
SLUG: a new target of lymphoid enhancer factor-1 in human osteoblasts

Journal of PathologyJ Pathol 2008; 214: 25–37Published online 31 October 2007 in Wiley InterScience(www.interscience.wiley.com) DOI: 10.1002/path.2254
Original Paper
The basal-like breast carcinoma phenotype is regulatedby SLUG gene expression
G Storci,1,2 P Sansone,1,3 D Trere,2 S Tavolari,4 M Taffurelli,5 C Ceccarelli,1,6 T Guarnieri,4 P Paterini,1,4
M Pariali,1 L Montanaro,2 D Santini,6 P Chieco1 and M Bonafe1,2*1Center for Applied Biomedical Research (CRBA), St Orsola-Malpighi Hospital, University of Bologna, Bologna, Italy2Department of Experimental Pathology, University of Bologna, Bologna, Italy3Department of Pharmacology and Toxicology, University of Bologna, Bologna, Italy4Department of Experimental Evolutionary Biology, University of Bologna, Bologna, Italy5Department of Surgical and Anaesthesiological Sciences, S Orsola-Malpighi Hospital, University of Bologna, Bologna, Italy6Department of Gastroenterology and Pathology, S Orsola-Malpighi Hospital, University of Bologna, Bologna, Italy
*Correspondence to:M Bonafe, Department ofExperimental Pathology,University of Bologna,Bologna, Italy.E-mail:[email protected]
No conflicts of interest weredeclared.
Received: 11 July 2007Revised: 28 August 2007Accepted: 6 September 2007
AbstractBasal-like breast carcinoma is an aggressive form of breast cancer, characterized by theabsence of oestrogen receptor and HER2 expression, the presence of cytokeratin 5 andepidermal growth factor receptor expression, and by the up-regulation of stem cell regulatorygenes. We show here that tumour tissues expressing high levels of SLUG mRNA show a basal-like breast carcinoma phenotype and that such tumours also express high levels of stemcell-regulatory genes, ie CD133, Bmi1. Further, we show that stem/progenitor cells, isolatedfrom ductal breast carcinoma and from normal mammary gland as mammospheres, expressSLUG, CD133, and Bmi1 mRNA and show a phenotype similar to that of basal-like breastcarcinoma. We also report that SLUG expression in tumour tissues correlates with that ofthe hypoxia survival gene carbonic anhydrase IX. In this regard, we report that the exposureof SLUG-negative/luminal-like MCF-7 cells to a hypoxic environment promotes the onset ofthe basal-like breast carcinoma phenotype, together with up-regulation of the SLUG gene,which in turn blunts oestrogen receptor-α and boosts carbonic anhydrase IX gene expression.Finally, we show that SLUG expression promotes the invasiveness of MCF-7 cells exposedto hypoxia and sustains the in vivo aggressiveness of hypoxia-selected, MCF-7-derived cellsin xenografts. These data indicate that SLUG gene expression is part of a hypoxia-inducedgenetic programme which sets up a basal/stem cell-like, aggressive phenotype in breastcancer cells.Copyright 2007 Pathological Society of Great Britain and Ireland. Published by JohnWiley & Sons, Ltd.
Keywords: SLUG; hypoxia; basal-like breast carcinoma; stem cells
Introduction
Breast cancer is the leading cause of female mortalityin the western world [1]. The lack of expression ofthe oestrogen receptor (ER) and HER2 protein, cou-pled with the expression of cytokeratin-5 (CK5) andepidermal growth factor receptor (EGFR), identifiesthe so-called ‘basal-like breast carcinoma phenotype’[2–7]. Some authors propose that the CK5-positivebasal cell compartment harbours the stem cells of themammary gland [8]. In keeping with this hypothe-sis, recent investigations have revealed that basal-likebreast carcinomas show a gene expression pattern sim-ilar to that of putative breast cancer stem cells [9,10].Hence, there are data in the literature supporting thenotion that a relationship exists between the basal andthe stem cell phenotype, in normal and tumour mam-mary tissue.
Aberrant and non-terminal differentiation is a hall-mark of malignant behaviour [11]. Recently, it hasbeen suggested that the poorly differentiated pheno-type in cancer cells might arise from breast stem cellswhose lineage commitment process has been arrested[12,13]. Alternatively, the lack of differentiation maybe the consequence of an epithelial/mesenchymal tran-sition (EMT), in which the down-regulation of differ-entiation genes is coupled with the up-regulation ofgenes expressed in immature cells [11,14,15]. Sucha process is crucial for stem/progenitor cells dur-ing embryonic development and promotes the inva-siveness of cancer cells [11,16]. EMT is mediatedby members of the Snail family of genes, such asSLUG [16,17], which is effective in inducing thedown-regulation of differentiation markers (such asE-cadherin) and the acquisition of a migrant pheno-type in a variety of cell types [14,16,17]. In recent
Copyright 2007 Pathological Society of Great Britain and Ireland. Published by John Wiley & Sons, Ltd.www.pathsoc.org.uk

26 G Storci et al
years, SLUG gene expression has been linked tothe biological aggressiveness of melanoma, and lung,colon, and ovarian cancer [18–21]. In breast cancer,SLUG gene expression correlates with the number ofinvaded lymph nodes or with metastatic disease, butfunctional data on such an association are still lacking[22,23]. Here we assessed SLUG mRNA expression inbreast cancer patients and its association with clinico-pathological features. Moreover, we explored the func-tion of the SLUG gene in vitro in breast cancer cells,as well as in stem/progenitor cells of the mammarygland expanded in vitro as mammospheres [24,25]. Wefound that SLUG gene expression identifies a subsetof breast tumours endowed with a basal/stem cell-likephenotype and, in our in vitro model, that SLUG geneexpression is functionally relevant for such a pheno-type.
Materials and methods
Patients
Sixty-seven consecutive patients (mean age 62 years;range 28–87 years) who had undergone surgical resec-tion for primary infiltrating breast carcinomas wereenrolled in the study. Tumours were histologicallyclassified according to the World Health Organisa-tion (WHO) criteria [26] as follows: 55 ductal, eightlobular, two mucoid, and two medullary carcino-mas. Tumours were histologically graded accordingto Elston and Ellis’s criteria [27]. Axillary node statuswas assessed after axillary node dissection. Follow-up data (disease free survival = 94 months, overallsurvival = 100 months) were available for 65 patients.Tissues were obtained after patients’ written informedconsent and the use of tissues was approved by thelocal ethics committee.
Cell culture
MCF-7 cells and hypoxia selected cells (Hypo-7) [25]were grown in RPMI-1640 medium supplementedwith 10% fetal bovine serum, 100 µg/ml streptomycin,100 IU/ml penicillin, and 2 mM L-glutamine (Euro-clone, Milan, Italy), kept at 37 ◦C in a humidified (5%CO2) atmosphere.
Generation of mammospheres (MSs)
Four fresh tumour specimens and two normal mam-mary gland tissues were obtained from patientsaffected by ductal breast carcinoma who underwentquadrantectomy or mastectomy. MSs were obtainedas previously described [24,25,28,29], except that theprotocol was down-scaled to handle sample tissuesof less than 1 g in weight. A representative part ofeach specimen was histologically examined to assessthe presence/absence of normal and tumour tissue inthe same tissue sample. Only homogeneous tissueswere used to generate MSs. All the procedures on
patients’ tissues had been approved by the local ethicscommittee and the patients gave written informed con-sent.
Immunofluorescence (IF)
Cultured cells were seeded onto glass coverslips at60% confluence, fixed with 2% paraformaldehyde,and permeabilized with 0.2% Triton X-100. Cellswere incubated with anti-cytokeratin-5 (CK5) antibody(Dako Cytomation, Glostrup, Denmark) and secondaryanti-rabbit antibody FITC conjugated (Santa CruzBiotechnology, Santa Cruz, CA, USA).
Boyden chamber invasion assay
Cell invasion was assessed in Matrigel-coated Boydenchambers (New Technologies Group, Italy). 1 × 105
MCF-7 or Hypo-7 cells, and 3 × 102 cells from dis-aggregated MSs were used in each assay. Invadingcells were stained with toluidine blue and scored asthe mean number of invaded cells per random opticalfield, in three independent experiments.
In vitro/in vivo RNA interference
SLUG siRNA and appropriate control, scramble(SCR) siRNAs, were purchased from Invitrogen(Carlsbad, CA, USA). SLUG/SCR siRNAs weretransfected into adherent MCF-7 and Hypo-7 cells(105 cells in a 3 cm2 well) at a concentration of1 µg/well, using Lipofectamine 2000 (Invitrogen).SLUG/SCR siRNA transfection in MSs was performedby mixing 0.5 µg of siRNA with in vitro JET-PEIreagent (Poly plus Trasfection, Illkirch, France) in a5 to 1 ratio.
Eight nude mice (6-week-old Balb/C females,Charles River Laboratories, Milan, Italy) were injectedwith 106 Hypo-7 cells in the abdominal cavity. After1 week, 10 µg of SLUG or SCR siRNA, mixed within vivo JET-PEI reagent (3 : 1 ratio), was intraperi-toneally administered to the animals. The proce-dure was repeated twice a week for 4 weeks. Theanimals were sacrificed and subjected to necropsy.Tissues were formalin-fixed, paraffin-embedded, andprocessed for immunohistochemistry. The procedureswere approved by the local veterinary ethics commit-tee.
RNA extraction and cDNA amplification
Total RNA was extracted using TRIzol reagent(Invitrogen). Reverse transcription was performed with2 µg of total RNA. The primers used to amplifythe target genes are listed in Supplementary Table 1,available online at http://www.interscience.wiley.com/jpages/0022-3417/suppmat/path.2254.html. The PCRprotocol was as follows: pre-denaturation step: 94 ◦C,2 min; 30 cycles of denaturation (94 ◦C, 1 min),
J Pathol 2008; 214: 25–37 DOI: 10.1002/pathCopyright 2007 Pathological Society of Great Britain and Ireland. Published by John Wiley & Sons, Ltd.

SLUG and basal-like breast carcinoma 27
Table 1. Association between the SLUG mRNA expressionlevel (absent, A; low, L; high, H) and clinical, histopathological,and bio-pathological characteristics of 67 breast cancer patients
SLUG mRNA expression levelMonte Carlo χ2
A L H (p value)
Age, years<50 8 7 3 0.928≥ 50 21 22 6
HistotypeDuctal 25 21 9Lobular 4 4 0 0.292Medullary 0 2 0Mucoid 0 2 0
Tumour size (pT)1 8 11 32 12 17 3 0.1463 4 0 24 5 1 1
Histological grade (G)12 2 2 1 0.2393 9 12 0
18 15 8
Lymph node involvement (pN∗)0 8 13 5 0.199≥1 21 14 4
ERNegative 8 8 7 0.014Positive 21 21 2
HER2 status†
Negative 12 13 8 0.036Positive 12 13 0
∗ In two patients, N-status was not available as axillary dissection wasnot performed owing to patient’s age.† Available in 58 patients only.
annealing (1 min), extension (72 ◦C, 1 min), finalextension (72 ◦C, 7 min).
Immunohistochemistry (IHC)
Breast tumour sections were stained using mono-clonal anti-EGFR (clone 31G7; Zymed Laborato-ries Inc, USA), ER and CK5 (clone 1D5, cloneD5/16B4; DakoCytomation), HER2 internal domain(clone CB11; BioGenex Laboratories, USA), CA-IX(clone M-75; kindly provided by J Pastorek, SlovakAcademy of Sciences, Bratislava, Slovak Republic),and Bmi1 (clone C-20; Santa Cruz). The immunopos-itive population was classified as negative (<10%) orpositive (≥10%), as previously described [30], andexpressed as the percentage of labelled area over thetotal neoplastic area in the section. CA-IX proteinexpression was evaluated using a semi-quantitativescore [31], where the final binomial positive/negativescore summarizes the frequency of positive neoplas-tic cells and their staining intensity. HER2 immunos-taining was assessed following the DakoCytoma-tion guidelines (0 and +1 = negative; +2 and +3 =positive). Basal-like breast tumours were diagnosed
as CK5/EGFR-positive, ER/HER2-negative tumours[2–7,32,33]. Tumour MS sections were probed withantibodies against ER, HER2, EGFR, CK5, CD133(clone AC-133; Myilteni Biotech, Gladbach, Ger-many), Bmi1 (clone C-20; Santa Cruz), vimentin(clone C-20; Santa Cruz), and SLUG (polyclonalrabbit antibody; Santa Cruz). Sections of collagen-embedded MCF-7 and Hypo-7 cells were assessedwith anti-EGFR, ER, and vimentin antibodies. Sec-tions of Hypo-7 xenografts were assessed with anti-CA-IX and SLUG antibodies.
Western blot (WB) analysis
Western blot analysis was performed as previouslydescribed [25]. Antibodies for WB analysis weredirected against EGFR, SLUG, CA-IX, and β-actin(Sigma). HRP-conjugated secondary antibodies werevisualized using the luminol/enhancer chemilumi-nescent substrate (Amersham Biosciences, Bucking-hamshire, UK).
Statistical analysis
Statistical analysis was performed using the SPSSstatistical software package (Statistical Package forSocial Science, SPSS Inc, Chicago, IL, USA). Cat-egorical data were analysed using the Monte Carlo χ2
test and Fisher’s exact test. Non-normally distributedvariables were analysed using the Kruskal–Wallisnon-parametric test. Survival analysis was analysedby the Kaplan and Meier method; differences in thesurvival curves were assessed by the log-rank test.
Results
SLUG mRNA expression in breast cancer tissues
RT-PCR analysis of SLUG mRNA levels in 67breast carcinomas revealed three distinct categories ofSLUG mRNA expression: absent (A-SLUG, n = 24);low (L-SLUG, n = 34); and high (H-SLUG, n = 9),(Figures 1A and 1B). When stratified according tothe patho-biological profile of breast cancer patients,the SLUG mRNA expression level correlated nega-tively with IHC expression of ER and HER2, whereasno significant association was found between SLUGmRNA expression and patient age, histological type,tumour size, histological grade or lymph node involve-ment (Table 1). These data suggest that a subgroup ofER/HER2-negative breast tumours show high levelsof SLUG mRNA expression.
Tumours with high levels of SLUG mRNA show abasal/stem cell-like phenotype
The lack of ER/HER2 expression in histologicallydefined ductal breast carcinomas has been associ-ated with the basal-like breast carcinoma phenotype[2–7,32]. Interestingly, SLUG gene up-regulation has
J Pathol 2008; 214: 25–37 DOI: 10.1002/pathCopyright 2007 Pathological Society of Great Britain and Ireland. Published by John Wiley & Sons, Ltd.

28 G Storci et al
Figure 1. SLUG mRNA level in breast cancer patients. (A) Bar-graph representation of SLUG mRNA level RT-PCR analysis in 67breast cancer patients: densitometric evaluation of SLUG mRNA expression level over beta-2 microglobulin (β2µ) mRNA level.Three categories of SLUG mRNA expression level were identified: absent (A), low (L), and high (H). The median level of theSLUG/β2µ mRNA ratio is reported. (B) RT-PCR analysis of the SLUG and β2µ mRNA level in representative cases of H- (n = 7),L- (n = 12), and A- (n = 12) SLUG mRNA-expressing breast cancer tissues
been recently identified in cell lines showing a basal-like breast carcinoma phenotype [34]. IHC analysis oftumour samples revealed that two hallmarks of basal-cell phenotype, ie CK5 and EGFR, were overrep-resented among H-SLUG mRNA-expressing tumours(Figure 2A). Similarly, RT-PCR analysis revealed thatCK5 and EGFR mRNAs were detectable in all H-SLUG mRNA-expressing tumours (Figure 2B), whichalso expressed high levels of vimentin mRNA, amarker of basal-like breast carcinoma [35](Figure 2B).
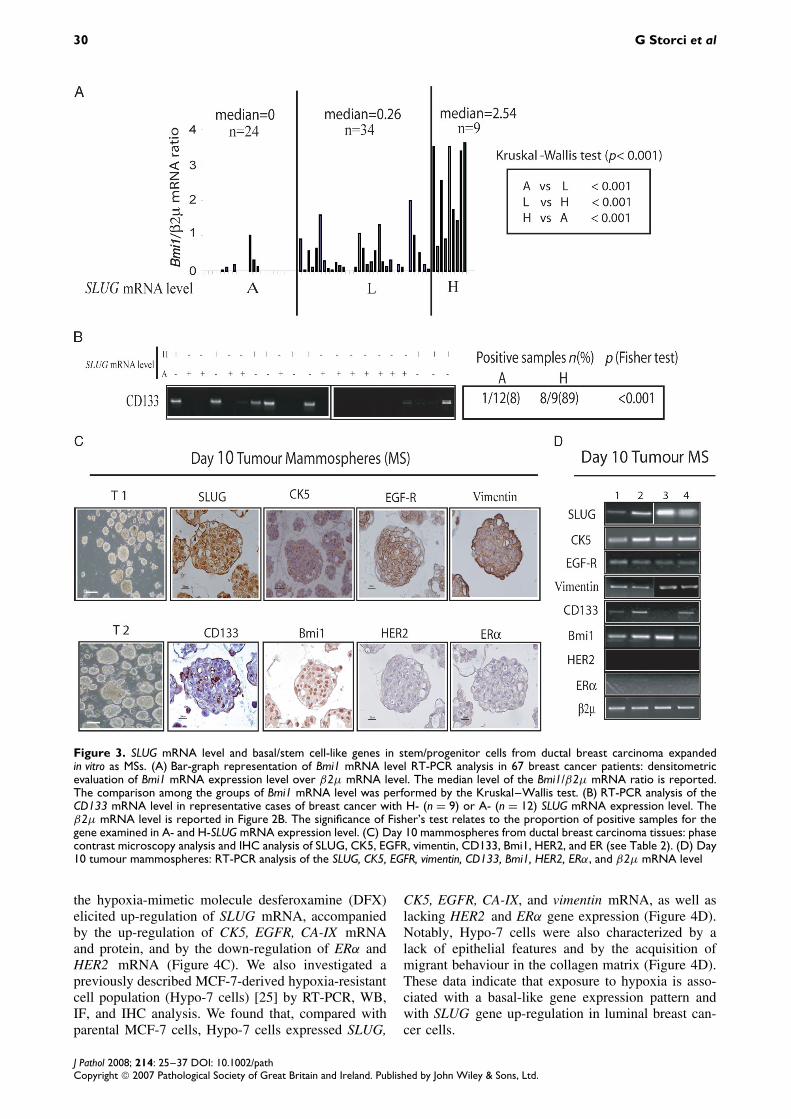
SLUG mRNA expression in tumourstem/progenitor cells from ductal breastcarcinoma
Basal tumours have been named due to their sim-ilarity to the basal cells, wherein stem cells have
been proposed to be harboured [8]. Accordingly,these tumours have been assumed to be endowedwith stem cell-like features [9,10,34]. In keeping withthis hypothesis, we found that H-SLUG-expressingtumour tissues also expressed high levels of the stemcell regulatory gene Bmi1 mRNA [36] (Figure 3A).Moreover, the expression of the stem cell markerCD133 mRNA [37] was detected only in H-SLUGmRNA-expressing tumours (Figure 3B). To inves-tigate the relationship between SLUG mRNA andthe stem cell-like phenotype further, we isolatedstem/progenitor cells from ductal breast carcinomatissues by expanding these cells in vitro as mammo-spheres (MSs) [24,25,28,29] (Table 2). IHC analysisrevealed that MSs expressed SLUG protein and dis-closed a basal/stem cell-like gene expression profile,ie the presence of CK5, EGFR, vimentin, CD133,
J Pathol 2008; 214: 25–37 DOI: 10.1002/pathCopyright 2007 Pathological Society of Great Britain and Ireland. Published by John Wiley & Sons, Ltd.

SLUG and basal-like breast carcinoma 29
Figure 2. Tumours expressing high levels of SLUG mRNA show basal-like breast carcinoma features. (A) Bar-graph representationand representative pictures of IHC (arrows indicate normal tissue) of CK5 and EGFR in breast cancer patients, classified accordingto the SLUG mRNA expression categories (A, L, and H). Note that CK5 staining was performed in 54 patients. (B) RT-PCR analysisof the CK5, EGFR, vimentin, and β2µ mRNA level in representative cases of breast cancer with H- (n = 9) or A- (n = 12) level ofSLUG mRNA expression. The significance of Fisher’s test relates to the proportion of samples expressing the gene examined inA- and H-SLUG mRNA-expressing tumours
and Bmi1, and the absence of HER2 and ER proteinexpression (Figure 3C). RT-PCR analysis confirmedthe reproducibility of the above gene expression pat-tern in all the samples examined (Figure 3D). Agene expression profile similar to tumour MSs waseven observed in MSs obtained from normal mam-mary gland (see Supplementary Figure 1, availableat http://www.interscience.wiley.com/jpages/0022-3417/suppmat/path.2254.html). These data suggestthat SLUG mRNA-expressing stem/progenitor cellscan be expanded in vitro from ductal breast tumourtissues.
Hypoxia up-regulates SLUG gene expression andthe basal-like breast carcinoma phenotype inluminal-like MCF-7 cellsRecent data indicate that the expression of CA-IX,a hypoxia survival gene, is increased in basal-like
Table 2. Clinico-pathological characteristics of the ductalbreast carcinoma tissues employed to generate mammospheres(MSs)
Sample Age (years) pT pN G ER HER2 EGFR
T1 83 1 3 2 1 3 0T2 44 1 0 1 1 0 0T3 39 2 3 2 1 1 0T4 73 1 2 2 1 1 0
breast carcinoma tissues [38–40]. In agreement withthis finding, RT-PCR and IHC analysis revealed ahigh proportion of tumour tissues expressing CA-IX mRNA and protein among H-SLUG mRNA-expressing tumours (Figures 4A and 4B). These dataled us to search for a functional correlation betweenbasal-like phenotype and hypoxia. We found that theexposure of luminal-like MCF-7 breast cancer cells to
J Pathol 2008; 214: 25–37 DOI: 10.1002/pathCopyright 2007 Pathological Society of Great Britain and Ireland. Published by John Wiley & Sons, Ltd.
30 G Storci et al
Figure 3. SLUG mRNA level and basal/stem cell-like genes in stem/progenitor cells from ductal breast carcinoma expandedin vitro as MSs. (A) Bar-graph representation of Bmi1 mRNA level RT-PCR analysis in 67 breast cancer patients: densitometricevaluation of Bmi1 mRNA expression level over β2µ mRNA level. The median level of the Bmi1/β2µ mRNA ratio is reported.The comparison among the groups of Bmi1 mRNA level was performed by the Kruskal–Wallis test. (B) RT-PCR analysis of theCD133 mRNA level in representative cases of breast cancer with H- (n = 9) or A- (n = 12) SLUG mRNA expression level. Theβ2µ mRNA level is reported in Figure 2B. The significance of Fisher’s test relates to the proportion of positive samples for thegene examined in A- and H-SLUG mRNA expression level. (C) Day 10 mammospheres from ductal breast carcinoma tissues: phasecontrast microscopy analysis and IHC analysis of SLUG, CK5, EGFR, vimentin, CD133, Bmi1, HER2, and ER (see Table 2). (D) Day10 tumour mammospheres: RT-PCR analysis of the SLUG, CK5, EGFR, vimentin, CD133, Bmi1, HER2, ERα, and β2µ mRNA level
the hypoxia-mimetic molecule desferoxamine (DFX)elicited up-regulation of SLUG mRNA, accompaniedby the up-regulation of CK5, EGFR, CA-IX mRNAand protein, and by the down-regulation of ERα andHER2 mRNA (Figure 4C). We also investigated apreviously described MCF-7-derived hypoxia-resistantcell population (Hypo-7 cells) [25] by RT-PCR, WB,IF, and IHC analysis. We found that, compared withparental MCF-7 cells, Hypo-7 cells expressed SLUG,
CK5, EGFR, CA-IX, and vimentin mRNA, as well aslacking HER2 and ERα gene expression (Figure 4D).Notably, Hypo-7 cells were also characterized by alack of epithelial features and by the acquisition ofmigrant behaviour in the collagen matrix (Figure 4D).These data indicate that exposure to hypoxia is asso-ciated with a basal-like gene expression pattern andwith SLUG gene up-regulation in luminal breast can-cer cells.
J Pathol 2008; 214: 25–37 DOI: 10.1002/pathCopyright 2007 Pathological Society of Great Britain and Ireland. Published by John Wiley & Sons, Ltd.

SLUG and basal-like breast carcinoma 31
Figure 4. SLUG mRNA expression is associated with the basal-like breast carcinoma phenotype and is up-regulated by hypoxia.(A) Bar-graph representation and representative picture of IHC analysis of CA-IX in breast cancer patients classified according tothe SLUG mRNA expression level. Note that the analysis of CA-IX by IHC was available only in 54 patients. (B) RT-PCR analysisof CA-IX mRNA in representative cases of breast cancer with H- (n = 9) or A- (n = 15) SLUG mRNA expression level. (C) MCF-7cells exposed to desferoxamine (DFX 100/300 µM, 72 h): RT-PCR analysis of the SLUG, CK5, EGFR, CA-IX, ERα, HER2, and β2 µmRNA level; immunofluorescence (IF) analysis of the CK5 protein level (DAPI was used for nuclear counterstaining); and westernblot (WB) analysis of the CA-IX and β-actin protein level. (D) MCF-7 and Hypo-7 cells: RT-PCR analysis of the SLUG, CK5, EGFR,CA-IX, ERα, HER2, vimentin, and β2 µ mRNA level; WB analysis of the SLUG, EGFR, CA-IX, and β-actin protein level; IF analysisof CK5; and IHC analysis of EGFR, ER, and vimentin. Scale bar = 100 µm
J Pathol 2008; 214: 25–37 DOI: 10.1002/pathCopyright 2007 Pathological Society of Great Britain and Ireland. Published by John Wiley & Sons, Ltd.

32 G Storci et al
SLUG regulates CA-IX and ERα gene expression
To unravel a functional role for SLUG gene expres-sion in the breast basal-like phenotype, we adminis-tered a SLUG-specific siRNA to MCF-7 cells exposedto DFX, and to Hypo-7 cells. We observed thatSLUG siRNA elicited down-regulation of CA-IXmRNA, as well as up-regulation of ERα mRNA, com-pared with the control (SCR) siRNA transfected cells(Figures 5A and 5B). Conversely, the expression ofother genes associated with the basal-like phenotype,such as EGFR, CK5, and HER2, was not affectedby SLUG/SCR siRNA administration (see Figures 5Aand 5B). These data indicate that SLUG modulates asubset (ie CA-IX and ERα mRNA) of basal-like phe-notype genes.
SLUG modulates in vitro invasiveness and in vivoaggressiveness of SLUG-expressing breast cancercells
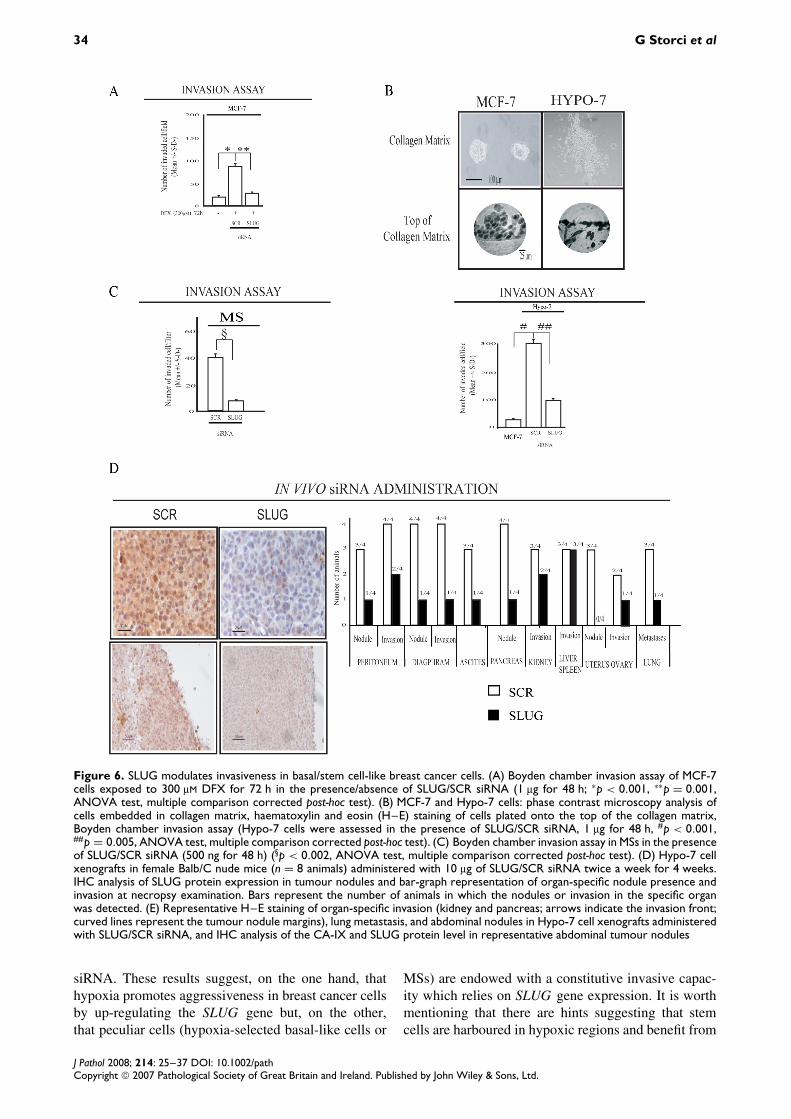
Basal-like carcinoma is a biologically aggressive andinvasive type of breast cancer [37,41,42]. In turn,SLUG induces an aggressive and invasive behaviourin cancer cells [16,19]. We observed that exposure to
DFX boosted the invasive capacity of MCF-7 cells(Figure 6A) and that the administration of SLUGsiRNA to DFX-treated MCF-7 cells induced a remark-able decrease in the invasive potential compared withSCR siRNA (Figure 6A). As reported in Figure 4D,we observed that Hypo-7 cells were endowed with aconstitutive migratory/invasive potential and that theadministration of SLUG siRNA induced a substan-tial decrease of their invasive potential compared withSCR siRNA (Figure 6B). Similarly, compared withSCR siRNA, SLUG siRNA administration decreasedthe invasive capacity of cells obtained from tumourMS dissociation (Figure 6C). We then evaluated theimpact of SLUG gene expression knock-down on thein vivo aggressiveness of Hypo-7 cells. We adminis-tered SLUG/SCR siRNA to Hypo-7 cell xenograftsin Balb/C nude mice. The capacity of SLUG siRNAto down-regulate SLUG gene expression in tumournodules was evaluated by IHC analysis of SLUGand E-cadherin gene expression, the latter being themost characterized SLUG down-regulated gene [17](Figure 6D and Supplementary Figure 2, available athttp://www.interscience.wiley.com/jpages/0022-3417/suppmat/path.2254.html). Necropsy examination of
Figure 5. SLUG modulates CA-IX and ERα gene expression. (A) RT-PCR analysis of the SLUG, CK5, EGFR, CA-IX, ERα, HER2, andβ2µ mRNA level in MCF-7 cells exposed to 100 or 300 µM DFX for 72 h in the presence of SLUG or SCR siRNA (1 µg for 48 h).(B) MCF-7 and Hypo-7 cells: RT-PCR analysis of the SLUG, CK5, EGFR, CA-IX, ERα, HER2, and β2µ, mRNA level in the presenceof SLUG or SCR siRNA (1 µg for 48 h)
J Pathol 2008; 214: 25–37 DOI: 10.1002/pathCopyright 2007 Pathological Society of Great Britain and Ireland. Published by John Wiley & Sons, Ltd.

SLUG and basal-like breast carcinoma 33
Hypo-7 xenografts revealed that the animals injectedwith the SLUG siRNA showed a reduced amountof nodules, a reduced degree of peritoneum anddiaphragm invasion, a lower degree of ascites, adiminished number of nodules and extent of invasionin abdominal organs (pancreas and kidney), reducedinvasion of pelvic organs (uterus and ovary), as wellas a reduction of lung metastasis (Figure 6D). Atmicroscopy analysis, a diminished degree of invasionin abdominal organs, such as the kidney and pancreas,as well as a reduced presence of lung metastases,was found in SLUG injected animals, compared withthose injected with SCR siRNA (Figure 6E). We alsoobserved that tumour nodules in the animals injectedwith SLUG siRNA showed well-defined and circum-scribed margins, whereas the tumour nodules in SCRsiRNA injected animals were surrounded by looselycohesive/spreading cells (Figure 6E). Notably, CA-IX and SLUG expression was high in the marginalzone of tumour nodules in SCR, but not SLUG,siRNA injected animals (Figure 6E). These data indi-cate that SLUG gene expression sustains the inva-sive/aggressive behaviour of basal-like breast cancercells.
Discussion
In the present work, we have shown that SLUGgene expression is associated with the basal-likebreast carcinoma phenotype [2–7]. This finding isin keeping with the results of a recent microarrayanalysis, which revealed that SLUG is overexpressedin breast cancer cell lines showing a basal-like breastcarcinoma phenotype [34]. In this investigation, wealso confirmed the previous evidence that the basal-like breast carcinoma phenotype is associated with theexpression of CA-IX, a hypoxia survival gene whoseexpression correlates with poor outcome in breastcancer patients [38–40].
These results prompted us to explore the connectionbetween the basal/stem-like phenotype and hypoxia.We proved that the exposure to hypoxia of SLUG-negative luminal-like breast cancer cells (MCF-7 cells)leads to up-regulation of SLUG gene expression andtriggers a basal-like carcinoma gene expression pat-tern. In line with this finding, Hypo-7 cells (selectedfrom MCF-7 cells exposed to hypoxia) showed con-stitutive up-regulation of the SLUG gene and a basal-like phenotype; this finding reinforces the notion thatthe basal-like phenotype is associated with a hypoxiaresponse. In addition, we also found that SLUG geneexpression is functionally relevant for the basal-likephenotype, being capable of regulating CA-IX and ERgene expression in MCF-7 cells exposed to a hypoxicenvironment and in MCF-7-derived hypoxia selectedcells. The absence of ER being a hallmark of basal-likebreast tumours [2–7,32,33], these data suggest thatSLUG is a chief orchestrator of the basal-like breastphenotype.
It has been recently proposed that basal-like breastcarcinoma is endowed with stem cellfeatures [9,10,32,34]. In keeping with this hypoth-esis, we have shown here that tumours expressinghigh levels of SLUG also display high mRNA lev-els of Bmi1 and CD133, two genes that have beenassociated with cancer stem cells in different tissues[36,37,41]. Interestingly, a recent microarray analy-sis detected CD133 expression in basal-like breastcarcinoma tumour samples [37,42]. Our results con-firm and expand on the concept that the basal-like breast carcinoma phenotype displays stem cellfeatures [9,10,34]. Moreover, these data allow theSLUG gene to be listed among those expressed bybasal/stem progenitor cells of the mammary gland.According to the above evidence, we have demon-strated here that tumour stem/progenitor cells prop-agated in vitro as MSs express SLUG mRNA andhigh levels of basal-like tumour markers (CK5 andEGFR), as well as Bmi1 and CD133 mRNA. Intrigu-ingly, we obtained the above data in MSs obtainedfrom ductal breast carcinomas showing a gene expres-sion pattern antithetic to that of basal-like breastcarcinomas: presence of ER (4 of 4); absence ofEGFR (4 of 4); and presence of HER2 (1 of 4).These data indicate that the stem/progenitor cellsfrom ductal breast carcinoma tissues show a basal-like breast carcinoma phenotype. Notably, we alsoshowed here that MSs obtained from the normalmammary gland express a basal/stem cell-like pat-tern (see Supplementary Figure 1, available onlineat http://www.interscience.wiley.com/jpages/0022-3417/suppmat/path.2254.html). These data suggestthat MSs may represent the in vitro counterpart of thebasal cell compartment, and thus reinforce the beliefthat stem/progenitor cells may be harboured in thebasal cell compartment [8,43]. The above data leadus to argue that the in vivo histological features ofductal breast carcinoma may be the consequence ofthe residual capacity of cancer stem/progenitor cellsto differentiate, whereas basal-like breast carcinomamay represent a tumour subtype in which the glandu-lar differentiation of cancer stem/progenitor cells doesnot manifest.
Data in the literature indicate that basal-like breasttumours are endowed with a peculiar aggressive/invasive behaviour [6,7]. We have provided here evi-dence that SLUG plays a primary role in such a biolog-ical characteristic, being responsible for the hypoxia-induced increase of the invasive potential of MCF-7 breast cancer cells. In addition, we have shownthat hypoxia-selected basal-like cells and tumour MSsexpress the SLUG gene in the absence of hypoxia andshow a constitutive capacity to invade the extracel-lular matrix, which depends on SLUG gene expres-sion. Such a modulatory effect of SLUG on malig-nancy is also evident in SLUG-expressing breastcancer cell xenografts, which show a remarkablereduction in tumour-spreading capacity and in inva-sive behaviour when administered with SLUG-specific
J Pathol 2008; 214: 25–37 DOI: 10.1002/pathCopyright 2007 Pathological Society of Great Britain and Ireland. Published by John Wiley & Sons, Ltd.
34 G Storci et al
Figure 6. SLUG modulates invasiveness in basal/stem cell-like breast cancer cells. (A) Boyden chamber invasion assay of MCF-7cells exposed to 300 µM DFX for 72 h in the presence/absence of SLUG/SCR siRNA (1 µg for 48 h; ∗p < 0.001, ∗∗p = 0.001,ANOVA test, multiple comparison corrected post-hoc test). (B) MCF-7 and Hypo-7 cells: phase contrast microscopy analysis ofcells embedded in collagen matrix, haematoxylin and eosin (H–E) staining of cells plated onto the top of the collagen matrix,Boyden chamber invasion assay (Hypo-7 cells were assessed in the presence of SLUG/SCR siRNA, 1 µg for 48 h, #p < 0.001,##p = 0.005, ANOVA test, multiple comparison corrected post-hoc test). (C) Boyden chamber invasion assay in MSs in the presenceof SLUG/SCR siRNA (500 ng for 48 h) (§p < 0.002, ANOVA test, multiple comparison corrected post-hoc test). (D) Hypo-7 cellxenografts in female Balb/C nude mice (n = 8 animals) administered with 10 µg of SLUG/SCR siRNA twice a week for 4 weeks.IHC analysis of SLUG protein expression in tumour nodules and bar-graph representation of organ-specific nodule presence andinvasion at necropsy examination. Bars represent the number of animals in which the nodules or invasion in the specific organwas detected. (E) Representative H–E staining of organ-specific invasion (kidney and pancreas; arrows indicate the invasion front;curved lines represent the tumour nodule margins), lung metastasis, and abdominal nodules in Hypo-7 cell xenografts administeredwith SLUG/SCR siRNA, and IHC analysis of the CA-IX and SLUG protein level in representative abdominal tumour nodules
siRNA. These results suggest, on the one hand, thathypoxia promotes aggressiveness in breast cancer cellsby up-regulating the SLUG gene but, on the other,that peculiar cells (hypoxia-selected basal-like cells or
MSs) are endowed with a constitutive invasive capac-ity which relies on SLUG gene expression. It is worthmentioning that there are hints suggesting that stemcells are harboured in hypoxic regions and benefit from
J Pathol 2008; 214: 25–37 DOI: 10.1002/pathCopyright 2007 Pathological Society of Great Britain and Ireland. Published by John Wiley & Sons, Ltd.

SLUG and basal-like breast carcinoma 35
Figure 6. Continued
the presence of hypoxia in their functioning and sur-vival [44,45]. Our data suggest that hypoxia may con-tribute to the maintenance of such an undifferentiatedphenotype by inducing a SLUG-mediated response.
In the literature, the worse clinical outcome ofbasal-like breast carcinoma patients is still contro-versial [46,47]. Moreover, there is seemingly a para-doxical dissociation between biological aggressivenessand chemo-sensitivity in basal-like breast carcinoma[6,7,37,48–50]. Indeed, in spite of the high grade of
histological dedifferentiation, basal-like breast carci-nomas seem to be more responsive to chemother-apy than ductal breast carcinomas [6,7,37,48–51].With the availability of the follow-up data, we testedwhether the SLUG gene expression level was associ-ated with survival in our patient set (see Supplemen-tary Figure 3, available at http://www.interscience.wiley.com/jpages/0022-3417/suppmat/path.2254.html). We found that tumours with a high SLUGmRNA expression level, despite their high histological
J Pathol 2008; 214: 25–37 DOI: 10.1002/pathCopyright 2007 Pathological Society of Great Britain and Ireland. Published by John Wiley & Sons, Ltd.

36 G Storci et al
grade, showed a favourable disease-free and overallsurvival, compared with those in which SLUG mRNAexpression was absent (see Supplementary Figure 3,available at http://www.interscience.wiley.com/jpages/0022-3417/suppmat/path.2254.html). Hence, thesedata support the notion that basal-like breast carcinomafeatures are associated with a favourable prognosis,even in patients who have undergone a heterogeneousset of therapy regimens.
Overall, the data reported in the present study indi-cate that SLUG is part of a hypoxia-induced geneticprogramme which confers a basal-like/aggressive phe-notype to cancer cells, and provide the first mechanis-tic insight into the basal-like breast cancer phenotype.
Acknowledgements
This work was supported by University of Bologna RFOFunds — ex 60%, the Cornelia Pallotti and Roberto PallottiFoundation (MB), and FIRB projects (PC). We also thankFondazione Cassa di Risparmio in Bologna for supporting theCenter for Applied Biomedical Research.
Supplementary material
Supplementary material may be found at the webaddress http://www.interscience.wiley.com/jpages/0022-3417/suppmat/path.2254.html
References
1. Landis SH, Murray T, Bolden S, Wingo PA. Cancer statistics.1999. CA Cancer J Clin 1999;49:8–31.
2. Andre F, Pusztai L. Molecular classification of breast cancer:implications for selection of adjuvant chemotherapy. Nature ClinPract Oncol 2006;3:621–632.
3. Perou CM, Sorlie T, Eisen MB, van de Rijn M, Jeffrey SS,Rees CA, et al. Molecular portraits of human breast tumours.Nature 2000;406:747–752.
4. Pusztai L, Mazouni C, Anderson K, Wu Y, Symmans WF.Molecular classification of breast cancer: limitations and potential.Oncologist 2006;11:868–877.
5. Nielsen TO, Hsu FD, Jensen K, Cheang M, Karaca G, Hu Z, et al.Immunohistochemical and clinical characterization of the basal-like subtype of invasive breast carcinoma. Clin Cancer Res2004;10:5367–5374.
6. Carey LA, Dees EC, Sawyer L, Gatti L, Moore DT, Collichio F,et al. The triple negative paradox: primary tumor chemosensitivityof breast cancer subtypes. Clin Cancer Res 2007;13:2329–2334.
7. Cleator S, Heller W, Coombes RC. Triple-negative breast cancer:therapeutic options. Lancet Oncol 2007;8:235–244.
8. Boecker W, Buerger H. Evidence of progenitor cells of glandularand myoepithelial cell lineages: cells in the human adult breastepithelium: a new progenitor (adult stem) concept. Cell Prolif2003;36:73–84.
9. Bertucci F, Cervera N, Birnbaum D. A gene signature in breastcancer. N Engl J Med 2007;356:1887–1888.
10. Liu R, Wang X, Chen GY, Dalerba P, Gurney A, Hoey T, et al.The prognostic role of a gene signature from tumorigenic breast-cancer cells. N Engl J Med 2007;356:217–226.
11. Thiery J. Epithelial–mesenchymal transitions in tumour progres-sion. Nature Rev 2002;2:442–454.
12. Al-Hajj M, Wicha MS, Benito-Hernandez A, Morrison SJ, ClarkeMF. Prospective identification of tumorigenic breast cancer cells.Proc Natl Acad Sci U S A 2003;100:3983–3988.
13. Dontu G, El-Ashry D, Wicha MS. Breast cancer, stem/progenitorcells and the estrogen receptor. Trends Endocrinol Metab2004;15:193–197.
14. Tripathi MK, Misra S, Chaudhuri G. Negative regulation ofthe expression of cytokeratins 8 and 19 by SLUG repressorprotein in human breast cells. Biochem Biophys Res Commun2005;329:508–515.
15. Martinez-Estrada OM, Culleres A, Soriano FX, Peinado H,Bolos V, Martinez FO, et al. The transcription factors Slug andSnail act as repressors of claudin-1 expression in epithelial cells.Biochem J 2006;394:449–457.
16. Barrallo-Gimeno A, Nieto MA. The Snail genes as inducers of cellmovement and survival: implications in development and cancer.Development 2005;132:3151–3161.
17. Conacci-Sorrell M, Simcha I, Ben-Yedidia T, Blechman J, Sav-agner P, Ben-Ze’ev A. Autoregulation of E-cadherin expressionby cadherin–cadherin interactions: the roles of β-catenin signal-ing, Slug, and MAPK. J Cell Biol 2003;163:847–857.
18. Shih JY, Tsai MF, Chang TH, Chang YL, Yuan A, Yu CJ, et al.Transcription repressor slug promotes carcinoma invasion andpredicts outcome of patients with lung adenocarcinoma. ClinCancer Res 2005;11:8070–8078.
19. Gupta PB, Kuperwasser C, Brunet JP, Ramaswamy S, Kuo WL,Gray JW, et al. The melanocyte differentiation program predis-poses to metastasis after neoplastic transformation. Nature Genet2005;37:1047–1054.
20. Shioiri M, Shida T, Koda K, Oda K, Seike K, Nishimura M,et al. Slug expression is an independent prognostic parameterfor poor survival in colorectal carcinoma patients. Br J Cancer2006;94:1816–1822.
21. Elloul S, Elstrand MB, Nesland JM, Trope CG, Kvalheim G,Goldberg I, et al. Snail, Slug, and Smad-interacting protein 1 asnovel parameters of disease aggressiveness in metastatic ovarianand breast carcinoma. Cancer 2005;103:1631–1643.
22. Come C, Magnino F, Bibeau F, De Santa Barbara P, Becker KF,Theillet C, et al. Snail and Slug play distinct roles during breastcarcinoma progression. Clin Cancer Res 2006;12:5395–5402.
23. Martin TA, Goyal A, Watkins G, Jiang WG. Expression ofthe transcription factors snail, slug, and twist and theirclinical significance in human breast cancer. Ann Surg Oncol2005;12:488–496.
24. Dontu G, Abdallah WM, Foley JM, Jackson KW, Clarke MF,Kawamura MJ, et al. In vitro propagation and transcriptionalprofiling of human mammary stem/progenitor cells. Genes Dev2003;17:1253–1270.
25. Sansone P, Storci G, Giovannini C, Pandolfi S, Pianetti S,Taffurelli M, et al. p66Shc/Notch-3 interplay controls self-renewaland hypoxia survival in human stem/progenitor cells of themammary gland expanded in vitro as mammospheres. Stem Cells2007;25:807–815.
26. WHO. The World Health Organization histologic typing of breasttumours (2nd edition). Am J Clin Pathol 1982;78:806–816.
27. Elston CW, Ellis IO. Pathological prognostic factors in breastcancer. I. The value of histological grade in breast cancer:experience from a large study with long-term follow-up.Histopathology 1991;19:403–410.
28. Dontu G, Jackson KW, McNicholas E, Kawamura MJ, Abdal-lah WM, Wicha MS. Role of Notch signaling in cell-fate determi-nation of human mammary stem/progenitor cells. Breast CancerRes 2004;6:R605–R615.
29. Ponti D, Costa A, Zaffaroni N, Pratesi G, Petrangolini G, Cora-dini D, et al. Isolation and in vitro propagation of tumorigenicbreast cancer cells with stem/progenitor cell properties. CancerRes 2005;65:5506–5511.
30. Chieco P, Jonker A, Van Noorden CJF (eds). Image Cytometry.BIOS: Oxford, 2001.
31. Reiner A, Neumeister B, Spona J, Reiner G, Schemper M,Jakesz R. Immunocytochemical localization of estrogen andprogesterone receptor and prognosis in human primary breastcancer. Cancer Res 1990;50:7057–7061.
J Pathol 2008; 214: 25–37 DOI: 10.1002/pathCopyright 2007 Pathological Society of Great Britain and Ireland. Published by John Wiley & Sons, Ltd.

SLUG and basal-like breast carcinoma 37
32. Abd El-Rehim DM, Pinder SE, Paish CE, Bell J, Blamey RW,Robertson JF, et al. Expression of luminal and basal cytokeratinsin human breast carcinoma. J Pathol 2004;203:661–671.
33. Livasy CA, Karaca G, Nanda R, Tretiakova MS, Olopade OI,Moore DT, et al. Phenotypic evaluation of the basal-like subtypeof invasive breast carcinoma. Mod Pathol 2006;19:264–271.
34. Charafe-Jauffret E, Ginestier C, Monville F, Finetti P, Ade-laide J, Cervera N, et al. Gene expression profiling of breastcell lines identifies potential new basal markers. Oncogene2006;25:2273–2284.
35. Rodriguez-Pinilla SM, Sarrio D, Honrado E, Moreno-Bueno G,Hardisson D, Calero F, et al. Vimentin and laminin expression isassociated with basal-like phenotype in both sporadic and BRCA1-associated breast carcinomas. J Clin Pathol 2007;60:1006–1012.
36. Prince ME, Sivanandan R, Kaczorowski A, Wolf GT, Kaplan MJ,Dalerba P, et al. Identification of a subpopulation of cells withcancer stem cell properties in head and neck squamous cellcarcinoma. Proc Natl Acad Sci U S A 2007;104:973–978.
37. Bertheau P, Turpin E, Rickman DS, Espie M, de Reynies A,Feugeas JP, et al. Exquisite sensitivity of TP53 mutant andbasal breast cancers to a dose-dense epirubicin–cyclophosphamideregimen. PLoS Med 2007;20:e90.
38. Garcia S, Dales JP, Charafe-Jauffret E, Carpentier-Meunier S,Andrac-Meyer L, Jacquemier J, et al. Poor prognosis in breastcarcinomas correlates with increased expression of targetableCD146 and c-Met and with proteomic basal-like phenotype. HumPathol 2007;38:830–841.
39. Makretsov NA, Huntsman DG, Nielsen TO, Yorida E, Pea-cock M, Cheang MC, et al. Hierarchical clustering analysisof tissue microarray immunostaining data identifies prognosti-cally significant groups of breast carcinoma. Clin Cancer Res2004;10:6143–6151.
40. Liu S, Dontu G, Mantle ID, Patel S, Ahn NS, Jackson KW,et al. Hedgehog signaling and Bmi1 regulate self-renewal ofnormal and malignant human mammary stem cells. Cancer Res2006;66:6063–6071.
41. O’Brien CA, Pollett A, Gallinger S, Dick JE. A human coloncancer cell capable of initiating tumour growth in immunodeficientmice. Nature 2007;445:106–110.
42. Brennan DJ, Jirstrom K, Kronblad A, Millikan RC, Landberg G,Duffy MJ, et al. CA IX is an independent prognostic marker inpremenopausal breast cancer patients with one to three positivelymph nodes and a putative marker of radiation resistance. ClinCancer Res 2006;12:6421–6431.
43. Boecker W, Moll R, Poremba C, Holland R, Van Diest PJ,Dervan P, et al. Common adult stem cells in the human breastgive rise to glandular and myoepithelial cell lineages: a new cellbiological concept. Lab Invest 2002;82:737–746.
44. Suda T, Arai F, Hirao A. Hematopoietic stem cells and their niche.Trends Immunol 2005;26:426–433.
45. Parmar K, Mauch P, Vergilio JA, Sackstein R, Down JD. Distri-bution of hematopoietic stem cells in the bone marrow according toregional hypoxia. Proc Natl Acad Sci U S A 2007;104:5431–5436.
46. Fulford LG, Reis-Filho JS, Ryder K, Jones C, Gillett CE, Hanby A,et al. Basal-like grade III invasive ductal carcinoma of the breast:patterns of metastasis and long-term survival. Breast Cancer Res2007;9:R4.
47. Sotiriou C, Neo SY, McShane LM, Korn EL, Long PM, Jaza-eri A, et al. Breast cancer classification and prognosis based ongene expression profiles from a population-based study. Proc NatlAcad Sci U S A 2003;100:10393–10398.
48. Sorlie T, Perou CM, Tibshirani R, Aas T, Geisler S, Johnsen H,et al. Gene expression patterns of breast carcinomas distinguishtumor subclasses with clinical implications. Proc Natl Acad Sci US A 2001;98:10869–10874.
49. Banerjee S, Reis-Filho JS, Ashley S, Steele D, Ashworth A,Lakhani SR, et al. Basal-like breast carcinomas: clinical outcomeand response to chemotherapy. J Clin Pathol 2006;59:729–735.
50. Rouzier R, Perou CM, Symmans WF, Ibrahim N, Cristofanilli M,Anderson K, et al. Breast cancer molecular subtypes responddifferently to preoperative chemotherapy. Clin Cancer Res2005;11:5678–5685.
51. James CR, Quinn JE, Mullan PB, Johnston PG, Harkin DP.BRCA1, a potential predictive biomarker in the treatment of breastcancer. Oncologist 2007;12:142–150.
J Pathol 2008; 214: 25–37 DOI: 10.1002/pathCopyright 2007 Pathological Society of Great Britain and Ireland. Published by John Wiley & Sons, Ltd.
Copyright © 2022 FDOKUMEN




















