
THE ELUCIDATION OF THE &INA M D A COMPLEX - TSpace
159
THE ELUCIDATION OF THE &INA MDA COMPLEX Raoul HarIey Bain A thesis submitted in confonnity with the requirements for the degree of Master of Science Graduate Department of Zoology University of Toronto O Copyright by Raoul Harley Bain 1998
-
Upload
khangminh22 -
Category
Documents
-
view
2 -
download
0
Transcript of THE ELUCIDATION OF THE &INA M D A COMPLEX - TSpace

THE ELUCIDATION OF THE &INA M D A COMPLEX
Raoul HarIey Bain
A thesis submitted in confonnity with the requirements for the degree of Master of Science Graduate Department of Zoology
University of Toronto
O Copyright by Raoul Harley Bain 1998

National Library 191 ofCanada Bibliothèque nationale du Canada
Acquisitions and Acquisitions et Bibliographic Services services bibliographiques 395 Wellington Street 395, rue Wellington Ottawa ON K I A ON4 Ottawa ON K1 A ON4 Canada Canada
The author has granted a non- L'auteur a accordé une licence non exclusive licence allowing the exclusive permettant à la National Library of Canada to Bibliothèque nationale du Canada de reproduce, loan, distribute or sell reproduire, prêter, distribuer ou copies of this thesis in microfonn, vendre des copies de cette thèse sous paper or electronic formats. la forme de microfiche/fïlm, de
reproduction sur papier ou sur format électronique.
The author retauis ownersship of the L'auteur conserve la propriété du copyright in this thesis. Neither the droit d'auteur qui protège cette thèse. thesis nor substantial extracts fkom it Ni la thèse ni des extraits substantiels may be printed or otherwise de celle-ci ne doivent être imprimés reproduced without the author' s ou autrement reproduits sans son permission. autorisation.

The Elucidation of the Rma livida Complex
Raoul Harley Bain, Master of Science Degree, 1998
Department of Zoology, University of Toronto
ABSTRACT
1 investigated the suspected polytypic nature of the green cascade frog, Rana livida, from
southeast Asia with a multi-step approach. My initial anatomical study of specimens from
Vietnam resulted in the recognition of three distinct species and five morphological groups. I *
tested the strength of these groupings with morphometrics and cellular DNA content analysis.
Congruence among the results of morphology, morphometrics, and cellular DNA content, led
me to suspect the presence of further cryptic species. 1 tested this hypothesis with allozyme
electrophoresis, resolving 15 presumptive loci. Fixed allelic differences suggested the
presence of four additional species. They also suggested that one of the new species may be
two distinct species, but the evidence collected here is equivocal. 1 elucidated seven species
of the Rana livida complex. 1 believe that much more cryptic diversity of this complex and
other cascade ranids await discovery. This highlights the need for rigorous taxonomic and
phylogenetic analyses of d l cascade ranids.

ACKNOWLEDGMENTS
1 would like to first thank my supervisor Bob Murphy. As rnost know, one is not part
of the Murphy lab, it is part of you. 1 thank him for al1 the mernorable experiences and
lessons taught (some knowingly, most not). I especially thank Bob for ROMMY. Though,
admittedly hesitant at first, it became more and more apparent to me, as we sweated in the
Pianetariurn, that it would be worth every drop (Oh!). I would also like to thank rny
cornmittee, Bob, Doug Currie, Hans Sues, and Dan Brooks. The cornments that Ham and
Dan lent to earlier drafts helped tremendously.
There are many whose technical help 1 gratefully acknowledge. For discussions of
things morphometric, I would like to thank Pedro Peres-Neto, Paul Galpern, and Debra
Yunnan. For teaching me allozyme electrophoresis, I thank Ross MacCulIoch, Darlene
Upton, Jinzhong Fu, and Bob. 1 thank Ross for performing the DNA Content analyses. I
thank Amy Lathrop for teaching me how to x-ray. Valuable assistance was also given in the
fonn of translations. For this I thank Chun-mei Huang, Jinzhong Fu, Wu Xiao-Chun, Johann
Lindell, Nasreen Rahman, and Amy's dictionaries. 1 thank Tanya Trepanier for allowing me
to use her laser pnnter. 1 also thank Bob and Amy for allowing me to use their photographs

in this thesis and in talks I have given. 1 would also like to thank Linda Ford of the Amencan
Museum of Natural History for her assistance.
1 would like to thank some colleagues in Vietnam. First, 1 thank Prof. Dr. Cao van
Sung, Director of the Instiîute of Ecology and Biological Resources (IEBR), Hanoi for dl of
his efforts. For invaluable field assistance, 1 would Mce to thank Dang Tuan Dat, vice
director of the Institute of Epidemiology, Ban Ma Thuot, Pham Duc Tien (IEBR), and, of
course Ms. Ho Thu Cuc also of the IEBR. I also thank Nikolai Orlov, Brad Hubley dong
with Amy, Bob, Doug, and students of the BI0 308 field course who helped collect
specimens. 1 wish to pointedly ignore acknowledging mefloquine.
I am lucky to have been part of the ROM. There are many who make this a great
place and 1 am grateful to have met thern. For their humour and wisdom 1 would like to
thank Doug Cume, Jon Barlow, Chris Darling, Burton Lim, Josh Feltharn, Alison Stuart,
Andy Bennett, John Swann, Marty Rouse, Judy Edwards-Davies, Antonia Guidotti, Colette
Baril, and Kevin Doyle. In a past life at the Museum, I was involved with Palaeobiology.
For tremendous experiences and special times with that department, 1 would tmly like to
thank, Tan Nicklin, Ted Ecclestone, Tim Fedak, Ryosuke Motani, Kevin Seymour, Catherine

Skrabec, Hans Sues, MicheIe Bobyn, and Ian Momson. I would also like to make special
mention to Chris McGowan for his support, his boyish awe, and devilish humour.
1 would like to acknowledge my friends, though I will surely Ieave some out. From
the Department of Zoology 1 thank Amaya Ortigosa, Paul GaIpern, Ian Dworkin, Pedro
Peres-Neto, Nasreen Rahman, Fernando Portella de Luna Marques, Kristy C h n a , and Sarah
Kalhok. Liz Tudor-Mulroney was exceptionally helpful in the grad office. A special thank-
you to my lab mates, past and present: Darlene Upton, Jinzhong Fu, and Amy Lathrop for
great times in Our various office-cubicIes around the ROM. Heartfelt thanks to Diana
Dobson, Ted Ecclestone, Tim Fedak, Kate Holmes, Blair Klayman, Mike Leibovici, Amy
Meckler, Jory Nash, Andrew Pifko, Max Westhead, Deana Vardy, and Ilana Zylbennan for
their support and encouragement (late night or otherwise).
1 thank my loving family. My sister Hilary and rny brothers Cale and Zachary aiways
help keep things in perspective and ensure that part of me stays twelve years old. My
parents, though not dways understanding why ("You're going where? To do what?), have
aIways supported me in the end ("lt would give us so much pleasure if you would let us buy
you a pair of hiking boots"). 1 am very thanldul. Of course, 1 know that none of this would
be possible without my grandparents.

Final thanks to Sarah Kalhok for al1 of her love and help. She has been my biggest
booster, my master typesetter, and my best critic the whole way through.
This project was supported in part by two University of Toronto Special Open
Masters Fellowships, and two Department of Zoology Pure and Applied Sciences Grants.
Field and lab work was supported by an NSERC (A3 148) and ROM Foundation Grants to
Robert W. Murphy as well as an American Museum of Natural History Collection Study
Grant to myself.
This thesis was catered by Blaze Catering.

Now to the very heart of wonder. Because species diversity was
created prior to humanity, and because we evolved within it, we have
never fathomed its limits. As a consequence, the living world is the
naturd domain of the most restless and paradoxicd part of the
human spirit. Our sense of wonder grows exponentially . . . .
E.O. Wilson
vii

TABLE OF CONTENTS
THE ELUCIDATION OF THE RANA LlVIDA CQMPLEX ........................................ i . . ABSTRACT. ..... . .... . .. .. ... .. .... ................ . . .. ... . . . ... . ........... .. . .... .. ... . .... .*. .. .....a. . .. .. . m... . . m.. . ...... . . . lI . . . ACKNOWLEDGMENTS . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . * . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . ........
TABLE OF CONTENTS. .... ..... ............................... . .............. ....................... . ........... viii
. . LIST OF TABLES . .......... ... . ... ..... ... .... .. .... .. .. ... ...... .. . .. .. . ..... . . ... . .... ... ... . . ...... ...................... XII
LIST OF FIGURES .. . . .. . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . .. . . . . . .. . . . . .. . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . xiv
LIST OF APPENDICES.. .. . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . -. . . . . . . . . . xv
CHAPTER 1:
RANA LIVTDA AND PTS IMPORTANCE TO BIODIVERSITY: ......... ..., ......... . 1 BACKGROUND ........................... .. ............................................... . .......................... . ....... . ........ 1 RANA LMDA: CONFUSION ABOUT SPECIES IDENTiTY . .................... . .................... ..... ......... 4
Rana livida ............................................ ....... .................. ,... ............ , ......................... , .,.............. 4
Rana grarninea.. . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . , . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 6
Rana 2epof.ipes ............................... . . .. . . ...................................................... . ........ , ....... 6
Rana sinica ..., , ........................................................... . ......................................................... 7 RANA LIVIDA: GENERIC AND SUBGENERIC CONFUSION ................................................... 8
Runa. .. . , .. . . , , . . . . . .. . . ., . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . .. . . .9 Hylarana . . . , . . . . . . . .. . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . .. . . . . . . , . . . . . . .. . . . . . . . . . . . . , . . . . . . . .. . , . . . . . . . . . . . . . .. . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . .9 Odorrana ........ .................................................. , ..................................................................... 11
Eburana ....................... , ......... , .....,...... . .......... ......... . ............................................................... 12
RANA LMDA: A SPECIES cOMPLEX? ............................... ................. . ......... . .... . ................ .... 13

........... RANA LNIDA: A SPECIES COMPLEX? .....................,......*........................................... 13
THIS STUDY .............................................................................................................................. 14 Species crmcepts ..................................................................................................................... 1 5
.................................................................................................... The theoretical species concept -15
................................................................................................... The operational species concept 15
Specieç delineation ............................................................................................................. 1 9
CHAPTER 2:
............................................. MORPHOLOGY AND MORPHOMETRICS 2 0
........................................................................................................................ INTRODUCTION 21
MATERIALS AND M ~ O D S ............................................................................................. 2 1
............................................................................................... General Collecting Procedures 21
............................................................................................................................ Morphology 25
Diagnosis of Rana livida sensu stricto ........................................................................................... î5
...................................................................................................................... Morphometrics -26
.................................................................................................................................... RESULTS 27
..................................................................................................................... Morphology 2 7
Morphotype 1 . Typical ................................................................................................................. 31
............................................................................................................. . Morphotype 2 . Black Egg 31
Morphotype 3 . SpeckIed ............................................................................................................... 32
..................................................................................................................... Morphotype 4 . Small 32
Morphotype 5 . Mottied ................................................................................................................. 32
Morphotype 6 . Southern Big-Eye ................................................................................................ 32
Morphotype 7 . Northem Big-Eye ................................................................................................ 33
Morphotype 8 - Large .................................................................................................................... 33 ....................................................................................................................... Morphometrics 33

.............................................................................................................................. DISCUSSION 53
........................................................................................................................... Morphology -53
Pigmented Eggs .............................................................................................................................. 53
Vomerine teeth ................................................................................................................................ 54
Vocal Sacs ........................................................................................................................................ 55
........................................................................................................................... Size Dimorphism -55
.............................................................. Taxonomie implications of the Rana livida cornplex 57
E b w a ............................................................................................................................................ 57
.......................................................................................................................................... Odorrana 58
CHAPTER 3:
................................... ...... ALLOZYME ELECTROPHORESIS .............................. 62
INTRODUCTION ...................................................................................................................... 63
............................................................................................. Population aggregate amlysis 6 3
...................................................................................................... A priori specieç groupings 65
... ........................................................................................... MATF, FUALS AND M ~ O D S ; 6 5
.................................................................................................................... DNA Con fe nt 6 6
........................................................................................................... Protein Electrophmesis 67
................................................................................................................................. RESULTS 7 2
DNA Content ....................................................................................................................... 72
........................................................................................................... Proiein Electrophoresis 73
............................................................................................. Population Aggregate Analysis 79
................................................................................................................ Within-group genotypes 79
Among-group genotypes in sympahy ......................................................................................... 79 Among-pups in allopatry ......................................................................................................... 80
.................................................................................................................... Phenetic amlyses 80

.................. ........................................*........*.............................,.......*.............. DISCU~SION .. 88
D NA content analysis ........................................................................................................... -88
An open question .................................................................................................................... 88
Taxonomie implications ......................................................................................................... 92
............................................................................................. Implications beyund fhis s f udy 92
CHAPTER 4:
SUMMARY ............................................................................................................................... 95
........................................................................................................ Implications of the study 96
Taxonomie ....................................................................................................................................... 96 Biodiversity ...................................................................................................................................... 96
REFERENCES ................... ... .................. ........ 97
APPENDICES ............................................................ .................. .............................. 103

LIST OF TABLES
Table 1.1 Current Taxonomie Status Of Rana livida ................... .. .................... 10
Table 2.1 Locality And Seasond Data For Collections ....................... ... ........ 22
. Table 2.2 Morphotypes And Their Defining Characters ... ................. ........... 28
Table 2.3 Coefficients Of The First Principal Component ........................ ........... 35
......................................................... Table 2.4 Reclassification Matrix For Females 36
Table 2.5 Female Factor Loadings .............................................................................. 37
.......................................................... Table 2.6 Reclassification Matrix For Males 42
.................................................................................. Table 2.7 Maie Factor Loadings 50
Table 2.8 Reclassification Ma& Females And Males .......................................... 51 ... .... Table 2.9 Female And Male Factor Loadings ..................... ..... ....................... 52

.......................................................................... Table 2.10 Female-Male Size Ratios 56
Table 3.1 Morphotypes And Species Investigated With Allozyme Electrophoresis
..................... ............ .............................................................................................. 6 8
............................. Table 3.2(A) Isozyme Systems Assayed ................................ 6 9
Table 3.2(B) Buffer Systems Used ........................................... ................... ............... 7 l
Table 3.3 T-Test Of DNA Contents Of Sympatnc Morphotypes And Species . 74
Table 3.4 Genotypes For The Rana livida Complex Based On Allozyme
Electrophoresis ............................................................................................................... 75
Table 3.5 Fixed Allelic Differences Among The Rana livida Complex .............. 78
............................................................... Table 3.6 Genetic Distance Using Nei's D 82
Table 3.7 Genetic Measures Using Rogers Distance And Similarity .................. 84
Table 3.8 Phylogenetic Codings Of Allozyme Data Of The Rana livida Complex

LIST OF FIGURES
Figure 1.1 Algorithm For Species Cornplex Elucidation ................... .................. . 18 Figure 2.0 Map Of The Socialist Republic Of Vietnam .................................. 2 4
Figure 2.1 Photographs Of Morphotypes Of Rana Zivida ..................... ........ 30
................................. Figure 2.2 Plot Of Firsi Two Factors Of A DFA Of Fernales 39
..................... Figure 2.3 Plot Of First And ThUd Factors Of A DFA Of Females 41
..... .............. Figure 2.4 Plot Of First Two Factors Of A DFA Of Males ..... ............. 45
Figure 2.5 Plot Of First And Third Factors Of A DFA Of Males
......................................... ..................................................................................... 47
Figure 2.6 Plot Of Second And Third Factors Of A DFA Of Males
Figure 2.7 Some Metastemal Elements Of The Rana livida Complex And Huia
nasica .................................... .... .................................................................................... 6 1
Figure 3.1 Unweighted Paired Group Analysis Phenogram Of The Rana livida
............................ ......................**............ Complex Based On Ailozyme Data .. ....... 86

LIST OF APPENDICES
Appendix A: Specimens Studied ............................................................................. 103 . Appendix B: Measurement means for each morphotype ......................... .... 110
............................ Appendïx C: Population Aggregates Analysis .................... .. 120
.............. ............... Appendix D: Redescription of the Rana livida complex .. .. 133

Chapter 1
Introduction:
Rana livida and its importance to biodiversity

Introduction
Background
A global biodiversity crisis has emerged, due primarily to the removal of habitat
for gains of timber and crops. Because the economic payoff of this destruction is
immediate, it has becorne necessary to point out the economic value of keeping these
habitats intact, through industries such as ecotourism or natural product mining. The
value of these resources lies in the high level of biodiversi~ or uniqueness of habitats and
their constituents. For this reason, it has become exceedingly important for biologists to
document the extent of global biodiversity, particularly in regions in imrnediate danger of
habitat loss. The challenges of this charge include the recognition of species, which can
be confounded by morphological similarity or geographic dispersion. This is especialiy
tnie in the tropical regions of southeast Asia, where rainforests are being destroyed at an
unprecedented rate.
One group that presents a major challenge regarding the identification of species
diversity in southeast Asia is that of the waterfall (cascade) ranids. These rainforest frogs
are especidly susceptible to deforestation, which is occurring throughout their range. In
Vietnam alone, primary forests are being removed at a rate of 10,000 - 200,000 hectares
a year (World Wide Fund for Nature, 1998). At the same tirne, these frogs hold
potentially valuable skin secretions, which may be used in products ranging from glues
(Anderson 1993, Tyler 1996), to antibiotics (Valigra 1994, Toledo and Jared 1995,
Coghlan 1996), to prophylactics (Hunton, 1996). Maintaining their habitat is extremely
important, because their secretions originate from their habitat and/or food source (Daly

et al. 1992, Daly et al. 1994). But understanding their diversity is cnticai, since the
secretions they ernit may vary from species to species. Unfortunately, the extent of their
diversity is unknown, since striking morphologicai and habitat sirnilarities have hindered
a taxonomie study for over 150 years (Frost 1985). This is especiaily true for the green
cascade frog, Rana livida (Blyth, 1855).
Rana livida is found in fast moving montane rivers and streams of the south Asian
mainland, from southem China to the eastern reaches of the Himalaya in India (Fellows
and Chi-hang 1997, Lazell and Goodyear 1988, Frost 1985 and references therein). This
species has the same basic body plan as al1 southeast Asian cascade ranids. Its
dorsoventrally compressed body, long powerful legs, and webbed feet allow individuals
to swim in very fast moving water with ease. The tips of the digits are greatly expanded
into toe pads, which dlow these frogs to cling to both rocks i n the torrent and trees in the
surrounding forest. Rana livida exhibits incredible sexual size dimorphism, with the
females being invariably larger than the males. The female bears white eggs. Rana livida
is well known for its odoriferous skin secretions that are highly toxic. These secretions
contain enough toxins to kill other amphibians within minutes (Pope 193 1, Taylor 1962,
Karsen et al. 1986, ROM field notes 1994-1 998 unpubl.). They even have varying effects
on humans, from mild skin irritations, to stinging pain on mucous membranes and open
wounds; they may even act as a cutaneous numbing agent. Nothing is known about the
chernical composition of the toxin(s) or its purpose. It could serve as a defence against
predators (R. livida has never been documented as part of the stomach contents of
sympatric snakes), or it could act as an antifungai agent (R. livida spends much of its time

being sedentary in the spray from rainforest waterfdls). The composition and role of skin
secretions is unknown for a11 of the cascade ranids that emit them.
There is much more that remains unknown about this species including aspects of
their life history traits, behaviour, and ontogeny. It is even unclear whether R. livida can
be recognized across its range. In the following section, I review the taxonomic history
of Rana livida to outline this confusion.
Rana livida: Confusion about species identity
The taxonomic literature on R. livida reveals the sarne problems inherent with
cascade ranids, multiple synonymies and mis-identifications, even among genera. Most
of this confusion has centered around two issues: the use of toe pads as a diagnostic
character, and the striking similaxity of cascade ranids of Southeast Asia.
Rana livida (Blyth, 1855) Type locality Tenasserium, Myanmar (formerly known as Burma). Type specimens
lost.
?Polypedates smaragdinus Blyth, 18% Polypedates chloronotus Günther, 1875 Rana chloronota Boulenger, 1882 Rana livida Boulenger, 1887 .
Rana (Hylarnna) livida Boulenger, 1920 Rana grarninea BouIenger 1900 Rana (Hylorana) sinica Ahl, 1925 Rana leporipes Werner, 1930 Odorrana livida Fei, Ye, and Huang, 1990 Rana (Eburana) livida Dubois, 1992

In 1852, Blyth described what he thought was a tree frog, due to its enomous toe
pads. He narned it Polypedates smaragdinus: "Length of the body 3 '/4 inches, hind limb
5 I / q inches. Wholly green above, changing in spirit to tivid blue; underparts white"
(Blyth 1852). It is not clear where the type specimen is. In 1855, Blyth described another
frog with expanded toe pads and named it P. lividus. The location of the type specimen of
this species is also unknown.
Anderson (1871) later described a series (the Jerdon series) of frogs in the British
Museum from Darjeeling, India that he believed to be P. smaragdinus. Günther (1875)
believed that Blyth's bnef and ambiguous description of P. smaragdinus did not
constitute sufficient evidence to assign any series to P. smaragdinus. Thus, Günther
(1875) redescribed Anderson's series, naming it P. chloronatus. In 1882, Boulenger
reevaluated this sarne series and assigned P. chloronatus to the genus Rana based the
extensive webbing on the hind foot. When Boulenger (1887) described another series
(the Fea series) of amphibians from Buma, and he decided that it was "beyond doubt to
belong to the species named Polypedates lividus by Blyth, and also to be inseparable
from Günther's P. chloronotus." Boulenger mentioned P. smaragdinus in his brief
discussion of synonymy, but, like Günther before him, he did not believe that Blyth's
description of P. smaragdinus was referable to anything. He therefore chose to
synonymise P. chloronata and P. lividus under the narne Rana livida. In subsequent
publications Boulenger included P. smargdinus as a possible senior synonym to R. livida
(Boulenger 1890, 1920). This nomenclatural arnbiguity has never been investigated.

Rana graminea (Boulenger, 1899)
Type locality, Hainan Island,'China Type Specimens in the British Museum
In 1899, Boulenger described a new species of frog, Ranu graminea, from the
Chinese island of Hainan. In 1920, the same species was collected in Vietnam. The
description of R. graminea very closely resembled that of R. livida (Boulenger 1890,
1920), except that the 2 male specimens of R. grarninea were found to have slightly
larger tympani, a concave (versus oblique) loreal region, a first finger that was smaller
than the second, smaller toe pads, smooth (versus Iaterally granulate) skin, and a weak
dorsolateral fold. Smith (1930) synonymised Boulenger's R. graminea with R. liviùa
based on 2 male specimens from the Nakon Sritamarat Mountains in India. However,
many subsequent workers continued to recognize R. graminea (Pope 193 1, Boring 1932,
Bourret 1939). In 1942, Bourret considered R. graminea a "northern variety" of R. livida.
He stated that the two forms were distinct based upon the presence of a weak dorsolateral
fold in R. graminea.
Rana leporipes (Werner, 1930)
Type Locdity: southern China (no firther information given). Type specimens not
designated
In 1930, Werner described a new member of the "Rana livida group", Rana
leporipes, from southern China. He noted that this frog differed from R. livida in that its
legs were more slender and lacked any banding pattern, its discs were smaller, and it had

a weak dorsolateral fold. Bourret (1942) synonyrnised R. Zeparipes with R livida
graminea. However, the description of R' leporipes lacks some important diagnostic
characters and reports a great variation in snout-vent length (S2mm-102rnm). In addition,
there was no mention of egg colour or egg morphology. It was not clear how the lone
male was sexed, and there is no mention of either nuptial excressences or vocal sacs. The
male specimen of R. leporipes is significantly larger than those of R. livida, as
summarized by Boulenger (1920) (93 mm vs. an average of 50 mm). This would
nomally be cause for recognition of a new species. One can only assume that Bourret
(who did not acknowledge whether or not he had seen the type senes of Rana leporipes),
interpreted the fernales described by Werner as covering a range of varying degrees of
mahirity, and that the male was incomectly identified. 1
Rana sinica (Ahl, 1925)
Type Iocality China. Type specimen not designated.
Bourret (1 942) also synonyrnised R. sinica (Ah1 1925) with R. livida. In his
description, Ah1 provided a clear definition of his new species. As a cascade ranid, Ml 's
species shares many features with R. livida. However, there are enough distinct
differences between the two to make Bourret's synonymy dubious: Rana sinica has a
head longer than it is wide, which is opposite of the condition in R. livida; R. sinica has
finger 1 smaller than finger II, and finger III smaller than the snout, whereas al1
descriptions of R. livida indicate the opposite conditions; R. sinica has large, round black

dots on its sides, whereas R. Zivida does not; the obvious white lip line in R. Zivida is not
mentioned in the det@led description of R. sinica; and finaily, R. sinica has a tympanum
that is covered by a layer of skin, but R. livida has a very distinct tympanum. Bourret's
(self-adrnittedly) questionable synonymy of R. sinica with R. livida is inappropriate.
Though the current taxonomy is stable, Taylor (1962) suggested veriQing it with
a study of al1 known series of R. livida. This has never been done, primarily because
most of the type specimens cannot be traced. For this reason, major systematic works
have continued to uphold Bourret's (1943) taxonomy, which recognizes the validity of
Rana livida as a taxon even though R. sinica appears to be very different (Liu 1950; Liu
and Hu 1961; Taylor 1962; Frost 1985; Yang 1991 a; Zhao and Adler 1993; Fe et al.,
1990). The historical confusion and current stability of R. livida at the species level is
rnirrored by the relative historicat stability and current confusion of its higher taxonomic
position.
Rana CividQ: Generic and subgeneric confusion
The current generic and subgeneric placement of Rana livida is unstable.
Although long regarded as being in the genus Rana, R. livida is variously placed in the
subgenus Eburana, Hylarana, and Odorrana. The latter two taxa are variously ranked at
the generic by some workers and at the subgeneric level by others. Confusion of the
higher taxonomy stems from the fact that Rana livida exhibits diagnostic characters of

each taxon: the fuiiy webbed and finnistemal condition and webbed feet of the R m ,
enlarged toepads of the Hylarana, odoriferous secretions of the Odorramz, and the white
eggs of Eburana. Here, 1 review the higher level taxonomy that includes R. livida and
discuss the validity of the inclusion of Rana livida within each of them (Table 1.1).
Rana Linneaus, 1758
Range cosmopolitan. Type species Ranu temporania (Linneaus, 1758)
The concept of the genus Rana has changed since its original description by
Linneaus (1758). M e n Boulenger (1887) realized that Polypedates lividus was actually
a Rana, he did so because the latter genus was already recognized by a suite of diagnostic
characters that is still recognized today. Boulenger moved P. lividus into Rana based on
its extensive webbing and t-shaped phalange. The genenc position of R. livida rernained
. stable for almost a century.
Hylarana Tschudi, 1838 Hylorana Günther, 1864
Range Africa, Asia. Type species Hyla eruthrae (Schlegel, 1827)
Hylarana Tschudi, 1838 was erected as a genus based on the presence of a
horseshoe-shaped circummarginal groove on the ventral surface of very large toe pads.

Table 1.1. Current taxonomic status of RaM livi& (Blyth 1855). Boulenger (1920) placed it
in the subgenus Hylarana. Fei et al. (1990) placed it in a new genus, Ohrranu. Other workers
have since believed that Odorrana deserves subgeneric status, due to its equivocal genus-level
characters (Matsui 1994, Zhao 1994, Inger 1996). Dubois (1992) rearranged the taxonomic
hierarchy by placing R. livida into the new subgenus Eburana, which was in the section
.Hylarana, and subsection Hylarana of the genus Rana. This taxonomic arrangement has corne
under spirited criticism (see text).
Rana lirtida (Blyth, 1855)
&na (Hylarana) livida
Odorrana livida
Rana (Odowuna) liuida
Rana
Hylarana (secf ion)
Hylaram (subsec fion)
Eburana (subgenus)
R. livida
(Boulenger, 1920)
( Fei et a1.1990)
(Matsui 1994, Zhao 1994, Inger,
1996)
(Dubois, 1992)

However, the plasticity of this character, both intra-specifically and interspecifically, has
led to much taxonornic instability. Grooves can range from being very distinctive to
absent. Although Boulenger (1 882) originally believed that Hylarana should not be
recognized at all, he subsequeniy nvised and refined the definition of it as a subgenus
(Boulenger 1920). He included R. livida in that t aon based on the presence of ventral
grooves of the large disks. Confusion regarding the status of Hylarana continues to the
present day. Biologists working on the African fauna rank it at the generic level, whereas
workers on the Asian fauna consider Hylarana a subgenus of Rana (Frost 1985).
Currently, workers refer to R. livida as a member of the subgenus Hylarana (Bourret
1942), or ignore subgeneric status (Taylor 1962, Lazell et. al. 1988, Zhao and Adler
1993).
Odorrana Fei, Ye, and Huang, 1990 Range: southeast Asia. Type species Rana margaratea (Liu, 1950)
Fei et al. (1990) placed 12 species of odoriferous Hylarana in a new genus,
Odorrana. The genus was defined primarily by extemal morphology and diagnosed by
stemal elements: "omostemum extremely small, base not forked; central sternum long
and slender, base thick, [xiphistemum] much larger than ornosternum, deepIy notched
posteriorly" (Fei et al. 1990). The authors neglected to include odoriferous skin
secretions in their description, even though it was the basis of the taxonornic etymology.
This new genus represented the first revision of the Hylarana since Boulenger redefined
that taxon in 1920. Boulenger (1920) had allied R. livida with the Rana (Hylarana)

chalconata group. However, later karyological studies suggested that R. livida had a
closer affinity with members of the Rana (Warana) anàersonii group (Li and Wang
1985). Ln response, Fei et al. (1990) grouped the entire Rana andersonii group and R
livida into the Odorrana. Neither Yang (1991a) nor Zhao and Adler (1993) recognized
this new genus when cataloguing Rana livida as part of the amphibian fauna of China.
Dubois (1992) believed that the diagnostic characters were insufficient to gant Odorrana
generic status, so he subsumed it as a subgenus of Rana. Zhao (1994) agreed with this
subgeneric status, as many of the 'diagnostic' characters for Odorrana are shared by
other members of the genus Rana.
Eburana Dubois, 1992 Range: southeast Asia, Japanese archipelago. Type species: Rana narina
(Stejneger, 190 1)
In his provisional classification of the farnily Ranidae, Dubois (1992) erected the
subsection Hylarana, which was composed of ten subgenera, including revisions of
Hylarana and Odorrana, and a new subgenus Eburana. The subsection Hylarana was
defined by the lack of humera1 glands in the males. Dubois removed R. livida from
Odorrana, placing it into a new subgenus, Eburana. The new subgenus (composed of R.
lividu, R. swinhoana, and R. narina) was defined by the l a r d dentical formula (4-5/4),
the absence of nuptial spines on the chest of the males, and the absence of pigmented
eggs-
This classification has since come under spirited criticism, due to both
philosophical and empirical probkms. The philosophical weaknesses of Dubois' work
stems from the phenetic basis of the classification, a cavalier treatment of characters

inferred from the literature, and an inadequate analysis of variance of the defrnng
characters (Emerson .and Berrigan 1993, Matsui et al. 1993, Matsui et al. 1995, Inger
1996). Dubois used larval dentical formula to place R. livida in Eburana, even though its
tadpoles are unknown. Empiricaliy, karyotypic evidence suggests that R. livida shares a
doser affinity with Odorrana (sensu Fei et al. 1990) than with other Eburana (Wei et
al. 1992; Matsui 1994; Matsui et al. 1995). Furthemore, R. livida is the only member of
Eburana to have odoriferous skin secretions.
Rana livida has been included in al1 of these reclassification schemes and current
workers relegate this species to any one of the three subgenera Hylarana, Odomana, or
Eburana. Several workers have been wary of accepting the validity of Odorrana as new
genus due to its equivocal characters (Dubois 1992, Zhao 1994, Matsui 1995, and hger
1996). Their collective recommendation of recognizing Odorrana as a subgenus is
foIlowed here. Several workers have also raised doubts about the validity of Eburana.
Though a phylogenetic analysis or further rnorphological evidence is necessary before
Eburana cm be declared invalid, the skepticism remains warranted.
Rana livida: A species complex?
Taylor (1962) noted that differences among populations of Rana livida were
marked, implying that this taxon may be a species complex. For this reason, he believed
that a review of al1 available series (including the type specimens) was a necessary first
step for testing the veracity of the current taxonomy. Comparative exarnination of the
species has been complicated by several factors. First, the type series of R. livîda, R.

leporipes, and R. sinieu cannot be located, as the original descriptions do not include
specimen accession numbers. Second, currently recognized synonyms were collected
from disparate parts of the huge geographic range of R. livida. This presents a logistical
problem for viewing original material, since collections are scattered throughout the
world. It also presents a conceptual problem, since the disparate sampling areas make it
unclear whether or not the variation is due to geographic variation or speciation. These
problems have prevented anyone from investigating the validity of the single species
designation of this taxon, until now. Beginning in 1994, field crews from the Royal
Ontario Museum (ROM) collected specimens of R. livida from three localities in northem
Vietnam. Differences in size and colour patterns of both sympatric and allopatric
specimens were apparent. Subsequent collections from other localities within Vietnam
resulted in the discovery of additional morphotypes. With the discovery of populations
of R. livida that exhibited evident morphological differences within a very small area of
its geographic range, a detailed study of this taxon is now possible.
This Study
In this study 1 endeavour to determine whether or not R. livida is a single species
or a complex of species. If it is a complex, then it suggests that there is diversity present,
undiscovered or unrecognized by Western science. It is also possible that the junior
synonymies could apply to one or more of the morphotypes identified in the present
study .

Species concepts
Because the objective of my study is to delineate species boundaries, it is
necessary to define rny concept of what constitutes a species. Mayden (1997) outlined
one approach to this enigrnatic problem. The cmx of the species problern, he argues, lies
in the dual nature of the biologicd classification system, whereby it documents both
hierarchical historical relations and evolutionary distinctiveness. It is unclear which
criterion is optimal for establishing a classification: the descriptive nature of the
relationships or the prescriptive nature of the distinctiveness. Mayden suggests that,
because the two criteria can never be fully reconciled with each other, a hierarchy of the
two concepts should bc used for a species definition: one concept is used for the
theoretical basis and the other subordinate for the operational basis.
The theoretical concept
Only one species concept can serve as the theoretical basis for our understanding
of the nature of species: the evolutionary species concept (Wiley 198 1). It is the only
species concept that recognizes and accommodates al1 known types of biological
diversity (see also Frost and Hillis 1990, Frost and Kluge 1994, Mayden and Wood
1995). This concept defines a species as a "single lineage of ancestor-descendant
populations which maintains its identity from other such lineages and which has its own
evolutionary trajectories and historical fate" (Wiley, 198 1 , p 25). The evolutionary
species concept, however, does not give any operational clues for recognizing these
lineages.

The operationul concept
The operationai species concept used in this shidy is the phylogenetic species
concept (PSC) sensu Nixon and Wheeler (1990). This is also known as the diagnosable
concept, because it recognizes species as "... the smallest aggregation of populations
(sexual) or lineages (asexual) diagnosable by a unique combination of character States in
comparable individuals (sernaphoronts) " (Nixon and Wheeler 1 990). This concept is
well suited for this study because it is consistent with the evolutionary species concept,
and it provides an operational tool for diagnosing species boundaries. One of the
drawbacks of the PSC is that it can result in the unnecessary oversplitting of species
(Frost and Hillis 1990). The problem is that species are defined in tenns of the smallest
cluster of individuals. Davis and Nixon (1990, 1992) suggest that one fixed difference
between populations constitutes a boundary between species. However, this parameter is
too loosely defined, as micro-lineages could exhibit fixed differences within a few
generations (Wake and Schneider 1998 and extensive references thersin). Frost and
Hillis (1990) highlight this point with their example of the absurdity of a new species for
each jar of fruit flies. I have endeavoured to correct for this problem by imposing stricter
constraints on the diagnosis of a species boundary. This was accomplished by using a
rnulti-faceted approach to finding diagnosable characters for species delineation (Figure
1.1).

Figure 1.1 - The three stage algonthm for elucidating evolutionary species in a species complex.
Fust, specimens are grouped morphologically. Those groups whose members show significant
differences are separated as distinct species. Those groups whose differences are considered
equivocal at a species level are placed into morphological groups (morphotypes). Next, the
relative strength of those morphotypes is tested by mnning a discriminant function analysis
(DFA) and cellular DNA content analysis on them. Morphotypes that are consistent with a DFA,
and have discreet Ievels of cellular DNA content are considered strong candidates for distinct
species. These make compelling a priori groupings for the third stage of the investigation,
aliozyme electrophoresis. Diagnostic characters from an electrophoretic study are used to
support previously established species boundaries and suggest new ones.

Many Species
Morphology
Identify some distinct species Identifv more distinct species Significant morphological Significant differences in
differences indicative of distinct evolutionary
lineages
gene flow indicative of distinct evolutionary
lineages

Species delineation
The first part of the investigation involves a morphological study. Specimens of
the ROM collection from Vietnam were divided into different morphologicai groups.
These morphotypes were then compared with descriptions from the literature and with
independently collected series from southeastern China and India. If evident
morphological differences were found, then species boundaries were established on the
basis of morphology alone. Second, the remaining morphologically recognized groups
were tested with discriminant functions analyses @FA) and DNA content andysis.
While DFA groupings and DNA content are inappropriate to use as diagnosable
characters of species, they can be highly suggestive of species boundaries. More
importantly, they validate a priori groupings for use in the third part of the investigation:
allozyme eIectrophoresis. Allozymes are used to find diagnostic characters in the form of
fixed allelic differences. The above analyses were used in concert to discover and
delineate species boundaries within R. livida. This study is the first of its kind for the
cascade ranids.

Chapter 2
Morphology and Morphometrics

Introduction
Cryptic species can be 'indistinguishable' morphologically but differ in other biological
characteristics. The Ievel of morphological similarity may Vary. The creation and
subsequent synonymy of 'new species' of Rana livida from specimens collected from
many disparate locales, and housed in institutions ail over the world is reason enough to
suspect that a carefûl comparative morphological s tudy, even arnong geographicail y close
populations, has never been done. Recent collections acquired by Royal Ontario Museum
(ROM) have resulted in series of R. livida from a small area within the species
geographic range. Populations within Vietnam exhibit evident morphological differences
from each other, suggesting that rnorphological studies will play an important role in
elucidating species diversity within the cornplex.
Materiais and Methods
General Collecihg Procedures
Specimens were collected by field crews of the Royal Ontario Museum (ROM)
from several localities within Vietnam at various times of the year from 1994-1997
(Table 2.1, Figure 2.0). Most frogs were collected at night and euthanised using MS-222
within 24 hours of collection. Frogs were first fixed in 90% ethanol for 24 hours, and
then preserved in 75% ethanol.

Table 2.1 Locality and seasonal data for collections of Rana livida. Where possible, exact
coordinates and months are given. Missing data suggest that the information was not
available.
Locality Coordinates, Season If known (Years)
Tarn Dao,
Vinh Phu Province, Vietnam
Ba Be National Park,
Cao Bang Province, Vietnam
Pac Ban, Nha Hang Nature Reserve, A
Vietnam Tuyen Quang Province
Sa Pa
Lao Cai Province, Vietnam
Khe Moi River, Con Cuong
Nghe An Province, Vietnam
Tram Lap, Gia Lai Province, Vietnam
Hong Kong
People's Republic of China
Darjeeling province India
ChWungan Hsien and Yenping Fujien Province, People's Republic of China
21°27'N. 105'39'~ October (1 994) May-June (1995, '96,'97)
October (1994) May (1 995)
Oc tober (1 994) June (1995)
July (1993) Juty (1997)
Apnl-Sep (1931)

Figure 2.0 Map of the Socialist Republic of Vietnam. Collection sites are indicated with stars
and bold letters.


Morphology
A total of 222 adult specimens of Rana livida were examined from locdities in
Vietnam, Darjeeling, India, and Hong Kong, China (Appendix A). The sex of each frog
was recorded and d l measurements were made to the nearest .O5 mm with digital calipers
(Appendix A). Extemal measurements were: snout-vent length (SVL); snout length;
orbit diameter; inter-orbital distance; tympanum diameter; tympanum-eye distance;
finger II disk length; finger I I disk width; toe DI disk length; toe IIl disk width.
Osteological measurernents were made on x-rays taken with a faxitron at the ROM. The
measurements were as follows: head length - posterior edge of otoccipital to tip of
premaxillary; head width - between left and right articulations of the quadratojugal and
rnaxillary; finger length - tip of the distal phalanx to the base of the proximal phalange of
finger II; hand length - distal end of radio-ulna to tip of distal phalanx of If; foot Iength -
distal end of tibia to tip of distal phalanx of III; tibia1 length - length of tibia. As well,
some specimens that had been partially dissected for tissue sampling, were examined for
sternal sstnicture.
Diagnosis of Rana livida sensu stricto
Al1 specimens were cornpared with the diagnoses from Boulenger (1920),
Bourret (1942), and Fei et aL(1990). The snout-vent length ranges from 45-50 mm for
mdes and 90- 1 0 0 m for fernales. Vomerine teeth are present in rows that are oblique to
the intemal choanae. A white lip line extends across the upper lip ending in a glandule
just anterior to the tympanum. The feet are fûlly webbed to the distal phalange and

brown in colour. The skin is smooth donally. becoming increasingly granulated laterally.
No dorsolateral folds are present. The dorsum is deep green. The legs are banded. The
xiphistemum is widely crescent-shaped, with a deep posterior notch. Males have paired
subgular extemal vocal sacs. Females bear white eggs.
Specirnens consistent with this diagnosis were considered part of the Typical
group, whereas those not conforming to the diagnosis were put into separate
morphological groups (referred to here as morphotypes). These morphotypes were then
compared with two synonyrnous forms of R. livida: R. grarninea and R. sinica. Rana
graminsa was distinguished by the presence of weak dorsolateral folds, larger tympanum
and small toe pads, whereas Rana sinica was noted for its large snout-vent Iength (males,
66 mm), and a layer of skin overlying its tympanum. The description of the third
synonymised form, R. leporipes, is too vague for any meaningful comparison. If the
morphotype was not congruent with one of the synonymous groups or the Typical form,
it was considered to be new. In addition, a series of R. livida (AMNH 29973-29991,
28543-5,28612) originally identified as R. graminea from Fujien, China (Pope 1931)
was compared with the descriptions of the synonymous forms and new morphotypes.
Morphometrics
Morphometric analyses were perforrned on morphotypes based on the initial
anatomical analyses. These analyses follow recornrnendations from Tabachnick (1989),
Hair et al. (1992), and Stat Soft (1995). Raw morphometric data were log-transformed.
Data were then tested for normality using categorized probability plots. Variables were

tested for heterogeneity using serial correlation plots. This was also used to detect
redundancies among the variables. The morphometry was designed to factor out the
effect of size on the data set. A pooled within-group correlation-variance matrix was
used in a principal component's analysis (PCA) . The first principal component (PC1)
was used as a size component and regressed against each log-transformed variable. The
resulting residuals were used in a pooled-within group correlation-variance matrix for a
discriminant functions analysis @FA). A priori classification error rates were al1 made
proportional to the group size. The strength of the morphotypes was tested by
reclassification methods. Where possible, large groups were sub-divided, with part of the
grmp being used in the initial DFA, and the other part being used in a reclassification.
As well, the specimens from hd ia were included in a reclassification. Plots of the first
three discriminant functions were used as visual aids in the separation of morphotypes.
Sexes were treated both separately and together in the DFA. Al1 analyses were perfonned
on STATISTICA ver. 5.1 (StatSoft, 1995).
Results
Morphulogy
Eight groups were distinguished using morphological differences (Table 2.2,
Figure 2.1). These groups are refemed to as rnorphotypes or operational taxonomie units
(OTUs) for the purpose of this discussion. Summaries of al1 the rneasurement means are
shown in Appendix B. The Typical morphotype was composed of specimens that did not
differ from the diagnosis.

Table 2.2 List of morphotypes and their defining morphologicai characters. Refer to Appendix 1
for a key to locality abbreviations.
M o ~ h o t ~ P Localities Defining Characters
Typical form
B lac k Egg fom
Large fonn
Northern Big Eye
Southern Big Eye
Mottle
Speckle
PB, B, TD, KM conforms to diagnosis of R. livida
sensu stricto
PB, B. TD, KM Black Eggs
Webs Marbled White and Black
Large body size
Dorsum shagreened, not srnooth
Bulging Eye
Brown Dorsun
Bulging Eye
Brown Dorsum
Males slightly Iarger
Brown night colour
Green day colour
Males large, females small
Heavy granules on fi&
Vocal dits
Very small body size
White dorso-lateral granules
Males with large tympanum
(314 diarneter of orbit)

Figure 2.1 Photographs of different morphotypes of Rana livida studied here. The
Typicai form (a) known from Darjeeling, India, China (Hong Kong, and Fujien Province), and
severai locales in Vietnam (Ba Be, Cao Bang Prov., Sa Pa, Lao Cai Prov., Pac Ban, Tuyen
Quang Prov. , Tarn Dao, Vinh Phu Prov., Khe Moi River, Nghe An Prov.). The BIack Egg form
(b) is found in northem Vietnam, (Ba Be, Tarn Dao, Pac Ban, Khe Moi River). Two forms, the
Speckled form (c) and the Small form (d), are known only from the far north of Vietnam: Sa Pa.
The Mottled form (e) and the southern Big Eye f o m (f) are h o w n from the centrai highlands of
Vietnam (Tram Lap, Gia Lai Province). The northem Big Eye fonn (g) is known from Pac Ban.
The Large fom is known from the Khe Moi River (h).


Morphotype 1- 'Typical": 57 males. 37 females (Figure 2. la). Refer to diagnosis above.
This morphotype found throughout northern Vietnam. Males have a mean SVL of 46.35
mm (40.46-53.08 mm), whereas fernales average 92.53 mm (80.35-101.74 mm). Ratio of
head length: head width is 0.97 for males, 0.84 for females.
No distinct morphological differences were found between the Chinese specimens
originally identified as R. grarninea (AMMH 29973-29991,28543-5,28612) and the
Typical morphotype of R. livida. The 15 Chinese males had a mean SVL of 46.7 mm
(4I.65-53.5mm), whereas the 4 females had a mean of 97.1 mm (94.75- 10.15 mm).
There was no dorsolateral fold present, nor was there a difference in the tympanum : eye (
0.75 for the Chinese specimens, 0.73 for those from Vietnam). The toe pads of the
Chinese specimens, however, were smaller at 1.70 mm (1.3-1.9 mm) than those of the
Typical form, 1.82 mm (1.61-3.44 mm), as described by Boulenger (1920). Both of the
Indian specimens were also of the Typical form. They had SVL's of 46.47 and 46.99
mm.
The remaining seven morphotypes differed morphologically from the Typical
morphotype and R. livida synonyms R. sinica and R. graminea. The characters that
distinguish the seven groups are outlined below (Table2.2):
Morphotype 2- "Black Egg": 1 1 females (Figure 2.1 b). This fonn is found
throughout northern Vietnam except the far north. Only fernales are known. A white iip
line is absent, and vertical lip bands are present. The fully webbed feet are marbled with
black and white to the toes. The skin is shagreened and mottled brown. This was
recognized as a new species due to the presence of black eggs (in contrast to the white
eggs borne by the Typical morphotype).

Morphotype 3- "Speckled": 3 males, 6 fernales (Figure 2. lc). This fom,
recognized as a distinct species, is found only in the far north, Lao Cai Province,
Vietnam. Males and females exhibit a lower level of sexual size dimorphism than the
other forms, with SVLs 57.03 mm (54.68 - 58.93 mm) and 90.24 mm (74.33-86.81 mm)
respectively. Dorsum has green speckles on smooth brown skin, which becomes heavily
granulated at the flanks and pelvis. The feet are fully webbed, but only to the second
phalanx of the middle toe. This form is unique in the presence possession of vocal slits,
whereas ail oîher forms have paired extemal sacs.
Morphotype 4 - "Small": 15 males, I femaIe (Figure 2. ld). This f o m is found
only in the far north, Lao Cai Province, Vietnam. It is also regarded as a separate species,
varying significantly from the Typical morphotype. Mernbers of this species are very
small, snout-vent length 35.52 mm (32.00-38.10 mm) and 55.66 mm in the male and
female respectively. Vomerine teeth are absent. White dorsollaterai glandules run dong
the back, and a large white spot appears on each flank. The xiphistemum is forked, as in
frogs of the genus Huia.
Morphotype 5 - "Mottled": 21 males, 6 females (Figure 2. le) This morphotype is
known only from the Central Highlands of southem Vietnam. The frogs are small, SVL
43.21 mm (39.18 - 45.86 mm) for males and 87.58 mm (80.40-99.57 mm) for females.
Colour varies from brown at night to green in daylight. The subarticular tubercles are not
pronounced, as in al1 other forms.
Morphotype 6- "Southern Big-Eye": 13 males, 10 females (Figure 2. lf) This
morphotype is found only in the Central Highlhds of southern Vietnam, and its
description closely matches that of Morphotype 7. The SVL of the males and females are

50.25 mm (42.52-54.60 mm) and 92.68 mm (83.35-98.72 mm) respectively. The skin is
variable; it c m be either shagreened or smooth; it can be spotted andor sofidly coloured;
and it can be brown or green in colour. Leg bands are absent. The eye bulges noticeably.
Morphotype 7- "Northem Big-Eye": 3 males (Figure 2.Ig). This has been found
in the Nah Hang Nature Reserve, Pac Ban in northem Vietnam. Only males are known.
The SVL is Iarger than that of the Typical morphotype at 52.29m.m (48.57-55.17 mm).
The shagreened skin is spotted brown-olive. The eye bulges visibly.
Morphotype 8- "Large": 10 females (Figure 2. lh). Found in north-central
Vietnam. Only females are known. The SVL is very large at 100.3 mm (93.59-105.34
mm). The white lip line is absent. The skin is shagreened and is olive-brown. A deep
supratympanic fold is present, unlike the condition of the Typical.
Morphometries
Morphometric anaIyses was performed on female specimens of the Typical,
Mottled, Speckled, and Large morphotypes, and on male specirnens of the Typical,
Mottle, Southern Big Eye, and Small morphotypes. Srnall sample sizes precluded the
analysis of the rernaining specimens. Black Egg and the series fiom China were excluded
from the analyses, as osteological measurements were not available. The sarnple size of
Typical was the only one large enough to be divided into two subsets for reclassification.
Al1 other rnorphotypes were included both in the classification and reclassification using
DFA.

Categorical probability plots show normal fnquency distribution among the log-
transfonned data. A high correlation between finger length and hmd Iength, and &O
between toe length and foot length was detected in serial comlation plots. Both the
finger and toe length data were omitted fiom the analysis as they were redundant
variables. The loadings of PC1 clearly show that it represents a size factor (Table 2.3).
A DFA of female specimens indicated strong, detectable patterns of
morphological variation in the data. The analysis resulted in an associated F-statistic of
6.86 (p<.0001). ~eclassificatioi of al1 four OTUs was performed at a rate of 90% or
higher (Table 2.4). The discriminant loadings indicate that tympanum size has the
greatest effect on Factor 1, while SVL has the greatest effect on Factor 2, and finger pad
width had the greatest load on Factor 3 (Table 2.5). A plot of Factor 1 vs. Factor 2 shows
clear differentiation among three morphotypes: Typical, Large, and Speckled (Figure
2.2). The Speckled morphotype has a centroid within the 95% confidence limit of both
the Large and Typical groups. A plot of Factor 3 vs. Factor 1 shows separation of the
three groups (Figure 2.3).
A DFA of the four male morphotypes resulted in an associated F-statistic of 9.64
(pc.0001). The Typical and Small morphotypes were found to be very distinct, as the
correct reclassification scores were 97% for the former and 100% for the latter (Table
2.6). The two Indian specirnens were reclassified as members of the Typical morphotype.
The reclassification score of Mottled was also robust at 84%, well above the a priori error
rate. Al1 of the incorrectly classified Mottled OTU's were instead classified as Southern
Big Eye. The percent reclassification of Southern Big Eye, 54%, was well above the a
priori expected error rate of 14%, but was still equivocal with the

Table 2.3 Coefficients of the fmt principal component (PC1) for a principal components
analysis (PCA) of 17 variables. Refer to Appendix 1 for variable abbreviations. AU variables
exhibit high scores, indicating that PC1 is strongly correlated with size, and, as such, serves as a
good size factor.
Variable PC1
SVL 0.982 HDL 0.963 HDW 0.970 FGR 0.962 HND 0.968 TL 0.845 FT 0.8 12 TIB 0.983 LOD 0.782 EYE 0.772 TEYE 0.932 TMP 0.697 SNT 0.897 FPL 0.9 17 FPW 0.896 TPL 0.93 1 TPW 0.923 Exp1.Va.r 13 .765 PrpTotl 0.8 10

Table 2.4 Redassification matrix for f d e morphotypes of the Rmia 1mUla complex
from a discriminant functions analysis (DFA). The following groups were anaiyzed: the
Typical forrn, the southem Big Eye form, the Speckle fom and the k g e form. The
redassification score was over 90% for each group.
Typical (F) 97.83 45 1 O
s Big Eye (F) 100.00 O 10 O
S~eckle (F) 100.00 O O 6
Large 0 90.00 1 O O
Total 97.22 46 11 6

Table 2.5 Factor loadings of female morphotypes of the Ram livida cornplex from a
discriminant functions analysis (DFA) for 14 variables. The foIlowing groups were
andyzed: the Typicai form, the.southem Big Eye form, the Speckle fomi and the Large
form. Refer to appendix 1 for variable abbreviations. The loadings indicate that
tyrnpanum size had the greatest effect on Factor 1, finger pad width and head width
had the greatest effect on factor 2, and head width and orbit diameter had the greatest
influence on factor three.
Variable Factor 1 Factor 2 Factor3
HDL
m w HND
FT
TIB
IOD
EYE
TEYE
TMP
SNT
FPL
FF'W
TPL
TPW

Figure 2.2 A plot of the firçt two factors resdting from a discniminant function analysis
(DFA) on female morphotypes of the Runa livida complex. The legend on the graph
indicates which symbol represents each morphotype: the Typical form, the southern Big
Eye fonn, the Speckled fom, and the Large form. Note the unclear separation of the
Typical and Mottled forms in this graph.

Female Factor Plot
- /-----,- .
Typical SEye Speckled
-6 -4 -2 0 2 4 6 8 + Large
Factor 1

Figure 2.3 A plot of the fust and third factors resulting from a discriminant function
analysis (DFA) on f e d e morphotypes of the Rana livida cornplex. The legend on the
graph indicates which symbol represents each morphotype: the Typical form, the
southern Big Eye form, the Speckle form, and the Large form. Note the clearer
separation of the Typical and southern Big Eye forms versus that found in Figure 2.2.

Female Factor Plot
Typical SEye Speckied Large
Factor 1

Table 2.6 Reclassification ma& for male morphotypes of the &na liuîdu complex from a
discriminant functions analysis (DFA). The foliowing groups were analyzed: the
Typical f o m the s&them Big Eye form, the Mottle form and the Small forrn. The
reclassification score was.over 85% for all of the morphotypes except the southern Big
Eye for, which only had a 36% success rate of reclassification. All of the incorrect
rdassifications for the southern Big Eye form were of the sympatric Mottle form,
indicating their morphometric similarity.
Percent Typ (M) sEye (M) Mott (M) Smaü 0
TYP (M) 97.22 35 1 O O
SEye (M) 36.36 1 4 6 O
Mott (M) 84.2 1 O 3 16 O
Small (M) 100.00 O O O 12
Total 85.90 36 8 22 12

Mottle morphotype. The similarity among the two sympatric morphotypes from
the south is reflected in the factor plot of the groups (Figures 2.4,2.5,2.6). In no
combination of the three factor plots could Southern Big Eye and MottIed be reasonably
separated. The factor loadings indicate that the first factor is most indicative of
tympanum diameter, while the second is based rnainly on the SVL, and the third factor is
indicative of finger pad with and head length (Table 2.7).
A DFA of the combined male-female data set resulted in an F-statistic of 6.78
(p<.Oûûl). Reclassification of al1 OTUs were comparable to those carried out on separate
data sets (Table 2.8). Al1 groupings were morphologically distinct , with the exception of
Southern Big Eye, which was reclassified as Mottled 54% of the time. The reverse was
not found to be true, as Mottled was clâssified correctly 84% of the time, well above the
0.13 a priori error rate. The discriminant loadings were such that tympanum had the
greatest effect on factor 1, snout-vent length had the greatest effect on factor 2, and finger
pad length, dong with toe pad length and orbit diameter had the greatest effect on factor
3 (Table 2.9).

Figure 2.4 A plot of the first two factors resulting from a discriminant function analysis
(DFA) on d e morphotypes of the Rana livida cornplex. The legend on the graph
indicaies which syrnbol represents each morphotype: the Typical forrn, the southern Big
Eye form, the Mottle form, and the S d fom. Note the uncIear resolution of the
Typical and Mottle forms.

Male Factor Plot
Typical SEye Mottled Small
Factor 1

Figure 2.5 A plot of the first and third factors resuiting fkom a discriminant function
analysis (DFA) on male morphotypes of the Rana livida complex. The legend on the
graph Uidicates which symbol represents each morphotype: the Typical form, the
southern Big Eye form, the Mottle form, and the S d fonn. Note the uncleat
resolution of the Typical and Mottle forms, as in Figure 2.4.

Male Factor Plot
Typical
SEye (Ml Mottled
-10 -8 -6 -4 -2 O 2 4 6 A Small
Factor 1

Figure 2.6 A plot of the second and third factors resulting from a discriminant function
analyçis (DFA) on male morphotypes of the Rana livida complex. The legend on the
graph indicates which symbol represents each morphotype: the Typical fom, the
southern Big Eye form, the Mottle form, and the Small form. Note the unclear
resolution of the Typical and Mottle forms, as in Figures 2.4 and 2.5.

Male Factor Plot
Typical SEye Mo ttled Smaii
Factor 2

Table 2.7 Factor loadings of male morphotypes of the Ranu ZiW complex from a
discriminant functionç analysis (DFA) for 14 variables. Refer to appendix I for variable
abbreviations. The loadings indicate that tympan- size had the 'greatest effect on
Factor 1, orbit diameter had the greatest effect on factor 2, and finger pad width, head
length and tympanum diameter had the greatest influence on factor three.
variables Factor 1 ~actor 2 Factor 3
HDL
HDW
HM>
FT
TIB
IOD
EYE
TEYE
TMP
SNT
FPL
FPW
TF'L
TPW

Table 2.8 Redassification ma& for fernale and male morphotypes of the Rana limda
complex from a combined discriminant functions analysis (DFA). The.foUowing groups
were analyzed: the Typical form (male and female), the southern Big Eye form (male
and f e d e ) , the Speckle form(female), the Large form (female), the Mottle form (male),
and the S m d form (male). The reclassification scores were comparable with scores
when sexes were treated separately (Table 2.4 and 2.6), although the overall success is
slightly Iower.
Percent T(M) T(F) sE(M) SE(??) Mtt 0 Sp(F) L g o Sm(M)
Typical (M)
Typical (F')
SEye (M)
SEye (FI
Mottle (M)
Speckle (F)
Large (FI Smail (M)
To ta1

52
Table 2.9 Factor loadings of female and maïe morphotypes of the Rmia liv& complex
from a combined disaiminant factor analysis (DFA) for 14 variables. Refer to appendu
1 for variable abbr&iations. The foilowing groups were analyzed: .the Typical form
(male and female), the southern Big Eye form (male and f e d e ) , the Speckle
fonn(female), the Large form (female), the Mottle form (male), and the Small form
(male). The loadings indicate that tympanum size had the greatest effect on Factor 1,
orbit diameter and finger pad width had the greatest effect on factor 2, and head width
and tibia1 length had the greatest influence on factor three.
Variables Factor 1 Factor 2 Factor 3
HDL
HDW
HND
FI'
TLB
IOD
EYE
TEYE
TMP
SNT
FPL
FPW
TPL
TPW

Discussion
Morphology
Initial anatomical analyses elucidated the presence of three distinct species, here
referred to as the BIack Egg species, the Small species, and the SpecMed species. In
addition to these species, morphometric anaiysis suggests three additional morphotypes
among Rana livida complex frogs. The Typical morphotype is the only one that had a
large enough sample size to be split into two subsets: one for establishing the
discriminant factors and one to test the reclassification. Members of the subset tested for
reclassification were correctly assigned to the Typical morphotype. Al1 of the other
foms were reclassified using the same data set, which introduces an optimistic b i s .
Regardless, most of these forms were reclassified with 90% accuracy. The failure of the
DFA to consistently recognize Southern Big Eye and Mottled as distinct groups, even in
light of the optimistic bias of the test, ernphasizes their great degree of morphological
similarity. Though the sirnilarities in the overall body fonn of the entire complex are
striking, some of the differencesare distinct.
Pigmented Eggs. The BIack Egg morphotype differs from the rest of the group
in having pigmented eggs. This important morphological difference is also indicative of
a behavioural difference with regard to egg deposition sites. Amphibian eggs that lack
melanin are deposited out of the sunlight, whereas those with melanin require sunlight

and thus are deposited in the sun (Dueuman and Trueb 1986 p. 114). Frogs of the
Amolops group of genera, cascade ranids of southeast Asia that also bear white eggs, are
known to lay their egg masses in the shade, on submerged rocks in fast moving waters
(Pope 1930, Liu 1950, ROM field notes unpubl.). Egg deposits of frogs referable to the
Rana livida group (including the Black Egg form) have not been reported. However it is
predicted that the Black Egg morphotype lays its eggs such that they are exposed to
sunlight, whereas the white egg bearing forrns Iay their eggs out of the sunlight.
Vomerine teeth. The Small form found in the far north of Vietnam has many
morphological differences from other Rana livida group species. Its small size, white
dorsolaterd granules, and large lateral white spot are indicative of a distinct species. One
character that stands out, however, is the condition of the vomerine teeth. Ali of the other
species and morphotypes of the Rana livida group exhibit paired sets of vomerine teeth
on the upper palate, whereas these are absent from the Small species. This is indicative
of a different mode of prey capture (Duellman and Trueb 1986 pp 232-240). Anurans
capture their prey with sticky tongues and then manipulate them into a position such that
they can be swallowed whole. Once captured, large and vigorous prey c m be easily
manipulated by being pressed against the vomerine teeth. Frogs without vomerine teeth
cannot have the same range of prey as those with vomerine teeth. Instead, they rely on ..
food sources that are small and docile enough to be swallowed with minimal handling.
This implies that the Small species has a diet that differs from those of other Rana livida
group mernbers.

Vocal Sacs. The Speckled morphotype is distinct from the rest of the group by
the presence of vocal slits. A11 other members bear paired, subgular, external vocal sacs.
The Rana livida Typical form is known to have a high-pitched chirp-like cd1 (Boulenger
1920, Pope 1931, Bourret, 1942). This call is very effective because high-frequency
sounds are captured more easily among the low, loud rumble of the fast moving torrents.
The catl of the Speckled morphotype is unknown, though the distinct difference in vocal
sac morphology must result in a different vocalization. This, in tum, must result in
distinctive mate recognition patterns, as voicing is a key element in anuran mate
recognition (DueIlman and Trueb 1986, pp 87-107). If the Speckled morphotype are not
capable of a high-pitched call, it could indicate that its breeding sites are away from fast
moving water.
Size DimorpPzism. The Speckled form is also unique in that it exhibits the lowest
degree of sexual size dimorphism among the group (Table 2.10). It is unclear how this
relates to any behavioural changes from the remainder of the Rana Zivida group. One
theory is that it may impact breeding habits. Southeast Asian cascade ranids have marked
sexual dimorphism in size, with females attaining lengths that can be more than twice
those of their male counterparts (Boulenger 1920, Boumt 1942. Frost 1985 and citations
therein, Matsui and Matsui 1990). These species breed in torrents where females must
swim with ampIexing males on their backs. The burden of canying this extra load has
favoured larger females andor smaller males, with a trend for even greater size
dimorphism arnong Iarger females (Matsui 1994). Presumably, this is because larger
females experience greater drag in water. Any added burden, such as a male
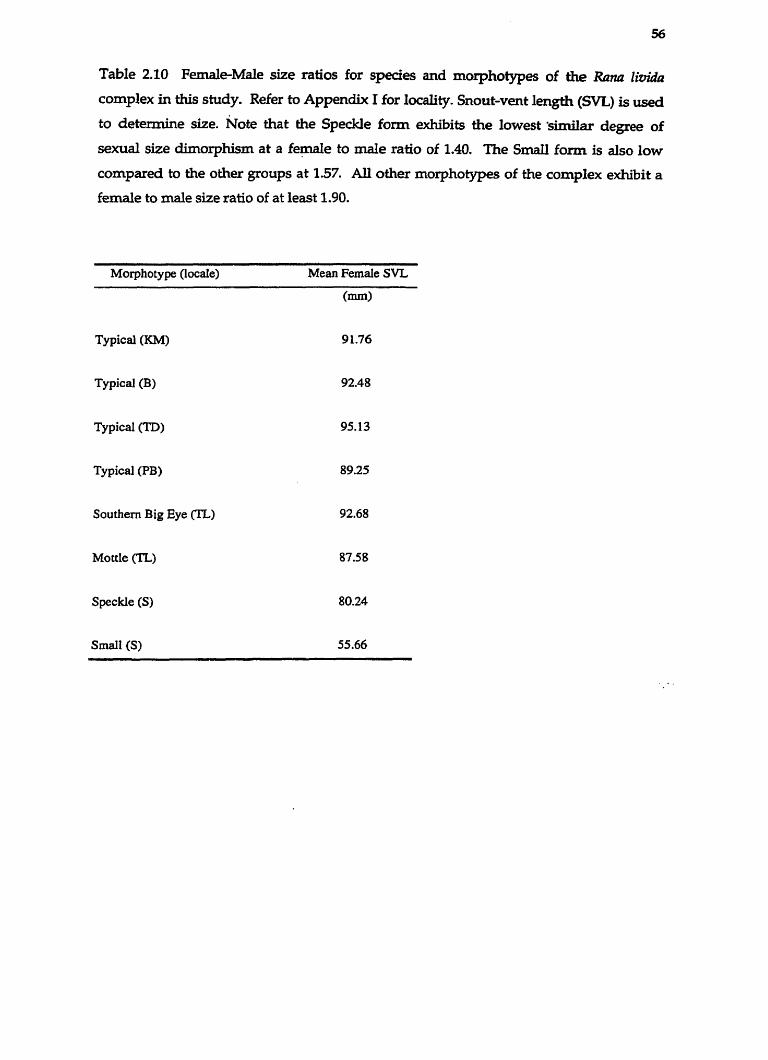
Table 2.10 FernaleMale size ratios for species and morphotypes of the Rrma l i e
compiex in this study. Refer to Appendix 1 for locaüty. Snout-vent length (ÇVL) is used
to determine size. Note that the Spedde form exhibits the lowest ~ i d a r degree of
sema1 size dimorphism at a f e d e to male ratio of 1.40. The S d form is also low
compared to the other groups at 1.57. AU other morphotypes of the complex exhibit a
fernale to male size ratio of at least 1.90.
Morphotype (locale) Mean Female SVL
Typicd (KM) 9 1.76
Typical (B) 92.48
Typical (TD) 95.13
Typicd (PB) 89.25
Southern Big Eye (TL) 92.68
Mottle (TL) 87.58
Speckle (S) 80.24
Smail (S) 55.66

passenger, will have a greater effect on their s w i d n g performance than on a femde
with a relatively srnaller body. Swirnming in torrents while in amplexus has been
O bserved arnong frogs of the Rana livida group (Pope 193 1, Bourret 1942, Liu 1950).
The low degree of sexual dimorphism in the Speckled fonn may suggest that they do not
breed in fast moving water. However, there have been no studies to determine a value of
intersexud size ratio that would be too low for effective swirnming in fast water, while in
amplexus. The possibility of niche segregation to reduce inter-sexual competition or
phylogenetic constraints are equally plausible explanations for sexual dimorphism in
body size.
Taxonomie implications of the Rana livida complex
The ehcidation of the Rana lividu complex both highlights and adds to the
historical taxonornic confusion. None of the new species match diagnoses of R.
leporipes, R. gramineu, or R. sinica and so these names remain junior synonyms of the
Rana livida complex. The complex also puts two of the higher level taxa, the subgenera
Eburana (Dubois 1992) and Odorrana (sensu Fei et al. 1990), into question.
Eburana
Dubois (1992) erected the subgenus Eburana based on three characters, including
the unpigmented eggs borne by the fernale. The classification has already been criticized
for its phenetic basis (Emerson and Berrigan 1993), its questionable characters ( Inger
1996), and its apparent inaccuracy (Matsui, et al. 1993, Matsui 1995). The discovery of
black eggs in the Black Egg species of the Rana livida cornplex adds further doubt to

validity of this subgenus. Regardless. the presence of pigmented eggs of the Black Egg
species reduces the number of known or consistent diagnostic characters to one: the
absence of nuptial spines on the chest of the males. This character is so widespread
among ranids that it has no diagnostic value at the subgeneric level. The Rana livida
group clearly does not belong to Eburana.
Odorrana
Fei et al. (1990) diagnosed R. livida and al1 species of Odorrana as having a
metastemal cartilage that is 'deeply notched posteriorly'. The Small species of the Rana
livida complex exhibits a sternal cartilage that is forked rather than notched (Figure2.7).
This is a condition found in Huia nasica, a member of the Amolops group of genera.
Amolops comprise another group of cascade ranids of southeast Asia (Inger 1966, Yang
199 1 b). This character presents a potential systematic problem, because sternal structures
are presently used to diagnose these groups.
There is a dose association of R. livida with the Amolops group of genera
(especially H. nasica). Although placed in a different genus, H. nasica shares an
overlapping range, habitat, and morphological similarities with members of the Rana
livida complex. Both taxa possess great1y expanded toe pads, white eggs, tremendous
size dimorphism, paired extemal vocal sacs, and a high chirp-like cal1 (Boulenger 1920, . .
Pope 193 1, Bourret 1942, ROM field notes unpub.). The adults are differentiated by the
prominent snout and brown colouration of H. nasica. The tadpole of H. nasica possesses
a sucker disk on its abdomen, which helps it to attach itself to the rocky bottoms of fast
rivers they inhabit. Unfortunately, tadpoles of the &na livida complex remain unknown.

This brings us to the problem of the forked sternum of the SrnaIl species. A bief
survey of the Rana Zivida complex show that there is an amay of metastemal f o m
(Figure 2.7). While differences seem clear at opposite ends of the spectmm, it is unclear
where the sternal element changes from king 'deeply notched' to 'forked'. Furthemore,
males of W. nasica exhibit the characteristic forked metastemum, whereas the females
exhibit a round metastemum that does not invaginate to form any kind of notch. The first
description of H. nasica to mention sternal structure refers only to males (Yang 199 lb).
A detailed study is beyond the scope of this project, as it would involve dissections of
specimens, and, therefore, possible type material. At this tirne it remains unclear as to
which genus the Small form of the Rana livida complex belongs, accepting current
generic limits. This highlights the need for a rigorous phyIogenetic analysis of the Rana
iivida compIex and the cascade ranids of southeast Asia. The new species will be
described, but no forma1 names will be given to them until this paper is published
(Appendix D).

Figure 2.7. %me metastenial elernents of m d e r s of the Rana livida complex and Huia
nasicn. Note how the condition of elements of the males and females of the same species
(or morphotype) are markedly different. The variation in the notch reveals a gradient of
shapes, rather than discrete d t s . This does not represent a comprehensive study of the
sternal elements of the Rana iivida complex.


Chapter 3
AUozyme Electrophoresis

Introduction
Odoriferous ranid frogs of the Rana livida complex exhibit evident differences
across their range. Members of the complex recently collected from Vietnam are grouped
into eight major rnorphological groups (Chapter II). Three of these groups are
morphologicalIy distinct species. The remaining five groups have characters that are
equivocal for species delimitation, so they are here referred to as morphotypes. A
morphometnc analysis using discriminant functions @FA) supported only four of the
five morphotypes. Because of the overlap in the morphotypes in the DFA and the
potentially labile nature of the characters that define them, 1 sought an independent
approach to further investigate species boundaries of the complex, allozyme
electrophoresis.(Richardson et al. 1986, Murphy et al. 1996). This method ailowed me to
further elucidate the species boundaries in frogs of the Rana livida complex.
Population aggregate analysis
Murphy et al. (1996) point out the advantages of using isozymes as diagnostic
markers for species boundaries (sensu Davis and Nixon 1992) in the fom of fixed allelic
differences. .Fixed differences not only imply the absence of gene flow, they are also
unique characters, which is evidence for lineage independence. Species boundaries can
thus be detected by studying fixed characters of populations in a population aggregate
analysis (Davis and Nixon 1992).

Fixed allelic differences arnong sympatric species indicate lineage independence.
The presence of two or more loci with fixed allelic differences among populations is
usually considered strong evidence of species boundaries (Richardson et al. 1986,
Murphy et al. 1996). Congruence between matornical and dozyme data provide
unequivocal evidence for species recognition. Populations that show no diagnostic
characters in the forrn of fixed alleles are considered conspecific.
Undersampling of specimens within populations can lead to over-estimation of
species diversity, because alleles may appear to be either fixed or absent. Currently a
minimum sample size of 3-5 for species delineation studies is standard (Richardson et al.
1986, Baverstock and Moritz 1996). An attempt was made to make the sample size for
each population of this study (Le. each morphotype per locality) at least five. Those
populations with a sample size that was less than three were considered to be identical to
allopatric members of the same morphotype.
Undersampling a species across its range can also lead to an overestimation of
species richness. Two distant populations may have no gene flow between thern and thus
exhibit fixed allelic differences. However, they may still interact with geographically
intermediate populations. Consequently, distant populations have different alleles each
with a very high frequency, and the populations in between carry both alleles at lower
frequencies, Failure to sample intermediate populations may result in two species being
recognized rather than a single polyrnorphic species. This is of particular importance to
the study of the Rana livida group, due to its large range. Sampling strategy helped takes
this factor into account, as populations of these frogs within Vietnam constitute a smdl
portion of each species range.

A priori species groupings
Traditionally, a p h r i groupings for electrophoresis consist of groups whose
diagnostic characters are equivocal for species delineation (Platz 1993, Green et al.
1997). While that method is applied in this analysis, 1 dso test the relative support of
these groups with cellular DNA content data. The use of DNA content in estimating
species assumes that conspecifics share equal levels of cellular D M (MacCulloch et al.
1996). Groups that show significantly different levels of DNA likely represent distinct
species. DNA content can be helpful in flagging cryptic species diversity, but it cannot
be used as a sole arbiter of species boundaries, as two different species may share the
same level of DNA content. When distinct groups also exhibit discrete levels of DNA a
species boundary is suggested. This evidence is even more compelling if the
morphotypes are sympatric. In this study, 1 expand upon the traditional methods of
electrophoretic analysis by establishing my a priori groups using mo~phology,
morphometrics, and DNA content. These groupings constitute a much stronger
hypothesis for testing species boundaries using isozymes.
Materials and Methods
Specimens of the Rana livida complex were collected by field crews of the Royal
Ontario Museum (ROM) from Hong Kong, People's Republic of China, and six localities
within Vietnam: Sa Pa (Lao Cai Province), Ba Be National Park (Cao Bang Province),
Tarn Dao (Vinh Phu Province), Pac Ban, Nah Hang Nature Reserve (Tuyen Quang
Province), the Khe Moi River, at Con Cuong ( Nghe An Province), and Tram Lap (Gia

Lai Province) (Appendix A). AI1 Vietnamese localities are northem, except Tram Lap,
which is in the Central Highlands. The species represented were the Black Egg, Smdl,
and Speckle species, defined in Chapter II (Appendix A). In addition, five morphotypes
were included in the study series, the Large form (Khe Moi), the northem Big Eye fonn
(Pac Ban), the southern Big Eye .fom (Tram Lap), the Mottled forrn (Tram Lap), and the
Typical form (Hong Kong, Ba Be, Tarn Dao, Pac Ban, and Khe Moi). Collections were
made at various times through the year from 1994-1998 (Table 2.1). Al1 frogs were
collected at night and euthanised within 18 hours using MS-222. Frogs were first fixed in
90% ethano1 and then preserved in 75% ethanol. Samples of blood, Iiver, leg muscle, and
heart were taken from some specimens. Blood samples were suspended in a freezing
solution (Vindelov et al. 1982, Murphy et al. 1997). Al1 tissue samples were stored in
liquid nitrogen in the field and subsequently stored at -80'~ at the ROM.
DNA Content
Cellular DNA content data were gathered from 1 10 specimens using flow
cytometry (Murphy et al. 1997) (Appendix A). Erythrocytes in whoIe blood were used in
the procedure. Blood samples were stained with propidium iodide. DNA content
analysis was carried out with a Becton Dickinson FACScan or a Becton Dickson
FACScalibur flow cytometer in the Faculty of Medicine, Department of Immunology,
University of Toronto between 1995 and 1997. Both systems are equipped with an argon
laser, operated at a wavelength of 488 nm at 15 mW. Fiuorescence at wavelengths
between 560 and 640 nm was collected and digitally transferred to a histogram of mean
DNA content concentration. Each sarnple was measured simultaneously with a known

standard (either Nerodia sipedon. or Bufo woodhousi fowlerî7. No samples were tested
on both fiow cytometers. DNA content data of morphotypes were compared using t-tests,
to determine whether or not they exhibited distinct levels of DNA from each other.
Protein Electrophoresis
Allozyme variation was examined for fixed allelic differences in 64 specimens of
R. livida cornplex frogs from 6 localities within Vietnam and 6 specimens from Hong
Kong (Appendix A). Al1 five morphotypes (Large form, Northern Big Eye form,
Southern Big Eye form, Mottled form, and the Typical form) were used as a priori
groupings in the electrophoretic study.
Three distinct species of the Rana livida cornplex (Black Egg species, Smdl
species, and Speckled species), as well as a population of HuiQ nasica (n = 5) from
Vietnam were inc1uded for cornparison. Sample sizes of each group (and each
population) varied according to specimen availability (Table 3.1). Sarnples of Iiver,
muscle, and heart were homogenized for use in starch gel electrophoresis. Al1 procedures
follow protocols of Murphy et al. (1996). Electrophoretic conditions were tested for
optimal resolution of 53 histochernicals on 11 buffer systems using a subset of the
samples (Table 3.2). Nomenclature follows Murphy and Crabtree (1985). Following this
pilot study, al1 electrophoresis was performed on the sarne lot of 11.5% hydrolyzed potato
starch (Connaught, lot #5 14-3).
Species boundaries of the samples were examined with a population aggregation
analysis, modified from Davis and Nixon (1992). Genotypes of individuals within the
same population (sympatric morphotypes) were surnmarized in a population profile.

Table 3.1 Morphotypes and comparative species investigated with allozyme
electrophoresls. Refer to Appendix 1 for locality abbreviations.
M o r ~ h o t ~ ~ e Locality Sample Species for Localities Sample Size Camparison Size
Typical form HK 6 Black Egg PB 4
Typical form B 5 Black Egg B 1
Typical form PB 4 B lack Egg TD 1
Typicai form KM 5 B lack Egg KM 5
Typical form TD 5 Small S 3
Large form KM 5 SpecMe S 3
Northern Big Eye PB 3 Huia nasica TD 5
Southern Big Eye TL 10

Table 3.2a Isozyme systems assayed. Abbreviations and enzyme commission numbers
(E.C.) are in parentheses.
Isozymes assayed AlcohoI dehydrogenase (ADH, E.C. 1.1.1.1) 6-Phophohctokinase (PFK, E.C. 2.7.1 1) Acid Phosphatase (ACP E.C. 3.1.3.2) Aconitate Hydratase (ACOH, E.C. 4.2.1.3) Adenosine Deaminase (ADA E.C. 3.5.4.4) Adenylate Kinase (AK E.C.2.7.4.3) Alanine Aminotransferase ( M T E.C. 2.6.1.2) Alanopine Dehydrogenase (ALPDH E.C. 1 S. 1.17) Alcohol Dehydrogenase (ADH, E.C. 1.1.1 .l) a-L-Arabinofuranosidase (ARAB E.C. 3.2.1.55) Aspartate Aminotransferase (AAT, E.C. 2.6.1.1) Catalase (CAT E.C. 1.1 1.1.6) Creatine Kinase (CK, E.C. 2.7.2.3) Dipeptidase-A (PEPA, E.C. 3.4.13.1 1) Dipeptidase-B (PEPB, E.C. 3.4.13.1 1) Fructose-biphosphate Aldolase (FBA, E.C. 4.1.2.13) Fumarase (FtJMH, E.C. 4.2.1.2) p- Galactosidase (P GAL E.C. 3.2.1 .23) General Proteins (GP) Glucose-6-phosphate Dehydrogenase (G6PDH E.C. 1.1.1.49) Glucose-6-phosphate isomerase (GPI, E.C. 5.3.1.9) p- Glucosidase (P -GLUS E.C. 3.2.1.21) N-AcetyI- P-Glucosaminidase (P-Ga E.C. 3.2.1.30) P-Glucuronidase (P -Glur E.C. 3.2.1.3 1) Glucose dehydrogenase (GCDH, E.C. 1.1.1.1 1 8) Glutamate dehydrogenase (GTDH, E.C. 1.4.1.3 Glutamate dehydrogenase (NADP) (GTDHP, E.C. 1.4.1.4) Glyceraldehyde-3-phosphate Dehydrogenase (GAPDH, E.C. 1.2.1.12) Gl ycerate Dehydrogenase (GLYDH, E.C. 1.1.1 .Z9) Glycerol-3-phosphate Dehydrogenase (G3PDH, E.C. 1.1.1.8) Guanylate Kinase (GUK, E.C. 2.7.4.8) Hexokinase (HK, E.C. 2.7.1.1) D-ZHydroxy-Acid Dehyârogenase (HADH E.C. 1.1.99.6) (S)-2-Hydroxy-Acid Oxidase (HAOX E.C. 1.1.3.15) A-Iditol Dehydrogenase (A-IDDH, E.C. 1.1.1.14) Isocitrate Dehydrogenase (IDH, E.C. 1.1.1.42) &Lactate Dehydrogenase (LDH, E.C. 1.1.1.27) MaIate Dehydrogenase (MDH, E.C. 1.1.1.37) Mannose-6-Phosphate Isomerase (MPI, E.C. 5.3.1.8) a-Mannosidase (a-MAN, E.C. 3.2.1.24) Oc tan01 Dehydrogenase (ODH, E.C. 1.1.1.73) Peptidase (PEPF, E.C. 3.4.-.-) Phosphoglucomutase (PGM, E.C. 5.4.2.2) Phosphogluconate Dehydrogenase (PGDH, E.C. 1.1.1.44,

Table 3.2a continued
Phosphoglycerate Kinase (PGK, E.C. 2.7.2.3) Proline Dipeptidase (PEPD, E.C. 3.4.13.9) Purine-Nucleoside Phosphorylase (PNP, E.C. 2.4.2.1) Pyroline-5-Carboxylate Dehydrogenase (PCDH, E.C. 1.5.1.12) Pyruvate Kinase (PK, E.C. 2.7.1.40) Succinate Dehydrogenase (SUDH, E.C. 1.3.99.1) Superoxide Dismutase (SOD, E.C. 1.15.1.1) Uridine Kinase (UK, E.C. 2.7.1.48) Xanthine Dehydrogenase (XHD, E.C. 1.1.1.204)

Table 3.2b Buffer systems used for aiiozyme elecbrophoresis. Abbreviatiom axe the
parentheses.
Buffer Systems
Borate continuous pH 8.6 (Sack)
Borate discontinuous pH 8.0 (B8)
Tris-Citrate Ii pH 8.0 (TC8)
Tris-Citrate IiI pH 7.0(WTC) .
Tris-HCI pH 8.2 (THC1)
Lithium-Boratems-Citrate pH 8.1 (LiOR)
Tris-CitrateBorate pH 8.7 (Plk)
Tris-Ci trate-EDTA pH 7.0 (TC-EDTA)
Tris-Borate-EDTA pH 8 .O EBT)
Amine-Citrate (Propanol) pH 7.5 (CT 7.5)
Amine-Citrate (Morpholine) pH 6.3 (CT 6.3)

Groups of populations were subsequently compared for fixed allelic differences.
The absence of fixed differences among populations indicated that the groups in question
were an aggregate of the same species. The presence of two or more fixed differences
between populations was indicative of distinct species. Populations with a sampk size of
one or two specimens were included with allopatric populations of their own
morphotypes, regardless of their allelic profiles.
Alï specimens were also evaluated using a variety of genetic distance and
similarity dgorithrns: minimum distance (Nei 1972), unbiased minimum distance (Nei
1978), genetic distance (Rogers 1972, and genetic similarity (Rogers 1972). An
unweighted paired group analysis (UPGMA) phenogram was generated as a visual aid for
understanding these distances. The matrices and phenogram were generated with the
program BIOSYS ver 1.7 (Swofford and Selander 198 1).
Results
DNA Content
While there were 1 10 specimens analyzed for DNA content, only sympatric
morphotypes were analyzed with the t-test resulting in 73 specimens being used for
cornparisons (Appendix A). In Sa Pa, the Speckled species had a mean DNA content
level of 14.86 pg (f 0.50), whereas the Small species had a level of 13.26 pg (f 0.42).
These two differed significantIy (t=8.40, p< 0.0001). In Pac Ban, the Typical form had a
level of 13.05 pg (f 0.54) and the Northern Big-eye

form had a level of 14.43 pg (f 0.06). These two also differed significantly (t = 3.59,pq
0.007). In the Khe Moi River, the Typical form had 12.45 pg (f 0.33) of DNA per cell,
whereas the sympatric Large form had 13.98 pg (k 0.14). These two forms differed
significantly (t= 13.00, p < 0.0001). The two fonns from the Central Highlands also had
distinct levels, with the Green form exhibiting 12.10 pgfcell (f 0.1 l), and the Southem
Big-Eye form showing levels of 13.02 pgkell (k 0.10). These two differed significantly
from each other (t = 7.02, p < 0.0007). T-tests indicated that al1 sympatric morphotypes
exhibited significantly distinct levels of DNA from each other (TabIe 3.3). Consequently,
congruence of DNA content with morphotypes is indicative of their strength as a priori
groups for electrophoretic studies.
Protein Electrophoresis
Fifteen presumptive enzyme loci were resolved on 6 buffer systems (Table 3.4).
Two of these, Ak-A and Gtdh-A, were monomorphic in aII samples and populations. The
13 remaining loci were polyrnorphic and they provided evidence of differentiation at the
species level for three of the morphotypes: the Typical form, the Southem Big Eye form,
and the Mottled fonn (Table 3.5). Although the allozyme data provided substantial
evidence for the recognition of new species, the sampIe sizes were too low to make
inferences about population substructure. The complete results of the population
aggregate analysis can be found in Appendix C,

Table 3.3 T -test of DNA contents of sympatric morphoSrpes. AU t-statiçtics are
significant, indicating that each of the groups compared have distinct ievels of DNA
from each other.
Locale Morphotype Sarnple Size Mean DNA Content t-statistic (n) (pg) P (.995)
Sa Pa Spec kled 7 S m d 8
Pac Ban Typical 6 n Big Eye 3
Khe Moi Typical Large
Tram Lap Mottled 14 12.1 s Big Eye 21 13.02 7.02 0.0006

Table 3.4 Allozyme frequenciesat aii resolved presumptive loci of the Rana livida
cornplex. Lociality data appear below morphotype/species name with çample sizes in
parentheses. Refer to Appendix 1 for locality abbxeviations. Enzyme systems are listed
in the Ieft-hand column, with the buffers used to resolve them in parentheses. Enzyme
and buffer abbreviations are referred to in Table 3.2a and 3.2b respectively. Alleles are
indicated with lower case letters. Missing data is indicated by a backslash (/).


s V
A : .

Table 3.5 Matrix of fixed allelic differences among species and morphotypes of the
Rana Zivida cornplex..
Typicai Large nBig Eye sBig Eye Mottled Smail Speckied Black Egg H. nasica
Typical da
Large 2 d a
nBig Eye 2 1 d a
sBig Eye 2 5 4 d a
Monled 2 5 7 7 d a
Srnall 5 8 8 6 7 da
Speckled 5 4 4 6 6 10 d a
BlackEgg 4 5 5 5 5 7 6 d a
H. nasica 5 6 6 6 7 5 5 5 d a

Population Aggregate Analysis
Within-group genotypes
Two of the groups were sarnpled from multiple populations: the Typical fom had
five populations, and the Black Egged species had four (Appendix C). Neither of these
groups exhibited fixed differences among their populations. Sample sizes of the Black
Egg populations from Pac Ban and Tam Dao were too small (n = 1) to be used in the
population aggregate analysis. Any unique alleles that these two populations exhibited
were ignored.
Among-group genotypes in sympatry
Fixed allelic differences in eight of the loci (mAat-A, Ck-C, Gtdh-A, Ldh-B,
rnMdh-A, SM&-A, Pgm-A, Fumh-A) diflerentiated 5 of the a prion' morphotypes from
everything eIse (Table 3.5). The SmaIl morph from Sa Pa exhibited fixed dlelic
differences at three loci, rnAat-
A, mMdh-A, and sMdh-A. The Southem Big Eye form exhibited differences at
Ck-C and Pgm-A. The Speckled form from Sa Pa had one difference at Gtdh-A, as did
the Large form from the Khe Moi River at Fumh-A. As well, the Black Egg species had
a fixed aiIele at Ldh-B.
Many fixed differences corresponded with the a priori morphological groups. The
two forms found in Sa Pa, the Small and Speckled species exhibited fixed allelic
differences at 10 loci, mAat-A, Ck-C, Gpi-A, Gtdh-A, micdh-A, Ldh-A, Ldh-B, mMdh-
A, sMdh-A, and Fumh-A. The two foms found in the south, Southem Big Eye and
Mottled, were also distinct, in having seven loci with fixed differences, Ck-C, Ldh-B,

SM&-A, Pep-A, Pgm-A, sSod-Al, and Fumh-A. Huia nasica and the Typical form at
Tarn Dao exhibited six fixed differences between hem: rnAat-A, Ck-C, rnIcdh-A, Ldh-A,
sSod-A, Fufnh-A. The Northem Big Eye form and the sympatric Typical population from
Pac Ban exhibited fixed differences at four loci, mAat-A, Ck-C, Ldh-B, sSod-A, and
Fumh-A.
The Khe Moi River area has three sympatric morphotypes: The Typical form, the
Large f o m and the Black Egg species. All three could be distinguished from each other
with alternate fixed alleles at Fumh-A. The Big form was aIso discernible with a fixed
difference at sSod-A, while the Black Egg species was distinct at three loci: Ldh-B, Ck-
C, and mAat-A.
Arnong-group genovpes in allopatry
Al1 allopatric groups exhibited at least one fixed allelic difference from each other
(Table 3.5). Two pairwise allopatric cornparisons appeared to be particularly important:
that between the northem versus Southern Big Eye forms, and Northem Big Eye fonn
versus Large form. The two Big Eye forms exhibited four fixed allelic differences, Ck-C,
Gpi-A, Pgrn-A, and sSod-A. In contrast, only one fixed alIeIic difference (Fumh-A)
occurs between the Large form and the northern Big Eye form.
Phenetic analyses
Phenetic analyses were canied out on those populations of the Rana livida
complex that showed resolution of al1 15 loci in the allozyme electrophoretic study.
Because the s-SOD-a locus of the Srnall species could not be resolved in this study, and a

technical problem (absence of a histochemical) prevented the resolution of the Ak-a locus
in three of the populations of the Black Egg species (Ba Be, Tarn Dao, and Pac Ban),
these populations were omitted from the phenetic analyses.
A matrix showing two measures of genetic distance, as established by minimum
genetic distance (Nei 1972) and the unbiased minimum distance (Nei 1978) (Table 3.6)
generally reflects the population aggregate analysis numbers of fixed allelic differences
among groups (Appendix C). The genetic distance between two known species was
consistently higher than .3 10 for the minimum distance, and -209 for the unbiased
rneasure. Populations of the Typical form al1 exhibited distances no greater than ,060 for
the minimum distance and .O75 for the unbiased distance.
A matrix showing another measure of genetic distance (Rogers 1972) and genetic
similarity (Rogers 1972) reflects the results of the fixed allelic difference matrix and the
Nei's distances (Table 3.7). The genetic distance between two known species was never
below .330 and the similarity was never below .460. Populations of the Typical form
exhibited differences no greater than .l8O and similarities no lower than 0.860. The
UPGMA phenogram was generated by using Rogers genetic distance (1972) (Figure 3.1).
It shows the Hong Kong, Pac Ban and Tarn Dao populations of the Typical f o m as being
most similar to each other. The Khe Moi Typical population is the most different from
the remainder of this morphotype. The Large form and the Northern Big Eye form are
aIso similar to each other.

Table 3.6 Genetic distance measures of members of the Rana livida complex. Minimum
genetic distance measures (Nei 1972) are shown below the diagonal, while unbiased
muUmum genetic distance measures (Nei 1978) are shown above the diagonal.

Population
Typical B
Typical HK
Typical PB .
Typical TD
Typical KM
Mottle TL
Specide S
BlackEgg KM
Large KM
nBigEye PB
sBigEye TL
H nasica TD
TY PHK
0.001
XXX
0.004
0.003
0.069
0.327
0.333
0.4
0.2 18
0.27 I
0.272
0.443
TY PTD
0.002
0.00 1
O
XXX
0.06 1
0.316
0.336
0.398
0.22
0.274
0.26
0.435
TYPKM
0.054
0.058
0.047
0.048
XXX
0,221
0.339
0.353
O. 1 88
0.238
0.27 1
0.4
Mott TL
0.314
0.326
. 0.312
0.315
0.209
XXX
0.46
0.602
0.378
0.464
0.446
0.485
Speck S BlkEgg KM Large KM
0.334
0.333
0.336
0,335
0.327
0.46
XXX
0.467
0.284
0.27 1
0.439
0.4 18
0.37 1
0.394
0.39
0.39
0.335
0.595
0.461
XXX
0.44
0.404
0.4 1
0.311
0.208
0.21 1
0.2 14
0.213
0.169
0.37 1
0.278
0.427
XXX
0.093
0.337
0.425
nEye PB
0.258
0.269
0.27 1
0.27 1
0.225
0.462
0.269
0.396
0.085
XXX
0.287
0.386
sEye TL
0.242
0.269
0.253
0.256
0.256
0.442
0.436
0.401
0.327
0.282
XXX
0.398
H nas TI)
0,423
0.433
0.422
0.424
0.378
0.474
0.408
0.295
0.408
0.374
0.385
XXX

Table 3.7 Genetic distance measures of members of the Rana li.oida cornplex. Genetic
distance (Rogers 1972) is shown below the diagonal, whte genetic similarity (Roger
1972) is shown above the diagonal.

Population
Typical B
Typical NK
Typical PB
Typical TD
Typical KM
Mottle TL
Speckle S
BlackEgg KM
Large KM
nBigEye PB
sBigEye TL
H m i c a TD
TY PB
XXX
0.048
0.065
0.062
O. 18
0.365
0.382
0.448
0.29 1
0.302
0.308
OS 14
TYPHK
0.952
XXX
0.017
0.013
0.154
0.33
0.333
0.442
0.253
0.283
0.312
0.493
TY PPB
0.935
0.983
XXX
0.003
O. 137
0.323
0.35
0.439
0.269
0.3
0.304
0.486
TYPTD
0.938
0.987
0.997
XXX
O. 14
0.324
0.347
0.438
0.266
0.297
0.305
0.487
TYPKM
0.82
0.846
0.863
0.86
XXX
0.292
0.408
0.44
0.277
0.336
0.363
0.497
Mott TL
0.635
0.67
0.677
0.676
0.708
XXX
0.463
0.63 1
0.4 16
0.48
0.482
0.538
Speck S
0.618
0.667
0.65
0.653
0.592
0.537
XXX
0.509
0.319
0.283
472
0.477
BlkEgg KM Large KM
0.552
0.558
0.561
0.562
0.56
0.369
0.49 1
XXX
0.509
0.459
0.475
0.394
0.709
0.747
0.73 1
0.734
0.723
0.584
0.681
0.49 1
XXX
O. 149
0.396
0.496
nEye PB
0.698
0.7 17
0.7
0.703
0.664
0.52
0.7 17
0.54 1
0.85 1
XXX
0.317
0.434
sEye TL
0.486
0.688
0.696
0.695
0.637
0.5 18
0.528
0.525
0,604
0.683
XXX '
0.46
H nas TD
0.423
0.507
0.5 14
O S 13
0.503
0.462
0.523
0.606
0.504
0.566
0.54
XXX

Figure 3.1 Unweighted paired group analysis (UPGMA) phenogram of members of the
Rana livùia complex based on Rogers genetic distance (1972). This visual representation
of the distance ma& shows that populations of the Typical form are the most similar to
each other.

TYP KM Large nEye sEye Speckle Mottle Black H. nasica

Discussion
DNA content analysis
Variation among DNA content data constitutes a potential source of error. The
greatest source of variation among results of DNA content analysis cornes from the daiiy
operation of the flow cytometer (Fisher et al. 1994). These variations are due both to the
subtle fluctuations of mechanicd parameters (such as the laser intensity and alignment, or
the flow of the sample through the cytometer), and prepatatory staining parameters (such
as the staining time or concentration relative to the size of the sample). DNA content for
this study was analyzed over three years using two flow cytometers. No sarnples were
m n on both cytometers, over different times, thus accuracy cannot be guaranteed. For
this reason, only those samples analyzed in the sarne run were compared with each other.
This resulted in the sympatric sarnples collected in the same year being compared. The
cost of this precaution is that DNA content data cannot be used with allopatric
populations.
An open question
Two of the morphotypes (the Northern Big Eye forrn and the Large form) are
distinct from the rest of the group, each exhibiting at least two fixed differences from al1
other rnembers of the cornplex. Several factors rnake it unclear whether or not they are
distinct from each other.
First, the dl-male series of the northern Big Eye form and the dl-femde series of
the Large form are separated by only one fixed allele (Table 3.5). While this minor level

of separation suggests population differentiation, it is not strong enough evidence to
recognize a species boundary. As well, the close similarity of the two groups is exhibited
by the small genetic distance (D) (Table3.6 , Figure 3.1). The genetic distance between
them is most comparable to those found arnong populations of the Typical fonn. Thus,
these populations appear to be conspecific. This evidence canot be used to delineate
species, because there is no threshold distance above which one can confidently proclaim
a species boundary, and below which one can claim conspecificity. Furthemore,
distance measures can be increased or decreased according to the number of alleles
included in the study. The locus number and the locus selection heavily influence the
distance measure, making it a largely arbitrary measure. Using genetic distances as an
arbiter of species boundaries is a phenetic method, and is, therefore, not consistent with
the evolutionary species concept followed in this study.
Two factors make it particularly difficult to use morphology to distinguish
between these two morphotypes: the small sarnple size (n=3) of the Big Eye forrn, and
the single sex sarnpling of each population. Owing to the various leve1s of sexual
dimorphism within the Rana livida cornplex, morphological cornparisons between the
Large and Northem Big Eye forms are of little help. Because the two populations are not
sympatric, cellular DNA content data can neither support nor refute the hypothesis that
the two forms represent distinct species. It is likely that these two morphotypes represent
males and females of the same species. The presence of a single fixed difference
suggests that a species boundary between these two groups must be left as an open
question, but that a conservative estimate, that of one species being present, be adopted
until further studies can be done;

We need specimens of Northern Big Eye fom females and Large fom males to
determine whether or not a species boundary exists between them. In the absence of
these specimens, a phylogenetic analysis of the Ranu livida group would help resolve
their status. If Northern Big Eye male specimens and Large female specimens are not
monophyletic with respect to each other, then they can be considered separate species. A
phylogeny of the group could also be used to test the species boundaries proposed here.
It would be expected that populations of the species would be monophyletic.
My morphological and allozyme data are insuficient for a phylogenetic
reconstruction. Coding the morphological data would yield a matrix that contained only
plesiomorphies or autapomorphies, al1 phylogenetically uninformative characters. A tree
based on allozymes is also problematic. In order to resolve al1 of the nodes of a tree
using allozymes, it is usually necessary to have three times as many loci as there are taxa
(Murphy et al. 1996). That is not the case here, as there are only 15 prospective loci for
as many as eight taxa. Even if this guideline is ignored, the aIlozyme data from this study
yields six potentially informative characters, which lack significant structure (Table 3.7).
A permutation tail probability (Faith and Cranston 1991) resulted in an insignificant
value (p = 0.460,999 replicates). Random trees analysis generated 45 trees that had
fewer branch lengths than the most parsimonious set of trees and a gl tree distribution
statistic (Hillis and Hulsenbeck, 1992) of 0.399. These evaluations al1 suggest that the
data do not differ from random, and thus are not suitable for phylogenetic analyses. DNA
sequences might be the best alternate choice for phylogenetic analysis of this group,
owing to the large potential database they offer (Hillis et al 1996).

Table 3.8 Phylogenetic codings based on allozyme data of the Ram livida complex.
Enzyme systems are shown on the left (refer to Table 3.2a for abbreviations). AU spdes
of the Ram lioida complex, as weU as H. Mnca and a hypothetical ancktor (HA) are
included. The Large and northern Big Eye forms, while considered to be a single
species were included as separate taxa. Aileles present are shown as lower case letters,
while the phylogenetic codulgs are shown in numbers below the alleles. Ordered
characters are indicated with (ord) at the end of the row, while unordered characters are
indicated with (u/o) at the end of the row.
HA Small Typ Mottie Large Neye SEye Speckled BIack H.
Ck-C cc abd a ad d b O O 3 1 2 2 3 d o
ord Gpi-A bfg ab dc a a db
O O 1 2 1 1 2 adf adf 2 2
ord
ord
d o
d o
d o
ord .
ord
d o
Mh-B a acd b d d ad O O 1 2 3 3 3
Mme a abc a a ab abd O O 2 O 0 2 2
abc a 1 O

Taxonomie Iniplications
ResuIts of the anatomical and allozyme investigations established that &na livida
is a complex of at least seven species. Ln naming new species of this complex, one would
have to designate one species as Rana Zivida. The type locality of Rana livida is
unknown, given as Tenassarium, Myanrnar, in Blyth's original description (1855). The
two Dajeeling specimens used in this study are the close representatives from the type
region. These are both of the Typical fonn, which is known to occur in northern Vietnam
and southeast China (Chapter 12). It is reasonable to assume that the Typical form is what
was originally described as R. Zivida, and that it inhabits a large portion of the range of
the Rana livida complex. Though the new species are characterized here (Appendix D), 1
will not assign forma1 names of any Runa livida complex frog until 1 publish formal
descriptions.
hnplications beyond this study
The elucidation of a species complex, where for 150 years it was believed only to
be one species highlights our current underestimation of species diversity in southeast
Asia, particularly Vietnam. Members of the Rana livida complex, as currently
understood, have been studied in detail from a small area within their geographic range,
suggesting that more undetected species are present. The confusing taxonomic history of
other cascade ranids (particularly members of Odorma , sensu Fei et al.) suggests that
these species also need to be carefully re-evaluated. This requires a rigorous taxonomic
approach, with a clear definition an acceptable diagnosis of species boundaries. The

integrated approach exemplified by this study is necessary to te& out cryptic diversity.
This study also exemplifies the need for the further biological understanding of the
complex. Little is known about the ecology or the behaviour of the group. This
information would surely be helpfil in species delineation, because al1 of these species
occupy the same habitat. Thus, they rnust occupy different ecological niches.
My study also highlights the need to understand the evolutionary history of the
cascade ranids. No phylogeny of the group bas been proposed, and so it is unclear
whether or not any of the generic or subgeneric groupings used today are valid. Species
identification and elucidation of groups of species is the necessary first step, and their
independent evolutionary history can be evident through their morphology, ethology,
ecology, and genetics. These, in turn, cm be used to understand their phylogenetic
relationships.

Chapter 4
Summary

Species recognition of the green cascade fiog has been problematic since it was
first described by Blyth (1855). 1 sought a multi-faceted approach to species recognition,
in order to detexmine whether or not R. Zivida was polytypic. My combined evidence
from morphology (Chapter II), morphometrics (Chapter Ii), DNA content (Chapter III),
and allozyrne electrophoresis (Chapter Ill) indicate that Rana Zivida is a species complex
(polytypic species). Among the specimens studied in this investigation of Rana Zivida
from Vietnam, no Iess than seven distinct species could be discerned. Species 1, the
Typical fom is found throughout the southeast Asian range of the group. Species 2, the
Black Egg form is easily distinguishable, since al1 other species of the complex bear
white eggs. Species 3, the Speckled form, is also distinguished by its heavy granulation
and vocal slits. The rest of the complex exhibits smooth or shagreened skin and males
have paired extemal subgular vocal sacs. Species 4, the Smdl form, is morphologicaHy
distinct due to its small size, white dorsolateral granules, forked metasternum, and
absence of vomenne teeth. Species 5, the Mottled form, is visually disfinguishable by its
ability to change its day and night colouration. Species 6, the southem Big Eye form,
though morphologically very sirnilar to the Mottle form, is distinct by the larger size of
the males and its bulging eye. Species 7, the Large form, is distinguished from al1 other . -
species by its brown colouration. It may be cornprised of two species, pending further
investigation.

Implications of the study
Taxonomie
The elucidation of the Rana livida complex both adds to and resolves some of the
historicd taxonomie confision. The pigmented eggs of the Black Egg form certainly
makes the presence of Rana livida within the Eburana (Dubois 1992) inappropriate, as
one of the three diagnostic characters of this subgenus is white eggs. The forked
metastemm of the Small form puts to question the validity of the metasternum as a
diagnostic character of Odorram , a subgenus in which Rana livida has also been placed.
These characters, as well as the vomerine teeth and dorsolateral granuIation of the Smail
species, and the vocal slits of the Speckled species, puts to question the monophyletic
nature of the complex. This highlights the need for a phylogenetic analysis.
Seven new species of the Rana livida complex were detected within a few
localities in Vietnam. This represents a very small area of the southeast Asian range of
the complex and suggests that there are yet undetected and undiscovered species. This is
also likely tme for other cascade ranids of the region, since they share two of the sarne
features as the Rana livida comp.lex: a large distribution and great morphological
similarity.

References
Ahl, E. (1925). "Ueber vernachlibsigte Merkmale bei Froschen." Sitzuwsberiçbte der Gesellschaft naturforschender Freunde ni Be& 40-45.
Anderson, 1. (1993). "Keep taking the frog skin." New Scientist 140: 10.
Anderson, J. (1871). "On some Indian Reptiles." Proceediw of the Zoologid Society of Londo~ Feb 21,1871: 208.
Baverstock, P. R. and C. Moritz (1996). Project Design. in Molecular Systematics. D. M. Hillis, C. Moritz and B. K. Mable. Sunderland, Mass., Sinauer and Associates. 2nd: 655.
Blyth, E. (1852). "Proceeding of the Society for April." Journal of the Asiatic Socieh . . Benval 21: 35 1-356.
Blyth , E. (1855). Journal of the Asiatic Society. BengaIZS(7): 732.
Boulenger, G. A. (1882). Catalogue of the Batrachia. Salentia S. Ecaudata in @ Collection of the British Museum. London, England, Trustees of the British Museum.
iles and batrach Boulenger, G. A. (1 887). An account of the r e ~ t ians obtained iq Tenasserium by M.L. Feae. of the Genoa Civic Museum.
Boulenger, G. A. (1890). Fauna of British India. includirg Cevlon and Buma; Re~tiiia and Batrac hia. London, Taylor and Francis.
Boulenger, G. A. (1899). "On the reptiles, batrachians, and fishes collected by the late Mr. John Whitehead in the interior of Hainan." Proceedines of the Zoolo~ical Society of London: 159- 172.
Boulenger, G. A. (1920). "A monograph of the south Asian, Papuan, Melanesian, and Australasian frogs of the genus Rana." Rec_ords 20: 1 - 126.
Bourret, R. (1942). Les Batrachians de L'Indochine. Hanoi, Vietnam, Gouvernment general de L'Indochine, Hanoi, Vietnam.
Coghlan, A. (1996). "Peptides punch it out with superbugs." New Scientist 150: 20.
Daly, 6. W., M. Garraffo, T.F. Spande, C. JaramilIo, A.S. Rand (1984). "Dieta. source for skin aikaioids from neotropical poison dart frogs (Dendrobatidae)?" Journal of Chemical Ecoloq 20(4): 943-955.

Daly, J. W., S. 1. Secunda, H.M. G-0, T.F. Spande, A. Wisnieski, J.F. Cover Jr. (1994). "An uptake systern for dietary alkaloids in poison frogs (Dendrobatidae)." Toxicoq 6: 657-663.
Davis, J. 1. and K. C. Nixon (1992). "Populations, genetic variation, and the delimitation . . of phylogenetic species." SYslte.matic 41(4): 42 1-435.
Dubois, A. (1992). "Notes sur la classification des Ranidae (Amphibiens: Anoures)." Bulletin Mensuel de la Societe b e . Par& 61(10): 305-352.
Duellman, W. (1 993). . Lawrence, KA, AIlen Press.
Dueiiman, W. and L. Trueb (1986). Biology of the Aruphibians . . , Johns Hopkins Press, Baltimore, Md.
Emerson, S. and D. Bemgan (1993). "Systernatics of southeast Asian ranids: multiple origins of voicelessness in the subgenus Limnonectes (Fitzinger)." Herpetoloeic~(22-3 1).
Faith, D. P. and P. S. Cranston (1991). "Could a cladogram this short have arisen by chance alone? On permutation tests for cladistic structure." Cladistics 7: 1-28.
Fei, L., C. Ye, et al. (1990). Keys to chinese Arnmibians . . . Chongqing, Chongqing
Branch, Publishing House of Science and Technology Materials.
Fellows, J. R. and Chi-hang, Hau (1997). A Faunal Srirvev of Nine Forest Reserves in Tro~ical South China. with a Review of Conservation Priorities in the R e m . Hong Kong, China, Kadoorie Farm and Botanic Garden, Hong Kong.
Fisher, S. K., C. E. Dallas, et al. (1994). "Sources of error associated with sample collection and preparation of nucleated blood cells for flow cytometric analyses." Çellulai: Biolo&&&&&gy 10: 145-153.
Frost, D.R. (1985). Amphibian Soec . . ies of the World. Lawrence, KA, Allen Press.
Frost, D. R. and D. M. Hillis (1990). "Species in Concept and Practice." Herpetologica 46( 1): 87- 104.
Frost, D. R. and A. G. Kluge (1994). "A consideration of epistemology in systernatic biology, with special reference to species." Wistic_s lO(2): 259-294.
Green, D., H. Kaiser, T.F. Sharbel, J. Kearsley, KR. McAllister (1997). "Cryptic species of spotted frogs, Rana pretiosa cornplex, in western North Arnerica" Copeia 1997(1): 1- 8.

Gunther, A. (1875). "Third report on the collections of Indian amphibians and reptiles obtained by the British Museum." R o c e e d i ~ of the Zoologjcal Society of Londqn: 567- 577, pls. Lm-LXVL
Hair, J. F., R. E. Anderson, et al. (1995). Multivariate Data A-. Englewood Cliffs, NJ, Prentice Hall.
Hillis, D. M. and J. P. Hulsenbeck (1992). "Signal, noise, and reliability in molecular phylogenetic analysis." Journal of Heredity 83(l89- 195).
Hillis, D. M., B. K. Mable, et al. (1996). Application of rnolecular systematics. Moleculq Svstematic~. D. M. Hillis, B. K. Mable and C. Moritz. Sunderland. MA, Sinauer and Associates: 5 15-543.
Hunton, N. K. (1996). "Al1 natural A I D S protection?"echnology Review 99: 18-24.
Inger, R. F. (1966). "Systematics and Zoogeography of the Arnphibia of Bomeo." Pieldiana: Zoolo~y 52(November, 1966): 188- 19 1,364.
Inger, R. F. (1 996). "Commentary on a proposed classification of the family Ranidae." H e m e t o l o a 52(2): 24 1 -246.
. . Karsen, S. J., M. W.-n. Lau, et al. (1986). H o n ~ K o n ~ Am~hibians and Reptiles. Hong Kong, Urban Council.
Lazell, J. D., Jr., 1;. Goodyear, et al. (1988). "Rana livida (green cascade frog) density." erpetolo~ical Review 19(2): 36.
Li, W.-H. and Z.-Y. Wang (1985). "Kary otypoe of Rana livida." Acta Hepetoloaical . . 4(1): 56. English
Linnaeus, C. (1758). Svstema Naturæ Fer Regna Tria Naturæ. Secundum Classes, Ordines. Gga
. . era, Sûecies. cum Characte fferentixs- Svnonvm Decima. Holmiae (=Stockholm), Laurenti Salvi.
Liu, C. (1950). Am~hibians of Western Chinq. Chicago.
Liu, C., and S. H u (1961). Çhinese T-ss m h i b i a n s . . . Peking, Science Press.
MacCulloch, R. D., D. E. Upton, R.W. Murphy (1996). "Trends in nuclear DNA content among amphibians and reptiles." m a r a t i v e Biochemistrv and P- 113B: 601- 605.

Matsui, M. (1994). "A taxonornic study of the Rana narina complex, with description of three new species (AmphibirRanidae)." Zoologbal Journal of the Lm Socieu 11: 385415.
Matsui, M., Hidetoshi Ota, Michael Lau, and Anthony Bogadek (1995). "Cytotaxonomic studies of three ranid species (Amphibia:Anura) from Hong Kong." &panese Journal of Brpetology 16(i): 12- 17.
Mayden, R., L. and R. M. Wood (1995). Svste&cs. s~ecies conce~ts. and the m i f i c a n t unit in biodiversity and on biology. Amencan
Fisheries Society Symposium, American Fisheries Society.
Mayden, R. L. (1997). A hierarchy of species concepts: the denouement in the saga of the . . species problem. S~ecies: The Units of Biodiversia. M. F. CIaridge, H. A. Dawah and M. R. Wilson, Chapman and Hail.
Murphy, R. W. and C. B. Crabtree (1985). "Evolutionary aspects of isozyme patterns, number of loci, and tissue-specific gene expression in the prairie rattlesnake, Crutalus virais viridis." I.l[erpetolo~ica 41: 45 1-470.
Murphy, R. W. and K. D. Doyle (1998). "Phylophenetics: frequencies and polymorphic characters in genealogical estimation." Svsternatic Biolo
Murphy, R. W., L. A. Lowcock, C. Smith, I.S.Darevsky, N.I. Orlov, R.D. Upton, D.E. Upton (1997). 'Flow cytometry in biodiversity surveys: methods, utility, and . . constraints." A&@~ibta-Reptilia 18(1): 1 - 13.
Murphy, R. W., J. W. Sites Jr., D.G. Buth, CH. Haufier (1996). Proteins: Isozyme electrophoresis. MolecuIar Systemati~. D. M. Hillis, C. Moritz and B. K. Mable. Sunderland, Sinauer Associates: 4 1 - 120.
Nei, M. (1972). "Genetic distance between populations." h e r i c a n N a t u r u 106: 283- 292.
Nei, M. (1978). "Estimation of average heterozygosity and genetic distance from a small number of individuals." Genetics 89: 583-590.
Nixon, K. C. and Q. D. Wheeler (1990). "An amplification of the phylogenetic species concept." Cladistics 6: 21 1-223.
Platz, W. B. (1993). "Rana subaquavocalis, a remarkable new species of leopard kog (Rana pipens complex) from southeastern Arizona that calls under water." Journal of fiemetoloa 27(2): 154- 162.
Pope, C. H. (193 1). "Notes on amphibians from Fukien, Hainan, and other parts of China." Bull. Amer. Mus. Nat. Hist, 61(8): 397-61 1, 10 plates, folding table.

Richardson, B. J., P. Baverstock, R., et al. (1986). Allozvme electr for a-1 svstmtics and population s tudie~ North Ride, NSW, A-ayd
esis: A
Press.
Rogers, J. S. (1972). "Measures of genetic similarity and genetic distance." Stud~es . . 7213: 145-153.
Smith, M. A. (192 1). "New or Me-known reptiles and batrachians from southern Annam (Indo-China)." P r o c e e d i ~ of the ZooIoeical Society of J.ondon: 423-440, pls. 1-II, figs. 1-2,
Smith, M. A. (193 1). The Fauna of Bnti& India Including: Cevlon and Burma: . . . . ewt-ia. Volume 1. London, Taylor and Francis.
Statsoft (1996). Statistica ver. 5.0. Tulsa, OK.
Swofford, D. and R. B. Selander (1981). BIOSYS 1.7: A cornputer program for the analysis of allelic variation in genetics. Urbana-Champaign, University of Illinois.
~abachnick, B. and L. S. Fidell(1989). IJ&g Multivanate Stat . . istics. New York, Harper
and Row.
Taylor, E. H. (1962). 'The amphibian fauna of Thailand." IJniv. Kansas Sci. Bulk 4318): 265-599, errata.
Toledo, R., C. and C. Jared (1995). "Cutaneous granular glands and ambhibian venorns." Comparative B i o c w and Phvsialogv 111(1): 1-29.
Tschudi, J. J. v. (1 838). "Classification der Batrachier, mit Berucksichtigung der Fossilen Thiere dieser Abtheilung der Reptilien." petitpierre. Neuchâtel, 99: (2) pages, 6 plates with page of explanation.
Tyler, M. (1996). New Drugs from Old Frog Skins. Kevnote Address. Society for the Study of Amphibians and ~ e ~ t i l e s , 39th Annual Meeting, Lawrence, Kansas.
Valigra, L. (1994). "Engineering the future of antibiotics." New Scientist 142: 25-27.
Vindelov, L., 1. J. Christensen, et al. (1982). "Long term storage of sarnples for flow cytometric DNA analysis." 3: 3 17-322.
Wake, D. B. and C. J. Schneider (1998). "Taxonomy of the plethodontid genus Ensatina." Hemetologica 54(2): 279-298.
Wei, G., N. Xu, et al. (1993). "Karyotype, C-band and Ag-NORs study of three stink frogs." Asiatic Herpetological Researa 5: 45-50.

Werner, F. (1930). "RUM Ieporipes a new species of fmg from south China." Science Journd 9(1+2): 45-47, p1.49.
Wiley, E. 0. (1981). Phvuenetics. New York, John Wiley and Sons.
WWF (1998). "World Wide Fund for Nature Global Network."b (website).
Yang, D. (1991). "Phylogenetic systematics of the Amolops group of ranid frogs of southeastem Asia and the Greater Sunda Islands." Fieldiana 63.
Yang, D.-T., Ed. (1% 1). -an-Fauna of Yunna~. Beijing, China Forestry Publishing House.
Zhao, E. and K. Adler, Eds. (1993). Heqetolo~v of Chiea. Contributions to Herpetology. St. Louis, MO, SSAR.
Zhao, E.-M. (1994). "A historical review and evduation of the partitioning of the ranid genus Rana (Amphibia : Saientia)." Sichuan Journalof(3): 1 1 1 - 1 15.

Appendix A:
Specimens studied

Appendix A
Appendk A. Specimens used in this study. A total of 227 specimens were used in this
investigation. The first column of the table identifies specimen number (Catalogue #). Those
whose museum catalogue numbers begin with 'A' are from the American Museum of Natural
History (AMNH), dl others are from the Royal Ontario Museum (ROM). The next column
identifies the morphotype or species to which the individual belongs (group). Abbreviations for
this column are as follows: Huia nasica (H. nasica); RUM livida morphotypes: 1) Typical fonn
2) Black Egg form, 3) Speckled form 4) Small form 5) Mottled form 6) southern Big Eye form (s
Big Eye), 7) northem Big Eye form (n Big Eye), 8) Large form. The next column identifies
Iocality and temporal data. Localities and abbreviations are as follows: Darjeeling Region, India
(Id 1882); Buoen Loi, Gia Lai Province, Vietnam (BL); Tram Lap, Gia Lai Province, Vietnam
(TL); Khe Moi River, n e z Con Cuong, Nghe An Province, Vietnam; Sa Pa, Lao Cai Province,
Vietnam (Sap); Tarn Dao, Vinh Phu Province, Vietnam (TD); Ba Be Nationai Park, Cao Bang
Province, (Ba Be); Hong King, People's Republic of China (HK); Nha Hang Nature Reserve, Pac
Ban, Tuyen Quang Province, Vietnam (PB); Fujien Province, People's Republic of China (Fuj).
Numbers beside locality abbreviations indicate year of collection, Refer to Table 2.1 for those
without dates indicated. The next three columns indicate the whether or not the specimen was
included (+) in morphornetric analyses (Morph), cellular DNA content analyses (DNA Con), or
Allozyme electrophoresis (Alloz). The final 17 columns are measurement data in mm.
Abbreviations for measurements taken are as follows: snout-vent length (SVL), head lengih
(HL), head width (HdW), finger length (Fgr), hand Iength.'(Hnd), toe Iength (TL), foot length
(Ft), tibial length (Tib), inter-orbita1 distance (IOD), orbit diameter (Eye), distance between
tympanum and eye (T-Eye), tympanum diameter (Tmp), snout length (Snt), finger pad length
(FpL), finger pad width (FpW), toe pad length (TpL), toe pad width (TpW).
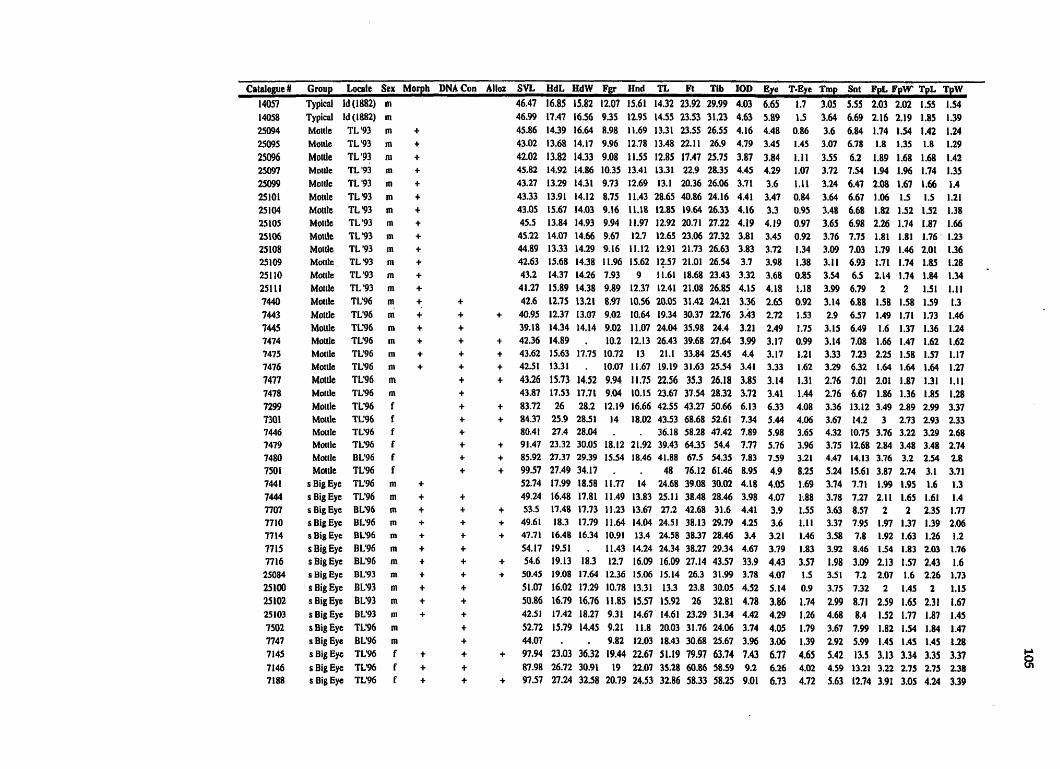
Catalogue# Group Locale Sex M o ~ h DNACon Alloz SVL HdL HdW F p Hnd TL Pt Tib IOD Eye T-Eye Tmp Snt FpL FpW TpL TpW
14057 Typical Id (1882) m 46.47 16.85 15.82 12.07 15.61 14.32 23.92 29.99 4.03 6.65 1.7 3.05 5.55 2.03 2.02 1.55 1.54 pical al ld(1882) m Motrle TL'93 m Mottle TL'93 m Mdile TL'93 m Mottlc TL'93 m MotUc TL '93 m Motde TL'93 m Mottlc TL '93 m Moiile TL '93 m Molile TL93 m Motile TL '93 m Mottle TL 9 3 m Moiile TL '93 m Motile TL93 m Mottlc TL96 m Motile TL'96 m Moule TL96 m Moule TL'96 m Motile TL96 m Motile TL96 m Motlle TL96 m Motile TL96 m Motile TL96 f Mottlc TL96 f Moitlc TL'96 f Moitlc TL96 f Motile BL'96 f Mollie TL96 f
s Big Eye TL'96 m s Big Eye TL96 m s Big Eye BL'96 m s BigEyc BC96 m s Big Eyc BL96 m s Big Eye BL'96 m s Big Eye BL'96 m s Big Eye BL'93 m sBig Eye BL'93 m s Big Eye BL'93 m sBig Eye BL'93 m s Big Eye TL96 m s Big Eye BL96 m s Big Eyc TL96 f s Big Eye TL96 f s Big Eye TL'96 f

Catalogue# Croup Locslc Sex Morph DNACon Alloz SVL HdL HdW Fgr Hnd TL Ft Tib IOD Eye T-Eyc Tmp Snl FpL FpW TpL TpW + + 92.58 29.11 33.32 19.98 24.3 49.96 78.29 62.25 8.9 5.87 5.2 3.84 14.16 3.16 3.13 3.6 3.21 s Big Eye TC96 f
s Big Eye TL96 s Big Eye ELY6 sBig Eye TC96 sBig Eye TL96 s Big Eye TL96 s Big Eyc TL96
Large KM94 Large KM94 Large KM94 iauge KM94 Large KM'94 Large KM94 Lnrge KM94 Large KM94 Large KM94 targt K M ~ S L q e KM95
n Big Eye PB '96 n Big Eye PB '96 nBig Eye PB96 Spccklt Sap95 Speckle Sap95 Speckle Sap'95 SpecWe Sap'95 Speckle Sap95 SpecLle Sap95 Speckle Sap'95 Specklc Sad95 Speckie Sap95 Small Sap'9S Small Sap'95 Small Sap'95 Small Sap'95 Srnall Sap'95 Small Sap'95 Small Sap95 Small Sap95 Small Sap'95 Small Sap'95 Small Sap'95 Small Sap'95 Smdl Sap'95 Small Sap95 Small Sap'95

CsîalogueU Group Locale Sur Morph DNA Con Alloz SVL HdL HdW Fgr End TL Ft Tib IOD Eyc T-Eye Tmp Snt FpL FpW TpL TpW 19047 Srnail Sap'95 f 55.66 14,08 17.75 13.91 15.54 30.17 47.01 33.59 5.59 5.59 2.43 2.4 7.76 2.84 2.84 228 1.9
Typicnl Typical Typical Typical Typid Typiatl Typical Typical Typicnl Typical Typical Typical Typical Typical Typicnl Typical Typical Typical Typical Typical Typical Typical Typicai Typical Typical Typical Typical Typical Typical Typical Typical Typical Typical Typical Typical Typical Typical Typical Typical Typifnl Typicol Typical Typical Typical
TU95 TD'95 TD'95 TD'95 Tû'95 ~ ~ ' 9 s TD95 TU95 TV95 TD95 TV95 T m 5 TD'96 TDP6 TV96 Tû'96 Tû'96 TV96 TiY95 TV95 TU95 TD95 TU96 TU96 TU96 T m 6 TU96 TD96 TD96 TD'96 TU96 TU96 BaBe BaBe BaBc BaBe BaBe BaBc BaBc BaBe BoBe BaBe BaBc BaDe

Catalogue # Group Locale Sex Morph DNA Con Alloz SM. HdL HdW Fgt Hnd TL Ft Tib IOD Eye T-Eye Tmp Snt FpL FpW TpL TpW 26362 Typical BoBe m + + + 47.69 17.08 17.1 11.4 13.6 23.34 36.32 28,8 10.19 8.1 2.64 3.79 7.3 2.37 2.37 1.85 1.9
Typical Typical Typical Typical Typical Typical Typical Typical Typical Typicol Typid Typicol Typical Typical Typicnl ~ ~ ~ i c n l Typical Typical Typid Typical Typical Typical Typical Typical Typical Typical Typical Typical Typical Typical Typical Typicnl Typical Typical Typical Typical Typical Typical Typicol Typical Typical Typical Typical Typical
BaBe rn + + BaBe m + + BaBe m + + BaBe m + + BaBe m + + BnBe m + + HKP6 f + + + HK1% m + + + HK'96 m + 4. + HKP6 m + + + HK96 m + + + PB '96 f + + PB96 f + + + PB'96 f + + PB'96 f + PB96 f + PBY6 f + PB'96 f + PB'96 f + PB '96 f + PB '96 ni + 4
PB'96 m + + + PB '96 m + + + PB96 m + + PB96 m + PB96 m + + PB'96 m + KM94 f + + KM94 f + + KM? f + + KM94 f + + KM94 f + + KM94 f + KM'94 f t KM94 f + KM94 f + KM'94 f + KM'94 f + KM'94 f KM95 f + KM'94 f KM94 m + + KM94 m + KM94 m +


Appendix B: Measurement means for each morphotype

111
Appendix B: Measurement means for each morphotype
Variable
India Male SVL HDL HDW FGR Hm3 TL FI' TIB IOD EYE
TEYE TMP SNT P L FPW 'PL TPW
Mottle Male SVL, HDL HDW FGR H m TL FT
TIB IOD EYE TEYE TMP SNT . 21 FPL 21 FPW 21 TPL 21 TPW 21
Minimum
46.47 16.85 15.82 9.35 12.95 14.32 23.53 29.99 4.03 5.89 1 .SO 3.05 5.55 2.03 2.02 1.55 1.39
39.18 12.37 13 .O7 7.93 9 .O0 11.61 17.47 22.76 3.21 2.49 0.84 2.76 6.20 1 .O6 1.35 1.31 1.1 2
Maximum
46.99 17.47 16.56 12.07 15.61 14.55 23.92 3 1.23 4.63 6.65 1.70 3 . a 6.69 2.16 2.19 1-85 1.54
45.86 17.53 17.75 1 1.96 15.62 28.65 40.86 28.35 4.79 4.48 1.75 3.99 7.75 2.26 2.00 2.01 1.66

Mottle Fem SVL HDL HDW . FGR HM> TL Fr TIB IOD EYE TEYE TMP SNT FPL FPW TPL TPW
s BigEye Male SVL HDL HDW FGR HND TL FT m IOD EYE TEYE TMP SNT FPL. FPW TPL TF'W
s BigEye Fem S m HDL HDW FGR HND TL FT
TIB

IOD EYE TEYE TMP SNT FPL FPW TPL TPW
Large SVL HDL, HDW FGR mm n FI' TIB IOD EYE TEYE TMP S m FPL FPW TPL TPW
TYP Fem (KM) SVL HDL HDW FGR ImD TL FT TIB IOD EYE TEYE TMP S m FPL FPW TF'L
TYP Male O SVL HDL, HDW FGR HND TL FT
T I . IOD EYE
TEYE TMP SNT FPL FPW TPL. TPW
Speck Fem SVL HDL HDW FGR IIND TL FT m IOD EYE
TEYE TMP SNT FPL FPW TPL TPW
Speck Male SVL 3 HDL 3 HDW 3 FGR 3 HND 3 TL 3 FT 3

TIB IOD EYE
TEYE TMP S m FPL FPW TPL TPW
Small male SVL M>L HDW FGR HND n FT
TIB IOD EYE
TEYE TMP SNT FPL FPW TPL TPW
TYF Fem (TW SVL HDL m w FGR HND TL FT
TIB IOD EYE
TEYE TMP SNT FPL FPW TPL

TYP Male SVL HDL HDW FGR HND TL FT
TIB IOD EYE
TEYE TMP SNT FPL FTW TPL TPW
TYP Fem (B) SVL HDL HDW FGR HND TL FT TIB IOD EYE
TEYE TMP SNT FPL FPW TF'L TPW

FT TIB IOD EYE TEYE TMP SNT FPL FPW TPL TPW
Typ Fem (HK) SVL H m HDW FGR HND TL FT TIB IOD EYE
TEYE TMP S m FPL FPW TPL TPW
Typ Male (HK) SVL HDL HDW FGR HND TL FT
TIB IOD EYE
TEYE TMP SNT FPL FF'W
TPL TPW
TYP Fem (PB) SVL HDL HDW FGR HND TL FT TLB IOD EYE TEYE TMP S m FPL FPW TPL TPW
TYP Mak (PB) SVL HDL HDW FGR HND TL FT TIB IOD EYE
TEYE TMP SNT FPL FPW TPL TPW
n BigEye Male SVL HDL HDW FGR HND

TL FT
TIB IOD EYE
TEYE TMP SNT P L FPW TPL TPW
B lack Egg
SVL HDL HDW FGR HND TL FT
TIB IOD EYE
TEYE TMP SNT FPL FPW TPL TPW

Appendix C: Population Aggregates Analysis Results

Appendix C. Population aggregate analysis, Stage 1. Presence/absence of 14 attributes among
(loci) from each of 15 populations of Rana livida complex frogs and one population of Huia
nasica. Numbers across the top refer to polymorphic loci and their alieles (refer to Table 3.2a
for enzyme abbreviations): 2 (rnAat-A), 3 (Ck-C), 4 (Gpi-A), 6 (mICDH-A), 7 Ldh-A, 8 (Ldh-
B), 9 (rnMdh-A), 10 (sMdh-A), I l ( M e ) , 12 (Pep-A), 13 (Pgm-A), 14 (sSod-A), 15 (Fumh-A).
The presence of an allele by ail individuals of the population is indicated with a '1' at the bottom
of the allele column, and absence is indicated with a '0'. AlleIes that are not expressed by the
whole population are shown as a plus minus (k), with a plus sign (+) indicating presence and a
minus sign (-) indicating absence in an individuai. Individuals with unresolved loci were
indicated with a backslash (0.






Appendix C. Popdation aggregate analysis Stage il. Populations profiles
representing presence @), absence (O), and fixed presence (1) of 14 attributes among
(loci) from each of 15 populations of Rana Zipidn complex frogs and one population of
Huia nasica. Numbers across the top refer to polymorphic loci and their alleles
(abbreviations follow Table 3.6). M y s i s indicates presence of four new species based
in the presence of fixed d e i c differences. The Large and northern Big Eye forms
exhibit one fbced delic difference from each other, insuffiàent for speàes delineation,
but suggesting population substructuring.


O O O - H fl


f l o o o
f l r r . 9
O O 0'0
0 0 0 0
0 0 0 0
r v - r c .

Appendix D: Redescription of the Rana livida complex

Appendix D: Redescription of the Rana Iivida complex
Rana liziida (Blyth, 1855)
?Polypedates smaragdinus Blyth, 1852 Polypedates chloronotus Günther, 1875 Rana chloronota Boulenger, 1 882 Rana livida Boulenger, 1 887 Rana (Hylarana) livida Boulenger, 1920 Rana graminea Boulenger 1900 Rana (Hylorana) sinica Ahl, 1925 Rana leporâpes Werner, 1930 O d o m a livida Fei, Ye, and Huang, 1990 Rana (Eburana) livida Dubois, 1992
Species 1 - Typical
Diagnosis
Rana livida considered to be a member of the subgenus Odorrana (sensu Fei et al.
1990), characterized by: (1) body dorso-ventrally compressed; (2) snout-vent length ranges from
45-50 mm for males and 90-1Wmm for females. (3) vomerine teeth are present in rows oblique
to the interna1 choanae; (4) white lip iine extends across the upper lip, ending in a glandule
anterior to the tympanum; (5) head bluntly rounded, head width exceeding head length; (6)
tympanum circular, distinct, approximately 36% of eye diameter for males, 32% for females,
siight supratympanic fold; (7) dorsal skin smooth, becoming grandar laterally, lacking
dorsolateral folds; (8) dorsum bright green, sometimes with black spots; forelimbs and
hindlirnbs; (9) feet fully webbed to the distal phalanx, webbing brown; (10) Fmger and toe pads
greatly enlarged (finger iII > 3 mm); (1 1) subarticular tubercles and an interna metatarsal
tubercle distinct, conicai; 12) distal phalanges T-shaped; (12) omostemum small, unforked at
base, while metasternum large, deeply notched posteriorly; (13) ernits an odoriferous secretion.
(14) males with paired subgular external vocal sacs; (15) females bear white eggs (2mm in
diameter).

Rana livida resembles Huia nasica and several members of the subgenus Odomcu~a
(sensu Fei et al 1990): R. grahami, R. schmackerii, and R. undersonii. Al1 of these frogs are
found in forested montane rivers of southeast Asia and share sirnilar morphologues. Huia m i c a
differs from R. livida by having a solid brown dorsum, a head that is longer than it is wide,
females with a shorter snout vent length (67 mm for A. nasica, while 90-100 mm for R. .livida).
Al1 members of the subgenus Odorrana mentioned above differ from R. livida in their
pigmented eggs, and absence of a smooth, green dorsum. Rana schnuzckeni is heavily spotted
on its dorsum. Rana grahami aiso differs from R. livida in that it is very granular IateraiIy and
lacks toe pads. Both in life and preservation, R. livida bears a closest resernblance to R.
andersonii. Rana andersonii differs from R. livida by having an olive-brown dorsum and eggs
with melanin.
Description
Adult female (ROM M64 14). Head much wider than long, head 1ength:head width 1.01.
Head width 33% of SVL; head length 34% of SVL; snout short, protruding beyond margin of
lower jaw, acutely rounded in dorsal view, bluntly rounded in profite; eye large, prominent, 41%
of snout; eyelid broader than inter-orbital distance. Top of head flat; canthus rostraiis rounded;
loreal region concave; lip flared slightly anterior to orbit; nostri1 about 3/4 distance from eye to
tip of snout. Slight supratyrnpanic fold, curving posteroventrally fiom the tympanum to point
above the rnaxilla; tympanum circular, distinctly visible, separated from eye by distance equal to
that of tympanum diameter, which itself is 5 1 % of diameter of the eye. Choanae ovoid;
vomerine dentigerous processes prominent, oblique, posterornedial to choanae, each bearing
numerous teeth. Tongue cordifoxm, distinctly notched posteriorly, free for approximately 2/3 of
its length.
Skin on dorsum smooth, becorning shagreened with iight granulations laterally; dennal
folds absent; small tubercles posteroventral to tyrnpanum; prominent granules on flanks and
antenor to cloaca; cioacal opening un-modified, directed posteriorly, at upper level of thighs.
Forearms moderately robust; ringers moderately short, slender, hands 28% of SVL,
relative lengths of fingers lI<IdV<II[I, disks greatly expanded, relative pad size m>lV>bII, pad
lengh (4.18 mm, digit III) exceeds pad width (3.73 mm, dig it m), horseshoe-shaped

circummarginal grooves present ventrally; terniinal phalanx t-shaped; subarticular tubercles
conical. Hind limbs moderately robust; tibia length 62% of SVL; foot length 45% of SVL;
relative toe lengths IdIcUicVdV; inner tarsal fo1d absent; feet fully webbed to distal phalange,
lateral skin flap on V; toes long, slender, with large , rounded triangular disks, relative pad size
I=II=m>IV>>V, pad length (IV) 4.09 mm, pad width (IV) 2.30mm, each with ventral
horseshoe-shaped circummarginal grooves; subarticular tubercles prominent and conical;
metatarsal tubercle on I ovoid, long.
Colour in preservative: Dorsum Iivid blue. White lip line; loreal region black;
tympanum beige with dark brown centre ring. Dorsal surfaces of limbs banded; webbing brown.
Flanks grey with some white rnottling posteroventrally. Cloaca1 region black, thighs rnarbled
brown-yellow. Ventrum creamy white, ventral surfaces of limbs creamy yeiiow with black
mottling.
Colour in life: Dorsum green, flanks yellow-grey; white lip line; loreal brown; iris gold;
top 1/3 red, dorsal limbs brown with black banding; thighs brown with bIack marbling; webbing
marbled white on dark brown; venter creamy white.
Distribution and Ecology
Rana livida is known frorn forested montane river systems across southeast Asia. The
rivers range fiom slow moving to extremely fast and deep. Specimens from this study were
found on boulders and logs, both in and around the water. They were aiso found in neighbouring
forests. No stomachs were dissected, but x-rays showed ingestion of large invertebrates, and
small vertebrates including a megophryid. The tadpoles of this species are unknown.
Species 2 - B h k Egg
Diagnosis
Differs fonn that of the ~ypical form in that: dorsal skin shagreened, becoming granula.
laterally, lacking dorsolated folds; dorsum brown with black blotches; tympanum ovoid,

distinct, approximately 52% of eye diameter, with slight supratympanic folci; lip bauds black,
extending vertically across mandible; forelimbs and hindlimbs banded upon indistinct blotches,
bands continuing to dibtal tips of external two digits; laterd skin flaps prominent on fingers II,
III, and IV; fmger and toe pads moderately enlarged; females bare black eggs ( 2 m in
diameter); males unknown.
This species c m easily be distinguished from ail other cascade ranids by its large size,
brown and black colouration, and its black eggs.
Description
Adult fernaie (ROM #29534). Differs from that of the Typical form in: Head
1ength:head width 0.82; head width 35% of SVL; head length 28% of SVL. Tympanum 5 1 % of
diameter of the eye. Skin on dorsum shagreened with heavy granulations, becorning leathery
upon dcohol preservation. Wind limbs rnoderately robust; tibia length 60% of SVL; foot length
63% of SVL.
Colour in preservative: Dorsum brown with black blotches. Distinct black bands cross
from upper to lower lips, ventral to the jaw. Dorsal surfaces of limbs banded with black on
brown to distal ends of digits; webbing distinctly rnarbIed white on black. Flanks grey with
more prominent black bIotches posteroventrally.
Colour in life: Dorsum and flanks brown with srnall black bIotches; vertical black Iip
bars; loreal brown; iris gold; top 1/3 red, dorsal limbs brown with black banding; webbing
marbled white on dark brown; venter crearny white.
Distribution and Ecology
As in Species 1 - the Typical form. This species is only known from four forested
montane river systems in northern Vietnam. X-rays reveaied ingestion of large invertebrates,
including a small freshwater crab. The type specimen is the only one bearing fully developed
ova. It was found in October, suggesting a fdl breeding season, This was corroborated by the
presence of undeveloped ova (too smdl to be measured) in two other specimens (ROM 26358,

29529) collected in the spring. No males, and therefore no calls, were associateci with this
species.
Species 3 - Speckled
Diagnosis
Differs from that of the Typical form in that: snout-vent length ranges of males 54 - 59
mm for males; skin heavily granulated; dorsum bright green, with deep black spots; finger and
toe pads modestly edarged (finger III < 3 mm); d e s with intemal vocal slits.
This species closely resernbles Rana grahami. The two are distinguishable since R.
grahami males are much larger (70 -80 mm), do not have enlarged toe pads, and females bear
pigmented eggs.
Description
Adult female (ROM #19055). As in the description of the Typical form with the
following exceptions. Head 1ength:head widtb .80; head width 37% of SVL; head length 30% of
SVL. Finger pad length (2.83 mm) greater than finger pad width (2.0 mm). Toe pad length (IV)
3.27 mm, pad width (IV) 3.26 mm. Skin bumpy with Iarge granules. Males with intemal vocal
slits.
Colour in preservative: Dorsum iivid blue and grey with bIack spots.
Distribution and Ecology
This species is only known from the far north of Vietnam. The mountains are the
highest in the country. Xt is the only part of Vietnam to have snow in the winter rnonths.

The Small species diiTers from the diagnosis of the Typical form in that: snout-vent
length ranges from 32-38 mm for males and 55.66 for the single fernale specimen; vomerine
teeth are absent; tympanum approximately 37% of eye diameter for d e s , 43% for the female;
small white dorsolateral glandules; dorsum green without any spots; sides with two large white
spots; metasternum forked.
This specimen is noticeably smaller than d l other cascade ranids of southeast Asia. Its
white dorsolateral glandules make it unmistalcable from other species.
Description
Adult female (ROM #lgON). The description follows that of the Typical form with the
following exceptions: Head 1ength:head width .79; head width 32% of SVL; head length 28% of
SVL; snout short, blunt. Eye very large, prominent, 72% of snout. Tympanum 43% of diameter
of the eye. Vomerine dentigerous processes absent. SrnaIl white dorsolaterd glandules. Fhger
pad length equals pad width (2.84 mm, dig it III). Toe pad length (IV) 2.28 mm, pad width (IV)
1.9 mm.
Colour in life and in preservative: As in the Typical form.
Distribution and Ecology
This species is only known from the far north of Vietnam. The mountains are the
highest in the country. It is the only part of Vietnam to have snow in the winter rnonths.
The forked stemum differs from that of the other rnembers of the complex and the
subgenus in which they are recognized, Odorrana. This type of stemum more closely resernbles
that of Hue Nabisco. It is unclear whether or not this characters has littie diagnostic power

above the species level. It is possible that this species is not of the genus RaM at dl. Frogs of
the Annupolis group of genera are defined by the sucker disks on the tadpoles. Because larvae of
al1 species of Rana livida are unknown, as are the phylogenetic relationships of the cascade
ranids if southeast Asia, it is unclear whether the Small fonn (or the entire cornplex, for that
matter) belong to the genus Rana.
Species 5 - Mottled
This species dfiers from the diagnosis of the Typical fonn in that: snout-vent length
ranges from 39-45 mm for males and 80-100mm for females; tympanum large approximately
69% of eye diameter for females, 95% for males; dorsum bright green in daylight, brown at
night. Subarticula. tubercles are not pronounced.
Description ,.
Adult fernale (ROM #7301). as in the Typical form with the following exceptions: Head
1ength:head width -91.; eye large, prominent, 38% of snout; tympanum is 67% of diameter of the
eye.
Colour in preservative: As in Typical form.
Colour in Me: Variable. Night colour typically brown, day colour green. Appearance is
variable, as the Mottle form cm appear to be identical to the Typicai fonn, completely green,
completely brown, or any combinations of the two colours.
Distribution and Ecology
The Mottle form is known only from the Central Highlands, Gia Lai Province, Vietnam.
It inhabits forested montane river systems. Specimens from this study were found on or near
rapids or waterfalls.

Cornmen ts
This species is the only form known to change colours from night and day.
Species 6 - Southern Big Eye
Diagnosis
The diagnosis of this species differs from that of the Typical form in that: eye bulges
noticeably in life; tympanurn very large, 88% of eye diameter for males, 74% for females; skin
variable in texture, either shagreened or smooth dorsally; dorsum variable in colour fonn bright
green to brown; legs unbanded.
This fonn greatly resembles the M o d e species of the Rana livida complex. The two
species are sympatric. Males of the southem Big Eye form are noticeably larger than their
Mottle form counterparts. The Mottle form is known to have day and night colouration, but it is
unclear whether or not the Big Eye form shows the same capability.
Description
AduIt female (ROM #7 145). The description of the southern Big Eye form is as that of
the Typical form, with the following exceptions: Head 1ength:head width .63; head width 34% of
SVL; head length 24% of SVL; eye large, prominent, 50% of snout; tympanum very large, ,
distinctly visible, 80% of diarneter of the eye. Skin on dorsum variable, sometimes smooth
sometimes shagreened, with light granulations laterally. 4
Finger pad length (3.13 mm, digit IXI) is less than pad width (3.34 mm, dig i t III). Feet
large, foot length 82% of SVL; toe pad length (IV) 3.35 mm, pad width (IV) 3.37 mm.
Colour in preservative: Variable colour in preservative, either as in the Typical form or
olive to brown.

Colow in life: Variable h m light green to dark brown.
Distribution and Ecology
The southern Big Eye form is known only from the Central Highlands, Gia Lai Province,
Vietnam. It inhabits forested montane river systems. Specimens from this study were found on
or near rapids or waterfalls Rana livida is known fiom forested montane river systerns across
southeast Asia. The rivers range from slow moving to extremely fast and deep. It is found
sympatrically with the MottIe form.
As in the Typical form, with the following exceptions: Snout-vent length ranges from
48-55 mm for males and 93-105mxi1 for females; white lip line is absent tympanum enorrnous for
males at, 115% of eye diameter, 52% for females; dorsal skin shagreened; dorsum olive to
brown sometimes with black spots;
This species (particularly the female) is remarkably simiiar to the Typical form. The two
species can be can be distinguished as the Large form is generally larger, has shagreened, not
smooth dorsal skin, and is usually brown , sometimes olive, but never bright green, as in the
Typical form of Rana livida.
Description
Adult female (ROM #12999). Follows that of the Typical form with the following
exceptions: Head lengtkhead width .80. Head width 38% of SVL; head length 27% of SVL;
eye very large, prorninent, 73% of snout. Skin on dorsum shagreened, becoming increasingly
granular lateraliy. Disks greatly expanded, fmger pad length (3.04 mm, digit III) greatly
exceeds pad width (2.55 mm, dig it III. Toe pad length (IV) 3.18 mm, pad width (IV) 2.39 mm.
CoIour in preservative: Dorsum grey to olive, otherwise, as in the Typical fom.

Colour in life: Dorsum olive - brown. White lip line is absent. Otherwise, as in Typical
form
Distribution and Ecology
This species is known from no& and north-central Vietnam. It is found in identicai
habitat as the Typical form.
Comments
There is a possibility that this species is polytypic. This fonn is only known from two
localities, Nha Hang Nature Reserve near Pac Ban, Tuyen Quang Province, Vietnam, and the
Khe Moi River near Con Cuong, Nghe An Province, Vietnam. The series collected at the Khe
Moi is dl-female, while the one collected at Pac Ban is ail-male, making a morphologicai
assessrnent very chdenging. Both sexes are notable for their large size and brown colour. As
well, they could only be distinguished by one fked allele with ailozyme electrophoresis. Each of
these populations showed at least two fked allelic differences with every other species of the
Rana Zivida complex and Huia nasica. The question of a species boundary must be left as an
open question, due to the lack of evidence to support the single fixed difference between the
male population (referred in this study as the northern Big Eye form) and the fernale population
(referred to in this study as the Large form).Error! Bookmark not defined.




















