
The contribution of biological nitrogen fixation for sustainable agricultural systems in the tropics
13
Soil Biel. Biochem. Vol. 29, No. 516, 787-799, pp. 1997 0 1997 Elsevier ScienceLtd. Al1rights reserved Printed in Great Britain PII: SOO38-0717(%)00221-0 0038-0717/97 $17.00 + 0.00 THE CONTRIBUTION OF BIOLOGICAL NITROGEN FIXATION FOR SUSTAINABLE AGRICULTURAL SYSTEMS IN THE TROPICS ROBERT M. BODDEY,‘* JOÀO CARLOS DE MORAES SÁ ,2 BRUNO J. R. ALVES’ and SEGUNDO URQUIAGA’ ‘EMBRAPA-CNPAB, Seropédica, Rio de Janeiro, 23851-970, Brazil and *Fundaçäo ABC, Caixa Postal 892, Ponta Grossa, 84001-970, Paraná, Brazil (Accepted 1 July 1996) Summary-The pressing need to increase food production in the tropics to feed the burgeoning popu- lation of the Third World requires that crop yields ha-’ must be increased without prejudicing the resource base for future generations. Biological nitrogen fixation (BNF), especially that associated with legumes, has great potential to contribute to productive and sustainable agricultural systems for the tro- pits, but more research is required to investigate how biologically fixed N, and the increased BNF con- tributions resulting from research innovations, can be incorporated into viable agricultural systems to increase crop or pasture yields and to substitute N fertiliser inputs. A majority of the soils of the humid and semi-humid tropics have mineral fractions composed of 1:1 lattice clays or sesquioxides of relatively low capacity to retain nutrients (CEC) and water (WHC). It is the soil organic matter (SOM) which has high CEC (after liming) and WHC, and soils under undisturbed climax vegetation are usually high in organic matter which is responsible for their fertility. The key to the long term fertility of such soils is to maintain their soil organic matter by the preservation of crop residues and the selection of suitable crop rotations or fallows. In this review we examine several types of agricultural systems utilised in the tropics ranging from pastures, ley cropping, zero-til1 rotations as wel1 as green manuring and discuss the management options that can be adopted to preserve their agricultural productivity through the strategie use of legumes in these systems, and their effects on pasture and crop yields. The introduction of forage legumes into tropical pastures can increase and sustain their productivity, with only modest inputs of lime and P and K fertilisers. Similarly, crop and pasture rotations (ley cropping) maintain SOM and soil fertility and crop yields can benefit greatly from the introduction of pasture legumes into the ley. Continuous cropping under minimum or zero tillage can maintain soil cover, and stimulate the retention of SOM, such that nutrient losses are often minimal, and legume derived N can be efficiently transferred to subsequent crops. The options for the resource-poor small-holder to efficiently utilise bio- logically fixed N as a N supply for cereal grains are more limited and need more attention from researchers as wel1 as less neglect from government organisations. The addition of lime and P fertiliser in modest quantities in many under-developed regions could make large contributions to increased crop yields. If such modest fertiliser inputs were to be combined with suitable crop rotations including green manure or grain legume crops, larger increases in crops yields could be achieved on a sustainable basis, but in many regions agricultural extension services are non-existent and poor farmers have little access to even these basic chemical inputs. 0 1997 Elsevier Science Ltd INTRODUCIION If the developing countries of the tropics are to meet their demand for food over the next 30 years, when their human population is expected to increase from approximately 4 to 7 billion, food crop yields wil1 have to increase by a similar pro- portion. Large areas of sub-Saharan Africa and Latin America (notably the 200 million ha of the Brazilian Cerrado), but virtually none in South and Centra1 Asia, are suitable for agriculture but uncul- *Author for correspondence: Fax: + 55-21-682-1230; e- mail: [email protected]. 787 tivated. While it is inevitable that a certain pro- portion of this land wil1 be needed for crop production, it is stil1 necessary that agricultural yields be increased in a sustainable manner in al1 regions of the tropics already under cultivation (Borlaug and Dowswell, 1994). Twenty years ago Greenland (1975) suggested five basic principles of soil management essential for sustainable agricultural production: (1) chemical nutrients removed by crops must be replenished, (2) the physical condition of the soil must be maintained,
Transcript of The contribution of biological nitrogen fixation for sustainable agricultural systems in the tropics

Soil Biel. Biochem. Vol. 29, No. 516, 787-799, pp. 1997 0 1997 Elsevier Science Ltd. Al1 rights reserved
Printed in Great Britain PII: SOO38-0717(%)00221-0 0038-0717/97 $17.00 + 0.00
THE CONTRIBUTION OF BIOLOGICAL NITROGEN FIXATION FOR SUSTAINABLE AGRICULTURAL
SYSTEMS IN THE TROPICS
ROBERT M. BODDEY,‘* JOÀO CARLOS DE MORAES SÁ ,2 BRUNO J. R. ALVES’ and SEGUNDO URQUIAGA’
‘EMBRAPA-CNPAB, Seropédica, Rio de Janeiro, 23851-970, Brazil and *Fundaçäo ABC, Caixa Postal 892, Ponta Grossa, 84001-970, Paraná, Brazil
(Accepted 1 July 1996)
Summary-The pressing need to increase food production in the tropics to feed the burgeoning popu- lation of the Third World requires that crop yields ha-’ must be increased without prejudicing the resource base for future generations. Biological nitrogen fixation (BNF), especially that associated with legumes, has great potential to contribute to productive and sustainable agricultural systems for the tro- pits, but more research is required to investigate how biologically fixed N, and the increased BNF con- tributions resulting from research innovations, can be incorporated into viable agricultural systems to increase crop or pasture yields and to substitute N fertiliser inputs. A majority of the soils of the humid and semi-humid tropics have mineral fractions composed of 1: 1 lattice clays or sesquioxides of relatively low capacity to retain nutrients (CEC) and water (WHC). It is the soil organic matter (SOM) which has high CEC (after liming) and WHC, and soils under undisturbed climax vegetation are usually high in organic matter which is responsible for their fertility. The key to the long term fertility of such soils is to maintain their soil organic matter by the preservation of crop residues and the selection of suitable crop rotations or fallows. In this review we examine several types of agricultural systems utilised in the tropics ranging from pastures, ley cropping, zero-til1 rotations as wel1 as green manuring and discuss the management options that can be adopted to preserve their agricultural productivity through the strategie use of legumes in these systems, and their effects on pasture and crop yields. The introduction of forage legumes into tropical pastures can increase and sustain their productivity, with only modest inputs of lime and P and K fertilisers. Similarly, crop and pasture rotations (ley cropping) maintain SOM and soil fertility and crop yields can benefit greatly from the introduction of pasture legumes into the ley. Continuous cropping under minimum or zero tillage can maintain soil cover, and stimulate the retention of SOM, such that nutrient losses are often minimal, and legume derived N can be efficiently transferred to subsequent crops. The options for the resource-poor small-holder to efficiently utilise bio- logically fixed N as a N supply for cereal grains are more limited and need more attention from researchers as wel1 as less neglect from government organisations. The addition of lime and P fertiliser in modest quantities in many under-developed regions could make large contributions to increased crop yields. If such modest fertiliser inputs were to be combined with suitable crop rotations including green manure or grain legume crops, larger increases in crops yields could be achieved on a sustainable basis, but in many regions agricultural extension services are non-existent and poor farmers have little access to even these basic chemical inputs. 0 1997 Elsevier Science Ltd
INTRODUCIION
If the developing countries of the tropics are to meet their demand for food over the next 30 years, when their human population is expected to increase from approximately 4 to 7 billion, food crop yields wil1 have to increase by a similar pro- portion. Large areas of sub-Saharan Africa and Latin America (notably the 200 million ha of the Brazilian Cerrado), but virtually none in South and Centra1 Asia, are suitable for agriculture but uncul-
*Author for correspondence: Fax: + 55-21-682-1230; e- mail: [email protected].
787
tivated. While it is inevitable that a certain pro- portion of this land wil1 be needed for crop production, it is stil1 necessary that agricultural yields be increased in a sustainable manner in al1 regions of the tropics already under cultivation (Borlaug and Dowswell, 1994).
Twenty years ago Greenland (1975) suggested five basic principles of soil management essential for sustainable agricultural production:
(1) chemical nutrients removed by crops must be replenished,
(2) the physical condition of the soil must be maintained,

788 Robert M. Boddey et UI
(3) there must be no build up of weeds, pests or diseases.
(4) there must be no increase in soil acidity or toxic elements,
(5) soil erosion must be controlled to be equal to or less than the rate of soil genesis.
In most tropical agricultural systems today it is obvious that most of these principles are not being adhered to. In addition to the physical degradation of the soil and widespread erosion in the tropics, in many areas more nutrients are being removed from the soil than are added. It is estimated that an aver- age of between 20 and 70 kg ha-’ of nitrogen and potassium are exported from agricultural land in sub-Saharan Africa, Asia and Latin America annually (Giller and Cadisch, 1995). Of course, it is not only necessary to replace the nutrients removed from the system as agricultural produce, or lost via leaching and to the atmosphere (when residues are burned for example), but also to ensure that their availability for crop growth is maintained.
While low levels of crop production can be main- tained for many years, or perhaps indefinitely, using systems where no inorganic fertilisers are used, the urgent need to increase crop productivity means that they cannot be dispensed with. Although it is possible to produce high yields of staple cereal grains using organic fertilisers (e.g. animal wastes), they are rarely available in sufficient quantities for application on a large scale. For example, a pro- ductive Bruchiariu pasture grazed at 3 animals ha-’ (with a live weight gain of 1 kg ha-’ d-‘) could pro- duce approximately 40 kg N y-’ as dung (and 40 kg as urine; Boddey et al., 1995a) so even if this dung could be stored without N loss, 2.5 ha of pasture would be required to provide 100 kg N ha-’ for a cereal crop.
It is obvious therefore that increased use of inor- ganic fertilisers, as wel1 as other agrochemicals, wil1 be essential to increase crop yields in these countries. However, use at the levels presently applied in Europe, the US, Japan and some of the Eastern Asian countries is unlikely to be sustainable even if they are available at prices which farmers can afford. For example, N fertiliser is produced mainly from natura1 gas, the price of which is clo- sely tied to that of crude oil, the present market price of which does not reflect the long term avail- ability of this resource (46 years of known reserves according to a 1985 Shell report (Shell, 1985)). Large increases in oil and natura1 gas, and hence N fertiliser, prices followed by decreasing fertiliser use by farmers (particularly many highly productive Asian rite farmers) leading to reversal of some of the enormous gains in crop yields achieved during the Green Revolution, is an alarming prospect. While some fertiliser and other agro-chemical inputs are essential, it is desirable, as has been suggested
by Scholes et al. (1994), that the working concept of a sustainable agriculture for the tropical develop- ing countries should exclude the excessive reliance on the use of mineral fertilisers, farm energy and crop protection chemicals.
In this review we consider what management options are available within this concept to develop sustainable agricultural production systems for the tropics, and how nitrogen derived from biological nitrogen fixation (BNF) can be introduced into these systems. The use of Nz-fixing trees is not dis- cussed as this is the subject of other papers pre- sented at this Symposium.
EFFICIENT USE OF PLANT NUTRIENTS AS A KEY TO SUSTAINABLE CROP PRODUCTION
The native climax vegetation of the forests and savannahs of the tropics are sustainable, since not only a smal1 fraction of their biomass is usually removed as fruits and leaves by native fauna, but also because of the dense, usually perennial, mat of roots which efficiently Capture soluble nutrients released from decomposing organic materials. It is this latter characteristic of these ecosystems, of vir- tually closed nutrient cycles, which contrasts starkly with many of the existing agricultural practises adopted in the tropics. Many of the soils of the humid and sub-humid tropics are low in clay con- tent, and in most, this clay is of the 1:l type or ses- quioxide material which in either case have very low cation exchange capacity (CEC), water holding capacity and aggregate stability. These properties, which are those responsible for soil fertility, are de- rived mainly from the organic matter of the soil which is replenished by plant litter and decaying roots. Frequent ploughing, which is a characteristic of the modern farming practices often adopted in these regions, causes the break up of soil structure and the oxidation of soil organic matter (SOM), with a consequent gradual loss of soil fertility. This is wel1 illustrated by data from the region of the West of Bahia (Barreiras) in Brazil, where it was shown that more than half of the SOM was lost in just 3 years after the conversion of native Cerrado to conventional annual planting of soybean (Fig. 1; Da Silva et al., 1994). Crop production in this area of 8 million ha was only started in the last 15 years and in 70% of the area, where the soils contain less than 20% clay, it was estimated that the fa11 in SOM content has been responsible for a decrease in soil CEC of between 53 and 61% within just 5 years of introducing soybean.
It follows therefore that if a productive and sus- tainable agriculture is to be practised in the acid soils of low clay content which predominate in the humid and sub-humid tropics, practises must be adopted which maintain or increase SOM reserves.
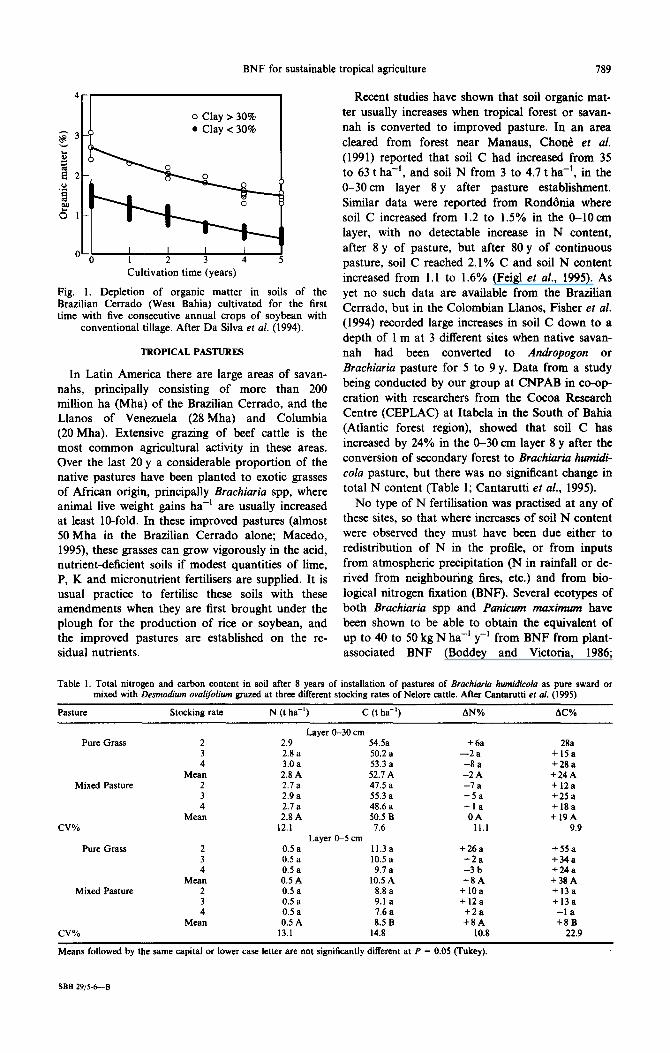
BNF for sustainable tropical agriculture 789
0 Clay > 30% ??Clay < 30%
-0-I Cultivation time (years)
Fig. 1. Dcpletion of organic matter in soils of the Brazilian Cerrado (West Bahia) cultivated for the first time with five consecutive annual crops of soybean with
conventional tillage. After Da Silva er al. (1994).
TROPICAL PASTURES
In Latin America there are large areas of savan- nahs, principally consisting of more than 200 million ha (Mha) of the Brazilian Cerrado, and the Llanos of Venezuela (28 Mha) and Columbia (20 Mha). Extensive grazing of beef cattle is the most common agricultural activity in these areas. Over the last 20 y a considerable proportion of the native pastures have been planted to exotic grasses of African origin, principally Bruchiuriu spp, where animal live weight gains ha-’ are usually increased at least lO-fold. In these improved pastures (almost 50 Mha in the Brazilian Cerrado alone; Macedo, 1995), these grasses can grow vigorously in the acid, nutrient-deficient soils if modest quantities of lime, P, K and micronutrient fertilisers are supplied. It is usual practice to fertilise these soils with these amendments when they are first brought under the plough for the production of rite or soybean, and the improved pastures are established on the re- sidual nutrients.
Recent studies have shown that soil organic mat- ter usually increases when tropical forest or savan- nah is converted to improved pasture. In an area cleared from forest near Manaus, Chonè et al. (1991) reported that soil C had increased from 35 to 63 t ha-‘, and soil N from 3 to 4.7 t ha-‘, in the 0-30 cm layer 8 y after pasture establishment. Similar data were reported from Rondônia where soil C increased from 1.2 to 1.5% in the O-10cm layer, with no detectable increase in N content, after 8 y of pasture, but after 80 y of continuous pasture, soil C reached 2.1% C and soil N content increased from 1.1 to 1.6% (Feigl et af., 1995). As yet no such data are available from the Brazilian Cerrado, but in the Colombian Llanos, Fisher et al. (1994) recorded large increases in soil C down to a depth of 1 m at 3 different sites when native savan- nah had been converted to Andropogon or Bruchiuriu pasture for 5 to 9 y. Data from a study being conducted by our group at CNPAB in co-op- eration with researchers from the Cocoa Research Centre (CEPLAC) at Itabela in the South of Bahia (Atlantic forest region), showed that soil C has increased by 24% in the 0-30 cm layer 8 y after the conversion of secondary forest to Bruchiuria humidi- cola pasture, but there was no significant change in total N content (Table 1; Cantarutti et al., 1995).
NO type of N fertilisation was practised at any of these sites, so that where increases of soil N content were observed they must have been due either to redistribution of N in the profile, or from inputs from atmospheric precipitation (N in rainfall or de- rived from neighbouring fires, etc.) and from bio- logica1 nitrogen fixation (BNF). Several ecotypes of both Bruchiuriu spp and Punicum maximum have been shown to be able to obtain the equivalent of up to 40 to 50 kg N ha-’ y-’ from BNF from plant- associated BNF (Boddey and Victoria, 1986;
Table 1. Total nitrogen and carbon content in soil after 8 years of installation of pastures of Erachiaria hmidicola as pure sward or mixed with Desmodium ovalifoliutn grazed at thm different smeking rates of Nelore cattle. After Cantarutti er al. (1995)
Pasture Stocking rate N (t ha-‘) C (t ha-‘) AN% AC%
Pure Grass 2 3 4
Mean Mixed Pasture 2
3 4
Mean CV%
Pure Grass 2 3 4
Mean Mixed Pasture 2
3 4
Mean CV%
Layer 0-30 cm 2.9 54.5a 2.8 a 50.2 a 3.0a 53.3 a 2.8 A 52.1 A 2.1 a 47.5 a 2.9 a 55.3 a 2.1 a 48.6 a 2.8 A 50.5 B
12.1 1.6 Layer 0-5 cm
0.5 a 11.3a 0.5 a 10.5 a 0.5 a 9.7 a 0.5 A 10.5 A 0.5 a 8.8 a 0.5 a 9.1 a 0.5 a 1.6 a 0.5 A 8.5 B
13.1 14.8
+6a 28a -2 a +15a -8 a +28a -2A +24A -la +12a +5a +25a +la +18a OA +19A
11.1 9.9
+26a +2a -3b +SA
+lOa +12a
+2a +8A
10.8
+55a +34a +24a +38A +13a +13a
-1 a +8B
22.9
Means followed by the same capita1 or lower case letter are not significantly different at P = 0.05 (Tukey).
SBB 29/S-6-B
790 Robert M. Boddey et al
Miranda and Boddey, 1987), but contributions in actual pastures under grazing have not been esti- mated.
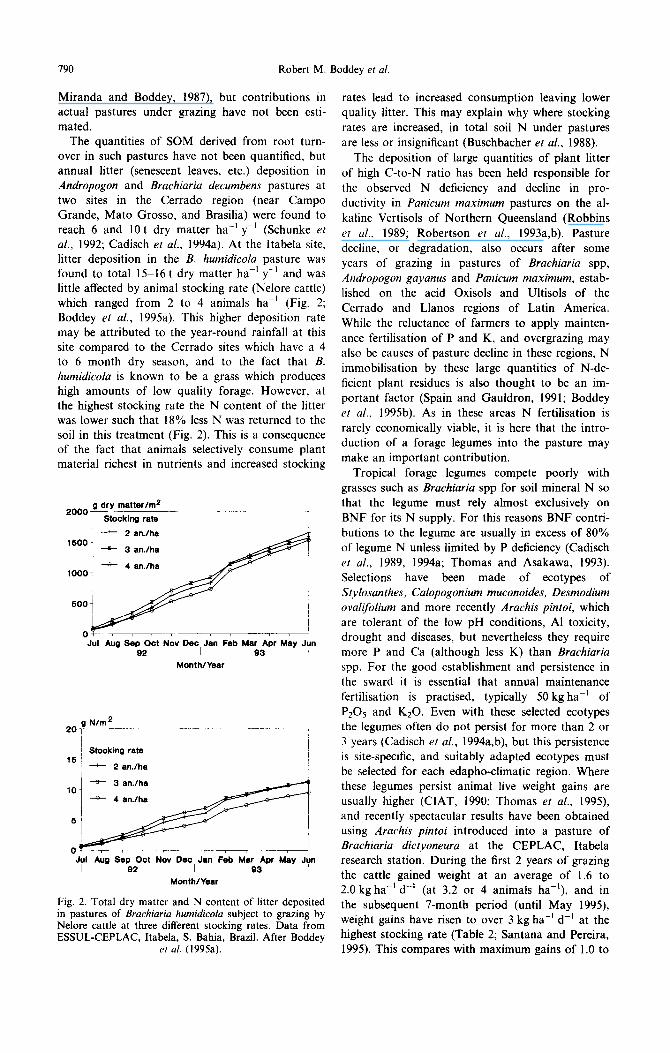
The quantities of SOM derived from root turn- over in such pastures have not been quantified, but annual litter (senescent leaves, etc.) deposition in Andropogon and Brachiaria decumbens pastures at two sites in the Cerrado region (near Campo Grande, Mato Grosso, and Brasilia) were found to reach 6 and 10 t dry matter ha-’ y-’ (Schunke et al., 1992; Cadisch et al., 1994a). At the Itabela site, litter deposition in the B. humidicolu pasture was found to total 15-16 t dry matter ha-’ y-’ and was little affected by animal stocking rate (Nelore cattle) which ranged from 2 to 4 animals ha-’ (Fig. 2; Boddey et al., 1995a). This higher deposition rate may be attributed to the year-round rainfall at this site compared to the Cerrado sites which have a 4 to 6 month dry season, and to the fact that B. humidicola is known to be a grass which produces high amounts of low quality forage. However, at the highest stocking rate the N content of the litter was lower such that 18% less N was returned to the soil in this treatment (Fig. 2). This is a consequente of the fact that animals selectively consume plant material richest in nutrients and increased stocking
2000 g dry matter/m2
Stocking rate
-+ 2 an./ha 1600
-+ 3 an.lha
+ 4 an.fha 1000
600 /
Jrl Aug Sep Oct NW Dec, Jan Feb Mar Apr May Jy 92 93
Month/Year
-g 3 an./ha 10
-+ 4 an.lha
Aug Sep Oot Nov Dec, Jan Feb Mar Apr May Jy 92 9.3
Fig. 2. Total dry matter and N content of litter deposited in pastures of Brachiaria humidicola subject to grazing by Nelore cattle at three different stocking rates. Data from ESSUL-CEPLAC, Itabela, S. Bahia, Brazil. After Boddey
et al. (1995a).
rates lead to increased consumption leaving lower quality litter. This may explain why where stocking rates are increased, in total soil N under pastures are less or insignificant (Buschbacher et al., 1988).
The deposition of large quantities of plant litter of high C-to-N ratio has been held responsible for the observed N deficiency and decline in pro- ductivity in Panicum maximum pastures on the al- kaline Vertisols of Northern Queensland (Robbins et al., 1989; Robertson et al., 1993a,b). Pasture decline, or degradation, also occurs after some years of grazing in pastures of Bruchiaria spp, Andropogon gayanus and Panicum maximum, estab- lished on the acid Oxisols and Ultisols of the Cerrado and Llanos regions of Latin America. While the reluctance of farmers to apply mainten- ante fertilisation of P and K, and overgrazing may also be causes of pasture decline in these regions. N immobilisation by these large quantities of N-de- ficient plant residues is also thought to be an im- portant factor (Spain and Gauldron, 1991; Boddey et al., 1995b). As in these areas N fertilisation is rarely economically viable, it is here that the intro- duction of a forage legumes into the pasture may make an important contribution.
Tropical forage legumes compete poorly with grasses such as Brachiaria spp for soil mineral N so that the legume must rely almost exclusively on BNF for its N supply. For this reasons BNF contri- butions to the legume are usually in excess of 80% of legume N unless limited by P deficiency (Cadisch <-‘t al., 1989, 1994a; Thomas and Asakawa, 1993). Selections have been made of ecotypes of Stylosanthes, Calopogonium muconoides, Desmodium ovalifolium and more recently Arachis pintoi, which are tolerant of the low pH conditions, Al toxicity, drought and diseases, but nevertheless they require more P and Ca (although less K) than Brachiaria spp. For the good establishment and persistente in the sward it is essential that annual maintenance fertilisation is practised, typically 50 kg ha-’ of PZOs and KzO. Even with these selected ecotypes the legumes often do not persist for more than 2 or 3 years (Cadisch et al., 1994a,b), but this persistente is site-specific, and suitably adapted ecotypes must be selected for each edapho-climatic region. Where these legumes persist animal live weight gains are usually higher (CIAT, 1990; Thomas er al., 1995), and recently spectacular results have been obtained using Arachis pintoi introduced into a pasture of Brachiaria dictyoneura at the CEPLAC, Itabela research station. During the first 2 years of grazing the cattle gained weight at an average of 1.6 to 2.0 kg ha-’ d-’ (at 3.2 or 4 animals ha-‘), and in the subsequent 7-month period (until May 1995), weight gains have risen to over 3 kg ha-’ d-’ at the highest stocking rate (Table 2; Santana and Pereira, 1995). This compares with maximum gains of 1.0 to
BNF for sustainable tropical agriculture 791
Table 2. Live weight gains of Nelor cattle grazing on a mixed pasture of Brachiario dicryoneura and Arachis repens in the Atlantic forest regio”, Southem Bahia, Brazil. Data from Santana and Pereira (1995)
Period Stocking rate (an ha-‘) Live weight gain
Nov. 92-May 93 (182 days) 1.6 2.4 3.2 4.0
May 93-Feb 94 (286 days) 1.6 2.4 3.2 4.0
Oct 94May 95 (205 days) 1.6 2.4 3.2 4.0
549 571 575 494 499 590 509 502 688 697 688
g ha-‘d-’ 879
1368 1841 1978 791
1416 1629 2010 1100 1674 2201 3194
1.2 kg ha-’ d-’ on other pure Brachiaria pastures at this site (Pereira et al., 1992; Boddey et al., 1995a).
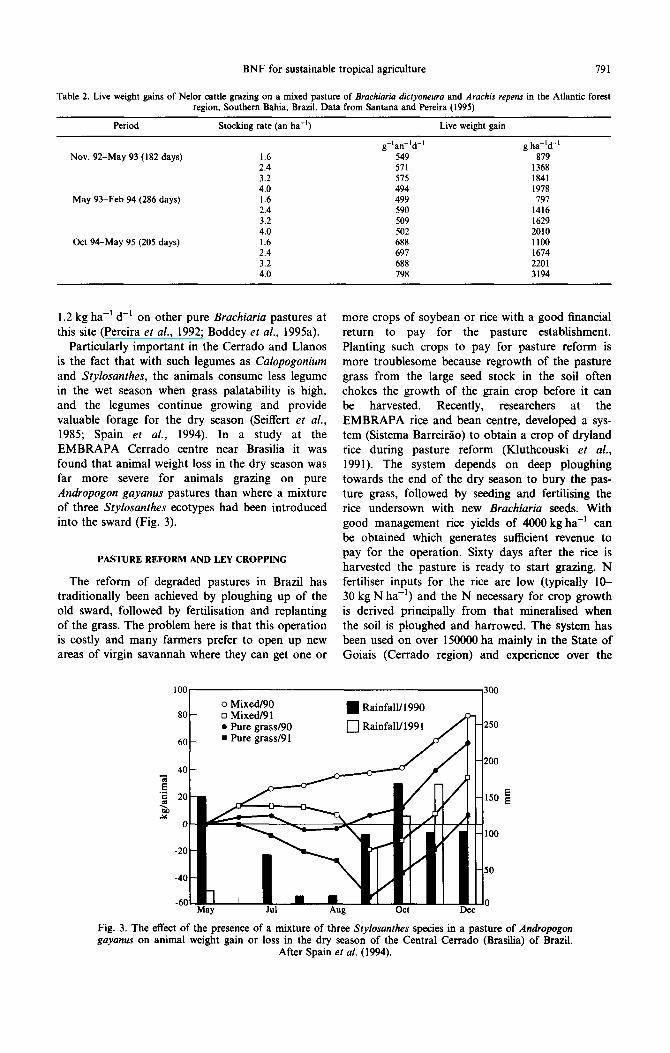
Particularly important in the Cerrado and Llanos is the fact that with such legumes as Calopogonium and Stylosanthes, the animals consume less legume in the wet season when grass palatability is high, and the legumes continue growing and provide valuable forage for the dry season (Seiffert et al., 1985; Spain et al., 1994). In a study at the EMBRAPA Cerrado centre near Brasilia it was found that animal weight loss in the dry season was far more severe for animals grazing on pure Andropogon gayanus pastures than where a mixture of three Stylosanthes ecotypes had been introduced into the sward (Fig. 3).
PASTURE REFORM AND LEY CROPPING
The reform of degraded pastures in Brazil has traditionally been achieved by ploughing up of the old sward, followed by fertilisation and replanting of the grass. The problem here is that this operation is costly and many farmers prefer to open up new areas of virgin savannah where they can get one or
more crops of soybean or rite with a good financial return to pay for the pasture establishment. Planting such crops to pay for pasture reform is more troublesome because regrowth of the pasture grass from the large seed stock in the soil often chokes the growth of the grain crop before it can be harvested. Recently, researchers at the EMBRAPA rite and bean centre, developed a sys- tem (Sistema Barreirao) to obtain a crop of dryland rite during pasture reform (Kluthcouski et al., 1991). The system depends on deep ploughing towards the end of the dry season to bury the pas- ture grass, followed by seeding and fertilising the rite undersown with new Brachiaria seeds. With good management rite yields of 4000 kg ha-’ can be obtained which generates sufficient revenue to pay for the operation. Sixty days after the rite is harvested the pasture is ready to start grazing. N fertiliser inputs for the rite are low (typically l(r 30 kg N ha-‘) and the N necessary for crop growth is derived principally from that mineralised when the soil is ploughed and harrowed. The system has been used on over 150000 ha mainly in the State of Goiais (Cerrado region) and experience over the
80
60
o Mixed/90 ??Mixed/91 ??Pure grassl90 ??Pure grassl91
??RainfaWl990 _
0 Rainf al111991 d AI1
fl250
-60 May Ja1 Aag Oct Dec
0
Fig. 3. The effect of the presence of a mixture of three Stylosanthes species in a pasture of Andropogon guyanus on animal weight gain or loss in the dry season of the Centra1 Cerrado (Brasilia) of Brazil.
After Spain er al. (1994).

192 Robert M. Boddey at ui.
last 9 y indicates that the process should be repeated every 5 to 7 y. At present studies are being undertaken to determine if repeated use of this sys- tem wil1 deplete SOM reserves as there is no appar- ent N input to balance possible leaching losses when the soil is cultivated and before the rite or grass roots are established. The suggested solution is to include a forage legume in this system to fìll this shortfall.
As the evidente suggests that SOM increases under continuous pastures, and mechanica1 cultiva- tion leads to SOM loss, a sustainable system of SOM management would logically appear to be the alternation between a few years of pasture followed by a few years of arable cropping. Such ley crop- ping systems were used for centuries in Britain and elsewhere in Europe, for example in the Norfolk four-course rotation. Their use has been widespread in Southern Australia in areas with a temperate or Mediterranean climate where legumes such as lucerne (alfalfa) and other Medicugo spp are reported to promote more enduring benefits to sub- sequent crops than does N fertiliser application (Lloyd et al., 1991). There has been httle adoption of ley cropping in the tropical regions of Australia although studies conducted on the heavy clay soils of subtropical Queensland (Dalal et al., 1994) and the Oxisols and Ultisols of the tropical regions of Northern Australia (Jones et al., 1991), showed that with suitable management grazed legume leys can produce high animal weight gains, and the residual effect of the legume N for the succeeding grain crop was the equivalent of up to 90 kg of fertiliser N ha-‘.
In the developing countries of the tropics few stu- dies appear to have been conducted, although long term trials have been initiated recently at CIAT (Columbia) and at two EMBRAPA centres sites in the Cerrado (CPAC, Brasilia and CNPGC, Campo Grande, MS). In ah these studies interest has focused on the residual effect of the legume in the sward on subsequent grain yields and in the Eastern Llanos of Colombia it was found that rite yield fol- lowing a 10 y-old grass-alone Brachiaria pasture was 1.3 t ha-‘, but this was increased to 3.0 t ha-’ where kudzu (Pueraria phaseoloides) was present in the pasture, but rite yields immediately after native savannah did not exceed 2 t ha-’ even when ferti- hsed with 50 kg P and 80 kg N ha-’ (Fig. 4).
The problems with pastures, either pure grass or mixed grass legume swards, is that they do not gen- erate sufficient revenues on a ha-’ basis to generate wealth for peasant small-holders. Similarly, ley cropping systems suit large landowners who can divide their land such that they continuously pro- duce grain crops. To resolve the probfems of smal1 farmers whether in tropical Africa, Asia or Latin America, cropping systems which can be used to produce grain or other cash crops al1 year round,
Rite yield (ka) n
0 40 80 0 40 80 (kgN/ha)
Rite after Savanna
Rite after Rite after B. decumbens B. decumbens +
i? phaseoloides
Fig. 4. Rite yields foilowing cultivation of native savan- nah. a grass-alone pasture of Brachiaria decumhens or a mixed pasture of B. decumhens and Pueraria phaseoloides in the Eastern Llanos (savannah) of Colombia. After Vera
e/ al. (1992).
or at least annually, without degrading the soil are required.
ZERO TILLAGE AND CROP ROTATION
One practice which can preserve soil organic mat- ter while continuing to produce grain crops annually, is the use of zero tillage. Much of the State of Parana in southern Brazil, was only opened up for agriculture over the last 30 to 40 years. As winters are mild with only a few dozen days of frost in this region, it was found that there was a very profitable combination of soybean or maize in the summer followed by wheat in the winter. However, this 2-crop rotation involved ploughing and preparing the seed bed twice a year, and in the regions with hghter soils, such as the Campos Gerais region, SOM reserves were quickly depleted leading to decreasing crops yields, increasing fertili- ser use and severe erosion problems on sloping lands.
Initial trials on using zero tillage were made by agronomists from the EMBRAPA wheat centre (CNPT) and the Agronomie Institute of Paraná (IAPAR) starting in the 1970s. Many problems were encountered including the lack of suitable ma- chinery for planting into the crop residues as wel1 as weed control and pest problems. It took some years before suitable crop rotations were established and good seed drills and effective herbicides became available. The increasing demand by farmers stimu- lated the development and distribution of the ma- chinery and chemicals by national and multinational companies. Today these problems have largely been solved through continued efforts by EMBRAPA and IAPAR and in the Campos Gerais region by research performed by the

BNF for sustainable tropical agriculture 793
Fundaçao ABC, a private foundation fimded by agricultural co-operatives of the region.
Today in this region the most common rotation is of 3 y duration starting year I-forage oats (Avena strigosa) in winter, followed by soybean (summer); year 2-wheat (winter), soybean (sum- mer); year 3-oats (winter), maize (summer). The soybean residues are of low C-to-N ratio and decompose rapidly, so to maintain soil cover and build up SOM, a winter graminaceous crop is required. Wheat every winter was found to cause the build up of root diseases and pests of this crop and the remaining straw was stil1 insufficient to arrive at the 4 to 5 ha-’ deemed necessary to keep year-round soil cover (Sá, 1993). The forage oats is not grown to maturity but only to the immature (milky) grain stage (so that there is no subsequent seed germination to produce a weed problem) and then a knife roller is passed to fel1 the crop to pro- duce a surface mulch. As long as the oats is only lightly grazed (after complete soil cover), or not grazed at all, the inclusion of the oats in the ro- tation increases the quantity of surface mulch, so that there is year-round soil cover which prevents erosion, conserves soil moisture, reduces surface soil temperatures and inhibits weed growth.
A recent study performed at the Fundaçäo ABC investigated the response of the maize crop to N fertiliser, and the substitution of the 3rd year oat crop by a green manure crop (vetch, I/iciu sntivu), over a period of 7 y of this no-til1 rotation started on an area previously used for unfertilised pasture. In the study, three areas were used with the three different years of the rotation, respectively, such that there was a maize crop in one of the areas every year. Even where no green manure was included in the rotation, grain yields were high (mean of al1 7 y 7860 kg ha-‘) even when no N ferti- liser was added. The mean grain N content was between 1.3 and 1.4%, so that more than 100 kg N ha-’ were removed from the field in grain every 3 y in the absente of N fertiliser or green manure. Soil organic matter (Walkley Black) increased in the 0-20 cm layer from 3.1 to 4.0% during the study, so that this 100 kg N ha-’ gained by the system during the rotation is a minimum figure. The only legume in this system was the soy- bean, which although it can obtain a very large pro- portion of N from BNF, also has a very high proportion of shoot N (in this case 7S-80%) in the grain which was exported from the system. Several authors have stressed that for this reason soybean is unlikely to contribute significant N to subsequent crops (Giller et al., 1994; Jensen and Castellanos, 1994). Assuming that BNF inputs to the maize or other cereals were negligible, the most likely expla- nation for this N input would be from soybean roots and nodules left in the soil. In any case these data show clearly that at no time during the ro-
140 l_
120
: 100 After oats
; *O 60
ò 40
d 20
0 88 89
UePr 91 92
Fig. 5. Nitrogen fertilisation for maximum economie yield of maize over 5 y in a zero-tillage rotation of soybean with wheat and oats (Avena strigosu), or with the oat crop pre- ceding the maize, replaced by vetch (Vicia sativa). Data
from Fundaçäo ABC, Ponta Grossa, Paraná, Brazil.
tation were N losses high and this system was highly efficient in conserving this, and presumably other, nutrients.
For each year a N response curve for the maize was computed to assess N fertiliser requirement for maximum economie yield. The data displayed in Fig. 5 show that even without the green manure, the N requirement of the maize declined each year. The explanation for this is that the SOM content is increasing but nearing equilibrium, so that each year there is less net immobilisation of N (C and other nutrients) in the SOM of the soil. The data also show that the replacement of the oats, which immediately preceded the maize crop, by vetch, halved the N requirement of the maize for maxi- mum yield.
The success of this system is such that across the 150000 ha of zero tillage on the co-operatives which fund the Fundaçäo ABC, maize yields were 7.2 t ha-’ (compared with a national average of 1.4) in 1994 with a mean addition of 80 kg N fertiliser ha-‘. Mean soybean and wheat yields in this area were 3.3 and 3.0 t ha-‘, respectively, last year (Brazilian averages: soybean 1.7 and wheat 1.5 t ha-‘).
Zero tillage systems are spreading rapidly in Brazil and it estimated that this has reached almost 5 million ha (almost 10% of al1 cropped land) and the fastest area of expansion is into the tropical areas, particularly the Cerrado. In these areas win- ter crops are substituted by others depending on the climate. In the savannah regions dry season crops such as millets or sorghums are used and pigeon pea (Cujunus cajun) is sometimes used as a dry sea- son legume. If farmers have the organisation, capi- tal, access to fertilisers and pesticides and they are wel1 served by competent private or public research organisations, large areas of the semi-arid, sub- humid or even humid tropics could produce high and sustainable yields using zero tillage techniques. However, for the poorer countries, or for the

794 Robert M. Boddey et al.
poorer regions in countries like Brazil, where small- holders lack education, infrastructure and even the smallest surplus to invest in their land, such systems are inappropriate. For these small-holders and sub- sistence farmers, simple crop rotations including legumes and intercropping of legumes with cereals are the systems most used at present, but their total benefit as an N source for tropical agriculture is as yet very limited.
GREEN MANURES FOR THE SMALL FARMER
Dry land crops
In many regions of the tropics the slash and burn system has been practised for centuries, and there is no question that if land is returned to bush for a sufficient number of years before being cleared again for cropping, the system is sustainable (Greenland, 1975). Legumes in the bush fallow are important in restoring N fertility, and deep-rooting trees recover other nutrients leached below the root- ing depth of the arable crops. However, the increased pressure on available land means that these systems are no longer sustainable and the farmers need alternatives to increase crop yields on ever-decreasing areas of land. In many parts of Asia, such as India, China and Indonesia, where population densities have been traditionally high and land use has been more intensive, the use of green manures, as wel1 as animal and human wastes, has been widespread. The most important crop in these areas is wetland rite and the use of green manures in this crop is discussed below. The great diversity of different systems used and investi- gated by research teams cannot be covered in this review, but just a few genera1 points are made.
The planting of a grain legume crop can give very variable residual N-benefit to a succeeding cer- eal crop. This wil1 depend on the amount of N fixed by the legume, the amount of N left behind in the field after removing the legume grain, and the efficiency of use of this residual N. This problem was discussed with respect to several grain legumes by Giller et al. (1994) and Jensen and Castellanos (1994), and, as mentioned before, legumes such as soybean which have very high harvest indices are unlikely to contribute much N to the succeeding crop. Obviously, farmers would not accept varieties of grain legumes which produced lower grain yields but leave more N in residues for the subsequent crop, but Jensen and Castellanos (1994) suggest that plant breeders could pay less attention to high harvest index if total grain yield was the same or higher. For legumes which naturally have a low or moderate harvest index, such as pigeon pea or groundnut, the benefits to subsequent cereal crops can be very significant (Kumar Rao et al., 1983; Dakora et al., 1987; McDonagh et al., 1993). In the field study of McDonagh et al. (1993) on groundnut
followed by maize conducted on a sandy soil in NE Thailand, the contribution of BNF to the ground- nut varieties (shoots) ranged from 100 to 130 kg N ha-‘, and the incorporation of the groundnut residues increased maize grain yields on average from 1930 to 2830 kg ha-’ and total shoot N content from 35 to 51 kg N ha-‘, equivalent to the addition of 75 kg N ha-’ as urea.
Naturally, much larger contributions of N are to be expected from crops planted solely as green man- mes and a wide range of legumes have been used for a wide range of subsequent crops (Lathwell, 1990). One problem is that legume residues can decompose quickly as soon as they are incorporated into the soil and may release organic acids or other compounds detrimental to germination and seedling growth of the following crop. Furthermore, a large proportion on the N in green manure residues may be released before the roots of the next crop are established to absorb it (Cornforth and Davis, 1968; Yadvinder-Singh et al., 1992) and large losses of N via leaching and even volatilisation from such residues have been reported (Costa et al., 1990; Janzen and McGinn, 1991). There are several sol- utions to this problem; one is to use legumes with a high fibre, lignin or polyphenol content which degrade slowly, releasing N at the time when the subsequent crop requires it. The rate of release of N from crop residues is closely related to their C- to-N ratio and content of lignin and polyphenol such that a close linear relationship of N release (mineralisation) with the (lignin + polyphenol)-to- N ratio has been reported by several authors (Fox et al., 1990; Thomas and Asakawa, 1993; Handayanto et al., 1994). In al1 cases, however, it is necessary that farmers are willing to invest time or money on planting a crop with no immediate econ- omic return and this is rarely the case if they can use the land for a grain or fodder crop at this time. Better acceptance is likely in regions where it is too dry for a grain or forage crop to be planted. Furthermore, if seeds of the green manure are cheap and readily available and planting costs are very low, this wil1 also increase the chances of adoption. The use of oats as the winter crop utilised in the zero tillage system described above, was easily adopted for these reasons, and planting is achieved by just scattering the seeds on the soil sur- face, with no tillage or fertiliser. Suitable drought- resistant legume manures may include pigeon pea (Cajanas cajan) or certain Stylosanthes ecotypes although these crops are not suitable for seeding on the soil surface.
Intercropping is regarded as another alternative method for supplying a cereal crop with N fixed by the accompanying legume. While intercropping often makes more efficient use of available land, and greater yield stability (Willey, 1979), the evi- dence that the cereal crop can benefit from a contri-

BNF for sustainable tropical agriculture 795
bution of biologically fixed N derived from the intercropped legume is often weak. Tropical grasses and cereals usually compete much better for soil mineral N than legumes, hence on an area basis the yield and N accumulation of the cereal crop may be higher in the intercrop than in monoculture, but this is often at the expense of the legume. In this case the legume wil1 often compensate for the lack of soil N which was taken up by the cereal, by increasing its contribution of BNF (Rerkasem and Rerkasem, 1988). The evidente that there is transfer of N from cereal (or grass) to legume in intercrops (or mixed pastures), has largely been derived from the use of the “N isotope dilution technique, where it must be assumed that the “N enrichment of soil N accumulated by the cereal or grass crop is the same when it is in monoculture as when in mixture with the legume. Unless the soil N is completely uniformly labelled with “N both in space and with time, this is unlikely to be the case (Papastylianou, 1988; Abaidoo and Van Kessel, 1989; Chalk and Smith, 1994). Direct methods of measuring N trans- fer from legume to grass by labelling legume leaves with “N have failed to show very significant N transfer either in pastures (Ledgard et al., 1985) or a bean plus maize intercrop (Giller et al., 1991). While N transfer from legume to intercropped cer- eal may be negligible, if the legume is not adversely affected by shading or competition for nutrients other than N, and fixes more nitrogen than it would do in monoculture, then the overall BNF contribution to the legume plus cereal intercrop is greater than if the two crops had been grown separ- ately.
Wetland rite
Wetland rite is the most important cereal crop in the Third World. It is not surprising therefore that a great deal of research has been performed on green manures for this crop at the International Rite Research Institute (IRRI, Philippines) as wel1 as many other institutions. Within the term green manures can be included cyanobacteria and Azolla as wel1 as legumes.
Cyanobacteria (blue-green algae) occur naturally in wetland rite fields and sometimes form very evi- dent blooms. At maximum such blooms were reported to have 480 kg of dry matter ha-’ and contain between 10 and 20 kg N although N turn- over may be considerably higher than these figures (Roger and Reynaud, 1982; Roger and Watanabe, 1986). Some considerable effort has been expended, especially in India, to develop inoculation of cyano- bacteria as a technology to supply biologically fixed N to rite (Venkataraman, 1981). This program in India had sporadic results sometimes because of poor inoculant quality and often because native populations of cyanobacteria were not replaced by
the inoculated strains, or the latter were no more effective (Giller and Wilson, 1991).
The use of the Nz-fixing water fern Atolla under experimental conditions has showed that it can ac- cumulate from 40 to 90 kg N ha-’ in a period of 30 to 46 days (Watanabe, 1982) of which more than 80% is derived from BNF (Yoneyama et al., 1987; Watanabe et al., 1991). Several network studies have been made on the use of Azolla in both Asia and elsewhere (Watanabe, 1982; Kumarasinghe and Eskew, 1993). For 12 sites in Asia the growth of Azolla before or after transplanting of the rite, with or without incorporation, was shown to increase rite yields on average from 2.6 to 3.1 or 3.2 t ha-‘, equivalent to the addition of 30 kg N ha-’ as urea. The efficiency of utilisation of the Azolla-N has been shown to be very similar to that of urea but it is difficult to find a method of producing much more than 15 to 20 t ha-’ of Azolla (fresh weight) which contain 30 to 40 kg N within the time and space available in normal wetland rite cropping. The main limiting agronomie factors for the use of Azolla are the necessity for good phosphorus avail- ability, insect pests and inhibition of growth by high temperatures. In a few areas, such as the region of South Cotabato in the Philippines, where soils have sufficient available P and there is little insect attack, it has been reported to be a cheaper source of N than urea (Kikuchi et al., 1984) but most rite growing areas of the tropics have P-de- ficient soils so that only where N fertiliser is not available, or very expensive, is the use of Azolla likely to be adopted. Its use has been traditional in some regions of China and Vietnam, but in recent years the area where Azolla is used in these countries has been decreasing. The reasons for this are wel1 explained by Roger and Watanabe (1986) and include the increasing availability of mineral N fertilisers, difficulties in maintaining and distributing inoculum from season to season and the labour- intensive nature of this technology. One other ad- vantage of the use of Azolla has recently been reported: the presence of a dense mat of Azolla on the surface of the flood water has been shown to lower surface pH, and has been shown to reduce losses of fertiliser and soil N via ammonia volatilis- ation and increase N fertiliser recovery (Kumarasinghe and Eskew, 1991; Vlek, 1995).
There is a considerable tradition of the use of legume green manures in several rite-growing areas of the World. In the cooler regions of China the dryland legume Astragalus sinicus was planted after rite and then the residues ploughed in at the time of land preparation for the next rite crop. In war- mer parts of the tropics, forage legumes such as Calopogonium SPP, siratro, Clitoria SPP> Desmodium or Stylosanthes have been used (Beri et al., 1989; Becker et al., 1995). To use these legumes it is necessary that the farmer does not wish to uti-
796 Robert M. Boddey et al.
lise the land for another grain or cash crop outside of the rite-growing season, and is content to only use the area for grazing. With the ever increasing pressure on land, and increasing availability of N fertiliser, it is not surprising that the area of rite where such green manure legumes are used has decreased considerably in recent years. There are a few studies in the tropics which show that such rite yields can be increased from approximately 3 t ha-’ to 5 or 6 t ha-’ by the incorporation of green man- ures such as sun-hemp (Crotalaria juncea), cowpea or mung bean, if the farmer can spare 40 to 60 days of his growing season before transplanting his rite (Morris et al., 1986; Beri et al., 1989).
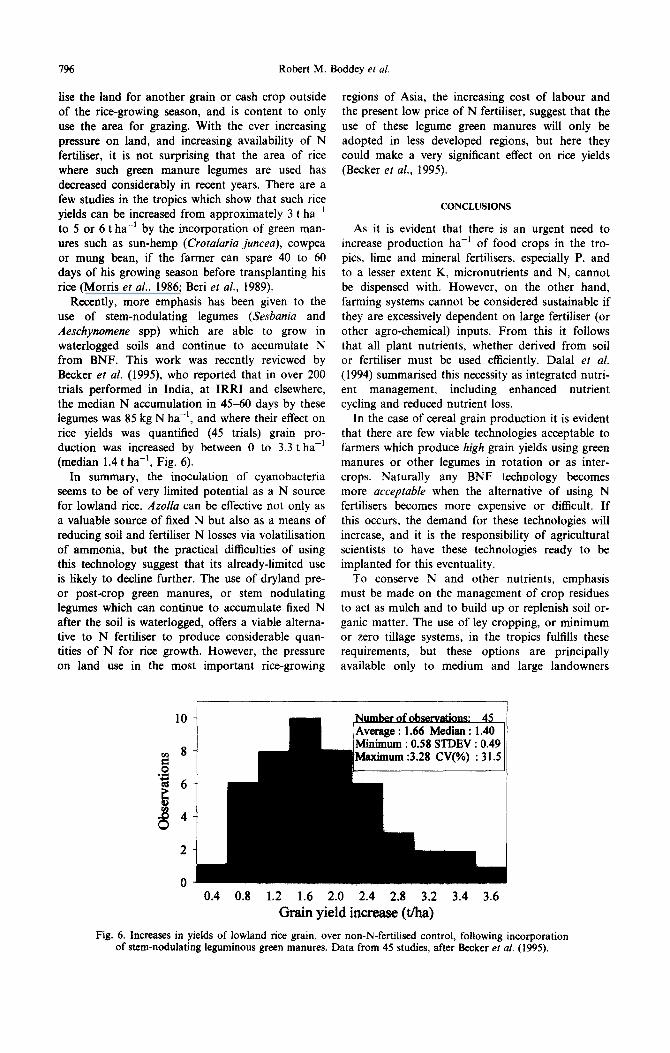
Recently, more emphasis has been given to the use of stem-nodulating legumes (Sesbania and Aeschynomene spp) which are able to grow in waterlogged soils and continue to accumulate N from BNF. This work was recently reviewed by Becker et al. (1995), who reported that in over 200 trials performed in India, at IRRI and elsewhere, the median N accumulation in 45-60 days by these legumes was 85 kg N ha-‘, and where their effect on rite yields was quantified (45 trials) grain pro- duction was increased by between 0 to 3.3 t ha-’ (median 1.4 t ha-‘, Fig. 6).
In summary, the inoculation of cyanobacteria seems to be of very limited potential as a N source for lowland rite. Azolln can be effective not only as a valuable source of fixed N but also as a means of reducing soil and fertiliser N losses via volatilisation of ammonia, but the practica1 difficulties of using this technology suggest that its already-limited use is likely to decline further. The use of dryland pre- or post-crop green manures, or stem nodulating legumes which can continue to accumulate fixed N after the soil is waterlogged, offers a viable alterna- tive to N fertiliser to produce considerable quan- tities of N for rite growth. However, the pressure on land use in the most important rite-growing
regions of Asia, the increasing tost of labour and the present low price of N fertiliser, suggest that the use of these legume green manures wil1 only be adopted in less developed regions, but here they could make a very significant effect on rite yields (Becker et al., 1995).
CONCLUSIONS
As it is evident that there is an urgent need to increase production ha-’ of food crops in the tro- pits, lime and mineral fertilisers, especially P, and to a lesser extent K, micronutrients and N, cannot be dispensed with. However, on the other hand, farming systems cannot be considered sustainable if they are excessively dependent on large fertiliser (or other agro-chemical) inputs. From this it follows that al1 plant nutrients, whether derived from soil or fertiliser must be used efficiently. Dalal et al. (1994) summarised this necessity as integrated nutri- ent management, including enhanced nutrient cycling and reduced nutrient 10s~.
In the case of cereal grain production it is evident that there are few viable technologies acceptable to farmers which produce high grain yields using green manures or other legumes in rotation or as inter- crops. Naturally any BNF technology becomes more acceptable when the alternative of using N fertilisers becomes more expensive or difficult. If this occurs, the demand for these technologies wil1 increase, and it is the responsibility of agricultural scientists to have these technologies ready to be implanted for this eventuality.
To conserve N and other nutrients, emphasis must be made on the management of crop residues to act as mulch and to build up or replenish soil or- ganic matter. The use of ley cropping, or minimum or zero tillage systems, in the tropics fulfills these requirements, but these options are principally available only to medium and large landowners
10
0 0.4 0.8 1.2 1.6 2.0 2.4 2.8 3.2 3.4 3.6
Grain yield increase (tha) Fig. 6. Increases in yields of lowland rite grain, over non-N-ferti1ise.d control, following incorporation
of stem-nodulating leguminous green manures. Data from 45 studies, after Becker er al. (1995).

BNF for sustainable tropical agriculture 191
cal forage-legumes at two levels of P:K supply. Field Crops Research 22, 181-194.
Cadisch G., Carvalho E. F., Suhet A. R., Vilela L., Soares W., Spain J. M., Urquiaga S., Giller K. E. and Boddey R. M. (1994a) The importante of legume Nz-fixation in sustainability of pastures in the cerrados of Brazil. In Proceedings of the International Grassland Congress, pp. 1915-1918. Palmerston North, New Zealand.
Cadisch G., Schunke R. M. and Giller K. E. (1994b) Nitrogen cycling in a pure grass pasture and a grass- legume mixture on a red latosol in Brazil. Tropical Grasslands 28, 43-52.
Cantarutti R. B., Braga J. M., Boddey R. M. and Resende C. de P. (1995) Caracterizaçäo do status de nitrogènio em solo sob pastagem de Brachiaria humidicola pura e consorciada com Desmodium ovalifolium CV. Itabela. Proceedings of the XXV Congresso Brasileiro do Ciência do Solo, pp. 733-735, Viçosa, MG, Brazil, 23-29 July.
Chalk P. M. and Smith C. J. (1994) 15N isotope dilution methodology for evaluating the dynamics of biologically fixed N in legume-non-legume associations. Biology and Fertility of Soils 17, 80-84.
Cho& T., Andreux F., Correa J. C., Volkoff B. and Cerri C. C. (1991) Changes in organic matter in an oxisol from the Centra1 Amazonian forest during eight years as pasture, determined by 13C isotopie composition. In Diversity of Environmental Biogeochemistry (J. Berthelin, Ed.), pp. 397405. Elsevier, New York.
CIAT (1990) CIAT Tropical Pastures Annual Report 1989, Working Document No. 70, CIAT, Cali.
Cornforth 1. S. and Davis J. B. (1968) Nitrogen transform- ations in tropical soil. 1. The mineralization of nitrogen- rich organic materials added to soil. Tropical Agriculture 45,211-221.
Costa F. J. S. A., Bouldin D. R. and Suhet A. R. (1990) Evaluation of N recovery from mucuna placed on the surface or incorporated in a Brazilian oxisol. Plant and Soill24,91-96.
Dalal R. C., Strong W. M., Weston E. J., Cahill M. J., Cooper J. E., Lehane K. J., King A. J. and Gaffney J. (1994) Evaluation of forage and grain legumes, no-til1 and fertilisers to restore fertility degraded soils. In Transaction of the I5th World Congress of Soil Science, Vol. 5a, pp. 62-74. International Society of Soil Science, Acapulco, Mexico.
Dakora F. D., Aboyinga R. A., Mahama Y. and Apaseku J. (1987) Assessment of Nz-fixation in groundnut (Arachis hypogaea) and cowpea (Vigna unguiculata L. Walp) and their relative N contribution to a succeeding maize crop in Northem Ghana. MZRCEN Journal of Applied Mierobiology and Biotechnology 3, 389-399.
Da Silva J. E., Lemainski J. and Resck D. V. S. (1994) Perdas de matéria orgânica e suas relaçöes com a capa- cidade de troca catiônico em solos da regiäo de Cerrados do oeste Baiano. Revista Brasileira de Ciência do Solo 18, 541-547.
Feigl B. J., Melillo J. and Cerri C. C. (1995) Changes in the origin and quality of soil organic matter after pas- ture introduction in Rondônia (Brazil). Plant and Soil 175,21-29.
Fisher M. J., Rao 1. M., Ayarza M. A., Lascano C. E., Sanz J. I., Thomas R. J. and Vera R. R. (1994) Carbon storage by introduced deep-rooted grasses in the South American savannas. Nature, Londen 371,236-238.
Fox R. H., Myers R. J. K. and Vallis 1. (1990) The nitro- gen mineralization rate of legume residues in soils as influenced by their polyphenol, lignin and nitrogen con- ten&. Plants and Soil 129, 251-259.
Giller K. E. and Cadisch G. (1995) Future benefits from biological nitrogen fixation: An ecological approach to agriculture. Plant and Soill74,255-211.
who have access to fertiliser, pesticides, machinery and an agricultural extension service of some kind. For the small-holders and subsistence farmers of less developed regions of the tropics, govemment neglect of rural areas often means that such exten- sion services are non-existent and even basic agri- cultural inputs such as lime and phosphate fertilisers are difficult to obtain even if farmers can afford them. However, as crop yields in these regions are invariably very low, modest quantities of basic fertiliser inputs to diminish Al toxicity and increase P availability could have dramatic results. Given some degree of infrastructural improvement, the use of crop rotations which supply N from green manures or grain legumes while maintaining or increasing SOM reserves, can improve farm income and food supply on a sustainable basis.
Acknowledgements-The studies on N cycling in pastures were financed by the European Community International Scientific Co-operation Programme and the Fundaçäo Banco do Brasil. RMB and SU gratefully acknowledge research fellowships form the Brazilian National Research Council (CNPq).
REFERENCES
Abaidoo R. C. and Van Kessel C. (1989) “N-uptake, Nz- fixation and rhizobial interstrain competition in soybean and bean, intercropped with maize. Soil Biology and Biochemistry 21, 155-159.
Becker M., Ladha J. K. and Ali M. (1995) Green manure technology: Potential, usage, and limitations. A case studv for lowland rite. Plant and Soi1174. 181-194.
Beri V:, Meelu 0. P. and Khind C. S. (1986) Biomass pro- duction, N accumulation, symbiotic effectiveness and mineralization of green manures in relation to yield of wetland rite Tropical Agriculture 66, 11-16.
Boddey R. M. and Victoria R. L. (1986) Estimation of biologica nitrogen fixation associated with Brachiaria and Paspalum grasses using ‘SN-labelled organic matter and fertilizer. Plant and Soil90, 265-292.
Boddey R. M., Resende C. de P., Pereira J. M., Cantarutti R. B., Alves B. J. R., Ferreira E., Richter M., Cadisch G. and Urquiaga S. (1995) The nitrogen cycle in pure grass and grass/legume pastures: Evaiuation of pasture sustainability. In Nuclear Techniques in Soil-Plant Studies for Sustainable Agriculture and Environmental Preservation, pp. 307- 319. FAO/IAEA, Vienna, Austria.
__
Boddey R. M., Rao 1. M. and Thomas R. J. (1996) Nutrient cycling and environmental impact of Brachiaria pastures. In Brachiaria: The Biology, Agronomy and Immovement (J. W. Miles, B. L. Maass and C. B. do Ville, Eds), pp. 72-86. Publication 259, CIAT, Cali Colombia.
Borlaug N. E. and Dowswell C. R. (1994) Feeding a human population that increasingly crowds a fragile planet. In Keynote Lecture Supplement to the Transactions of the 15th World Congress of Soil Science. International Society of Soil Science, Acapulco, Mexico.
Buschbacher R., Uhl C. and Serräo E. A. S. (1988) Abandoned pastures in the eastem Amazonia. 11. Nutrient stocks in the soil and vegetation. Journal of Ecology 76, 682-699.
Cadisch G., Sylvester-Bradley R. and Nösberger J. (1989) 15N-based estimation of nitrogen fixation by eight tropi-

798 Robert M. Boddey et al
Giller K. E. and Wilson K. J. (1991) Nitrogen Fiwation tn Tropical Cropping Systems. CAB International, Wallingford.
Giller K. E., Ormesher J. and Awah F. M. (1991) Nitrogen transfer from Phaseolus bean to intercropped maize measured using “N-enrichment and “N-isotope dilution methods. Soil Biology and Biochemistry 23, 339-346.
Giller K. E., McDonagh J. F. and Cadisch G. (1994) Can biological nitrogen fixation sustain agriculture in the tropics? In Soil Science and Sustainable Land Management in the Tropics (J. K. Syers and B. L. Rimmer, Eds), pp. 173-191. CAB International, Wallingford.
Greenland D. J. (1975) Bringing the green revolution to the shifting cultivator. Science 190, 841-844.
Handayanto E., Cadisch G. and Giller K. E. (1994) Nitrogen release from prunings of legume hedgerow trees in relation to quality of the prunings and incu- bation method. Plant and Soil 160, 237-248.
Janzen H. H. and McGinn S. M. (1991) Volatile loss of nitrogen during decomposition of legume green manure. Soìl Biology and Biochemìstry 23, 29 1-297.
Jensen E. S. and Castellanos J. 2. (1994) The role of grain legumes in nitrogen cycling of low input sustainable agro-ecosystems, In Transactions of the 15th World Congress of Soil Science, Vol. 5a. pp. 32245. International Society of Soil Science. Acapulco, Mexico.
Jones R. K., Dalgliesh N. P., Dimes J. P. and McCown R. L. (1991) Sustaining multiple production systems 4. Ley pastures in crop-livestock systems in the semi-arid tropics. Tropical Grasstands 23, I89- 196.
Kikuchi M., Watanabe 1. and Haws L. D. (1984) Economie evaluation of Asolla use in rite production. In Organic Matter and Rite. pp. 5699592. IRRI, Los Baiìos, Philippines.
Kluthcouski J., Pacheco A. R., Teixeira S. M. and Oliveira E. T. (1991) Renovaçäo de pastagens do Cerrado com arroz. I.-Sistema Barreiräo.. Boletim Técnico No. 33. EMBRAPA-CNPAF. Goiânia, GO, Brazil.
Kumar Rao J. V. D. K., Dart P. J. and Sastry P. V. S. S. (1983) Residual effect of pigeonpea (Cajanus cajan) on yield and nitrogen response of maize. Experimental Agriculture 19, 131-141.
Kumarasinghe K. S. and Eskew D. L. (Eds) (1993) Isotopic Studies of Azolla and Nitrogen Fertilization of Rite. Kluwer Academie Publishers, Dordrecht.
Lathwell D. J. (1990) Legume green manures: principles for management based on recent research, Trop Soils Bulletin, No. 90-01. North Carolina State University, NC, USA.
Ledgard S. F., Freney J. R. and Simpson J. R. (1985) Assessing nitrogen transfer from legumes to associated grasses. Soil Biology and Biochemistry 17, 575-577.
Lloyd D. L., Smith K. P., Clarkson N. M., Weston E. J. and Johnson B. (1991) Sustaining multiple pro- duction systems 3. Ley pastures in the subtropics. Tropical Grasslands 23, 181-188.
Macedo M. C. M. (1995) Pastagens no ecossistema Cerrados: Pesquisa para o desenvolvimento sustentável. In Anais do Simpósio sobre Pastagens nos Ecosistemas Brasileiros: Pesquisas Para 0 Desenvolvimento Sustentavel (R. P. de Andrade, A. de 0. Barcellos and C. M. C. da Rocha, Eds), pp. 28-62. Sociedade Brasileira de Zootecnia, Universidade Federal de Viçosa, Viçosa, MG, Brazil.
McDonagh J. F., Toomsan B., Limpinuntana V. and Giller K. E. (1993) Estimates of the residual nitrogen benefit of groundnut to maize in Northeast Thailand. Plant and Soil154, 267-277.
Miranda C. H. B. and Boddey R. M. (1987) Estimation of biological nitrogen fixation associated with 11 ecotypes of Panicum maximum grown in nitrogen-15 labeled soil. Agronomy Journal79, 558-563.
Morris R. A., Furoc R. E. and Dizon M. A. (1986) Rite responses to a short-duration green manure 1. Grain yield. Agronomy Journal78, 409-412.
Papastylianou 1. (1988) The “N methodology in estimat- ing NS fixation by vetch and pea grown in pure stand or in mixtures with oat. Plant and Soil 107, 183-188.
Pereira J. M., Nascimento D. Jr, Santana J. R., Cantarutti R. B. and Leäo M. 1. (1992) Teor de proteína bruta e digestibildade in vitro de matéria seca da forragem disponível e da dieta selecionada por bovinos em pasta- gen de Brachiaria humidicola (Rendle) Schweickt, em monocultivo ou consorcioado com leguminosas, subme- tidas a diferentes taxas de lotaçäo. Revista da Sociedade Brasileira de Zootecnia 21, 104-117.
Rerkasem K. and Rerkasem B. (1988) Yields and nitrogen nutrition of intercropped maize and ricebean (Vigna umbellata [Thunb.] Ohwi and Ohashi). Plant and Soil 108, 151-162.
Robertson F. A., Myers R. J. K. and Saffigna P. G. (1993a) Distribution of carbon and nitrogen in a long-term cropping system and permanent pasture sys- tem. Australian Journal of Agricultural Research 44, 1323-1336.
Robertson F. A., Myers R. J. K. and Saffigna P. G. (1993b) Carbon and nitrogen mineralisation in culti- vated and grassland soils in subtropical Queensland. Australian Journal of Soil Research 31, 611-619.
Robbins G. B., Bushell J. J. and McKeon G. M. (1989) Nitrogen immobihzation in decomposing litter contrib- utes to productivity decline in ageing pastures of green panic (Panicum maximum var. trichoglume). Journal of Agricultural Science 113, 40 1406.
Roger P. A. and Reynaud P. A. (1982) Free-living blue- green algae in tropical soils. In Microbiology of Tropical Soils and Plant Productivity (Y. R. Dommergues and H. G. Diem, Eds), pp. 147-168. Martinus Nijhoff, The Hague.
Roger P. A. and Watanabe 1. (1986) Technologies for uti- hzing biological nitrogen fixation in wetland rite: poten- tialities, current usage and limiting factors. Fertilizer Research 9, 39-77.
Sá, J. C. M. (1993) Manejo da fertilidade do solo no plan- tio direto. Boletim da Fundaçäo ABC, Castro, Paraná, Brazil. 96~.
Santana J. R. de and Pereira J. M. (1995) Efeito do pas- tejo sobre a persistência, produçäo e qualidade da con- sorciaçäo Brachiaria dictyoneura Stapf. CIAT 6133 e Arachis repens. Relatório CEPECICEPLAC. Itabuna, BA, Brazil.
Scholes M. C., Swift M. J., Heal 0. W., Sanchez P. A., Ingram J. S. 1. and Dalal R. (1994) Soil fertility research in response to the demand for sustainability, In The Management of Tropical Soil Feríility (P. L. Woomer and M. J. Swift, Eds), pp. 1-14. John Wiley, Chichester.
Schunke R. M., Cadisch G., Santos J. C.- C. dos and Boddev R. M. 11992) Mineralizacäo da matéria orsä- . , nica do solo em pastagem de Brachiaria decumbens adu- bada com fósforo, In Anais la Reunión Sabanas, Red International de Evaluacìón de Pastos Tropicales (RIEPTJ (E. A. Pizarro, Ed.), pp. 455458. Documento de Trabalho No. 117., CIA?, Cah.
Seiffert N. M.. Zimmer A. H.. Schunke R. M. and Miranda C. H. B. (1985) Reciclagem de nitrogênio em pastagem consorciada de Calopogonium mucunoides com Brachiaria decumbens. Pesquisa Agropecuária Brasileira 20, 5299544.
Spain J. M. and Gauldron R. (1991) Degradacion y reha- bilitacion de pasturas. In Establecimiento y Renovacion

BNF for sustainable tropical agriculture 199
de Pasturas: Conceptos. Experiencias y Enfoque de la Investigacion (C. E. Lascano and J. M. Spain, Eds), pp. 269-283. Red International de Evaluacion de Pastos Tropicales-Sexta Reunion del Comite Asesor. CIAT, Cali.
Spain J. M., Leite G., Vilela L., Gomide C. and Rocha C. M. da (1994) The effect of grazing systems and legumes on productivity and stability of pastures in the Brazilian Cerrados. In Proceedings of the International Grassland Congress. Palmerston North, New Zealand, 1993.
Thomas R. J. and Asakawa N. M. (1993) Decomposition of leaf litter from tropical forage grasses and legumes. Soil Biology and Biochemistry 25, 1351-1361.
Thomas R. J., Fisher M. J., Ayarza M. A. and Sanz J. 1. (1995) The role of forage grasses and legumes in maintaining the productivity of acid soils in Latin America. In Soil Management-Experimental Basis for Sustainability and Environmental Quality (R. Lal and B. A. Stewart, Eds), pp. 61-83. CRC Press, Boca Raton, Florida, USA.
Venkataraman G. S. (1981) Blue-green algae for rite pro- duction: a manual for its promotion. Soils Bulletin, 46, FAO, Rome.
Vera R. R., Thomas R. J., Sanint L. R. and Sanz J. 1. (1992) Development of sustainable ley-farming sys- tems for the acid soil savannas of tropical America.
Anais da Academia Brasileira de Ciências 64(l), 105- 125.
Vlek P. L. G., Diakite M. Y. and Mueller H. (1995) The role of Azolla in curbing ammonia volatilization from flooded rite systems. Fertilizer Research 42, 165-174.
Watanabe 1. (1982) Azolla-Anabaena symbiosis-its physi- ology and use in tropical agriculture. In Microbiology of Tropical Soils and Plant Productivity (Y. R. Dommergues and H. G. Diem, Eds), pp. 1688185. Martinus Niijhoff, The Hague.
Watanabe I., Yoneyama T., Talukdar H. and Ventura W. (1991) The contribution of atmospheric N2 to Azolla spp grown on flooded soil. Soil Science and Plant Nutrition 37, 101-110.
Willey R. W. (1979) Intercropping-its importante and research needs. Part 1, Competition and yield advan- tages. Field Crop Abstracts 32, 1-10.
Yadvinder-Singh, Bijay-Singh and Khind C. S. (1992) Nutrient transformations in soils amended with green manures. Advances in Soil Science 20, 237-309.
Yoneyama T., Ladha J. K. and Watanabe 1. (1987) Nodule bacteroids and Anabaena: natura1 “N enrich- ment in the legume-Rhizobium and Azolla-Anabaena symbiotic systems. Journal of Plant Physiology 127, 251-259.




















