
Dermal and pulmonary absorption of propan-1-ol and propan-2-ol from hand rubs
Upload
independentCategory
view
0download
0
The dermal microenvironment induces the expression of thealternative activation marker CD301/mMGL in mononuclearphagocytes, independent of IL-4/IL-13 signaling
Marcel Dupasquier,* Patrizia Stoitzner,† Hui Wan,* Denise Cerqueira,* Adri van Oudenaren,*Jane S. A. Voerman,* Kaori Denda-Nagai,‡ Tatsuro Irimura,‡ Geert Raes,§ Nikolaus Romani,†
and Pieter J. M. Leenen*,1
*Department of Immunology, Erasmus MC, Rotterdam, the Netherlands; †Department of Dermatology, InnsbruckMedical University, Innsbruck, Austria; ‡Laboratory of Cancer Biology and Molecular Immunology, Graduate Schoolof Pharmaceutical Sciences, University of Tokyo, Tokyo, Japan; and §Laboratory of Cellular and MolecularImmunology, Department of Molecular and Cellular Interactions, Vlaams Interuniversitair Instituut voorBiotechnologie, Vrije Universiteit Brussel, Brussels, Belgium
Abstract: Recently, we have shown that mononu-clear phagocytes comprise the majority of inter-stitial cells in the mouse dermis, as indicated bytheir phenotypic and functional characteristics. Inparticular, these cells express the mouse macro-phage galactose-/N-acetylgalactosamine-specificlectin (mMGL)/CD301, identified by the monoclo-nal antibody ER-MP23, as well as other macro-phage markers. As expression of mMGL is inducedby IL-4 or IL-13 and is therefore a marker ofalternatively activated macrophages, we askedwhether dermal mononuclear phagocytes are gen-uinely alternatively activated. We observed thatthese cells expressed, next to mMGL, two otheralternative activation markers, namely, the man-nose receptor/CD206 and Dectin-1. Yet, as thisexpression profile was similar in IL-4 receptor �knockout mice, neither IL-4 nor IL-13 signalingappeared to be required for this phenotype. Wealso found that Langerhans cells (LC), whichshowed only a low level of mMGL in the epidermis,up-regulated mMGL expression upon migrationthrough the dermis, allowing these cells to inter-nalize limited amounts of mMGL ligands. LC iso-lated from epidermal preparations did not showthis up-regulation when cultured in standard me-dium, but whole skin-conditioned medium did stim-ulate mMGL expression by LC. The vast majorityof mMGL molecules was present in the cytoplasm,however. LC, which arrived in skin-draining lymphnodes, quickly down-regulated mMGL expression,and dermally derived cells retained significantmMGL levels. Taken together, these data suggestthat the dermal microenvironment induces mono-nuclear phagocyte subpopulations to expressmMGL and possibly other markers of alternativelyactivated macrophages, independent of IL-4/IL-13signaling. J. Leukoc. Biol. 80: 838–849; 2006.
Key Words: dermis � C-type lectin � Langerhans cells � macrophages
INTRODUCTION
Mononuclear phagocytes are functionally versatile cells, whichperform key roles in the immune system. They are traditionallyseparated into macrophages, the professional phagocytic cells,and dendritic cells (DC), the professional antigen-presentingcells. Nevertheless, it is becoming increasingly clear thatmononuclear phagocytes perform many more functions thanjust taking up pathogens and presenting peptides to T cells [1].They orchestrate inflammatory and immune responses and areintimately involved in maintaining tissue homeostasis by cell-cell interactions and by the production of myriad mediators andeffector molecules. As a consequence, tissue damage is oftenconnected with this function. Conversely, mononuclear phago-cytes also contribute to tissue repair by breaking down gran-ulation tissue and stimulating the production of new extracel-lular matrix (ECM) or by producing such components them-selves [2]. As these functions contradict each other, they needto be controlled tightly. Environmental signals, such as cyto-kines, are now recognized to play pivotal roles in imposingspecific phenotypes and functions in mononuclear phagocytes[3, 4]. Specifically, a dichotomy of M1 and M2 macrophages,modeled after the Th1 and Th2 paradigm of Th cells, wassuggested after the discovery that IL-4 stimulation drives mac-rophages into an alternatively activated state, which is con-nected with immunosuppression and tissue repair [5]. Thisstate contrasts the classically activated state, which is obtainedafter stimulation with IFN-�, and associates with microbicidalactivity and tissue degradation. However, distinguishing justtwo activation states is clearly an oversimplification [3, 4, 6].Exposure of macrophages to IL-10, TGF-�, or corticosteroids,for example, leads to a state that is connected with immuno-suppression, comparable with alternatively activated macro-phages, but those macrophages express phenotypes and func-
1 Correspondence: Department of Immunology, Erasmus MC, Dr. Molewa-terplein 50, NL-3015 GE Rotterdam, The Netherlands. E-mail: [email protected]
Received October 4, 2005; revised May 20, 2006; accepted June 12, 2006;doi: 10.1189/jlb.1005564.
838 Journal of Leukocyte Biology Volume 80, October 2006 0741-5400/06/0080-838 © Society for Leukocyte Biology

tions that differ clearly from IL-4-treated macrophages [7, 8].As a consequence, it has been suggested that only macro-phages stimulated with IL-4 or IL-13, which share a receptorchain and lead to similarly activated cells, should be calledalternatively activated [6].
In vivo, it is difficult to demonstrate the functions of indi-vidual, differently activated macrophage populations. Conse-quently, identification of specific phenotypic markers, whichdistinguish between differentially stimulated macrophages, hasbeen a major goal in the field. The mannose receptor (MR)/CD206 was initially found to serve this purpose for identifica-tion of alternatively activated macrophages [5]. More recently,other cell surface molecules, in particular, the �-glucan recep-tor/Dectin-1 [9] and the mouse macrophage galactose-/N-acetylgalactosamine-specific C-type lectins 1 and 2 (mMGL1and mMGL2)/CD301a and CD301b [10], have been describedas alternative activation markers of macrophages. It is inter-esting that we previously found that mMGL/CD301 expressiontypifies mononuclear phagocytes associated with connectivetissue environments, such as the dermis [11] or the pancreaticsepta [12]. Therefore, we asked whether these cells mightrepresent a population of alternatively activated macrophagesin the tissue. We show in this report that although dermalmononuclear phagocytes do express mMGL/CD301, MR/CD206, and Dectin-1, they are not alternatively activatedmacrophages in the strict sense, as they also express thesereceptors in IL-4 receptor � (IL-4R�) knockout (KO) mice, inwhich cells are unable to respond to IL-4 and IL-13. Moreover,we demonstrate that epidermal Langerhans cells (LC) alsoup-regulate the expression of mMGL in response to dermalfactors. Together, these results suggest that the dermal micro-environment stimulates mononuclear phagocytes to expressmarkers of alternative activation, independent of IL-4/IL-13signaling.
MATERIALS AND METHODS
Animals
Female C57BL/6J mice were obtained from Harlan (Horst, the Netherlands)and used between 10 and 16 weeks of age. IL-4R� KO mice were generatedas described [13] and back-crossed for nine generations onto the C57BL/6background.
Antibodies, conjugates, and other reagents
The following antibodies have been used in this study: rat anti-mMGL1 and-mMGL2, clone ER-MP23, IgG2a, as hybridoma culture supernatant or bio-tinylated, and rat control IgG2a, clone PH2-99, were generated in our lab. Ratanti-mouse Langerin (Clone 929F3, IgG1, purified or Alexa Fluor 488-labeled)was kindly provided by Dr. Sem Saeland (INSERM U 503/IFR, Lyon, France);purified rat anti-MR (Clone MR5D3, IgG2a) and rat anti-Dectin-1 (Clone2A11, IgG2b) were from Hycult Biotechnology (Uden, the Netherlands); andphycoerythrin (PE)-labeled rat anti-I-A/I-E (Clone M5/114.15.2, IgG2b), PE-labeled rat anti-CD86 (Clone GL1, IgG2a), allophycocyanin (APC)-labeled ratanti-CD11b (Clone M1/70, IgG2b), and APC-labeled streptavidin (SA) werefrom BD PharMingen (San Diego, CA). FITC- and HRP-labeled rabbit anti-ratIgs were from Dako (Glostrup, Denmark), and Texas Red-labeled SA andPE-labeled goat anti-rat Igs were from Caltag (San Francisco, CA). Culturemedium was RPMI 1640 (Cambrex, Verviers, Belgium), supplemented with10% FCS (HyClone, Kansas City, MO), 100 U/ml penicillin, and 100 �g/mlstreptomycin (both from Cambrex). FITC was from Sigma Chemical Co. (St.
Louis, MO), whereas biotinylated �-N-acetylgalactosaminide (�-GalNAc),�-GalNAc, �-N-acetylglucosaminide (�-GlcNAc), and LewisX (LeX)-conju-gated polyacrylamide carriers were obtained from GlycoTech (Rockville, MD).
Solid-phase binding assay
Adsorption of the purified, soluble recombinant mMGL1 [14] or mMGL2 [15]onto ELISA plates (655061, Greiner, Kremsmunster Austria) was carried outby adding 100 �l protein solution [2.5 �g/ml in Dulbecco’s PBS (DPBS)] toeach well and incubating the plates for 18 h at 4°C. After blocking nonspecificbinding using 3% BSA in DPBS for 2 h at room temperature, a dilution seriesof ER-MP23 hybridoma culture supernatant or purified control antibodies(stock concentration: 10 �g/ml) in DPBS plus 1% BSA was added. Afterincubation for 2 h at room temperature, wells were washed three times withDPBS to remove unbound materials, and then 100 �l HRP-conjugated goatanti-rat IgG solution (0.75 �g/ml in DPBS) was added to detect boundmaterials. After incubation for 1 h at room temperature, the wells were washedthree times with DPBS. Subsequently, 100 �l 1 mM 2,2�-amino-bis(3-ethyl-benzthiazoline-6-sulfonic acid; ABTS) solution containing 0.34% H2O2 in 0.1M sodium citrate buffer (pH 4.3) was added, and the absorbance was measuredat 405 nm on a microplate reader (Model 550, Bio-Rad, Hercules, CA).
To detect blocking ability of the ER-MP23 antibody, 300 ng recombinantmMGL1 or mMGL2 was adsorbed per well onto plates. Inhibition of nonspe-cific binding and subsequent washing was done as described before. Thereaf-ter, increasing concentrations of ER-MP23 or a rat control antibody wereadded to the wells and incubated for 2 h at room temperature. Subsequently,biotinylated LeX polymers (for mMGL1) or �-GalNAc polymers (for mMGL2)were added to the wells and incubated for 1.5 h at 4°C. Bound polymers weredetected by incubation with 100 �l HRP-conjugated SA solution (1.25 �g/mlin DPBS) for 0.5 h at 4°C and visualized with ABTS as before.
Binding of desialylated erythrocytes tomacrophage cell lines
Macrophage cell lines RAW309Cr.1 (expressing significant levels of mMGL)and RAW264.7 (mMGL–/low), grown in a 96-well culture plate (Nunc, Ros-kilde, Danmark), were preincubated with 50 �g/ml-purified ER-MP23, 200mM galactose, or 200 mM mannose for 15 min at room temperature. Thereafter,human erythrocytes, desialylated by neuraminidase treatment as described[16], were added and incubated for 30 min at 37°C. Subsequently, plates werewashed five times with PBS plus Ca2� and Mg2�, remaining erythrocytes werelysed with 10 mg/ml Na2CO3, and hemoglobin absorbance was measured at414 nm on a Titertek Multiscan (Flow Labs, Redwood City, CA).
Staining of tissue sections
Back-skin or skin-draining lymph nodes were frozen in Tissue-Tek O.C.T.compound (Sakura Finetek, Zoeterwoude, the Netherlands) and cut into 6�m-thick sections on a cryostat (Leitz, Wetzlar, Germany). Subsequently,sections were fixed for 4 min (skin) or 10 s (lymph nodes) in acetone,rehydrated in PBS, pH 7.8, plus 0.05% Tween-20 (Fluka, Buchs, Switzerland),incubated with avidin/biotin blocking kit (Vector Labs, Burlingame, CA),blocked with 10% normal rabbit serum, and incubated with purified antibod-ies. Thereafter, antibody binding was detected with FITC (for Langerin)- orHRP-labeled anti-rat Igs (for mMGL, MR, and Dectin-1). HRP-labeled anti-bodies were visualized by incubating slides with nickel-3,3�-diaminobenzidinetetrahydrochloride (Ni-DAB; Sigma Chemical Co.) plus 1% H2O2 for 3 min.Slides were subsequently embedded in Entellan mounting media (Merck,Darmstadt, Germany). For fluorescence stainings, sections were subsequentlyblocked with 5% normal rat serum and incubated with biotinylated ER-MP23antibodies. After visualization with Texas Red-labeled SA, sections wereembedded with Vectashield mounting media, which contained 4�,6-diamidino-2-phenylindole (DAPI) to counterstain nuclei (Vector Labs). Incubation stepswere all performed in the dark at room temperature for 30 min; sections werewashed twice between incubations with PBS supplemented with 0.05% Tween-20. Thereafter, sections were examined using a Zeiss Axioscop or a ZeissAxioplan 2 fluorescence microscope (Zeiss, Gottingen, Germany).
Epidermal cell and lymph node cell suspensions
Ears of killed mice were cleaned with 70% ethanol and cut off at the base.Thereafter, they were split in dorsal and ventral halves by means of strong
Dupasquier et al. mMGL as dermis-induced mononuclear phagocyte marker 839

forceps; both halves were incubated at 37°C in 0.8% trypsin (Merck) in PBSfor 25 min (cartilage-free half) or 45 min. Epidermis was peeled off and shakenin a water bath for 30 min, and cell suspension was filtered through a 70-�mnylon cell strainer (BD Falcon, Bedford, MA). The resulting epidermal cellsuspensions contained 1–3% LC. Epidermal cells were stained directly for flowcytometry or cultured for 1 day in culture medium, supplemented with 4 ng/mlmurine GM-CSF (BioSource, Camarillo, CA). In some experiments, epidermalcells were cultured for 1 day in conditioned medium obtained from a Day-3skin explant culture (see below), which had also been supplemented withGM-CSF. Cells were stained for ER-MP23-bio, followed by SA-APC andI-A/I-E-PE to detect extracellular mMGL or only for I-A/I-E-PE. Thereafter,cells were fixed with 2% paraformaldehyde (PFA; Merck), permeabilized with0.25% saponin (Sigma Chemical Co.), and subsequently, stained with anti-Langerin-Alexa Fluor 488 antibodies or to detect intracellular mMGL, withanti-Langerin and ER-MP23-bio antibodies, followed by SA-APC.
Axillary, brachial, and inguinal skin-draining lymph nodes were collectedfrom untreated mice or from mice, which were treated with 250 �l 1% FITCin 1:1 acetone:dibutylphtalate (Sigma Chemical Co.) on their shaved back skin24 h before they were killed. Skin-draining lymph nodes were cut into smallpieces and subsequently filtered through a 70-�m nylon cell strainer. Cellswere fixed and permeabilized as described above. Thereafter, they were stainedwith purified anti-Langerin antibodies, which were visualized with PE-labeledanti-rat Igs. Cells were subsequently stained with ER-MP23-bio and SA-APC.Fluorescence of all cells was measured on a FACSCalibur (Becton Dickinson,San Jose, CA) and analyzed using WinMDI 2.8 software.
Skin explant cultures
Dorsal and ventral ear halves were cultured on 1 ml culture medium in 24-wellplates for 1, 2, 3, or 4 days. Cells, which had emigrated out from the explants,were collected afterward and stained for flow cytometry as described forepidermal cell suspensions. For analysis of emigrant cells by confocal micros-copy, cells were allowed to adhere for 6 h on Lab-Tek II chamber slides(Nunc). Thereafter, they were fixed and permeabilizied with PFA and saponinand stained for Langerin-AF488 and ER-MP23-bio, followed by SA-TxR, asdescribed for tissue slides. Slides were analyzed with a Zeiss LSM 510 Metaconfocal microscope. For the endocytosis experiments, 10 �g/ml biotinylated�-GalNAc, �-GalNAc, or �-GlcNAc polymers were added to the culturedmedium as indicated during the 2-day culture period. In preliminary experi-ments, we determined that matured LC and mMGL� dermal-derived cellsexpressed similar high levels of CD86 (data not shown). Therefore, collectedemigrated cells were fixed, permeabilized, and stained with anti-Langerin-Alexa Fluor 488, CD86-PE, and SA-APC to detect the endocytosed sugarpolymers. In some wells, 50–100 �g/ml-purified ER-MP23 anti-mMGL anti-bodies were added during the culture period to block binding of the glycosy-lated polymers (no difference in blocking efficiency was observed between thetwo concentrations). For the emigration experiments, 100 �g/ml purifiedER-MP23 antibodies or control rat IgG2a antibodies were added to the mediumduring the 2-day culture period. Thereafter, cells were fixed, permeabilized,and stained with anti-Langerin-Alexa Fluor 488 and CD11b-APC. Cells wereanalyzed by flow cytometry as indicated above.
RESULTS
ER-MP23 binds mMGL1 and mMGL2
The antimononuclear phagocyte mAb ER-MP23 previouslydescribed [12, 17] appears to recognize the mMGL/CD301.This notion is based on the following observations. In prelim-inary experiments, we found that ER-MP23 precipitated a43-kDa antigen from a J774-1.6 macrophage cell line homog-enate. Protein sequencing revealed the nonblocked N terminusMEYENLQNIRIE, which corresponds to the described N ter-minus of the mouse macrophage mMGL1 [18], with the excep-tion of two amino acids (underscored). Recently, mMGL2 hasbeen discovered, which shows 79.0% nucleotide identity and91.5% amino acid identity to mMGL1 [15]. As shown in
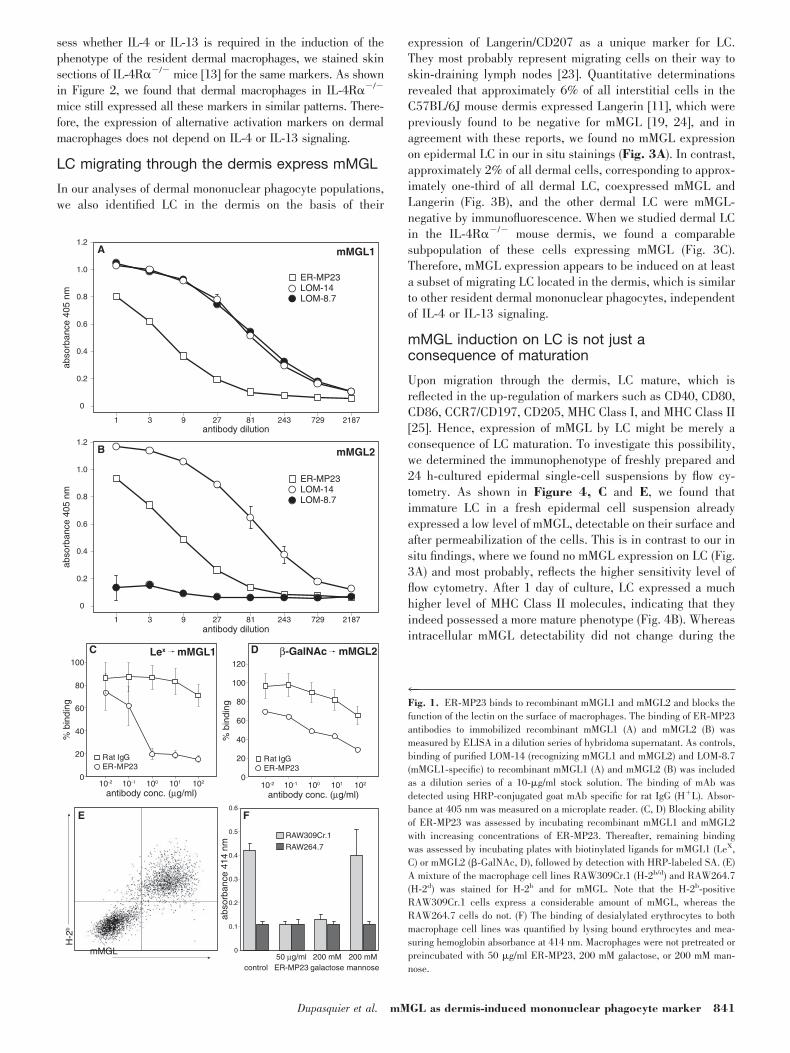
Figure 1, A and B, we found now by ELISA assays withrecombinant mMGL1 and mMGL2 that ER-MP23 bound toextracellular domains of mMGL1 and mMGL2. We confirmedthis finding by positive flow cytometry stainings of Chinesehamster ovary cell clones, which had been transfected withfull-length mMGL1 or mMGL2 (data not shown). To investigatewhether ER-MP23 interferes with the carbohydrate ligand-binding function of the lectins, we incubated immobilized,recombinant mMGL1 and mMGL2 with increasing concentra-tions of ER-MP23. Thereafter, we assessed the remainingsugar-binding capacity of the two recombinant lectins. Asshown in Figure 1, C and D, ER-MP23 could indeed inhibit thebinding of ligands to mMGL1 and mMGL2, although the inhi-bition of mMGL1 ligand binding is possibly more efficient. Toconfirm that this blocking also occurs at the cellular level, weincubated two macrophage cell lines, RAW309Cr.1 andRAW264.7, being high and low mMGL-expressing cell lines,respectively (Fig. 1E), with desialylated erythrocytes. Desialy-lation of erythrocytes exposes nonreduced terminal galactoseresidues at N-linked sugar chains of erythrocytes, therebyforming high-affinity ligands for the mMGL lectins [15]. Asshown in Figure 1F, erythrocyte binding could indeed beobserved for the RAW309Cr.1 but not for the RAW264.7 cells.This binding could be inhibited by preincubating the macro-phages with galactose but not with mannose. Moreover, prein-cubation with ER-MP23 also inhibited erythrocyte binding toRAW309Cr.1 cells, thus confirming that ER-MP23 blocks thebinding of carbohydrate ligands to the mMGL lectins. Proba-bly, ER-MP23 recognizes an epitope inside the carbohydraterecognition domains of mMGL1 and mMGL2.
Dermal mononuclear phagocytes expressmarkers of alternatively activated macrophages
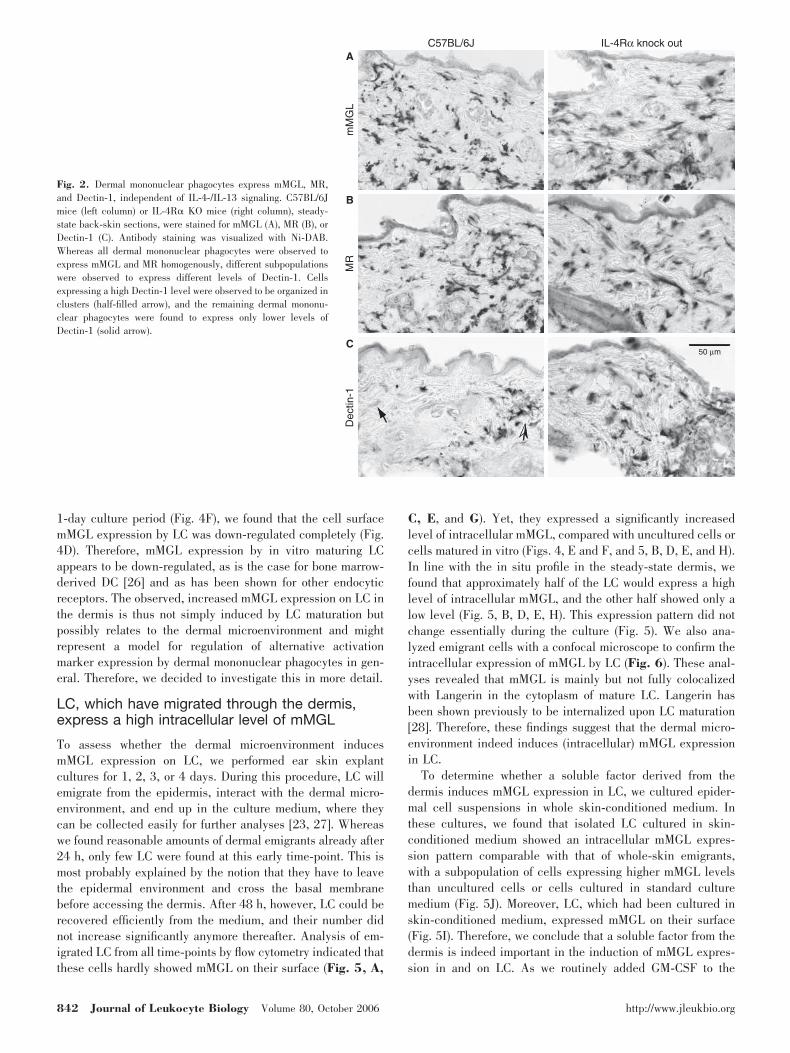
Previously, we characterized antigen-presenting cell popula-tions in the mouse dermis and observed that the majority of alldermal cells represents mononuclear phagocytes expressingmMGL [11, 15, 19]. As the mMGL lectins have been identifiedas markers for alternatively activated macrophages [10], weapproached the question of whether resident dermal mononu-clear phagocytes might represent alternatively activated mac-rophages. To that end, we stained skin sections for two otherwell-established markers of alternatively activated macro-phages, the MR/CD206 [5, 20] and the �-glucan receptor/Dectin-1 [9]. As shown in Figure 2, we found that a similarlylarge population of dermal cells also expressed MR (Fig. 2B).Linehan [21] identified these cells recently as mononuclearphagocytes. Dectin-1 staining showed a less uniform profile,indicating that this marker was expressed at a high level onlyby clusters of cells in the dermis (Fig. 2C), reminiscent of theexpression of MHC Class II in the ear dermis [11]. Neverthe-less, all cells were observed to express at least a low level ofDectin-1. In the ear dermis, we found that MHC Class II andhigh Dectin-1 expression were indeed colocalized on the samecells (data not shown). Therefore, we conclude that dermalmononuclear phagocytes indeed express multiple markers ofalternatively activated macrophages, a phenotype that can beinduced by IL-4 or IL-13 signaling [6]. Macrophages that lackthe IL-4/IL-13 common receptor � chain (IL-4R�) cannotdevelop into alternatively activated macrophages [22]. To as-
840 Journal of Leukocyte Biology Volume 80, October 2006 http://www.jleukbio.org
sess whether IL-4 or IL-13 is required in the induction of thephenotype of the resident dermal macrophages, we stained skinsections of IL-4R��/� mice [13] for the same markers. As shownin Figure 2, we found that dermal macrophages in IL-4R��/�
mice still expressed all these markers in similar patterns. There-fore, the expression of alternative activation markers on dermalmacrophages does not depend on IL-4 or IL-13 signaling.
LC migrating through the dermis express mMGL
In our analyses of dermal mononuclear phagocyte populations,we also identified LC in the dermis on the basis of their
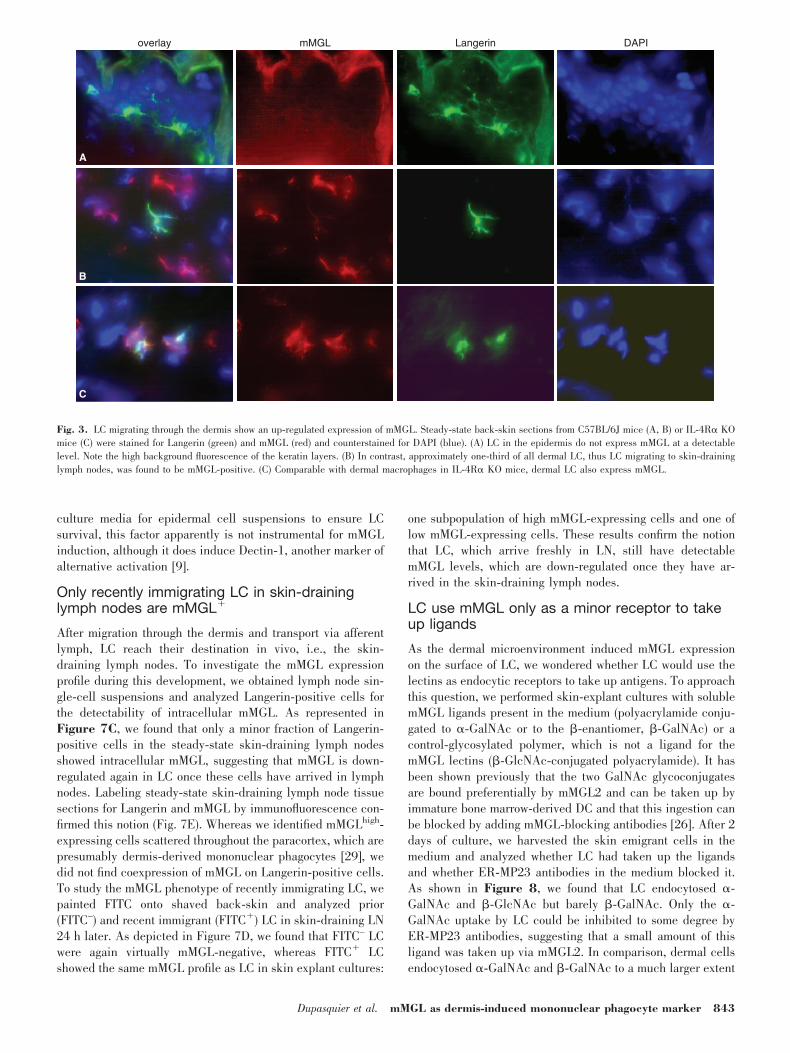
expression of Langerin/CD207 as a unique marker for LC.They most probably represent migrating cells on their way toskin-draining lymph nodes [23]. Quantitative determinationsrevealed that approximately 6% of all interstitial cells in theC57BL/6J mouse dermis expressed Langerin [11], which werepreviously found to be negative for mMGL [19, 24], and inagreement with these reports, we found no mMGL expressionon epidermal LC in our in situ stainings (Fig. 3A). In contrast,approximately 2% of all dermal cells, corresponding to approx-imately one-third of all dermal LC, coexpressed mMGL andLangerin (Fig. 3B), and the other dermal LC were mMGL-negative by immunofluorescence. When we studied dermal LCin the IL-4R��/� mouse dermis, we found a comparablesubpopulation of these cells expressing mMGL (Fig. 3C).Therefore, mMGL expression appears to be induced on at leasta subset of migrating LC located in the dermis, which is similarto other resident dermal mononuclear phagocytes, independentof IL-4 or IL-13 signaling.
mMGL induction on LC is not just aconsequence of maturation
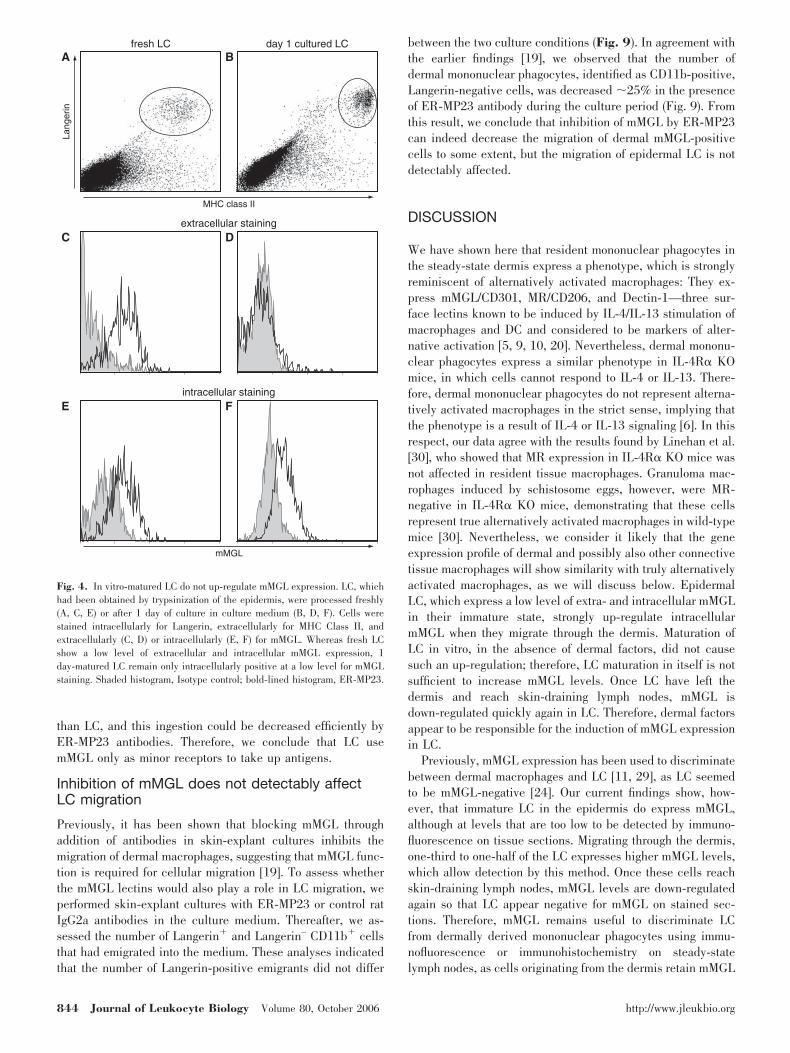
Upon migration through the dermis, LC mature, which isreflected in the up-regulation of markers such as CD40, CD80,CD86, CCR7/CD197, CD205, MHC Class I, and MHC Class II[25]. Hence, expression of mMGL by LC might be merely aconsequence of LC maturation. To investigate this possibility,we determined the immunophenotype of freshly prepared and24 h-cultured epidermal single-cell suspensions by flow cy-tometry. As shown in Figure 4, C and E, we found thatimmature LC in a fresh epidermal cell suspension alreadyexpressed a low level of mMGL, detectable on their surface andafter permeabilization of the cells. This is in contrast to our insitu findings, where we found no mMGL expression on LC (Fig.3A) and most probably, reflects the higher sensitivity level offlow cytometry. After 1 day of culture, LC expressed a muchhigher level of MHC Class II molecules, indicating that theyindeed possessed a more mature phenotype (Fig. 4B). Whereasintracellular mMGL detectability did not change during the
control ER-MP23 galactose mannose50 µg/ml
RAW309Cr.1
RAW264.7
200 mM 200 mM0
0.1
0.2
0.3
0.4
0.5
0.6
abso
rban
ce 4
14 n
m
E F
C D
0
0.2
0.4
0.6
0.8
1.0
1.2
abso
rban
ce 4
05 n
mH
-2b
mMGL
antibody dilution
B mMGL2
1 3 9 27 81 243 729 2187
1 3 9 27 81 243 729 2187
0
0.2
0.4
0.6
0.8
1.0
1.2
abso
rban
ce 4
05 n
m
antibody dilution
ER-MP23
A mMGL1
LOM-14 LOM-8.7
ER-MP23 LOM-14 LOM-8.7
Lex mMGL1
0
20
40
60
80
100
antibody conc. (µg/ml) antibody conc. (µg/ml)
% b
indi
ng
Rat IgGER-MP23
Rat IgGER-MP23
10210110010-110-2
β-GalNAc mMGL2
0
20
40
60
80
100
120
% b
indi
ng
10210110010-110-2
4Fig. 1. ER-MP23 binds to recombinant mMGL1 and mMGL2 and blocks thefunction of the lectin on the surface of macrophages. The binding of ER-MP23antibodies to immobilized recombinant mMGL1 (A) and mMGL2 (B) wasmeasured by ELISA in a dilution series of hybridoma supernatant. As controls,binding of purified LOM-14 (recognizing mMGL1 and mMGL2) and LOM-8.7(mMGL1-specific) to recombinant mMGL1 (A) and mMGL2 (B) was includedas a dilution series of a 10-�g/ml stock solution. The binding of mAb wasdetected using HRP-conjugated goat mAb specific for rat IgG (H�L). Absor-bance at 405 nm was measured on a microplate reader. (C, D) Blocking abilityof ER-MP23 was assessed by incubating recombinant mMGL1 and mMGL2with increasing concentrations of ER-MP23. Thereafter, remaining bindingwas assessed by incubating plates with biotinylated ligands for mMGL1 (LeX,C) or mMGL2 (�-GalNAc, D), followed by detection with HRP-labeled SA. (E)A mixture of the macrophage cell lines RAW309Cr.1 (H-2b/d) and RAW264.7(H-2d) was stained for H-2b and for mMGL. Note that the H-2b-positiveRAW309Cr.1 cells express a considerable amount of mMGL, whereas theRAW264.7 cells do not. (F) The binding of desialylated erythrocytes to bothmacrophage cell lines was quantified by lysing bound erythrocytes and mea-suring hemoglobin absorbance at 414 nm. Macrophages were not pretreated orpreincubated with 50 �g/ml ER-MP23, 200 mM galactose, or 200 mM man-nose.
Dupasquier et al. mMGL as dermis-induced mononuclear phagocyte marker 841
1-day culture period (Fig. 4F), we found that the cell surfacemMGL expression by LC was down-regulated completely (Fig.4D). Therefore, mMGL expression by in vitro maturing LCappears to be down-regulated, as is the case for bone marrow-derived DC [26] and as has been shown for other endocyticreceptors. The observed, increased mMGL expression on LC inthe dermis is thus not simply induced by LC maturation butpossibly relates to the dermal microenvironment and mightrepresent a model for regulation of alternative activationmarker expression by dermal mononuclear phagocytes in gen-eral. Therefore, we decided to investigate this in more detail.
LC, which have migrated through the dermis,express a high intracellular level of mMGL
To assess whether the dermal microenvironment inducesmMGL expression on LC, we performed ear skin explantcultures for 1, 2, 3, or 4 days. During this procedure, LC willemigrate from the epidermis, interact with the dermal micro-environment, and end up in the culture medium, where theycan be collected easily for further analyses [23, 27]. Whereaswe found reasonable amounts of dermal emigrants already after24 h, only few LC were found at this early time-point. This ismost probably explained by the notion that they have to leavethe epidermal environment and cross the basal membranebefore accessing the dermis. After 48 h, however, LC could berecovered efficiently from the medium, and their number didnot increase significantly anymore thereafter. Analysis of em-igrated LC from all time-points by flow cytometry indicated thatthese cells hardly showed mMGL on their surface (Fig. 5, A,
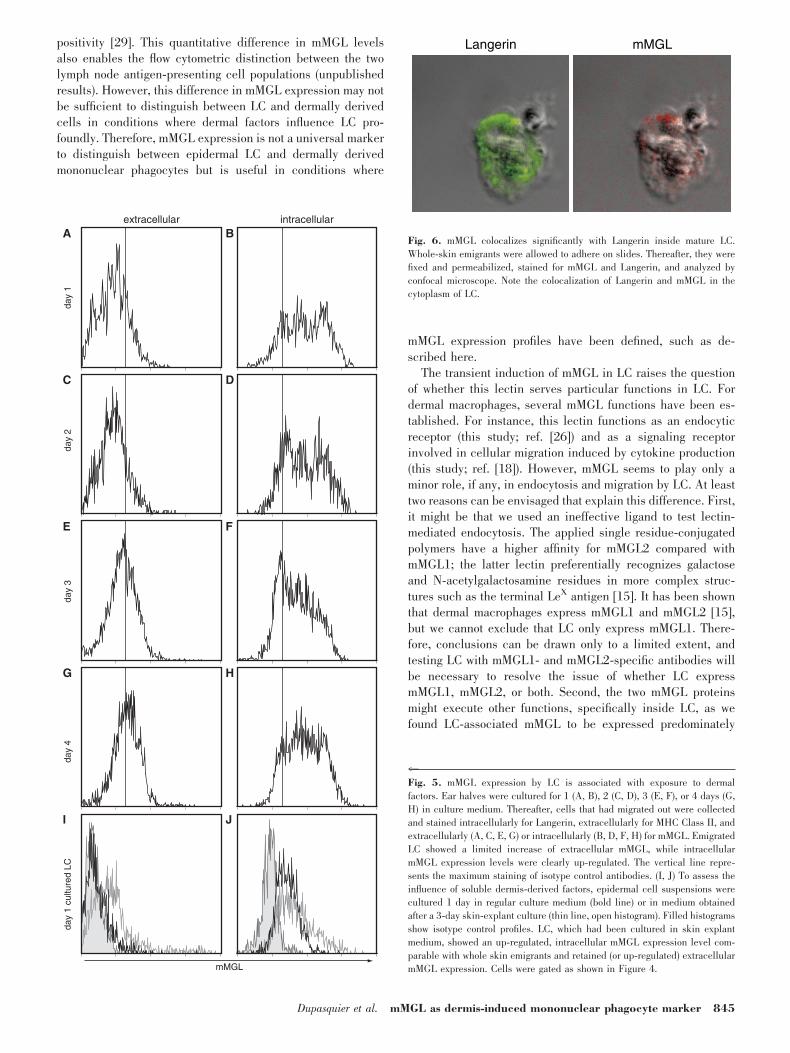
C, E, and G). Yet, they expressed a significantly increasedlevel of intracellular mMGL, compared with uncultured cells orcells matured in vitro (Figs. 4, E and F, and 5, B, D, E, and H).In line with the in situ profile in the steady-state dermis, wefound that approximately half of the LC would express a highlevel of intracellular mMGL, and the other half showed only alow level (Fig. 5, B, D, E, H). This expression pattern did notchange essentially during the culture (Fig. 5). We also ana-lyzed emigrant cells with a confocal microscope to confirm theintracellular expression of mMGL by LC (Fig. 6). These anal-yses revealed that mMGL is mainly but not fully colocalizedwith Langerin in the cytoplasm of mature LC. Langerin hasbeen shown previously to be internalized upon LC maturation[28]. Therefore, these findings suggest that the dermal micro-environment indeed induces (intracellular) mMGL expressionin LC.
To determine whether a soluble factor derived from thedermis induces mMGL expression in LC, we cultured epider-mal cell suspensions in whole skin-conditioned medium. Inthese cultures, we found that isolated LC cultured in skin-conditioned medium showed an intracellular mMGL expres-sion pattern comparable with that of whole-skin emigrants,with a subpopulation of cells expressing higher mMGL levelsthan uncultured cells or cells cultured in standard culturemedium (Fig. 5J). Moreover, LC, which had been cultured inskin-conditioned medium, expressed mMGL on their surface(Fig. 5I). Therefore, we conclude that a soluble factor from thedermis is indeed important in the induction of mMGL expres-sion in and on LC. As we routinely added GM-CSF to the
A
B
Cm
MG
LM
RD
ectin
-1
IL-4Rα knock outC57BL/6J
50 µm
Fig. 2. Dermal mononuclear phagocytes express mMGL, MR,and Dectin-1, independent of IL-4-/IL-13 signaling. C57BL/6Jmice (left column) or IL-4R� KO mice (right column), steady-state back-skin sections, were stained for mMGL (A), MR (B), orDectin-1 (C). Antibody staining was visualized with Ni-DAB.Whereas all dermal mononuclear phagocytes were observed toexpress mMGL and MR homogenously, different subpopulationswere observed to express different levels of Dectin-1. Cellsexpressing a high Dectin-1 level were observed to be organized inclusters (half-filled arrow), and the remaining dermal mononu-clear phagocytes were found to express only lower levels ofDectin-1 (solid arrow).
842 Journal of Leukocyte Biology Volume 80, October 2006 http://www.jleukbio.org
culture media for epidermal cell suspensions to ensure LCsurvival, this factor apparently is not instrumental for mMGLinduction, although it does induce Dectin-1, another marker ofalternative activation [9].
Only recently immigrating LC in skin-draininglymph nodes are mMGL�
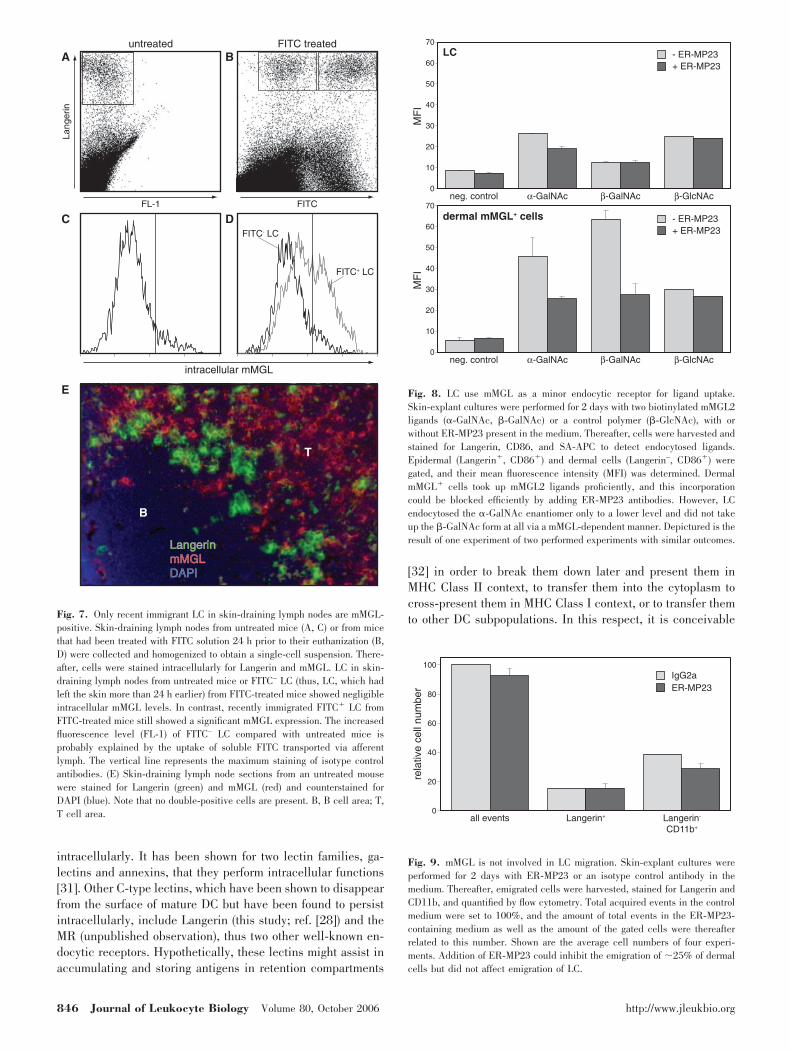
After migration through the dermis and transport via afferentlymph, LC reach their destination in vivo, i.e., the skin-draining lymph nodes. To investigate the mMGL expressionprofile during this development, we obtained lymph node sin-gle-cell suspensions and analyzed Langerin-positive cells forthe detectability of intracellular mMGL. As represented inFigure 7C, we found that only a minor fraction of Langerin-positive cells in the steady-state skin-draining lymph nodesshowed intracellular mMGL, suggesting that mMGL is down-regulated again in LC once these cells have arrived in lymphnodes. Labeling steady-state skin-draining lymph node tissuesections for Langerin and mMGL by immunofluorescence con-firmed this notion (Fig. 7E). Whereas we identified mMGLhigh-expressing cells scattered throughout the paracortex, which arepresumably dermis-derived mononuclear phagocytes [29], wedid not find coexpression of mMGL on Langerin-positive cells.To study the mMGL phenotype of recently immigrating LC, wepainted FITC onto shaved back-skin and analyzed prior(FITC–) and recent immigrant (FITC�) LC in skin-draining LN24 h later. As depicted in Figure 7D, we found that FITC– LCwere again virtually mMGL-negative, whereas FITC� LCshowed the same mMGL profile as LC in skin explant cultures:
one subpopulation of high mMGL-expressing cells and one oflow mMGL-expressing cells. These results confirm the notionthat LC, which arrive freshly in LN, still have detectablemMGL levels, which are down-regulated once they have ar-rived in the skin-draining lymph nodes.
LC use mMGL only as a minor receptor to takeup ligands
As the dermal microenvironment induced mMGL expressionon the surface of LC, we wondered whether LC would use thelectins as endocytic receptors to take up antigens. To approachthis question, we performed skin-explant cultures with solublemMGL ligands present in the medium (polyacrylamide conju-gated to �-GalNAc or to the �-enantiomer, �-GalNAc) or acontrol-glycosylated polymer, which is not a ligand for themMGL lectins (�-GlcNAc-conjugated polyacrylamide). It hasbeen shown previously that the two GalNAc glycoconjugatesare bound preferentially by mMGL2 and can be taken up byimmature bone marrow-derived DC and that this ingestion canbe blocked by adding mMGL-blocking antibodies [26]. After 2days of culture, we harvested the skin emigrant cells in themedium and analyzed whether LC had taken up the ligandsand whether ER-MP23 antibodies in the medium blocked it.As shown in Figure 8, we found that LC endocytosed �-GalNAc and �-GlcNAc but barely �-GalNAc. Only the �-GalNAc uptake by LC could be inhibited to some degree byER-MP23 antibodies, suggesting that a small amount of thisligand was taken up via mMGL2. In comparison, dermal cellsendocytosed �-GalNAc and �-GalNAc to a much larger extent
overlay mMGL Langerin DAPI
A
B
C
Fig. 3. LC migrating through the dermis show an up-regulated expression of mMGL. Steady-state back-skin sections from C57BL/6J mice (A, B) or IL-4R� KOmice (C) were stained for Langerin (green) and mMGL (red) and counterstained for DAPI (blue). (A) LC in the epidermis do not express mMGL at a detectablelevel. Note the high background fluorescence of the keratin layers. (B) In contrast, approximately one-third of all dermal LC, thus LC migrating to skin-draininglymph nodes, was found to be mMGL-positive. (C) Comparable with dermal macrophages in IL-4R� KO mice, dermal LC also express mMGL.
Dupasquier et al. mMGL as dermis-induced mononuclear phagocyte marker 843
than LC, and this ingestion could be decreased efficiently byER-MP23 antibodies. Therefore, we conclude that LC usemMGL only as minor receptors to take up antigens.
Inhibition of mMGL does not detectably affectLC migration
Previously, it has been shown that blocking mMGL throughaddition of antibodies in skin-explant cultures inhibits themigration of dermal macrophages, suggesting that mMGL func-tion is required for cellular migration [19]. To assess whetherthe mMGL lectins would also play a role in LC migration, weperformed skin-explant cultures with ER-MP23 or control ratIgG2a antibodies in the culture medium. Thereafter, we as-sessed the number of Langerin� and Langerin– CD11b� cellsthat had emigrated into the medium. These analyses indicatedthat the number of Langerin-positive emigrants did not differ
between the two culture conditions (Fig. 9). In agreement withthe earlier findings [19], we observed that the number ofdermal mononuclear phagocytes, identified as CD11b-positive,Langerin-negative cells, was decreased �25% in the presenceof ER-MP23 antibody during the culture period (Fig. 9). Fromthis result, we conclude that inhibition of mMGL by ER-MP23can indeed decrease the migration of dermal mMGL-positivecells to some extent, but the migration of epidermal LC is notdetectably affected.
DISCUSSION
We have shown here that resident mononuclear phagocytes inthe steady-state dermis express a phenotype, which is stronglyreminiscent of alternatively activated macrophages: They ex-press mMGL/CD301, MR/CD206, and Dectin-1—three sur-face lectins known to be induced by IL-4/IL-13 stimulation ofmacrophages and DC and considered to be markers of alter-native activation [5, 9, 10, 20]. Nevertheless, dermal mononu-clear phagocytes express a similar phenotype in IL-4R� KOmice, in which cells cannot respond to IL-4 or IL-13. There-fore, dermal mononuclear phagocytes do not represent alterna-tively activated macrophages in the strict sense, implying thatthe phenotype is a result of IL-4 or IL-13 signaling [6]. In thisrespect, our data agree with the results found by Linehan et al.[30], who showed that MR expression in IL-4R� KO mice wasnot affected in resident tissue macrophages. Granuloma mac-rophages induced by schistosome eggs, however, were MR-negative in IL-4R� KO mice, demonstrating that these cellsrepresent true alternatively activated macrophages in wild-typemice [30]. Nevertheless, we consider it likely that the geneexpression profile of dermal and possibly also other connectivetissue macrophages will show similarity with truly alternativelyactivated macrophages, as we will discuss below. EpidermalLC, which express a low level of extra- and intracellular mMGLin their immature state, strongly up-regulate intracellularmMGL when they migrate through the dermis. Maturation ofLC in vitro, in the absence of dermal factors, did not causesuch an up-regulation; therefore, LC maturation in itself is notsufficient to increase mMGL levels. Once LC have left thedermis and reach skin-draining lymph nodes, mMGL isdown-regulated quickly again in LC. Therefore, dermal factorsappear to be responsible for the induction of mMGL expressionin LC.
Previously, mMGL expression has been used to discriminatebetween dermal macrophages and LC [11, 29], as LC seemedto be mMGL-negative [24]. Our current findings show, how-ever, that immature LC in the epidermis do express mMGL,although at levels that are too low to be detected by immuno-fluorescence on tissue sections. Migrating through the dermis,one-third to one-half of the LC expresses higher mMGL levels,which allow detection by this method. Once these cells reachskin-draining lymph nodes, mMGL levels are down-regulatedagain so that LC appear negative for mMGL on stained sec-tions. Therefore, mMGL remains useful to discriminate LCfrom dermally derived mononuclear phagocytes using immu-nofluorescence or immunohistochemistry on steady-statelymph nodes, as cells originating from the dermis retain mMGL
Lang
erin
MHC class II
fresh LC
extracellular staining
intracellular staining
day 1 cultured LCA B
C D
mMGL
E F
Fig. 4. In vitro-matured LC do not up-regulate mMGL expression. LC, whichhad been obtained by trypsinization of the epidermis, were processed freshly(A, C, E) or after 1 day of culture in culture medium (B, D, F). Cells werestained intracellularly for Langerin, extracellularly for MHC Class II, andextracellularly (C, D) or intracellularly (E, F) for mMGL. Whereas fresh LCshow a low level of extracellular and intracellular mMGL expression, 1day-matured LC remain only intracellularly positive at a low level for mMGLstaining. Shaded histogram, Isotype control; bold-lined histogram, ER-MP23.
844 Journal of Leukocyte Biology Volume 80, October 2006 http://www.jleukbio.org
positivity [29]. This quantitative difference in mMGL levelsalso enables the flow cytometric distinction between the twolymph node antigen-presenting cell populations (unpublishedresults). However, this difference in mMGL expression may notbe sufficient to distinguish between LC and dermally derivedcells in conditions where dermal factors influence LC pro-foundly. Therefore, mMGL expression is not a universal markerto distinguish between epidermal LC and dermally derivedmononuclear phagocytes but is useful in conditions where
mMGL expression profiles have been defined, such as de-scribed here.
The transient induction of mMGL in LC raises the questionof whether this lectin serves particular functions in LC. Fordermal macrophages, several mMGL functions have been es-tablished. For instance, this lectin functions as an endocyticreceptor (this study; ref. [26]) and as a signaling receptorinvolved in cellular migration induced by cytokine production(this study; ref. [18]). However, mMGL seems to play only aminor role, if any, in endocytosis and migration by LC. At leasttwo reasons can be envisaged that explain this difference. First,it might be that we used an ineffective ligand to test lectin-mediated endocytosis. The applied single residue-conjugatedpolymers have a higher affinity for mMGL2 compared withmMGL1; the latter lectin preferentially recognizes galactoseand N-acetylgalactosamine residues in more complex struc-tures such as the terminal LeX antigen [15]. It has been shownthat dermal macrophages express mMGL1 and mMGL2 [15],but we cannot exclude that LC only express mMGL1. There-fore, conclusions can be drawn only to a limited extent, andtesting LC with mMGL1- and mMGL2-specific antibodies willbe necessary to resolve the issue of whether LC expressmMGL1, mMGL2, or both. Second, the two mMGL proteinsmight execute other functions, specifically inside LC, as wefound LC-associated mMGL to be expressed predominately
extracellular
day
1da
y 2
day
3da
y 4
day
1 cu
lture
d LC
intracellularA B
mMGL
C D
E F
I J
G H
Langerin mMGL
Fig. 6. mMGL colocalizes significantly with Langerin inside mature LC.Whole-skin emigrants were allowed to adhere on slides. Thereafter, they werefixed and permeabilized, stained for mMGL and Langerin, and analyzed byconfocal microscope. Note the colocalization of Langerin and mMGL in thecytoplasm of LC.
4Fig. 5. mMGL expression by LC is associated with exposure to dermalfactors. Ear halves were cultured for 1 (A, B), 2 (C, D), 3 (E, F), or 4 days (G,H) in culture medium. Thereafter, cells that had migrated out were collectedand stained intracellularly for Langerin, extracellularly for MHC Class II, andextracellularly (A, C, E, G) or intracellularly (B, D, F, H) for mMGL. EmigratedLC showed a limited increase of extracellular mMGL, while intracellularmMGL expression levels were clearly up-regulated. The vertical line repre-sents the maximum staining of isotype control antibodies. (I, J) To assess theinfluence of soluble dermis-derived factors, epidermal cell suspensions werecultured 1 day in regular culture medium (bold line) or in medium obtainedafter a 3-day skin-explant culture (thin line, open histogram). Filled histogramsshow isotype control profiles. LC, which had been cultured in skin explantmedium, showed an up-regulated, intracellular mMGL expression level com-parable with whole skin emigrants and retained (or up-regulated) extracellularmMGL expression. Cells were gated as shown in Figure 4.
Dupasquier et al. mMGL as dermis-induced mononuclear phagocyte marker 845
intracellularly. It has been shown for two lectin families, ga-lectins and annexins, that they perform intracellular functions[31]. Other C-type lectins, which have been shown to disappearfrom the surface of mature DC but have been found to persistintracellularly, include Langerin (this study; ref. [28]) and theMR (unpublished observation), thus two other well-known en-docytic receptors. Hypothetically, these lectins might assist inaccumulating and storing antigens in retention compartments
[32] in order to break them down later and present them inMHC Class II context, to transfer them into the cytoplasm tocross-present them in MHC Class I context, or to transfer themto other DC subpopulations. In this respect, it is conceivable
Lang
erin
FL-1 FITC
FITC+ LC
FITC- LC
untreated
intracellular mMGL
FITC treatedA B
B
T
C D
E
LangerinLangerinmMGLmMGLDAPIDAPI
Fig. 7. Only recent immigrant LC in skin-draining lymph nodes are mMGL-positive. Skin-draining lymph nodes from untreated mice (A, C) or from micethat had been treated with FITC solution 24 h prior to their euthanization (B,D) were collected and homogenized to obtain a single-cell suspension. There-after, cells were stained intracellularly for Langerin and mMGL. LC in skin-draining lymph nodes from untreated mice or FITC– LC (thus, LC, which hadleft the skin more than 24 h earlier) from FITC-treated mice showed negligibleintracellular mMGL levels. In contrast, recently immigrated FITC� LC fromFITC-treated mice still showed a significant mMGL expression. The increasedfluorescence level (FL-1) of FITC– LC compared with untreated mice isprobably explained by the uptake of soluble FITC transported via afferentlymph. The vertical line represents the maximum staining of isotype controlantibodies. (E) Skin-draining lymph node sections from an untreated mousewere stained for Langerin (green) and mMGL (red) and counterstained forDAPI (blue). Note that no double-positive cells are present. B, B cell area; T,T cell area.
neg. control
dermal mMGL+ cells
LC - ER-MP23+ ER-MP23
α-GalNAc β-GalNAc β-GlcNAc
neg. control α-GalNAc β-GalNAc β-GlcNAc0
10
20
30
40
50
60
70
MF
I
0
10
20
30
40
50
60
70
MF
I
- ER-MP23+ ER-MP23
Fig. 8. LC use mMGL as a minor endocytic receptor for ligand uptake.Skin-explant cultures were performed for 2 days with two biotinylated mMGL2ligands (�-GalNAc, �-GalNAc) or a control polymer (�-GlcNAc), with orwithout ER-MP23 present in the medium. Thereafter, cells were harvested andstained for Langerin, CD86, and SA-APC to detect endocytosed ligands.Epidermal (Langerin�, CD86�) and dermal cells (Langerin–, CD86�) weregated, and their mean fluorescence intensity (MFI) was determined. DermalmMGL� cells took up mMGL2 ligands proficiently, and this incorporationcould be blocked efficiently by adding ER-MP23 antibodies. However, LCendocytosed the �-GalNAc enantiomer only to a lower level and did not takeup the �-GalNAc form at all via a mMGL-dependent manner. Depictured is theresult of one experiment of two performed experiments with similar outcomes.
all events Langerin+ Langerin-
CD11b+
0
20
40
60
80
100
rela
tive
cell
num
ber
IgG2aER-MP23
Fig. 9. mMGL is not involved in LC migration. Skin-explant cultures wereperformed for 2 days with ER-MP23 or an isotype control antibody in themedium. Thereafter, emigrated cells were harvested, stained for Langerin andCD11b, and quantified by flow cytometry. Total acquired events in the controlmedium were set to 100%, and the amount of total events in the ER-MP23-containing medium as well as the amount of the gated cells were thereafterrelated to this number. Shown are the average cell numbers of four experi-ments. Addition of ER-MP23 could inhibit the emigration of �25% of dermalcells but did not affect emigration of LC.
846 Journal of Leukocyte Biology Volume 80, October 2006 http://www.jleukbio.org

that the mMGL lectins are only expressed briefly on the LCsurface, are quickly internalized again, and are delivered thento retention compartments. Our findings of extracellular mMGLexpression on in vitro-matured LC in skin-conditioned mediumwould fit into this picture. Future determinations of othermMGL functions might shed more light on the functions ofmMGL expression in LC, in particular, and in mononuclearphagocytes in general.
Only recently, mMGL expression was found to be inducedupon alternative activation of macrophages in vitro and in vivoby the Th2-associated inflammatory cytokines IL-4 and/orIL-13 [10]. In the steady-state mouse, however, mMGL isexpressed almost exclusively by connective tissue macro-phages but not by other populations including splenic macro-phages, Kupffer cells, or peripheral blood monocytes (ref. [24];unpublished observations). Hence, also, the connective tissuemicroenvironment appears to provide factors that induce theexpression of mMGL and most probably, other markers onresident mononuclear phagocytes and cells in transit. We haveshown that this dermis-associated mMGL expression by mac-rophages and LC was independent of IL-4 and IL-13, exclud-ing these as factors responsible for imposing an alternative-likeactivation phenotype.
Can inflammatory mediators other than IL-4 and IL-13 beheld responsible for induced mMGL expression in migratingLC? Some of our experimental conditions, in which we foundmMGL in and on LC, are clearly connected with inflammation,such as in vivo skin painting with FITC and adjuvant or in vitroskin explant cultures. However, different arguments pleaagainst a predominant role of inflammatory mediators in dermalmMGL induction in LC. First, we have found that LC in theundisturbed steady-state dermis are also mMGL-positive insitu. Second, obtaining epidermal cell suspensions activateskeratinocytes to produce an array of different mediators, in-cluding proinflammatory factors such as GM-CSF [33]. In invitro studies, it has been shown already that GM-CSF iscapable of inducing the expression of Dectin-1 on macrophages[9]. However, as we generally included GM-CSF in our epi-dermal cell suspension cultures to ensure LC viability andfound no mMGL induction, we can also exclude this cytokine.Only when epidermal cell suspensions were cultured in wholeskin-conditioned medium, up-regulation of mMGL in and onLC was observed. In line with this, we regard the detectablemMGL level in recent immigrant FITC� LC in skin-draininglymph nodes a reflection of the relative immaturity and recentdermal sojourn of inflammatory LC [34] rather than an enhanc-ing effect of inflammatory mediators on mMGL expression.Taken together, connective tissue factors present in the steady-state and not inflammation-related mediators seem to be mostimportant for the induction of mMGL expression in LC.
In skin-draining lymph nodes, mMGL expression can befound on two subpopulations of mononuclear phagocytes. Cellsthat have immigrated from the dermis are present in theparacortical T cell area and express a high level of mMGL (ref.[29] and unpublished observations), and macrophages in sub-capsular and trabecular sinuses have only low levels of mMGL[24]. As these latter cells filter the lymph fluid, it seems likelythat these cells are exposed to the dermal factors and/or theirbreakdown products, which are brought into the lymph nodes
via the afferent lymph flow. Therefore, subcapsular sinus mac-rophages might be induced to express mMGL in a similar wayas LC, which have been cultured in skin-conditioned medium.Resident mononuclear phagocytes in the dermis will be ex-posed constantly to a high concentration of the dermal factors.Although intriguing, the notion that this exposure induces theexpression of mMGL and possibly also other alternative acti-vation markers on dermal resident cells remains to be proven.As the dermis predominately consists of ECM, we tested var-ious ECM components for their ability to induce mMGL inpreliminary in vitro experiments. Nevertheless, culturing bonemarrow-derived DC on collagen type I (ECM component of thedermis), fibronectin (ECM component of the epidermal base-ment membrane and the dermis), or collagen type VII (ECM ofthe epidermal basement membrane) did not induce an in-creased mMGL expression in these cells, whereas IL-4 aspositive control treatment did (unpublished observations).Therefore, the identity of the dermal factors, which induce theexpression of mMGL, and possibly other alternative activationmarkers on mononuclear phagocytes in the steady-state dermisand lymph nodes remains unknown.
Alternatively activated macrophages are often associatedwith tissue remodeling or tissue repair, thus with the break-down and de novo synthesis of ECM components [6, 7]. Despitethe fact that the dermal mononuclear phagocytes are not IL-4/IL-13-activated, a major role of these cells in connective tissuemaintenance is likely. We have shown recently that the ma-jority of dermal interstitial cells are macrophages and notfibroblasts as widely believed. Synthesis of ECM componentsby macrophages is well-documented [2], and as such, thenotion that the majority of interstitial cells synthesizes theECM of the dermis might nevertheless turn out to be true.Alternatively activated macrophages are also associated withtolerance and suppression of immune responses. The dermisand other connective tissues might thus be places where animmunosuppressive microenvironment is supported. Tumor-associated macrophages, which mostly reside in and regulatesupporting connective tissue [35], have been shown consis-tently to express mMGL [36–38]. It is feasible that the ECMproduction stimulated by the tumor generates a microenviron-ment, which activates tumor-infiltrating macrophages along analternative route with related connective tissue, maintaining animmune-suppressive function. Identifying the factors that in-duce macrophages to adopt phenotypes similar to alternativelyactivated macrophages and counteracting these might thus alsobenefit cancer treatments.
To summarize, we have shown that resident dermal mono-nuclear phagocyte subpopulations express several markers ofalternatively activated macrophages, for which IL-4 or IL-13signaling is not required. In addition, LC migrating through thedermis in vivo or cultured in vitro in medium from skin-explantcultures contain higher mMGL levels than immature, epider-mal LC or LC cultured in standard medium. LC, which arrivefreshly in skin-draining lymph nodes, still show intracellularmMGL, whereas LC, which have remained for a prolonged timein the lymph nodes, did not. Therefore, the expression ofmMGL in LC was linked closely to the exposure to the dermalmicroenvironment and factors derived thereof.
Dupasquier et al. mMGL as dermis-induced mononuclear phagocyte marker 847

ACKNOWLEDGMENTS
This study was supported by the J. A. Cohen Institute/Inter-universitary Research Institute for Radiopathology and Radio-protection (IRS; Leiden, The Netherlands). P. S. and N. R.were supported by Grant P-14949 of the Austrian ScienceFund, and G. R. was supported by a grant from the “Institutefor Promotion of Innovation by Science and Technology inFlanders” (IWT-Vlaanderen) for “Generisch Basisonderzoekaan de Universiteiten” (IWT-GBOU). We express our appre-ciation for the contribution of the following colleagues to therealization of this paper: Dr. Timo van den Berg for his help inthe erythrocyte binding studies, Dr. Sem Saeland for makingthe anti-Langerin antibodies available, Dr. Frank Brombacherfor the permission to use material from IL-4R� KO mice, Dr.Gert van Cappellen for help with the confocal microscope, andTar van Os for preparation of the (photo)graphs in this work.
REFERENCES
1. Nikolic, T., Leenen, P. J. (2005) Histiocyte function and development inthe normal immune system. Chapter 3. In Histiocytic Disorders of Childrenand Adults (S. Weitzman, R. M. Egeler, eds.), Cambridge, UK, CambridgeUniversity Press, 40–65.
2. Lindblad, W. J. (1998) Collagen expression by novel cell populations inthe dermal wound environment. Wound Repair Regen. 6, 186–193.
3. Hume, D. A., Ross, I. L., Himes, S. R., Sasmono, R. T., Wells, C. A.,Ravasi, T. (2002) The mononuclear phagocyte system revisited. J. Leukoc.Biol. 72, 621–627.
4. Stout, R. D., Suttles, J. (2004) Functional plasticity of macrophages:reversible adaptation to changing microenvironments. J. Leukoc. Biol. 76,509–513.
5. Stein, M., Keshav, S., Harris, N., Gordon, S. (1992) Interleukin 4 potentlyenhances murine macrophage mannose receptor activity: a marker ofalternative immunologic macrophage activation. J. Exp. Med. 176, 287–292.
6. Gordon, S. (2003) Alternative activation of macrophages. Nat. Rev. Im-munol. 3, 23–35.
7. Gratchev, A., Kzhyshkowska, J., Utikal, J., Goerdt, S. (2005) Interleukin-4and dexamethasone counterregulate extracellular matrix remodeling andphagocytosis in type-2 macrophages. Scand. J. Immunol. 61, 10–17.
8. Porcheray, F., Viaud, S., Rimaniol, A-C., Leone, C., Samah, B., Dereud-dre-Bosquet, N., Dormont, D., Gras, G. (2005) Macrophage activationswitching: an asset for the resolution of inflammation. Clin. Exp. Immunol.142, 481–489.
9. Willment, J. A., Lin, H. H., Reid, D. M., Taylor, P. R., Williams, D. L.,Wong, S. Y., Gordon, S., Brown, G. D. (2003) Dectin-1 expression andfunction are enhanced on alternatively activated and GM-CSF-treatedmacrophages and are negatively regulated by IL-10, dexamethasone, andlipopolysaccharide. J. Immunol. 171, 4569–4573.
10. Raes, G., Brys, L., Dahal, B. K., Brandt, J., Grooten, J., Brombacher, F.,Vanham, G., Noel, W., Bogaert, P., Boonefaes, T., Kindt, A., van denBergh, R., Leenen, P. J., de Baetselier, P., Ghassabeh, G. H. (2005)Macrophage galactose-type C-type lectins as novel markers for alterna-tively activated macrophages elicited by parasitic infections and allergicairway inflammation. J. Leukoc. Biol. 77, 321–327.
11. Dupasquier, M., Stoitzner, P., van Oudenaren, A., Romani, N., Leenen,P. J. (2004) Macrophages and dendritic cells constitute a major subpopu-lation of cells in the mouse dermis. J. Invest. Dermatol. 123, 876–879.
12. Geutskens, S. B., Otonkoski, T., Pulkkinen, M. A., Drexhage, H. A.,Leenen, P. J. (2005) Macrophages in the murine pancreas and theirinvolvement in fetal endocrine development in vitro. J. Leukoc. Biol. 78,845–852.
13. Barner, M., Mohrs, M., Brombacher, F., Kopf, M. (1998) Differencesbetween IL-4R �-deficient and IL-4-deficient mice reveal a role for IL-13in the regulation of Th2 responses. Curr. Biol. 8, 669–672.
14. Yamamoto, K., Ishida, C., Shinohara, Y., Hasegawa, Y., Konami, Y.,Osawa, T., Irimura, T. (1994) Interaction of immobilized recombinantmouse C-type macrophage lectin with glycopeptides and oligosaccharides.Biochemistry 33, 8159–8166.
15. Tsuiji, M., Fujimori, M., Ohashi, Y., Higashi, N., Onami, T. M., Hedrick,S. M., Irimura, T. (2002) Molecular cloning and characterization of a novelmouse macrophage C-type lectin, mMGL2, which has a distinct carbohy-drate specificity from mMGL1. J. Biol. Chem. 277, 28892–28901.
16. Fearon, D. T. (1978) Regulation by membrane sialic acid of �1H-depen-dent decay-dissociation of amplification C3 convertase of the alternativecomplement pathway. Proc. Natl. Acad. Sci. USA 75, 1971–1975.
17. Jansen, A., Homo-Delarche, F., Hooijkaas, H., Leenen, P. J., Dardenne,M., Drexhage, H. A. (1994) Immunohistochemical characterization ofmonocytes-macrophages and dendritic cells involved in the initiation ofthe insulitis and �-cell destruction in NOD mice. Diabetes 43, 667–675.
18. Sato, M., Kawakami, K., Osawa, T., Toyoshima, S. (1992) Molecularcloning and expression of cDNA encoding a galactose/N-acetylgalac-tosamine-specific lectin on mouse tumoricidal macrophages. J. Biochem.(Tokyo) 111, 331–336.
19. Chun, K. H., Imai, Y., Higashi, N., Irimura, T. (2000) Migration of dermalcells expressing a macrophage C-type lectin during the sensitization phaseof delayed-type hypersensitivity. J. Leukoc. Biol. 68, 471–478.
20. Martinez-Pomares, L., Reid, D. M., Brown, G. D., Taylor, P. R., Stillion,R. J., Linehan, S. A., Zamze, S., Gordon, S., Wong, S. Y. (2003) Analysisof mannose receptor regulation by IL-4, IL-10, and proteolytic processingusing novel monoclonal antibodies. J. Leukoc. Biol. 73, 604–613.
21. Linehan, S. A. (2005) The mannose receptor is expressed by subsets ofAPC in non-lymphoid organs. BMC Immunol. 6, 4.
22. Herbert, D. R., Holscher, C., Mohrs, M., Arendse, B., Schwegmann, A.,Radwanska, M., Leeto, M., Kirsch, R., Hall, P., Mossmann, H., Claussen,B., Forster, I., Brombacher, F. (2004) Alternative macrophage activation isessential for survival during schistosomiasis and downmodulates T helper1 responses and immunopathology. Immunity 20, 623–635.
23. Stoitzner, P., Holzmann, S., McLellan, A. D., Ivarsson, L., Stossel, H.,Kapp, M., Kammerer, U., Douillard, P., Kampgen, E., Koch, F., Saeland,S., Romani, N. (2003) Visualization and characterization of migratoryLangerhans cells in murine skin and lymph nodes by antibodies againstLangerin/CD207. J. Invest. Dermatol. 120, 266–274.
24. Mizuochi, S., Akimoto, Y., Imai, Y., Hirano, H., Irimura, T. (1997) Uniquetissue distribution of a mouse macrophage C-type lectin. Glycobiology 7,137–146.
25. Romani, N., Holzmann, S., Tripp, C. H., Koch, F., Stoitzner, P. (2003)Langerhans cells—dendritic cells of the epidermis. APMIS 111, 725–740.
26. Denda-Nagai, K., Kubota, N., Tsuiji, M., Kamata, M., Irimura, T. (2002)Macrophage C-type lectin on bone marrow-derived immature dendriticcells is involved in the internalization of glycosylated antigens. Glycobi-ology 12, 443–450.
27. Larsen, C. P., Steinman, R. M., Witmer-Pack, M., Hankins, D. F., Morris,P. J., Austyn, J. M. (1990) Migration and maturation of Langerhans cellsin skin transplants and explants. J. Exp. Med. 172, 1483–1493.
28. Douillard, P., Stoitzner, P., Tripp, C. H., Clair-Moninot, V., Ait-Yahia, S.,McLellan, A. D., Eggert, A., Romani, N., Saeland, S. (2005) Mouselymphoid tissue contains distinct subsets of Langerin/CD207 dendriticcells, only one of which represents epidermal-derived Langerhans cells.J. Invest. Dermatol. 125, 983–994.
29. Sato, K., Imai, Y., Irimura, T. (1998) Contribution of dermal macrophagetrafficking in the sensitization phase of contact hypersensitivity. J. Immu-nol. 161, 6835–6844.
30. Linehan, S. A., Coulson, P. S., Wilson, R. A., Mountford, A. P., Brom-bacher, F., Martinez-Pomares, L., Gordon, S. (2003) IL-4 receptor signal-ing is required for mannose receptor expression by macrophages recruitedto granulomata but not resident cells in mice infected with Schistosomamansoni. Lab. Invest. 83, 1223–1231.
31. Wang, J. L., Gray, R. M., Haudek, K. C., Patterson, R. J. (2004) Nu-cleocytoplasmic lectins. Biochim. Biophys. Acta 1673, 75–93.
32. Lutz, M. B., Rovere, P., Kleijmeer, M. J., Rescigno, M., Assmann, C. U.,Oorschot, V. M., Geuze, H. J., Trucy, J., Demandolx, D., Davoust, J.,Ricciardi-Castagnoli, P. (1997) Intracellular routes and selective retentionof antigens in mildly acidic cathepsin D/lysosome-associated membraneprotein-1/MHC class II-positive vesicles in immature dendritic cells.J. Immunol. 159, 3707–3716.
33. Mann, A., Breuhahn, K., Schirmacher, P., Blessing, M. (2001) Keratino-cyte-derived granulocyte-macrophage colony stimulating factor acceler-ates wound healing: Stimulation of keratinocyte proliferation, granulationtissue formation, and vascularization. J. Invest. Dermatol. 117, 1382–1390.
34. Geissmann, F., Dieu-Nosjean, M. C., Dezutter, C., Valladeau, J., Kayal, S.,Leborgne, M., Brousse, N., Saeland, S., Davoust, J. (2002) Accumulationof immature Langerhans cells in human lymph nodes draining chronicallyinflamed skin. J. Exp. Med. 196, 417–430.
848 Journal of Leukocyte Biology Volume 80, October 2006 http://www.jleukbio.org

35. Mantovani, A., Allavena, P., Sica, A. (2004) Tumor-associated macro-phages as a prototypic type II polarized phagocyte population: role intumor progression. Eur. J. Cancer 40, 1660–1667.
36. Oda, S., Sato, M., Toyoshima, S., Osawa, T. (1988) Purification and charac-terization of a lectin-like molecule specific for galactose/N-acetyl-galac-tosamine from tumoricidal macrophages. J. Biochem. (Tokyo) 104, 600–605.
37. Imai, Y., Akimoto, Y., Mizuochi, S., Kimura, T., Hirano, H., Irimura, T.(1995) Restricted expression of galactose/N-acetylgalactosamine-specific
macrophage C-type lectin to connective tissue and to metastatic lesions inmouse lung. Immunology 86, 591–598.
38. Biswas, S. K., Gangi, L., Paul, S., Schioppa, T., Saccani, A., Sironi, M.,Bottazzi, B., Doni, A., Bronte, V., Pasqualini, F., Vago, L., Nebuloni,M., Mantovani, A., Sica, A. (2006) A distinct and unique transcrip-tional program expressed by tumor-associated macrophages: defectiveNF-{}B and enhanced IRF-3/STAT1 activation. Blood 107, 2112–2122.
Dupasquier et al. mMGL as dermis-induced mononuclear phagocyte marker 849
Copyright © 2022 FDOKUMEN




















