
The Arabidopsis ascorbate peroxidase 3 is a peroxisomal membrane-bound antioxidant enzyme and is...
10
RESEARCH PAPER The Arabidopsis ascorbate peroxidase 3 is a peroxisomal membrane-bound antioxidant enzyme and is dispensable for Arabidopsis growth and development Savitha Narendra 1 , Sujatha Venkataramani 1 , Guoxin Shen 1 , Jing Wang 1, *, Vijaya Pasapula 1 , Yun Lin 2 , Dmytro Kornyeyev 1 , A. Scott Holaday 1 and Hong Zhang 1,† 1 Department of Biological Sciences, Texas Tech University, Lubbock, TX 79409, USA 2 Department of Crop Sciences, University of Illinois, Urbana, IL 61801, USA Received 21 February 2006; Accepted 20 May 2006 Abstract Ascorbate peroxidase (APX) exists as several isoforms that are found in various compartments in plant cells. The cytosolic and chloroplast APXs appear to play important roles in antioxidation metabolism in plant cells, yet the function of peroxisomal APX is not well studied. In this study, the localization of a putative peroxisomal membrane-bound ascorbate peroxidase, APX3 from Arabidopsis, was confirmed by studying the green fluorescent protein (GFP)–APX3 fusion protein in transgenic plants. GFP–APX3 was found to co-localize with a reporter protein that was targeted to peroxisomes by the peroxisomal targeting signal 1. The function of APX3 in Arabidopsis was investigated by analysing an APX3 knockout mutant under normal and several stress conditions. It was found that loss of function in APX3 does not affect Arabidopsis growth and development, suggesting that APX3 may not be an important antioxidant enzyme in Arabidopsis, at least under the conditions that were tested, or the function of APX3 could be compensated by other antioxidant enzymes in plant cells. Key words: Antioxidation, hydrogen peroxide scavenging, peroxisomal membrane targeting, reactive oxygen species, subcellular localization. Introduction Reactive oxygen species (ROS, e.g. O ÿ 2 and H 2 O 2 ) are produced as by-products of plant cellular metabolism (Asada and Takahashi, 1987; Foyer and Noctor, 2000). On the one hand, ROS serve as signalling molecules in plant redox signal transduction (Foyer and Noctor, 2005), and on the other hand ROS are harmful to plant cellular metabolism (Mittler, 2002; Apel and Hirt, 2004). Under stress conditions, ROS can be overproduced in plant cells, which creates a condition called oxidative stress that can damage cellular components including DNA, proteins, and membrane lipids (Mittler, 2002; Apel and Hirt, 2004). Plants have developed efficient antioxidant systems that can protect plants from oxidative stress (Asada, 1999; Mittler et al., 2004). These antioxidant systems include antioxidant molecules such as ascorbic acid, glutathione, a-tocopherol, and carotenoids, and antioxidant enzymes such as ascorbate peroxidase (APX), catalase, glutathione reductase (GR), and superoxide dismutase (Noctor and Foyer, 1998; Apel and Hirt, 2004). Antioxidant molecules can interact with ROS directly, thereby removing ROS from cells. However, they are more effective in serving as electron donors to scavenge ROS in reactions carried out by some antioxidant enzymes such as APX and GR (Noctor and Foyer, 1998; Apel and Hirt, 2004). For ex- ample, APX uses ascorbic acid as the electron donor to scavenge H 2 O 2 efficiently in plant cells (Asada, 1992, 1999; Shigeoka et al., 2002). APX exists in several isoforms that are found in various compartments of plant cells (Panchuk et al., 2002; * Present address: USDA-ARS Cropping Systems Research Laboratory, Lubbock, TX 79415, USA y To whom correspondence should be addressed. E-mail: [email protected] Abbreviations: APX, ascorbate peroxidase; ER, endoplasmic reticulum; GFP, green fluorescent protein; PTS1, peroxisomal targeting signal 1; RFP, red fluorescence protein; ROS, reactive oxygen species. Journal of Experimental Botany, Vol. 57, No. 12, pp. 3033–3042, 2006 doi:10.1093/jxb/erl060 Advance Access publication 26 July, 2006 ª The Author [2006]. Published by Oxford University Press [on behalf of the Society for Experimental Biology]. All rights reserved. For Permissions, please e-mail: [email protected] by guest on July 13, 2014 http://jxb.oxfordjournals.org/ Downloaded from
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of The Arabidopsis ascorbate peroxidase 3 is a peroxisomal membrane-bound antioxidant enzyme and is...

RESEARCH PAPER
The Arabidopsis ascorbate peroxidase 3 is a peroxisomalmembrane-bound antioxidant enzyme and is dispensablefor Arabidopsis growth and development
Savitha Narendra1, Sujatha Venkataramani1, Guoxin Shen1, Jing Wang1,*, Vijaya Pasapula1, Yun Lin2,
Dmytro Kornyeyev1, A. Scott Holaday1 and Hong Zhang1,†
1 Department of Biological Sciences, Texas Tech University, Lubbock, TX 79409, USA2 Department of Crop Sciences, University of Illinois, Urbana, IL 61801, USA
Received 21 February 2006; Accepted 20 May 2006
Abstract
Ascorbate peroxidase (APX) exists as several isoforms
that are found in various compartments in plant cells.
The cytosolic and chloroplast APXs appear to play
important roles in antioxidation metabolism in plant
cells, yet the function of peroxisomal APX is not well
studied. In this study, the localization of a putative
peroxisomal membrane-bound ascorbate peroxidase,
APX3 from Arabidopsis, was confirmed by studying
the green fluorescent protein (GFP)–APX3 fusion
protein in transgenic plants. GFP–APX3 was found
to co-localize with a reporter protein that was targeted
to peroxisomes by the peroxisomal targeting signal 1.
The function of APX3 in Arabidopsis was investigated
by analysing an APX3 knockout mutant under normal
and several stress conditions. It was found that loss of
function in APX3 does not affect Arabidopsis growth
and development, suggesting that APX3 may not be an
important antioxidant enzyme in Arabidopsis, at least
under the conditions that were tested, or the function
of APX3 could be compensated by other antioxidant
enzymes in plant cells.
Key words: Antioxidation, hydrogen peroxide scavenging,
peroxisomal membrane targeting, reactive oxygen species,
subcellular localization.
Introduction
Reactive oxygen species (ROS, e.g. O�2 and H2O2) are
produced as by-products of plant cellular metabolism(Asada and Takahashi, 1987; Foyer and Noctor, 2000).On the one hand, ROS serve as signalling molecules inplant redox signal transduction (Foyer and Noctor, 2005),and on the other hand ROS are harmful to plant cellularmetabolism (Mittler, 2002; Apel and Hirt, 2004). Understress conditions, ROS can be overproduced in plant cells,which creates a condition called oxidative stress that candamage cellular components including DNA, proteins,and membrane lipids (Mittler, 2002; Apel and Hirt, 2004).Plants have developed efficient antioxidant systems thatcan protect plants from oxidative stress (Asada, 1999;Mittler et al., 2004). These antioxidant systems includeantioxidant molecules such as ascorbic acid, glutathione,a-tocopherol, and carotenoids, and antioxidant enzymessuch as ascorbate peroxidase (APX), catalase, glutathionereductase (GR), and superoxide dismutase (Noctor andFoyer, 1998; Apel and Hirt, 2004). Antioxidant moleculescan interact with ROS directly, thereby removing ROSfrom cells. However, they are more effective in serving aselectron donors to scavenge ROS in reactions carried outby some antioxidant enzymes such as APX and GR(Noctor and Foyer, 1998; Apel and Hirt, 2004). For ex-ample, APX uses ascorbic acid as the electron donor toscavenge H2O2 efficiently in plant cells (Asada, 1992, 1999;Shigeoka et al., 2002).
APX exists in several isoforms that are found in variouscompartments of plant cells (Panchuk et al., 2002;
* Present address: USDA-ARS Cropping Systems Research Laboratory, Lubbock, TX 79415, USAy To whom correspondence should be addressed. E-mail: [email protected]: APX, ascorbate peroxidase; ER, endoplasmic reticulum; GFP, green fluorescent protein; PTS1, peroxisomal targeting signal 1; RFP, redfluorescence protein; ROS, reactive oxygen species.
Journal of Experimental Botany, Vol. 57, No. 12, pp. 3033–3042, 2006
doi:10.1093/jxb/erl060 Advance Access publication 26 July, 2006
ª The Author [2006]. Published by Oxford University Press [on behalf of the Society for Experimental Biology]. All rights reserved.For Permissions, please e-mail: [email protected]
by guest on July 13, 2014http://jxb.oxfordjournals.org/
Dow
nloaded from

Shigeoka et al., 2002; Chew et al., 2003). The cytosolicAPXs play important roles in antioxidant metabolism inplant cells. The loss-of-function mutation in APX1 thatencodes a cytosolic APX in Arabidopsis leads to lowerphotosynthetic rates, slower growth, and delayed floweringunder normal growth conditions in comparison with thesecharacteristics of wild-type plants (Pnueli et al., 2003).Under high-intensity light, the chloroplast H2O2-scavengingsystem is unable to keep pace with H2O2 production, andH2O2 accumulates, causing massive protein oxidation inAPX1-deficient cells (Davletova et al., 2005). These datasuggest that cytosolic APXs, particularly APX1, play a keyrole in protecting chloroplasts under light stress conditions.Chloroplast APXs also play important roles in photosyn-thesis under stress conditions (Asada, 1999). For example,the loss of a thylakoid membrane-bound APX (tAPX) inhexaploid wheat leads to a reduction in CO2 assimilationand photosynthetic electron transport under moderate lightintensities (Danna et al., 2003). Overexpression of a geneencoding a tAPX in tobacco or in Arabidopsis significantlyincreases tolerance to oxidative stress caused by the ap-plication of methylviologen and by chilling temperaturewith high-intensity light (Yabuta et al., 2002), and to nitricoxide-induced cell death (Murgia et al., 2004). It appearsthat with normal levels of cytosolic APXs, the tAPX isa limiting factor of antioxidant systems under oxidativestress in chloroplasts. Interestingly, a chloroplast stromalAPX that is also targeted to mitochondria does not appearto be important in the presence of cytosolic APXs in plantantioxidation metabolism, because a knockout mutant ofthe stromal/mitochondrial APX does not display any vis-ible phenotype or difference in photosynthetic rate whencompared with wild-type plants under controlled condi-tions (Davletova et al., 2005).
In comparison with the rich literature on the functions ofcytosolic and chloroplast APXs, relatively little is knownabout the function of peroxisomal/glyoxysomal APXs(pAPX or gAPX). Although APXs have been found inperoxisomal and glyoxysomal membranes for many years(Yamaguchi et al., 1995; Bunkelman and Trelease, 1996;Ishikawa et al., 1998), their function was mainly specula-tive based on the functions of this organelle (Corpas et al.,2001; del Rio et al., 2002), not derived from experimen-tation. Recently, two genes encoding putative peroxisomalmembrane-bound APX were isolated from Arabidopsisand barley, and their roles in plant antioxidation metab-olism were assessed by overexpression in transgenic plants.Overexpression of APX3 that encodes a putative peroxi-somal membrane-bound APX in Arabidopsis leads toincreased tolerance against aminotriazole treatment intransgenic tobacco plants (Wang et al., 1999). Amino-triazole is an irreversible inhibitor of peroxisomal catalase(Amory et al., 1992), which explains why APX3-overexpressing plants did better in the presence of amino-triazole, because APX3 becomes the main enzyme that
scavenges H2O2 produced in photorespiratory metabolismin peroxisomes in the absence of active catalase. Further-more, overexpression of APX3 leads to higher seed pro-duction under moderate drought conditions in tobaccoplants (Yan et al., 2003). When a putative peroxisomalmembrane-bound APX from barley was overexpressed inArabidopsis, an increased tolerance to higher temperaturetreatment was observed (Shi et al., 2001). These dataindicate that the increased level of APX in peroxisomalmembranes appears to increase stress tolerance in plants.However, these studies did not address the question as tohow important is the peroxisomal APX to cellular anti-oxidation metabolism.
One objective of this research was to obtain evidence thatthe Arabidopsis APX3 is indeed a peroxisomal membrane-bound antioxidant enzyme. A further objective was to studythe possible function of APX3 by analysing the perfor-mance of APX3 knockout mutants under conditions thatwould favour oxidative stress. The APX3-deficient plantsdid not display any major abnormalities in growth anddevelopment under normal growth condition, or under thestress conditions that were tested here, suggesting thatperhaps APX3 is not absolutely required for Arabidopsisgrowth and development.
Materials and methods
Construction of GFP–APX3 fusion construct and
Arabidopsis transformation
The green fluorescent protein (GFP) coding sequence was amplifiedby polymerase chain reaction (PCR) using primers gFP-3 and gFP-5,digested with enzymes BamHI and XbaI, and subcloned into thevector pGEM-3Z (Promega, Madison, WI, USA) to form theintermediate vector pGEM-3Z-GFP-B. The full-length APX3 wasamplified by PCR from a cDNA library with primers APX3-18 andAPX3-19, digested with BamHI, and then subcloned into the C-terminal end of GFP in the intermediate vector pGEM-3Z-GFP-B toform the vector pGEM-3Z-GFP-APX3. The correct orientation of theDNA insert was confirmed by PCR using GFP-specific and APX3-specific oligonucleotides, and sequenced for accuracy. The vectorpGEM-3Z-GFP-APX3 was digested with enzymes XbaI and SacI(partial digestion due to an internal SacI site), and the GFP–APX3fusion fragment was then subcloned into pBI121 (Jefferson et al.,1987) by replacing the ‘GUS’ gene to form the transforming vectorthat was transformed into the Agrobacterium tumefaciens GV3101.The Arabidopsis transformation was conducted according to the pro-tocol of Clough and Bent (1998). The sequences of oligonucleotidesused for cloning are listed as follows: APX3-18, 59-TAGTCG-GATCCTTACTTCATCCTCTTCCGAATC-39; APX3-19, 59-CA-GTCGGATCCTCATGGCTGCACCGATT-39; gFP-3, 59-CAGCT-GAGCTCGGATCCCTTTGTATAGTTCATCCATGC-39; and gFP-5,59-CGTACTCTAGAATGAGTAAAGGAGAAGAACT-39.
RNA isolation and hybridization
Total RNAs were isolated from 3-week-old Arabidopsis plantsusing the TRIzol reagent from Invitrogen (Carlsbad, CA, USA),separated by electrophoresis (10 lg per lane), blotted to a Nylonmembrane, and hybridized with various probes. Hybridization wascarried out according to the method of Church and Gilbert (1984)
3034 Narendra et al.
by guest on July 13, 2014http://jxb.oxfordjournals.org/
Dow
nloaded from

using probes labelled by random priming. The washing conditionswere as follows: once for 10 min in 0.5% bovine serum albumin(BSA), 1 mM EDTA, 40 mM NaHPO4 (pH 7.2), and 5.0% SDS at64 8C; then four times (10 min each) in 0.1 mM EDTA, 40 mMsodium phosphate (pH 7.2), and 1% SDS at 64 8C. The same filterwas used for hybridizations with probes APX3 and PolyUb consec-utively. The condition for stripping the filter was as follows: twice(15 min each) in 2 mM TRIS (pH 8.2), 2 mM EDTA (pH 8.0),and 0.08% SDS. The polyubiquitin gene [Arabidopsis BiologicalResources Center (ABRC) number ATIS 0348] was obtainedfrom ABRC at Ohio State University, Columbus, OH, USA.
Immunoblot analysis
Plants at the 5–6 leaf stage were ground to a fine powder in liquidnitrogen. The powder was transferred to a fresh tube containing 200ll of 50 mMNaHPO4 (pH 7.0) and centrifuged at 15 000 g for 10 minat 4 8C. The supernatant that contained mainly cytosolic proteinswas discarded, and the pellet was re-suspended in 200 ll of 50 mMsodium phosphate (pH 7.0) and 200 ll of 23 SDS buffer [125 mMTRIS–HCl, 2% SDS, 20% glycerol, 200 mM dithiothreitol (DTT),0.02% bromophenol blue, pH 6.8]. The resuspended pellet was boiledin a water bath for 10 min and centrifuged at 15 000 g for 10 min at4 8C. The supernatant was transferred to a fresh tube, and the proteinconcentration in the supernatant was determined by using theBradford assay (Bradford, 1976). Proteins from GFP–APX3 trans-genic plants, APX3 knockout plants, and wild-type plants weresubjected to electrophoresis in a 12% SDS–polyacrylamide gel. Theconditions for blotting and colour development were the same asdescribed previously (Yan et al., 2003), except that the antibody usedwas anti-APX3 antibody.
Protoplast isolation
Protoplasts were isolated using a modified method of Sheen (2002).Large rosette leaves from 3-week-old seedlings were rinsed, blotteddry, piled one on top of another, and sliced into 0.5–1 mm strips.The stripswere transferred to a 25ml flask containing the enzyme solu-tion (1% cellulase, 0.25%macerozyme, 0.4 Mmannitol, 20 mMKCl,20 mM MES, 10 mM CaCl2, 5 mM mercaptoethanol, 0.1% BSA,pH 5.7) and subjected to continuous shaking at 40 rpm for 90 min.After this step, the speed was increased to 80 rpm for 1 min to aidin the complete release of protoplasts. The enzyme solution waspassed through a 70 lm nylon mesh and centrifuged at 60 g for5 min. The pellet was washed twice with wash and incubation solution(0.5 M mannitol, 4 mM MES, 20 mM KCl, pH 5.7) and thenresuspended in 100 ll of the wash and incubation solution.
Creation of transgenic plants that express GFP–APX3 and
RFP–PTS1 fusion proteins
Five independent transgenic lines that express GFP–APX3 fusionprotein at high levels (based on western blot analysis) were separatelycrossed to a line that expresses red fluorescent protein (RFP)–peroxisomal targeting signal 1 (PTS1) fusion protein (Lin et al.,2004), and the F1 progeny from these crosses were analysed for thepresence of both GFP–APX3 and RFP–PTS1 fusion proteins. Allfive F1 progeny gave rise to similar results under the fluorescencemicroscope.
Arabidopsis growth and stress treatments
Arabidopsis seeds were sown on MS medium and kept at 4 8C for5 d before being allowed to grow at room temperature for 7 d, andthen they were divided into three groups: one for the normal growthcondition, one for chilling temperature treatment, and one for heattreatment. For the normal growth condition, plants were grown underwhite light (60 lE m2 s�1) at 22 8C for 21 d before harvesting for
biomass analysis. For chilling temperature treatment, plants were putinside a refrigerator with white light (50 lEm2 s�1) and a temperatureof 8 8C for 33 d before harvesting for biomass analysis. For heattreatment, plants were put in a growth chamber with white light(200 lE m2 s�1) and a temperature cycle of 40 8C for 3 h and 22 8Cfor 21 h d�1 for 25 d before harvesting for biomass analysis.
Seed germination experiment
Sterilized seeds of wild-type plants and the APX3 knockout mutantwere planted on MS plates that contained either 50 or 100 mM NaClfor germination. The percentage of germination was calculated as theratio of seeds that germinated to total seeds planted 7 d afterstratification.
Fluorescence microscopy analysis
The transgenic plants that express the GFP–APX3 fusion protein ata high level selected from western blot analysis were used forfluorescence microscopy analysis. Protoplasts made from GFP–APX3 transgenic and wild-type plants were analysed with anOlympus BX-50 fluorescence microscope. The flower petals androot tissues from F1 transgenic plants that expressed both GFP–APX3and RFP–PTS1 fusion proteins were used directly for localizationstudies using a fluorescence microscope. The GFP images werethen taken using a fluorescence microscope equipped with an exciterfilter (HQ 470/40), a dichroic mirror (Q495LP), and a barrier filter(HQ 525/50). The red fluorescence images of the RFP–PTS1 fusionprotein and chloroplasts were taken using an exciter filter (BP530/550), a dichroic mirror (DM570), and a barrier filter (BA590).The overlay images were created using the Simple PCI software(version 4.0). The black and white images for the same fields wereobtained using white light on the same microscope.
Chlorophyll fluorescence measurement
Detached dark-acclimated leaves were placed in the temperature-controlled chamber of an oxygen electrode (Hansatech, Kings Lynn,Norfolk, UK) and illuminated by actinic light (photon flux density =350 lmol m�2 s�1) at room temperature (25 8C) and ambient CO2.Then the temperature was quickly (within 5 min) increased up to 40or 38 8C. Chlorophyll fluorescence emission from the leaves wasmeasured with a pulse amplitude-modulated fluorometer (PAM 101/103, Heinz Walz GmbH, Effeltrich, Germany) through a port in thechamber at different times during the light exposure. Saturating lightpulses, 2 s in duration, were provided by a KL 1500 light source(Schott, Wiesbaden, Germany). The experimental protocol describedby Schreiber et al. (1986) and the nomenclature of van Kooten andSnel (1990) were used. Quantum efficiency of electron transport inphotosystem II complexes (UPSII) was estimated as described byGenty et al. (1989). The following formula was used:
UPSII = ðF9m � FÞ=F9mwhere F9m is maximal chlorophyll fluorescence yield by the saturat-ing pulse light under actinic light, and F is the level of fluorescencejust before application of a saturating flash.
Results
APX3 is predicted to be a peroxisomalmembrane-bound enzyme
Although APX3 from Arabidopsis shows the highestsequence similarity to the cotton pAPX and pumpkinpAPX throughout its entire length (Zhang et al., 1997),including the transmembrane domain at the C-terminus
Subcellular localization and function of APX3 3035
by guest on July 13, 2014http://jxb.oxfordjournals.org/
Dow
nloaded from

(Bunkelmann and Trelease, 1996; Nito et al., 2001), APX3was never experimentally determined to be localized toglyoxysomal or peroxisomal membranes. The cotton pAPXis localized to the peroxisomal membrane (Mullen et al.,1999), and the sequence that targets cotton pAPX to theperoxisomal membrane is composed of a C-terminal trans-membrane domain followed by a few basic amino acidresidues (Mullen et al., 2000, 2001). This sequence wasdefined as mPTS (i.e. the targeting signal of peroxisomalmembrane-bound proteins) (Mullen et al., 2000, 2001).
Since the C-terminal sequence of APX3 resembles themPTS from cotton pAPX, it is predicted that APX3 isperoxisomal membrane-bound.
Compared with other Arabidopsis APXs, APX3 is mostsimilar to APX5 (Panchuk et al., 2002). Like APX3, APX5also has a C-terminal transmembrane domain followed bya few positively charged amino acid residues (Fig. 1). Itappears that there are only two APXs in Arabidopsis thatare likely to be peroxisomal membrane-bound. The thirdAPX that contains a C-terminal transmembrane is the
Fig. 1. Sequence comparison of ArabidopsisAPX proteins. Amino acid residues that are completely conserved are in dark blue, those that share>75%homology are in red, and those that share>50% homology are in light blue. The transmembrane domain found at the C-terminal end of APX3 or APX5is underlined, and the positively charged amino acid residues following the transmembrane domain are marked with asterisks.
3036 Narendra et al.
by guest on July 13, 2014http://jxb.oxfordjournals.org/
Dow
nloaded from

thylakoid membrane-bound APX in Arabidopsis (i.e. tAPXin Fig. 1). The other five APX isoforms in Arabidopsisdo not have transmembrane domains (Fig. 1). APX1 andAPX2 are cytosolic proteins, APX4 may be a peroxisomalmatrix protein (Panchuk et al., 2002) or a chloroplastprotein (Chew et al., 2003), APX6 may be a cytosolicprotein (Panchuk et al., 2002; Chew et al., 2003), andfinally the stromal APX (i.e. sAPX) can also be targetedto mitochondria (Chew et al., 2003). In addition, two moreAPXs were found in the Arabidopsis genome (Narendra,2005): APX7 (At1g33660) and an unknown APX(At1g43220). APX7 is a very small APX and its subcellularlocalization is not known. The unknown APX has noexpressed sequence tag (EST) sequences associated withit; therefore it might be a pseudogene (Narendra, 2005).
APX3 is localized to the peroxisomal membrane
In order to localize APX3, it was fused to the C-terminalside of the GFP and the subcellular localization of the GFP–APX3 fusion protein was studied. More than 30 transgenicplants were created and it was found that most of themexpressed the GFP–APX3 fusion transcript at high levels(Fig. 2A). The steady-state levels of the GFP–APX3 fusionprotein were then studied by western blot analysis in linesthat contained high levels of the transgene transcript. Asexpected, all high expression lines at the transcript levelalso expressed GFP–APX3 fusion protein (Fig. 2B).
The subcellular localization of GFP–APX3 fusion proteinwas analysed in those high expression lines. Fluorescencemicroscopy was used to observe the green fluorescencesignal in protoplasts made from high expression lines. Thedata clearly indicated that the GFP–APX3 fusion protein
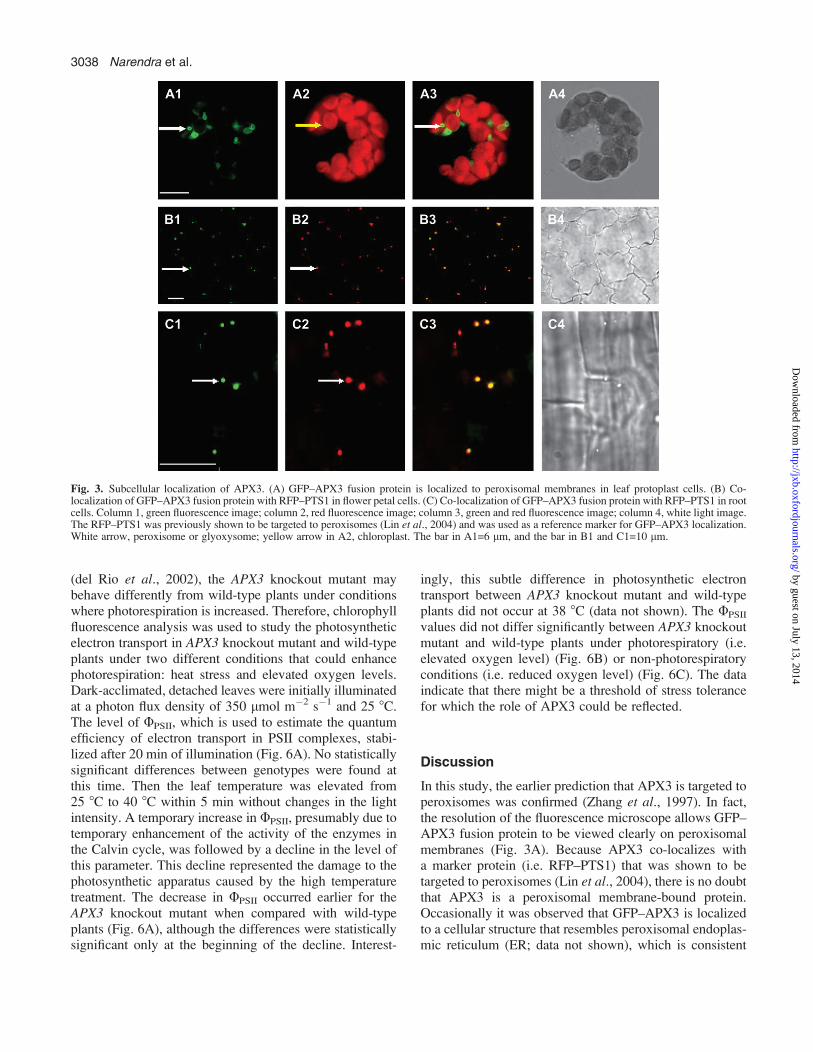
was found in the membranes of organelles that appeared tobe peroxisomes (Fig. 3A). To prove that the organelles towhich GFP–APX3 was targeted were indeed peroxisomes,a reference marker line was used that expressed RFP thatwas tagged with PTS1. The RFP–PTS1 fusion protein waspreviously shown to be targeted to peroxisomes by Linet al. (2004). GFP–APX3 lines were crossed to RFP–PTS1lines to obtain F1 plants that expressed both fusion proteins,and then the localization of these two fusion proteins wasdetermined using fluorescence microscopy. To avoidthe red fluorescence signal coming from chlorophylls, cellsthat did not contain chloroplasts were used. It was foundthat the two fusion proteins co-localized to the sameorganelles, namely peroxisomes in petal cells (Fig. 3B)or glyoxysomes in root cells (Fig. 3C). Based on thesedata, it was concluded that APX3 is localized to theperoxisomal/glyoxysomal membranes.
APX3 is dispensable for Arabidopsis growth anddevelopment
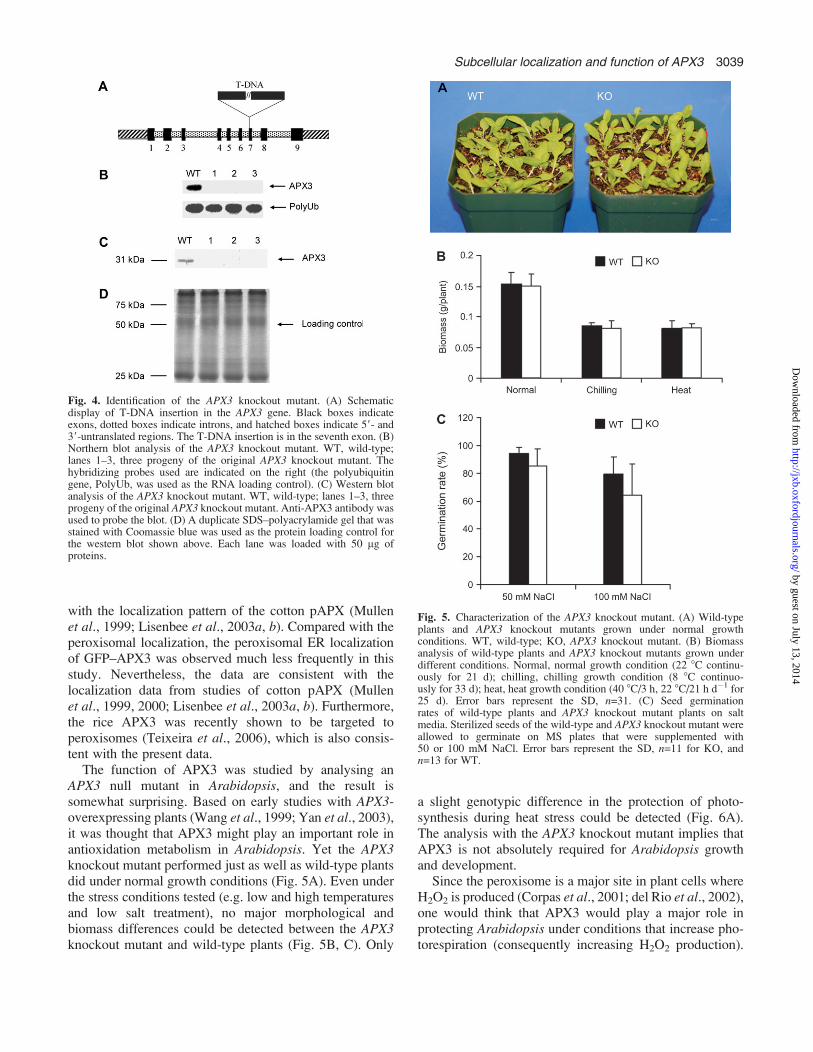
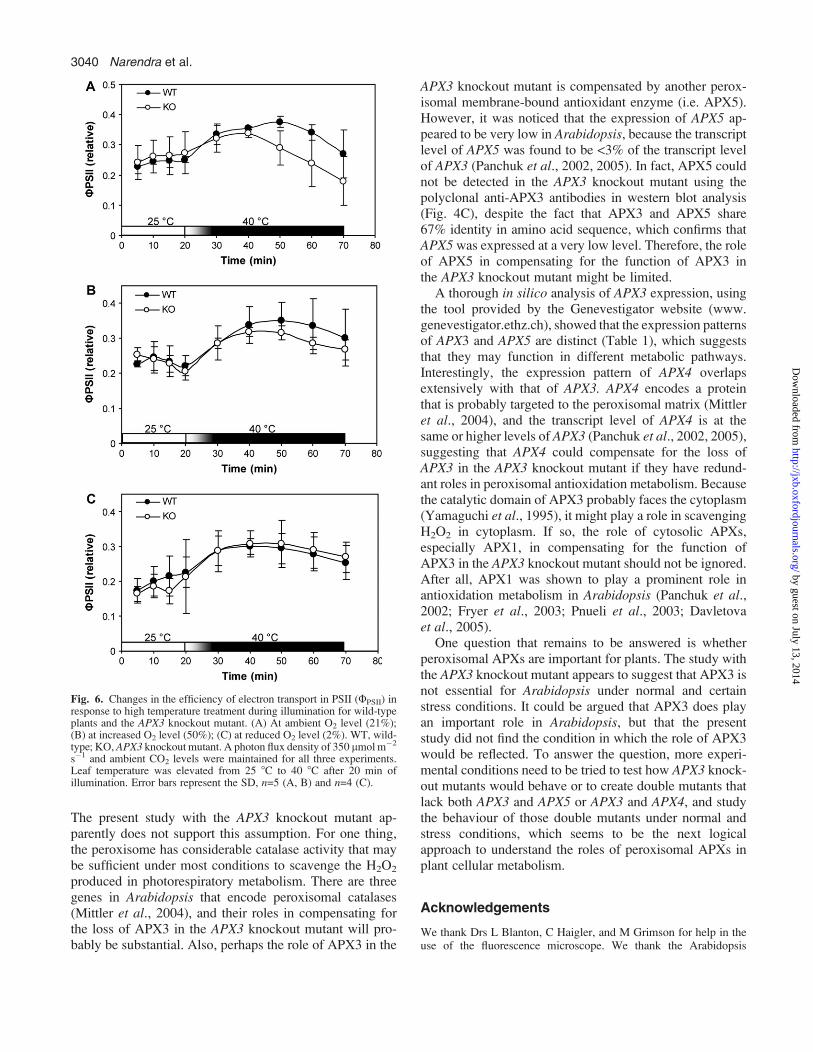
Previously it was demonstrated that overexpression ofAPX3 increases photosynthetic tolerance to drought stress(Yan et al., 2003) and oxidative stress (Wang et al., 1999)in transgenic plants. Therefore, it ws thought that APX3might play an important role in plant cellular metabolism.To explore the function of APX3 further, an APX3knockout mutant was studied that was obtained from theABRC at Ohio State University. This mutant was due toa T-DNA insertion in the seventh exon (Fig. 4A), whichleads to a null phenotype at the transcript and protein level(Fig. 4B, C). The size of the T-DNA insert is larger than5 kb, which might result in highly unstable transcriptsand disrupt the translation process. Despite several trials,no APX3 transcript was ever detected from this mutant.Consequently, no APX3 protein could be detected bywestern blot analysis (Fig. 4B). The phenotypes and growthbehaviours of the APX3 knockout mutant were examinedunder normal and temperature stress conditions. No majormorphological and biomass differences were detected be-tween APX3 knockout mutant and wild-type plants underthese conditions (Fig. 5A, B). Since H2O2 is produced fromperoxisomes during lipid mobilization (via fatty acidb-oxidation) in seed germination, the seed germinationrates of APX3 knockout mutant and wild-type plants werecompared under salt stress conditions. Again no significantdifferences were observed between APX3 knockout mu-tant and wild-type plants (Fig. 5C). The data indicatethat APX3 is dispensable for Arabidopsis under normalgrowth conditions and the stress conditions tested.
APX3 knockout plants display an altered response toheat treatment
Since peroxisomal APX is probably involved in scaveng-ing H2O2 produced during photorespiratory metabolism
Fig. 2. Identification of GFP–AXP3-overexpressing lines. (A) Northernblot analysis of GFP–APX3 transgenic plants. WT, wild-type; lanes A–O,15 independent GFP–APX3 transgenic lines. The endogenous APX3transcript and the GFP–APX3 transgene transcript are marked on the right.An APX3 cDNA fragment was used as the probe, and the polyubiquitingene (Poly Ub) was used as the RNA loading control in the northernblot experiments. (B) Western blot analysis of GFP–APX3 transgenicplants. WT, wild-type; lanes 1–6, six independent GFP–APX3 transgenicplants, which correspond to lanes A, C, E, H, K, and M, respectively, ofthe northern blot shown above. The GFP–APX3 fusion protein andthe endogenous APX3 are marked on the right. Each lane was loaded with50 lg of proteins. The polyclonal anti-APX3 antibodies were used in thewestern blot experiment.
Subcellular localization and function of APX3 3037
by guest on July 13, 2014http://jxb.oxfordjournals.org/
Dow
nloaded from
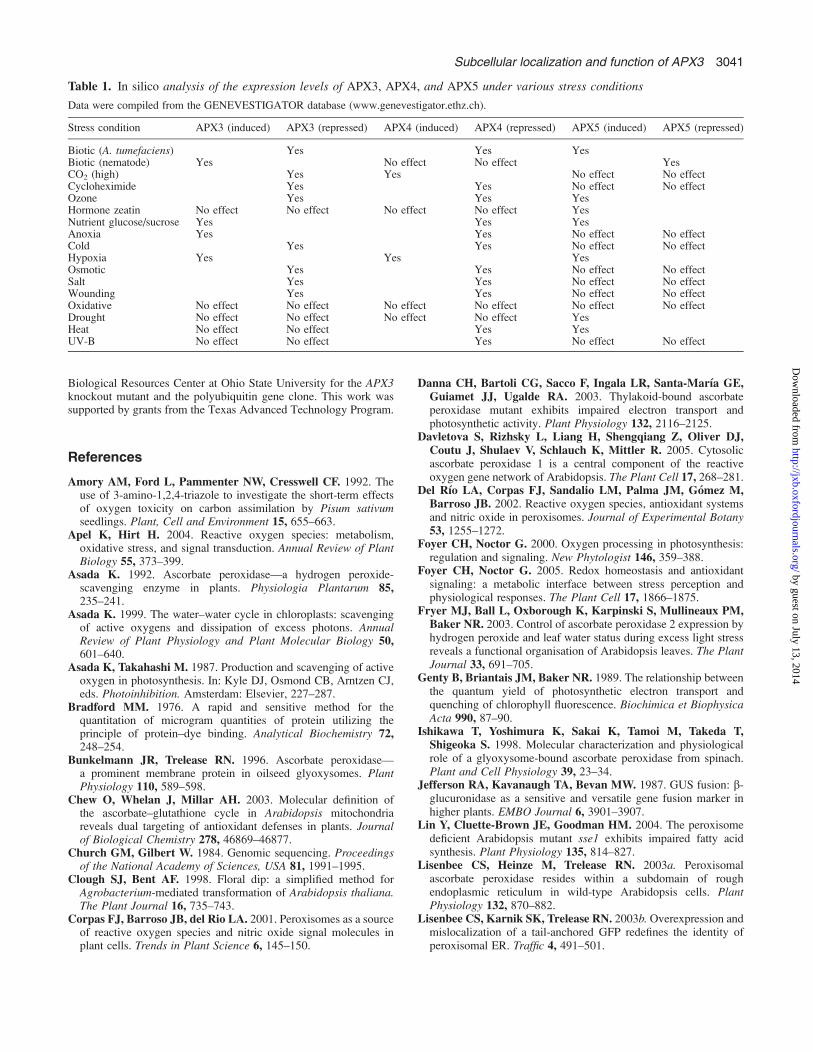
(del Rio et al., 2002), the APX3 knockout mutant maybehave differently from wild-type plants under conditionswhere photorespiration is increased. Therefore, chlorophyllfluorescence analysis was used to study the photosyntheticelectron transport in APX3 knockout mutant and wild-typeplants under two different conditions that could enhancephotorespiration: heat stress and elevated oxygen levels.Dark-acclimated, detached leaves were initially illuminatedat a photon flux density of 350 lmol m�2 s�1 and 25 8C.The level of UPSII, which is used to estimate the quantumefficiency of electron transport in PSII complexes, stabi-lized after 20 min of illumination (Fig. 6A). No statisticallysignificant differences between genotypes were found atthis time. Then the leaf temperature was elevated from25 8C to 40 8C within 5 min without changes in the lightintensity. A temporary increase in UPSII, presumably due totemporary enhancement of the activity of the enzymes inthe Calvin cycle, was followed by a decline in the level ofthis parameter. This decline represented the damage to thephotosynthetic apparatus caused by the high temperaturetreatment. The decrease in UPSII occurred earlier for theAPX3 knockout mutant when compared with wild-typeplants (Fig. 6A), although the differences were statisticallysignificant only at the beginning of the decline. Interest-
ingly, this subtle difference in photosynthetic electrontransport between APX3 knockout mutant and wild-typeplants did not occur at 38 8C (data not shown). The UPSII
values did not differ significantly between APX3 knockoutmutant and wild-type plants under photorespiratory (i.e.elevated oxygen level) (Fig. 6B) or non-photorespiratoryconditions (i.e. reduced oxygen level) (Fig. 6C). The dataindicate that there might be a threshold of stress tolerancefor which the role of APX3 could be reflected.
Discussion
In this study, the earlier prediction that APX3 is targeted toperoxisomes was confirmed (Zhang et al., 1997). In fact,the resolution of the fluorescence microscope allows GFP–APX3 fusion protein to be viewed clearly on peroxisomalmembranes (Fig. 3A). Because APX3 co-localizes witha marker protein (i.e. RFP–PTS1) that was shown to betargeted to peroxisomes (Lin et al., 2004), there is no doubtthat APX3 is a peroxisomal membrane-bound protein.Occasionally it was observed that GFP–APX3 is localizedto a cellular structure that resembles peroxisomal endoplas-mic reticulum (ER; data not shown), which is consistent
Fig. 3. Subcellular localization of APX3. (A) GFP–APX3 fusion protein is localized to peroxisomal membranes in leaf protoplast cells. (B) Co-localization of GFP–APX3 fusion protein with RFP–PTS1 in flower petal cells. (C) Co-localization of GFP–APX3 fusion protein with RFP–PTS1 in rootcells. Column 1, green fluorescence image; column 2, red fluorescence image; column 3, green and red fluorescence image; column 4, white light image.The RFP–PTS1 was previously shown to be targeted to peroxisomes (Lin et al., 2004) and was used as a reference marker for GFP–APX3 localization.White arrow, peroxisome or glyoxysome; yellow arrow in A2, chloroplast. The bar in A1=6 lm, and the bar in B1 and C1=10 lm.
3038 Narendra et al.
by guest on July 13, 2014http://jxb.oxfordjournals.org/
Dow
nloaded from
with the localization pattern of the cotton pAPX (Mullenet al., 1999; Lisenbee et al., 2003a, b). Compared with theperoxisomal localization, the peroxisomal ER localizationof GFP–APX3 was observed much less frequently in thisstudy. Nevertheless, the data are consistent with thelocalization data from studies of cotton pAPX (Mullenet al., 1999, 2000; Lisenbee et al., 2003a, b). Furthermore,the rice APX3 was recently shown to be targeted toperoxisomes (Teixeira et al., 2006), which is also consis-tent with the present data.
The function of APX3 was studied by analysing anAPX3 null mutant in Arabidopsis, and the result issomewhat surprising. Based on early studies with APX3-overexpressing plants (Wang et al., 1999; Yan et al., 2003),it was thought that APX3 might play an important role inantioxidation metabolism in Arabidopsis. Yet the APX3knockout mutant performed just as well as wild-type plantsdid under normal growth conditions (Fig. 5A). Even underthe stress conditions tested (e.g. low and high temperaturesand low salt treatment), no major morphological andbiomass differences could be detected between the APX3knockout mutant and wild-type plants (Fig. 5B, C). Only
a slight genotypic difference in the protection of photo-synthesis during heat stress could be detected (Fig. 6A).The analysis with the APX3 knockout mutant implies thatAPX3 is not absolutely required for Arabidopsis growthand development.
Since the peroxisome is a major site in plant cells whereH2O2 is produced (Corpas et al., 2001; del Rio et al., 2002),one would think that APX3 would play a major role inprotecting Arabidopsis under conditions that increase pho-torespiration (consequently increasing H2O2 production).
Fig. 4. Identification of the APX3 knockout mutant. (A) Schematicdisplay of T-DNA insertion in the APX3 gene. Black boxes indicateexons, dotted boxes indicate introns, and hatched boxes indicate 59- and39-untranslated regions. The T-DNA insertion is in the seventh exon. (B)Northern blot analysis of the APX3 knockout mutant. WT, wild-type;lanes 1–3, three progeny of the original APX3 knockout mutant. Thehybridizing probes used are indicated on the right (the polyubiquitingene, PolyUb, was used as the RNA loading control). (C) Western blotanalysis of the APX3 knockout mutant. WT, wild-type; lanes 1–3, threeprogeny of the original APX3 knockout mutant. Anti-APX3 antibody wasused to probe the blot. (D) A duplicate SDS–polyacrylamide gel that wasstained with Coomassie blue was used as the protein loading control forthe western blot shown above. Each lane was loaded with 50 lg ofproteins.
Fig. 5. Characterization of the APX3 knockout mutant. (A) Wild-typeplants and APX3 knockout mutants grown under normal growthconditions. WT, wild-type; KO, APX3 knockout mutant. (B) Biomassanalysis of wild-type plants and APX3 knockout mutants grown underdifferent conditions. Normal, normal growth condition (22 8C continu-ously for 21 d); chilling, chilling growth condition (8 8C continuo-usly for 33 d); heat, heat growth condition (40 8C/3 h, 22 8C/21 h d�1 for25 d). Error bars represent the SD, n=31. (C) Seed germinationrates of wild-type plants and APX3 knockout mutant plants on saltmedia. Sterilized seeds of the wild-type and APX3 knockout mutant wereallowed to germinate on MS plates that were supplemented with50 or 100 mM NaCl. Error bars represent the SD, n=11 for KO, andn=13 for WT.
Subcellular localization and function of APX3 3039
by guest on July 13, 2014http://jxb.oxfordjournals.org/
Dow
nloaded from
The present study with the APX3 knockout mutant ap-parently does not support this assumption. For one thing,the peroxisome has considerable catalase activity that maybe sufficient under most conditions to scavenge the H2O2
produced in photorespiratory metabolism. There are threegenes in Arabidopsis that encode peroxisomal catalases(Mittler et al., 2004), and their roles in compensating forthe loss of APX3 in the APX3 knockout mutant will pro-bably be substantial. Also, perhaps the role of APX3 in the
APX3 knockout mutant is compensated by another perox-isomal membrane-bound antioxidant enzyme (i.e. APX5).However, it was noticed that the expression of APX5 ap-peared to be very low in Arabidopsis, because the transcriptlevel of APX5 was found to be <3% of the transcript levelof APX3 (Panchuk et al., 2002, 2005). In fact, APX5 couldnot be detected in the APX3 knockout mutant using thepolyclonal anti-APX3 antibodies in western blot analysis(Fig. 4C), despite the fact that APX3 and APX5 share67% identity in amino acid sequence, which confirms thatAPX5 was expressed at a very low level. Therefore, the roleof APX5 in compensating for the function of APX3 inthe APX3 knockout mutant might be limited.
A thorough in silico analysis of APX3 expression, usingthe tool provided by the Genevestigator website (www.genevestigator.ethz.ch), showed that the expression patternsof APX3 and APX5 are distinct (Table 1), which suggeststhat they may function in different metabolic pathways.Interestingly, the expression pattern of APX4 overlapsextensively with that of APX3. APX4 encodes a proteinthat is probably targeted to the peroxisomal matrix (Mittleret al., 2004), and the transcript level of APX4 is at thesame or higher levels of APX3 (Panchuk et al., 2002, 2005),suggesting that APX4 could compensate for the loss ofAPX3 in the APX3 knockout mutant if they have redund-ant roles in peroxisomal antioxidation metabolism. Becausethe catalytic domain of APX3 probably faces the cytoplasm(Yamaguchi et al., 1995), it might play a role in scavengingH2O2 in cytoplasm. If so, the role of cytosolic APXs,especially APX1, in compensating for the function ofAPX3 in the APX3 knockout mutant should not be ignored.After all, APX1 was shown to play a prominent role inantioxidation metabolism in Arabidopsis (Panchuk et al.,2002; Fryer et al., 2003; Pnueli et al., 2003; Davletovaet al., 2005).
One question that remains to be answered is whetherperoxisomal APXs are important for plants. The study withthe APX3 knockout mutant appears to suggest that APX3 isnot essential for Arabidopsis under normal and certainstress conditions. It could be argued that APX3 does playan important role in Arabidopsis, but that the presentstudy did not find the condition in which the role of APX3would be reflected. To answer the question, more experi-mental conditions need to be tried to test how APX3 knock-out mutants would behave or to create double mutants thatlack both APX3 and APX5 or APX3 and APX4, and studythe behaviour of those double mutants under normal andstress conditions, which seems to be the next logicalapproach to understand the roles of peroxisomal APXs inplant cellular metabolism.
Acknowledgements
We thank Drs L Blanton, C Haigler, and M Grimson for help in theuse of the fluorescence microscope. We thank the Arabidopsis
Fig. 6. Changes in the efficiency of electron transport in PSII (UPSII) inresponse to high temperature treatment during illumination for wild-typeplants and the APX3 knockout mutant. (A) At ambient O2 level (21%);(B) at increased O2 level (50%); (C) at reduced O2 level (2%). WT, wild-type; KO, APX3 knockout mutant. A photon flux density of 350 lmol m�2
s�1 and ambient CO2 levels were maintained for all three experiments.Leaf temperature was elevated from 25 8C to 40 8C after 20 min ofillumination. Error bars represent the SD, n=5 (A, B) and n=4 (C).
3040 Narendra et al.
by guest on July 13, 2014http://jxb.oxfordjournals.org/
Dow
nloaded from
Biological Resources Center at Ohio State University for the APX3knockout mutant and the polyubiquitin gene clone. This work wassupported by grants from the Texas Advanced Technology Program.
References
Amory AM, Ford L, Pammenter NW, Cresswell CF. 1992. Theuse of 3-amino-1,2,4-triazole to investigate the short-term effectsof oxygen toxicity on carbon assimilation by Pisum sativumseedlings. Plant, Cell and Environment 15, 655–663.
Apel K, Hirt H. 2004. Reactive oxygen species: metabolism,oxidative stress, and signal transduction. Annual Review of PlantBiology 55, 373–399.
Asada K. 1992. Ascorbate peroxidase—a hydrogen peroxide-scavenging enzyme in plants. Physiologia Plantarum 85,235–241.
Asada K. 1999. The water–water cycle in chloroplasts: scavengingof active oxygens and dissipation of excess photons. AnnualReview of Plant Physiology and Plant Molecular Biology 50,601–640.
Asada K, Takahashi M. 1987. Production and scavenging of activeoxygen in photosynthesis. In: Kyle DJ, Osmond CB, Arntzen CJ,eds. Photoinhibition. Amsterdam: Elsevier, 227–287.
Bradford MM. 1976. A rapid and sensitive method for thequantitation of microgram quantities of protein utilizing theprinciple of protein–dye binding. Analytical Biochemistry 72,248–254.
Bunkelmann JR, Trelease RN. 1996. Ascorbate peroxidase—a prominent membrane protein in oilseed glyoxysomes. PlantPhysiology 110, 589–598.
Chew O, Whelan J, Millar AH. 2003. Molecular definition ofthe ascorbate–glutathione cycle in Arabidopsis mitochondriareveals dual targeting of antioxidant defenses in plants. Journalof Biological Chemistry 278, 46869–46877.
Church GM, Gilbert W. 1984. Genomic sequencing. Proceedingsof the National Academy of Sciences, USA 81, 1991–1995.
Clough SJ, Bent AF. 1998. Floral dip: a simplified method forAgrobacterium-mediated transformation of Arabidopsis thaliana.The Plant Journal 16, 735–743.
Corpas FJ, Barroso JB, del Rio LA. 2001. Peroxisomes as a sourceof reactive oxygen species and nitric oxide signal molecules inplant cells. Trends in Plant Science 6, 145–150.
Danna CH, Bartoli CG, Sacco F, Ingala LR, Santa-Marıa GE,Guiamet JJ, Ugalde RA. 2003. Thylakoid-bound ascorbateperoxidase mutant exhibits impaired electron transport andphotosynthetic activity. Plant Physiology 132, 2116–2125.
Davletova S, Rizhsky L, Liang H, Shengqiang Z, Oliver DJ,Coutu J, Shulaev V, Schlauch K, Mittler R. 2005. Cytosolicascorbate peroxidase 1 is a central component of the reactiveoxygen gene network of Arabidopsis. The Plant Cell 17, 268–281.
Del Rıo LA, Corpas FJ, Sandalio LM, Palma JM, Gomez M,Barroso JB. 2002. Reactive oxygen species, antioxidant systemsand nitric oxide in peroxisomes. Journal of Experimental Botany53, 1255–1272.
Foyer CH, Noctor G. 2000. Oxygen processing in photosynthesis:regulation and signaling. New Phytologist 146, 359–388.
Foyer CH, Noctor G. 2005. Redox homeostasis and antioxidantsignaling: a metabolic interface between stress perception andphysiological responses. The Plant Cell 17, 1866–1875.
Fryer MJ, Ball L, Oxborough K, Karpinski S, Mullineaux PM,Baker NR. 2003. Control of ascorbate peroxidase 2 expression byhydrogen peroxide and leaf water status during excess light stressreveals a functional organisation of Arabidopsis leaves. The PlantJournal 33, 691–705.
Genty B, Briantais JM, Baker NR. 1989. The relationship betweenthe quantum yield of photosynthetic electron transport andquenching of chlorophyll fluorescence. Biochimica et BiophysicaActa 990, 87–90.
Ishikawa T, Yoshimura K, Sakai K, Tamoi M, Takeda T,Shigeoka S. 1998. Molecular characterization and physiologicalrole of a glyoxysome-bound ascorbate peroxidase from spinach.Plant and Cell Physiology 39, 23–34.
Jefferson RA, Kavanaugh TA, Bevan MW. 1987. GUS fusion: b-glucuronidase as a sensitive and versatile gene fusion marker inhigher plants. EMBO Journal 6, 3901–3907.
Lin Y, Cluette-Brown JE, Goodman HM. 2004. The peroxisomedeficient Arabidopsis mutant sse1 exhibits impaired fatty acidsynthesis. Plant Physiology 135, 814–827.
Lisenbee CS, Heinze M, Trelease RN. 2003a. Peroxisomalascorbate peroxidase resides within a subdomain of roughendoplasmic reticulum in wild-type Arabidopsis cells. PlantPhysiology 132, 870–882.
Lisenbee CS, Karnik SK, Trelease RN. 2003b. Overexpression andmislocalization of a tail-anchored GFP redefines the identity ofperoxisomal ER. Traffic 4, 491–501.
Table 1. In silico analysis of the expression levels of APX3, APX4, and APX5 under various stress conditions
Data were compiled from the GENEVESTIGATOR database (www.genevestigator.ethz.ch).
Stress condition APX3 (induced) APX3 (repressed) APX4 (induced) APX4 (repressed) APX5 (induced) APX5 (repressed)
Biotic (A. tumefaciens) Yes Yes YesBiotic (nematode) Yes No effect No effect YesCO2 (high) Yes Yes No effect No effectCycloheximide Yes Yes No effect No effectOzone Yes Yes YesHormone zeatin No effect No effect No effect No effect YesNutrient glucose/sucrose Yes Yes YesAnoxia Yes Yes No effect No effectCold Yes Yes No effect No effectHypoxia Yes Yes YesOsmotic Yes Yes No effect No effectSalt Yes Yes No effect No effectWounding Yes Yes No effect No effectOxidative No effect No effect No effect No effect No effect No effectDrought No effect No effect No effect No effect YesHeat No effect No effect Yes YesUV-B No effect No effect Yes No effect No effect
Subcellular localization and function of APX3 3041
by guest on July 13, 2014http://jxb.oxfordjournals.org/
Dow
nloaded from

Mittler R. 2002. Oxidative stress, antioxidants and stress tolerance.Trends in Plant Science 7, 405–410.
Mittler R, Vanderauwera S, Gollery M, Van Breusegem F.2004. Reactive oxygen gene network of plants. Trends in PlantScience 9, 490–498.
Mullen RT, Lisenbee CS, Miernyk JA, Trelease RN. 1999.Peroxisomal membrane ascorbate peroxidase is sorted to a mem-branous network that resembles a subdomain of the endoplasmicreticulum. The Plant Cell 11, 2167–2186.
Mullen RT, Trelease RN. 2000. The sorting signals for peroxisomalmembrane-bound ascorbate peroxidase are within its C-terminaltail. Journal of Biological Chemistry 275, 16337–16344.
Mullen RT, Flynn CR, Trelease RN. 2001. How are peroxisomesformed? The role of the endoplasmic reticulum and peroxins.Trends in Plant Science 6, 256–261.
Murgia I, Tarantino D, Vannini C, Bracale M, Carravieri S,Soave C. 2004. Arabidopsis thaliana plants overexpressingthylakoidal ascorbate peroxidase show increased resistance toparaquat-induced photo-oxidative stress and to nitric oxide-induced cell death. The Plant Journal 38, 940–953.
Narendra S. 2005. The subcellular localization and function ofAPX3 in Arabidopsis. PhD thesis, Texas Tech University,Lubbock, TX, USA.
Nito K, Yamaguchi K, Kondo M, Hayashi M, Nishimura M.2001. Pumpkin peroxisomal ascorbate peroxidase is localized onperoxisomal membranes and unknown membranous structures.Plant and Cell Physiology 42, 20–27.
Noctor G, Foyer CH. 1998. Ascorbate and glutathione: keepingactive oxygen under control. Annual Review of Plant Physiologyand Plant Molecular Biology 49, 249–279.
Panchuk II, Volkov RA, Schoffl F. 2002. Heat stress- and heatshock transcription factor-dependent expression and activity ofascorbate peroxidase in Arabidopsis. Plant Physiology 129,838–853.
Panchuk II, Zentgraf U, Volkov RA. 2005. Expression of theApx gene family during senescence of Arabidopsis thaliana.Planta 222, 926–932.
Pnueli L, Liang H, Rozenberg M, Mittler R. 2003. Growthsuppression, altered stomatal responses, and augmented inductionof heat shock proteins in cytosolic ascorbate peroxidase (Apx1)-deficient Arabidopsis plants. The Plant Journal 34, 187–203.
Schreiber U, Schliwa U, Bilger W. 1986. Continuous recording ofphotochemical and non-photochemical chlorophyll fluorescencequenching with new modulation fluorometer. PhotosynthesisResearch 10, 51–62.
Sheen J. 2002. A transient expression assay using Arabidopsismesophyll protoplasts. http://genetics.mgh.harvard.edu/sheenweb/.
Shi WM, Muramoto Y, Ueda A, Takabe T. 2001. Cloning ofperoxisomal ascorbate peroxidase gene from barley and enhancedthermotolerance by overexpressing in Arabidopsis thaliana. Gene273, 23–27.
Shigeoka S, Ishikawa T, Tamoi M, Miyagawa Y, Takeda T,Yabuta Y, Yoshimura K. 2002. Regulation and function ofascorbate peroxidase isoenzymes. Journal of Experimental Botany53, 1305–1319.
Teixeira FK, Menezes-Benavente L, Galvao VC, Margis R,Margis-Pinheiro M. 2006. Rice ascorbate peroxidase gene familyencodes functionally diverse isoforms localized in different sub-cellular compartments. Planta (in press).
Van Kooten O, Snel JFH. 1990. The use of chlorophyll fluorescencenomenclature in plant stress physiology. Photosynthesis Research25, 147–150.
Wang J, Zhang H, Allen RD. 1999. Overexpression of anArabidopsis putative peroxisomal ascorbate peroxidase gene intobacco increases protection against oxidative stress. Plant andCell Physiology 40, 725–732.
Yabuta Y, Motoki T, Yoshimura K, Takeda T, Ishikawa T,Shigeoka S. 2002. Thylakoid membrane-bound ascorbate perox-idase is a limiting factor of antioxidative systems under photo-oxidative stress. The Plant Journal 32, 915–925.
Yamaguchi K, Mori H, Nishimura M. 1995. A novel isozyme ofascorbate peroxidase localized on glyoxysomal and leaf peroxi-somal membranes in pumpkin. Plant and Cell Physiology 36,1157–1162.
Yan J, Wang J, Tissue D, Holaday AS, Allen R, Zhang H. 2003.Photosynthesis and seed production under water-deficit conditionsin transgenic tobacco plants that overexpress an Arabidopsisascorbate peroxidase gene. Crop Science 43, 1477–1483.
Zhang H, Wang J, Allen RD, Nickel U, Goodman HM. 1997.Cloning and expression of an Arabidopsis gene encoding a putativeperoxisomal ascorbate peroxidase. Plant Molecular Biology 34,967–971.
3042 Narendra et al.
by guest on July 13, 2014http://jxb.oxfordjournals.org/
Dow
nloaded from




















