
The Chicken Anemia Virus-Derived Protein Apoptin Requires Activation of Caspases for Induction of...
8
10.1128/JVI.74.15.7072-7078.2000. 2000, 74(15):7072. DOI: J. Virol. M. Noteborn A. A. A. M. Danen-van Oorschot, A. J. van der Eb and M. H. Cells for Induction of Apoptosis in Human Tumor Apoptin Requires Activation of Caspases The Chicken Anemia Virus-Derived Protein http://jvi.asm.org/content/74/15/7072 Updated information and services can be found at: These include: REFERENCES http://jvi.asm.org/content/74/15/7072#ref-list-1 at: This article cites 46 articles, 26 of which can be accessed free CONTENT ALERTS more» articles cite this article), Receive: RSS Feeds, eTOCs, free email alerts (when new http://journals.asm.org/site/misc/reprints.xhtml Information about commercial reprint orders: http://journals.asm.org/site/subscriptions/ To subscribe to to another ASM Journal go to: on October 16, 2014 by guest http://jvi.asm.org/ Downloaded from on October 16, 2014 by guest http://jvi.asm.org/ Downloaded from
-
Upload
independent -
Category
Documents
-
view
2 -
download
0
Transcript of The Chicken Anemia Virus-Derived Protein Apoptin Requires Activation of Caspases for Induction of...

10.1128/JVI.74.15.7072-7078.2000.
2000, 74(15):7072. DOI:J. Virol. M. NotebornA. A. A. M. Danen-van Oorschot, A. J. van der Eb and M. H. Cellsfor Induction of Apoptosis in Human TumorApoptin Requires Activation of Caspases The Chicken Anemia Virus-Derived Protein
http://jvi.asm.org/content/74/15/7072Updated information and services can be found at:
These include:
REFERENCEShttp://jvi.asm.org/content/74/15/7072#ref-list-1at:
This article cites 46 articles, 26 of which can be accessed free
CONTENT ALERTS more»articles cite this article),
Receive: RSS Feeds, eTOCs, free email alerts (when new
http://journals.asm.org/site/misc/reprints.xhtmlInformation about commercial reprint orders: http://journals.asm.org/site/subscriptions/To subscribe to to another ASM Journal go to:
on October 16, 2014 by guest
http://jvi.asm.org/
Dow
nloaded from
on October 16, 2014 by guest
http://jvi.asm.org/
Dow
nloaded from

JOURNAL OF VIROLOGY,0022-538X/00/$04.0010
Aug. 2000, p. 7072–7078 Vol. 74, No. 15
Copyright © 2000, American Society for Microbiology. All Rights Reserved.
The Chicken Anemia Virus-Derived Protein Apoptin RequiresActivation of Caspases for Induction of Apoptosis
in Human Tumor CellsA. A. A. M. DANEN-VAN OORSCHOT,1,2 A. J. VAN DER EB,1† AND M. H. M. NOTEBORN1,2*
Department of Molecular Cell Biology, Leiden University Medical Center,1 and Leadd BV,2 Leiden, The Netherlands
Received 26 October 1999/Accepted 26 April 2000
The chicken anemia virus protein Apoptin has been shown to induce apoptosis in a large number oftransformed and tumor cell lines, but not in primary cells. Whereas many other apoptotic stimuli (e.g., manychemotherapeutic agents and radiation) require functional p53 and are inhibited by Bcl-2, Apoptin actsindependently of p53, and its activity is enhanced by Bcl-2. Here we study the involvement of caspases, animportant component of the apoptotic machinery present in mammalian cells. Using a specific antibody, activecaspase-3 was detected in cells expressing Apoptin and undergoing apoptosis. Although Apoptin activity wasnot affected by CrmA, p35 did inhibit Apoptin-induced apoptosis, as determined by nuclear morphology. Cellsexpressing both Apoptin and p35 showed only a slight change in nuclear morphology. However, in most of thesecells, cytochrome c is still released and the mitochondria are not stained by CMX-Ros, indicating a drop inmitochondrial membrane potential. These results imply that although the final apoptotic events are blocked byp35, parts of the upstream apoptotic pathway that affect mitochondria are already activated by Apoptin. Takentogether, these data show that the viral protein Apoptin employs cellular apoptotic factors for induction ofapoptosis. Although activation of upstream caspases is not required, activation of caspase-3 and possibly alsoother downstream caspases is essential for rapid Apoptin-induced apoptosis.
Although many viruses encode apoptotic inhibitors, a num-ber of viruses have been found to carry genes specifying apo-ptosis-inducing proteins (35, 39, 41). Apoptin, a 13.6-kDa pro-tein encoded by the chicken anemia virus, is one such geneproduct. In cell culture, expression of Apoptin is sufficient toinduce apoptosis (27). Interestingly, Apoptin only induces apo-ptosis in transformed or tumor-derived cells and not in normaldiploid or primary cells of human or rodent origin (9; Y.Zhuang, unpublished results). In contrast to most chemother-apeutic agents, Apoptin induces apoptosis in cells lacking func-tional p53 or overexpressing Bcl-2 (47, 48). When cotrans-fected, Bcl-2 even enhances Apoptin activity (8, 10). In orderto understand how Apoptin induces apoptosis, further insightinto the involvement of known apoptotic effectors is required.
Several observations indicate that the mitochondria play animportant role in the commitment to programmed cell death(13, 15, 19). Many apoptotic stimuli (e.g., Bax, oxidants, andhigh Ca21) induce a loss of mitochondrial membrane integrity.Following a drop in the mitochondrial inner membrane poten-tial (DCm), it is thought that either a permeability transitionpore opens or the outer mitochondrial membrane is physicallydisrupted. In either case, this results in release of proapoptoticmolecules from the intermembrane space, such as procaspases(24, 36), apoptosis-inducing factor (37), and cytochrome c,which can act as a cofactor for caspase activation (22). Disrup-tion of the mitochondrial membrane also leads to a drop incellular ATP and production of reactive oxygen species, al-though this seems to occur relatively late in apoptosis (15).Antiapoptotic Bcl-2 family members, like Bcl-2 and Bcl-xL,which block cytochrome c release from mitochondria and in-
hibit opening of the permeability transition pore, can com-pletely rescue cells from cell death induced by many differentstimuli (1, 31, 42). However, not all apoptotic stimuli are in-hibited by Bcl-2. It has been proposed that there is also amitochondrion-independent pathway, feeding directly into thecaspase cascade, which is not inhibited by Bcl-2 (33).
Caspases play a major role in the execution phase of apo-ptosis (7, 14, 28, 40) by cleaving a large number of proteins,which in turn leads to the typical morphology of apoptosis.Among these substrates are cytoskeletal and structural pro-teins, DNA repair enzymes, transcription factors, protein ki-nases, and proteins involved in cell cycle regulation (C. Strohand K. Schulze-Osthoff, Editorial, Cell Death Differ. 5:997–1000, 1998). Also, some of the antiapoptotic Bcl-2 family mem-bers have been found to be cleaved by caspases (5, 6). Caspasesall cleave after an aspartic acid residue. Specificity is largelydetermined by the tetrapeptide directly N terminal to thecleavage site (26). Caspases exist as inactive zymogens in thecell which become activated upon proteolytic cleavage by othercaspases or by autocatalysis. Functionally, they can be dividedinto initiator (upstream) and effector (downstream) caspases.Different apoptotic signals activate different initiator caspases,in turn activating the effector caspases, resulting in a cascade ofcaspase activation. Cleavage of procaspases can be regulatedby self-oligomerization (44), compartmentalization (24, 36),the availability of cofactors like cytochrome c (22), and thepresence of cellular inhibitors (11, 32). It has been shown thatcaspases can activate cytosolic factors, e.g., Bid, which inducethe release of cytochrome c from mitochondria, possibly actingas an amplification loop during apoptosis (2, 21, 23).
For viruses, blocking apoptosis is a way to circumvent thecellular defense mechanism against viral infection, and manyof them have evolved their own caspase inhibitors, like CrmAfrom cowpox virus (30) and p35 from baculovirus (4). How-ever, caspase inhibitors have also been found in mammals; forexample, the IAP (inhibitor of apoptosis) family has both
* Corresponding author. Mailing address: Leadd BV, Wassenaarse-weg 72, 2333 AL Leiden, The Netherlands. Phone: 31 (0)71 527 8736.Fax: 31 (0)71 527 1736. E-mail: [email protected].
† Present address: Radiation Genetics and Chemical Mutagenesis,Leiden University Medical Center, Leiden, The Netherlands.
7072
on October 16, 2014 by guest
http://jvi.asm.org/
Dow
nloaded from

mammalian and viral homologs (11, 32, 34). Inhibition ofcaspase activation blocks the appearance of apoptotic mor-phology, illustrating the important role of caspases in the ex-ecution phase of apoptosis. However, blocking caspases doesnot necessarily lead to cell survival. In several cases, the apo-ptotic morphology is inhibited but clonogenicity is lost, andeventually the cells still die, albeit more slowly (16, 25). Theseresults imply that the commitment to undergo programmedcell death is made upstream of the activation of the caspasecascade.
In this study, we used several inhibitors of caspases withdifferent specificities to determine the involvement of caspasesand mitochondrial factors in Apoptin-induced apoptosis. Ap-optin-induced apoptosis exhibits remarkable specificity fortransformed or tumorigenic cells. Therefore, determining howApoptin interacts with the cell’s normal apoptotic pathways isimportant. Evidence is provided that Apoptin can indeed uti-lize the cell’s effector caspases yet remains independent of theinitiator caspase pathways.
MATERIALS AND METHODS
Cell culture. The human osteosarcoma cell line Saos-2, which lacks functionalp53, was cultured in Dulbecco’s modified Eagle’s medium containing 10% fetalcalf serum, penicillin, and streptomycin (Life Technologies, Rockville, Md.). Oneday prior to transfection, the cells were seeded in dishes containing uncoatedglass slides. At the time of transfection, the cells were 30 to 50% confluent.
Plasmids and transfection. The plasmids pCMV-VP3, encoding Apoptin,pCMV-p53, encoding p53, and pCMV-neo-Bam, the empty control plasmid,have been described previously (27, 47). The cDNAs for CrmA and p35 (kindlydonated by D. J. Pickup and L. K. Miller, respectively) were each subcloned intothe BamHI site of pCMV and confirmed by restriction enzyme analysis andsequencing, generating pCMV-CrmA and pCMV-p35. Expression of CrmA frompCMV-CrmA was shown by an in vitro transcription-translation assay (S. Olijs-lagers, unpublished results). Expression of p35 in Saos-2 cells transfected withpCMV-p35 was shown by Western blot analysis using a specific antibody againstp35 (kindly donated by L. K. Miller).
phGFP-S65T (Clontech, Palo Alto, Calif.) was used to generate phGFP-VP3,fusing Apoptin to the C terminus of green fluorescent protein (GFP), under thecontrol of a cytomegalovirus promoter. The plasmid was tested by sequencing,and expression of the fusion protein was confirmed by Western blot analysis. Intransfection assays, GFP-Apoptin had the same localization and activity as wild-type Apoptin in tumor cells (A. van Zon, unpublished results). pcDNA3.1/MycHis/LacZ (Invitrogen, Carlsbad, Calif.) encodes LacZ with both a myc tagand a His tag attached to the C terminus. All plasmids were purified with Jetstarmaxiprep columns (Genomed, Bad Oeyenhausen, Germany). Saos-2 cells weretransfected by the CaPO4-method as described previously (43), with 5 to 6 mg ofplasmid DNA per 6-cm-diameter dish or 3 mg per well of a 6-well plate or3.5-cm-diameter dish. In cotransfections, the ratio of pCMV-VP3 to pCMV-CrmA or pCMV-p35 was always 1:2.
Immunofluorescence assays and antibodies. Two to 5 days after transfection,cells were fixed with 80% acetone for 10 min and kept at 220°C until furtherstaining. For the antibody staining, the cells were first incubated with phosphate-buffered saline (PBS) plus 0.05% Tween 20 (PBS-Tween) plus 5% normal goatserum (NGS) for 30 min, incubated with the first antibody in PBS-Tween plus5% NGS for 1 h, washed with PBS-Tween, and incubated with the secondantibody in PBS-Tween plus 5% NGS for 1 h. Finally, the cells were washed withPBS-Tween and embedded in 90% glycerol–0.1 M Tris (pH 8.0) containing 2.3%Dabco (diazabicyclo-[2,2,2]-octane) to prevent quenching of the signal and 1 mgof DAPI (2,4-diamidino-2-phenylindole)/ml to stain the DNA. The cells wereanalyzed by fluorescence microscopy for expression of the transfected protein,and nuclear morphology indicating the apoptotic state of the cell was determinedby DAPI staining. Staining with anti-cytochrome c was done in essentially thesame way, except that fixation of the cells was done with 50% methanol–50%acetone for 5 min, incubation with the first antibody was done for 3 h instead of1 h, and 5% NGS was replaced with 3% bovine serum albumin.
For propidium iodide (PI) exclusion, cells were first incubated with 5 mg ofPI/ml in the medium for 20 min, washed with PBS, and then fixed with formal-dehyde-methanol-acetone (sequential incubation with 1% formaldehyde for 10min, 100% cold methanol for 5 min, and 80% cold acetone for 2 min). In thestaining experiments with the mitochondrion-specific dye CMX-Ros, cells werewashed with PBS, stained with 100 nM Mito Tracker Red CMX-Ros (MolecularProbes, Eugene, Oreg.) in PBS for 30 min, washed with PBS, and fixed withformaldehyde-methanol-acetone.
The mouse monoclonal antibody CVI-CAV-111.3 was used to detect Apoptin(supernatant was diluted 1:3) (9). A rabbit polyclonal antibody, RaVP3-C, raisedagainst part of the C terminus of Apoptin, was used in double stainings (dilution,
1:200). p53 expression was detected with the mouse monoclonal antibody DO-1(dilution, 1:100) (Santa Cruz Biotechnology, Santa Cruz, Calif.). The mousemonoclonal antibody 6H2.B4 was used to detect cytochrome c (dilution, 1:200),and a rabbit polyclonal antibody was used to detect active caspase-3 (dilution,1:200) (both from Pharmingen, San Diego, Calif.). Second antibodies were con-jugated either to fluorescein isothiocyanate or to rhodamine (dilution, 1:100)(Jackson ImmunoResearch Laboratories, West Grove, Pa.).
Subcellular fractionation and Western blot analysis. Subcellular fractionationwas performed essentially as described by Juin et al. (18). Saos-2 cells were grownin 15-cm-diameter dishes and transfected with FuGENE 6 (Roche MolecularBiochemicals, Indianapolis, Ind.) according to the manufacturer’s protocol. Thecells were washed with PBS and then scraped and centrifuged at 2,000 3 g for 5min. The cells were then resuspended in cell extraction buffer (300 mM sucrose,10 mM HEPES [pH 7.4], 50 mM KCl, 5 mM EGTA, 5 mM MgCl2, 1 mMdithiothreitol, 1 mM phenylmethylsulfonyl fluoride, 100 mM cytochalasin B), lefton ice for 30 min, and homogenized by 50 strokes in a Dounce homogenizer.Unbroken cells and nuclei were pelleted by centrifugation for 5 min at 2,000 3g. Heavy membranes were removed from the resulting supernatant by centrifu-gation for 5 min at 13,000 3 g. The resulting supernatant was the crude cytosolicfraction. All samples were frozen in liquid nitrogen and stored at 280°C.
For Western blotting, 30 mg of protein was loaded in each lane of a sodiumdodecyl sulfide–15% polyacrylamide gel, separated by electrophoresis, and elec-troblotted onto Immobilon-P membranes (Millipore, Bedford, Mass.). The blotswere then incubated with the monoclonal antibody 7H8.2C12 (Pharmingen)(1:1,000) to detect cytochrome c, and positive signals were visualized by en-hanced chemiluminescence (Amersham, Piscataway, N.J.) according to the man-ufacturer’s protocol.
RESULTS
Apoptin requires activation of downstream caspases forrapid induction of apoptosis. The viral inhibitors CrmA andp35 show different specificities for the various caspases. In vitrostudies of binding kinetics indicate that CrmA mainly repressesupstream caspases, e.g., caspases 1 and 8, and has little effecton the more downstream caspases 3, 6, and 7 (12, 46). Com-parable studies of p35 show that it is a more general caspaseinhibitor which inhibits both upstream and downstreamcaspases (45). To determine which caspases are involved inApoptin-induced apoptosis, plasmids encoding these inhibitorswere cotransfected with Apoptin in Saos-2 cells, a human tu-mor cell line which lacks endogenous p53. Transfection withp53 was used as a positive control for apoptosis induction inthese cells. Induction of apoptosis was scored by analysis ofnuclear morphology by DAPI staining. Intact nuclei arestained evenly, but apoptotic nuclei are often fragmented andshow irregular or weak DNA staining caused by condensationand fragmentation of the DNA (9, 38). Four days posttrans-fection, coexpression of CrmA did not inhibit Apoptin activity,whereas p53-induced apoptosis was inhibited by coexpressionof CrmA by approximately 50% (Fig. 1). In contrast, in thepresence of p35, Apoptin-induced apoptosis was almost com-pletely abolished (Fig. 2a). Only a background level of apo-ptosis was observed, similar to that seen with overexpression ofthe nonapoptotic control protein Desmin or LacZ (reference 9and data not shown), which is most likely caused by the trans-fection method. In parallel experiments, p35 also almost com-pletely inhibited p53-induced apoptosis, as expected (Fig. 2b).Incubation for up to 4 days posttransfection with 50 mMzVAD-fmk, a synthetic, broad-spectrum caspase inhibitor witha relatively high affinity for upstream caspases (12), partiallyinhibited p53-induced apoptosis but had no effect on Apoptin(data not shown). Taken together, these results indicate thatactivation of downstream, but not upstream, caspases is re-quired for rapid Apoptin-induced apoptosis.
Caspase-3 is a downstream caspase that plays a central rolein the execution of programmed cell death. Therefore, the nextstep was to determine whether caspase-3 is activated in Apop-tin-expressing cells. Saos-2 cells were transfected with Apoptin,fixed 5 days later, and stained with an antibody specific foractive caspase-3. Among the Apoptin-expressing cells, active
VOL. 74, 2000 APOPTIN REQUIRES DOWNSTREAM CASPASES 7073
on October 16, 2014 by guest
http://jvi.asm.org/
Dow
nloaded from

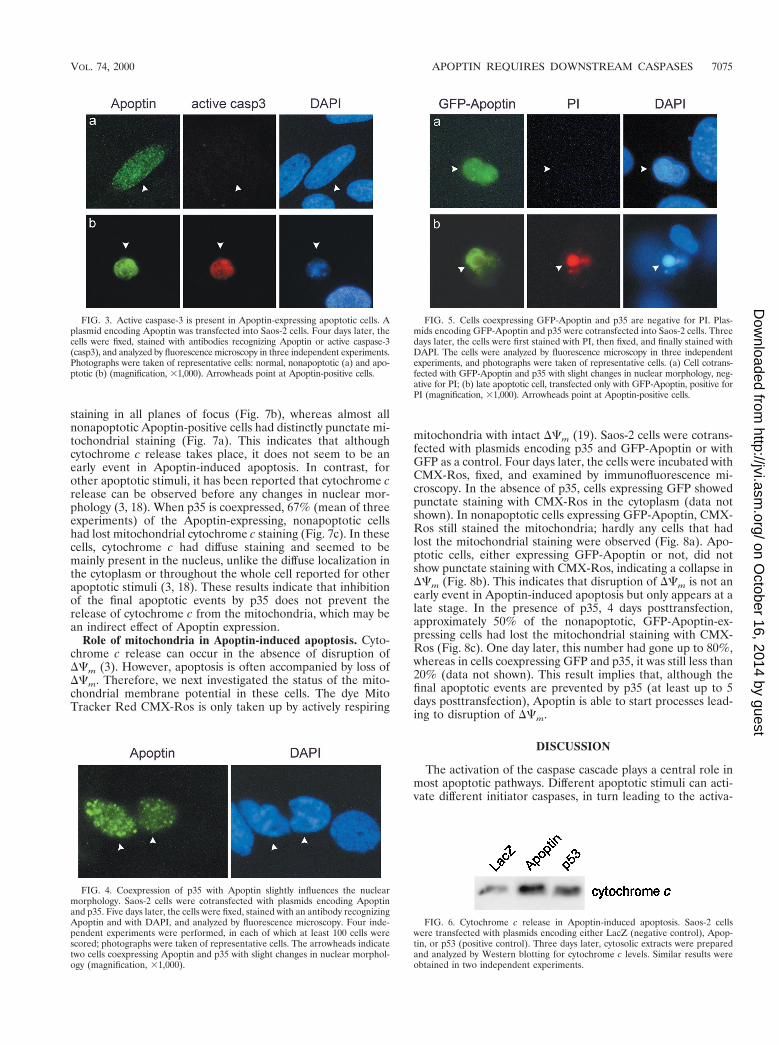
caspase-3 could be detected in all apoptotic cells but was ob-served in only 1% of the nonapoptotic cells containing Apoptin(Fig. 3). As expected, upon coexpression of p35, no activecaspase-3 was seen (data not shown). These results show di-rectly that caspase-3 is activated in Apoptin-induced cell death.Furthermore, the absence of active caspase-3 in nonapoptotic,Apoptin-positive cells indicates that Apoptin expression pre-cedes caspase-3 activation.
Caspase inhibition does not prevent all aspects of Apoptin-induced apoptosis. Whereas coexpression of p35 with eitherApoptin or p53 diminished the number of Saos-2 cells showingthe morphology typical of apoptosis at 5 days posttransfection(shrinkage of the nucleus, condensation and breakdown ofDNA, and nuclear blebbing), some slight changes in nuclearmorphology were still observed in 58% of the cells (Fig. 4). Thenuclei of these cells no longer had smooth surfaces but ap-peared to be dented, and the DNA seemed to be slightlycondensed. This was not observed in cells coexpressing p35 andLacZ (data not shown). Although p35 strongly inhibits Apop-tin- or p53-induced apoptosis for up to 5 days posttransfection,it appears to be slowing down apoptosis rather than blocking itcompletely. Others have reported that caspase inhibition pre-vents certain aspects of apoptosis but does not prevent the cellsfrom dying eventually (16, 25). It is possible that the cellscoexpressing p35 and Apoptin with slightly irregular nucleiwere dying already. To determine the condition of these cells,we next studied the integrity of the cell membrane by lookingat PI exclusion. For this purpose, Saos-2 cells were cotrans-fected with plasmids encoding p35 and GFP-Apoptin, a fusionprotein of GFP and Apoptin which behaves in the same way aswild-type Apoptin in human tumor cells (van Zon, unpub-lished). Three days posttransfection, the cells were analyzedfor PI exclusion. Cells expressing GFP-Apoptin, either with orwithout p35, did not stain with PI, indicating that the cellmembrane was still intact. Cells showing the slight morpholog-ical nuclear changes when coexpressing p35 also did not stainwith PI (Fig. 5a). Only late apoptotic cells were positive for PIstaining (Fig. 5b). These data show that the cells in whichapoptosis is impaired by p35 still have an intact cell membrane,despite the irregular characteristics of their nuclei.
Apoptin induces cytochrome c release, which is not inhibitedby p35. Release of cytochrome c from mitochondria is a well-known event in apoptosis which is often required for activationof downstream caspases. Therefore, we investigated whethercytochrome c is released from mitochondria during Apoptin-induced apoptosis. Saos-2 cells were transfected with plasmidsencoding Apoptin or with LacZ as a negative control. Expres-sion of p53 was used as a positive control for apoptosis induc-tion. Three days later, cytosolic extracts were prepared bysubcellular fractionation and analyzed by Western blotting. Incells transfected with Apoptin or p53, the levels of cytochromec were increased compared to those in cells transfected withLacZ (Fig. 6), which indicates that cytochrome c is releasedfrom mitochondria during Apoptin-induced apoptosis. In or-der to determine the status of the mitochondria on a single-celllevel, we examined the release of cytochrome c from mitochon-dria in cells expressing Apoptin by immunofluorescence mi-croscopy. Saos-2 cells were cotransfected with plasmids encod-ing Apoptin, fixed 5 days later, and stained with specificantibodies for cytochrome c and Apoptin. All Apoptin-ex-pressing apoptotic cells had lost mitochondrial cytochrome c
FIG. 1. CrmA inhibits p53-induced but not Apoptin-induced apoptosis.Saos-2 cells were transfected with plasmids encoding Apoptin or p53, togetherwith a plasmid encoding CrmA or an empty control plasmid (neo). Four daysafter transfection, the cells were fixed, stained with antibodies recognizing Ap-optin or p53, and analyzed by fluorescence microscopy. The percentage of apop-totic cells among those expressing Apoptin or p53 was determined by evaluatingthe DAPI staining. The values shown are the means of three independentexperiments 1 standard deviations; at least 100 cells were scored per experiment.
FIG. 2. p35 inhibits both Apoptin-induced and p53-induced apoptosis.Saos-2 cells were transfected with plasmids encoding Apoptin or p53, togetherwith a plasmid encoding p35 or an empty control plasmid (neo). At several timepoints after transfection, cells were fixed, stained with specific antibodies, andanalyzed by fluorescence microscopy. The percentage of apoptotic cells amongthose expressing Apoptin or p53 was determined by evaluating the DAPI stain-ing. The values shown are the means of three independent experiments 6standard deviations; at least 100 cells were scored per experiment.
7074 DANEN-VAN OORSCHOT ET AL. J. VIROL.
on October 16, 2014 by guest
http://jvi.asm.org/
Dow
nloaded from
staining in all planes of focus (Fig. 7b), whereas almost allnonapoptotic Apoptin-positive cells had distinctly punctate mi-tochondrial staining (Fig. 7a). This indicates that althoughcytochrome c release takes place, it does not seem to be anearly event in Apoptin-induced apoptosis. In contrast, forother apoptotic stimuli, it has been reported that cytochrome crelease can be observed before any changes in nuclear mor-phology (3, 18). When p35 is coexpressed, 67% (mean of threeexperiments) of the Apoptin-expressing, nonapoptotic cellshad lost mitochondrial cytochrome c staining (Fig. 7c). In thesecells, cytochrome c had diffuse staining and seemed to bemainly present in the nucleus, unlike the diffuse localization inthe cytoplasm or throughout the whole cell reported for otherapoptotic stimuli (3, 18). These results indicate that inhibitionof the final apoptotic events by p35 does not prevent therelease of cytochrome c from the mitochondria, which may bean indirect effect of Apoptin expression.
Role of mitochondria in Apoptin-induced apoptosis. Cyto-chrome c release can occur in the absence of disruption ofDCm (3). However, apoptosis is often accompanied by loss ofDCm. Therefore, we next investigated the status of the mito-chondrial membrane potential in these cells. The dye MitoTracker Red CMX-Ros is only taken up by actively respiring
mitochondria with intact DCm (19). Saos-2 cells were cotrans-fected with plasmids encoding p35 and GFP-Apoptin or withGFP as a control. Four days later, the cells were incubated withCMX-Ros, fixed, and examined by immunofluorescence mi-croscopy. In the absence of p35, cells expressing GFP showedpunctate staining with CMX-Ros in the cytoplasm (data notshown). In nonapoptotic cells expressing GFP-Apoptin, CMX-Ros still stained the mitochondria; hardly any cells that hadlost the mitochondrial staining were observed (Fig. 8a). Apo-ptotic cells, either expressing GFP-Apoptin or not, did notshow punctate staining with CMX-Ros, indicating a collapse inDCm (Fig. 8b). This indicates that disruption of DCm is not anearly event in Apoptin-induced apoptosis but only appears at alate stage. In the presence of p35, 4 days posttransfection,approximately 50% of the nonapoptotic, GFP-Apoptin-ex-pressing cells had lost the mitochondrial staining with CMX-Ros (Fig. 8c). One day later, this number had gone up to 80%,whereas in cells coexpressing GFP and p35, it was still less than20% (data not shown). This result implies that, although thefinal apoptotic events are prevented by p35 (at least up to 5days posttransfection), Apoptin is able to start processes lead-ing to disruption of DCm.
DISCUSSION
The activation of the caspase cascade plays a central role inmost apoptotic pathways. Different apoptotic stimuli can acti-vate different initiator caspases, in turn leading to the activa-
FIG. 3. Active caspase-3 is present in Apoptin-expressing apoptotic cells. Aplasmid encoding Apoptin was transfected into Saos-2 cells. Four days later, thecells were fixed, stained with antibodies recognizing Apoptin or active caspase-3(casp3), and analyzed by fluorescence microscopy in three independent experiments.Photographs were taken of representative cells: normal, nonapoptotic (a) and apo-ptotic (b) (magnification, 31,000). Arrowheads point at Apoptin-positive cells.
FIG. 4. Coexpression of p35 with Apoptin slightly influences the nuclearmorphology. Saos-2 cells were cotransfected with plasmids encoding Apoptinand p35. Five days later, the cells were fixed, stained with an antibody recognizingApoptin and with DAPI, and analyzed by fluorescence microscopy. Four inde-pendent experiments were performed, in each of which at least 100 cells werescored; photographs were taken of representative cells. The arrowheads indicatetwo cells coexpressing Apoptin and p35 with slight changes in nuclear morphol-ogy (magnification, 31,000).
FIG. 5. Cells coexpressing GFP-Apoptin and p35 are negative for PI. Plas-mids encoding GFP-Apoptin and p35 were cotransfected into Saos-2 cells. Threedays later, the cells were first stained with PI, then fixed, and finally stained withDAPI. The cells were analyzed by fluorescence microscopy in three independentexperiments, and photographs were taken of representative cells. (a) Cell cotrans-fected with GFP-Apoptin and p35 with slight changes in nuclear morphology, neg-ative for PI; (b) late apoptotic cell, transfected only with GFP-Apoptin, positive forPI (magnification, 31,000). Arrowheads point at Apoptin-positive cells.
FIG. 6. Cytochrome c release in Apoptin-induced apoptosis. Saos-2 cellswere transfected with plasmids encoding either LacZ (negative control), Apop-tin, or p53 (positive control). Three days later, cytosolic extracts were preparedand analyzed by Western blotting for cytochrome c levels. Similar results wereobtained in two independent experiments.
VOL. 74, 2000 APOPTIN REQUIRES DOWNSTREAM CASPASES 7075
on October 16, 2014 by guest
http://jvi.asm.org/
Dow
nloaded from

tion of downstream effector caspases. Here we studied the roleof caspases in apoptosis induced by expression of Apoptin inhuman tumor cells. Apoptin-induced cell death is not affectedby CrmA but is clearly inhibited by p35. The data presentedhere indicate that Apoptin does not require the activation ofthe upstream caspases 1 and 8 but that activation of one ormore downstream caspases is necessary for the rapid inductionof apoptotic cell death. Previously, it had been shown thatp53-induced apoptosis can be inhibited by coexpression of p35in insect cells (29); here we show that this is also the case inhuman cells.
In immunofluorescence studies, activation of caspase-3could be demonstrated in Apoptin-expressing apoptotic cells.In contrast, in cells expressing Apoptin that still had a normalnuclear morphology, active caspase-3 was hardly ever ob-served. These results prove that caspase-3 is activated duringApoptin-induced cell death, but this activation seems to occurat a late stage. In a caspase-3-negative human breast cancercell line, MCF7 (17, 20), Apoptin does not induce cell death inthe same rapid way as in other tumor cell lines and the mor-phological changes typical of apoptosis are not observed (M.Noteborn, unpublished results). This again suggests thatcaspase-3 activation is necessary for rapid Apoptin-inducedapoptosis. However, Apoptin-expressing MCF7 cells did notlook entirely normal either: they displayed condensed DNAand appeared to be dying. Activation of other downstreamcaspases present in MCF7 cells, e.g., 6 and 7, could eventuallylead to cell death. Similarly, the possibility that in Saos-2 cellsother downstream caspases are activated by Apoptin in addi-tion to caspase-3 cannot be excluded.
In cells in which Apoptin-induced apoptosis was inhibited bycoexpression of p35, a slight change in nuclear morphology was
observed. These cells still had intact cell membranes, as deter-mined by PI exclusion. However, in most of these cells, themitochondrial membrane integrity was disrupted: the mito-chondria were no longer stained with CMX-Ros or antibodyagainst cytochrome c. In almost all Apoptin-expressing cellsthat still had normal nuclear morphology (in the absence ofp35), no loss of mitochondrial staining by either CMX-Ros oranti-cytochrome c could be detected, indicating that in thesecells, the DCm is not disrupted. Coexpression of p35 with LacZdid not have any effect on the nuclear morphology, and coex-pression of p35 with GFP had only a minor effect on CMX-Rosstaining (data not shown). Therefore, the effect on nuclearmorphology and mitochondrial membrane integrity cannot beascribed to overexpression of p35 in these cells but is primarilycaused by overexpression of Apoptin. It had been reportedearlier that inhibition of caspases does not prevent cytochromec release caused by, e.g., UVB irradiation or staurosporine (3).Furthermore, in a number of cases, inhibition of caspases pre-vents certain morphological aspects of apoptosis, but eventu-ally the cells still die (16, 25). Similarly, in cells coexpressingApoptin and p35, the slight nuclear changes and loss of mito-chondrial membrane integrity suggest that, in the end, thesecells may not survive.
From the data obtained here, it cannot be concluded thatmitochondria are crucial for Apoptin-induced apoptosis. Ifthey are, it is a late event, which in the absence of p35 ispractically only observed in apoptotic Apoptin-expressing cellsand could be an effect rather than a cause of apoptosis induc-tion. Furthermore, we have shown previously that Bcl-2, whichis thought to inhibit apoptosis by preventing disruption of DCmand release of cytochrome c, does not inhibit Apoptin-induced
FIG. 7. Cytochrome c release in Apoptin-induced apoptosis. Saos-2 cellswere cotransfected with plasmids encoding Apoptin and p35 or with an emptycontrol. Five days later, the cells were fixed and stained with an antibody recog-nizing Apoptin or cytochrome c and with DAPI. The cells were analyzed byfluorescence microscopy in four independent experiments; at least 100 cells wereexamined per experiment. Photographs were taken of representative cells. Cellstransfected only with Apoptin which are nonapoptotic (a) or apoptotic (b) and acell cotransfected with Apoptin and p35 with changed nuclear morphology (c)are shown. Arrowheads point at Apoptin-positive cells. Magnification, 31,000.
FIG. 8. Staining of mitochondria with CMX-Ros. Saos-2 cells were cotrans-fected with plasmids encoding GFP-Apoptin and p35 or with an empty control.Four or 5 days later, the cells were first stained with CMX-Ros and then fixedand stained with DAPI. The cells were analyzed by fluorescence microscopy inthree independent experiments, in each of which at least 100 cells were exam-ined, and photographs were taken of representative cells. A nonapoptotic (a) andan apoptotic (b) cell transfected only with GFP-Apoptin and a cell coexpressingGFP-Apoptin and p35 with changed nuclear morphology (c) are shown. In thelast cell, GFP-Apoptin is also present in the cytoplasm, which occurred moreoften in the presence of p35 than in its absence. Arrowheads point at Apoptin-positive cells. Magnification, 31,000.
7076 DANEN-VAN OORSCHOT ET AL. J. VIROL.
on October 16, 2014 by guest
http://jvi.asm.org/
Dow
nloaded from

cell death in Saos-2 cells. Rather, overexpression of Bcl-2 evenaccelerates Apoptin activity (8, 10). A possible explanationcould be that Apoptin expression leads to cleavage of Bcl-2 byactivated caspases, resulting in a proapoptotic form of Bcl-2(5). However, a mutant form of Bcl-2 which can no longer becleaved by caspases can accelerate Apoptin activity to the sameextent as wild-type Bcl-2 in Saos-2 cells (data not shown).
In this study, we have shown that cell death induction byApoptin involves the activation of caspase-3 and possibly otherdownstream caspases, which also play an essential role in apo-ptosis induced by many other stimuli. Inactive procaspases arepresent in most cells, both normal, nontransformed cells andtransformed or tumorigenic cells (7, 14, 28, 40). However,Apoptin only induces apoptosis in transformed or tumorigeniccells. We propose two possible explanations for these phenom-ena. First, Apoptin may be differentially modified in tumorcells so that it can exert its apoptotic activity. Second, thecellular decision machinery that decides whether to enter ap-optosis may be different in nontransformed and transformed ortumorigenic cells. It is conceivable that expression or absenceof expression of one or more genes in transformed or tumor-igenic cells causes them to respond to certain stimuli by un-dergoing apoptosis. Previous data suggest that the difference inApoptin activity is correlated with a difference in localization:in normal cells, it is found mainly in the cytoplasm, whereas intransformed or tumorigenic cells it is located predominantly inthe nucleus (9). We hypothesize that once Apoptin is in thenucleus (either modified or not), it generates an apoptoticsignal that eventually activates downstream caspases, leadingto apoptosis.
In summary, we have shown that activation of caspases,especially caspase-3, is necessary for apoptosis induction byApoptin. The role of mitochondria is still unclear, but they donot seem to be crucial for Apoptin activity. Studies of themodification of Apoptin, its binding factors, and its localizationin normal versus tumor cells are in progress. Better under-standing of the mechanism of Apoptin-induced apoptosis willbe helpful in its use as an antitumor agent and may alsoprovide information on the aspects of a cell that determine itstransformed or tumorigenic state.
ACKNOWLEDGMENTS
We thank D. J. Pickup, L. K. Miller, and J. M. Hardwick for dona-tion of plasmids and/or antibody, S. J. Olijslagers and A. van Zon fortechnical assistance, B. van de Water for helpful discussions, and J. L.Rohn for critical review of the manuscript and stimulating discussions.
This work was supported by grants from the Dutch Cancer Society,the Netherlands Ministry of Economic Affairs, and Schering AG.
REFERENCES1. Adams, J. M., and S. Cory. 1998. The Bcl-2 protein family: arbiters of cell
survival. Science 281:1322–1326.2. Bossy-Wetzel, E., and D. R. Green. 1999. Caspases induce cytochrome c
release from mitochondria by activating cytosolic factors. J. Biol. Chem.274:17484–17490.
3. Bossy-Wetzel, E., D. D. Newmeyer, and D. R. Green. 1998. Mitochondrialcytochrome c release in apoptosis occurs upstream of DEVD-specificcaspase activation and independently of mitochondrial transmembrane de-polarization. EMBO J. 17:37–49.
4. Bump, N. J., M. Hackett, M. Hugunin, S. Seshagiri, K. Brady, P. Chen, C.Ferenz, S. Franklin, T. Ghayur, and P. Li. 1995. Inhibition of ICE familyproteases by baculovirus antiapoptotic protein p35. Science 269:1885–1888.
5. Cheng, E. H., D. G. Kirsch, R. J. Clem, R. Ravi, M. B. Kastan, A. Bedi, K.Ueno, and J. M. Hardwick. 1997. Conversion of Bcl-2 to a Bax-like deatheffector by caspases. Science 278:1966–1968.
6. Clem, R. J., E. H. Cheng, C. L. Karp, D. G. Kirsch, K. Ueno, A. Takahashi,M. B. Kastan, D. E. Griffin, W. C. Earnshaw, M. A. Veliuona, and J. M.Hardwick. 1998. Modulation of cell death by Bcl-XL through caspase inter-action. Proc. Natl. Acad. Sci. USA 95:554–559.
7. Cryns, V., and J. Yuan. 1998. Proteases to die for. Genes Dev. 12:1551–1570.(Erratum, 13:371, 1999.)
8. Danen-Van Oorschot, A. A. A. M., A. I. Den Hollander, S. Takayama, J. C.Reed, A. J. Van der Eb, and M. H. M. Noteborn. 1997. BAG-1 inhibitsp53-induced but not Apoptin-induced apoptosis. Apoptosis 2:395–402.
9. Danen-Van Oorschot, A. A. A. M., D. F. Fischer, J. M. Grimbergen, B. Klein,S. Zhuang, J. H. Falkenburg, C. Backendorf, P. H. Quax, A. J. Van der Eb,and M. H. M. Noteborn. 1997. Apoptin induces apoptosis in human trans-formed and malignant cells but not in normal cells. Proc. Natl. Acad. Sci.USA 94:5843–5847.
10. Danen-Van Oorschot, A. A. A. M., Y. Zhang, S. J. Erkeland, D. F. Fischer,A. J. Van der Eb, and M. H. M. Noteborn. 1999. The effect of Bcl-2 onApoptin in ‘normal’ vs transformed human cells. Leukemia 13(Suppl. 1):S75–S77.
11. Deveraux, Q. L., R. Takahashi, G. S. Salvesen, and J. C. Reed. 1997. X-linked IAP is a direct inhibitor of cell-death proteases. Nature 388:300–304.
12. Garcia-Calvo, M., E. P. Peterson, B. Leiting, R. Ruel, D. W. Nicholson, andN. A. Thornberry. 1998. Inhibition of human caspases by peptide-based andmacromolecular inhibitors. J. Biol. Chem. 273:32608–32613.
13. Green, D., and G. Kroemer. 1998. The central executioners of apoptosis:caspases or mitochondria? Trends Cell Biol. 8:267–271.
14. Green, D. R. 1998. Apoptotic pathways: the roads to ruin. Cell 94:695–698.15. Green, D. R., and J. C. Reed. 1998. Mitochondria and apoptosis. Science
281:1309–1312.16. Hirsch, T., P. Marchetti, S. A. Susin, B. Dallaporta, N. Zamzami, I. Marzo,
M. Geuskens, and G. Kroemer. 1997. The apoptosis-necrosis paradox.Apoptogenic proteases activated after mitochondrial permeability transitiondetermine the mode of cell death. Oncogene 15:1573–1581.
17. Janicke, R. U., M. L. Sprengart, M. R. Wati, and A. G. Porter. 1998.Caspase-3 is required for DNA fragmentation and morphological changesassociated with apoptosis. J. Biol. Chem. 273:9357–9360.
18. Juin, P., A. O. Hueber, T. Littlewood, and G. Evan. 1999. c-Myc-inducedsensitization to apoptosis is mediated through cytochrome c release. GenesDev. 13:1367–1381.
19. Kroemer, G., N. Zamzami, and S. A. Susin. 1997. Mitochondrial control ofapoptosis. Immunol. Today 18:44–51.
20. Li, F., A. Srinivasan, Y. Wang, R. C. Armstrong, K. J. Tomaselli, and L. C.Fritz. 1997. Cell-specific induction of apoptosis by microinjection of cyto-chrome c. Bcl-xL has activity independent of cytochrome c release. J. Biol.Chem. 272:30299–30305.
21. Li, H., H. Zhu, C. J. Xu, and J. Yuan. 1998. Cleavage of BID by caspase 8mediates the mitochondrial damage in the Fas pathway of apoptosis. Cell94:491–501.
22. Liu, X., C. N. Kim, J. Yang, R. Jemmerson, and X. Wang. 1996. Induction ofapoptotic program in cell-free extracts: requirement for dATP and cyto-chrome c. Cell 86:147–157.
23. Luo, X., I. Budihardjo, H. Zou, C. Slaughter, and X. Wang. 1998. Bid, a Bcl2interacting protein, mediates cytochrome c release from mitochondria inresponse to activation of cell surface death receptors. Cell 94:481–490.
24. Mancini, M., D. W. Nicholson, S. Roy, N. A. Thornberry, E. P. Peterson,L. A. Casciola-Rosen, and A. Rosen. 1998. The caspase-3 precursor has acytosolic and mitochondrial distribution: implications for apoptotic signaling.J. Cell Biol. 140:1485–1495.
25. McCarthy, N. J., M. K. Whyte, C. S. Gilbert, and G. I. Evan. 1997. Inhibitionof Ced-3/ICE-related proteases does not prevent cell death induced byoncogenes, DNA damage, or the Bcl-2 homologue Bak. J. Cell. Biol. 136:215–227.
26. Nicholson, D. W., and N. A. Thornberry. 1997. Caspases: killer proteases.Trends Biochem. Sci. 22:299–306.
27. Noteborn, M. H. M., D. Todd, C. A. Verschueren, H. W. de Gauw, W. L.Curran, S. Veldkamp, A. J. Douglas, M. S. McNulty, A. J. Van der Eb, andG. Koch. 1994. A single chicken anemia virus protein induces apoptosis.J. Virol. 68:346–351.
28. Nunez, G., M. A. Benedict, Y. Hu, and N. Inohara. 1998. Caspases: theproteases of the apoptotic pathway. Oncogene 17:3237–3245.
29. Prikhod’ko, G. G., Y. Wang, E. Freulich, C. Prives, and L. K. Miller. 1999.Baculovirus p33 binds human p53 and enhances p53-mediated apoptosis.J. Virol. 73:1227–1234.
30. Ray, C. A., R. A. Black, S. R. Kronheim, T. A. Greenstreet, P. R. Sleath, G. S.Salvesen, and D. J. Pickup. 1992. Viral inhibition of inflammation: cowpoxvirus encodes an inhibitor of the interleukin-1 beta converting enzyme. Cell69:597–604.
31. Reed, J. C. 1998. Bcl-2 family proteins. Oncogene 17:3225–3236.32. Roy, N., Q. L. Deveraux, R. Takahashi, G. S. Salvesen, and J. C. Reed. 1997.
The c-IAP-1 and c-IAP-2 proteins are direct inhibitors of specific caspases.EMBO J. 16:6914–6925.
33. Scaffidi, C., S. Fulda, A. Srinivasan, C. Friesen, F. Li, K. J. Tomaselli, K. M.Debatin, P. H. Krammer, and M. E. Peter. 1998. Two CD95 (APO-1/Fas)signaling pathways. EMBO J. 17:1675–1687.
34. Seshagiri, S., and L. K. Miller. 1997. Baculovirus inhibitors of apoptosis(IAPs) block activation of Sf-caspase-1. Proc. Natl. Acad. Sci. USA 94:13606–13611.
35. Shen, Y., and T. E. Shenk. 1995. Viruses and apoptosis. Curr. Opin. Genet.Dev. 5:105–111.
VOL. 74, 2000 APOPTIN REQUIRES DOWNSTREAM CASPASES 7077
on October 16, 2014 by guest
http://jvi.asm.org/
Dow
nloaded from

36. Susin, S. A., H. K. Lorenzo, N. Zamzami, I. Marzo, C. Brenner, N. Laro-chette, M. C. Prevost, P. M. Alzari, and G. Kroemer. 1999. Mitochondrialrelease of caspase-2 and -9 during the apoptotic process. J. Exp. Med.189:381–394.
37. Susin, S. A., H. K. Lorenzo, N. Zamzami, I. Marzo, B. E. Snow, G. M.Brothers, J. Mangion, E. Jacotot, P. Costantini, M. Loeffler, N. Larochette,D. R. Goodlett, R. Aebersold, D. P. Siderovski, J. M. Penninger, and G.Kroemer. 1999. Molecular characterization of mitochondrial apoptosis-in-ducing factor. Nature 397:441–446.
38. Telford, W. G., L. E. King, and P. J. Fraker. 1992. Comparative evaluationof several DNA binding dyes in the detection of apoptosis-associated chro-matin degradation by flow cytometry. Cytometry 13:137–143.
39. Teodoro, J. G., and P. E. Branton. 1997. Regulation of apoptosis by viralgene products. J. Virol. 71:1739–1746.
40. Thornberry, N. A., and Y. Lazebnik. 1998. Caspases: enemies within. Science281:1312–1316.
41. Tschopp, J., M. Thome, K. Hofmann, and E. Meinl. 1998. The fight of virusesagainst apoptosis. Curr. Opin. Genet. Dev. 8:82–87.
42. Tsujimoto, Y. 1998. Role of Bcl-2 family proteins in apoptosis: apoptosomesor mitochondria? Genes Cells 3:697–707.
43. Van der Eb, A. J., and F. L. Graham. 1980. Assay of transforming activity oftumor virus DNA. Methods Enzymol. 65:826–839.
44. Yang, X., H. Y. Chang, and D. Baltimore. 1998. Autoproteolytic activation ofpro-caspases by oligomerization. Mol. Cell 1:319–325.
45. Zhou, Q., J. F. Krebs, S. J. Snipas, A. Price, E. S. Alnemri, K. J. Tomaselli,and G. S. Salvesen. 1998. Interaction of the baculovirus anti-apoptotic pro-tein p35 with caspases. Specificity, kinetics, and characterization of thecaspase/p35 complex. Biochemistry 37:10757–10765.
46. Zhou, Q., S. Snipas, K. Orth, M. Muzio, V. M. Dixit, and G. S. Salvesen.1997. Target protease specificity of the viral serpin CrmA. Analysis of fivecaspases. J. Biol. Chem. 272:7797–7800.
47. Zhuang, S. M., A. Shvarts, A. G. Jochemsen, A. A. A. M. van Oorschot, A. J.Van der Eb, and M. H. M. Noteborn. 1995. Differential sensitivity to Ad5E1B-21kD and Bcl-2 proteins of apoptin-induced versus p53-induced apo-ptosis. Carcinogenesis 16:2939–2944.
48. Zhuang, S. M., A. Shvarts, H. van Ormondt, A. G. Jochemsen, A. J. Van derEb, and M. H. M. Noteborn. 1995. Apoptin, a protein derived from chickenanemia virus, induces p53-independent apoptosis in human osteosarcomacells. Cancer Res. 55:486–489.
7078 DANEN-VAN OORSCHOT ET AL. J. VIROL.
on October 16, 2014 by guest
http://jvi.asm.org/
Dow
nloaded from




















