
Impact de l'acidification des océans sur les épibiontes des feuilles de posidonies (Posidonia...
36
Master d’Océanographie – Biologie Ecologie Marine BEM – Delille Jeremy Institut Pythéas – Université d’Aix-Marseille Laboratoire d’Océanographie de Villefranche sur mer Stage réalisé de janvier à juin 2014 Tuteurs : Frédéric Gazeau Victoria Diaz-Castañeda Erin Cox
Transcript of Impact de l'acidification des océans sur les épibiontes des feuilles de posidonies (Posidonia...

Master d’Océanographie – Biologie Ecologie Marine BEM –
Delille Jeremy
Institut Pythéas – Université d’Aix-Marseille
Laboratoire d’Océanographie de Villefranche sur mer
Stage réalisé de janvier à juin 2014
Tuteurs :
Frédéric Gazeau
Victoria Diaz-Castañeda
Erin Cox

CONTENU
Introduction ................................................................................................................................ 4
Acidification de l’océan global .............................................................................................. 4
Photosynthèse ......................................................................................................................... 5
Calcification ........................................................................................................................... 5
Modèle d’étude ....................................................................................................................... 6
Objectifs de l’étude ................................................................................................................ 7
Hypothèses ............................................................................................................................. 8
Matériel et méthodes .................................................................................................................. 9
Lieu d’étude et matériel biologique........................................................................................ 9
Traitements post-prélèvement .............................................................................................. 10
Paramètres de l’expérience ................................................................................................... 10
Mesures analytiques et protocole ......................................................................................... 12
Mesures des taux de calcification en incubation .................................................................. 14
Mesures de fin d’expérience................................................................................................. 15
Statistiques ........................................................................................................................... 19
Résultats ................................................................................................................................... 21
Mesures des réservoirs ......................................................................................................... 21
Taux de calcification ............................................................................................................ 22
Masse calcifiée ..................................................................................................................... 23
Recouvrement ....................................................................................................................... 24
Discussion ................................................................................................................................ 28
Conclusion ................................................................................................................................ 32
Bibliographie ............................................................................................................................ 33

Je tiens à remercier toutes les personnes dont la présence a été indispensable au bon
déroulement de ce stage.
Merci à Frédéric Gazeau de m’avoir offert son écoute avec le sourire, en prenant le temps
pour lire, orienter et diriger ce travail.
Merci à Victoria Diaz-Castañeda pour avoir partagé ses connaissances en diversité biologique
et m’avoir fait profiter de son expérience dans la recherche scientifique.
Merci à Erin Cox pour m’avoir encadré, conseillé et avoir su gérer mes diverses activités de
plongée, mesures et bien entendu rédaction.
Merci à Stefano Schenone et Lidia Urbini pour avoir toujours été là durant l’expérience, avoir
pu répondre à la moindre demande durant leurs stages de Master, eux aussi, et avoir fait rêver
de l’Italie.
Merci à Samir Alliouane d’avoir été prêt à expliquer, former, réparer et bien sûr discuter
durant les activités de laboratoire.
Merci à Paul Mahacek et Arnaud Le Fur pour leur côté ingénieurs ingénieux, pour avoir su
faire profiter de leur savoir-faire pratique et leur connaissance du matériel utilisé.
Merci à Jean-Pierre Gattuso pour sa disponibilité et son implication à tout moment.
Merci à Amanda Elineau pour m’avoir indiqué comment me servir de ZooScan et à tous dans
ce bureau partagé pour m’avoir supporté durant de longues ‘séances de scan’.
Merci à toutes les personnes de l’Observatoire Océanologique de Villefranche sur Mer
(bâtiment ‘Jean Maetz’, cantine, etc.). Ce côté ‘grande famille’ a ses attraits qui vont me
manquer.
Et encore merci à ceux dont j’ai pu croiser le chemin et dont la liste serait un peu longue à
énumérer ici. Merci encore de votre écoute et de votre soutien.
En couverture :
L’herbier de P. oceanica face à la Station marine de Villefranche sur mer
(-10 m ; photographie de Delille J.)
Images issues de ZooScan (après 4 semaines de traitement)
a) plant # 9, à pH : 7,4 ; b) plant # 5, à pH : 7,8 ; c) plant # 7, à pH : 8,1
Indications de légende dans le texte

4
INTRODUCTION
Acidification de l’océan global
Actuellement, de nombreuses références au réchauffement climatique constaté sont entendues
dans les médias. Les émissions de dioxyde de carbone (CO2) sont identifiées comme étant en
grande partie responsables des augmentations de température atmosphérique mises en évidence
de 0,1 degré Celsius (°C) par décennie depuis 1950 (IPCC AR5 intergovernmental panel on
climate change, 5th attested report – panel intergouvernemental sur le changement climatique,
cinquième rapport attesté, 2014). Ce CO2 agit de concert avec les autres ‘gaz à effet de serre’ en
faisant écran au rayonnement solaire réfléchi depuis la surface terrestre, piégeant une partie
supplémentaire de la chaleur dans les basses couches de l’atmosphère terrestre. Parmi ces gaz, le
CO2 est un des plus à craindre de par sa conservation pour des milliers d’années avec des taux
moyens de 390,44 ± 0,16 ppm (parties par million ; + 40 % depuis 1750 ; Canadell et al., 2011)
ayant dépassé le seuil des 400 ppm en avril 2014 (NOAA National Oceanic and Atmospheric
Administration, station de Mauna Loa, Hawaï, Etats Unis d’Amérique in ‘Trends in atmospheric
carbon dioxide’ [en ligne]). Tout le CO2 émis par les activités humaines ne reste pas dans
l’atmosphère. En effet, l’océan global (toutes masses d’eau de mer en contact avec l’atmosphère)
puise, chaque jour, 25 millions de tonnes de CO2 correspondant à environ 25 à 30 % des
émissions anthropogéniques. Ce phénomène limite en partie la concentration de ce gaz à effet de
serre dans l’atmosphère par une séquestration naturelle qui contribue à modérer leur impact sur
le réchauffement global (Sabine et al., 2004 ; Khatiwala et al., 2013).
Le CO2 étant un polluant acide, son absorption dans l’eau de mer conduit au processus appelé
‘acidification des océans’, dont l’augmentation du taux de CO2 atmosphérique est ainsi la cause
principale (Doney et al., 2009). La référence au terme de polluant est logique (‘altéragène qui
au-delà d’un certain seul ou en cas de potentialisation développe des impacts négatifs’ selon
l’AFNOR, association française de normalisation) car il est avéré que l’apport de CO2 peut
causer une pollution1 avec des effets mesurables et observables.
1 ‘(…) l’introduction directe ou indirecte, par l’homme, de substances ou d’énergie dans le milieu marin (...)
lorsqu’elle a ou peut avoir des effets nuisibles tels que dommages aux ressources biologiques et à la faune et la
flore marine, risques pour la santé de l’homme, entrave aux activités maritimes (...), altération de la qualité de
l’eau de mer du point de vue de son utilisation et dégradation des valeurs d’agrément.‘ (Convention
internationale sur le droit de la mer, Montego-Bay, 1982 ; article 1).

5
Lorsque le CO2 se dissout dans l’eau de mer (équation 1), il réagit avec les molécules d’eau
(H2O) pour former l’acide carbonique (H2CO3 ; équation 2). Une partie de cet acide persiste dans
l’eau de mer mais la majorité se dissocie en ions hydrogène (H+) et ions bicarbonate (HCO3
-) et
ions carbonate (CO32-
; équation 3). L’augmentation de la concentration en H+ a pour effet de
réduire le pH (pH = -log10
[H+]) ainsi que la concentration en ions carbonatedisponibles, car ils
s’associent aux ions H+ pour former HCO3
- (résumé en équation 4).
(1) CO2 (atmosphérique) ⇌ CO2 (aqueux)
(2) CO2 (aqueux) + H2O ⇌ H2CO3
(3) H2CO3 ⇌ H+ + HCO3
- ⇌ 2 H+ + CO3
2-
(4) CO2 + H2O + CO32-
→ 2 HCO3-
Les pressions partielles en CO2 (pCO2) atmosphérique sont prévus être à 550 ppm environ pour
2050 et 900 ppm environ en 2100, menant à une diminution de pH dans les couches supérieures
de l’océan de 0,3 ou 0,4 unité par rapport aux mesures actuelles (IPCC AR5, 2014).
L’acidification induite par l’augmentation de la pCO2 (‘acidification des océans’ AO ou OA
ocean acidification) pourrait agir sur les organismes marins, modifiant leur environnement
chimique avec des effets positifs ou négatifs, directement par la modification du pH ainsi qu’au
travers de changements de concentration des éléments (e.g. [CO2] ↗, [CO32-
]↘).
Photosynthèse
L’augmentation attendue de [CO2]pourrait favoriser la fixation du CO2 lors de la photosynthèse.
Il a été montré dans la dernière décennie que l’élévation de [CO2] aurait un effet positif sur
certaines espèces de phanérogames marines (‘plantes à fleurs’, aujourd’hui désignées sous le
terme Magnoliophytes) ou ‘algues’ benthiques (groupe polyphylétique ; Andersson et al., 2011).
À l’inverse, les taux de photosynthèse de la majorité des espèces phytoplanctoniques étudiées
jusqu’à aujourd’hui ne seraient que faiblement augmentés (Riebell & Tortell, 2011). Ceci
s’explique car le carbone n’est pas un élément limitant pour la photosynthèse, puisque la
majorité des espèces a développé la capacité à utiliser HCO3-, forme de carbone la plus
abondante de l’eau de mer, à l’aide de l’enzyme anhydrase carbonique leur permettant de
convertir ces ions en CO2 (Falkowski & Raven, 1997 ; Tortell et al., 1998).
Calcification
Le phénomène de calcification se produit chez un grand nombre d’êtres vivants en mer : ‘algues
calcaires’, coraux, coquillages, annélides, foraminifères, échinodermes, crustacés et bryozoaires.

6
En grande majorité, les organismes dits ‘calcaires’ utilisent le calcium Ca2+
et les ions carbonate
pour former le carbonate de calcium CaCO3 (équation 5)2.
(5) Ca2+
+ CO32-
→ CaCO3
Une diminution de la disponibilité en ions carbonate entrainerait une diminution sensible des
taux de calcification et serait une menace pour la survie de nombreuses espèces (Kroeker et al.,
2013). Les effets de l’acidification des océans sur les organismes calcaires et non-calcaires ont
été communément étudiés durant la dernière décennie à partir d’expériences menées sur le court
terme et sur des organismes mis hors milieu naturel pour des facilités de gestion.
Modèle d’étude
Les épibiontes (organismes, animaux et végétaux, vivant à la surface d’un support vivant) ou
épiphytes3utilisent souvent comme support les plantes et les ‘algues’ marines. Une partie de ces
épibiontes est mobile, tandis que d’autres épibiontes sont sans déplacement depuis leur support.
Beaucoup de ces organismes épibiontes sont à squelette, coquille et/ou parties externes formés
de CaCO3.
La posidonie Posidonia oceanica (Linnaeus) Delile (Figure 1) est un Magnoliophyte marin
présentant pourtant de façon prédominante une reproduction végétative (Boudouresque et al.,
2012). La zone de croissance des feuilles est située à leur base : les feuilles juvéniles (moins de 5
centimètre (cm) de longueur) et les feuilles intermédiaires sont dépourvues de gaine basale (ou
pétiole) alors que les feuilles dites adultes en sont pourvues. Après un temps (5 à 8 mois en
moyenne, jusqu’à 13 mois) les limbes des feuilles tombent, tandis que le pétiole reste fixé au
rhizome. P. oceanica représente un ingénieur d’écosystème4 et est une espèce-clé5 d’écosystème
littoral photophile. En effet, les plants de P. oceanica regroupés en dense population peuvent
constituer un habitat abrité préférentiel et une source de nourriture (par P. oceanica elle-même
ou en abritant certaines espèces-proies). Cet écosystème constituése nomme ‘herbier’.
2 Le CaCO3 existe majoritairement sous 2 formes cristallines particulières (aragonite et calcite).
3 ‘Epiphytes’ est parfois utilisé pour désigner seulement les organismes végétaux sur surface déterminée.
Étymologiquement, ce devrait être compris comme ‘à la surface d'un végétal’, du grec έπί ‘sur’ et φυτόν
‘végétal’. En raison de cette grande disparité, le terme ‘épibiontes’, plus généraliste, sera choisi ici.
4 ‘Espèce qui contrôle la disponibilité en ressources (autre qu’elle-même) pour d’autres espèces en provoquant des
changements physiques dans le matériel biotique ou abiotique’ (Lawton, 1994).
5 ‘Espèce dont l'impact sur le fonctionnement de l’écosystème est plus important que ce que l’on pourrait en
déduire de son abondance’ (Bond, 2001).

7
Les feuilles étant caduques et les rhizomes restant fixés, les débris de ces organismes mobiles et
surtout les restes de ces organismes immobiles forment un sédiment autochtone (Pergent et al.,
2012) à la base des herbiers. Ces restes peuvent s’accumuler en hauteur au fur et à mesure de la
croissance de l’herbier, formant une ‘matte’.
Figure 1 : P. oceanica
(Photographie de Delille J. en rade de Villefranche sur mer, 10 m de profondeur ;
d'après l'illustration de Berlioz T. – wikihydro.developpement-durable.gouv.fr)
Objectifs de l’étude
Les épibiontes des feuilles de P. oceanica étant principalement des organismes calcaires, ceux-ci
peuvent potentiellement voir leurs taux de calcification diminués en contact avec l’eau de mer
acidifiée.
Peud’études ont cependant été réalisées sur ces organismes et la majorité en a été faite en milieu
naturellement acidifié par des rejets de CO2 d’origine volcanique (Martin et al., 2008 ;
Donnaruma et al., 2013) concluant à des effets importants en comparant les communautés
établies près du site d’Ischia (Italie) à des distances différentes depuis la source de CO2.
Ces études sont intéressantes car les communautés ont été soumises à des niveaux de pH bas
depuis des milliers d’années mais les résultats sont difficilement interprétables car le pH est
supposé augmenter avec la distance à la source d’acidification mais varie fortement en fonction
des courants et de l’intensité des rejets. De plus ces étudesétaient grandement dépendantes des
émissions de CO2 d’origine volcanique, variables au cours du temps et ne reflétaient pas les
niveaux de pH ambiant projetés pour la fin de ce siècle en Méditerranée.

8
Dans le cadre de cette étude conjointe6, des plants de P. oceanica ont été mis en culture dans des
mésocosmes (milieu semi-naturel) pour cinq semaines à trois niveaux de pH différents : 1) un
pH de 8,1 considéré comme référence ou ‘ témoin’ de l’expérience. Ce pH est celui usuellement
relevé au Point B (Baie de Villefranche sur Mer ; Figure 2) à profondeurs comparables, avec
capteurs de précision (Gattuso, non publié) et est supposé être uniforme en tout point dans la
rade (sauf conditions particulières), 2) un pH de 7,8 ou pH supposé intervenir dans les 90
prochaines années suite à une baisse de 0,3 à 0,4 du pH océanique (IPCC WGI, AR5, 2014) et 3)
un pH de 7,4 ou pH ‘extrême’. Ce dernier niveau de pH a été choisi car, dans les conditions de
salinité et de température de l’étude, il correspond au seuil au-delà duquel l’eau de mer est
légèrement sous-saturée au regard de l’aragonite (une des formes de CaCO3 la plus soluble).
Quantifier ces changements et attester des possibilités de réaction du recouvrement épibionte à
l’acidification programmée des océans peut fournir une base pour la prise de décision afin de
préserver la biodiversité telle qu’elle est connue dans les herbiers de P. oceanica.
L’étude s’est donc portée sur le limbe des feuilles, partie aisément mise potentiellement à
disposition pour la chaine alimentaire et pour lesquels la présence des épibiontes semble être un
critère de choix (Tomas et al., 2006 ; Gacia et al., 2009).
Pour la première fois, trois aspects liés à ces épibiontes ont été considérés et mesurés sur les
plants exposés aux trois niveaux de pH : 1) à plusieurs reprises durant la culture, les taux de
calcification ont été mesurés à partir de la technique de l’anomalie d’alcalinité (Smith & Key,
1975), 2) à la fin de la culture, les masses des épibiontes ont été quantifiées et finalement, 3) les
surfaces et la diversité de recouvrement ont été ont été estimées à partir d’une technique
d’imagerie.
Hypothèses
Les hypothèses de travail établies au début de l’expérience étaient que : 1) les surfaces de
recouvrement par ces épibiontes et leur croissance (liée au taux de calcification) diminuent avec
la baisse du pH et 2) les populations présentes peuvent réagir de manières variées à la diminution
du pH avec des modifications possibles de la communauté.
6 Conjointe, car Stefano Schenone, étudiant en Master de l’Université de Gènes (Italie), a étudié les réponses
photosynthétiques des P. oceanica dans les mêmes conditions en parallèle de cette étude ici présentée.

9
MATERIEL ET METHODES
Lieu d’étude et matériel biologique
La collecte ou prélèvement initial a été effectuée devant la station marine dans la baie de
Villefranche sur mer (Alpes Maritimes, France métropolitaine ; Figure 2), le 3 février 2014, en
réalisation unique par 12 m de profondeur.
Figure 2 : Carte figurant la localisation en rade de Villefranche sur mer
(Coordonnées du point B -WGS 84- : 43° 41,10' N - 7° 18.94' E. ; données issues du
SHOM)
Il a été choisi de prélever des plantsde P. oceanica avec morceau de rhizome à racines
apparentes, si possible attachés ensemble (pour éviter les épanchements des lésions).
Le prélèvement a eu lieu en plongée autonome : les plants ont été placés dans un filet sous-marin
et récupérés de façon ponctuelle et aléatoire en site reconnu comme représentatif de l’herbier
pour éviter les perturbations répétées (qui rendraient le site non-représentatif). La méthode de
collecte choisie est le hasard à paramètre sélectionné (ou aléatoire stratifié) de plants faisant 40
cm de longueur maximale de feuille. Ainsi, ce sont supposément des individus d’âges similaires,
compatibles avec la longueur des feuilles en moyenne depuis la pointe à la garde du pied à cette

10
période de l’année qui ont été prélevés. Le recouvrement foliaire par les épibiontes a été estimé
visuellement similaire en moyenne sur les différents plants.
Traitements post-prélèvement
Ces échantillons ont pu être marqués à la calcéine pendant 48 heures (h) à une concentration de
60 mg/L-1
environ (milligrammes par litre) pour identifier les variations de potentiel de
croissance des espèces calcifiées.
La calcéine C30H26N2O13 est un colorant qui fluoresce à λ = 495/515 nm (nanomètres) de
longueur d’onde. Ce colorant biologique aux concentrations considérées est sans effets
physiologiques avérés, et sert comme marquage des structures calcifiées. Les résultats sont
encore en cours de traitement et ne seront pas présentés dans ce document.
Paramètres de l’expérience
Acclimatation
Après le marquage a débuté une période d’acclimatation dans les récipients pourune semaine à
pH naturel (~8,1) avant la mise en conditions de pH contrôlé.
Conditions de culture
La pièce où s’est déroulée l’expérience a été régulée à 14 °C (degré Celsius), température proche
de celle du milieu naturel afin d’éviter le réchauffement rapide des petits volumes.
L’éclairement a été régulé de façon conforme aux durées constatées à l’extérieur : 9,5 heures (h)
illuminées par jour, ce qui est typique à cet endroit pour les mois d’hiver. Un faisceau
d’éclairage (Marin Day T5 1450 80 Watt Light Bank) a été placé à environ 15 cm au-dessus des
récipients, ce qui produit un éclairement énergétique (irradiance) de 50 µmol photon.m-2
.s-1
au
fond des récipients à 120 µmol photon.m-2
.s-1
à la surface.
Paramètres biologiques
Afin d’être compatible au mieux avec la densité d’individus présenteau sein de l’herbier, cinq
plants de P. oceanica ont été groupés par échantillon (sample). Chaque échantillon a été placé en
récipient contenant de l’eau de mer à pH contrôlé et cet ensemble est ici nommé ‘ récipient’.
Quatre échantillons ont été considérés similairement pour chaque traitement (réplicats). Au total,
60 plants de P. oceanica ont été incubés (5 plants x 3 traitements x 4 réplicats).
Quinze plants supplémentaires ont été utilisés pour tester les méthodes et mesures ou procéder à
l’identification des épibiontes majoritaires avant incubation. Une table d’identification par

11
critères et des méthodes de comptage préférentielles ont alors été mises en place.
Les rhizomes ont été attachés à une grille plastifiée pour faciliter la prise de tous les pieds à la
fois lors des incubations réalisées par la suite. Les feuilles ont été laissées libres dans l’eau en
mouvement.
Un mélange stable sable/sédiment issus du lieu d’étude a été ajouté au fond de chaque récipient.
Ce mélange a été filtré par une maille de 1 mm² (millimètre carré) afin d’éviter la présence de
macroorganismes hors sujet d’étude.
Récipients
Les récipients d’étude sont 12 cylindres de 18 cm de diamètre pour 33 cm de haut. Ainsi ceci
forme un volume total estimé de 8,4 L pour 0,8 L environ de sédiment, soit 7,6 L d’eau de mer
circulant dans chaque récipient.
Ces cylindres ont été conservés dans un grand bac à eau circulante, alimenté en eau de mer
ambiante qui est acheminée depuis point de prélèvement en mer (conditions identiques au lieu de
collecte) avec débit élevé et évacuation aisée. Ce bac a permis le maintien en température des
récipients. Ceux-ci ont été placés de façon aléatoire dans le système expérimental (Figure 3) pour
éviter les effets d’influence de la position du lieu de culture.
Cette même eau de provenance extérieure a été distribuée dans des réservoirs de 160 L qui ont
été maintenus en circulation par pompes et dont le surplus de remplissage a été évacué au moyen
de tuyaux de gros diamètre.
La distribution d’eau a été faite par gravité dans les récipients reliés à ces réservoirs (débit
variable selonles récipients pour un débit moyen de 0,5 L par minute (L.min-1
).
Régulation du pH
Deux des trois réservoirsont été maintenus aux niveaux de pH prédéfinis (7,4 et 7,8) par bullage
de CO2 pur (Air Liquide©) contrôlé par une ordinateur d’aquariologie IKS (Internes
Kontrollensystem–ComputerSystem GmbH). Les électrodes de ces systèmes ont été
intercalibrées tous les deux jours, par une comparaison entre leurs mesuresetcelles obtenues avec
des électrodes en verre (Metrohm®). Celles-ci sont calibrées sur l’échelle totale avec des
tampons salins (2-amino-2-hyroxymethyl-1,3-propanediol ou TRIS et 2-aminopyridine ou
AMP).

12
Figure 3 : Schématisation (a) et photographie (b) du montage expérimental avec les
récipients contenant les plants de P. oceanica (4 réplicats par traitement de pH).
T °C : Température ; CO2 : dioxyde de carbone (sous forme gazeuse)
Mesures analytiques et protocole
Les pH dans les 12 récipients ont été mesurés trois fois par semaine avec les électrodes en verre
et suivant le protocole détaillé ci-dessus. De plus, des prélèvements ont été réalisés pour mesures
de contrôle de carbone inorganique dissous CT et d’alcalinité totale AT. Il est à noter
qu’uniquement un réservoir a été échantillonné pour l’ATcar il est établi que l’ajout de CO2 n’a
pas d’influence sur ce paramètre (Wolf-Gladrow et al., 2007).

13
Tous les prélèvements ont été effectués suivant les protocoles établis (Dickson et al., 2007).
Pour les mesures d’AT, les échantillons de 500 mL ont été filtrés sur des filtres GF/F de 47 mm
de diamètre (Whatmann®) dès prélèvement.
Pour la mesure de la concentration en CT, des échantillons de 60 mL prélevés sans bulles ont été
directement stockés dans des bouteilles brunes en verre jusqu’à analyse.
Afin de limiter l’évolution des paramètres physico-chimique de l’eau prélevée par action
microbienne, un empoisonnement au chlorure de mercure (HgCl2) a été réalisé (ajout de 100 µL
par échantillon) après chaque prélèvement. Enfin, les échantillons ont été conservés à
température ambiante en environnement sombre jusqu’au jour de l’analyse.
Analyse de l’alcalinité totale en cellule ouverte
L’AT d’un échantillon d’eau de mer est définie comme l’équivalence entre le nombre de moles
d’ions hydrogène et l’excès de d’accepteurs de protons sur les donneurs de protons par
kilogramme (kg). Les bases sont ici les acides faibles (avecforce ionique nulle et constante de
dissociation K ≤ 10-4,5
à 25 °C) et les acides les acides forts (K ≥ 10-4,5
).
(6) AT = [HCO3-] + 2 [CO3
2-] + [B(OH)4
-] + [OH
-] + [HPO4
2-] + 2 [PO4
3-] + [SiO(OH)3
-] +
[NH3] + …– [H+]L – [HSO4-] – [HF] – [H3PO4] – …
Dans cette formule, [H+]L représente la concentration en ions hydrogène libres et les points de
suspension traduisent la présence d’ions en quantités négligeables dans l’eau de mer.
Pour la mesure, une quantité connue d’eau de mer à analyser est placée dans un bécher ouvert à
l’air libre et titrée par l’ajout d’acide chlorhydrique (HCl) tout en mesurant les variations de pH.
Ce bécher ouvert fait supposer le fait que la concentration de CT (donc la concentration en ions
bicarbonate [HCO3-]) est d’environ 0 dans cette gamme de pH après dégazage du CO2 (seule
forme de carbone inorganique persistant à ce niveau de pH). Une électrode en verre permet de
mesurer les valeurs électromagnétiques (electromagnetic field ou champ électromagnétique) tout
au long du titrage. L’AT est déduite du volume équivalent et des valeurs de pH suivant une
approche mathématique. Pour de plus amples détails, la lecture du ‘Guide to best practises for
ocean CO2 measurements’ (Dickson et al., 2007), chapitre 4 - SO3b est conseillée.
Les mesures ont été effectuées en triplicats, à partir de sous-échantillons de 50 mL par titrage
potentiométrique (à 22 °C) à l’aide d’un Dosimat (Metrohm®
), d’HCl à 0,1 mol/L-1
, d’une sonde
de température (Metrohm®) et d’une électrode pH (Metrohm
®) calibrée sur l’échelle totale à
partir de tampons TRIS. La qualité des mesures a été testée de façon régulière à partir du titrage

14
d’échantillons d’eau de mer et comparés à des standards (Dickson, Batch 132 ; précision à ± 1
µmol.kg-1
du standard).
Les concentrations de CT ont été mesurées en triplicat sur des échantillons de 1,2 mL à l’aide
d’un analyseur de carbone inorganique dissous (AIRICA, Marianda©). Le principe de cette
mesure est d’acidifier l’échantillon avec un acide fort (acide phosphorique, 3 mol.L-1
) afin de
transformer tout le carbone inorganique dissous en CO2 puis, après bullage de l’échantillon avec
un gaz neutre (azote ultra pur, AirLiquide©), de mesurer la quantité de CO2 dégazée avec un
analyseur de gaz infrarouge (LI-6262, Licor©).
Les autres paramètres de la chimie des carbonates ont été calculés avec le module SeaCarb
2.4.10 sous R, mis à jour en cours d’expérience (Lavigne et al., 2011).
Mesures des taux de calcification en incubation
Système d’incubation
Pour les 5 plants contenus dans chaque récipient, des incubations ont été réalisées pendant 7 h
dans des incubateurs supplémentaires à 3 reprises pendant l’expérience : au début de
l’expérimentation après marquage (première mesure, t0), après 2 semaines de culture en milieu à
pH contrôlé (t2S) et après 4 semaines de culture dans ce même milieu (t4S).
Pour chaque session d’incubation (Figure 5), un bac de grande taille a été installé dans la
chambre à température régulée. Il a été rempli d’eau circulante et placé dans des conditions
identiques au milieu de culture (éclairement et température). Les trois incubateurs étaient des
cylindres en plexiglas de 14,3 cm de diamètre sur 31,7 cm de hauteur, placés dans cette eau afin
d’être maintenus en température constante pour le temps d’incubation. Les plants extraits des
récipients initiaux ont été placés en eau traitée au pH similaire (ambiant/contrôle pH : 8,1/pH :
7,8/pH : 7,4) régulés par un second ordinateur (IKS).
Avant placement, la grille avec les rhizomes attachés a été agitée pour évacuer le sédiment
superficiel. Les incubations ont été réalisées, pour chaque période (t0, t2S et t4S), sur 4 jours afin
d’incuber tous les réplicats d’expérience. Avant et après incubations, des échantillons ont été
prélevés afin de mesurer AT. Les échantillonnages et les mesures d’AT ont été réalisés de la
même manière que décrit ci-dessus.

15
Estimation des taux de calcification
Les ΔAT (différences en AT ; en μmol.kg-1
), après 7 h entre les mesures de l’état initial et l’état
final de chaque incubation, sontconsidérées comme proportionnelles à la quantité de carbonate
de calcium CaCO3 ayant précipité en faisant l’hypothèse que dans le cadre de nos incubations
seules la précipitation et la dissolution de CaCO3 influencent de façon significative l’AT.
Comme 1 mole de CO32-
est consommée pour 1 mole de CaCO3 produite, les taux de
calcification G peuvent être calculés en μmol.kg-1
.h-1
(équation 8), avec une durée d’incubation
(t) de 7 h.
(7) G = -0,5 ΔAT/t
Ces taux de calcification ont ensuite été exprimés par unité de masse sèche de feuille (voir plus
loin le protocole utilisé) en tenant en compte de la densité de l’eau de mer aux conditions de
température (14,5 °C) et de salinité (valeur moyenne de 37,5) pour un volume des cylindres
d’incubation de 5 L environ). La variation relative des taux de calcification (%), entre
l’incubation initiale (t0) et les suivantes (t2S et t4S) a été ensuite calculée et utilisée dans les tests
présentés plus loin.
Mesures de fin d’expérience
Les 15 plants supplémentaires initialement prélevés ont été étudiés pour établir les bases et
méthodes d’identification utilisées après la période de culture.
Recouvrement
Pour mesurer l’abondance par pourcentage de recouvrement (%), une observation directe sur 60
plants est souvent peu réalisable devant l’urgence en fin d’expérience. L’idée de prendre des
Figure 4 : Photographie de l’installation des récipients d’incubation, en
configuration de mesure des taux de calcification.
Les mesures pour l’échelle exacte sont notées dans le texte.

16
photographies numériques des feuilles a semblé judicieuse. En effet, le gain de temps de prise de
données sur un organisme marin hors de l’eau de mer est appréciable.
Pour produire ces photographies à traiter, l’utilisation de microscope optique a montré des
avantages, mais un problème de taille est survenu lors de l’observation de feuilles de P.
oceanica : leur longueur justement, forçant leur découpe pour observation centimètre par
centimètre. Afin de ne pas détruire l’échantillon tout en restant à grande résolution de capture,
l’utilisation d’un scanner spécialement conçu s’est révélée idéale : le ZooScan (Hydroptic).
ZooScan est un scanner d’échantillons biologiques à efficience reconnue (Gorsky et al., 2010)
constitué d’un corps protégé des actions de l’eau et d’un système de prise de vues à grande
résolution. Sa surface de table de captage est assez importante pour scanner les feuilles d’un
plant complet de P. oceanica avec une résolution de 2400 dpi (dot per inch ou pixel par pouce, 1
pouce = 2,54 cm)
Les feuilles ont ainsi été séparées de la base racinaire, scannées par plant et traitées par la suite
selon les récipients et les traitements (Figure 6).
Traitement des images
Les images obtenues ont été traitées par la suite avec ImageJ (Rasband, 1997-2014) et le module
de comptage spécifique (plug-in Cell counter) sur les intersections d’une grille superposée (grille
systématique séquentielle aléatoire). Les organismes situés à cette intersection ont été identifiés
au mieux possible selon la qualité d’image et l’état dans lequel ils étaient. Un tableau a été ainsi
réalisé traduisant les observations avec l’aide des données à disposition (Van Der Ben, 1971 ;
Jacquemart & Demoulin, 2006).
Le choix de résolution (écartement des points) aété fait après essais successifs à différentes
mesures de surface. Il a pu être ainsi constaté qu’une résolution de grille de un point par
centimètre carré était véritablement trop importante pour prendre en compte les organismes ici
étudiés dans un microcosme.
Une résolution de 1 point par 0,1 cm² a semblé être pertinente pour apprécier l’abondance en
réduisant le nombre de points à étudier. Le recouvrement est comparable entre les deux faces de
ces feuilles placées en microcosmes. De même, se limiter aux feuilles les plus longues supposées
être les plus vieilles m’a paru ne pas exprimer l’abondance complète dans l’herbier ou le
microcosme. Cette feuille âgée a pu être tombée auparavant ou cette longue feuille signifier une
croissance rapide. Il a été choisi de traiter les données concernant le plant entier quand celui-ci
est sélectionné afin d’avoir une surface de recouvrement, une abondance directement mesurée

17
par une face sur un plant, feuilles âgées ou juvéniles comprises, mettant en valeur les restes de
feuilles par la même voie. Une seule face a donc été étudiée pour des raisons de temps.
Figure 5 : Acquisition par ZooScan (images des plants # 4 tirés du récipient # 9 –
pH : 7,4) et traitement par ImageJ–Cellcounter avec la grille utilisée présentée en (a)
et son traitement d’identification (b).
Deux plants par récipient, soit 8 plants par traitement pour un total de 24 plants de 5-6 feuilles
sur 18 cm de longueur en moyenne sur une face, ont été observés aléatoirement de cette façon,
pour un total de 36 000 points environ. Ce grand nombre a permis ainsi de limiter les erreurs
d’identifications relatives à l’image obtenue et permet de montrer la surface de recouvrement en
figurant un cinquième représentatif de la surface totale mise en culture. Des difficultés de
reconnaissance précise sur une image numérique même à grande résolution ont conduit à des
regroupements par catégories observables. La présence de ces regroupements a été quantifiée et
rapportée à la surface de feuilles étudiée sous la forme d’un pourcentage de couverture. Ces
pourcentages sont alors comparables entre plants et entre récipients (la moyenne des
pourcentages tirés de ces plants qu’ils contiennent) selon les groupes retenus après les multiples
observations et essais successifs de détermination pratique (Tableau 1). Le blanchiment des
algues a été considéré comme une marque de la possible mort de l’organisme (Anthony et al.,
2008).
Masse sèche de CaCO3
Après les mesures de diversité et de recouvrement foliaire, les feuilles ont été râpées avec un
stylet anguleux pour récupérer les épibiontes recouvrants.

18
Tableau 1 : Groupes identifiés visuellement en images tirées de ZooScan
Groupe identifié Référence visuelle
(échelle variable, non indicative)
Algues encroûtantes calcifiées ‘vivantes’
(Crustose corraline algae ‘alive’ CCA-A)
Algues encroûtantes calcifiées ‘dissolution–décès’
(Crustose corraline algae ‘dissolved–dead’ CCA-D)
Bryozoaires à test calcifié
(Calcareous bryzoans C bry)
Annélides polychètes Spirorbis sp.
La bande orange représente le marquage à la calcéine
(Spirobis)
Foraminifères calcaires
(calcareous forams C-form)
Biofilm – Organismes unicellulaires agglomérés non identifiés
(Biofilm)
Phéophycées Dictyota sp.
(Dictyota)
Algues filamenteuses non identifiées
(filamentous algae Unknown filamentous)
Céramiales
(Ceramiales)
Bryozoaire Electra posidoniae
(Electra)
Hydrozoaires non identifiés
(Hydroids Hydroids)
Spongiaires non identifiés
(Sponges)

19
Ce volume est collecté dans des tubes de 2 mL (Eppendorf), séché en étuve (60 °C pendant 6
jours) et repesé pour obtenir le poids sec (dry weight d. w.) d’organismes recouvrants. En
présence d’HCl, le CaCO3 forme du CO2, de l’eau H2O et du chlorure de calcium CaCl2
(équation 8).
(8) CaCO3 + 2 HCl → CO2 + H2O + CaCl2
Après acidification avec 2 mL d’HCl 5 %, dégazage du CO2 à l’air libre, rinçage à l’eau milliQ
(extrêmement filtrée) et séchage en étuve, le poids calcifié (CaCO3) a été déduit par différence.
Statistiques
L’ergodicité7 et la stationnarité
8 des données a été évaluée compatible avec le fait que les
analyses statistiques sur faible échantillonnage soient reportées à l’herbier entier.
Des analyses de variances ont été réalisées (voir plus bas). Dans le cas de différences
significatives observées, des tests complémentaires (post-hoc) ont été réalisés pour comparer les
traitements par paires (Tuckey ou Dunns suivant si les tests sont paramétriques ou non
paramétriques).
Taux de calcification
La différence relative de taux de calcification calculée entre les mesures prises lors des
incubations (t2S et t4S) et les mesures initiales (t0) pourles3 traitements a été prise comme base
de calcul.
Une analyse de variance à 2 facteurs pour mesures répétées (2-way repeated measures ANOVA)
a été utilisée en considérant le temps de traitement (‘Semaine’) et le pH comme facteurs
possibles dont la relative influence est quantifiée par la valeur de significativité p (ou valeur-p).
Un facteur de significativité inférieur à 5 % (soit p = 0,05) est considéré comme montrant une
réelle influence du facteur sur les mesures répétées.
Masse de CaCO3
Le test de normalité (Shapiro-Wilk ; p < 0,050) ayant échoué sur ces valeurs, une analyse de
variance à un facteur (pH) sur les rangs (non paramétrique : Kruskal-Wallis) a été réalisée afin
de tester les différences entre les niveaux de pH.
7 Relative à l’hypothèse ergodique : la valeur moyenne prise pour une partie du système à un temps donné
correspond à la moyenne de l’ensemble des valeurs au cours du temps, observer une partie revient à pouvoir
conclure sur l’ensemble 8 Le processus sous-jacent ne change pas avec le temps au cours de l’expérience. L’espérance et la variance sont
constantes au cours du temps.

20
Pourcentages de couverture
Le test de normalitéa été passé avec succès (Shapiro-Wilk ; p = 0,449) et donc une analyse de
variance à 1 facteur a été réalisée avec de tester des différences entre les niveaux de pH.
Les statistiques multivariées (multivariate statistics) ont été utilisées pour tester l’influence du
traitement (pH) sur la composition et l’abondance des épibiontes.
Les groupes ont été identifiés à un niveau taxonomique compatible avec les observations
(Tableau 1 et description correspondante) pour 2 plants par récipient.
Les pourcentages de couverture par groupe ont été ensuitetransformés par racine carréepour
stabiliser les variances, en faisant moins peser les espèces les plus communes sur le résultat tout
en prenant en compte la répartition par parcelles des communautés épibiontes.
Avec le logiciel PRIMER (Clarke & Gorley, 2006 ; PRIMER v6.15) et selon l’indice de Bray-
Curtis (S17), la comparaison entre traitements et récipients a permis de construire une table de
similarité. Une analyse des similarités (ANOSIM ou analysis of similarities) à 1 facteur (pH)
pour 999 permutations a pu être réalisée pour tester statistiquement des différences entre les
assemblages résultant des traitements par les différents pH. La similarité est indiquée par la
valeur R (Clarke, 1993) variant de +1 à -1 (groupes totalement différents), la valeur 0 indiquant
une similarité complète.
Une représentation en nMDS (non metric multidimentional scaling) a été faite à partir des
résultats obtenus pour mettre en évidence de possibles regroupements.
Une analyse SIMPER (ou similarity percentage) a pu être utilisée pour identifier quels groupes
contribuaient majoritairement aux structures observées pour chaque traitement et pour indiquer
quels groupes étaient les plus impactés.

21
RESULTATS
Mesures des réservoirs
Les températures (Temp. °C), AT et CT mesurées dans les réservoirs pendant les périodes de
culture à pH régulé ont été relativementstables (Tableau 2).
Tableau 2 : Mesures moyennes d’alcalinité totale (AT), de carbone inorganique total
(CT), de température (Temp.), et de saturation de l’eau de mer au regard de
l’aragonite ou de la calcite (Ω) en moyenne dans les réservoirs (pH : 8,1/7,8/7,4) ±
écart-type.
Réservoirs pH : 8,1 pH : 7,8 pH : 7,4
AT (µmol.kg-1
) 2557,1 ± 5,8 2557,1 ± 5,8 2557,1 ± 5,8
CT (µmol.kg-1
) 2264,6 ± 13,8 2461,1 ± 40,5 2584,0 ± 34,3
Temp. (°C) 14,7 ± 0,3 14,7 ± 0,3 14,7 ± 0,3
Ω aragonite 3,1 ± 0,1 1,4 ± 0,3 0,6 ± 0,2
Ω calcite 4,9 ± 0,2 2,2 ± 0,4 1,1 ± 0,3
Les pH des réservoirs ont été maintenus constants au mieux durant le temps de l’expérience et
les récipients n’ont connu que des variations minimes de pH de culture.
Ces mesures de pH dans les récipients restent groupées, indiquant bien qu’aucune mesure faite
ne peut être écartée pour des biais expérimentaux (Figure 7).
Figure 6 : Mesures de pH au cours du temps dans les 12 récipients et
les 3 réservoirs correspondants

22
Taux de calcification
Pour tous les plants et à tous niveaux de pH, le taux de calcification a diminué entre l’état initial
d’expérience (après période d’acclimatation) et les périodes suivantes (Tableau 3).
Les changements relatifs de taux de calcification par rapport à la mesure faite en t0 sont
présentés en Figure 7. Dans les récipients à pH : 8,1 de contrôle, les taux de calcification en
semaine 2 ont baissé (changement relatif = -1,2 avec erreur standard e.s. de 0,7) puis ont remonté
en semaine 4 (changement relatif = -0,5 e.s. 0,06). Pour le pH : 7,8 la diminution a été similaire
au contrôle en semaine 2 mais s’est maintenue en semaine 4. Les plus fortes diminutions de
calcification ont été observées pour les récipients exposés à un pH : 7,4 et se sont maintenues en
semaine 4. Une dissolution nette à pH : 7,4 est observée par des valeurs négatives en taux de
calcification.
Tableau 3 : Taux de calcification G pour les groupes à pH de traitement. Valeur
initiales : t0 ; après 2 semaines : t2S ; après 4 semaines : t4S
G < 0 signifie une décalcification en cours, G > 0 signifie une calcification en cours.
Un impact de la mise en culture est noté sur le groupe à pH : 8,1 (valeurs ~nulles en
moyenne à 2 semaines).
Groupe G(t0) μmol.kg-1
.h-1
G(t2S) μmol.kg-1
.h-1
G(t4S) μmol.kg-1
.h-1
pH : 7,4 1,19 -1,21 -1,17
pH : 7,8 1,16 0,10 0,20
pH : 8,1 0,90 0,05 0,55
En semaine 2, en raison de la forte variabilité du contrôle en récipient # 7, aucune différence
significative avec les 2 autres traitements n’a pu être mise ne évidence. Il existe une différence
nette et significative entre les traitements 7,8 et 7,4 (p = 0,02). Les 3 traitements sont
significativement différents les uns des autres (p = 0,004 entre pH : 7,4 et pH : 7,8 ; p = 0,0005
entre pH : 7,4 et 8,1 ; p = 0,01 entre pH : 7,8 et pH : 8,1) en semaine 4, avec des diminutions
relatives des taux de calcification par rapport à la période pré-régulation du pH, qui sont plus
accentuées avec la diminution du pH.

23
Figure 7 : Changement relatif de taux de calcification par traitement (pH) au cours
du temps d’expérience. Le pH : 7,4 montre une nette décalcification. Les barres
représentent l’erreur standard (e.s.) et les lettres les différences entre les groupes.
L’analyse des variances montre que le pH semble a une influence majeure sur ces changements
relatifs en taux de calcification (Tableau 4 ; p = 0,007) selon les taux calculés (Tableau 3). Ces 3
groupes de pH seront donc traités individuellement.
Tableau 4 : Facteurs de significativité sur les changements relatifs de taux de
calcification (ANOVA à 2 facteurs pour des mesures répétées).
Facteur Facteur de significativité
Semaine p = 0,576
pH p = 0,007 **
pH/Semaine p = 0,400
Masse calcifiée
Les plants à pH : 8,1 ont montré une masse d’épibiontes grandement calcifiés en moyenne ±
erreur standard (35,5 ± 5,5 % de la masse sèche totale) alors que les plants maintenus à pH :
7,8sont légèrement moins calcifiés (29,1 ± 5,0 %). Le groupe à pH : 7,4 est très détaché (17,74 ±
2,0 %). Le test de Kruskal-Wallis a en effet montré une différence statistique entre les
traitements (p = 0,024).

24
Une comparaison post-hoc multiple (test de Tukey) a montré une différence uniquement entre les
groupes extrêmes (pH : 8,1 et pH : 7,4 ; p < 0,05), ceci étant représenté par les lettres sur la
Figure 8.
Figure 8 : Contribution (%) de la matière carbonatée (CaCO3) à la matière sèche des
épibiontes en fonction du pH de culture (moyenne ± erreur standard)
Recouvrement
De nombreuses jeunes pousses étaient présentes, peu recouvertes du fait de leur jeune âge hors
milieu naturel. Ces valeurs ont ainsi pu présenter une variabilité élevée selon les conditions de
croissance dans les différents récipients.
La structure communautaire globale a différé selon les pH utilisés (ANOSIM à 1 facteur ; Global
R = 0,80 ; p = 0,001). L’analyse par paires a montré que les communautés étaient distinctes entre
elles selon les traitements (p = 0,03 dans les 3 cas ; R = 0,64 entre pH : 8,1 et pH : 7,8 ; R = 1,00
entre pH : 8,1 et pH : 7,4 ; R = 0,96 entre pH : 7,8 et pH : 7,4).
La similarité figurée en nMDS (Figure 9) a montré que les récipients sous le même traitement
étaient ‘plus similaires’ entre eux qu’avec les récipients des autres traitements. Ces groupes de
pH ont donc pu être considérés spécifiquement.
Ce total de recouvrement (tous groupes confondus) a été mesuré à 21,5 ± 3,0 % (moyenne ±
écart-type) pour le groupe à pH : 8,1 alors que le groupe à pH : 7,4 a montré un recouvrement
cumulé de 6,6 ± 2,0 %. Le groupe à pH : 7,8 montre un recouvrement cumulé intermédiaire
(13,4 ± 2,4 %). Ces valeurs de total de recouvrement sont fortement distinctes du fait du pH (p =
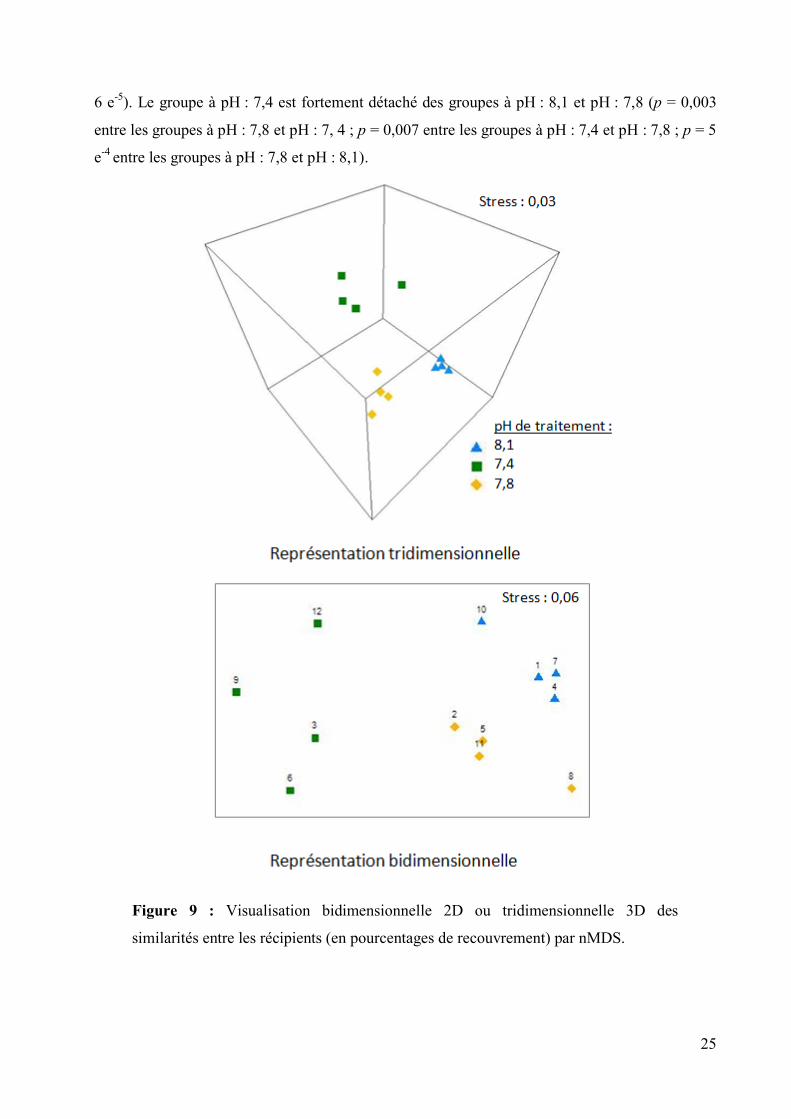
25
6 e-5
). Le groupe à pH : 7,4 est fortement détaché des groupes à pH : 8,1 et pH : 7,8 (p = 0,003
entre les groupes à pH : 7,8 et pH : 7, 4 ; p = 0,007 entre les groupes à pH : 7,4 et pH : 7,8 ; p = 5
e-4
entre les groupes à pH : 7,8 et pH : 8,1).
Figure 9 : Visualisation bidimensionnelle 2D ou tridimensionnelle 3D des
similarités entre les récipients (en pourcentages de recouvrement) par nMDS.
26
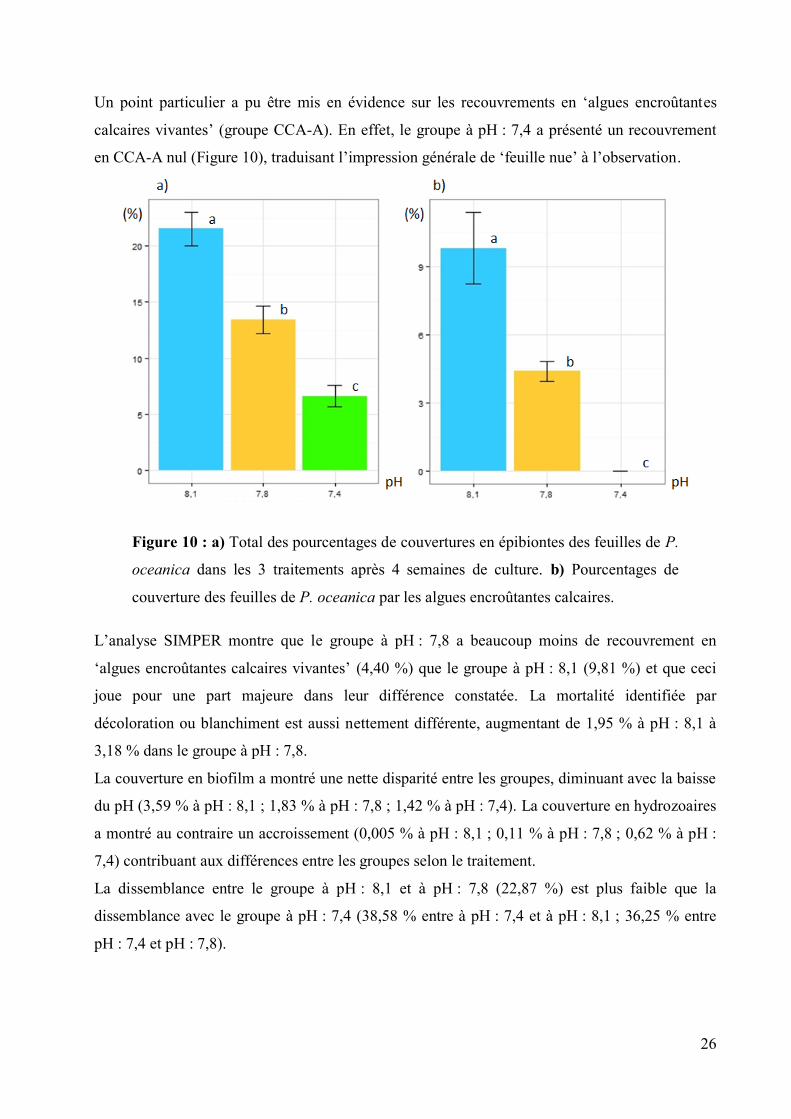
Un point particulier a pu être mis en évidence sur les recouvrements en ‘algues encroûtantes
calcaires vivantes’ (groupe CCA-A). En effet, le groupe à pH : 7,4 a présenté un recouvrement
en CCA-A nul (Figure 10), traduisant l’impression générale de ‘feuille nue’ à l’observation.
Figure 10 : a) Total des pourcentages de couvertures en épibiontes des feuilles de P.
oceanica dans les 3 traitements après 4 semaines de culture. b) Pourcentages de
couverture des feuilles de P. oceanica par les algues encroûtantes calcaires.
L’analyse SIMPER montre que le groupe à pH : 7,8 a beaucoup moins de recouvrement en
‘algues encroûtantes calcaires vivantes’ (4,40 %) que le groupe à pH : 8,1 (9,81 %) et que ceci
joue pour une part majeure dans leur différence constatée. La mortalité identifiée par
décoloration ou blanchiment est aussi nettement différente, augmentant de 1,95 % à pH : 8,1 à
3,18 % dans le groupe à pH : 7,8.
La couverture en biofilm a montré une nette disparité entre les groupes, diminuant avec la baisse
du pH (3,59 % à pH : 8,1 ; 1,83 % à pH : 7,8 ; 1,42 % à pH : 7,4). La couverture en hydrozoaires
a montré au contraire un accroissement (0,005 % à pH : 8,1 ; 0,11 % à pH : 7,8 ; 0,62 % à pH :
7,4) contribuant aux différences entre les groupes selon le traitement.
La dissemblance entre le groupe à pH : 8,1 et à pH : 7,8 (22,87 %) est plus faible que la
dissemblance avec le groupe à pH : 7,4 (38,58 % entre à pH : 7,4 et à pH : 8,1 ; 36,25 % entre
pH : 7,4 et pH : 7,8).
27
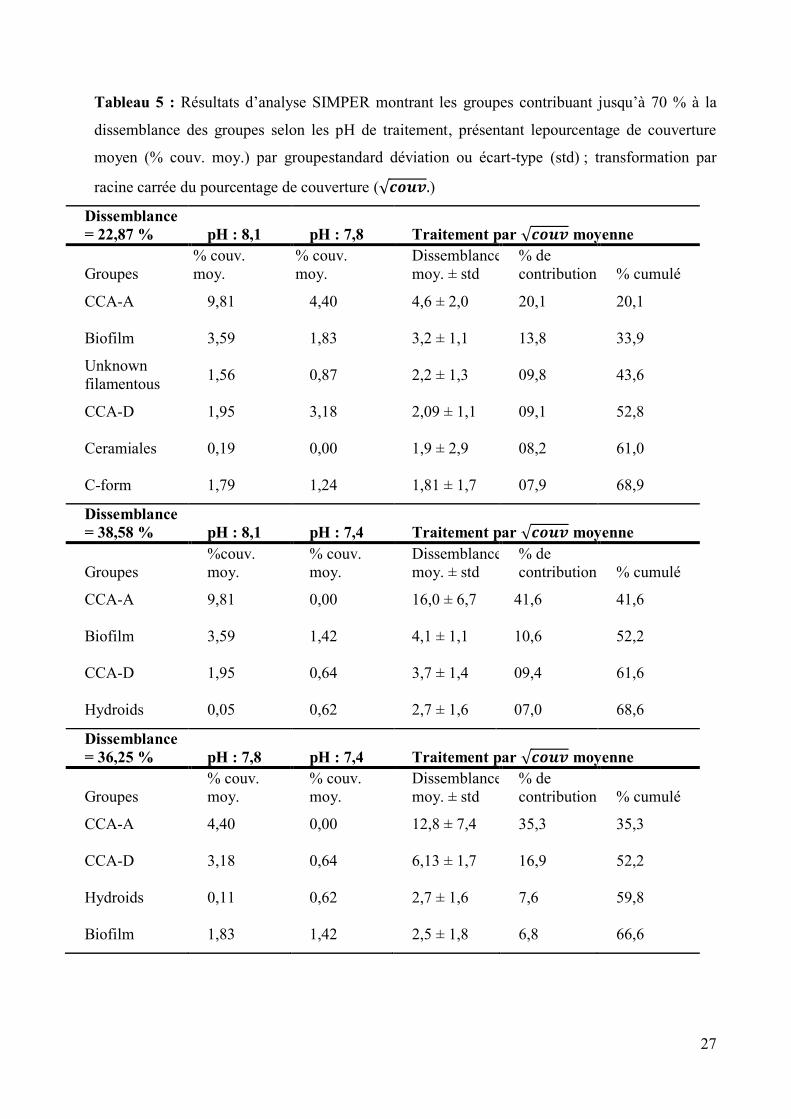
Tableau 5 : Résultats d’analyse SIMPER montrant les groupes contribuant jusqu’à 70 % à la
dissemblance des groupes selon les pH de traitement, présentant lepourcentage de couverture
moyen (% couv. moy.) par groupestandard déviation ou écart-type (std) ; transformation par
racine carrée du pourcentage de couverture (√ )
Dissemblance =
= 22,87 % pH : 8,1 pH : 7,8 Traitement par √ moyenne
Groupes
% couv.
moy.
% couv.
moy.
Dissemblance
moy. ± std
% de
contribution % cumulé
CCA-A 9,81 4,40 4,6 ± 2,0 20,1 20,1
Biofilm 3,59 1,83 3,2 ± 1,1 13,8 33,9
Unknown
filamentous 1,56 0,87 2,2 ± 1,3 09,8 43,6
CCA-D 1,95 3,18 2,09 ± 1,1 09,1 52,8
Ceramiales 0,19 0,00 1,9 ± 2,9 08,2 61,0
C-form 1,79 1,24 1,81 ± 1,7 07,9 68,9
Dissemblance =
= 38,58 % pH : 8,1 pH : 7,4 Traitement par √ moyenne
Groupes
%couv.
moy.
% couv.
moy.
Dissemblance
moy. ± std
% de
contribution % cumulé
CCA-A 9,81 0,00 16,0 ± 6,7 41,6 41,6
Biofilm 3,59 1,42 4,1 ± 1,1 10,6 52,2
CCA-D 1,95 0,64 3,7 ± 1,4 09,4 61,6
Hydroids 0,05 0,62 2,7 ± 1,6 07,0 68,6
Dissemblance =
= 36,25 % pH : 7,8 pH : 7,4 Traitement par √ moyenne
Groupes
% couv.
moy.
% couv.
moy.
Dissemblance
moy. ± std
% de
contribution % cumulé
CCA-A 4,40 0,00 12,8 ± 7,4 35,3 35,3
CCA-D 3,18 0,64 6,13 ± 1,7 16,9 52,2
Hydroids 0,11 0,62 2,7 ± 1,6 7,6 59,8
Biofilm 1,83 1,42 2,5 ± 1,8 6,8 66,6

28
DISCUSSION
Cette expérience commune montre les premiers résultats significatifs en milieu contrôlé
concernant le recouvrement par les épibiontes de P. oceanica en herbier à pH réalistes et
maintenus.
Elle étude associe pour la première fois des mesures de taux de calcification et recouvrement
spécifique, basée sur l’utilisation d’une technique généralement utilisée pour des échantillons
zooplanctoniques, le ZooScan.
La composition en épiphytes sur les feuilles de P. oceanica est un indicateur sensible de
dégradation environnementale (Martinez-Crego et al., 2010) car les réponses des organismes à
l’augmentation de pCO2 sont très variables (Noisette et al., 2013).
La diminution des taux de calcification pour le pH : 8,1 après 2 ou 4 semaines de culture par
rapport aux mesures initiales a montré que la mise en culture expérimentale a eu un effet négatif.
Le récipient # 7 a montré des impacts particuliers, et un plus fort nombre de réplicats aurait pu en
diminuer encore l’influence sur les résultats obtenus. C’est un biais de la culture en laboratoire
qui a été relativisé par le traitement statistique.
Une baisse du taux de calcification peut avoir plusieurs causes : une faible décalcification du fait
d’une dissolution (rejet de Ca2+
dans le milieu) ou une non-utilisation du Ca2+
contenu dans l’eau.
Les taux de calcification mesurés semblent être corrélés aux pourcentages de recouvrement
observés pour chaque pH de traitement. La dissolution observée sur les limbes a dû être de faible
ampleur pendant le temps d’incubation, mais avoir joué un effet accumulé pendant les semaines
de culture, limitant la production de calcaire.
La couverture en ‘algues encroûtantes calcaires’ a donc été constatée comme étant la plus
impactée par cette diminution de pH (Figure 11 ; Martin et al., 2008 ; Donnarumma et al., 2014).
Il a été observé avec le total de couvertures (somme des pourcentages de couverture par groupes)
une décroissance plus ou moins proportionnelle avec l’abaissement du pH jusqu’à atteindre 0 %
de recouvrement en ‘algues encroûtantes calcaires’ à pH : 7,4 dans les relevés faits, par
dissolution (observée parfois sous la loupe binoculaire) ayant eu lieu au cours des semaines de
traitement. L’analyse de similarité a montré que le groupe à pH : 7,8 a beaucoup moins de
recouvrement en ‘algues encroûtantes calcaires’ (7,58 %) que le groupe à pH : 8,1 (11,76 %),
‘mortes’ ou ‘vivantes’, ce qui met en évidence un processus en cours avec l’augmentation de la
mortalité constatée.

29
Figure 11 : ‘Algues encroûtantes calcaires’ (récipient # 11 – plant # 3 – pH : 7,8)
Dissolution en cours, changement chromatique ou mortalité constatée ?
Une décroissance dans la couverture en Biofilm (qui avait due se constituer lors des semaines de
culture en aquarium, conditions connues de tout aquariologiste) et pas de nette augmentation de
la couverture en ‘algues filamenteuses’ ont été constatées mais n’entrent pas en rapport avec les
observations en milieu naturel (biofilm épais et algues filamenteuses présentes en quantité selon
les relevés de Donnarumma et al., 2014). Ceci peut être dû à un biais de méthode. Le biofilm
présent est un agglomérat d’individus microscopiques qui a été considéré comme étant une seule
entité pour l’étude de recouvrement. Selon l’échelle considérée, leur observation est plus ou
moins aisée, dépendant du traitement des plants (séchage avant passage au ZooScan) car ce
biofilm est peu fixé. Le biofilm a de plus changé de consistance selon les observations faites.
Pour le pH : 8,1 le biofilm était aisément retiré, alors qu’il semblait être collant à pH : 7,8. Ces
deux types de biofilm observés doivent traduire un changement de population microscopique à
conditions de pH diminué.
Par contre, les couvertures en hydrozoaires non identifiés (Hydroids) semblent potentiellement
augmenter avec la diminution de pH. Concernant Electra posidoniae, les couvertures à pH : 8,1
et pH : 7,8 sont similaires, mais l’augmentation est effective à pH : 7,4. Les algues encroûtantes
étant très rares à pH : 7,4beaucoup du recouvrement a semblé être dû à Electra posidoniae et aux
Spongiaires, seuls observables sans attention particulière.
Les études précédentes sur les communautés hétérogènes des épibiontes (Van der Ben, 1971 ;
Romero, 1988 ; Bouzama, 1995) ont montré une évolution saisonnière importante, e.g. une
augmentation de la biomasse des phéophycées dressées non calcifiées au printemps. Durant notre
période d’étude (hiver, 4 semaines) et hors milieu naturel, les changements relatifs de population

30
observés ne peuvent être dus à un changement saisonnier en cours d’installation et peuvent être
inférés au traitement différentiel de pH.
Cette nette diminution pourrait avoir influence sur les taux de photosynthèse en libérant de
l’espace de captage sur le limbe (Cebrian, 1999) certains auteurs (Hurd et al., 2011 ; Noisette et
al., 2013) ont montré que les réponses physiologiques à l’augmentation de pCO2 seraient faibles
en termes de taux de photosynthèse pour les algues encroûtantes calcaires du fait de leurs
mécanismes de concentration du carbone (carbon concentration mechanisms CCM ; Giordano et
al., 2005) et leur possibilité de résistance à un faible pH du fait de l’abaissement local du pH par
photosynthèse. Ces résultats sont grandement dépendants du sujet d’étude, souvent spécifique.
Les épibiontes non-calcifiés ou à faible calcification n’ont pas présenté ici d’impact négatif et
certains (e.g. foraminifères) semblent montrer une résistance élevée au changement de pH sur
cette période expérimentale. La résistance au changement de pH semble être à relier avec la
composition des organes externes. L’aragonite présente une saturation (Ω aragonite) plus faible
que celle de la calcite. La proportion (différant selon les espèces) et la résistance de ces
composants minéraux à des abaissements de pH peut influer sur leur tenue par les pH considérés.
Des études spécifiques sont à prendre en compte. En effet, certaines ‘algues calcaires’ présentent
une proportion significative de calcite magnésienne (de 10 à 20 % de la masse sèche ; Gazeau,
communication personnelle). A ces concentrations, la calcite magnésienne est encore plus
soluble que l’aragonite, l’impact sur les espèces présentant une proportion de calcite
magnésienne importante pourrait être majeur. Par contre, les foraminifères benthiques sont
souvent formés uniquement de calcite pure et donc sont beaucoup moins sujets à dissolution de
leur exosquelette.
Concernant le pH : 7,8 (qui est celui présentant le plus d’intérêt dans cette étude du fait de sa
prévisibilité dans quelques décennies), la mesure du taux de calcification par l’anomalie
d’alcalinité a permis de mettre en évidence une diminution du taux de calcification par rapport au
pH contrôle (pH : 8,1) après 4 semaines bien que ceci n’était pas statistiquement démontré du
fait d’un faible nombre de réplicats après 2 semaines. De même, la mesure de masse de CaCO3 a
montré une tendance, mais non significative. Par contre, la méthode basée sur les analyses
d’images et les pourcentages de recouvrement (total ou par groupes) semble être plus précise et
montrer un impact majeur des niveaux de pH sur les communautés d’épibiontes (comme
présenté pour les ‘algues calcaires’ par Hall-Spencer et al., 2008).

31
La quantité et la qualité des épibiontes présents sur les feuilles de P. oceanica influe sur les
choix alimentaires des phytophages avec un apport d’azote supplémentaire (Tomas et al., 2005).
Les crustacés, gastéropodes et ‘poissons’ inféodés à l’herbier de posidonies consomment souvent
ces organismes épibiontes (Gacia et al., 2009). Cette production primaire des épibiontes est ainsi
actuellement intégrée dans les réseaux trophiques et la matière organique des herbiers de
posidonies peut de ce fait être rendue disponible aux non phytophages (Alcoverro et al., 1997).
Un changement de pCO2 peut ainsi causer des changements directs par action sur la physiologie9
des organismes, mais aussi par action indirecte du fait des relations alimentaires.
Des changements alimentaires adaptatifs des consommateurs peuvent apparaitre, tout comme des
disparitions du fait de consommations ciblées actuellement devenues impossibles.
Les processus évolutifs de sélection naturelle dans la population prennent un temps certain,
relativement long pour les macroorganismes habituellement. Le changement climatique global et
surtout les processus d’acidification des océans dans les valeurs ici testées (ph : 7,8) sont
supposés intervenir avant la fin du siècle. Les organismes issus d’espèces actuellement adaptés
aux modifications constantes de leur environnement, en faible nombre comparé à la diversité
actuelle, pourraient alorsvoir augmenter leur participation aux cycles biogéochimiques dans les
herbiers de P. oceanica.
9 N’oublions pas que l’échelle de pH est une échelle logarithmique et qu’un changement de 1 unité
équivaut à une forte augmentation de cette valeur dans la concentration en ions [H+]. Ceux-ci peuvent
avoir une action en réactifs chimiques, spécifiques, ou en compétition avec les actifs usuels de certaines réactions, voire même modifier des paramètres physiques du fait de leur concentration.

32
CONCLUSION
La mesure du taux de calcification a mis en évidence une nette diminution des taux de
calcificationdes échantillons à pH : 7,8 allant jusqu’à la décalcification à 7,4. La mesure de
masse de CaCO3 a montré une tendance à la diminution avec la baisse du pH. Les échantillons
maintenus à pH : 7,8 ont présenté des valeurs intermédiaires, traduisant un impact des conditions
de pH prévues pour les prochaines décennies sur le recouvrement en épibiontes des feuilles de P.
oceanica.
Les ‘algues encroûtantes calcaires’ ont présenté à un pH de 7,8 une baisse significative de leur
pourcentage de recouvrement. Leur dissolution a été constatée à un pH de 7,4. Le recouvrement
en foraminifères n’a pas montré de changement significatif aux niveaux de pH étudiés. Par
contre, celui des hydrozoaires a augmenté avec la diminution du pH. Les communautés
d’épibiontes ont montré des différences entre pH : 8,1 et 7,8 avec un total de recouvrement
diminuant selon la baisse du pH, jusqu’à une très faible couverture à pH : 7,4.
La composition des herbiers de P. oceanica pourrait subir des modifications importantes dans le
siècle en cours par impact direct ou indirect de l’acidification anthropique des océans. Les études
menées en laboratoire ne peuvent retranscrire tous les effets possibles du fait du grand nombre
d’interactions possibles. Des observations et des études in situen environnement contrôlé telles
que celles réalisées dans le cadre du projet eFOCE (European Free-Ocean Carbon dioxide
enrichment Experiment ; www.efoce.eu) sont de ce fait déterminantes pour appréhender les
changements futurs.

33
BIBLIOGRAPHIE
Alcoverro T., Duarte C. M., Romero J. (1997) The influence of herbivores on Posidonia oceanica epiphytes. Aquat.
Bot. 56 : 93-104
Andersson A. J., Mackenzie F. T., Gattuso J.-P. (2011) Effects of ocean acidification on benthic processes,
organisms and ecosystems. In Gattuso J.-P., Hansson L., eds, Ocean Acidification. Oxford U. Press : 122-153
Anthony K. R. N., Kline D. J., Diaz-Pulido G., Dove S., Hoegh-Guldberg O. (2008) Ocean acidification causes
bleaching and productivity loss in coral reef builders. Proc. Nat. Acad. Sci. U. S. A. 105, 17442-17446
Boudouresque C. F., Bernard G., Bonhomme P., Charbonnel E., Diviacco G., Meinesz A., Pergent G., Pergent-
Martini C., Ruitton S., Tunesi L. (2012) Protection and conservation of Posidonia oceanica meadows. RAMOGE &
RAC/SPA publisher, Tunis : 1-202
Canadell J. G., Le Quéré C., Raupach M., Field C. B., Buitenhuis E. T., Clais P., Conway T. J., Gillett N. P.,
Houghton R. A., Marland G. (2007) Contributions to accelerating atmospheric CO2 growth from economic activity,
carbon intensity, and efficiency of natural sinks. In PNAS (Proceedings of the National Academy of Sciences) [en
ligne] (consulté le 28 mai 2014)
Cebrian J., Enriquez S., Fortes M., Agawin N., Vermaat J. E., Duarte C. M. (1999) Epiphyte accrual on Posidonia
oceanica Delile leaves Implications for light absorption. Botanica : 123-128
Clarke K. R. (1993) Non-parametric multivariate analyses of changes in community structure. Aust. J. Ecol. 18 :
117-143
Clarke K. R., Gorley R. N. (2006) PRIMER v6 : User Manual/Tutorial. PRIMER-E, Plymouth
Dickson A. G., Sabine C. & Christian J. (2007) Guide to best Practices for oceanic CO2 measurements. PICES
Series Publication 3, IOCCP Report n° 8 : 191 pp.
Doney S. C., Fabry V. J., Feely R. A. & Kleypas J. A. (2009) Ocean acidification : the other CO2 problem. Annual
Review of Marine Science 1, n° 1 : 169-92
Donnarumma L., Lombardi C., Cocito S. & Gambi M. C. (2013) Settlement pattern of Posidonia oceanica epibionts
along a gradient of ocean acidification : an approach with mimics. Med. Mar. Sci.[en ligne] (consulté le 20 mai
2014)
Falkowski P. G, Raven J. A. (1997) Aquatic photosynthesis. Malden, Mass : Blackwell Science : 375 pp.
Gorsky G., Ohman M. D., Picheral M., Gasparini S., Stemmann L., Romagnan J. B., Cawood A., Pesant S., Garcia-
Comas C. & Prejger F. (2010) Digital zooplankton image analysis using the ZooScan integrated system. Journal of
Plankton Research, vol. 32, n° 3 : 285-303
Giordano M., Beardall J., Raven J. A. (2005) CO2 concentrating mechanisms in algae : mechanisms, environmental
modulation, and evolution. Annu. Rev. Plant Biol. 56 : 99-131
Hall-Spencer J. M., Rodolfo-Metalpa R., Martin S., Ransome E., Fine M., Turner S. M., Rowley S. J., Tedesco D.,
Buia M. (2008) Volcanic carbon dioxide vents show ecosystem effects of ocean acidification. Nature 454 : 96-99
Hurd C. L., Cornwall C. E., Currie K., Hepburn C. D., McGraw C. M., Hunter K. A. & Boyd P. W. (2011)
Metabolically induced pH fluctuations by some coastal calcifiers exceed projected 22nd
century ocean acidification :
a mechanism for differential susceptibility? Global Change Biology 17 : 3254-3262
IPCC (2013) Working Group I Contribution to the IPCC fifth assessment report (AR5) Climate change 2013 : the
physical science basis. Intergovernmental Panel on Climate Change, Genève, Suisse

34
Jacquemart J. & Demoulin V. (2006) Inventaire des macroalgues épiphytes des feuilles de Posidonia oceanica (L.)
Delile dans la baie de La Revellata (Calvi, Corse), Lejeunia, Revue de Botanique, n° 181[en ligne] (consulté le 20
mai 2014)
Khatiwala S., Tanhua T., Mikaloff Fletcher S., Gerber M., Doney S. C., Graven H. D., Gruber N. et al. (2013)
Global ocean storage of anthropogenic carbon. Biogeosciences 10, no. 4 : 2169–91
Kroeker K. J., Kordas R. L., Crim R., Hendriks I. E., Ramajo L., Singh G. S., Duarte C. M. & Gattuso J.-P. (2013)
Impacts of ocean acidification on marine organisms : quantifying sensitivities and interaction with warming. Global
Change Biology vol. 19, n°6 : 1884-1896
Lavigne H., Epitalon J.-M. & Gattuso J.-P. (2011) Seacarb : seawater carbonate chemistry with R. R package
version 3.0. (consulté le 20 mai 2014)
Martin S., Rodolfo-Metalpa R., Ransome E., Rowley S., Buia M. C., Gattuso J.-P. & Hall-Spencer J. (2008) Effects
of naturally acidified seawater on seagrass calcareous epibionts. Biology Letters 4, n° 6 : 689-92
Martinez-Crego B., Prado P., Alco verro T., Romero J. (2010) Composition of epiphytic leaf community of
Posidonia oceanica as a tool for environmental biomonitoring. Estuarine, Coastal and Shelf Science 88-2 : 199-208
NOAA Trends in atmospheric carbon dioxide[en ligne] (consulté le 20 mai 2014)
Noisette F., Egilsdöttir H., Davoult D., Martin S. (2013) Physiological responses of three temperate coralline algae
from contrasting habitats to near-future ocean acidification. Journal of Experimental Marine Biology and Ecology,
448 : 179-187
Pergent G. et al. (2012) Les herbiers de Magnoliophytes marines de Méditerranée : résilience et contribution à
l’atténuation des changements climatiques. G land, Suisse et Malaga, Espagne. IUCN : 80 pp.
Rasband W. S. (1997-2014) ImageJ. U. S. National Institutes of Health, Bethesda, Maryland, USA (consulté le 20
mai 2014)
Riebesell U., Tortell P. D. (2011) Effects of ocean acidification on pelagic organisms and ecosystems. In Gattuso J.-
P., Hansson L., eds, Ocean Acidification. Oxford U. Press : 99-116
Romero J. (1988) Biomass seasonal variation of leaf epiphytes of Posidonia oceanica along a depth gradient in
Medes Islands (Girona). Oecol. Aquat.9 : 19-25
Sabine C. L. (2004) The Oceanic Sink for Anthropogenic CO2. Science 305, n° 5682 : 367-71
Smith S. V., Key G. S. (1975) Carbon dioxide and metabolism in marine environments. Limnol. Oceanogr. 20(3) :
493-495
Tomas F., Álvarez-Cascos D., Turon X., Romero J. (2006) Differential element assimilation by sea urchins
Paracentrotus lividus in seagrass beds : implications for trophic interactions. Marine Ecology, Progress Series 306 :
125-131
Tomas F., Turon X., Romero J. (2005) Effects of herbivores on a Posidonia oceanica seagrass meadow : importance
of epiphytes. Marine Ecology, Progress Series 287 : 115-125
Tortell P. D., Reinfelder J. R. and Morel F. M. M. (1997) Active uptake of bicarbonate by diatoms. Nature 390 :
243-244
Van der Ben D. (1971) Les épiphytes des feuilles de Posidonia oceanica Delile sur les côtes françaises de la
Méditerranée. Institut royal des sciences naturelles de Belgique, Mémoire n° 168 : 100-101
Wolf-Gladrow D. A, Zeebe R. E. (2001) CO2 in seawater : equilibrium, kinetics, isotopes. Elsevier Oceanography
Series 65, Amsterdam, Pays-Bas : 346 pp.

Les épibiontes sont des participants prédominants à la mise en circulation de la biomasse passant
par l’écosystème des herbiers de Posidonia oceanica (L.) Delile, qui présente le plus de
biodiversité en Méditerranée. De nombreux épibiontes sont à dominante calcaire et peuvent être
sensibles à l’acidification des océans, en particulier du fait de la baisse de disponibilité des ions
carbonate. Dans l’étude conjointe menée au Laboratoire d’Océanographie de Villefranche sur
mer (Alpes Maritimes, France) les plants de P. oceanica et leurs épibiontes ont été maintenus en
culture à pH régulé (8,1/7,8/7,4) pendant 5 semaines. Les réactions des épibiontes foliaires ont
été quantifiées, par 1) une observation de plants entiers pour la surface de couverture de chaque
groupe biologique, 2) une mesure de masse totale ou calcaire des épibiontes et 3) des mesures de
taux de calcification par la méthode d’anomalie d’alcalinité. Les taux de calcification et les
mesures de recouvrement des feuilles à un pH de 7,8 s’établissent à des valeurs intermédiaires
entre les valeurs de contrôle mesurées à pH 8,1 (actuel) et une dissolution effective des algues
encroûtantes calcaires par pH : 7,4 avec une forte réduction du recouvrement foliaire. Ceci
traduit un impact négatif possible de l’acidification prévue sur les communautés des herbiers de
P. oceanica, grandement dépendants des épibiontes de ces feuilles, et les moyens doivent être
donnés pour éviter d’atteindre le pH de 7,4 aux effets importants sur les communautés liées.
......................................................................................................................................................
The biomass cycling through the seagrass Posidonia oceanica (L.) Delile meadows is often
made available through epibionts. Many of them are calcareous and potentially sensitive to
ocean acidification, especially related to the decrease in availability of carbonate ions.
In this collaborative study conducted at the ‘Laboratoire d’Océanographie de Villefranche sur
mer’ (Alpes Maritimes, France), shoots of P. oceanica and their epibionts were kept alive for 5
weeks at different pH levels (8.1/7.8/7.4). The response of the leaf epibiont community was
quantified by 1) determining the percent cover per group over the entire surface of the shoot, 2)
examining differences in calcareous epibionts mass and 3) measuring the relative change in
calcification rates with the alkalinity anomaly method. Calcification rates at pH of 7.8 are shown
to be intermediate between values measuredat a pH of 8.1 and at pH of 7.4 (beneath the seawater
saturation rate). At a pH of 7.4, the calcareous encrusting algae, the dominant taxa on leaves, are
particularly impacted anddissolve. The predicted change in pH from 8.1 to 7.8 could result in
moderate dissolution and alterations to the epibiont community while a more extreme forecast, a
pH of 7.4 would result in dramatic changes to the epibiont community and thus greater impact on
seagrass meadows. This study helps scientist better predict the possible negative impact of
acidification on P. oceanica meadows, which is known to rely on epibiont community.

Résumé
Abstract




















