
The differentiation antigen NY-BR-1 is a potential target for antibody-based therapies in breast...
8
The differentiation antigen NY-BR-1 is a potential target for antibody-based therapies in breast cancer Inka Seil 1 , Claudia Frei 2 , Holger S€ ultmann 3 , Shirley K. Knauer 1 , Knut Engels 4 , Elke J€ ager 5 , Kurt Zatloukal 6 , Michael Pfreundschuh 7 , Alexander Knuth 8 , Yao Tseng-Chen 9 , Achim A. Jungbluth 10 , Roland H. Stauber 1,11 and Dirk J€ ager 2 * 1 Georg-Speyer-Haus, Institute for Biomedical Research, Frankfurt, Germany 2 Medical Oncology, National Center for Tumor Diseases, University Hospital Heidelberg, Heidelberg, Germany 3 Division of Molecular Genome Analysis, German Cancer Research Center, Heidelberg, Germany 4 Department of Pathology, University Hospital of Frankfurt, Germany 5 II. Medizinische Klinik, Krankenhaus Nordwest, Frankfurt, Germany 6 Department of Pathology, University of Graz, Graz, Austria 7 Universit € atsklinikum des Saarlandes, Homburg/Saar, Germany 8 Department of Oncology, University Hospital Zurich, Switzerland 9 Department of Pathology, Weill Medical College of Cornell University, New York, NY 10 Ludwig Institute for Cancer Research, New York Branch at Memorial Sloan-Kettering Cancer Center, New York, NY 11 Department of Otorhinolaryngology, Molecular and Cellular Oncology, University Hospital of Mainz, Germany Antibody-based cancer immunotherapy relies on the identification and characterization of target antigens and the development of potent antibodies recognizing the target. Here we report the expression analysis and molecular characterization of the differ- entiation antigen NY-BR-1, which we previously identified by using the SEREX (serological analysis of recombinant cDNA expression libraries) method. Corroborating methodologies, including mRNA quantitation and immunoblotting show that NY- BR-1 is strongly expressed in >70% of 129 breast tumors. Appli- cation of a NY-BR-1 specific antibody demonstrated NY-BR-1 expression in primary and metastastic breast cancers. In contrast, most of the breast cancer cell lines tested, expressed only low NY- BR-1 levels. Importantly, confocal microscopy revealed that ectopically expressed NY-BR-1 localizes to the cytoplasm and the cell membrane. NY-BR-1 localization in breast cancer specimens was also confirmed by immunohistochemistry. Bioinformatic anal- ysis and deletion mutagenesis further show that NY-BR-1 mem- brane localization is mediated by 2 cis-active membrane targeting domains. Biochemical surface labeling and FACS analysis of live cells further characterize NY-BR-1 as a transmembrane protein, which can be specifically recognized by the anti-NY-BR-1 anti- body on the surface of vital cells. The strong expression of NY- BR-1 in breast tumors, its cytoplasmic and membrane localization and accessibility to an ectopically applied antibody now suggest to pursue NY-BR-1 as a potential target for antibody-based therapies in breast cancer patients. ' 2007 Wiley-Liss, Inc. Key words: breast cancer; tumor antigen; immunotherapy Breast cancer is a worldwide health problem affecting more than 10% of women in the western world. 1 Recently, the rates of metastasis and mortality in breast cancer patients have decreased as a result of early diagnosis by mammographic screening, and the use of systemic adjuvant therapy. However, there is still a need to develop novel therapeutic strategies and to improve our under- standing in breast cancer development and disease progression. Recent clinical success with antibodies in cancer therapy has rein- vigorated interest in the identification and validation of novel anti- gens for antibody-based therapy, mostly cell surface proteins (reviewed in Ref. 2). Antigen-specific immunotherapeutic approaches, both cell- mediated and antibody-based, rely on the identification of target antigens expressed in cancer cells. The most promising target anti- gens are those that show tumor specific expression and homoge- nous distribution throughout the tumor. 3 In general, tumor antigens that are accessible to systemic or locally applied antibodies are considered as the most promising targets for antibody-based immunotherapy (reviewed in Refs. 4 and 5). Among various strat- egies used for target identification, 6 identification of antigens by serological expression cloning (SEREX) 7 resulted in the discovery of several tumor-associated antigens. Although no biological func- tions have been described for most of these antigens, some of these tumor antigens were already evaluated as targets and are currently successfully exploited in clinical cancer-immunotherapy trials. 8,9 We previously used SEREX to identify a novel breast cancer differentiation antigen, designated as NY-BR-1. 10,11 In prelimi- nary studies, NY-BR-1 mRNA was detected predominantly in breast cancers, 11–13 and bioinformatic analysis annotated NY-BR- 1 as a potential transcription factor. NY-BR-1 protein was shown to be expressed in normal breast epithelia and in the majority of primary breast cancers. 14,15 It has been shown that NY-BR-1- derived HLA-A2-restricted epitopes are being processed and pre- sented and can be recognized by CD8 T cells. 10 To further understand the biological function of NY-BR-1 and its potential for therapeutic approaches, we systematically analyzed NY-BR-1 protein expression and intracellular localization in vitro as well as in vivo. Our in vitro data provide the first evidence that NY-BR-1 encodes a membrane protein accessible to a generated monoclonal antibody. Thus, our study now positions NY-BR-1 in the line of potential target antigens for antibody-based immuno- therapy in addition to T-cell-based approaches in breast cancer. Material and methods Patient material Tissue, pleural effusions and serum samples were obtained from the University Hospital of Zurich, the Krankenhaus Nordw- est Frankfurt, the University Hospital of Homburg and the Depart- ments of Pathology University of Graz and Frankfurt. Patients’ Inka Seil’s current address is: Medical Oncology, National Center for Tumor Diseases, University Hospital Heidelberg, Heidelberg, Germany The last two authors contributed equally to this work. Received 21 July 2006; Accepted after revision 19 December 2006 DOI 10.1002/ijc.22620 Publishedonline28February2007inWileyInterScience(www.interscience. wiley.com). This article contains supplementary material available via the Internet at http://www.interscience.wiley.com/jpages/0020-7136/suppmat Grant sponsor: EU FP6 BRECOSM; Grant sponsor: BMBF; Grant num- bers: NGFN 01GS0451, N1KR-S12T22; Grant sponsor: Cancer Research Institute, New York. *Correspondence to: Medical Oncology, National Center for Tumor Diseases, University Hospital Heidelberg, Heidelberg, Germany. Fax: 49-6221-56-7225. E-mail: [email protected] Int. J. Cancer: 120, 2635–2642 (2007) ' 2007 Wiley-Liss, Inc. Publication of the International Union Against Cancer
Transcript of The differentiation antigen NY-BR-1 is a potential target for antibody-based therapies in breast...

The differentiation antigen NY-BR-1 is a potential target for antibody-based
therapies in breast cancer
Inka Seil1, Claudia Frei2, Holger S€ultmann3, Shirley K. Knauer1, Knut Engels4, Elke J€ager5, Kurt Zatloukal6,Michael Pfreundschuh7, Alexander Knuth8, Yao Tseng-Chen9, Achim A. Jungbluth10,Roland H. Stauber
1,11and Dirk J€ager2*
1Georg-Speyer-Haus, Institute for Biomedical Research, Frankfurt, Germany2Medical Oncology, National Center for Tumor Diseases, University Hospital Heidelberg, Heidelberg, Germany3Division of Molecular Genome Analysis, German Cancer Research Center, Heidelberg, Germany4Department of Pathology, University Hospital of Frankfurt, Germany5II. Medizinische Klinik, Krankenhaus Nordwest, Frankfurt, Germany6Department of Pathology, University of Graz, Graz, Austria7Universit€atsklinikum des Saarlandes, Homburg/Saar, Germany8Department of Oncology, University Hospital Zurich, Switzerland9Department of Pathology, Weill Medical College of Cornell University, New York, NY10Ludwig Institute for Cancer Research, New York Branch at Memorial Sloan-Kettering Cancer Center, New York, NY11Department of Otorhinolaryngology, Molecular and Cellular Oncology, University Hospital of Mainz, Germany
Antibody-based cancer immunotherapy relies on the identificationand characterization of target antigens and the development ofpotent antibodies recognizing the target. Here we report theexpression analysis and molecular characterization of the differ-entiation antigen NY-BR-1, which we previously identified byusing the SEREX (serological analysis of recombinant cDNAexpression libraries) method. Corroborating methodologies,including mRNA quantitation and immunoblotting show that NY-BR-1 is strongly expressed in >70% of 129 breast tumors. Appli-cation of a NY-BR-1 specific antibody demonstrated NY-BR-1expression in primary and metastastic breast cancers. In contrast,most of the breast cancer cell lines tested, expressed only low NY-BR-1 levels. Importantly, confocal microscopy revealed thatectopically expressed NY-BR-1 localizes to the cytoplasm and thecell membrane. NY-BR-1 localization in breast cancer specimenswas also confirmed by immunohistochemistry. Bioinformatic anal-ysis and deletion mutagenesis further show that NY-BR-1 mem-brane localization is mediated by 2 cis-active membrane targetingdomains. Biochemical surface labeling and FACS analysis of livecells further characterize NY-BR-1 as a transmembrane protein,which can be specifically recognized by the anti-NY-BR-1 anti-body on the surface of vital cells. The strong expression of NY-BR-1 in breast tumors, its cytoplasmic and membrane localizationand accessibility to an ectopically applied antibody now suggest topursue NY-BR-1 as a potential target for antibody-based therapiesin breast cancer patients.' 2007 Wiley-Liss, Inc.
Key words: breast cancer; tumor antigen; immunotherapy
Breast cancer is a worldwide health problem affecting morethan 10% of women in the western world.1 Recently, the rates ofmetastasis and mortality in breast cancer patients have decreasedas a result of early diagnosis by mammographic screening, and theuse of systemic adjuvant therapy. However, there is still a need todevelop novel therapeutic strategies and to improve our under-standing in breast cancer development and disease progression.Recent clinical success with antibodies in cancer therapy has rein-vigorated interest in the identification and validation of novel anti-gens for antibody-based therapy, mostly cell surface proteins(reviewed in Ref. 2).
Antigen-specific immunotherapeutic approaches, both cell-mediated and antibody-based, rely on the identification of targetantigens expressed in cancer cells. The most promising target anti-gens are those that show tumor specific expression and homoge-nous distribution throughout the tumor.3 In general, tumor antigensthat are accessible to systemic or locally applied antibodies areconsidered as the most promising targets for antibody-based
immunotherapy (reviewed in Refs. 4 and 5). Among various strat-egies used for target identification,6 identification of antigens byserological expression cloning (SEREX)7 resulted in the discoveryof several tumor-associated antigens. Although no biological func-tions have been described for most of these antigens, some of thesetumor antigens were already evaluated as targets and are currentlysuccessfully exploited in clinical cancer-immunotherapy trials.8,9
We previously used SEREX to identify a novel breast cancerdifferentiation antigen, designated as NY-BR-1.10,11 In prelimi-nary studies, NY-BR-1 mRNA was detected predominantly inbreast cancers,11–13 and bioinformatic analysis annotated NY-BR-1 as a potential transcription factor. NY-BR-1 protein was shownto be expressed in normal breast epithelia and in the majority ofprimary breast cancers.14,15 It has been shown that NY-BR-1-derived HLA-A2-restricted epitopes are being processed and pre-sented and can be recognized by CD8 T cells.10
To further understand the biological function of NY-BR-1 andits potential for therapeutic approaches, we systematically analyzedNY-BR-1 protein expression and intracellular localization in vitroas well as in vivo. Our in vitro data provide the first evidence thatNY-BR-1 encodes a membrane protein accessible to a generatedmonoclonal antibody. Thus, our study now positions NY-BR-1 inthe line of potential target antigens for antibody-based immuno-therapy in addition to T-cell-based approaches in breast cancer.
Material and methods
Patient material
Tissue, pleural effusions and serum samples were obtainedfrom the University Hospital of Zurich, the Krankenhaus Nordw-est Frankfurt, the University Hospital of Homburg and the Depart-ments of Pathology University of Graz and Frankfurt. Patients’
Inka Seil’s current address is: Medical Oncology, National Center forTumor Diseases, University Hospital Heidelberg, Heidelberg, Germany
The last two authors contributed equally to this work.Received 21 July 2006; Accepted after revision 19 December 2006DOI 10.1002/ijc.22620Publishedonline28February2007inWileyInterScience(www.interscience.
wiley.com).
This article contains supplementary material available via the Internet athttp://www.interscience.wiley.com/jpages/0020-7136/suppmatGrant sponsor: EU FP6 BRECOSM; Grant sponsor: BMBF; Grant num-
bers: NGFN 01GS0451, N1KR-S12T22; Grant sponsor: Cancer ResearchInstitute, New York.*Correspondence to: Medical Oncology, National Center for Tumor
Diseases, University Hospital Heidelberg, Heidelberg, Germany.Fax: 49-6221-56-7225. E-mail: [email protected]
Int. J. Cancer: 120, 2635–2642 (2007)' 2007 Wiley-Liss, Inc.
Publication of the International Union Against Cancer

and institutional ethics approval was obtained for all material usedin this study, and samples were processed anonymously. All caseswere diagnosed histopathologically and staged according to theTNM classification. Tissue specimens were flash frozen in liquidnitrogen and stored until extraction of mRNA after removal ofportions needed for pathological diagnosis. Histological analysesensured that tumor specimen contained >70% tumor tissue and<10% necrotic debris, and samples not meeting these criteriawere rejected.
RNA extraction, reverse transcription and quantitative PCR
Total RNA was purified from patient material or cells usingTRIzol1 reagent (Invitrogen Life Technologies, Karlsruhe, Ger-many) as described.16 Changes in mRNA levels were comparedby reverse transcription (RT) and subsequent quantitative real-time PCR analysis (qRT-PCR) as described.16 The expression ra-tio (R) of the target gene NY-BR-1 is calculated using the equa-tion: Ratio5 2(CT b-actin -CT NY-BR-1). SYBR green was used as flu-orescent dye. Alternatively, qualitative RT-PCR was performed asdescribed.11 Primers were: NY-BR-1, 50-CAAGAGCTCTG-CAGTGTGAGATTG-30 (sense) and 50-CTGGTATTGGTGTTT-CAGTGTGGC-30 (antisense); b-actin, 50-GCGGGAAATC-GTGCGTGACATT-30 (sense) and 50-GATGGAGTTGAAGG-TAGTTTCGTG-30 (antisense).
Probe preparation and microarray expression analysis
Custom-designed cDNA arrays consisting of 8,441 breast-spe-cific cDNA clones17 were hybridized in duplicate with Cy-5 la-beled tumor sample cDNA as described.18 The Universal HumanReference RNA (UHRR; Stratagene, La Jolla, CA), labeled withCy-3, served as a reference. On the array, NY-BR-1 was repre-sented by the RZPD-clone ANKRD30A (UniGene Hs.373787).Image quantification, data analysis and normalization were per-formed as described, and NY-BR-1 expression levels in tumorswere normalized to UHRR.18,19 Values from duplicate spots andduplicate microarrays for each cDNA clone were averaged.
Plasmids
pc3NY-BR-1_aa1/1397 encodes full length NY-BR-1.10 Theplasmid was constructed by PCR amplification and cloned intopcDNA 3 (Invitrogen Life Technologies, Karlsruhe, Germany).Tagged as well as untagged full length and deletion NY-BR-1expression constructs (Table I) were constructed by PCR cloning asdescribed.20 pc3FLAG_NY-BR-1, pc3FLAG_NY-BR-1_1/367,encodes N-terminal fusions of the FLAG-epitope and the indicatedNY-BR-1 amino acids (aa). pc3NY-BR-1_GFP, pc3NY-BR-1_609/1125_GFP and pc3NY-BR-1_908/1397_GFP encode NY-BR-1_GFP fusion proteins. pNY-BR-1_908/1397_RevM10_BFPencodes a fusion of the NY-BR-1 aa 908-1397 and the HIV-1RevM10BL_BFP fusion protein.21 pc3_Survivin_GFP encodessurvivin-GFP, and pc3FasL_FLAG encodes a FLAG-epitopetagged version of the Fas-ligand. The bacterial expression con-structs pET24a-NY-BR-1_851/928 expresses the His-tagged NY-BR-1 aa 851-928 from pET24a (EMDBioscience, San Diego, CA).
Cell culture, immunofluorescence and imaging of cells
Cell lines used in the study (Supplementary Table SII) weremaintained under conditions recommended by the American TypeCulture Collection and transfected as described.21 Peritoneal andpleural effusions were obtained at disease recurrence from patientsdiagnosed with breast cancer. Effusion cells were gained by cen-trifugation (1,200g, 10 min, 4�C) and, in the case of macroscopicblood contamination, subjected to density gradient separation overFicoll–Hypaque (Sigma Aldrich, Munich, Germany). Immunoflu-orescence and microscopy using a Leica TCS SL LSM confocalmicroscope (Leica Microsystems GmbH, Wetzlar, Germany) oran inverted fluorescence microscope (AxioVert200; Carl ZeissAG, G€ottingen, Germany) was carried out according to standardprocedures.21,22 Staining was performed using the monoclonalanti-FLAG antibody M2 (1:500, Sigma Aldrich, Munich,Germany) or a generated mouse monoclonal anti-NY-BR-1 anti-body (1:100). For surface staining of live cells, cells were incubatedwith primary antibodies at 37�C in the presence of NaN3 prior tofixation. Rhodamine conjugated wheat germ agglutinin (WGA)(Invitrogen Life Technologies, Karlsruhe, Germany) was used forstaining of the cell membrane, and nuclei were marked withTO-PRO1-3 iodide (Invitrogen Life Technologies, Karlsruhe,Germany).21 Internalization experiments were carried out by incu-bating transfected 293T cells with the monoclonal antibody withor without Protein A (Sigma Aldrich, Munich, Germany) to allowantibody complex formation. Cells were fixed at different timepoints (30, 60, 90, 120 and 180 min), permeabilized and antigen–antibody complexes were stained with a FITC or Cy3 conjugatedanti-mouse antibody (1:500, Dianova, Hamburg, Germany). Apolyclonal anti-Lamp-1 antibody (1:500, Sigma Aldrich, Munich,Germany) served as an endosomal marker. To determine the aver-age intracellular localization of the respective proteins, at least100 fluorescencent cells from 2 separate images were examined.The number of cells exhibiting cytoplasmic (C), plasma mem-brane (P) or nuclear (N) fluorescence was counted, and standarddeviations were determined.
Generation of NY-BR-1 specific monoclonal antibodies
The generation of anti-NY-BR-1#2 mAb was previouslydescribed.15 Briefly, a recombinant His-tagged peptide (NY-BR-1aa 851-928), showing no homologies to other known proteins, wascloned into pET24a and purified from E. coli by Ni-NTA Sephar-ose chromatography (EMD Bioscience, San Diego, CA) accordingto the manufacturer’s recommendations. The recombinant peptidewas used to immunize BALB/c mice for the generation of mono-clonal antibody. A positive hybridoma clone was identified byELISA and immunoblot analysis using the His-tagged NY-BR-1aa 851-928 peptide as described.15
Immunoblotting and antibodies
Immunoblotting of whole cell lysates was performed accordingto the standard protocols20. Tissue specimens were homogenizedusing an Ultra-Turrax T25 homogenizer (IKA Labortechnik) in0.5 ml modified RIPA buffer (50 mM Tris-HCl, pH 7.4, 150 mM
TABLE I – SUMMARY OF NY-BR-1 EXPRESSION CONSTRUCTS AND NY-BR-1’S INTRACELLULAR LOCALIZATION FOLLOWING ECTOPIC EXPRESSION
Plasmid Protein expressed (NY-BR-1 aa)Cellular localization
P/C C N Cell lines tested
pc3NY-BR-1 f.l., NY-BR-1 (aa 1-1397) 11 1 2 293, H, M7pc3FLAG-NY-BR-1 f.l., FLAG-NY-BR-1 (aa 1-1397) 11 1 2 293, H, M7pc3FLAG-NY-BR-1_1-367 FLAG-NY-BR-1 (aa 1-367) 11 1 2 293, Hpc3NY-BR-1-GFP f.l., NY-BR-1-GFP (aa 1-1397) 11*P 1*1 2*2 293, H, M7, M10pc3NY-BR-1_609/1125-GFP NY-BR-1-GFP (aa 609-1125) 2 11 1 293, H, M7pc3NY-BR-1_908/1397-GFP NY-BR-1-GFP (aa 908-1397) 11 1 2 293, H, M7pc3NY-BR-1_908/1397-RevM10BL-BFP NY-BR-1-RevM10BL-BFP (aa 908-1397) 11 1 2 293, HpET24a-NY-BR-1_851/928 63His_NY-BR-1 (aa 851-928) n.d.
P/C, plasma membrane/cytoplasm; C, cytoplasm; N, nucleus; 293, 293T cells; H, HeLa cells; M7, MCF-7 cells; M10, MCF-10A cells; f.l.,full length; *, living cells.
2636 SEIL ET AL.

NaCl, 0.1% SDS, 1 mM EDTA, 1% Na-deoxycholate, 1% TritonX-100) supplemented with protease inhibitor cocktail (RocheDiagnostics, Mannheim, Germany). Subsequently, the lysateswere centrifuged (30 min, 13,000g, 4�C) and the soluble and theinsoluble membrane fractions were subjected to immunoblot anal-ysis. The following antibodies were used: the anti-NY-BR-1 anti-body (NYmAb) (1:1,000), anti-GFP antibody (1:1,000; BD Bio-sciences, Heidelberg, Germany), anti-ErbB2 (1:500; Santa CruzBiotechnology, Heidelberg, Germany), anti-b actin (1:1,000;Sigma Aldrich, Munich, Germany), anti-FLAG antibody M2(1:500, Sigma Aldrich, Munich, Germany) or serum from breastcancer patients (1:100). Mouse IgG (1:500; Sigma Aldrich, Mu-nich, Germany) served to control unspecific binding, and appropri-ate secondary antibodies were used.
Flow cytometry analysis
For cell surface protein detection, fixed or unfixed cells wereincubated with saturating amounts of the respective antibodies,and analyzed according to standard protocols as described22. Thefollowing antibodies were used: anti-NY-BR-1 NYmAb (1:200),anti-survivin antibody (1:1,000; Novus Biologicals, Littleton, CO)and APC-conjugated goat anti-mouse Ig (1:100; BD Pharmingen,San Diego, CA).
Immunohistochemistry
All tissue samples were fixed in 4% buffered formalin for 12 hr.The presence of breast carcinoma was confirmed on a conven-tional haematoxylin-eosin stain. Immunohistochemical staining ofFFPE NY-BR-1 transfectants and patient tissues were performedaccording to standard procedures.15,23 Briefly, sections weremounted on Superfrost PlusTM glass slides (Microm, Walldorf,Germany) and dewaxed through graded alcohols. The tyramineamplification technique (CSA-Kit; DakoCytomation GmbH, Ham-burg, Germany) was used and antigen retrieval was performed bypretreatment in a pressure cooker with EDTA buffer (1 mM, pH8.0). Expression of NY-BR-1 was highlighted using the monoclo-nal a-NY-BR-1 antibody (1:600; 2 lg/mL). For visualization, theEnVision1 detection system (DakoCytomation GmbH, Hamburg,Germany) conjugated with horseradish peroxidase and 3,3 diami-nobenzidine as the chromogen was used according to the manu-facturers guidelines. Sections were counterstained with haematox-ylin and examined by light microscopy. The intensity of the im-munostaining in tumor cells and surrounding cells was evaluatedindependently by 2 pathologists blinded and scored semiquantita-tively as: �, negative; 1, weak; 11, moderate and 111,strongly positive.
Cell surface protein biotinylation
The cell surface proteins of live confluent 293T transfectantswere labeled at 4�C with membrane-impermeable EZ-Link Sulfo-NHS-SS-Biotin (Perbio, Bonn, Germany). After several washingsteps with TBS, whole cell lysates (input) were subjected to pre-cipitation with neutravidin beads and eluted with 50 mM DTT in13SDS-PAGE sample buffer according to the manufacturer’sinstructions. As a control, nonbiotinylated cell extracts were incu-bated with neutravidin beads. Proteins were identified by immuno-blotting.
Results
NY-BR-1 is overexpressed in most breast cancers
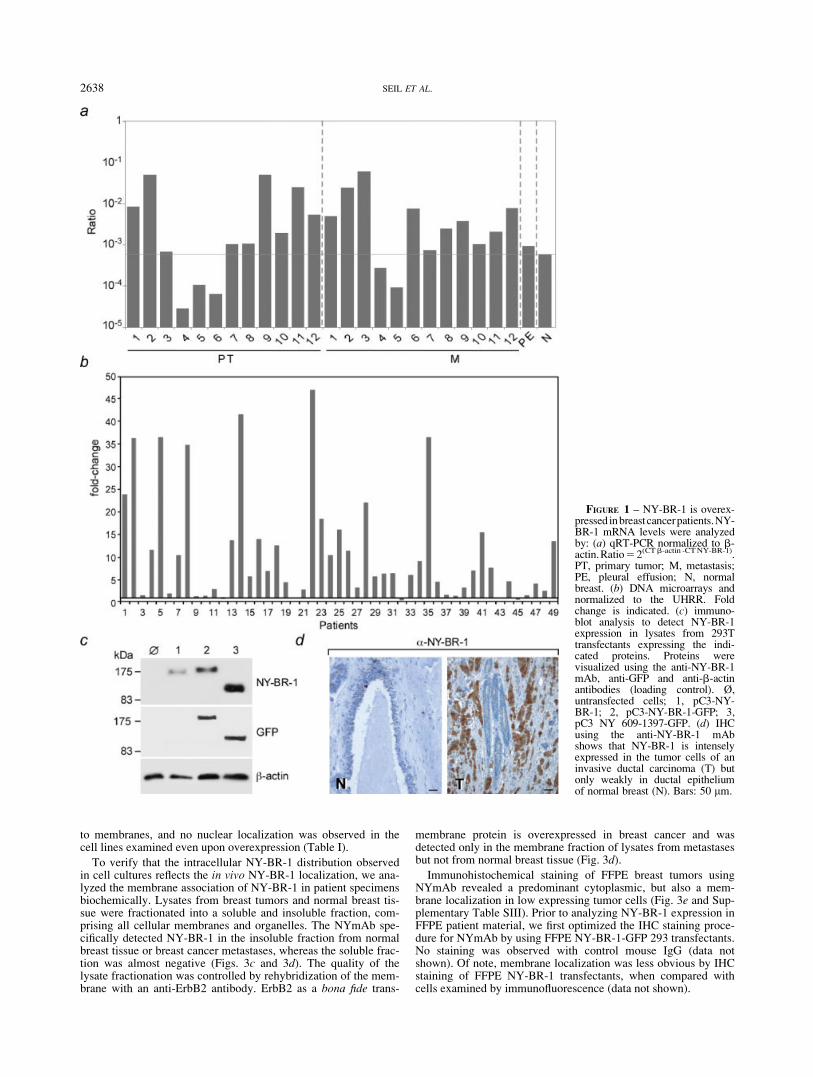
To systematically examine the expression of NY-BR-1 in breastcancer patients, we analyzed 129 patients, who underwent surgeryfor breast cancer. NY-BR-1 mRNA levels in patients from Ger-many, Switzerland and Austria were analyzed by microarray aswell as by quantitative and qualitative RT-PCR analysis. NY-BR-1 was found overexpressed in primary tumors (PT) as well as inbreast cancer metastasis (M) in 80% of the patients analyzed byqRT-PCR (20 out of 25 patients) (Fig. 1a). Microarray analysis of
49 patients demonstrated that in comparison to the UniversalHuman Reference RNA, NY-BR-1 was upregulated >2-fold in69% of the patients (34 out of 49 patients) (Fig. 1b, Supplemen-tary Table SI for patient characteristics). Likewise, 78% of thepatients examined by conventional RT-PCR analysis (43 out of55) were positive for NY-BR-1 RNA (Fig. 2a). No significantNY-BR-1 expression was detectable in normal tissues or carcino-mas from the endometrium and colon (data not shown). The fre-quency of NY-BR-1 mRNA expression in breast cancer is thushighly consistent using various methodologies and in differentpatient populations.
Next, NY-BR-1 protein expression was investigated using themonoclonal anti-NY-BR-1 antibody (NYmAb), raised againstrecombinant NY-BR-1 aa 851-928. NYmAb specifically recog-nized NY-BR-1 aa 851-928 in immunoblot analysis (data notshown). To demonstrate that NYmAb can also be used for thedetection of NY-BR-1 in eukaryotic systems, we examined itsspecificity by immunoblot analysis of lysates from 293T transfec-tants. As shown in Fig. 1c, NYmAb specifically reacts with NY-BR-1, NY-BR-1-GFP and NY-BR-1_609/1397-GFP. Stainingwith anti-GFP antibodies served to confirm the identity of thedetected proteins, and no unspecific proteins were recognized inlysates from NY-BR-1 negative 293T cells. Since NYmAb alsodetected NY-BR-1-GFP protein in formalin-fixed paraffin-embed-ded (FFPE) 293T transfectants (data not shown), we examined theNY-BR-1 expression by immunohistochemistry (IHC) in patientmaterial. NY-BR-1 was found strongly expressed in breast tumorcells, whereas normal ductal epithelium of the mammary glandstained only weakly for NY-BR-1 (Fig. 1d).
NY-BR-1 can induce antibody response in breast cancer patients
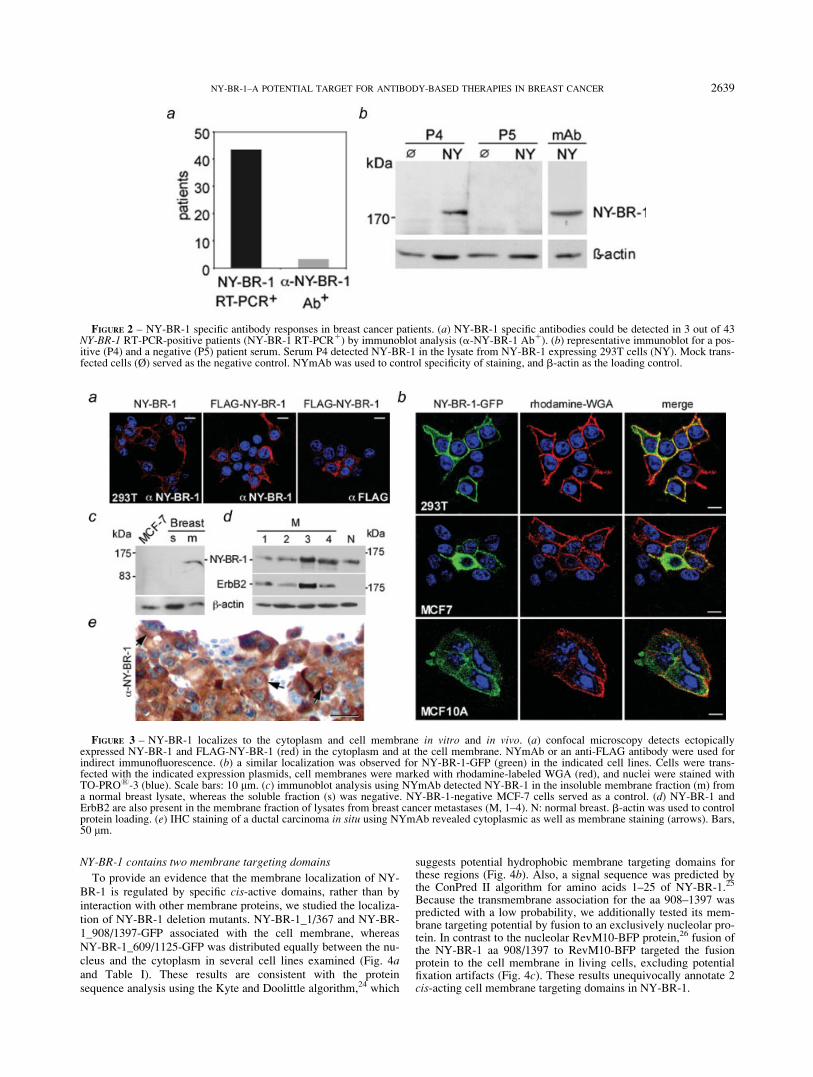
Since NY-BR-1 was identified by SEREX, we confirmed theimmunogenicity of endogenous NY-BR-1, by analyzing the fre-quency of spontaneous NY-BR-1 directed antibody responses in43 breast cancer patients, tested positive for NY-BR-1 by RT-PCR(Fig. 2a). NY-BR-1 specific antibodies were detected in 7% of thepatients (3 out of 43) and no immune response was detected nei-ther in NY-BR-1 negative patients nor in 12 healthy donors (notdepicted). So far, we have not systematically examined spontane-ous humoral immune responses in a larger cohort of healthy indi-viduals. Figure 2b shows a representative western blot for a posi-tive (P4) and a negative (P5) patient serum. Of note, NY-BR-1positive sera also detected NY-BR-1 in fixed 293T transfectantsby FACS analysis (not depicted).
NY-BR-1 localizes to the cytoplasm and the cell membrane
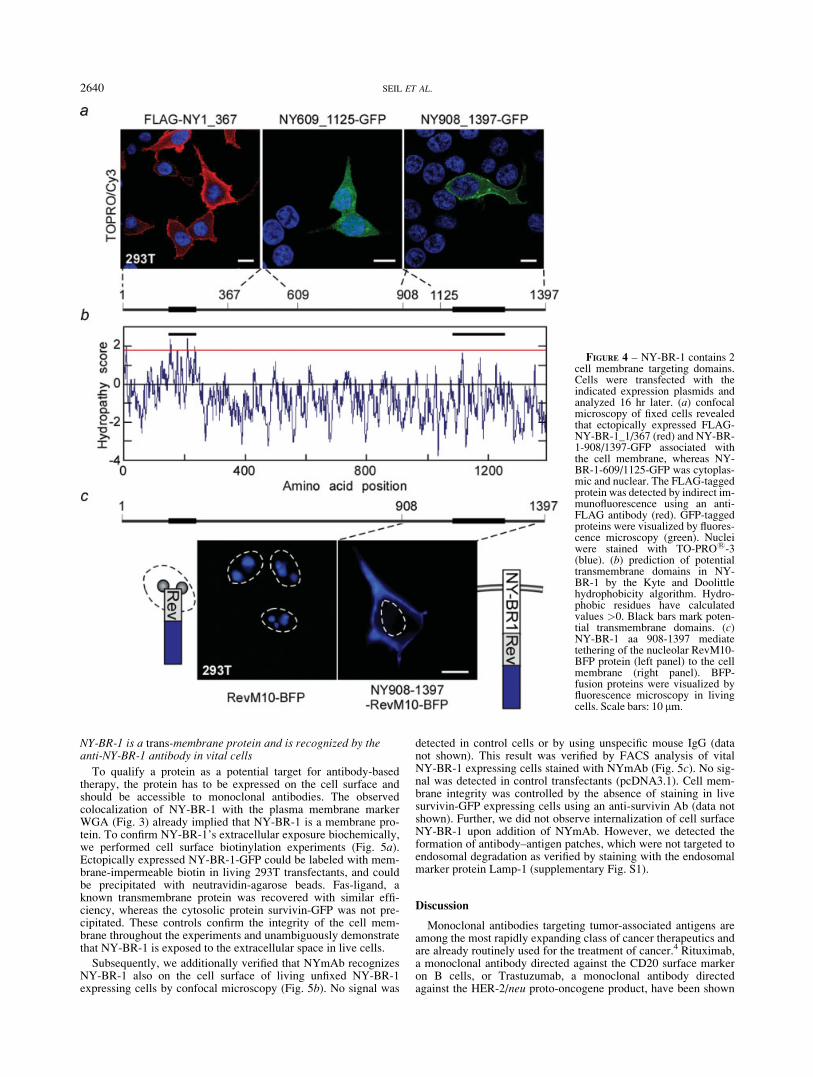
Cell culture models are valuable tools to dissect the biologicalfunctions of proteins. Thus, we first analyzed the expression of en-dogenous NY-BR-1 in several breast cancer and other tumor celllines by qRT-PCR and western blot. Surprisingly, in contrast tothe patient samples, NY-BR-1 expression was almost undetectablein the cell lines investigated, showing only low expression by RT-PCR in 3 out of 11 breast cancer cell lines (MDA-MB-453,BT474, T47D), and in none of the other cell lines tested (Supple-mentary Table SII). Since to date no appropriate cell line is suita-ble to study NY-BR-1’s subcellular localization, cells ectopicallyexpressing tagged or untagged full length or deletion NY-BR-1proteins were utilized. Interestingly, confocal fluorescence andindirect immunofluorescence microscopy showed that NY-BR-1,FLAG-NY-BR-1 (Fig. 3a) and NY-BR-1-GFP (Fig. 3b) localizeto the cytoplasm with a predominant membrane staining in livingor fixed cells (Table I). Cell membrane localization was controlledby marking the plasma membrane with rhodamine labeled WGA.Of note, cells that express low amounts of NY-BR-1, resemblingmore physiological expression levels, displayed a pronounced cellmembrane staining, while in strong expressors the cytoplasmaticstaining was more prominent. In the cytoplasm, full length NY-BR-1 is not homogeneously distributed, but appears to be attached
2637NY-BR-1–A POTENTIAL TARGET FOR ANTIBODY-BASED THERAPIES IN BREAST CANCER
to membranes, and no nuclear localization was observed in thecell lines examined even upon overexpression (Table I).
To verify that the intracellular NY-BR-1 distribution observedin cell cultures reflects the in vivo NY-BR-1 localization, we ana-lyzed the membrane association of NY-BR-1 in patient specimensbiochemically. Lysates from breast tumors and normal breast tis-sue were fractionated into a soluble and insoluble fraction, com-prising all cellular membranes and organelles. The NYmAb spe-cifically detected NY-BR-1 in the insoluble fraction from normalbreast tissue or breast cancer metastases, whereas the soluble frac-tion was almost negative (Figs. 3c and 3d). The quality of thelysate fractionation was controlled by rehybridization of the mem-brane with an anti-ErbB2 antibody. ErbB2 as a bona fide trans-
membrane protein is overexpressed in breast cancer and wasdetected only in the membrane fraction of lysates from metastasesbut not from normal breast tissue (Fig. 3d).
Immunohistochemical staining of FFPE breast tumors usingNYmAb revealed a predominant cytoplasmic, but also a mem-brane localization in low expressing tumor cells (Fig. 3e and Sup-plementary Table SIII). Prior to analyzing NY-BR-1 expression inFFPE patient material, we first optimized the IHC staining proce-dure for NYmAb by using FFPE NY-BR-1-GFP 293 transfectants.No staining was observed with control mouse IgG (data notshown). Of note, membrane localization was less obvious by IHCstaining of FFPE NY-BR-1 transfectants, when compared withcells examined by immunofluorescence (data not shown).
FIGURE 1 – NY-BR-1 is overex-pressed inbreastcancerpatients.NY-BR-1 mRNA levels were analyzedby: (a) qRT-PCR normalized to b-actin.Ratio52(CTb-actin -CTNY-BR-1).PT, primary tumor; M, metastasis;PE, pleural effusion; N, normalbreast. (b) DNA microarrays andnormalized to the UHRR. Foldchange is indicated. (c) immuno-blot analysis to detect NY-BR-1expression in lysates from 293Ttransfectants expressing the indi-cated proteins. Proteins werevisualized using the anti-NY-BR-1mAb, anti-GFP and anti-b-actinantibodies (loading control). Ø,untransfected cells; 1, pC3-NY-BR-1; 2, pC3-NY-BR-1-GFP; 3,pC3 NY 609-1397-GFP. (d) IHCusing the anti-NY-BR-1 mAbshows that NY-BR-1 is intenselyexpressed in the tumor cells of aninvasive ductal carcinoma (T) butonly weakly in ductal epitheliumof normal breast (N). Bars: 50 lm.
2638 SEIL ET AL.
NY-BR-1 contains two membrane targeting domains
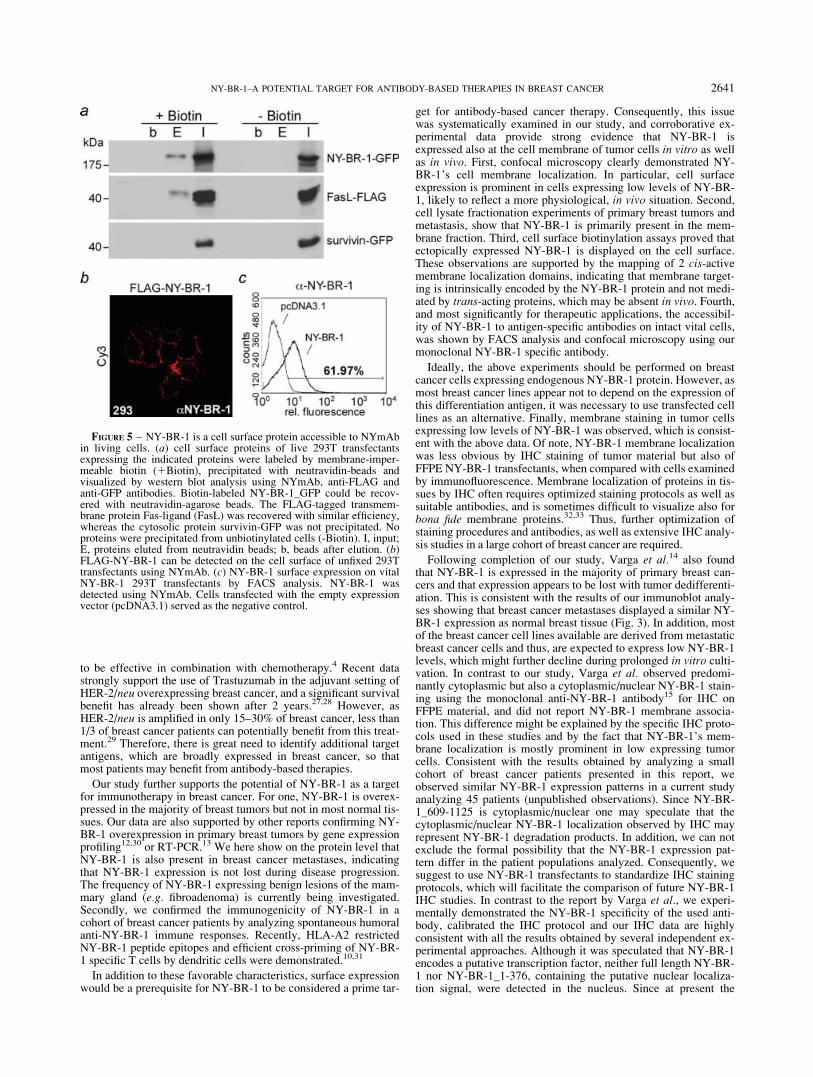
To provide an evidence that the membrane localization of NY-BR-1 is regulated by specific cis-active domains, rather than byinteraction with other membrane proteins, we studied the localiza-tion of NY-BR-1 deletion mutants. NY-BR-1_1/367 and NY-BR-1_908/1397-GFP associated with the cell membrane, whereasNY-BR-1_609/1125-GFP was distributed equally between the nu-cleus and the cytoplasm in several cell lines examined (Fig. 4aand Table I). These results are consistent with the proteinsequence analysis using the Kyte and Doolittle algorithm,24 which
suggests potential hydrophobic membrane targeting domains forthese regions (Fig. 4b). Also, a signal sequence was predicted bythe ConPred II algorithm for amino acids 1–25 of NY-BR-1.25
Because the transmembrane association for the aa 908–1397 waspredicted with a low probability, we additionally tested its mem-brane targeting potential by fusion to an exclusively nucleolar pro-tein. In contrast to the nucleolar RevM10-BFP protein,26 fusion ofthe NY-BR-1 aa 908/1397 to RevM10-BFP targeted the fusionprotein to the cell membrane in living cells, excluding potentialfixation artifacts (Fig. 4c). These results unequivocally annotate 2cis-acting cell membrane targeting domains in NY-BR-1.
FIGURE 2 – NY-BR-1 specific antibody responses in breast cancer patients. (a) NY-BR-1 specific antibodies could be detected in 3 out of 43NY-BR-1 RT-PCR-positive patients (NY-BR-1 RT-PCR1) by immunoblot analysis (a-NY-BR-1 Ab1). (b) representative immunoblot for a pos-itive (P4) and a negative (P5) patient serum. Serum P4 detected NY-BR-1 in the lysate from NY-BR-1 expressing 293T cells (NY). Mock trans-fected cells (Ø) served as the negative control. NYmAb was used to control specificity of staining, and b-actin as the loading control.
FIGURE 3 – NY-BR-1 localizes to the cytoplasm and cell membrane in vitro and in vivo. (a) confocal microscopy detects ectopicallyexpressed NY-BR-1 and FLAG-NY-BR-1 (red) in the cytoplasm and at the cell membrane. NYmAb or an anti-FLAG antibody were used forindirect immunofluorescence. (b) a similar localization was observed for NY-BR-1-GFP (green) in the indicated cell lines. Cells were trans-fected with the indicated expression plasmids, cell membranes were marked with rhodamine-labeled WGA (red), and nuclei were stained withTO-PRO1-3 (blue). Scale bars: 10 lm. (c) immunoblot analysis using NYmAb detected NY-BR-1 in the insoluble membrane fraction (m) froma normal breast lysate, whereas the soluble fraction (s) was negative. NY-BR-1-negative MCF-7 cells served as a control. (d) NY-BR-1 andErbB2 are also present in the membrane fraction of lysates from breast cancer metastases (M, 1–4). N: normal breast. b-actin was used to controlprotein loading. (e) IHC staining of a ductal carcinoma in situ using NYmAb revealed cytoplasmic as well as membrane staining (arrows). Bars,50 lm.
2639NY-BR-1–A POTENTIAL TARGET FOR ANTIBODY-BASED THERAPIES IN BREAST CANCER
NY-BR-1 is a trans-membrane protein and is recognized by theanti-NY-BR-1 antibody in vital cells
To qualify a protein as a potential target for antibody-basedtherapy, the protein has to be expressed on the cell surface andshould be accessible to monoclonal antibodies. The observedcolocalization of NY-BR-1 with the plasma membrane markerWGA (Fig. 3) already implied that NY-BR-1 is a membrane pro-tein. To confirm NY-BR-1’s extracellular exposure biochemically,we performed cell surface biotinylation experiments (Fig. 5a).Ectopically expressed NY-BR-1-GFP could be labeled with mem-brane-impermeable biotin in living 293T transfectants, and couldbe precipitated with neutravidin-agarose beads. Fas-ligand, aknown transmembrane protein was recovered with similar effi-ciency, whereas the cytosolic protein survivin-GFP was not pre-cipitated. These controls confirm the integrity of the cell mem-brane throughout the experiments and unambiguously demonstratethat NY-BR-1 is exposed to the extracellular space in live cells.
Subsequently, we additionally verified that NYmAb recognizesNY-BR-1 also on the cell surface of living unfixed NY-BR-1expressing cells by confocal microscopy (Fig. 5b). No signal was
detected in control cells or by using unspecific mouse IgG (datanot shown). This result was verified by FACS analysis of vitalNY-BR-1 expressing cells stained with NYmAb (Fig. 5c). No sig-nal was detected in control transfectants (pcDNA3.1). Cell mem-brane integrity was controlled by the absence of staining in livesurvivin-GFP expressing cells using an anti-survivin Ab (data notshown). Further, we did not observe internalization of cell surfaceNY-BR-1 upon addition of NYmAb. However, we detected theformation of antibody–antigen patches, which were not targeted toendosomal degradation as verified by staining with the endosomalmarker protein Lamp-1 (supplementary Fig. S1).
Discussion
Monoclonal antibodies targeting tumor-associated antigens areamong the most rapidly expanding class of cancer therapeutics andare already routinely used for the treatment of cancer.4 Rituximab,a monoclonal antibody directed against the CD20 surface markeron B cells, or Trastuzumab, a monoclonal antibody directedagainst the HER-2/neu proto-oncogene product, have been shown
FIGURE 4 – NY-BR-1 contains 2cell membrane targeting domains.Cells were transfected with theindicated expression plasmids andanalyzed 16 hr later. (a) confocalmicroscopy of fixed cells revealedthat ectopically expressed FLAG-NY-BR-1_1/367 (red) and NY-BR-1-908/1397-GFP associated withthe cell membrane, whereas NY-BR-1-609/1125-GFP was cytoplas-mic and nuclear. The FLAG-taggedprotein was detected by indirect im-munofluorescence using an anti-FLAG antibody (red). GFP-taggedproteins were visualized by fluores-cence microscopy (green). Nucleiwere stained with TO-PRO1-3(blue). (b) prediction of potentialtransmembrane domains in NY-BR-1 by the Kyte and Doolittlehydrophobicity algorithm. Hydro-phobic residues have calculatedvalues >0. Black bars mark poten-tial transmembrane domains. (c)NY-BR-1 aa 908-1397 mediatetethering of the nucleolar RevM10-BFP protein (left panel) to the cellmembrane (right panel). BFP-fusion proteins were visualized byfluorescence microscopy in livingcells. Scale bars: 10 lm.
2640 SEIL ET AL.
to be effective in combination with chemotherapy.4 Recent datastrongly support the use of Trastuzumab in the adjuvant setting ofHER-2/neu overexpressing breast cancer, and a significant survivalbenefit has already been shown after 2 years.27,28 However, asHER-2/neu is amplified in only 15–30% of breast cancer, less than1/3 of breast cancer patients can potentially benefit from this treat-ment.29 Therefore, there is great need to identify additional targetantigens, which are broadly expressed in breast cancer, so thatmost patients may benefit from antibody-based therapies.
Our study further supports the potential of NY-BR-1 as a targetfor immunotherapy in breast cancer. For one, NY-BR-1 is overex-pressed in the majority of breast tumors but not in most normal tis-sues. Our data are also supported by other reports confirming NY-BR-1 overexpression in primary breast tumors by gene expressionprofiling12,30 or RT-PCR.13 We here show on the protein level thatNY-BR-1 is also present in breast cancer metastases, indicatingthat NY-BR-1 expression is not lost during disease progression.The frequency of NY-BR-1 expressing benign lesions of the mam-mary gland (e.g. fibroadenoma) is currently being investigated.Secondly, we confirmed the immunogenicity of NY-BR-1 in acohort of breast cancer patients by analyzing spontaneous humoralanti-NY-BR-1 immune responses. Recently, HLA-A2 restrictedNY-BR-1 peptide epitopes and efficient cross-priming of NY-BR-1 specific T cells by dendritic cells were demonstrated.10,31
In addition to these favorable characteristics, surface expressionwould be a prerequisite for NY-BR-1 to be considered a prime tar-
get for antibody-based cancer therapy. Consequently, this issuewas systematically examined in our study, and corroborative ex-perimental data provide strong evidence that NY-BR-1 isexpressed also at the cell membrane of tumor cells in vitro as wellas in vivo. First, confocal microscopy clearly demonstrated NY-BR-1’s cell membrane localization. In particular, cell surfaceexpression is prominent in cells expressing low levels of NY-BR-1, likely to reflect a more physiological, in vivo situation. Second,cell lysate fractionation experiments of primary breast tumors andmetastasis, show that NY-BR-1 is primarily present in the mem-brane fraction. Third, cell surface biotinylation assays proved thatectopically expressed NY-BR-1 is displayed on the cell surface.These observations are supported by the mapping of 2 cis-activemembrane localization domains, indicating that membrane target-ing is intrinsically encoded by the NY-BR-1 protein and not medi-ated by trans-acting proteins, which may be absent in vivo. Fourth,and most significantly for therapeutic applications, the accessibil-ity of NY-BR-1 to antigen-specific antibodies on intact vital cells,was shown by FACS analysis and confocal microscopy using ourmonoclonal NY-BR-1 specific antibody.
Ideally, the above experiments should be performed on breastcancer cells expressing endogenous NY-BR-1 protein. However, asmost breast cancer lines appear not to depend on the expression ofthis differentiation antigen, it was necessary to use transfected celllines as an alternative. Finally, membrane staining in tumor cellsexpressing low levels of NY-BR-1 was observed, which is consist-ent with the above data. Of note, NY-BR-1 membrane localizationwas less obvious by IHC staining of tumor material but also ofFFPE NY-BR-1 transfectants, when compared with cells examinedby immunofluorescence. Membrane localization of proteins in tis-sues by IHC often requires optimized staining protocols as well assuitable antibodies, and is sometimes difficult to visualize also forbona fide membrane proteins.32,33 Thus, further optimization ofstaining procedures and antibodies, as well as extensive IHC analy-sis studies in a large cohort of breast cancer are required.
Following completion of our study, Varga et al.14 also foundthat NY-BR-1 is expressed in the majority of primary breast can-cers and that expression appears to be lost with tumor dedifferenti-ation. This is consistent with the results of our immunoblot analy-ses showing that breast cancer metastases displayed a similar NY-BR-1 expression as normal breast tissue (Fig. 3). In addition, mostof the breast cancer cell lines available are derived from metastaticbreast cancer cells and thus, are expected to express low NY-BR-1levels, which might further decline during prolonged in vitro culti-vation. In contrast to our study, Varga et al. observed predomi-nantly cytoplasmic but also a cytoplasmic/nuclear NY-BR-1 stain-ing using the monoclonal anti-NY-BR-1 antibody15 for IHC onFFPE material, and did not report NY-BR-1 membrane associa-tion. This difference might be explained by the specific IHC proto-cols used in these studies and by the fact that NY-BR-1’s mem-brane localization is mostly prominent in low expressing tumorcells. Consistent with the results obtained by analyzing a smallcohort of breast cancer patients presented in this report, weobserved similar NY-BR-1 expression patterns in a current studyanalyzing 45 patients (unpublished observations). Since NY-BR-1_609-1125 is cytoplasmic/nuclear one may speculate that thecytoplasmic/nuclear NY-BR-1 localization observed by IHC mayrepresent NY-BR-1 degradation products. In addition, we can notexclude the formal possibility that the NY-BR-1 expression pat-tern differ in the patient populations analyzed. Consequently, wesuggest to use NY-BR-1 transfectants to standardize IHC stainingprotocols, which will facilitate the comparison of future NY-BR-1IHC studies. In contrast to the report by Varga et al., we experi-mentally demonstrated the NY-BR-1 specificity of the used anti-body, calibrated the IHC protocol and our IHC data are highlyconsistent with all the results obtained by several independent ex-perimental approaches. Although it was speculated that NY-BR-1encodes a putative transcription factor, neither full length NY-BR-1 nor NY-BR-1_1-376, containing the putative nuclear localiza-tion signal, were detected in the nucleus. Since at present the
FIGURE 5 – NY-BR-1 is a cell surface protein accessible to NYmAbin living cells. (a) cell surface proteins of live 293T transfectantsexpressing the indicated proteins were labeled by membrane-imper-meable biotin (1Biotin), precipitated with neutravidin-beads andvisualized by western blot analysis using NYmAb, anti-FLAG andanti-GFP antibodies. Biotin-labeled NY-BR-1_GFP could be recov-ered with neutravidin-agarose beads. The FLAG-tagged transmem-brane protein Fas-ligand (FasL) was recovered with similar efficiency,whereas the cytosolic protein survivin-GFP was not precipitated. Noproteins were precipitated from unbiotinylated cells (-Biotin). I, input;E, proteins eluted from neutravidin beads; b, beads after elution. (b)FLAG-NY-BR-1 can be detected on the cell surface of unfixed 293Ttransfectants using NYmAb. (c) NY-BR-1 surface expression on vitalNY-BR-1 293T transfectants by FACS analysis. NY-BR-1 wasdetected using NYmAb. Cells transfected with the empty expressionvector (pcDNA3.1) served as the negative control.
2641NY-BR-1–A POTENTIAL TARGET FOR ANTIBODY-BASED THERAPIES IN BREAST CANCER

biological function of NY-BR-1 is completely unknown furtherstudies to elucidate the biological role of NY-BR-1 applying in-ducible cell culture and transgenic animal models are urgentlyneeded.
In conclusion, our study provides the first evidence that NY-BR-1 may represent a potential target antigen for antibody-basedtherapy in breast cancer. Its frequent expression in breast cancerpatients, its cell surface localization and accessibility to the mono-clonal antibody mark NY-BR-1 as an attractive target. Studies toevaluate the efficacy of our antibody in NY-BR-1 expressing tu-mor models are thus warranted. Treatment protocols targeting
NY-BR-1 by antibody and T cells can be envisioned as a promis-ing approach to treat breast cancer patients.
Acknowledgements
The authors thank Silke Deckert and Gert Carra for excellenttechnical assistance, Martin Z€ornig for advice with confocal mi-croscopy and for providing the FasL-FLAG expression constructand Bernd Groner for support. This work was supported by theEU FP6 BRECOSM (S.R.), the BMBF (NGFN 01GS0451/N1KR-S12T22 to S.R.) and the Cancer Research Institute, New York.
References
1. Levin M. Breast cancer: issues in risk reduction, screening and sur-veillance, early identification and therapy planning. Drug News Per-spect 2003;16:395–8.
2. Carter P, Smith L, Ryan M. Identification and validation of cell sur-face antigens for antibody targeting in oncology. Endocr Relat Cancer2004;11:659–87.
3. Jager E, Jager D, Knuth A. Antigen-specific immunotherapy and can-cer vaccines. Int J Cancer 2003;106:817–20.
4. Adams GP, Weiner LM. Monoclonal antibody therapy of cancer. NatBiotechnol 2005;23:1147–57.
5. Jager D, Knuth A. Antibodies and vaccines–hope or illusion? Breast2005;14:631–5.
6. Chen YT, Scanlan MJ, Venditti CA, Chua R, Theiler G, Stevenson BJ,Iseli C, Gure AO, Vasicek T, Strausberg RL, Jongeneel CV, Old LJ,et al. Identification of cancer/testis-antigen genes by massively parallelsignature sequencing. Proc Natl Acad Sci USA 2005;102:7940–5.
7. Chen YT. Cancer vaccine: identification of human tumor antigens bySEREX. Cancer J 2000;6(Suppl 3):S208–S217.
8. Jager E, Gnjatic S, Nagata Y, Stockert E, Jager D, Karbach J, Neu-mann A, Rieckenberg J, Chen YT, Ritter G, Hoffman E, Arand M,et al. Induction of primary NY-ESO-1 immunity: CD81 T lympho-cyte and antibody responses in peptide-vaccinated patients with NY-ESO-11 cancers. Proc Natl Acad Sci USA 2000;97:12198–203.
9. Simpson AJ, Caballero OL, Jungbluth A, Chen YT, Old LJ. Cancer/testis antigens, gametogenesis and cancer. Nat Rev Cancer 2005;5:615–25.
10. Jager D, Karbach J, Pauligk C, Seil I, Frei C, Chen YT, Old LJ, KnuthA, Jager E. Humoral and cellular immune responses against the breastcancer antigen NY-BR-1: definition of two HLA-A2 restricted peptideepitopes. Cancer Immun 2005;5:11.
11. Jager D, Stockert E, Gure AO, Scanlan MJ, Karbach J, Jager E, KnuthA, Old LJ, Chen YT. Identification of a tissue-specific putative tran-scription factor in breast tissue by serological screening of a breastcancer library. Cancer Res 2001;61:2055–61.
12. Jiang Y, Harlocker SL, Molesh DA, Dillon DC, Stolk JA, HoughtonRL, Repasky EA, Badaro R, Reed SG, Xu J. Discovery of differen-tially expressed genes in human breast cancer using subtracted cDNAlibraries and cDNA microarrays. Oncogene 2002;21:2270–82.
13. Nissan A, Jager D, Roystacher M, Prus D, Peretz T, Eisenberg I,Freund HR, Scanlan M, Ritter G, Old LJ, Mitrani-Rosenbaum S. Mul-timarker RT-PCR assay for the detection of minimal residual diseasein sentinel lymph nodes of breast cancer patients. Br J Cancer 2006;94:681–5.
14. Varga Z, Theurillat JP, Filonenko V, Sasse B, Odermatt B, JungbluthAA, Chen YT, Old LJ, Knuth A, Jager D, Moch H. Preferential nu-clear and cytoplasmic NY-BR-1 protein expression in primary breastcancer and lymph node metastases. Clin Cancer Res 2006;12:2745–51.
15. Jager D, Filonenko V, Gouth I, Frosina D, Eastlake-Wade S, CastelliS, Varga Z, Moch H, Chen YT, Busam KJ, Seil I, Old L, et al. NY-BR-1 is a differentiation antigen of the mammary gland. Appl Immu-nohistochem Mol Morphol, in press.
16. Schlingemann J, Habtemichael N, Ittrich C, Toedt G, Kramer H,Hambek M, Knecht R, Lichter P, Stauber RH, Hahn M. Patient-basedcross-platform comparison of oligonucleotide microarray expressionprofiles. Lab Invest 2005;85(8):1024–39.
17. Schneider J, Buness A, Huber W, Volz J, Kioschis P, Hafner M,Poustka A, Sultmann H. Systematic analysis of T7 RNA polymerasebased in vitro linear RNA amplification for use in microarray experi-ments. BMC Genomics 2004;5:29–38.
18. Sultmann H, von Heydebreck A, Huber W, Kuner R, Buness A, VogtM, Gunawan B, Vingron M, Fuzesi L, Poustka A. Gene expression inkidney cancer is associated with cytogenetic abnormalities, metastasisformation, and patient survival. Clin Cancer Res 2005;11:646–55.
19. Buness A, Huber W, Steiner K, Sultmann H, Poustka A. arrayMagic:two-colour cDNA microarray quality control and preprocessing. Bio-informatics 2005;21:554–6.
20. Knauer SK, Carra G, Stauber RH. Nuclear export is evolutionarilyconserved in CVC paired-like homeobox proteins and influences pro-tein stability, transcriptional activation, and extracellular secretion.Mol Cell Biol 2005;25:2573–82.
21. Knauer SK, Moodt S, Berg T, Liebel U, Pepperkok R, Stauber RH.Translocation biosensors to study signal specific nucleo-cytoplasmictransport, protease activity and protein interactions. Traffic 2005;6:1–13.
22. Stauber RH, Horie K, Carney P, Hudson EA, Tarasova NI, GaitanarisGA, Pavlakis GN. Development and applications of enhanced greenfluorescent protein mutants. BioTechniques 1998;24:462–71.
23. Lubomierski N, Plotz G, Wormek M, Engels K, Kriener S, Trojan J,Jungling B, Zeuzem S, Raedle J. BRAF mutations in colorectal carci-noma suggest two entities of microsatellite-unstable tumors. Cancer2005;104:952–61.
24. Kyte J, Doolittle RF. A simple method for displaying the hydropathiccharacter of a protein. J Mol Biol 1982;157:105–32.
25. Arai M, Mitsuke H, Ikeda M, Xia JX, Kikuchi T, Satake M, ShimizuT. ConPred II: a consensus prediction method for obtaining trans-membrane topology models with high reliability. Nucleic Acids Res2004;32:390–3.
26. Knauer SK, Stauber RH. Development of an autofluorescent translo-cation biosensor system to investigate protein-protein interactions inliving cells. Anal Chem 2005;77:4815–20.
27. Emens LA. Trastuzumab: targeted therapy for the management ofHER-2/neu-overexpressing metastatic breast cancer. Am J Ther 2005;12:243–53.
28. Vogel CL, Reddy JC, Reyno LM. Efficacy of trastuzumab. CancerRes 2005;65:2044.
29. Slamon DJ, Leyland-Jones B, Shak S, Fuchs H, Paton V, BajamondeA, Fleming T, Eiermann W, Wolter J, Pegram M, Baselga J, Norton L.Use of chemotherapy plus a monoclonal antibody against HER2 formetastatic breast cancer that overexpresses HER2. N Engl J Med2001;344:783–92.
30. Radvanyi L, Singh-Sandhu D, Gallichan S, Lovitt C, Pedyczak A,Mallo G, Gish K, Kwok K, Hanna W, Zubovits J, Armes J, Venter D,et al. The gene associated with trichorhinophalangeal syndrome inhumans is overexpressed in breast cancer. Proc Natl Acad Sci USA2005;102:11005–10.
31. Groh V, Li YQ, Cioca D, Hunder NN, Wang W, Riddell SR, Yee C,Spies T. From the cover: efficient cross-priming of tumor antigen-spe-cific T cells by dendritic cells sensitized with diverse anti-MICAopsonized tumor cells. Proc Natl Acad Sci USA 2005;102:6461–6.
32. Boor PK, de Groot K, Waisfisz Q, Kamphorst W, Oudejans CB,Powers JM, Pronk JC, Scheper GC, van der Knaap MS. MLC1: anovel protein in distal astroglial processes. J Neuropathol Exp Neurol2005;64:412–19.
33. Bei R, Budillon A, Masuelli L, Cereda V, Vitolo D, Di Gennaro E,Ripavecchia V, Palumbo C, Ionna F, Losito S, Modesti A, Kraus MH,et al. Frequent overexpression of multiple ErbB receptors by head andneck squamous cell carcinoma contrasts with rare antibody immunityin patients. J Pathol 2004;204:317–25.
2642 SEIL ET AL.




















