
Seasonal Dynamics of a Microtidal Pocket Beach with Posidonia oceanica Seabeds (Mallorca, Spain
Upload
independentCategory
view
0download
0
P.S.Z.N .I : Marine Ecology, 13 (1) : 69-83 (1992)
Accepted : May 24, 1991© 1992 Paul Parey Scientific Publishers, Berlin and HamburgISSN 0173-9565
The Detritic Compartment in aPosidonia oceanica Meadow :Litter Features, Decomposition Rates,and Mineral Stocks
JAVIER ROMERO', GÉRARD PERGENT2, CHRISTINE PERGENT-MARTIN2,MIGUEL-ANGEL MATEO' & CÉCILE REGNIER2
1 Departamento de Ecologfa, Facultad de Biologfa, Universidad de Barcelona,Diagonal 645, E-08028 Barcelona, Spain.
2 Laboratoire de Biologie Marine et d'Ecologie du Benthos, Faculté des Sciencesde Luminy, Université Aix-Marseille II, F-13288 Marseille Cedex 9, France .
With 4 figures and 8 tables
Key words: Seagrass, Posidonia oceanica, detritus, decomposition, mineral stocks,nutrient cycling .
Abstract . The ecosystem associated to the Mediterranean seagrass Posidonia oceanica shows a cleardistinction in two subcompartments regarding turnover time : aboveground and belowground .Aboveground parts (leaves) are highly dynamic, and most of the leaf material is decomposed orexported in less than one year, representing a net loss of nutrients . In contrast, belowgroundbiomass (roots and rhizomes) has a turnover time of the order of centuries, with a consequentaccumulation of organic matter in the sediment . The accumulation rates for the single elements rankin the order C > N > P . This ecosystem may be considered as a sink for biogenic elements .
Problem
The functioning of an ecosystem implies the existence of a certain amount ofdead organic matter resulting from the budget in space and time betweenproduction and decay rates . The standing dead organic matter (necromass) hasbeen recognized as a key compartment in terrestrial ecosystems, as regards bothtrophic aspects and nutrient cycling, and there is an extensive literature dealingwith these topics (OLSON, 1963 ; HUNT, 1977; SWIFT et al., 1979; MELILLO et al.,1982; SMITH, 1982; MELILLO et al ., 1989) . In the aquatic environment, researchhas mostly concentrated on the kinetics of the decomposition processes (BROCKet al ., 1985 ; GODSHALK & WETZEL, 1978 ; PELLIKAAN, 1984; VALIELA et al., 1982 ;for a review, see HARRISON, 1989) and, to some extent, on topics related to therole of detritus as a food source (ODUM & DE LA CRUZ, 1967 ; for a review, seeLEVINTON et al., 1984) . Nevertheless, attempts to include these aspects and thenecromass compartment in a general ecosystem model are scarce (PEL-
U. S . Copyright Clearance Center Code Statement : 0173-9565/92/1301-0069$02 .50/0

70
LIKAAN & NIENHUIS, 1988) . This may be due to the fact that in macrophyte-dominated systems, only a minor part of the produced material is recycled insitu ; the bulk is exported, especially in marine communities (FENCHEL, 1977),resulting in a spatial uncoupling between production and decay .
Consequently, within the framework of a major cooperative project on theMediterranean ecosystem dominated by the seagrass Posidonia oceanica (L .)DELILE, we have attempted to gain some insight into the necromass compart-ment. The first stage of our investigation was focused on two basic objectives :(i) To estimate the amount of necromass, especially leaf litter, associated withthe Posidonia bed, and its spatio-temporal variability .(ii) To estimate the decay rates of the plant material .
These questions are of major importance for understanding the functioning ofthe Posidonia ecosystem, since they relate to basic ecological processes, i . e .,nutrient cycling, export of plant material, and detritivore food chains .
Material and Methods
1 . Estimation of litter stocks
Samples were taken using a suction device, as described previously in PERGENT-MARTINI et al . (1991),inside a 35 X 35 cm quadrat. A bathymetric transect in Lacco Ameno (see COLANTONI et al., 1982,and MAZZELLA et al., 1989 for a general description of the site) was sampled every three months(July 88, October 88, January 89, April 89, and July 89) at 1, 2, 5, 10, 20, and 30m depth . At eachdepth and on each date, three samples (replicates) - randomly selected over an area of ca .100-200 ml - were taken . The samples included both the dead material lying on the sediment and thesediment itself to a depth of 2-4 cm . The number of Posidonia shoots inside the quadrat wasrecorded .
The samples were washed immediately with running seawater, and both the sediment and all thecoarse material coming from the belowground parts of the plant (mainly old leaf bases andrhizomes) were discarded . The remainder was sorted into two fractions :(i) Coarse leaf litter (> 0 .8 cm) and (ii) Fine litter (0 .1-0 .8 cm) .
The coarse fraction is made up of blade material in a more or less decomposed stage, while thefine litter is more heterogeneous, with variable amounts of tiny leaf debris and fibers resulting fromleaf base decay .
Both fractions were weighed separately after drying at 90'C until constant weight .
2. Decay experiments
Classic litter bag experiments were performed at 5 and 20 m depth (see the early works of FALCONERet al ., 1933; BOCOCK et al., 1960; for the aquatic environment see, for example, GODSHALK &WETZEL, 1978; JOSSELYN et al., 1986 ; BROCK et al ., 1985) . Material for the incubation was collectedfrom living plants ; the oldest, senescent leaf of each shoot was chosen in order to simulate a naturalleaf abscission . A known amount (30 g of fresh weight) of this material was enclosed in a number ofnet bags (mesh size : 0 .1 cm) . The bags were then placed in the meadow under the foliar canopy,attached close to the sediment, and collected at increasing time intervals (1, 2, 3, 4 weeks ; 2, 3, 6months) Two series of bags were placed at both 5 m and 20 m in July 1988 . Unfortunately, the bagsat 5 m were lost 1 month later . Two more series were them initiated in October 1989 . At eachsampling time, three bags were collected at random and taken to the laboratory . There, the sampleswere washed, the fauna was eliminated, and the foliar debris sorted into two fractions (coarse andfine), then dessicated, and weighed in the same way as for the litter .
ROMERO, PERGENT, PERGENT-MARTINI, MATEO & REGNIER
72
2. Decay experiments
A weight loss with time was evident in the incubated material. This loss followeda simple negative exponential model, with a high goodness of fit . Althoughsimple exponential models have often been criticised, other models that havebeen proposed to fit decomposition data (GODSHALK & WETZEL, 1978 ; WIEDER&LANG, 1982) did not, in our case, increase the goodness of fit or reveal anyspecial biological significance .
Decay rates (computed as k, the exponent of the model) appear in Table 1 .Weight loss was more rapid at 5 m than at 20 m for equivalent periods . Aseasonal trend also appeared, with higher decomposition rates in July/Januarythan in October/April . The remaining material inside the bags consisted mainlyof coarse fraction; the fine fraction accounted for no more than 10 % of totalweight in all cases .
ROMERO, PERGENT, PERGENT-MARTINI, MATEO & REGNIER
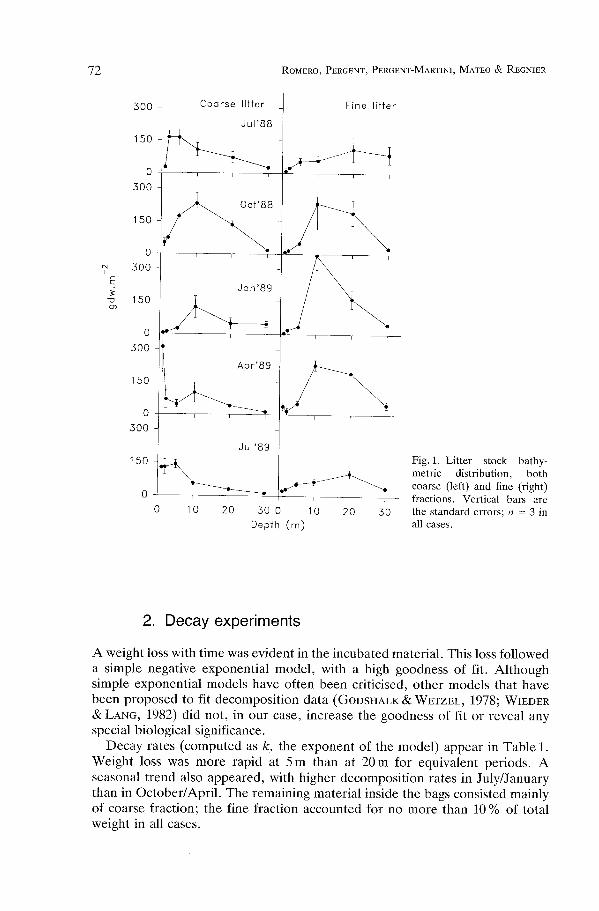
Fig . 1 . Litter stock bathy-metric distribution, bothcoarse (left) and fine (right)fractions . Vertical bars arethe standard errors ; n = 3 inall cases .

The detritic compartment in a Posidonia oceanica meadow
71
3 . Belowground parts
Samples of rhizomes + roots were taken using a diver-held corer (as described in PIRG, 1983) at 5and 20m depth, penetrating around 20 cm in the sediment . Three replicates were taken per depth .The samples were washed carefully and sorted into the following fractions :(i) living rhizomes, (ii) dead rhizomes, (iii) living roots, (iv) dead roots, (v) fine undifferentiatedfraction (0 .8-0 .1 cm) - and then weighed as described above . Fractions (i) and (ii) included theattached leaf bases .
The criteria used in fractioning the belowground plant parts are described in FRANCOUR (1990) .An approach to the decay dynamics of some belowground parts was carried out using lepido-
chronology (see PERGENT et al., 1989). When Posidonia oceanica leaves die, only the blade fallsaway; the leaf sheathing base remains attached to the rhizome after leaf abscission, and is then oftenincorrectly termed a scale . As BouDouREsQuE et al. (1983) and PERGENT et al . (1983) demonstrated,the occurrence of annual fluctuations in leaf base thickness makes it possible to date long sections oforthotropic rhizomes . This constitutes a valuable tool for establishing the exact year of formation ofa given section of the rhizome or a given leaf base . Consequently, the decay rate of the associatedtissues can easily be inferred .
4 . Chemical elementary composition
In order to assess the turnover of the various biogenic elements in the different necromasssubcompartments, the elementary composition (carbon, nitrogen, and phosphorus) was determinedfor subsamples - dried at a maximum temperature of 70'C - of the following items :(i) litter from 5 and 20m depth (both fine and coarse), (ii) material from the decay experiments(5 and 20m), (iii) belowground parts, (iv) lepidochronological samples .
The subsamples were ground to a fine powder and processed through a Carlo-Erba autoanalyzerto determine C and N content . P content was analyzed using ICP (Induction Coupling Plasma) aftera wet acid digestion following JACKSON (1970) and DELGADO (1986), modified to a micro-wavedigestor by MATEO & SABATÉ (1989) .
Results
1 . Litter stocks
Litter stock bathymetric distributions (both coarse and fine) are summarized inFig . 1 . Coarse litter followed a basic pattern, showing an intermediate depthmaximum, except in April, and, in terms of seasonal variation, a summer-autumn maximum and a winter minimum .
The fine fraction is heterogeneous and includes small leaf fragments andfibers resulting from leaf base decay . The relative proportion of fibers increasedwith depth . This heterogeneity results in less clear spatio-temporal patterns . Ingeneral, maximum stocks were found at greater depths than for the coarsefraction, and seasonal maxima were shifted towards winter. The relative propor-tions of coarse/fine in the total litter stock clearly decreased with depth .
The few literature data on seagrass litter stocks seem to confirm the quantita-tive importance of this compartment (GALLAGHER et al., 1984 ; PEL-LIKAAN & NIENHUIS, 1988) .

The detritic compartment in a Posidonia oceanica meadow
73
Table 1 . Decay rates in the different litter bag experiments . The dates of start and end of theexperiment are given, as well as the average temperature during the incubation period and the initialN-concentration in the enclosed plant material . Experiments n and n' represent decay ratescomputed over different time intervals using the same data set in order to allow comparisons .
3. Belowground parts: roots and rhizomes
Data for biomass and necromass of the belowground compartment are shown inTable 2. Variability was very high due to the heterogeneity of the meadow, butin all samples large stocks of dead material were registered . No living parts werefound below 20 cm under the sediment surface . The fine, undifferentiatedfraction showed high values and included the fine litter sampled with the suctiondevice plus the fine fraction buried in the sediment . Values were higher at 5 mthan at 20 m due to the higher density of the meadow near the surface .
Table 2 . Standing stocks of both bio- and necromass (g dw • M-2) at 5 and 20 m depth . Standarderrors are given and, in all cases, n = 3 .
4. Belowground parts : lepidochronological data
Sheath decay is shown in Fig . 2 . Since there is a certain variability in the numberof leaves produced per year (see PERGENT et al ., 1989), the data has beennormalized for an ideal average sheath . From these data, and assuming again a
74 ROMERO, PERGENT, PERGENT-MARTINI, MATEO & REGNIER
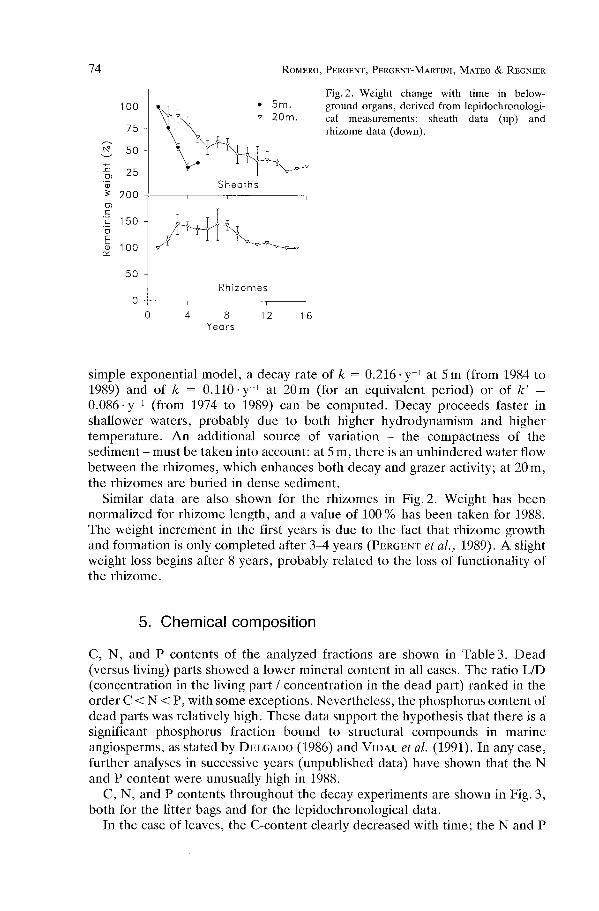
Fig .2 . Weight change with time in below-ground organs, derived from lepidochronologi-cal measurements : sheath data (up) andrhizome data (down) .
simple exponential model, a decay rate of k = 0 .216 • y- I at 5 m (from 1984 to1989) and of k = 0.110-y -1 at 20m (for an equivalent period) or of k' =0.086-y -1 (from 1974 to 1989) can be computed. Decay proceeds faster inshallower waters, probably due to both higher hydrodynamism and highertemperature. An additional source of variation - the compactness of thesediment - must be taken into account: at 5 m, there is an unhindered water flowbetween the rhizomes, which enhances both decay and grazer activity ; at 20 m,the rhizomes are buried in dense sediment .
Similar data are also shown for the rhizomes in Fig . 2 . Weight has beennormalized for rhizome length, and a value of 100 % has been taken for 1988 .The weight increment in the first years is due to the fact that rhizome growthand formation is only completed after 3-4 years (PERGENT et al., 1989). A slightweight loss begins after 8 years, probably related to the loss of functionality ofthe rhizome .
5 . Chemical composition
C, N, and P contents of the analyzed fractions are shown in Table 3 . Dead(versus living) parts showed a lower mineral content in all cases . The ratio L/D(concentration in the living part / concentration in the dead part) ranked in theorder C < N < P, with some exceptions . Nevertheless, the phosphorus content ofdead parts was relatively high . These data support the hypothesis that there is asignificant phosphorus fraction bound to structural compounds in marineangiosperms, as stated by DELGADO (1986) and VIDAL et al. (1991) . In any case,further analyses in successive years (unpublished data) have shown that the Nand P content were unusually high in 1988 .
C, N, and P contents throughout the decay experiments are shown in Fig . 3,both for the litter bags and for the lepidochronological data .
In the case of leaves, the C-content clearly decreased with time ; the N and P

The detritic compartment in a Posidonia oceanica meadow
Table 3 . Carbon, nitrogen, and phosphorus content in differents parts (living and dead) of theplant. Values are expressed in % of the element relative to the dry weight . The SEM is within therange 5-20 % of the mean, with n between 3 and 10 . The data are from material collected in July at20 m .
Fig . 3 . C, N, and P content indifferent parts of the plantduring the decompositionprocess . Left : data from litterbag experiments (note thatthe experiment at 20 m beganin July, that at 5 m began inOctober) . Right : lepido-chronological data . Verticalbars are the standard errors .
75
content decreased at 20 m and remained fairly constant at 5 m . For the below-ground parts, there was also a slight carbon loss with time . N and P contentsdecrease sharply in the rhizomes during the first three years, while growth is stillproceeding, but remain constant in leaf sheaths .
Discussion
1 . Litter stocks: models
Bathymetric distribution of litter depends upon the budget of inputs (i. e ., fromleaf fall) and outputs (i . e ., export and decay) . Assuming that decay rates are

76
constant with depth (or that its variation is well below the bathymetric variationdue to the other two factors), the following model can account for the observedlitter distribution :
L =10 • (exp [- p • z])(1 - exp [- h • z])
[equ. 1]
where to is a scale constant (i . e ., the theoretical litter input at 0 meters), p is therate of decrease of leaf (and subsequently, litter) production with depth, and h isthe attenuation rate of the hydrodynamism . The equation was fitted to our data(only coarse litter) using an iterative method (LEATHERBARROW, 1982) . Theestimation of the parameters of equ . 1 are summarized in Table 4 . The fit of the
Table 4. Estimated parameters for the model of litter stock bathymetric distribution (for explana-tion, see text) .
ROMERO, PERGENT, PERGENT-MARTINI, MATEO & REGNIER
model to our data is good (the predicted values are within 1 SE interval aroundthe mean in 80 % of cases, and of 2 SE in all cases) . Although the model issimple, the parameters seem to be quite realistic . The p rate gives a roughestimate of how Posidonia production decreases with depth, and its value isclose to the d rate from the following equation :
Dz = Do. exp (- d • z)
[equ. 2]
where Dz is shoot density (shoots . m-2) at depth z . Fitting this exponentialmodel to our data on shoot density (Fig . 4) gives an estimation of d near0.06 • m-I . The h rate is of the same order as the proposed hydrodynamiccoefficient for Lacco Ameno in various previous works (GAMBI, 1986 ; GAMBI et
al., 1989) . The ability of the model to predict litter stocks led us to conclude thatbathymetric distribution of litter is in effect controlled by both production (beddensity) and export (hydrodynamism) .
Fig . 4 . Variation in meadow density withdepth . Vertical bars are the standard errors ;n = 12 in all cases .

The detritic compartment in a Posidonia oceanica meadow
77
In the absence of litter export, bathymetric variation in litter stocks is onlydue to variation of inputs from leaf fall . Leaf fall data can be obtained fromBUIA et al . (1992), assuming that the weight of fallen leaves is the differencebetween primary production (estimated by the ZIEMAN method) and biomassincrease . From this, and taking into account the decay rates estimated, a litteraccumulation value can be computed under the hypothesis of zero export, that is
L'(i) = F(i) • exp (- k . t/2) + L(i-t) • exp (- k . t)
[equ . 3]
where L'(i) is the predicted standing litter at time i, F(i) is the weight of leafmaterial fallen between times i and i-t, t is the time interval between consecu-tive samplings, k is the decay rate and L(i-t) is the standing litter (observed) attime i-t. Consequently, export per unit surface between i and i-t is
E(i) = L'(i) - L(i)
[equ. 4]
This model was run using monthly estimates of leaf fall and our data of standinglitter; the results are shown in Table 5 .
Table 5 . Export rate at different depths, expressed as a percentage of the annual foliar (blades only)primary production .
Although this simple model overlooks some basic aspects of thesephenomena (decay rate variations on both a seasonal and a bathymetric basis,different structures of the meadow, effects of the fauna, discontinuity of exportevents), the values in Table 5 are probably a good indication of the order ofmagnitude of the organic material produced in the meadow and exported, andthus recycled elsewhere. Most of this material (70 %) is exported at shallowdepths (1-2m), while the export rate is fairly constant from 5 to 30m . For thewhole Posidonia oceanica meadow of Lacco Ameno, we can consider that 50of the leaf material produced is exported ; this is not far from Orr's (1980)estimation . Compared to data for other plants, this value is relatively high :1-30 % in Zostera marina (BACH et al ., 1986), 10-50 % in Spartina (DAY et al .,1973 ; VALIELA et al., 1978 ; HOPKINSON & HOFFMAN, 1984) ; 12 % in Posidoniaaustralis (KIRKMAN & REID, 1979) ; 1 % in Thalassia testudinum (ZIEMAN et al .,1979) ; 90 % in Syringodium filiforme (ZIEMAN et al ., 1979) ; 20 % in mangroves(LUGO et al., 1976). Since Posidonia oceanica decay is not lower than in otherspecies, this high export rate may be attributed to the geomorphology of thezone (open shore) .
In general, the export process has been emphasized from the point of view ofits role as a nutritional input for adjacent ecosystems ; yet it also represents a netloss of nutrients for the seagrass ecosystem (see the last section of this paper) .

78
2. Decay experiments
Our leaf material decomposition rates are within the range of published data onaquatic angiosperms (HARRISON, 1989), although comparisons are difficultbecause of different methods, experiment durations, pretreatments, etc . Thedata in Table 1, although not conclusive, indicate one main source of variationassociated with depth ; this can be interpreted as hydrodynamism, since decay isgreater at 5 than at 20 m (for the same time intervals and the same season) . Theobserved seasonal trend can be attributed both to temperature and to thequality of the decaying material (N content of July experiments : 1 .33 % ; Ncontent of October experiments : 0 .68 %), although it should be recalled that Ncontent is not a universal estimator of decay rates . In effect, while the PosidoniaN content is low compared to other seagrasses (HARRISON, 1989 ; DUARTE, 1990),the decay rates are intermediate .
The dynamics of N and P release during decay can be summarized as follows(see Fig . 3 and Table 3) . A first loss takes place during leaf senescence (11 % N,30 % P) . How these elements are lost before leaf absicission remains unknown ;both an early leaching or a retranslocation process can occur (see FRESI &SAGGIOMO, 1980; PIRG, 1985 ; PIRC & WOLLENWEBER, 1988) . Further losses occurduring decomposition (37 % N, 35 % P relative to the concentration of the livingmaterial), until a plateau is reached (see Fig . 3 ; material from July at 20 m) thatcan be interpreted as the refractory fraction (52 % N, 35 % P) . However, whenthe initial material has a low nutrient content (see Fig . 3 ; material from October,5 m), no further losses occur during decomposition, but the plateau is reachedwith very similar values .
No nitrogen accumulation during decay was recorded . This seems to be ageneral feature of seagrasses (HARRISON, 1989), with some exceptions (JOSSELYN& MATHIESON, 1980 ; RICE & TENORE, 1981) . This is a key factor in establishingwhether or not leaf litter acts as a sink for nitrogen (as for mangroves : RICE,1982). Although data on P release during decay are much scarcer, the existenceof a refractory fraction seems to be a universal feature of vascular plants . Thisrefractory fraction can be estimated as the P content in the tissues after a certaintime of decay expressed as a percentage of the P content of the fresh material .Data in the literature indicate that this refractory fraction can represent between15 and 90 % of the initial P concentration, depending on the species and on thedecay conditions (SWIFT et al., 1979; KLUMP & VAN DER VALK, 1984 ; PELLIKAAN,1984; VAN DER VALK & ATTIWILL, 1984 ; FORÉS et al., 1988; MENÉNDEZ et al.,1989) . The existence of such a fraction allows a net export capacity, either to thesediment or to adjacent ecosystems .
3 . Mineral stocks
The data presented in this paper allow mineral stocks of different compartmentsof the system to be estimated; for the sake of brevity, we only present data from20m (Table 6) . The ratio dead/living (in terms of total weight) is around 3 .1 .This is quite high compared with terrestrial ecosystems, being of the same orderof magnitude as for boreal forests (OLSON, 1963 ; RODIN & BAZILEVICH, 1967) . It
ROMERO, PERGENT, PERGENT-MARTINI, MATEO & REGNIER
The detritic compartment in a Posidonia oceanica meadow
79
Table 6 . Biomass and mineral stocks in the different parts of the plant . Dry weight in gDW • M-2 . C,N, P: g of element . M-2 . The data are from the 20 m station . For leaves and litter, maximum yearlyvalues are reported .
would be even higher with sampling to a greater sediment depth . This ratiodrops if we consider single elements C (2 .6), N (1 .3), and P (around 1 .4) . Thesevalues are obviously related to turnover rates of the different elements .
Using PB data from OTT (1980) and ROMERO (1985), the turnover time of thedifferent necromass subcompartments (that is, the standing necromass of agiven subcompartment divided by its yearly production) for each of the threeelements can be estimated (Table7) . The turnover time is low for the above-ground parts, on the order of months . For the belowground part, the turnovertime is much longer, from years to centuries . As shown in Table 7, both N and Precycle faster than C .
Even our rough estimations at this preliminary stage of work demonstratethat the Posidonia ecosystem acts as a sink for the various biogenic elements . Ineffect, amounts of the order of 25 g C . M-2 . y-1 (and associated elements : seeTable 8) are diverted to a very low turnover subcompartment (rhizomes plusroots). In contrast, the leaf subcompartment is much more dynamic, anddecomposition and export lead to low residence time of old leaf material withinthe meadow . Since both immobilization and export represent net losses ofbiogenic elements for the system (summarized in Table 8), we conclude that atleast 35 % of the plant production is new production (in the sense of DUGDALE &GOERING, 1967), and will need an external nutrient supply to proceed .
Table 7 . Turnover rates (year - ') of the different parts of the plant for the different elements (C, N,and P) . Turnover rate for a given subcompartment is calculated as the annual production of thesubcompartment divided by its maximum annual biomass . Data from the 20m station .

80
Table 8. Summary of nutrient losses for the Posidonia ecosystem (g of element .m2 . y') . Therequirements have been estimated using data of primary production and data in Tables 3 and 6 .
Summary
This paper represents a preliminary attempt to investigate the detritic compart-ment of a Mediterranean seagrass ecosystem (Posidonia oceanica) . The amountof dead organic matter (necromass) within the system has been evaluated,taking into account both leaf litter and dead rhizomes and roots, as well as themineral stocks (C, N, and P) it represents . Decay rates of leaves and rhizomeshave been estimated, and turnover time for each element within each subsystemderived. The leaf compartment is highly dynamic, and within one year all thematerial produced is either decomposed or exported ; yearly export can beestimated as 50 % of the produced leaf material for the whole meadow .Turnover time of the belowground biomass is higher, of the order of centuries .The belowground parts act as a sink for biogenic elements, annually divertingfrom the general nutrient cycling up to 25 g C, 0 .4 g N, and 0 .02 g P . M-2 . Takinginto account both export and burial, we conclude that at least 35 % of the yearlyplant production must be supported by external nutrient inputs, a value that alsorepresents new production .
Acknowledgements
This work was supported by EEC grant EV4V-0139-B . Field work was carried out at Ischia, wherelaboratory and diving facilities were kindly provided by Dr . LUCIA MAZZELLA and her staff at theLaboratorio di Ecologia del Benthos de la Stazione Zoologica di Napoli . Thanks are also due toP. BULTEEL and P . COULON for their help . The Spectroscopy Service of the University of Barcelona,from where ELIONOR PELFORT deserves a special mention, gave technical support for the phosphorusanalysis .
References
BACH, S . D ., G . W. THAYER & M. W. LACROIX, 1986 : Export of detritus from eelgrass (Zosteramarina) beds near Beaufort, North Carolina, USA. Mar . Biol . Prog . Ser ., 28 : 265-278 .
BOCOCK, K . L ., O . J . GILBERT, C. K. CAPSTICK, D . C . TURNER, J . S . WARD & M . J . WOODMAN, 1960 :Changes in leaf litter when placed on the surface of soils with contrasting humus types . J . SoilSci., 11: 1-9 .
BOUDOURESQUE, C . F ., A. CROUZET & G . PERGENT, 1983 : Un nouvel outil au service de l'étude desherbiers à Posidonia oceanica : la l épidochronologie . Rapp . P . V. Réun. Comm. Int . Explor .Sci . Médit ., Monaco, 28 (3) : 111-112 .
ROMERO, PERGENT, PERGENT-MARTINI, MATEO & REGNIER

The detritic compartment in a Posidonia oceanica meadow 81
BROCK, T . C . M ., J . J . BooN & B . G . P . PAFFEN, 1985 : The effects of the season and of waterchemistry on the decomposition of Nymphaea alba L . ; weight loss and pyrolysis massspectrometry of the particulate matter . Aquat . Bot ., 22: 197-229 .
BETA, M . C ., V . Zupo & L. MAZZELLA, 1992 : Primary production and growth dynamics of Posidoniaoceanica . P .S .Z.N .I : Marine Ecology, 13 (1) : 1-15 .
COLANTONI, P ., P . GALLIGNANI, E . FRESI & F . CINELLI, 1982 : Patterns of Posidonia oceanica (L .)DELILE bed around the island of Ischia (Gulf of Naples) and in adjacent waters . P .S .Z.N .I :Marine Ecology, 4 (2) : 151-159 .
DAY, J . A ., W . G . SMITH, P . R. WAGNER & W. C . STOWE, 1973 : Community structure and carbonbudget of a salt-marsh and shallow bay estuarine system in Louisiana . Centre for WetlandResources, Louisiana State University ; 411 pp .
DELGADO, 0., 1986 : Contenido de f6sforo de los tejidos de faner6gamas marinas del Mediterràneooccidental y su relaci6n con la dinàmica de cada especie. Oecol. Aquat ., 8: 139-151 .
DUARTE, C., 1990 : Seagrass nutrient content . Mar . Ecol . Prog . Ser ., 67: 201-207 .DUODALE, R . C . & J . J . GOERING, 1967 : Uptake of new and regenerated forms of nitrogen in
primary production . Limnol . Oceanogr., 12 : 196-206 .FALCONER, G . J ., J . W. WRIGHT & H . W. BEALL, 1933 : The decomposition of certain types of fresh
litter under field conditions . Am . J . Bot ., 20 : 196-203 .FENCHEL, T ., 1977 : Aspects of decomposition of seagrasses . In : C . P. MCROY & C . HELFFERICH,
(Eds .), Seagrasses ecosystems : a scientific perspective . Dekker, New York : 132-145 .FORÉS, E., M. MENÉNDEZ & A . CoMIN, 1988 : Rice straw decomposition in rice-field soil . Plant Soil,
109 : 145-146 .FRANCOUR, P., 1990 : Dynamique de l'écosystème à Posidonia oceanica dans le Parc National de
Port-Cros . Analyse des compartiments matte, litière, fauna vagile, échinodermes et poissons .Thèse Troisième Cycle. Univ . Aix-Marseille 11 ; 278 pp .
FRESI, E . & V. SAGGIOMO, 1980 : Phosphorus uptake and transfer in Posidonia oceanica (L.) DELILE .Rapp. P .V. Réun. Comm . Int . Explor . Sci . Médit ., Monaco, 27 (2) : 178-188 .
GALLAGHER, J . L., H . V. KIRBY & K. V . SKIRVIN, 1984 : Detritus processing and mineral cycling inseagrass (Zostera) litter in an Oregon salt marsh . Aquat . Bot ., 20 : 97-108 .
GAMBI, M . C ., 1986 : The filed method of `plaster balls' for water movement estimates applied toPosidonia oceanica beds. Rapp . P . V. Réun. Comm . Int . Explor . Sci . Médit ., Monaco, 30 (2) :1-2 .M. C, BUIA, E. CASOLA & M . SCARDI, 1989: Estimates of water movement in Posidoniaoceanica beds : a first approach . II . International Workshop on Posidonia oceanica beds, 2 :175-179 .
GODSHALK, G . L . & R . G . WETZEL, 1978 : Decomposition of aquatic angiosperms . II . Particulatecomponents, Aquat . Bot ., 5 : 301-327 .
HARRISON, P . G ., 1989 : Detrial processing in seagrass systems : a review of factors affecting decayrates, remineralization and detritivory . Aquat . Bot ., 23 : 263-288 .
HOPKINSON, C . S . & F . A. HOFFMAN, 1984 : The estuary extended - a recipient system study ofestuarine outwelling in Georgia . In : V . S . KENNEDY (Ed .), The estuary as a filter . AcademicPress, New York : 313-330 .
HUNT, H . W ., 1977 : A simulation model for decomposition in grasslands . Ecology, 58 : 469-484 .JACKSON, M . L ., 1970 : Anâlisis qufmico del suelo . Omega, Barcelona ; 662 pp .JOSSELYN, M . N . & A . C . MATHIESON, 1980 : Seasonal influx and decomposition of autochthonous
macrophyte litter in a north temperate estuary . Hydrobiologia, 71: 197-208 .M.FONSECA, T. NIESEN & R . LARSON, 1986 : Biomass, production and decomposition of adeepwater seagrass, Halophila decipiens OSTENFELD. Aquat . Bot ., 25: 47-61 .
KIRKK4N, H. & D . D. REID, 1979 : A study of the role of a seagrass Posidonia australis in the carbonbudget of an estuary . Aquat . Bot ., 7 (2) ; 173-183 .
KLUMPP, D. W. & VAN DER VALK, 1984 : Nutritional quality of seagrasses (Posidonia australis andHeterozostera tasmanica) : comparison between species and stages of decomposition . Mar . Biol .Letters, 5 : 67-83 .
LEATHERBARROW, E . J ., 1982 : Enzfitter : User's manual . Elsevier, Amsterdam ; 91 pp .LEVINTON, J . S ., T . S . BIANCHI & S . STEWART, 1984 : What is the role of particulate organic matter in
benthic invertebrate nutrition? Bull . Mar. Sci ., 35: 270-282.LUGO, A . E ., M. SELL & S . C . SNEDAKER, 1976 : Mangrove ecosystem analysis . In : B . C . PATTERN
(Ed .), Systems analysis and simulation in ecology . Academic Press, New York ; 4: 114-145 .

82 ROMERO, PERGENT, PERGENT-MARTINI, MATEO & REGNIER
MATEO, M . A . & S . SABATÉ, 1989 : Digestion de material vegetal por via hdmeda mediante métodosclâsicos modificados y microondas como fuente de energfa . Documentos de trabajo . Dpto . deEcologfa, Facultad de Biologfa, Universidad de Barcelona ; 21 pp .
MAZZELLA, L ., M . B . SCIPIONE & M . C. BUIA, 1989 : Spatio-temporal distribution of algal and animalcommunities in a Posidonia oceanica meadow . P .S .Z.N .I : Marine Ecology, 10 (2) : 107-129 .
MELILLO, J . M., J . D . ABER & J . F . MURATORE, 1982: Nitrogen and lignin control of hardwood leaflitter decomposition dynamics . Ecology, 63: 621-626 .
--, J . D . ABER, A . E . LINKINS, A . RICCA, B . FRY & K . J . NADELHOFFER, 1989 : Carbon and nitrogendynamics along the decay continuum : plant litter to soil organic matter . Plant Soil, 115 :189-198 .
MENÉNDEZ, M ., E . FORÉS & A. COMIN, 1989 : Ruppia cirrhosa - Decomposition in a coastaltemperate lagoon as affected by macroinvertebrates . Arch . Hydrobiol., 117 : 39-48 .
ODUM, E . P. & A . A. DE LA CRUZ, 1967 : Particulate organic detritus in a Georgia salt marshestuarine ecosystem . In : G . H . LAUFF (Ed .), Estuaries . American Association for the Advance-ment of Science, 83.
OLSON, J . S ., 1963 : Energy storage and the balance of producers and decomposers in ecologicalsystems . Ecology, 44 (2) : 322-331 .
OTT, J . A., 1980 : Growth and production in Posidonia oceanica (L .) DELILE. P .S .Z.N .I : MarineEcology, 1 : 47-64 .
PELLIKAAN, J . C ., 1984 : Laboratory experiments on eelgrass (Zostera marina L.) decomposition .Neth . J . Sea Res ., 18 : 360-383 .& P. H . NIENHUIS, 1988 : Nutrient uptake and release during growth and decomposition ofeelgrass, Zostera marina L., and its effects on the nutrient dynamics of lake Grevelingen .Aquat . Bot ., 30: 189-214 .
PERGENT, G ., C . F . BOUNDOURESQUE & A . CROUZET, 1983 : Variations cycliques dans les écailles desrhizomes orthotropes de Posidonia oceanica. Trav. Sci . Parc . Nat. Port-Cros, 9: 107-148 .C. F. BOUDOURESQUE, A . CROUZET & A. MEINESZ, 1989 : Cyclic changes along Posidoniaoceanica rhizomes (lepidochronology) : present state and perspectives . P .S .Z.N .I : MarineEcology, 10 : 221-230 .
PERGENT-MARTINI, C., G. PERGENT & J . ROMERO, 1991 : The litter in the Posidonia oceanica eco-system : preliminary results . In : B . F . KEEGAN (Ed .), Space and time series data analysis incoastal benthic ecology . Commission of the European Communities : 541-547 .
PIRG, H ., 1983 : Belowground biomass of Posidonia oceanica (L .) DELILE and its importance to thegrowth dynamics . Proceedings International Symposium on Aquatic Macrophytes. Nijmegen,18-23 Sept ., 1983 : 77-181 .1985 : Growth dynamics in Posidonia oceanica (L .) DELILE . I . Seasonal changes of solublecarbohydrates, starch, free amino acids, nitrogen and organic anions in different parts of theplant . P .S .Z.N .I : Marine Ecology, 6 (2) : 141-165 .& B. WOLLENWEBER, 1988: Seasonal changes in nitrogen, free amino acids, and C/N ratio inMediterranean seagrasses . P .S.Z.N .I : Marine Ecology, 9 (2) : 167-179 .
RICE, D . L ., 1982 : The detritus nitrogen problem : new observations and perspectives from organicgeochemistry . Mar . Ecol. Prog . Ser ., 9 : 153-162 .& K. R . TENDRE, 1981 : Dynamics of carbon and nitrogen during the decomposition of detritusderived from estuarine macrophytes . Estuarine Coastal Shelf Sci ., 13 : 681-690 .
RODIN, L. E. & W . I . BAZ1LEW1CH, 1967: Production and mineral cycling in terrestrial vegetation .Oliver & Boyd, Edinburgh ; 288 pp .
ROMERO, J., 1985 : Estudio ecol6gico de las faner6gamas marinas de la costa catalana : producci6nprimaria de Posidonia oceanica (L .) DELILE en ]as islas Medes . Tesis doctoral, Facultad deBiologfa, Universidad de Barcelona ; 266 pp .
SM1TH, O . L ., 1982 : Soil microbiology : a model of decomposition and nutrient cycling . C .R.C . Press,Boca Raton, Florida ; 273 pp .
SWIFT, M . J ., O . W. HEAL & J . M. ANDERSON, 1979 : Decomposition in terrestrial ecosystems .Blackwell Scientific Publications, Oxford ; 372 pp .
VALIELA, I ., J . M . TEAL, S . VOLKMAN, D . SCHAFER & E . J . CARPENTER, 1978 : Nutrient and particu-late fluxes in a salt-marsh ecosystem : tidal exchanges and inputs by precipitation and ground-water . Limnol . Oceanogr ., 23 : 798-812 .B. HOWES, R . HOWARTH, A . GIBLIN, K . FOREMAN, J . M . TEAL, J . E . HOBBLE, 1982 : Regulationof primary production and decomposition in a salt marsh ecosystem . In : B . GoPAL, R . E .

The detritic compartment in a Posidonia oceanica meadow 83
TURNER, R . G . WETZEL & D. F . WHIGHAM (Eds .), Wetlands, ecology and management . Nat .Inst . of Ecology, India : 151-168 .
VAN DER VALK, A. G. & P . M. ATTIWILL, 1984: Decomposition of leaf and root litter of Aviceniamarina at Westernport Bay, Victoria, Australia . Aquat . Bot ., 18: 205-221 .
VIDAL, M., O . DELGADO & J. ROMERO, 1991 : El fôsforo en praderas de fanerôgamas : datospreliminares sobre su concentraci6n en tejidos vegetales y en el sedimento . Proc . IberianSymposium on Marine Benthos, 5 (1) : 463-472 .
W1EDER, R. K. & G . E . LANG, 1982 : A critique of the analytical methods used in examiningdecomposition data obtained from litter bags . Ecology, 63 (6) : 1636-1642 .
ZIEMAN, J . C ., G. W. THAYER, M . B . ROBLEE & R . T . ZIEMAN, 1979 : Production and export ofseagrasses from a tropical bay . In : R . J . LIVINGSTON (Ed .), Ecological Processes in Coastal andMarine Systems . Plenum Press, New York; 21-33 .
Copyright © 2022 FDOKUMEN




















