
Histamine, Histamine Receptors, and their Role in Immunomodulation: An Updated Systematic Review
Upload
iberoamericanaCategory
view
0download
0
THE DISTRIBUTION OF HISTAMINE H,-RECEPTORS IN THE RAT BRAIN: AN AUTORADIOGRAPHIC
STUDY
J. M. PALACIOS. J. K. WAMSLE~ and M. J. KUHAR
Departments of Pharmacology and Experimental Therapeutics and Psychiatry and Behavioral Sciences.
Johns Hopkins University School of Medicine. Baltimore. Maryland 21205. U.S.A.
Abstract--The localization of histamine H,-receptors in the rat braln was studied at the light micro-
scopic level by quantitative autoradiography. Receptors were labeled i/l vitro with [3H]mepyramine. The
autoradiograms revealed a widespread distribution of these receptors throughout the brain. Areas with
high receptor concentrations are: the bed nucleus of the stria terminalis. polymorphic layer of the hilus
of the area dentata. ventromedial nuclei of the hypothalamus. pontine nuclei and other nuclei in the
pans. the nucleus tractus solitarii and the dorsal motor nucleus of the vagus nerve. Possible relationship!,
of the distributions of histamine H,-receptors with (a) specific anatomical systems. (b) the known
distribution of histaminergic terminals and (c) the central pharmacological effects of histamine and
antihistaminics are discussed.
HISTAMINE is possibly a neurotransmitter in the mam- malian brain, and most of the biochemical com-
ponents of a putative histaminergic synapse have been
characterized in the rat brain (SCHWARTZ. 1977;
SCHWARTZ. BARRIN, GARBARG. LLORENS, PALACIOS &
POLLARD, 1978; GREEN, JOHNSON & WEINSTEIN, 1978;
SNYDER 8~ TAYLOR, 1972; SCHWARTZ. BARBI&,
BAUDRY, GARBARG, MARTRES & VERDIERE, 1979;
TAYLOR, 1975). In addition, histamine receptors of the
HI and Hz types are also present in brain (SCHWARTZ
et al., 1978; GREEN et al., 1978; SCHWARTZ, BARBIN,
DUCHEMIN, GARBARG, PALACIOS, QUACH & ROSE,
1980; SCHWARTZ, 1979).
The recent use of C3H]mepyramine has allowed di- rect binding studies of histamine HI-receptors. The
c3H]mepyramine binding sites in the brain (CHANG.
TRAN & SNYDER, 1978 ; HILL. EMSON & YOUNG, 1978;
TRAN. CHANC & SNYDER. 1978; CHANG, TRAN &
SNYDER. 1979a) and peripheral (HILL, YOUNG & MAR-
RIAN, 1977; CHANG. TRAN & SNYDER, 1979/1) tissues
have all the properties of a pharmacologically rele-
vent H, -receptor.
We have recently described a procedure that allows
the ‘in oitro’ labeling and subsequent autoradiographic
localization of neurotransmitter receptors in mounted
tissue sections (YOUNG & KUHAR. 1979a). The con- ditions for the autoradiographic localization of
HI-receptors using [3H]mepyramine have also been described (PALACIOS. YOI!NG & KUHAR, 1979). In this paper, we present the detailed anatomical distribution of H,-histamine receptors in the rat brain, the species
where most of the available neurochemical data on the histaminergic system has been obtained.
EXPERIMENTAL PROCEDURES
The overall procedure for the ‘;)I oifro’ labeling of recep-
tors and the specific conditions for the USC of [3H]mepyr-
amine have been described (Yorluc & KVHAR. iY79rr;
PALACIOS c’r crl.. 1979). Briefly, male Sprague-Da\\ Ic> rat5
(18@250 g) were killed under anesthesia with pentobarbital
(60 mg;kg) by perfusion with phosphate buffered salmc: containing 0. I”,, formaldehyde. The brains were removed.
the appropriate regions were mounted on microtomc
chucks and frozen in liquid nitrogen for storage. Six to ten
micron sections were cut on a Harris Cryostat Microtomc
and thaw mounted onto subbed microscope slides. Tissues
from at least 3 different animals were pooled in each e+
periment. The results reported in this stud) came from the
observation of tissue from X rats In 3 Independent e\perl-
men&. Human postmortem tissue samples were obtained
at autopsy from a 50 year-old male (3 h post mortem: sud-
den death with no drug history) and a 22 qear-old male
(4 h postmortem: stab wound with no recent drug hIstory)
in cooperabon with the Medical Examiner’s Olfice in BaItI-
more. Maryland. HI-histamine receptors were labeled as
described previously. Mounted tissues were mcubatcd in
sodium potassium phosphate buffer (0.3 M. pH 7.5, 2 4 C),
containing 5 nM C3H]mepyramine (28.3 Ci’mmol, New
England Nuclear Corp., Boston, Mass.). For obtaming
blank or non-specific binding, the incubation medium con-
tained also 2,uM triprolidine (Burroughs Wellcomc). The
incubation time was 40 min at ice-water bath temperature
At the end of the incubation. the tissues were rinsed hy ;I
rapid in and out dipping in buffer and a subsequent IO min
washing period in fresh buffer. Salts were eliminated by ;I
rapid dipping in water and the tissues dried rapid11 (~30 s) under a current of cold. dry air. For autoradiograph!.
emulsion-coated coverslips were attached with glut to one
end of the slide. The assemblies were protected \tlth a
square piece of Teflon and secured with paper chpb. The
emulsion-coated coverslips were allowed IO esposc for 6
weeks. after which the autoradiograms were developed and
tissues stained as previously descrlbcd (Yor 1~; 6i I<( IIAK.
1979a). Autoradiograms were observed and photographed
under both bright-field and dark-field illumination using
an Olympus JM (Olympus Optical Company. Jupm) microscope and a Leitz Ortholus (Leitr. Wctflar. (;er-
many) microscope equipped with a Hinsch-Goldman Bou
(Bunton Instrument Company. Rockville, Maryland.
U.S.A.). Slides were examined by two independent ob-
servers.

16 d. M. PALACIOS. .I K. WAWSll 1 and %I J. KI IIAK
, 1 r’ 12 4 8 16
‘H- MEPYRAMINE (nM)
FG. 1. Saturation of [‘H]mepyramine binding to rat brain mounted tissue sections. Ten micron sections (at the levels A 3CCU-A 4000 of the atlas of KONIG & KLIPPEL. 1963) were incubated with increasing [‘Hlligand concentrations as described in the text. Blank values were obtained by adding 2 nM triprolidine to the incubation. The points in the lines are the mean of three separate samples and the
experiment was replicated twice with similar results.
Grain denstttes were determmed by counting grams in a grid containing eyepiece using a loOX oil immersion ob~rc-
tivc on a Zeiss (Zeiss. West Germany) dual-viewing bmocu- lar microscope.
RESULTS
Characteristics oJ the binding of [‘H]mepyrumine to
rut bruin mounted tissue sections
Biochemical studies of the binding of [-‘H]mepyr-
amine to brain membranes from different species have
revealed considerable species differences not only in
the regional distribution but also in the characteristics
of the binding (CHANG el al., 1979~: PALAC‘IOS VI (I/..
1979). As our previous studies were done mostly in
guinea-pig tissues, the characteristics of the binding of
[‘Hlmepyramine to rat brain mounted tissue sections
were investigated. Figure I shows a typical saturation
curve of C3H]mepyramine to mounted rat brain
sections containing hypothalamus. hippocampus,
thakdmus and cortex. As reported for the binding to
membranes, the affinity of [3H]mepyramine for the
H,-receptor was lower in the rat brain (Ku approx
5 nM) than in the guinea-pig (K,, = 0.5 nM).
Autorudiographic studies
Virtually all areas of gray matter in the brain
showed a slight increase in grain density over that
seen in white matter areas. Thus increase could not be
identified on control slides (those incubated with
[ ‘H]mcpyraminc and triprolidine) and thus should be
regarded as specific binding sites for the H, antagon- ist. Areas containing concentrations of autoradio-
graphic grains were classified as follow~s
HI& (41 70 grams per 529 quart microns of
11sslle)
Modcratc (30 39 grains~j29 pm’ ) Low (20 29 grains329 pm’)
Very low I < 20 grains529 pm’)
C‘nntrol or displaced tissue UIUCS were consistently
less than IO grains/529 grn’.
C‘ortc,.\ c~rld hasa/ qanylitr. Lnmma IV of the cerebral
cortex vcas consistently labeled throughout the forc-
brain showing most prominently as a wide band in
lateral aspect or the cortex (Fig. 21. The grain density
in lamina IV varied from moderate in temporal areas
(Fig. 7) to IOU in other regions. .4 very low level of
grains uust slightly above Icv~l\ seen over white
matter) were spread evenly throughout the caudate-
putamen (Fig. 3). Moderate levels of grain densities
were seen in rostra1 regions of the forebrain just dor-
sal to the rhinal sulcus and 111 .I circular area just
ventral to the forceps minor (not shown). Moderate to
low levels of grains could bc Jsmonstrated in the
pyriform cortex and surrounding the claustrum
(Figs 3. 12). The receptor levels In these areas were
moderate in rostra1 regions of the Corebrain, becoming
progressively lower in more caudal areas.
Lirnhrt trnd oJfirc.ror!. urns. l‘hc bed nucleus of the
strta terminalis showed one 01 the highest grain
densities 111 portions above the anterior commissure.
while demonstrating only modcrate levels below the
lcvcl of this commissurc (Fig. 12). High levels of
receptors could also be seen in the nucleus of the
tractus diagonalis and the organum vasculosum
Iamina terminalis (not shown) low levels of auto-
radiographic grains could hc we:11 m the lateral septal
nucleus (Fig. I?). The hippocampus had a h1gl1 concentratron oi
rcccptors in its most rostra1 tip Proceeding caudally.
high levels of grains occupied only the polymorphic
and molecular layer ol’ CA3 extending into the poly-
morphic layer of the hilus of the area dentata (Fig. 6).
Large numbers of grains could also bc seen in the
ventral part of the subiculum (the grain density was
moderate dorsally) of the hippocampal formation and
III the prcsubicular cortex (Fig. :i. The umygdala had a wide \;uiation in receptor
dcnsitics. The medial amygdaloid nucleus showed
moderate grain densities rostrally and high levels in
caudal areas (Figs 4.7). The pars lateralis of the basal
amygdnloid nucleus showed this same distribution
with low to moderate grain densities while the pars
medialrs mamtained moderate densities of label throughout ns extent. Receptor icvels in the cortical
amygdaloid nucleus were also low rn rostra1 portions
of the amygdala. becoming moderate in more caudal

Autorad~ography of histamine H,-receptors in rat brain 17
ievels. LOW levels of HI-receptors were seen through-
out the central amygdaloid nucleus and the pars POS- terior of the lateral amygdaloid nucleus. The pars anterior of the lateral amygdaloid nucleus contained
very few autoradiographic grains. Low levels of antoradiographic grains were local-
ized over the lamina pyramidalis, polymorphi~a and plexiformis OF the tuberculum olfactorium and in the lateral nucleus of the olfactory tract (Fig. 12). Very low levels were seen over the lamina granularis externa and lamina molecularis of the olfactory bulb (not shown).
r~za~a~l~s. There was a paucity of autoradiographi~ grains throughout the thalamus. Moderate levels were found only in the nucleus periventricularis of the tbalamus (Fig. 5). Low grain levels coufd be seen in the nucleus reuniens, nucleus rhomboideus, reticular nucleus of the thalamus. nucleus subparafas~icularis and the nucleus posteromedjanus thalami (Fig. 13). The rest of the thalamic nuclei contained a very low amount of uniformly distributed autoradiographic grains.
~.v~~thfl~~i~~s and su~t~aiam~s. The highest auto- radiographic grain densities for me~yram~ne binding sites in the hypothalamus occurred in the supraoptic and suprachiasmatic nuclei, in all parts of the ventro- medial nucleus, and in the nucleus premammillaris ventralis (Figs 3, 4. 5, 13). Moderate grain densities were identified in the nucleus preopticus medialis, nu- cleus preo~ticus periventrjcularis aad in the nucleus paraventricularis (filiformis); (Fig. 5). Moderate levels of grains were seen enveloping the lateral and dorsal surfaces of the posterior mammillary nucleus (not shown). The dorsa1 premammiiiary nucleus and the nucleus periventricularis fhypothalami) were also moderately labeled with mepyramine. Low levels of autoradiographic grains could be identified over the nucleus preopticus magnocellularis” nucleus preop- ticus lateralis, caudal portions of the anterior hypo- thalamic nucleus, the lateral and posterior hypothaia- mic nuclei, both divisions of the nucleus dorso- media& the arcuate nucleus and infundibulum, the lateral preI~amrni~~ary nucleus, subthalamic nucleus and caudal portions of the zona incerta (Figs 13, 14).
Midbrairl ad metuthalurnus. The nucleus tractus optici basalis (Gillian) displayed moderate levels of autoradiographic grains (not shown). Other areas of the midbrain were low or very low in grain levels. Areas of iow grain densities included the nucleus tractus optici-pars medialis, the ventral and dorsal nuclei of the lateral geniculste body, the pars caudalis of the Edinger-Westphal nucleus, the nucleus of the oculomotor nerve, the stratum gr~~urn superficiale of the superior colliculus. caudal portions of the inferior
colliculus, the nucleus linearis oralis, the red nucleus, rostra1 portions of the periaquaductal gray matter. the interpeduncular nucleus and the zona compacta of the substantia nigra. Very low levels of autoradio- graphic grains could be seen in parts of the zona reti- culata of the substantia nigra.
Pens. Areas displaying high grain concentrations include the pontine nuclei, the nucleus reticularis teg- menti pontis, nucleus suprageniculatus facialis. nu- cleus of the facial nerve (VII), the medial vestibular nucleus and the nucleus raphe magnus (Figs 10. 14). Moderate amounts of autoradio~ra~hic grains were jdent~~ed in the nuclei on the Noor of the IV ventricle including the dorsal raphe nucleus. the mesencephalic nucleus of the trigeminal nerve, the locus coeruleus, nucleus tegmenti dorsalis lateralis and the nucleus tegmenti dorsalis (Gudden); (Fig. 9). Other areas with this concentratian of grain density include the super- ior central nucleus. the dorsal nucleus of the lateral lemniscus, nucleus tegmenti ventralis (Gudden). nu- cleus originis of the trigeminal nerve (V). external preo~ivary nucleus. rostra1 portions of the nucleus of the trapezoid body, spinal nucleus of the trigeminal nerve-pars dorsomedialis. the superior olivary nucleus and the superior vestibular nucleus (Figs 10. 14). A low grain density was seen in the nucleus of the troch- lear nerve (IV). ventral nucleus of the lateral lem- r&us, nucle~~s raphe pontis. spread along the corpus trapezoideum, the dorsal and ventral cochlear nuclei, the dorsal and ventral nucleus par~~brachiaI~s and the lateral vestibukr nucleus. The nucleus reticuk~ris pontis oralis contained very low levels of autoraciio- graphic grains.
~~~u~~~. High autoradjographic grain concen- trations were identified over the nucleus tractus soli- tarii and the dorsal motor nucleus of the vagus nerve (X). Moderate grain densities were seen over the nu- cleus of the hypogiossal nerve (XII), the nucleus ambi- guus, the nucleus reticularis parvocellularis, nucleus raphe pallidus and the lateral reticular nucleus (Figs 10, 14). Low levels were seen over the inferior olivary nucleus and accessory nuclei and the nucleus intercalatus. The dorsal and ventral medullary reticu- lar nuclei and the spinai nucleus of the tripeminal nerve contained very low levels of autoradiographic grains.
Crrrh&m. Only control levels of autoradiographic grains could be ~denti~ed in the cerebell~im, but this observation seems to be peculiar to the rat {Fig. I I ). Our investi~~~tions of the guinea-pig cerebellum demonstrated a large concentration of grains over the molecular layer. We have also examined human ccre- bellum and found mepyramine binding sites conccn- trated in the molecular layer although with a density much lower than that seen in the guinea-pig (Pai.,,rc- CIOS Vt al., 1979).
DISCUSSION
In these studies, we have localized with quanti- tative, light microscopic auto~dio~raphic methods, HI-histamine receptors in rat brain. More specifical]y,
we are Utilizing [‘H]mepyramine binding which has been extensively characterized kinetically and phar- macologically in homogenates of brain and in our mounted tissue sections (CHANG YT ai._ 1978; HI~.L cf

J. M. P~~I.AC‘IOS. J. K. WAMSL~,S and M J Kt ISAH
FIG. 3. A. LOH. powr‘r photomtcrograph 01 111~ a~rtorad,ugraplttc pram distrihutton over rostr.tf t,,rcbr;tttt Dark-field illumination causes the @rains to appear u bite on the coverslip against (he dark ),ackpro,,nd
Of the liSSw. !‘&to the very 10% concentratton ol’ grains (hut still htghcr than background) tfvcI most <,f the tissue. In this scctton rhc ~l~~~~str~~nl t(‘) t up. tfe&hx! 15, h;cvr!ly .I 1Ou ~~~n~~ntr~iti~fl of H,-reCeptors. Hhife the media) prcoptto nutieus tpoml cont;nned moderate icveh. and (he d,~,~,f pnr(,cn cf the bed nucleus of the stria (crminali~ IS() conratrts high lc\c)s of H,-rcccptors, bar I mm. U. Brtght-field photomicrograph of it t~ssuc section stamcd with pyrontn Y. Note (hc )~!_x(uJ~ (rf (hc
supraoptic nucleus (SO). C‘. Dark-field photomicrograph of’ H allowng visualtration of (he d($(r(nu(ion of
autoradiographic grains. Note the htgh ILSYCI of gratns QX~ aver the supr;ioptlc nuclcu, ,arrou ).
&,I ICW If
FIG. 1. A. Histamine (H, ) rcceptctr~ Ivcal~xd ;tutaradlu@rap2ltl~lll! II, Ixtrth of the tft;li;ttnw. h~poth&.
mus and amygdala. High levels of grains can he seen (11 the wntromcdtal hypo(n;(lamus ;h\ma) wtth
somewhat more moderate Icvrls tn 11~ ,tmygdal;t B;u I mm. B. Higher magntlication dark-field photomicrograph of the amygdaloid complex as seen 111 A, C. Bright-field photomicrograph of the same
area seen in B indicating the approximate localiration of various amygdaloid nuclei. Bar = 100 11.
Fig. 5. ‘Two 10% p~wur dark-field photomicrogntphs J~prctmp the ;rtituradiograph(c gr,(tn ~iIstr(hu(i(?n
dyer sections taken through the thalamus. Xote the l~theling of the midline th;tl;tmic nuclet .tnd man\ of
the nuclei of the l~~p~)ti~~~l~~~n~~s. Bar I mm.
FIG. 6. Various coronal sections (4 parts 01 (hc Ittppocampal (ormatron under dark-ticld I..\. ( . ii\ i111d bright-field (B. D. F) illuminatton demonstrattng the H ,-rcccptorc as stun from <cctions (ithcn progrubs.
ing from a rostra1 to a more caudal area, Phntomtcrtrgrnph I! demonstra(c\ l~gltt being rellcctcd from
pyramidal cells (white arrow) of the htppoc:tmpu\ ;tntl fmnule ceils thlsck arrow) ot’ the gyt :I\ dcnt~ttus.
G shows the distrihuti(~n of non-specific grains (01s tissue wits rncuhatecl in ~‘H]n~ep~ramine and unlabeled tripr~lidine). Note the ‘tissue light’ is still present m thtr, control pht~tomicr~~gr~ph and does
not indicate labeling. Bar = 100~~
PI<;. 7. Dark-held photomicrograph demon\tr;tttng the localr~at~~~ 01 ti ,-rcccpt~>rs in the c‘iudal hrppo-
campus and adjacent areas. Note the high ct~nccntration of’ rcccptors in the subiculum (S) and prcsubt- cular cortex. Also note the high concentration of receptors in Iamina IV of the cortex tit this level
(appears in the upper right-l~;tnd corner 01’ the pl~crt~,rnlcropraph). This area corresponds IO arca 41 01
KRIW (KRIFI;. 1946) and is related to (hc ~orttcal projecttons ctf the auditory system. Bat I mm. 1%.
Dark-field and bright-held (C) pi~c,tomicrogr;cplls ticm~mstratmg the high concentration ~)f .tutor;tdto-
graphic grams in the medial ;tmygdaloitl nr~clcu\ (urn) :thout mtdw;t> through the .~nrygdal;t
Il;lr IIHI ,,
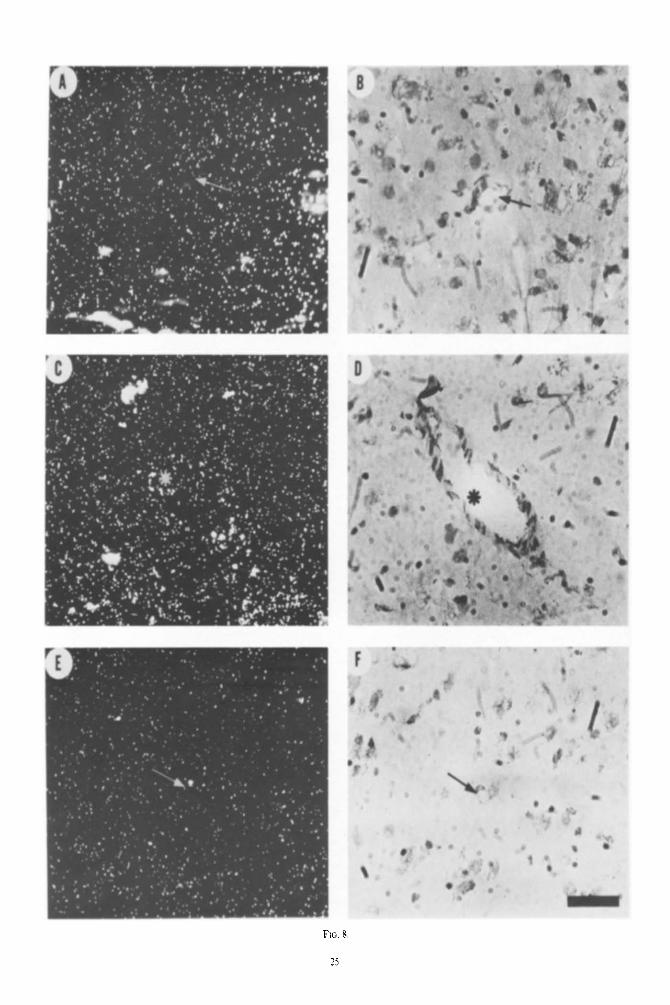
Ftc;. 8. Photomtcrographs showing l-l,-recq)turb in rcgwnx near Mood vessels. .A and f.3 demonstrate I~O
apparent association of grains with an arteriole tarro\c I f‘ sl~ows the gram dtstrihution i)ver <I vrnulc
seen in D (asterisk). E! dcmorstmtcs gratn\ nc,tr :t c;tpillar> ntrrow) as seen in F Hat st t ,c.
Ftc;. 0. Series of photorntcr~~~r~~ph~ taken lwm labeled \tructurc:.\ near the tloor (II’ the IV ventrtcle. r\
represents total binding areas of ),‘fiJmep!rammc while ( slio\bs non-spacific iire;ir. I312 giunulc cell
layer of the nodulus (N) of the cerebellum and the locu:, coerulcus (Icl demonstrate reflected ‘tissue light’
in C. Only this reflected light is seen m the nodulu\ ((I .\ hut the locus coeruleu\ she\\\ .t moderate
specific grain tlenslty ;I% well a\ the reflected light. Bar = I(K) (1
FIG. IO. A. Ph(~tomicro~r~iph of IIIC If, fic’cpt4x 10~;ili~rtio11 111 thr: pontmc regron t+( the ~r~~irtstctll.
Note the high density of grams in Ltrci(\ WLT the pontm~ nuctc~ rpo) and the nuchXts retlcuiaris tegmetlti
pontis (rtp). The dorsal raphc nucleus (rd) I \ I;ilreled rr~~&r:itely .tnd the supcrtor ccntml ni:clcus (ncz) is
labeled somewhat ;tt a lower )evcl. B. Control d,trk-field photomicrograph of .t SW~IOII X~JacUlt tcl the
one seen in ,A. Grains in this picture indicate non-spccttie htndtng 01 the [3H]mcpvramtttc. Bar z- 500 I$
C. section of brain stem tissue at the Ic\~l ~3f the mcdt;tl \cstihular nucleus (vm) r\~rtoradropriipllcc grams can be seen throughout this nuclem as sell as the nucleus cd nerve WI (nvll). Note the intense
labeling in the nucleus raphe magnus (rm) and the low labeling m the cochlear nuclet of the auditory
system (cod. cov). Bar = 1 mm. D. Section through the brainstem at the level of the area postrema (aP). The nucleus of nerve XII (nXlI) and the nucleus tractus solitarti (nts) are heavily labeled with [‘HIme- pyramine in this photomicrograph. lheir location can be determined by viewing the tissue underneath
the emulsion-coated coverslip ustng bright-field illumination as seen in E Bar = 10(111.
~tc;. 11. Dark-field and correspondrng hrrght-tield photomicrographs from rat (A. B). gutnea-ptg (c fl) and human (b, F) cerebcfta. The motccuhtc Ltyer tm) of the puiooa-pip cerehe))Um dcrn~)l~str~itcs man?
I-f,_receptors uhile that or the human shrnvs CMI~ ;t sltght collcetltraliot~ and the mo)ecuhtr htyer of the rat shows no f{ ,.rcceptors at ;rll. Agatn. in these pltot~,micrographs the closeness cl 1))~ c&s In the
grenule cell layer (g) causes arti(Xtua) ((shut! rcllcctancc vvhtch can be seen cvcnly thrtrrhutcd tn thts
layer in all three specimens. p = layer of Purkinte cells. w = white matter. Bar .= 100 jr

FIG. 2. A. Photomicrograph of the autoradiographic grain distribution over a tissue section taken from parietal cortex (corticat area 2 of KRIEG, 1946). This tissue section had been incubated in [3H]mepyr- amine with 2 PM triprolidine and indicates non-specific binding. B. Adjacent tissue section, but incu- bated in C3H]mepyramine alone indicating total binding. C. Brightfield photomicrograph showing the tissue as it appears below the grain distribution on the coverslip seen in B. The Roman numerals demarcate the approximate levels of the cerebral cortex. Note the apparent specific increase in
HI-receptors in lamina IV as seen in B. CC, corpus callosum. Bar = 200 p.
t9


FIG. 4.



Fiti. 7.
24


‘?


Abbreviations used in Figures 12, 13, 14
a
abl
abm
ac
ace
ala
alp
am
amb
ar
ci
COd
CO\
cP cu
dcgi
&r hdd
hl
hp
hpv hvmc
hvml
hvmm
io
iod
iom
IC
Ih
IIV
mh
npd npv
nrp ntd
ntdl
ntm
nts
ntv,
ntVd
nVl1
nX
nXI1
P
pd
Pf
Pot
pom poma
POP post
Pv
Pvr pvs
rd
re r&i
rh
rl
Nucleus accumbens
Nucleus amygdaloideus basalis, pars lateralis
Nucleus amygdaloideus basalis, pars medialis
Nucleus amygdaloideus centralis
Nucleus amygdaloideus corticalis
Nucleus amygdaloideus lateralis, pars anterior
Nucleus amygdaloideus lateralis pars posterior
Nucleus amygdaloideus medialis
Nucleus ambiguus
Nucleus arcuatus
Colliculus inferior
Nucleus cochlearis dorsalis
Nucleus cochlearis ventralis
Nucleus caudatus putamen
Nucleus cuneatus
Nucleus dorsalis corporis geniculati lateralis
Nucleus gracilis
Nucleus dorsomedialis (hypothalami), pars dor-
salis
Nucleus lateralis (hypothalami)
Nucleus posterior (hypothalami)
Nucleus periventricularis (hypothalami)
Nucleus ventromedialis (hypothalami), pars cen-
tralis
Nucleus ventromedialis (hypothalami), pars later-
alis
Nucleus ventromedialis (hypothalami). pars
medialis
Nucleus olivaris inferior
Nucleus accessorius olivaris dorsalis
Nucleus accessorius olivaris medialis
Locus coeruleus
Nucleus habenulae lateralis
Nucleus lemnisci lateralis ventralis
Nucleus medialis habenulae
Nucleus parabrachialis dorsalis
Nucleus parabachialis ventralis
Nucleus reticularis paramedianus
Nucleus tegmenti dorsalis lateralis
Nucleus tegmenti dorsalis Gudden
Nucleus tractus mesencephali
Nucleus tractus sohtarii
Nucleus tegmenti ventralis Gudden
Nucleus tractus spinalis nervi trigemini pars dor-
somedialis
Nucleus originis nervi facialis
Nucleus originis dorsalis vagi
Nucleus originis nervi hypoglossi
Nucleus pretectalis
Nucleus premamillaris dorsalis
Nucleus parafascicularis
Nucleus preopticus lateralis
Nucleus preopticus medialis
Nucleus preopticus magnocellularis
Nucleus preopticus periventricularis
Nucleus preopticus. pars suprachiasmatica
Nucleus premamillaris ventralis
Nucleus perventricularis rotundocelfularis
Nucleus periventricularis stellatocellularis Nucleus raphe dorsalis
Nucleus reuniens
Nucleus reticularis gigantocellularis
Nucleus rhomboideus
Nucleus reticularis lateralis
rm
ro
rpc
rpo rpoo
rtp sl
sm
St
sut
td
11
tlP tml
tmm
tpo tr
ts
tv
tvd
tvm
tvp
VI
vm
CAA
CA1
cc
CL
co
CPF
FC
FLM
FMP
FR
GD
GP
HI
LL
LM
P
PC1
PCM
PCS
S
SM
SO
ST
TO
TOL
TS
TSTH
TSV
TULC
TULl
TULP
ZI
IV
V
Nucleus raphe magus
Nucleus raphe obscurus
Nucleus reticularis parvocellularis
Nucleus raphe pontis
Nucleus reticularis pontis oralis
Nucleus reticularis tegmenti pontis
Nucleus septi lateralis
Nucleus septi medialis
Nucleus interstitialis striae terminalis
Nucleus subthalamicus
Nucleus tractus diagonalis (Broca)
Nucleus lateralis thalami
Nucleus lateralis thalami, pars posterior
Nucleus medialis thalami, pars lateralis
Nucleus mediahs thalami, pars medialis
Nucleus posterior thalami
Nucleus reticularis thalami
Nucleus triangularis septi
Nucleus ventralis thalami
Nucleus ventralis thalami, pars dorsomedialis
Nucleus ventralis medialis thalami, pars magno-
cellularis
Nucleus ventralis medialis thalami, pars parvocel-
lularis
Nucleus vestibularis lateralis
Nucleus vestibularis medialis
Commissura anterior, pars anterior
Capsula interna
Crus cerebri
Claustrum
Chiasma opticum
Cortex piriformis
Columna fornicis
Fasciculus cuneatus
Fasciculus longitudinalis
Fasciculus medialis prosencephali
Fasciculus retroflexus
Gyrus dentatus
Globus pallidus
Hippocampus
Lemniscus lateralis
Lemniscus medialis
Tractus corticospinalis
Pedunculus cerebellaris inferior
Pedunculus cerebellaris medius
Pedunculus cerebellaris superior
Subiculum
Stria medullaris thalami
Stratum opticum colliculi superioris
Stria terminalis
Tractus opticus
Tractus olfactorius lateralis
Tractus solitarius
Tractus striohypothalamicus
Tractus spinalis nervi trigemini
Tuberculum olfactorium, pars corticalis, lamina
pyramidalis
Tuberculum olfactorium, pars interna, lamina polymorphica
Tuberculum olfactorium, pars corticalis, lamina plexiformis
Zona incerta
Nervus trochlearis
Nervus trigeminalis
29

__. CPF
TOL
Autoradiogr~phy of histamine HI-receptors in rat brain 31
hdd hp; or’ hkm hvm’ FtYp To am
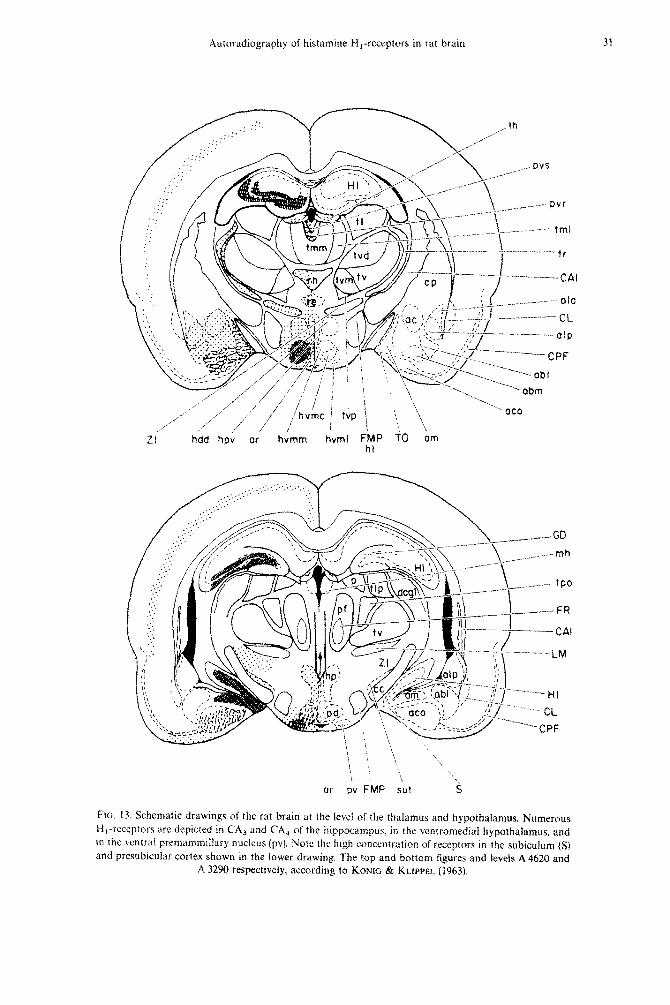
Ftc. 13. Schematic drawings of the rat brain at the level of the thalamus and hypothalamus, Numerous H,-receptors arc depicted in CA3 and CA4 of the hippoc~lmpus. in the ventromedial hypothalamus. and in the ventral premamm~llary nucleus (pv). Note the high concentr;ttion of receptors in the subiculum (S) and presubicular cortex shown in the lower drawing. ‘The top and bottom figures and levels A 4620 and
A 3290 respectively, according to Kowrc & KLIPPEL (19631.

1. M. 1'a~nrrc.k~. .f. K. WAMSCEY and M. J. KUHAK
ic
ntm
nfdi
PCS
FiM
FIG. 14. Schematic drawings of three levels of brainstem showing the distnborion of auturadiographx grains fbflowkg fabeiing with @-Qmepyramine. Dense receptor cancentrations are represented in rhe nuckus reticularis tegmenti pantis frtp) the nudei of the pooa, the media3 v&b&r nucleirs fvm). nuchs of the facial nerve fnVII), nucfeua tractus solitarii (nts) and in the dorsal motor nucleus of thr vagus nerve f&X). The top, middle and bottom schematics are lavcls P1.5, P3.9 and P6.5 aLXordiflg to

Autoradiography of histamine HI-receptors in rat brain 33
&.. 197X; TRAN rf al., 1978; CHANG et al., 1979~; PAC,AUOS et a/., 1979). It is important to point out that this site is not the only site at which histamine acts. For example, the I-$,-receptor is not labeled to any significant extent under our conditions (BUCK, DUNCAN, DURANT, GANELLIN & PARX~NS, 1972). However, the mepyramine binding site has all of the characteristics of a physiologically and pharmacologi- cally relevant H,-receptor (CHANG et d., 1978; HILL et d., 1978; TRAN er ul.. 1978; CHANG et ~1.. I9790: Ho+t er al., 1977; CHAWS et ui'., 197983. Also, in these studies, we have more or less focused on the areas of the brain showing the highest densities of receptor. However, it seems worth cautioning that even areas with low densities of receptor may be important in some cases for histaminergic transmission or for the action of histaminergic drugs. At present. no corre- lation has been established between density of rccep- tors and physiological response.
An important consideration is the possibility that all H,-receptors are not neuronal. There is abundant biochemical, anatomical and electrophysiologica~ data suggesting that histamine is a n~urotransmitter in the brain and this provides substantial support for the notion that H,-receptors are localized to neurons in synaptic areas (SCHWARTZ er ~1.. 1979h; SCWWARTZ et d., 1978; GREEN et (II., 1978; SCWWAIUZ et ul., 1979~; TAYLOR, 1975). On the other hand, some studies suggest the ~ocaliz~ltion of H,-receptors to non-neuro- nal sites. For example. preparations of micro-blood vessels from brain have elevated densities of W,-receptors (PER~IXKA. MOSKOWI~Z, REINHARD & SNYDER, 1980). Also, studies involving several lesions of the hippocampus and caudate nuclei, i.e. lesions causing a loss of input as well as kainate lesions which destroy cell bodies, did not cause a loss of ~,-receptor binding (CWANG, TRAN & SNYDER, 1980).
Our light microscopic studies do not provide suf- ficient resolution to settle this issue (Fig. 8). Accord- ingly, our results should be kept in perspective given these possibitities.
We observed wide variations in the densities of H,-receptor in the different regions of the rat brain, Some of these areas containing high densities could be linked together on the basis of function. For example, many components of the auditory system had sj~ificant densities of receptors. These areas in- clude the ventral and dorsal cochlear nuclei, the nu- clei of the trapezoid body, the superior olive, the ven- tral and dorsal nuclei of the lateral lemniscus. the inferior colliculus and elevated grain levels in lamina IV of the cortex in the temporal areas. Another sys- tem having high densities of receptors in various parts is the limbic system. Areas with high densities of receptors include parts of the amygdala and hippo- campus. Areas associated with the limbic system hav- ing high densities include parts of the hypothalamus
and &&ate cortex and b4 nucleus of the stria terminalis.
We have observed elevated densities of I-i,-histamine receptors in some of the same areas where we have observed several other types of recep tors. For example. lamina IV of the cerebra1 cortex in the rat has elevated densities of opiate receptors (ATWEH & KUNAR, 1977~; YC)UNC~ & KUHAR, 1979a). benzodiazepine receptors (YOUNC; & KU~AR, 1979h: Youuc & KUtiAR. 19SOa), neurotcnsin receptors (YOUNG & KUWAR, 197%). alpha-l adrenergic recep- tors (Yotr~c; & KUWAR, 19794: YOON~; & KUIIAR, l%Ohf, muscarinic receptors (WAMX.EY et cl!., in press) and 7%.aminobutyrate receptors (PALAUOS. YOUNG & KUHAR, 1980). Also, the nucleus tractus solitarii has high densities of opiate receptors (ATWEH & KUHAR, 197763, as well as other receptors (YoI:N(; & KUNAR. 1979rf: YCRJ~~G & KUHAR, 1980h; WAMSLEY pf ok in
press). The per~v~ntricu~ar nucleus of the thalamus also has been found to have elevated densities of several receptors including opiate (ATWEW
& KUHAR, 1977c), beta-adrenergic receptors ( PALA- CIOS & KUHAR, 1480~) as weH as several other recep- tors. Why certain areas of the brain are receptor ‘hot spots’ is an interesting question. No obvious explana- tion is apparent and there need not be any special significance to this. But these areas are likely to show a lot of drug interactions.
Detailed information on the hist~~miI~ergic inner- vation in some areas of the rat brain has been obtained by microchemical analysis of small tissue samples and lesion studies (SCHWAREY, 1977:
SCHWARTZ ef ui., 197X). The presence of a widespread inn~r~ti~~ in the forebrain was deduced from the decrease in histidine decarboxylase activity (a marker for histaminergic terminals) after medial forebrain bundle lesions (GARBARC;, BARBIN, FECZR &
SCHWARTZ, 1974; BARBIN, HIRSCH, GARBARC; &
SCHWARTZ, 1975; GARBARG, BARBIN, BISCMOFF. POL-
LARD & S~~WAR~2. 19761. The ce!! bodies of this ascending pathway are thought to be localized in the upper midbrain (SCHWARTZ er ul.. 197X).
While most of the areas where we observed signif& cant concentrations of histamine H,-receptors poss- ibly receive hist~~minergic innerv~lt~on, there is striking difference in the relative conce~ltration of both pre- and postsyn~~ptic ‘markers”, These disparities were &O observed in our initial study in the guinea-pig brain (PALACIOS cf al. 1979). The bed nucleus of the stria term&a& is one of the areas with very high receptor concentration. One of the highest histjdine decarboxylase c(~ncentr~~tions in the brain is afso found in this nuclt~s. However. while the terminals Seem to be concentrated in the ventral part of the nucleus. the receptors are much more dense in the dorsal part (BEN Am, Lr: GALL LA SALLI:. &,RBIN,
SCQwART2 & GARRARG. 1977).

J. M. PALACIOS. J. K. WAMSLFW and M. J. KUHAR
-_.________ _~._ __ _.________.._~___ Wwdinc decarbosylase ‘Specific’ [3H~Mep~ramjn~
activity grain density \dpm;h!mg tissue, IO’)’ (per 529 pz of tissue)
-..--.-. __.__- -_-.. _- ._.. I___-.._-._____-__-.__ _. Hyporhalamws
Nucleus Supraopticus 4.3 41.5 + 2.0 Nucleus Suprachiasmaticus 6.3 42.8 + 0.5 Nucleus Arcuatus 6.3 IX.3 + 0.x Nucleus Paraventricularis 5.3 25.9 + 0.9 Nucleus Dorsomedialis 8.1 17.7 + 2.h Posterior Hypothalamus 77 21.5 * 2.3 Nucleus Premammillaris ventralis 8.j 42.5 i 1.9
,4l~y~da~fl Medial Nucleus 2.6 JO.1 * 1.7 Central Nucleus 2.3 14.9 * 0.6 Basomedial Nucleus 1.9 28.8 * 0.8 Basolateral Nucleus 1.1 16.7 _t 0.7 Cortical Nucleus I.0 29.4 * I.i Posterior Lateral Nucleus 1.1 14.0 * 0.8 Anterior Lateral Nucleus I.0 X.6 f 0.4
- .-
’ Histidine decarhoxylase values are from POLLARD tlr (I/. (1976) for the hypothala. mus and from BE~V ARI et al. (1977) for the amygdala.
The hippocampal formation has a relatively low concentration of histidine decarboxylase but is rich in receptors. Mepyramine binding sites within this area are mostly concentrated in the CA, and CA4 regions where the histaminergic innervation appears to be low (BARBIN, GARBARG, SCHWARTZ & STORM-MATHI-
SEN, 19%). Again, in the cases of the amygdala and hypothala-
mus, there is a lack of correspondence between ter- minal and receptor densities (Table 1). Both the medial and central nuclei of the amygdala are rich in histidine decarboxylase activity (BEN ARI et ul., 1977), but only the medial nucleus has a high density of H,-receptors. The hypothalamus. the brain region with the highest levels of histamine and histidine d~arboxy~ase, is also very rich in HI-receptors. although there is not a good correlation between histaminergic innervation and receptor concentration in the different hypothalamic nuclei (BROWNSTEIN,
SAAVEDRA, PALKOVITS & AXELROD, 1974; POLLARD,
BISCHOFF, LLORENS & SCHWARTZ, 1976).
Several hypotheses could be postulated to explain these discrepancies. In the case of the histamine&z system. a second class of receptors, Hz, that is not revealed by our method, and is also present in the brain (SCHWARTZ, 1979; GREEN et at.. 1978) has to be considered. On the other hand, one would not expect a total correlation between levels of neurotr~smitter and receptor, but rather a correlation between nerve terminals and receptors. At present, however, the detailed anatomical distribution of histaminergic ter- minals is unknown.
Possible pharmncoloyical rrlevance
Classic antihistamine drugs are widely used thera- peutic agents (DOUGLAS, 1975). We have observed some interesting relationships between the pharmaco-
logic responses to antihistamines and the distribution of HI-receptors in this study. We propose. at least as a first hypothesis. that many of the areas showing high densities of receptors are those where antihis- tamine drugs exert their therapeutic effects (Table 2). For example, antihistamines are used in the treatment of motion sickness. Motion sickness is due to over- stimulation of the vestibular system (DOUGLAS, 1975: JAJU & WANG, 1971: MOSEY. 1970). The vestibular nuclei in the floor of the fourth ventricle in the medulla have high densities of receptors and may very well be the site where antihistamines alleviate and counteract motion sickness. It is interesting that the anticholinergic properties of antihistaminic drugs were postulated to be responsible for the antimotion sickness effects since the vestibular nuclei are affected by anticholinergic drugs. which are also useful in treating motion sickness (DOUC;LAS. 1975; MONEY. 1970). Recently, WC have found high densities of mus-
carinic receptors in the vestibular nuclei as well (WAMSLEY et cl/.. in press). This suggests that anti- histaminics and anticholinergics are both antimotion sickness drugs because of action at separate receptors which are located in the same anatomical area
Antihistamine drugs are also well known for their sedative properties (FAIF~~Y)LD, 1978). Related to this. we have observed high levels of receptors in the raphc nucleus of the midbr~n. an area associated with sleep
(JOUET, 1969). We have also noted e!evaccd densities of receptors in lamina IV of the cortex. Severai other drugs with sedative properties such as opiates. mus- carinic drugs and alpha 1 -adrenergic drugs also have elevated levels of receptors in lamina IV of the cerebral cortex (WAMSLEY, ZARRIN, RIKDSALL & KUHAR. 1980:
YOUNG & KWHAR, 1979d: YOUNG & KUHAR, 1980h). It is possible that these receptors could interfere with cortical arousal from thalamic input. When antihis-

Autoradiography of histamine Hi-receptors in rat brain 35
TAAE 2. TENTATIVE CORRELATION BETWEEN AREAS WITH RECEPTORS AND PHYSIO- LOGICAL EFFECTS OF HISTAMINE AND ANTIHISTAMINE DRUGS
Drug Action Anatomical Area
,~f~~ihistai~j~es Antimotion Sickness Sedation and sleep
Vestibular Nuclei Dorsal Midbrain Raphe nucleus and cerebral cortex
Histumirw Suppression of Food Intake
Release of ADH Production of Thirst
Prolactin Secretion Hypothermia Increase Blood Pressure
and Heart Rate Retching
Ventromedial Hypothalamus Lateral Hypothalamus Supraopti~ Nucleus Supraoptic Nucleus Posterior Hypothalamic Areas Basal Hypothalamus Anterior Hypothalamus Nucleus Tractus Solitarii Hypothalamus Nucleus Amhiguus
tamines are given in high doses, convulsions can occur (WYNGAARDEN & SEEVERS, 1951). Areas often
associated with epileptiform activity that have high
levels of receptor include the hipp~ampus, amygdala
and cerebral cortex. Additional effects have been observed in experi-
mental animals after injection of histamine and anti-
histaminics by various routes (CALCUTT, 1976; SCHWARTZ et ul., 1979~; GREEN et of., 1978;
SCHWARTZ, 1979). Histamine, when injected intracere- broventricularfy causes a depression of food intake
(CLINESCHMIDT & LOTTI, 1973). Related to this we ob- served very high densities of histamine receptors in
the ventromedial hypothalamus which is thought to be the satiety center controlling food intake. These effects could also be caused by histamine receptors in
the lateral hypothalamus which has been shown to be involved in the control of feeding behavior (STEVEN-
SON, 1969). Injection of histamine also causes a release of anti-
diuretic hormone probably via an H,-receptor (BHAR- GAVA, 1974; BHARGAVA, KULS~ES~A, SANTHAKU- MARI & SRIVASTAVA, 1973; BENNETT & PERT, 1974;
HOFFMAN & SCHMID, 1978; TUOMISTO & ERIKSSON, 1979; DOGTEROM, VAN WIMERSMA GREIDANUS & DE WIED, 1976). Related to this, we observed high
densities of receptors in the supraoptic nuclei, an area with ceil bodies containing antid~uretic hormone. As- sociated with this may be the observation that injec- tion of histamine causes thirst in animals (GERALD & STERN, 1969: GERALD & MAICKEL, 1972; LEIBOWITZ, 1973). One must also consider the posterior hypo-
thalamus. an area with elevated receptor levels, to be involved in the observations that histamine causes thirst since stimulation of this area causes animals to seek water (STEVENSON, 1969)
The hypothermia observed after histamine injection could be due to the H,-receptors located in the preoptic-anterior hypothalamus area (BREZENOFF &
LOMAX. 1970; BRIMBLECOMBE & CALCLJTT. 1974: cos-
TENTIN, BOULU & SCHWARTZ, 1973; GREEN, Cox &
LOMAX, 1976). The increase in blood pressure and tachycardia observed after histamine injection (for a
review, see OWEN, 1977, see abo FINCH & HICKS,
1976) may very well be mediated by the high densities
of receptors in the nucleus tractus solitarii and the
posterior hypothalamus. It is also possible that the
retching and swallowing induced by intra-cerebroven- tricular injection of histamine (FELI)BERC & SHER-
wooa, 1954) are mediated by the high densities of receptors in the nucleus ambiguus, an area mediating
these functions. While we are not aware of any
reports that histamine or antihistaminics affect audi-
tion, we would expect there to be some effect on this sensory system given the extensive localization of
receptors to auditory areas as described above. Many electrophysiologic studies have found specific
histamine responses in many of the areas mentioned above (for a review, see HAAS & WOLF. 1977;
SCHWARTZ, PALACIOS, BARBIN, QUACH, GARBARG,
HAAS & WOLF, 1979). These observations support, not only the notion that these receptors are functional,
but that they are localized to neurons (rather than
nonneuronal sites) in many of these areas. While the effects of HI-antihistamine applied by iontophoresis
are difficult to interpret because of prominent local anesthetic effects, recent studies with cultured hypo-
thalamic neurons strongIy support the presence of functional H,-receptors in these hypothalamic neurons (GELLER, 1976).
Ackrlowtm~~c~,nrt,ts--The authors acknowledge the techni- cal assistance of Mrs. NAOMI TAYLOR, the clerical assist- ance of Ms. DARLENE WEIMER and MRS. MARY FLUTKA, and the support of USPHS Grant MH 25951. M. J. KUHAR is the recipient of an RCDA Type II Award MH 00053. J. K. WAMSLEY is the recipient of postdoctoral fellowship HD 05739 and J. M. PALACI~S is the recipient of a Fogarty International Fellowship TW 02583.

36 J. M. PALACWS. .I. K. W.AMSLEY and M. J. KCHAR
REFERENC’ES ATWEH S. F. & KUHAR M. J. (1977~) Autorad~ographic localization of opiate receptors in rat brain. 111. The telencepha-
Ion. Brain Res. 1.34, 393,405.
ATWEA S. F. & KUHAR M. J. (1977b) Autoradiographi~ localization of opiate receptor in rat brain. 1. 7’ht spinal cord and lower medulla. Bruin Res. 124, 5368.
ATwEH S. F. & KUHAR M. J. (1977(,) Autor~diograpbi~ localization of opiate receptors in rat brain. II. The brainstem. Brain Res. 129, 1. 12.
BARBIN G., GARBARG M., SCHWARTZ J. C. & STORM-MATHSEN J. (1976) Histamine synthesizing aflcrrnt~ to the hippo-
campal region. J. Neurochem. 26, 259-263.
BARBIN G., HIRSCH J. C.. GARBARC M. & S(.HWARTZ. J. C. (1975) Decrease in histamine content ;rnd decarboxylase
activities in an isolated area of the cerebral cortex of the cat. Brain Res. 92, 170_174.
BE”: ARI Y.. Lk GALL. LA SALLE G.. BARRIN G.. ScfiwAK.rz J. C. Cy: GARRARG M. (1977) Histamine s~n~h~si~ing afferents
within the am~daloid comples and bed nucleus of the stria termina~is in the rat. Br&n Re,s. 138, 185 2%+.
FW-WTT C. T. & PEKI‘ A. (1974) Antidiur~sis produced by injections of histamine into the cat supraoptic ntrcicus. Braitt Res. 78, ISI-156.
BHARC~AVA K, P. (1974) Some neuropharmacological studies with histamine. J. P~~urt~l~c~~. 5, (SuppI. ! + ?;, BHAKGAVA K. P., KUi..SHRESU%A V. K., SANTI~AIUMARI G. & SRIVASTAVA Y. P. (1973) Mechanism of }listamine~indu~eci
antidiuretic response. 81: J. Plzurnluc: 47. 700 706.
&.AC.R J. W., Dt:N(‘Ar W. A. M.. DL’KAS’I’ C’. J., GANELI.I\: t’. R. & PARSO&S E. M. (1972) Definition anti ~ntagonlsm of histamine Hz-receptors. Nature, Lorrd. 236, 3X5 340.
BKEZENOYF H. E. & LOMAX P. (1970) Temperature changes following microinjection of histamine in&o the thermoregufat. ory ccntres of the rat. Ezp&~tltrrr 26, 51 .51.
&UMHLEC’OMBE R. W. & CALCWT‘ c. R. (1974) The involvement of H, and Hz-histamine receptors in the hypothermic
response to intra~erebroventrictilariy-injected histamine and related drugs. J, P~tartnucffi. 5, (Suppl. 7) 1 I.
~R~wN~~~N M. f., SAAVEDRA J. M., PALKOVITS M. & AXELROD J. (1974) Histamine content of by~tbalamjc nuclei of the rat. Bruin Rrs. 77, 151--l%.
CALCUTT C. R. (1976) The role of histamine in the brain. G‘rtt. ~~zur~ffc. 7, I5 -75.
C~~ANC; R. S. L., THAN V. T. & SNYI>ER S. H. (19783 Histamine fit-receptors in brain labeled with ~~H~mepyrarni~e, &r.
f. P~~ur~l~c. 48. 463.464.
CHANC R. S. L.. TRAN V. T. 8: SNYDER S. H. (1979ti) ~leterogeneity of histamine Hz-receptors: species variations in
[‘Hlmepyramine binding of brain membranes. .I, Nrurochem. 32, 1653--1663.
CNANG R. S. L.. TRAM V. T. & SN~II?ER S. H. (19798) Characteristics of histamine HI-receptors in periphmal tissues labeled
with c3H]mepyr~mine. J. Pharmac. rsy. Tl~rr. 209, 4.77.,443.
CHANG R. S. L.. TRAN V. T. & S~YY!>ER S. H. (1980) Neurotransmitter receptors iocalizations: In ben~~~d~a~epine~ GABA.
beta-adrenergic and histamine H,-receptor binding. Elrcrin Res. 190, 95-I 10. CL.INESCHMIDT B. V. & LOTTI V. J. (IY73) Histamine: intraventricular injection suppresses ingestive hehnviour of the cat.
Arc/is hr. Pharntocodyn. T’hir. 206, 28&29X.
~CXTEUTIN J.. Bn%%u P. & SCHWARTZ J. C. (1073) Ph;~rmaco~ogi~a~ studies on the role of histamine in tf?ermoregu~at~#n.
.4ge/tts und A&Oll.S 3. 177.
DOGI+EROM J.. VAN ~~~E~SMA GR~I~A~US TJ. B. & DE WZED D. (19761 Histamine as an extremely potent releaser of
vasopressin in the rat. ~~pfr~~nr~a 32, 659-660.
DOWLAS W. W. (197.5) Histamine and antihis~dmines; ~-hydr~)~ytryptam~ne and antagonists. In ^I’/!<, ~~~~~u~o~u~~~u~
Basis ~~~~~~~[~p~‘~~ric.~. 5th edition (eds GW~MAN L. S. & GILMAN A.), pp. 59%629, MacMi~ian Ptiblisbi~g Co.. New York. FAINGOLD C. L. (1978) Antihistaminics as central nervous system depressants. In handbook of ~~p~rime~r~~ F~urrn~c~~~g~
feds EICHLER 0. & FARAH A.), Vof. 18, Part 2. pp. 561..573, Springer-Verlag, Berlin.
FELDBERG W. & SHERWOOD S. L. (1954) tnjection of drugs into the lateral ventricle of the cat. .I. Pltpioi., Lund. t23.
148-- 167‘ FINCH L. & Hrclts P. E. (19761 fnvofvement of h~pot~~~~~drnic histamine receptors in the central cardiovascuhtr actions of
histamine. Nrurophrrmrucolo(~p 16, 21 I.- 218. GARBARC M,, BI\KI~I~ G., BISU~OFI. S.. POI.LARI) H. & SC~~WAKT% J. c‘. (1976) Dual localization 01 histamine in an
ascending neuronal pathway and in non-neuronal cells evidenced by lesions in the lateral hypothalamic area. Brain Res.
$06, 333-34s. GAR~AR~~ M., Bnaatu G.. f;r.~~a J. & SCHWAIW~ J. c‘. (1974) ~istaminer~i~ pathway in rat brain evidenced by lesions of
the medial forebrain bundte. Scioicr N. E If%, X.33, X75. GI:LLEK f-I. M. (1676) Effects of some putative netirotr~~nsmitt~rs on unit activity of tuberaf h~pothaiam~~ neurons irt &ru.
Hririir Res. 108. 413 430. &,RALI) M. C. & MA~CKI:L R. P. (1972) Studies on the role of brain histamine in behaviour. Br. J. Pharnrw. 44.46247l.
C;~:KALJ> M. (:. & STI:RN W. C. (1969) Interactions of histamine and antihistamines with behaviourai systems. Fe& f’roc.
Fedri Am. SCW rs,t. Hial. 27. 272. GREEN M. D.. Cox 3. & LOMAX P. (1976) Histamine H, and I+,-receptors in the central thermor~guiator~ pathway’s of the
rat. J. Neurosci. Res. I, 353 39.
GKEI:N J. P., Jolr~so~ C. L. & WEINSTEIN H. (197X) Histamine as a neurotransmitter. In PsycBuphffrf~~urclnt/oy~~ A &‘f~.*r- ation of Progress (eds LWTON M. A., DIMA~C~O A. & KILLAM K. F.), pp. 319.“332. Raven Press, New York.
HAAS H. L. & WOLF P. (1977) Central actions of histamine: microelectrophoretic studies. Brain Res. 122, 269-279.
HIU S. J., EMSON P. c‘. & YUtJVC J. M. (197X) The binding of [~H]mep~~amine to histamine H,-receptors in guinea”pig
brain. 1. ;~~~~t~~t~~?~tf~. 31, 997.. 10114.

Autoradiography of histamine Hi-receptors in tat brain 37
f-fl~~ S. J., YQI:NC; J. M. & MARRIAU D. H. (1977) Specific binding of C3H]mepyramine to histamine H,-receptors in
jntcstinal smooth muscle. Nururr. Lotril 270, X-363. H(>r-_rllt~h W. E. & S~MII) P. Cr. (1978) Cardiovascular and antidiuret~c cfTects of central histamine. Lifi* Sci. 22,
1709-171‘3.
JAJ~ B. P. & Wn,uc; S. C. (1971) EfTects o~d~pl~enhyd~mi~~e and dimenhydrin~ite on ves&~b~lar neuronal activity oi‘ cat: A search for the IUCLZS of their ~~ntimotion sickness action. J. ~~iu~i~i~~. rasp. That. 175. 71% ‘724.
Jot VI:T M. (1969) Biogcnic amines and the states of sleep. %i~wt~. N. I< 163, 32--41.
KONI(; J. F, R. & KI.IPPTI. R. A. (1963) The Rtrt Bmin: A Stcwotczsic~ Atlus. R. E. Krieger Publishing Co.. New York.
KKIIX; W. J. S. (1946) Connections of the Cerebral Cortex. J. C’onrp. .Yeuro/. 84. 277 323.
I.,t~tnow~~z S. F. (1973) Histamme: a stimulating effect on drinking behaviour in the rat. BrcCrl RPS. 63, 440- 444.
MoNt,Y K. E. (1970) Motion Sickness. Physiol. Rer. 50. I 30. Owriu D. A. A. fl977) Histamine receptors in the cardiovascular system. Gnr. Phc~rrmc. 8, 141- 156.
PAI.ACXE J. M. & KUBAR M. J. (1980) Beta-adrenergic receptor localization by light microscopic autoradiography.
ScG37tf~~ N. I! 208, 137X.- 1380.
PA~.ACIOS J. M., Youwc III W. S. & KUHAK M. J. (1979) Autoradiographi~ localization of HI-histamine receptors in brain
using [~Hlmepyramine: Preliminary studies. Eur. J. P~~~~~~~i~. 58, 295-304. PALACIOS J. M., YOUN(; III W. S. & K~JHAR M. J. (1980) Autoradiographi~ localization of gamma-alninobutyrjc acid
receptors in rat cerebellum, Proc. naln. A&. Sri. U.S.A. 77, 670674.
PALKO~ITS M. & JAC‘OWO~ITZ D. M. (1974) Topographic atlas of ~~techoiamines and acetylcholiflesterasc-fnntaining
neurons in the rat brain. II. Hindhr~~in. J. Conrp. Ncwo~. 157. 29.43.
PI-KOL:T~A S. .I.. MosKowtTz M. A.. REIKHAR~ J. F. & SNurJt K S. H. (19801 ~eurotransmitter Receptor Binding in Bovine
Cerebral Microvessels. S&,jrcc,, N. X 208, 6 I O-6 I?. POLLAKI) H.. Btsctcocr: S.. LI.ORENS C. & SCRWARTZ J. C. (1976) Histamine and histidine decarboxylase in discrete nuclei
ol’ rat hypothalamus and the evidence f’or mast-cells in the median eminence. Erairl Rex 118, 509-513.
Smw.w~% .I. C. (1977) fjistaminergic mechanisms in brain. A. Rev. Phurmu~. Tozicoi. 17, 325-339.
S~I-~WAKT~ J. C. (1979) Histamine Receptors in Brain. L@ Sci. 25. 895-912. St.rrw~nrz J. C., BARBIN G.. BAU~RV M., GARBARC; M.. MAKTRES M. P. & VEnnn%E M. 1197911) Metabolism and function
of histamine in the brain. In Curretrt Deoeloptnonrs in Ps~c,hopftartnircoi~~ teds ES~MAN B. & VALZEL~~E L.), Vol. 5. pp.
173-261. Spectrum Publ., New York. SCHWARTZ J. C., BARBIN G.. DU~HEMIN A. M., GARBARG M., PALACI~S J. M., QUACH T. T. & ROSE C. (1980) Histamine
receptors in brain: Characterization by binding studies and bio~hemicaf effects. In R~~eptor.~~~r ~eu~~~r~l~~s~irt~~s nnd
&pride ff~}ri?~~~es @eds PEPEU G., KL!HAR M. J. & ENNA S. J.), pp. 169-182. Raven Press, New York.
ScrfwAarz J. C.. BARHIN Ci.. GAKBARG M.. LLORE~S C.. PALA( J. M. & POLLARD l-1. (1978) ~istaminer8~~ Systems in
Brain. In .itriro~rc.(*s irr ~~~~~~~J?~~~~~~~~~ a& ~~z~l~~~p~~~~~j~.s (ed SIMON P.) pp. 171- I80. Pergamon Press. Oxford.
SCHWARTZ J. C.. PALATIOS J. M.. BARBIN G.. QUASH T. T., GARRARC~ M., HAAS W. L. & WOLF P. (1979h) Histamine
receptors in mammalian brain: Characters and modifications studied electrophysiologically and biochemically. In
Nisrrrrnine Receptors (ed. YELLIN T. 0.). pp. 165-184. Spectrum Publications, New York.
SNYDER S. H. & TAYLOR K. M. (1972) Histamine in the brain: a neurotransmitter? In Perspectires in Netrrophurmclcolt)~~~~
a tribute to Julius Axefrod ted. SNYDER S. H.). pp. 43-73. Oxford University Press. New York.
STEVENSON J. A. F. (1969) Neural control of food and water intake. In The ~~p~rku~u~~s. (eds HAYMAKER W., AN~XRSC~N
E. & NANT~ W. J. H.). pp. 524-621. Charles C. Thomas Publishers, Springfield, IL.
TAYLOR K. M. (1975) Brain Histamine. In ~undb~~~ qf ~s~~b~)phar~~ff~~~~~~ (eds IVERSEN L. L., IVERSEN S. D. 6t SNYDER
S. H.), Vol. 3, pp. 327-379. Plenum Press. New York.
TRAN V. T.. CHANT R. S. L. & SNYDER S. H. (1978) Histamine Hr-receptors identi~ed in mammalian brain with
r3H]mepyramine. Proc. nutn. Accrd. Sci. L~.S.A. 75, 6290-6294.
TLKX++ISTO L. & ER~KSS~~ L. t 1979) Antid~uresis induced by inf~lsiol~s of histamine into the brain ventricles ol’ conscious hydrated goats. FIX J. ~~?~4~/~?u~. 54, IYI-201.
WAMSL~Y J. K.. I.EWE M. S., YOUNG III W. S. & Kt:n~a M. J. ($981) Autoradiographi~ localization of muscarinia cholinergic receptors in rat brain. J. Nrurosc?., in press.
WAMsLt:V J. K.. ZARntN M. A., BIRIXALL N. J. M. & KUHAR M. J. (1980) Muscarinic cholinergic receptors: Autoradio- graphic localization of high and low affmity agonist binding sites, Brcrin Res. 200, 1-12.
WYNC~AARDEN J. B. & SEEVERS M. H. (1951) The toxic effects of antihistaminic drugs. J. Am. med. dss, 145, 277-282. YWNC; III W. S. & KUHAR M. J. (1979a) A new method for receptor autoradiography: [3H]opioid receptor labeling in
mounted tissue sections. Bruit, Ras. 179, 255-270.
YO~JNG 111 W. S. & K<jHAn M. J. (19796) Autoradiographic localization of benzodiazepine receptors in the brain of humans and animals. .l’urt,rt~. Loud. 280, 39.3395.
YOUIVC; III W. S. & KUHA~C M. J. (1979~) Neurotensin receptors: Autoradiographic localization in rat CNS. our. 1. ~~~~~~~U~,. 59, 161 --I 63.
Yr)t!~; I?f W. S. d Kt1ff.m M. J. (~979d) ~o~dren~rgic alpha-l and alpha-l receptors: Atitor~ldi~~gr~~h~c V~suaii~~ti~~~. Ew. J. Pha!w?nC. 59, 3 1 7 3 19.
Y()~‘HG IfI W. S. & KUHAK M. J. fi980u) Radiohist~hemical localization of benzodiazepine receptors in rat brain, ,I, P/trrri,r~rc. c.cp. T-her. 212. 327-346.
YrrtI~ IfI W. S. & KllHAn M. J. (198Oh) Noradrenergic alpha-l and alpha-2 receptors: Light microscopic autoradjo graphic localization. Proc. mm. Actrd. Sci. U.S.A. 77, I 696” I 700.
Copyright © 2022 FDOKUMEN




















