
Gene therapy in the Cornea: 2005–present
22
Gene therapy in the Cornea: 2005epresent Rajiv R. Mohan a, b, c, * , Jonathan C.K. Tovey a, b , Ajay Sharma a, b , Ashish Tandon a, b a Harry S. Truman Memorial Veterans’ Hospital, 800 Hospital Drive, Columbia, MO 65201, USA b Mason Eye Institute, University of Missouri, 1 Hospital Drive, Columbia, MO 65212, USA c College of Veterinary Medicine, University of Missouri, 1600 East Rollins Street, Columbia, MO 65212, USA article info Article history: Available online 28 September 2011 Keywords: Cornea Gene therapy AAV Nanoparticles Decorin Corneal scarring Corneal neovascularization abstract Successful restoration of vision in human patients with gene therapy affirmed its promise to cure ocular diseases and disorders. The efficacy of gene therapy is contingent upon vector and mode of therapeutic DNA introduction into targeted cells/tissues. The cornea is an ideal tissue for gene therapy due to its ease of access and relative immune-privilege. Considerable progress has been made in the field of corneal gene therapy in last 5 years. Several new gene transfer vectors, techniques and approaches have evolved. Although corneal gene therapy is still in its early stages of development, the potential of gene-based interventions to treat corneal abnormalities has begun to surface. Identification of next generation viral and nanoparticle vectors, characterization of delivered gene levels, localization, and duration in the cornea, and significant success in controlling corneal disorders, particularly fibrosis and angiogenesis, in experimental animal disease models, with no major side effects have propelled gene therapy a step closer toward establishing gene-based therapies for corneal blindness. Recently, researchers have assessed the delivery of therapeutic genes for corneal diseases and disorders due to trauma, infections, chemical, mechanical, and surgical injury, and/or abnormal wound healing. This review provides an update on the developments in gene therapy for corneal diseases and discusses the barriers that hinder its utilization for delivering genes in the cornea. Published by Elsevier Ltd. Contents 1. Introduction ....................................................................................................................... 44 2. Gene therapy vehicles for the cornea ................................................................................................. 44 2.1. Viral vectors .................................................................................................................. 44 2.1.1. Adenovirus ........................................................................................................... 44 2.1.2. Adeno-associated virus ................................................................................................. 45 2.1.3. Retrovirus and lentivirus ................................................... ............................................ 46 2.2. Nonviral vectors .............................................................................................................. 47 2.2.1. Microinjection technique ................................................... ............................................ 47 2.2.2. Electroporation ........................................................................................................ 47 2.2.3. Sonoporation .......................................................................................................... 48 2.2.4. Gene gun ............................................................................................................ 48 2.2.5. Controlled corneal dehydration ................................................ ......................................... 48 2.2.6. Laser ................................................................................................................. 49 2.2.7. Chemicals ............................................................................................................ 49 2.3. Next generation AAV and nanoparticle vectors .................................................................................... 49 2.3.1. Tyrosine-mutant AAV vectors ................................................. .......................................... 49 2.3.2. Nanoparticle vectors ................................................................................................... 50 2.3.3. Dendrimer vectors ..................................................................................................... 52 3. Tissue-selective gene therapy tools ..................................................... ............................................. 52 * Corresponding author. Mason Eye Institute, School of Medicine, University of Missouri-Columbia,1 Hospital Dr., Columbia, MO 65212, USA. Tel.: þ1 573 884 1449; fax: þ1 573 814 6551. E-mail address: [email protected] (R.R. Mohan). Contents lists available at SciVerse ScienceDirect Progress in Retinal and Eye Research journal homepage: www.elsevier.com/locate/prer 1350-9462/$ e see front matter Published by Elsevier Ltd. doi:10.1016/j.preteyeres.2011.09.001 Progress in Retinal and Eye Research 31 (2012) 43e64
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Gene therapy in the Cornea: 2005–present

at SciVerse ScienceDirect
Progress in Retinal and Eye Research 31 (2012) 43e64
Contents lists available
Progress in Retinal and Eye Research
journal homepage: www.elsevier .com/locate/prer
Gene therapy in the Cornea: 2005epresent
Rajiv R. Mohan a,b,c,*, Jonathan C.K. Tovey a,b, Ajay Sharma a,b, Ashish Tandon a,b
aHarry S. Truman Memorial Veterans’ Hospital, 800 Hospital Drive, Columbia, MO 65201, USAbMason Eye Institute, University of Missouri, 1 Hospital Drive, Columbia, MO 65212, USAcCollege of Veterinary Medicine, University of Missouri, 1600 East Rollins Street, Columbia, MO 65212, USA
a r t i c l e i n f o
Article history:Available online 28 September 2011
Keywords:CorneaGene therapyAAVNanoparticlesDecorinCorneal scarringCorneal neovascularization
* Corresponding author. Mason Eye Institute, Schoofax: þ1 573 814 6551.
E-mail address: [email protected] (R.R
1350-9462/$ e see front matter Published by Elseviedoi:10.1016/j.preteyeres.2011.09.001
a b s t r a c t
Successful restoration of vision in human patients with gene therapy affirmed its promise to cure oculardiseases and disorders. The efficacy of gene therapy is contingent upon vector and mode of therapeuticDNA introduction into targeted cells/tissues. The cornea is an ideal tissue for gene therapy due to its easeof access and relative immune-privilege. Considerable progress has been made in the field of cornealgene therapy in last 5 years. Several new gene transfer vectors, techniques and approaches have evolved.Although corneal gene therapy is still in its early stages of development, the potential of gene-basedinterventions to treat corneal abnormalities has begun to surface. Identification of next generationviral and nanoparticle vectors, characterization of delivered gene levels, localization, and duration in thecornea, and significant success in controlling corneal disorders, particularly fibrosis and angiogenesis, inexperimental animal disease models, with no major side effects have propelled gene therapy a stepcloser toward establishing gene-based therapies for corneal blindness. Recently, researchers haveassessed the delivery of therapeutic genes for corneal diseases and disorders due to trauma, infections,chemical, mechanical, and surgical injury, and/or abnormal wound healing. This review provides anupdate on the developments in gene therapy for corneal diseases and discusses the barriers that hinderits utilization for delivering genes in the cornea.
Published by Elsevier Ltd.
Contents
1. Introduction . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 442. Gene therapy vehicles for the cornea . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 44
2.1. Viral vectors . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 442.1.1. Adenovirus . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 442.1.2. Adeno-associated virus . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 452.1.3. Retrovirus and lentivirus . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 46
2.2. Nonviral vectors . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 472.2.1. Microinjection technique . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 472.2.2. Electroporation . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 472.2.3. Sonoporation . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 482.2.4. Gene gun . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 482.2.5. Controlled corneal dehydration . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 482.2.6. Laser . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 492.2.7. Chemicals . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 49
2.3. Next generation AAV and nanoparticle vectors . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 492.3.1. Tyrosine-mutant AAV vectors . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 492.3.2. Nanoparticle vectors . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 502.3.3. Dendrimer vectors . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 52
3. Tissue-selective gene therapy tools . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 52
l of Medicine, University of Missouri-Columbia, 1 Hospital Dr., Columbia, MO 65212, USA. Tel.: þ1 573 884 1449;
. Mohan).
r Ltd.

R.R. Mohan et al. / Progress in Retinal and Eye Research 31 (2012) 43e6444
3.1. Vector engineering . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 523.2. Custom vector-delivery techniques . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 53
4. Gene therapy updates in various corneal disorders . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 544.1. Corneal graft rejection . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 544.2. Corneal scarring and wound healing . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 554.3. Corneal neovascularization . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 564.4. Corneal alkali burn . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 574.5. Other corneal disorders . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 58
5. Lessons learned from corneal genetic studies: humans and animals . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 596. Ocular gene therapy clinical trials: current status . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 597. Future directions: regenerative medicine, nanomedicine and other approaches . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 598. Conclusions . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 60
Acknowledgments . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 61References . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 61
1. Introduction
Gene therapy has advanced in leaps and bounds since itsintroduction to medicine almost 20 years ago as a query on genetherapy in the United States National Library of Medicine databasePubMed yields over 128,000 articles from almost all medicaldisciplines. In the field of ophthalmology, gene therapy has shownastounding success. Recently, a breakthrough was attained in therestoration of vision in human patients suffering from Leber’scongenital amaurosis using gene replacement therapy in the retinalpigment epithelium (Bainbridge et al., 2008; Hauswirth et al., 2008;Maguire et al., 2008, 2009; Simonelli et al., 2010). This plainly givescredence to the potential of gene therapy to cure diseases of theocular system and prevent blindness.
Current treatments for corneal disorders include various phar-macological agents and surgical approaches depending on the cause,extent, and type of corneal damage. Conventional drug therapies totreat corneal disorders provide only short-term relief, requirerepeated application, cause side effects, and are often ineffective.More importantly they do not correct the cause of the problem butmerelysuppress symptoms.Conversely, gene therapyfixes the rootofthe problem, provides long-term cure, and does not require repeatedapplications or clinic visits. However, at present no gene therapymodalities are available for corneal diseases. The cornea is an idealtissue for gene therapy due to its ease of access and relative immune-privilege. As it is a transparent tissue and devoid of blood vessels, itcan be readilymonitored visually. These properties also allow for theadministration of therapeutic genes into corneal cells with relativeease. The cornea is well suited for conducting ex vivo gene therapystrategies because it can bemaintained in culture for a long time. Thearduous task of developing novel gene-based modalities for thecornea has greatly improved due to increased comprehension ofacquired and inherited corneal diseases in terms of molecularmechanisms and pathogenesis. Numerous approaches employingdifferent viral andnonviralvectors and techniques to introducegenesinto the cornea in vitro, ex vivo, and in vivo have been tested. Amongviral vectors, adenovirus, adeno-associated virus (AAV), retrovirus,and lentivirus vectors have been found to efficiently transport genesinto corneal tissue. However, concerns over safety and immunoge-nicityhave limited theiruse.Nonviralvectors includingplasmidDNA,lipids, polymers, and nanoparticles are generally safe but often foundless efficient than their viral counterparts. Various physical tech-niques such as topical administration, gene gun, electroporation,intrastromal injection, and iontophoresis have beenused to augmentdelivery of both viral and nonviral vectors. However, none of thesevectors or techniques is ideal, and each has its benefits and short-comings. Hereinwe provide a comprehensive reviewof corneal gene
therapy approaches tested in the past six years and a brief generaloverview of vectors. A detailed overview of gene therapy vectors andtheir mode of action can be found in our previous corneal genetherapy review (Mohan et al., 2005).
2. Gene therapy vehicles for the cornea
2.1. Viral vectors
Viruses have been used since the dawn of gene transfer tech-nology to deliver genes into different cells and tissues. Viruses wereused as a vector in about 70% of gene therapy clinical trials (Younget al., 2006). Adenovirus (AV), adeno-associated virus (AAV),retrovirus, and lentivirus have been found to efficiently transportgenes into the cornea. Nevertheless, each of these vectors has itsown limitations. Adenovirus and retrovirus can successfully delivergenes into the cornea for short periods of time with mild-to-severeinflammatory responses. However, both of these vectors are oflimited use for corneal gene therapy because of their inability totransduce low/non-dividing cells such as corneal endothelium andkeratocytes, and induction of immune reactions. AAV and disabledlentivirus vectors offer better alternatives for delivering genes intocorneal keratocytes and endothelium because of their ability totransduce slow/non-dividing cells and ability to provide long-termtransgene expression. The origin of lentivirus vectors (equineinfectious anemia virus and HIV) remains a major concern andsignificantly dampens enthusiasm for its use in human patients.Among viral vectors, AAV appears to be a good choice for cor-neal gene therapy because of its potency and safety profile.Recombinant AAV vectors have shown great promise for oculargene therapy and restoring vision in patients with no major sideeffects.
2.1.1. AdenovirusRecombinant forms of AV have been engineered and utilized in
gene transfer studies in the past (Mohan et al., 2005). In sum, first-generation AV vectors lack the E1 gene region rendering themunable to replicate although they can proliferate in cell lines thatprovide E1 gene product. Second-generation AV vectors lack E1, E2,and E4 viral genes leading to less immunogenicity than first-generation vectors. In third-generation AV vectors, the AV viralgenome is absent and only ITR sequences and packaging genes arepresent thus giving the name gutless or high-capacity vectors.Third-generation AV vectors are able to carry larger gene insertsand are less immunogenic but require a helper virus. In order toreduce helper virus contamination, Cre/lox-P system and 293 cellsstably transfected with Cre recombinase are utilized and have been

R.R. Mohan et al. / Progress in Retinal and Eye Research 31 (2012) 43e64 45
shown to decrease helper virus contamination to <0.01% (Palmerand Ng, 2005). However, even this minimal level of contamina-tion may be detrimental in clinical scenarios where large doses arerequired.
Multiple studies have examined the efficiency of AV vectors forcorneal gene therapy (Mohan et al., 2005). AV vector was foundmost efficient in transducing murine cornea compared to equineimmunodeficiency, lenti and bacculovirus vectors (Beutelspacheret al., 2005). Recently, AV vectors have been shown to delivergenes successfully into mouse corneal stroma and endotheliumunder control of cytomegalovirus immediate-early promoter(CMV) (Yu et al., 2007). The Ritter group has also reported cytokineor growth factor gene transfer with AV vector in rodent corneas(Ritter et al., 2007; Gong et al., 2007a,b). AV vectors provide short-lived transgene expression and also demonstrate moderate tosevere immune reaction apart from other side effects (Yu et al.,2007; Ritter et al., 2007; Gong et al., 2007a,b; Sharma et al.,2010a). Recombinant AV vectors have potential use in cornealgene therapy to over-express proteins in the corneal epitheliumparticularly in scenarios such as diabetes mellitus where transientgene expression is desirable. However, the ephemeral nature of AVvectors discourages its application in gene-based treatment forinherited gene defects (Saghizadeh et al., 2010).
2.1.2. Adeno-associated virusAmong 110 identified AAV serotypes, to date only serotypes
1e10 have been used for gene therapy. AAV vectors have demon-strated high transduction efficiency and long-term transgeneexpression in the retina, cornea, and many non-ocular tissuesin vivo (Alexander and Hauswirth, 2008; Gao et al., 2005; Suraceand Auricchio, 2008 and references therein). AAV2 has beentested more in depth compared to other AAV serotypes for genetherapy although each serotype has shown unique transductionpatterns for various cells/tissues. AAV6, -8 and -9, especially, havethe ability to mediate whole-body gene transfer efficiently (Ghoshet al., 2007; Pacak et al., 2006). The variation in gene transfer bydifferent AAV serotypes is likely due to the interactions betweenhost-cell receptor and viral capsid (Van Vliet et al., 2008). Structuralvariations in the capsid region of different AAV serotypes enableeach serotype to bind different cell surface receptors. For instance,AAV serotypes 4, 5, and 6 use sialic acid (Wu et al., 2006) whereasAAV 8 and 9 use laminin receptors to enter cells (Akache et al.,2006). This led to the development of hybrid AAV vectors asa variety of pseudopackaged vectors were produced using trans-encapsidation of the AAV2 genome into the capsid of AAV1e9.These next generation hybrid vectors AAV2/1e9, displayed muchgreater transduction efficiency compared to AAV2/2 in oculartissues of various animal models (Surace and Auricchio, 2008). It isimportant to note that a great deal of variation in transductionefficiency, tissue preference, first transgene appearance timing, andduration was observed in studies performed with various hybridrecombinant AAV vectors (Surace and Auricchio, 2008). Lebherzet al. (2008) also evaluated the gene delivery efficiency andexpression stability of AAV2/7, 2/8, 2/9, with AAV2/1, 2/2, 2/5 usingenhanced green fluorescent protein (EGFP) and rhesus erythro-poietin as transgenes. After vector introduction into the eye viaintravitreal and subretinal injection, the authors demonstrated thatAAV2/7 and 2/8 possess superior long-term transduction ability (upto 6 months) in both retinal and anterior chamber tissues includingthe iris, trabecular meshwork, and cornea, (Lebherz et al., 2008).Subsequent studies with AAV2/6, AAV2/7, AAV2/8, and AAV2/9vectors further reinforced these findings for retinal tissue (Suraceand Auricchio, 2008). In a similar fashion, AAV2/8 and 2/9demonstrated 5e100 fold superior transduction efficiency in non-ocular tissues such as heart (Wang et al., 2005), brain (Broekman
et al., 2006), skeletal muscle (Wang et al., 2005), lung (Limberisand Wilson, 2006), and liver (Gao et al., 2006). We compared theefficiency of AAV2/6, AAV2/8, and AAV2/9 serotypes for cornealgene delivery using in vitro, in vivo, and ex vivo models to testa hypothesis that the relative efficiency of AAV-mediated genetransfer in the cornea depends on serotype. In our first study, thecellular tropism and transduction efficiency of AAV2/6, AAV2/8, andAAV2/9 vectors encoding alkaline phosphatase (AP) were testedusing primary human corneal fibroblasts cultures. We found thetested serotypes efficiently transduced human corneal fibroblastsbut they differed in transduction efficiency. AAV2/6 displayed30e50 fold higher transduction efficiency compared to AAV2/8 orAAV2/9 (Fig. 1). A remarkably positive and important finding wasthat none of the tested AAV serotypes induced significant cell deathor loss of cellular viability reaffirming that AAV vectors are safe forthe cornea (Sharma et al., 2010b). Interestingly, our subsequentstudies with these vectors using mouse cornea in vivo and humancornea ex vivo showed very different transduction profilescompared to our in vitro investigations (Sharma et al., 2010c).Contrary to in vitro findings, the order of transduction efficiency forthe three tested vectors was found to be AAV2/9�AAV2/8>AAV2/6 (Fig. 2). These findings are not completely surprising as pastliterature reports imply that in vitro gene delivery may differ fromin vivo transduction (Lipkowitz et al., 1999; Richter et al., 2000).However, the important take home message from these studies isthat in vivo testing of gene therapy vectors for the cornea isimportant before making any conclusive inferences about vectorefficacy. The one thing that remained unchanged during in vitro,ex vivo and in vivo testing was the safety profile of tested AAVserotypes. None of the three tested serotypes caused any significantside effects such as cell death, loss of cellular viability, inflammationand/or noticeable immune reaction in the cornea as measured withTUNEL and CD11b or F4/80 immunostaining (Fig. 3). Recently,transgene expression in the corneal epithelium was detected up to8 months with AAV2 in the mouse eye suggesting that rAAV-delivered transgenes can persist for several months, and possiblyyears, in vivo (Alexander and Hauswirth, 2008; Lai et al., 2007).Thus, using AAV2/5 vector and a mouse model we performed time-dependent long-term animal experiments to characterize deliveredtransgene expression and duration for the cornea in vivo. Ourongoing studies showed different levels of transgene expression inthe cornea in vivo up to 10 months following topical vector appli-cation (Mohan et al., unpublished data).
The alterations in function, host range, and tissue tropismshown by different AAV serotypes may be due to a degree ofsequence variation within the capsid region (Van Vliet et al., 2008).Furthermore, the differences in transduction profile are likelyrelated to distinct uptake and intracellular trafficking mechanismsof the various serotypes. In addition, different serotype-specific ITRelements may also influence transgene expression. Various studiesreport that the use of different serotypes or cross-packagingexpands the tissue tropism of rAAV vectors. Moreover, selectiveuse of serotypes might allow for targeting specific tissues. Forexample, an rAAV2 genome packaged in an AAV5 capsid (AAV2/5)transduced photoreceptors and retinal pigment epithelial cells inthe eye (Gao et al., 2005; Surace and Auricchio, 2008) whereasAAV2-encapsidated vector transduced primarily photoreceptors,and an AAV1-encapsidated vector transduced mainly retinalpigment epithelial cells. Additionally, the utilization of differentAAV serotypes may overcome the problems associated with vectorre-administration and preexisting immunity to rAAV2 in humans(Adriaansen et al., 2006).
AAV vectors have emerged to the forefront in ocular genetherapy as they have been used in a ground-breaking clinical trial totreat retinal diseases. Despite the proven potency and safety of AAV
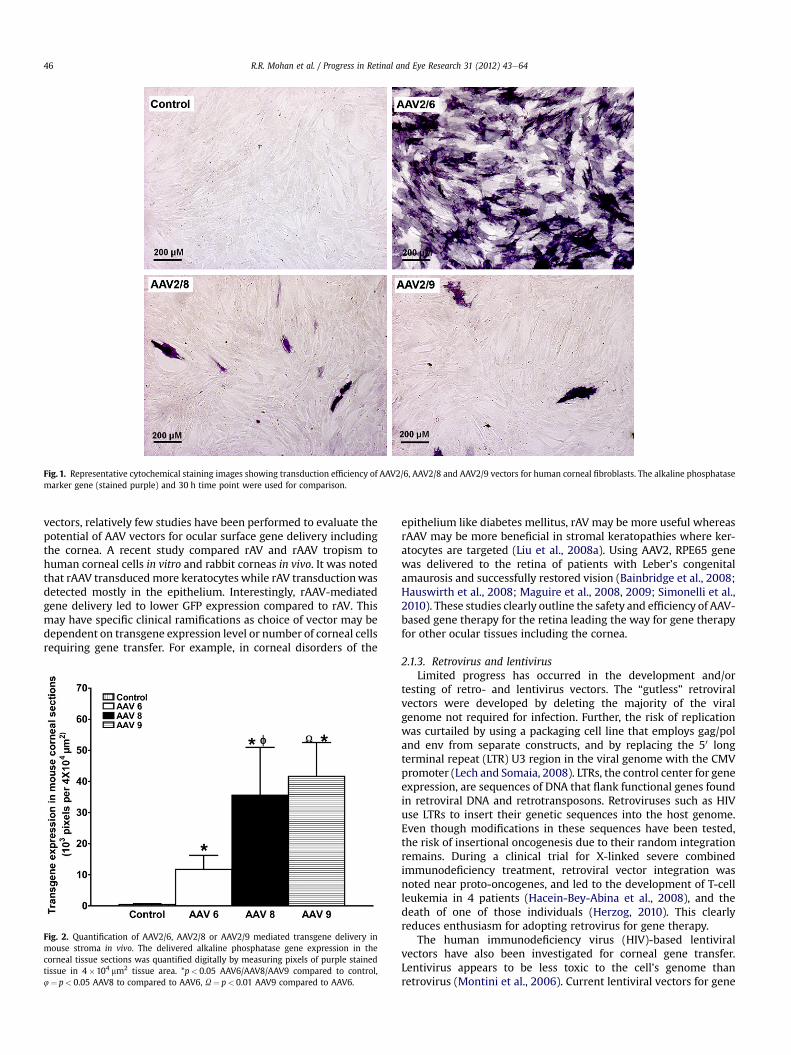
Fig. 1. Representative cytochemical staining images showing transduction efficiency of AAV2/6, AAV2/8 and AAV2/9 vectors for human corneal fibroblasts. The alkaline phosphatasemarker gene (stained purple) and 30 h time point were used for comparison.
R.R. Mohan et al. / Progress in Retinal and Eye Research 31 (2012) 43e6446
vectors, relatively few studies have been performed to evaluate thepotential of AAV vectors for ocular surface gene delivery includingthe cornea. A recent study compared rAV and rAAV tropism tohuman corneal cells in vitro and rabbit corneas in vivo. It was notedthat rAAV transducedmore keratocytes while rAV transductionwasdetected mostly in the epithelium. Interestingly, rAAV-mediatedgene delivery led to lower GFP expression compared to rAV. Thismay have specific clinical ramifications as choice of vector may bedependent on transgene expression level or number of corneal cellsrequiring gene transfer. For example, in corneal disorders of the
Fig. 2. Quantification of AAV2/6, AAV2/8 or AAV2/9 mediated transgene delivery inmouse stroma in vivo. The delivered alkaline phosphatase gene expression in thecorneal tissue sections was quantified digitally by measuring pixels of purple stainedtissue in 4�104 mm2 tissue area. *p< 0.05 AAV6/AAV8/AAV9 compared to control,4¼ p< 0.05 AAV8 to compared to AAV6, U¼ p< 0.01 AAV9 compared to AAV6.
epithelium like diabetes mellitus, rAV may be more useful whereasrAAV may be more beneficial in stromal keratopathies where ker-atocytes are targeted (Liu et al., 2008a). Using AAV2, RPE65 genewas delivered to the retina of patients with Leber’s congenitalamaurosis and successfully restored vision (Bainbridge et al., 2008;Hauswirth et al., 2008; Maguire et al., 2008, 2009; Simonelli et al.,2010). These studies clearly outline the safety and efficiency of AAV-based gene therapy for the retina leading the way for gene therapyfor other ocular tissues including the cornea.
2.1.3. Retrovirus and lentivirusLimited progress has occurred in the development and/or
testing of retro- and lentivirus vectors. The “gutless” retroviralvectors were developed by deleting the majority of the viralgenome not required for infection. Further, the risk of replicationwas curtailed by using a packaging cell line that employs gag/poland env from separate constructs, and by replacing the 50 longterminal repeat (LTR) U3 region in the viral genome with the CMVpromoter (Lech and Somaia, 2008). LTRs, the control center for geneexpression, are sequences of DNA that flank functional genes foundin retroviral DNA and retrotransposons. Retroviruses such as HIVuse LTRs to insert their genetic sequences into the host genome.Even though modifications in these sequences have been tested,the risk of insertional oncogenesis due to their random integrationremains. During a clinical trial for X-linked severe combinedimmunodeficiency treatment, retroviral vector integration wasnoted near proto-oncogenes, and led to the development of T-cellleukemia in 4 patients (Hacein-Bey-Abina et al., 2008), and thedeath of one of those individuals (Herzog, 2010). This clearlyreduces enthusiasm for adopting retrovirus for gene therapy.
The human immunodeficiency virus (HIV)-based lentiviralvectors have also been investigated for corneal gene transfer.Lentivirus appears to be less toxic to the cell’s genome thanretrovirus (Montini et al., 2006). Current lentiviral vectors for gene

R.R. Mohan et al. / Progress in Retinal and Eye Research 31 (2012) 43e64 47
therapy lack the entire viral genome, and contain only the pack-aging signal and the cis-acting elements, including the LTR. In anattempt to generate safer lentiviral vectors, the U3 region of the 30
LTR has been deleted. In recently generated lentiviral vectors, theendogenous promoter has been replaced with a CMV/LTR hybridpromoter in an effort to improve safety. Currently, glycoprotein ofvesicular stomatitis virus is the most commonly used env proteinfor lentiviral pseudotyping (Lech and Somaia, 2008). Not long ago itwas reported that transcriptionally active LTRs are chief contribu-tors to genotoxicity and that self-inactivating LTRs decrease theoncogenic potential thus improving the safety of lentiviral vectors(Montini et al., 2009). Clinically, lentiviral vectors have yielded noreports of serious adverse events (Williams and Coster, 2010).Selective studies have been performed to evaluate the efficacy oflentiviral vectors for corneal gene therapy during the past 6 years.Beutelspacher et al. (2005) compared self-inactivating HIV-1 andEquine Infectious Anemia Virus (EIAV) transduction efficiency inmurine, rabbit, and human corneal endothelial cells and found EIAVto be more effective than HIV-1 for corneal endothelial genetherapy. Another study reported that a single injection of HIV-based lentivirus into the anterior chamber of the rodent eye effi-ciently delivered transgene into the corneal endothelium (Challaet al., 2005). Parker et al. (2007) demonstrated 80e90% trans-duction of rat, bovine, and human corneal endothelial cells withlentiviral vector expressing enhanced yellow fluorescent proteinunder control of the Simian virus type 40 early promoter.Bemelmans et al. (2009) demonstrated efficient transduction ofkeratocytes up to 3 weeks in a pig model by injecting HIV-1expressing GFP into a stromal pocket created with a femtosecondlaser.
2.2. Nonviral vectors
The introduction of plasmid DNA expressing therapeutic genesinto target cells without the use of viruses falls under the broadcategory of nonviral gene transfer methods. Nonviral gene therapyis considered safer than viral gene therapy due to low toxicity,immunogenicity, and pathogenicity. Additionally, plasmid vectorproduction is straightforward and cost-effective. Nevertheless, lowtransfection efficiency is a major challenge. Many strategies havebeen developed and are in the preclinical pipeline to improveplasmid delivery in cells. A brief description of popular techniquesis given below.
Fig. 3. Representative images showing AAV9 effects on immune reaction (measured by Cd11treatment), few Cd11bþ and TUNELþ cells were observed in AAV-treated mouse stroma. Theapoptosis. AAV6 and AAV8 showed similar results (data not shown). Nuclei are stained blu
2.2.1. Microinjection techniqueIntroduction of plasmid via microinjection has led to successful
delivery of genes including GFP, interleukin (IL)18, Flt23k, endo-statin, MMP14, and vasohibin into various cells of the cornea (Kimet al., 2005; Jani et al., 2007; Lai et al., 2007; Galiacy et al., 2011;Zhou et al., 2010; Sharma A, et al. IOVS 2010;51:ARVO E-Abstract2839). Microinjections targeting different layers of the cornea havebeen performed at various anatomic locations and include intra-stromal, subconjunctival, and directly into the anterior chamber(Singh et al., 2005; Kim et al., 2005; Lai et al., 2007; Zhou et al.,2010; Yu et al., 2007; Sharma A, et al. IOVS 2010;51:ARVO E-Abstract 2839). A recent study performed in mice demonstratedthat bevacizumab treatment given via intraocular injection wasmore efficacious than the subconjunctival (periocular) route(Dratviman-Storobinsky et al., 2009). Intrastromal injection may beappropriate in the treatment of acute corneal diseases as short-lived gene expression was detected in the cornea following intra-stromal injection (Hao et al., 2010). Conversely, subconjunctivalinjection showed long-term stable transgene expression and mayhelp to avoid the endophthalmitis and cataract formation associ-ated with intracameral injection (Kuo et al., 2009). Microsurgicaltechniques also offer a feasible method of exploring gene functionin corneal disorders. For example, Kuo et al. (2008) showedsuccessful transgene delivery in the cornea in vivo using intra-stromal lamellar implantation of a partially dried p-bFGFeSAINT-18complex composed of SAINT-18 and plasmid vector encodingreporter or FGF2 gene. This corneal gene transfermethod permittedlocalized transgene delivery in the cornea. It is thought that exceptfor topical and subconjunctival administration, all other genedelivery strategies to the cornea are invasive and compromisecorneal integrity (Cheng et al., 2007).
2.2.2. ElectroporationElectroporation, also known as electrogenetherapy or electro-
permeabilization, makes use of high-intensity electrical pulses toform transient pores in the cell membrane and is useful for genedelivery in both cultured eye cells and ocular surface tissues in vivo.An advantage is large DNA constructs can be transported into cellsalthough specialized equipment is necessary (Williams and Coster,2010). However, very few studies have been reported in the corneasince 2005. Electroporation has the ability to deliver foreign genesinto the corneal epithelium as well as keratocytes (Zhou and Dean,2007). Electrical current of 200 V/cm did not cause trauma, corneal
b) and cell death (measured by TUNEL) in mouse corneas. Like control corneas (no AAVTUNEL þ cells detected in the corneal epitheliumwere because of its replenishment viae with DAPI.

R.R. Mohan et al. / Progress in Retinal and Eye Research 31 (2012) 43e6448
edema, or inflammation but introduced transgene at low levels.Higher electrical current resulted in enhanced gene transfer butalso led to considerable corneal damage. Electrical current cancause irreversible tissue damage as a result of thermal heating orCa2þ influx due to disruption of cell membranes. Electricallyassisted gene delivery to the endothelium of ex vivo human corneaswas recently described (He et al., 2010). Using custom-designedelectrodes, two reporter genes, EGFP and beta-galactosidase(bgal), were successfully transported into human corneas inorgan culture using eight 1-Hz 100-ms pulses of 125 mA squarecurrent. Although efficiency was much lower than viral vector, lowcell death and no remarkable change in tight junction integrity ofendothelial cells show its potential clinical application (He et al.,2010). The electrotransfer of glyceraldehyde-3-phosphate dehy-drogenase (GAPDH) small interfering RNA (siRNA) and dextranmacromolecules into mice corneal epithelium in vivo using ionto-phoresis and electroporation individually and in combination hasbeen reported (Hao et al., 2009). Although both iontophoresis andelectroporation independently delivered macromolecules into thecornea, iontophoresis was found to be more efficient and thecombination of iontophoresis followed by electroporation wasmore effective than both methods alone (Hao et al., 2009).
2.2.3. SonoporationSonoporation employs ultrasound waves to create pores in the
plasma membrane in order to deliver DNA to the nucleus. Ultra-sound is effective for cell transfection in vitro and in vivo (Liu et al.,2006). Transfection efficiency of this approach is dependent on thetransducer frequency, acoustic pressure, output strength, and pulseduration of ultrasound treatment in addition to the use of contrastagents such as microbubbles. Microbubbles were generated ascontrast agents to not only enhance imaging but also improve genedelivery efficiency by boosting cell permeability (Mitragotri, 2005).Ultrasound-targeted microbubble destruction may hold greatpotential as a site-specific gene transfer approach and has been usedsuccessfully in AAV-mediated gene transfection of human RPE cellsin vitro and rat retina in vivo (Li et al., 2009a,b). Microbubblesgenerally measure approximately 3 mm in diameter and arecomposed of a shell which houses a gas core. Available commer-cially, microbubbles differ in shell composition (i.e. albumin,galactose, lipid, polymers) and gas core (i.e. air, perfluorocarbon,nitrogen) (Lindner, 2004). A notable advantage of microbubbles isthat they may be targeted to specific areas of interest. One way toaccomplish this is the addition of a tiered polymer coat to the shelland subsequent covalent (i.e. carbodiimide-mediated amide) ornoncovalent (i.e. biotineavidin) coupling of targeting ligands to thepolymer coat (Klibanov, 2006). The precise mechanism by whichgene transfection is enhanced by ultrasound-mediated destructionof carrier microbubbles is not clear but may include shellfragmentation-induced cellular microporation and micro-environment changes brought about by high-velocity pressure jetsor heat and free radical production (Lindner, 2004). Ultrasoundfacilitates gene transfer byway of passive diffusion (Gao et al., 2007).The Sakamoto group demonstrated successful gene delivery intorabbit cornea in vitro and in vivo by combining ultrasound andmicrobubbles (Sonoda et al., 2006). The central cornea was injectedwith plasmid mixed with perflutren protein and a probe was placedon the ocular surface for ultrasound (1 MHz, 120 s, 50% duty cycle,1e2W/cm2) exposure. The ultrasound and microbubbles greatlyincreased transduction efficiency into keratocytes without tissuedamage. GFP was expressed in the stroma for up to 30 days and waslimited to the area exposed to ultrasound (Sonoda et al., 2006).However, multiple safety concerns of microbubble use includinginstability in serum and infection risk of components such as humanalbumin limit its use. Subsequent studies entailed the utilization of
ultrasound with a newly developed contrast agent, a bubble lipo-some made of polyethylenglycol modified liposome containingperfluoropropane gas. Sonoporation coupled with the bubble lipo-some showed enhanced transgene delivery into rabbit cornealepithelial cells and rat subconjunctiva (Yamashita et al., 2007).Although these studies outline the potential of sonoporation forcorneal gene therapy, many parameters remain to be investigated.
2.2.4. Gene gunGene gun is a ballistic (also called bioballistic) gene transfer
method. It utilizesmicron-sized biologically inert heavymetal (gold,silver or tungsten) particles and mechanical or macroprojectile(centripetal, magnetic or electrostatic) force. The bombarding ofDNA-coated particles on cells/tissues with high velocity results ingene transfer. Gene delivery with gene gun depends onmany factorssuch as amount of DNA-coated on particles, temperature, amount ofcells, amount of force, number of DNA-coated particles, etc. Shallowpenetration of particles, substantial cell damage, uncontrolled genetransfer, high cost, access to internal organs, etc. are few amongmany limitations of this method. Successful gene delivery in thecornea with gene gun has been reported (Hao et al., 2010). Genessuch as IL4 and IL10 plasmid DNA and opioid growth factor receptor(OGFr) have been introduced into the corneal epithelium with thismethod (Bauer et al., 2006; Zagon et al., 2006). Zagon and cohortsshowed delayed corneal abrasion healing after gene gun-mediateddelivery of sense OGFr cDNA in rat eyes. Conversely, they foundthat antisense OGFr cDNA over-expression by the same deliverymethod led to accelerated corneal wound healing (Zagon et al.,2006). This study established the autocrine behavior of the OGF-OGFr axis, its regulatory role in ocular surface wound healing, andits potential in gene therapy approaches for corneal diseases wherewound healing is impaired such as diabetic keratopathy. In addition,it highlighted the gene gun technique as a valuable tool in exam-ining the role of genes in epithelial abnormalities (Zagon et al.,2006). Another research group targeted plasmids expressing IL10and IL4 to the mouse cornea in order to determine immunemodulation of HSV-1 infection. The expression of delivered geneswas limited to corneal epithelial cells and attenuated the clinicalcourse of infection although small numbers of F4/80þ and CD11bþcells were noted with treatment (Bauer et al., 2006). Thus the genegunmethodmay be of great benefit in the focal treatment of cornealepithelial disorders as minimal transfection occurs in neighboringtissues. However, the technique, in its present form, is in need ofadditional optimization before being adopted for use in the clinic(Bauer et al., 2006). Furthermore, due to high corneal epithelial cellturnover, transgene expression is transient.
2.2.5. Controlled corneal dehydrationRecently we reported that controlled corneal dehydration
increases vector absorption inmouse and rabbit corneas in vivo andhuman cornea ex vivo using a hair dryer following epithelialremoval. Vectors expressing marker gene were used to study theeffect of corneal drying on gene transfer. As evident from the datapresented in Fig. 4, increased corneal drying with a hair dryershowed increased transgene delivery in the mouse cornea in vivo.However, excessive dehydration may have a negative impact oncorneal integrity. Detection of significantly higher transgenedelivery in vivo after 50 s of corneal drying with moderate changesin corneal morphology affirmed the promise of this simple tech-nique for corneal gene therapy. High transgene delivery was alsoobserved after 30 s of drying without jeopardizing cornealmorphology and moderate gene delivery was noted after 10 or 20 sof drying with no altered corneal morphology. This study revealedthat controlled corneal drying modulates gene delivery in thecornea presumably due to a change in corneal hydration. Thus, we

Fig. 4. Representative H&E staining images showing histology of mouse corneas subjected to air-drying and collected 14 days after AAV8 application. A: 0 s air-drying. B: 50 s air-drying. Mouse corneas subjected to 50 s drying showed moderate or higher levels of morphological changes in anterior stroma. C: shows digital quantification of delivered markergene expression detected at day 14 in mouse corneas in vivo. These corneas received 2 ml of AAV8 vector immediately after 0, 20, 30, or 50 s of drying. *p< 0.05 as compared to 20 s;4 p< 0.05 as compared to 30 s.
R.R. Mohan et al. / Progress in Retinal and Eye Research 31 (2012) 43e64 49
postulated that administration of efficient vector employing simpleminimally invasive techniques is a novel approach for deliveringtherapeutic levels of genes in the cornea in vivo (Mohan et al.,2010b).
2.2.6. LaserRecently, a stromal pocket technique involving the use of the
femtosecond laser to introduce genes into the pig cornea ex vivowas reported (Bemelmans et al., 2009). A stromal pocket 110 mm indepth was produced with a femtosecond laser and lentiviral vectorexpressing GFP was injected. Histology of corneal tissue performed5 days after vector application showed wound closure and markergene expression in the cells (most likely keratocytes) around thecorneal pocket. Interestingly, the levels of transgene expressionnoted at day-5 remained up to 3 weeks (Bemelmans et al., 2009).This method facilitates gene delivery into cells in a specific, tar-geted corneal region. However, femtosecond laser is known toinduce intensewound healing and infiltration of inflammatory cellsin the rabbit cornea (Netto et al., 2007). Thus, the next step wouldentail the translation of this procedure from an ex vivo model to anin vivo model and the establishment of a safety profile.
2.2.7. ChemicalsScores of natural and synthetic chemicals have been tested to
introduce genes into corneal cells. In this review we provide a shortdescription of more widely tested lipids and polymers for deliveringgenes in the cornea. Amongmany lipids dioleoyltrimethylammoniumchloride (DOTMA), dioleoylphosphatidyl-ethanolamine (DOPE),1,2-dioleoyl-3-trimethylammonium-propane (DOTAP), dimethyldioctadecyl ammonium bromide (DDAB), 3b-[N-(N0,N0-dimethyla-minoethane) carbamoyl]cholesterol (DC-cholesterol), and N-methyl-4(dioleyl) methylpyridinium chloride (SAINT-2) showed promise forcorneal gene therapy. A transfection solution prepared from fivedifferent lipids showed 7e17% transgene delivery in corneal endo-thelial cells in vitro (Dannowski et al., 2005). Our laboratory formu-lated transfection solutions manipulating ratios of DOPE and DDAB,and observed 12e15% gene delivery in human corneal fibroblastsin vitro, and rodent and rabbit stroma in vivo after corneal
administration via a defined delivery technique (Mohan et al.,unpublished data). Gene delivery in cultured human epithelial cellsand in rabbit corneal epithelium in vivo has also been reported withDOTAP/DOPE transfection mixture (Toropainen et al., 2007). Aformulation of liposomes and transferrin was shown to modulatechemokine expressionwith viralmacrophage inflammatory protein IIin a murine model of corneal allograft rejection (Pillai et al., 2008).Transferrinwas used to enhance transfection efficiency by promotingendocytosis. Among cationic lipids/polymers, polyethyleneimine(PEI), poly-lactide-co-glycolide (PLGA), polylactic acid (PLA), and chi-tosan have been evaluated for corneal gene transfer. PEI showedsignificant transgene delivery in cultured human corneal epithelialcells and rabbit cornea in vivo (Hornof et al., 2008; Yuan et al., 2006).Chitosan, a naturally occurring aminopolysaccharide resulting fromalkaline deacetylation of chitin, has been examined widely for genetransfer because of its nontoxic nature at high concentrations, highavailability, low cost, and biodegradable character. Several studieshave reported successful delivery of genes in ocular tissues, includingthe cornea, using chitosan (de la Fuente et al., 2008a,b). Nonetheless,many challenges such as poor transfection, significant immune reac-tion, cell-targeting, etc. remain to be resolved for its wider applicationas a useful delivery system (Xu et al., 2010).
2.3. Next generation AAV and nanoparticle vectors
To overcome the obstacles presented by conventional viral andnonviral vehicles many new vectors considered next generationwere developed. Discussing all of these is beyond the scope of thisreview. Herein we focus our attention on tyrosine-mutant AAV,nanoparticles, and dendrimers which have shown promise forcorneal gene therapy in preclinical animal studies.
2.3.1. Tyrosine-mutant AAV vectorsThe AAV capsid surface is a fundamental element involved in
host receptor binding, cellular uptake, and intracellular trafficking(Van Vliet et al., 2008) thus affecting transduction efficiency. Traf-ficking within the target cell renders the AAV vector susceptible tonatural cellular degradation mechanisms such as the

R.R. Mohan et al. / Progress in Retinal and Eye Research 31 (2012) 43e6450
ubiquitineproteasome pathway (Douar et al., 2001). Phosphoryla-tion of AAV surface exposed tyrosine residues marks the vector forubiquitination and subsequent proteasome-mediated degradationprior to entering the nucleus (Petrs-Silva et al., 2009). A recentstudy presented evidence that phosphorylation of surface exposedtyrosine residues by epidermal growth factor receptor proteintyrosine kinase (EGFR-PTK) significantly reduces transductionefficiency of both single-stranded and self-complementary AAV2vectors by w68% and w74%, respectively. In addition, intracellulartrafficking is retarded due to AAV ubiquitination followed bydegradation mediated by proteasomes (Zhong et al., 2008). Site-directed mutagenesis of each of the seven AAV2 capsid tyrosineresidues (Y252, Y272, Y444, Y500, Y700, Y704, and Y730) byphenylalanine residue substitution leads to increased vectortransduction and transgene expression by circumventing EGFR-PTKphosphorylation and the ubiquitineproteasome pathway in humancells in vitro and murine hepatocytes in vivo (Zhong et al., 2008).These so-called next generation tyrosine-mutant AAV2, AAV8, andAAV9 vectors have several-fold higher transgene delivery in retinalcells after subretinal or intravitreal injection compared to theirwild-type counterparts (Petrs-Silva et al., 2009). These results arealso clinically applicable in the context of vector dose. AAV2 vectorshave been used in many Phase I/II clinical trials (Zhong et al., 2008).However, relatively large vector doses are required to attain ther-apeutic levels and may trigger an immune reaction. Neutralizingantibody levels are thought to be proportional to virus dose andpreexisting antibodies may reduce transduction efficiency uponvector re-administration (Petry et al., 2008). Tyrosine-mutant AAVvectors were shown to decrease vector dosage needed for trans-duction. A potent tyrosine-mutant AAV2 Y444F vector showedsuperior transduction even at 10,000-fold lower titer compared towild-type AAV2. The lower AAV dose also leads to a decreasedimmune response (Petrs-Silva et al., 2009). Double, triple, andquadruple tyrosine-mutants have been generated to evaluatepotential augmentation of AAV transduction efficiency. In bonemarrow-derived primary murine cells and human mesenchymalstem cells, a triple AAV2 mutant consisting of Y444F, Y500F, andY730F mutations increased transduction efficiency by almost 130-fold and significantly improved viral intracellular trafficking (Liet al., 2010). Our laboratory examined the efficacy of tyrosine-mutant AAV2, AAV8, and AAV9 vectors in delivering genes to thestroma and tyrosine-mutant AAV8 to the endothelium of themousecornea in vivo. In our studies, we utilized hair dryer-based topicalapplication technique for achieving targeted delivery into themouse stroma (Mohan et al., unpublished data) and specialized
Fig. 5. Representative stereomicroscopy (A) and immunocytochemistry in tissue sections (Bof mouse eye in vivo. Dispensing of vector via custom delivery-technique fetched selectivecells expressing GFP gene stained green.
microinjection techniques to introduce the AAV8 vector encodingGFP into the anterior chamber of mice eyes (Sharma A, et al. IOVS2010;51:ARVO E-Abstract 2839). All the three tested tyrosine-mutant AAV serotypes showed significantly higher transgenedelivery in the mouse stroma in vivo compared to their corre-sponding wild type in preliminary studies (Mohan et al., unpub-lished data). The tyrosine-mutant AAV8 vector demonstratedsubstantially greatermarker gene delivery than thewild-type AAV8vector (Fig. 5) into the corneal endothelium of mouse eye in vivowith no apparent adverse effects. Stereomicroscopy, immunocy-tochemistry and slit-lamp biomicroscopy were used to analyze theefficacy and safety of screened vectors. Our recent studies showingsubstantial targeted gene delivery into corneal endothelium areparticularly exciting as it has potential as an effective therapy forendothelial dystrophies in the future.
2.3.2. Nanoparticle vectorsThe National Institutes of Health coined the term nanoparticles
(NPs) for materials 1e100 nm in size used in medicine for diseasediagnosis and therapy. The advances in nanotechnology havesupplied a large reservoir of NPs that may well revolutionize thefuture of medicine. Smaller than 100 nm in size, NPs are minusculeand thus able to access anywhere within the in vivo realm, as livingcells are 10,000e20,000 nm in diameter (Sharma et al., 2010a).Additionally, the ability of NPs to multiplex and incorporatea myriad of ligands such as DNA, antibodies, peptides, and probescan give rise to an array of therapeutic modalities. Internalization ofNPs in mammalian cells is carried out by phagocytosis, macro-pinocytosis, clathrin- or caveolae-mediated endocytosis, and otherclathrin- and caveolae-independent endocytic pathways (Ragusaet al., 2007; Hillaireau and Couvreur, 2009).
Many studies reported gene transfer in non-ocular cells withmetallic (gold), polymeric (PEI, poly-L-lysine, PLGA), or hybrid(metallic-polymeric) NPs (Ragusa et al., 2007; Ghosh et al., 2008).Limited studies investigated the prospective of NPs for deliveringgenes in the eye, including the cornea (Cai et al., 2008; de la Fuenteet al., 2008a,b; Sharma et al., 2011). An in vivo study showedmarkergene delivery in the retina, lens, trabecular meshwork, and corneawith PEG-substituted-30mer lysine NPs (Cai et al., 2008). These NPsprovided short-term transgene expression, and may necessitaterepeated applications for sustained effect. NPs made from PLGAhave been shown to deliver significant transgene in cultured rabbitconjunctival epithelial cells (Qaddoumi et al., 2004). Chitosan NPshave been shown to transfect up to 15% of human corneal epithelialand normal human conjunctival cells in vitro without affecting
) showing tyrosine-mutant AAV8-mediated transgene delivery in corneal endotheliumgene delivery into corneal endothelium. Cell nuclei were stained blue with DAPI while
R.R. Mohan et al. / Progress in Retinal and Eye Research 31 (2012) 43e64 51
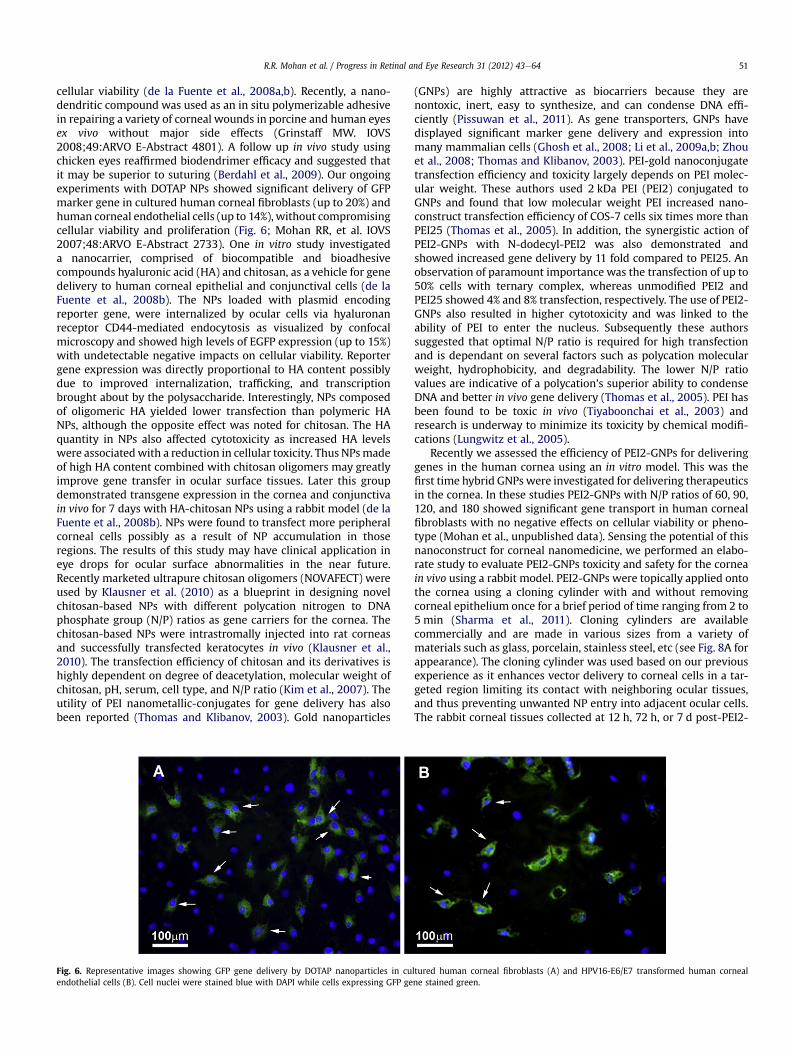
cellular viability (de la Fuente et al., 2008a,b). Recently, a nano-dendritic compound was used as an in situ polymerizable adhesivein repairing a variety of corneal wounds in porcine and human eyesex vivo without major side effects (Grinstaff MW. IOVS2008;49:ARVO E-Abstract 4801). A follow up in vivo study usingchicken eyes reaffirmed biodendrimer efficacy and suggested thatit may be superior to suturing (Berdahl et al., 2009). Our ongoingexperiments with DOTAP NPs showed significant delivery of GFPmarker gene in cultured human corneal fibroblasts (up to 20%) andhuman corneal endothelial cells (up to 14%), without compromisingcellular viability and proliferation (Fig. 6; Mohan RR, et al. IOVS2007;48:ARVO E-Abstract 2733). One in vitro study investigateda nanocarrier, comprised of biocompatible and bioadhesivecompounds hyaluronic acid (HA) and chitosan, as a vehicle for genedelivery to human corneal epithelial and conjunctival cells (de laFuente et al., 2008b). The NPs loaded with plasmid encodingreporter gene, were internalized by ocular cells via hyaluronanreceptor CD44-mediated endocytosis as visualized by confocalmicroscopy and showed high levels of EGFP expression (up to 15%)with undetectable negative impacts on cellular viability. Reportergene expression was directly proportional to HA content possiblydue to improved internalization, trafficking, and transcriptionbrought about by the polysaccharide. Interestingly, NPs composedof oligomeric HA yielded lower transfection than polymeric HANPs, although the opposite effect was noted for chitosan. The HAquantity in NPs also affected cytotoxicity as increased HA levelswere associatedwith a reduction in cellular toxicity. Thus NPsmadeof high HA content combined with chitosan oligomers may greatlyimprove gene transfer in ocular surface tissues. Later this groupdemonstrated transgene expression in the cornea and conjunctivain vivo for 7 days with HA-chitosan NPs using a rabbit model (de laFuente et al., 2008b). NPs were found to transfect more peripheralcorneal cells possibly as a result of NP accumulation in thoseregions. The results of this study may have clinical application ineye drops for ocular surface abnormalities in the near future.Recently marketed ultrapure chitosan oligomers (NOVAFECT) wereused by Klausner et al. (2010) as a blueprint in designing novelchitosan-based NPs with different polycation nitrogen to DNAphosphate group (N/P) ratios as gene carriers for the cornea. Thechitosan-based NPs were intrastromally injected into rat corneasand successfully transfected keratocytes in vivo (Klausner et al.,2010). The transfection efficiency of chitosan and its derivatives ishighly dependent on degree of deacetylation, molecular weight ofchitosan, pH, serum, cell type, and N/P ratio (Kim et al., 2007). Theutility of PEI nanometallic-conjugates for gene delivery has alsobeen reported (Thomas and Klibanov, 2003). Gold nanoparticles
Fig. 6. Representative images showing GFP gene delivery by DOTAP nanoparticles in cuendothelial cells (B). Cell nuclei were stained blue with DAPI while cells expressing GFP ge
(GNPs) are highly attractive as biocarriers because they arenontoxic, inert, easy to synthesize, and can condense DNA effi-ciently (Pissuwan et al., 2011). As gene transporters, GNPs havedisplayed significant marker gene delivery and expression intomany mammalian cells (Ghosh et al., 2008; Li et al., 2009a,b; Zhouet al., 2008; Thomas and Klibanov, 2003). PEI-gold nanoconjugatetransfection efficiency and toxicity largely depends on PEI molec-ular weight. These authors used 2 kDa PEI (PEI2) conjugated toGNPs and found that low molecular weight PEI increased nano-construct transfection efficiency of COS-7 cells six times more thanPEI25 (Thomas et al., 2005). In addition, the synergistic action ofPEI2-GNPs with N-dodecyl-PEI2 was also demonstrated andshowed increased gene delivery by 11 fold compared to PEI25. Anobservation of paramount importance was the transfection of up to50% cells with ternary complex, whereas unmodified PEI2 andPEI25 showed 4% and 8% transfection, respectively. The use of PEI2-GNPs also resulted in higher cytotoxicity and was linked to theability of PEI to enter the nucleus. Subsequently these authorssuggested that optimal N/P ratio is required for high transfectionand is dependant on several factors such as polycation molecularweight, hydrophobicity, and degradability. The lower N/P ratiovalues are indicative of a polycation’s superior ability to condenseDNA and better in vivo gene delivery (Thomas et al., 2005). PEI hasbeen found to be toxic in vivo (Tiyaboonchai et al., 2003) andresearch is underway to minimize its toxicity by chemical modifi-cations (Lungwitz et al., 2005).
Recently we assessed the efficiency of PEI2-GNPs for deliveringgenes in the human cornea using an in vitro model. This was thefirst time hybrid GNPs were investigated for delivering therapeuticsin the cornea. In these studies PEI2-GNPs with N/P ratios of 60, 90,120, and 180 showed significant gene transport in human cornealfibroblasts with no negative effects on cellular viability or pheno-type (Mohan et al., unpublished data). Sensing the potential of thisnanoconstruct for corneal nanomedicine, we performed an elabo-rate study to evaluate PEI2-GNPs toxicity and safety for the corneain vivo using a rabbit model. PEI2-GNPs were topically applied ontothe cornea using a cloning cylinder with and without removingcorneal epithelium once for a brief period of time ranging from 2 to5 min (Sharma et al., 2011). Cloning cylinders are availablecommercially and are made in various sizes from a variety ofmaterials such as glass, porcelain, stainless steel, etc (see Fig. 8A forappearance). The cloning cylinder was used based on our previousexperience as it enhances vector delivery to corneal cells in a tar-geted region limiting its contact with neighboring ocular tissues,and thus preventing unwanted NP entry into adjacent ocular cells.The rabbit corneal tissues collected at 12 h, 72 h, or 7 d post-PEI2-
ltured human corneal fibroblasts (A) and HPV16-E6/E7 transformed human cornealne stained green.
R.R. Mohan et al. / Progress in Retinal and Eye Research 31 (2012) 43e6452
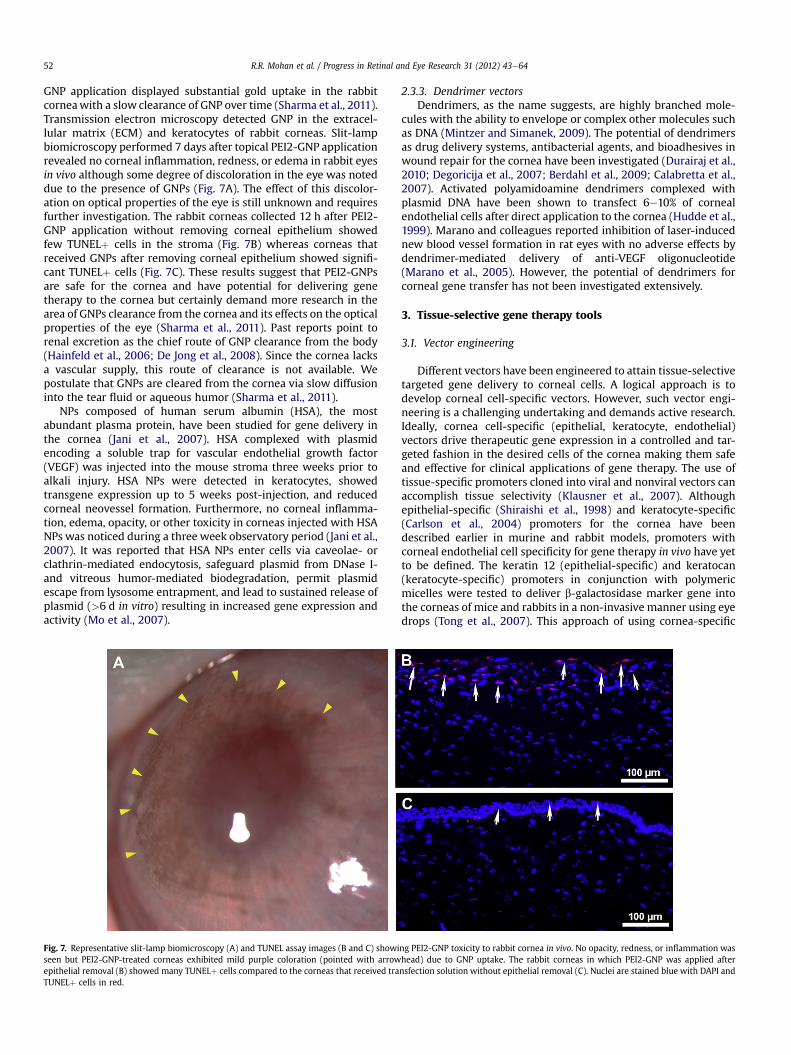
GNP application displayed substantial gold uptake in the rabbitcorneawith a slow clearance of GNP over time (Sharma et al., 2011).Transmission electron microscopy detected GNP in the extracel-lular matrix (ECM) and keratocytes of rabbit corneas. Slit-lampbiomicroscopy performed 7 days after topical PEI2-GNP applicationrevealed no corneal inflammation, redness, or edema in rabbit eyesin vivo although some degree of discoloration in the eye was noteddue to the presence of GNPs (Fig. 7A). The effect of this discolor-ation on optical properties of the eye is still unknown and requiresfurther investigation. The rabbit corneas collected 12 h after PEI2-GNP application without removing corneal epithelium showedfew TUNELþ cells in the stroma (Fig. 7B) whereas corneas thatreceived GNPs after removing corneal epithelium showed signifi-cant TUNELþ cells (Fig. 7C). These results suggest that PEI2-GNPsare safe for the cornea and have potential for delivering genetherapy to the cornea but certainly demand more research in thearea of GNPs clearance from the cornea and its effects on the opticalproperties of the eye (Sharma et al., 2011). Past reports point torenal excretion as the chief route of GNP clearance from the body(Hainfeld et al., 2006; De Jong et al., 2008). Since the cornea lacksa vascular supply, this route of clearance is not available. Wepostulate that GNPs are cleared from the cornea via slow diffusioninto the tear fluid or aqueous humor (Sharma et al., 2011).
NPs composed of human serum albumin (HSA), the mostabundant plasma protein, have been studied for gene delivery inthe cornea (Jani et al., 2007). HSA complexed with plasmidencoding a soluble trap for vascular endothelial growth factor(VEGF) was injected into the mouse stroma three weeks prior toalkali injury. HSA NPs were detected in keratocytes, showedtransgene expression up to 5 weeks post-injection, and reducedcorneal neovessel formation. Furthermore, no corneal inflamma-tion, edema, opacity, or other toxicity in corneas injected with HSANPs was noticed during a three week observatory period (Jani et al.,2007). It was reported that HSA NPs enter cells via caveolae- orclathrin-mediated endocytosis, safeguard plasmid from DNase I-and vitreous humor-mediated biodegradation, permit plasmidescape from lysosome entrapment, and lead to sustained release ofplasmid (>6 d in vitro) resulting in increased gene expression andactivity (Mo et al., 2007).
Fig. 7. Representative slit-lamp biomicroscopy (A) and TUNEL assay images (B and C) showiseen but PEI2-GNP-treated corneas exhibited mild purple coloration (pointed with arrowepithelial removal (B) showed many TUNELþ cells compared to the corneas that received traTUNELþ cells in red.
2.3.3. Dendrimer vectorsDendrimers, as the name suggests, are highly branched mole-
cules with the ability to envelope or complex other molecules suchas DNA (Mintzer and Simanek, 2009). The potential of dendrimersas drug delivery systems, antibacterial agents, and bioadhesives inwound repair for the cornea have been investigated (Durairaj et al.,2010; Degoricija et al., 2007; Berdahl et al., 2009; Calabretta et al.,2007). Activated polyamidoamine dendrimers complexed withplasmid DNA have been shown to transfect 6e10% of cornealendothelial cells after direct application to the cornea (Hudde et al.,1999). Marano and colleagues reported inhibition of laser-inducednew blood vessel formation in rat eyes with no adverse effects bydendrimer-mediated delivery of anti-VEGF oligonucleotide(Marano et al., 2005). However, the potential of dendrimers forcorneal gene transfer has not been investigated extensively.
3. Tissue-selective gene therapy tools
3.1. Vector engineering
Different vectors have been engineered to attain tissue-selectivetargeted gene delivery to corneal cells. A logical approach is todevelop corneal cell-specific vectors. However, such vector engi-neering is a challenging undertaking and demands active research.Ideally, cornea cell-specific (epithelial, keratocyte, endothelial)vectors drive therapeutic gene expression in a controlled and tar-geted fashion in the desired cells of the cornea making them safeand effective for clinical applications of gene therapy. The use oftissue-specific promoters cloned into viral and nonviral vectors canaccomplish tissue selectivity (Klausner et al., 2007). Althoughepithelial-specific (Shiraishi et al., 1998) and keratocyte-specific(Carlson et al., 2004) promoters for the cornea have beendescribed earlier in murine and rabbit models, promoters withcorneal endothelial cell specificity for gene therapy in vivo have yetto be defined. The keratin 12 (epithelial-specific) and keratocan(keratocyte-specific) promoters in conjunction with polymericmicelles were tested to deliver b-galactosidase marker gene intothe corneas of mice and rabbits in a non-invasive manner using eyedrops (Tong et al., 2007). This approach of using cornea-specific
ng PEI2-GNP toxicity to rabbit cornea in vivo. No opacity, redness, or inflammation washead) due to GNP uptake. The rabbit corneas in which PEI2-GNP was applied afternsfection solution without epithelial removal (C). Nuclei are stained blue with DAPI and
R.R. Mohan et al. / Progress in Retinal and Eye Research 31 (2012) 43e64 53
promoters in plasmids has significantly enhanced transgeneexpression in targeted corneal cells (Tong et al., 2007). Anotherapproach for targeted gene therapy in the cornea includes usage ofan inducible strong promoter with a switch-on and -off mechanism(Williams and Coster, 2010). These inducible promoters regulateactivation and duration of gene expression, which is triggered inthe presence of a supplementary factor that is either releasedendogenously under particular physiological conditions (e.g.ischemia) or administered exogenously (Bainbridge et al., 2006).Bitransgenic mouse lines that over-express b-galactosidase in thecorneal epithelium upon induction with doxycycline have beencharacterized in the past (Chikama et al., 2005). A gene-targetingconstruct with an internal ribosomal entry site-reverse tetracy-cline transcription activator cassette was introduced into thekeratin 12 gene (Krt12) to generate a knock-in Krt12rtTA/þ mouseline. These knock-in mice were then bred with tet-O-LacZ reportermice to produce Krt12rtTA/þ/tet-O-LacZ bitransgenic mice. Doxy-cycline ingestion by these bitransgenic mice increased cornealreporter gene expression 15-fold. In addition, b-galactosidaseenzyme activity was detected 24 h post-antibiotic induction, lev-eled out at 2 d, and returned to basal levels 4 wks after doxycyclinewas eliminated from the diet (Chikama et al., 2005). Not only arestudies of this nature extremely useful in expounding signalingmechanisms of different growth factors and cytokines, but also inclarifying the roles of various corneal genes under homeostatic andpathologic conditions (Chikama et al., 2005). More recently, Parkeret al. (2009) studied a glucocorticosteroid-inducible promoter(GRE5) in a lentiviral vector encoding IL10 in A549 cells, and ovineand human corneas in vitro (Parker et al., 2009). A549 cells culturedwith dexamethasone displayed a 30e40-fold increase in IL10compared to controls while ovine and human corneas demon-strated 9e10-fold increase as quantified with enzyme-linkedimmunosorbent assay. This study outlines the efficacy ofa steroid-inducible promoter in corneal gene transfer and paves theway for its application in future studies involving themodulation oftransgene expression in donor corneal allografts (Parker et al.,2009). Even now, no ideal non-leaky cell-specific or induciblevectors are available for corneal gene therapy although consider-able progress has been made in this area.
3.2. Custom vector-delivery techniques
Clinicians routinely perform simple surgical procedures such asepithelial scrape, microinjection, etc in the cornea in the generaleye clinic to treat corneal abnormalities. We postulated a few yearsago that an appropriate combination of vector and vector-deliverytechniques could be used for developing targeted gene therapies.The ideal vector-delivery technique would involve minimal cornealinjury during and/or following vector application and have theability to introduce the vector to a precise location in a controlled
Fig. 8. Representative image showing various minimally invasive defined vector-delivery tecapillary (C) for targeted gene therapy approaches for the cornea.
manner. Methods such as intracameral delivery, intrastromalapplication, topical, and subconjunctival administration have allshown promising outcomes. However, with the exception of topicalapplication and subconjunctival injection, all corneal gene transferapproaches have been deemed invasive with damaging impacts onthe integrity of the ocular surface (Cheng et al., 2007). Even thoughtopical application is the most acceptable approach to delivertherapeutics to the eye due to its non-invasive nature, preservationof corneal integrity, and minimal systemic side effects, the use ofeye drops containing macromolecules including genes for cornealdelivery is ineffective without opening tight junctions to enhanceepithelial layer permeability (Cheng et al., 2007; Hao et al., 2010).Our recent studies demonstrated that vector-delivery techniquesplay a role in gene delivery in the cornea, and could be used as a toolto reduce contact of vector to unwanted tissue and enhance target-specificity (Mohan et al., 2010b). A few strategies we have used forthis purpose are shown in Fig. 8 and include cloning cylinder (A)outlined previously, soaked circular sponge (B), and glass-capillary(C). It is well known that corneal permeability and hydration affectcorneal transparency (Maurice, 1984). The dehydration of thecornea with a hair dryer is a conventional method to treat Fuchs’dystrophy in human patients (Bainbridge et al., 2008). We incor-porated this technique currently in clinical use to dehydrate thecornea in a controlled fashion to increase vector absorption (Mohanet al., 2010b). After removing the epithelium via gentle scrapingwith a beaver blade, a technique commonly used in refractive lasersurgery, a state of corneal dehydration was promoted by applyinga hair dryer once, twice, thrice, or five times to blow warm air onthe ocular surface for 10 s with 5 s intervals. Balanced salt solution(BSS) or AAV8 vector encoding alkaline phosphatase gene was thenimmediately applied topically for 2 min with a custom-designedcloning cylinder to mouse and rabbit corneas in vivo and humancornea ex vivo. We found a statistically significant BSS/vectorabsorption ranging from 14 to 27% in corneas dried for 20, 30, and50 s. AAV8-mediated transgene delivery was also increased withlonger exposure to the hair dryer. However, corneas that under-went prolonged drying for 50 s demonstrated amplified infiltrationof activated granulocytes as detected with CD11b immunostainingwhereas 30 s of hair dryer-supplied warm air did not trigger severeimmune reaction or jeopardize corneal morphology (Mohan et al.,2010b).
It has been long known that corneal epithelial injury inducesapoptosis of keratocytes, inflammation, and wound healing in thecornea (Mohan et al., 2003; Wilson et al., 2002; Jester et al., 1999).Several researchers have demonstrated a cascade of cellular eventsincluding cytokine and growth factor release following epithelialinjury leading to varied levels of corneal wound healing response(Mohan et al., 2003;Wilson et al., 2002; Jester et al., 1999). Scores ofclinical and animal studies have found distinctly different woundhealing responses in the cornea after surgical, mechanical, or
chniques developed using cloning cylinder (A), soaked circular sponge (B), and glass-

R.R. Mohan et al. / Progress in Retinal and Eye Research 31 (2012) 43e6454
chemical injury. Many investigators including us have reported thatinjury to the corneal epithelium causes keratocyte apoptosis in theanterior stroma. However, no studies were performed to examinewhether the intensity of mechanical injury dictates the amount ofkeratocyte death in the anterior stroma. Recently, we hypothesizedthat careful, gentle removal of the corneal epithelium with a #64surgical blade under controlled conditions induces clinicallyinsignificant keratocyte apoptosis and wound healing in the corneain vivo contrary to rough corneal epithelium removal with a #64blade under undefined conditions. To test the hypothesis we per-formed a very simple experiment using a rabbit model. In the samerabbit eye, 5 mm central corneal epthelium was gently removedfollowing defined condition and 4 mm peripheral corneal epithe-lium was removed in a rough manner with no defined conditionswith a #64 surgical blade. Fig. 9 shows the TUNEL assay of therabbit cornea collected 24 h after euthanasia. The detection of a fewTUNELþ cells in the central cornea where the epithelium wasremoved by defined gentle technique and several TUNELþ cells inthe peripheral cornea where the epithelium was removed byundefined rough technique strongly support our hypothesis.Additional support to our hypothesis comes from our recent hairdryer study in which we observed almost no inflammatoryresponse in corneas subjected to up to 3 rounds of 10 s dehydrationas a comparable quantity of inflammatory cells were seen inmurinecorneas that did not experience hair dryer-assisted dehydration(Fig. 4, Mohan et al., 2010b). Thus we hold that the administrationof an efficient vector via defined vector-delivery technique cansuccessfully deliver therapeutic genes in the corneal stroma in vivowithout compromising corneal integrity or function. More researchis required to fully define parameters and techniques that minimizeside effects, improve safety, and augment targeted delivery oftherapeutic genes into keratocytes or the stroma in vivo with AAVvectors.
Delivering genes precisely into the corneal endothelium isa huge challenge and a major limitation for developing genetherapy for corneal dystrophies such as Fuchs’ dystrophy, a leadingcause of corneal blindness with no effective treatments available.To address this challenge we developed minimally invasivemicrosurgical techniques based on the principle of air pressureforce like a water gun as shown in Fig. 10. In our studies we foundthese techniques improved target-specificity significantly potenti-ating transgene delivery into mouse corneal endothelium in vivo.The newly optimized microsurgical procedures derived frommicroinjection technique are simple and may be used routinely inophthalmology but may require practice to attain proficiency likeall other surgical techniques.
Fig. 9. Representative TUNEL assay images showing effects of epithelial removal on keratoccorneas. Few TUNELþ cells were noted in customized-gentle scrape technique (A) compareand TUNELþ cells in red.
4. Gene therapy updates in various corneal disorders
Trauma, injury and/or infections to the eye lead to cornealdysfunction and vision impairment. Corneal disorders are the 3rdleading cause of blindness in the world according to the WorldHealth Organization (World Health Organization, 2011; Whitcheret al., 2001). Eight million people in the world, including 1.5million children, are blind due to corneal abnormalities (Whitcheret al., 2001). Corneal disease and disorders have a broad range ofpathology leading to diverse outcomes. Haze, scarring, abnormalwound healing, new blood vessel ingrowth, conjunctivitis, dry eye,graft failure, endothelial defects, and Fuchs’ dystrophy are preva-lent corneal problems among many acquired and genetic cornealdiseases. Gene therapy holds great promise for treating as well aspreventing corneal diseases and disorders. In the field of cornealgene therapy emphasis is more on treating common cornealdisorders instead of curing genetic defects. Here we provide anoverview of the progress made in the field of corneal gene therapywith a brief description about the disease condition/cause.
4.1. Corneal graft rejection
Keratoplasty is currently used for treating many cornealdiseases. According to the Eye Bank Association of America morethan 42,600 corneal transplants were performed in the UnitedStates in 2010 alone (Eye Bank Association of America, 2010).Although perhaps the most successful transplant procedure, it isnot without blemish. Corneal graft rejection due to immunologicalreaction is the chief cause of failure despite the cornea enjoying animmune-privilege status (Williams and Coster, 2010). Post-operative complications and lack of good quality donor corneasare the other concerns. Gene therapy approaches have been testedto improve allograft survival by delivering various therapeuticgenes to modulate cellular transport, apoptosis, angiogenesis, andwound healing in vivo (Williams and Coster, 2010). In 2006, theLarkin group reported the delivery of indoleamine 2,3-dioxygenase (IDO) in murine corneal endothelium with lenti-virus vector (Beutelspacher et al., 2006). An increase in IDO mRNAand protein levels was detected and IDO expression was found tobe intracellular and restricted to the endothelial layer of thecornea. Ironically corneal endothelial pathology accounts foralmost 48% of penetrating keratoplasty. Once activated, IDO, anintracellular enzyme, induced by pro-inflammatory cytokines likeinterferon (IFN) and TNF, leads to the immunoregulatory catabo-lism of tryptophan via opening of the essential amino acid’s indolering forming N-formylkynurenine and subsequently L-kynurenine
yte death by custom (A) and non-custom (B) epithelial scrape techniques in the rabbitd to non-optimized epithelial removal technique (B). Nuclei are stained blue with DAPI

Fig. 10. Representative images showing technique utilized for achieving preferential gene delivery into mouse corneal endothelium in vivo.
R.R. Mohan et al. / Progress in Retinal and Eye Research 31 (2012) 43e64 55
(Higuchi and Hayaishi, 1967). The IDO-mediated diminution oftryptophan, in addition to the extracellular release of tryptophancatabolites, is believed to halt activated T cells in the cell cycle’s G1phase encouraging immune tolerance (Munn et al., 1999) andapoptosis essential to the survival of transplanted corneas. Full-thickness corneal grafts were also used by the Larkin group tostudy vector-mediated IDO over-expression (Beutelspacher et al.,2006). The study demonstrated significant prolongation ofcorneal allograft survival after IDO transduction of excised donorcornea ex vivo prior to transplantation compared to GFP and non-IDO-transduced control corneas (Beutelspacher et al., 2006). Thisdemonstrates the potential of IDO gene therapy to thwart cornealallograft rejection. A year later in 2007, the Dana group reportedthe use of a lentivirus vector to over-express four anti-apoptoticgenes (bcl-xL, bcl-2, survivin and p35) in a rodent corneal trans-plantation model. They found that lentivirus-mediated transfer ofbcl-xL gene was not only effective at inhibiting apoptosis in thecorneal endothelium in vitro, but also significantly enhancedcorneal graft survival in vivo as a 90% success rate was observed at8 weeks post-transplantation compared to 40% in uninfectedcontrols and 30% in reporter gene controls (Barcia et al., 2007).More recently, the same group studied lentivirus-driven bcl-xLand p35 gene delivery in both primary human endothelial cellsand human corneas, and found both genes displayed anti-apoptotic properties in the corneal endothelium although p35had greater anti-apoptotic efficiency than bcl-xL (Fuchsluger et al.,2011). It is also believed that gene therapy at earlier steps like thesensitization of host to foreign antigen may be more efficacious(Williams and Coster, 2010). Other potential gene therapyapproaches include utilizing IL4, IL10, CTLA4-Ig, p40-IL12, viralMIPII, and nerve growth factor (Williams and Coster 2010).
Another avenue for the utilization of gene therapy with regardto corneal transplantation involves corneal endothelial cell densitymaintenance in corneal tissue procured and stored at eye banks.Recombinant adenovirus-driven over-expression of transcriptionfactor E2F2 ex vivo demonstrated increased endothelial cell countin rabbit and human corneas (Joyce et al., 2004; McAlister et al.,2005). Endothelial cells, normally amitotic, progressed throughthe cell cycle fromG1 to S phase in significant quantities accountingfor the boost in the monolayer cell density. E2F2 expression wasshort lived with expression dropping off 7 d following a 2 h vectorexposure (McAlister et al., 2005). This has great potential in notonly the long-term preservation of corneas but also the donation ofcorneas with low endothelial density previously consideredunsuitable for keratoplasty involving endothelial replacement. Inaddition, the benefits of the vector-mediated transfer of E2F2 mayprove beneficial in the treatment of corneal endotheliopathies.
4.2. Corneal scarring and wound healing
Wound healing plays a central part in the maintenance ofcorneal transparency and thus normal vision. Corneal injury,regardless of cause, may ignite a dysregulated wound healingresponse that commonly leads to corneal fibrosis and loss of visualfunction. Corneal wound healing is an extremely intricate processunder the regulation of several cytokines and growth factors.Among several cytokines transforming growth factor beta (TGFb)has been identified as a chief player in the formation of a myofi-broblasts and opacity in the cornea. Thus, it has become a primetarget for gene therapy aimed at preventing corneal scarring andother corneal disorders caused by TGFb. Many investigators sharethe thought that impeding TGFb or its signal transduction repre-sents a powerful strategy to modulate uncontrolled corneal healingand prevent or even cure corneal scarring. Recently, using anin vitro model our laboratory demonstrated that decorin over-expression in human corneal fibroblasts significantly inhibitsTGFb-driven keratocyte differentiation to myofibroblasts withoutcompromising cellular viability (Mohan et al., 2010a). Decorin,a member of the small leucine-rich proteoglycan family, isexpressed in the corneal stroma and plays an important role inwound healing and structural support in the cornea (Mohan et al.,2011a). Using laser capture technique, we found that photo-refractive keratectomy-induced laser injury to the cornea does notalter decorin significantly but dramatically increases TGFb levels inthe rabbit cornea, in vivo (Tandon et al., 2010). This observationconcurs with our hypothesis that AAV-mediated decorin genetherapy can effectively reduce corneal scarring in vivo. Indeed, ina very recent study we detected significant inhibition of cornealscarring in the rabbit eye (59e73%; p< 0.001 or <0.01) with noadverse effects at one-month time point with AAV5-mediateddecorin gene therapy (Mohan et al., 2011c). This study was thefirst to demonstrate the therapeutic potential of decorin for treat-ing corneal diseases via targeted gene therapy (Fig. 11). Sensing thetranslational potential of AAV-mediated decorin gene therapy forcorneal scarring we are studying whether long-term targeted over-expression of decorin causes any complication to the eye or changesin corneal function or transparency. The clinical and slit-lampexamination in the eyes of live rabbit at up to 4 months showed nosign of adverse effects indicating that decorin gene therapy is safefor patients (Mohan et al., unpublished data). The pending histo-logical, immunochemical, molecular and transmission electronmicroscopy studies will validate this notion.
Venturing to block specific targets in TGFb signaling is anotherapproach used by scientists to develop corneal fibrosis treatment.Numerous studies have shown that TGFb uses the Smad pathway

R.R. Mohan et al. / Progress in Retinal and Eye Research 31 (2012) 43e6456
to relay its signal to the nucleus leading to fibrosis (Tandon et al.,2010). The translocation of Smad2 and Smad4 proteins duringcorneal wound healing in vivo has been demonstrated by per-forming keratectomy and epithelial debridement wounds inSpragueeDawley rats (Hutcheon et al., 2005). This study moti-vated us to perform in vitro studies with Smad2, Smad3, Smad4,and Smad7 siRNA to examine whether TGFb in human cornealfibroblast cells relays its signal via the Smad pathway. The resultsof our study showed that TGFb-mediated keratocyte trans-differentiation to myofibroblasts occurs via Smad signaling andthat Smad7 is an attractive target to prevent myofibroblastformation. Our findings are in accord with many literature reportsas Smad7 gene therapy has been reported to inhibit fibrosis inmany non-ocular tissues such as kidneys, peritoneum, liver, etc.(Chen et al., 2011; Dooley et al., 2003). Recently, Galiacy et al.(2011) used gene therapy strategies to hinder excessive collagendeposition in corneal scar development. Using a recombinantAAV-based vector (AAV2/8) the researchers over-expresseda specific fibril collagenase, matrix metalloproteinase (MMP) 14,to prevent collagen deposition and subsequent scarring followingincisional injury to the cornea in mice. After microdissectinga 0.75 mm full-thickness button from the central corneal region,a single injection of 10 ml PBS into the stroma in control mice wasperformed (vector was injected 10 h prior to incisional injury inthe case of AAV2/8-GFP and AAV2/8-MMP14). This previouslyoutlined model encourages the fibroproliferative response duringrepair including the development of alpha smooth muscle actin(aSMA)-expressing myofibroblasts from stromal keratocytes(Stramer et al., 2003). The investigators demonstrated that a singleintrastromal injection with recombinant AAVeMMP14 mediatedmurine stromal keratocyte gene expression. This was evident withdecreased mRNA expression of aSMA and type III collagen, twokey gene markers of corneal fibrosis (Galiacy et al., 2011). MMP,like TGFb, also has important implications in other processes suchas neovessel formation.
Fig. 11. Representative stereomicroscopy (A, B) and a-smooth muscle actin immunostainingin rabbits in vivo. AAV5 viral titer (6.5�1012 mg/ml) was topically applied onto the cornea
4.3. Corneal neovascularization
Neovascularization may occur in any of the layers of the corneafollowing ocular trauma, infection, injury, etc. and leads to cornealopacity. The sprouting of new blood vessels in the cornea from thelimbus is closely linked to the inflammatory response and posesa major risk for corneal allograft rejection (Klausner et al., 2007).The mechanism of neovascular formation and regression istremendously complex involving a slew of cytokines, growthfactors, and cell types. However, it is widely accepted that vascularendothelial growth factor (VEGF) plays a pivotal role in the devel-opment of new blood vessels. A well-designed study performed bythe Ambati group reported that the avascular phenotype of thecornea is due, at least in part, to soluble VEGF receptor-1(sVEGFR-1)or sflt-1, an endogenous trap for VEGF (Ambati et al., 2006). Thuspatients with aniridia and mutations in Paired box gene 6 (PAX6)lack sflt-1 and display spontaneous corneal neovascularization(CNV). Gene therapy with sflt-1 may greatly benefit patients withthese types of corneal disturbances. Five years earlier in 2001, Laiand cohorts conducted a study in a rat model showing the inhibi-tion of CNV with recombinant adenovirus-mediated delivery ofsflt-1 (Lai et al., 2001). Gene therapy utilizing sflt-1 may certainlyprove beneficial in averting VEGF-induced neovascularization inthe setting of corneal graft rejection and other corneal disorders. Avariety of gene therapy methodologies have been probed to stemcorneal angiogenesis using experimental models and differenttransgenes that target VEGF such as decorin, angiostatin, PEDF,vasohibin-1 and VEGF receptors Flt-1 and Flk-1 (Cheng et al., 2007;Jani et al., 2007; Lai et al., 2007; Oh et al., 2010; Kuo et al., 2009; Yuet al., 2007; Zhou et al., 2010; Mohan et al., 2011b). Adenovirus-mediated gene transfer of soluble VEGF receptor-2, sFlk-1 wasshown to inhibit VEGF-stimulated proliferation of murine andhuman umbilical vein endothelial cells (Yu et al., 2007). The samestudy demonstrated that CNV development in rats in vivo followingcauterization is suppressed by anterior chamber injection of
(C, D) images showing corneal haze inhibition by targeted AAV5-decorin gene therapyonce for 2 min after PRK.

R.R. Mohan et al. / Progress in Retinal and Eye Research 31 (2012) 43e64 57
adenovirus delivery of sFlk-1 (Yu et al., 2007). Immunohisto-chemistry studies of corneas from these animals showed localiza-tion of protein production in the stroma and endothelium 14 daysfollowing cauterization compared to absent sFlk-1 expression incontrols (adenovirus-null and adenovirus-GFP). This stronglysuggests that adenovirus gene therapy has the ability to transducecorneal cells of the endothelial and stromal layers with sFlk-1,a vital element in the development of angiogenesis (Yu et al., 2007).
Gene therapy strategies using endostatin have also shownpromise in treating CNV. Endostatin, a naturally occurring fragmentof collagen type XVIII, blocks endothelial cell adhesion, migrationand proliferation, and inhibits apoptosis (O’Reilly et al., 1997). Miceeyes treated via single subconjunctival injection with recombinantAAV encoding endostatin following silver nitrate cauterization toinduce CNV showed significant inhibition of corneal angiogenesis.Endostatin released from the conjunctiva suppressed the migratinglimbal vasculature, was detected at 4 days post-injection, andretained stable levels for more than 8 months. Interestingly nodifference in immune response (measured by CD4 and CD8lymphocyte staining)was noticed inmice treatedwith recombinantAAV-GFP injection, PBS injection, and no injection (Lai et al., 2007).
Another gene therapy approach to obstruct VEGF-induced CNVdevelopment utilized intrastromal injection of intraceptors linkedto the endoplasmic reticulum retaining peptide KDEL. Plasmidsencoding for specific VEGF binding domains of sFlt-1, Flt23K andFlt24K, coupled to KDEL were delivered via pCMV vector and wereshown to interrupt VEGF signaling in vitro and in vivo. Thesepeptides offset intracellular VEGF as they are not liberated from thecell into the extracellular milieu. Interestingly a variation in VEGFdownregulation was seen between the two intraceptors withFlt23K suppressing injury induced CNV by 66.8% and Flt24K byalmost 50%, possibly due to differences in protein configuration(Singh et al., 2005). The same group then tested the ability of Flt23Kto impede corneal neovessel growth using albumin nanoparticlesand showed CNV regression up to 40% at 5 weeks post intrastromalinjection in a murine model (Jani et al., 2007).
Recently Chen and cohorts evaluated gene therapy withmultiple genes known to have antivasculogenic activity in a murinemodel of CNV (Chen et al., 2010). Retrovirus encoding murine sFlk-1, sTie2, and endostatin genes was used in cell cultures of humanumbilical vein endothelial cells (HUVEC) as well as in vivo studiesinvolving subconjunctival injection of vector into corneas of C57B1/6 mice with chemically induced CNV. Studies in vitro showeddecreased HUVEC proliferation with obvious additive effects whentwo or three genes were combined (i.e. sFlk-1/sTie2, msFlk-1/endostatin, sTie2/endostatin, sFlk-1/sTie2/endostatin). However,although HUVEC migration was also decreased by these genesindividually, the additive effect seen in proliferation was absenthighlighting the various pathways involved in these intricateprocesses of CNV. For the in vivo segment of the study, genecombination showed efficient CNV inhibition specifically down-regulating levels of IL-1b, MMP-9, and VEGF as measured bywestern blotting and RT-PCR. In addition, the combinations of sFlk-1/endostatin and sFlk-1/sTie2/endostatin were found to signifi-cantly decrease nuclear factor (NF)-kB phosphorylation and superrepressor inhibitor kappa Ba (IkBa) expression without regard tototal NF-kB quantity (Chen et al., 2010). This has direct implicationsto CNV as NF-kB leads to VEGF expressionwhile IkBa inhibits NF-kBby maintaining it in an inactive complex in the cytoplasm (Jo�skoand Mazurek, 2004; Karin, 1999).
Combining genes to thwart CNV is not a new concept. Forexample, an almost decade old report from Casey Eye Instituteevaluated lentivirus transfer of a fusion protein in an allogenictransplantationmodel of CNV. Lentivirus was employed to transfectrabbit corneal buttons ex vivo with a fusion gene made up of
endostatin and the kringle 5 domain of plasminogen. This smallspecific plasminogen fragment is known to display robust inhibi-tion of angiogenesis. Corneas transduced with the endostatin andkringle 5 fusion protein showed decreased proliferation andmigration of endothelial cells and subsequently less post-operativeneovascularization and inflammation in comparison to controls(Murthy et al., 2003). Other kringle elements have also beenassessed for their antiangiogenic properties. Angiostatin isa product of proteolytic cleavage that comprises the first fourkringle domains of plasminogen. One group examined the role ofsubconjunctival injection of AAV carrying angiostatin in a rat modelof alkali-induced CNV. The researchers illustrated corneal genetransduction of angiostatin and a post-operative day 7 meanCNV area of 15.83� 0.55 mm2 in untreated controls and6.87� 2.23 mm2 in angiostatin treated animals reflecting 56.6%abatement of CNV due to angiostatin (Cheng et al., 2007).
In another study, mouse corneas were transfected with DNAplasmid encoding IL18 using intrastromal injection. The angiostaticproperty of IL18 is well known making it potentially useful in thetreatment of ocular disorders associated with CNV such as herpeskeratitis. The investigators showed a decrease in VEGF productionin IL18 treated eyes and subsequently a weakened immunoin-flammatory response compared to controls. Furthermore, bFGFinduced MBE cell lines treated with IL18 demonstrated diminishedVEGF production and gene expression in contrast to cells incubatedwith OVA (Kim et al., 2005).
Vasohibin-1, an endothelium-derived negative feedbackmodulator of neovessel formation characterized by Watanabe, hasalso been studied in gene therapy studies as a means to inhibit CNV(Watanabe et al., 2004; Zhou et al., 2010). A 2010 report by Zhouand colleagues described the subconjunctival injection of adeno-virus encoding vasohibin-1 that resulted in significant CNVreduction. It was speculated that the antiangiogenic nature ofvasohibin-1 may be related to the downregulation of VEGF type 2receptor (Zhou et al., 2010).
Other gene transfer studies have assessed the utility of CD36(Mwaikambo et al., 2006), pigment epithelium derived factor (Kuoet al., 2009), and GA binding protein (Yoon et al., 2009) amongother approaches to treating CNV. Future experiments may includevector-mediated delivery of inhibitors of angiogenesis derived frombasement membrane such as neostatin, restin, arresten, canstatin,and tumstatin (Ellenberg et al., 2010). Recently, our laboratorydetected significant CNV reduction in rabbit eyes after targeteddecorin gene therapy using AAV5 vector (Mohan et al., 2011b). CNVin rabbit eyes was induced by VEGF implantation and AAVexpressing decorin or GFP gene was topically applied for twominutes on the rabbit cornea one day after VEGF implantation. Asshown in Fig. 12, targeted decorin delivery in the stroma showed(52e66% p< 0.05, p< 0.001 or p< 0.01) reduction at various stagesof CNV in rabbit model (Mohan et al., 2011b).
4.4. Corneal alkali burn
Alkali burn injury to the cornea is amajor clinical entity that leadsto sight-threatening sequelae such as corneal scarring, neovesselformation, and ulceration. Scores of preclinical studies have beendedicated to finding ways to halt the progression of these negativeaftereffects of alkali burn. In2007 theSaikagroupoutlined theuseofadenovirus to introduceperoxisomeproliferator-activated receptor-g (PPARg) to keratocytes in a mouse alkali burn model (Saika et al.,2007). Gene therapy employing PPARg, a nuclear hormonereceptor with immunomodulatory functions, has been evaluatedextensively for non-ocular tissues (Lewis et al., 2008; Klotz et al.,2009; Kulkarni et al., 2011). In the cornea, a faint expression ofPPARg was noted via immunohistochemistry in the undamaged
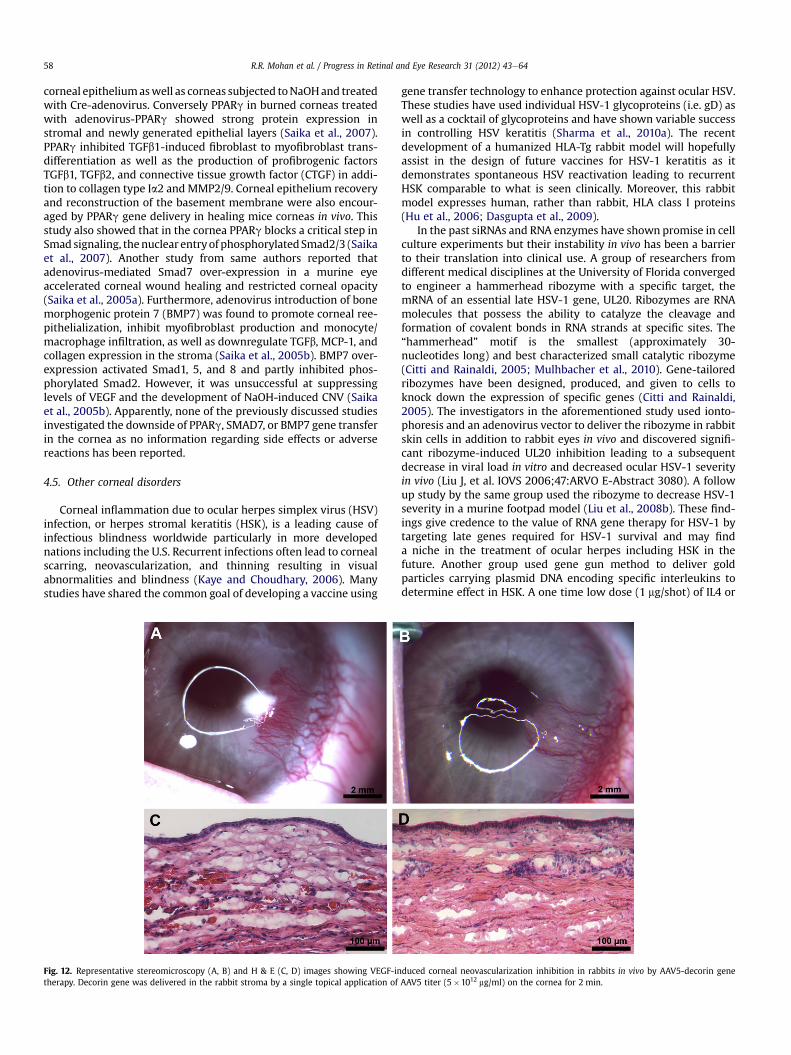
R.R. Mohan et al. / Progress in Retinal and Eye Research 31 (2012) 43e6458
corneal epitheliumaswell as corneas subjected toNaOHand treatedwith Cre-adenovirus. Conversely PPARg in burned corneas treatedwith adenovirus-PPARg showed strong protein expression instromal and newly generated epithelial layers (Saika et al., 2007).PPARg inhibited TGFb1-induced fibroblast to myofibroblast trans-differentiation as well as the production of profibrogenic factorsTGFb1, TGFb2, and connective tissue growth factor (CTGF) in addi-tion to collagen type Ia2 and MMP2/9. Corneal epithelium recoveryand reconstruction of the basement membrane were also encour-aged by PPARg gene delivery in healing mice corneas in vivo. Thisstudy also showed that in the cornea PPARg blocks a critical step inSmad signaling, thenuclear entryof phosphorylatedSmad2/3 (Saikaet al., 2007). Another study from same authors reported thatadenovirus-mediated Smad7 over-expression in a murine eyeaccelerated corneal wound healing and restricted corneal opacity(Saika et al., 2005a). Furthermore, adenovirus introduction of bonemorphogenic protein 7 (BMP7) was found to promote corneal ree-pithelialization, inhibit myofibroblast production and monocyte/macrophage infiltration, as well as downregulate TGFb, MCP-1, andcollagen expression in the stroma (Saika et al., 2005b). BMP7 over-expression activated Smad1, 5, and 8 and partly inhibited phos-phorylated Smad2. However, it was unsuccessful at suppressinglevels of VEGF and the development of NaOH-induced CNV (Saikaet al., 2005b). Apparently, none of the previously discussed studiesinvestigated the downside of PPARg, SMAD7, or BMP7 gene transferin the cornea as no information regarding side effects or adversereactions has been reported.
4.5. Other corneal disorders
Corneal inflammation due to ocular herpes simplex virus (HSV)infection, or herpes stromal keratitis (HSK), is a leading cause ofinfectious blindness worldwide particularly in more developednations including the U.S. Recurrent infections often lead to cornealscarring, neovascularization, and thinning resulting in visualabnormalities and blindness (Kaye and Choudhary, 2006). Manystudies have shared the common goal of developing a vaccine using
Fig. 12. Representative stereomicroscopy (A, B) and H & E (C, D) images showing VEGF-intherapy. Decorin gene was delivered in the rabbit stroma by a single topical application of
gene transfer technology to enhance protection against ocular HSV.These studies have used individual HSV-1 glycoproteins (i.e. gD) aswell as a cocktail of glycoproteins and have shown variable successin controlling HSV keratitis (Sharma et al., 2010a). The recentdevelopment of a humanized HLA-Tg rabbit model will hopefullyassist in the design of future vaccines for HSV-1 keratitis as itdemonstrates spontaneous HSV reactivation leading to recurrentHSK comparable to what is seen clinically. Moreover, this rabbitmodel expresses human, rather than rabbit, HLA class I proteins(Hu et al., 2006; Dasgupta et al., 2009).
In the past siRNAs and RNA enzymes have shown promise in cellculture experiments but their instability in vivo has been a barrierto their translation into clinical use. A group of researchers fromdifferent medical disciplines at the University of Florida convergedto engineer a hammerhead ribozyme with a specific target, themRNA of an essential late HSV-1 gene, UL20. Ribozymes are RNAmolecules that possess the ability to catalyze the cleavage andformation of covalent bonds in RNA strands at specific sites. The“hammerhead” motif is the smallest (approximately 30-nucleotides long) and best characterized small catalytic ribozyme(Citti and Rainaldi, 2005; Mulhbacher et al., 2010). Gene-tailoredribozymes have been designed, produced, and given to cells toknock down the expression of specific genes (Citti and Rainaldi,2005). The investigators in the aforementioned study used ionto-phoresis and an adenovirus vector to deliver the ribozyme in rabbitskin cells in addition to rabbit eyes in vivo and discovered signifi-cant ribozyme-induced UL20 inhibition leading to a subsequentdecrease in viral load in vitro and decreased ocular HSV-1 severityin vivo (Liu J, et al. IOVS 2006;47:ARVO E-Abstract 3080). A followup study by the same group used the ribozyme to decrease HSV-1severity in a murine footpad model (Liu et al., 2008b). These find-ings give credence to the value of RNA gene therapy for HSV-1 bytargeting late genes required for HSV-1 survival and may finda niche in the treatment of ocular herpes including HSK in thefuture. Another group used gene gun method to deliver goldparticles carrying plasmid DNA encoding specific interleukins todetermine effect in HSK. A one time low dose (1 mg/shot) of IL4 or
duced corneal neovascularization inhibition in rabbits in vivo by AAV5-decorin geneAAV5 titer (5�1012 mg/ml) on the cornea for 2 min.

R.R. Mohan et al. / Progress in Retinal and Eye Research 31 (2012) 43e64 59
IL10 plasmid DNA was found to significantly abrogate the clinicalcourse of HSK when administered at 300 psi helium pressure withminimal alterations in immune response systemically. IL6, shownto play a role in HSK development (Banerjee et al., 2004), was alsodownregulated. In addition, gene gun utilization did not lead tosignificant corneal damage as ascertained by histology and clinicalexamination (Bauer et al., 2006).
Diabetic disease of the cornea is a common occurrence inpatients with type I and II diabetes mellitus and can lead to sight-threatening corneal neuropathy and epitheliopathy. Currently, notreatment exists to thwart the progression or cure the disease andtreatment regimens are largely symptomatic. Saghizadeh et al.(2010) recently used recombinant AV to over-express the hepato-cyte growth factor (HGF) receptor c-met proto-oncogene in cornealepithelium of diabetic human corneas in vitro. Their past studiesoutlined disturbances in HGF/c-met signaling in diabetic corneas asc-met expression was reduced in the face of upregulated HGFexpression (Saghizadeh et al., 2005). With the introduction ofexogenous c-met via recombinant AV vector they reportedimproved reestablishment of HGF signaling and subsequentrestoration of marker protein patterns and enhanced woundhealing (Saghizadeh et al., 2010).
Mucopolysaccharidosis (MPS) is a set of metabolic disorderscharacterized by the inability to breakdown glycosaminoglycans(GAG) due to lysosomal enzyme deficiency. GAGs thus accumulatein lysosomes systemically as well as in the eyes (Ashworth et al.,2006). MPS VII, or Sly disease, is due to b-glucuronidase (GUSB)insufficiency leading to the stockpiling of GAGs (dermatan, hep-aran, and chondroitin sulfate) while MPS VI, or MaroteauxeLamysyndrome, results from aryl N-acetylgalactosamine-4-sulfatasedeficiency leading to accumulation of dermatan sulfate(Ashworth et al., 2006). Pathology is common in the ocular systemand results in visual impairment due to opacification of the corneaand disorders of other ocular tissues (Ashworth et al., 2006). Whilelysosomal accumulation occurs in the corneal endothelium, it istheir storage in keratocytes that causes corneal opacity (Williamsand Coster, 2010). Systemic gene therapy using a retroviral vectorhas been shown to avert GAG accrual and ensuing corneal cloudingdue to MPS using neonatal canines (Wang et al., 2006; Traas et al.,2007; Ponder and Auricchio, 2010). Even 6.5e8 years followingtherapy, little or no corneal opacity was detected in dogs with MPSVII (Ponder and Auricchio, 2010). Gene therapy may have a role asan alternative therapeutic strategy for MPS disorders by intro-ducing genes that encode for specific enzymes, such as GUSB,clearing lysosome build up in the eye and other tissues.
5. Lessons learned from corneal genetic studies: humans andanimals
Advances in molecular biology have led to the identification ofnumerous genes and gene mutations associated with dystrophicdiseases of the cornea. Differentmodes ofMendelian inheritance areevident in corneal dystrophies likely due to alterations in severalgenes includingCHST6,KRT3, TGFBI, andUBIAD1 (Klintworth, 2009).TheTGFb inducedgene (TGFBI), commonlyknownasbig-h3, encodesthe TGFBI protein (TGFBIp or keratoepithelin). Changes in thegenomic sequence of TGFBI have been linked to several dystrophiesof the corneal stroma such as Lattice dystrophy type I, Reis-Bucklerdystrophy, Thiel-Behnke corneal dystrophy, granular cornealdystrophy type I, and granular corneal dystrophy II (Avellino cornealdystrophy) (Runager et al., 2008; Klintworth, 2009). Indeed 38different mutations in the TGFBI gene are responsible for variouscorneal dystrophies, the majority being missense mutations(Runager et al., 2008). TGFBIp functions as a cell adhesion moleculeacting byway of integrins (a1b1, a3b1, avb3, etc) and elements of the
ECMincludingproteoglycans (i.e. decorin),fibronectin, andcollagens(Runager et al., 2008). The precise function of these interactionsremains to be identified although TGFBIp may play a role as a regu-latorymodulator between the ECM and cells found in its supportingmeshwork (Runager et al., 2008). Dystrophic corneas containaggregates of unknownmakeup although increasedTGFBIphas beendetected in addition to clusterin, a proaggregation protein, in Fuchsendothelial corneal dystrophy (Runager et al., 2008; Jurkunas et al.,2009). A recent report identified the two corneal dystrophies,gelatinous drop-like corneal dystrophyand spheroidal degeneration,in the same patient (Zhang and Yao, 2010). Upon molecular geneticexamination a new missense mutation in the tumor associatedcalcium signal transducer 2 (TACSTD2) gene was unearthed (Zhangand Yao, 2010). Earlier it was found that alterations in the UbiA pre-nyltransferase domain containing 1 gene are responsible forSchnyder corneal dystrophy (Weiss et al., 2007). The corneal endo-thelial defect was found to be caused, at least in part, by variations inthe transcription factor 4 (TCF4) gene (Baratz et al., 2010). Theknowledge of gene localization is paramount in creating an animalmodel of disease and treatment approaches via gene therapy.Previously itwas thought that type I and IImacular corneal dystrophywere due to genetic aberrations of carbohydrate sulfotransferase 6(CHST6) in the epithelial layer (Abbruzzese et al., 2004) and thatlimbal stem-cell therapy might possibly offer benefits to patients interms of therapy. It was recently shown however, that CHST6 alongwith keratan sulfate are chiefly the stromal products of keratocytesunderscoring the need for therapeutic modalities, including genetherapy, to target stromal keratocytes (Di Iorio et al., 2010).
Nonetheless, the lack of in vivo animal models has sharplyhindered progress in the development of gene therapy options forpatients suffering from corneal dystrophies (Weiss, 2010).Furthermore, the pathogenesis and localization of genes respon-sible for many of these disorders has yet to be elucidated. Recently,an animal model mimicking human corneal dystrophy was re-ported by Chen et al. (2010). A mouse model of congenital stromalcorneal dystrophy, caused by a faulty decorin gene leading tocorneal clouding, was created utilizing a Cre-on approach. It wasalso surmised that mutations in decorin accounting for the disor-ganized stromal architecture may be influenced by class II SLRPslumican and keratocan (Chen S, et al. IOVS 2010;51:ARVO E-Abstract 3832). It is conceivable that keratocytes may induce anincrease in GAG chain and SLRP production to offset deficiencies instromal ECM GAGs or SLRPs brought about by genetic causes. Thetissue-selective targeted gene-based therapeutic approaches weare attempting to define for the cornea have great potential forcorneal genetic dystrophies.
6. Ocular gene therapy clinical trials: current status
Currently 20 gene therapy trials for eye disease are listed on TheJournal of Gene Medicine Clinical Trial Website. Out of 20 genetherapy clinical trials, 18 are directed toward retinal disorders, 1 forglaucoma, and 1 for the cornea. The lone corneal gene therapyPhase I/II clinical trial is focused on evaluating the safety and effi-cacy of a matrix-targeted retroviral vector bearing a dominantnegative cyclin G1 construct (Mx-dnG1) as adjunctive interventionfor superficial corneal opacity/corneal scarring. The findings of theclinical trial assessing gene therapy for diseases of the cornea havenot yet been published.
7. Future directions: regenerative medicine, nanomedicineand other approaches
The future of gene therapy for the cornea is very promising andinvolves different branches of medicine including regenerative

R.R. Mohan et al. / Progress in Retinal and Eye Research 31 (2012) 43e6460
medicine and nanotechnology. Indeed, nanotechnology will lead tothe development of regenerative medicine, microsensors andfeedback apparatus, improved imaging in vivo, and artificial vision(Zarbin et al., 2010a). Regenerative nanomedicine is a new field inmedicine gaining widespread attention. It entails the use of nano-particles containing regulatory molecules such as gene transcrip-tion factors that allow for cellular reprogramming in vivo (Zarbinet al., 2010a). Nanoparticles may not only serve as vectors fortherapeutic gene delivery but may also act as biocarriers of geneswhich may assist in the diagnosis and monitoring of cornealdisease. This process of coupling disease diagnosis with targetedtreatment is termed theragnostics and has enormous potential inthe ocular system including the cornea. For example, oxidativestress has been implicated in numerous inflammatory, metabolic,traumatic, and iatrogenic corneal diseases (Shoham et al., 2008). Bycoupling a therapeutic gene such as peroxidase, superoxide dis-mutase, or catalase (as well as a marker gene like GFP) to nano-particles capable of activating antioxidant response element, onemay produce a nano-device with dual diagnosticetherapeuticeffects enabling corneal cells to treat themselves following oxida-tive damage (Zarbin et al., 2010b).
Fagerholm and colleagues recently reported the results ofa clinical study in Sweden involving corneal regeneration withrecombinant human collagen. Recombinantly produced humancollagen type III was cross-linked with 1-ethyl-3-(3-dimethylaminopropyl) carbodiimide (EDC) and N-hydrox-ysuccinimide (NHS) and fashioned into substitutes with humancorneal dimensions. This biomimetic substitute was then trans-planted into patients following anterior lamellar keratoplasty. Theinvestigators found that a cell-free corneal substitute imitating thecornea’s unique ECM can successfully provide support for endog-enous restoration of corneal epithelium, stroma, and nerves. Ofnote was the absence of inflammation, neovascularization, orrejection that plagues recipients of human donor corneas(Fagerholm et al., 2009). Although the study had a small patientpopulation (n¼ 10), it clearly highlights the safety profile ofrecombinant human collagen and cross-linking methods(Fagerholm et al., 2009). Preliminary results from the studydemonstrate improved visual acuity, ocular surface quality, andcorneal sensitivity in patients with a biomimetic corneal substitute.These corneal substitutes may prove beneficial in circumstancesnecessitating short-term or emergency corneal replacements,where availability of human donor corneas is poor or nonexistentand where prostheses are unfavorable (Fagerholm et al., 2009). Inthe future these biomimetic corneas may serve as efficientpermanent corneal replacements and a viable option to humancorneal tissue in transplantation. These substitutes in combinationwith single or multiple gene therapy may find a niche in thetreatment of recurrent CNV and other disorders of the ocularsurface.
Cell-based therapy for corneal diseases has also been examined.A study from the Funderburgh group (Du et al., 2009) used stemcells isolated from adult human corneal stroma to rectify stromalopacity due to SLRP lumican deficiency in mice via intrastromalinjection of cells. Stem cells subsisted inwild-type mice for monthswithout host-cell fusion or provoking an immune T-cell response.Human stromal stem cells introduced into the stroma of lumican-null mice with corneal opacity (similar in nature to that of scartissue due to disorganized stromal collagen) led to rectification offibrillar collagen defects and stromal thickness and restoration ofcorneal transparency (Du et al., 2009). Thus stem-cell therapyshows proof of principle in treating corneal disease through theregeneration of corneal tissue in a safe and efficacious mannersimilar to organogenesis. The results of a remarkable clinical studyin Italy involving autologous limbal cell treatment were recently
reported (Rama et al., 2010). Currently limbal stem cells, a group ofcells responsible for corneal epithelial renewal and regeneration,are transplanted as a treatment for limbal stem-cell deficiency(Pellegrini et al., 2009). 112 patients with burn-related cornealdestruction associated with limbal stem-cell deficiency weretreated with autologous limbal stem cells cultivated on fibrin.Corneal transparency was restored in 76.6% of eyes up to 10 yearsafter corneal transplantation with limbal stem-cell grafts identi-fying limbal stem-cell cultures as a source of transplantable cells fortreating corneal burns (Rama et al., 2010).
Future studies involving gene therapy for corneal diseases mayinclude non-coding RNA (ncRNA). ncRNA includes several types offunctional RNAmolecules not translated into proteins that play keyroles in several cellular processes such as transfer RNA, ribosomalRNA, small interfering RNAs, microRNAs, PIWI-associated RNAs,small nucleolar RNAs, promoter-associated RNAs, and telomerespecific small RNAs. It has been shown that ncRNAs play a part inoncogenesis and may be a factor in molecular aberrations of otherdiseases (Galasso et al., 2010).
In the recent past, toll like receptors (TLRs) have gained atten-tion in the cornea as their activation sets off a multifacetedsignaling pathway instigating innate and adaptive immunitygenerating inflammatory cytokines and co-stimulatory molecules(Redfern and McDermott, 2010). The production of chemokinesencourages corneal infiltration of macrophages and neutrophilsresulting in the functional loss of corneal epithelium, stroma, andendothelium leading to the loss of corneal transparency and visualactivity (Kanwar et al., 2010). The modulation of TLR signaling viagene therapy methods may play a vital role in blocking immune-mediated corneal graft rejection as well as aberrant wound heal-ing following corneal infection.
The future will reveal animal models of human corneal diseasein which to test highly efficient viral or nonviral vectors with low-immunogenicity and sustained transgene expression in combina-tion with cell-based therapy and regenerative approaches. Pres-ently the cornea lags behind the retina in gene therapy for variousreasons including lack of animal models of inherited disease forexperimentation. Currently available technologies are likelynontoxic and effective, at least in the short term, with little furtherdevelopment (Weiss, 2010).
8. Conclusions
The last five to six years have brought about great progress andadvancement in the field of corneal gene therapy. The increasedunderstanding of the molecular mechanisms and course ofinherited and acquired corneal diseases have allowed for theemergence of targets for gene-based therapy. The testing of variousviral and nonviral vectors and multiple delivery systems has beenthe key to the success observed in various models of cornealdisease. We along with other investigators have examined theefficacy of gene therapy using in vitro, ex vivo, and in vivo animal(equine, murine, canine, etc) and human corneal tissues, identifiedseveral efficient AAV serotypes and nanoparticles, and definedsimpleminimally invasive vector-delivery techniques for deliveringgenes precisely into targeted cells (keratocytes and endothelium) ofthe cornea in vivo. Furthermore, using a combination of vector anddelivery techniques, we have defined tissue-selected targeted genetherapy approaches for the cornea. Although much of the cornealgene therapy investigations have been accomplished in animals,there is a dearth of research that has been translated fully frombench to bedside including the clinic. The progress in establishingcorneal gene therapy clinical trials has been meager compared togene therapy trials conducted in other ocular tissues. Even thoughthe cornea is easily accessible for gene transfer, it is lagging far

R.R. Mohan et al. / Progress in Retinal and Eye Research 31 (2012) 43e64 61
behind the retina in terms of clinical application of gene therapy forhuman patients. The future is bright however, as novel innovationsand treatment approaches have proved effective in animal models(i.e. rabbits) that closely resemble the human cornea in terms ofanatomy, wound healing response, and function.
Over the past few years, much effort has been made to developstrategies for effective delivery of DNA to the nucleus of target cells.The most pertinent question concerning the success of genetherapy is “how to introduce therapeutic genes into target cells/tissues?” Vectors are important tools for introducing foreign genesinto cells and are one of the twomajor determinants for the successof gene therapy, the other being gene delivery technique. Severalviral and nonviral vectors have been developed and each has itsown advantages as well as limitations. In addition, many vector-delivery techniques have been developed to enhance vector effi-ciency and achieve target-specificity. With the discovery of animalmodels of human corneal disease and the combination of genetherapy approaches with stem-cell therapy and regenerativemedicine, the cure for corneal blindness may well become a realitywithin the next decade.
Acknowledgments
The work was supported from the RO1EY17294 (R.R.M.)National Eye Institute, NIH, Bethesda, MD, NEI/NIH RO1EY17294S2Diversity Supplement (J.C.K.T.), 1I01BX00035701 (R.R.M.) VeteranHealth Affairs, Washington, DC, and an Unrestricted Research toPrevent Blindness, New York, NY grants.
References
Abbruzzese, C., Kuhn, U., Molina, F., Rama, P., De Luca, M., 2004. Novel mutations inthe CHST6 gene causing macular corneal dystrophy. Clin. Genet. 65, 120e125.
Adriaansen, J., Vervoordeldonk, M.J., Tak, P.P., 2006. Gene therapy as a therapeuticapproach for the treatment of rheumatoid arthritis: innovative vectors andtherapeutic genes. Rheumatology (Oxford) 45, 656e668.
Akache, B., Grimm, D., Pandey, K., Yant, S.R., Xu, H., Kay, M.A., 2006. The 37/67-kilodalton laminin receptor is a receptor for adeno-associated virus serotypes8, 2, 3, and 9. J. Virol. 80, 9831e9836.
Alexander, J.J., Hauswirth, W.W., 2008. Adeno-associated viral vectors and theretina. Adv. Exp. Med. Biol. 613, 121e128.
Ambati, B.K., Nozaki, M., Singh, N., Takeda, A., Jani, P.D., Suthar, T., Albuquerque, R.J.,Richter, E., Sakurai, E., Newcomb, M.T., Kleinman, M.E., Caldwell, R.B., Lin, Q.,Ogura, Y., Orecchia, A., Samuelson, D.A., Agnew, D.W., St Leger, J., Green, W.R.,Mahasreshti, P.J., Curiel, D.T., Kwan, D., Marsh, H., Ikeda, S., Leiper, L.J.,Collinson, J.M., Bogdanovich, S., Khurana, T.S., Shibuya, M., Baldwin, M.E.,Ferrara, N., Gerber, H.P., De Falco, S., Witta, J., Baffi, J.Z., Raisler, B.J., Ambati, J.,2006. Corneal avascularity is due to soluble VEGF receptor-1. Nature 443,993e997.
Ashworth, J.L., Biswas, S., Wraith, E., Lloyd, I.C., 2006. Mucopolysaccharidoses andthe eye. Surv. Ophthalmol. 51, 1e17.
Bainbridge, J.W., Tan, M.H., Ali, R.R., 2006. Gene therapy progress and prospects: theeye. Gene Ther. 13, 1191e1197.
Bainbridge, J.W., Smith, A.J., Barker, S.S., Robbie, S., Henderson, R., Balaggan, K.,Viswanathan, A., Holder, G.E., Stockman, A., Tyler, N., Petersen-Jones, S.,Bhattacharya, S.S., Thrasher, A.J., Fitzke, F.W., Carter, B.J., Rubin, G.S., Moore, A.T.,Ali, R.R., 2008. Effect of gene therapy on visual function in Leber’s congenitalamaurosis. N. Engl. J. Med. 358, 2231e2239.
Banerjee, K., Biswas, P.S., Kim, B., Lee, S., Rouse, B.T., 2004. CXCR2�/� mice showenhanced susceptibility to herpetic stromal keratitis: a role for IL-6-inducedneovascularization. J. Immunol. 172, 1237e1245.
Baratz, K.H., Tosakulwong, N., Ryu, E., Brown, W.L., Branham, K., Chen, W., Tran, K.D.,Schmid-Kubista, K.E., Heckenlively, J.R., Swaroop, A., Abecasis, G., Bailey, K.R.,Edwards, A.O., 2010. E2-2 protein and Fuchs’s corneal dystrophy. N. Engl. J. Med.363, 1016e1024.
Barcia, R.N., Dana, M.R., Kazlauskas, A., 2007. Corneal graft rejection is accompaniedby apoptosis of the endothelium and is prevented by gene therapy with bcl-xL.Am. J. Transplant. 7, 2082e2089.
Bauer, D., Lu, M., Wasmuth, S., Li, H., Yang, Y., Roggendorf, M., Steuhl, K.P.,Heiligenhaus, A., 2006. Immunomodulation by topical particle-mediatedadministration of cytokine plasmid DNA suppresses herpetic stromal keratitiswithout impairment of antiviral defense. Graefes Arch. Clin. Exp. Ophthalmol.244, 216e225.
Bemelmans, A.P., Arsenijevic, Y., Majo, F., 2009. Efficient lentiviral gene transfer intocorneal stroma cells using a femtosecond laser. Gene Ther. 16, 933e938.
Berdahl, J.P., Johnson, C.S., Proia, A.D., Grinstaff, M.W., Kim, T., 2009. Comparison ofsutures and dendritic polymer adhesives for corneal laceration repair in anin vivo chicken model. Arch. Ophthalmol. 127, 442e447.
Beutelspacher, S.C., Ardjomand, N., Tan, P.H., Patton, G.S., Larkin, D.F., George, A.J.,McClure, M.O., 2005. Comparison of HIV-1 and EIAV-based lentiviral vectors incorneal transduction. Exp. Eye Res. 80, 787e794.
Beutelspacher, S.C., Pillai, R., Watson, M.P., Tan, P.H., Tsang, J., McClure, M.O.,George, A.J., Larkin, D.F., 2006. Function of indoleamine 2,3-dioxygenase incorneal allograft rejection and prolongation of allograft survival by over-expression. Eur. J. Immunol. 36, 690e700.
Broekman, M.L., Comer, L.A., Hyman, B.T., Sena-Esteves, M., 2006. Adeno-associatedvirus vectors serotyped with AAV8 capsid are more efficient than AAV-1 or -2serotypes for widespread gene delivery to the neonatal mouse brain. Neuro-science 138, 501e510.
Cai, X., Conley, S., Naash, M., 2008. Nanoparticle applications in ocular gene therapy.Vision Res. 48, 319e324.
Calabretta, M.K., Kumar, A., McDermott, A.M., Cai, C., 2007. Antibacterial activities ofpoly(amidoamine) dendrimers terminated with amino and poly(ethyleneglycol) groups. Biomacromolecules 8, 1807e1811.
Carlson, E.C., Liu, C.Y., Yang, X., Gregory, M., Ksander, B., Drazba, J., Perez, V.L., 2004.In vivo gene delivery and visualization of corneal stromal cells using anadenoviral vector and keratocyte-specific promoter. Invest. Ophthalmol. Vis. Sci.45, 2194e2200.
Challa, P., Luna, C., Liton, P.B., Chamblin, B., Wakefield, J., Ramabhadran, R.,Epstein, D.L., Gonzalez, P., 2005. Lentiviral mediated gene delivery to theanterior chamber of rodent eyes. Mol. Vis. 11, 425e430.
Chen, H.Y., Huang, X.R., Wang, W., Li, J.H., Heuchel, R.L., Chung, A.C., Lan, H.Y., 2011.The protective role of Smad7 in diabetic kidney disease: mechanism andtherapeutic potential. Diabetes 60, 590e601.
Chen, P., Yin, H., Wang, Y., Mi, J., He, W., Xie, L., Wang, Y., 2010. Multi-gene targetedantiangiogenic therapies for experimental corneal neovascularization. Mol. Vis.16, 310e319.
Cheng, H.C., Yeh, S.I., Tsao, Y.P., Kuo, P.C., 2007. Subconjunctival injection ofrecombinant AAV-angiostatin ameliorates alkali burn induced corneal angio-genesis. Mol. Vis. 13, 2344e2352.
Chikama, T., Hayashi, Y., Liu, C.Y., Terai, N., Terai, K., Kao, C.W., Wang, L., Hayashi, M.,Nishida, T., Sanford, P., Doestchman, T., Kao, W.W., 2005. Characterization oftetracycline-inducible bitransgenic Krt12rtTA/þ/tet-O-LacZ mice. Invest. Oph-thalmol. Vis. Sci. 46, 1966e1972.
Citti, L., Rainaldi, G., 2005. Synthetic hammerhead ribozymes as therapeutic tools tocontrol disease genes. Curr. Gene Ther. 5, 11e24.
Dannowski, H., Bednarz, J., Reszka, R., Engelmann, K., Pleyer, U., 2005. Lipid-medi-ated gene transfer of acidic fibroblast growth factor into human cornealendothelial cells. Exp. Eye Res. 80, 93e101.
Dasgupta, G., Chentoufi, A.A., Nesburn, A.B., Wechsler, S.L., BenMohamed, L., 2009.New concepts in herpes simplex virus vaccine development: notes from thebattlefield. Expert Rev. Vaccines 8, 1023e1035.
De Jong, W.H., Hagens, W.I., Krystek, P., Burger, M.C., Sips, A.J., Geertsma, R.E., 2008.Particle size-dependent organ distribution of gold nanoparticles after intrave-nous administration. Biomaterials 29, 1912e1919.
de la Fuente, M., Seijo, B., Alonso, M.J., 2008a. Bioadhesive hyaluronanechitosannanoparticles can transport genes across the ocular mucosa and transfectocular tissue. Gene Ther. 15, 668e676.
de la Fuente, M., Seijo, B., Alonso, M.J., 2008b. Novel hyaluronic acid-chitosan nano-particles for ocular gene therapy. Invest. Ophthalmol. Vis. Sci. 49, 2016e2024.
Degoricija, L., Johnson, C.S., Wathier, M., Kim, T., Grinstaff, M.W., 2007. Photo cross-linkable biodendrimers as ophthalmic adhesives for central lacerations andpenetrating keratoplasties. Invest. Ophthalmol. Vis. Sci. 48, 2037e2042.
Di Iorio, E., Barbaro, V., Volpi, N., Bertolin, M., Ferrari, B., Fasolo, A., Arnaldi, R.,Brusini, P., Prosdocimo, G., Ponzin, D., Ferrari, S., 2010. Localization andexpression of CHST6 and keratan sulfate proteoglycans in the human cornea.Exp. Eye Res. 91, 293e299.
Dooley, S., Hamzavi, J., Breitkopf, K., Wiercinska, E., Said, H.M., Lorenzen, J., TenDijke, P., Gressner, A.M., 2003. Smad7 prevents activation of hepatic stellatecells and liver fibrosis in rats. Gastroenterology 125, 178e191.
Douar, A.M., Poulard, K., Stockholm, D., Danos, O., 2001. Intracellular trafficking ofadenoassociated virus vectors: routing to the late endosomal compartment andproteasome degradation. J. Virol. 75, 1824e1833.
Dratviman-Storobinsky, O., Lubin, B.C., Hasanreisoglu, M., Goldenberg-Cohen, N.,2009. Effect of subconjuctival and intraocular bevacizumab injection onangiogenic gene expression levels in a mouse model of corneal neo-vascularization. Mol. Vis. 15, 2326e2338.
Du, Y., Carlson, E.C., Funderburgh, M.L., Birk, D.E., Pearlman, E., Guo, N., Kao, W.W.,Funderburgh, J.L., 2009. Stem cell therapy restores transparency to defectivemurine corneas. Stem Cells 27, 1635e1642.
Durairaj, C., Kadam, R.S., Chandler, J.W., Hutcherson, S.L., Kompella, U.B., 2010.Nanosized dendritic polyguanidilyated translocators for enhanced solubility,permeability, and delivery of gatifloxacin. Invest. Ophthalmol. Vis. Sci. 51,5804e5816.
Ellenberg, D., Azar, D.T., Hallak, J.A., Tobaigy, F., Han, K.Y., Jain, S., Zhou, Z.,Chang, J.H., 2010. Novel aspects of corneal angiogenic and lymphangiogenicprivilege. Prog. Retin. Eye Res. 29, 208e248.
Eye Bank Association of America 2010 Statistical Report. URL: http://www.restoresight.org/about-us/understanding-donation/cornea-donation-transplantation-statistics/(accessed on 23.09.11).

R.R. Mohan et al. / Progress in Retinal and Eye Research 31 (2012) 43e6462
Fagerholm, P., Lagali, N.S., Carlsson, D.J., Merrett, K., Griffith, M., 2009. Cornealregeneration following implantation of a biomimetic tissue-engineeredsubstitute. Clin. Transl. Sci. 2, 162e164.
Fuchsluger, T.A., Jurkunas, U., Kazlauskas, A., Dana, R., 2011. Corneal endothelial cellsare protected from apoptosis by gene therapy. Hum. Gene Ther. 22, 549e558.
Galasso, M., Elena Sana, M., Volinia, S., 2010. Non-coding RNAs: a key to futurepersonalized molecular therapy? Genome Med. 2, 12.
Galiacy, S.D., Fournié, P., Massoudi, D., Ancèle, E., Quintyn, J.C., Erraud, A., Raymond-Letron, I., Rolling, F., Malecaze, F., 2011. Matrix metalloproteinase 14 over-expression reduces corneal scarring. Gene Ther. 18, 462e468.
Gao, G., Vandenberghe, L.H., Wilson, J.M., 2005. New recombinant serotypes of AAVvectors. Curr. Gene Ther. 5, 285e297.
Gao, G., Lu, Y., Calcedo, R., Grant, R.L., Bell, P., Wang, L., Figueredo, J., Lock, M.,Wilson, J.M., 2006. Biology of AAV serotype vectors in liver-directed genetransfer to nonhuman primates. Mol. Ther. 13, 77e87.
Gao, X., Kim, K.S., Liu, D., 2007. Nonviral gene delivery: what we know and what isnext. AAPS J. 9, E92e104.
Ghosh, A., Yue, Y., Long, C., Bostick, B., Duan, D., 2007. Efficient whole-body trans-duction with trans-splicing adeno-associated viral vectors. Mol. Ther. 15,750e755.
Ghosh, P.S., Kim, C.K., Han, G., Forbes, N.S., Rotello, V.M., 2008. Efficient genedelivery vectors by tuning the surface charge density of amino acid-functionalized gold nanoparticles. ACS Nano. 2, 2213e2218.
Gong, N., Pleyer, U., Vogt, K., Anegon, I., Flügel, A., Volk, H.D., Ritter, T., 2007a. Localoverexpression of nerve growth factor in rat corneal transplants improvesallograft survival. Invest. Ophthalmol. Vis. Sci. 48, 1043e1052.
Gong, N., Pleyer, U., Volk, H.D., Ritter, T., 2007b. Effects of local and systemic viralinterleukin-10 gene transfer on corneal allograft survival. Gene Ther. 14,484e490.
Hacein-Bey-Abina, S., Garrigue, A., Wang, G.P., Soulier, J., Lim, A., Morillon, E.,Clappier, E., Caccavelli, L., Delabesse, E., Beldjord, K., Asnafi, V., MacIntyre, E., DalCortivo, L., Radford, I., Brousse, N., Sigaux, F., Moshous, D., Hauer, J.,Borkhardt, A., Belohradsky, B.H., Wintergerst, U., Velez, M.C., Leiva, L.,Sorensen, R., Wulffraat, N., Blanche, S., Bushman, F.D., Fischer, A., Cavazzana-Calvo, M., 2008. Insertional oncogenesis in 4 patients after retrovirus-mediatedgene therapy of SCID-X1. J. Clin. Invest. 118, 3132e3142.
Hainfeld, J.F., Slatkin, D.N., Focella, T.M., Smilowitz, H.M., 2006. Gold nanoparticles:a new X-ray contrast agent. Br. J. Radiol. 79, 248e253.
Hao, J., Li, S.K., Liu, C.Y., Kao, W.W., 2009. Electrically assisted delivery of macro-molecules into the corneal epithelium. Exp. Eye Res. 89, 934e941.
Hao, J., Li, S.K., Kao, W.W., Liu, C.Y., 2010. Gene delivery to cornea. Brain Res. Bull. 81,256e261.
Hauswirth, W.W., Aleman, T.S., Kaushal, S., Cideciyan, A.V., Schwartz, S.B., Wang, L.,Conlon, T.J., Boye, S.L., Flotte, T.R., Byrne, B.J., Jacobson, S.G., 2008. Treatment ofleber congenital amaurosis due to RPE65 mutations by ocular subretinalinjection of adeno-associated virus gene vector: short-term results of a phase Itrial. Hum. Gene Ther. 19, 979e990.
He, Z., Pipparelli, A., Manissolle, C., Acquart, S., Garraud, O., Gain, P., Thuret, G., 2010.Ex vivo gene electrotransfer to the endothelium of organ cultured humancorneas. Ophthalmic Res. 43, 43e55.
Herzog, R.W., 2010. Gene therapy for SCID-X1: round 2. Mol. Ther. 18, 1891.Higuchi, K., Hayaishi, O., 1967. Enzymic formation of d-kynurenine from dtrypto-
phan. Arch. Biochem. Biophys. 120, 397e403.Hillaireau, H., Couvreur, P., 2009. Nanocarriers’ entry into the cell: relevance to drug
delivery. Cell. Mol. Life Sci. 66, 2873e2896.Hornof, M., de la Fuente, M., Hallikainen, M., Tammi, R.H., Urtti, A., 2008. Low
molecular weight hyaluronan shielding of DNA/PEI polyplexes facilitates CD44receptor mediated uptake in human corneal epithelial cells. J. Gene Med. 10,70e80.
Hu, J., Peng, X., Schell, T.D., Budgeon, L.R., Cladel, N.M., Christensen, N.D., 2006. AnHLA-A2.1-transgenic rabbit model to study immunity to papillomavirus infec-tion. J. Immunol. 177, 8037e8045.
Hudde, T., Rayner, S.A., Comer, R.M., Weber, M., Isaacs, J.D., Waldmann, H.,Larkin, D.F., George, A.J., 1999. Activated polyamidoamine dendrimers, a non-viral vector for gene transfer to the corneal endothelium. Gene Ther. 6,939e943.
Hutcheon, A.E., Guo, X.Q., Stepp, M.A., Simon, K.J., Weinreb, P.H., Violette, S.M.,Zieske, J.D., 2005. Effect of wound type on Smad 2 and 4 translocation. Invest.Ophthalmol. Vis. Sci. 46, 2362e2368.
Jani, P.D., Singh, N., Jenkins, C., Raghava, S., Mo, Y., Amin, S., Kompella, U.B.,Ambati, B.K., 2007. Nanoparticles sustain expression of Flt intraceptors in thecornea and inhibit injury-induced corneal angiogenesis. Invest. Ophthalmol.Vis. Sci. 48, 2030e2036.
Jester, J.V., Petroll, W.M., Cavanagh, H.D., 1999. Corneal stromal wound healing inrefractive surgery: the role of myofibroblasts. Prog. Retin. Eye Res. 18,311e356.
Jo�sko, J., Mazurek, M., 2004. Transcription factors having impact on vascularendothelial growth factor (VEGF) gene expression in angiogenesis. Med. Sci.Monit. 10, RA89eRA98.
Joyce, N.C., Harris, D.L., McAlister, J.C., Ali, R.R., Larkin, D.F.P., 2004. Overexpressionof the transcription factor E2F2 induces cell cycle progression in rabbit cornealendothelial cells. Invest. Ophthalmol. Vis. Sci. 45, 1340e1348.
Jurkunas, U.V., Bitar, M., Rawe, I., 2009. Colocalization of increased transforminggrowth factor-beta-induced protein (TGFBIp) and Clusterin in Fuchs endothelialcorneal dystrophy. Invest. Ophthalmol. Vis. Sci. 50, 1129e1136.
Kanwar, J.R., Mohan, R.R., Kanwar, R.K., Roy, K., Bawa, R., 2010. Applications ofaptamers in nanodelivery systems in cancer, eye and inflammatory diseases.Nanomedicine (Lond.) 5, 1435e1445.
Karin, M., 1999. How NF-kappaB is activated: the role of the IkappaB kinase (IKK)complex. Oncogene 18, 6867e6874.
Kaye, S., Choudhary, A., 2006. Herpes simplex keratitis. Prog. Retin. Eye Res. 25,355e380.
Kim, B., Lee, S., Suvas, S., Rouse, B.T., 2005. Application of plasmid DNA encoding IL-18 diminishes development of herpetic stromal keratitis by antiangiogeniceffects. J. Immunol. 175, 509e516.
Kim, T.H., Jiang, H.L., Jere, D., Park, I.K., Cho, M.H., Nah, J.W., Choi, U.J., Akaike, T.,Cho, C.S., 2007. Chemical modification of chitosan as a gene carrier in vitro andin vivo. Prog. Polym. Sci. 32, 726e753.
Klausner, E.A., Peer, D., Chapman, R.L., Multack, R.F., Andurkar, S.V., 2007. Cornealgene therapy. J. Control Release 124, 107e133.
Klausner, E.A., Zhang, Z., Chapman, R.L., Multack, R.F., Volin, M.V., 2010. Ultrapure chi-tosan oligomers as carriers for corneal gene transfer. Biomaterials 31, 1814e1820.
Klibanov, A.L., 2006. Microbubble contrast agents: targeted ultrasound imaging andultrasound-assisted drug-delivery applications. Invest. Radiol. 41, 354e362.
Klintworth, G.K., 2009. Corneal dystrophies. Orphanet. J. Rare Dis. 4, 7.Klotz, L., Hucke, S., Thimm, D., Classen, S., Gaarz, A., Schultze, J., Edenhofer, F.,
Kurts, C., Klockgether, T., Limmer, A., Knolle, P., Burgdorf, S., 2009. Increasedantigen cross-presentation but impaired cross-priming after activation ofperoxisome proliferator-activated receptor gamma is mediated by up-regulation of B7H1. J. Immunol. 183, 129e136.
Kulkarni, A.A., Thatcher, T.H., Olsen, K.C., Maggirwar, S.B., Phipps, R.P., Sime, P.J.,2011. PPAR-g ligands repress TGFb-induced myofibroblast differentiation bytargeting the PI3K/Akt pathway: implications for therapy of fibrosis. PLoS One6, e15909.
Kuo, C.N., Yang, L.C., Yang, C.T., Chen, M.F., Lai, C.H., Chen, Y.H., Chen, C.H., Chen, C.H.,Wu, P.C., Kou, H.K., Tsai, J.C., Hung, C.H., 2008. A novel vector system for genetransfer into the cornea using a partially dried plasmid expressing 18 basicfibroblast growth factor-synthetic amphiphile INTeraction-18 (SAINT-18)complex. Curr. Eye Res. 33, 839e848.
Kuo, C.N., Yang, L.C., Yang, C.T., Lai, C.H., Chen, M.F., Chen, C.Y., Chen, C.H., Wu, P.C.,Kou, H.K., Chen, Y.J., Hung, C.H., Tsai, C.B., 2009. Inhibition of corneal neo-vascularization with plasmid pigment epithelium-derived factor (p-PEDF)delivered by synthetic amphiphile Interaction-18 (SAINT-18) vector in anexperimental model of rat corneal angiogenesis. Exp. Eye Res. 89, 678e685.
Lai, C.M., Brankov, M., Zaknich, T., Lai, Y.K., Shen, W.Y., Constable, I.J., Kovesdi, I.,Rakoczy, P.E., 2001. Inhibition of angiogenesis by adenovirus-mediated sFlt-1expression in a rat model of corneal neovascularization. Hum. Gene Ther. 12,1299e1310.
Lai, L.J., Xiao, X., Wu, J.H., 2007. Inhibition of corneal neovascularization withendostatin delivered by adeno-associated viral (AAV) vector in a mouse cornealinjury model. J. Biomed. Sci. 14, 313e322.
Lebherz, C., Maguire, A., Tang, W., Bennett, J., Wilson, J.M., 2008. Novel AAV sero-types for improved ocular gene transfer. J. Gene Med. 10, 375e382.
Lech, P., Somaia, N.V., 2008. Retrovirus vectors. In: Benigni, A., Remuzzi, G. (Eds.),Gene Therapy for Renal Diseases and Transplantation. Karger, Basel, pp. 30e46.
Lewis, J.D., Lichtenstein, G.R., Deren, J.J., Sands, B.E., Hanauer, S.B., Katz, J.A.,Lashner, B., Present, D.H., Chuai, S., Ellenberg, J.H., Nessel, L., Wu, G.D., Rosi-glitazone for Ulcerative Colitis Study Group, 2008. Rosiglitazone for activeulcerative colitis: a randomized placebo-controlled trial. Gastroenterology 134,688e695.
Li, D., Li, P., Li, G., Wang, J., Wang, E., 2009a. The effect of nocodazole on thetransfection efficiency of lipid-bilayer coated gold nanoparticles. Biomaterials30, 1382e1388.
Li, H.L., Zheng, X.Z., Wang, H.P., Li, F., Wu, Y., Du, L.F., 2009b. Ultrasound-targetedmicrobubble destruction enhances AAV-mediated gene transfection in humanRPE cells in vitro and rat retina in vivo. Gene Ther. 16, 1146e1153.
Li, M., Jayandharan, G.R., Li, B., Ling, C., Ma, W., Srivastava, A., Zhong, L., 2010. High-efficiency transduction of fibroblasts and mesenchymal stem cells by tyrosine-mutant AAV2 vectors for their potential use in cellular therapy. Hum. GeneTher. 21, 1527e1543.
Limberis, M.P., Wilson, J.M., 2006. Adeno-associated virus serotype 9 vectorstransduce murine alveolar and nasal epithelia and can be readministered. Proc.Natl. Acad. Sci. U.S.A. 103, 12993e12998.
Lindner, J.R., 2004. Microbubbles in medical imaging: current applications andfuture directions. Nat Rev Drug Discov 3, 527e532.
Liu, J., Saghizadeh, M., Tuli, S.S., Kramerov, A.A., Lewin, A.S., Bloom, D.C.,Hauswirth, W.W., Castro, M.G., Schultz, G.S., Ljubimov, A.V., 2008a. Differenttropism of adenoviruses and adeno-associated viruses to corneal cells: impli-cations for corneal gene therapy. Mol. Vis. 14, 2087e2096.
Liu, J., Lewin, A.S., Tuli, S.S., Ghivizzani, S.C., Schultz, G.S., Bloom, D.C., 2008b.Reduction in severity of a herpes simplex virus type 1 murine infection bytreatment with a ribozyme targeting the UL20 gene RNA. J. Virol. 82, 7467e7474.
Lipkowitz, M.S., Hanss, B., Tulchin, N., Wilson, P.D., Langer, J.C., Ross, M.D.,Kurtzman, G.J., Klotman, P.E., Klotman, M.E., 1999. Transduction of renal cellsin vitro and in vivo by adeno-associated virus gene therapy vectors. J. Am. Soc.Nephrol. 10, 1908e1915.
Liu, Y., Yang, H., Sakanishi, A., 2006. Ultrasound: mechanical gene transfer into plantcells by sonoporation. Biotechnol. Adv. 24, 1e16.
Lungwitz, U., Breunig, M., Blunk, T., Gopferich, A., 2005. Polyethylenimine basednon-viral gene delivery systems. Eur. J. Pharm. Biopharm. 60, 247e266.

R.R. Mohan et al. / Progress in Retinal and Eye Research 31 (2012) 43e64 63
McAlister, J.C., Joyce, N.C., Harris, D.L., Ali, R.R., Larkin, D.F., 2005. Induction ofreplication in human corneal endothelial cells by E2F2 transcription factorcDNA transfer. Invest. Ophthalmol. Vis. Sci. 46, 3597e3603.
Maguire, A.M., Simonelli, F., Pierce, E.A., Pugh Jr., E.N., Mingozzi, F., Bennicelli, J.,Banfi, S., Marshall, K.A., Testa, F., Surace, E.M., Rossi, S., Lyubarsky, A.,Arruda, V.R., Konkle, B., Stone, E., Sun, J., Jacobs, J., Dell’Osso, L., Hertle, R.,Ma, J.X., Redmond, T.M., Zhu, X., Hauck, B., Zelenaia, O., Shindler, K.S.,Maguire, M.G., Wright, J.F., Volpe, N.J., McDonnell, J.W., Auricchio, A., High, K.A.,Bennett, J., 2008. Safety and efficacy of gene transfer for Leber’s congenitalamaurosis. N. Engl. J. Med. 358, 2240e2248.
Maguire, A.M., High, K.A., Auricchio, A., Wright, J.F., Pierce, E.A., Testa, F., Mingozzi, F.,Bennicelli, J.L., Ying, G.S., Rossi, S., Fulton, A., Marshall, K.A., Banfi, S., Chung, D.C.,Morgan, J.I., Hauck, B., Zelenaia, O., Zhu, X., Raffini, L., Coppieters, F., De Baere, E.,Shindler, K.S., Volpe, N.J., Surace, E.M., Acerra, C., Lyubarsky, A., Redmond, T.M.,Stone, E., Sun, J., McDonnell, J.W., Leroy, B.P., Simonelli, F., Bennett, J., 2009. Age-dependent effects of RPE65 gene therapy for Leber’s congenital amaurosis:a phase 1 dose-escalation trial. Lancet 374, 1597e1605.
Marano, R.J., Toth, I., Wimmer, N., Brankov, M., Rakoczy, P.E., 2005. Dendrimerdelivery of an anti-VEGF oligonucleotide into the eye: a long-term study intoinhibition of laser-induced CNV, distribution, uptake and toxicity. Gene Ther. 12,1544e1550.
Maurice, D.M., 1984. Cornea and sclera. In: Davson, D. (Ed.), The Eye. AcademicPress, London, pp. 1e158.
Mintzer, M.A., Simanek, E.E., 2009. Nonviral vectors for gene delivery. Chem. Rev.109, 259e302.
Mitragotri, S., 2005. Healing sound: the use of ultrasound in drug delivery and othertherapeutic applications. Nat. Rev. Drug Discov. 4, 255e260.
Mo, Y., Barnett, M.E., Takemoto, D., Davidson, H., Kompella, U.B., 2007. Humanserum albumin nanoparticles for efficient delivery of Cu, Zn superoxide dis-mutase gene. Mol. Vis. 13, 746e757.
Mohan, R.R., Hutcheon, A.E., Choi, R., Hong, J., Lee, J., Mohan, R.R., Ambrósio Jr., R.,Zieske, J.D., Wilson, S.E., 2003. Apoptosis, necrosis, proliferation, and myofi-broblast generation in the stroma following LASIK and PRK. Exp. Eye Res. 76,71e87.
Mohan, R.R., Sharma, A., Netto, M.V., Sinha, S., Wilson, S.E., 2005. Gene therapy inthe cornea. Prog. Retin. Eye Res. 24, 537e559.
Mohan, R.R., Gupta, R., Mehan, M.K., Cowden, J.W., Sinha, S., 2010a. Decorintransfection suppresses profibrogenic genes and myofibroblast formation inhuman corneal fibroblasts. Exp. Eye Res. 91, 238e245.
Mohan, R.R., Sharma, A., Cebulko, T.C., Tandon, A., 2010b. Vector delivery techniqueaffects gene transfer in the cornea in vivo. Mol. Vis. 16, 2494e2501.
Mohan, R.R., Tovey, J.C., Gupta, R., Sharma, A., Tandon, A., 2011a. Decorinbiology, expression, function and therapy in the cornea. Curr. Mol. Med. 11,110e128.
Mohan, R.R., Tovey, J.C., Sharma, A., Schultz, G., Cowden, J., Tandon, A., 2011b.Targeted decorin gene therapy delivered with adeno-associated viruseffectively retards corneal neovascularization in vivo. PLoS One, in press.
Mohan, R.R., Tandon, A., Sharma, A., Cowden, J.W., Tovey, J.C., 2011c. Significantinhibition of corneal scarring in vivo with tissue-selective, targeted AAV5decorin gene therapy. Invest Ophthalmol Vis Sci. 52, 4833e4841.
Montini, E., Cesana, D., Schmidt, M., Sanvito, F., Ponzoni, M., Bartholomae, C., SergiSergi, L., Benedicenti, F., Ambrosi, A., Di Serio, C., Doglioni, C., von Kalle, C.,Naldini, L., 2006. Hematopoietic stem cell gene transfer in a tumor-pronemouse model uncovers low genotoxicity of lentiviral vector integration. Nat.Biotechnol. 24, 687e696.
Montini, E., Cesana, D., Schmidt, M., Sanvito, F., Bartholomae, C.C., Ranzani, M.,Benedicenti, F., Sergi, L.S., Ambrosi, A., Ponzoni, M., Doglioni, C., Di Serio, C., vonKalle, C., Naldini, L., 2009. The genotoxic potential of retroviral vectors isstrongly modulated by vector design and integration site selection in a mousemodel of HSC gene therapy. J. Clin. Invest. 119, 964e975.
Mulhbacher, J., St-Pierre, P., Lafontaine, D.A., 2010. Therapeutic applications ofribozymes and riboswitches. Curr. Opin. Pharmacol. 10, 551e556.
Munn, D.H., Shafizadeh, E., Attwood, J.T., Bondarev, I., Pashine, A., Mellor, A.L., 1999.Inhibition of T cell proliferation by macrophage tryptophan catabolism. J. Exp.Med. 189, 1363e1372.
Murthy, R.C., McFarland, T.J., Yoken, J., Chen, S., Barone, C., Burke, D., Zhang, Y.,Appukuttan, B., Stout, J.T., 2003. Corneal transduction to inhibit angiogenesisand graft failure. Invest. Ophthalmol. Vis. Sci. 44, 1837e1842.
Mwaikambo, B.R., Sennlaub, F., Ong, H., Chemtob, S., Hardy, P., 2006. Activation ofCD36 inhibits and induces regression of inflammatory corneal neo-vascularization. Invest. Ophthalmol. Vis. Sci. 47, 4356e4364.
Netto, M.V., Mohan, R.R., Medeiros, F.W., Dupps Jr., W.J., Sinha, S., Krueger, R.R.,Stapleton, W.M., Rayborn, M., Suto, C., Wilson, S.E., 2007. Femtosecond laser andmicrokeratome corneal flaps: comparison of stromal wound healing andinflammation. J. Refract. Surg. 23, 667e676.
O’Reilly, M.S., Boehm, T., Shing, Y., Fukai, N., Vasios, G., Lane, W.S., Flynn, E.,Birkhead, J.R., Olsen, B.R., Folkman, J., 1997. Endostatin: an endogenous inhibitorof angiogenesis and tumor growth. Cell 88, 277e285.
Oh, J.Y., Roddy, G.W., Choi, H., Lee, R.H., Ylöstalo, J.H., Rosa Jr., R.H., Prockop, D.J.,2010. Anti-inflammatory protein TSG-6 reduces inflammatory damage to thecornea following chemical and mechanical injury. Proc. Natl. Acad. Sci. U.S.A.107, 16875e16880.
Pacak, C.A., Mah, C.S., Thattaliyath, B.D., Conlon, T.J., Lewis, M.A., Cloutier, D.E.,Zolotukhin, I., Tarantal, A.F., Byrne, B.J., 2006. Recombinant adeno-associated
virus serotype 9 leads to preferential cardiac transduction in vivo. Circ. Res.99, e3e9.
Palmer, D.J., Ng, P., 2005. Helper-dependent adenoviral vectors for gene therapy.Hum. Gene Ther. 16, 1e16.
Parker, D.G., Kaufmann, C., Brereton, H.M., Anson, D.S., Francis-Staite, L., Jessup, C.F.,Marshall, K., Tan, C., Koldej, R., Coster, D.J., Williams, K.A., 2007. Lentivirus-mediated gene transfer to the rat, ovine and human cornea. Gene Ther. 14,760e767.
Parker, D.G., Brereton, H.M., Klebe, S., Coster, D.J., Williams, K.A., 2009. A steroid-inducible promoter for the cornea. Br. J. Ophthalmol. 93, 1255e1259.
Pellegrini, G., Rama, P., Mavilio, F., De Luca, M., 2009. Epithelial stem cells in cornealregeneration and epidermal gene therapy. J. Pathol. 217, 217e228.
Petrs-Silva, H., Dinculescu, A., Li, Q., Min, S.H., Chiodo, V., Pang, J.J., Zhong, L.,Zolotukhin, S., Srivastava, A., Lewin, A.S., Hauswirth, W.W., 2009. High-effi-ciency transduction of the mouse retina by tyrosine-mutant AAV serotypevectors. Mol. Ther. 17, 463e471.
Petry, H., Brooks, A., Orme, A., Wang, P., Liu, P., Xie, J., Kretschmer, P., Qian, H.S.,Hermiston, T.W., Harkins, R.N., 2008. Effect of viral dose on neutralizing anti-body response and transgene expression after AAV1 vector re-administration inmice. Gene Ther. 15, 54e60.
Pillai, R.G., Beutelspacher, S.C., Larkin, D.F., George, A.J., 2008. Expression of thechemokine antagonist vMIP II using a non-viral vector can prolong cornealallograft survival. Transplantation 85, 1640e1647.
Pissuwan, D., Niidome, T., Cortie, M.B., 2011. The forthcoming applications of goldnanoparticles in drug and gene delivery systems. J. Control. Release 149, 65e71.
Ponder, K.P., Auricchio, A., 2010. Gene therapy for ocular problems in mucopoly-saccharidosis: an experimental and promising approach with benefits in animalmodels e a review. Clin. Exp. Ophthalmol. 38, 43e51.
Qaddoumi, M.G., Ueda, H., Yang, J., Davda, J., Labhasetwar, V., Lee, V.H., 2004. Thecharacteristics and mechanisms of uptake of PLGA nanoparticles in rabbitconjunctival epithelial cell layers. Pharm. Res. 21, 641e648.
Ragusa, A., García, I., Penadés, S., 2007. Nanoparticles as nonviral gene deliveryvectors. IEEE Trans. Nanobiosci. 6, 319e330.
Rama, P., Matuska, S., Paganoni, G., Spinelli, A., De Luca, M., Pellegrini, G., 2010.Limbal stem-cell therapy and long-term corneal regeneration. N. Engl. J. Med.363, 147e155.
Redfern, R.L., McDermott, A.M., 2010. Toll-like receptors in ocular surface disease.Exp. Eye Res. 90, 679e687.
Richter, M., Iwata, A., Nyhuis, J., Nitta, Y., Miller, A.D., Halbert, C.L., Allen, M.D., 2000.Adeno-associated virus vector transduction of vascular smooth muscle cellsin vivo. Physiol. Genom. 2, 117e127.
Ritter, T., Yang, J., Dannowski, H., Vogt, K., Volk, H.D., Pleyer, U., 2007. Effects ofinterleukin-12p40 gene transfer on rat corneal allograft survival. Transpl.Immunol. 18, 101e107.
Runager, K., Enghild, J.J., Klintworth, G.K., 2008. Focus on molecules: transforminggrowth factor beta induced protein (TGFBIp). Exp. Eye Res. 87, 298e299.
Saghizadeh, M., Kramerov, A.A., Tajbakhsh, J., Aoki, A.M., Wang, C., Chai, N.N.,Ljubimova, J.Y., Sasaki, T., Sosne, G., Carlson, M.R., Nelson, S.F., Ljubimov, A.V.,2005. Proteinase and growth factor alterations revealed by gene microarrayanalysis of human diabetic corneas. Invest. Ophthalmol. Vis. Sci. 46, 3604e3615.
Saghizadeh, M., Kramerov, A.A., Yu, F.S., Castro, M.G., Ljubimov, A.V., 2010.Normalization of wound healing and diabetic markers in organ cultured humandiabetic corneas by adenoviral delivery of c-Met gene. Invest. Ophthalmol. Vis.Sci. 51, 1970e1980.
Saika, S., Ikeda, K., Yamanaka, O., Miyamoto, T., Ohnishi, Y., Sato, M., Muragaki, Y.,Ooshima, A., Nakajima, Y., Kao, W.W., Flanders, K.C., Roberts, A.B., 2005a.Expression of Smad7 in mouse eyes accelerates healing of corneal tissue afterexposure to alkali. Am. J. Pathol. 166, 1405e1418.
Saika, S., Ikeda, K., Yamanaka, O., Flanders, K.C., Nakajima, Y., Miyamoto, T.,Ohnishi, Y., Kao, W.W., Muragaki, Y., Ooshima, A., 2005b. Therapeutic effects ofadenoviral gene transfer of bone morphogenic protein-7 on a corneal alkaliinjury model in mice. Lab. Invest. 85, 474e486.
Saika, S., Yamanaka, O., Okada, Y., Miyamoto, T., Kitano, A., Flanders, K.C., Ohnishi, Y.,Nakajima, Y., Kao, W.W., Ikeda, K., 2007. Effect of overexpression of PPARgammaon the healing process of corneal alkali burn in mice. Am. J. Physiol. Cell.Physiol. 293, C75eC86.
Sharma, A., Ghosh, A., Siddappa, C., Mohan, R.R., 2010a. Ocular surface: genetherapy. In: Besharse, J., Dana, R., Dartt, D.A. (Eds.), Encyclopedia of the Eye.Elsevier, pp. 185e194.
Sharma, A., Ghosh, A., Hansen, E.T., Newman, J.M., Mohan, R.R., 2010b. Transductionefficiency of AAV 2/6, 2/8 and 2/9 vectors for delivering genes in human cornealfibroblasts. Brain Res. Bull. 81, 273e278.
Sharma, A., Tovey, J.C., Ghosh, A., Mohan, R.R., 2010c. AAV serotype influences genetransfer in corneal stroma in vivo. Exp. Eye Res. 91, 440e448.
Sharma, A., Tandon, A., Tovey, J.C., Gupta, R., Robertson, J.D., Fortune, J.A.,Klibanov, A.M., Cowden, J.W., Rieger, F.G., Mohan, R.R., 2011. Polyethylenimine-conjugated gold nanoparticles: gene transfer potential and low toxicity in thecornea. Nanomedicine PMID: 21272669.
Shiraishi, A., Converse, R.L., Liu, C.Y., Zhou, F., Kao, C.W., Kao, W.W., 1998. Identifi-cation of the cornea-specific keratin 12 promoter by in vivo particle-mediatedgene transfer. Invest. Ophthalmol. Vis. Sci. 39, 2554e2561.
Shoham, A., Hadziahmetovic, M., Dunaief, J.L., Mydlarski, M.B., Schipper, H.M., 2008.Oxidative stress in diseases of the human cornea. Free Radic. Biol. Med. 45,1047e1055.

R.R. Mohan et al. / Progress in Retinal and Eye Research 31 (2012) 43e6464
Simonelli, F., Maguire, A.M., Testa, F., Pierce, E.A., Mingozzi, F., Bennicelli, J.L.,Rossi, S., Marshall, K., Banfi, S., Surace, E.M., Sun, J., Redmond, T.M., Zhu, X.,Shindler, K.S., Ying, G.S., Ziviello, C., Acerra, C., Wright, J.F., McDonnell, J.W.,High, K.A., Bennett, J., Auricchio, A., 2010. Gene therapy for Leber’s congenitalamaurosis is safe and effective through 1.5 years after vector administration.Mol. Ther. 18, 643e650.
Singh, N., Amin, S., Richter, E., Rashid, S., Scoglietti, V., Jani, P.D., Wang, J., Kaur, R.,Ambati, J., Dong, Z., Ambati, B.K., 2005. Flt-1 intraceptors inhibit hypoxia-induced VEGF expression in vitro and corneal neovascularization in vivo.Invest. Ophthalmol. Vis. Sci. 46, 1647e1652.
Sonoda, S., Tachibana, K., Uchino, E., Okubo, A., Yamamoto, M., Sakoda, K.,Hisatomi, T., Sonoda, K.H., Negishi, Y., Izumi, Y., Takao, S., Sakamoto, T., 2006.Gene transfer to corneal epithelium and keratocytes mediated by ultrasoundwith microbubbles. Invest. Ophthalmol. Vis. Sci. 47, 558e564.
Stramer, B.M., Zieske, J.D., Jung, J.C., Austin, J.S., Fini, M.E., 2003. Molecular mech-anisms controlling the fibrotic repair phenotype in cornea: implications forsurgical outcomes. Invest. Ophthalmol. Vis. Sci. 44, 4237e4246.
Surace, E.M., Auricchio, A., 2008. Versatility of AAV vectors for retinal gene transfer.Vision Res. 48, 353e359.
Tandon, A., Tovey, J.C., Sharma, A., Gupta, R., Mohan, R.R., 2010. Role of transforminggrowth factor Beta in corneal function, biology and pathology. Curr. Mol. Med.10, 565e578.
The Journal of Gene Medicine Clinical Trial Website. update June 2010 Available at:http://www.abedia.com/wiley/index.html (accessed 23.05.11).
Thomas, M., Klibanov, A.M., 2003. Conjugation to gold nanoparticles enhancespolyethylenimine’s transfer of plasmid DNA into mammalian cells. Proc. Natl.Acad. Sci. U.S.A. 100, 9138e9143.
Thomas, M., Ge, Q., Lu, J.J., Chen, J., Klibanov, A.M., 2005. Cross-linked small poly-ethylenimines: while still nontoxic, deliver DNA efficiently to mammalian cellsin vitro and in vivo. Pharm. Res. 22, 373e380.
Tiyaboonchai, W., Woiszwillo, J., Middaugh, C.R., 2003. Formulation and charac-terization of DNAepolyethylenimineedextran sulfate nanoparticles. Eur. J.Pharm. Sci. 19, 191e202.
Tong, Y.C., Chang, S.F., Liu, C.Y., Kao, W.W., Huang, C.H., Liaw, J., 2007. Eye dropdelivery of nano-polymeric micelle formulated genes with cornea-specificpromoters. J. Gene Med. 9, 956e966.
Toropainen, E., Hornof, M., Kaarniranta, K., Johansson, P., Urtti, A., 2007. Cornealepithelium as a platform for secretion of transgene products after transfectionwith liposomal gene eyedrops. J. Gene Med. 9, 208e216.
Traas, A.M., Wang, P., Ma, X., Tittiger, M., Schaller, L., O’donnell, P., Sleeper, M.M.,Vite, C., Herati, R., Aguirre, G.D., Haskins, M., Ponder, K.P., 2007. Correction ofclinical manifestations of canine mucopolysaccharidosis I with neonatal retro-viral vector gene therapy. Mol. Ther. 15, 1423e1431.
Van Vliet, K.M., Blouin, V., Brument, N., Agbandje-McKenna, M., Snyder, R.O., 2008.The role of the adeno-associated virus capsid in gene transfer. Methods Mol.Biol. 437, 51e91.
Wang, B., O’Malley, T.M., Xu, L., Vite, C., Wang, P., O’Donnell, P.A., Ellinwood, N.M.,Haskins, M.E., Ponder, K.P., 2006. Expression in blood cells may contribute tobiochemical and pathological improvements after neonatal intravenous genetherapy for mucopolysaccharidosis VII in dogs. Mol. Genet. Metab. 87, 8e21.
Wang, Z., Zhu, T., Qiao, C., Zhou, L., Wang, B., Zhang, J., Chen, C., Li, J., Xiao, X., 2005.Adeno-associated virus serotype 8 efficiently delivers genes to muscle andheart. Nat. Biotechnol. 23, 321e328.
Watanabe, K., Hasegawa, Y., Yamashita, H., Shimizu, K., Ding, Y., Abe, M., Ohta, H.,Imagawa, K., Hojo, K., Maki, H., Sonoda, H., Sato, Y., 2004. Vasohibin as anendothelium-derived negative feedback regulator of angiogenesis. J. Clin.Invest. 114, 898e907.
Weiss, J.S., Kruth, H.S., Kuivaniemi, H., Tromp, G., White, P.S., Winters, R.S., Lisch, W.,Henn, W., Denninger, E., Krause, M., Wasson, P., Ebenezer, N., Mahurkar, S.,Nickerson, M.L., 2007. Mutations in the UBIAD1 gene on chromosome short arm
1, region 36, cause Schnyder crystalline corneal dystrophy. Invest. Ophthalmol.Vis. Sci. 48, 5007e5012.
Weiss, J.S., Fifth ARVO/Pfizer Ophthalmics Research Institute Conference WorkingGroup, 2010. Corneal dystrophies: molecular genetics to therapeutic inter-ventiondFifth ARVO/Pfizer Ophthalmics Research Institute Conference. Invest.Ophthalmol. Vis. Sci. 51, 5391e5402.
Whitcher, J.P., Srinivasan, M., Upadhyay, M.P., 2001. Corneal blindness: a globalperspective. Bull. World Health Org. 79, 214e221.
Williams, K.A., Coster, D.J., 2010. Gene therapy for diseases of the cornea e a review.Clin. Exp. Ophthalmol. 38, 93e103.
Wilson, S.E., Mohan, R.R., Hong, J., Lee, J., Choi, R., Liu, J.J., Mohan, R.R., 2002.Apoptosis in the cornea in response to epithelial injury: significance to woundhealing and dry eye. Adv. Exp. Med. Biol. 506, 821e826.
World Health Organization Report, fact sheet 282, April 2011. Visual Impairmentand Blindness. URL: http://www.who.int/mediacentre/factsheets/fs282/en/index.html (accessed 23.09.11).
Wu, Z., Miller, E., Agbandje-McKenna, M., Samulski, R.J., 2006. Alpha2,3 and alpha2,6 N-linked sialic acids facilitate binding and transduction by adenoassociatedvirus types 1 and 6. J. Virol. 80, 9093e9103.
Xu, Q., Wang, C.H., Pack, D.W., 2010. Polymeric carriers for gene delivery: chitosanand poly(amidoamine) dendrimers. Curr. Pharm. Des. 16, 2350e2368.
Yamashita, T., Sonoda, S., Suzuki, R., Arimura, N., Tachibana, K., Maruyama, K.,Sakamoto, T., 2007. A novel bubble liposome and ultrasound-mediated genetransfer to ocular surface: RC-1 cells in vitro and conjunctiva in vivo. Exp. EyeRes. 85, 741e748.
Yoon, K.C., Bae, J.A., Park, H.J., Im, S.K., Oh, H.J., Lin, X.H., Kim, M.Y., Lee, J.H., Lee, S.E.,Ahn, K.Y., Kim, K.K., 2009. Subconjunctival gene delivery of the transcriptionfactor GA-binding protein delays corneal neovascularization in a mouse model.Gene Ther. 16, 973e981.
Young, L.S., Searle, P.F., Onion, D., Mautner, V., 2006. Viral gene therapy strategies:from basic science to clinical application. J. Pathol. 208, 299e318.
Yu, H., Wu, J., Li, H., Wang, Z., Chen, X., Tian, Y., Yi, M., Ji, X., Ma, J., Huang, Q., 2007.Inhibition of corneal neovascularization by recombinant adenovirus-mediatedsFlk-1 expression. Biochem. Biophys. Res. Commun. 361, 946e952.
Yuan, J., Chen, J.Q., Zhou, S.Y., Liu, Z.G., Wang, Z.C., Gu, J.J., 2006. The transfection andexpression of IL-1ra gene to the rabbit cornea in situ via cation polymermediation. Zhonghua Yan Ke Za Zhi 42, 686e693.
Zagon, I.S., Sassani, J.W., Malefyt, K.J., McLaughlin, P.J., 2006. Regulation of cornealrepair by particle-mediated gene transfer of opioid growth factor receptorcomplementary DNA. Arch. Ophthalmol. 124, 1620e1624.
Zarbin, M.A., Montemagno, C., Leary, J.F., Ritch, R., 2010a. Nanomedicine inophthalmology: the new frontier. Am. J. Ophthalmol. 150, 144e162.
Zarbin, M.A., Montemagno, C., Leary, J.F., Ritch, R., 2010b. Nanotechnology inophthalmology. Can. J. Ophthalmol. 45, 457e476.
Zhang, B., Yao, Y.F., 2010. Gelatinous drop-like corneal dystrophy with a novelmutation of TACSTD2 manifested in combination with spheroidal degenerationin a Chinese patient. Mol. Vis. 16, 1570e1575.
Zhong, L., Li, B., Jayandharan, G., Mah, C.S., Govindasamy, L., Agbandje-McKenna, M.,Herzog, R.W., Weigel-Van Aken, K.A., Hobbs, J.A., Zolotukhin, S., Muzyczka, N.,Srivastava,A., 2008. Tyrosine-phosphorylationofAAV2vectors anditsconsequenceson viral intracellular trafficking and transgene expression. Virology 381, 194e202.
Zhou, R., Dean, D.A., 2007. Gene transfer of interleukin 10 to the murine corneausing electroporation. Exp. Biol. Med. (Maywood) 232, 362e369.
Zhou, S.Y., Xie, Z.L., Xiao, O., Yang, X.R., Heng, B.C., Sato, Y., 2010. Inhibition of mousealkali burn induced-corneal neovascularization by recombinant adenovirusencoding human vasohibin-1. Mol. Vis. 16, 1389e1398.
Zhou, X., Zhang, X., Yu, X., Zha, X., Fu, Q., Liu, B., Wang, X., Chen, Y., Chen, Y., Shan, Y.,Jin, Y., Wu, Y., Liu, J., Kong, W., Shen, J., 2008. The effect of conjugation to goldnanoparticles on the ability of low molecular weight chitosan to transfer DNAvaccine. Biomaterials 29, 111e117.




















