The effects of altered maxillary growth on patterns of mandibular rotation in a pig model
8
The effects of altered maxillary growth on patterns of mandibular rotation in a pig model Nathan E. Holton a,b, * , Christina L. Nicholas b,c , Steve D. Marshall a , Robert G. Franciscus b , Thomas E. Southard a a Department of Orthodontics, The University of Iowa, Iowa City, IA 52242, USA b Department of Anthropology, The University of Iowa, Iowa City, IA 52242, USA c Dows Research Institute, The University of Iowa, Iowa City, IA 52242, USA 1. Introduction A significant proportion of variation in human cranioman- dibular form is arrayed along the vertical dimensions of the facial skeleton. 1–3 As an important contributing factor, the height of the lower facial skeleton is markedly influenced by patterns of mandibular rotation during ontogeny. The mor- phogenetic influences on mandibular rotation are complex and multifactorial as evidenced by the various rotational patterns documented in classic implant studies. 4 In more general terms however, mandibular rotation can be divided a r c h i v e s o f o r a l b i o l o g y 6 0 ( 2 0 1 5 ) 9 3 3 – 9 4 0 a r t i c l e i n f o Article history: Accepted 21 February 2015 Keywords: Mandibular rotation Integration Sus scrofa a b s t r a c t Objectives: A thorough understanding of influence of maxillary growth on patterns of man- dibular rotation during development is important with regard to the treatment of skeletal discrepancies. In the present study, we examined whether experimentally altered maxillary position has a significant influence on patterns of mandibular rotation in a pig model. Design: Maxillary growth was altered in a sample of n = 10 domestic pigs via surgical fixation of the circummaxillary sutures. We compared the experimental group to control and surgical sham samples and assessed the effects of altered maxillary growth on mandibular form using geometric morphometric techniques. We tested for significant differences in mandibular shape between our samples and examined axes of morphological variation. Additionally, we examined whether altered mandibular shape resulting from altered maxillary position was predictably associated with morphological changes to the condylar region. Results: There was a statistically significant difference in mandibular shape between the experimental and control/sham groups. As a result of vertical displacement of the snout, mandibles in the experimental sample resulted in greater anterior rotation when compared to the control/sham pigs. Variation in rotation was correlated with morphological changes in the condyle including the shape of the articular surface and condylar orientation indicative of greater anterior mandibular rotation. Conclusions: Vertical displacement of the maxilla had a significant effect on mandibular shape by encouraging anterior mandibular rotation. This result has important implications for understanding the effects of altered mandibular posture on condylar remodeling the treat- ment of skeletal discrepancies such as the correction of hyperdivegent mandibular growth. # 2015 Elsevier Ltd. All rights reserved. * Corresponding author at: Department of Orthodontics, The University of Iowa, Iowa City, IA 52242, USA. Tel.: +1 319 384 4786. E-mail address: [email protected] (N.E. Holton). Available online at www.sciencedirect.com ScienceDirect journal homepage: http://www.elsevier.com/locate/aob http://dx.doi.org/10.1016/j.archoralbio.2015.02.019 0003–9969/# 2015 Elsevier Ltd. All rights reserved.
Transcript of The effects of altered maxillary growth on patterns of mandibular rotation in a pig model

The effects of altered maxillary growth on patternsof mandibular rotation in a pig model
Nathan E. Holton a,b,*, Christina L. Nicholas b,c, Steve D. Marshall a,Robert G. Franciscus b, Thomas E. Southard a
aDepartment of Orthodontics, The University of Iowa, Iowa City, IA 52242, USAbDepartment of Anthropology, The University of Iowa, Iowa City, IA 52242, USAcDows Research Institute, The University of Iowa, Iowa City, IA 52242, USA
a r c h i v e s o f o r a l b i o l o g y 6 0 ( 2 0 1 5 ) 9 3 3 – 9 4 0
a r t i c l e i n f o
Article history:
Accepted 21 February 2015
Keywords:
Mandibular rotation
Integration
Sus scrofa
a b s t r a c t
Objectives: A thorough understanding of influence of maxillary growth on patterns of man-
dibular rotation during development is important with regard to the treatment of skeletal
discrepancies. In the present study, we examined whether experimentally altered maxillary
position has a significant influence on patterns of mandibular rotation in a pig model.
Design: Maxillary growth was altered in a sample of n = 10 domestic pigs via surgical fixation of
the circummaxillary sutures. We compared the experimental group to control and surgical
sham samples and assessed the effects of altered maxillary growth on mandibular form using
geometric morphometric techniques. We tested for significant differences in mandibular
shape between our samples and examined axes of morphological variation. Additionally, we
examined whether altered mandibular shape resulting from altered maxillary position was
predictably associated with morphological changes to the condylar region.
Results: There was a statistically significant difference in mandibular shape between the
experimental and control/sham groups. As a result of vertical displacement of the snout,
mandibles in the experimental sample resulted in greater anterior rotation when compared
to the control/sham pigs. Variation in rotation was correlated with morphological changes
in the condyle including the shape of the articular surface and condylar orientation
indicative of greater anterior mandibular rotation.
Conclusions: Vertical displacement of the maxilla had a significant effect on mandibular shape
by encouraging anterior mandibular rotation. This result has important implications for
understanding the effects of altered mandibular posture on condylar remodeling the treat-
ment of skeletal discrepancies such as the correction of hyperdivegent mandibular growth.
# 2015 Elsevier Ltd. All rights reserved.
Available online at www.sciencedirect.com
ScienceDirect
journal homepage: http://www.elsevier.com/locate/aob
1. Introduction
A significant proportion of variation in human cranioman-
dibular form is arrayed along the vertical dimensions of the
facial skeleton.1–3 As an important contributing factor, the
* Corresponding author at: Department of Orthodontics, The UniversiE-mail address: [email protected] (N.E. Holton).
http://dx.doi.org/10.1016/j.archoralbio.2015.02.0190003–9969/# 2015 Elsevier Ltd. All rights reserved.
height of the lower facial skeleton is markedly influenced by
patterns of mandibular rotation during ontogeny. The mor-
phogenetic influences on mandibular rotation are complex
and multifactorial as evidenced by the various rotational
patterns documented in classic implant studies.4 In more
general terms however, mandibular rotation can be divided
ty of Iowa, Iowa City, IA 52242, USA. Tel.: +1 319 384 4786.

a r c h i v e s o f o r a l b i o l o g y 6 0 ( 2 0 1 5 ) 9 3 3 – 9 4 0934
into two categories, i.e., forward rotation and backward
rotation, which are tied to a larger integrated suite of
morphological features of the mandible.2–4
Variation in mandibular rotation is influenced by the
amount and direction of mandibular condylar growth during
development.5,6 Increased anterior growth at the condyles
produces greater forward mandibular rotation whereas
greater posterior condylar growth results in a greater degree
of backward mandibular rotation.4
While numerous factors likely affect the direction of
condylar growth, variation in mandibular posture has a
significant influence.7 Changes in mandibular posture and
thus condylar position alter the biomechanical environment
of the temporomandibular joint thereby affecting the growth
of the condylar cartilage.8 While the precise influence of
altered mandibular posture on condylar growth is incom-
pletely understood, experimental studies have demonstrated
the effects of postural variation on the condylar growth and
resulting gross phenotypic changes in mandibular form.9,10
For example, posterior relocation of the glenoid fossa relative
to the mandibular condyle in a rabbit model results in
significant morphological changes in the shape of the condylar
surface and an increase in mandibular length.11,12 Sugiyama
et al.13 found that altering the vertical relationship between
the maxilla and mandible using a plate bonded to the
maxillary molars in a rat model resulted in altered condylar
growth associated with greater posterior mandibular rotation.
In a similar fashion, Ferrari and Herring14 examined the effects
of bite blocks on craniomandibular growth using a sample of
miniature pigs. In addition to a number of morphological
changes in the maxillary region, the mandible exhibited a
pattern that suggests increased posterior rotation including a
larger gonial angle (see,14 Fig. 2) as well as significant changes
to the condylar surface.
Whereas many experimental studies have examined the
influence of functional appliances that encourage posterior
condylar growth and thus backward mandibular rotation, less
is understood about the morphological effects of develop-
mental modifications that encourage anterior rotation of the
mandible. This is particularly important from an orthodontic
perspective given that treatment of hyperdivergent patients
with retrognathic mandibles via functional appliances
achieves dental correction but is unable to correct associated
vertical and sagittal skeletal discrepancies.15 Moreover,
maxillary impaction surgery producing autorotation of the
mandible in growing patients does not appear to inhibit
mandibular growth or affect the long-term rotational pattern
of the mandible.16 A limited number of studies, however, have
documented that anterior mandibular rotation induced by
molar intrusion results in favorable skeletal changes in
hypodivergent patients such as increased chin projection,
reduced facial height and a decreased gonial angle.15,17,18
In the present study, we examined whether experimentally
altered maxillary position has a significant influence on
patterns of mandibular rotation in a pig model. This is a
continuation of our research examining the influence of rigid
plate fixation of the circummaxillary sutures on facial growth
and development.17–19 Previously we found that sutural
fixation affects maxillary morphology such that pigs with
restricted sutural growth exhibited shorter and more dorsally
rotated (i.e., superiorly displaced) snouts. Using geometric
morphometric techniques, we report how altered vertical
maxillary position affects the growth of the mandible by
addressing the following research questions. First, to what
degree does variation in vertical maxillary position affect the
pattern of mandibular rotation in our experimentally modified
pigs when compared to normal, non-experimentally modified
sample of control pigs? Second, if the pattern of rotation is
altered as a result of the vertical displacement of the maxilla,
how is this pattern reflected in the morphology of the condylar
region?
2. Materials and methods
Ten female Sus scrofa sibship cohorts, each consisting of three
individuals, were allocated to one of three trial groups (i.e.,
experimental, sham and control). Surgical procedures are
described in detail in Holton et al.18 Briefly, in the experimental
group (n = 10), rigid miniplates were bilaterally affixed across
the zygomaxillary, frontonasal and nasomaxillary sutures at
two months of age. The sham group (n = 10) underwent an
identical surgical procedure but received only microscrew
implantation. The control group (n = 10) underwent no surgical
procedure. All animals were euthanized following four months
of post-surgical growth (i.e., six months of age). One control pig
did not survive to age 6 months, and one sham pig was
damaged during post-mortem processing resulting in a total of
n = 28 pigs available for analysis (i.e., n = 10 control, n = 9 sham
and n = 9 control). Since there were no significant differences
between our control and sham pigs, these were combined into
a single control group for all analyses. The University of Iowa
Institutional Animal Care and Use Committee approved all
procedures. For comparison, CT images illustrating the gross
morphological differences between an experimental pig and a
control pig are found in Fig. 1.
To assess variation in mandibular form, we collected a
series of k = 25 two-dimensional traditional and semiland-
marks from CT images that were imported into tpsDIG 2.12
software.20 We selected landmarks that adequately repre-
sented the mandibular corpus, mandibular ramus and
condylar process including the articular surface of the condyle
(Fig. 2). All landmarks were superimposed and scaled using
generalized Procrustes analysis (GPA). This method translates
objects to a common centroid origin, scales for size, and
rotates objects using a least-squares criterion. In the case of
our semilandmarks, a sliding semilandmark analysis was also
conducted (as a component of the GPA). Sliding semilandmark
analysis superimposes the semilandmarks by sliding them
along the curve to a consensus position that minimizes the
summed squared deviations for each given landmark across
the sample. This renders the semilandmarks homologous,
allowing semilandmarks henceforth to be treated the same as
standard landmarks in further analyses.21
With regard to our first research question, (i.e., effect of
maxillary variation on mandibular rotation), we first tested for
significant differences in overall mandibular shape between
our experimental and control trial groups using Procrustes
ANOVA. Next, we used principal components analysis (PCA) of
Procrustes scaled shape variables to examine axes of variation
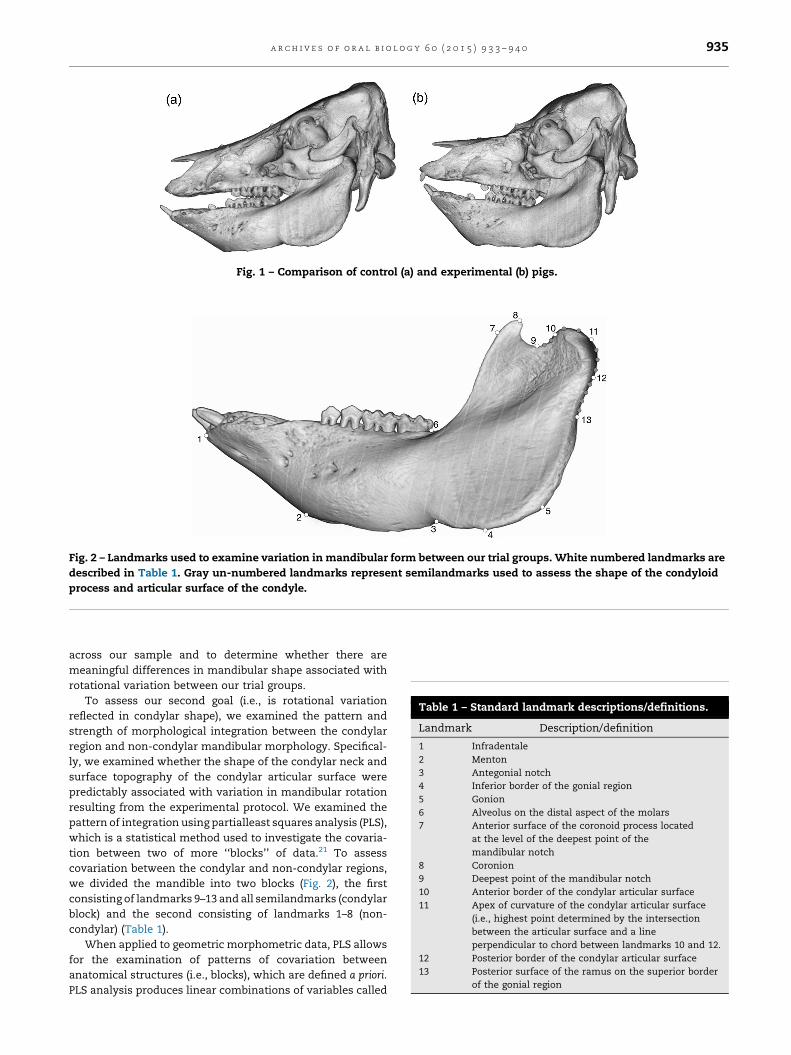
Fig. 1 – Comparison of control (a) and experimental (b) pigs.
Fig. 2 – Landmarks used to examine variation in mandibular form between our trial groups. White numbered landmarks are
described in Table 1. Gray un-numbered landmarks represent semilandmarks used to assess the shape of the condyloid
process and articular surface of the condyle.
Table 1 – Standard landmark descriptions/definitions.
Landmark Description/definition
1 Infradentale
2 Menton
3 Antegonial notch
4 Inferior border of the gonial region
5 Gonion
6 Alveolus on the distal aspect of the molars
7 Anterior surface of the coronoid process located
at the level of the deepest point of the
mandibular notch
8 Coronion
9 Deepest point of the mandibular notch
10 Anterior border of the condylar articular surface
11 Apex of curvature of the condylar articular surface
(i.e., highest point determined by the intersection
between the articular surface and a line
perpendicular to chord between landmarks 10 and 12.
12 Posterior border of the condylar articular surface
13 Posterior surface of the ramus on the superior border
of the gonial region
a r c h i v e s o f o r a l b i o l o g y 6 0 ( 2 0 1 5 ) 9 3 3 – 9 4 0 935
across our sample and to determine whether there are
meaningful differences in mandibular shape associated with
rotational variation between our trial groups.
To assess our second goal (i.e., is rotational variation
reflected in condylar shape), we examined the pattern and
strength of morphological integration between the condylar
region and non-condylar mandibular morphology. Specifical-
ly, we examined whether the shape of the condylar neck and
surface topography of the condylar articular surface were
predictably associated with variation in mandibular rotation
resulting from the experimental protocol. We examined the
pattern of integration using partialleast squares analysis (PLS),
which is a statistical method used to investigate the covaria-
tion between two of more ‘‘blocks’’ of data.21 To assess
covariation between the condylar and non-condylar regions,
we divided the mandible into two blocks (Fig. 2), the first
consisting of landmarks 9–13 and all semilandmarks (condylar
block) and the second consisting of landmarks 1–8 (non-
condylar) (Table 1).
When applied to geometric morphometric data, PLS allows
for the examination of patterns of covariation between
anatomical structures (i.e., blocks), which are defined a priori.
PLS analysis produces linear combinations of variables called

Fig. 3 – Scatter plot of individual principal component
scores for the first two principal components. White
circles = experimental pigs; gray circles = control pigs.
a r c h i v e s o f o r a l b i o l o g y 6 0 ( 2 0 1 5 ) 9 3 3 – 9 4 0936
singular axes, which maximize covariation between the
blocks of data. The strength of the covariation between the
singular axes is measured as the correlation (r) of individual
singular warp scores along the singular axes. We additionally
assessed the strength of the relationship between the two
blocks using the RV coefficient,22,23 which is a multivariate
analogue of the coefficient of determination (r2). Statistical
significance for both the PLS analysis and RV coefficient were
assessed using permutation tests (n = 999) to determine
whether the strength of the relationship between the two
blocks using both measures is greater than would be expected
to occur by chance.21,24 All analyses were conducted using the
Geomorph package in R.25
Fig. 5 – Superimpositions of mean experimental (black) and me
superimposed along condylar landmarks. (b) Mean shapes sup
Fig. 4 – Thin-plate splines illustrating morphological variation a
the positive end of PC1 is shown in (a) while shape variation a
3. Results
There was a statistically significant difference in mandibular
shape between the experimental and control groups as
indicated by the result of the Procrustes ANOVA (ss = 0.0078;
df = 1; P = 0.01). Mandibular shape differences between trial
groups were also reflected in the results of the PCA. The
experimental and control groups were largely separated along
PC1 (26.9%) although there was some overlap especially with
regard to one control pig that fell at the extreme end of the
experimental trial group’s morphospace (Fig. 3). PC1 primarily
described variation in the angular relationship between the
mandibular ramus and corpus. As a result of the surgically
induced dorsal orientation of the snout, mandibular shape in
the experimental group tended to exhibit a pattern consistent
with greater anterior mandibular rotation. As evidenced by the
thin-plate spline in Fig. 4a, the experimental pigs were
characterized by anterior displacement of mandibular ramus
landmarks (including the coronoid and condyloid processes)
and superior displacement of the anterior corpus landmarks.
This resulted in a more acute gonial angle and a relative
reduction in the overall anterior–posterior dimensions of the
mandible. The control pigs (Fig. 4b), in contrast, exhibited a
mandibular shape consistent with relatively less anterior
mandibular rotation. In particular, the control pigs were
characterized by a more posteriorly oriented ramus and an
inferior displacement of the anterior corpus resulting in a
larger gonial angle and relatively longer anterior–posterior
mandibular dimensions.
Morphological variation between the experimental and
control groups is further illustrated in Fig. 5 in which trial
group mean shapes have been superimposed at the mandibu-
lar condyle (Fig. 5a) and mandibular plane (Fig. 5b). We note
an control (gray) mandibular shapes. (a) Mean shapes
erimposed along the mandibular plane.
long the first principal component. Mandibular shape along
long the negative end of PC1 is shown in (b).

a r c h i v e s o f o r a l b i o l o g y 6 0 ( 2 0 1 5 ) 9 3 3 – 9 4 0 937
that unlike the Procrustes superimposition used for the
statistical analyses, these superimpositions are used for
illustrative purposes only (i.e., they were not used for
quantitative analysis). When superimposed at the mandibular
condyle, the mean experimental mandibular shape clearly
exhibits a dorsal rotation of the corpus following the dorsal
reorientation of the experimental trial group pigs. In addition,
there is an inferior displacement of the gonial region relative
to superior ramus. When superimposed along the mandibular
plane, the mandibular ramus of the mean experimental shape
is anteriorly displaced and the condylar region is more
vertically oriented when compared to the oblique orientation
of the mean control shape.
The results of our PLS analysis (Fig. 6a and b) reveal that
there is a strong significant correlation between the morphol-
ogy of the condylar and non-condylar regions across our
experimental and control trial groups (r = 0.90; P < 0.001). The
strength of the pattern of covariation between the two blocks
is further evident from the RV coefficient (RV = 0.53; P < 0.001)
which falls to the far right of the distribution of RV coefficients
calculated from randomly permuted landmark combinations,
indicating that the two blocks are integrated.
Correlated patterns of morphological variation along the
first singular axes are illustrated in Fig. 6c and d. These results
are similar to the morphological variation arrayed along PC1.
As the mandibular ramus becomes more vertically oriented
relative to the mandibular corpus, resulting in a smaller gonial
Fig. 6 – Results of the PLS analysis. (a) Scatter plot of PLS scores
(block 2). White circles = experimental pigs; gray circles = contro
correlations. The arrow indicates the correlation of the condyla
illustrating patterns of morphological covariation between the c
(black landmarks).
angle, there is a tendency for the condylar process to become
more anteriorly displaced. Moreover, the condylar articular
surface is rounded indicating greater vertical positioning of
the superior aspect of the condyle. In contrast, a more oblique
ramus relative to corpus is associated with a more posteriorly
displaced condylar process with a flattened articular surface
that is similarly posteriorly displaced.
4. Discussion and conclusions
The effects of altered mandibular posture via protrusive
functional appliances are well documented in both human
subjects,6,26 and experimental animal models.9,10,13,14 These
studies have shown that mandibular protrusion results in
altered mandibular condylar development and greater posteri-
or rotation of the mandible. However, despite the large number
of studies that have focused on the effects of protrusion, far
fewer studies have examined growth modifications that
encourage anterior rotation of the mandible. In the present
study, we examined the influence of experimentally altered
mandibular posture on mandibular form via superior reposi-
tioning of the maxilla in a pig model. Pigs are well regarded as
the best non-primate model for the human temporomandibular
joint,26–28 and as such, the results from our analysis are
particularly relevant to understanding mandibular growth
dynamics in humans. Ultimately, the results of our analysis
for the corpus/ramus (block 1) and the condyloid process
l pigs. (b) Distribution of randomly permuted PLS
r and non-condylar blocks. (c and d) Thin-plate splines
ondylar block (gray landmarks) and non-condylar block

a r c h i v e s o f o r a l b i o l o g y 6 0 ( 2 0 1 5 ) 9 3 3 – 9 4 0938
are important for a more thorough understanding of the
influence of mandibular posture on patterns of rotation, the
potential utility of applied anterior mandibular rotation for the
correction of hyperdivergent mandibular growth, and a broader
understanding of the integrative effects of maxillary form on
the growth of the mandible.
The most apparent changes in mandibular form associated
with superior displacement of the maxilla were found in the
gonial region. The morphological differences between our
experimental and control trial groups were largely consistent
with the suite of mandibular features commonly associated
with anterior and posterior mandibular rotation in human
subjects.2,3,29 As a result of maxillary growth modification,
there was a considerable reduction in the gonial angle in the
experimental trial group. This resulted from the dorsal
reorientation of the mandibular corpus relative to the ramus
and mirrored the reorientation of the snout documented in
our previous analysis.18 Coupled with the reorientation of the
snout and reduced gonial angle, there was a relative increase
in mandibular ramus height. This is evident in the patterns of
thin-plate spline deformation in the inferior gonial region, and
in the inferior displacement of the lower border of the gonial
region relative to the condyles when superimposing the mean
experimental and control trial group shapes at the condyles.
This result is consisted with Sugiyama et al.13 who found that
protrusive appliances resulted in a reduction in mandibular
ramus height in their rat model.
Increased anterior rotation of the mandible in our
experimental trial group was significantly correlated with
morphological changes in the condylar region. The results of
our partial least squares analysis indicate that anterior
mandibular rotation is associated with a more vertically
oriented condyloid process. This is evident both in the position
of the condyloid process relative to the mandibular ramus and
the shape of the condylar articular surface. This suggests that
altered mandibular posture associated with superior displace-
ment of the maxilla may have resulted in greater vertical
condylar growth in contrast to the control sample to produce
the altered mandibular morphology. In general, this result
follows previous experimental animal studies that have
documented significant adaptive chances in condylar growth
and remodeling via altered mandibular posture using protru-
sive appliances.9,10,13,30
The effects of altered maxillary position on the morphology
of the mandibular condyle suggest that the experimentally
induced change in mandibular posture had a significant effect
on the mechanical environment of the temporomandibular
joint8 resulting in a change in condylar shape. Indeed, an
alteration in the mechanical environment of the condylar
region resulting from distraction osteogenesis also results in
an alteration in the shape of articular surface of the
condyle.31,32 Rafferty et al.32 documented in a pig model that
the distraction side condyle exhibited a decrease and
reorientation in mechanical strain. This was associated with
greater convexity of the condyle resulting from an increase in
mineralization in the central region of the articular surface.
Previous studies have documented that variation in man-
dibular posture has a significant influence on the anterior–
posterior dimensions of the mandible. This is evidenced
by increased mandibular length resulting from the use of
protrusive appliances in both experimental animal mod-
els14,33,34 and human samples.35 In a similar fashion our
experimental sample, with increase anterior rotation exhibited
a relative reduction in mandibular length as evidenced by thin-
plate spine deformation in our principal components and
partial least squares analyses. The effects of altered posture on
mandibular length have lead researchers to suggest that
variation in condylar position has a significant influence on
the rates of growth the condylar cartilages.33,34 Araujo et al.6
have argued, however, that mandibular length variation is more
likely the result of the direction of condylar growth rather than
changes in the overall magnitude. Given the nature of the
sample used in the present study (i.e., static vs. longitudinal
comparisons) we are unable to directly assess whether reduced
mandibular length is the result of a reduction in growth at the
condyle. Nevertheless, the reduction in mandibular length in
our experimental sample is at least partly a function of shape
differences associated with the reorientation of the corpus and
reduced gonial angle. As such, this result is consistent with the
conclusions drawn by Araujo et al.6
The changes in mandibular morphology in our experimen-
tal trial group suggest that orthodontic treatments designed to
encourage anterior rotation of the mandible may be useful for
skeletal correction of hyperdivergent patients. The use of
protrusive appliances and headgear normally used to treat
hyperdivergent patients often produces changes in dental
arch relationships but does not correct increased posterior
mandibular rotation associated with hyperdivergent mor-
phology.15,36 Treatments that encourage anterior mandibular
rotation, on the other hand, are expected to increase vertical
condylar growth thereby resulting in skeletal correction of
hyperdivergent mandibular growth. Buschang et al.15 for
example, have shown that molar intrusion using miniscrew
implants in Class II hyperdivergent patients can produce
favorable skeletal changes in the mandible during treatment.
This includes increased vertical condylar growth, a reduction
in the gonial angle and a flatter mandibular plane. The results
of our analysis provide important experimentally derived
evidence that further underscores the influence of altered
mandibular posture on encouraging anterior rotation of the
mandible.
The results of our analysis indicate that experimentally
altered growth in the maxillary region via ridged plate fixation
of the circummaxillary sutures has a significant influence on
the growth of both the anterior and posterior aspects of the
mandible. While our understanding of the patterns of
integration between the cranium and mandible are incom-
plete, generally speaking, previous researchers have docu-
mented a relative modularity of the anterior and posterior
mandibular regions both animal models37,38 and in humans39.
Following Enlow’s Counterpart Principle,2 the mandibular
corpus forms the inferior component of the anterior facial
column and as such, its form is influenced by the integrated
growth of the ethmomaxillary complex. Variation in the
mandibular ramus, on the other hand, is influenced to a
greater degree by the development of the posterior facial
column. Indeed, recent studies have underscored the close
morphological relationship between the mandibular ramus
and middle cranial fossa suggesting that these structures form
an integrated ‘‘petroso-mandibular unit.’’40 In spite of the

a r c h i v e s o f o r a l b i o l o g y 6 0 ( 2 0 1 5 ) 9 3 3 – 9 4 0 939
relative modularity of the anterior and posterior regions of the
mandible, our results underscore that morphological variation
in the maxillary region results in significant morphological
changes across the anterior and posterior components of the
mandible. As such, while the there is a level of modularity
between the corpus and ramus, morphological changes that
affect the posture of the mandible have a more global effect on
mandibular form.
Ultimately, a more thorough understanding of the devel-
opmental relationship between the cranium and mandible are
integral for understanding the precise influence of larger
cranial growth dynamics on mandibular form. This is
particularly important given that recent work by Alcaron
et al.29 found that the strength and pattern of cranial–
mandibular integration varies with facial type (i.e., dolicho-
facial vs. brachyfacial). Moreover, their results indicate that
morphological integration between the cranium and mandible
has a strong influence on the manifestation of skeletal
malocclusion. Nevertheless, a significant amount of variation
in skeletal malocclusion is independent of cranial–mandibular
integration. This has significant implications for the success-
ful treatment of skeletal discrepancies using orthopedic
appliances given that the response to treatment may be
affected by the relative level of cranial and mandibular
integration. As such, a greater understanding of the influence
of maxillary growth on mandibular form is important with
regard to the etiology of skeletal discrepancies, especially
given that skeletal discrepancies are predictably associated
with patterns of mandibular rotation.
Funding
This research was funded by an internal University of Iowa
OVPR BSFP Developmental Biology Grant #50255004.
Competing interest
None declare.
Ethical approval
Approval was given by The University of Iowa Institutional
Animal Care and Use Committee.
Acknowledgements
We would like to thank the editor and reviewer for their
helpful comments regarding this manuscript. This research
was funded by a University of Iowa OVPR BSFP Developmental
Biology Grant.
r e f e r e n c e s
1. Sassouni V, Nanda S. Analysis of dentofacial verticalproportions. Am J Orthod 1964;50(11):801–23.
2. Enlow DH, Hans MG. Essentials of facial growth. Philadelphia/London/New York: W.B. Saunders Company; 1996.
3. Bastir M, Rosas A. Facial heights: evolutionary relevance ofpostnatal ontogeny for facial orientation and skullmorphology in humans and chimpanzees. J Hum Evol2004;47(5):359–81.
4. Bjork A. Prediction of mandibular growth rotation. Am JOrthod 1969;55(6):585–99.
5. Buschang PH, Gandini LG. Mandibular skeletal growth andmodeling between 10 and 15 years of age. Eur J Orthod2002;24(1):69–79.
6. Araujo AM, Buschang PH, Melo ACM. Adaptive condylargrowth and mandibular remodeling changes with bionatortherapy – an implant study. Eur J Orthod 2004;26(5):515–22.
7. Buschang PH, Jacob H, Carrillo R. The morphologicalcharacteristics, growth, and etiology of the hyperdivergentphenotypes. Sem Orthod 2013;19(4):212–26.
8. Owtad P, Park JH, Shen G, Potres Z, Darendeliler MA. Thebiology of TMJ growth modification: a review. J Dent Res2013;92(4):315–21.
9. Woodside DG, Altuna G, Harvold E, Herbert M, Metaxas A.Primate experiments in malocclusion and bone induction.Am J Orthod 1983;83(6):460–8.
10. McNamara JA, Bryan FA. Long-term mandibular adaptationsto protrusive function: an experimental study in Macacamulatta. Am J Orthod 1987;92(2):134–44.
11. Kantomaa T. Effect of increased posterior displacement ofthe glenoid fossa on mandibular growth: a methodologicalstudy on the rabbit. Eur J Orthod 1984;6(1):15–24.
12. Pirttiniemi P, Kantomaa T, Tuominen M. Increased condylargrowth after experimental relocation of the glenoid fossa.J Dent Res 1993;72(9):1356–9.
13. Sugiyama H, Lee K, Imoto S, Sasaki A, Kawata T, YamaguchiT, et al. Influences of vertical occlusal discrepancies oncondylar responses and craniofacial growth in growing rats.Angle Orthod 1999;69(4):356–64.
14. Ferrari CS, Herring SW. Use of a bite-opening appliance inthe miniature pig: modification of craniofacial growth. ActaAnat 1995;154(3):205–15.
15. Buschang PH, Carrillo R, Rossouw PE. Orthopedic correctionof growing hyperdivergent, retrognathic patients withminiscrew implants. J Oral Maxillofac Surg 2011;69(3):754–62.
16. Mojdehi M, Buschang PH, English JD, Wolford LM.Postsurgical growth changes in the mandible of adolescentswith vertical maxillary excess growth pattern. Am J OrthodDentofac Orthop 2001;119(2):106–16.
17. Southard TE, Franciscus RG, Fridrich KL, Nieves MA, KellerJC, Holton NE, et al. Restriction of facial bone growth usingskeletal fixation: a preliminary study. Am J Orthod DentofacOrthop 2006;130(2):218–23.
18. Holton NE, Franciscus RG, Nieves MA, Marshall SD, ReimerSB, Marshall SD, et al. Sutural growth modification in Susscrofa as a model for the evolution of modern humancraniofacial anatomy. J Anat 2010;216(1):48–61.
19. Holton NE, Franciscus RG, Marshall SD, Southard TE, NievesMA. Nasal septal and premaxillary developmentalintegration: Implications for facial reduction in genus Homo.Anat Rec 2011;294(1):68–78.
20. Rohlf FJ. TPSDig2 version 2.16. NY: Department of Ecologyand Evolution, State University of New York at Stony Brook;2010.
21. Zelditch ML, Swiderski DL, Sheets HD, Fink WL. Geometricmorphometrics for biologists: a primer. New York: ElsevierAcademic Press; 2012.
22. Escoufier Y. Le traitement des variables vectorielles.Biometrics 1973;29(4):751–60.
23. Klingenberg CP. Morphometric integration and modularityin configurations of landmarks: tools for evaluating a priorihypotheses. Evol Dev 2009;11(4):405–21.

a r c h i v e s o f o r a l b i o l o g y 6 0 ( 2 0 1 5 ) 9 3 3 – 9 4 0940
24. Rohlf FJ, Corti M. Use of two-block partial least-squares tostudy covariation in shape. Syst Biol 2000;49(4):740–53.
25. Adams D, Otarola-Castillo E. Package ‘geomorph’: geometricmorphometric analysis of 2D/3D landmark data. 2012.
26. Scheman P. Anthropoid comparisons of the anatomy ofexternal pterygoid muscles of the fetal and adult domesticpig. J Dent Res 1967;46(6):1337–43.
27. Bermejo A, Gonzalez O, Gonzalez JM. The pig as an animalmodel for experimentation on the temporomandibulararticular complex. Oral Surg Oral Med Oral Pathol1993;75(1):18–23.
28. Herring SW. Animal models and temporomandibulardisorders: how to choose. In: Sesle BJ, Bryant PS, Dionne RA,editors. Temporomanidbular disorders and related painconditions. Seattle: IASP Press; 1985. p. 323–8.
29. Alarcon JA, Bastir M, Garcıa-Espona I, Menendez-Nunez M.Morphological integration of mandible and cranium:orthodontic implications. Arch Oral Biol 2014;59(1):22–9.
30. Petrovic AG, Stutzman J, Oudet C. Control processes in thepostnatal growth of the mandible. In: McNamara JA, editor.Determinants of mandibular form and growth. Monograph No 4.Ann Arbor: Center for Human Growth and Development,University of Michigan; 1975. p. 101–54.
31. Thurmuller P, Troulis MJ, Rosenberg A, Kaban LB. Changesin the condyle and disc in response to distractionosteogenesis of the minipig mandible. J Oral Maxillofac Surg2002;60(11):1327–33.
32. Rafferty KL, Sun Z, Egbert M, Bakko DW, Herring SW.Changes in growth and morphology of the condylefollowing mandibular distraction in minipigs: overloadingor underloading? Arch Oral Biol 2007;52(10):967–76.
33. McNamara JA, Howe RP, Dischinger TG. A comparison ofthe Herbst and Frankel appliances in the treatment ofClass II malocclusion. Am J Orthod Dentofac Orthop1990;98(2):134–44.
34. Toth LR, McNamara JA. Treatment effects produced by thetwin-block appliance and the FR-2 appliance of Frankelcompared with an untreated Class II sample. Am J OrthodDentofac Orthop 1999;116(6):597–609.
35. Op Heij DG, Callaert H, Opdebeeck HM. The effect of theamount of protrusion built into the bionator on condylargrowth and displacement: a clinical study. Am J OrthodDentofac Orthop 1989;95(5):401–9.
36. LaHaye MB, Buschang PH, Alexander RG, Boley JC.Orthodontic treatment changes of chin position in Class IIdivision 1 patients. Am J Orthod Dentofac Orthop2006;130(6):732–41.
37. Atchley WR, Hall BK. A model for development andevolution of complex morphological structures. Biol Rev1991;66(2):101–57.
38. Klingenberg CP, Mebus K, Auffray J-C. Developmentalintegration in a complex morphological structure: howdistinct are the modules in the mouse mandible? Evol Dev2003;5(5):522–31.
39. Wellens HLL, Kuijpers-Jagtman Halazonetis DJ. Geometricmorphometric analysis of craniofacial variation, ontogenyand modularity in a cross-sectional sample of modernhumans. J Anat 2013;222(4):397–409.
40. Bastir M, Rosas A, Kuroe K. Petrosal orientation andmandibular ramus breadth: evidence for an integratedpetroso-mandibular developmental unit. Am J PhysAnthropol 2004;123(4):340–50.