
The complex Robertsonian system of Dichroplus pratensis (Melanoplinae, Acrididae). II. Effects of...
15
Heredity 64 (1990) 145—159 Received 31 May 1989 The Genetical Society of Great Britain The complex Robertsonian system of Dichroplus pratensis (Melanoplinae, Acrid idae). II. Effects of the fusion polymorphisms on chiasma frequency and distribution Claudio J. Bidau*t Laboratorio de Genética, Departmento de Ciencias Biológicas, Facultad de Ciencias Exactas y Naturales, Universidad de Buenos Aires, Ciudad Universitaria, 1428 Buenos Aires, Repüblica Argentina. The effects of a complex series of Robertsonian polymorphisms on chiasma frequency and distribution were analysed in natural populations of the grasshopper Dichroplus pratensis which has a standard karyotype of 2n =19 (XOd) telocentrics. Populations are usually polymorphic for one to three of seven distinct fusions between the six large (L1—L6) autosomes. The study revealed that: 1. Standard males have an essentially proximal-distal pattern of chiasma distribution. Interstitial chiasmata are less frequent. 2. Fused males have significantly fewer total and proximal chiasmata than standards from the same population but show an increase in interstitial tnd distal chiasmata. 3. The former is due to the fact that centric fusions, either homozygons or heterozygous, produce a decrease in total chiasma frequency of the involved chromosomes and a redistribution of chiasmata resulting in a sharp reduction of proximal chiasmata and an itccse in interstitial and distal ones. In this respect, there are no significant differences between fusion trivalents and corresponding fusion bivalents. The same chromosomes in the basic homozygous state, have a typical standard chiasma pattern. 4. The effects of each fusion are intrachromosomal and although the same chiasma repatterning occurs in all seven fusions, total reduction in chiasma frequency depends on the telocentrics involved since a highly significant positive correlation exists between telocentric length and chiasma frequency. Since all chromosome arms tend to have a minimum of one chiasma, the effects of a fusion are more marked in longer autosomes. 5. Comparisons of populations that differ for number and frequency of fusions, showed a highly significant negative correlation between mean number of different fusions per male (per population) (F) and total chiasma frequency per cell (Xta). The same type f correlation exists between F and proximal Xta and F and between-cell variance of totalX. frequency, while a positive correlation was found between F and mean interstitial chiasmata. 6. According to 5, intrachromosomal recombination within populations decreases as number and frequency of fusions increase. This effect is added to the instant reduction of interchromosomal recombination produced by the combination of two linkage groups into one. These results suggest that redistribution of chiasma patterns is due to a direct effect of the rearrangements. Furthermore, the changes in chiasma patterns are discussed in relation to the maintenance of the polymorphisms since trivalents require the elimination of proximal and interstitial chiasmata for balanced orientation and segregation. This was corroborated by the finding of a population in which three fusions were fixed. Here, proximal and interstitial chiasmata in the fused chromosomes are significantly more frequent than expected. Finally, the modifications of recombination potential are discussed relative to the possible adaptive role of these widespread polymorphisms and to a model of chromosomal evolution for the species. * Member of the Carrera del Investigador CientIfico y Tec- INTRODUCTION nológico, CONICET. t Present address: Departamento de Genética Facultad de Ciencias Exactas, Qulmicas y Naturales, Universidad Nacional The effects of chromosomal rearrangements on de Misiones, Felix de Azara 174, 3300 Posadas, Misiones, chiasma frequency and localisation have been Repüblica Argentina. extensively studied and reviewed in animals and
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of The complex Robertsonian system of Dichroplus pratensis (Melanoplinae, Acrididae). II. Effects of...

Heredity 64 (1990) 145—159 Received 31 May 1989The Genetical Society of Great Britain
The complex Robertsonian system ofDichroplus pratensis (Melanoplinae,Acrididae).II. Effects of the fusion polymorphismson chiasma frequency and distributionClaudio J. Bidau*t Laboratorio de Genética, Departmento de Ciencias
Biológicas, Facultad de Ciencias Exactas y Naturales,Universidad de Buenos Aires, Ciudad Universitaria,1428 Buenos Aires, Repüblica Argentina.
The effects of a complex series of Robertsonian polymorphisms on chiasma frequency and distribution were analysed innatural populations of the grasshopper Dichroplus pratensis which has a standard karyotype of 2n =19 (XOd)telocentrics. Populations are usually polymorphic for one to three of seven distinct fusions between the six large(L1—L6) autosomes. The study revealed that:1. Standard males have an essentially proximal-distal pattern of chiasma distribution. Interstitial chiasmata are lessfrequent.2. Fused males have significantly fewer total and proximal chiasmata than standards from the same population butshow an increase in interstitial tnd distal chiasmata.3. The former is due to the fact that centric fusions, either homozygons or heterozygous, produce a decrease in totalchiasma frequency of the involved chromosomes and a redistribution of chiasmata resulting in a sharp reduction ofproximal chiasmata and an itccse in interstitial and distal ones. In this respect, there are no significant differencesbetween fusion trivalents and corresponding fusion bivalents. The same chromosomes in the basic homozygous state,have a typical standard chiasma pattern.4. The effects of each fusion are intrachromosomal and although the same chiasma repatterning occurs in all sevenfusions, total reduction in chiasma frequency depends on the telocentrics involved since a highly significant positivecorrelation exists between telocentric length and chiasma frequency. Since all chromosome arms tend to have aminimum of one chiasma, the effects of a fusion are more marked in longer autosomes.5. Comparisons of populations that differ for number and frequency of fusions, showed a highly significant negativecorrelation between mean number of different fusions per male (per population) (F) and total chiasma frequency percell (Xta). The same type f correlation exists between F and proximal Xtaand F and between-cell variance of totalX.frequency, while a positive correlation was found between F and mean interstitial chiasmata.6. According to 5, intrachromosomal recombination within populations decreases as number and frequency of fusionsincrease. This effect is added to the instant reduction of interchromosomal recombination produced by the combinationof two linkage groups into one.
These results suggest that redistribution of chiasma patterns is due to a direct effect of the rearrangements.Furthermore, the changes in chiasma patterns are discussed in relation to the maintenance of the polymorphisms sincetrivalents require the elimination of proximal and interstitial chiasmata for balanced orientation and segregation. Thiswas corroborated by the finding of a population in which three fusions were fixed. Here, proximal and interstitialchiasmata in the fused chromosomes are significantly more frequent than expected. Finally, the modifications ofrecombination potential are discussed relative to the possible adaptive role of these widespread polymorphisms and to amodel of chromosomal evolution for the species.
* Member of the Carrera del Investigador CientIfico y Tec- INTRODUCTIONnológico, CONICET.t Present address: Departamento de Genética Facultad deCiencias Exactas, Qulmicas y Naturales, Universidad Nacional
The effects of chromosomal rearrangements onde Misiones, Felix de Azara 174, 3300 Posadas, Misiones, chiasma frequency and localisation have beenRepüblica Argentina. extensively studied and reviewed in animals and

146 C. J. BIDALJ
plants. Instances of B chromosomes, interchanges,Robertsonian translocations, inversions and super-numerary heterochromatin affecting chiasmataand recombination, have been reported. Intra- andinterchromosomal effects have been demon-strated (Hewitt, 1979; Parker et a!., 1982; Sperlichand Pfriem, 1986; White, 1973).
A distinction must be made however, betweenthe effects of spontaneous and polymorphicrearrangements since in many cases, the survivaland establishment of a stable polymorphic condi-tion or conversely, its transience and the con-sequent elimination or fixation of a given karyo-morph may depend on a reorganisation of thechiasma patterns in the involved chromosomes.Such is the case of centric fusions. In a smallnumber of cases of spontaneous fusions studiedin grasshoppers, no modifications of chiasma pat-terns were apparent in the resultant trivalents(Colombo, 1987; Kayano and Nakamura, 1960;Lopez-Fernandez et a!., 1984; Southern, 1967;Teoh and Yong, 1983). However, in the few casesof fusion polymorphisms studied in depth, therearranged chromosomes showed modifications inchiasma frequency and localisation which allowstable convergent orientation and normal segrega-tion of trivalents in heterozygotes (Bidau, 1984;Bidau and Hasson, 1984; Bidau and Mirol, 1988;Colombo, 1987, 1988; Hewitt and Schroeter, 1968).In some cases however, chromosomes may be pre-adapted to exist in a polymorphic state if they havelow frequencies and distal localisation of chias-mata as in the Coleopteran Chilocorus (Smith andVirkki, 1978). However, this is not always so sincetrivalents of hybrids between C. tricyclus and C.hexacyclus show high frequencies of non-conver-
gent orientation despite strong distal localisationof chiasmata (Smith, 1966).
The modifications mentioned above have anadditional secondary effect which is the reductionof intrachromosomal (as well as interchromo-somal) recombination and this may be ofevolutionary significance since it could allow themaintenance of adaptive supergenes.
The grasshopper Dichroplus pratensis Bruneroffers a unique opportunity to study these effects.This South American melanopline is widespreadin Argentina, Uruguay and Southern Brazil(Liebermann, 1963) and comprises severalchromosomal races that differ with respect to num-ber and frequency of at least seven different fusionsthat involve six pairs of large autosomes. With asingle exception, all populations already sampledare polymorphic for one to three fusions. Karyo-types, geographic distribution of polymorphismsand meiotic behaviour of heterozygotes havealready been described (Bidau, 1984, 1988; Bidauand Mirol, 1988).
In this paper the relationship of the fusionpolymorphisms of D. pratensis to changes inchiasma frequency and localisation are studied inseveral Argentine populations.
MATERIAL AND METHODS
This study is based on 297 males of D. pratensiscollected by the author and collaborators at thelocalities shown in table 1. A much larger numberof males has been cytologically studied, especiallyat Sierra de la Ventana, being the subject of forth-coming papers; however, some data of these are
Table 1 Origin of the populations whose chiasma characteristics are described in table VII. All populations are polymorphic forthe involved fusions except C in which three fusions have become fixed. Populations K and SV belong to hybrid zones whichresult from the overlapping of chromosome races that differ for fusions with monobrachial homologies
Locality Department Province Collection date Centric fusionsPer centstandard males
Gaiman (G) Biedma Chubut Jan '83 5/6 930P. Madryn (PM) Biedma Chubut Jan '83 1/4, 5/6 770Km 784 (K) Villarino B. Aires Feb '83 1/6, 3/4, 5/6 150El Condor (EC) A. Alsina Rio Negro Jan '83 2/4, 5/6 40Tortuguitas (T) G. Sarmiento B. Aires Feb '82 1/6, 3/4 00Necochea (N) Necochea B. Aires Jan '85 1/6, 3/4 00Balcarce (B) Balcarce B. Aires Jan '85 1/6, 3/4 00La LoberIa (LL) A. Alsina RIo Negro Jan '83 2/4, 5/6 0.0Tandil 1 (Ta 1) Tandil B. Aires Jan '85 1/6, 3/4 0•0Tandil 2 (Ta 2) Tandil B. Aires Jan '85 1/6, 3/4 00Sierra de la Ventana (SV) Tornquist B. Aires Feb '83 1/2, 3/4, 5/6, 1/6 00La Florida (LF) Pringles San Luis Jan '84 1/6, 3/4, 2/5 0.0Monte Hermoso (MH) C. Dorrego B. Aires Jan '83 1/2, 3/4, 5/6 00Cerro (C) Tornquist B. Aires Jan '86 1/2, 3/4, 5/6 00

CENTRIC FUSIONS AND CHIASMA FREQUENCY 147
incorported in this paper. All males were processedin the field. Testes were fixed in 3: 1 alcohol-aceticand squashed in lacto-propionic orcein.
Chiasmata were scored at MI since orientationof bivalents and trivalents at this stage allows easyidentification of chiasma position. Ten cells werescored per male and chiasmata classified asproximal (P), interstitial (I) and distal (D) accord-ing to their location in the first (proximal), secondor third portion of the chromosome or chromo-some arm when the latter are divided in equal parts.
RESULTS
The karyotype and fusion polymorphismsof ft pratensis
The Robertsonian system of the species has alreadybeen described (Bidau, 1984, 1986, 1988). Thekaryotype comprises in its standard form, six pairsof large telocentric autosomes (L1—L6), three smalltelocentrics (S7—S9) of which S7 is the megamericbivalent and S8 carries the single standard NOR,and one ((3) or two (?) medium-sized telocentricX chromosomes. L-L6 chromosomes can beunambiguously recognised by distinctive length.Only L6 can be identified by heterochromatincontent since it usually has a C-positive hetero-chromatic block at its telomeric end. The six Lautosomes are involved in fusions and sevendifferent fusions have been identified (1/2; 1/4;1/6; 2/4; 2/5; 3/4; 5/6). Some populations arepolymorphic for a single fusion, some for two andsome for three although not necessarily the sameones since the same telocentric is involved indifferent fusions. Hybrid zones between chromo-somal races have been identified and within them,popultions may be polymorphic for four fusionswith monobrachial homologies (Bidau, 1988).
Chiasma frequency and localisation in standardand fused males
Until the present, no completely standard (st)population has been found in the sampled areas.Indeed, st individuals are very rare (Bidau, 1988)due to the numbers and frequencies of the fusionspresent. Only two populations showed highfrequencies of St males: Puerto Madryn (PM) andGaiman (0) (table 1), both from Chubut and about80 km apart. The first one is polymorphic forfusions 1/4 and 5/6. In Gaiman, a single 5/6heterozygote was found among 14 males sampled.The standard individuals from these populationswere assumed to display the typical chiasma pat-
terns of non-fused individuals of all populationsstudied although variations between populationsprobably do occur. Nevertheless, st's from PM andG were very similar regarding chiasma frequencyand localisation (table 2) (see also Bidau, 1984).
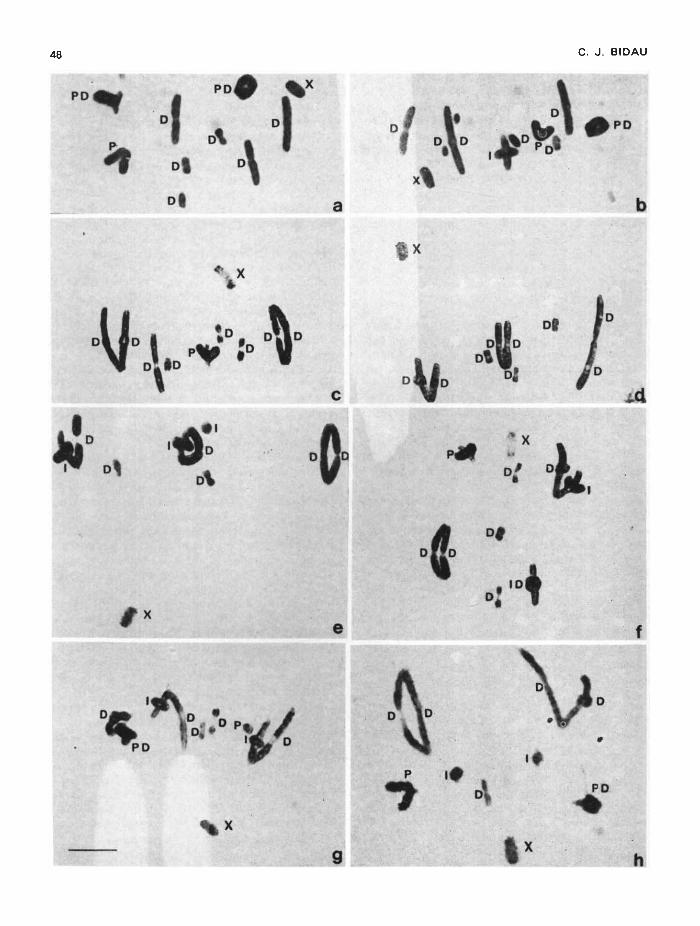
Fig. 1 shows chiasma patterns of several distinctfusion karyomorphs. The S bivalents never formmore than one chiasma that may be P, I or Dalthough the latter predominante (fig. 1 (a)-(h)).Telocentric L bivalents never form more than twochiasmata (fig. 1(a)—(c), (f), (h)). MonochiasmateL bivalents usually have a P or a D chiasma (fig.1(a)—(c), (f), (h)), seldom an I one (fig. 1(b)).Bichiasmate telocentric L bivalents are veryfrequently P/D (fig. 1(a), (b), (h)), sometimes I/D(fig. 1(f)) and only rarely P/I. A summary ofchiasma frequencies and distribution in telocentricL bivalents of standard males is given in table 2.Chiasma conditions are radically altered in meta-centric fused chromosomes as discussed below (fig.1(c)—(h)). There is a highly significant correlationbetween chiasma frequency per bivalent andchromosome length (fig. 2; table 3) within the Lgroup. The correlation does not hold for the Sbivalents since there must always be a chiasmapresent for proper segregation.
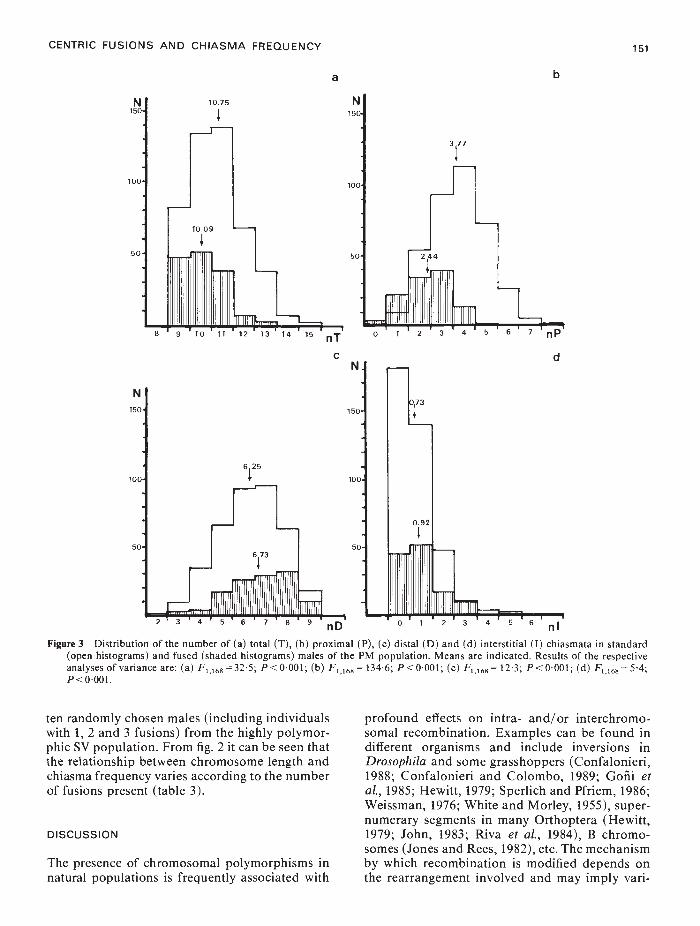
Frequencies of total (T), P, I and D chiasmatain 47 St males from PM are shown in fig. 3(a)-(d).These are not significantly different from those ofthe st's from 0. That is, standards have a mainlyP/D distribution, I chiasmata being rare (Bidau,1984). In PM, 15 males carrying fusions (all ofthem heterozygotes) wre found: nine had the 1/4fusion, five the 5/6 fusion and the remaining onewas a double 1/4, 5/6 heterozygote. These males,when considered jointly showed significantly fewertotal chiasmata than the St males from the samepopulation (fig. 3(a)). This was mainly due to asignificant decrease in P chiasmata (fig. 3(b)). Dand I chiasmata were, on the contrary, higher inheterozygotes (fig. 3(c)— (d)). Chiasma frequenciesand distribution of individual 1-1.4-4 and 5-5.6-6trivalents are summarised in table 4.
In order to test for possible interchromosomaleffects, mean cell chiasma frequencies (X) exclud-ing chromosomes 1 and 4 in one case, and 5 and6 in the other, were calculated in ten randomlychosen non-fused males. The same was done withnine 1/4 and five 5/6 heterozygotes. The chiasmafrequencies thus calculated were not significantlydifferent in both cases (non fused, X =786vs.1/4, X =771; F1189 = 156; P>005; non-fused,X = 830 vs. 5/6, X=856; F1 149= F92; P> 0.05).These results suggest that the effect of the fusionson shiasma frequency are exerted solely upon the
9 %x
•PDDr
4
ba
ix
C DJD
a
D'D3
D%
D DD
144
• UPD•PD
I Dli
4D$ D
I.
D(_JDDtID
x
pa4
eo:I4
I.7I.
0
a
PD
a.
f
h
4
48 C. J. BIDAU
CENTRIC FUSIONS AND CHIASMA FREQUENCY 149
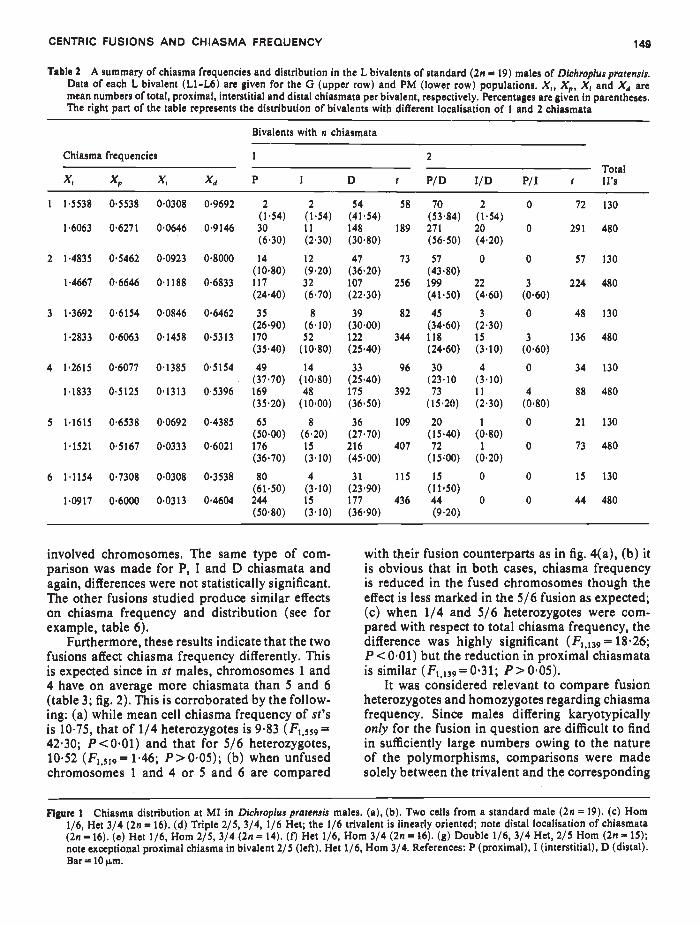
Table 2 A summary of chiasma frequencies and distribution in the L bivalents of standard (2n = 19) males of Dichroplus pratensis.Data of each L bivalent (L1-L6) are given for the G (upper row) and PM (lower row) populations. X, Xi,, X and X4 aremean numbers of total, proximal, interstitial and distal chiasmata per bivalent, respectively. Percentages are given in parentheses.The right part of the table represents the distribution of bivalents with different localisation of 1 and 2 chiasmata
Bivalents with n chiasmata
Chiasnia frequencies 1 2Total
D I P/D l/D P/I I II'sX, X,, X1 X P 1
1 15538 05538 00308 09692 2 2 54 58 70 2 0 72 130
(1'54) (154) (4154) (53'84) (l'54)16063 06271 00646 09146 30 ii 148 189 271 20 0 291 480
(630) (230) (30.80) (56•50) (420)2 1•4835 O'5462 00923 08000 14 12 47 73 57 0 0 57 130
(l080) (9.20) (36'20) (4380)14667 0'6646 01188 06833 117 32 107 256 199 22 3 224 480
(24'40) (6.70) (22.30) (41.50) (460) (060)3 1'3692 0'6154 00846 06462 35 8 39 82 45 3 0 48 130
(26.90) (6'IO) (30'OO) (3460) (2'30)12833 0'6063 0'1458 0'5313 170 52 122 344 118 15 3 136 480
(3540) (1080) (2S40) (24.60) (3.10) (060)4 12615 0'6077 01385 05154 49 14 33 96 30 4 0 34 130
(3770) (1080) (2540) (23'lO (310)l'1833 05125 01333 05396 169 48 175 392 73 11 4 88 480
(3520) (1000) (3650) (15.20) (2'30) (0.80)
5 1'1615 06538 00692 04385 65 8 36 109 20 1 0 21 130
(5000) (620) (27'70) (15.40) (0.80)
11521 0'5167 0'0333 06021 176 15 216 407 72 1 0 73 480
(36.70) (310) (45.00) (15.00) (020)
6 11154 0'7308 0'0308 0'3538 80 4 31 115 15 0 0 15 130
(61'SO) (3.10) (2390) (11.50)
1'0917 0'6000 00313 0'4604 244 15 177 436 44 0 0 44 480
(50.80) (310) (36'90) (9.20)
involved chromosomes. The same type of corn- with their fusion counterparts as in fig. 4(a), (b) itparison was made for P, I and D chiasmata and is obvious that in both cases, chiasma frequencyagain, differences were not statistically significant. is reduced in the fused chromosomes though theThe other fusions studied produce similar effects effect is less marked in the 5/6 fusion as expected;on chiasma frequency and distribution (see forexample, table 6).
Furthermore, these results indicate that the two
(c) when 1/4 and 5/6 heterozygotes were corn-pared with respect to total chiasma frequency, thedifference was highly significant (F1,139 = 1826;
fusions affect chiasma frequency differently. This P <0.01) but the reduction in proximal chiasmatais expected since in st males, chromosomes 1 and4 have on average more chiasmata than 5 and 6(table 3; fig. 2). This is corroborated by the follow-
is similar (F,,139=0'31; P>0'05).It was considered relevant to compare fusion
heterozygotes and homozygotes regarding chiasmaing: (a) while mean cell chiasma frequency of st'sis 10'75, that of 1/4 heterozygotes is 983 (F1,559 =
frequency. Since males differing karyotypicallyonly for the fusion in question are difficult to find
42'30; P<0'Ol) and that for 5/6 heterozygotes, in sufficiently large numbers owing to the nature10'52 (F1,519= 1.46; P>0.05); (b) when unfusedchromosomes 1 and 4 or 5 and 6 are compared
of the polymorphisms, comparisons were madesolely between the trivalent and the corresponding
FIgure 1 Chiasma distribution at MI in Dichroplus prazensis males. (a), (b). Two cells from a standard male (2n 19). (c) Horn1/6, Het 3/4 (2n = 16). (d) Triple 2/5, 3/4, 1/6 Het; the 1/6 trivalent is linearly oriented; note distal localisation of chiasmata(2n= 16). (e) Het 1/6, Horn 2/5, 3/4 (2n= 14). (1) Het 1/6, Horn 3/4 (2n = 16). (g) Double 1/6, 3/4 Met, 2/5 Hom (2n 15);
note exceptional proximal chiasma in bivalent 2/5 (left). Het 1/6, Horn 3/4. References: P (proximal), I (interstitial), D (distal).Bar = 10m.

150 C. J. BIDAU
.6-
08
0.7
i=iI11// 9 0 II '2 113 '4 l'5 l'6 '7 I'S RL
Figure 2 Relationship between relative L chromosome length (RL) and chiasma frequency per chromosome or chromosome armin ten standard males (*), ten randomly chosen males from a polymorphic population for fusions 1/2, 3/4 and 5/6 (0) andten males from a population with the same fusions fixed (•). Regression equations and their significance are given in table 3.
fusion bivalent. The fact that no interchrornosomaleffects have been detected (see above) was con-sidered. The comparisons were made usingindividuals from the SV, LF and EC populations.As can be seen in tables 5 and 6, no significantdifferences between Hets. and Horns, are apparent(interstitial chiasmata are however, more frequentin fusion bivalents than in trivalents).
These results suggest that the recombinationcapacity of a given population (hence its potentialability for the liberation of genetic variability)depends on the quality, number and frequency ofthe fusions present.
Inter-population comparisons
It is worth noting that interchromosomal recombi-nation is instantly reduced as a consequence of acentric fusion. Two independent linkage groups
are combined into one thus reducing the numberof elements that are randomly transmitted to thegametes. Chiasma frequency reduction is addedto this effect producing a generalised decrease inrecombination.
To test this prediction, the mean number ofdifferent fusions per individual (F) was calculatedfor each population (table 7). Mean cell total,proximal, interstitial and distal chiasma frequencyand inter cell variance of total chiasma frequency,were plotted against F. The results of the regressionanalyses (figs. 5 and 6) show that T, P and I chiasmafrequency and inter-cell variance depend sig-nificantly upon F.
A further confirmation of this effect wasobtained when populations SV, PM and C werecompared for chiasma frequency on each Lchromosome or chromosome arm. Ten males fromeach sample were considered: ten st's from PM,ten triple homozygotes (1/2, 3/4, 5/6) from C and
Table 3 Chiasma frequency per L chromosome or chromosome arm based on ten randomly chosen standard males (a) from PMpopulation, ten from a polymorphic population (b) (SV population polymorphic for 1/2, 3/4 and 5/6 fusions) and ten from amonomorphic population (c) (C population; fusions 1/2, 3/4 and 5/6 fixed)
L chromosome or chromosome arm
1 2 3 4 5 6
Mean relative length (RL) 1785 1467 1368 12-20 1093 972
Mean chiasma frequency perL chromosome or chromosomearm (±SE)
(a)(b)(c)
149±005133±0051-08±003
140±005126±0-04102±002
125±004112±003104±002
120±0-04103±002098±003
115±004118±004095±003
105±002106±0020-94±003
(a) RL vs. . Y=054+0-055X; F(1,4)=9370; P<001.(b) RL vs. X. Y=074±O032X; F(1,4)=6-96; P<001.(c) RL vs. X. Y=076+0018X; F(1,4)=78-86; P<001.
x
I 5
I 4-
3.
I 2
I I'
I 0.
09-
CENTRIC FUSIONS AND CHIASMA FREQUENCY 151
a b
Figure 3 Distribution of the number of (a) total (T), (b) proximal (P), (c) distal (D) and (d) interstitial (I) chiasmata in standard(open histograms) and fused (shaded histograms) males of the PM population. Means are indicated. Results of the respectiveanalyses of variance are: (a) F1168 = 325; P <0001; (b) F1168 = 1346; P<0001; (c) F1168 = 123; P <0001; (d) F1168 = 54;P < 000 1.
ten randomly chosen males (including individualswith 1, 2 and 3 fusions) from the highly polymor-phic SV population. From fig. 2 it can be seen thatthe relationship between chromosome length andchiasma frequency varies according to the numberof fusions present (table 3).
DISCUSSION
The presence of chromosomal polymorphisms innatural populations is frequently associated with
profound effects on intra- and/or interchromo-somal recombination. Examples can be found indifferent organisms and include inversions inDrosophila and some grasshoppers (Confalonieri,1988; Confalonieri and Colombo, 1989; Goñi eta!., 1985; Hewitt, 1979; Sperlich and Pfriem, 1986;Weissman, 1976; White and Morley, 1955), super-numerary segments in many Orthoptera (Hewitt,1979; John, 1983; Riva et a!., 1984), B chromo-somes (Jones and Rees, 1982), etc. The mechanismby which recombination is modified depends onthe rearrangement involved and may imply van-
1075 N
3
j77
N
C d
6,2 5
nD 0•1 23 •4 ni
152
C.
J. B
IDA
U
a
I S
XL4
7
3. A5_6
9
b
—
— , 6 6
c 6666 S
o a r ri
4rs
n —
——
0
0
— o
0 0
ii a >
, . a V
ao U
0 •a
LH
V
V.2
o 0 vU
0 0 .2
_9
. . a V
V
—
a 0.
a 0
0
V
U U
U
U—
'ii' '-0 V
8 .0
a . o E
V
< V
I j a-
en - .2 a 0 •_ F
-
oo
-
- F
—
- 9
10
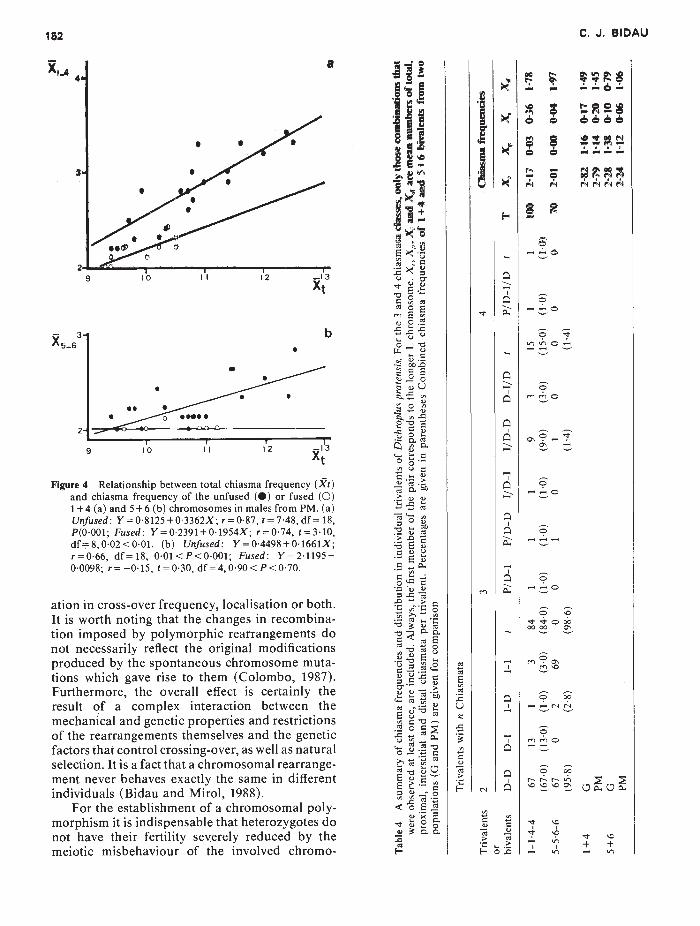
II
12
—13 xt
Figure
4 R
elationship
between
total
chiasma
frequency
(It) and
chiasma
frequency
of the
unfused
(S)
or
fused
(0) 1 + 4 (a)
and
5 +
6 (b)
chromosom
es
in m
ales
from
PM.
(a)
Unfused:
Y=
0-8125-1-0-3362X;
r 0-87,
t 7-48,
df=
18,
P(0-001;
Fused:
Y=
0-2391
+01954X
;
r=0-74,
1=310, df=
8,0-02<0-01.
(b)
Unfused:
Y=
0-4498+0-1661X
; r=0-66,
df=18,
0-01<P<
0•00l;
Fused:
Y=
2-1195—
0-0098;
r=
—015,
(=0-30,
df=4,
090<
P<0-70.
ation
in cross-over
frequency,
localisation
or
both. It is w
orth
noting
that
the
changes
in recom
bina-
tion
imposed
by
polymorphic
rearrangements
do
not
necessarily
reflect
the
original
modifications produced
by
the
spontaneous
chromosom
e
muta-
tions
which
gave
rise
to them
(Colom
bo,
1987). Furthermore,
the
overall
effect
is certainly
the
result
of
a com
plex
interaction
between
the
mechanical
and
genetic
properties
and
restrictions of the
rearrangements
themselves
and
the
genetic
factors
that
control
crossing-over,
as
well
as
natural
selection.
It is a fact
that
a chrom
osomal
rearrange-
ment
never
behaves
exactly
the
same
in different
individuals
(Bidau
and
Mirol,
1988). For
the
establishment
of a
chromosom
al
poly- morphism
it is indispensable
that
heterozygotes
do
not
have
their
fertility
severely
reduced
by
the
meiotic
misbehaviour
of
the
involved
chromo-
a a 8 a .0 .0
0 V a
1-
0 a-
000 0000
a
en
O
—
—
r-1
c-
en
en
0 000
a- '7
+
+
—
—
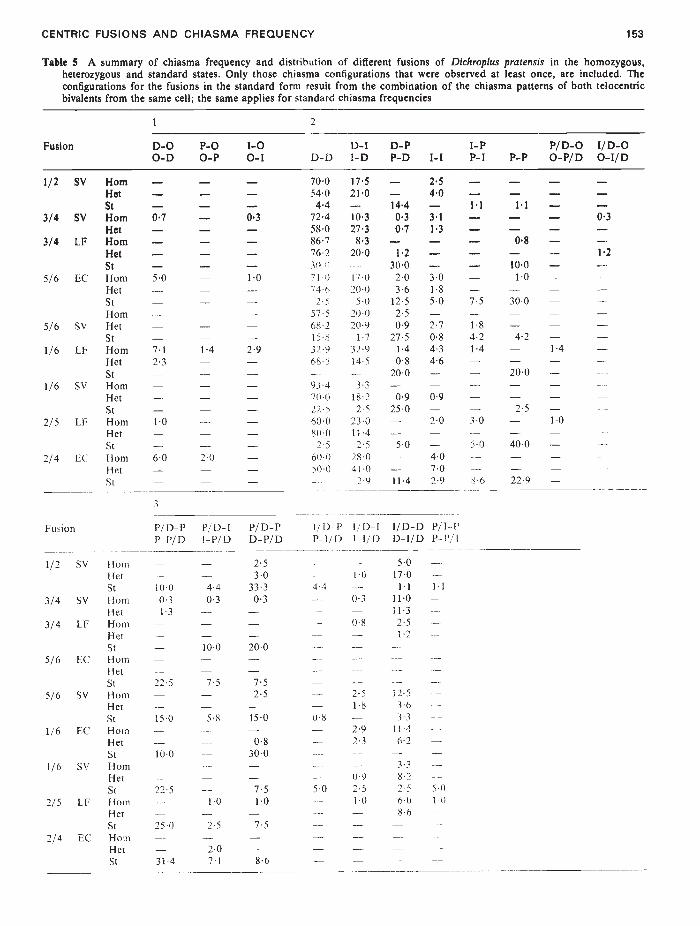
CENTRIC FUSIONS AND CHIASMA FREQUENCY
Fusion P/D-P P11)-I
1/2 SV HornHet —St 100
3/4 SV Horn 03Het 13
3/4 LF Horn —Het —St
5/6 EC Horn —HetSt 225
5/6 SV Horn —Het —St 150
1/6 I/C Horn —Het —St 100
1/6 SV HornHet —St 225
2/5 LF HornHet —St 250
2/4 EC Horn —-
Het —St 314
—— 104.4— 03
08
— 25— 1808 ——- 29- 23
— 0950 25— 1°0
153
Table 5 A summary of chiasma frequency and distribution of different fusions of Dichroplus pratensis in the homozygous,heterozygous and standard states. Only those chiasma configurations that were observed at least once, are included. Theconfigurations for the fusions in the standard form result from the combination of the chiasma patterns of both telocentricbivalents from the same cell; the same applies for standard chiasma frequencies
Fusion
1 2
D-O P-O 1-0 D-1 D-P I-P P/D-O I/D-OO-D 0-P 0-I D-D 1-D P-D 1-I P-I P-P O-P/D 0-l/D
1/2 SV Horn — — — 700 175 — 2'5 — — — —Het — — — 540 21'O — 40 — — —St — — — 4.4 — 14'4 — 1.1 1'I — —
3/4 SV Horn 0'7 — 0'3 724 103 O'3 31 — — — 0•3Het — — — 580 273 07 1'3 — — — —
3/4 LF Horn — — — 867 83 — — — 0'S — —Het — — — 762 200 12 — — — 12St — — — 300 -- .— 100 —.—
5/6 I/C Horn 50 — 10 712 170 20 30 — 10 — —I-let — — — 74-6 200 36 18 — — — —St — — — 2-5 5-0 12-5 5-0 7-5 300 — —Horn — — — 57-5 20(1 25 — — — — —
5/6 SV Het — — — 68-2 20-9 0-9 2-7 1-8 — — —St — — — 15-S 1-7 27-5 08 42 4-2 — —
1/6 LF Horn 71 1-4 2-9 32-9 32-9 14 43 14 — 14 —
Het 23 — — 68-5 14-5 0-8 46 — — — —-
St — — — —- —--- 20-0 — — 200 — ---
1/6 SV Horn — — — 93.4 3.3Het — — — 70-0 18-2 09 09 — — — —--
St — — — 22-5 2-5 25-0 — — 2-5 — —--
2/5 I/F Horn 10 — — 60-0 23-0 — 20 3-0 — 1-0 -—-
Het — — — 82-0 11-4St — — — 2-5 2-5 50 — 5-0 400 — —
2/4 I/C Horn 6-0 2-0 — 60-0 280 — 4-0 -—- —Het — — — 52-0 41-0 — 7-0 — — — -—-
St — — — —-- 2-9 114 29 5-6 22-9 — -
P/ D- PP—I'/D I-P/I) D—P/D
4.40-3
10-0
7.5
5-8
1-0
2-5
2-07-1
2530
33.30-3
200
7.52-5
150
0-830-0
7.51-0
7.5
8-6
1/ft-P l/D—I 1/D—D P/I--PP-I/i) I—I/I) D—I/D P—P/I
5-0 ——
1701-1 1-1
11-0 —11-32-51-2 —
125 —36 -—
3.3114 —62 —
3.38-22-5 5-060 1-08-6
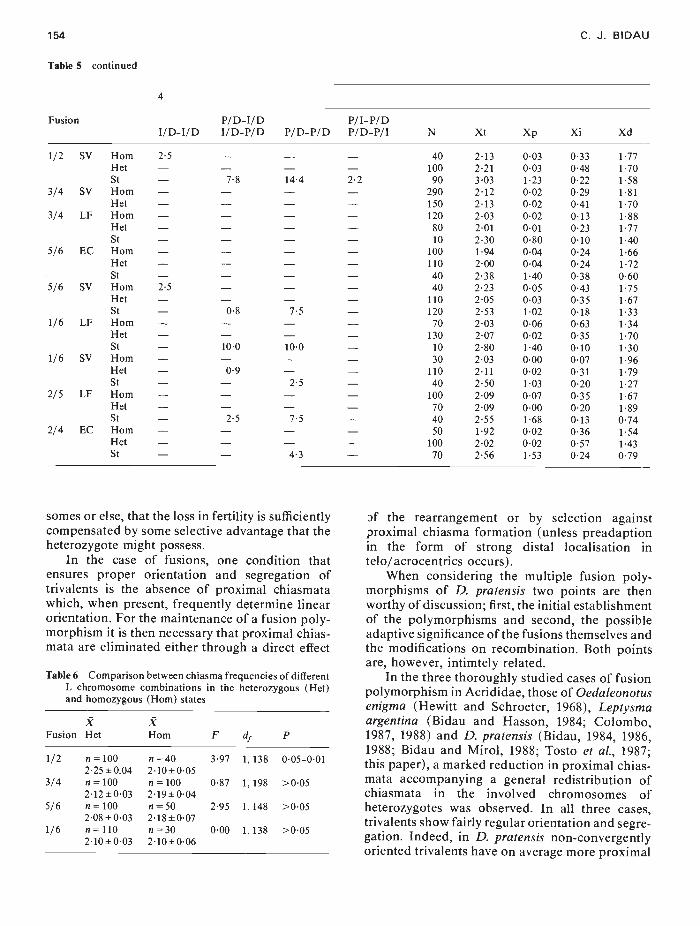
154 C. J. BIDAU
Table 5 continued
Fusion
4
I/D-I/DP/D-I/DI/D-P/D P/D-P/D
P/I-P/DP/D-P/I N Xt Xp Xi Xd
1/2 SV Horn 2-5 — — — 40 213 003 033 177Het — — — — 100 2-21 0-03 0-48 170St — 7-8 144 22 90 3-03 1-23 022 1-58
3/4 5V Horn — — — — 290 2-12 0-02 029 1-81Het — — — — 150 2-13 002 0-41 1-70
3/4 LF Horn — — — — 120 203 0-02 013 188Het — — — — 80 2-01 001 023 1-77St — — — — 10 2-30 0-80 0-10 1-40
5/6 EC Horn — — — — 100 194 0-04 0-24 1-66Het — — — — 110 2-00 0-04 0-24 172St — — — — 40 2-38 1-40 0-38 060
5/6 SV Horn 2-5 — — — 40 2-23 0-05 043 1-75Het — — — — 110 205 003 0-35 1-67St — 0-8 7-5 — 120 253 1-02 018 133
1/6 LF Horn — — — — 70 2-03 006 0-63 134Het — — — — 130 207 0-02 0-35 1-70St — 10-0 10-0 — 10 280 1-40 0-10 1-30
1/6 SV Horn — — — — 30 2-03 0-00 0-07 196Het — 09 — — 110 2-11 0-02 031 1-79St — — 2-5 — 40 2-50 1-03 020 127
2/5 LF Horn — — — — 100 2-09 007 0-35 167Het — — — — 70 2-09 0-00 020 189St — 2-5 7-5 — 40 2-55 168 013 0-74
2/4 EC Horn — — — — 50 1-92 0-02 0-36 1-54Het — — — — 100 2-02 0-02 0-57 1-43St — — 43 — 70 2-56 1-53 0-24 0-79
somes or else, that the loss in fertility is sufficientlycompensated by some selective advantage that theheterozygote might possess.
In the case of fusions, one condition thatensures proper orientation and segregation oftrivalents is the absence of proximal chiasmatawhich, when present, frequently determine linearorientation. For the maintenance of a fusion poly-morphism it is then necessary that proximal chias-mata are eliminated either through a direct effect
Table 6 Comparison between chiasrna frequencies of differentL chromosome cornbinations in the heterozygous (Het)and homozygous (Horn) states
Fusion?Het
?Horn F d P
1/2 n = 100 n = 40 3-97 1, 138 0-05—0-012-25±0.04 2-10±0-05
3/4 n=1002-12±0-03
n=1002-19±0-04
0-87 1,198 >0-05
5/6 n = 100 n = 50 2-95 1, 148 >0-052-08±0-03 2-18±0-07
1/6 n 1102-10±0•03
n =302-10±0-06
0-00 1,138 >0-05
Df the rearrangement or by selection againstproximal chiasma formation (unless preadaptionin the form of strong distal localisation intelo/acrocentrics occurs).
When considering the multiple fusion poly-morphisms of D. pratensis two points are thenworthy of discussion; first, the initial establishmentof the polymorphisms and second, the possibleadaptive significance of the fusions themselves andthe modifications on recombination. Both pointsare, however, intimtely related.
In the three thoroughly studied cases of fusionpolymorphism in Acrididae, those of Oedaleonotusenigma (Hewitt and Schroeter, 1968), Leptysmaargentina (Bidau and Hasson, 1984; Colombo,1987, 1988) and D. pratensis (Bidau, 1984, 1986,1988; Bidau and Mirol, 1988; Tosto et a!., 1987;this paper), a marked reduction in proximal chias-mata accompanying a general redistribution ofchiasmata in the involved chromosomes ofheterozygotes was observed. In all three cases,trivalents show fairly regular orientation and segre-gation. Indeed, in D. pratensis non-convergentlyoriented trivalents have on average more proximal
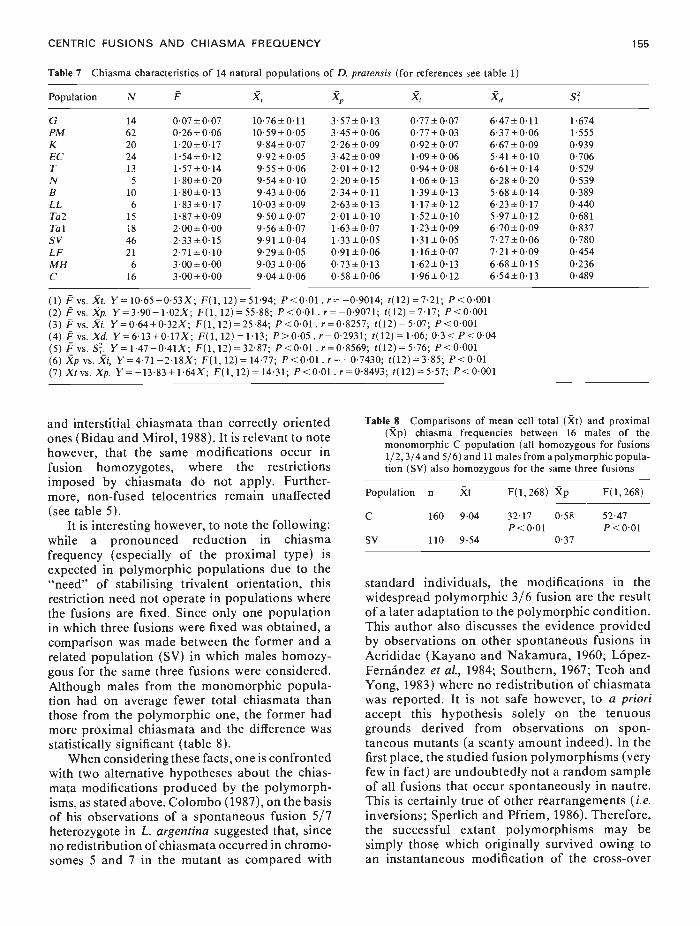
CENTRIC FUSIONS AND CHIASMA FREQUENCY 155
Table 7 Chiasma characteristics of 14 natural populations of D. pratensis (for references see table 1)
Population N F X, X X, Xd SG 14 007±007 1076±011 357±013 077±007 647±011 1674PM 62 0•26±006 1059±005 345±006 077±003 637±006 1555K 20 120±017 984±007 226±009 092±007 667±009 0939EC 24 154±012 992±005 342±009 1'09±006 541±0'lO 0706T 13 1•57±014 955±006 2•01±012 0•94±008 6•61±014 0529N 5 180±020 954±010 220±015 106±013 628±020 0539B 10 180±013 943±0•06 234±011 139±013 568±014 0389LL 6 183±017 1003±009 263±013 117±012 623±017 0440Ta2 15 187±009 950±007 201±010 152±010 597±012 0681Tal 18 200±000 956±007 163±007 123±009 670±009 0837SV 46 2'33±015 991±004 133±005 131±005 727±006 0780LF 21 271±010 9•29±005 091±006 116±007 721±009 0454MH 6 300±000 903±0'06 073±013 162±013 668±0'lS 0236C 16 3•00±000 904±006 058±006 196±012 654±013 0489
(1) P vs. Xi. Y= 1065 —053X; F(1, 12) = 5194; P <001 . r= —09014; 1(12) = 721; P<0001(2) Pvs. .p. Y=390—102X; F(l,12)=5588; P<001 . r=—09071; i(12)=717; P<0001(3) F vs. Xi Y=064+032X; F(l, 12)=2584; P<001 . r=08257; i(12)= 507; P<0001(4) Pvs. Xd. Y=613+017X; F(1, 12)=113; P>005. r=02931; t(12)=106; 03<P<004(5) F vs. S. Y= 147 —041X; F(1, 12) =3287; P<001 .r= 08569; 1(12) = 576; P<0001(6) .p vs. Ri, Y=471 —218X; F(1, 12) = 1477; P<001 . r= —07430; i(12) = 385; P<001(7) Xi vs. Xp. Y —1383+ 164X; F(1, 12) = 1431; P<001 . r= 08493; 102) = 557; P<0001
and interstitial chiasmata than correctly orientedones (Bidau and Mirol, 1988). It is relevant to notehowever, that the same modifications occur infusion homozygotes, where the restrictionsimposed by chiasmata do not apply. Further-more, non-fused telocentrics remain unaffected(see table 5).
It is interesting however, to note the following:while a pronounced reduction in chiasmafrequency (especially of the proximal type) isexpected in polymorphic populations due to the"need" of stabilising trivalent orientation, thisrestriction need not operate in populations wherethe fusions are fixed. Since only one populationin which three fusions were fixed was obtained, acomparison was made between the former and arelated population (SV) in which males homozy-gous for the same three fusions were considered.Although males from the monomorphic popula-tion had on average fewer total chiasmata thanthose from the polymorphic one, the former hadmore proximal chiasmata and the difference wasstatistically significant (table 8).
When considering these facts, one is confrontedwith two alternative hypotheses about the chias-mata modifications produced by the polymorph-isms, as stated above. Colombo (1987), on the basisof his observations of a spontaneous fusion 5/7heterozygote in L. argentina suggested that, sinceno redistribution of chiasmata occurred in chromo-somes 5 and 7 in the mutant as compared with
Table 8 Comparisons of mean cell total (it) and proximal(Xp) chiasma frequencies between 16 males of themonomorphic C population (all homozygous for fusions1/2,3/4 and 5/6) and 11 males from a polymorphic popula-tion (SV) also homozygous for the same three fusions
Population n Xt F(1, 268) Xp F(1, 268)
C 160 904 3217P<0•01
058 5247P<00l
SV 110 954 037
standard individuals, the modifications in thewidespread polymorphic 3/6 fusion are the resultof a later adaptation to the polymorphic condition.This author also discusses the evidence providedby observations on other spontaneous fusions inAcrididae (Kayano and Nakamura, 1960; Lopez-Fernández et a!., 1984; Southern, 1967; Teoh andYong, 1983) where no redistribution of chiasmatawas reported. It is not safe however, to a prioriaccept this hypothesis solely on the tenuousgrounds derived from observations on spon-taneous mutants (a scanty amount indeed). In thefirst place, the studied fusion polymorphisms (veryfew in fact) are undoubtedly not a random sampleof all fusions that occur spontaneously in nautre.This is certainly true of other rearrangements (i.e.inversions; Sperlich and Pfriem, 1986). Therefore,the successful extant polymorphisms may besimply those which originally survived owing toan instantaneous modification of the cross-over
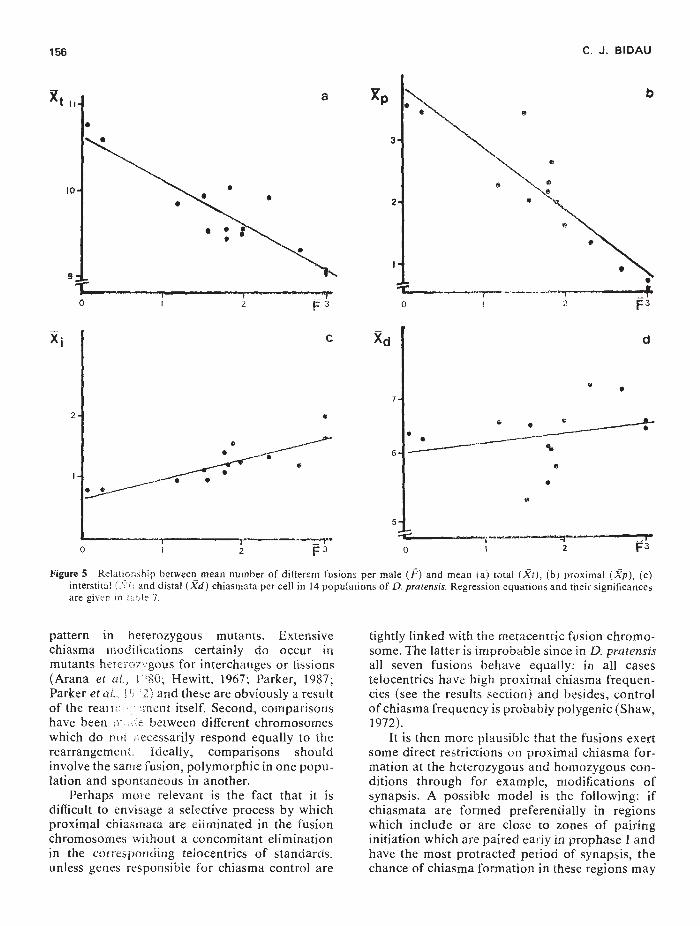
156 C. J. BIDAU
I I2
—,2 F
Figure 5 Relahonship between mean number of different fusions per male (F) and mean (a) total (.i), (b) proximal (gp), (c)interstital (X and distal (Xd) chiasmata per cell in 14 populations of D. pratensis. Regression equations and their sigriificancesare givrn fl le 7.
pattern in heterozygous mutants. Extensivechiasma modifications certainly do occur inmutants hetero7vgous for interchanges or fissions(Arana et a1. SO; Hewitt, 1967; Parker, 1987;Parker et aL, 2) and these are obviously a resultof the rearn ment itself. Second, comparisonshave been between different chromosomeswhich do n'.'t necessarily respond equally to therearrangement. Ideally, comparisons shouldinvolve the same fusion, polymorphic in one popu-lation and spontaneous in another.
Perhaps more relevant is the fact that it isdifficult to envisage a selective process by whichproximal chiasmaa are eiiminated in the fusionchromosomes without a concomitant eliminationin the corresponding telocentrics of standards,unless genes responsible for chiasma control are
tightly linked with the metacentric fusion chromo-some. The latter is improbable since in D. pratensisall seven fusions behave equally: in all casestelocentrics have high proximal chiasrna frequen-cies (see the results section) and besides, controlof chiasma frequency is probably polygenic (Shaw,1972).
It is then more plausible that the fusions exertsome direct restrictions on proximal chiasma for-mation at the heterozygous and homozygous con-ditions through for example, modifications ofsynapsis. A possible model is the following: ifchiasmata are formed preferentially in regionswhich include or are close to zones of pairinginitiation which are paired early in prophase I andhave the most protracted period of synapsis, thechance of chiasma formation in these regions may
Io
3
S
b
0
xi
a
C Xd
7.
0
6
0
2
d
•0
0
S .S
5.
0

CENTRIC FUSIONS AND CHIASMA FREQUENCY 157
a
— 0
Figure 6 Relationship between (a) F and mean betweenceli variance of total chiasmata (S) in 14 populations of 1). prafnthand (b) mean frequency of interstitial (Xi) ano proximal (Xp) cniasmasa in the same populations. Regression equations andtheir significance are given in table 7.
well be increased. Then, if unfused telocentricshave both ends attached to the nuclear membrane,they can initiate pairing from either or both ends,which could explain the P/D chiasma distribution,In fused chromosomes however, four original endsare replaced by two and the number of early pairingregions is consequently reduced and shifted todistal ends only. This would explain both thereduction in total chiasrna frequency and thereduction in proximal chiasmata in fusionheterozygones and iiornozygotes.
Turning to the possible adaptive significanceof the D. pratensis's polymorphisms one is temptedto relate their maintanence to the restrictionsimposed upon recombination. Fusions are not aseffective suppressors of recombination as inver-sions bum a fusion instantly reduces interchromo-somal recombination by bringing together two for-merly independent linkage groups, usually accom-panied by a suppression of chiasmata in proximalportions of chromosomes as discussed above.These facts could allow the conservation ofcoadapted supergenes partially or completely pro-tected from recombination. Thus the maintenanceof the polymorphisms would become adaptivesince fixation could, by eliminating the segregationrestrictions of heterozygotes, result in an increasein crossing-over in regions previously protected(as in the C population described in Results) andthe disruption of supergenes.
Two hypothese relate polymorphic rearrange-ments and supergenes, the postadaptation and thepreadaptation hypotheses (Sperlich and Pfriem,1986). In the first case, the newly arisen fusion
could be initially successful because of favourableposition effects. The need for stabilising segrega-tiort in heterozygotes could then, by restrictingchiasma formation, allow the surging of super-genes. That is, the decrease in recombination beinga byproduct of the establishment of the poly-morphism would not be the initial cause of itsmaintenance. In the second case one couldenvisage a situation in which some coadaptationalready exists and any fusion that involves suchgene arrangements should be favoured since itwould protect them from recombination.
A model proposed for the chromosomal evol-ution of D. pratensis (Bidau, 1984, 1988) can beexpanded when the effects of the fusions on re-combination are considered. It proposes that rangeexpansion of the species occurred mainly in themonomorphic standard form since these popula-tions have more recombination potential and thus,a greater capacity for liberating genetic variabilitywhich allows the production of more gene arrange-ments to be selected for in order that the populationbecomes adapted to a new environment. Once anew colony adapts successfully to its new habitat,the fortuituous appearance of one or more fusionsthat include the adaptive gene combinations wouldgenerate the polymorphic condition to preservethem as explained above.
It is possible to relate this model to the central-marginal model of evolutionary biology as it hasbeen applied for example to Drosophila (Brussard,1984; Sperlich and Pfriem, 1986). The modelassumes that populations belonging or near to thecentre of a species' range, have certain characteris-
r2 2
3
0
b
2S
• 0
a
0
2
S
a 10 15 20

158 C. J. BIDAU
tics such as high density, high levels of genetic andphenotypic variation, etc. In contrast, peripheralpopulations are isolated, sparse, have low levelsof chromosomal polymorphism, etc. A number ofhypotheses concerning the inversion polymorph-isms in Drosophila as related to this model havebeen put forward (Brussard, 1984).
When considering the distribution of fusionpolymorphisms in D. pratensis it was shown that,at least in the sampled area, no clear pattern isevident (Bidau, 1988). Certainly, some fusions arewidespread and others restricted to particularlocalities, and populations are generally very poly-morphic. Exceptions however, do exist; twoPatagonian populations have exceptionally highfrequencies of standard individuals (PM and G)and are consequently the less polymorphic onesand those with the highest recombination indexes.Despite the wide ecological tolerance displayed bythe species, these populations are the best candi-dates for being considered marginal (in the eco-logical sense) since environmental conditions inthese areas are certainly more strenuous (seeBidau, 1984).
These populations, having more recombinationpotential, could liberate more genetic variability(if available) that would be of advantage in a new,unexploited environment. The PM and G popula-tions would then agree with Carson's (1959) modelin having low polymorphism and high recombina-tion. It is however relevant to discuss somemorphological data of D. pratensis (Bidau andTosto, 1986). Although the putative marginalpopulations seem to be phenotypically more vari-able than those presumed to be central (or optimalin he ecological sense), no statistically significantcorrelation was found between degree of poly-morphism and morphological variability. This isin contrast with the report of Westerman (1983)on Phaulacridium in which a negative correlationbetween recombination potential and phenotypicvariability was demonstrated. He produced a veryapt explanation: those populations with high cross-over frequencies liberate much genetic variabilitywhich is subjected to selection very rapidly andconsequently, rapidly lost. Populations with alower recombination potential would preserve theextant genetic variability. In Westerman's (1983)case however, low recombination populationswere the peripheral ones. A further case of acentral-marginal distribution of a fusion poly-morphism in a grasshopper was reported byColombo (1988).
The case of D. pratensis is puzzling. On theone side, polymorphism is lower and recombina-
tion higher in marginal populations; on the other,phenotypic variation shows no significant differen-ces. The latter could be related to the fact thatallozyme variability is not correlated with chromo-somal variability (Brussard, 1984). It is relevanthowever to note that although central populationsmay be chromosomally more polymorphic, thetype of polymorphism conditions the propositionsof the central-marginal model: central populationsmay be highly polymorphic for B chromosomeswhich raise chiasma frequency, or inversions orfusions that decrease it. Hence, the chromosomalsituation is possibly related not only tomodifications of recombination but to other effectsdue to the rearrangements themselves.
The hypothesis that the PM and G populationsare recent invaders is appealing but by no meansdefinitive.
Acknowledgements I am fully indebted to Dr CristinaD'Aiutolo and my sons, Claudio Jr and Pablo, for help in thecollection and processing of the grasshoppers. lam also indeb-ted to my collaborators, Miss Daniela Tosto and Miss PatriciaMirol for their patient work, useful comments and good com-pany. Thanks are also due to my colleague and friend DrPablo Colombo for the critical review of the manuscript andmany stimulating discussions. I also wish to thank an anony-mous reviewer whose corrections and suggestions improvedthe manuscript substantially.
This work was supported by grants from CONICET andUniversidad de Buenos Aires.
REFERENCES
ARANA, P., SANTOS, J. L. AND GIRALDEZ, R. 1980. Chiasmainterference and centromere co-orientation in a spon-taneous translocation heterozygote of Euchorthippus pul-vinatus gallicus (Acrididae: Orthoptera). Chromosoma, 78,327—340.
BIDAU, c. i. 1984. Ph.D. Thesis, Universidad de Buenos Aires.BIDAU, c. i. 1986. Geographic distribution of chromosome
polymorphisms in Dichroplus pratensis Bruner (Melano-plinae, Acrididae) in Argentina. Proc. IV Triennial Meet.,Pan Amer. Acridol. Soc., Saskatoon, 165-166.
BIDAU, C. .j. 1988. Zonas hIbridas en ortópteros: el ejemplo deDichroplus pratensis (Acrididae). Revista Soc. Entomol.Argentina (In press).
BIDAU, C. J. AND HASSON, E. R. 1984. Population cytology ofLeptysma argentina Bruner (Leptysminae, Acrididae).Genetica, 62, 161-175.
BIDAU, C. J. AND MIROL, i'. 1988. Orientation and segregationof Robertsonian trivalents in Dichroplus pro tensis(Acrididae). Genome, 30, 947—955.
BIDAU, C. J. AND TOSTO, D. 1986. Análisis de la variabilidadmorfológica de razas cromosómicas de Dichroplus pratensisen relación a la capacidad potencial de recombinacjón.XVII Congr. Arg. Genet., RIo Cuarlo, 65.
BRUSSARD, i. F. 1984. Geographic patterns and environmentalgradients: the central marginal model in Drosophilarevisited. Ann. Rev. Ecol. Syst., 15, 25—64.

CENTRIC FUSIONS AND CHIASMA FREQUENCY 159
CARSON, H. L. 1959. Genetic conditions that promote or retardthe formation of species. Cold Spring Harbor Symp. Quant.BioL, 24, 87-103.
COLOMBO, P. C. 1987. Effects of centric fusions on chiasmafrequency and position in Leptysma argentina. 1. Spon-taneous and stable polymorphic centric fusions. Genetica,72, 171—179.
COLOMBO, P. C. 1988. Polimorfismos cromosómicos enOrthoptera. Revista Soc. Entomol. Argentina (In press).
CONFALONIERI, V. A. 1988. Effects of chromosome polymorph-isms on chiasma conditions in Trimerotropis pallidipennis(Oedipodinae: Acrididae), Genetica 76, 171—179.
CONFALONIERI, V. A. AND COLOMBO, P. C. 1989. Inversionpolymorphism in Trimerotropis pallidipennis (Orthoptera):clinal variation along an altitudinal gradient. Heredity, 62,107—I 12.
GONI, B., DE VAIO, E. S., BELTRAMI, M., LEIRA, M. S., CRIVEL,M., PANZERA, F., CASTELLANOS. P. AND BASSO, A. 1985.Geographic patterns of chromosomal variation in popula-tions of the grasshopper Trimerotropis pallidipennis fromSouthern Argentina. Can. J. Genet. Cytol., 27, 259—271.
HEwITT, 0. M. 1967. An interchange which raises chiasmafrequency. Chromosoma, 21, 285-295.
HEWITT, G. M. 1979. Orthoptera. Grasshoppers and crickets.In Animal cytogenetics. 3. Insecta 1. (B. John, ed). GebruderBorn traeger, Berlin-Stuttgart.
HEWITT, G. M. AND SCHROETER, 0. L. 1968. Populationcytology of Oedaleonotus. 1. The karyotypic facies ofOedaleonotus enigma (Scudder). Chromosoma, 25, 121-140.
JOHN, B. The role of chromosome change in the evolution ofOrthopteroid insects. In Sharma, A. K. and A. S. (Eds)Chromosomes in the Evolution of Eukaryotes. Vol. 1, CRCPress, Boca Raton, pp. 1-110.
JONES, R. N. AND REES, H. 1982. B chromosomes. AcademicPress, London-New York
KAYANO, H. AND NAKAMURA, K. 1960. Chiasma studies instructural hybrids. V. Heterozygotes for a centric fusionand a translocation in Acrida lata. Cytologia, 25, 476-480.
LIEBERMANN, .j. 1963. Contribucibn al conocimiento deDichroplus pratensis (Orth., Acrid.) IDIA, 191, 23—28.
LOPEZ-FERNANDEZ. C., RUFAS, J. S., GARCIA DE LA VEGA, C.AND GOSALVEZ, j. 1984. Cytogenetic studies on Chort-hippus jucundus (Fisch.) (Orthoptera). III. The meioticconsequences of a spontaneous centric fusion. Genetica,63, 3-7.
PARKER, i. S. 1987. Increased chiasma frequency as a result ofchromosome rearrangement. Heredity, 58, 87-94.
PARKER, J. S., PALMER, R. W., WHITEHORN, M. A. F. ANDEDGAR, L.A. 1982. Chiasma frequency effects of structuralchromosome change. Chromosoma, 85, 673-686.
RIVA, E., FOX, D. P., GIRALDEZ, R. AND SANTOS, J. L. 1984.Chiasma frequency and distribution in the presence andabsence of supernumerary chromosome segments in thegrasshopper, Euchorthippus pulvinatus gallicus. Heredity,53, 101-106.
SHAW, D. D. 1972. Genetic and environmental components ofchiasma control. II. The response to selection in Schis-tocerca. Chromosoma, 37, 297—308.
SMITH, S. G. 1966. Natural hybridization in the coccinellidgenus Chilocorus. Chromosoma, 18, 380-406.
SMITH, S. G. AND VIRKKI. N. 1978. Coleoptera. In John, B.(Ed) Animal Cytogenetics 3. Insecta 5. Gebruder Born-traeger, Berlin-Stuttgart.
SOUTHERN, D. 1. 1967. Spontaneous chromosome mutationsin Truxaline grasshoppers. Chromosoma, 22, 241-257.
SPERLICH, D. AND PFRIEM, P. 1986. Chromosomal polymorph-isms in natural and experimental populations. In TheGenetics and Biology of Drosophila, vol. 3e, chapter 37,Academic Press, London, pp. 257-309.
TEOH, S. B. AND YONG, M. s. 1983. A spontaneous fusionheterozygote in the tropical grasshopper, Valanga nigri-cornis (Burmeister). Caryologia, 36, 165-173.
TOSTO, D., MIROL, P. AND BIDAU, C. J. 1987. Aspectos de lasrelaciones entre las fusiones céntricas y Ia distribución delos quiasmas en Dichroplus pratensis (Melanoplinae,Acrididae). XVIII Congr. Arg. Genet., Buenos Aires, 21.
WEISSMAN, D. B. 1976. Geographical variability in the pen-centric inversion system of the grasshopper Trimerotropispseudofasciata. Chromosoma, 55, 325-347.
WESTERMAN, M. 1983. Chiasma frequency and variabilityof morphological characters in populations of two grass-hopper species. Heredity, 51, 501—506.
WRITE, M. J. D. 1973. Animal cytology and evolution. 3rd Edit.Cambridge Univ. Press, London-New York
WHITE, M. J. D. AND MORLEY, F. H. W. 1955. Effects of pen-centric rearrangements on recombination in grasshopperchromosomes. Genetics, 40, 604-619.




















