
The engine of thought is a hybrid: Roles of associative and structured knowledge in reasoning
75
RUNNING HEAD: Knowledge and Inductive Reasoning The engine of thought is a hybrid: Roles of associative and structured knowledge in reasoning Aimée K. Bright Queen Mary University of London Aidan Feeney Queen’s University Belfast Author Note Aimée K. Bright, School of Biological and Chemical Sciences, Psychology Division, Queen Mary University of London; Aidan Feeney, Department of Psychology, Queen’s University Belfast This research was supported by an ESRC PhD studentship awarded to Aimée K. Bright Correspondence concerning this article should be addressed to Aimée K. Bright, School of Biological and Chemical Sciences, Psychology Division, Queen Mary University of London, E1 4NS, London, United Kingdom. Email: [email protected] Word Count: 15,041
Transcript of The engine of thought is a hybrid: Roles of associative and structured knowledge in reasoning

RUNNING HEAD: Knowledge and Inductive Reasoning
The engine of thought is a hybrid: Roles of associative and structured knowledge in
reasoning
Aimée K. Bright
Queen Mary University of London
Aidan Feeney
Queen’s University Belfast
Author Note
Aimée K. Bright, School of Biological and Chemical Sciences, Psychology Division,
Queen Mary University of London; Aidan Feeney, Department of Psychology, Queen’s
University Belfast
This research was supported by an ESRC PhD studentship awarded to Aimée K.
Bright
Correspondence concerning this article should be addressed to Aimée K. Bright,
School of Biological and Chemical Sciences, Psychology Division, Queen Mary University
of London, E1 4NS, London, United Kingdom.
Email: [email protected]
Word Count: 15,041

Knowledge and Inductive Reasoning
Abstract
Across a range of domains in psychology different theories assume different mental
representations of knowledge. For example, in the literature on category-based inductive
reasoning, certain theories (e.g. Rogers & McClelland, 2004; Sloutsky & Fisher, 2008)
assume that the knowledge upon which inductive inferences are based is associative whereas
others (e.g Heit & Rubinstein, 1994; Kemp & Tenenbaum, 2009; Osherson, Smith, Wilkie,
Lopez, & Shafir, 1990) assume that knowledge is structured. In this paper we investigate
whether associative and structured knowledge underlie inductive reasoning to different
degrees under different processing conditions. We develop a measure of knowledge about the
degree of association between categories and show that it dissociates from measures of
structured knowledge. In Experiment 1 participants rated the strength of inductive arguments
whose categories were either taxonomically or causally related. A measure of associative
strength predicted reasoning when people had to respond fast, whereas causal and taxonomic
knowledge explained inference strength when people responded slowly. In Experiment 2, we
also manipulated whether the causal link between the categories was predictive or diagnostic.
Participants preferred predictive to diagnostic arguments except when they responded under
cognitive load. In Experiment 3, using an open-ended induction paradigm, people generated
and evaluated their own conclusion categories. Inductive strength was predicted by
associative strength under heavy cognitive load, whereas an index of structured knowledge
was more predictive of inductive strength under minimal cognitive load. Together these
results suggest that associative and structured models of reasoning apply best under different
processing conditions, and that the application of structured knowledge in reasoning is often
effortful.
Keywords: inductive reasoning, associative, causal knowledge, categories

Introduction
Inductive reasoning has been referred to as the engine of thought because it allows us
to use what we already know to reduce our uncertainty about the way the world is. To ensure
that our inductive inferences are potent but sufficiently constrained, it is crucial that we select
the appropriate background knowledge. For example, if you eat shrimps and get ill, in future
you might avoid eating shrimps and other members of the shellfish family. However, if you
realize that the shrimps were past their expiration date, you might instead avoid eating meat,
dairy or vegetables past their expiration date, but happily eat fresh shrimps in future. As yet,
our understanding about the form taken by the knowledge that drives our inductive inferences
and how reasoners select the most appropriate knowledge with which to constrain a particular
inference is relatively poor. In this paper we will argue that the inductive engine of thought is
a hybrid, taking both associative and structured knowledge as input. Because associative
knowledge can influence the reasoning process relatively automatically and effortlessly,
whereas the influence of structured knowledge is more cognitively demanding and time-
consuming, environmental conditions are likely to determine which type of knowledge is
most important for a particular inference. In a series of experiments where processing
conditions were manipulated, we examined whether associative and structured knowledge
have dissociable effects on inductive reasoning.
In this paper we will focus on category-based inductive reasoning as it has been the
focus of much recent attention, and there are well specified models of category-based
induction which make different representational assumptions. In a category-based inductive
argument participants are asked to evaluate how strongly knowing that members of one
category possess a property supports the conclusion that members of another category do. For
example, knowing that shrimp possess property A how likely is it that mussels also possess
that property? Although early studies of category based induction (e.g. Osherson et al., 1990;

Rips, 1975) tended to use properties about which participants were unlikely to possess
background knowledge, subsequent studies showed the importance of the type of property in
the argument (see Heit & Rubenstein, 1994; Osherson, et al., 1993). More recent work
(Coley, 2012; for a review see Vasilyeva & Coley, 2010) has emphasized the different types
of knowledge made available by taxonomic (genes, cells) versus ecological properties
(infections, diseases).
Although the basic category-based inductive reasoning paradigm appears simple it
has generated quite a number of models (for a review see chapters in Feeney & Heit, 2007).
Because models of category-based reasoning appear to make contrasting representational
suppositions, which means that knowledge will be represented in only one form, there is a
pressing need for synthesis. We will consider the implications of our results for other types of
inductive reasoning and for a variety of modeling frameworks in the General Discussion. We
will begin with a review of literature highlighting the difference between associative and
structured knowledge and the role each type of knowledge plays in the reasoning process. We
then review current theories of category-based induction, broadly distinguishing them by the
contrasting emphasis they place on structured and associative knowledge. We then argue that
a process-level account can resolve the apparent dichotomy between frameworks which
emphasize different types of knowledge. This account predicts that the extent to which
associative and structured knowledge serve as input to reasoning processes will depend on
processing conditions.
Associative and Structured Knowledge
There are multiple types of knowledge that result from perception, learning and
reasoning. Here we want to focus on associative and structured knowledge. Broadly
speaking, associative knowledge develops as a result of co-occurrence, similarity or
contiguity (e.g. Kruschke, 1992; McClelland & Rumelhart, 1985; Rescorla & Wagner, 1972).

What we will refer to as structured knowledge captures the relations that exist between things
in the world. These relations can be causal, taxonomic, functional, spatial or take some other
form. Although the term “relational” is commonly used to refer to the type of knowledge we
wish to consider here (see Halford, Wilson, & Phillips, 2010), as “relational” theories of
category-based induction often posit the existence of complex structures of relations between
categories, we prefer the term “structured knowledge”. However, we mean the term
“structured” to refer to both complex (Kemp & Tenenbaum, 2009) and relatively simple
(Halford et al., 2010) relational knowledge.
There have been numerous attempts to define the characteristics of structured or
relational knowledge (see Halford et al., 2010; Markman, 1999). From our point of view,
perhaps the most important is that categories or objects are bound to a particular role by their
involvement in a particular relation (see Holyoak, 2012). Thus, in the context of induction,
the relation provides a mechanism by which the two categories involved in the relation come
to share a property. For example, we may think it likely that horses and cows share a gene
because their relative positions in the taxonomic hierarchy mean that they stand in close
relation to one another, but think it unlikely that horses and eagles will share a gene because
the taxonomic relation between them is not strong (see Osherson et al., 1990; Shafto, Kemp,
Bonawitz, Coley, & Tenenbaum, 2008). Similarly, we may think it likely that rabbits will
have a disease given that carrots have it, because their involvement in a food web (see Shafto
et al., 2008) may provide an unseen causal mechanism for disease transmission. This last
example clearly illustrates the importance of role-binding as carrots serve as the cause and
rabbits possessing the disease as the effect. These roles stay the same even when we reason
diagnostically i.e. when we consider the likelihood that carrots have a disease given that
rabbits have it (see Fernbach, Darlow, & Sloman, 2011).

Associative and structured knowledge appear to differ in a number of ways. First, the
knowledge types appear to have different developmental trajectories. Andrews, Halford and
Boyce (2012) demonstrated that children between 4 and 6 years could learn conditional
discriminations without awareness of the conditional relation. They argued that awareness of
the relation indicated relational rather than associative knowledge. Awareness of the
relational structure increased with age and was associated with a measure of fluid
intelligence. This suggests that the ability to represent and make use of structured relational
knowledge is an effortful process that develops with age.
Evidence for a second difference between the knowledge types comes from
comparative studies. Results in this area are contentious, and some authors have suggested
for example, that baboons in the wild are sensitive to hierarchical structure (Bergman,
Beehner, Cheney, & Seyfarth, 2003). However, Penn, Holyoak and Povenilli (2008) have
argued that whilst humans are not the only animals to possess associative knowledge,
possession of knowledge in the form of symbolic relational structure is unique to humans.
Furthermore, they argue that the extant evidence on animal’s sensitivity to hierarchical and
causal relations (see Bergman et al., 2003; Seed, Tebbich, Emery, & Clayton, 2006) suggests
that animals do not possess human-like sensitivity to unseen causal mechanisms or to
hierarchical relational structure (for a recent alternative view see Taylor, Miller, & Gray,
2013).
A third difference between these knowledge types is that requesting participants to
use one or other type of knowledge leads to different patterns of performance. For example,
Fenker, Waldman and Holyoak (2005) showed that there is a causal asymmetry effect in
reaction times when people evaluate the existence of causal relations between pairs of
categories (e.g. spark & flame). Reaction times were faster when the categories were
presented in a predictive (spark flame) compared to a diagnostic order (flame spark).

However, the asymmetry effect disappeared for evaluations of an association between the
same category pairs. This suggests that relations in semantic memory can be represented
either by explicitly encoded higher-order structural relations, or by simpler associations
between entities which cannot be reduced to one another. This also implies that the same
information can be encoded in different forms even if it is redundant (Sun, 1994; 2002).
In the rest of this paper we will consider whether the different types of process which
operate over associative and structured knowledge might have dissociable effects on
inductive reasoning. This possibility has not been considered before, but as we will see in the
following review, it could offer a way of synthesising apparently contradictory theoretical
frameworks for explaining category-based induction.
Associative Knowledge for Inductive Reasoning
The rationale for assuming that associative knowledge must play a central role in
inductive reasoning comes from a vast developmental literature (Colunga & Smith, 2005;
French, Mareschal, Mermillod, & Quinn, 2004; Jones & Smith, 2002; Sloutsky, Kloos, &
Fisher, 2007). These studies suggest that early induction is heavily influenced by the
statistical properties inherent in the environment such as co-occurrence and statistical
distribution of perceptual features. For example, Sloutsky and Fisher (2008) found that
associative learning mechanisms could explain young children’s flexible reasoning patterns
in the absence of higher-level, conceptual knowledge.These researchers also assume that
there is only a gradual and developmentally late transition from exclusive reliance on
associative knowledge to the use of category membership as a basis for induction (Sloutsky
& Fisher, 2008). This transition is largely seen as the product of explicit instruction and
learning about general characteristics of categories (Fisher & Sloutsky, 2005). However,
despite category representations being supplemented and expanded by top-down knowledge,

proponents of associative approaches advocate that adult induction is still heavily influenced
by similarity (Sloman, 1993, 1998) and associations in semantic memory (Rogers &
McClelland, 2004; Sloman, 1998). For example, Sloman’s (1993) Feature-Based Model
(FBM) explains generalizations in terms of the extent to which features which are associated
with different categories are simultaneously activated. The more associated features they
have in common, the more willing people should be to infer that the categories share a
property. Without reference to category membership, this neural network model can explain
several inductive reasoning phenomena. Sloman (1998) does not preclude the possibility that
assessment of similarity and inference process can at times reflect a more effortful process
which draws on knowledge about stable category hierarchies. However, he does suggest that
the default mode of category-based induction reflects a predominantly intuitive thought
process, requiring no processing effort or reference to class inclusion relations, especially
when people lack relevant knowledge, are under time pressure or have not been explicitly
instructed to carefully consider their response.
Smith and DeCoster (2000) link this associative view to underlying memory
processes. A slow-learning system gradually forms stable representations of the environment
based on the accumulation of similar experiences. When similar situations or objects are
encountered in the future, there is an automatic activation of these knowledge representations,
which deliver information and shape behaviour without the need for deliberate or effortful
thought. Thus, this kind of reasoning predominantly delivers responses which encode
statistical properties of the environment.
Smith and DeCoster’s suggestions about the nature of associative induction resonate
strongly with the characteristics of connectionist networks. A connectionist account of
category-based induction is given by Rogers and McClelland’s (2004) Parallel Distributed
Processing (PDP) model. It assumes that semantic information about an instance is stored as

an internal representation encoded by the pattern of distributed activation across processing
units. Knowledge is refined by gradually adjusting the weight of connections between
concepts (e.g. sparrows) and attributes (e.g. wings, chirp, small) for different kinds of
relations (e.g. has, can, is). This means that generalizations from one instance to another will
be strong to the extent that the activated distributed representations of the two instances
overlap via their shared attributes. Note that McClelland and colleagues (McClelland et al.,
2010) demonstrate that a structure can emerge though the gradual adjustments made to the
connections between concepts. However, this is not equivalent to having structured
knowledge about how the concepts are interrelated (Halford et al., 2010). The emergent
structures in connectionist networks represent connections between concrete concepts and
instances, that is, the nodes cannot act as abstract place holders that describe higher-order
theoretical interrelationships between concepts. Thus, theories which emphasize such higher
level theories and interrelationships would appear to stand in direct opposition to frameworks
emphasizing associative knowledge.
Structured Knowledge in Induction
Osherson et al’s (1990) Similarity-Coverage Model (SCM) was one of the earliest
models to emphasize the importance of knowledge about stable taxonomic structure when
evaluating categorical arguments. In contrast to Sloman’s (1993) feature-based
conceptualization of similarity, Osherson et al’s (1990) notion of similarity relies far more on
explicit knowledge about category relations between category instances. Thus, inductive
evaluations reflect the weighted sum of two primary parameters, similarity and coverage.
Similarity refers to the maximum average similarity between the premise and conclusion
categories. Coverage assumes that people have knowledge about stable category hierarchies
and refers to the degree to which the premise categories cover the category space of the
inclusive superordinate category.

The coverage component of the model gives rise to the diversity effect (for a
discussion see Heit, Hahn, & Feeney, 2005), whereby dissimilar premise categories act as
stronger evidence than similar premise categories. Although this phenomenon can be
explained by Sloman’s (1993) model, other evidence suggests that certain characteristics of
this effect are not explicable in terms of associative knowledge, instead requiring reference to
structured knowledge. For example, Feeney (2007) showed that only a minority of people
manifest the diversity effect when they reason about specific (e.g. rabbits) rather than general
(e.g. mammals) conclusions, and that this was related to cognitive ability. Developmental
evidence by Rhodes, Brickman and Gelman (2008) showed that even when a diverse
sampling strategy is explicitly suggested as a basis for making generalisations, children under
the age of eight years still prefer the non-diverse choice. It appears that in order to show
adult-like inductive reasoning, there must be a qualitative developmental shift in how
children use their category knowledge and acquire an appreciation of group-level properties
to make inductive inferences. For example, Lopez and colleagues (Lopez, Gelman, Gutheil,
& Smith, 1992) showed that several developmental changes seemed to depend on knowledge
about stable category hierarchies. The primary aim of these researchers was to identify
separate components which comprise the coverage parameter in Osherson et al’s (1990)
model. However, their findings are also consistent with the idea that the emergence of
computationally more complex inductive reasoning phenomena lags behind those that are
thought to rely on computationally simpler associative processes. For example, in the case of
the diversity effect for specific arguments, children first have to generate an inclusive
superordinate category before calculating coverage, a process that requires knowledge about
structured taxonomic relations between categories. Lopez et al. (1992) showed that 9-year-
old children exhibit sensitivity to diversity with general arguments, but performed at chance
levels with specific arguments.

Together these findings suggest that generating the superordinate category which
includes both the premise and conclusion category before being able to assess coverage (e.g.
having to generate the inclusive superordinate category mammal when reasoning from the
premise categories dog and sheep to the conclusion category buffalo) involves a
computationally demanding processing step that relies on knowledge about stable category
hierarchies. In contrast, this superordinate category is already available when reasoning
about general conclusions (e.g. reasoning from the premise categories dog and sheep to the
conclusion category mammal). If sensitivity to diversity was based exclusively on associative
knowledge, one would not expect differences based on cognitive ability and changing
developmental trajectories, nor should there be a dissociation between specific and general
arguments.
Accounts which draw on associative knowledge also have no straightforward means
of explaining effects that arise from considering underlying higher-order interrelationships
between categories. For example, Tenenbaum and Kemp (2009) and Shafto et al. (2008) have
demonstrated that inductive reasoning about causal disease transmission can be dissociated
from inductive inferences about physiological properties. Such dissociations suggest that the
context or property people are reasoning about prompts them to draw on different and most
relevant sources of structured knowledge. This dissociation is explained with reference to
theory-based Bayesian models, in which a domain-general Bayesian inference engine
operates over a domain-specific theoretical model. These models can represent people’s
structured knowledge about hierarchical taxonomic relations or causal links between species.
Evidence that people make extensive use of structured knowledge that might violate
similarity principles comes from cross-cultural research (e.g. Lopez, Atran, Coley, Medin &
Smith, 1997) and research on experts (e.g. Medin, Lynch, Coley & Atran, 1997; Proffitt,
Coley and Medin, 2000; Shafto & Coley, 2003). For example, Proffit and colleagues (2000)

looked at patterns of inductive inferences about tree diseases across three different types of
tree experts. Depending upon their speciality, they exhibited a variety of different reasoning
strategies, including the context-sensitive use of ecological and causal knowledge.
Knowledge structures that capture causal knowledge are especially interesting, as they
have several unique characteristics not shared by other knowledge structures. Asymmetry is
one of their most distinguishing features. That is, causes must always precede, or at least
coincide, with their effects. Causal structures can also vary in their complexity, ranging from
simple cause-effect relations such as unidirectional causal chains, to more elaborate common
cause models (Pearl, 2000). People do seem to make use of these abstract relations to
evaluate inductive arguments, as their inferences are highly sensitive to the direction of the
causal link between category features (Rehder, 2009), as well as the direction of the causal
link between species when making generalizations about novel properties (Medin, Coley,
Storms, & Hayes, 2003) and disease transmission (Shafto et al., 2008). For example, people
are more likely to endorse the argument that ‘lions have retinum’ given that ‘gazelles have
retinum’ than the diagnostic inference that ‘gazelles have retinum’ given that ‘lions have
retinum’ (Medin, et al., 2003), thus demonstrating a robust causal asymmetry effect which
can be formally derived using Bayes rule (Shafto et al., 2008). These causal asymmetry
effects have recently been generalised to children as young as five years of age (Bright &
Feeney, 2014). Again, it is hard to see how approaches relying exclusively on associative
knowledge might cogently explain such effects.
Reconciling Theories: The Importance of Processing Conditions
On the surface it appears that approaches placing divergent emphasis on different
types of knowledge are incompatible. However, there are strong arguments for the roles of
associative and structured knowledge in inductive reasoning, and there is a suggestion of

individual differences in the extent to which structured knowledge plays a role (Feeney,
2007). Particularly given its importance during development, it is likely that in adults,
associative knowledge influences category-based inductive inference with relative ease and
computational simplicity. In contrast there appear to be computational challenges involved in
drawing on structured knowledge. Thus, one way to resolve the apparent dichotomy between
theories which emphasize different kinds of knowledge is to argue that the inductive engine
of thought is a hybrid, taking different types of input depending on environmental conditions.
As we have seen, there are good reasons for ascribing different processing
characteristics to structured and associative knowledge, meaning that each is more likely to
power the inductive engine under different environmental conditions. This in turn allows us
to delineate and predict circumstances in which each type of knowledge is more likely to be
used during the inference process. For example, Rehder (2009) explicitly suggests that the
use of structured knowledge relies on an elaborate, analytical thought process, whereas
associative knowledge influences inductive reasoning fairly automatically and without much
cognitive effort. Rehder (2009) taught participants about the causal links between features of
artificial categories. In line with assumption that people draw on extensive casual knowledge,
he demonstrated various phenomena, such as a casual asymmetry effect. However, he also
found that there was a substantial minority of people whose patterns of inferences did not
adhere to those predicted by his causal-based generalization model. Instead, they seemed to
rely more on non-directional associations between category features. This suggests that
induction can either be driven by structured knowledge based on sometimes complex
relations between categories, or on associative knowledge based on temporal and/or spatial
contiguity between the categories. Whereas selecting the former type of knowledge during
the inference process might require effortful processing and availability of cognitive

resources, the latter type of knowledge may influence inductive reasoning relatively
automatically and with little processing cost.
Although ours will be the first investigation of the availability of associative and
structured knowledge to reasoning processes, there have been suggestions that some
knowledge structures may be less available to reasoning processes than others. Shafto,
Baldwin and Coley (2007) examined the processing conditions under which taxonomic and
causal knowledge impact on inductive reasoning. In general, people’s reasoning is context-
sensitive, that is, they will draw on relevant taxonomic knowledge when reasoning about
anatomical properties, but rely more heavily on causal knowledge when reasoning about
disease transmission (see also Heit & Rubinstein, 1994; Kemp & Tenenbaum, 2009; Shafto et
al. 2008). Shafto et al. (2007) showed that taxonomic knowledge predicts reasoning about
genes regardless of processing constraints whereas causal knowledge predicts reasoning
about diseases only when processing is unconstrained. Although our primary focus will be on
the contrast between structured and associative knowledge, by using Shafto et al’s paradigm
in our first experiment, we will also be able to carry out a conceptual replication of their
study of causal and taxonomic structured knowledge. Because they did not control for
associative knowledge about relations between categories, it is unclear whether their results
will hold once degree of association has been controlled for.
Our Experimental Strategy
We had four aims in the experiments to be described here. First we wanted to show
that measures of associative and structured knowledge about the relations between categories
are dissociable. Second we wished to show that both types of knowledge influence judgments
of inductive strength. Such a demonstration would be entirely novel in the context of the
literature on induction where researchers, because of their adherence to one or other model,

have assumed that only one representational form of knowledge drives inductive reasoning.
Third, we wished to examine the processing conditions under which each type of knowledge
exerts most influence on the inductive reasoning process. Showing that the type of knowledge
which is most important differs by processing condition would help to resolve conflict
between apparently contrasting models. Thus, although our experiments focus on category-
based inductive inference, our results are relevant to a number of more general questions such
as whether it is possible to dissociate the effects on reasoning of associative and structured
knowledge, whether it is possible to delineate when different representational forms of
knowledge play the biggest role in reasoning, and whether the effects of causal knowledge on
reasoning are relatively effortless and automatic or cognitively demanding. Finally, our
fourth aim was to examine whether, once degree of association between the categories in the
arguments has been controlled for, as claimed by Shafto et al. (2007), taxonomic knowledge
is more impervious to manipulations of processing resources than is causal knowledge.
Our overall experimental strategy rests on a contrast between the effects of associative
and structured knowledge on inductive reasoning. We use the term associative as it has been
used by other researchers in the literature on inductive reasoning (see Sloman, 1996; Sloutsky
& Fisher, 2008), to refer to computations that reflect temporal contiguity or similarity.
However, in the literature on priming, distinctions are made between associative and featural
overlap accounts of priming effects (see Hutchison, 2003). In that literature, the term
associative is used to refer to accounts where concepts are represented by nodes in a network
and nodes are associated with each other (see Collins & Loftus, 1975). Distributed accounts
(e.g. Masson, 1995), on the other hand, like Sloman’s (1993) and Rogers & McClelland’s
(2004) accounts of inductive reasoning, assume that concepts are represented by patterns in
the weights of connections between the features that are activated when the concept is
encountered. The simplest explanation for priming under such accounts is in terms of the

overlap between the features that are associated with the priming concept and the concept that
is primed. This explanation is very similar to the mechanism said to underlie inference in
associative models of inductive reasoning. Thus, Sloman’s (1993) and Rogers &
McClelland’s (2004) models might better be described as feature overlap rather than
associative models. However, individual categories are represented associatively in these
models, even if inferences are computed via a process of feature overlap.
As will become apparent below, we measured the associative relation between the
categories in the arguments that participants evaluated simply by asking other participants to
rate the strength of the association between the categories. It is likely that such questions are
more transparent to participants than questions referring to shared features. Feature-overlap is
often interpreted in the literature on inductive reasoning, but not the literature on priming (see
Hutchison, 2003), as a measure of similarity, so it might be argued that we should have
explicitly asked about similarity. However, as feature overlap can occur due to similarity, or
temporal contiguity, we decided to measure degree of association instead. In addition, there
are arguments (see Medin, Goldstone & Gentner, 1993) that the nature of a similarity
calculation may be time dependent; perceptual similarity, for example, may be more
automatically calculated than structural similarity. As degree of association is, by definition,
the automatic output of an associative process, it is a better concept for our present purposes.
Pretest: Dissociating types of knowledge. One problem with testing the idea that
dissociable types of knowledge can be selected in category-based inductive reasoning is how
to a) measure and b) dissociate different types of knowledge. With regards to structured
knowledge, one can simply ask participants whether or not they believe that the categories
are taxonomically, ecologically or causally related. On the face of it, indexing associative
knowledge is also simple: we can simply ask participants to rate the strength of the
association which they believe exists between pairs of categories. One obvious objection to

this method is that participants may be drawing on the same underlying knowledge to
perform both tasks. However, if they are drawing on different sources of knowledge then we
would expect the measures not to be strongly associated with each other. Moreover, because
associative knowledge can be based on co-occurrence we might expect ratings of associative
strength to correlate strongly with objective measures of co-occurrence whilst our measures
of structured knowledge should correlate much less strongly with an objective measure of co-
occurrence. Note that for co-occurring category pairs which are also visually similar, an
objective measure of co-occurrence will likely underestimate the degree of association
between the categories. This should serve to make any evidence we find for dissociation
between associative and structured knowledge more impressive.
Strength of association. To measure strength of association between category pairs,
we told 20 participants that we were interested in their beliefs about how strongly associated
two categories (e.g. “rabbits and carrots”; “salmon and herring”; “robin and grass”, etc.)
were on a scale from 1 (unrelated) to 9 (very strongly associated). They were asked to think
about all kinds of possible associations and the instructions emphasized that we wanted them
to give their first intuitive response. In total they rated the strength of association between
270 category pairs.
Conditional co-occurrence. We wanted to compare our subjective ratings to a more
objective association criterion. Thus, for all the category pairs for which we had obtained
participant association ratings, we examined their co-occurrence frequencies in natural
language. The rationale is that words which frequently co-occur in natural language should
have a stronger associative relationship (Spence & Owens, 1990). We conducted a proximity
search, specifying that the categories must co-occur in any order within a window of 6
consecutive words. We chose the search engine Exalead, which explicitly supports proximity
searches for specified distances (word 1 NEAR/ X word 2). Once we derived the co-

occurrence measure, we recorded the Exalead word frequency of each individual category.
Using Heylighen’s (2001) formula below, we calculated the conditional co-occurrence.
In this equation, represents the probability that a text contains both words
w1 and w2, represents the probability that it contains w1 on its own. To calculate the
conditional probability, one can simply count the number of times w1 and w2 co-occur and
divide this by the number of times w1 occurs by chance in the same text sample. As we were
not interested in potential asymmetries, we repeated the calculation for w2 and took the mean
conditional co-occurrence for each category pair. This was correlated with our subjective
association measures. The correlation between the subjective measure and the objective
measure was .66.
The size of this correlation suggests that our measure of associative strength is
sensitive to at least one objective measure of the strength of an association: the extent to
which the associated entities co-occur in the world. Of course, this correlation on its own
does not demonstrate that associative strength and structured knowledge are dissociated.
Thus, for each of the experiments reported in this paper we calculated, for the items used in
that experiment, the strength of the correlation between beliefs about structured relations
(collected for each experiment in a post-test, see Appendices 1 & 2), the objective measure of
co-occurrence and the measure of associative strength. These results are presented in Table 1
and will be discussed alongside the results for each experiment. To preview, the results show
that whereas the measure of association correlates strongly with the measure of co-
occurrence, our measures of structured knowledge do not correlate strongly with the measure

of associative strength nor with the measure of co-occurrence. Thus, our measures of
associative and structured knowledge are dissociated.
Experiment 1
In Experiment 1 we ran a conceptual replication of the study by Shafto et al (2007).
Shafto et al (2007) asked people to evaluate the strength of inductive inferences about cell or
disease properties in members of categories known to be taxonomically or causally related.
People were either asked to respond immediately or to delay their response. After they had
completed the inference part of the experiment, participants’ beliefs about the taxonomic and
causal relations between the categories in each argument were post-tested. In a series of
regression analyses by item, results of the post-tests were used to predict ratings of inductive
strength. Whereas beliefs about taxonomic relatedness predict argument strength regardless
of processing conditions, beliefs about causal relatedness only predicted strength judgements
for disease arguments and only in the delayed condition. Shafto et al (2007) concluded that
taxonomic knowledge was more available to reasoning processes than knowledge about
causal relations between categories. We departed from Shafto et al’s (2007) method in a
number of crucial ways. First, we chose taxonomically and causally related category pairs so
that the degree of association between the categories in the argument was controlled for.
Second, we included ratings of associative strength as a predictor in the regression analyses.
These changes allow us to address all of our aims. First, by examining correlations between
our measures of associative and structured knowledge and our objective co-occurrence
measure, we can examine whether associative and structured knowledge are dissociable.
Second, the results of the regression analyses will help us to establish whether associative and
structured knowledge predict reasoning. Third, by incorporating a response time
manipulation, we will be able to examine the degree to which each type of knowledge affects
reasoning under different processing conditions. Finally, by controlling for degree of

association across arguments, and including association as a predictor in our regression
analyses, we will be able to examine whether Shafto et al’s (2007) finding that taxonomic
knowledge is more available to reasoning processes than causal knowledge holds up when
associative strength is controlled for.
Method
Design
The experiment had a 2 (relation: causal or taxonomic) by 2 (property: cells or
diseases) by 2 (timing: fast or slow) by 4 (list) mixed design, with list and timing as between-
subjects variables.
Participants
The 40 participants were from Durham University, and received course credit for their
participation. The mean age was 24.2 years (SD = 5.8 years).
Materials and Procedure
There were 20 items in total. Items consisted of a base category, a causally related
target category and a taxonomically related target category. Causally related targets were
always from different superordinate categories, for example, plants and animals, or mammals
and reptiles. In contrast, taxonomically related pairs were always from the same
superordinate taxonomic category. For example, the causal target for the base category fly
would be frog and the taxonomic target would be ant. We ensured that the degree of
association between the base and the taxonomic (M = 4.86, SD = 1.13) and causal targets (M
= 4.73, SD = 1.01) was approximately equal; t(19) = .81, p = .43 (for association ratings for
both of the arguments in each item, see Appendix 1). We also confirmed that people did
perceive a causal link between the casually related category pairs. In a pre-test, 10 Durham
University students had previously rated the strength of the possible causal link between two

categories on a scale from 1 (unrelated) to 9 (very strong causal relation). Causal strength
ratings for the selected pairs ranged from 3.4 to 6.9 with a median of 4.5 (SD = 1).
For each of the 20 items, we created a taxonomic and a causal induction problem,
resulting in a total of 40 inductive arguments. Participants were told that the base category
had either a blank disease, such as disease 3dfT, or blank cells, such as cells T78. The
participants reasoned about different items for cells and diseases, so each person received 40
problems. Content was counterbalanced across participants in an incomplete Latin-square
design resulting in four different lists.
Participants’ task was to evaluate the likelihood that the cells/disease would be
present in the conclusion category on a scale from 1 (very unlikely) to 9 (very likely).
The premise and conclusions were presented simultaneously and appeared in a red
font. Participants could only enter their response once the font changed from red to green. In
the fast condition, the font changed from red to green after one second. Participants were
instructed to try and respond as quickly as possible. In the slow condition, the font only
changed color after 10 seconds and people could not enter their response before this time had
elapsed. In this condition, participants were instructed to take their time and think carefully
about each item before typing a response.
Post-test. The post-test assessed people’s beliefs about taxonomic and causal
relatedness. For each of the 40 category pairs (20 causal, 20 taxonomic) about which people
had made inferences in the main task, they were asked two questions, resulting in a total of
80 questions. One question asked them whether they believed that the two categories were
from the same biological class. The second question was whether the two categories were
part of the same food chain. Order of presentation was completely randomized. Participants
could respond with YES, NO or DON’T KNOW, but were instructed to use the third option

sparingly, as the emphasis was on their intuitions and beliefs rather than on factual
correctness.
Results
Pre- and Post-Testing
Our post-test confirmed that participants were largely aware of the purported relations
between the category pairs. For the causally related items, the causal endorsement proportion
was .71 (SD = .24), whereas the taxonomic endorsement proportion was .23 (SD = .11). For
the taxonomically related items, the taxonomic endorsement proportion was .75 (SD = .19),
whereas the causal endorsement proportion was .21 (SD = .25). To ensure that our pre-test
measure of associative strength and our post-test measures of structured knowledge taken at
the end of this experiment are dissociated, we calculated the magnitude of the correlation
between the pre-test and post-test measures across all 40 unique category pairs. As may be
seen in Table 1, the correlations between taxonomic and associative knowledge and between
causal and associative knowledge were small and non-significant. To obtain further evidence
for a dissociation we also calculated the correlation between our pre- and post-test measures
of knowledge and the objective measure of conditional co-occurrence. Once again, the
correlation was large and statistically significant in the case of the associative measure and
small and non-significant for the structured measures. The measure of associative knowledge
dissociates from the measures of taxonomic and causal knowledge obtained from the
participants in this experiment.
Analysing Inductive Strength Ratings
Mean inductive strength scores were calculated for the 10 problems representing the
unique property by relation combination, resulting in 4 means for each participant. These
were subjected to a 2 (relation: causal or taxonomic) by 2 (property: disease or cell) by 2

(timing: fast or slow) by 4 (list) mixed-design ANOVA, with list and timing as between-
subject variables. For the analysis by items, inductive strength ratings were averaged across
subjects rather than items and were analysed with a 2 (property) by 2 (timing) by 2 (relation)
mixed-design ANOVA, with relation as between-items variable. In what follows we will
report only effects that were significant in the items and subjects analyses. Cell means can be
found in Table 2. There were no main or interaction effects involving list (all p’s >. 07), so
this counterbalancing variable will not be mentioned any further.
The only significant effect was a two-way interaction between property and relation,
FS (1, 32) = 23.17, p < .0005, η2
p = 0.42, FI (1, 38) = 36.17, p < .0005, η2
p =0.49, suggesting
that people showed some context-sensitive reasoning. Bonferroni post hoc tests showed that
when reasoning about cells, people rated taxonomic inferences (M = 5.01, SE =.2)
significantly stronger than causal inferences (M = 3.79, SE = .22, p < .0005, effect size d =
.9). When reasoning about diseases, people rated causal inferences slightly higher (M = 4.32,
SE = .26) than taxonomic inferences (M = 3.89, SE = .17) although this difference was not
significant (p = .16, effect size d = .3). None of the other higher-order interactions were
significant (all p’s > .08). This result replicates selectivity effects that have been reported in
the literature (see Heit & Rubenstein, 1994).
Finally, the effect of timing was significant by items FI (1, 38) = 18.24, p < .0005, η2
p
= 0.32, and was approaching significance for subjects, FS (1, 32) = 2.82, p = .1, η2
p= .08.
Thus, people gave higher inductive strength ratings when they responded slowly (M = 4.48,
SE = 0.18) than when they responded quickly (M = 4.03, SE = 0.18). Crucially however,
timing did not interact with any of the other variables. This is in sharp contrast to Shafto et
al’s (2007) finding that people who were asked to respond quickly gave lower inductive
strength ratings when evaluating ecological/causal arguments, whereas there was no effect of
timing on taxonomic arguments. Thus, once strength of association had been equated

between knowledge domains, there was no evidence from this paradigm to support Shafto et
al.’s (2007) contention that taxonomic knowledge is more available to reasoning processes
than causal knowledge.
Using Associative and Structured Knowledge to Predict Reasoning
We ran hierarchical regression analyses to see how associative knowledge, as well as
how relevant domains of structured knowledge (causal and taxonomic beliefs) influence
category-based inferences under different timing conditions. For this purpose, mean inductive
strength ratings and taxonomic and causal endorsement proportions were averaged across
participants to predict inductive strength ratings for each item when people responded quickly
or slowly and for the two different properties. Strength of association derived from the pre-
test was entered in the first block of the regression analysis, followed by taxonomic and
causal endorsement proportions in the second block.
Inferences about cells. Shown in Table 3, inferences about cells in the fast condition
(R = .64) were predicted by strength of association (beta = .51, t = 3.76, p = .001) and were
negatively predicted by causal knowledge (beta = -.34, t = -2.5, p = .048). Taxonomic
knowledge was not a significant predictor of inductive strength in the fast condition (beta =
.11, t = .65, p = .52). However, adding the structured knowledge coefficients did explain
some additional variance above strength of association on its own (R2
Change: 16.7%, F (2,
36) = 5.09, p = .011).
In the slow condition (R = .72), strength of association was not a significant predictor
of inductive strength (beta = .19, t = 1.52, p = .14). Inductive inferences were however
predicted by taxonomic knowledge (beta = .48, t = 3.48, p = .001), and were negatively
predicted by causal knowledge (beta = -.31, t = -2.29, p = .028). Given that we had selected
causal targets that were always from different superordinate categories (i.e. taxonomically

unrelated) it is not surprising that believing in the existence of a causal link was a negative
predictor of inferences about cells. Adding the structured knowledge predictors in a second
block of the regression analysis accounted for significantly more variance in inductive
strength ratings than strength of association on its own when people responded slowly ( R2
Change: 44.2%, F(2, 36) = 16.33, p < .0005).
Comparing the regression weights (Cohen, 1983) across the two timing conditions
showed that the beta weight for associative strength was significantly larger in the in the fast
than in the slow condition (Z = -2.48, p =.007, one-tailed), whereas the beta weight for
taxonomic knowledge was larger in the slow condition, as predicted (Z = 2.56, p =.005, one-
tailed). For causal knowledge there was no significant difference between the beta weights
across the two timings conditions (Z = 0.17, p =.43, one-tailed).
Inferences about diseases. As may be seen in Table 3, reasoning about diseases in
the fast condition (R= .59) was significantly predicted by strength of association (beta = .45, t
= 3.13, p = .003). When entered in the second block, causal knowledge was also a significant
predictor (beta = .35, t = 2.04, p = .049), whereas taxonomic knowledge was not a significant
predictor (beta = .08, t = .44, p = .67). Together, adding these two structured knowledge
variables accounted for a non-significant amount of additional variance (R2
Change: 9.6%, F
(2, 36) = 2.64, p = .085).
In contrast, reasoning about diseases in the slow condition (R= .68) was no longer as
strongly predicted by strength of association (beta = .24, t = 1.8, p = .08). However, inductive
strength was strongly predicted by causal knowledge (beta = .61, t = 4.34, p < .0005), and
also by taxonomic knowledge (beta = .34, t = 2.33, p = .026). Adding the structured
knowledge predictors in a second block accounted for significantly more variance in

inductive strength ratings than strength of association on its own (R2 Change: 28.1%, F(2, 36)
= 9.64, p < .0005).
A comparison of regression weights across fast and slow conditions showed that the
associative strength beta weight was slightly but not significantly larger in the fast than in the
slow condition (Z = 1.25, p =.1, one-tailed). As predicted though, the beta weight for causal
knowledge (Z = 2.1, p =.018, one-tailed) was significantly larger in the slow condition.
Furthermore, the beta weight for taxonomic knowledge was also significantly larger in the
slow condition (Z = 1.9, p =.029, one-tailed). This might be explained in terms of people’s
belief that taxonomic relations also offer a good explanation for shared diseases (e.g. due to
genetic susceptibility), an effect we have observed previously in adults but not in younger
children (Bright & Feeney, 2014).
Timing Manipulation Check
To ensure that our timing manipulation had been effective, we compared mean
reaction times across the two timing conditions from stimulus onset. As we did not provide
an actual response deadline, one risk is that people might not have responded as soon as
possible, but continued to think about the problem, making their response reaction times
indistinguishable from the slow group. However, our analysis showed that people in the fast
condition responded after an average of 4.4 seconds (SD = 1.26), which was significantly
faster than response time in the slow condition (M = 12.0 seconds, SD = 1.89, t (38) = 14.97,
p < .0005). Thus, our timing manipulation was effective in eliciting diverging response times.
Discussion
We had four aims in running Experiment 1. First, we wished to show that measures of
associative and structured knowledge dissociate. The results clearly show that the experiment
has achieved that aim. Measures of associative and structured knowledge were not

significantly correlated. In addition, the subjective measure of association was strongly
correlated with an objective measure of category co-occurrence whereas the correlations
between measures of structured knowledge and the objective measure of co-occurrence were
much smaller and non-significant. All of these findings suggest that our measures are tapping
into different types of knowledge.
The second aim of the experiment was to show that both associative and structured
knowledge act as input to inductive reasoning. It is clear from our results that associative
knowledge and knowledge about structured relations are both related to judgements of
inductive strength.
With regards to our third aim, of delineating the conditions under which each type of
knowledge exerts its biggest influence, the results of this experiment offer evidence that
associative and structured knowledge have different effects dependent on the processing
conditions. Thus, associative knowledge predicted inference strength when people were
encouraged to respond quickly but not when they responded slowly. On the other hand, the
strongest effects of our measures of structured knowledge were observed when people
responded slowly.
Our fourth aim was to examine whether Shafto et al’s (2007) finding that taxonomic
knowledge is more available to reasoning processes than causal knowledge could be
replicated once strength of association had been controlled for. In fact, the results of this
suggest that the reverse can sometimes be the case. When people reasoned about diseases in
the fast timing condition in this experiment, causal knowledge was a significant predictor of
inferential strength. On the other hand, taxonomic knowledge was not a significant predictor
in any analysis of strength ratings provided by participants when they responded quickly.

Although our results suggest that once associative strength is controlled for,
taxonomic knowledge is no more available to reasoning processes than causal knowledge, we
did use different causal relations than did Shafto et al. and it is possible that this difference
may be important in explaining the differences between our results. The causal relations in
our experiment were based on predation whereas Shafto et al used shared habitat relations
and it is possible that the former are more available to reasoning processes than the latter.
Experiment 2
Experiment 2 was designed to experimentally test the claim that structured knowledge
has a greater effect on reasoning processes under favourable processing conditions. We
presented participants with inductive arguments containing one of three different types of
structured relation. One was taxonomic, a second was causal predictive, both as before. The
third was a causal diagnostic relation. For example, participants might learn that eagles have
a disease and be asked to consider how likely it is that mice also have the disease. We
predicted a causal asymmetry effect, that is, causal inferences should be stronger for
categories with a predictive causal link than for those with a diagnostic causal link (Medin et
al., 2003). Previous work (e.g. Shafto et al., 2008) shows that asymmetry effects are most
likely when the property in the argument may be transmitted from members of one category
in the argument to members of the other, so we expected to observe asymmetry effects with
disease properties rather than with cell properties.
To manipulate processing conditions, we used a secondary task paradigm in which
people had to remember simple or complex dot patterns (De Neys, 2006) whilst making their
inductive strength evaluations. Remembering the simple patterns should only minimally
affect working memory resources, whereas remembering the complex patterns places a much
heavier burden on people’s mental resources. If the retrieval of a causal knowledge structure

and assignment of causal roles is indeed mediated by an effortful process, one might expect
this causal asymmetry effect to be weakened when people are under heavy cognitive load.
Method
Design
The experiment had a 3 (relation: causal predictive, causal diagnostic, taxonomic) by
2 (load: heavy or light) by 2 (property: cells or infection) by 4 (list) mixed design, with list
and load as between-subjects manipulations.
Participants
There were 40 participants who were volunteers from Queen’s University Belfast and
Durham University. Participants received either course credit or were paid £5 for their time.
The mean age was 23.3 years (SD = 6.2 years).
Materials and Procedure
In total there were 16 items. For each item, we created four induction problems: A
causal predictive argument and its taxonomic counterpart, as well as a causal diagnostic
argument and its taxonomic counterpart.
In predictive causal induction problems people were told that the base category
(prey/plant) had either novel cells or a novel infection. The taxonomic counterpart was
created by keeping the same prey/plant base category but replacing the conclusion category
with a taxonomically related category of equivalent associative strength. In contrast to
Experiment 1, we changed the property from diseases to infections in an attempt to
discourage participants from using of taxonomic knowledge when reasoning about properties
that could be causally transmitted and thus demonstrate clearer selectivity effects.
As a variation of causal structure, for each of these items we then reversed the order
of the categories for the predictive causal problems (base category: predator/consumer →

conclusion category: prey/plant), resulting in a diagnostic causal induction problem. This was
matched again with a taxonomic induction problem by keeping the predator/consumer base
category but substituting the conclusion category for a taxonomically related alternative.
Thus, there were a total of 64 problems. For example, participants might be presented with
one of the following induction problems:
Mice have infection 9TT7. (causal predictive/ infection)
How likely is it that Eagles have infection 9TT7?
Mice have infection 7rR4. (taxonomic / infection)
How likely is it that Squirrels have infection 7rR4?
Eagles have 45T-cells. (causal diagnostic/ cells)
How likely is it that Mice have 45T-cells?
Eagles have e2T-cells. (taxonomic / cells)
How likely is it that Parrots have e2T-cells?
Based on the pre-test ratings of strength of association between category pairs, we
ensured that the strength of association was identical for the causal category pairs (M = 4.98,
SD = 1.21) and their taxonomic counterparts (M = 4.94, SD = 1.14; t (31) = .25, p =.81).
Association ratings for each item are presented in Appendix 2.
For each problem, participants reasoned either about a predictive or diagnostic causal
category pair and its taxonomic match. This meant that each participant rated 32 inductive
arguments. The items for which participants reasoned about cells or infections in a predictive
or diagnostic direction was counterbalanced in an incomplete Latin square design, resulting

in four lists. We ensured that there were an equal number of strongly and weakly associated
items in both the cell and infection condition.
Before rating each inductive argument, participants were shown a 4 by 4 grid with 4
dots for 2 seconds. In the heavy load condition, the dots were displayed in a random order
with the constraint that they could never form a straight line or diagonal. In contrast, in the
light load condition, the dots always formed a straight line or diagonal, placing minimal
burden on working memory. Once participants had read the induction question, they entered
their response on the keyboard by pressing a number between 1 and 9. They then saw an
empty matrix and had to try and recall the location of the four dots by pressing on an
appropriate box with the mouse cursor. Participants received no feedback on their recall
performance
Post-test. The post-test was identical to the one used in Experiment 1. Participants
answered two questions about the 32 category pairs, assessing their beliefs about biological
group membership, as well as their beliefs about causal relatedness.
Results
Analysis of Inductive Strength Ratings
We calculated 6 mean inductive strength ratings for each participant and analysed
these with a 3 (relation: predictive, diagnostic or taxonomic) by 2 (property: cells or
infection) by 2 (cognitive load: heavy or light) by 4 (list) mixed-design ANOVA, with load
and list as between-subjects variables. For the analysis by items we used a 2 (property) by 2
(load) by 3 (relation) mixed-design ANOVA, with relation as a between-items variable.
There was no significant main effect of list, F (3,32) = 0.92, p = .44, η2
p= 0.08, and
none of its interactions with other variables were significant (all p’s > .33), so no further
reference is made to this counterbalancing variable. There was a significant main effect of

relation, FS (1.66, 53.03)1
= 6.09, p = .007, η2
p= 0.16, FI (2, 61) = 7.22, p = .002, η2
p= 0.19.
Post-hoc comparisons showed that there was a difference in inductive strength ratings for
causal predictive (M = 3.90, SE = .24) and causal diagnostic inferences (M = 3.33, SE = .23, p
= .038, effect size d = .42), but no significant difference between causal predictive and
taxonomic inductive strength ratings (M = 4.3, SE = .25, p = .42, effect size d = .19). The
difference between diagnostic and taxonomic inferences was significant (p = .005, effect size
d = .54).
There was a significant interaction between property and relation, FS1 (1.97, 62.87) =
5.48, p = .007, η2
p= 0.15, FI (2, 61) = 11.08, p < .0005, η2
p= 0.27. When people made
inferences about cells, taxonomic inferences (M = 4.50, SE = .27) were significantly stronger
than both causal predictive inferences (M = 3.50, SE = .24, p =.017, effect size d = .47) and
causal diagnostic inferences (M= 3.26, SE = .27, p = .001, effect size d = .60), whereas there
was no difference between the latter two types of relation (p = .94, effect size d = .16).When
reasoning about infections, people rated causal predictive inferences (M= 4.30, SE = .33)
significantly higher than causal diagnostic inferences (M= 3.39, SE = .30, p = .025, effect size
d = .45). However, there was no difference between causal predictive and taxonomic
inductive strength ratings (M= 4.11, SE = .25, p = .99, effect size d = .07), or between
taxonomic inferences and causal diagnostic inferences (p = .5, effect size d = .32).
There was a significant three-way interaction between property, load and relation, FS
(1.97, 62.87) 1 = 3.22, p = .047, η
2p= 0.09; FI (2, 61) = 3.95, p = .024, η
2p= 0.12. The cell
means across all conditions are shown in Table 4. To further examine this interaction and to
explore our a-priori prediction that the causal asymmetry effect would be less evident under
cognitive load, we carried out two separate 2 (causal relation: predictive versus diagnostic)
by 2 (load: heavy versus light) mixed-model ANOVAs, one for cells and the other for
infections.

Inductive strength ratings when reasoning about infections. For infections, there
was a significant main effect of causal relation, FS (1, 38) = 8.99, p = .005, η2
p= 0.19, FI (1,
30) = 7.59, p = .01, η2
p = 0.2. However, the main effect of causal relation was modulated by
the significant interaction between load and causal relation, FS (1, 38) = 4.04, p = .051, η2
p=
0.1, FI (1, 30) = 7.77, p = .009, η2
p = 0.21, presented in Figure 1. Bonferroni post-hoc tests
showed that people who were not burdened by a heavy secondary memory load were able to
take causal structure into account. Indicative of a significant causal asymmetry effect,
inferences with a causal predictive relation (M = 5.01, SE = .44) were rated much stronger
than inferences with a diagnostic causal link (M = 3.49, SE = .41, p = .001, effect size d =
.73). In contrast, the causal asymmetry effect was absent when people had to contend with a
heavy secondary memory load. Thus, inductive strength ratings were almost identical for
causal diagnostic inferences (M = 3.29, SE = .41) and causal predictive inferences (M = 3.49,
SE = .41, p = .49, effect size d = .17).
This finding suggests that the causal asymmetry effect arises because people consider
how the underlying causal structure might influence the distribution of properties such as
infections or diseases. When people are cognitively burdened, they may not be able to take
the underlying causal structure into account, instead forcing them to rely on the existence of
an association between the categories.
Inductive strength ratings when reasoning about cells. As expected, for inferences
about cells, there was no effect of causal relation, FS (1, 38) = 1.05, p = .31, η2
p= 0.03, FI (1,
30) = 0.97, p = .33, η2
p= 0.03. People rated inferences about category pairs with a predictive
causal relation (M = 3.50, SE = .24) the same as when the relation between the two categories
was diagnostic (M = 3.26, SE = .27).

The main effect of load was non-significant, FS (1, 38) = .007, p = .93, η2
p< .001, FI
(1, 30) = 0.2, p = .88, η2
p= 0.001, as was the interaction between load and causal relation, FS
(1, 38) = 1.29, p = .26, η2
p = 0.03, FI (1, 30) = 0.99, p = .33, η2
p = .180.03.
Pre-and Post-Testing
Across items, causally related categories attracted a causal endorsement proportion of
.74 (SD = .21) and a taxonomic endorsement proportion of .11 (SD = .18). In contrast,
taxonomically related categories received a taxonomic endorsement proportion of .73 (SD =
.18) and a causal endorsement proportion of .23 (SD = .10).
To ensure that our pretest measure of associative strength and our post-test measures
of taxonomic and causal knowledge taken at the end of this experiment are dissociated, we
calculated the magnitude of the correlation across all 64 unique category pairs between the
pre-test and post-test measures. As may be seen in Table 1, the correlations between
structured knowledge and associative knowledge were small and non-significant. The
correlations between subjective and objective measures of associative knowledge were large
and significant whereas the correlations between structured knowledge and the objective
measure of associative knowledge were small and non-significant. As before, our pre- and
post-test measures of different types of knowledge were dissociated.
Secondary Task Analysis
In a dual task paradigm, dissociable effects of the secondary task can reflect strategic
trade-offs between primary and secondary tasks (Hegarty, Shah, & Miyake, 2000). To guard
against the possibility that the effect of memory load on causal inferences reflected such a
strategic trade-off, we calculated the number of correctly recalled dots separately for the trials
preceding each of the unique property by load by relation conditions. A 2 (relation: causal
predictive, causal diagnostic or taxonomic) by 2 (property: cell of infection) by 2 (load:

heavy versus light) by 4 (list) mixed-design ANOVA, with list and load as between-subjects
variables showed that the only significant difference in the number of dots recalled was
between the two load conditions, F (1, 31) = 17.71, p < .0005, η2
p = 0.36. In the heavy load
condition, participants recalled a mean of 3.2 dots (SE = .1), whereas they recalled on average
3.8 dots (SE = .1) when they were only under a light memory burden. This suggests that
people were consistent in how they allocated their mental resources to the primary and
secondary tasks across all problems and verified that the more complex patterns were harder
to remember than the simple dot patterns.
Discussion
The results from Experiment 2 strongly suggest that the influence on inductive
reasoning of different types of knowledge is dependent upon processing conditions. Thus, the
causal asymmetry effect was not observed when participants were under heavy cognitive
load. This finding supports our contention that drawing on structured knowledge is mediated
by an effortful, time-consuming process, unlike the use of associative knowledge, which
influences the reasoning process relatively automatically.
As might be expected given other findings in the literature, the results of Experiment
2 also contain evidence of inductive selectivity (see Heit & Rubenstein, 1994). Thus, people
were sensitive to causal direction only when reasoning about infections, and found arguments
containing taxonomically related categories more convincing only when reasoning about
cells.
Experiment 3
Thus far we have shown that both associative and structured knowledge drive
reasoning processes but to different extents under different processing conditions. The
experimental task used in both of the experiments described so far required people to evaluate

inductive arguments. One disadvantage of this method is that people may lack the relevant
structured knowledge (see Coley, Shafto, Stepanova, & Barraff, 2005), forcing them to rely
on associative strategies that they wouldn’t normally use. In addition, such a task may not be
very similar to how we make everyday inductive inferences. In real life we are likely to learn
new facts about categories and then generate other categories to which those new facts may
generalise. In the final experiment to be described here we asked our participants to carry out
an inductive generation task of this sort. One disadvantage of such a task is that it gives us
less control over our materials than we have exerted in Experiments 1 and 2. Thus, because
we cannot control the categories which people generate, it may not be possible to carry out all
of the analyses we performed on the results of the earlier experiments. Nonetheless, to make
the strongest argument possible about the involvement of associative and structured
knowledge in inductive reasoning, it is important to generalise our findings to a range of
different paradigms.
In Experiment 3, people were told that a base category had a novel infection or cells
and were asked to infer which other category was most likely to also have the infection or
cells. Based on the previous two experiments, we predicted that when people were under
heavy cognitive load, they would generate conclusion categories which were more strongly
associated with the base category. In contrast, we expected more complex structured
knowledge to guide inferences when people were under light load. Because separate groups
of participants rated the degree of association and the existence of structured relations
between the categories in the arguments generated by a first group of reasoners, the
experiment had three phases, induction generation, associative rating, and structured relation
rating.
Method - Induction Generation Phase

Design
In this experiment, we manipulated whether people generated an open-ended
inference under heavy or light load..
Participants
Twenty-three students (M age = 24.2 years) from Durham University received course
credit for their participation. 11 were randomly assigned to the heavy load condition, whereas
12 completed the inductive task under minimal cognitive load.
Materials and Procedure
Participants (the reasoners) were presented with 20 base categories and told that each
category had a novel property, either an infection (e.g. has infection 5y5u) or cells (e.g. has 3-
yu-cells). We did not expect the two properties to differ in how structured versus associative
knowledge might affect the inference drawn. However, we included this variation to try and
encourage people to think about why a conclusion category might share the property with the
base rather than simply recalling a strong association. Participants were then asked to
generate ONE other category that they believed was most likely to also have the property. For
example, people would read the following generative induction problem:
Weasels have 4Ou-cells / infection 4Ou.
Which other category is most likely to also have 4Ou-cells/ infection 4OU?
People wrote down their response and also gave a rating of inductive strength on a scale from
1 (very unlikely to share the property) to 9 (highly likely to share the property).
Akin to Experiment 2, participants completed a secondary task (4 by 4 dot matrix),
either under heavy (complex 4-dot configuration) or light cognitive load (simple 4-dot

configuration) conditions. Participants had to memorize the location of the dots, then
complete the inference task before attempting to recall the location of the dots in an empty 4
by 4 matrix.
Method - Association Rating Phase
Participants
Participants consisted of a group of 92 students (the raters) from Durham University
(M age = 22.4 years) who had not taken part in the first phase. They completed the task at the
beginning of a lecture.
Materials and Procedure
Each individual participant’s 20 category pairs from phase 1 were transcribed onto an
association rating sheet and interspersed with 15 weakly associated distracter items.
Participants in phase 2 received one of 23 different sheets (approximately 4 participants per
sheet) and were asked to rate the strength of association on a scale from 1 (unrelated) to 9
(very highly associated) between the 35 category pairs. They were instructed to respond as
fast as possible, based on the first intuitive answer that came to mind.
Method - Structured Relation Ratings Phase
To identify different types of structured relations, the experimenter and a second blind
coder rated whether there was a taxonomic and/or causal or interaction-based relationship
between the 20 category pairs participants had generated in phase 1. Table 5 below shows an
example of the different types of relation.
Category pairs were assigned 0 if there was no apparent link between the base and the
generated category (e.g. alligator → soil), 1 if they were taxonomically related (e.g. zebra →
horse), 1 if they were related through a causal link or ecological interaction (e.g. hawk →

mouse) and 2 if there was both a taxonomic and interaction-based relation between the
categories (e.g. cod → shark). Concordance rate across the two primary coders was 67%.
Disagreements were resolved through discussion with two further colleagues.
Results
Relations between Association Ratings, Index of Structured Relations and Co-
occurrence
To create an associative strength measure we averaged the mean strength of
association scores attached to each reasoner’s 20 category pairs across the four raters from
phase 2. For each participant’s 20 category pairs, we also calculated the web-based
conditional co-occurrence using the formula described in Experiment 1. To measure the
degree to which each the categories in each argument were structurally related, we used the
structured relation measure described above.
For each of the 23 participants we calculated the correlation between the associative
strength and structured knowledge measures across their 20 category pairs, the correlation
between the measure of structured knowledge and the measure of co-occurrence, and the
correlation between the measure of associative knowledge and the measure of co-occurrence.
We then compared these three correlation coefficients across the 23 reasoners using a
repeated-measures one-way ANOVA, which was significant, F (2, 44) = 20.81, p < .0001, η2
p
= 0.49. As expected, Bonferroni post-hoc tests showed that the mean correlation coefficient
between association ratings and co-occurrence (M Kendall’s τ =.41, SD = .12) was
significantly larger than the mean correlation coefficient between association ratings and the
index of structured knowledge (M Kendall’s τ = .14, SD = .20; p = .045, effect size d = 1.25)
and the mean correlation coefficient between web-based co-occurrence and structured
knowledge (M Kendall’s τ = .23, SD = .18; p = .048, effect size d = 0.77). These results

suggest, as did the results of Experiments 1 and 2, that our measures of structured and
associative knowledge are dissociated.
Comparing Strength of Association and Structured Knowledge across Processing
Conditions
We predicted that people under heavy load would generate conclusion categories that
were more strongly associated (M = 6.42, SD = 1.09) than those generated under light load
(M = 5.96, SD = 1.08). This prediction was confirmed with a between-subjects t-test across
participants who had assigned a strength of association rating to each category pair, t (89) =
2.01, p = .048, effect size d = 0.42.
In contrast, we expected structured knowledge to be important under favourable
reasoning conditions. For each reasoner, we counted the number of structural relations across
all items. A score of 0 would mean that there were no discernable structural relation between
base and conclusion category and 40 would mean that there were multiple relations between
every generated category pair. A between-subjects t-test on the sum of structured relations
showed that there were more structured relations under light load (M = 24.25, SD = 1.29)
than under heavy load conditions (M = 20.63, SD = 2.5), t (21) = 4.41, p < .0005, effect size d
= 1.8.
Relations between Inductive Strength Ratings, Structured Relations and Associative
Strength
In line with our second and third aims - to show that both structured and associative
knowledge influence inductive reasoning and to delineate the importance of each type of
knowledge under contrasting processing conditions - we used the associative strength

measure and the index of structured relations described above to predict inductive strength
ratings.
For each participant who had taken part in phase 1, we carried out a regression
analysis, using associative strength and structured relation measures to predict his/her
inductive strength ratings. The beta weights were analysed with a 2 (load: heavy or light) by
2 (type of beta weight: associative versus structured relation) mixed-design ANOVA, with
type of beta weight as the repeated-measures variable.
There was no significant main effect of type of beta weight, F(1, 21) = .068, p = .80,
η2
p = 0.003 and no main effect of load, F(1, 21) = 3.22, p = .09, η2
p = 0.13. However, there
was a significant cross-over interaction between beta weight type and load, F(1, 21) = 6.53, p
= .018, η2
p = 0.24. This is illustrated below in Figure 2. Bonferroni posthoc tests showed that
when participants were under heavy memory load, the associative strength beta weight (M
beta= .20, SE = .07) was larger than the structured relation beta weight (M beta = .02, SE =
.02), although this difference was not quite statistically significant due to the small number of
participants in this condition (p = .065, effect size d = 1.2). A one-sample t-test confirmed
that associative strength beta weight was significantly above zero, t(10) = 3.79, p = .004, but
that the structured relation beta weight was not significantly above zero, t(10) = 0.45, p = .66.
The pattern was reversed when participants were not under a heavy cognitive load.
Thus, the structured relation beta weight (M beta = .28, SE = .05) was slightly but not
significantly larger in magnitude than associative strength beta weight (M beta = .11, SE =
.07, p =.11, effect size d = .62). However, the one-sample t-test showed that whereas the
structured relation beta weight was significantly above zero, t(11) = 5.24, p <.0005, the
associative strength beta weight was not statistically different from zero, t(11) = 1.56, p = .15.

Across the two load conditions, the associative strength beta weight was slightly but
not significantly larger for reasoners who generated their inferences under heavy load
compared to those who were only under light load (p = .44, effect size d = .32). In contrast,
the mean structured relation beta weights were significantly larger when participants were
only under minimal cognitive load compared to those who had generated their inferences
under heavy cognitive load (p = .001, effect size d = 1.6).
Discussion
The results of Experiment 3 support and extend the conclusions suggested by our
earlier experiments. Thus, the results suggest that measures of associative and structured
knowledge dissociate, and that both types of knowledge drive reasoning, but to different
extents depending on processing conditions. Because we have used a generation paradigm in
Experiment 3, our conclusions can be generalised beyond situations where participants
simply make a judgement about an argument, to situations where inductive targets are
retrieved from memory prior to the operation of a judgement process. It is unclear exactly
how associative and structured knowledge are used to retrieve targets from memory, and this
would be an interesting topic for further study.
As mentioned at the outset, one of the disadvantages of the generation paradigm is
that we lose control of the arguments that participants evaluate. Accordingly, we have been
unable to present an analysis comparing taxonomic and causal inferences. Although
participants tended to generate more taxonomically related targets under low load,
preliminary analysis revealed that categories which were taxonomically related were rated as
more strongly associated than categories that only shared more causal interaction-based
relations. The nature of the paradigm meant that this confound could not have been avoided,
and as the confound makes it impossible to draw inferences about availability differences

between taxonomic and causal knowledge from the final experiment, we have not presented
the analysis here. However, this should not detract from our central conclusion that structured
knowledge drives open-ended induction under favourable processing conditions, whereas
associative knowledge is more influential under unfavourable conditions. This underscores
our main conclusions from Experiment 1 and 2, but using a completely different paradigm
that makes different demands on the reasoner.
It is also important to compare the results of the regression analyses from this
experiment to the regression results from Experiments 1. Of course, because the analyses of
Experiment 1 were calculated by item whereas those in Experiment 3 were averaged across
participants, and we had no control over the relations between premise and conclusion
category in Experiment 3, a direct comparison is not possible. Nonetheless, there are some
interesting conclusions to be drawn. The effects of associative knowledge are particularly
interesting. One response to our arguments that associative and structured knowledge are
dissociated could be that structured knowledge is simply more complex than associative
knowledge but that these two types of knowledge are not qualitatively different. However, in
this case we would expect associative knowledge to always act as input to inductive
reasoning. Our results suggest otherwise; when cognitive resources are available, associative
knowledge is not a particularly good predictor of inductive reasoning. Associative knowledge
was not a significant predictor of reasoning about cells or diseases in the delayed conditions
of Experiment 1, and the average beta weight for associative reasoning in the light load
condition of Experiment 3 did not differ significantly from zero. This suggests that when
resources are available, participants are selective in the type of knowledge that they draw
upon for reasoning.

General Discussion
The results described in this paper make several important contributions to what we
know about cognition in general, and inductive reasoning in particular. We have devised a
simple method to measure the strength of the association between two categories. By
showing that results using this method are strongly related to an objective measure of
association, we have established its validity, and we have shown that it dissociates from
measures of people’s knowledge of various structured relationships that might hold between a
pair of categories. Using our measures of knowledge, we have shown that in many
circumstances, associative and structured knowledge influence inductive reasoning but that
this depends upon processing environment. It is also clear from our results that structured
knowledge has more influence on inductive inferences when processing conditions are
favourable, whereas associative knowledge has more effect on reasoning under adverse
processing conditions. Of course, these results have implications for theories of inductive
reasoning, but they are also relevant to a range of questions about theories of cognition more
generally.
Implications for Theories of Inductive Reasoning
Different models of induction make different assumptions about whether knowledge
is represented in associative or structured form, and our results show that both associative and
structured knowledge influence reasoning under different processing conditions. We can see
no way for structured (e.g. Osherson et al., 1990; Kemp & Tenenbaum, 2009) or associative
(Rogers & McClelland, 2004) theories of reasoning to account for all of our findings.
Although the structured Bayesian approach copes very impressively with the effects of
different types of structured knowledge which seem to be evoked by manipulations of the
property reasoned about, it assumes that all knowledge is structured. Our results contradict

this assumption. A measure of the extent to which the categories in the argument are
associated, which itself is dissociated from measures of knowledge about more structured
relations between the categories, predicts inductive strength judgements. Contrary to the
predictions of connectionist accounts which hold that all knowledge is associative, those
measures of structured knowledge also predict reasoning performance, particularly under
favourable processing conditions. Our results suggest that to fully capture people’s
judgements of argument strength, hybrid models will be required.
The effects of timing and secondary task manipulations observed in our experiments
have particular implications for structured Bayesian accounts of reasoning. Rational analyses
of cognition (see Anderson, 1990) of the kind exemplified by Causal Bayes nets accounts of
reasoning (e.g. Kemp & Tenenbaum, 2009; Shafto et al., 2008) tend not to consider the
processes underlying participants’ judgements of inferential strength (although see Griffiths,
Vul, & Sanborn, 2012). For example, Shafto et al. (2008) show how a structured Bayesian
account of induction predicts causal asymmetry effects similar to the ones observed here, but
do not speculate on the processes underlying such effects. Our results place processing
constraints on Shafto et al.’s model as we show that one of its key predictions is confirmed
only when people’s processing resources are relatively uncompromised. This finding raises
interesting questions about how reasoning consistent with a Bayesian analysis might be
instantiated in the mind to which we will return below.
Implications for Theories of Cognition
Structured versus associative accounts. As well as carrying implications for
theories of inductive reasoning, our results also bear on recent debates about theories of
cognition more generally. For example, because our experiments were designed to contrast
the effects of structured and associative knowledge on inductive reasoning, their results have

implications for conclusions about the relations between specific structured and connectionist
accounts of thinking (see Griffiths, Chater, Kemp, Perfors, & Tenenbaum, 2010; McClelland
et al., 2010). There has been recent discussion about whether structured Bayesian (see Kemp
& Tenenbaum, 2009) and connectionist (see Rogers & McClelland, 2004) accounts of
semantic cognition are complementary (see Griffiths et al., 2012). One argument is that they
are because they operate at different levels of analysis (see Kemp & Tenenbaum, 2008).
Another view is that these models exemplify different frameworks for explaining complex
cognition (see Rogers & McClelland, 2008). Regardless of the level at which they are cast or
whether they are radically different due to the nature of their ontological commitments, these
theories seem likely to account for different patterns in our data. Thus, a connectionist
account should easily be able to predict effects of the strength of the association between the
categories in the argument on strength judgements whereas it will have more difficulty in
predicting effects of knowledge about shared category or food chain membership. A
structured Bayesian model, on the other hand cannot explain the effects of associative
strength that we have observed. Thus, our results suggest that the accounts are
complementary because neither on its own is likely to be able to account for all of the data.
On the other hand, the argument that the accounts are complementary because each explains
the data, but at different levels (see Kemp & Tenenbaum 2008), is incompatible with our
claim that neither account will be able to explain all of the data.
Dual process theories. Our results also have implications for claims that there are
two processes for thinking (Chaiken & Trope, 1999; Evans, 2008). Such claims have been
widely made in the literatures on social and cognitive psychology (for recent reviews see
Evans & Stanovich, 2013; Evans, 2008), and have also been made about inductive reasoning
specifically (see Feeney, 2007; Feeney & Heit, 2007; Evan Heit & Hayes, 2011; Rotello &
Heit, 2009; Sloman, 1998). Although there is some disagreement about the characteristics

that have been ascribed to these processes, Type 1 processes are generally held to be fast,
automatic and independent of general cognitive resources such as working memory. They are
commonly equated with the operation of associative processes (see Morewedge &
Kahneman, 2010; Sloman, 1996). Type 2 processes are slow and sequential, and they process
structured representations in a rule-based manner (Sloman, 1996). On the face of it, our
results are relatively straightforward to interpret in a dual process framework: Whereas
associative knowledge influences inductive inferences in a fairly automatic and effortless
manner, akin to the operation of Type 1 processes described in dual process theories, for
structured knowledge to influence reasoning often requires the availability of time and mental
resources, both hallmarks of Type 2 processes.
Dual process theories have their critics (see Keren, 2013; Kruglanski, 2013;
Kruglanski & Gigerenzer, 2011; Osman, 2013) and even proponents of the approach
acknowledge that finding effects of a secondary task or response time manipulation can be
inconclusive (see Lieberman, 2008). Our results seem to be particularly powerful evidence
for a dual process approach. In addition to obtaining more standard effects of secondary task
and timing manipulation on reasoning performance, we have shown that the extent to which
two dissociated measures of knowledge predict reasoning performance is dependent on
processing conditions. There are good a priori reasons for expecting that associative or Type
1 processes will take associative knowledge as input and that Type 2 processes will take
structured knowledge as input. We have shown not only that reasoning performance is
influenced by the imposition of a secondary load or time limit, but that the influence of
knowledge which we expect to inform Type 2 processing is decreased under unfavourable
processing conditions. This finding, that the degree to which structured knowledge informs
reasoning is dependent on processing conditions, is highly novel evidence for the dual
process position.

Relations between Bayesian and dual-process accounts. It is interesting in this
context to consider the implications of our data for ideas about the relation between Bayesian
and dual-process accounts of thinking. As we have already pointed out, our data constrain
psychological theorising about structured Bayesian accounts. They turn out to do so in an
unexpected way. Although the primary goal of a rational analysis is not a process-level
specification of performance, there has been some speculation about the relation between
Bayesian inference and dual process theories of thinking. For example, Oaksford and Chater
(2001) speculated that the probabilistic approach offers a computational-level account of
Type 1 processes for thinking, which they claim to be much more widely used than are Type
2 processes. Consistent with this claim is Chater, Tenenbaum and Yuille’s (2006) tentative
suggestion that the cognitive system draws on a set of heuristic strategies to approximate a
Bayesian solution, rather than engaging in explicit probabilistic computations. Certainly, if
one considers the complexity of the solutions that must be approximated, then it seems
unlikely that slow, sequential, working memory intensive processes are responsible.
Recent work on how people make inductive inferences about objects when their
categorisation is uncertain appears to support these views. Chen, Ross and Murphy (2013)
asked participants to make an inductive inference about an object that could belong to one of
two categories. Previous findings show that, contrary to the Bayesian account of this task (see
Anderson, 1990), people consider only the most likely category when making their prediction
(e.g. Murphy & Ross, 1992). In a verbally presented task, participants displayed a tendency
to ignore the alternative category. However, in an isomorphic speeded, action-based task,
consistent with a Bayesian account, people took both possible categorisations into
consideration. To account for their data, Chen et al. appeal to the distinction between implicit
and explicit cognition which is often made when contrasting Type 1 and Type 2 processes
(see Evans, 2007). They argue that participants are precluded from using working memory by

the speeded action-based task, and so must fall back on implicit (or Type 1) processing which
approximates a Bayesian solution.
At first inspection, our findings - that the causal asymmetry effect is not observed
under conditions of load, and that causal knowledge best predicts reasoning under favourable
processing conditions – seem to run counter to the suggestions described above. That is, they
suggest that performance in line with Bayesian accounts is more likely under conditions
favourable to Type 2 processing. However, one way in which the two sets of results may be
reconciled depends on recognising the constraints placed by the reasoning context on
decisions about which knowledge is most relevant for a particular inference. Once sufficient
time is available reasoners may attempt to suppress or delay a response that is driven by
associative knowledge in order for structured knowledge to play a role. Thus, it may not be
the application of structured knowledge that requires Type 2 processing, but the inhibition of
associative knowledge. Such a possibility is consistent with findings suggesting that Type 2
processes are often required to inhibit the output of Type 1 processes (see De Neys & Van
Gelder, 2009; Handley, Capon, Beveridge, Dennis, & Evans, 2004; Lieberman, Hariri,
Jarcho, Eisenberger, & Bookheimer, 2005).
It is possible that inhibitory control may play a role in our experimental results also.
This suggestion is consistent with classic ideas about the interplay between automatic and
controlled processes in semantic priming (see Neely, 1977) and reading (Gernsbacher &
Faust, 1991). In order for the relevant structured knowledge to dominate responding,
participants may need to inhibit or suppress the effects of quickly processed associative
knowledge. That such control processes are known to take time and effort is consistent with
our secondary task and timing findings. Assuming a role for control processes also allows for
the claim that once the relevant structured knowledge has been selected, the psychological

instantiation of Kemp and Tenenbaum’s structured Bayesian model draws on relatively
effortless processes.
Causal Asymmetry Effects
As we pointed out in the introduction, our experiments also speak more generally to
accounts of asymmetry effects in causal thinking. The original demonstration that people
treated prediction inferences from cause to effect differently to diagnostic inferences from
effect to cause, was provided by Tversky and Kahneman (1980). They showed that people
thought it more likely that a daughter would have blue eyes given that her mother did than
that a mother would have blue eyes given that her daughter did. Tversky and Kahneman
(1980) argued that it is easier or more natural to go from cause to effect than from effect to
cause. Recently however, Fernbach et al. (2011) questioned this account and provided
evidence that Tversky and Kahneman’s example does not produce an asymmetry effect under
certain experimental conditions. They also suggest that, in the majority of cases, a cause
provides more evidence for an effect than an effect does for a cause, and suggest that the
reasoning of participants in earlier experiments (e.g. Medin et al., 2003) may have been
consistent with the predictions of a Bayesian analysis. Our results are relevant to this debate
because we have shown that causal asymmetry effects are not observed when participants are
placed under heavy cognitive load.
In this context, there are two possible interpretations of our results. First, it may be
that, as Tversky and Kahneman argued, it is easier to reason from cause to effect than from
effect to cause. A second interpretation is that people are better Bayesians in less challenging
environments. Under this interpretation, if a Bayesian analysis holds that predictive
arguments are often stronger than diagnostic arguments, then we might expect to observe that
people are better able to distinguish one from the other in favourable processing conditions. .

Our data seems more consistent with this set of predictions, although it is probably too early
to come to a definite conclusion and because there are arguments that people are better
Bayesians in more challenging environments (see Chen, Ross & Murphy, 2013), it is not
clear that this second interpretation coheres with other findings and explanations in the
literature.
Future Research
One important direction for future research will be in determining the importance of
individual differences to inductive reasoning. Dual process accounts equate individual
differences in performance with variability in the operation of Type 2 processes and Feeney
(2007) has already shown associations between cognitive ability and inductive reasoning. The
discussion here suggests that other sources of differences between individuals might predict
inductive reasoning performance. For example, individual differences in inhibitory control
might predict performance on certain inductive reasoning problems. Of course there are
different types of inhibitory control so it will be interesting to see whether inductive
reasoning is best predicted by the ability to inhibit prepotent responses based on associative
knowledge (see Handley et al., 2004) or by individual differences in the ability to inhibit
associative knowledge itself (see Burgess & Shallice, 1996). Should the ability to inhibit
semantic knowledge turn out to predict inductive reasoning performance, then this will have
theoretical consequences for debates about the merits of Bayesian and associative accounts as
well as for assumptions about the type of processes that approximate structured Bayesian
models of reasoning.
We also hope that our method, of using dissociated measures of different types of
knowledge to predict performance, will be taken up by researchers in other areas of
psychology. We can certainly envision possible uses for the method in the study of semantic

priming or recognition memory. Our method is also likely to be useful in the study of how
reasoning develops. For example, an important question for future research concerns the
relative importance of associative and causal knowledge (see Hayes & Thompson, 2007) in
children’s inductive reasoning. It would also be extremely interesting to examine more
generally the extent to which an objective measure of association and children’s own
structured knowledge predicts their pattern of inductive projections. Such a study would be
particularly helpful in resolving recent debates about the role of associative and structured
knowledge in children’s reasoning (Hayes, McKinnon, & Sweller, 2008; Sloutsky & Fisher,
2004; Wilburn & Feeney, 2008).
Conclusions
There is currently much debate over the relative merits of, and relations between,
Bayesian and associative approaches to cognition (see Jones & Love, 2011; McClelland et
al., 2010). The results we have described here suggest that accounts derived from these
approaches will not suffice to explain inductive reasoning. Each approach makes different
assumptions about how knowledge is represented and our experiments show that associative
and structured knowledge differentially affect reasoning depending on processing conditions.
In addition, whether causal asymmetry effects, which can only be explained by accounts that
allow for structured knowledge, are observed is also dependent on processing conditions.
These results suggest that either a hybrid theory or some entirely new account that takes into
account different representational formats in semantic memory will be required in order to
capture the richness of human inductive inference.

References
Anderson, J. R. (1990). The adaptive character of thought: Erlbaum.
Andrews, G., Halford, G. S., & Boyce, J. (2012). Conditional discrimination in young children: The
roles of associative and relational processing. Journal of Experimental Child Psychology,
112(1), 84-101.
Bergman, T. J., Beehner, J. C., Cheney, D. L., & Seyfarth, R. M. (2003). Hierarchical classification
by rank and kinship in baboons. Science, 302(5648), 1234-1236.
doi:10.1126/science.1087513
Bright, A.K. & Feeney, A. (2014). Children’s causal knowledge and the development of inductive
reasoning. Journal of Experimental Child Psychology, 122, 4861.
Burgess, P. W., & Shallice, T. (1996). Response suppression, initiation and strategy use following
frontal lobe lesions. Neuropsychologia, 34(4), 263-272.
Chaiken, S., & Trope, Y. (1999). Dual-process theories in social psychology (reader): Guilford Press.
Chater, N., Tenenbaum, J. B., & Yuille, A. (2006). Probabilistic models of cognition: where next?
Trends in Cognitive Sciences, 10(7), 292-293. doi: 10.1016/j.tics.2006.05.008
Chen, S. Y., Ross, B. H., & Murphy, G. L. (2013). Implicit and explicit processes in category-based
induction: is induction best when we don't think? Journal of Experimental Psychology:
General, No Pagination Specified. doi: 10.1037/a0032064
Cohen, A. (1983). Comparing regression-coefficients across subsamples - A study of the statistical
test. Sociological Methods & Research, 12(1), 77-94.
Coley, J. D. (2012). Where the wild things are: informal experience and ecological reasoning. Child
Development, 83(3), 992-1006. doi: 10.1111/j.1467-8624.2012.01751.x
Coley, J. D., Shafto, P., Stepanova, O., & Barraff, E. (2005). Knowledge and category-based
induction. In W. K. Ahn, Goldstone, R. L., Love, B. C., Markman, A. B. and Wolf, P. (Ed.),
Categorization inside and outside the laboratory: Essays in honor of Douglas L. Medin.
Washington, D.C.: American Psychological Association.

Collins, A. M., & Loftus, E. F. (1975). A spreading activation theory of semantic processing.
Psychological Review, 82, 407-428.
Colunga, E., & Smith, L. B. (2005). From the lexicon to expectations about kinds: A role for
associative learning. Psychological Review, 112(2), 347-382. doi: 10.1037/0033-
295x.112.2.347
De Neys, W. (2006). Automatic-heuristic and executive-analytic processing during reasoning:
Chronometric and dual-task considerations. Quarterly Journal of Experimental Psychology,
59(6), 1070-1100. doi: 10.1080/02724980543000123
De Neys, W., & Van Gelder, E. (2009). Logic and belief across the lifespan: the rise and fall of belief
inhibition during syllogistic reasoning. Developmental Science, 12(1), 123-130. doi:
10.1111/j.1467-7687.2008.00746.x
Evans, J. S. B., & Stanovich, K. E. (2013). Dual-process theories of higher cognition advancing the
debate. Perspectives on Psychological Science, 8(3), 223-241.
Evans, J. S. B. T. (2007). On the resolution of conflict in dual process theories of reasoning. Thinking
& Reasoning, 13(4), 321-339. doi: 10.1080/13546780601008825
Evans, J. S. B. T. (2008). Dual-processing accounts of reasoning, judgment, and social cognition.
Annual Review of Psychology, 59, 255-278. doi: 10.1146/annurev.psych.59.103006.093629
Feeney, A. (2007). How many processes underline category-based induction? Effect of conclusion
specificity and cognitive ability. Memory & Cognition, 35, 1830-1839.
Feeney, A., & Heit, E. (2007). Inductive reasoning. Cambridge: Cambridge University Press.
Fenker, D. B., Waldmann, M. R., & Holyoak, K. J. (2005). Accessing causal relations in semantic
memory. Memory & Cognition, 33(6), 1036-1046.
Fernbach, P. M., Darlow, A., & Sloman, S. A. (2011). Asymmetries in predictive and diagnostic
reasoning. Journal of Experimental Psychology-General, 140(2), 168-185. doi:
10.1037/a0022100
Fisher, A. V., & Sloutsky, V. M. (2005). When induction meets memory: Evidence for gradual
transition from similarity-based to category-based induction. Child Development, 76(3), 583-
597.

French, R. M., Mareschal, D., Mermillod, M., & Quinn, P. C. (2004). The role of bottom-up
processing in perceptual categorization by 3-to 4-month-old infants: Simulations and data.
Journal of Experimental Psychology-General, 133(3), 382-397. doi: 10.1037/0096-
3445.133.3.382
Gernsbacher, M. A., & Faust, M. E. (1991). The mechanism of suppression - a component of general
comprehension skill. Journal of Experimental Psychology-Learning Memory and Cognition,
17(2), 245-262. doi: 10.1037/0278-7393.17.2.245
Griffiths, T. L., Chater, N., Kemp, C., Perfors, A., & Tenenbaum, J. B. (2010). Probabilistic models of
cognition: exploring representations and inductive biases. Trends in Cognitive Sciences,
14(8), 357-364. doi: 10.1016/j.tics.2010.05.004
Griffiths, T. L., Vul, E., & Sanborn, A. N. (2012). Bridging levels of analysis for probabilistic models
of cognition. Current Directions in Psychological Science, 21(4), 263-268. doi:
10.1177/0963721412447619
Halford, G. S., Wilson, W. H., & Phillips, S. (2010). Relational knowledge: the foundation of higher
cognition. Trends in Cognitive Sciences, 14(11), 497-505. doi: 10.1016/j.tics.2010.08.005
Handley, S. J., Capon, A., Beveridge, M., Dennis, I., & Evans, J. S. B. T. (2004). Working memory,
inhibitory control and the development of children's reasoning. Thinking & Reasoning, 10(2),
175-195. doi: 10.1080/13546780442000051
Hayes, B. K., McKinnon, R., & Sweller, N. (2008). The development of category-based induction:
Reexamining conclusions from the induction then recognition (ITR) paradigm.
Developmental Psychology, 44(5), 1430-1441. doi: 10.1037/0012-1649.44.5.1430
Hayes, B. K., & Thompson, S. P. (2007). Causal relations and feature similarity in children's
inductive reasoning. Journal of Experimental Psychology-General, 136(3), 470-484. doi:
10.1037/0096-3445.136.3.470
Hegarty, M., Shah, P., & Miyake, A. (2000). Constraints on using the dual-task methodology to
specify the degree of central executive involvement in cognitive tasks. Memory & Cognition,
28(3), 376-385.

Heit, E., Hahn, U., & Feeney, A. (2005). Defending Diversity. In W. K. Ahn, R. L. Goldstone, B. C.
Love, A. B. Markman & P. Wolff (Eds.), Categorization inside and outside of the laboratory:
Essays in honor of Douglas L. Medin (pp. 87-99): APA.
Heit, E., & Hayes, B. K. (2011). Predicting reasoning from memory. Journal of Experimental
Psychology. General, 140(1), 76.
Heit, E., & Rubinstein, J. (1994). Similarity and property effects in inductive reasoning. Journal of
Experimental Psychology-Learning Memory and Cognition, 20(2), 411-422.
Heylighen, F. (2001). Mining associative meanings from the web: from word disambiguation to the
global brain: Standaard Publishers.
Holyoak, K. (2012). Analogy and relational reasoning. In K. Holyoak & R. G. Morrison (Eds.), The
Oxford handbook of thinking and reasoning (pp. 234-259). New York: Oxford University
Press.
Holyoak, K. J., Lee, H. S., & Lu, H. J. (2010). Analogical and category-based inference a theoretical
integration with bayesian causal models. Journal of Experimental Psychology-General,
139(4), 702-727. doi: 10.1037/a0020488
Hutchison, K. A. (2003). Is semantic priming due to association strength or feature overlap? A
microanalytic review. Psychonomic Bulletin & Review, 10(4), 785-813.
Jones, M., & Love, B. C. (2011). Bayesian fundamentalism or enlightenment? On the explanatory
status and theoretical contributions of Bayesian models of cognition. Behavioral and Brain
Sciences, 34(04), 169-188.
Jones, S. S., & Smith, L. B. (2002). How children know the relevant properties for generalizing object
names. Developmental Science, 5(2), 219-232.
Kemp, C., & Jern, A. (2009). A taxonomy of inductive problems. Paper presented at the Proceedings
of the 31st Annual Conference of the Cognitive Science Society.
Kemp, C., & Tenenbaum, J. B. (2009). Structured statistical models of inductive reasoning.
Psychological Review, 116(1), 20-58. doi: 10.1037/a0014282
Keren, G. (2013). A tale of two systems a scientific advance or a theoretical stone soup? Commentary
on Evans & Stanovich (2013). Perspectives on Psychological Science, 8(3), 257-262.

Kruglanski, A. W. (2013). Only one? The default interventionist perspective as a unimodel—
Commentary on Evans & Stanovich (2013). Perspectives on Psychological Science, 8(3),
242-247.
Kruglanski, A. W., & Gigerenzer, G. (2011). Intuitive and deliberate judgments are based on common
principles. Psychological Review, 118(1), 97-109. doi: 10.1037/a0020762
Kruschke, J. K. (1992). Alcove - an exemplar-based connectionist model of category learning.
Psychological Review, 99(1), 22-44.
Lieberman, M. D., Hariri, A., Jarcho, J. M., Eisenberger, N. I., & Bookheimer, S. Y. (2005). An fMRI
investigation of race-related amygdala activity in African-American and Caucasian-American
individuals. Nature Neuroscience, 8(6), 720-722. doi: 10.1038/nn1465
Lopez, A., Atran, S., Coley, J. D., Medin, D. L., & Smith, E. E. (1997). The tree of life: Universal and
cultural features of folkbiological taxonomies and inductions. Cognitive Psychology, 32(3),
251-295.
Lopez, A., Gelman, S. A., Gutheil, G., & Smith, E. E. (1992). The development of category-based
induction. Child Development, 63(5), 1070-1090.
Masson, M. E. J. (1995). A distributed memory model of semantic priming. Journal of Experimental
Psychology: Learning, Memory, & Cognition, 21, 3-23
Markman, A. B. (1999). Knowledge Representation. Mahwah, NJ: Lawrence Erlbaum Associates.
McClelland, J. L., Botvinick, M. M., Noelle, D. C., Plaut, D. C., Rogers, T. T., Seidenberg, M. S., &
Smith, L. B. (2010). Letting structure emerge: connectionist and dynamical systems
approaches to cognition. Trends in Cognitive Sciences, 14(8), 348-356. doi:
10.1016/j.tics.2010.06.002
McClelland, J. L., & Rumelhart, D. E. (1985). Distributed memory and the representation of general
and specific information. . Journal of Experimental Psychology: General, 114, 159-197.
Medin, D. L., Coley, J. D., Storms, G., & Hayes, B. K. (2003). A relevance theory of induction.
Psychonomic Bulletin & Review, 10(3), 517-532.
Medin, D. L., Goldstone, R. L., & Gentner, D. (1993). Respects for similarity. Psychological Review,
100(2), 254.

Medin, D. L., Lynch, E. B., Coley, J. D. &Atran, S. (1997). Categorization and reasoning among tree
experts: do all roads lead to Rome? Cognitive Psychology, 32(1), 49-96.
Morewedge, C. K., & Kahneman, D. (2010). Associative processes in intuitive judgment. Trends in
Cognitive Sciences, 14(10), 435-440. doi: 10.1016/j.tics.2010.07.004
Neely, J. H. (1977). Semantic priming and retrieval from lexical memory - Roles of inhibition-less
spreading activation and limited-capacity attention. Journal of Experimental Psychology-
General, 106(3), 226-254. doi: 10.1037//0096-3445.106.3.226
Oaksford, M., & Chater, N. (2001). The probabilistic approach to human reasoning. Trends in
Cognitive Sciences, 5(8), 349-357.
Osherson, D. N., Smith, E. E., Wilkie, O., Lopez, A., & Shafir, E. (1990). Category-based induction.
Psychological Review, 97(2), 185-200.
Osman, M. (2013). A Case Study Dual-Process Theories of Higher Cognition—Commentary on
Evans & Stanovich (2013). Perspectives on Psychological Science, 8(3), 248-252.
Pearl, J. (2000). Causality : models, reasoning, and inference. Cambridge, U.K. ; New York:
Cambridge University Press.
Penn, D. C., Holyoak, K. J., & Povinelli, D. J. (2008). Darwin's mistake: Explaining the discontinuity
between human and nonhuman minds. Behavioral and Brain Sciences, 31(2), 109-+. doi:
10.1017/s0140525x08003543
Proffitt, J. B., Coley, J. D., & Medin, D. L. (2000). Expertise and category-based induction. Journal of
Experimental Psychology-Learning Memory and Cognition, 26(4), 811-828.
Rehder, B. (2009). Causal-Based Property Generalization. Cognitive Science, 33(3), 301-344. doi:
10.1111/j.1551-6709.2009.01015.x
Rescorla, R. A., & Wagner, A. R. (1972). A theory of Pavlovian conditioning: Variations in the
effectiveness of reinforcement and nonreinforcement. . In A. H. Black & W. F. Prokasy
(Eds.), Classical conditioning II: Current research and theory (pp. 64-99). New York:
Appleton-Century-Crofts. .
Rhodes, M., Gelman, S.A., & Brickman, D. (2008). Developmental changes in the consideration of
sample diversity in inductive reasoning. Journal of Cognition and Development, 9, 112-143.

Rips, L. J. (1975). Inductive judgments about natural categories. Journal of Verbal Learning and
Verbal Behavior, 14(6), 665-681.
Rogers, T. T., & McClelland, J. L. (2004). Semantic cognition : a parallel distributed processing
approach. Cambridge, Mass. ; London: MIT.
Rogers, T. T., & McClelland, J. L. (2008). A simple model from a powerful framework that spans
levels of analysis. Behavioral and Brain Sciences, 31(6), 729-749. doi:
10.1017/s0140525x08006067
Rotello, C. M., & Heit, E. (2009). Modeling the effects of argument length and validity on inductive
and deductive reasoning. Journal of Experimental Psychology-Learning Memory and
Cognition, 35(5), 1317-1330. doi: 10.1037/a0016648
Seed, A. M., Tebbich, S., Emery, N. J., & Clayton, N. S. (2006). Investigating physical cognition in
rooks, Corvus frugilegus. Current Biology, 16(7), 697-701. doi: 10.1016/j.cub.2006.02.066
Shafto, P., & Coley, J. D. (2003). Development of categorization and reasoning in the natural world:
Novices to experts, naive similarity to ecological knowledge. Journal of Experimental
Psychology-Learning Memory and Cognition, 29(4), 641-649
Shafto, P., Coley, J. D., & Baldwin, D. (2007). Effects of time pressure on context-sensitive property
induction. Psychonomic Bulletin & Review, 14(5), 890-894.
Shafto, P., Kemp, C., Bonawitz, E. B., Coley, J. D., & Tenenbaum, J. B. (2008). Inductive reasoning
about causally transmitted properties. Cognition, 109(2), 175-192.
Sloman, S. A. (1993). Do simple associations lead to systematic reasoning. Behavioral and Brain
Sciences, 16(3), 471-472.
Sloman, S. A. (1996). The empirical case for two systems of reasoning. Psychological Bulletin,
119(1), 3-22.
Sloman, S. A. (1998). Categorical inference is not a tree: The myth of inheritance hierarchies.
Cognitive Psychology, 35(1), 1-33.
Sloutsky, V. M., & Fisher, A. V. (2004). Induction and categorization in young children: A similarity-
based model. Journal of Experimental Psychology-General, 133(2), 166-188.

Sloutsky, V. M., & Fisher, A. V. (2008). Attentional learning and flexible induction: How mundane
mechanisms give rise to smart behaviors. Child Development, 79(3), 639-651.
Sloutsky, V. M., Kloos, H., & Fisher, A. V. (2007). When looks are everything: Appearance
similarity versus kind information in early induction. Psychological Science, 18(2), 179-185.
Smith, E. R., & DeCoster, J. (2000). Dual-process models in social and cognitive psychology:
Conceptual integration and links to underlying memory systems. Personality and Social
Psychology Review, 4(2), 108-131.
Spence, D. P., & Owens, K. C. (1990). Lexical co-occurrence and association strength. Journal of
Psycholinguistic Research, 19(5), 317-330.
Sun, R. (1994). Integrating rules and connectionism for robust commonsense reasoning. New York:
Wiley.
Sun, R. (2002). Duality of the mind: A bottom-up approach toward cognition. Mahwah, NJ: Erlbaum.
Taylor, A. H., Miller, R., & Gray, R. D. (2013). Reply to Boogert et al.: The devil is unlikely to be in
association or distraction. Proceedings of the National Academy of Sciences of the United
States of America, 110(4), E274-E274. doi: 10.1073/pnas.1220564110
Tversky, A. & Kahneman, D. (1980). Causal schemata in judgements under uncertainty. In M.
Fishbein (Ed.), Progress in social psychology (pp 49-72). Hillsdale, HJ: Erlbaum.
Vasilyeva, N., & Coley, J. (2010). Generating inductive inferences: premise relations and property
effects. In B. H. Ross (Ed.), The Psychology of Learning and Motivation (Vol. 53, pp. 183-
226). Burlington: Academic Press.
Wilburn, C., & Feeney, A. (2008). Do development and learning really decrease memory? On
similarity and category-based induction in adults and children. Cognition, 106(3), 1451-1464.
doi: 10.1016/j.cognition.2007.04.018

Footnotes
1. Degrees of freedom adjusted for non-sphericity using Greenhouse-Geiser corrections.

Acknowledgements
The research described in this paper was supported by a postgraduate studentship awarded to
the first author by the Economic and Social Research Council and were carried in the
Department of Psychology at Durham University. The details of Experiment 3 were
presented at the 32nd
Annual Conference of the Cognitive Science Society. The authors wish
to acknowledge Dave Knight for his assistance in programming the experiments.

Table 1: Correlations between Mean Association Ratings, Co-Occurrence and Structured
Knowledge Measures in Experiments 1 and 2
Z-score
Exalead Co-
Occurrence
Causal
Beliefs
Taxonomic
Beliefs
Experiment 1
(40 Items)
Mean Association
Rating
Spearman’s rho .63 .14 .16
p <.0005 .41 .31
Z-score Exalead
Co-Occurrence
Spearman’s rho -.15 .06
p .36 .70
Causal Beliefs Spearman’s rho -.54
p <.0005
Experiment 2
(64 Items)
Mean Association
Rating
Spearman’s rho .59 .20 .001
p <.0005 .11 .99
Z-score Exalead
Co-Occurrence
Spearman’s rho .04 ..07
p .75 .56
Causal Beliefs Spearman’s rho -.80
p <.0005

Table 2: Mean Inductive Strength Ratings (and Standard Deviations) across the Two Timing
Conditions in Experiment 1
Cells Infections
Timing Condition Slow Fast Slow Fast
Relation
Causal 3.9 (1.4) 3.6 (1.4) 4.6 (1.6) 4.1 (1.7)
Taxonomic 5.3 (1.3) 4.7 (1.2) 4.1 (1.0) 3.7 (1.2)

Table 3: Summary Regression Results and Beta Weight Differences in Experiment 1
Expt 1 Beta
Weights
Fast
Cell
Slow
Cell
Diff Z-
score
p-
valuea
Fast
Disease
Slow
Disease
Diff Z-
score
p-
valuea
Strength of
Association .51
** .19
ns 2.48 .007 .45
** .24
ns 1.25 .10
Causal
Knowledge -.34
* -.31
* 0.17 .43 .35
** .61
** 2.10 .018
Taxonomic
Knowledge .11
ns .48
** 2.56 .005 .08
ns .34
** 1.90 .029
R2 Change 16.7 %
* 44.2 %
** 9.6 %
ns 28.1 %
**
* p < .05
** p < .01
ns p > .05
a right-tailed p-value

Table 4: Mean Inductive Strength Ratings (and Standard Deviations) for Cells and
Infections across the Load Conditions for the Three Types of Relation in Experiment 2
Property
Cells Infections
Load Light Heavy Light Heavy
Relation
Causal Predictive 3.4 (1.6) 3.6 (1.4) 5.0 (2.2) 3.6 (1.7)
Causal Diagnostic 3.4 (1.9) 3.1 (1.5) 3.5 (1.7) 3.3 (2.0)
Taxonomic 4.4 (1.5) 4.6 (1.7) 4.2 (1.5) 4.0 (1.8)

Table 5: Coding Scheme for Structured Relations in Experiment 3
Taxonomic Relationship
Category
Membership
Both categories belong to the same class or category
(e.g. carrot & parsnip)
Physiological
Similarity
Both categories are similar with respect to specific
organs or systems (e.g. bat & bird)
Causal/ Interaction-Based Relationship
Similar Habitat Both categories share similar or the same habitat
(e.g. trout & shrimp)
Behavioural
Interaction
Both categories interact via some aspect of behaviour
(e.g. monkey & tree)
Food Chain
Interaction
Both categories interact with respect to diet or eating,
i.e. one category eats or is eaten by the other (e.g.
heron & fish)

Figures
Figure 1: Causal Asymmetry Effect (Mean Inductive Strength Rating and Standard Error
Bars) across Load Conditions when reasoning about Infections in Experiment 2
Figure 2: Beta Weights (and Standard Error Bars) across the Two Load Conditions in
Experiment 3

1.0
2.0
3.0
4.0
5.0
6.0
Light Load Heavy Load
Predictive
Diagnostic


Appendix 1: Post-Test Endorsement Proportions and Strength of
Association in Experiment 1
Proportion who
believe
Categories are
Causally
Related
Proportion
who believe
Categories
are from same
Taxonomic
Group
Strength of
Association
Z-score
Conditional
Co-
Occurrence
Causal
Acorn & Squirrel 0.95 0.75 6.1 -0.1
Butterfly & Swallow 0.58 0.00 4.7 -0.54
Cabbage & Snail 1.00 0.05 4.1 -0.63
Carrot & Rabbit 0.90 0.05 6.1 0.56
Fly & Frog 0.85 0.25 5.0 -0.32
Grass & Sheep 0.90 0.00 6.9 -0.22
Grasshopper & Toad 0.67 0.45 3.8 -0.59
Krill & Penguin 0.86 0.08 3.6 -0.42
Meerkat & Eagle 0.69 0.11 3.4 -0.59
Mouse & Buzzard 0.84 0.87 3.8 -0.61
Penguin & Orca 0.70 0.08 4.8 -0.42
Rabbit & Fox 0.93 0.03 6.0 -0.12
Salmon & Grizzly 0.93 0.50 5.9 1.33
Scorpion & Eagle 0.35 0.11 4.5 -0.57
Shrimp & Salmon 0.55 0.05 5.3 4.32
Snail & Frog 0.30 0.10 4.5 -0.32
Snail & Hedgehog 0.62 0.43 4.3 -0.56
Squirrel & Cat 0.13 0.11 3.9 2.04
Toad & Snake 0.51 0.05 3.8 -0.28
Wheat & Mouse 0.90 0.25 4.4 -0.64

Mean 0.71 0.21 4.7 0.07
Taxonomic
Acorn & Walnut 0.21 0.98 6.30 -0.14
Butterfly & Ant 0.43 0.83 4.70 -0.5
Cabbage & Rapeseed 0.08 0.77 3.70 -0.54
Carrot & Radish 0.20 0.98 6.40 1.85
Fly & Ant 0.05 0.75 5.20 -0.45
Grass & Bamboo 0.30 0.95 6.70 -0.05
Grasshopper & Lice 0.27 0.63 3.70 -0.64
Krill & Woodlice 0.06 0.27 2.80 -0.35
Meerkat & Badger 0.13 0.87 4.40 -0.55
Mouse & Squirrel 0.19 0.83 4.40 -0.08
Penguin & Duck 0.48 0.80 4.30 -0.46
Rabbit & Squirrel 0.26 0.90 5.70 -0.46
Salmon & Herring 0.34 0.90 6.60 2.03
Scorpion & Spider 0.23 0.60 5.70 0.11
Shrimp & Barnacle 0.33 0.69 4.70 -0.48
Snail & Squid 0.13 0.78 3.20 0.12
Snail & Limpet 0.29 0.35 5.60 -0.5
Squirrel & Prairie Dog 0.28 0.70 4.30 -0.21
Toad & Salamander 0.20 0.64 5.00 0.46
Wheat & Bamboo 0.16 0.93 3.80 -0.48
Mean 0.23 0.75 4.9 -0.07

Appendix 2: Post-Test Endorsement Proportions and Strength of Association in
Experiment 2
Proportion who
believe
Categories are
Causally
Related
Proportion
who believe
Categories
are from same
Taxonomic
Group
Strength of
Association
Z-score
Conditional
Co-
Occurrence
Causal
Acorn & Squirrel 0.98 0.00 6.1 -.25
Butterfly & Swallow 0.57 0.08 4.7 -.55
Cabbage & Snail 0.92 0.00 4.1 -.60
Carrot & Rabbit 0.95 0.05 6.1 .19
Flower & Butterfly 0.71 0.00 6.4 1.30
Fly & Frog 0.92 0.05 5.0 -.40
Frog & Heron 0.72 0.03 4.3 -.53
Grass & Sheep 0.98 0.00 6.9 -.33
Grasshopper & Toad 0.72 0.16 3.8 -.58
Krill & Penguin 0.68 0.11 3.6 -.46
Meerkat & Eagle 0.64 0.06 3.4 -.57
Shrimp & Salmon 0.44 0.57 5.3 2.70
Snail & Hedgehog 0.67 0.03 4.3 -.56
Squirrel & Cat 0.21 0.54 3.9 1.18
Tree & Bark Beetle 0.90 0.03 7.3 3.32
Wheat & Mouse 0.85 0.05 4.4 -.61
Mean 0.74 0.11 5.0 0.20
Taxonomic
Acorn & Pecan 0.27 0.50 5.8 -.50
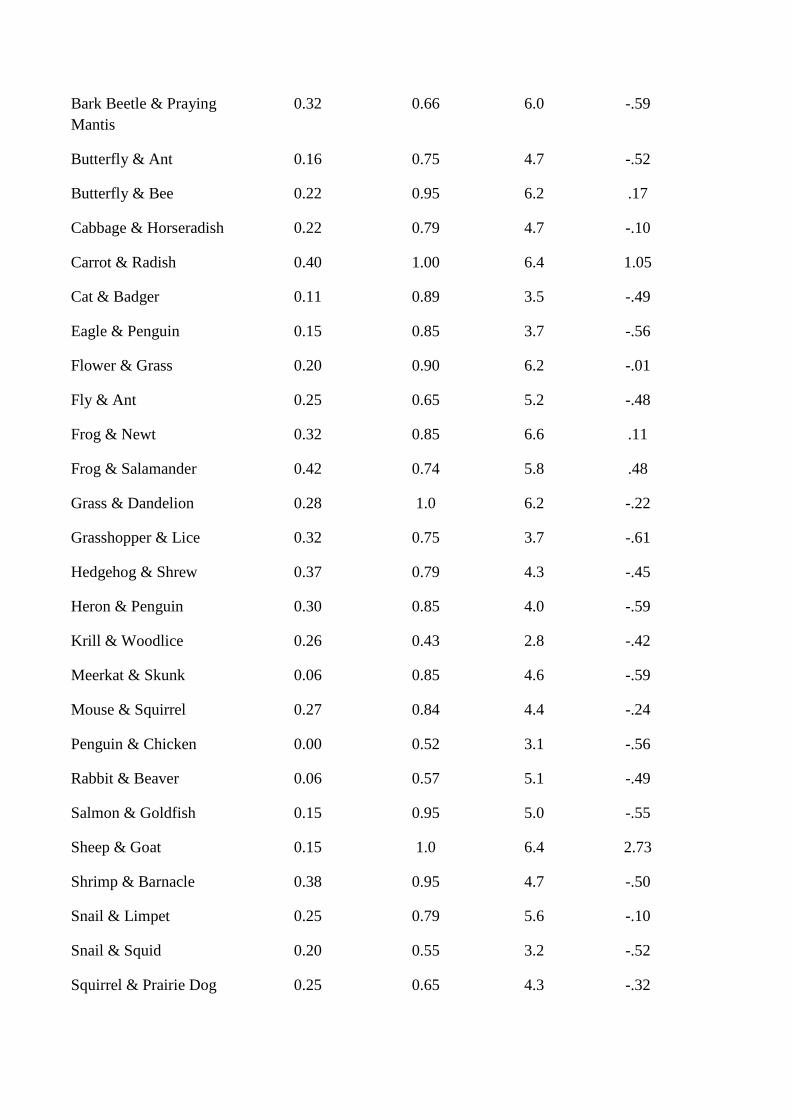
Bark Beetle & Praying
Mantis
0.32 0.66 6.0 -.59
Butterfly & Ant 0.16 0.75 4.7 -.52
Butterfly & Bee 0.22 0.95 6.2 .17
Cabbage & Horseradish 0.22 0.79 4.7 -.10
Carrot & Radish 0.40 1.00 6.4 1.05
Cat & Badger 0.11 0.89 3.5 -.49
Eagle & Penguin 0.15 0.85 3.7 -.56
Flower & Grass 0.20 0.90 6.2 -.01
Fly & Ant 0.25 0.65 5.2 -.48
Frog & Newt 0.32 0.85 6.6 .11
Frog & Salamander 0.42 0.74 5.8 .48
Grass & Dandelion 0.28 1.0 6.2 -.22
Grasshopper & Lice 0.32 0.75 3.7 -.61
Hedgehog & Shrew 0.37 0.79 4.3 -.45
Heron & Penguin 0.30 0.85 4.0 -.59
Krill & Woodlice 0.26 0.43 2.8 -.42
Meerkat & Skunk 0.06 0.85 4.6 -.59
Mouse & Squirrel 0.27 0.84 4.4 -.24
Penguin & Chicken 0.00 0.52 3.1 -.56
Rabbit & Beaver 0.06 0.57 5.1 -.49
Salmon & Goldfish 0.15 0.95 5.0 -.55
Sheep & Goat 0.15 1.0 6.4 2.73
Shrimp & Barnacle 0.38 0.95 4.7 -.50
Snail & Limpet 0.25 0.79 5.6 -.10
Snail & Squid 0.20 0.55 3.2 -.52
Squirrel & Prairie Dog 0.25 0.65 4.3 -.32

Squirrel & Prairie Dog 0.30 0.65 4.3 -.32
Swallow & Parrot 0.10 0.75 5.6 -.60
Toad & Salamander 0.25 0.75 5.0 .12
Tree & Bush 0.15 0.95 7.1 -.34
Wheat & Bamboo 0.15 0.85 3.8 -.51
Mean 0.23 0.78 4.9 -.20




















