A migration of grasshoppers, particularly Diabolocatantops axillaris (Thunberg) (Orthoptera:...
21
Bull. ent. Res. 78, 251-271 251 Published 1988 A migration of grasshoppers, particularly Diabolocatantops axillaris (Thun- berg) (Orthoptera: Acrididae), in the West African Sahel D. R. REYNOLDS and J. R. RILEY Tropical Development and Research Institute, Radar Entomology Unit, Royal Signals & Radar Establishment, Malvern, Worcs., WR14 ILL, UK Abstract A dense layer of large insects in windborne, migratory flight was observed by radar in the Tilemsi Valley in Mali between about 23.00 and 02.00 h on 10-11 October 1978. The volume density, height of flight, area density, displacement speed and direction, orientation and migration rate were deter- mined for the overflying insects. Light-trap catches and radar signatures provided strong evidence that the pest grasshopper Diabolocatantops axillaris (Thunberg) contributed to the overflying layer. Other species contributing probably included Oedaleus senegalensis (Krauss) and possibly Ochrilidia spp. For D. axillaris, the migration can be regarded as a search for overwin- tering sites by adults in reproductive diapause, and thus is an example of C. G. Johnson's Class III migration. Estimated trajectories placed the prob- able source areas of the overflying grasshoppers in the Gourma, about 150 km west-south-west of the radar site. Migration direction was approxi- mately downwind, but the grasshoppers showed a degree of common orien- tation towards the east-south-east, which added a southwards component to their displacement. At 02.30 h on the same night, a very dense line-con- centration associated with a wind-shift moved across the radar site, and insects still in flight probably became entrained in this wind convergence zone and added to the line-concentration. Other evidence of long-distance, windborne migration in D. axillaris was adduced from records of captures on ships at sea, mainly off the West African coast. The consequences of downwind displacement and concentration for grasshopper ecology and pest management are discussed. The migration behaviour of D. axillaris and other grasshopper species probably reduces migration losses by the efficient location of new habitats and forms an essential part of life-history strategies for survival in a sahelian environment. Introduction There is steadily accumulating evidence that many species of grasshoppers in the Sahel zone of Africa undertake extensive migrations at night. As well as the circumstantial evidence obtained from ground studies (see references in Riley & Reynolds, 1983), there are the direct, quantitative observations recently made with radar (Schaefer, 1976; Riley & Reynolds, 1979, 1983) which reveal the spectacular nature of many of these mass movements, with vast numbers of individuals traversing distances of hundreds of kilo- metres in a few hours. Studies of the movement of sahelian grasshoppers are of interest
-
Upload
independent -
Category
Documents
-
view
3 -
download
0
Transcript of A migration of grasshoppers, particularly Diabolocatantops axillaris (Thunberg) (Orthoptera:...

Bull. ent. Res. 78, 251-271 251Published 1988
A migration of grasshoppers, particularly Diabolocatantops axillaris (Thun-berg) (Orthoptera: Acrididae), in the West African Sahel
D. R. REYNOLDS and J. R. RILEY
Tropical Development and Research Institute, Radar Entomology Unit, Royal Signals & RadarEstablishment, Malvern, Worcs., WR14 ILL, UK
AbstractA dense layer of large insects in windborne, migratory flight was observed
by radar in the Tilemsi Valley in Mali between about 23.00 and 02.00 h on10-11 October 1978. The volume density, height of flight, area density,displacement speed and direction, orientation and migration rate were deter-mined for the overflying insects. Light-trap catches and radar signaturesprovided strong evidence that the pest grasshopper Diabolocatantops axillaris(Thunberg) contributed to the overflying layer. Other species contributingprobably included Oedaleus senegalensis (Krauss) and possibly Ochrilidiaspp. For D. axillaris, the migration can be regarded as a search for overwin-tering sites by adults in reproductive diapause, and thus is an example ofC. G. Johnson's Class III migration. Estimated trajectories placed the prob-able source areas of the overflying grasshoppers in the Gourma, about150 km west-south-west of the radar site. Migration direction was approxi-mately downwind, but the grasshoppers showed a degree of common orien-tation towards the east-south-east, which added a southwards component totheir displacement. At 02.30 h on the same night, a very dense line-con-centration associated with a wind-shift moved across the radar site, andinsects still in flight probably became entrained in this wind convergencezone and added to the line-concentration. Other evidence of long-distance,windborne migration in D. axillaris was adduced from records of captures onships at sea, mainly off the West African coast. The consequences ofdownwind displacement and concentration for grasshopper ecology and pestmanagement are discussed. The migration behaviour of D. axillaris andother grasshopper species probably reduces migration losses by the efficientlocation of new habitats and forms an essential part of life-history strategiesfor survival in a sahelian environment.
IntroductionThere is steadily accumulating evidence that many species of grasshoppers in the Sahel
zone of Africa undertake extensive migrations at night. As well as the circumstantialevidence obtained from ground studies (see references in Riley & Reynolds, 1983), thereare the direct, quantitative observations recently made with radar (Schaefer, 1976; Riley &Reynolds, 1979, 1983) which reveal the spectacular nature of many of these massmovements, with vast numbers of individuals traversing distances of hundreds of kilo-metres in a few hours. Studies of the movement of sahelian grasshoppers are of interest

2 5 2 D. R. REYNOLDS and J. R. RILEY
because they contribute to an understanding of how migration patterns allow the exploit-ation of the sahelian environment with its erratic, shifting and seasonally scarce resourcesand, conversely, how this environment has shaped the species' life-history strategies, ofwhich migration behaviour forms an essential part (Dingle, 1986; Taylor, 1986).
Consideration of migration is also important for the management of several species ofsahelian grasshopper pests, because they can rapidly invade crop areas from long distances,at densities which exceed the economic injury level, and so can devastate the crop beforecontrol measures can be implemented; they may therefore need to be controlled beforethey enter the area at risk (Joyce, 1981). Additionally, it has been suggested (Joyce, 1976,1983) that severe crop losses usually occur after aerial concentration of the windbornemigrant grasshoppers by convergent wind-fields (see Rainey, 1976). Control of potentiallydamaging pest concentrations before they reach crops is the method successfully used inthe case of locusts (Gunn & Rainey, 1979), and in principle could be used againstgrasshoppers. Even if this 'strategic' control proves impracticable, elucidation of theseasonal changes in distribution of sahelian grasshopper species will be useful if it at leastenables prediction of the crop areas most at risk so that the (often meagre) control facilitiesmay be deployed in the most effective and timely manner.
The main species considered in this paper, the catantopine grasshopper Diabolocatan-tops axillaris (Thunberg) (previously Catantops axillaris, see Jago, 1984), is found through-out the savanna of tropical Africa, the Arabian peninsula and southern Iran, and iscommon on the thorn scrub of the African Sahel. In the Sahel, it is a pest of millet,sorghum, cassava, Guinea corn and other crops (Joyce, 1952; Popov, 1976; Cheke et al.1980; Fishpool & Popov, 1984), and although it is less serious than the Senegalesegrasshopper, Oedaleus senegalensis (Krauss), it is occasionally of substantial econonicimportance (COPR, 1982).
Until recently, evidence of migration in D. axillaris has been largely circumstantial.Joyce (1952) in Sudan and Davey et al. (1959) in Mali caught large numbers at light at theend of the rainy season (September-October), and they deduced that the species under-takes extensive night-time migrations at this time. Joyce (1952) also mentioned possiblemovements of D. axillaris to "distant and isolated" irrigated gardens in April and May,before the beginning of the rains. With the advent of radar entomology, some directobservations of the migrating grasshoppers have been possible. For example, D. axillarisvery probably contributed to the southward migration of a dense aerial concentration ofgrasshoppers observed overflying two radars 100 km apart in Mali in October (Riley &Reynolds, 1983), and also in October, the species has been caught by aircraft net at 450 mabove ground in Sudan during a dense southward migration observed by radar (Schaefer,1976).
In this paper, new radar evidence is presented of long-range nocturnal migration by D.axillaris in the sahelian zone of Mali, and instances enumerated of D. axillaris captured onships at sea, which demonstrate that this species certainly has the potential for long-distance migration.
MethodsAn X-band (3-2-cm-wavelength) scanning radar, was operated at Tin Aouker (16°48'N,
0°08'E) (Fig. 7) in the Tilemsi Valley, about 65 km north of Gao, Mali, from 9 to 22October, 1978. A survey of the site indicated that 69% of the ground was bare, 29%covered by Cenchrus biflorus grass (mainly drying or dried-out) and 2% covered byCitrullus colocynthis; there were occasional scattered Acacia bushes. The radar (describedin Riley & Reynolds (1983)) was equipped with an antenna which produces a narrow (1-8°)beam, which can be pointed at any desired elevation and scanned about the zenith. Insectsof D. axillaris size were detected out to a maximum range of 1-7-2 km, and their presencewas registered on a Plan Position Indicator (PPI) screen, which was photographed on 16-mm film (one frame per revolution of the antenna). Radar signatures (Riley, 1974;Schaefer, 1976; Drake & Farrow, 1983) were recorded from individual targets flying

GRASSHOPPER MIGRATION IN THE SAHEL 253
through the stationary beam. These signals were subsequently analysed to extract theinsect wing-beat frequencies, which form an aid to identification.
A light trap, situated near the radar, consisted of a 150-W ultraviolet light bulbsuspended above a 1-2 x 1-2-m water bath made of polyethylene sheet dug into the soil.The trap was emptied at intervals throughout the night, and each species counted andsexed. The reproductive status of females was determined, and the tegmen lengths of bothsexes were measured on a sample of the commoner grasshopper species from the trap.Estimates of the mean wing-beat frequency and approximate 95% confidence limits (forcomparison with the radar-derived wing-beat frequencies), were calculated from themeasurements of tegmen length using the relationships given in Riley & Reynolds (1983).
ResultsRadar observations of insect aerial density and displacement direction on 10-11 October1978
The variation in the aerial density of insects detected by the radar at 150 m aboveground level during the night of 10-11 October is shown in Fig. la. After a fairly densedusk take-off (starting at ca 17.55 h), the numbers overflying the radar declined from 19.15until 22.00 h. The insects were displacing towards the east-north-east (60-90°) during thisperiod (Fig. \b). At 22.15 h, a concentration of insects forming a line-echo on the PPIscreen crossed the radar moving towards 300°, and there was a corresponding change in thedisplacement direction (towards 300° at 90 m and 350° at 480 m) until about 22.30 h. From23.30 h onwards, increased aerial densities of large insects flew over the radar, displacingto the east or east-south-east. Densities of this magnitude (up to almost 1/104 m3) wereunusual this late in the night at Tin Aouker, and it is these insects which form the mainfeature of interest of the present paper. The vertical density profile of the insects wasstratified, with a fairly strong layer between about 240 and 350 m above the ground (Figs 2& 3a), although many individuals were flying lower than this. The layer became less well-defined after 00.16 h, and its altitude decreased (Fig. 3b). The insects in the layer showedsome degree of common orientation (Riley, 1975) towards 115-125°. Coincident with theoverflight of this layered concentration, large numbers of grasshoppers, especially D.axillaris, were observed arriving at the light trap (see below). After about 01.30 h, thelayering was no longer clear and aerial densities declined somewhat (Fig. 3c).
At 02.27 h, a dense (at least 3 insects/103 m3) line-echo, with its axis aligned 140-320°,moved across the radar towards 225° at about 6-5 m/s. As was usually observed, the line-echo occurred at a meso-scale front or wind-shift line; in this case, the insects displacedtowards approximately 110° before (and above) the line-echo, and towards the south-westor south within and behind the echo. Surface winds at the radar site changed abruptly fromlight (<1 m/s) south-westerlies to 4 m/s north-north-easterlies at 02.20 h, and then backedto 2-3 m/s north-westerlies at about 02.30 h and remained thus for the rest of the night.Surface dew point fell slightly from 21°C at 02.00 h to 19°C at 03.00 h. It was deduced thatthe wind-shift line was associated with a rain-producing, meso-scale disturbance becausemiddle-level rain echo was seen on the radar up to 5-6 km above ground level (a.g.l.) at02.40-03.00 h after the passage of the line-echo, although no rain reached the ground nearthe radar site. By 02.55 h, insect densities had decreased again to values seen before thearrival of the line-echo, and displacement was towards the south-south-east. Densities nowgenerally continued to decline, and displacement direction was maintained towards thesouth-east until dawn, when insect aerial activity fell rapidly, as was usual, to the extremelylow levels seen just after dawn.
Light-trap catchesThe overwhelming majority of the larger-sized insects caught in the light trap at Tin
Aouker were grasshoppers or gryllids (Riley & Reynolds, 1983). The commonest grass-hoppers in the habitat, O. senegalensis and Acrotylus longipes (Charpentier), were also themost abundant species in the trap, contributing 40 and 16%, respectively, of the 18.00-

254 D. R. REYNOLDS and J. R. RILEY
TIN AOUKER 10-11 Oct. 1978
Layer extendsdown to 150m
Line l e v e l
echo
Denseline echo
Surface winddirection
Fig. la.—Variation in volume density (numbers per 107 m3) of insects in the 143-172-mheight interval, during the night of 10-11 October 1978 at Tin Aouker; densities werederived from counts of targets on the PPI in the range interval 914-1097 m (1000-1200 yd)at 9° beam elevation angle. Fig. Ib.—Variation in insect displacement direction at ca 160 mabove the ground during the night of 10-11 October 1978, determined from the radar PPIscreen; also shown are wind arrows giving wind speed and direction at 2 m above the
ground (see Fig. 6 for key to wind speeds).
21.00 h total catch for all nights at Tin Aouker (n = 15). On the other hand, D. axillarisnumbers were generally low during the first three hours (18.00-21.00 h) of light-trapping,averaging only 2-1% of the total acridoids caught. The catches for the period 18.00-22.55 hon the night of 10-11 October (Table I) show this clearly, with D. axillaris representing aminuscule proportion of the total. In the next period (23.55-03.05 h), however, during the
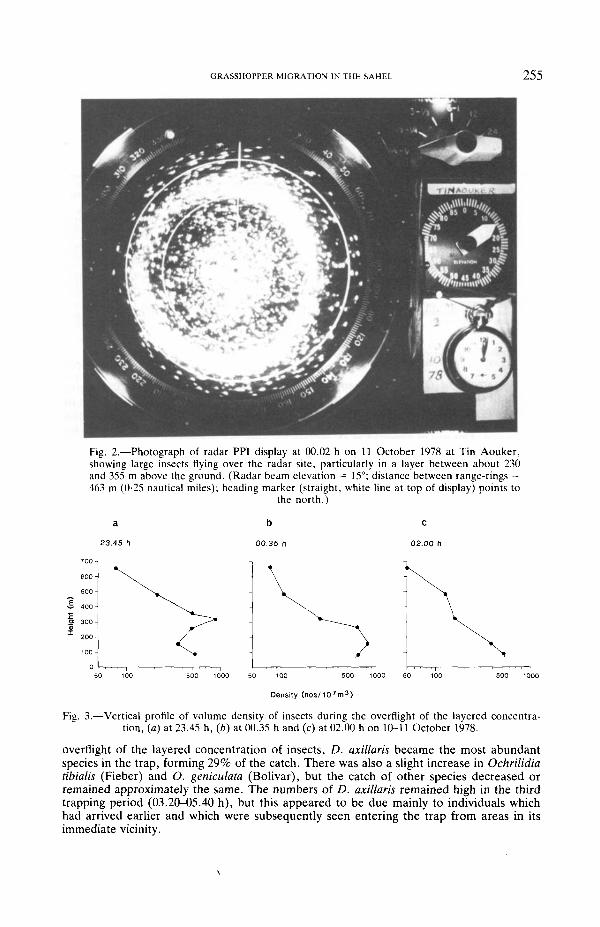
GRASSHOPPER MIGRATION IN THE SAHEL 255
Fig. 2.—Photograph of radar PPI display at 00.02 h on 11 October 1978 at Tin Aouker,showing large insects flying over the radar site, particularly in a layer between about 230and 355 m above the ground. (Radar beam elevation = 15°; distance between range-rings =463 m (0-25 nautical miles); heading marker (straight, white line at top of display) points to
the north.)
00.35 h
700 -
600-
500-
400-
300-
200-
100-
050 500 1000 50 100 500 1000 50 100
Density (noa/107m3)
Fig. 3.—Vertical profile of volume density of insects during the overflight of the layered concentra-tion, (a) at 23.45 h, (b) at 00.35 h and (c) at 02.00 h on 10-11 October 1978.
overflight of the layered concentration of insects, D. axillaris became the most abundantspecies in the trap, forming 29% of the catch. There was also a slight increase in Ochrilidiatibialis (Fieber) and O. geniculata (Bolivar), but the catch of other species decreased orremained approximately the same. The numbers of D. axillaris remained high in the thirdtrapping period (03.20-05.40 h), but this appeared to be due mainly to individuals whichhad arrived earlier and which were subsequently seen entering the trap from areas in itsimmediate vicinity.
256 D. R. REYNOLDS and J. R. RILEY
TABLE I. Light-trap catches
i
Grasshopper species
Diabolocatantops axillarisOedaleus senegalensisOchrilidia gracilisPyrgomorpha conica
(Olivier)Ochrilidia tibialisStenohippus epacromioides
(Krauss)Ochrilidia geniculataAiolopus simulatrixAcrolylus longipesAcrotylus blondeli SaussureOther grasshopper species
of grasshoppers at Tini Aoukerduring the night of10-11 October 1978
Time p18.00-2
Averagenumber caught
per hour*
0-68116-437-3
40-94-6
18-96-7
13-577-919-87-4
334-0
leriod2.55 h
Numbersas %
of total
0-233-810-8
11-91-3
5-51-93-9
22-75-82-2100
Time period23.55-03.05 h
Averagenumber caught
per hour*
90-471-433-2
27-218-6
18-314-212-310-72-5
11-7311-0
Numbersas %
of total
29-223-010-7
8-76 0
5-94-63-93-40-83-8100
for three periods
Time period03.20-05.40 h
Averagenumber caught
per hour*
40-354-417-1
9-011-6
9-412-07-3
16-33-4
11-6
192-4
Numbersas %
of total
20-928-38-9
4-76-0
4-96-23-88-51-86 0
100
* If sub-periods different from 1 hour, sub-period totals corrected.
All samples of D. axillaris taken from the light trap during the 23.55-03.05-h periodhad hard cuticles, but their wings were intact and untattered. It was thus deduced that thegrasshoppers were undertaking post-teneral migration (Johnson, 1969). All the femaleshad undeveloped ovaries with no vitellogenesis, and there was no sign that any hadoviposited. This was as expected as D. axillaris has only one generation a year, survives thedry season in imaginal diapause and does not oviposit until the beginning of the next year'srainy season (Joyce, 1952; Fishpool & Popov, 1984); thus at the end of the rains inOctober, the population would be composed of young immature adults.
Wing-beat frequenciesThe distribution of all the wing-beat frequencies (n = 3362) derived from radar
signatures recorded during the whole observational period at Tin Aouker (Fig. 4) fellwithin the range of wing-beat frequencies estimated for the common grasshopper speciesfound at this site (see Riley & Reynolds, 1983), and thus supported the hypothesis thatmost of the larger-sized insects in flight at heights scanned by the radar were indeedgrasshoppers. The peak frequencies (26-32 Hz) in the signature distribution (Fig. 4)corresponded well with the wing-beat frequency range of Oedaleus senegalensis (approxi-mately 26-28 Hz for females and approximately 30-32 Hz for males (Riley & Reynolds,1983 & unpublished data)), so it seems very likely that the commonest grasshopper speciesin the trap was the insect most frequently detected by the radar. (The size of O.senegalensis generally tended to decrease during the observational period, so the tegmenlengths of individuals in the trap on the night of 9-10 October (Table II) were slightlylonger (about 1-2 mm) than those measured later in the month (Riley & Reynolds,unpublished), thus producing a corresponding increase of about 1 Hz in the mean wing-beat frequency.)
The distributions of radar-derived wing-beat frequencies for the night of the 10-11October, both before the arrival of the insect layer and during its passage, are shown inFig. 5a & b. Before applying a jr2 test to see whether there was a significant differencebetween these distributions, the range of wing-beat frequencies was restricted to thatoccupied by the grasshopper species found in the trap on 10-11 October, viz. 23-44 Hz(Table II), thus excluding non-grasshopper targets. Using this range, it was found thatthere was a statistically significant difference between the distribution for 18.17-22.27 h(before the arrival of the layer) and that for 23.24-00.35 h (during the passage of the layer)(f = 59-77, 15 d.f., F<0-001), with the 23.24-00.35 distribution having more wing-beat

GRASSHOPPER MIGRATION IN THE SAHEL 257
TABLE II. Wing-beat frequencies of grasshoppers common in the Tin Aouker lighttrap on 10-11 October 1978, estimated from tegmen length measurements,
by the relationships given in Riley & Reynolds (1983)
Species
Diabolocatantops axillarisOedaleus senegalensisOchrilidia gracilisPyrgomorpha conicaOchrilidia tibialis*Stenohippus epacromioides*^Ochrilidia geniculata*Aiolopus simulatrixAcrotylus longipes
Males
Mean tegmenlength (mm) ± 95%
confidence limits"
Estimated wing-beat frequency(Hz) (with 95%
confidence limits)
30-7723-9417-97161717-0814-7517-2118-1318-38
±0-77± 0-37+ 0-77± 0-57±0-54±0-26±0-43± 0-58± 0-50
26-330-937-740-539-043-038-8
25-9-26-8)30-5-31-2)36-7-38-8)39-5-41-538-2-39-942-5-43-538-2-39-5
37-5 36-7-38-337-2 (36-5-37-9)
Mean tegmenlength (mm) ± 95%
confidence limits3
FemalesEstimated wing-beat frequency
(Hz) (with 95%confidence limits)
35-9331-7625-9319-3422-4020-0328-1921-0420-94
±0-83±0-51± 0-83± 0-61± 0-67±0-30±0-73±0-70± 0-75
23-825-729-535-932-635-127-934-034-1
23-4-24-1)25-4-25-9)28-9-30-2)35-2-36-7)31-9-33-2)34-7-35-4)27-5-28-4)33-2-34-9)33-3-34-9)
" Tegmen lengths are derived from a sample of the grasshoppers caught in the light trap on 10-11 October 1978,except for species marked * where tegmen lengths were taken from Riley & Reynolds (1983).
t Referred to as Leva nigrovitlata (Krauss) in Riley & Reynolds (1983).
frequencies than expected in the 23-27-Hz class and fewer of the higher frequencies (above34 Hz).
The wing-beat frequency distribution for the period before the arrival of the layer wasthen compared with the accumulated distribution for all nights at Tin Aouker, and theywere found to be very similar (except for the presence in the former distribution of a fewmore wing-beat frequencies in the 41^44-Hz band). On the other hand, the distribution forthe period during the passage of the layer was significantly different (^ = 68-60, 16 d.f.,P<0-001) from the accumulated Tin Aouker distribution, with the peak of the formeroccurring at lower frequency (23-27 Hz). So it appears that the species composition of theinsects flying earlier in the evening of 10 October was rather typical for the Tin Aoukerarea, but the composition of the layer was distinctly different and contained a greaterproportion of a larger-sized species.
The estimated range of wing-beat frequencies of the D. axillaris caught in the trap onthe 10-11 October was 23-27 Hz, and the only other common grasshoppers contributing tothis range were females of O. senegalensis. Signatures in this 23-27-Hz interval comprised43% of those in the whole grasshopper band (23-44 Hz) during the overflight of the layer,and D. axillaris and females of O. senegalensis between them comprised a very similarpercentage (41%) of the grasshopper catch during the corresponding light-trapping period.This similarity suggests that the species composition of the insects flying in the layer wasvery like that of insects caught in the light trap, at least as regards the presence of D.axillaris, and it seems reasonable to conclude that the trap was catching individuals whichhad descended from the layer at the end of their migratory flight and were either flyingnear the ground or had landed in the vicinity of the trap.
Radar signatures recorded during a short session between 04.14 and 04.20 h on 11October, from insects flying after the passage of the layer and the line-echo, contained veryfew wing-beat frequencies below 30 Hz, i.e. very few signatures attributable to D. axillaris.
It is not possible to use radar signatures by themselves to assess which of the smallergrasshopper species contributed to the layer because the wing-beat frequencies of thedifferent species overlapped. However, females of Ochrilidia tibialis and O. geniculatawere possible candidates as their numbers increased in the trap during the period of theoverflight, and their wing-beat frequencies could be accommodated within the range of theappropriate signature distribution (Fig. 5b). (The estimated wing-beat frequencies of malesof O. tibialis and O. geniculata seemed too high to have produced the recorded signatures,but less than 10% of the specimens caught were males.) Signature evidence indicated thatO. gracilis (Krauss) and Aiolopus simulatrix (Walker) could have formed a minor com-ponent of the overflying layer.

258 D. R. REYNOLDS and J. R. RILEY
Tin Aouker 1978 Total
S 12-V
1 8 -a.
.9 4 -
No. of signatures-3362
A
0 10 20 30 40 50 60 70 80 90 100
Wing-beat frequency (Hz)
Fig. 4.—Histogram of the distribution of all the wing-beat frequencies obtained from the accumulated radar
signatures recorded at the Tin Aouker site.
Tin Aouker 10-11 Oct. 1978a
18.17-22.27 h10-1
No. of signatures = 306
o 00 10 20 30 40 50 60 70 80 90 100
Wing-beat frequency (Hz)
* 20-
23.24-00.35 h
No. of signatures-164
0 10 20 30 40 50 60 70 80 90 100Wing-beat frequency (Hz)
Fig. 5.—Histograms showing the distribution of wing-beat frequencies obtained from radar signatures re-corded at Tin Aouker on the night of 10-11 October1978 (a) between 18.17 and 22.27 h, i.e. before thearrival of the insect layer, and (b) between 23.24 and
00.35 h, i.e. during the passage of the layer.
Area densities and migration rates
The area density, i.e. the total number of insects flying above a unit ground area, canbe estimated by integrating the observed volume densities over the height interval between

Area densities(nos per hectare)
For height interval For height interval88-660 m
221234219211201221124
204
ground to 660 m
262291280245249242179250
GRASSHOPPER MIGRATION IN THE SAHEL 259
the ground and the flight ceiling (Drake & Farrow, 1983). Area densities calculated in thisway for the overflying concentration for various times between 23.30 and 02.00 h are givenin Table III. Two sets of area densities are shown, one calculated for the interval between88 and 660 m (for which volume densities were available) and one for the interval betweenthe ground and 660 m (which used, in addition, an extrapolated value for aerial densities atground level). There was little practical difference between these two sets of values, andthe average area density during the overflight period was about 200-250 individuals/ha. It isinteresting to note that even in this fairly dense migration, the area densities are 2-3 ordersof magnitude lower than the ground densities often found during grasshopper outbreaks oncrops, and this emphasizes the necessity for a postulated concentrating mechanism toexplain the magnitude of the outbreak densities.
TABLE III. Area densities of insects for varioustimes during the overflight of the layered
concentration
Time
23.3023.4500.0000.2500.3500.5002.00
Average
TABLE IV. Total migration rates for various timesduring the overflight of the layered con-centration, estimated by integrating thefluxes through aim2 'window' at each
altitude
Total migration rate(nos per metre per minute)
For height interval For height intervalTime 88-660 m ground to 660 m
15-016-415-714-313-912-98-3
The estimated number of insects crossing a line perpendicular to thedisplacement direction during the period from 23.30 to 02.00 h was 1632/mover the 88-660 m height interval and 1932/m in the ground to 660 m heightinterval.
Another quantity of interest is the net rate at which the insects at all heights werecrossing a unit line perpendicular to the direction of migration, termed 'the total migrationrate' by Drake & Farrow (1983). This is estimated by first calculating the 'flux' (i.e. therate at which insects pass through a unit area at right-angles to the direction of displace-ment) from the volume densities and displacement speeds at various heights, and thenintegrating the flux measurements over the whole height profile. Estimates of totalmigration rates, over the height interval from ground level to 660 m, ranged between 16/mper minute at 23.45 h and 8/m per minute at 02.00 h (Table IV). (For comparison,
23.3023.4500.0000.2500.3500.5002.00
12-613-412-712-711-611-86 1

2 6 0 D. R. REYNOLDS and J. R. RILEY
Drake & Farrow (1983) recorded total migration rates at about midnight in Chortoicetesterminifera (Walker) of from <l/m per minute on nights of weak migration up to 45/m perminute during a major overflight.) During the whole period of the present overflight (from23.30 to 02.00 h) about 1-9 million grasshopper-sized insects crossed each kilometre of animaginary line on the ground.
Flight trajectories and the source area of the overflying layer
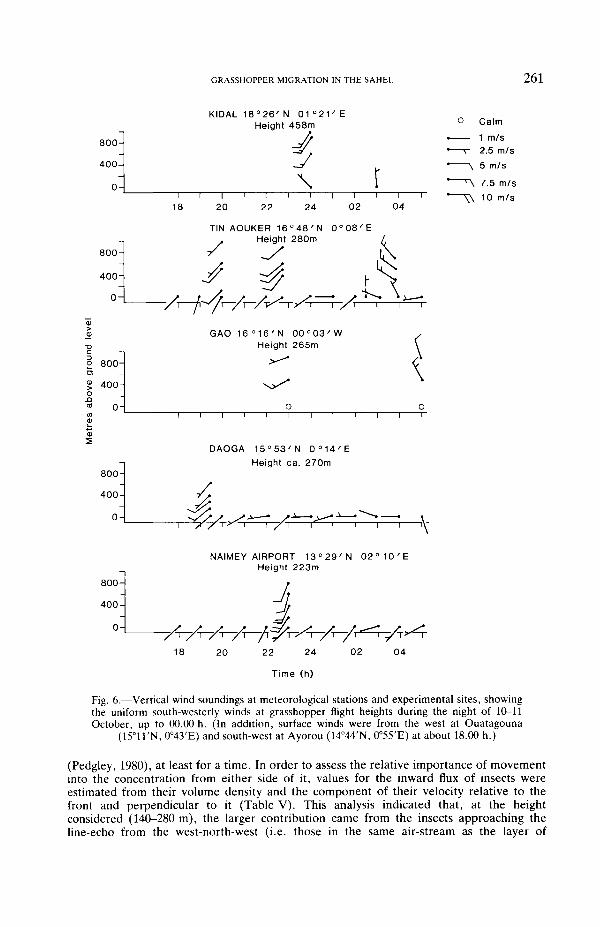
In order to identify the source area of the insects in the layered concentration, back-trajectories were constructed on the simple assumption that the insects' mean displacementspeeds and directions up-range of the radar site changed with time in the same way asthose measured at the radar. This method becomes progressively less accurate the furtherthe trajectory is carried from the radar, but it is unlikely to be subject to serious error overthe range (150 km) of the trajectories we plotted, because the wind-field was reasonablyuniform. (Wind direction at the insects' flight height appeared to be consistently from thesouth-west during the first half of the night at stations from Kidal to Niamey (Fig. 6).) Inthe trajectory calculations, grasshoppers were assumed to have taken-off at 18.30 h, i.e.during the time of the mass take-off at dusk observed every evening on the radar. We havenot observed, nor found evidence from the literature for, mass take-off of grasshopperslater in the night.
A back-trajectory for insects crossing the radar at 01.00 h on 11 October (i.e. about themiddle of the period of overflight of the layer) is shown in Fig. 7. The difference betweenthe general east-north-eastward direction of the trajectories (before midnight), and thedirection of the wind, which was blowing towards the north-east (Fig. 6), is attributable tothe contribution of the insects' air speed, which was directed towards the east-south-east.
During the night, the direction of the insects' displacement seen on the radar changedprogressively from east-north-eastward to east-south-eastward (and later towards thesouth-east), and this was reflected in the trajectories. This change was consistent with awidespread veer in wind direction from south-west to north-west as a cyclonic circulation(subsequently identified on synoptic charts) approached from the east, within thesouthward-moving Intertropical Convergence Zone. A corresponding change in insectdisplacement direction was observed by another entomological radar situated at Daoga(100 km to the south, see Fig. 7), where displacement was towards the north-east from19.00 h, changing to eastwards after 20.00 h and remaining thus until at least 22.30. Therewas thus much support from meteorological and radar observations for the validity of theback-trajectory calculations.
The start point (take-off point) of a trajectory for a group of insects overflying the radaridentifies the estimated source for those insects, and series of back-trajectory startingpoints can be joined to form a locus-line of sources. Two locus-lines for grasshoppersarriving at the radar between 23.30 and 01.00 h on 10-11 October, calculated to allow forthe variability of the displacement directions observed on the radar, are shown in Fig. 7.Together, the lines enclose the estimated source area of the migrating grasshoppers, whichwas in the Gourma (the area within the bend of the River Niger), particularly to the northand north-west of Doro (Fig. 7). Within this area are several wadis and depressions (e.g.In Elouan, Doukay, In Dirdene and Tegabaline) which receive run-off water from thesurrounding slightly higher ground and thus form seasonal wet areas with abundantvegetation. It is not known whether these harboured grasshopper populations, but it seemslikely that they did. Control against grasshopper outbreaks at densities of 30/m2 wasnecessary during October 1978 in the "region de Gossi" (O.C.La.Lav., 1978), whichindicates that conditions in the area were generally suitable for grasshopper development.
Concentration of flying insects by wind convergenceThe insects still in flight (below about 300 m) at 02.30 h, were observed to be displaced
towards the very dense, south-westward-moving line-echo which crossed the radar at thattime (Fig. 8), and they probably became trapped in this meso-scale wind-convergence zone
GRASSHOPPER MIGRATION IN THE SAHEL 261
800-
400-
0-
800-
400-
o-
o 800-
S 400-
o-
KIDAL 18°26' N 01 ° 2 1 ' EHeight 458m
\
18 20 22I I I
24 02 04
TIN AOUKER 1 6 ° 4 8 ' N 0 ° 0 8 ' EHeight 280m
GAO 16 ° 1 6 ' N 0 0 ° 0 3 ' WHeight 265m
\
"1 1 r
DAOGA 1 5 ° 5 3 ' N 0 o 1 4 ' E
Height ca. 270m
NAIMEY AIRPORT 1 3 ° 2 9 ' N 0 2 ° 1 0 ' EHeight 223m
18 04
Fig. 6.—Vertical wind soundings at meteorological stations and experimental sites, showingthe uniform south-westerly winds at grasshopper flight heights during the night of 10-11October, up to 00.00 h. (In addition, surface winds were from the west at Ouatagouna
(15°11'N, 0°43'E) and south-west at Ayorou (14°44'N, 0°55'E) at about 18.00 h.)
(Pedgley, 1980), at least for a time. In order to assess the relative importance of movementinto the concentration from either side of it, values for the inward flux of insects wereestimated from their volume density and the component of their velocity relative to thefront and perpendicular to it (Table V). This analysis indicated that, at the heightconsidered (140-280 m), the larger contribution came from the insects approaching theline-echo from the west-north-west (i.e. those in the same air-stream as the layer of

262 D. R. REYNOLDS and J. R. RILEY
Seasonal wet lands(Wadis.daias)
Fig. 7.—Typical back-trajectory (solid line) showing the estimated flight-path of insectstaking-off at 18.30 h on 10 October and crossing the Tin Aouker radar site at 01.00 h on 11October 1978. Locus-lines (dashed) join the estimated sources of insects in the layercrossing the radar between 23.00 and 01.00 h on 10-11 October. The two locus-lines showthe effect of the spread in the observed displacement directions, and together they enclosethe most likely source area. (The times on the locus-lines are the arrival times at the radar
of the insects taking-off from that source point.)
grasshoppers which earlier overflew the radar) rather than from those insects approachingfrom the north behind the line-echo. Thus, the overflying grasshoppers probably made themajor contribution to the high insect densities observed in the line-echo.
An approximate estimate of the time taken for the concentration to form can be madeby assuming that the insects entering the line-echo (from both sides) remain in a volume ofunit vertical cross-section and length equal to the distance (ca 1500 m) across the densepart of the line-echo. Then the time t required to accumulate the observed volume density(0-003/m3) is given by:
?(s) = volume density x distance across line-echo/flux
The estimated time (t) for the line-concentration to form was approximately 3 h, andthis was of the right order of magnitude for an event associated with a meso-scaleconvergence phenomenon. The estimate is rather crude because the insects within theconcentration were probably not completely constrained against upward or lateralmovement as envisaged here. Leakage of insects out of the concentration would tend toextend the time taken for its formation. On the other hand, the fluxes into the concentra-tion earlier in the evening may well have been greater, because of the higher insectdensities, than the ones measured by us.
As the grasshoppers were tending to land at this time of the night, the aerial
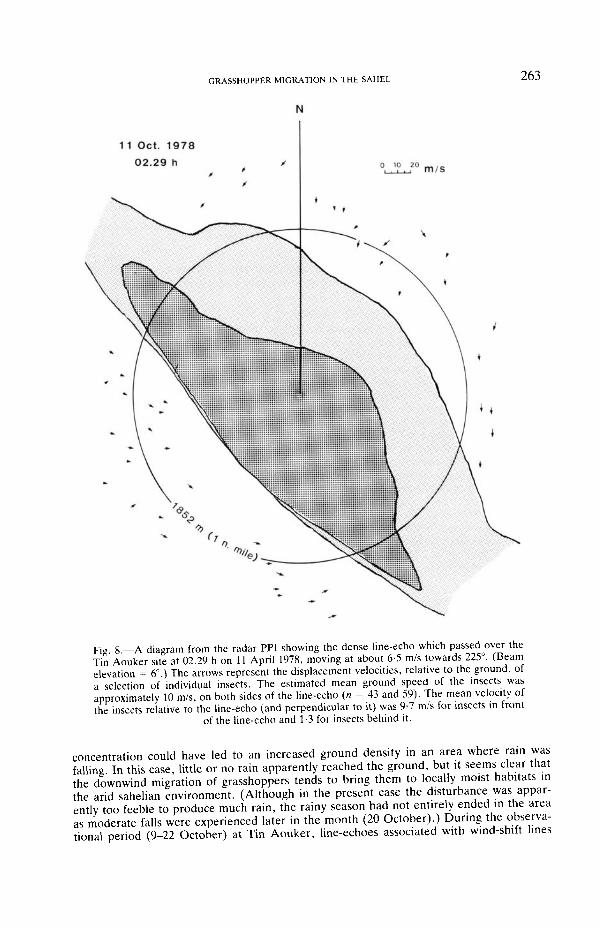
GRASSHOPPER MIGRATION IN THE SAHEL
N
263
11 Oct. 197802.29 h 0 10 20 m/s
• *
F i g 8 —A diagram from the radar PPI showing the dense line-echo which passed over theTin Aouker site at 02.29 h on 11 April 1978, moving at about 6-5 m/s towards 225°. (Beamelevation = 6°.) The arrows represent the displacement velocities, relative to the ground, ofa selection of individual insects. The estimated mean ground speed of the insects wasapproximately 10 m/s, on both sides of the line-echo (n = 43 and 59). The mean velocity ofthe insects relative to the line-echo (and perpendicular to it) was 9-7 m/s for insects in front
of the line-echo and 1-3 for insects behind it.
concentration could have led to an increased ground density in an area where rain wasfalling In this case, little or no rain apparently reached the ground, but it seems clear thatthe downwind migration of grasshoppers tends to bring them to locally moist habitats inthe arid sahelian environment. (Although in the present case the disturbance was appar-ently too feeble to produce much rain, the rainy season had not entirely ended in the areaas moderate falls were experienced later in the month (20 October).) During the observa-tional period (9-22 October) at Tin Aouker, line-echoes associated with wind-shift lines

Before line-echo
9-73-6 x 10-5
(at 02.26 h)
3-5 x 10~4
1-2 X(at 02
1-6 x
Behind
10"4
.34 h)
10-4
line-echo
1-35-2 x(at 02
6-9 x
10"5
.50 h)
10-'
2 6 4 D. R. REYNOLDS and J R. RILEY
TABLE V. Estimation of the insect flux (at a height of ca 140-280 m) into the line-concentration crossing the radar site at 02.27 h on 11 October 1978
Insect velocity relativeto line-echo (m/s)
Volume density(insccts/m3)
Flux(insects/m2/s)
The contribution of insects coming from west (and entering front of line-echo) is between two times and five timesgreater than the contribution of insects coming from north (and entering back of line-echo).
were seen on the radar at a frequency of more than one per night on average, althoughmost of them were not closely associated with rain echo within radar range.
Catches of Diabolocatantops axillaris on shipsIn years when grasshopper numbers are high in West Africa, various species have been
caught on ships approximately 100-200 km off the coast of Mauritania and Senegal inSeptember-October (TDRI archives, unpublished data; see also Ritchie, 1978). Infor-mation relating to D. axillaris captures at sea, is given in Table VI and Fig. 9. Examinationof synoptic-scale winds for stations in Mauritania and Senegal (M. Tucker, pers. comm.)reveals that in each case the D. axillaris individuals probably migrated on the strong north-easterly or easterly winds recorded near the surface (examples 2, 3 & 4 in Fig. 9) or nearerthe 850-mbar level (ca 1500 m) in the case of example 1. The flight times from the nearestcoast would be about 5-7 h, although if the insects had been on the wing since the dusktake-off (ca 18.30 h) until they were first seen on the ship, they would have been in flightfor about 14 h and could come from much further inland (see trajectories, Fig. 9). TableVI also includes an example of D. axillaris caught in the Gulf of Oman. In this case, thegrasshoppers probably flew on winds from the west, and if they had taken-off just afterdusk (19.30 h) they would have covered about 175 km when first seen on the ship. Atentative source area between the foothills of the Al Hajar al Gharbi Mountains and theBatinah coast near Suhar (24°22'N, 56°45'E), was indicated.
Although flight over the sea may stimulate insects to continue flying for unusually longperiods (Wolf et al., 1986), these records certainly demonstrate the potential for long-distance movement in D. axillaris.
DiscussionOne of the major problems of radar entomology is establishing the specific identity of
the insect targets detected flying over the radar (Riley, 1980). There is often little evidenceat ground level of the dense migrations observed by radar (Schaefer, 1976; Reid et al.,1979), and we have generally found, in studies in the Sahel, that the aerial density ofgrasshopper-like targets was poorly correlated with grasshopper numbers in light traps(Reynolds & Riley, unpublished). Similarly, Farrow (1979) found that light traps mostlycatch immigrant acridids, while emigrants, and migrants 'in transit' at altitude, were littleattracted. We were thus fortunate that in the present study there were enough immigrantscaught in the light trap to indicate the species composition of the migration observed by theradar. The radar-measured wing-beat frequencies were similar to those expected for thiscomposition, and this provided strong supporting evidence that certain grasshopper speciestaken at light were the same as those flying overhead. Thus we were able to conclude thatD. axillaris contributed to the overflying layer, together with other grasshopper species,quite probably Oedaleus senegalensis, and possibly, females of Ochrilidia tibialis, femalesof O. geniculata, O. gracilis and A. simulatrix.
D. axillaris and the other grasshoppers arriving at the radar site had probably beenflying continuously for about 6 h, an inference consistent with the fact that catches of D.

TA
BL
E V
I.
Som
e re
cord
s of
Dia
bolo
cata
ntop
s ax
illar
is c
augh
t abo
ard
ship
s of
f th
e W
est
Afr
ican
coa
st a
nd i
n th
e G
ulf
ofO
man
^
Shi
p
1 M
V S
hong
a
2 E
ssos
aba
Dat
e
29.ix
.74
Tim
e(G
MT
)
13.3
0
Pos
itio
n:A
tlan
tic
Oce
an
19°0
6'N
, 17
°52'
W
18.x
.75
09. (
X)
18°0
8'N
, 18
°(K
)'W
Win
ds a
t sh
ip
Dir
ecti
on
N b
y W
NE
3 M
V G
orji
stan
28
.x.7
1 08
.35
16°4
7'N
, 17
°45'
W
E b
y N
14.3
0 17
°53'
N,
17°4
3'W
20.x
.74
08.0
0 13
°48'
N,
17°4
2'W
N
NE
15.(K
) (a
t 08
.(X)
hrs)
4 SS
Act
6
5 M
V C
lan
Mac
lcod
5.
xi.7
4 12
.00
25°3
5'N
, 16
°23'
W
NE
by
N
Gul
f of
Om
an6
Bri
tish
25
.V.7
7 05
.00
24°2
2'N
, 58
°15'
ES
cien
tist
12
.00
23°2
5'N
, 59
°1 l
'E
t In
all
of t
hese
exa
mpl
es,
the
spec
imen
s w
ere
late
r po
siti
vely
id
enti
fied
.*
Dis
tanc
es f
rom
th
e po
siti
on w
hen
gras
shop
per s
wer
e fi
rst
sigh
ted.
Spe
ed
For
ce 4
(5-5
-7-9
m
/s)
12 k
nots
(6-2
m/s
)
For
ce
2-3
(1-6
-5-4
m
/s)
For
ce 5
(8-0
-10-
7 m
/s)
Oth
er s
peci
esca
ught
Rem
arks
Gr o
up o
f ab
out
20-3
0ob
serv
ed o
n bo
ard.
F
lyin
gsi
ngly
and
see
med
inde
pend
ent
of e
ach
othe
r .
Shi
p go
ing
sout
h, '
iocu
sts"
appe
arin
g on
boa
rd.
Dis
tanc
e to
near
est
land
2(K
1 km
140
km
205
km
Ana
crid
ium
A
ppea
ring
on
boar
d fr
om
180
kmm
elan
orho
don
08.3
5 to
14.
30 h
. S
ever
al
140
km(W
alke
r)
seen
fly
ing
roun
d sh
ip a
t08
.35
h.
Oed
aleu
s S
een
in f
light
, so
me
land
ing
85 k
mse
nega
lens
Ls
on d
eck.
90
km
—
1 in
divi
dual
cap
ture
d, n
o —
othe
rs s
een.
Pos
sibl
y on
shi
pfo
r so
me
tim
e as
rat
her
far
nort
h fo
r D
. ax
illa
ris.
N
otpl
otte
d.
Ana
crid
ium
"L
ocus
ts"
land
ing
on b
oard
. 15
0 km
*m
elan
orho
don
Win
ds l
ikel
y to
be
from
80
km
*W
/WN
W r
athe
r th
an N
or
S.
(130
km
)
Dir
ecti
on
tone
ares
tla
nd 25°
78° 50°
75°
85°
70°
100-
110°
20°
25-7
0°
270°
190°
(N)
o >o > o JO 2 a so > 5 5 H X m to ON

266 D. R. REYNOLDS and J. R. RILEY
i / r
Fig. 9.—Map showing the positions of four ships offWest Africa on which Diabolocatantops axillaris speci-mens were captured. The numbers refer to the indi-vidual cases documented in Table 6. Where thegrasshoppers were appearing at the ship over a periodof several hours, the initial and final positions of theship are joined by a solid line. The dashed lines repre-sent tentative back-trajectories from the position of theship when the grasshoppers were first sighted, assuming
take-off was at 18.30 h the previous evening.
axillaris on ships have shown that some individuals of this species are certainly capable offlights of at least 5-7 hours' duration. For the migration observed on the radar, a 6-h flightin the winds at flight altitude would have enabled the grasshoppers to cover distances of upto about 150 km, and it seems that nightly migration distances of this order are quitecommon in acridids of semi-arid areas. For example, Schaefer (1976) in the Sudandeduced, from the decline in area-densities through the night, that grasshoppers flying afterthe dusk peak period covered on average about 150 km in five hours' flight. In Australia,Drake & Farrow (1983) estimated from calculated trajectories that a proportion of the C.

GRASSHOPPER MIGRATION IN THE SAHEL 267
terminifera population studied regularly flew 150-200 km, and in Mali, Riley & Reynolds(1983) demonstrated that Oedaleus senegalensis and other species migrated at least 100 km.On some nights in the Sahel, when a low-level jet-stream wind (Drake, 1985) developsabove the surface temperature inversion, wind speeds of 60 km/h can occur, so thatgrasshoppers could cover distances of over 300 km by midnight (Riley & Reynolds, 1979 &unpublished).
D. axillaris spends the dry season as an adult in reproductive diapause or quiescenceand does not oviposit until the start of the next rains (May-June) (Joyce, 1952; Davey etal., 1959; Fishpool & Popov, 1984). Thus, a major problem faced by this species is thesurvival of the dry season (October-May). It seems probable that the extensive migrationsin October, such as the one reported here, enable the newly fledged adults to findlocalized, moister areas with suitable vegetation for food and cover in the increasingly aridSahel zone. A tendency for D. axillaris to become aggregated in the moister habitats as thedry season progressed was noted by Joyce (1952), but substantial mortality neverthelessoccurred during this period. D. axillaris has been taken at light throughout the wintermonths (Fishpool & Popov, 1984), which implies that some movement occurs in the middleof the dry season (perhaps as initially favourable habitats dry up), but migration at thistime seems much less extensive than in October (G. B. Popov, pers. comm.). Althoughthis impression could be partly caused because of the decrease in population size as the dryseason progresses, there may nevertheless be a real reduction in flight activity due to, forexample, the presence of suitable conditions (vegetation, etc.) in the immigration area, oras a response to seasonal cues such as the lower temperatures and humidities or the (albeitslight) shortening of daylength. Some decrease in flight activity would be expected merelyas a result of the December-January night-time temperatures often falling below thethreshold for flight. Whatever the mechanism, it appears to be reversed at the beginning ofthe next rainy season, when there seems to be an increase in D. axillaris migration (Joyce,1952, Popov, pers. comm.). Flights at this time presumably enable the grasshoppersactively to seek suitable oviposition sites in areas where the first sporadic rains of theseason have fallen (instead of having to wait passively for rain as in the case of specieswhich overwinter as diapausing eggs). D. axillaris thus shows the Class III migration(migration to and from imaginal diapause sites) of Johnson (1969).
D. axillaris, and probably many other species of sahelian grasshopper, are adapted touse the night-time synoptic-scale winds as their primary means of long-distance migration(Riley & Reynolds, 1979, 1983; Schaefer, 1976; Rainey, 1976, 1979). These wind-borneflights are an efficient behavioural strategy for finding suitable overwintering or breedingsites because the grasshoppers tend to be carried towards low-level wind convergencezones (usually the Intertropical Convergence Zone (ITCZ) or more intense meso-scaleconvergence zones associated with this) where rain is more likely to fall (cf. Rainey, 1951,1963, 1976). In the present case, for example, rain was seen on the radar, although little ornone reached the ground in the vicinity of the experimental site. Flight on synoptic-scalewinds means that grasshopper migration is often in directions largely dictated by the windrather than in a particular geographical direction. Far from the ITCZ, the night-time windsare likely to be strong and directionally uniform, and consequently the grasshoppers coverlarge distances at high ground speeds during a night's migration. Conversely, in the oftenlight and variable winds near the ITCZ, the insects would not displace far and would tendto remain near the convergence zone (as shown by Rainey (1963) for locust swarms). Thusthe distribution of the grasshoppers tends to change seasonally with the movements of theITCZ. During the present study, early in October, the ITCZ was to the north of the site,and the grasshoppers' displacement had a slight northwards component while being mainlyto the east. Later in October and in November, migration generally tends to carry thegrasshoppers southwards on the north-easterly or northerly winds which set in after thesouthward retreat of the ITCZ (e.g. Riley & Reynolds, 1979, 1983). At the beginning ofthe next rainy season, migratory flights on winds associated with the advancing ITCZwould tend to displace grasshoppers northwards again, enabling the species to recolonizethe vast area of the Sahel just as it is becoming favourable. There is increasing evidence

268 D. R. REYNOLDS and J R. RILEY
from ground surveys that populations of many sahelian grasshopper species do, in fact,move seasonally northwards and southwards with the ITCZ (Popov, 1976 & unpublished;Lecoq, 1978a & b; Duranton et al., 1979). The northward movements at the start of therains are much less well-documented than southward movements in autumn, partly becausethe northward-moving populations are much smaller. However, N. D. Jago (pers. comm.)has observed a mass influx of grasshoppers (mainly O. senegalensis and catanopinesincluding D. axillaris) to light at Mourdiah (14°28'N, 7°28'W) in the sahelian zone of Mali,on the evening of 30 June, 1986. The first heavy rains of the season had fallen in the areathe night before, and the grasshoppers almost certainly came from the south as no sizeablepopulations were present in the area or to the north of it.
Another consequence of the downwind displacement of flying grasshoppers is that theymay become concentrated, at least temporarily, in the intense horizontal convergence atmeso-scale wind-shift lines particularly of the 'density current' type (e.g. A. simuL rix instorm outflows in the Sudan (Schaefer, 1976), D. axillaris, O. senegalensis and otherspecies in wind-shift lines in Mali (Riley & Reynolds, 1983 & this paper) and grasshoppersand other insects in the sea-breeze front at Canberra, Australia (Drake, 1982)). If thegrasshoppers land before these aerial concentrations disperse, very dense ground popula-tions will result, which could be a hazard to crops in the manner suggested by Joyce (1976,1983).
Besides downwind displacement, other behaviour may well have evolved to assistsahelian grasshoppers to locate areas where rain has fallen. For example, weatherdisturbances, such as line-squalls, appeared to promote increased night flight activity(shown by increased catches at light) in O. senegalensis and other grasshoppers in the WestAfrican Sahel (L. D. C. Fishpool, pers. comm.; cf. Clark, 1971).
Nocturnally-migrating sahelian grasshoppers often show a measure of common orien-tation (Riley & Reynolds, 1986). In strong winds, the mean direction of orientation (ofseveral species including D. axillaris) is sometimes directly downwind, so that the insects'air speed is added to the wind speed (e.g. Riley & Reynolds, 1983). Thus their displace-ment speed is maximized and the energy required to cover a given distance minimized. Onother occasions, orientation is (as here) systematically off-wind. In these cases, the anglebetween the mean orientation direction and the direction towards which the wind isblowing appears to be always less than 90°, so that although the insects drift laterally due tothe wind, they avoid gross backwards displacement. The off-wind headings observed by ususually added a small but systematic southerly bias to the wind-induced displacement. Thesignificance of the off-wind headings is not clear; while the headings may merely resultfrom a 'mistake' in wind-direction finding, they might alternatively represent an attempt tobias movement towards a particular, ecologically advantageous, geographical direction(generally to the south in autumn) while retaining most of the advantages of wind-bornedisplacement. Schaefer (1976) reported common orientation towards the south-south-west(in not strongly-opposed winds) for A. simulatrix in the Sudan Gezira, which was said to beadaptive behaviour facilitating migration of the grasshoppers to their overwintering areas.Recently, Gibo (1986) has reported complex orientation responses to wind direction inmigrating monarch butterflies, Danausplexippus (L.), in North America, which seem to bedesigned to utilize wind power as much as is possible commensurate with escaping fromnorthern latitudes and attaining the overwintering sites in Mexico.
In light winds, at least in the middle Niger delta of Mali, unidentified insects, probablyacridids, orientated towards a particular geographical direction (25-50°) in a variety ofwind directions, producing displacement directions towards 300-35° (Riley & Reynolds,1986). Distances covered per evening in these conditions were much less than in strongwinds, but the direction of movement remained more under the insects' control. A north-eastwards geographical orientation in the middle Niger delta may facilitate the colonizationof habitats progressively uncovered from south-west to north-east in this area by theretreating floodwaters in December-February. For example, populations of Locusta migra-toria migratorioides (Reiche & Fairmaire), which do not diapause, follow the retreatingflood (against the prevailing north-easterly winds) and breed on the areas uncovered

GRASSHOPPER MIGRATION IN THE SAHEL 269
(Farrow, 1975): the orientation mechanism described above may be more adaptive in thesecircumstances than rapid downwind displacement which would carry the acridids out of themiddle Niger area. Thus, it would appear that while the main mode of migration insahelian grasshoppers involves rapid, downwind displacement, there may also be shorter-range movements, not particularly related to wind direction, under certain circumstances.These non-wind-related orientations would permit detailed exploration of habitats within acircumscribed area. Specialized orientation mechanisms in strongly flying migrants maysubstantially reduce mortality during migration, as indeed has been suggested by R. A.Farrow (in Hughes, 1979) for L. m. migratorioides in the middle Niger area, and the ideathat losses during insect migration are less than previously supposed is generally gainingground (Rankin, 1985).
The apparently obligate long-range migrations of sahelian grasshoppers, such as the onedocumented here, are a necessary adaptation to the temporary and fragmented nature ofhabitats in the semi-arid sahelian zone, where the spatial and temporal distribution ofrainfall is shifting and erratic, particularly at the beginning and end of the rains.'Somewhere else' will usually tend to be more favourable than the present habitat, so thatthe risks associated with migration and the finding of a suitable new habitat are less thanthose inherent in 'staying put' (Southwood, 1977; Dingle, 1980). The high mobility of thegrasshoppers greatly facilitates migration to new habitats as these become available andhelps to ensure that all areas favourable for D. axillaris survival will be found andexploited. High mobility may also help evade the build-up of natural enemies and diseases.A strong indication that migratory flight in grasshoppers of the Sahel and Sudan savanna isecologically essential is shown by the fact that complete Sightlessness appears not to haveevolved in these areas, but only in those species occurring in the more stable habitats southof 12°N (i.e. in the Guinea savanna and forest zones) (Fishpool & Popov, 1984). Inaddition to the patchiness of the habitat due to spatial and temporal variability of therainfall, the sahelian grasshoppers have to contend with the general deterioration of theenvironment in the dry season. The majority of sahelian grasshoppers have responded byevolving a period of diapause or quiescence, either in the adult (like D. axillaris) or in theegg stage (Fishpool & Popov, 1984; Lecoq, 1978a). Solbreck (1978) has pointed out thatwhere the present habitat is deteriorating at the same time as other habitats are tending todeteriorate, then migration would be expected to be of the Class III type, followed bydiapause, rather than Class I type (migration by young adults quickly followed bybreeding). Thus, the evolved life-history stratagem of D. axillaris entails breeding not only'elsewhere' but also 'much later' (Southwood, 1977; Solbreck, 1978; Dingle, 1980).
AcknowledgementsThis study formed part of a joint research project undertaken by the Centre for
Overseas Pest Research (now the TDRI) and the Organisation Commune de LutteAntiacridienne et de Lutte Antiaviare. We gratefully acknowledge the assistance of theproject staff, particularly Mr G. B. Popov, Dr L. D. C. Fishpool, Mr S. Coulibaly and MrP. M. S. Ward. For their collaboration and active participation in the field work, we thankDrs E. A. Bernays and R. F. Chapman (formerly of TDRI), Dr M. J. Farmery (formerlyof York University), and Drs N. D. Jago and J. M. Ritchie (TDRI). We are particularlyindebted to Mr A. D. Smith (TDRI) for maintaining the radars under difficult conditions.We thank Mr Popov, Dr Jago and Dr Fishpool for discussions of sahelian grasshoppermigration, and Dr R. C. Rainey and Dr R. A. Farrow for their comments on a draft of thepaper. We are also indebted to Miss S. M. Green for statistical advice, to Mrs G. Barfieldand Mr J. McNeil for assistance with the analysis of the radar data, and to Mr D. E.Pedgley and Mr M. R. Tucker for the synoptic weather analysis.
ReferencesCHEKE, R. A., FISHPOOL, L. D. C. & FORREST, G. A. (1980). Oedaleus senegalensis (Krauss)
(Orthoptera: Acrididae: Oedipodinae): an account of the 1977 outbreak in West Africa andnotes on eclosion under laboratory conditions.—Acrida 9, 107-132.

2 7 0 D R. REYNOLDS and J. R RILEY
CLARK, D. P. (1971). Flights after sunset by the Australian plague locust, Chortoicetes terminifera(Walk.), and their significance in dispersal and migration.—Aust. J. Zool. 19, 159-176.
COPR (CENTRE FOR OVERSEAS PEST RESEARCH) (1982). The locust and grasshopper manual.—690 pp. London, Cent. Overseas Pest Res.
DAVEY, J. T., DESCAMPS, M. & DEMANGE, R. (1959). Notes on the Acrididae of the French Sudan,with special reference to the Central Niger Delta.—Bull. Inst. fr. Afr. Noire (A) 21, 68-112565-600.
DINGLE, H. (1980). Ecology and evolution of migration.—pp. 1-101 in Gauthreaux, S. A. (Ed.).Animal migration, orientation, and navigation.—387 pp. New York, Academic Press.
DINGLE, H. (1986). Evolution and genetics of insect migration.—pp. 11-26 in Danthanarayana, W.(Ed.). Insect flight: dispersal and migration.—289 pp. Berlin, Springer-Verlag.
DRAKE, V. A. (1982). Insects in the sea-breeze front at Canberra: a radar study.—Weather, Lond.37, 134-143.
DRAKE, V. A. (1985). Radar observations of moths migrating in a nocturnal low-level jet.—Ecol.Enlomol. 10, 259-265.
DRAKE, V. A. & FARROW, R. A. (1983). The nocturnal migration of the Australian plague locust,Chortoicetes terminifera (Walker) (Orthoptera: Acrididae): quantitative radar observationsof a series of northward flights.—Bull. ent. Res. 73, 567-585.
DURANTON, J. F., LAUNOIS, M., LAUNOIS-LUONG, M. H. & LECOQ, M. (1979). Biologie et ecologiede Catantops haemorrhoidalis en Afrique de l'Ouest (Orthopt. Acrididae).—Annls Soc. ent.Fr. (N.S.) 15, 319-343.
FARROW, R. A. (1975). The African migratory locust in its main outbreak area of the Middle Niger:quantitative studies of solitary populations in relation to environmental factors.—Locustano. 11, 198 pp.
FARROW, R. A. (1979). Population dynamics of the Australian plague locust, Chortoicetes terminifera(Walker), in central western New South Wales. I. Reproduction and migration in relation toweather.—Aust. J. Zool. 27, 717-745.
FISHPOOL, L. D. C. & POPOV, G. B. (1984). The grasshopper faunas of the savannas of Mali, Niger,Benin and Togo.—Bull. Inst. Fond. Afr. Noire (A) 43, 275-410.
GIBO, D. L. (1986). Flight strategies of migrating monarch butterflies (Danaus plexippus L.) insouthern Ontario.—pp. 172-184 in Danthanarayana, W. (Ed.). Insect flight: dispersal andmigration.—289 pp. Berlin, Springer-Verlag.
GUNN, D. L. & RAINEY, R. C. (Eds) (1979). Strategy and tactics of control of migrant pests.—Phil.Trans. R. Soc. (B) 287, 245-488.
HUGHES, R. D. (1979). Movement in population dynamics.—pp. 14-34 in Rabb, R. L. & Kennedy,G. G. (Eds). Movement of highly mobile insects: concepts and methodology in research.—456 pp. Raleigh, North Carolina, N. Carolina St. Univ.
JAGO, N. D. (1984). The alate genera of East African Catantopinae (Orthoptera, Acridoidea)including revision of the genus Catantops Schaum.—Trans. Am. ent. Soc. 110, 295—387.
JOHNSON, C. G. (1969). Migration and dispersal of insects by flight.—763 pp. London, Methuen.JOYCE, R. J. V. (1952). The ecology of grasshoppers in East Central Sudan.—Anti-Locust Bull. no.
11, 96 pp.JOYCE, R. J. V. (1976). Insect flight in relation to problems of pest control.—pp. 135-155 in Rainey,
R. C. (Ed.). Insect flight.—287 pp. Oxford, Blackwell (Symp. R. Ent. Soc. no. 7).JOYCE, R. J. V. (1981). The control of migrant pests.—pp. 209-229 in Aidley, D. J. (Ed.). Animal
migration.—264 pp. Cambridge, Univ. Press.JOYCE, R. J. V. (1983). Aerial transport of pests and pest outbreaks.—EPPO Bull. 13 (2), 111-119.LECOQ, M. (1978a). Biologie et dynamique d'un peuplement acridien de zone soudanienne en
Afrique de l'Ouest (Orthoptera, Acrididae).—Annls Soc. ent. Fr. (N.S.) 14, 603-681.LECOQ, M. (1978fc). Les deplacements par a grande distance chez les acridiens des zones sahelienne
et soudanienne en Afrique de l'Ouest.—C. r. hebd. Seanc. Acad. Sci., Paris (D) 286, 419—422.
O.C.LA.LAV. (ORGANISATION COMMUNE DE LUTTE ANTIACRIDIENNE ET DE LUTTE ANTIAVIAIRE)(1978). Signalisation Acridienne, October 1978.—6 pp. Dakar, O.C.La.Lav. (Bull. no. 10).
PEDGLEY, D. E. (1980). Weather and airborne organisms.—Tech. Notes Wld met. Org. no. 173,115 pp.
POPOV, G. B. (1976). The 1974 outbreak of grasshoppers in western Africa.—pp. 35-43 in Smith,R. F. & Schlegel, D. E. (Eds). Report of the Sahel Crop Pest Management Conference, heldat the invitation of the Agency for International Development, United States Department ofState, Washington DC, USA, December 11-12, 1974. Berkeley, Univ. California (for UnitedStates Agency for International Development).

GRASSHOPPER MIGRATION IN THE SAHEL 271
RAINEY, R. C. (1951). Weather and the movements of locust swarms: a new hypothesis.—Nature,Lond. 168, 1057-1060.
RAINEY, R. C. (1963). Meteorology and the migration of desert locusts: applications of synopticmeteorology in locust control.—Tech. Notes Wld met. Org. no. 54, 115 pp. (Also as Anti-Locust Mem. no. 7.)
RAINEY, R. C. (1976). Flight behaviour and features of the atmospheric environment.—pp. 75-112in Rainey, R. C. (Ed.). Insect flight.—287 pp. Oxford, Blackwell (Symp. R. Ent. Soc.no. 7).
RAINEY, R. C. (1979). Dispersal and redistribution of some Orthoptera and Lepidoptera by flight.—Mitt, schweiz. ent. Ges. 52, 125-132.
RANKIN, M. A. (1985). Introduction.—pp. 1-4 in Rankin, M. A. (Ed.). Migration: mechanisms andadaptive significance.—868 pp. Port Aransas, Texas, Marine Sci. Inst. (Contributions inMarine Science, supplement vol. 7).
REID, D. G., WARDHAUGH, K. G. & ROFFEY, J. (1979). Radar studies of insect flight at Benalla,Victoria, in February 1974.—Tech. Pap. Div. Ent. C.S.I.R.O. Aust. no. 16, 21 pp.
RILEY, J. R. (1974). Radar observations of individual desert locusts (Schistocerca gregaria (Forsk.)(Orthoptera, Locustidae)).—Bull. ent. Res. 64, 19-32.
RILEY, J. R. (1975). Collective orientation in night-flying insects.—Nature, Lond. 253, 113-114.RILEY, J. R. (1980). Radar as an aid to the study of insect flight.—pp. 131-140 in Amlaner, C. J.,
Jr. & Macdonald, D. W. (Eds). Handbook on biotelemetry and radio tracking.—804 pp.Oxford, Pergamon Press.
RILEY, J. R. & REYNOLDS, D. R. (1979). Radar-based studies on the migratory flight of grasshoppersin the middle Niger area of Mali.—Proc. R. Soc. (B) 204, 67-82.
RILEY, J. R. & REYNOLDS, D. R. (1983). A long-range migration of grasshoppers observed in thesahelian zone of Mali by two radars.—J. Anim. Ecol. 52, 167-183.
RILEY, J. R. & REYNOLDS, D. R. (1986). Orientation at night by high-flying insects.—pp. 71-87 inDanthanarayana, W. (Ed.). Insect flight: dispersal and migration.—289 pp. Berlin, Springer-Verlag.
RITCHIE, J. M. (1978). Melanism in Oedaleus senegalensis and other oedipodines (Orthoptera,Acrididae).—Jnlnat. Hist. 12, 153-162.
SCHAEFER, G. W. (1976). Radar observations of insect flight.—pp. 157-197 in Rainey, R. C. (Ed).Insect flight.—287 pp. Oxford, Blackwell (Symp. R. Ent. Soc. no. 7).
SOLBRECK, C. (1978). Migration, diapause, and direct development as alternative life histories in aseed bug, Neacoryphus bicrucis.—pp. 195-217 in Dingle, H. (Ed.). Evolution of insectmigration and diapause.—284 pp. New York, Springer-Verlag.
SOUTHWOOD, T. R. E. (1977). Habitat, the templet for ecological strategies?—J. Anim. Ecol. 46,337-365.
TAYLOR, L. R. (1986). The four kinds of migration.—pp. 265-280 in Danthanarayana, W. (Ed.).Insect flight: dispersal and migration.—289 pp. Berlin, Springer-Verlag.
WOLF, W. W., SPARKS, A. N., PAIR, S. D., WESTBROOK, J. K. & TRUESDALE, F. M. (1986). Radar
observations and collections of insects in the Gulf of Mexico.—pp. 221-234 in Danthanaray-ana, W. (Ed.). Insect flight: dispersal and migration.—289 pp. Berlin, Springer-Verlag.
(Received 1 September 1987)
© C A B International, 1988