
Antibiotic efficacy is linked to bacterial cellular respiration
Upload
independentCategory
view
2download
0
The effects of elevated CO2 on root respiration rates oftwo Mojave Desert shrubs
N A O M I M . C L A R K *1 , M A R T H A E . A P P L E w and R O B E R T S . N O WA K *
*Department of Natural Resources and Environmental Sciences, University of Nevada, Reno, Reno, NV 89557, USA, wDepartment
of Biological Sciences, Montana Tech of the University of Montana, Butte, MT 59701, USA
Abstract
Although desert ecosystems are predicted to be the most responsive to elevated CO2, lownutrient availability may limit increases in productivity and cause plants in deserts to allocatemore resources to root biomass or activity for increased nutrient acquisition. We measuredroot respiration of two Mojave Desert shrubs, Ambrosia dumosa and Larrea tridentata, grownunder ambient (� 375 ppm) and elevated (� 517 ppm) CO2 concentrations at the NevadaDesert FACE Facility (NDFF) over five growing seasons. In addition, we grew L. tridentataseedlings in a greenhouse with similar CO2 treatments to determine responses of primary andlateral roots to an increase in CO2. In both field and greenhouse studies, root respiration wasnot significantly affected by elevated CO2. However, respiration of A. dumosa roots o1 monthold was significantly greater than respiration of A. dumosa roots between 1 and 4 months old.For both shrub species, respiration rates of very fine (o1.0 mm diameter) roots weresignificantly greater than those of fine (1–2 mm diameter) roots, and root respiration decreasedas soil water decreased. Because specific root length was not significantly affected by CO2 andbecause field minirhizotron measurements of root production were not significantly different,we infer that root growth at the NDFF has not increased with elevated CO2. Furthermore, otherstudies at the NDFF have shown increased nutrient availability under elevated CO2, whichreduces the need for roots to increase scavenging for nutrients. Thus, we conclude thatA. dumosa and L. tridentata root systems have not increased in size or activity, and increasedshoot production observed under elevated CO2 for these species does not appear to beconstrained by the plant’s root growth or activity.
Keywords: Ambrosia dumosa, atmospheric CO2, Larrea tridentata, Mojave Desert, root respiration,
specific root length
Received 18 May 2009; revised version received 28 August 2009 and accepted 7 September 2009
Introduction
The recent rise in atmospheric CO2 concentrations
resulting from fossil fuel burning and deforestation is
the most widespread, documented, and observed ex-
ample of human influence on the earth’s atmosphere
(Vitousek et al., 1997). The magnitude of this effect is
expected to be greatest in desert ecosystems, where
elevated atmospheric CO2 concentrations are thought
to increase plant water use efficiency (Melillo et al.,
1993). Indeed, deserts have among the greatest in-
creases in aboveground productivity in response to
CO2 enrichment (Nowak et al., 2004) with the greatest
responses occurring during wet years (Smith et al., 2000;
Housman et al., 2006).
At the Nevada Desert FACE Facility (NDFF), above-
ground productivity and photosynthesis have increased
with elevated CO2 in the Mojave Desert (Smith et al.,
2000; Housman et al., 2006). With increases in carbon
assimilation under elevated CO2 concentrations, below-
ground resources are hypothesized to restrict plant
growth, especially in the water and nutrient limited
Mojave Desert (Titus et al., 2002). Therefore, alterations
in aboveground physiology likely influence the alloca-
tion of recent photosynthate to roots. For example,
plants may increase carbon allocation to root systems
for nutrient or water uptake, mycorrhizal symbioses or
further soil exploration. These investments can lead to
greater carbon losses via root turnover, exudation, and
respiration (Hungate et al., 1997). Because rhizosphere
processes are strongly related to nutrient uptake and
cycling, these activities must be adequately understood
1Current Address: Department of Plant Sciences, University of
California at Davis, Davis, CA 95616, USA.
Correspondence: Robert S. Nowak, tel. 1 775 784 1656, fax 1 775
784 4789, e-mail [email protected]
Global Change Biology (2010) 16, 1566–1575, doi: 10.1111/j.1365-2486.2009.02075.x
1566 r 2009 Blackwell Publishing Ltd

to better predict ecosystem changes such as plant
growth and carbon storage with elevated CO2.
Although both root structure and function influence
rhizosphere processes, more studies have focused on
root biomass responses to elevated CO2 than on root
physiological processes. Plant roots typically display a
pronounced increase in growth under enhanced CO2
conditions (Rogers et al., 1994; Hodge & Millard, 1998;
George et al., 2003), but this response depends on species
(Gorissen, 1996; Cotrufo & Gorissen, 1997). Larrea triden-
tata, a dominant shrub in the Mojave Desert and other hot
deserts of western North America, has not exhibited
consistent changes in belowground biomass in response
to elevated CO2 in controlled growth chamber and green-
house experiments. Obrist & Arnone (2003) found ele-
vated CO2 increased the total biomass of L. tridentata,
including the roots, while others have reported no in-
crease in belowground biomass (BassiriRad et al., 1997;
Huxman et al., 1999). Unfortunately, in most root biomass
studies, distinctions are not generally made on the rela-
tive contributions of active and non-active root to bio-
mass values, and these results can be misleading. For
example, root tips constitute a small portion of root mass,
but are the primary site of many physiological processes
(Norby, 1994; Cotrufo & Gorissen, 1997). Therefore,
changes in belowground biomass must be interpreted
cautiously, and root physiology should be further re-
searched to gain a clearer understanding of how below-
ground processes respond to elevated CO2.
Previous studies investigating the influences of CO2 on
root respiration in various plant species range widely in
their results. Increases (Janssens et al., 1998; Edwards &
Norby, 1999), no change (Tjoelker et al., 1999; Yoder et al.,
2000; George et al., 2003), and decreases (BassiriRad et al.,
1996) have all been reported. By examining the effects of
elevated temperature and CO2 concentrations on sugar
and red maples, Edwards & Norby (1999) found that the
respective rhizospheres of these species respond differ-
ently to these particular environmental variables. Because
average study site temperatures and species composition
differ among experiments, these factors may be respon-
sible for varying responses of root respiration to elevated
CO2. The lack of a universal response to elevated CO2
highlights the need for experiments testing CO2 effects in
specific systems.
At the NDFF, soil respiration rates are significantly
greater with elevated CO2 (A. de Soyza et al., unpub-
lished data). Because we have not observed increased
root growth (Phillips et al., 2006) or mycorrhizae (Clark
et al., 2009) under elevated CO2 in the Mojave Desert at
the NDFF, we hypothesized that plants at the NDFF
increased carbon allocation to root activity rather than
biomass with elevated CO2, thereby contributing to
greater soil respiration. Our objective was to determine
the effects of elevated CO2 on the root respiration rates
of Ambrosia dumosa and L. tridentata, two dominant
shrubs in the Mojave Desert, during both greenhouse
and field experiments. In this study, we examined fine
roots, which are the sites for uptake of most nutrients
and water and thus are most likely to respond to
increased carbon input.
Materials and methods
Greenhouse experiment
CO2 treatment. We grew L. tridentata seedlings under
two different CO2 concentrations (360 and 550 ppm) in
the Frits Went Greenhouse at the University of Nevada,
Reno. We did not include A. dumosa in this experiment
due to the difficulty in cultivating this species. The
greenhouse has two climate-controlled bays and is fitted
with a CO2 injection and scrub system allowing the two
bays to be maintained at different CO2 concentrations.
Night : day temperatures were set to 20 : 37 1C to mimic
those of a Mojave Desert spring and summer. To avoid
confounding bay effects, CO2 treatments and their
corresponding plants were transferred between bays
throughout the experiment.
L. tridentata seeds were placed in cheesecloth slings
under a dripping water faucet for 2 days to leach away
germination inhibitors. The seeds germinated under their
respective CO2 treatment in water-saturated, sterilized
sand in transparent, domed trays that maintained high
humidity. After 4 weeks, seedlings were either harvested
or transplanted to 5.5 L PVC pots filled with sand. Once
transplanted in September 2001, seedlings were given
100 mL H2O every 4 days. In addition, pots were
supplemented with 50 mL of 1/4 strength Hoagland’s
solution once per week until plants were harvested after
10 months.
Harvests. We harvested 16 seedlings per CO2 treatment
at 4 weeks (Fall 2001) and harvested the remainder of
the seedlings (20 plants per treatment) at 10 months
(Summer 2002). We measured root respiration as a func-
tion of oxygen consumption with a Clark-style oxygen
electrode (Hansatech Oxytherm, Hansatech Instru-
ments Ltd, Norfolk, UK). Immediately after harvest,
excised roots were placed in an aqueous physiological
buffer (10 mM MES, 1 mM CaSO4), and then root
respiration was measured for 10–15 min at 20 1C. We
measured the length of excised roots, and following
respiration measurements, we determined dry root
mass to calculate specific root length (SRL; m root g�1
dry mass). Although root diameter classes were defined
as fine (1–2 mm diameter) and very fine (o1 mm
diameter), almost all primary and lateral roots were in
E L E VA T E D C O 2 A N D D E S E R T S H R U B R O O T R E S P I R AT I O N 1567
r 2009 Blackwell Publishing Ltd, Global Change Biology, 16, 1566–1575

the very fine root diameter class for these very young
seedlings. Finally, we measured total dry mass of
aboveground and belowground biomass, and then com-
puted the root mass fraction (i.e. the proportion of total
biomass that was root mass). Roots were then combusted
and analyzed with a Perkin Elmer Instruments (Shelton
CT, USA) Model 2400 Series II CHNS/O Analyzer for
carbon and nitrogen content.
Statistical analyses. For 4-week-old seedlings, plant mass
and root mass fraction were analyzed using a one-way
ANOVA (PROC GLIMMIX, SAS 9.1) with CO2 concentra-
tion as the main effect. Root mass, respiration, and SRL
were analyzed with a two-way split-plot ANOVA (root
type�CO2) to determine CO2 effects on primary and
lateral roots. The individual plant was treated as the
experimental unit with root type as a split-plot treat-
ment. Data were transformed to meet ANOVA assump-
tions using the Box-Cox family of transformations
((yl�1)/l), with l selected using PROC TRANSREG
(SAS 9.1).
For 10-month-old seedlings, root and shoot biomass,
root mass fraction, very fine root respiration, and very
fine root SRL were analyzed with ANOVA (PROC
GLIMMIX, SAS 9.1). The main effect was CO2 concen-
tration. Again, data were transformed to meet ANOVA
assumptions using the Box-Cox family of equations
with the best l determined by PROC TRANSREG (SAS
9.1). We conducted similar analyses on carbon, nitrogen,
and C : N ratios of very fine and fine roots. All percent
carbon and nitrogen data were transformed using
arcsine square root, and C : N ratios were transformed
using log10.
Field experiment
Site description. The NDFF, 970 m asl on Frenchman Flat
at the Nevada Test Site, Nevada, USA, is a research
facility designed to study responses of an undisturbed
Mojave Desert ecosystem to increasing atmospheric
CO2 using free air CO2 enrichment (FACE) technology
(Jordan et al., 1999). Temperature extremes range from
48 1C to �19 1C, and precipitation is sporadic, averaging
� 140 mm annually. At the NDFF, which began
operation in April 1997 and ran continuously until
July 2007, six 23 m diameter study plots were used in
our study: three elevated CO2 FACE plots (CO2 set
point of � 550 ppm; average CO2 concentration
across all dates during 2001–2006 was � 517 ppm)
and three control rings fumigated at ambient CO2
(� 375 ppm) (http://www.unlv.edu/Climate_Change_
Research/). The soils are well drained Aridisols on an
alluvial fan with low soil carbon (0.18–1.8%) and
nitrogen (0.01–0.08%) concentrations (Jordan et al.,
1999). Vegetation is codominated by L. tridentata, an
evergreen shrub that grows over 1 m in height, and
A. dumosa, a smaller, drought-deciduous shrub.
Root observation boxes. We accessed roots of L. tridentata
and A. dumosa via small root observation boxes measur-
ing 60 cm� 45 cm� 30 cm with removable 50 cm� 20 cm
Plexiglas front windows that were perpendicular to but
located below the soil surface. Root boxes were installed
in 2000 by digging a square pit into the soil near the base
of two plants per species per plot and then placing a root
box in each soil pit (for a total of 24 boxes). The square pit
was slightly larger in cross-sectional area than the root
box to allow the root box to easily fit into the soil pit.
When inserting the box into the pit, the Plexiglas front
window was placed against undisturbed soil near the
base of the target plant, and then native soil (sieved to
remove gravel and rocks) was used to backfill any gaps
between the box and existing soil. Boxes were tightly
fitted with a lid that was constructed from a metal
frame and doubled layer of dense carpeting that was
sandwiched between metal hardware cloth. The carpet
helped minimize disturbance to the plant’s rooting
environment by: (1) intercepting sunlight such that the
interior of the root boxes stayed dark and roots were not
unnaturally affected by light; (2) providing thermal
insulation to help maintain natural soil temperatures
for roots growing near the root box; (3) absorbing rain
so that run-off from the box lid to the surrounding soil
did not occur, which would artificially increase soil
water content for surrounding roots; and (4) allowing
some water from large rain events to drip through the
interior of the box to deeper soils so that deeper soil
under the root box was not artificially dry. Styrofoam
also was placed against the Plexiglas window when not
in use to help maintain ambient soil temperature and a
dark environment. Boxes were accessed from moveable
platforms to avoid soil disturbance. Baseline tracings of
existing roots were made on sheets of transparent mylar
fitted to the Plexiglas windows each November before
the start of the growing season. Roots were monitored
and new roots were traced in date-specific colors at 2–4
week intervals until major root growth ceased in May or
June. The age of an individual root at the date of harvest
was determined from these tracings.
Respiration measurements. We measured individual
2–4 cm long excised end-of-root segments of known
age and diameter class. Excised roots were placed in
an aqueous physiological buffer (10 mM MES, 1 mM
CaSO4) in order to equilibrate for the 10 min
immediately after harvest. Respiration was measured
with a Hansatech Oxytherm Clark-style oxygen
electrode for 10–15 min at 20 1C by placing one root
1568 N . M . C L A R K et al.
r 2009 Blackwell Publishing Ltd, Global Change Biology, 16, 1566–1575

segment in oxygenated buffer in the cuvette of the
Oxytherm. We dried root samples for 48 h at 60 1C to
determine dry mass. Roots of two diameter classes
(very fine o1.0 mm diameter; fine 1–2 mm diameter)
were harvested in 2001, 2002, and 2003, but only very
fine roots were sampled in 2005 and 2006. Root age
varied from o1 month to � 4 months for roots
measured in 2001–2003, but only young roots
(o1 month) were sufficiently present in 2005 and
2006. Lengths and dry mass of roots were used to
calculate SRL (m root g�1 dry mass). Roots were
combusted in a Perkin Elmer Instruments Model 2400
Series II CHNS/O Analyzer to determine carbon and
nitrogen content.
Statistical analysis. Roots younger than 1 month were
categorized as ‘young’ for comparisons across years.
‘Old’ roots were defined as those roots older than
1 month. For each species, we conducted two
statistical analyses for root respiration and SRL. The
first analysis consisted only of data from 2001 to 2003
and included roots of different diameter and age classes
because fine roots and older roots were only abundant
enough for harvest in 2001, 2002, and 2003. Data for
2001–2003 were analyzed with a three-factor, split-plot
ANOVA using PROC GLIMMIX (SAS 9.1). The factors in
the analysis were CO2, diameter, and age. Age and
diameter were treated as split-plot treatments within
the main CO2 plots, and the ANOVA model included all
two- and three-way interaction terms. Data were
transformed to meet ANOVA assumptions using the
Box-Cox family of transformations with the best ldetermined by PROC TRANSREG (SAS 9.1). Owing to
the scarcity of fine roots, we were unable to consider
sampling date in this analysis, but pooled data across
years. Each plot was considered the unit of replication,
and the average respiration rate of roots from each root
box for very fine and fine roots between 2001 and 2003
was considered a subsample of each plot.
Because young, very fine roots were abundantly
available for harvest in 2005–2006, the second analysis
of root respiration and SRL included data from all
young, very fine roots across all years. We employed a
two-factor split-plot ANOVA (PROC GLIMMIX SAS 9.1)
for data analyses. We treated CO2 as the whole plot
factor and sampling date as the split-plot factor.
Individual root boxes (two per species per plot) were
considered to be subsamples. CO2 and sampling date
were the main effects, and the two-way ANOVA model
included the interaction term. Data were Box-Cox
transformed using the best l as determined by PROC
TRANSREG (SAS 9.1).
Results
Greenhouse experiment
Four-week-old seedlings. Although elevated CO2
significantly increased total plant biomass production
(P 5 0.010), it did not significantly affect root chara-
cteristics of 4-week-old L. tridentata seedlings (Table 1).
L. tridentata seedlings grown under elevated CO2 were
40% larger than those under ambient conditions.
Almost 60% of this increase in total biomass was due
to a significant increase in shoot biomass with elevated
CO2 (P 5 0.049). Although average primary and lateral
root biomass also increased with elevated CO2, these
differences were not significant (P 5 0.253). Root mass
fraction (the proportion of total biomass being root)
also did not respond significantly to elevated CO2
Table 1 Means and standard errors for parameters measured on 4-week-old Larrea tridentata seedlings grown in a greenhouse
under ambient and elevated CO2
Parameter
Ambient CO2 Elevated CO2
Mean (� SE) Mean (� SE)
Total plant biomass (mg) 6.717a (� 0.715) 9.634b (� 0.773)
Root mass fraction 0.240 (� 0.043) 0.358 (� 0.040)
Lateral root biomass (mg) 0.591 (� 0.195) 1.236 (� 0.322)
Primary root biomass (mg) 1.091 (� 0.202) 1.751 (� 0.266)
Shoot biomass (mg) 5.110a (� 0.536) 6.785b (� 0.685)
Lateral root respiration (mmol O2 min�1 m�1) �0.0303 (� 0.0161) �0.0171 (� 0.0038)
Primary root respiration (mmol O2 min�1 m�1) �0.0092 (� 0.0028) �0.0116 (� 0.0016)
Lateral root specific root length (m g�1) 59.83 (� 17.84) 72.85 (� 27.32)
Primary root specific root length (m g�1) 63.62 (� 13.01) 39.93 (� 4.69)
Means within a row followed by a different letter are significantly different.
E L E VA T E D C O 2 A N D D E S E R T S H R U B R O O T R E S P I R AT I O N 1569
r 2009 Blackwell Publishing Ltd, Global Change Biology, 16, 1566–1575

(P 5 0.080) but tended to increase with elevated CO2.
Lateral root respiration tended to be greater than
primary root respiration, but this difference was also
not significant (P 5 0.083). Neither CO2 (P 5 0.713) nor
its interaction with root type (P 5 0.484) significantly
affected root respiration. SRL was not significantly
affected by CO2 (P 5 0.332), root type (P 5 0.614), or
their interaction (P 5 0.714).
Ten-month-old seedlings. Root and shoot biomass as well
as root mass fractions of 10-month-old L. tridentata
seedlings were significantly affected by the CO2
treatments. Root biomass averaged 200% greater with
elevated CO2 (Po0.001; Table 2). Shoot biomass was
also greater with the elevated CO2 treatment (P 5 0.020),
although the relative increase (50%) was smaller than
that for root biomass. Consequently, root mass fractions
were significantly greater with elevated CO2 (P 5 0.012).
Very fine root carbon and nitrogen concentrations
(Table 2) were not significantly affected by CO2
(P 5 0.344 and 0.657, respectively), and consequently,
C : N ratios also were not influenced (P 5 0.606). Fine
root carbon (P 5 0.105) and nitrogen (P 5 0.681)
concentrations and C : N ratios (P 5 0.296) also were
not significantly affected by CO2 treatment.
Very fine root respiration was not significantly
affected by CO2 treatment (P 5 0.707; Table 2). Averaged
across CO2 treatments, L. tridentata seedlings’ root
respiration was �0.0140 � 0.0014mmol O2 m�1 min�1. In
addition, very fine root SRL of L. tridentata seedlings was
not significantly affected by CO2, (P 5 0.610). Averaged
across CO2 treatments, SRL of L. tridentata seedling very
fine roots was 159 � 17 m g�1.
Field experiment
Root respiration. Root respiration rates of adult A. dumosa
and L. tridentata growing in the field at the NDFF were
not significantly affected by CO2 treatment (Table 3,
Figs 1 and 2). CO2 interactions with age class and dia-
meter class were also not significant for both species
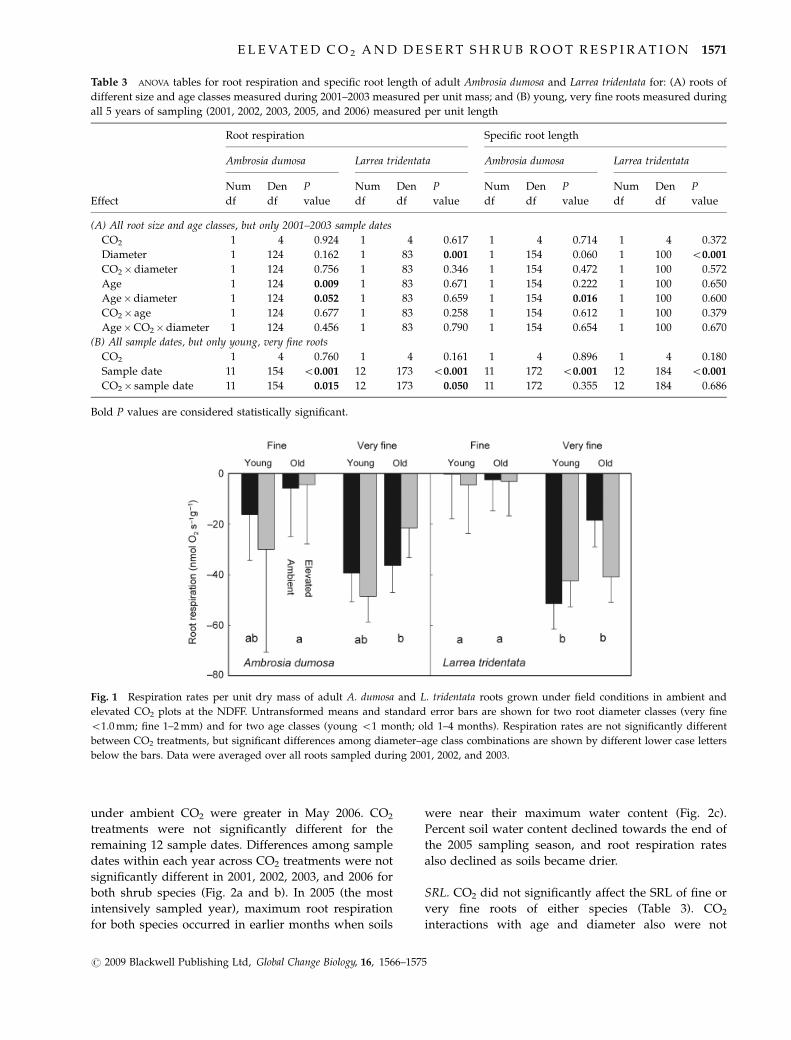
(Table 3A). For A. dumosa roots, age and age�diameter
were significant in the ANOVA. Averaged over both CO2
treatments and both root diameter classes, respiration
rates of young (o1-month-old) A. dumosa roots were
significantly greater than those of old (1–4 months old)
roots (Fig. 1). Although this ranking of means occurred
for both fine and very fine roots, mean comparisons
from the significant age�diameter interaction term did
not show significant differences between age classes
within each diameter class. However, for old A. dumosa
roots, fine roots had significantly lower respiration rates
than very fine roots. For L. tridentata, diameter was
the only significant effect in the ANOVA. Very fine
L. tridentata roots had greater respiration rates than
fine roots. Root respiration averaged across all CO2
treatments, ages, and diameters was �0.018 � 0.002
and �0.017 � 0.002mmol O2 min�1 m�1 for A. dumosa
and L. tridentata, respectively, on a unit length basis.
When expressed on a per mass basis, root respiration
rates averaged �25.3 � 5.7 and �20.5 � 7.5 nmol
O2 s�1 g�1 for A. dumosa and L. tridentata, respectively.
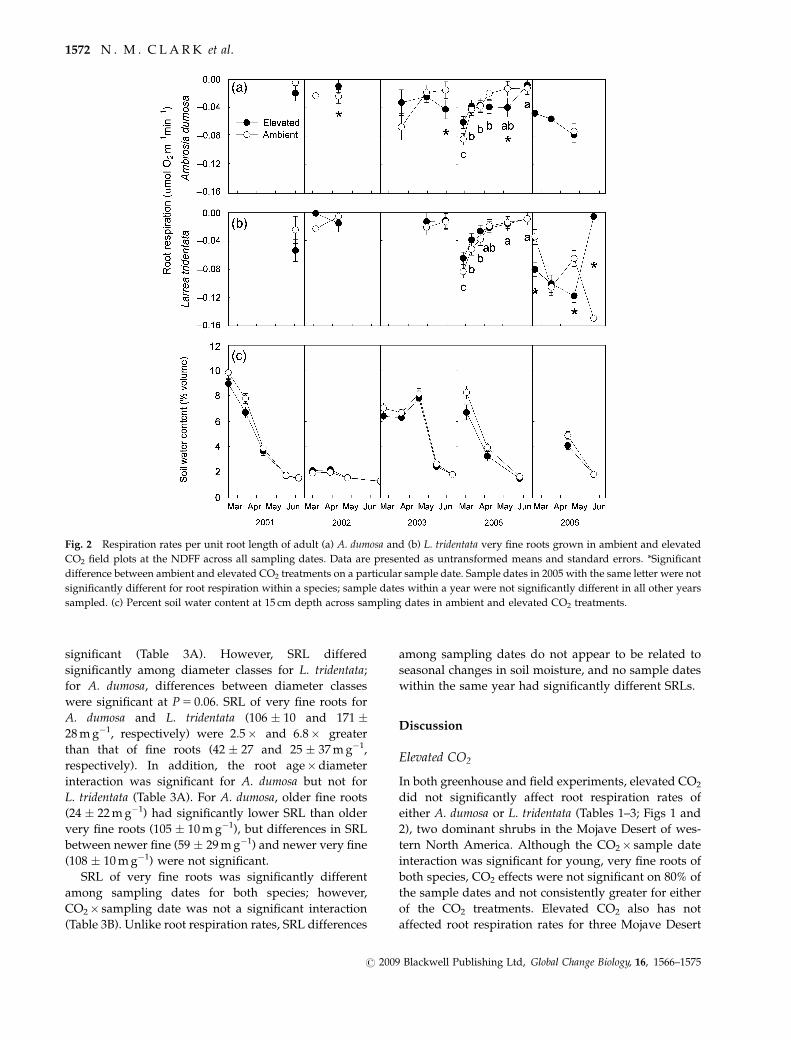
Although the overall main effect of CO2 was
not significant for very fine root respiration of both
A. dumosa and L. tridentata, rates significantly differed
among sampling dates and between CO2 treatments on
some sampling dates (Table 3B). For A. dumosa,
significant differences between CO2 treatments
occurred on only three of 15 sample dates (Fig. 2a).
On two of those dates (May 2003 and May 2005),
respiration of very fine roots for plants grown under
elevated CO2 was significantly greater than that under
ambient CO2, but respiration rates under ambient CO2
were greater on the other sample date (April 2002).
Significant differences between CO2 treatments for
L. tridentata only occurred in 2006 (Fig. 2b), and as for
A. dumosa, plants grown under elevated CO2 had
significantly greater root respiration for two sample
dates (February and April 2006) but respiration rates
Table 2 Means and standard errors for biomass fractions and
carbon and nitrogen contents of 10-month-old Larrea tridentata
seedlings in ambient and elevated CO2 treatments
Measurement
Ambient CO2 Elevated CO2
Mean (� SE) Mean (� SE)
Biomass
Root (g) 2.08a (� 0.23) 4.56b (� 0.54)
Shoot (g) 1.50a (� 0.15) 2.01b (� 0.15)
Root mass fraction 0.573a (� 0.026) 0.668b (� 0.025)
Carbon content
Very fine roots 28.5 (� 2.3) 31.5 (� 2.2)
Fine roots 31.1 (� 3.0) 37.3 (� 2.0)
Nitrogen content
Very fine roots 0.83 (� 0.07) 0.87 (� 0.07)
Fine roots 0.65 (� 0.08) 0.69 (� 0.06)
C : N ratio
Very fine roots 35.0 (� 1.7) 37.3 (� 2.6)
Fine roots 51.0 (� 3.1) 56.5 (� 4.0)
Root respiration
(mmol O2
min�1 m�1)
�0.0136 (� 0.0022) �0.0143 (� 0.0019)
Specific root length
(m g�1 root)
154 (� 28) 163 (� 22)
Means within a row followed by a different letter are signifi-
cantly different.
1570 N . M . C L A R K et al.
r 2009 Blackwell Publishing Ltd, Global Change Biology, 16, 1566–1575
under ambient CO2 were greater in May 2006. CO2
treatments were not significantly different for the
remaining 12 sample dates. Differences among sample
dates within each year across CO2 treatments were not
significantly different in 2001, 2002, 2003, and 2006 for
both shrub species (Fig. 2a and b). In 2005 (the most
intensively sampled year), maximum root respiration
for both species occurred in earlier months when soils
were near their maximum water content (Fig. 2c).
Percent soil water content declined towards the end of
the 2005 sampling season, and root respiration rates
also declined as soils became drier.
SRL. CO2 did not significantly affect the SRL of fine or
very fine roots of either species (Table 3). CO2
interactions with age and diameter also were not
Table 3 ANOVA tables for root respiration and specific root length of adult Ambrosia dumosa and Larrea tridentata for: (A) roots of
different size and age classes measured during 2001–2003 measured per unit mass; and (B) young, very fine roots measured during
all 5 years of sampling (2001, 2002, 2003, 2005, and 2006) measured per unit length
Effect
Root respiration Specific root length
Ambrosia dumosa Larrea tridentata Ambrosia dumosa Larrea tridentata
Num
df
Den
df
P
value
Num
df
Den
df
P
value
Num
df
Den
df
P
value
Num
df
Den
df
P
value
(A) All root size and age classes, but only 2001–2003 sample dates
CO2 1 4 0.924 1 4 0.617 1 4 0.714 1 4 0.372
Diameter 1 124 0.162 1 83 0.001 1 154 0.060 1 100 o0.001
CO2�diameter 1 124 0.756 1 83 0.346 1 154 0.472 1 100 0.572
Age 1 124 0.009 1 83 0.671 1 154 0.222 1 100 0.650
Age�diameter 1 124 0.052 1 83 0.659 1 154 0.016 1 100 0.600
CO2� age 1 124 0.677 1 83 0.258 1 154 0.612 1 100 0.379
Age�CO2�diameter 1 124 0.456 1 83 0.790 1 154 0.654 1 100 0.670
(B) All sample dates, but only young, very fine roots
CO2 1 4 0.760 1 4 0.161 1 4 0.896 1 4 0.180
Sample date 11 154 o0.001 12 173 o0.001 11 172 o0.001 12 184 o0.001
CO2� sample date 11 154 0.015 12 173 0.050 11 172 0.355 12 184 0.686
Bold P values are considered statistically significant.
Fig. 1 Respiration rates per unit dry mass of adult A. dumosa and L. tridentata roots grown under field conditions in ambient and
elevated CO2 plots at the NDFF. Untransformed means and standard error bars are shown for two root diameter classes (very fine
o1.0 mm; fine 1–2 mm) and for two age classes (young o1 month; old 1–4 months). Respiration rates are not significantly different
between CO2 treatments, but significant differences among diameter–age class combinations are shown by different lower case letters
below the bars. Data were averaged over all roots sampled during 2001, 2002, and 2003.
E L E VA T E D C O 2 A N D D E S E R T S H R U B R O O T R E S P I R AT I O N 1571
r 2009 Blackwell Publishing Ltd, Global Change Biology, 16, 1566–1575
significant (Table 3A). However, SRL differed
significantly among diameter classes for L. tridentata;
for A. dumosa, differences between diameter classes
were significant at P 5 0.06. SRL of very fine roots for
A. dumosa and L. tridentata (106 � 10 and 171 �28 m g�1, respectively) were 2.5� and 6.8� greater
than that of fine roots (42 � 27 and 25 � 37 m g�1,
respectively). In addition, the root age�diameter
interaction was significant for A. dumosa but not for
L. tridentata (Table 3A). For A. dumosa, older fine roots
(24 � 22 m g�1) had significantly lower SRL than older
very fine roots (105 � 10 m g�1), but differences in SRL
between newer fine (59 � 29 m g�1) and newer very fine
(108 � 10 m g�1) were not significant.
SRL of very fine roots was significantly different
among sampling dates for both species; however,
CO2� sampling date was not a significant interaction
(Table 3B). Unlike root respiration rates, SRL differences
among sampling dates do not appear to be related to
seasonal changes in soil moisture, and no sample dates
within the same year had significantly different SRLs.
Discussion
Elevated CO2
In both greenhouse and field experiments, elevated CO2
did not significantly affect root respiration rates of
either A. dumosa or L. tridentata (Tables 1–3; Figs 1 and
2), two dominant shrubs in the Mojave Desert of wes-
tern North America. Although the CO2� sample date
interaction was significant for young, very fine roots of
both species, CO2 effects were not significant on 80% of
the sample dates and not consistently greater for either
of the CO2 treatments. Elevated CO2 also has not
affected root respiration rates for three Mojave Desert
Fig. 2 Respiration rates per unit root length of adult (a) A. dumosa and (b) L. tridentata very fine roots grown in ambient and elevated
CO2 field plots at the NDFF across all sampling dates. Data are presented as untransformed means and standard errors. *Significant
difference between ambient and elevated CO2 treatments on a particular sample date. Sample dates in 2005 with the same letter were not
significantly different for root respiration within a species; sample dates within a year were not significantly different in all other years
sampled. (c) Percent soil water content at 15 cm depth across sampling dates in ambient and elevated CO2 treatments.
1572 N . M . C L A R K et al.
r 2009 Blackwell Publishing Ltd, Global Change Biology, 16, 1566–1575

grasses, Bromus madritdensis ssp. rubens, Achnatherum
hymenoides, and Pleuraphis rigida (Yoder et al., 2000).
Although similar results have been observed in other
ecosystems (Tjoelker et al., 1999; George et al., 2003),
contrary results have also been reported (BassiriRad
et al., 1996; Janssens et al., 1998; Edwards & Norby,
1999), making generalizations on how elevated CO2
affects root respiration difficult. Nitrogen contents of
A. dumosa and L. tridentata roots sampled from the
elevated CO2 plots at the NDFF are not significantly
different from those from ambient CO2 plots (R.S. Nowak
et al., unpublished data), and L. tridentata root nitrogen
also was not affected in our greenhouse experiment after
10 months of CO2 treatment (Table 2). These similar
nitrogen contents may help explain the similar root
respiration rates, which are highly correlated with nitro-
gen concentrations in root tissue across ecosystems
(Burton et al., 2002).
Even when conceptually scaled up to the collective
root systems of A. dumosa and L. tridentata in our field
study, our results indicate that root respiration rates of
the entire root system were not greater with elevated
CO2. Minirhizotron images at the NDFF do not
show significant changes in root length of A. dumosa
or L. tridentata with elevated CO2 treatment (Phillips
et al., 2006). Because we did not observe a change in SRL
of either species (Table 3), we can also infer that below-
ground fine root biomass for these two species is also
unaffected by the elevated CO2 treatment.
Contrary to the field results at the NDFF, where root
length of adult shrub communities treated with elevated
CO2 for up to 10 years were not significantly different
(Phillips et al., 2006; S. Ferguson et al., unpublished data),
L. tridentata root systems for 10-month-old greenhouse
seedlings were larger under elevated CO2 (Table 2).
However, the increase in root biomass after 10 months
may be short-lived and restricted to the early develop-
mental periods of seedlings. For example, Obrist &
Arnone (2003) noted that stimulation of root growth in
L. tridentata with elevated CO2 declined after 100 days
until their final harvest at 178 days. Greater root invest-
ments in early developmental stages with elevated CO2
may be an attempt to increase the likelihood of seedling
establishment. Indeed, at the NDFF, early survival of L.
tridentata seedlings was greater in the elevated CO2 plots
(Housman et al., 2003); however, this effect did not
continue with severe drought.
Compared with species found in other ecosystems,
our measured rates of root respiration for A. dumosa and
L. tridentata are slightly higher. Fine (1–2 mm diameter)
A. dumosa and L. tridentata roots respire at average rates
of �14 and �3 nmol O2 s�1 g�1, respectively (Fig. 1).
Unfortunately, no known studies have reported respira-
tion rates of desert shrubs for comparison with our
study. In other ecosystems, sugar maple roots of similar
diameter have respiration rates between �1 and
�10 nmol O2 s�1 g�1 (Pregitzer et al., 1998); our measure-
ments for A. dumosa are slightly higher than these for
sugar maple whereas L. tridentata falls within the range.
Very fine roots (o1 mm, although frequently o0.5 mm
diameter) of A. dumosa and L. tridentata under field
conditions respire at average rates of �36 and
�38 nmol O2 s�1 g�1, respectively; the average rate for
10-month-old L. tridentata under greenhouse conditions
is �37 nmol O2 s�1 g�1. Sugar maple roots o0.5 mm
diameter collected in Michigan respired at rates
between �5 and �20 nmol O2 s�1 g�1 (Pregitzer et al.,
1998), which are rates lower than those we measured for
two desert shrubs. Various gymnosperm and angios-
perm tree roots o1 mm diameter from a variety of
North American forests have respiration rates between
�2 and �8 nmol O2 s�1 g�1 (Burton et al., 2002), and
three Typha species have respiration rates ranging from
�1 to �7 nmol O2 s�1 g�1 (Matsui Inoue & Tsuchiya,
2008). Thus, A. dumosa and L. tridentata have greater
respiration rates than those reported for other woody
and herbaceous species. One possible explanation for
our observations of higher rates may be because the
time between root collection and respiration measure-
ment in our study was minimal (o20 min after harvest)
whereas time between collection and measurement for
other studies were up to 4 h (Pregitzer et al., 1998;
Burton et al., 2002).
Sample date
In the field experiment, both shrubs had reduced res-
piration rates at the end of the growing season in 2005
(Table 3, Fig. 2a and b). During 2005, soil water content
also was much lower than at the beginning of the year
(Fig. 2c). In this year, we found a significant, positive
relationship between soil moisture and root respiration
(result not shown). Although 2003 had a similar trend
as 2005, 2003 had too few data points to reliably conduct
a regression analysis. We also were not able to combine
data across years because differences among years in
other factors that affect root respiration (such as pre-
vailing soil temperatures, plant phenological stage at
the time of measurement, root age characteristics, etc.)
differed among years and because soil water and root
respiration were not always measured on the same
dates. Nonetheless, the relationship between soil water
and root activity is well documented in many systems
(e.g. Burton et al., 1998). It is important to note that
although we observed a decline in respiration rates of
L. tridentata and A. dumosa with low soil moisture, water
availability at this site is highly confounded with tem-
perature and phenological differences. To accurately
E L E VA T E D C O 2 A N D D E S E R T S H R U B R O O T R E S P I R AT I O N 1573
r 2009 Blackwell Publishing Ltd, Global Change Biology, 16, 1566–1575

determine the importance of soil moisture on root activ-
ity, a manipulative experiment should be implemented
with varying water availability throughout the season to
account for temperature and phenological effects.
Root diameter
Varying root diameters of A. dumosa and L. tridentata had
significantly different respiration rates (Table 3). Very fine
roots of both species had greater respiration rates than
fine roots, and this effect was not surprising given that
the finest roots are typically responsible for nutrient
uptake (Pregitzer et al., 1998; Desrochers et al., 2002).
However, when roots are classified by branching order,
clearer relationships between root form and function are
reported (e.g. Pregitzer et al., 2002). Indeed, when we
compared primary vs. lateral roots in our 4-week-old
L. tridentata seedlings, we found a trend toward greater
lateral root respiration (Table 1). Owing to our field
sampling technique, we could not always determine
branching order, although the majority of harvested very
fine roots appeared to be first branching order.
Root age
Root respiration rates were not significantly different
between age classes for L. tridentata but were for
A. dumosa, with older roots having lower respiration
rates than young roots (Table 3). Root nutrient uptake is
typically thought to decline with age (Eissenstat &
Yanai, 1997), which would decrease root respiration,
and grape, apple and citrus roots have all exhibited a
decline in root respiration with age (Bouma et al., 2001;
Volder et al., 2005). Although our results for A. dumosa
are consistent with these previous observations, it is not
clear why L. tridentata did not show similar trends.
Because of limited root availability and collection
feasibility, we defined root age as a categorical variable
(young or old) rather than numerical (exact age), with
the cutoff value of 1 month due to sampling constraints
(1 month was the approximate time interval between
root samplings). However, because 75% of L. tridentata
roots observed in the field with minirhizotrons survived
for over 1 year (S. Ferguson et al., unpublished data),
defining root age as older or younger than 1 month may
not have provided the resolution needed to detect root
age effects.
Conclusions
Our results from both the greenhouse and field experi-
ments suggest that root systems of L. tridentata and
A. dumosa have not altered carbon use for respiration
processes nor have they grown in size to compensate for
any nutrient limitations imposed by carbon fertilization.
Indeed, cumulative resin nitrogen and soil enzyme
activities are greater with elevated CO2 (Billings et al.,
2004; Jin & Evans, 2007), which indicate a change in
microbial community composition or activity with ele-
vated CO2 in the Mojave Desert. This carbon-limited
microbial community (Schaeffer et al., 2003) may be
stimulated by an increase in available carbon from litter
or root exudation. Increased aboveground productivity
with elevated CO2 (Smith et al., 2000; Nowak et al., 2004;
Housman et al., 2006) likely increases plant litter inputs
into the ecosystem (Weatherly et al., 2003). Root exu-
dation under elevated CO2 may also be altered, and
changes in the composition of root exudates could
consequently influence the heterotrophic communities
residing there (Jones et al., 2004). However, root exuda-
tion responses to elevated CO2 have not been investi-
gated for these Mojave Desert species and should be
investigated in future studies to clarify the potential for
plant-mediated shifts in soil carbon fluxes.
Acknowledgements
Funding for this research was provided by US DOE Office ofScience (DE-FG02-03ER63650 and DE-FG02-03ER63651) with addi-tional support from DOE National Nuclear Security Administra-tion/Nevada Operations Office, Brookhaven National Laboratory,and the Nevada Agricultural Experiment Station. We would like tothank Christina Wells, Vicki Longozo, Renee Richards, AndrewYoung, Dave Bryla, Iker Aranjuelo Michelana, Scot Ferguson, JamesKim, Eric Hoskins, and Christopher Holmes for help with the fieldand greenhouse studies and Kim Allcock and George Fernandezwith statistical analyses.
References
BassiriRad H, Reynolds JF, Virginia RA, Brunelle MH (1997) Growth and
root NO3� and PO4
�3 uptake capacity of three desert species in response
to atmospheric CO2 enrichment. Australian Journal of Plant Physiology,
24, 353–358.
BassiriRad H, Tissue DT, Reynolds JF, Chapin FS (1996) Response of
Eriophorum vaginatum to CO2 enrichment at different soil temperatures:
effects on growth, root respiration and PO4�3 uptake kinetics. New
Phytologist, 133, 423–430.
Billings SA, Schaeffer SM, Evans RD (2004) Soil microbial activity and N
availability with elevated CO2 in Mojave Desert soils. Global Biogeo-
chemical Cycles, 18, GB1011.
Bouma TJ, Yanai RD, Elkin AD, Hartmond U, Flores-Alva DE, Eissenstat
DM (2001) Estimating age-dependent costs and benefits of roots with
contrasting life span: comparing apples and oranges. New Phytologist,
150, 685–695.
Burton AJ, Pregitzer KS, Ruess RW, Hendrick RL, Allen MF (2002) Root
respiration in North American forests: effects of nitrogen concentration
and temperature across biomes. Oecologia, 131, 559–568.
Burton AJ, Pregitzer KS, Zogg GP, Zak DR (1998) Drought reduces root
respiration in sugar maple forests. Ecological Applications, 8, 771–778.
Clark NM, Rillig MC, Nowak RS (2009) Arbuscular mycorrhizal fungal
abundance in the Mojave Desert: seasonal dynamics and impacts of
elevated CO2. Journal of Arid Environments, 73, 834–843.
1574 N . M . C L A R K et al.
r 2009 Blackwell Publishing Ltd, Global Change Biology, 16, 1566–1575

Cotrufo MF, Gorissen A (1997) Elevated CO2 enhances below-ground C
allocation in three perennial grass species at different levels of N
availability. New Phytologist, 137, 421–431.
Desrochers A, Landhausser SM, Lieffers VJ (2002) Coarse and fine root
respiration in aspen (Populus tremuloides). Tree Physiology, 22, 725–732.
Edwards NT, Norby RJ (1999) Below-ground respiratory responses of
sugar maple and red maple saplings to atmospheric CO2 enrichment
and elevated air temperature. Plant and Soil, 206, 85–97.
Eissenstat DM, Yanai RD (1997) The ecology of root lifespan. Advances in
Ecological Research, 27, 1–60.
George K, Norby RJ, Hamilton JG, DeLucia EH (2003) Fine-root respira-
tion in a loblolly pine and sweetgum forest growing in elevated CO2.
New Phytologist, 160, 511–522.
Gorissen A (1996) Elevated CO2 evokes quantitative and qualitative
changes in carbon dynamics in a plant/soil system: mechanics and
implications. Plant and Soil, 187, 289–298.
Hodge A, Millard P (1998) Effect of elevated CO2 on carbon partitioning
and exudates release form Plantago lanceolata seedlings. Physiologia
Plantarum, 103, 280–286.
Housman DC, Naumburg E, Huxman TE, Charlet TN, Nowak RS, Smith
SD (2006) Increases in desert shrub productivity under elevated CO2
vary with water availability. Ecosystems, 9, 374–385.
Housman DC, Zitzer SF, Huxman TE, Smith SD (2003) Functional ecology
of shrub seedlings after a natural recruitment event at the Nevada
desert FACE Facility. Global Change Biology, 9, 718–728.
Hungate BA, Holland EA, Jackson RB, Stuart Chapin F III, Mooney HA,
Field CB (1997) The fate of carbon grasslands under carbon dioxide
enrichment. Nature, 338, 576–579.
Huxman KA, Smith SD, Neuman DS (1999) Root hydraulic conductivity
of Larrea Tridentata and Helianthus annuus under elevated CO2. Plant,
Cell and Environment, 22, 325–330.
Janssens IA, Crookshanks M, Taylor G, Ceulemans R (1998) Elevated
atmospheric CO2 increases fine root production, respiration, rhizo-
sphere respiration and soil CO2 efflux in scots pine seedlings. Global
Change Biology, 4, 871–878.
Jin V, Evans RD (2007) Elevated CO2 affects microbial carbon substrate use
and N cycling in Mojave Desert soils. Global Change Biology, 13, 1–12.
Jones DL, Hodge A, Kuzyakov Y (2004) Plant and mycorrhizal regulation
of rhizodeposition. New Phytologist, 163, 459–480.
Jordan DN, Zitzer SF, Hendry GR et al. (1999) Biotic, abiotic and perfor-
mance aspects of the Nevada Desert Free-Air CO2 Enrichment (FACE)
Facility. Global Change Biology, 5, 659–668.
Matsui Inoue Y, Tsuchiya T (2008) Interspecific differences in radial oxygen
loss from the fine roots of three typha species. Limnology, 9, 207–211.
Melillo JM, McGuire AD, Kicklighter DW, Moore B III, Vorosmarty CJ,
Schloss AL (1993) Global climate change and terrestrial net primary
production. Nature, 363, 234–240.
Norby RJ (1994) Issues and perspectives for investigating root responses
to elevated atmospheric carbon dioxide. Plant and Soil, 165, 9–20.
Nowak RS, Ellsworth DS, Smith SD (2004) Tansley review: functional
responses of plants to elevated atmospheric CO2 - Do photosynthetic
and productivity data from FACE experiments support early predic-
tions? New Phytologist, 162, 253–280.
Obrist D, Arnone JA III (2003) Increasing CO2 accelerates root growth and
enhances water acquisition during early stages of development in
Larrea tridentata. New Phytologist, 159, 175–184.
Phillips DL, Johnson MG, Tingey DT, Catricala CE, Hoyman TL, Nowak
RS (2006) Effects of elevated CO2 on fine root dynamics in a Mojave
Desert community: a FACE study. Global Change Biology, 12, 61–73.
Pregitzer KS, Deforest JL, Burton AJ, Allen MF, Ruess RW, Hendrick RL
(2002) Fine root architecture of nine North American trees. Ecological
Monographs, 72, 293–309.
Pregitzer KS, Lakowski MJ, Burton AJ, Lessard VC, Zak DR (1998)
Variation in sugar maple root respiration with root diameter and soil
depth. Tree Physiology, 18, 665–670.
Rogers HH, Runion GB, Krupa SV (1994) Plant responses to atmospheric
CO2 enrichment with emphasis on roots and the rhizosphere. Environ-
mental Pollution, 83, 155–189.
Schaeffer SM, Billings SA, Evans RD (2003) Responses of soil nitrogen
dynamics in a Mojave Desert ecosystem to manipulations in soil carbon
and nitrogen availability. Oecologia, 134, 547–553.
Smith SD, Huxman TE, Zitzer SF et al. (2000) Elevated CO2 increases
productivity and invasive species success in an arid ecosystem. Nature,
408, 79–82.
Titus JH, Nowak RS, Smith SD (2002) Soil resource heterogeneity in the
Mojave Desert. Journal of Arid Environments, 52, 269–292.
Tjoelker MG, Oleksyn J, Reich PB (1999) Acclimation of respiration
to temperature and CO2 in seedlings of boreal tree species in rela-
tion to plant size and relative growth rate. Global Change Biology, 49,
679–691.
Vitousek PM, Mooney HA, Lubchenco J, Melillo JM (1997) Human
domination of Earth’s ecosystems. Science, 277, 494–499.
Volder A, Smart DR, Bloom AJ, Eissenstat DM (2005) Rapid decline in
nitrate uptake and respiration with age in fine lateral roots of grape:
implications for root efficiency and competitive effectiveness. New
Phytologist, 165, 493–502.
Weatherly HE, Zitzer SF, Coleman JS, Arnone JA III (2003) In situ litter
decomposition and litter quality in a Mojave Desert ecosystem: effects
of elevated CO2 and interannual climate variability. Global Change
Biology, 9, 1223–1233.
Yoder CK, Vivin P, Defalco LA, Seemann JR, Nowak RS (2000) Root
growth and function of three Mojave Desert grasses in response
to elevated atmospheric CO2 concentration. New Phytologist, 145,
245–256.
E L E VA T E D C O 2 A N D D E S E R T S H R U B R O O T R E S P I R AT I O N 1575
r 2009 Blackwell Publishing Ltd, Global Change Biology, 16, 1566–1575
Copyright © 2022 FDOKUMEN




















