization of the Fas ligand was re-ported,25 we also tested a second downstream PCRprimer that gave a different-sized PCR product formRNA and genomic amplifications (Table 1). Ampli-fication product of the expected size for mRNA (750bp), but not genomic, amplification was detected (notshown) with the second primer set. Fas ligand proteinwas detected by immunohistochemistry in corneal epi-thelial and endothelial, but not keratocyte, cells (Fig.5). Thus, although Fas ligand mRNA was detected inprimary cultures of corneal stromal fibroblasts, Fasligand protein was not detected in keratocytes in fresh-frozen human cornea.
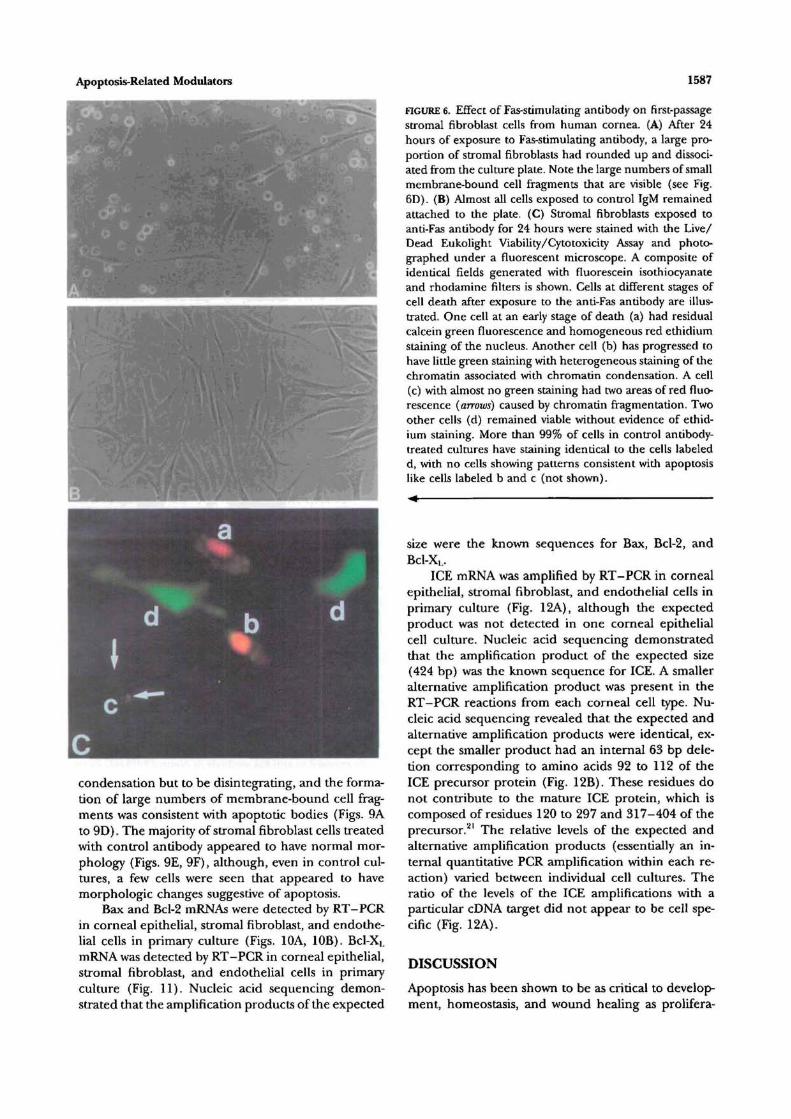
After 12 to 24 hours of exposure to anti-Fas anti-body and depending on the donor, a large proportionof first-passage stromal fibroblast cells had roundedup and dissociated from the culture plate (Fig. 6A).Few stromal fibroblasts rounded up and dissociatedin flasks treated with control IgM (Fig. 6B). In a repre-sentative experiment with cells plated at 1 X 104 cells/cm2, 75% ± 8% (SD) of stromal fibroblast cells ex-posed to anti-Fas antibody had dissociated and 1 % ±1% exposed to control antibody had dissociated at 24hours. The difference was statistically significant (P— 0.0002). Results were similar in five experimentsperformed with stromal fibroblasts from different do-nors. There appeared to be no difference in the re-sponse of stromal fibroblasts to die anti-Fas antibodywith varying plating densities between 5 X 102 to 1X 105 cells/cm8. The Live/Dead Eukolight Viability/Cytotoxicity Assay (Molecular Probes) demonstratedthat cells rounding up and dissociating from the platein response to the anti-Fas antibody were dead or dy-ing (Fig. 6C). In addition, fluorescence microscopywith this assay showed that many cells exposed to Fas-stimulating antibody had chromatin condensationand fragmentation consistent with apoptosis.
Internucleosomal DNA fragmentation was de-tected when human stromal fibroblasts were exposedto Fas-stimulating antibody, but not to control anti-body (Fig. 7). DNA fragments less than approximately1500 bp could not be detected in three separate exper-iments.
Toluidine blue-stained stromal fibroblast cells ex-posed to anti-Fas antibody had cell shrinkage, bleb-bing with formation of membrane-bound bodies, andcondensation and fragmentation of the chromatinconsistent with apoptosis (Figs. 8A to 8D). Controlantibody-treated stromal fibroblast cells maintainednormal cellular morphology with few cells showingchanges consistent with apoptosis (Figs. 8E, 8F).
Electron microscopy of pelleted stromal fibro-blasts that had been exposed to anti-Fas antibody re-vealed large numbers of cells with chromatin conden-sation and nucleosomal fragmentation (Figs. 9A to9C). There were also numerous membrane-bound cellfragments, many of which contained cell organelles(Figs. 9A to 9D). Many cells in the anti-Fas-treatedcultures were observed not only to have chromatin
A
FIGURE 5. Immunohistochemical detection of Fas ligand inhuman corneal cells. Fas ligand protein was detected inhuman corneal epithelial (A, between arrows) and endothelial(C, arrow) cells but not in keratocyte cells (A,C). Note thatin both epithelial and endothelial cells, a staining patternconsistent with Fas ligand association with the cell mem-brane was noted. The morphology of the human endothelialcells was distorted by cryostat sectioning, but this did notinfluence immunohistologic staining. Staining was reducedmarkedly when antibody was preincubated with Fas ligandantigen (B,D), demonstrating the specificity of the detectionin human corneal cells. Magnification, X200.
Apoptosis-Related Modulators
condensation but to be disintegrating, and the forma-tion of large numbers of membrane-bound cell frag-ments was consistent with apoptotic bodies (Figs. 9Ato 9D). The majority of stromal fibroblast cells treatedwith control antibody appeared to have normal mor-phology (Figs. 9E, 9F), although, even in control cul-tures, a few cells were seen that appeared to havemorphologic changes suggestive of apoptosis.
Bax and Bcl-2 mRNAs were detected by RT-PCRin corneal epithelial, stromal fibroblast, and endothe-lial cells in primary culture (Figs. 10A, 10B). Bcl-X^mRNA was detected by RT-PCR in corneal epithelial,stromal fibroblast, and endothelial cells in primaryculture (Fig. 11). Nucleic acid sequencing demon-strated that the amplification products of the expected
1587
FIGURE 6. Effect of Fas-stimulating antibody on first-passagestromal fibroblast cells from human cornea. (A) After 24hours of exposure to Fas-stimulating antibody, a large pro-portion of stromal fibroblasts had rounded up and dissoci-ated from the culture plate. Note the large numbers of smallmembrane-bound cell fragments that are visible (see Fig.6D). (B) Almost all cells exposed to control IgM remainedattached to the plate. (C) Stromal fibroblasts exposed toanti-Fas antibody for 24 hours were stained with the Live/Dead Eukolight Viability/Cytotoxicity Assay and photo-graphed under a fluorescent microscope. A composite ofidentical fields generated with fluorescein isothiocyanateand rhodamine filters is shown. Cells at different stages ofcell death after exposure to the anti-Fas antibody are illus-trated. One cell at an early stage of death (a) had residualcalcein green fluorescence and homogeneous red ethidiumstaining of the nucleus. Another cell (b) has progressed tohave little green staining with heterogeneous staining of thechromatin associated with chromatin condensation. A cell(c) with almost no green staining had two areas of red fluo-rescence (arrows) caused by chromatin fragmentation. Twoother cells (d) remained viable without evidence of ethid-ium staining. More than 99% of cells in control antibody-treated cultures have staining identical to the cells labeledd, with no cells showing patterns consistent wiui apoptosislike cells labeled b and c (not shown).
size were the known sequences for Bax, Bcl-2, andBcl-XL.
ICE mRNA was amplified by RT-PCR in cornealepithelial, stromal fibroblast, and endodielial cells inprimary culture (Fig. 12A), although the expectedproduct was not detected in one corneal epithelialcell culture. Nucleic acid sequencing demonstratedthat the amplification product of the expected size(424 bp) was die known sequence for ICE. A smalleralternative amplification product was present in theRT-PCR reactions from each corneal cell type. Nu-cleic acid sequencing revealed diat die expected andalternative amplification products were identical, ex-cept the smaller product had an internal 63 bp dele-tion corresponding to amino acids 92 to 112 of dieICE precursor protein (Fig. 12B). These residues donot contribute to die mature ICE protein, which iscomposed of residues 120 to 297 and 317-404 of theprecursor.21 The relative levels of the expected andalternative amplification products (essentially an in-ternal quantitative PCR amplification within each re-action) varied between individual cell cultures. Theratio of the levels of the ICE amplifications with aparticular cDNA target did not appear to be cell spe-cific (Fig. 12A).
DISCUSSION
Apoptosis has been shown to be as critical to develop-ment, homeostasis, and wound healing as prolifera-

1590
BAX BCL-2
Investigative Ophthalmology & Visual Science, July 1996, Vol. 37, No. 8
HSF HCE HCN
BAX BCL-2
BFIGURE 10. Bax and Bcl-2 mRNA expression in human cor-neal cells. (A) Reverse-transcription-polymerase chain reac-tion amplification products of the expected size for Bax(482 bp) and Bcl-2 (332 bp) mRNAs were detected in pri-mary human stromal fibroblast cells. For each primer pair,results are shown for cDNA prepared from cells from threedifferent donors (donors 1, 2 and 3). C = a simultaneouscontrol amplification without cDNA for each primer set. M= a 100 bp marker with sizes in bp (BP) indicated to theleft. (B) Bax and Bcl-2 mRNAs were detected by RT-PCRin primary cultures of human corneal epithelial (EPI) andendothelial (HCN) cells.
late corneal tissue organization.3 The distribution ofexpression of Fas-Fas ligand in the cornea suggeststhat such signaling could occur from the epithelialand endothelial cells to keratocyte cells by this system.The IL-1 -ILrl receptor system has been shown to havea similar pattern of cell expression in vivo and to in-duce comparable effects on corneal cells.3 Whetherthe Fas-Fas ligand and 1L-1-IL-1 receptor systemsfunction independently or are interrelated cannot bedetermined from the current study. If soluble Fas li-gand is released from epithelial cells after injury, it
• - * - 379 BP
200FIGURE U. Bcl-XL mRNA expression in human corneal cells.Primary cultures of human corneal epithelial (EPI), stromalfibroblast (SF), and endodielial (HCN) cells were evaluatedfor the production of Bcl-X|. mRNA by reverse-transcrip-tion-polymerase chain reaction. Each corneal cell typeyielded amplification bands of the expected size of 379 bpfor Bcl-XL ligand mRNA. M = a 100 bp marker with sizesin bp (BP) indicated to the left. C = a simultaneous controlreaction without added cDNA target.
BP M
.424 BPS361 BP
B
GM-CTC TCA GCA GAT C M ACA TCT GSA AAT TAC CTT AAT ATC CAALeu Ser Ala Asp Gin Thr Ser Gy Asn Tyr Leu Asn Met Gin
GAC TCT CAA GSA GTA CTT TCT TCC TTT CCA GCT CCT CAG GCA -Asp Ser Qn Gly Val Leu Ser Ser Phe Pro Ala Pro Gin Ala
FIGURE 12. Interleukin-1 beta converting enzyme (ICE) pro-duction in corneal cells. (A) Primary cultures of humanstromal fibroblast (HSF), corneal epithelial (HCE), and cor-neal endothelial (HCN) cells were evaluated for the produc-tion of ICE mRNA by reverse-transcription-polymerasechain reaction (RT-PCR). Each corneal cell type yieldedPCR product of the expected size for ICE (424 bp). Theexpected product was not, however, detected in one epithe-lial culture (donor 1). An alternative band (361 bp) wasdetected in each cell. Note the variability of the relativelevels of the expected and alternative products within indi-vidual cell cultures. (B) Partial sequences of the previouslyreported and alternative ICE mRNAs and expected transla-tion products. The 63-nucleotide sequence deleted withindie alternative 361 bp ICE mRNA amplification is under-lined. Note that this deletion will not produce a frame shiftin the sequence beyond the deletion and, therefore, theamino acid residues before and after the deletion shouldbe identical to the previously reported ICE precursor.21
could have a role in mediating apoptosis of underlyingkeratocytes in response to such injury.3 Autocrinefunction within the epithelium and endothelium alsois possible because these cells express both Fas andFas ligand. Recent experiments have demonstratedthat the Fas-stimulating antibody will trigger death ofprimary human corneal epithelial or endothelial cells,and the death of these cells is accompanied by elec-tron microscopic morphologic changes consistentwith apoptosis (Mohan R, Wilson SE, unpublisheddata, 1996). Brunner et al37 demonstrated simultane-ous expression of Fas and Fas ligand on T-cell hybrid-oma cells. These investigators showed that Fas and Fasligand interaction, resulting in apoptosis, could occuron a single cell. It is, however, unclear how these inter-actions occur on a single cell.37
Griffith and coworkersM also recently reportedthe expression of Fas ligand protein in corneal epithe-lial and endothelial cells, as well as many other cellsof the eye. Similar to our results, these investigatorsdid not detect Fas ligand production in keratocytecells. They suggested that Fas ligand produced by ocu-lar cells could stimulate apoptosis of inflammatorycells expressing Fas and that these interactions had arole in maintaining immune privilege within the cor-nea and other areas of the eye.38 Although their data

Apoptosis-Related Modulators 1591
convincingly demonstrate that these types of interac-tions may occur between corneal and immune cells,our demonstration of Fas mRNA and protein expres-sion in each corneal cell type and stimulation ofapoptosis in stromal fibroblasts by anti-Fas-stimulatingantibody suggests that the Fas-Fas ligand system couldbe involved in regulation of corneal cells.
The current study also demonstrated that mRNAcoding for several mediators (Bax, BCL-2, BCL-XL,and ICE) of the common final pathway of apoptosis(Fig. 1) are expressed in corneal epithelial, stromalfibroblast, and endothelial cells in primary culture.We have discovered a smaller alternative PCR amplifi-cation product for ICE that also is expressed in eachcorneal cell type. Nucleic acid sequencing revealedthat the expected and alternative ICE amplificationproducts were identical, except that the latter had aninternal 63 bp deletion corresponding to amino acids92 to 112 of the precursor protein (Fig. 11B). Theseresidues do not contribute to the mature ICE protein,which is composed of two subunits containing residues120 to 297 and 317 to 404 of the precursor.21 It isunknown whether the corresponding 21-amino aciddeletion from the ICE precursor protein contains sig-naling or other information that might have func-tional relevance to the corresponding alternative pro-tein. If the Bax, Bcl-2, Bcl-XL, and ICE proteins areexpressed in each corneal cell type, each cell is likelyto be competent to undergo apoptosis in response toappropriate signals. Studies that identify cell-specificsignals activating the final common pathway ofapoptosis are likely to provide important insights intothe normal physiology and pathophysiology of cornealcells.
Key Words
apoptosis, Bax, Bcl-2, cornea, Fas, Fas ligand, interleukin-1converting enzyme
Acknowledgments
The authors thank Alisdar McDowell and John Gabrovsekfor their expert assistance with electron microscopy.
References
1. Cohen JJ. Apoptosis: Physiologic cell death. J Lab ClinMed. 1994;124:761-765.
2. Hoffman B, Liebermann DA. Molecular controls ofapoptosis: Differentiation/growth arrest primary re-sponse genes, proto-oncogenes, and tumor suppressergenes as positive and negative modulators. Oncogene.1994;9:1807-1812.
3. Wilson SE, He Y-G, Weng J, Li Q, Vital M, ChwangEL. Epithelial injury induces keratocyte apoptosis: Hy-pothesized role for the interleukin-1 system in themodulation of corneal tissue organization. Exp Eye Res.1996;62:325-338.
4. Nakayasu K. Stromal changes following removal of
epithelium in rat cornea. Jpn J Ophthalmol. 1988;32:113-125.
5. Crosson CE. Cellular changes following epithelialabrasion. In: Beuerman RW, Crosson CE, KaufmanHE, eds. Healing Processes in the Cornea. Houston: GulfPublishing; 1989:3-14.
6. Campos M, Szerenyi K, Lee M, McDonnell JM,McDonnell PJ. Keratocyte loss after corneal deepithe-lialization in primates and rabbits. Arch Ophthalmol.1994; 112:254-260.
7. Hengartner MO, Horvitz HR. Programmed cell deathin Caenorhabditis elegans. Curr Opin Genet Dev. 1994;4:581-586.
8. Steller H. Mechanisms and genes of cellular suicide.Science. 1995; 267:1445-1449.
9. White K, Grether ME, Abrams JM, Young L, FarrellK, Steller H. Genetic control of programmed celldeath in Drosophila. Science. 1994;264:677-683.
10. Miura M, Zhu H, Rotello R, Hartwieg E, YuanJ. Induc-tion of apoptosis in fibroblasts by IL-1 beta-convertingenzyme, a mammalian homolog of the C. elegans celldeath gene ced-3. Cell. 1993; 75:653-660.
11. Tsujimoto Y, Croce CM. Analysis of the structure, tran-scripts, and protein products of bcl-2, the gene in-volved in human follicular lymphoma. Proc Natl AcadSci USA. 1986; 83:5214-5218.
12. Boise LH, Gonzalez-Garcia M, Postema CE, et al. Bcl-X, a bcl-2-related gene that functions as a dominantregulator of apoptotic cell death. Cell 1993; 74:597-608.
13. Oltvai ZN, Milliman CL, Korsmeyer SJ. Bcl-2 hetero-dimerizes in vivo with a conserved homolog, Bax, thataccelerates programed cell death. Cell. 1993; 74:609-619.
14. Wilson SE, He Y-G, Lloyd SA. EGF, EGF receptor,basic FGF, TGF beta-1, and interleukin-1 alpha mRNAin human corneal epithelial cells and stromal fibro-blasts. Invest Ophthalmol Vis Sci. 1992; 33:1756-1765.
15. Wilson SE, Schultz GS, Chegini N, WengJ, He Y-G.Epidermal growth factor, transforming growth factoralpha, transforming growth factor beta, acidic fibro-blast growth factor, basic fibroblast growth factor, andinterleukin-1 proteins in the cornea. Exp Eye Res.1994; 59:63-70.
16. Wilson SE, Lloyd SA, He Y-G. Glucocorticoid receptorand interleukin-1 receptor messenger RNA expressionin corneal cells. Cornea. 1994; 13:4-8.
17. Fabre EJ, Bureau J, Pouliquen Y, Lorans G. Bindingsites for human interleukin 1 alpha, gamma inter-feron and tumor necrosis factor on cultured fibro-blasts of normal cornea and keratoconus. Curr Eye Res.1991; 10:585-592.
18. Taverne J. Transgenic mice in the study of cytokinefunction. Int J Exp Pathol. 1993; 74:525-546.
19. Wilson SE, Walker JW, Chwang EL, He Y-G. Hepato-cyte growth factor (HGF), keratinocyte growth factor(KGF), their receptors, FGF receptor-2, and the cellsof the cornea. Invest Ophthalmol Vis Sci. 1993; 34:2544-2561.
20. Ng S-Y, Gunning P, Eddy R, et al. Evolution of thefunctional neta-actin gene and its multi-pseudogenefamily: Conservation of noncoding regions and chro-

1592 Investigative Ophthalmology & Visual Science, July 1996, Vol. 37, No. 8
mosomal dispersion of pseudogenes. Mol Cell Biol.1985; 5:2720-2732.
21. Cerretti DP, Hollingsworth LT, Kozlosky CJ, et al. Mo-lecular characterization of the gene for human in-terleukin-1 beta converting enzyme (IL1BC). Geno-mics. 1994; 20:468-473.
22. Itoh N, Yonehara S, Ishli A, et al. The polypeptideencoded by the cDNA for human cell surface antigenFas can mediate apoptosis. Cell. 1991;66:233-243.
23. Cheng J, Liu C, Koopman WJ, Mountz JD. Characteriza-tion of human Fas gene: Exon/intron organization andpromoter region. J Immunol. 1995; 154:1239-1245.
24. Alderson M. Fas ligand mediates activation inducedcell death in human T lymphocytes. / Exp Med.1995;181:7l-77.
25. Takahashi T, Tanaka M, Inazawa J, Abe T, Suda T,Nagata S. Human Fas ligand: Gene structure, chromo-somal location and species specificity. Int Immunol.1994;6:1567-1574.
26. Zhivotovsky B, Cedervall B, Jiang S, Nicotera P, Orren-ius S. Involvement of Ca2+ in the formation of highmolecular weight DNA fragments in thymocyteapoptosis. Biochem Biophys Res Commun. 1994; 202:120-127.
27. Michel-Salmin LA, Tosi-Couture E, Gautier A,McDowall AW, Dubochet J. Electron microscopy ofthe chromosomes of dinoflagellate Porocentrum viicans:Confirmation of bouligands liquid crystal hypothesis./ Ultrastruct Moke Struct Res. 1986; 97:10-30.
28. Zakeri ZF, Quaglino D, Latham T, Lockshin RA. De-layed internucleosomal DNA fragmentation in pro-grammed cell death. FASEBJ. 1993; 7:470-478.
29. Fini ME, Strissel KJ, Girard MT, Mays JW, RinehartWB. Interleukin 1 alpha mediates collagenase synthe-
sis stimulated by phorbol 12-myristate 13-acetate.Chem. 1994; 269:11291-11298.
30. Girard MT, Matsubara M, Fini ME. Transforminggrowth factor-beta and interleukin-1 modulate metal-loproteinase expression by corneal stromal cells. InvestOphthalmol Vis Sci. 1991; 32:2441-2454.
31. Nagata S, Golstein P. The Fas death factor. Science.1995;267:1449-1456.
32. Ni R, Tomita Y, Matsuda K, et al. Fas-mediatedapoptosis in primary cultured mouse hepatocytes. ExpCell Res. 1994; 215:332-337.
33. Oishi M, Maeda K, Sugiyama S. Distribution ofapoptosis-mediating Fas antigen in human skin andeffects of anti-Fas monoclonal antibody on human epi-dermal keratinocyte and squamous cell carcinoma celllines. Arch Dermatol Res. 1994;286:396-407.
34. Sayama K, Yonehara S, Watanabe Y, Miki Y. Expressionof Fas antigen on keratinocytes in vivo and inductionof apoptosis in cultured keratinocytes. / Invest Derma-tol. 1994; 103:330-334.
35. Tanaka M, Suda T, Takahashi T, Nagata S. Expressionof the functional soluble form of human fas ligand inactivated lymphocytes. EMBOJ. 1995; 14:1129-1135.
36. Kimura K, Wakatsuki T, Yamamoto M. A variantmRNA species encoding a truncated form of Fas anti-gen in the rat liver. Biochem Biophys Res Commun.1994; 198:666-674.
37. Brunner T, Mogil RJ, LaFace D, et al. Cell-autono-mous Fas (CD95)/Fas-ligand interaction mediates ac-tivation-induced apoptosis in T-cell hybridomas. Na-ture. 1995; 373:441-444.
38. Griffith TS, Brunner T, Fletcher SM, Green DR, Fer-guson TA. Fas ligand-induced apoptosis as a mecha-nism of immune privilege. Science. 1995; 270:1189-1192.
Copyright © 2022 FDOKUMEN




















