
tesis - idUS - Universidad de Sevilla
458
TESIS APLICACI ´ ON DE MODELOS DE REMODELACI ´ ON ´ OSEA INTERNA AL ESTUDIO DEL COMPORTAMIENTO DE IMPLANTES DENTALES por FRANCISCO JAVIER MART ´ INEZ REINA Ingeniero Industrial por la E.S.I. de Sevilla presentada en la ESCUELA SUPERIOR DE INGENIEROS INDUSTRIALES de la UNIVERSIDAD DE SEVILLA para la obtenci´ on del Grado de Doctor Ingeniero Industrial Sevilla, octubre de 2006
-
Upload
khangminh22 -
Category
Documents
-
view
0 -
download
0
Transcript of tesis - idUS - Universidad de Sevilla

TESIS
APLICACION DE MODELOS DE
REMODELACION OSEA INTERNA
AL ESTUDIO DEL COMPORTAMIENTO
DE IMPLANTES DENTALES
por
FRANCISCO JAVIER MARTINEZ REINA
Ingeniero Industrial por la E.S.I. de Sevilla
presentada en la
ESCUELA SUPERIOR DE
INGENIEROS INDUSTRIALES
de la
UNIVERSIDAD DE SEVILLA
para la obtencion del
Grado de Doctor Ingeniero Industrial
Sevilla, octubre de 2006


Agradecimientos
Estos agradecimientos no estan escritos siguiendo ningun orden de importancia. No ob-
stante lo anterior, el primero a quien quiero darle las gracias es a Dios. Dicho esto, si lees
tu nombre aquı es porque has hecho algo que me ha ayudado a acabar el trabajo, pero
no te preguntes si tu ayuda tiene mas valor que la del que esta unas lıneas mas arriba.
Simplemente lo tiene. Si no estas aquı, puede que me haya olvidado. Lo siento en ese caso.
En primer lugar, me gustarıa agradecer el apoyo de mis dos directores de tesis, por sus
consejos cientıficos, siempre utiles y pertinentes y por estar disponibles en todo momento
para solucionar cualquier problema. Tambien tengo que agradecerle su apoyo a mis com-
paneros del departamento y en especial a mis companeros de despacho, Merche y Carlos,
por soportar la musica que pongo y por soportarme a mı. A Alfredo quiero darles las gracias
por su inestimable y paciente ayuda con el Latex y otras cuestiones tecnicas.
A todos los miembros del Grupo de Mecanica de los Medios Continuos y Teorıa de
Estructuras del Departamento de Ingenierıa Mecanica de la Universidad de Zaragoza y en
particular a su jefe, D. Manuel Doblare. A todos ellos les agradezco su calida acogida, su
apoyo cientıfico, informatico y sobre todo moral.
I would also like to thank Dr. Heinz Pettermann from the University of Wien, for his
support during my stay in the Austrian Research Center Seibersdorf, and later, during his
visits to Seville.
A mi amigo Fernando. Nos hemos sobrepuesto juntos a las adversidades y al final lo
hemos conseguido. Enhorabuena tambien a ti.
A mis amigos (Alvarito, Balleto, Curro, Emilio, Gnomo, Juan, Julio, Lagui, Mario, Rojas,
Ignacio, Fernando) que ultimamente no me han visto mucho, pero que me saben comprender
y me apoyan.
A toda mi familia, que me ha soportado el mal humor que la tesis me causaba a veces.

4
A mis padres Carmen y Antonio; a mis hermanos Aguas-Santas y Antonio; a mis sobrinos
Manuel, Antonio y Marıa; a mis suegros Manoli y Pepe; cunados, tıos y primos.
Buena prueba de que estos agradecimientos no estan dispuestos por orden de importancia
es el hecho de que mi mujer venga la ultima. Gracias, Marina, por tu constante apoyo en
los momentos malos. Sin ti no hubiera podido acabar esto. Y por cierto, enhorabuena.
Sevilla, 6 de octubre de 2006

Indice general
1. Introduccion 1
1.1. ¿Protesis extraıble o fijas? . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2
1.2. Relacion entre estado tensional y reabsorcion osea . . . . . . . . . . . . . . . 5
1.3. Estado del arte de la aplicacion del MEF al estudio de implantes dentales . . 6
1.3.1. Modelos de mandıbula completa o de rebanada . . . . . . . . . . . . . 7
1.3.2. Aplicacion de cargas y condiciones de apoyo . . . . . . . . . . . . . . . 10
1.3.3. Compatibilidad de desplazamientos en la union implante-hueso. Grado
de osteointegracion . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 11
1.3.4. Propiedades mecanicas del hueso . . . . . . . . . . . . . . . . . . . . . 12
1.3.5. Remodelacion osea. ¿Por que se utiliza en este estudio? . . . . . . . . 13
1.4. Objetivos y descripcion de la tesis . . . . . . . . . . . . . . . . . . . . . . . . 14
2. Anatomıa funcional de la mandıbula 17
2.1. Introduccion . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 17
2.2. Huesos del craneo . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 18
2.3. Mandıbula . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 20
2.4. Articulacion temporomandibular . . . . . . . . . . . . . . . . . . . . . . . . . 21
2.5. Musculos que intervienen en la masticacion . . . . . . . . . . . . . . . . . . . 26
2.5.1. Musculos masticatorios . . . . . . . . . . . . . . . . . . . . . . . . . . 26
2.5.2. Otros musculos que intervienen en la masticacion . . . . . . . . . . . . 33
2.6. Movimientos libres de la mandıbula . . . . . . . . . . . . . . . . . . . . . . . . 36
2.6.1. Cierre y apertura . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 37
2.6.2. Protrusion y retrusion . . . . . . . . . . . . . . . . . . . . . . . . . . . 38
2.6.3. Desviacion lateral . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 39
i

II INDICE GENERAL
2.7. Movimientos masticatorios de la mandıbula . . . . . . . . . . . . . . . . . . . 40
2.7.1. Incision . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 40
2.7.2. Masticacion . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 41
2.8. Fuerzas desarrolladas por los musculos masticatorios . . . . . . . . . . . . . . 44
3. Modelo de EF de una mandıbula humana con protesis dental implantoso-
portada 47
3.1. Introduccion . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 47
3.2. Modelo de EF de la mandıbula con denticion completa . . . . . . . . . . . . . 48
3.3. Simulacion de la actividad masticatoria . . . . . . . . . . . . . . . . . . . . . 52
3.3.1. Simplificaciones realizadas para simular la actividad masticatoria . . . 53
3.3.2. Secuencias y patrones de masticacion . . . . . . . . . . . . . . . . . . . 57
3.4. Modelo implantado . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 58
3.5. Osteointegracion. Contacto en la interfase implante-hueso . . . . . . . . . . . 62
3.5.1. Aspectos biologicos de la osteointegracion . . . . . . . . . . . . . . . . 62
3.5.2. Simulacion numerica de la osteointegracion . . . . . . . . . . . . . . . 65
3.6. Submodelo . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 68
3.7. Secuencias de masticacion en el modelo implantado . . . . . . . . . . . . . . . 70
4. Biologıa osea 73
4.1. Introduccion . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 73
4.2. Composicion del hueso . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 74
4.2.1. Matriz osea . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 74
4.2.2. Descripcion cuantitativa de la composicion de la matriz osea . . . . . 75
4.2.3. Celulas oseas . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 76
4.3. Tipos de tejido oseo . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 80
4.3.1. Trabecular y cortical . . . . . . . . . . . . . . . . . . . . . . . . . . . . 80
4.3.2. Lamelar y fibroso . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 83
4.3.3. Primario y secundario . . . . . . . . . . . . . . . . . . . . . . . . . . . 84
4.4. Estructura del hueso . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 86
4.5. Remodelacion osea . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 88
4.5.1. Remodelacion osea externa . . . . . . . . . . . . . . . . . . . . . . . . 89
4.5.2. Remodelacion osea interna . . . . . . . . . . . . . . . . . . . . . . . . 90

INDICE GENERAL III
4.6. Propiedades mecanicas del hueso . . . . . . . . . . . . . . . . . . . . . . . . . 102
4.6.1. Aspectos generales . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 102
4.6.2. Influencia del contenido de mineral . . . . . . . . . . . . . . . . . . . . 106
4.6.3. Influencia de la porosidad . . . . . . . . . . . . . . . . . . . . . . . . . 107
4.6.4. Influencia de la microestructura . . . . . . . . . . . . . . . . . . . . . . 108
4.6.5. Fabric tensor . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 112
5. Modelos de remodelacion osea interna 117
5.1. Introduccion . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 117
5.2. Revision historica de los modelos de remodelacion osea interna . . . . . . . . 118
5.2.1. Primeros conceptos. Ley de Wolff . . . . . . . . . . . . . . . . . . . . . 118
5.2.2. Primeros modelos matematicos. Pauwels y Kummer . . . . . . . . . . 121
5.2.3. Teorıa de la elasticidad adaptativa de Cowin . . . . . . . . . . . . . . 123
5.2.4. Carter: el concepto de auto-optimizacion . . . . . . . . . . . . . . . . . 127
5.2.5. El modelo isotropo de Huiskes . . . . . . . . . . . . . . . . . . . . . . 128
5.2.6. Modelo de remodelacion osea interna isotropa de Stanford . . . . . . . 130
5.3. Modelos de remodelacion osea anisotropa . . . . . . . . . . . . . . . . . . . . 140
5.3.1. Modelo de remodelacion osea anisotropa de Stanford basado en la
energıa . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 141
5.3.2. Modelo de remodelacion osea anisotropa de Stanford basado en las
tensiones . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 147
5.3.3. Modelo de remodelacion osea anisotropa basado en la Mecanica del
dano . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 151
5.4. Otros modelos posteriores . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 159
5.4.1. Modelo de remodelacion basado en la optimizacion global de la estruc-
tura osea de Bagge . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 160
5.4.2. Modelo de las tensiones locales uniformes de Tsubota, Adachi y Tomita166
5.4.3. Modelo de remodelacion basado en la optimizacion global de la estruc-
tura osea de Rodrigues y Fernandes . . . . . . . . . . . . . . . . . . . 169
6. Obtencion de las propiedades mecanicas de una mandıbula humana 175
6.1. Introduccion . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 175
6.2. Distribucion de tensiones inicial . . . . . . . . . . . . . . . . . . . . . . . . . . 176

IV INDICE GENERAL
6.3. Distribucion de densidad osea y propiedades elasticas . . . . . . . . . . . . . . 181
6.3.1. Introduccion y resultados experimentales de referencia . . . . . . . . . 181
6.3.2. Simulacion de referencia . . . . . . . . . . . . . . . . . . . . . . . . . . 182
6.3.3. Influencia del patron de masticacion . . . . . . . . . . . . . . . . . . . 198
6.3.4. Influencia del parametro de anisotropıa ω . . . . . . . . . . . . . . . . 201
6.3.5. Influencia del estımulo de equilibrio . . . . . . . . . . . . . . . . . . . 204
6.3.6. Discusion de los resultados . . . . . . . . . . . . . . . . . . . . . . . . 208
7. Modelos mecanobiologicos 211
7.1. Introduccion . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 211
7.2. Revision de algunos modelos de remodelacion mecanobiologicos . . . . . . . . 212
7.2.1. Modelo de adaptacion osea de hueso esponjoso de Huiskes et al. . . . . 212
7.2.2. Modelo mecanobiologico de BMUs de Hernandez . . . . . . . . . . . . 216
7.2.3. Modelo de BMUs con activacion por desuso y dano de
Hazelwood . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 219
7.2.4. Modelo de fallo y reparacion probabilista de Taylor . . . . . . . . . . . 223
7.3. Modelo isotropo de remodelacion osea basado en la actividad de las BMUs . 227
7.3.1. Introduccion . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 227
7.3.2. Descripcion matematica de la actividad de las BMUs . . . . . . . . . . 230
7.3.3. Balance de masa osea . . . . . . . . . . . . . . . . . . . . . . . . . . . 234
7.3.4. Activacion de BMUs . . . . . . . . . . . . . . . . . . . . . . . . . . . . 239
7.4. Modelo anisotropo de BMUs . . . . . . . . . . . . . . . . . . . . . . . . . . . 241
7.4.1. Introduccion . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 241
7.4.2. Extension al caso anisotropo de las variables basicas . . . . . . . . . . 242
7.4.3. Actividad de las hemiosteonas. Variacion del tensor MIL en el hueso
trabecular . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 251
7.4.4. Actividad de las osteonas. Variacion del tensor MIL en el hueso cortical260
7.4.5. Balance local direccional de hueso y principio de acomodacion celular 263
7.4.6. Evolucion del dano en el caso anisotropo . . . . . . . . . . . . . . . . . 268
7.4.7. Propiedades elasticas del hueso . . . . . . . . . . . . . . . . . . . . . . 273
7.5. Aplicacion del modelo propuesto a algunos casos sencillos . . . . . . . . . . . 278
7.5.1. Simulacion isotropa . . . . . . . . . . . . . . . . . . . . . . . . . . . . 279
7.5.2. Simulacion anisotropa . . . . . . . . . . . . . . . . . . . . . . . . . . . 295

INDICE GENERAL V
7.5.3. Cambio de anisotropıa . . . . . . . . . . . . . . . . . . . . . . . . . . . 305
8. Analisis de implantes dentales 321
8.1. Introduccion . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 321
8.2. Implantes completamente osteointegrados . . . . . . . . . . . . . . . . . . . . 325
8.2.1. Secuencia S4. Densidad inicial 2.0g/cm3 en el hueso periimplantario 327
8.2.2. Secuencia S4. Densidad inicial 1.7g/cm3 en el hueso periimplantario 353
8.2.3. Secuencia S5. Densidad inicial 2.0g/cm3 en el hueso periimplantario 361
8.3. Influencia de la edad . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 368
8.4. Implantes parcialmente osteointegrados . . . . . . . . . . . . . . . . . . . . . 377
8.5. Resumen de los resultados . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 390
9. Conclusiones y desarrollos futuros 393
9.1. Resumen del trabajo realizado . . . . . . . . . . . . . . . . . . . . . . . . . . 393
9.2. Conclusiones . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 394
9.3. Aportaciones originales . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 407
9.4. Desarrollos futuros . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 408
A. Modelo de Stanford basado en la energıa: Revision. 413


Capıtulo 1
Introduccion
Que la esperanza de vida aumenta cada dıa es un hecho evidente, al menos en el mundo
desarrollado. En los Estados Unidos, claro exponente de esta porcion del mundo, el numero
de personas mayores de 65 anos representaban un 4 % de la poblacion total en 1900; a
mediados del siglo pasado ese porcentaje era del 8.1 %; actualmente es aproximadamente
del 13 % y a mediados de este siglo se preve que alcance el 21.8 % [238]. En Espana, segun
datos del INE, el 5.2 % de la poblacion tenıa mas de 65 anos en 1900, el 7.2 % en 1950,
actualmente el 15 % y se espera que en el 2030 ese porcentaje sea el 19.6 % [86]. Este hecho
tiene implicaciones muy importantes en economıa, polıtica, sociologıa, medicina y en la
tecnica en general. La sociedad actual demanda no solo vivir hasta una edad mas avanzada,
sino que ademas lo hagamos disfrutando de una mayor calidad de vida. La medicina debe
proporcionar ese bienestar razonable durante los anos de mas que ya nos proporciona con
sus avances. La ingenierıa tiene tambien mucho que aportar a este fin, como complemento
de la medicina. Son cada vez mas numerosas las ocasiones en que los medicos hacen uso
de herramientas y tecnicas experimentales y numericas mas familiares para los ingenieros.
En otras ocasiones, ambos trabajan conjuntamente, en estudios que se encuentran a medio
camino entre las dos disciplinas y que se incluyen en una de reciente creacion y enorme
futuro, que se ha dado en llamar Bioingenierıa o Ingenierıa Biomedica.
Uno de los campos donde la Bioingenierıa esta teniendo mayor relevancia es en el diseno
de protesis. La gran mayorıa de metodos que se emplean en esta tesis surgieron para dar
respuesta a este problema. En concreto, para mejorar las protesis de cadera usadas en
fracturas de la cabeza del femur. La osteoporosis, patologıa que esta detras de la mayorıa
1

2 Introduccion
de las fracturas de este tipo, se ve agravada, entre otros factores, por la edad, que tambien
influye de manera crucial en la perdida de dientes, problema que se pretende analizar aquı.
Si se hace caso de las previsiones anteriores, no es descabellado pensar que, en los proximos
anos, la implantacion de protesis dentales puede ser demandada al mismo nivel que otros
servicios que hoy dıa parecen basicos. Esta creciente necesidad de servicios relacionados con
la implantologıa no solo se debe al aumento de la esperanza de vida. Otros factores, esteticos,
psicologicos, de comodidad, etc. se pueden combinar con este y deben ser tenidos en cuenta
a la hora de elegir el tipo de protesis dental mas conveniente.
1.1. ¿Protesis extraıble o fijas?
Las protesis dentales que se utilizan en la actualidad en casos de edentulismo parcial o
total, se pueden clasificar en tres tipos segun el lugar donde se apoyan:
a) Mucosoportadas: se apoyan directamente sobre la mucosa (encıas y paladar).
b) Dentosoportadas: se apoyan directamente en dientes propios del paciente. En este caso
solo se aplican a pacientes parcialmente edentulos.
c) Implantosoportadas: se apoyan sobre implantes.
A veces, el apoyo no ocurre estrictamente sobre una zona y se habla entonces de protesis
mucodentosoportadas, mucoimplantosoportadas y dentoimplantosoportadas. Todas aquellas
que se apoyen sobre uno o mas implantes requieren de una intervencion quirurgica para
introducir dicho implante en el hueso maxilar y se basan en el fenomeno conocido como
osteointegracion.
Fue el Dr. Per-Ingvar Branemark quien acuno dicho termino en protesis dentales [38].
Segun el glosario de la AAID [3], osteointegracion es un contacto establecido entre el hueso
normal y remodelado y la superficie de un implante sin la interposicion de tejido conjuntivo
o no oseo. Conseguir una buena osteointegracion es primordial para asegurar el exito de
un implante. Depende de diversos factores, tales como: el material del que esta fabricado
el implante, su tratamiento superficial, la movilidad inicial, la higiene oral, la ausencia de
carga durante un cierto perıodo, denominado de curacion, ası como la duracion del mismo,
etc. De entre todos ellos, los unicos que son parametros asociados al diseno del implante
son el material (el titanio puro y sus aleaciones son los mas utilizados) y el tratamiento
superficial. Algunos autores establecen que una superficie de implante rugosa propicia una

1.1 ¿Protesis extraıble o fijas? 3
mejor osteointegracion que una superficie lisa [51,54,68,93,185,347,353]. Esta rugosidad se
consigue mediante diversos tratamientos superficiales, de entre los cuales el recubrimiento
de hidroxiapatita depositada mediante spray de plasma y el tratamiento con chorro de arena
y posterior atacado acido son los mas habituales.
A continuacion se analizan las ventajas que presentan las implantosoportadas, en las que
se centra esta tesis, frente a los otros dos tipos. Posteriormente, se analizaran sus inconve-
nientes, a modo de introduccion del problema planteado en este estudio. Dicho problema es
la perdida de hueso crestal periimplantario que puede convertirse a largo plazo en una de
las razones de fracaso del implante.
En primer lugar, hay que destacar las graves consecuencias anatomicas que trae consigo
el edentulismo por el desuso del hueso maxilar. Solo en las protesis implantosoportadas el
maxilar vuelve a recibir carga durante la masticacion, dentro de un rango fisiologico relativa-
mente normal. La perdida de dientes provoca una reabsorcion del hueso alveolar circundante
y da lugar a la formacion de rebordes edentulos atroficos. Con ello se reduce la anchura y
la altura del hueso de soporte de la protesis. En el caso de disponer sobre la mandıbula una
protesis mucosoportada, la reduccion de hueso hace disminuir la oposicion al desplazamiento
anterior de dicha protesis durante la funcion bucal o el habla, entorpeciendolas. En el caso
de las dentosoportadas, tampoco se puede evitar dicha reduccion de hueso. Tambien puede
reabsorberse el hueso basal, lo que a largo plazo trae como consecuencia una dehiscencia
del conducto mandibular y el agujero mentoniano, que pasa a servir de apoyo de la protesis
provocando dolor agudo y/o parestesia de las zonas inervadas por el nervio mandibular. La
reabsorcion del hueso basal aumenta tambien las posibilidades de sufrir una fractura de la
mandıbula. Los cambios faciales que se producen como consecuencia del envejecimiento se
ven favorecidos por la perdida de dientes y no se frenan con el uso de protesis no implantoso-
portadas. El aspecto prognatico, la disminucion de la altura facial, el adelgazamiento de los
labios, el aumento de la papada, entre otras, son alteraciones esteticas que se producen en
mayor o menor medida en pacientes con este tipo de protesis. Los trastornos citados, junto
con la disfuncion masticatoria y el aumento de las dificultades en el habla pueden provocar
graves trastornos psicologicos que van desde la simple verguenza por llevar la protesis al
rechazo total de la misma, pudiendo convertir al paciente en un autentico lisiado dental.
Entre las ventajas que proporcionan las protesis implantosoportadas cabe destacar las
siguientes: mantenimiento del hueso, mantenimiento de las dimensiones verticales de la

4 Introduccion
mandıbula, buena oclusion y aumento de la fuerza oclusal, mejora general de la funcion mas-
ticatoria, mayor estabilidad de la protesis, mejor fonetica, mejora de la estetica, etc. [238] y
en general mayor calidad de vida.
Desafortunadamente, no todo son ventajas y aunque el porcentaje de exito de las protesis
implantadas es superior y cada dıa mayor,1 hay ocasiones en que el implante no cumple su
mision de forma adecuada y es necesaria su extraccion. Las causas de rechazo en protesis
implantadas son casi exclusivamente de caracter clınico, a diferencia de las protesis no im-
plantadas, en las que, como acabamos de ver, factores psicologicos y de comodidad inter-
vienen tambien de manera crucial. Bruggenkate et al. [42], han llevado a cabo una revision
de lo escrito en torno al exito de protesis implantosoportadas, encontrando algunos estudios
que consideran ciertos criterios subjetivos de comodidad y satisfaccion del paciente, para
establecer que un implante ha fallado. Tambien se registran complicaciones debido a proble-
mas de integridad estructural o de funcionalidad de los elementos mecanicos que componen
la protesis [127], pero estas no se van a tener en cuenta aquı, porque dichos defectos pueden
ser facilmente corregidos. La retirada del implante debida a la concurrencia de una o varias
complicaciones clınicas, que a veces son consecuencia unas de otras, sigue siendo el criterio
mas utilizado para definir el fallo o fracaso del mismo. Este fracaso debe evitarse por el grave
perjuicio, economico y fisiologico, que se le causa al paciente. El economico es claro, dado
su elevado precio en comparacion con otro tipo de restauraciones, pero aun siendo grave,
no lo es tanto como el perjuicio fisiologico, ya que la extraccion del implante requiere un
tratamiento posterior consistente en la aplicacion de injertos oseos autogenos y/o sinteticos
para reponer el hueso perdido.
Se analizan a continuacion algunas de las complicaciones clınicas que, como se ha indica-
do, pueden conducir a la retirada del implante. La fundamental para valorar el exito o fracaso
de un implante es la ausencia de dolor durante la aplicacion de fuerzas verticales u horizon-
tales. Habitualmente, el dolor esta asociado con la movilidad, aunque a veces es necesario
retirar el implante solo por el dolor que produce sin que exista movilidad. El objetivo clınico
de todos los implantes es la fijacion rıgida de la protesis y por esta razon hay que garantizar
que no exista movilidad bajo la accion de fuerzas horizontales o verticales inferiores a 500g.
1En la decada de los 80 el porcentaje de exito estaba en torno al 84 % a largo plazo (que entonces se
consideraba mas de tres anos) [5, 9, 76]. Misch, en un trabajo posterior [238], de finales de la decada de los
90, establece que la supervivencia de los implantes debe valorarse sobre la base de un plazo de 10 anos. Hoy
dıa ese plazo ha sido ampliamente superado.

1.2 Relacion entre estado tensional y reabsorcion osea 5
Esto no garantiza la existencia de una interfase directa (osteointegracion) entre implante y
hueso en toda su longitud, aunque sı al menos en una parte. Steflik et al. [311] observaron
que la movilidad estaba relacionada con la presencia de un tejido conjuntivo fibroso alrede-
dor del cuerpo del implante, es decir, a la falta de osteointegracion. Los implantes con menos
de 0.5mm de desplazamiento horizontal pueden recuperar la rigidez de fijacion y la movil-
idad cero si se tratan adecuadamente y se retira temporalmente la carga, siendo mayores
las posibilidades de mejora si no se observo movilidad inicial [238]. Un implante con mas
de 1mm de movilidad horizontal o cualquier grado de movilidad vertical no puede entrar
en funcionamiento, para evitar una perdida continuada de hueso y el consecuente deterioro
del emplazamiento del implante. La perdida de hueso crestal, conducente a otros tipos de
lesiones periodontales, como la periimplantitis, que a su vez contribuye a acelerar la perdida
osea, o la gingivitis, es la causa mas comun del fracaso de implantes [264]. Esto se debe a
que dicha perdida osea provoca no solo las patologıas mencionadas, sino que habitualmente
esta detras de las otras causas que se han citado aquı: el dolor y la movilidad. En efecto,
una perdida osea a nivel crestal hace disminuir la estabilidad del implante, aumentando su
movilidad y esta es la principal causa de dolor, como ya se ha indicado. Se puede decir que
es la perdida osea crestal la causa que esta detras de todo el trabajo descrito en esta tesis.
1.2. Relacion entre estado tensional y reabsorcion osea
Se establece a continuacion, de forma muy resumida, la relacion existente entre la reab-
sorcion osea crestal y el estado de tensiones que sufre el hueso periimplantario durante la
actividad masticatoria.
La capacidad del hueso para adaptarse a la carga que soporta, ha sido objeto de discusion
e investigacion durante mas de un siglo, desde que Wolff enunciara en 1892 su famosa
teorıa, segun la cual la arquitectura osea esta ıntimamente relacionada con las direcciones
principales de tension [354]. Esta teorıa, que origino gran controversia en su epoca, fue
retomada en la decada de los sesenta del siglo pasado y reformulada de forma matematica.
Esto dio origen a una serie de modelos de remodelacion osea que relacionan la densidad y
las propiedades elasticas del hueso con la historia de carga a la que esta sometido. Dichos
modelos tienen una base comun y se pueden resumir, en pocas palabras, diciendo que una
actividad por encima de lo normal hace que el hueso aumente su densidad y con ello su
rigidez, y un desuso del hueso provoca la disminucion de la masa osea con la consecuente

6 Introduccion
perdida de rigidez.
Tomando esta idea como cierta, no parecen tener mucho sentido los numerosos analisis de
elementos finitos realizados en los ultimos anos, para modelar el hueso maxilar con implantes
durante la funcion masticatoria. Estos analisis, que se comentan mas adelante, esperaban
encontrar zonas de elevada tension en el cuello del implante que explicaran la reabsorcion
osea a nivel crestal mencionada en el apartado anterior, al modo en que se hace en cualquier
analisis estructural, en el que los puntos susceptibles de fallar se encuentran generalmente en
las zonas con tensiones mas altas. Esto no estarıa de acuerdo con la teorıa de remodelacion
osea, segun la cual en las zonas de mayor actividad es previsible un aumento de la densidad.
Sin embargo, la perdida osea a largo plazo en el cuello del implante esta corroborada clınica-
mente [96, 152, 165, 204, 276]. Lo que ocurre en realidad es que el aumento de la densidad
osea con la actividad, previsto por la mayorıa de modelos de remodelacion osea tiene un
lımite fisiologico: por encima de un determinado valor, incrementos en el nivel de actividad
no llevan asociados incrementos proporcionales de densidad y por tanto de la rigidez. Mas
bien al contrario, el aumento de la tension por encima de determinados valores provoca la
aparicion, crecimiento y coalescencia de microgrietas por fatiga, que no da tiempo a reparar
mediante los mecanismos de remodelacion osea y que pueden danar de forma irreversible la
zona sobrecargada, el cuello del implante en este caso. Valores de tension altos contribuyen
al aumento de rigidez del hueso y valores demasiado altos acaban produciendo reabsorcion,
de la misma forma que lo hace el desuso. Se deduce de lo anterior, que en el diseno de
una protesis, es fundamental conseguir una transferencia de carga lo mas uniforme posible
entre la protesis y el hueso y que el nivel de tension en este sea adecuado para mantener
la densidad del tejido oseo circundante [134,135]: que no haya zonas de tension baja donde
desaparezca el hueso por desuso, ni concentradores de tension donde lo haga por sobrecarga.
1.3. Estado del arte de la aplicacion del MEF al estudio
de implantes dentales
La necesidad de conocer el estado tensional en el hueso periimplantario, discutido en el
apartado anterior, es el que ha motivado la aplicacion del metodo de los elementos finitos
(MEF) al estudio de implantes dentales en los ultimos anos. Dicho metodo se ha aplicado
de forma exhaustiva para analizar la influencia que tienen diversos factores en las tensiones

1.3 Estado del arte de la aplicacion del MEF al estudio de implantes dentales 7
transmitidas al hueso alveolar circundante. Entre ellos, los mas analizados son los siguientes:
a) Forma del implante. Es habitual la comparacion entre varios disenos comerciales [66,
89, 285,286,304] y entre los implantes roscados y los cilındricos o impactados [206,317]. En
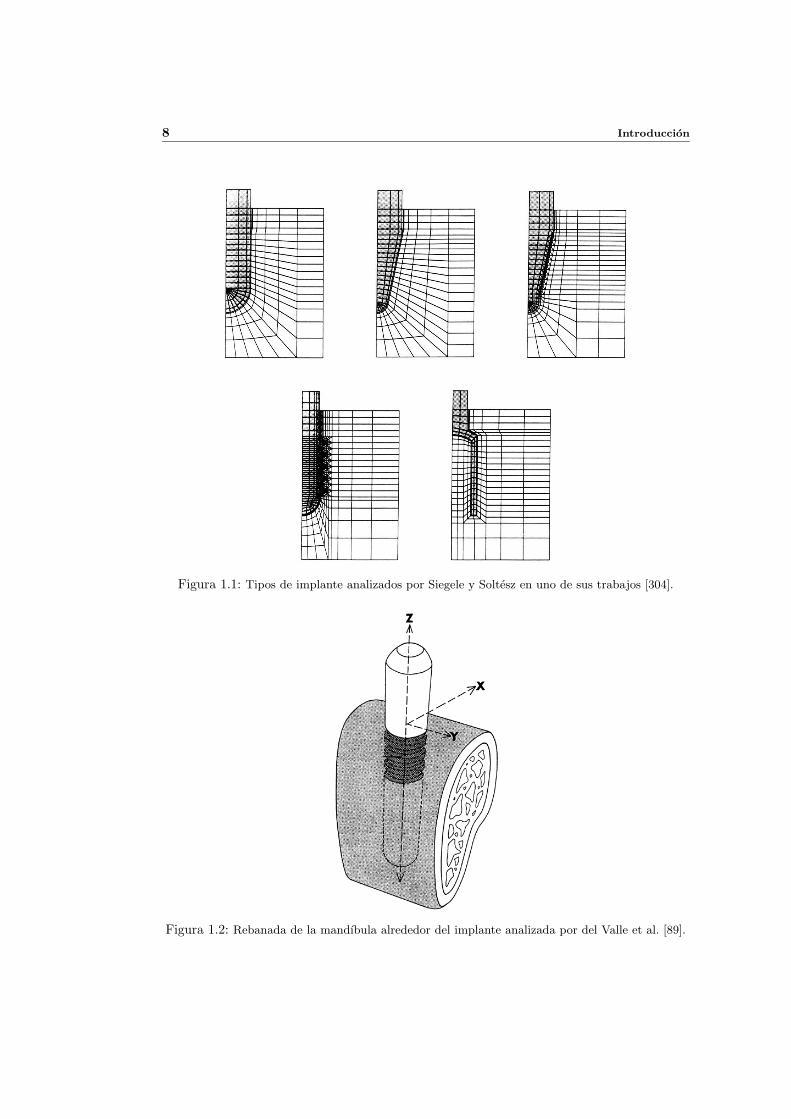
la figura 1.1 se muestran los cinco implantes analizados en uno de ellos [304].
b) Longitud y/o diametro [66,148,164,227,302,317,323].
c) Influencia de la forma de la protesis (numero y posicion de los implantes, implantes
aislados o unidos entre sı, longitud del voladizo, etc.) [187,202,227,230,302,358].
d) Recubrimiento del implante [4, 119, 229, 331], que afecta de manera muy directa a la
osteointegracion.
e) Altura de hueso disponible [141,227,259].
f) Calidad del hueso circundante [89,141,259,317,357].
g) Grado de osteointegracion. Se analiza en particular la union hueso-implante a la altura
del cuello, que depende del tipo de recubrimiento y que puede perderse como consecuencia de
las elevadas tensiones en esa zona [159,232,256,331,350]. Tambien se han realizado analisis
dinamicos y de respuesta en frecuencia para relacionar el grado de osteointegracion y la
respuesta dinamica de forma empırica, al igual que se hace en pruebas clınicas: se aplica una
percusion al implante y se relaciona el sonido que se produce con el grado de contacto en la
interfase [153,247,351].
h) Orientacion de la carga [67,118,164,231,246,302] y posicion de la misma en la protesis
[6, 60,203,227].
Como cualquier analisis numerico, todos los trabajos citados anteriormente presentan
limitaciones y simplificaciones, algunas de las cuales tienen una importancia relevante en los
resultados obtenidos. Tal es el caso de las que se analizan a continuacion y que deberıan ser
corregidas en la medida de lo posible.
1.3.1. Modelos de mandıbula completa o de rebanada
En los modelos anteriores en los que se analiza el diseno de la protesis, se suele modelar
la mandıbula completa; sin embargo, es una practica muy extendida discretizar tan solo
la region, que aquı se denomina rebanada, comprendida entre dos secciones transversales
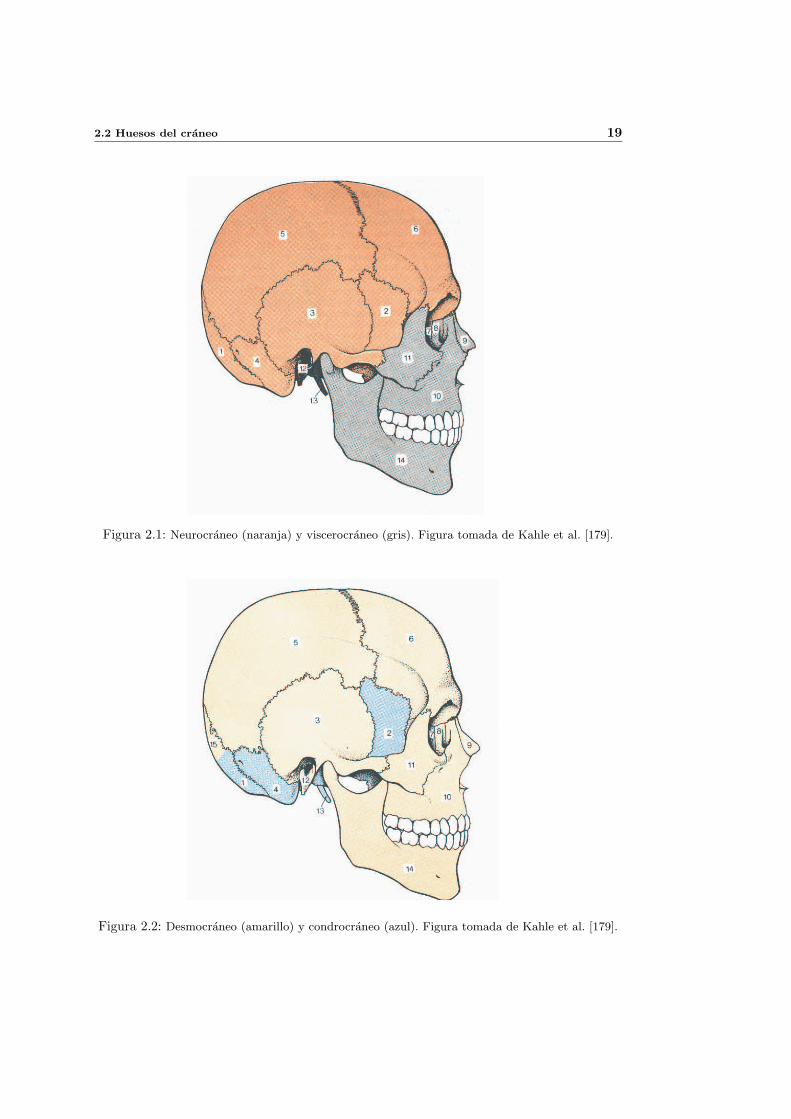
donde se coloca el implante. En la figura 1.2 se puede observar la rebanada de la mandıbula
analizada por del Valle et al. [89].
En los extremos de esta seccion se aplican unas condiciones de contorno en desplazamien-
8 Introduccion
1
Figura 1.1: Tipos de implante analizados por Siegele y Soltesz en uno de sus trabajos [304].
Figura 1.2: Rebanada de la mandıbula alrededor del implante analizada por del Valle et al. [89].

1.3 Estado del arte de la aplicacion del MEF al estudio de implantes dentales 9
tos y sobre el pilar o directamente sobre el implante se aplican las cargas de masticacion. La
validez de los resultados obtenidos con este modelo simplificado dependen en gran medida de
las condiciones de apoyo empleadas, que por lo general son de empotramiento en ambos ex-
tremos, aunque la casuıstica es bastante amplia y en algunos casos de validez muy discutible.
Por poner un ejemplo, son comunes los estudios que aprovechan la simetrıa de revolucion
de un implante concreto y construyen un modelo axisimetrico, que solo incluye una capa
de hueso concentrica al implante. Es obvia la simetrıa de revolucion de algunos implantes,
sin embargo, no existe en la mandıbula mas simetrıa que respecto al plano sinfisiario, en la
region de los incisivos: no hay ningun tipo de simetrıa en una rebanada y tampoco en las
cargas de masticacion, que justifiquen el empleo de modelos simetricos.
Tambien se pueden encontrar modelos bidimensionales de la seccion transversal de la
mandıbula. Estos pueden servir, en todo caso, como estudio preliminar, para estimar de
forma cualitativa la concentracion de tensiones en el cuello del implante, pero poco mas,
a la vista de la manifiesta tridimensionalidad del problema, que se podra comprobar en
el capıtulo de resultados. Meijer et al. [228] han comparado los resultados obtenidos con
tres modelos distintos que se pueden observar en la figura 1.3: un modelo 3D de media
mandıbula (aplicando simetrıa con respecto a la sınfisis mandibular), un modelo 3D de una
rebanada y por ultimo, un modelo 2D de una seccion de la rebanada. Comprueban que este
modelo 2D proporciona una distribucion de tensiones totalmente diferente al modelo 3D
de media mandıbula, que toman como referencia, mientras que el modelo 3D de la seccion
sı conduce a resultados similares. A la conclusion sobre la validez del modelo 3D de la
seccion, se llega despues de una comparacion no excesivamente rigurosa de los resultados.
Es muy discutible dicha validez, aunque generalmente aceptada, ya que el hecho de impedir
los desplazamientos en los extremos de la rebanada limita mucho la torsion y mucho mas la
flexion que soporta la mandıbula durante la actividad masticatoria. Un analisis bastante mas
riguroso de la distribucion de tensiones en una mandıbula completa (igualmente discutible
parece utilizar simetrıa cuando las cargas de masticacion raramente lo son), ha sido llevado
a cabo por Korioth et al. [186] y el analisis de sus resultados permite comprobar como,
efectivamente, los esfuerzos en la mandıbula son de flexion y torsion principalmente, razon
que deja serias dudas sobre la validez de los modelos de una rebanada de la mandıbula con
sus extremos empotrados. En las proximidades del implante, las tensiones obtenidas con el
modelo completo y con el modelo de rebanada seran tanto mas parecidas cuanto mas lo

10 Introduccion
sean los desplazamientos impuestos en los extremos (parece que impedirlos no es la mejor
opcion) y cuanto mas alejados esten estos extremos del implante, es decir, cuanto mayor sea
la seccion discretizada.
(a)
(b) (c)
1
Figura 1.3: Modelos de EF desarrollados por Meijer et al. [228]: (a) modelo 3D de media mandıbula;
(b) Modelo 3D de una rebanada y (c) modelo 2D de la rebanada.
1.3.2. Aplicacion de cargas y condiciones de apoyo
En cuanto a la forma de aplicar las cargas y las condiciones de apoyo, hay que reconocer
la limitacion que tienen los modelos de rebanada. Como ya se ha comentado, las condiciones
de apoyo se aplican a los extremos de la rebanada, que por lo general se empotran. Este
empotramiento no concuerda con los desplazamientos que se obtienen en las zonas de corte
y por tanto alteran los resultados. Tampoco es aceptable suponer otro tipo de apoyo sen-
cillo, distinto del empotramiento, ya que los desplazamientos de las zonas de corte son muy
particulares y dependen de las propiedades del hueso y del tipo de carga que soporte la
mandıbula.
En cuanto a las cargas, la unica posibilidad que cabe en los modelos de rebanada es

1.3 Estado del arte de la aplicacion del MEF al estudio de implantes dentales 11
aplicar las fuerzas oclusales como fuerzas externas actuando sobre la protesis o directamente
sobre el implante. En cambio, en los modelos de mandıbula completa es viable simular unas
condiciones de carga mas parecidas a la realidad [186,187,228]. En primer lugar, se restringen
los desplazamientos en la zona de apoyo de la mandıbula, la articulacion temporomandibular
(ATM), y en la zona de oclusion. En la ATM se establece el contacto entre el condilo y el
disco articular, y en la zona de oclusion, entre los dientes y el alimento. Como resultado de
ese contacto aparecen unas fuerzas, que representan, respectivamente, la reaccion en la ATM
y la fuerza con que se tritura el alimento en la zona occlusal. Estas ultimas son las que se
intentan simular en el modelo de rebanada, pero allı aplicadas como fuerzas externas. En los
modelos de mandıbula completa no es necesario hacer ninguna suposicion sobre la forma en
que estan repartidas las fuerzas de masticacion, como hacen los modelos de rebanada, que
usualmente suponen una distribucion uniforme. En los modelos de mandıbula completa, las
fuerzas de masticacion aparecen simplemente como reaccion a las fuerzas ejercidas por los
musculos masticatorios, que son las unicas aplicadas externamente.
1.3.3. Compatibilidad de desplazamientos en la union implante-
hueso. Grado de osteointegracion
El grado de osteointegracion influye de manera fundamental en el comportamiento del
implante y en particular en dos aspectos cruciales para su funcionalidad: el nivel de tensiones
que se generan en el hueso periimplantario y la resistencia de la union. Se han llevado a cabo
diversos estudios que analizan estos efectos y que comparan diferentes disenos (con y sin
recubrimiento o con distintos tipos de ellos) [229,256,331].
La principal peculiaridad de los estudios de elementos finitos que analizan la influencia
de la osteointegracion radica en las diferentes condiciones de compatibilidad en desplaza-
mientos que establecen entre los elementos de hueso e implante. Cuando se simula un caso
de osteointegracion optima, como se suele hacer en implantes con recubrimiento, se disponen
los elementos de cada material compartiendo los nodos de la cara de contacto. Sin embargo,
cuando se analizan implantes sin recubrimiento o de superficie pulida, donde la union es
mas debil, es normal suponer que en cuanto el implante entre en carga, dicha union se va a
romper, al menos donde las tensiones son mayores, esto es, en el cuello del implante. En esta
zona se aplican en ese caso condiciones de contacto unilateral estandar entre los elementos
de ambos materiales, y union perfecta en la zona donde la osteointegracion no se ha perdido.

12 Introduccion
Lo que ocurre en la interfase entre hueso e implante es mucho mas complicado que esta
simple dicotomıa entre perfectamente unido o completamente separado. Pueden aparecer
tejidos conectivos que son muy flexibles y permiten una cierta movilidad entre el hueso
y el implante, sin que haya separacion total. Ademas, de perderse la osteointegracion, lo
hara progresivamente, salvo en casos excepcionales de tipo traumatico, por lo que las condi-
ciones de la union variaran con el tiempo. En ese sentido cabe destacar los trabajos de Perez
et al. [267] sobre el deterioro de la union en implantes cementados de cadera. En cualquier
caso, una consideracion mas exhaustiva del proceso de osteointegracion requiere un analisis
complejo, que escapa a los objetivos de esta tesis, en la que se usara la simplificacion anterior:
perfectamente unido o completamente separado.
1.3.4. Propiedades mecanicas del hueso
El hueso es un material con unas propiedades mecanicas muy particulares. Es un ma-
terial heterogeneo y anisotropo, con un nivel de anisotropıa que ademas varıa de un hueso
a otro y dentro de un mismo hueso, de unas zonas a otras. Practicamente todos los huesos
del cuerpo humano presentan una capa externa de tejido oseo muy compacto, denominado
tambien cortical y un nucleo central formado por un tejido oseo mas poroso, denominado
trabecular o esponjoso. Ambos tipos de tejido tienen una microestructura radicalmente dis-
tinta, que hacen que sus caracterısticas mecanicas tambien lo sean. Sin embargo, la mayorıa
de los estudios anteriormente citados, por simplicidad, consideran el hueso como un material
isotropo y en todo caso asignan un modulo de Young distinto a cada tipo de tejido. Son
menos los que tienen en cuenta su anisotropıa [137,186,247,256] y menos aun los que asignan
unas propiedades elasticas distintas a cada zona de la mandıbula [186].
Otra de las caracterısticas del hueso es su heterogeneidad. La compleja distribucion de
poros hace completamente inviable modelar un hueso con un nivel de detalle microscopico.
A escala macroscopica se puede aplicar, sin embargo, la Mecanica de los Medios Continuos,
sin mas que promediar las propiedades a nivel microscopico para obtener las propiedades
macroscopicas. Las propiedades mecanicas a nivel macroscopico tambien se pueden obtener
mediante ensayos experimentales o incluso relacionarlas con otros parametros, como por
ejemplo la porosidad, mediante correlaciones obtenidas experimentalmente [26,84] con sufi-
ciente aproximacion. Sin embargo, a nivel mesoscopico, sigue existiendo heterogeneidad: la
porosidad y la orientacion de los poros cambian de un punto a otro. Es impensable tratar de

1.3 Estado del arte de la aplicacion del MEF al estudio de implantes dentales 13
obtener un modelo detallado de la distribucion de hueso alrededor del implante, sobre todo
atendiendo a su aplicabilidad, ya que esa distribucion es ademas caracterıstica de cada indi-
viduo. Una simplificacion casi obligada es considerar el hueso como un material homogeneo,
si acaso diferenciando entre distintas zonas de la mandıbula, y adoptar valores promedio
para diferentes individuos, provenientes de medidas experimentales que se pueden encontrar
en la literatura.
Otra particularidad del hueso, sin duda la que hace mas apasionante su estudio, es el
hecho de que se trata de un material vivo. Esto lo hace completamente distinto a todos los
demas materiales que se estudian tradicionalmente en Ingenierıa. Un material vivo adapta
sus propiedades al entorno en el que se encuentra. En concreto, el tejido oseo cambia su
masa y su organizacion interna y consecuentemente sus propiedades mecanicas en funcion
de la historia de cargas a las que se encuentra sometido. Este fenomeno de remodelacion
osea es fundamental para comprender el comportamiento del hueso a largo plazo.
En todos los estudios anteriores, las propiedades mecanicas del hueso son obtenidas
mediante medidas in vitro y permanecen invariables en el analisis a pesar de la actividad
biologica del hueso, que las puede modificar. Dicha actividad no esta contemplada, por tanto,
en tales estudios.
1.3.5. Remodelacion osea. ¿Por que se utiliza en este estudio?
La consideracion de la remodelacion osea en el modelo de comportamiento del hueso no
solo permite tener en cuenta el caracter vivo del tejido oseo, sino que ademas evita tener
que hacer algunas de las simplificaciones comentadas en los epıgrafes anteriores.
Como ya se ha indicado, el hueso es un tejido vivo que esta cambiando continuamente
su estructura interna como respuesta a una serie de senales, que pueden ser de caracter
externo o interno, tales como cambios hormonales, inmovilizacion, cargas derivadas de la
actividad del individuo, etc. A este proceso de cambio microestructural se le conoce como
remodelacion osea interna y ha sido ampliamente estudiado, con especial profusion en las
ultimas decadas. Diversos modelos matematicos, que seran objeto de un capıtulo posterior,
se han desarrollado para simularlo, permitiendo estimar las propiedades elasticas del hueso
y su dependencia de las mencionadas senales. Si esas senales pueden ser obtenidas mediante
una simulacion de elementos finitos, como ocurre por ejemplo en el caso de la carga, la
propia simulacion permite estimar los cambios estructurales. De esta forma, en el modelo

14 Introduccion
de elementos finitos, el hueso se comporta como si de un material vivo se tratara, al menos
desde el punto de vista mecanico, ya que adapta sus propiedades elasticas a la actividad a
la que se encuentra sometido.
Aprovechando esta capacidad de los modelos de remodelacion osea, diversos autores los
han utilizado para simular la formacion de la estructura interna de un determinado hueso.
Ası por ejemplo, Jacobs [166], aplica el modelo isotropo de Stanford [57] a la extremidad
proximal del femur; igualmente, Garcıa utiliza un modelo de remodelacion anisotropo [91],
basado en la mecanica del dano, en la extremidad proximal del femur y en la componente
acetabular de la pelvis [115]. En ambos casos se parte de una situacion que nada tiene que ver
con la realidad: hueso homogeneo y con una densidad osea promedio. Tras aplicar durante
un cierto tiempo las cargas normales de uso de cada hueso, se obtiene una distribucion de
masa osea razonablemente similar a la real. El modelo basado en la mecanica del dano es
capaz ademas, de predecir con bastante aproximacion la anisotropıa del hueso, partiendo de
un hueso inicialmente isotropo.
Si esto es posible hacerlo con la mandıbula, se podrıan superar dos de las principales
limitaciones, que como se comento anteriormente, tienen los estudios en que se aplica el
MEF al estudio de implantes dentales: la consideracion de la heterogeneidad del hueso a
nivel macroscopico y de unas propiedades elasticas del mismo considerado como anisotropo.
1.4. Objetivos y descripcion de la tesis
El objetivo final de esta tesis es el desarrollo de una herramienta que permita evaluar la
bondad de un determinado diseno de implante dental y/o si ese diseno es adecuado para un
determinado paciente. Lejos de alcanzar esta aun el objetivo de conocer a priori la duracion
de un implante en condiciones de osteointegracion y movilidad adecuadas, pero un pequeno
esfuerzo por conseguirlo se realiza en este trabajo.
Un objetivo previo, muy ligado al anterior puesto que es necesaria su consecucion para
conseguir aquel, es el de desarrollar un algoritmo numerico que simule los fenomenos mecanobi-
ologicos que intervienen en la remodelacion osea. Ya existen modelos que lo hacen relati-
vamente bien, pero estos, o bien no consiguen reproducir la anisotropıa del hueso de forma
correcta o bien no son capaces de simular la reabsorcion osea por sobrecarga. Ambos obje-
tivos se pretenden conseguir con el modelo propuesto.
La primera parte de la tesis tiene como objetivo discretizar una mandıbula humana, para

1.4 Objetivos y descripcion de la tesis 15
poder aplicar sobre ella las tecnicas numericas propuestas en los capıtulos siguientes.
El capıtulo 2 se dedica a describir brevemente la anatomıa funcional de la mandıbula.
Se analiza la articulacion temporomandibular y los musculos implicados en la actividad
masticatoria. Por ultimo, se estudia el proceso masticatorio: se describen los movimientos y
el papel de los musculos en cada una de las actividades que tienen lugar durante el mismo.
En el capıtulo 3 se detalla el proceso seguido para la obtencion del modelo de elementos
finitos. Se describe el proceso seguido para obtener la geometrıa y el proceso de mallado,
ası como las condiciones de contorno en carga y desplazamientos utilizadas para simular la
actividad masticatoria. Se describe el proceso seguido para la inclusion del implante ası como
las particularidades que tiene este modelo implantado.
El capıtulo 4 es una breve introduccion a la biologıa osea. Se estudiara la composicion del
tejido oseo y su estructura para continuar asimismo con una breve descripcion del proceso de
remodelacion osea. Se detalla como son las propiedades mecanicas del tejido oseo en general
y de la mandıbula en particular.
En el capıtulo 5 se realiza una revision de los modelos fenomenologicos desarrollados
hasta la fecha para simular el proceso biologico de remodelacion osea. Se describe con algo
mas de detalle el modelo anisotropo basado en la mecanica del dano, que sera el que se utilice
para obtener la distribucion inicial de densidades y propiedades mecanicas anisotropas de la
mandıbula.
Los resultados que proporcione esta primera fase del analisis se muestran en el capıtulo 6,
en el que ademas se comparan con los resultados experimentales encontrados en la literatura,
para de esta forma validar la aplicacion del modelo anisotropo basado en la mecanica del
dano.
Tras esta primera parte de la tesis, se dispone de un modelo de elementos finitos con una
estructura interna parecida a la real, que se denomina modelo completo, porque tiene todas
las piezas dentales naturales. En este modelo se sustituye el primer molar inferior derecho
por un implante dental, cuyo efecto sobre el hueso circundante es uno de los objetivos a
analizar en la segunda parte de esta tesis. El modelo implantado se denomina submodelo,
porque no incluye toda la mandıbula sino solo la region en torno al implante. Como se
obtiene este submodelo se explica en el capıtulo 3, dedicado al modelo de elementos finitos.
En el capıtulo 7 se hace una revision de los modelos mecanobiologicos de remodelacion
osea interna y se propone un nuevo modelo. En los modelos fenomenologicos se pierde, en

16 Introduccion
cierta medida, la vision biologica del problema. Una causa de caracter puramente mecanico,
como es la carga, produce un efecto biologico de creacion o desaparicion de tejido y reorde-
nacion del mismo. Este efecto se cuantifica desde un punto de vista mecanico, mediante la
variacion de las propiedades elasticas, sin que se consideren los mecanismos biologicos que lo
provocan, solo la relacion causa mecanica - efecto mecanico. En los modelos mecanobiologi-
cos, dichos mecanismos sı estan presentes y cuantificados de forma matematica, sin perder
de vista su trasfondo biologico. El modelo propuesto es capaz ademas de simular el proceso
de remodelacion osea interna en algunas situaciones en las que los modelos fenomenologicos
no funcionan y una de ellas, ya se ha comentado en este epıgrafe, es la reabsorcion osea por
sobrecarga en el cuello del implante.
En el capıtulo 8 se muestran los resultados de la aplicacion del modelo mecanobiologico
de remodelacion osea interna al submodelo implantado. Se comentan y discuten los resul-
tados de: perdida osea a nivel crestal, acumulacion de dano microestructural en el hueso
periimplantario, movilidad del implante, rigidez del tejido a largo plazo y grado de min-
eralizacion promedio del hueso circundante. Asimismo, se estudia la influencia de diversos
parametros en dichos resultados: la edad del paciente, el grado de osteointegracion a nivel
crestal, la densidad inicial del hueso en el que se implanta la protesis y el efecto que tiene
no masticar con el lado del implante despues de la operacion.
Por ultimo, en el capıtulo 9 se extraen una serie de conclusiones y se comenta la aplicacion
de las aportaciones originales de esta tesis a un posible desarrollo futuro dentro de este
campo.

Capıtulo 2
Anatomıa funcional de la
mandıbula
2.1. Introduccion
En este capıtulo se hace una somera descripcion anatomica de la mandıbula humana.
No se pretende alcanzar el detalle con el que lo harıa un anatomista, ya que el interes no se
centra en la descripcion misma de la mandıbula, sino en los musculos, huesos, organos, etc.
que intervienen en la masticacion. Sin sacrificio del rigor, se describe de una forma sencilla
como se produce la masticacion. De ahı el nombre que recibe el capıtulo.
El objetivo ultimo de este interes por entender la funcion de la mandıbula es la construc-
cion de un modelo de elementos finitos de la misma. Con dicho modelo se intentara estimar el
estado de tensiones que se produce en el hueso que la forma y para ello es necesario conocer
su geometrıa, las cargas y las condiciones de contorno del problema mecanico asociado. A
estos tres aspectos del problema pretende dar respuesta este capıtulo. La geometrıa esta di-
rectamente relacionada con la forma del hueso maxilar inferior o mandıbula; las cargas,
con los musculos masticatorios y las condiciones de apoyo, con los movimientos mastica-
torios y con la articulacion que une la mandıbula al hueso temporal, llamada articulacion
temporomandibular (ATM).
En primer lugar, se hace una descripcion general de los huesos de la cabeza, necesaria
para situar posteriormente las inserciones de algunos musculos masticatorios. Entre este
17

18 Anatomıa funcional de la mandıbula
grupo de huesos se dedica un apartado especial al maxilar inferior y la arcada dentaria, que
aunque no es propiamente un hueso se describe junto a la mandıbula. A continuacion, y
siguiendo con los huesos, se describe la ATM: los cuerpos que se articulan y los ligamentos
que controlan los movimientos relativos dentro de la articulacion. Sigue un apartado dedicado
a la descripcion de los musculos que se insertan en la mandıbula, de los cuales reciben un
tratamiento mas extenso los llamados musculos masticatorios. Por ultimo se habla de la
accion de estos musculos en los movimientos libres de la mandıbula y en los movimientos
masticatorios: como son estos movimientos, que musculos entran en juego en cada uno de
ellos y que fuerzas ejercen.
2.2. Huesos del craneo
El craneo constituye el esqueleto oseo de la cabeza, alberga el encefalo y los organos de
los sentidos, da soporte estructural a la cara y contiene los tramos iniciales de los tractos
gastrointestinal y respiratorio.
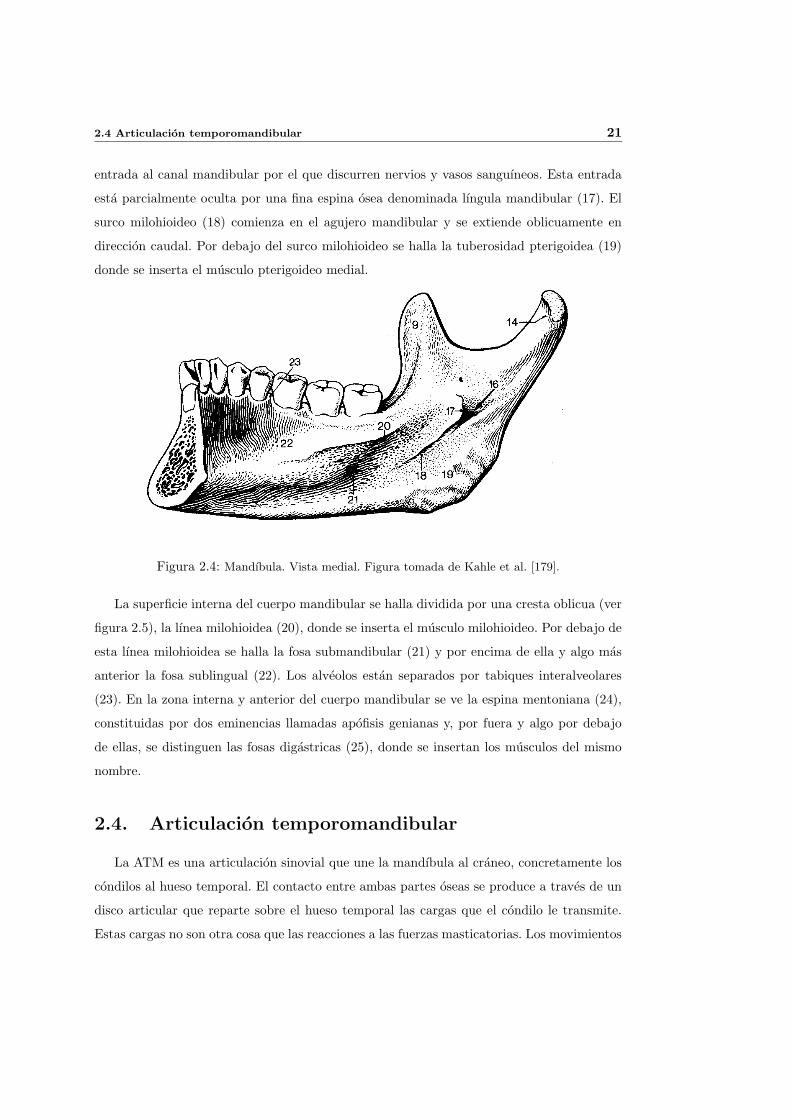
El craneo puede dividirse en neurocraneo y viscerocraneo. El primero, protege al encefalo
y se representa de color naranja en la figura 2.1. Comprende los siguientes huesos: el occip-
ital (1), los esfenoides (2), porcion escamosa (3) y porcion petrosa o mastoidea (4) de los
temporales, los parietales (5) y el frontal (6). El viscerocraneo, de color gris en la misma
figura, corresponde al esqueleto facial y comprende: los cornetes inferiores, el vomer, el hue-
so incisivo y los palatinos, que no aparecen en la vista lateral del craneo; el etmoides (7),
el lacrimal (8), el nasal (9), los maxilares superiores (10), los cigomaticos (11), la porcion
timpanica (12) y la apofisis estiloide (13) de los temporales y por fin el maxilar inferior o
mandıbula (14). Existe otro hueso en la cabeza, el hioides, que no pertenece a ninguna de
las dos partes comentadas. De hecho es el unico hueso del esqueleto que no se une a otro
hueso y tan solo sirve como insercion de diversos musculos que en su mayorıa se insertan
tambien en la mandıbula.
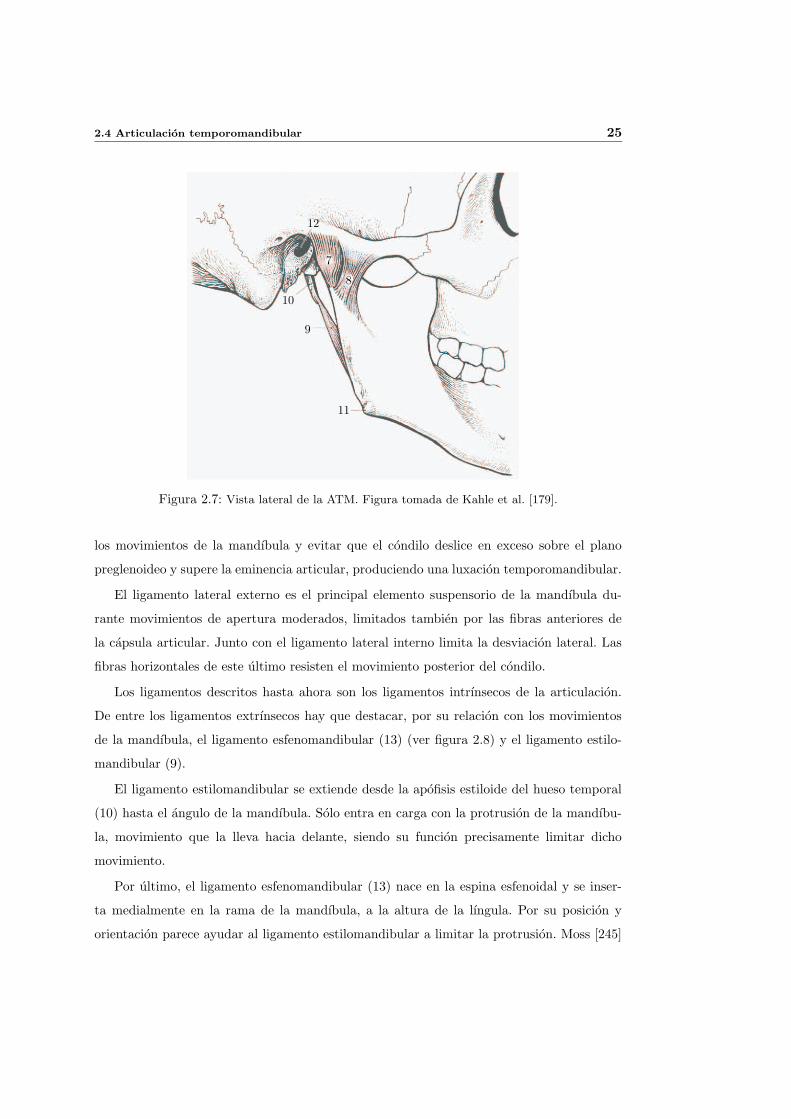
Existen fundamentalmente dos mecanismos de desarrollo del craneo que dan lugar al
condrocraneo, en azul en la figura 2.2 y al desmocraneo, en amarillo en dicha figura. En
el primer caso se produce una sustitucion de tejido cartilaginoso por tejido oseo, mientras
que en el segundo caso se producen condensaciones de tejido conjuntivo que se osifican
directamente. El maxilar inferior o mandıbula pertenece al desmocraneo y es por tanto un
hueso de origen membranoso.
2.2 Huesos del craneo 19
Figura 2.1: Neurocraneo (naranja) y viscerocraneo (gris). Figura tomada de Kahle et al. [179].
Figura 2.2: Desmocraneo (amarillo) y condrocraneo (azul). Figura tomada de Kahle et al. [179].

20 Anatomıa funcional de la mandıbula
2.3. Mandıbula
La mandıbula esta unida al resto del craneo unicamente por un par de articulaciones
sinoviales. Se origina a partir de un esbozo de tejido conectivo y en ella se distinguen un
cuerpo (1) (ver figura 2.3) y una rama ascendente (2) a cada lado. En el cuerpo de la
mandıbula del adulto puede apreciarse el proceso alveolar (3), en el que las raıces de los
dientes dan lugar a las eminencias alveolares (4). En la vejez, tras la perdida de los dientes
estos procesos alveolares sufren una regresion. En la parte anterior del cuerpo mandibular se
encuentra la protuberancia mentoniana (5) compuesta por dos prominencias pares llamadas
tuberculos mentonianos. En la cara externa, en la vertical del segundo premolar, se encuentra
el agujero mentoniano (6). La lınea oblicua (7) se extiende del cuerpo a la rama mandibular.
El angulo de la mandıbula (8) se forma en la union del cuerpo con la rama.
Figura 2.3: Mandıbula. Vista lateral. Figura tomada de Kahle et al. [179].
La rama de la mandıbula tiene dos apofisis: la apofisis coronoides (9) (ver figura 2.3) y la
apofisis condilar o condilo (10). En el condilo se aprecia un cuello (12) y una cabeza (13) con
la superficie articular. Entre ambas apofisis se extiende la incisura o escotadura mandibular
(11). En la cara interna de la cabeza del condilo, por debajo de la superficie articular,
se localiza una pequena depresion denominada fosa pterigoidea (14), donde se inserta el
musculo pteigoideo lateral. En la proximidad del angulo se identifica una superficie rugosa
conocida como tuberosidad maseterica (15) donde se inserta el musculo masetero. En la
cara interna de la mandıbula, en la rama, se situa el orificio mandibular (16), que sirve de
2.4 Articulacion temporomandibular 21
entrada al canal mandibular por el que discurren nervios y vasos sanguıneos. Esta entrada
esta parcialmente oculta por una fina espina osea denominada lıngula mandibular (17). El
surco milohioideo (18) comienza en el agujero mandibular y se extiende oblicuamente en
direccion caudal. Por debajo del surco milohioideo se halla la tuberosidad pterigoidea (19)
donde se inserta el musculo pterigoideo medial.
Figura 2.4: Mandıbula. Vista medial. Figura tomada de Kahle et al. [179].
La superficie interna del cuerpo mandibular se halla dividida por una cresta oblicua (ver
figura 2.5), la lınea milohioidea (20), donde se inserta el musculo milohioideo. Por debajo de
esta lınea milohioidea se halla la fosa submandibular (21) y por encima de ella y algo mas
anterior la fosa sublingual (22). Los alveolos estan separados por tabiques interalveolares
(23). En la zona interna y anterior del cuerpo mandibular se ve la espina mentoniana (24),
constituidas por dos eminencias llamadas apofisis genianas y, por fuera y algo por debajo
de ellas, se distinguen las fosas digastricas (25), donde se insertan los musculos del mismo
nombre.
2.4. Articulacion temporomandibular
La ATM es una articulacion sinovial que une la mandıbula al craneo, concretamente los
condilos al hueso temporal. El contacto entre ambas partes oseas se produce a traves de un
disco articular que reparte sobre el hueso temporal las cargas que el condilo le transmite.
Estas cargas no son otra cosa que las reacciones a las fuerzas masticatorias. Los movimientos

22 Anatomıa funcional de la mandıbula
Figura 2.5: Mandıbula. Vista posterior. Figura tomada de Kahle et al. [179].
relativos entre mandıbula y craneo esta limitados por una serie de ligamentos. Seguidamente
se hara una breve descripcion de las partes de la articulacion.
Disco articular
La ATM se halla dividida en dos partes por un disco articular (1) (ver figura 2.6). Las
superficies articulares son, de un lado la cabeza del condilo (2) y de otro la fosa mandibular
del hueso temporal o fosa glenoidea (3) con la eminencia articular (4).
El disco articular forma una envoltura movil para la cabeza del condilo. Tiene forma oval,
siendo su parte central considerablemente mas gruesa que su periferia. Esta directamente
unido al condilo, medial y lateralmente, en los polos del condilo, lo que le obliga a moverse
con el en la apertura y cierre de la boca.
Su porcion anterior esta formada por tejido fibroso con algunas celulas cartilaginosas
dispersas entre las fibras. La porcion posterior (5) es bilaminar. En su parte superior se fija
a la pared posterior de la fosa glenoidea y consiste en tejido laxo fibroelastico, mientras que
en la parte inferior se une al borde posterior de la cabeza del condilo y esta constituido por
tejido fibroso muy denso. Anteriormente, el disco se halla firmemente unido a la capsula
articular y a la porcion superior del musculo pterigoideo lateral en algunos individuos [233].
De esta forma la contraccion de esta porcion del musculo provoca un movimiento relativo
del disco respecto al condilo, en direccion anteromedial, que ocurre durante la apertura de

2.4 Articulacion temporomandibular 23
12
6
5
1
2
7
4
3
1
Figura 2.6: Seccion sagital de la ATM. Figura tomada de Hylander [160].
la boca.
La ausencia de irrigacion sanguınea e inervacion de la zona central del disco, ası como
de los tejidos que cubren las superficies articulares de la mandıbula y el hueso temporal, es
buena prueba de la considerable fuerza de reaccion que soporta esta zona de la articulacion.
Condilos
La superficie articular de la mandıbula comprende las caras superior y anterior de los
condilos. En un adulto el condilo tiene entre 15 y 20 mm de longitud y entre 8 y 10 mm de
ancho. Su eje largo es perpendicular a la rama mandibular y debido a la inclinacion de esta,
los ejes de los condilos izquierdo y derecho se cortan formando un angulo comprendido entre
145 y 160 grados. La superficie articular del condilo es fuertemente convexa vista desde un
lateral y no tanto vista desde frente y ademas esta visiblemente inclinada hacia delante.
Fosa glenoidea y eminencia articular
La fosa glenoidea o fosa mandibular (3) es la concavidad del temporal donde se aloja el
condilo. Su pared anterior se denomina eminencia articular (4) y su pared posterior, apofisis
postglenoidea. La parte superior de la fosa glenoidea es una pared osea muy delgada, lo cual

24 Anatomıa funcional de la mandıbula
puede indicar que no es una zona que soporte mucha carga.
La eminencia articular es la barra de hueso compacto que forma la raız posterior del arco
cigomatico y la pared anterior de la fosa mandibular. Tiene la apariencia de una silla de
montar: vista desde un lateral es convexa, mientras que vista de frente es ligeramente conca-
va, aunque el grado de convexidad y de concavidad es altamente variable de un individuo a
otro.
La pendiente anterior de la eminencia articular, denominada plano preglenoideo, de-
sciende suavemente desde la base del hueso temporal. Durante la apertura de la boca, el
condilo y el disco articular se mueven en direccion anterior por encima de la cresta de la
eminencia articular y sobre el plano preglenoideo. Cuando la boca se cierra disco y condi-
lo vuelven a alojarse en la fosa glenoidea viendose facilitado su movimiento por la suave
pendiente del citado plano.
El tejido fibroso que cubre la eminencia articular es bastante grueso y firme. Ademas,
dicha eminencia articular se caracteriza por la densidad del hueso que la compone. Estas
caracterısticas morfologicas refuerzan la hipotesis de que es en esta zona de la articulacion
donde se transmite la mayor parte de las reacciones entre la mandıbula y el hueso temporal.
Capsula articular y ligamentos
La ATM esta rodeada de una capsula articular (7) (ver figura 2.7) fibrosa reforzada en
algunas zonas por ligamentos. Superiormente se inserta en la region escamosa del temporal.
Posteriormente se inserta en la apofisis postglenoidea y en la fisura timpanoescamosa, que
une las porciones timpanica y escamosa del hueso temporal. Inferiormente se inserta en el
cuello del condilo, cerrando de esta forma la articulacion.
La capsula articular es bastante delgada anteromedial, medial y posteriormente y mas
gruesa anterolateral y lateralmente. Este refuerzo fibroso se conoce como ligamento lateral
externo o ligamento temporomandibular (8). De fibras oblicuas, se inserta en la parte pos-
terior del cuello del condilo y en el tuberculo articular, pequena protuberancia del hueso
temporal situada lateralmente respecto a la eminencia articular. Medialmente al ligamento
lateral externo se encuentra el ligamento lateral interno, otra porcion de fibras menos nu-
merosas que las anteriores y orientadas horizontalmente. Se insertan en el polo lateral del
condilo, en la parte posterior del disco y en el tuberculo articular.
La funcion principal de la capsula articular y de los dos ligamentos laterales es limitar
2.4 Articulacion temporomandibular 25
10
8
9
11
12
7
1
Figura 2.7: Vista lateral de la ATM. Figura tomada de Kahle et al. [179].
los movimientos de la mandıbula y evitar que el condilo deslice en exceso sobre el plano
preglenoideo y supere la eminencia articular, produciendo una luxacion temporomandibular.
El ligamento lateral externo es el principal elemento suspensorio de la mandıbula du-
rante movimientos de apertura moderados, limitados tambien por las fibras anteriores de
la capsula articular. Junto con el ligamento lateral interno limita la desviacion lateral. Las
fibras horizontales de este ultimo resisten el movimiento posterior del condilo.
Los ligamentos descritos hasta ahora son los ligamentos intrınsecos de la articulacion.
De entre los ligamentos extrınsecos hay que destacar, por su relacion con los movimientos
de la mandıbula, el ligamento esfenomandibular (13) (ver figura 2.8) y el ligamento estilo-
mandibular (9).
El ligamento estilomandibular se extiende desde la apofisis estiloide del hueso temporal
(10) hasta el angulo de la mandıbula. Solo entra en carga con la protrusion de la mandıbu-
la, movimiento que la lleva hacia delante, siendo su funcion precisamente limitar dicho
movimiento.
Por ultimo, el ligamento esfenomandibular (13) nace en la espina esfenoidal y se inser-
ta medialmente en la rama de la mandıbula, a la altura de la lıngula. Por su posicion y
orientacion parece ayudar al ligamento estilomandibular a limitar la protrusion. Moss [245]

26 Anatomıa funcional de la mandıbula
10
7
9
13
14
1
Figura 2.8: Vista medial de la ATM. Figura tomada de Hylander [160].
sugirio, por el contrario, que este ligamento nada tiene que ver con la ATM y que tan solo
protege los vasos sanguıneos y nervios que pasan a traves del agujero mandibular, de las
tensiones de traccion que tienen lugar en la apertura y cierre de la boca.
2.5. Musculos que intervienen en la masticacion
Cuatro potentes musculos se consideran responsables de la masticacion: masetero, tempo-
ral, pterigoideo lateral y pterigoideo medial. Estos cuatro, llamados musculos masticatorios,
junto con otros musculos de la cara, con la lengua, el paladar y el hueso hioides, funcionan
de forma coordinada durante la masticacion. A continuacion se realiza una breve descripcion
de los mismos, indicando su funcion, zonas de insercion y orientacion.
2.5.1. Musculos masticatorios
Masetero
El masetero (1) (ver figura 2.9) es el musculo mas potente del cuerpo humano. Con
forma de lamina rectangular, se extiende desde el arco cigomatico hasta la superficie inferior
del angulo de la mandıbula. Esta dividido en dos partes: una porcion superficial (2) y otra
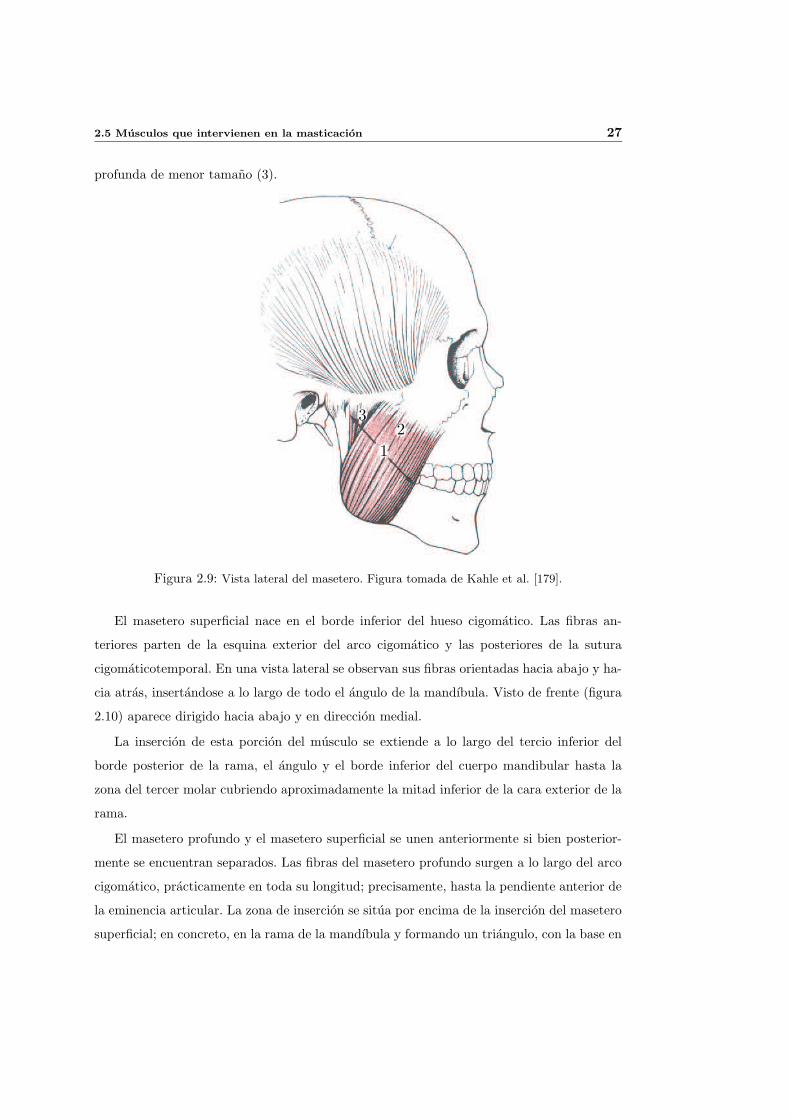
2.5 Musculos que intervienen en la masticacion 27
profunda de menor tamano (3).
1
2
3
1
Figura 2.9: Vista lateral del masetero. Figura tomada de Kahle et al. [179].
El masetero superficial nace en el borde inferior del hueso cigomatico. Las fibras an-
teriores parten de la esquina exterior del arco cigomatico y las posteriores de la sutura
cigomaticotemporal. En una vista lateral se observan sus fibras orientadas hacia abajo y ha-
cia atras, insertandose a lo largo de todo el angulo de la mandıbula. Visto de frente (figura
2.10) aparece dirigido hacia abajo y en direccion medial.
La insercion de esta porcion del musculo se extiende a lo largo del tercio inferior del
borde posterior de la rama, el angulo y el borde inferior del cuerpo mandibular hasta la
zona del tercer molar cubriendo aproximadamente la mitad inferior de la cara exterior de la
rama.
El masetero profundo y el masetero superficial se unen anteriormente si bien posterior-
mente se encuentran separados. Las fibras del masetero profundo surgen a lo largo del arco
cigomatico, practicamente en toda su longitud; precisamente, hasta la pendiente anterior de
la eminencia articular. La zona de insercion se situa por encima de la insercion del masetero
superficial; en concreto, en la rama de la mandıbula y formando un triangulo, con la base en
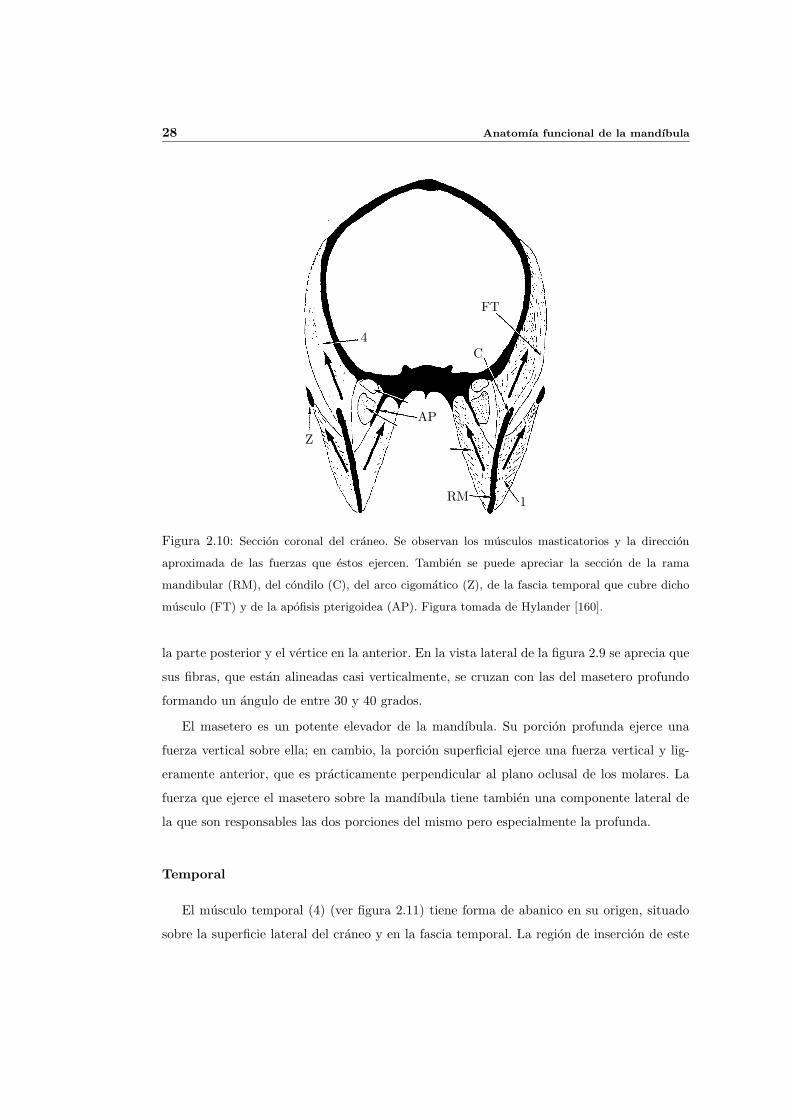
28 Anatomıa funcional de la mandıbula
1
4
RM
C
Z
AP
FT
Figura 2.10: Seccion coronal del craneo. Se observan los musculos masticatorios y la direccion
aproximada de las fuerzas que estos ejercen. Tambien se puede apreciar la seccion de la rama
mandibular (RM), del condilo (C), del arco cigomatico (Z), de la fascia temporal que cubre dicho
musculo (FT) y de la apofisis pterigoidea (AP). Figura tomada de Hylander [160].
la parte posterior y el vertice en la anterior. En la vista lateral de la figura 2.9 se aprecia que
sus fibras, que estan alineadas casi verticalmente, se cruzan con las del masetero profundo
formando un angulo de entre 30 y 40 grados.
El masetero es un potente elevador de la mandıbula. Su porcion profunda ejerce una
fuerza vertical sobre ella; en cambio, la porcion superficial ejerce una fuerza vertical y lig-
eramente anterior, que es practicamente perpendicular al plano oclusal de los molares. La
fuerza que ejerce el masetero sobre la mandıbula tiene tambien una componente lateral de
la que son responsables las dos porciones del mismo pero especialmente la profunda.
Temporal
El musculo temporal (4) (ver figura 2.11) tiene forma de abanico en su origen, situado
sobre la superficie lateral del craneo y en la fascia temporal. La region de insercion de este
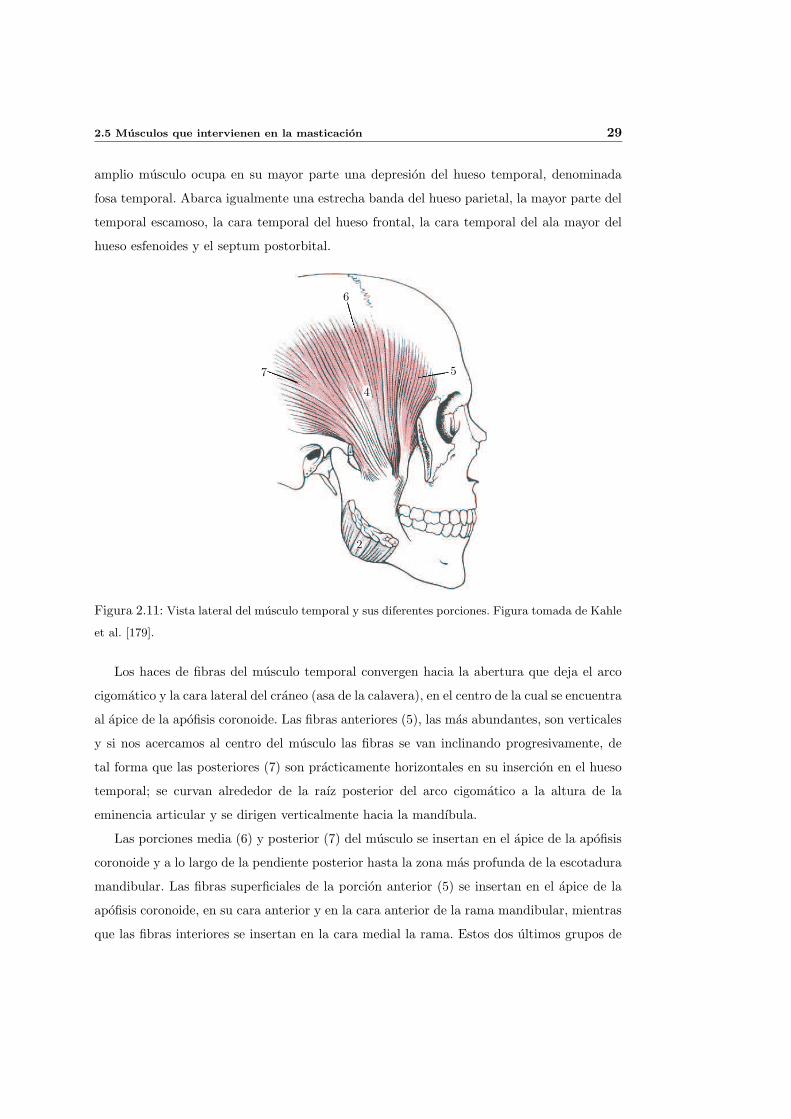
2.5 Musculos que intervienen en la masticacion 29
amplio musculo ocupa en su mayor parte una depresion del hueso temporal, denominada
fosa temporal. Abarca igualmente una estrecha banda del hueso parietal, la mayor parte del
temporal escamoso, la cara temporal del hueso frontal, la cara temporal del ala mayor del
hueso esfenoides y el septum postorbital.
4
5
6
7
2
1
Figura 2.11: Vista lateral del musculo temporal y sus diferentes porciones. Figura tomada de Kahle
et al. [179].
Los haces de fibras del musculo temporal convergen hacia la abertura que deja el arco
cigomatico y la cara lateral del craneo (asa de la calavera), en el centro de la cual se encuentra
al apice de la apofisis coronoide. Las fibras anteriores (5), las mas abundantes, son verticales
y si nos acercamos al centro del musculo las fibras se van inclinando progresivamente, de
tal forma que las posteriores (7) son practicamente horizontales en su insercion en el hueso
temporal; se curvan alrededor de la raız posterior del arco cigomatico a la altura de la
eminencia articular y se dirigen verticalmente hacia la mandıbula.
Las porciones media (6) y posterior (7) del musculo se insertan en el apice de la apofisis
coronoide y a lo largo de la pendiente posterior hasta la zona mas profunda de la escotadura
mandibular. Las fibras superficiales de la porcion anterior (5) se insertan en el apice de la
apofisis coronoide, en su cara anterior y en la cara anterior de la rama mandibular, mientras
que las fibras interiores se insertan en la cara medial la rama. Estos dos ultimos grupos de

30 Anatomıa funcional de la mandıbula
fibras envıan sendos tendones que se dirigen hacia el extremo posterior del proceso alveolar.
El interior, mas largo y fuerte, alcanza la zona del tercer molar inferior y el superficial
esta unido al borde anterior de la apofisis coronoide y la cara anterior de la rama.
La funcion principal del musculo temporal es elevar la mandıbula. Su forma de abanico
hace que la direccion de traccion varıe dependiendo de las fibras que se encuentren activas.
Parece logico pensar que las fibras posteriores, que estan alineadas horizontalmente,
ayuden en el movimiento de retrusion. Sin embargo estas fibras se curvan anteriormente
para insertarse en la muesca de la mandıbula en direccion vertical y es en esta direccion
como actuan sobre ella. Al pasar cerca de la eminencia articular es probable que realicen
una funcion de estabilizacion de la ATM. La porcion media de fibras, orientadas en direccion
oblicua, ejercen una fuerza que permite tanto la elevacion de la mandıbula como su retrusion.
La mayor parte de las fibras anteriores ejercen una traccion vertical. Las que se originan en
el septum postorbital tiran de la mandıbula hacia arriba y ligeramente hacia delante. Por
ultimo, las fibras anteriores profundas tiran hacia arriba y en direccion medial.
Pterigoideo medial
Este potente musculo rectangular, (8) en la figura 2.12, esta situado en la cara medial
de la rama mandibular y puede considerarse homologo del masetero a ese lado de la rama,
aunque es un poco mas pequeno y menos potente.
Tiene su origen en la fosa pterigoidea, depresion existente entre las laminas pterigoides del
hueso esfenoides. Las fibras mas profundas surgen de firmes tendones en la superficie medial
de la lamina pterigoide lateral. Anterior y lateralmente respecto a esta porcion profunda se
situa otro haz de fibras, que constituyen la porcion superficial del musculo y que se originan
en la cara lateral de la lamina pterigoide lateral y en la tuberosidad maxilar.
El pterigoideo medial discurre principalmente hacia abajo, hacia atras y en direccion
lateral para insertarse en la superficie medial del angulo de la mandıbula, en un area aprox-
imadamente triangular localizada entre dicho angulo y el surco milohioideo.
En una vista lateral la orientacion global de las fibras de este musculo es similar a
la porcion superficial del masetero y por tanto se considera un elevador de la mandıbula.
Sin embargo, a diferencia de este que desarrolla una fuerza con una pequena componente
lateral, el pterigoideo medial ejerce una fuerza con una componente medial, importante en
el movimiento de desviacion lateral.

2.5 Musculos que intervienen en la masticacion 31
4
8 10
11
1
Figura 2.12: Vista lateral del pterigoideo medial. Las dos porciones del pterigoideo lateral, (10)
y (11), ası como el temporal, (4), aparecen seccionados por claridad. Figura tomada de Kahle et
al. [179].
Pterigoideo lateral
Este musculo (9) tiene dos porciones, superior (10) e inferior (11), como se aprecia en la
figura 2.12, en las que aparecen seccionados, y en la figura 2.13.
La porcion superior tiene su origen en la superficie infratemporal del ala mayor del
hueso esfenoides. A partir de su origen las fibras de esta porcion se dirigen hacia atras,
horizontal y lateralmente. La porcion inferior, que es aproximadamente tres veces mayor
que la porcion superior, surge de la superficie exterior de la lamina pterigoidea lateral. Sus
fibras tambien se dirigen hacia atras y lateralmente, como las de la porcion superior, pero
en este caso no en direccion horizontal, sino hacia arriba, formando ambas un angulo de 45
grados aproximadamente.
Las dos porciones del pterigoideo lateral estan separadas en su origen pero se unen justo
enfrente de la ATM. Las fibras de la porcion superior se insertan principalmente en una
cavidad rugosa situada en la superficie anteromedial del cuello del condilo, llamada fosita
pterigoidea, pero una pequena porcion se inserta directamente en la parte anteromedial de

32 Anatomıa funcional de la mandıbula
4
9
10
11
1
Figura 2.13: Vista lateral de las dos porciones, superior (10) e inferior (11), del pterigoideo lateral
(9). Tambien se puede apreciar el pterigoideo medial: la porcion superficial, anterior, esta situ-
ada lateralmente a la porcion inferior del pterigoideo lateral y la porcion profunda esta situada
medialmente a la misma. Figura tomada de Kahle et al. [179].
la capsula y el disco articular de la ATM [346, 348]. Todas las fibras de la porcion inferior
se insertan en la fosita pterigoidea y su periferia.
Meyenberg [233] aprecia una cierta disparidad en la morfologıa del musculo de unos
individuos a otros. Este autor disecciono 25 ATMs y comprobo que, tal como se acaba de
decir, en todos los individuos las dos porciones del musculo se unıan enfrente de la ATM,
en contra de la opinion de algunos autores [129,151]. Sin embargo, en el 40 % de los casos la
porcion superior no se insertaba ni en el disco ni en la capsula articular, sino que lo hacıa
exclusivamente en la fosita pterigoidea. Esta disparidad siembra ciertas dudas sobre el papel
que juega el pterigoideo lateral durante la apertura de la boca, ya que es probable que no
ayude al movimiento relativo entre condilo y eminencia articular sino que su mision sea otra.
La fuerza resultante que ejerce sobre el condilo la porcion superior del musculo esta di-
rigida hacia delante y en direccion medial. Vista de lado, es practicamente perpendicular a
la superficie posterior de la eminencia articular y a la cara del condilo que esta enfrentada
a esta. Parece por tanto que esta porcion del musculo tiene una mision de estabilizacion del

2.5 Musculos que intervienen en la masticacion 33
condilo, al empujarlo contra la eminencia articular durante la masticacion y la incision.
La fuerza que desarrolla la porcion inferior sobre el condilo esta dirigida hacia delante,
medialmente y hacia abajo. Comparado con la porcion superior su direccion de traccion
se aproxima mas al plano tangente a la superficie articular de la ATM. La contraccion
bilateral de ambas porciones inferiores, derecha e izquierda, tira de los condilos y de los
discos, que de esta forma superan la eminencia articular en el movimiento de retrusion. La
contraccion unilateral de dicha porcion tira del condilo hacia abajo, hacia delante y hacia
dentro, produciendo una desviacion lateral.
Durante el cierre de la boca ambas porciones experimentan una contraccion excentrica
(estan en tension pero se alargan), ya que el condilo vuelve hacia atras. En estas condiciones,
el musculo pterigoideo hace de estabilizador de la articulacion controlando los movimientos
del condilo.
2.5.2. Otros musculos que intervienen en la masticacion
Aparte de los anteriores, denominados musculos masticatorios, en la masticacion inter-
vienen otros musculos que ayudan a la apertura de la boca y al movimiento de la lengua y
el hueso hioides, para la deglucion.
Digastrico
Tiene dos partes: vientre anterior (12) y vientre posterior (13), que se aprecian en la figura
2.14. El posterior tiene su origen en la escotadura mastoidea del hueso temporal (aparece
cortado en la figura) y el anterior en la fosa digastrica de la mandıbula. Ambos se unen en
el tendon intermedio (14), que se une al hueso hioides por una hoja de la fascia cervical
superficial (15).
Una de las misiones del digastrico es fijar el hueso hioides, junto con otros musculos que
se unen a este hueso, que sin embargo no esta del todo fijo durante la masticacion. Al tirar
de la mandıbula hacia atras y hacia abajo actua durante los movimientos de apertura y de
retrusion.
Milohiodeo
El musculo milohiodeo (16) forma un diafragma muscular que sirve de suelo de la boca.
Es un musculo plano, amplio de forma trapezoidal y esta localizado por debajo del vientre

34 Anatomıa funcional de la mandıbula
12
13
14
15
16
18
20
1
Figura 2.14: Musculos suprahioideos: vientres anterior (12) y posterior (13) del digastrico, milo-
hioideo (16) y estilohioideo (18). De este grupo muscular solo falta el genihioideo, que se representa
aparte, en la figura 2.15, por claridad. Figura tomada de Hylander [160].
anterior del digastrico (ver fig. 2.14), con el que a veces se funde. La base menor del trapecio
se une a la cara anterior del hueso hioides y la mayor a la mandıbula a lo largo de la lınea
milohioidea, hasta la sınfisis.
Su mision es elevar el hueso hioides y la lengua, ayudando ası a la deglucion. Tambien
puede actuar como depresor de la mandıbula.
Genihioideo
El genihioideo (17) es un musculo en forma de cordon que se origina en la apofisis geniana
y por abajo en el hueso hioides y se inserta en la cara lingual de la sınfisis mandibular, a
uno y otro lado de la linea sinfisaria (ver fig. 2.15).
Si la mandıbula esta cerrada su contraccion puede levantar ligeramente el hueso hioides
y la lengua llevandolas hacia delante. Por contra, si lo que esta fijo es el hueso hioides o bien
esta situado en su posicion mas baja, actua como depresor de la mandıbula.

2.5 Musculos que intervienen en la masticacion 35
16
17
1
Figura 2.15: Musculos milohioideo (16) y genihioideo (17). Figura tomada de Hylander [160].
Estilohioideo
El estilohioideo (18) es un musculo delgado y redondo que se origina en la apofisis estiloide
y se inserta en el asta mayor del hueso hioides. Antes de su insercion se divide en dos,
abrazando el tendon intermedio del digastrico (ver fig. 2.14).
Su funcion no esta muy clara y parece tener poca influencia en los movimientos de la
mandıbula, si acaso, podrıa actuar como elevador del hueso hioides.
Musculos infrahioideos
Cuatro musculos en forma de cordon forman el grupo muscular infrahioideo: el omo-
hioideo (19), el esternohioideo (20), el esternotiroideo (21) y el tirohioideo (22), que se
pueden ver en la figura 2.16.
Omohioideo
El omohioideo se origina en el borde superior de la escapula y se inserta en el asta mayor
del hioides. El esternotiroideo se origina en la parte superior del manubrio esternal y se
inserta en el cartılago tiroides. De ahı surge el tirohioideo, que es continuacion del ester-
notiroideo, insertandose en la superficie posterior del hioides. Por ultimo, el esternohioideo,
se origina en el manubrio del esternon y se inserta en la cara posterior del hioides.
Todos los musculos infrahioideos actuan de forma conjunta estabilizando y deprimiendo
36 Anatomıa funcional de la mandıbula
19
19
19
20
21
22
1
19
20
21
22
1
Figura 2.16: Grupo muscular infrahioideo: omohioideo (19), esternohioideo (20), esternotiroideo
(21) y tirohioideo (22). Figura tomada de Kahle et al. [179].
el hioides, cuyos movimientos ascendentes tambien quedan limitados por este grupo mus-
cular. Ası, junto al grupo de musculos suprahioideos (digastrico, milohioideo, genihioideo y
estilohioideo), controlan la posicion del hioides la lengua y la mandıbula durante la masti-
cacion.
2.6. Movimientos libres de la mandıbula
Los movimientos masticatorios de la mandıbula, que se analizan en un apartado poste-
rior, pueden entenderse mas facilmente si antes se analizan los movimientos libres. Ası se
denominan a los movimientos que se producen en ausencia de comida en la cavidad oral.
Los movimientos libres que se pueden realizar con la mandıbula son: cierre-apertura, pro-
trusion-retrusion y desviacion lateral.
Pero estos movimientos son combinacion de dos movimientos basicos: rotacion y traslacion.
La rotacion se produce alrededor de un eje que pasa por los condilos y provoca un movimien-
to relativo entre el condilo y el disco articular, en el compartimento inferior de la ATM. La

2.6 Movimientos libres de la mandıbula 37
traslacion, en cambio, se produce en el compartimento superior, por un movimiento relativo
entre disco y eminencia articular. La traslacion es posible tanto en direccion anteroposterior
como en direccion mediolateral.
2.6.1. Cierre y apertura
Descripcion del movimiento
Durante los movimientos de apertura y cierre de la boca se produce una traslacion y
rotacion combinadas. La traslacion lleva el disco y el condilo hacia delante y hacia abajo, a
lo largo de la pendiente posterior de la eminencia articular. Puede incluso llevarlos mas alla,
sobre el plano preglenoideo. La rotacion es amplia y permite una apertura de la boca entre
40 y 60mm.
En la primera fase del movimiento de apertura predomina la rotacion, que posteriormente
se combina con la traslacion hasta la apertura completa (ver fig. 2.17). El cierre comienza con
una traslacion hacia atras, hasta que se cierra un tercio de la apertura maxima, momento
en que se superponen traslacion y rotacion, para llevar a la mandıbula a la posicion de
reposo. Si se sigue cerrando la boca aun mas, hasta la posicion de oclusion, el movimiento
predominante en esta fase es el de rotacion [250].
1
2
Figura 2.17: Movimiento de apertura libre. En la primera fase (1) la traslacion es mucho menor
que en la segunda (2). Figura tomada de Hylander [160].

38 Anatomıa funcional de la mandıbula
Actividad muscular
La apertura se produce gracias a la gravedad, la relajacion de los musculos elevadores de
la mandıbula (masetero, temporal y pterigoideo medio) y por la accion conjunta del milo-
hioideo, el digastrico y el genihioideo. Los musculos infrahioideos mantienen la estabilidad
del hueso hioides.
Si la apertura es moderada y sin resistencia, el movimiento se produce simplemente por
efecto de la gravedad y la relajacion de los elevadores (solo fase de rotacion). En una apertura
mas amplia, la porcion inferior del pterigoideo lateral tira de los condilos hacia delante,
genihioideo y digastrico tiran hacia abajo y atras del menton, mientras que el milohioideo
hace lo propio con el cuerpo mandibular.
El cierre de la boca se produce por la accion de los musculos elevadores de la mandıbula.
Cuando la apertura ha sido amplia se produce primero una relajacion de los musculos que
abren la boca, relajacion que hace que los condilos vuelvan desde el plano preglenoideo hasta
la eminencia articular (fase de traslacion) y entonces se contraen los elevadores que terminan
el movimiento (fase de rotacion-traslacion).
2.6.2. Protrusion y retrusion
Descripcion del movimiento
Se denominan protrusion y retrusion a los movimientos que llevan la mandıbula hacia
delante y hacia atras. Son principalmente de traslacion y por lo tanto ocurren en el compar-
timento superior de la ATM, moviendose el disco y el condilo conjuntamente.
La retrusion puede producirse desde una posicion adelantada, de protrusion, y tambien
desde la posicion de reposo. En este ultimo caso la mandıbula puede moverse hacia atras
entre 1 y 2mm, estando limitado este movimiento por las fibras horizontales de la capsula
articular.
Actividad muscular
La protrusion se realiza, casi exclusivamente, por la contraccion de la porcion inferior del
pterigoideo lateral, que tira del condilo y el disco articular hacia delante. Tambien actuan,
aunque en menor medida, el masetero, el pterigoideo medial y los depresores de la mandıbu-
la (genihioideo, milohioideo y digastrico), que ayudan a estabilizar su posicion durante el

2.6 Movimientos libres de la mandıbula 39
movimiento.
En la retrusion intervienen las porciones media y posterior del musculo temporal, incli-
nadas respecto a la vertical, y los depresores.
2.6.3. Desviacion lateral
Este es el movimiento que se produce cuando se intenta mover la mandıbula hacia un lado.
El movimiento, que se esquematiza en la figura 2.18, consiste en una rotacion alrededor de un
eje vertical, situado inmediatamente detras del condilo hacia el que se mueve la mandıbula,
denominado condilo ipsilateral.
1
2
3
12
3
I C
LLE
Figura 2.18: Esquema del movimiento de desviacion lateral de la mandıbula. El condilo ipsilateral
(I) comienza girando alrededor de un eje vertical que pasa por su centro hasta que el ligamento
lateral externo (LLE) entra en tension, en la posicion 2. Figura tomada de Hylander [160].
El condilo del lado opuesto, o contralateral, se mueve hacia abajo por la eminencia
articular, anterior y medialmente. El condilo ipsilateral gira en primer lugar alrededor de un
eje que pasa por su centro. De esta forma el polo lateral de dicho condilo se desplaza hacia
atras, entre 1 y 2mm, hasta que el ligamento lateral externo entra en tension y no permite
que el condilo siga girando. A partir de ese instante solo se desplaza, en direccion lateral y
ligeramente hacia delante, guiado por dicho ligamento.
Este movimiento se consigue por la accion combinada del pterigoideo lateral del lado
contralateral, que tira del condilo correspondiente hacia delante, y la porcion media del

40 Anatomıa funcional de la mandıbula
temporal del lado ipsilateral, que estabiliza el otro condilo, impidiendo que se mueva en
exceso en direccion anterior.
2.7. Movimientos masticatorios de la mandıbula
Como se comento al principio del capıtulo, se analizan los movimientos masticatorios de
la mandıbula con el objeto de simular las cargas y condiciones de contorno que es necesario
aplicar a un modelo de EF que trate de estimar las tensiones que la actividad masticatoria
produce en la mandıbula.
Aunque existe un patron general de movimiento de la mandıbula durante la masticacion,
dicho movimiento puede variar con el individuo y la situacion, dependiendo de numerosos
factores como pueden ser: la forma y dimensiones, tanto de la mandıbula como de los dientes,
la edad y el estado de la denticion, el tipo de comida, el estado del bolo alimenticio, etc.
Los movimientos masticatorios son de dos tipos: un movimiento de corte, denominado in-
cision, que se realiza normalmente con caninos e incisivos y un movimiento de trituracion del
alimento, denominado masticacion, que se realiza exclusivamente con premolares y molares.
2.7.1. Incision
Este movimiento consta de tres fases [168, 303]: apertura, cierre y fase de corte. La fase
de cierre termina en el instante en el que los bordes incisales de los incisivos inferiores tocan
el alimento. Sin embargo esta fase no se produce siempre, ya que si el tamano del alimento
es demasiado grande, la fase de apertura termina con los incisivos tocando dicho alimento.
Las fases de apertura y cierre son identicas a los movimientos libres del mismo nombre,
que se analizaron en el apartado anterior. Sin embargo, la fase de corte es diferente al cierre
libre, aunque la boca se siga cerrando durante el corte. Durante esta fase la mandıbula
se mueve hacia atras y hacia arriba, deslizando los bordes incisales de los incisivos infe-
riores sobre la cara palatina de los incisivos superiores (accion de cizalla), que se detiene
cuando el elemento hace resistencia. Posteriormente se produce una elevacion acompanada
de oscilaciones en direccion anteroposterior hasta que el alimento se corta y la mandıbula
desciende.
La actividad muscular durante las fases de apertura y cierre tambien es identica a la que
tiene lugar durante la apertura y cierre libre de la boca. En la fase de corte se produce una

2.7 Movimientos masticatorios de la mandıbula 41
contraccion simetrica, mas o menos sincronizada, de los musculos elevadores de la mandıbu-
la, si bien sobre la actividad del temporal hay disparidad de opiniones.1 El pterigoideo lateral
tambien participa en la incision, para estabilizar la ATM. Esta estabilizacion es necesaria
para controlar posibles cambios bruscos en la direccion y magnitud de las fuerzas de mas-
ticacion. Por ejemplo, cuando el alimento se rompe repentinamente, el pterigoideo lateral
previene un desplazamiento brusco de los condilos hacia arriba y hacia atras.
2.7.2. Masticacion
El ser humano tiene diferentes patrones de masticacion: unilateral, si el alimento se
situa siempre en el mismo lado de la boca; bilateral simultaneo, si el alimento se mastica
con los dientes de ambos lados, y bilateral alternante, si se mastica solo con un lado, pero
alternando el lado entre masticaciones. Este ultimo es el patron mas habitual, 75 % segun
Manns y Dıaz [211], seguido del unilateral con un 15 % y el bilateral simultaneo, que siguen
el 10 % restante. Se analiza a continuacion la masticacion unilateral por ser la mas comun (la
realizan los individuos con patron unilateral y bilateral alternante, el 90 % de la poblacion).
El movimiento de masticacion es una sucesion de ciclos denominada secuencia mastica-
toria. Cada ciclo masticatorio se puede dividir en tres fases [147]: apertura, cierre y fase
oclusal. Las fases de apertura y cierre no son simetricas en la masticacion unilateral, como
se puede apreciar en la figura 2.19. En ella se representan los movimientos del punto de
contacto entre los incisivos durante una masticacion unilateral con el lado derecho.
Despues de completarse la fase de cierre comienza la fase oclusal, en la que se produce el
contacto entre la comida y las superficies oclusales de molares y/o premolares. A veces, la
boca se abre nuevamente, acabando ası la fase oclusal, antes de alcanzar el contacto directo
entre dientes. En caso contrario, la fase oclusal se denomina con contacto y se subdivide en
dos fases, esquematizadas en la figura 2.20. En la fase I el movimiento de la mandıbula tiene
una pequena componente ascendente (no se aprecia en la figura) que lleva a ambos maxilares
a la oclusion centrica [181,237]. En este instante existen dos posibilidades: o bien la boca se
abre y se interrumpe la fase oclusal o bien esta continua en la fase II [181,237], moviendose
la mandıbula respecto al maxilar superior en la direccion de la figura 2.20 y ligeramente
1MacDonald [207] entre otros, niega su participacion, mientras que Hylander y Johnson [161] demuestran
lo contrario. Lo que si confirman gran cantidad de estudios [197, 240, 334] es que la actividad del temporal
es nula durante un apriete con los incisivos, por lo que parece mas creıble que tambien lo sea durante la
incision.

42 Anatomıa funcional de la mandıbula
Figura 2.19: Trayectoria descrita por el punto de contacto del incisivo central inferior durante una
masticacion unilateral con el lado derecho. Figura tomada de Hylander [160].
hacia abajo. Si existe la fase II, antes de que esta comience, se produce una parada en la
posicion de oclusion centrica, que dura entre 50 y 200 milisegundos [121,160].
Antes de describir la actividad muscular en los ciclos masticatorios, conviene aclarar que
en la masticacion unilateral los terminos ipsilateral y contralateral hacen referencia al lado
en el que se situa el alimento y al lado opuesto, respectivamente.
La actividad muscular durante la masticacion unilateral no es simetrica, como tampoco
lo eran los movimientos. En la figura 2.21 se observa un registro electromiografico de la
actividad de los principales musculos implicados en la masticacion. Por encima de cada
lınea se representa la actividad del musculo ipsilateral correspondiente, por debajo la del
contralateral.
En la figura 2.21 se senalan con flechas los instantes en los que se produce la oclusion
centrica, (1), se pierde el contacto, (2), y se produce la maxima apertura de la boca (3).
Se observa como al final de la fase de cierre, cuando comienza la fase oclusal, aumenta la
actividad de los elevadores, alcanzandose el maximo casi simultaneamente en todos ellos
[239]. Este maximo ocurre durante la fase I, aproximadamente entre 40 y 80 milisegundos
antes del instante de maxima fuerza oclusal, que coincide con la oclusion centrica [128,162].
En el instante de maxima fuerza oclusal comienza tambien la actividad de los depresores de
la mandıbula (digastrico y milohiodeo son los unicos de los que se tienen datos) que entran
en tension a la vez que los elevadores se relajan y se abre rapidamente la boca [162].

2.7 Movimientos masticatorios de la mandıbula 43
III
OC
Figura 2.20: Vista paralela al plano oclusal de la mandıbula con la direccion de sus movimientos
respecto al maxilar inferior durante las fases I y II, en una masticacion con el segundo molar derecho.
OC hace referencia a la posicion de oclusion centrica. Figura tomada de Hylander [160].
La actividad del pterigoideo lateral en la masticacion es un tema bastante controvertido.
Segun Møller [239], las dos porciones actuan de forma independiente: la porcion superior
(lınea continua en la figura 2.21) actua durante la apertura y la inferior (lınea discontinua)
en el cierre.
Otro hecho destacable es la desigual contraccion de los musculos de uno y otro lado, si
exceptuamos el masetero profundo1 y el temporal. Ası, el masetero superficial y el pterigoideo
medio ipsilaterales son mas activos que los contralaterales en la masticacion unilateral [29,
239,240].
1La actividad de esta porcion del masetero no aparece registrada en los trabajos de Møller [239, 240], a
los que corresponde la figura 2.21. En [29] pueden encontrarse los valores maximos de su actividad aunque
sin indicacion del instante en el que ocurren.

44 Anatomıa funcional de la mandıbula
TA
TP
MS
PM
PL
DG
MH
1 2 3
200 ms
Cierre Apertura Cierre
Figura 2.21: Actividad electromiografica de los principales musculos implicados en una masticacion
unilateral: TA, temporal anterior; TP, temporal posterior; MS, masetero superficial; PM, pterigoideo
medio; PL, pterigoideo lateral; DG, digastrico y MH, milohioideo. Se senalan los siguientes instantes:
(1) oclusion centrica, (2) perdida de oclusion y (3) maxima apertura. Adaptado de [239].
2.8. Fuerzas desarrolladas por los musculos masticato-
rios
En la determinacion de la fuerza que es capaz de desarrollar un musculo durante una
actividad determinada se establecen dos hipotesis generalmente aceptadas:
a) La fuerza desarrollada por un musculo al contraerse es proporcional a la seccion del
mismo, siendo la constante de proporcionalidad un parametro fisiologico [138,275,278,340].
b) La contraccion del musculo es variable a lo largo del tiempo que dura una determinada
actividad [208,209,239].
De esta forma, la resultante de fuerzas que un musculo realiza en su contraccion, Mir se
suele escribir:
Mir = EMGMi · (XMi ·K) (2.1)
donde K es la referida constante fisiologica, XMi es la seccion del musculo y EMGMi su
actividad, medida por electromiografıa y que representa la proporcion entre la contraccion
instantanea del musculo y la contraccion maxima que este puede experimentar [275,339].
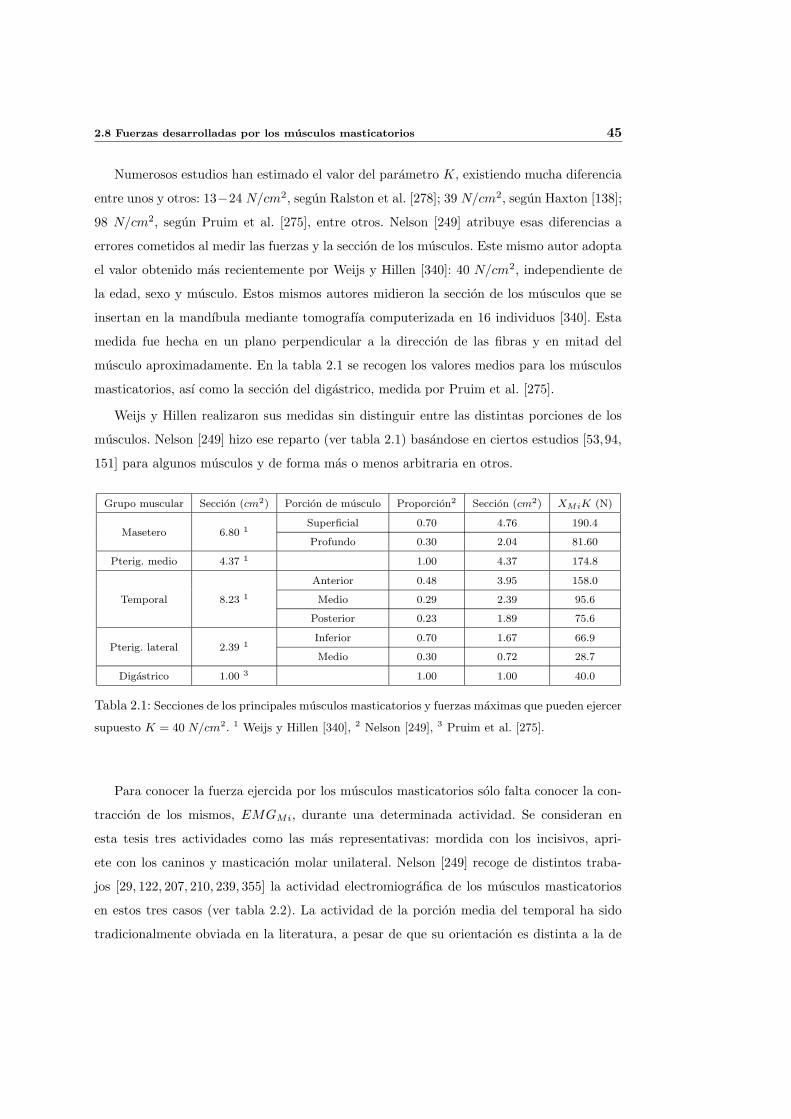
2.8 Fuerzas desarrolladas por los musculos masticatorios 45
Numerosos estudios han estimado el valor del parametro K, existiendo mucha diferencia
entre unos y otros: 13−24 N/cm2, segun Ralston et al. [278]; 39 N/cm2, segun Haxton [138];
98 N/cm2, segun Pruim et al. [275], entre otros. Nelson [249] atribuye esas diferencias a
errores cometidos al medir las fuerzas y la seccion de los musculos. Este mismo autor adopta
el valor obtenido mas recientemente por Weijs y Hillen [340]: 40 N/cm2, independiente de
la edad, sexo y musculo. Estos mismos autores midieron la seccion de los musculos que se
insertan en la mandıbula mediante tomografıa computerizada en 16 individuos [340]. Esta
medida fue hecha en un plano perpendicular a la direccion de las fibras y en mitad del
musculo aproximadamente. En la tabla 2.1 se recogen los valores medios para los musculos
masticatorios, ası como la seccion del digastrico, medida por Pruim et al. [275].
Weijs y Hillen realizaron sus medidas sin distinguir entre las distintas porciones de los
musculos. Nelson [249] hizo ese reparto (ver tabla 2.1) basandose en ciertos estudios [53,94,
151] para algunos musculos y de forma mas o menos arbitraria en otros.
Grupo muscular Seccion (cm2) Porcion de musculo Proporcion2 Seccion (cm2) XMiK (N)
Masetero 6.80 1Superficial 0.70 4.76 190.4
Profundo 0.30 2.04 81.60
Pterig. medio 4.37 1 1.00 4.37 174.8
Temporal 8.23 1
Anterior 0.48 3.95 158.0
Medio 0.29 2.39 95.6
Posterior 0.23 1.89 75.6
Pterig. lateral 2.39 1Inferior 0.70 1.67 66.9
Medio 0.30 0.72 28.7
Digastrico 1.00 3 1.00 1.00 40.0
Tabla 2.1: Secciones de los principales musculos masticatorios y fuerzas maximas que pueden ejercer
supuesto K = 40 N/cm2. 1 Weijs y Hillen [340], 2 Nelson [249], 3 Pruim et al. [275].
Para conocer la fuerza ejercida por los musculos masticatorios solo falta conocer la con-
traccion de los mismos, EMGMi, durante una determinada actividad. Se consideran en
esta tesis tres actividades como las mas representativas: mordida con los incisivos, apri-
ete con los caninos y masticacion molar unilateral. Nelson [249] recoge de distintos traba-
jos [29, 122, 207, 210, 239, 355] la actividad electromiografica de los musculos masticatorios
en estos tres casos (ver tabla 2.2). La actividad de la porcion media del temporal ha sido
tradicionalmente obviada en la literatura, a pesar de que su orientacion es distinta a la de
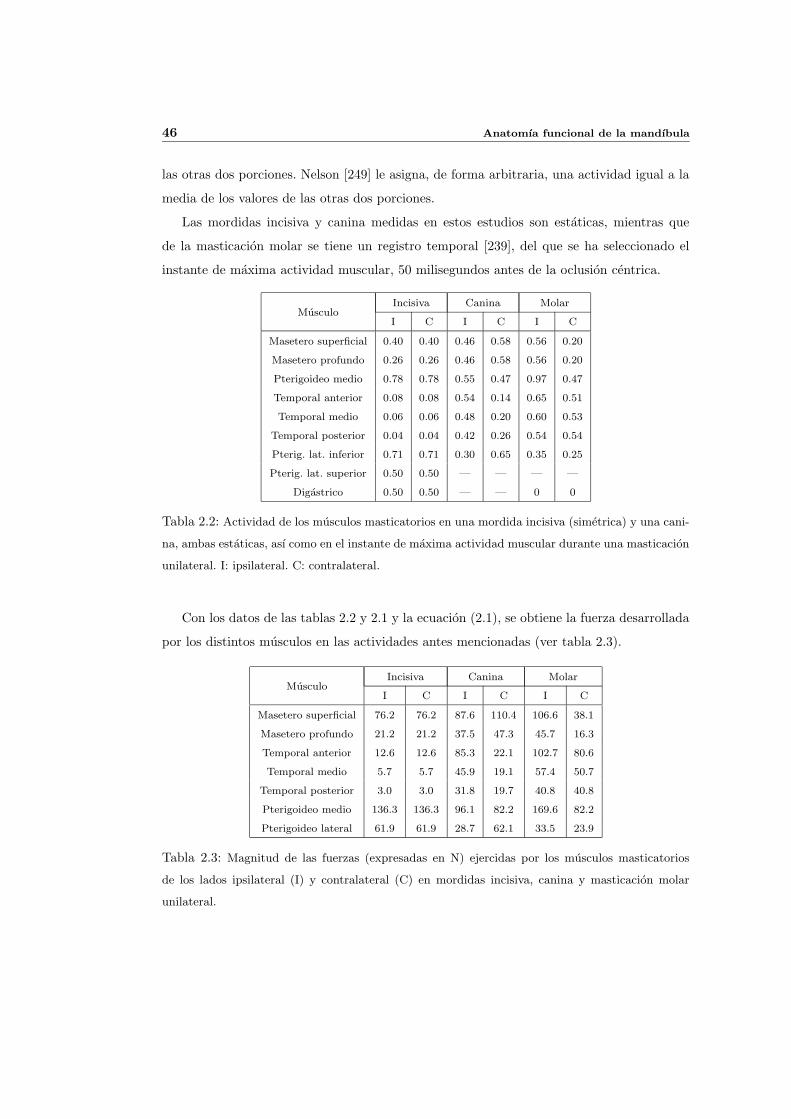
46 Anatomıa funcional de la mandıbula
las otras dos porciones. Nelson [249] le asigna, de forma arbitraria, una actividad igual a la
media de los valores de las otras dos porciones.
Las mordidas incisiva y canina medidas en estos estudios son estaticas, mientras que
de la masticacion molar se tiene un registro temporal [239], del que se ha seleccionado el
instante de maxima actividad muscular, 50 milisegundos antes de la oclusion centrica.
MusculoIncisiva Canina Molar
I C I C I C
Masetero superficial 0.40 0.40 0.46 0.58 0.56 0.20
Masetero profundo 0.26 0.26 0.46 0.58 0.56 0.20
Pterigoideo medio 0.78 0.78 0.55 0.47 0.97 0.47
Temporal anterior 0.08 0.08 0.54 0.14 0.65 0.51
Temporal medio 0.06 0.06 0.48 0.20 0.60 0.53
Temporal posterior 0.04 0.04 0.42 0.26 0.54 0.54
Pterig. lat. inferior 0.71 0.71 0.30 0.65 0.35 0.25
Pterig. lat. superior 0.50 0.50 — — — —
Digastrico 0.50 0.50 — — 0 0
Tabla 2.2: Actividad de los musculos masticatorios en una mordida incisiva (simetrica) y una cani-
na, ambas estaticas, ası como en el instante de maxima actividad muscular durante una masticacion
unilateral. I: ipsilateral. C: contralateral.
Con los datos de las tablas 2.2 y 2.1 y la ecuacion (2.1), se obtiene la fuerza desarrollada
por los distintos musculos en las actividades antes mencionadas (ver tabla 2.3).
MusculoIncisiva Canina Molar
I C I C I C
Masetero superficial 76.2 76.2 87.6 110.4 106.6 38.1
Masetero profundo 21.2 21.2 37.5 47.3 45.7 16.3
Temporal anterior 12.6 12.6 85.3 22.1 102.7 80.6
Temporal medio 5.7 5.7 45.9 19.1 57.4 50.7
Temporal posterior 3.0 3.0 31.8 19.7 40.8 40.8
Pterigoideo medio 136.3 136.3 96.1 82.2 169.6 82.2
Pterigoideo lateral 61.9 61.9 28.7 62.1 33.5 23.9
Tabla 2.3: Magnitud de las fuerzas (expresadas en N) ejercidas por los musculos masticatorios
de los lados ipsilateral (I) y contralateral (C) en mordidas incisiva, canina y masticacion molar
unilateral.

Capıtulo 3
Modelo de EF de una
mandıbula humana con protesis
dental implantosoportada
3.1. Introduccion
En este capıtulo se describe el modelo de elementos finitos de la mandıbula empleado
para las simulaciones de esta tesis. Se distinguen dos fases en la obtencion del modelo con
protesis implantosoportada, que se esquematizan en la figura 3.1. En primer lugar se obtiene
un modelo de EF de la mandıbula completa con la denticion tambien completa. Se describe el
proceso seguido para la obtencion de dicho modelo, a partir de la mandıbula de un cadaver,
varon, de unos 50 anos de edad y parcialmente edentulo. Posteriormente se describen las
cargas y condiciones de contorno que se aplican para simular la actividad masticatoria. Dicho
modelo con denticion completa se usa para estimar la distribucion de densidad osea y de
propiedades elasticas, como se describe en un capıtulo posterior.
En la segunda fase, se supondra que el primer molar derecho ha sido sustituido por un
implante dental osteointegrado similar al implante comercial IMZ r©. Dicha sustitucion se
traslada al modelo de EF, para la posterior simulacion del comportamiento del hueso peri-
implantario. Este hueso, que rodea al implante, se vera afectado por el cambio en la forma
de transferirse las cargas: en un diente natural se reparten gracias al ligamento periodontal,
47

48 Modelo de EF de una mandıbula humana con protesis dental implantosoportada
mientras que desde el implante al hueso se transfieren principalmente en el cuello del im-
plante. Conocer los cambios que experimenta el hueso periimplantario es uno de los objetivos
de este trabajo. Las propiedades del resto del hueso se importan desde el modelo de denticion
completa, donde han sido estimadas, en la primera fase del analisis. Para reducir el tiempo
de computacion, el modelo implantado se corta en torno a la zona de interes para reducir
el numero de elementos, aplicando para ello tecnicas de submodelado. Es este submodelo al
que se aplica el algoritmo de remodelacion osea propuesto en esta tesis.
Medida de una mandıbula real
Modelo de EF de lamandıbula completa
ρ0, isotropo
Cargas de masticacionModelo de remod. basado en
la Mec. del Dano (cap. 5)
Mandıbula con huesode ρ f y anisotropo
Submodelo, con implante enlugar del 1er molar derecho
Cargas de masticacionModelo de remod. propuesto
en esta tesis (cap. 7)
Maduracion del huesoperiimplantario
1a fase
2a fase
1
Figura 3.1: Esquema del proceso seguido en las simulaciones de esta tesis.
3.2. Modelo de EF de la mandıbula con denticion com-
pleta
El proceso seguido para obtener el modelo de elementos finitos de la mandıbula se de-
scribe brevemente a continuacion. Con una maquina de medicion por coordenadas (MMC) se
obtuvo la posicion de un buen numero de puntos en el contorno de una mandıbula humana.
La fotografıa de la figura 3.2 fue tomada durante el proceso de medicion. En ella se puede

3.2 Modelo de EF de la mandıbula con denticion completa 49
observar el palpador ası como las pinzas de que dispone la MMC para fijar las piezas que lo
requieran. La fijacion, a la altura de la rama mandibular izquierda, limito en gran medida
la zona a la que podıa acceder el palpador. Por esta razon, se opto por medir tan solo la
mitad derecha de la mandıbula, para efectuar mas adelante una operacion de simetrıa con
respecto al plano medio en la region sinfisaria. En la figura 3.3 se observa la nube de puntos
obtenida con la MMC.
Figura 3.2: Montaje de la mandıbula en la MMC.
Posteriormente, con la ayuda del software CATIA r©, se importo la posicion de todos
los puntos obtenidos con la MMC y se interpolo una superficie que pasara por ellos. Para
realizar la operacion de simetrıa es necesario conocer el plano medio sinfisario, lo que puede
hacerse solo de forma aproximada. La busqueda de este plano medio se hizo de forma inversa:
se probo con varias orientaciones y posiciones posibles hasta que la operacion de simetrıa dio
un resultado coherente con la geometrıa original. Esta coherencia se refiere en concreto a la
coincidencia de las siguientes medidas: distancia entre algunos puntos del angulo mandibular
de uno y otro lado, entre los condilos y entre las apofisis coronoides. En la figura 3.4 se observa
la superficie exterior de la mandıbula obtenida por interpolacion y cortada en el plano medio,
obtenido como se acaba de decir.
50 Modelo de EF de una mandıbula humana con protesis dental implantosoportada
Figura 3.3: Nube de puntos obtenidos con la MMC.
Figura 3.4: Superficie que pasa por la nube de puntos obtenidos con la MMC.

3.2 Modelo de EF de la mandıbula con denticion completa 51
Una vez obtenida la superficie exterior de la mandıbula, el volumen comprendido por
esta fue mallado con elementos hexaedricos lineales de 8 nodos, del tipo C3D8, de la librerıa
de elementos de ABAQUS r©, codigo comercial usado en este estudio.
Hay que indicar que el proceso de medicion por coordenadas se limito a la zona ocupada
por hueso basal. No se midieron los dientes, por faltar muchos de ellos, ni el proceso alveolar,
alterado precisamente por el edentulismo del individuo. Las coordenadas de los puntos del
proceso alveolar fueron extrapoladas a partir de las obtenidas en el hueso basal y la de los
dientes fue supuesta de forma aproximada, basandose en las dimensiones de algunas piezas
que conservaba la mandıbula. Entre los dientes y el hueso se interpuso una capa de elementos
de 0.2mm de espesor para simular el ligamento periodontal, de la misma forma en que lo
hacen Korioth et al. [186]. Como particularidad del modelo hay que senalar que se dispuso
el tercer molar del lado derecho, pero no ası el del lado izquierdo. Esto esta justificado dado
que no todas las personas desarrollan este molar, vulgarmente llamado muela del juicio. El
hecho de considerar solo un tercer molar, permite comparar su influencia sobre el hueso
proximo a el.
Una vez obtenida la malla de una mitad de la mandıbula se procedio a realizar la op-
eracion de simetrıa con respecto al plano medio sinfisario. El modelo de elementos finitos
completo tiene 77490 elementos y 88836 nodos y se puede observar en la figura 3.5.
Figura 3.5: Modelo de EF de la mandıbula completa

52 Modelo de EF de una mandıbula humana con protesis dental implantosoportada
De las propiedades mecanicas del tejido oseo se hablara en los siguientes capıtulos, pues
constituye uno de los aspectos fundamentales de esta tesis. Sin embargo, a los otros tejidos
y materiales presentes en el modelo, en concreto a la dentina1 y al ligamento periodontal, no
se le presta una dedicacion tan exhaustiva y simplemente se les supone un comportamiento
isotropo, elastico y lineal. En la tabla 3.1 se da el modulo de Young y el coeficiente de
Poisson de ambos materiales
Propiedades elasticas E(MPa) ν
Dentina [78] 17600 0.25
Ligamento periodontal [277,345] 3 0.45
Tabla 3.1: Propiedades mecanicas de los materiales presentes en el modelo aparte del hueso.
El comportamiento de ambos se supone isotropo, elastico y lineal.
3.3. Simulacion de la actividad masticatoria
La actividad masticatoria es el proceso mediante el cual el alimento es convertido en bolo
alimenticio antes de ser deglutido. El papel de ambos maxilares en este proceso es cortar
el alimento, si es necesario, y triturarlo hasta convertirlo en una pasta mientras se mezcla
con la saliva. El corte o incision, solo es necesario en determinados casos, dependiendo del
alimento y del empleo de utensilios para comer. Se ejecuta normalmente con los incisivos
y a veces con los caninos. La trituracion del alimento se ejecuta con molares y premolares,
mediante los ciclos masticatorios descritos en el capıtulo 2.
La incision es un proceso en el que la boca permanece casi cerrada y practicamente
inmovil. Sin embargo, en la masticacion se produce una sucesion de aperturas y cierres de la
boca con un ligero movimiento lateral y en direccion anteroposterior, en los ya referidos ciclos
masticatorios. Por tanto, simular con rigor la masticacion requiere un analisis dinamico,
bastante complejo por otra parte: la contraccion de los musculos masticatorios es variable
a lo largo del ciclo; durante la apertura y cierre de la boca el disco articular desliza por la
cavidad glenoidea del temporal, lo que requiere ademas resolver un problema de contacto;
entre los molares y el alimento no solo existe contacto sino ademas grandes deformaciones
conforme el alimento va siendo triturado. Por todo ello, se haran algunas simplificaciones
para simular la masticacion, simplificaciones que tambien se usaran para simular la incision.
1La capa de esmalte que recubre la parte del diente que queda por encima de la mucosa, es muy delgada
y no se ha incluido. Se ha supuesto, por tanto, que el diente se compone exclusivamente de dentina.
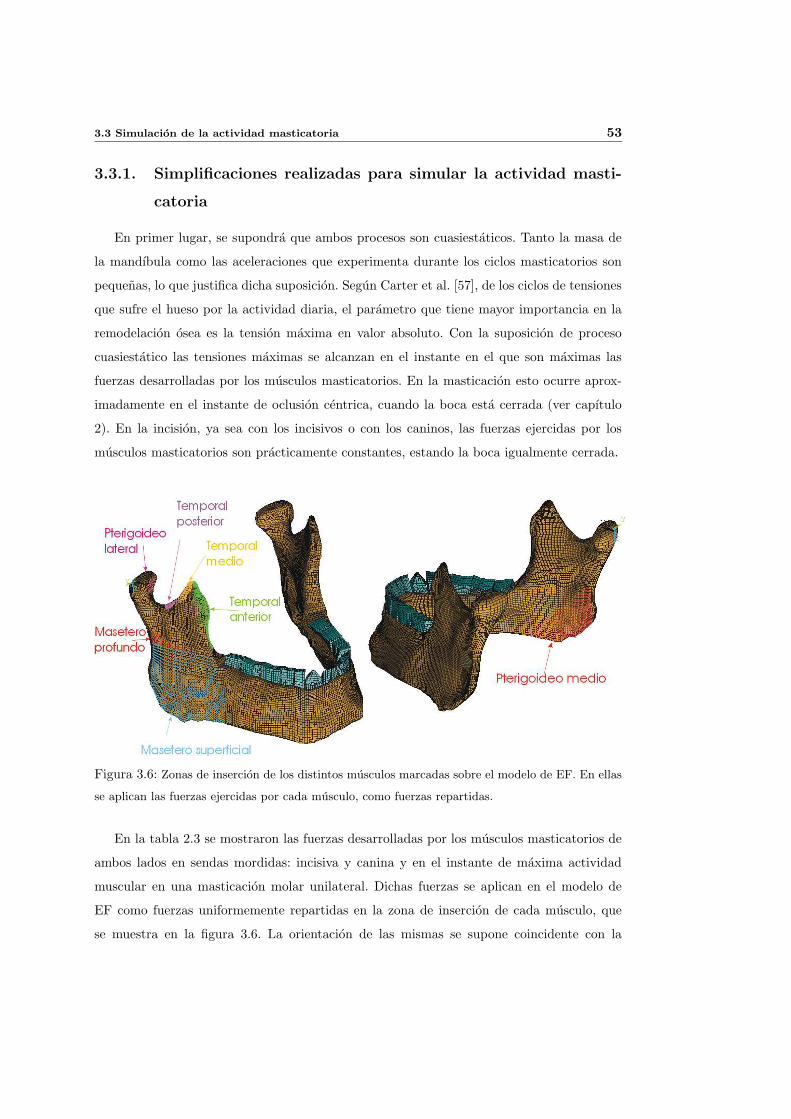
3.3 Simulacion de la actividad masticatoria 53
3.3.1. Simplificaciones realizadas para simular la actividad masti-
catoria
En primer lugar, se supondra que ambos procesos son cuasiestaticos. Tanto la masa de
la mandıbula como las aceleraciones que experimenta durante los ciclos masticatorios son
pequenas, lo que justifica dicha suposicion. Segun Carter et al. [57], de los ciclos de tensiones
que sufre el hueso por la actividad diaria, el parametro que tiene mayor importancia en la
remodelacion osea es la tension maxima en valor absoluto. Con la suposicion de proceso
cuasiestatico las tensiones maximas se alcanzan en el instante en el que son maximas las
fuerzas desarrolladas por los musculos masticatorios. En la masticacion esto ocurre aprox-
imadamente en el instante de oclusion centrica, cuando la boca esta cerrada (ver capıtulo
2). En la incision, ya sea con los incisivos o con los caninos, las fuerzas ejercidas por los
musculos masticatorios son practicamente constantes, estando la boca igualmente cerrada.
Figura 3.6: Zonas de insercion de los distintos musculos marcadas sobre el modelo de EF. En ellas
se aplican las fuerzas ejercidas por cada musculo, como fuerzas repartidas.
En la tabla 2.3 se mostraron las fuerzas desarrolladas por los musculos masticatorios de
ambos lados en sendas mordidas: incisiva y canina y en el instante de maxima actividad
muscular en una masticacion molar unilateral. Dichas fuerzas se aplican en el modelo de
EF como fuerzas uniformemente repartidas en la zona de insercion de cada musculo, que
se muestra en la figura 3.6. La orientacion de las mismas se supone coincidente con la
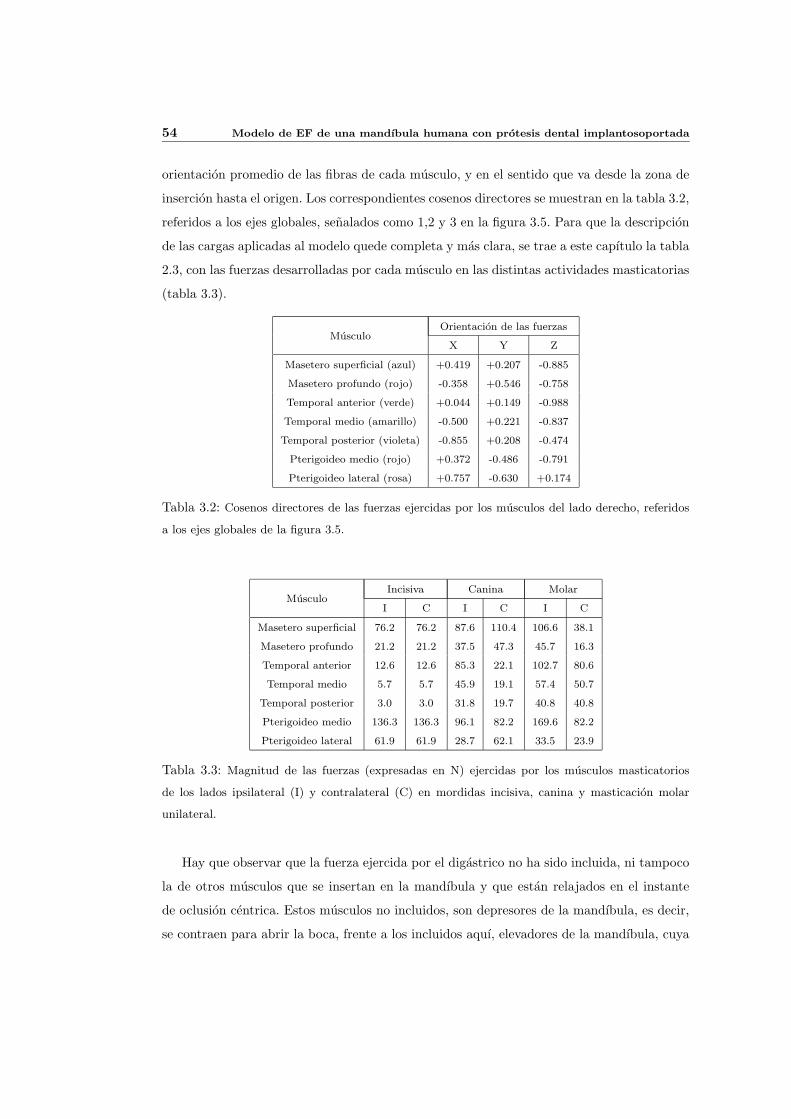
54 Modelo de EF de una mandıbula humana con protesis dental implantosoportada
orientacion promedio de las fibras de cada musculo, y en el sentido que va desde la zona de
insercion hasta el origen. Los correspondientes cosenos directores se muestran en la tabla 3.2,
referidos a los ejes globales, senalados como 1,2 y 3 en la figura 3.5. Para que la descripcion
de las cargas aplicadas al modelo quede completa y mas clara, se trae a este capıtulo la tabla
2.3, con las fuerzas desarrolladas por cada musculo en las distintas actividades masticatorias
(tabla 3.3).
MusculoOrientacion de las fuerzas
X Y Z
Masetero superficial (azul) +0.419 +0.207 -0.885
Masetero profundo (rojo) -0.358 +0.546 -0.758
Temporal anterior (verde) +0.044 +0.149 -0.988
Temporal medio (amarillo) -0.500 +0.221 -0.837
Temporal posterior (violeta) -0.855 +0.208 -0.474
Pterigoideo medio (rojo) +0.372 -0.486 -0.791
Pterigoideo lateral (rosa) +0.757 -0.630 +0.174
Tabla 3.2: Cosenos directores de las fuerzas ejercidas por los musculos del lado derecho, referidos
a los ejes globales de la figura 3.5.
MusculoIncisiva Canina Molar
I C I C I C
Masetero superficial 76.2 76.2 87.6 110.4 106.6 38.1
Masetero profundo 21.2 21.2 37.5 47.3 45.7 16.3
Temporal anterior 12.6 12.6 85.3 22.1 102.7 80.6
Temporal medio 5.7 5.7 45.9 19.1 57.4 50.7
Temporal posterior 3.0 3.0 31.8 19.7 40.8 40.8
Pterigoideo medio 136.3 136.3 96.1 82.2 169.6 82.2
Pterigoideo lateral 61.9 61.9 28.7 62.1 33.5 23.9
Tabla 3.3: Magnitud de las fuerzas (expresadas en N) ejercidas por los musculos masticatorios
de los lados ipsilateral (I) y contralateral (C) en mordidas incisiva, canina y masticacion molar
unilateral.
Hay que observar que la fuerza ejercida por el digastrico no ha sido incluida, ni tampoco
la de otros musculos que se insertan en la mandıbula y que estan relajados en el instante
de oclusion centrica. Estos musculos no incluidos, son depresores de la mandıbula, es decir,
se contraen para abrir la boca, frente a los incluidos aquı, elevadores de la mandıbula, cuya

3.3 Simulacion de la actividad masticatoria 55
actividad es maxima cuando se cierra la boca.
La segunda simplificacion sera no considerar ninguno de los problemas de contacto in-
volucrados: entre dientes y alimento, y en la articulacion temporomandibular, entre el condilo
y la cavidad articular del hueso temporal. En su lugar, se impiden los desplazamientos de
los nodos que entran en contacto, en las direcciones en las que las fuerzas de contacto son
apreciables, simulando ası dichas fuerzas como reacciones.
En el caso de la articulacion, el contacto no se establece directamente entre condilo y
cavidad articular, sino por medio del disco articular que se interpone entre ambos. Este tiene
una rigidez a compresion que varıa con la deformacion, siendo muy pequena al principio y
muy alta a partir de un cierto punto, cuando el disco se haya comprimido aproximadamente
0.5mm, segun un estudio de Chin et al. [63]. A partir de ahı la carga se transmite comple-
tamente a la cavidad articular sin que el disco se siga deformando. El hueso temporal es
muy rıgido, sobre todo en la region de la articulacion temporomandibular. Todo lo anterior
hace que tenga sentido la simplificacion adoptada aquı de impedir los desplazamientos en la
region articular del disco para simular el problema de contacto de la articulacion.
Un argumento similar puede usarse para justificar la simplificacion del contacto entre
los dientes y el alimento. Cuando la boca esta cerrada y se esta ejerciendo la maxima
fuerza muscular el alimento esta completamente comprimido y las fuerzas masticatorias se
transmiten a los dientes del maxilar superior. Se considera aquı que el maxilar superior
es muy rıgido y por tanto son nulos los desplazamientos de los nodos en contacto con el
alimento una vez que este se haya comprimido totalmente.
Mordidas incisiva y canina
Tanto en la mordida incisiva como en la canina, la boca permanece cerrada y la accion
de los musculos masticatorios empuja a los condilos, que estan en su posicion mas atrasada,
contra la superficie articular de la cavidad glenoidea. Se impiden todos los desplazamientos
de los nodos situados en la zona articular del disco para simular las fuerzas de contacto en
la articulacion. Las fuerzas masticatorias con las que los dientes cortan el alimento tienen
una componente vertical predominante y una componente transversal, perpendicular al eje
de la mandıbula, mas pequena. Esta ultima aparece por dos razones:
a) En el instante del corte se producen unos movimientos oscilatorios en direccion
transversal hasta que el alimento se rompe por cizalladura.

56 Modelo de EF de una mandıbula humana con protesis dental implantosoportada
b) Es usual ayudar al corte del alimento tirando de este hacia fuera con la mano.
Para simular estas fuerzas masticatorias, los desplazamientos en ambas direcciones, ver-
tical y transversal, son impedidos en los bordes correspondientes. En la mordida incisiva se
impiden los nodos de los bordes incisales de los cuatro incisivos inferiores, mientras que en la
mordida canina se impiden los nodos de los bordes incisales del segundo incisivo, del canino
y los bordes oclusales del primer premolar, del lado correspondiente.
Masticacion molar
Durante la masticacion, la boca esta continuamente abriendose y cerrandose y por tanto,
los condilos, deslizando sobre la cara anterior de la cavidad glenoidea hacia delante y hacia
atras, respectivamente [160]. El instante que se simula aquı corresponde al de maxima inter-
cuspidacion, tambien llamado oclusion centrica. En el la boca esta cerrada con los dientes
superiores enfrentados a los inferiores y los condilos en su posicion mas atrasada.
En los instantes previos a la oclusion centrica la mandıbula gira para terminar de cerrar
la boca y el condilo desliza hacia atras sobre el disco articular, al que a la vez empuja contra
la eminencia articular. Este problema de contacto con deslizamiento se sustituye por un
problema estatico sin contacto en el que se impiden los desplazamientos de los nodos en
la zona articular en direccion normal a dicho contacto y se permiten los desplazamientos
en direccion tangencial (ver figura 3.7). Con esto, se asegura la existencia de una reaccion
normal, pero es nula la reaccion tangencial, despreciando ası el rozamiento entre el disco
articular y el condilo. Tambien se permiten los desplazamientos en direccion lateral (normal
al plano del dibujo en la figura 3.7) y la normal se supone que forma 30 con la horizontal.
En los molares existe un ligero movimiento en direccion transversal al final de la fase I
(ver capıtulo 2), justo antes de llegar a la oclusion centrica. El alimento ofrece cierta resisten-
cia a ser deformado en esa direccion, resistencia que es vencida por el movimiento relativo de
los molares inferiores y superiores. Para simular esta componente de la fuerza masticatoria
se impiden los desplazamientos transversales en la cara oclusal del molar correspondiente.
Con todo, la componente principal de la fuerza masticatoria es la vertical, la que tritura el
alimento. Esta fuerza vertical aparece como reaccion, al impedir los desplazamientos verti-
cales de los nodos situados en la cara oclusal de los molares. Se simulan aquı masticaciones
unilaterales con el primer y segundo molar, de ambos lados, lo que conduce a cuatro casos de
carga diferentes. Sin embargo, en los datos de actividad muscular, tomados de Nelson [249],
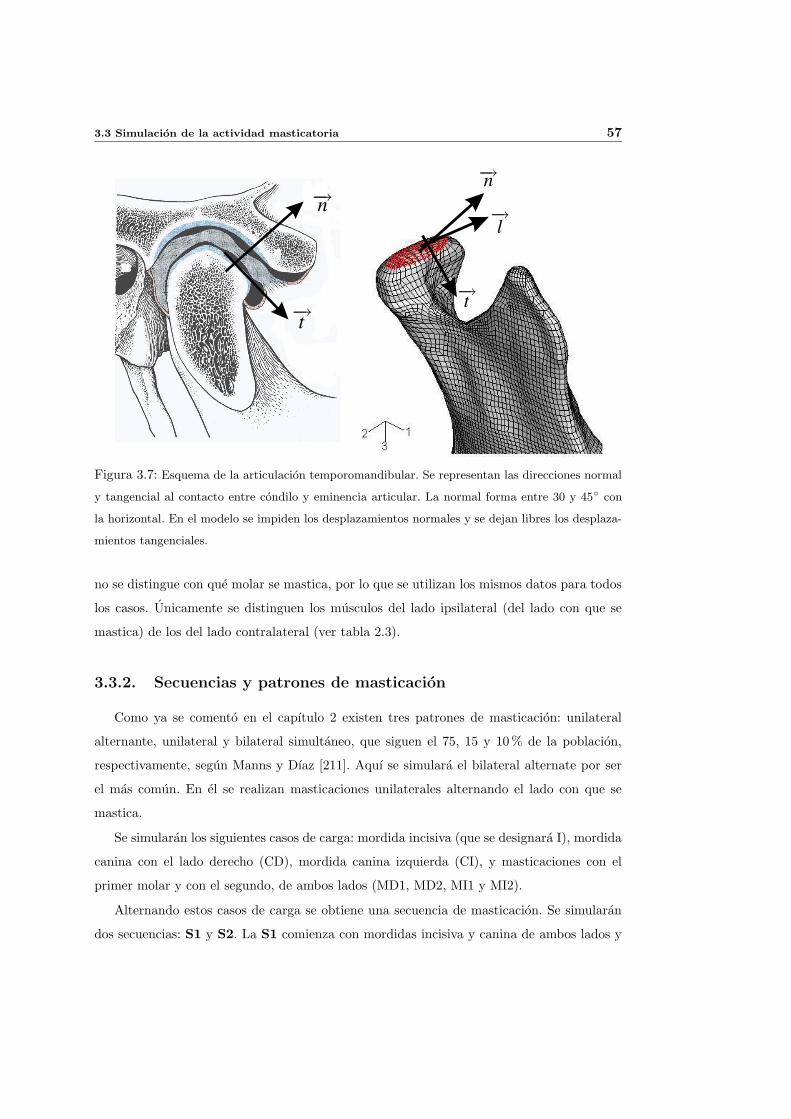
3.3 Simulacion de la actividad masticatoria 57
−→n
−→t
1
−→n
−→t
−→
l
1
Figura 3.7: Esquema de la articulacion temporomandibular. Se representan las direcciones normal
y tangencial al contacto entre condilo y eminencia articular. La normal forma entre 30 y 45 con
la horizontal. En el modelo se impiden los desplazamientos normales y se dejan libres los desplaza-
mientos tangenciales.
no se distingue con que molar se mastica, por lo que se utilizan los mismos datos para todos
los casos. Unicamente se distinguen los musculos del lado ipsilateral (del lado con que se
mastica) de los del lado contralateral (ver tabla 2.3).
3.3.2. Secuencias y patrones de masticacion
Como ya se comento en el capıtulo 2 existen tres patrones de masticacion: unilateral
alternante, unilateral y bilateral simultaneo, que siguen el 75, 15 y 10 % de la poblacion,
respectivamente, segun Manns y Dıaz [211]. Aquı se simulara el bilateral alternate por ser
el mas comun. En el se realizan masticaciones unilaterales alternando el lado con que se
mastica.
Se simularan los siguientes casos de carga: mordida incisiva (que se designara I), mordida
canina con el lado derecho (CD), mordida canina izquierda (CI), y masticaciones con el
primer molar y con el segundo, de ambos lados (MD1, MD2, MI1 y MI2).
Alternando estos casos de carga se obtiene una secuencia de masticacion. Se simularan
dos secuencias: S1 y S2. La S1 comienza con mordidas incisiva y canina de ambos lados y

58 Modelo de EF de una mandıbula humana con protesis dental implantosoportada
sigue con una serie de masticaciones unilaterales alternadas: I-CD-CI-MD1-MI1-MD2-MI2-
MD1-MI1-MD2-MI2-MD1-MI1-MD2-MI2. La secuencia comprende 15 casos de carga y se
repite indefinidamente para simular la actividad masticatoria del individuo.
La secuencia S2 comprende cuatro casos de carga, MD1-MI1-MD2-MI2, que nuevamente
se repiten de manera indefinida. En ella se han suprimido las mordidas por no ser muy
comunes con los habitos alimenticios actuales.
En la primera fase del analisis se utiliza el modelo de EF de denticion completa con las
cargas descritas anteriormente para estimar las propiedades elasticas de toda la mandıbula.
Para ello se usa el modelo de remodelacion osea fenomenologico desarrollado por Doblare y
Garcıa [91, 115]. Los resultados de esta primera fase del analisis se muestran en el capıtulo
6.
3.4. Modelo implantado
El primer molar se ha sustituido por una protesis dental, soportada por un unico implante
osteointegrado. El implante usado aquı no corresponde a ningun tipo comercial, aunque es
muy parecido al tipo IMZ r©, desarrollado por Kirsch [182].
Este implante, fabricado en titanio, es cilındrico, no roscado pero con la superficie rugosa
debido al recubrimiento de spray de plasma de titanio, que se utiliza para mejorar la osteoin-
tegracion. Su apice tiene forma semiesferica y esta ahuecado, de forma que cuatro vastagos
lo unen al resto del implante (ver figura 3.8). Este diseno pretende facilitar el crecimiento de
hueso en el hueco, para mejorar el anclaje del implante, si bien no parece facil que se pueda
conseguir, dado el nivel de cargas relativamente bajo que existe en esa zona, como se vera en
las simulaciones. Es precisamente en esta zona mas compleja donde se ha simplificado la
geometrıa del implante. El que se ha usado en esta tesis, no tiene ese vaciado y consiste
simplemente en un cilindro terminado en una cabeza semiesferica. Se ha hecho ası porque la
zona de interes es el cuello del implante, donde tiene lugar preferentemente la transferencia
de cargas y la perdida osea [256].
No es tan importante el apice, donde se ha simplificado la geometrıa, ya que ahı el nivel
de cargas es menor, como se ha dicho. Ademas, la parte de la carga que se transfiere aquı lo
hace por compresion en la superficie de la semiesfera, por lo que el vaciado que posee el
implante IMZ r©tiene poca influencia. En la figura 3.9 se muestra la malla de elementos
finitos del implante empleado aquı, para el que se usaron elementos lineales de 8 nodos, del

3.4 Modelo implantado 59
Figura 3.8: Cuatro implantes del tipo IMZ r©de diferentes longitudes.
tipo C3D8, al igual que en el resto de la mandıbula. El diametro del implante es 4 mm y
su longitud 17 mm, de las cuales 12 estan dentro del hueso y el resto queda por encima y
corresponden a las piezas de conexion entre el implante y la protesis.
Para simular la colocacion del implante se partio del modelo con denticion completa, con
las propiedades estimadas mediante dicho modelo y se elimino el primer molar derecho, su
ligamento periodontal y parte del hueso que lo rodea. En el hueco se coloco el implante y un
hueso periimplantario nuevo, sustituyendo al que se habıa eliminado. Este hueso periimplan-
tario tiene unas propiedades iniciales supuestas y uniformes en toda la region que ocupa,
hipotesis que precisa siquiera unas lıneas para su justificacion. Estimar con un mınimo grado
de aproximacion las propiedades elasticas del hueso periimplantario requiere de la simulacion
de una serie de mecanismos tan complejos que cada uno de ellos podrıa dar lugar a un estu-
dio tan amplio como el desarrollado en esta tesis. Ademas, hay que tener en cuenta que el
implante se coloca allı donde se ha perdido un diente natural. Por lo general, si ha pasado
cierto tiempo desde la perdida del diente, se produce remodelacion externa que rellena el
hueco dejado por el diente de un hueso mas o menos poroso. La fotografıa de la figura 3.10
muestra un detalle de la mandıbula usada en esta tesis y en concreto, del hueco dejado por el
primer molar derecho que se ha rellenado de un hueso de porosidad intermedia. El individuo

60 Modelo de EF de una mandıbula humana con protesis dental implantosoportada
Figura 3.9: Malla de elementos finitos del implante.
perdio el primer molar derecho, bastante tiempo antes de morir, suficiente como para tapar
el hueco. La simulacion del proceso de relleno del hueco permitirıa estimar las propiedades
de ese hueso como se hace para el resto de la mandıbula en la primera fase del analisis, pero
requerirıa considerar la remodelacion osea externa, objetivo no planteado en esta tesis. Se
supone que el proceso de relleno solo ha modificado el hueso en las proximidades del hueco
y que lejos de el se pueden usar las propiedades obtenidas en la primera fase del analisis,
con la dentadura completa.
De todas formas, aunque se hubiera simulado el proceso de relleno del hueco, hay que
tener en cuenta que el proceso quirurgico de implantacion afecta al hueso de manera im-
portante. Durante el postoperatorio ocurren una serie de procesos mas o menos complejos.
Como se ven afectadas las propiedades elasticas en este postoperatorio es un problema que
no se ha investigado aquı y que requiere un estudio muy exhaustivo. Dada la dificultad de
abordar el problema con todo rigor, se ha optado por suponer una distribucion de densidad y
propiedades elasticas inicial razonable y lo mas sencilla posible, esto es, uniforme e isotropa,
esperando que el modelo de remodelacion corrija esta inexactitud.
En la figura 3.11a se muestra un detalle del modelo de denticion completa con los ele-
mentos eliminados para la implantacion y en la figura 3.11b el hueso periimplantario y el
implante que lo sustituyen en el modelo implantado.
Con el modelo implantado y aplicando las cargas de masticacion descritas anteriormente
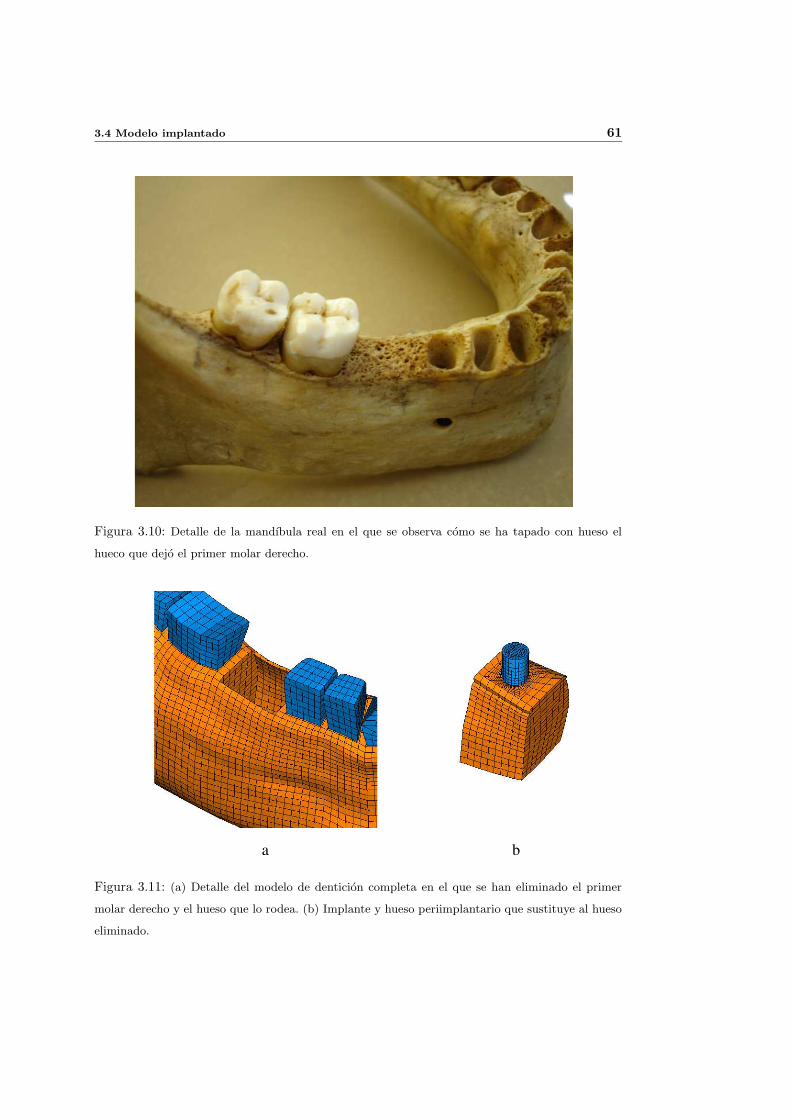
3.4 Modelo implantado 61
Figura 3.10: Detalle de la mandıbula real en el que se observa como se ha tapado con hueso el
hueco que dejo el primer molar derecho.
a b
1
Figura 3.11: (a) Detalle del modelo de denticion completa en el que se han eliminado el primer
molar derecho y el hueso que lo rodea. (b) Implante y hueso periimplantario que sustituye al hueso
eliminado.

62 Modelo de EF de una mandıbula humana con protesis dental implantosoportada
se simulara la evolucion del tejido periimplantario, cuya integridad es vital para el exito del
implante. Para este hueso se utiliza el modelo mecanobiologico de remodelacion osea desar-
rollado en esta tesis (ver capıtulo 7), mientras que para el resto del hueso, que permanece del
modelo de denticion completa, se continua usando el modelo de remodelacion de Doblare y
Garcıa [91,115].
3.5. Osteointegracion. Contacto en la interfase implante-
hueso
Despues de colocar el implante, durante el perıodo de cicatrizacion, y si las condiciones
son las adecuadas, se produce la union entre el tejido oseo y el implante. Esta union se de-
nomina osteointegracion y fue definida por Branemark como la “conexion directa de caracter
estructural y funcional, entre un hueso vivo y un implante soporte de carga” [37]. Fısica-
mente se simula en el modelo de EF como una union entre los elementos del implante y los
del hueso periimplantario.
3.5.1. Aspectos biologicos de la osteointegracion
Quiza sea este el momento de detenerse en analizar a grandes rasgos como es el proceso
de osteointegracion. En la tabla 3.4 se muestran las 4 fases en que lo divide Roberts [287] y
el perıodo aproximado en el que ocurren. En la figura 3.12 se muestra un esquema de dicho
proceso.
Fase Perıodo en el que transcurre
Fase 1: Formacion del callo semanas 0-6
Fase 2: Compactacion laminar del callo semanas 6-18
Fase 3: Remodelacion de la interfase semanas 6-18
Fase 4: Maduracion del hueso compacto semanas 18-54
Tabla 3.4: Cronologıa del desarrollo de la interfase en implantes endooseos insertados en hueso
cortical [287].
Para empezar, la fase de cicatrizacion comienza con la formacion de un callo oseo [287],
como sucede con cualquier tipo de fractura. Para que el callo de union pueda reducir eficaz-
mente un defecto oseo, se requiere la estabilidad relativa de los segmentos en aproximacion.

3.5 Osteointegracion. Contacto en la interfase implante-hueso 63
Por lo general, se prescribe un perıodo de cicatrizacion sin cargas (tecnica de insercion en
dos tiempos) para evitar una excesiva movilidad funcional durante la cicatrizacion. Posteri-
ormente, en la fase 2, el callo se remodela reduciendo su tamano y aumentando su rigidez,
en el proceso de compactacion laminar. El problema de crecimiento y remodelacion del callo
oseo es muy complejo y ha sido simulado numericamente por algunos autores [116,126], pero
no se tendra en cuenta aquı. Pero ademas, mientras el callo se esta compactando, comienza
el proceso de osteointegracion, con la remodelacion de la interfase de union entre hueso e
implante (como puede verse en la tabla 3.4, las fases 2 y 3 se solapan en el tiempo). Tras la
operacion muere aproximadamente 1mm de hueso compacto junto a la herida osea, aunque
se utilice una tecnica quirurgica optima. Esta interfase desvitalizada se remodela por la ac-
cion de BMUs1 que avanzan paralelamente al eje del implante. Estas BMUs avanzan a una
cierta distancia de la interfase, pero los cambios que ocurren mas cerca de la interfase no
son muy bien conocidos. Por ultimo en la fase 4, se produce la maduracion del hueso cir-
cundante que se adapta a los cambios mecanicos que produce la nueva forma de transferirse
las cargas. Es precisamente esta fase 4 la que se simula con el modelo de remodelacion osea
aquı propuesto.
Es interesante observar que el callo oseo hace que aumente temporalmente la seccion de
la capa de hueso compacto sobre la que se sustenta el implante. Este recrecimiento no se
ha considerado, al no tenerse en cuenta el proceso de crecimiento del callo. Por otro lado,
el hueso que se forma en la fase 2, por la compactacion del callo es un hueso primario,
practicamente isotropo [116, 126]. Esto justifica la suposicion que se hace en el capıtulo 8
para el hueso periimplantario, que se toma como inicialmente isotropo.
La osteointegracion se puede perder si las tensiones en la interfase son altas. La resistencia
de la union depende en gran medida de la superficie del implante. Ası, en superficies de
implante pulidas se han medido resistencias a cortante de la union entre 2 y 6 MPa [39,324].
La resistencia es superior si la superficie es rugosa, hasta 26 MPa segun Han et al. [133],
ya que la rugosidad mejora la ostoeintegracion. Con recubrimiento de hidroxiapatita se han
medido resistencias de la union de 34.3 MPa [183] y si la rugosidad se consigue con un
recubrimiento de spray de plasma de titanio, como el que presenta el implante IMZ r©, la
resistencia es de 25 MPa segun Steinemann et al. [313].
1En el capıtulo 4 se describe de forma detallada como se produce la remodelacion y que son las BMUs.
Se puede adelantar que son asociaciones temporales de celulas que renuevan el tejido oseo avanzando en una
cierta direccion.

64 Modelo de EF de una mandıbula humana con protesis dental implantosoportada
Hueso compacto originalTCP
TCP
BMU
Callo
Callo
Fase 1 Fase 2
Fase 3 Fase 4
1
Figura 3.12: Esquema de las fases de que consta el perıodo de cicatrizacion y estabilizacion de
implantes de titanio recubiertos de fosfato tricalcico (TCP). Tomado de Roberts et al. [287]

3.5 Osteointegracion. Contacto en la interfase implante-hueso 65
La superficie de los implantes IMZ r©es pulida en el cuello, con un aspecto brillante como
se puede observar en la figura 3.8, y rugosa en el resto. La zona del cuello del implante es
precisamente la que presenta tensiones mas altas [256], por lo que parece logico suponer
que la osteointegracion se perdera en la zona pulida en cuanto el implante entre en carga,
dada la pequena resistencia a cortante de dicha union. El cuello del implante modelado
aquı se considera pulido, hasta una profundidad de 1.7 mm y rugoso a partir de ahı, por
un recubrimiento similar al del implante IMZ r©. De esta forma, en todo el implante se va a
considerar una osteointegracion perfecta (hueso e implante comparten nodos en la interfase),
excepto en el cuello, en el que la osteointegracion se ha perdido por completo y por tanto es
preciso modelar el contacto entre las superficies de hueso e implante.
3.5.2. Simulacion numerica de la osteointegracion
Una osteointegracion completa se simula en el modelo de EF como una union rıgida entre
los elementos del implante y los del hueso periimplantario, que en la interfase comparten los
nodos. Si la osteointegracion se pierde, como puede ocurrir en la zona pulida, se supondra que
ocurre una separacion total de las caras de la interfase, que a partir de entonces solo pueden
transmitirse esfuerzos de compresion, y de cortadura hasta un cierto valor, es decir, contacto
unilateral estandar. No se simulan fenomenos tales como la degradacion de la union o la
acumulacion de dano, que se han simulado en otros tipos de protesis cementadas [267], ni
una posterior osteointegracion: o bien la union es perfecta, o bien no hay union, mas que de
contacto. Ademas, una zona que esta perfectamente osteointegrada no va a perder la union
y tampoco se puede osteointegrar una zona que este inicialmente separada.
En caso de falta de osteointegracion, hay que simular el contacto entre la superficie
del implante y la superficie del hueso no osteointegrado. Lo normal es usar un modelo de
contacto unilateral estandar: cuando las superficies entran en contacto (la distancia entre
las superficies se hace cero, d = 0 en la figura 3.14), cualquier presion puede transmitirse
entre ambas; las superficies se separan cuando la presion se anula (d > 0) y no se permite
la penetracion de una superficie dentro de otra (d < 0).
Pero en un caso como este, en el que una superficie es mucho mas rıgida que la otra, el
algoritmo anterior puede dar problemas de tipo numerico. El titanio es mucho mas rıgido
que el hueso y mas aun cuando el hueso de la interfase va acumulando dano y disminuyendo
su rigidez. Es ventajoso en estos casos relajar la restriccion impuesta por la ley de la figura

66 Modelo de EF de una mandıbula humana con protesis dental implantosoportada
p
d
Interpenetracion(d < 0): no permitida
Contacto (d = 0): cualquierpresion es posible
Separacion (d > 0): presion nula
1
Figura 3.13: Relacion presion - distancia entre las superficies (p − d) en el modelo de contacto
estandar.
3.13 y suavizar un tanto dicha curva. Se utiliza el algoritmo de contacto suavizado, que se
esquematiza en la figura 3.14. En este se permite una pequena penetracion y se aumenta
progresivamente la presion transmitida con la penetracion, en concreto de manera exponen-
cial. Se admite tambien que existe una pequena presion, antes incluso de que se llegue al
contacto, para que la variacion de la presion sea suave.
En la practica, una de las superficies de contacto se elige como “maestra” y la otra como
“esclava”. Se impide a los nodos de la superficie esclava penetrar dentro de la superficie
maestra (ver figura 3.15), mientras que los nodos de la superficie maestra sı pueden, en
cambio, penetrar dentro de la superficie esclava. La restriccion d = 0 entre los nodos esclavos
y la superficie maestra se impone mediante multiplicadores de Lagrange.
Lo normal es designar como superficie maestra a la superficie mas rıgida de las dos.
Ası pues, la superficie esclava sera la correspondiente al hueso y la superficie maestra la del
implante. En la figura 3.16 se muestra una seccion del modelo de EF en la zona del contacto.
En rojo se muestran los elementos del implante sobre los que se apoya la superficie maestra,
y en azul los elementos del hueso sobre los que se apoya la superficie esclava. Todos los nodos
de la interfase hueso-implante desde el senalado como n1 hacia abajo, son compartidos por
ambos materiales, simulando ası una osteointegracion perfecta. El nodo n2 de la superficie
esclava no puede penetrar dentro de la superficie roja, maestra.
Entre las dos superficies existe friccion que se simula mediante un modelo de rozamiento
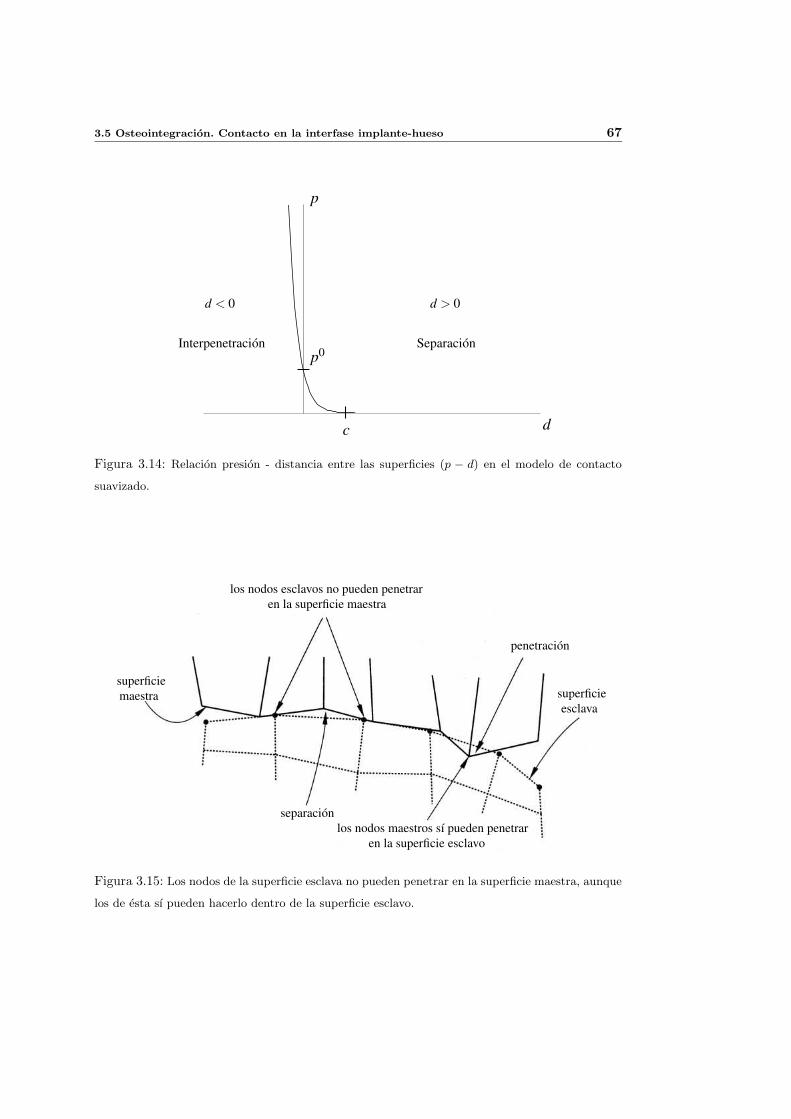
3.5 Osteointegracion. Contacto en la interfase implante-hueso 67
c
p0
p
d
d < 0
Interpenetracion
d > 0
Separacion
1
Figura 3.14: Relacion presion - distancia entre las superficies (p − d) en el modelo de contacto
suavizado.
superficiemaestra
los nodos esclavos no pueden penetraren la superficie maestra
separacionlos nodos maestros sı pueden penetrar
en la superficie esclavo
superficieesclava
penetracion
1
Figura 3.15: Los nodos de la superficie esclava no pueden penetrar en la superficie maestra, aunque
los de esta sı pueden hacerlo dentro de la superficie esclavo.

68 Modelo de EF de una mandıbula humana con protesis dental implantosoportada
n1n2
1
Figura 3.16: Seccion del modelo de EF en la zona del contacto hueso-implante. Se muestran en
rojo los elementos sobre los que se apoya la superficie maestra y en azul aquellos sobre los que se
apoya la superficie esclavo.
de Coulomb: ambas superficies pueden soportar tensiones tangenciales en la superficie de
contacto hasta un cierto valor, τcrit, sin que haya deslizamiento entre ambas. Si se alcanza
este valor crıtico comienza el deslizamiento, manteniendose constante la tension tangencial,
τcrit, que ademas es proporcional a la presion de contacto (τcrit = µ p). El coeficiente de
friccion, µ, usado aquı es de 0.4, igual al que usan Genna et al. [120] entre un implante
dental de titanio y el hueso periimplantario. En otros estudios numericos con implantes de
titanio en diferentes huesos dan valores de µ muy parecidos: entre 0.2 y 0.5 [125,169,268].
Para comparar el efecto que tiene en los implantes de tipo IMZ r©la perdida de osteoin-
tegracion en la zona pulida del cuello, se ha simulado otro tipo de implante con la misma
geometrıa pero recubierto de spray de plasma de titanio en toda su superficie. Para este caso
se supondra que la osteointegracion es perfecta en toda la interfase y no se va a romper al
entrar en carga, por lo que desaparece el problema de contacto.
3.6. Submodelo
Como ya se ha dicho, en el modelo implantado se utilizan dos modelos de remodelacion
osea: uno mecanobiologico, desarrollado en esta tesis, para el hueso periimplantario, y otro

3.6 Submodelo 69
fenomenologico, desarrollado por Doblare y Garcıa [91, 117], para el resto del hueso, que
permanece desde el modelo de denticion completa. Esto es ası por dos motivos. En primer
lugar porque los cambios que sufre el hueso por la colocacion del implante se limitan al
tejido periimplantario. En segundo lugar, por el elevado coste computacional del modelo
aquı desarrollado.
Esa misma razon ha llevado a reducir el numero de grados de libertad del modelo de
EF. Dicha reduccion, sin embargo, no afecta al tamano ni al orden de aproximacion de los
elementos, que siguen siendo del tipo C3D8. Lo que se hace en su lugar es analizar solo una
parte de la mandıbula, que se denominara submodelo. Este es el resultado de cortar el modelo
completo en torno a la region del implante, en dos secciones que se designaran fronteras de
corte. En la figura 3.17 se observa la malla de EF del submodelo, con las fronteras de corte
mesial (FC1) y distal (FC2).
FC1
FC2
2 premolar
2 molar
1
Figura 3.17: Malla del submodelo. Se muestran las fronteras de corte, FC1 y FC2.
La tecnica de submodelado consiste en analizar el modelo completo en primer lugar y
calcular los desplazamientos en las fronteras de corte bajo un determinado estado de cargas.
En segundo lugar, se aplican estos desplazamientos como condiciones de contorno en el
submodelo, junto con las cargas y restricciones de desplazamientos que originalmente se
hubieran aplicado sobre el. En virtud de la unicidad de la solucion del problema elastico, las

70 Modelo de EF de una mandıbula humana con protesis dental implantosoportada
tensiones, deformaciones y desplazamientos calculados de esta forma en el submodelo, son
los mismos que se habrıan obtenido al resolver el dominio completo.
Los distintos casos de carga simulados en el modelo de denticion completa proporcionan
desplazamientos en las fronteras de corte que varıan con el tiempo mientras lo hacen las
propiedades del hueso, estimadas en la primera fase del analisis. Sin embargo, cuando se
alcanza la convergencia, estos desplazamientos permanecen inalterables para cada caso de
carga. Se supone que la introduccion del implante no modifica de manera sustancial los de-
splazamientos en las fronteras de corte, si estas estan lo suficientemente alejadas del implante
como para no verse afectadas por la redistribucion de las cargas. Esto permite utilizar los
desplazamientos del modelo de denticion completa como condiciones de contorno del modelo
implantado.
3.7. Secuencias de masticacion en el modelo implantado
Como ya se ha dicho, el proceso de cicatrizacion y osteointegracion es muy complejo
y no se va a simular aquı. Por el contrario, la simulacion comienza una vez que se ha
alcanzado definitivamente la osteointegracion. Siguiendo la cronologıa dada por Roberts [287]
y expuesta con anterioridad, la simulacion comprenderıa la fase 4, de maduracion del hueso
circundante.
Una practica que es muy comun durante el perıodo que sigue a la colocacion del implante
es la de mantener sin carga la zona afectada durante un cierto tiempo (tecnica de insercion
en dos tiempos [238]). Con esta tecnica, en la primera fase quirurgica se coloca el implante en
el hueso y se tapa con un tornillo de cubierta (ver figura 3.18), que impide que el hueso, los
tejidos blandos o cualquier cuerpo extrano invada la zona de conexion hueca que el implante
tiene en su interior. Este tornillo queda tapado por la mucosa y por tanto no recibe presion
directa del alimento. En esta fase de cicatrizacion el implante recibe carga de forma indirecta,
ya que la flexion-torsion de la mandıbula durante la masticacion es inevitable, aunque ni
siquiera se mastique con ese lado.
Tras el perıodo de cicatrizacion, de unos tres a seis meses, dependiendo de si el implante
se coloca en el maxilar superior o en la mandıbula, se procede a la cirugıa de segunda fase,
en la que se coloca la extension transmucosa y el pilar, que sirve para retener la protesis.
En la figura 3.18 se muestran esquematicamente las piezas que se colocan durante cada fase
quirurgica. Tras la colocacion del pilar y la protesis el implante entra en funcionamiento y

3.7 Secuencias de masticacion en el modelo implantado 71
puede recibir carga directamente.
Cuerpo del
implante
Tornillo de cubierta
o de primera fase
Extension transmucosa
o de segunda fase
Pilar
1
Figura 3.18: (a) Piezas que se colocan en la primera fase quirurgica. (b) En la segunda fase se
sustituye el tornillo de cubierta por la extension transmucosa y el pilar. Tomada de Misch [238]
Se simularan dos tipos de carga: una correspondiente al perıodo funcional (secuencia
S3), en la que ya se mastica con el implante; y otra en la que no se mastica con ese lado
de la mandıbula (secuencia S4). Es necesario aclarar que este ultimo caso no pretende
simular el perıodo de cicatrizacion, ya que para ello habrıa que simular tambien el proceso de
osteointegracion, que no se hace en esta tesis, como ya se ha dicho antes (la osteointegracion
se supone ya terminada cuando comienza la simulacion). En ese caso, ¿cual es el objetivo de
simular la secuencia S4? Hay que observar antes el hecho de que la segunda fase quirurgica,
la de colocacion de la protesis, puede retrasarse como medida de seguridad, a la espera de
que se alcance una osteointegracion definitiva. Sin embargo, dicha osteointegracion puede
haberse alcanzado antes de que se coloque la protesis y, por tanto, antes de que el implante
empiece a recibir carga. Las razones por las que no se mastica con el lado del implante
pueden ser diversas: por recomendacion medica, hasta la colocacion de la protesis; por una
decision voluntaria del paciente, que a pesar de tener ya la protesis sigue sin masticar con
ese lado, por falta de confianza; o incluso por un acto involuntario, ya que el paciente ha

72 Modelo de EF de una mandıbula humana con protesis dental implantosoportada
cambiado sus habitos masticatorios tras la perdida del diente natural. ¿Que ocurre cuando
no se mastica con el lado del implante, a pesar de haberse conseguido ya la osteointegracion?.
La influencia que tiene ese comportamiento en el hueso periimplantario es lo que se pretende
analizar con la secuencia S4.
La secuencia S3 corresponde a un tipo de masticacion bilateral alternante con la siguiente
sucesion de masticaciones: (MI1-MI2-MImp-MD2)-(MI1-MI2-MImp-MD2)-. . . . En ella se
incluye el caso de carga MImp, que es identico al MD1 descrito anteriormente, pero esta
vez impidiendo los desplazamientos verticales y transversales de la superficie superior del
implante. Por ultimo, la secuencia S4 es una masticacion unilateral con el lado izquierdo,
con la sucesion de cargas (MI1-MI2)-(MI1-MI2)-. . .

Capıtulo 4
Biologıa osea
4.1. Introduccion
El tejido oseo es el tejido conjuntivo que forma los huesos. Tiene la matriz extracelular
mineralizada, lo que lo convierte en el unico tejido conectivo duro. Esta caracterıstica es
fundamental para algunas de las funciones esenciales de los huesos, entre las que destacan:
a) Constituyen un sistema estructural que protege los organos vitales del cuerpo. Por
ejemplo, el craneo, que protege el cerebro, o la caja toracica, que protege el corazon y los
pulmones.
b) Es la estructura soporte del sistema muscular, formando junto a el el aparato loco-
motor.
c) Tiene un papel crucial en la homeostasis del calcio y el fosforo, de los que actua como
reserva, a expensas de lo que dicte el sistema endocrino, que actua como regulador.
d) La medula osea que rellena sus poros es la principal fuente de globulos rojos y blancos
del cuerpo humano.
La forma de los huesos sigue unos estereotipos bastante definidos. Ası, independiente-
mente del animal al que pertenezca, se puede distinguir facilmente una tibia de un femur,
por ejemplo. Tambien resulta inmediato diferenciar el femur de un humano del de cualquier
otro primate. Todo esto parece indicar que los huesos son estructuras bastante estaticas
y definidas. Nada mas lejos de la realidad. Si se pasa a una escala microscopica, se po-
dra comprobar que los huesos son estructuras muy dinamicas, en continua evolucion, que
se adaptan a los cambios fisiologicos y funcionales a los que se les somete. Este proceso de
73

74 Biologıa osea
adaptacion, conocido como remodelacion osea, hace que, dentro de unos margenes, la forma
y propiedades mecanicas de los huesos sea muy variable de un individuo a otro y altamente
dependiente del tipo de actividad que el individuo desarrolla.
A continuacion se describe de forma somera la composicion del tejido oseo y su actividad
fisiologica, que permite entender como ocurre esta remodelacion osea y como esta determina
la forma y propiedades mecanicas del hueso.
4.2. Composicion del hueso
El tejido oseo es un componente vivo, formado por celulas de cuatro tipos (osteoclastos,
osteoblastos, celulas de borde y osteocitos) y una matriz osea extracelular.
4.2.1. Matriz osea
La matriz osea tiene una gran estabilidad, pudiendo permanecer inalterable y mante-
niendo su resistencia durante cientos de anos, despues de muerto el tejido.
Se distinguen en ella una componente inorganica o mineral, que representa aproximada-
mente el 65 % en peso, una componente organica, 20 %, y agua, el 10 % restante [46]. Estos
valores son aproximados y pueden variar de un tipo de hueso a otro (ver figura 4.1).
0 10 20 30 40 50 60 70 80 90 100
Esponjoso
Compactonormal
Compactohipo-mineralizado
%
poro (medula)
agua
mineral
colageno
otras
1
Figura 4.1: Composicion volumetrica aproximada de tres tipos de hueso.
La componente inorganica del hueso es principalmente hidroxiapatita, Ca10(PO4)6(OH)2,
en cristales de simetrıa hexagonal de dimensiones aproximadas 50 x 50 x 400 A. En algunos

4.2 Composicion del hueso 75
cristales hay sustituciones del ion fosfato por iones carbonato, bifosfato, fluoruro, citrato,
etc., segun la composicion de la medula osea circundante, con la que interaccionan. Esta
componente inorganica tiene dos funciones esenciales: confiere al hueso gran parte de su
resistencia y rigidez y sirve de reserva de iones. De hecho, el 90 % del calcio del cuerpo, el
85 % del fosforo y la mitad del sodio y el magnesio se encuentran en el hueso. La resistencia
mecanica que aporta la componente inorganica es principalmente resistencia a compresion,
ya que su resistencia a traccion es muy pequena y su fragilidad alta.
De la resistencia a traccion es responsable la componente organica, que ademas aporta
flexibilidad y le da forma al hueso. Esta constituida por colageno, principalmente de tipo
I, tambien presente en tendones y ligamentos, y en menor medida de tipo V y tipo XII. El
colageno es una proteına estructural que se organiza en fuertes fibras, de gran diametro en
el tipo I. Constituye el 90 % de la componente organica. El 10 % restante incluye proteogli-
canos y proteınas no colagenas. La funcion de los primeros no esta muy clara, pero podrıan
intervenir en el proceso de mineralizacion por su propiedad de fijadores del calcio. Entre
las proteınas no colagenas destacan la osteopontina, la osteonectina y la osteocalcina, esta
ultima segregada por los osteoblastos y con un papel importante en la mineralizacion del
nuevo hueso.
El agua que contiene el hueso se encuentra formando parte de la medula osea que rellena
los poros, o bien contenida en el tejido, en este caso ligada al colageno. El hueso recien
formado, denominado osteoide, esta constituido principalmente de componente organica y
agua, atrapada entre el colageno. Esta es sustituida por mineral de forma progresiva, en el
proceso denominado mineralizacion osea.
4.2.2. Descripcion cuantitativa de la composicion de la matriz osea
A continuacion se definen algunos parametros relacionados con la composicion de la
matriz osea que se utilizaran mas adelante. En primer lugar el volumen de hueso, Vt, se
descompone en volumen de matriz osea, Vb, y volumen de poros, Vv, ocupados por medula
osea:
Vt = Vb + Vv (4.1)
Se define la fraccion volumetrica de hueso, vb, como
vb =VbVt
(4.2)

76 Biologıa osea
y la porosidad, p, complementaria de la anterior,
p =VvVt
= 1− vb. (4.3)
La densidad aparente del hueso, ρb, es:
ρh =ρVb + ρvVv
Vt(4.4)
donde ρ es la densidad aparente de la matriz osea, y ρv la densidad de la medula.
La matriz osea se divide en componente organica, Vo, componente mineral, Vm y agua,
Va,
Vb = Vo + Vm + Va (4.5)
siendo la densidad de la matriz osea
ρ =ρoVo + ρmVm + ρaVa
Vb(4.6)
donde ρo, ρm y ρa son respectivamente las densidades de la componente organica, la com-
ponente mineral y el agua.
Si un especimen de hueso se seca en un horno hasta que se evapora todo el agua, su peso
se denomina “masa seca”, md,
md∼= ρoVo + ρmVm = mo +mm (4.7)
El signo en la anterior ecuacion no es de igualdad estricto porque en el horno se evapora
no solo el agua que ocupa los poros, contenida en la medula osea, sino tambien la que
se encuentra en el tejido, ligada al colageno. Si a continuacion se introduce el especimen
de nuevo en el horno y se mantiene a 800C durante 24 horas, se evapora la componente
organica quedando solo la “masa de ceniza”, mm = ρmVm. Se define la fraccion de ceniza
como la relacion entre la masa de ceniza y la masa seca
α =mm
mm +mo(4.8)
y suele estar en torno a 0.65± 0.03 [220].
4.2.3. Celulas oseas
Acaba de describirse brevemente de que se compone la matriz osea, pero no se ha habla-
do aun de quienes son los responsables de su formacion y mantenimiento. Son las celulas

4.2 Composicion del hueso 77
oseas. Existen cuatro tipos y se pueden dividir en dos categorıas: las que reabsorben hueso
(osteoclastos) y las que forman (osteoblastos) o han formado hueso (osteocitos y celulas de
borde).
Osteoclastos
Son las celulas responsables de la reabsorcion. Los osteoclastos maduros son celulas
multinucleadas que surgen de la fusion de otras, llamadas osteoclastos precursores. Estos se
encuentran en la porcion hemopoyetica de la medula osea y, bajo ciertas condiciones que
se explicaran mas adelante, pueden fusionarse entre sı y formar los osteoclastos, que tienen
de tres a veinte nucleos y gran numero de mitocondrias, que producen energıa suficiente
para la reabsorcion. En la cara del osteoclasto apoyada sobre la matriz osea, que tiene forma
ondulada, baja drasticamente el pH, de 7 a 4 aproximadamente [46], por la secrecion de acidos
que fabrica la propia celula. Este pH tan bajo hace que se disuelva el mineral. Posteriormente
se degrada la componente organica con enzimas, completandose ası la reabsorcion, que ocurre
a una velocidad de decenas de micras por dıa. Una vez completada, los osteoclastos mueren
por apoptosis, muerte celular programada que se conoce tambien como “suicidio celular”.
En la figura 4.2 se puede observar un grupo de osteoclastos reabsorbiendo hueso y creando
lo que se denomina una laguna de Howship, sobre hueso trabecular.
Osteoblastos
Son celulas cuboidales mononucleadas responsables de la formacion de hueso. Su princi-
pal funcion es segregar las proteınas que componen la matriz organica. En concreto, forman
el osteoide, sobre una superficie de hueso antiguo. El osteoide es la capa de tejido oseo que
se forma en primer lugar. Contiene colageno, proteınas no colagenas, proteoglicanos y agua,
pero no contiene mineral. Este aparece posteriormente desplazando al agua durante el pro-
ceso de mineralizacion, proceso en el que tambien intervienen los osteoblastos, controlando
el flujo electrolıtico desde la medula al osteoide. El espesor de la capa de osteoide crece
a una velocidad de 1 µm/dıa aproximadamente. En la figura 4.3 se observa una capa de
osteoblastos formando osteoide sobre la superficie osea.
Sus precursores son las celulas mesenquimales, que tambien son precursoras de las celulas
de grasa. Estas celulas mesenquimales provienen bien de la capa interna de la membrana
del periostio o bien del estroma de la medula osea, dependiendo de la zona donde ocurra
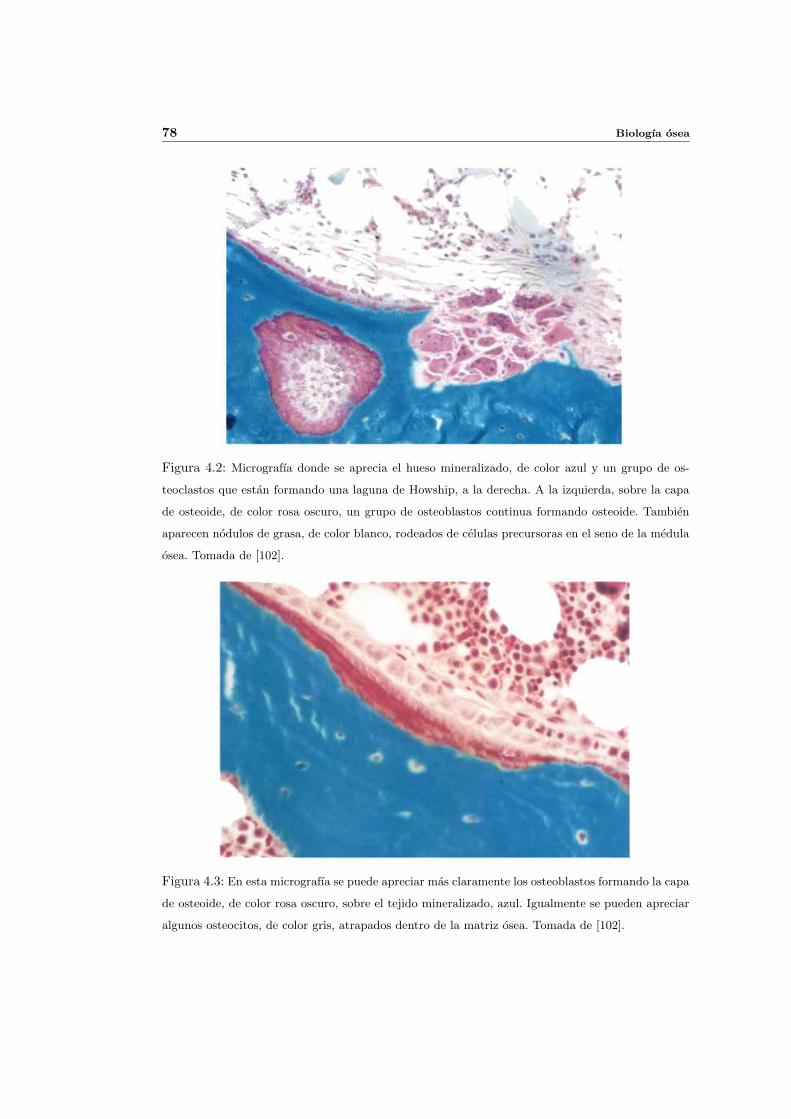
78 Biologıa osea
Figura 4.2: Micrografıa donde se aprecia el hueso mineralizado, de color azul y un grupo de os-
teoclastos que estan formando una laguna de Howship, a la derecha. A la izquierda, sobre la capa
de osteoide, de color rosa oscuro, un grupo de osteoblastos continua formando osteoide. Tambien
aparecen nodulos de grasa, de color blanco, rodeados de celulas precursoras en el seno de la medula
osea. Tomada de [102].
Figura 4.3: En esta micrografıa se puede apreciar mas claramente los osteoblastos formando la capa
de osteoide, de color rosa oscuro, sobre el tejido mineralizado, azul. Igualmente se pueden apreciar
algunos osteocitos, de color gris, atrapados dentro de la matriz osea. Tomada de [102].

4.2 Composicion del hueso 79
la formacion de osteoide. Que estas celulas se diferencien en osteoblastos y no en celulas de
grasa depende de diversos factores, entre ellos el estado tensional del entorno. El proceso de
diferenciacion de los osteoblastos dura entre 2 y 3 dıas.
Otra funcion de los osteoblastos es segregar determinadas sustancias que les permiten
comunicarse con otras celulas y entre ellas el RANK-ligando (RANK-L), que activa la fusion
de osteoclastos precursores. Tambien son receptores de ciertas hormonas como la vitamina
D, el estrogeno o la hormona paratiroidea (PTH).
Una vez que han cumplido su mision de depositar osteoide, al osteoblasto le pueden
suceder tres cosas: que quede encerrado en la matriz osea, en cuyo caso se convierte en un
osteocito; que permanezca sobre la superficie osea, diferenciandose en una celula de borde y
que sufra apoptosis.
Osteocitos
Mas del 90 % de las celulas oseas de una persona adulta son osteocitos. Son osteoblastos
que han quedado atrapados en la matriz osea, en huecos denominados lagunas. Al diferen-
ciarse de los osteoblastos, los osteocitos cambian su forma. Se estiran apareciendo en ellos
unas prolongaciones (ver figura 4.4), que se extienden por estrechos tuneles dentro la matriz
osea, denominados canalıculos (ver figura 4.6). Lagunas y canalıculos forman un complejo
entramado que permite la transferencia de mineral y otras sustancias dentro del hueso y a
los osteocitos comunicarse entre sı y con las celulas de borde.
Estas celulas son las encargadas del mantenimiento del hueso. Para ello, “sienten” la
deformacion mecanica en su entorno y emiten una senal de algun tipo cuando dejan de
sentirla, lo que puede ser un indicio de que existe un problema. Esta senal puede trasmitirse
por la red de canalıculos hasta llegar a las celulas de borde que inician la remodelacion si es
necesario.
Celulas de borde
Son celulas que, como los osteocitos, provienen de osteoblastos que han terminado su
actividad de secrecion de osteoide. A diferencia de aquellos, no quedan encerrados dentro
de la matriz osea, sino que se situan sobre su superficie cubriendola casi por completo y
cambiando su forma a otra mas plana y alargada.
Tienen un papel crucial en la homeostasis del calcio [261], trasfiriendolo de la matriz

80 Biologıa osea
Figura 4.4: Tres osteocitos con conexion entre sus prolongaciones. Aumento x1600. Tomado de
Jee [174].
osea a la medula si la concentracion en sangre desciende. Protegen la matriz osea de diver-
sos agentes quımicos presentes en la sangre que pueden disolver el mineral. Mantienen la
comunicacion con los osteocitos por medio de las prolongaciones de estos ultimos y tienen
receptores de hormonas (estrogeno, PTH) y otros “mensajeros quımicos”, que podrıan ini-
ciar la remodelacion, si la senal recibida de los osteocitos ası lo determina. Segun Miller y
Jee [236] pueden ser reactivadas para formar una capa de osteoblastos.
4.3. Tipos de tejido oseo
Atendiendo a diversos factores se puede dividir el tejido oseo en varios tipos. A contin-
uacion se veran algunos de ellos.
4.3.1. Trabecular y cortical
Esta es sin duda la forma mas comun de distinguir a los tejidos oseos y atiende a la
diferente porosidad de uno y otro tipo, por lo que tambien se suelen denominar esponjoso
y compacto respectivamente [45,219,251]. El hueso cortical o compacto es el hueso de baja

4.3 Tipos de tejido oseo 81
porosidad y el hueso trabecular es el de porosidad alta, y aunque en principio, la porosidad
puede variar entre 0 y 1, es difıcil encontrar un tejido con porosidad intermedia.
El hueso trabecular tiene una porosidad entre 0.75 y 0.95 y se encuentra en los huesos
cortos, en los huesos planos y en los extremos de los huesos largos. La estructura de este
hueso consiste en un conjunto de placas y barras de tejido mineralizado y duro, denominadas
trabeculas, de 200 µm de espesor caracterıstico, conectadas entre sı y que dejan poros,
ocupados por medula osea. La distribucion de estas trabeculas es muy variable, casi ortogonal
en unas ocasiones y aleatoria en otras, pero parece obedecer a un objetivo unico, resistir las
cargas a las que se encuentra sometido el hueso.1
El hueso cortical o compacto tiene una porosidad entre 0.05 y 0.1 y constituye el 80 %
del esqueleto humano. Se encuentra en la parte central de los huesos largos y envolviendo el
hueso trabecular en huesos largos, cortos y planos, a modo de corteza; de ahı su nombre. En
la figura 4.5 se muestra un corte de un hueso largo, en el que se distingue la capa de hueso
cortical (1) rodeando al hueso trabecular (2). La porosidad en el hueso cortical consiste en
huecos de tres tipos:
a) Canales de Havers, (5), aproximadamente paralelos al eje del hueso, de unos 50 µm de
diametro y que contienen nervios y capilares.
b) Canales de Volkmann, (6), mas cortos y dispuestos en direccion transversal, conectan los
canales de Havers entre sı y con la capa exterior del hueso, el periostio, (8). Tambien se
encuentran vascularizados.
c) Cavidades de reabsorcion. Son huecos temporales creados por los osteoclastos durante la
fase inicial de la remodelacion. Tienen un diametro aproximado de 200 µm .
Se denominan osteonas (3 en la figura 4.5) a los sistemas que forman la mayor parte del
hueso cortical. Su apariencia es la de una serie de cilindros enrollados, denominados lamelas,
alrededor de un canal de Havers. En la parte externa del hueso se disponen algunas lamelas
cilındricas concentricas, (7), y cubriendo estas, el periostio, (8), que contiene vasos y nervios,
(9).
Este se conecta a los osteocitos a traves de los canalıculos (ver figura 4.6), por donde
fluyen los nutrientes.
Las osteonas estan delimitadas por las lıneas de cementacion, que son finas capas de
componente organica que aıslan unas osteonas de otras. Cuando la matriz osea se fractura,
1Wolff establecio a finales del siglo XIX que para cumplir este objetivo las trabeculas se alinean con las
direcciones principales de tension, lo que se conoce como “Ley de Wolff” [354].
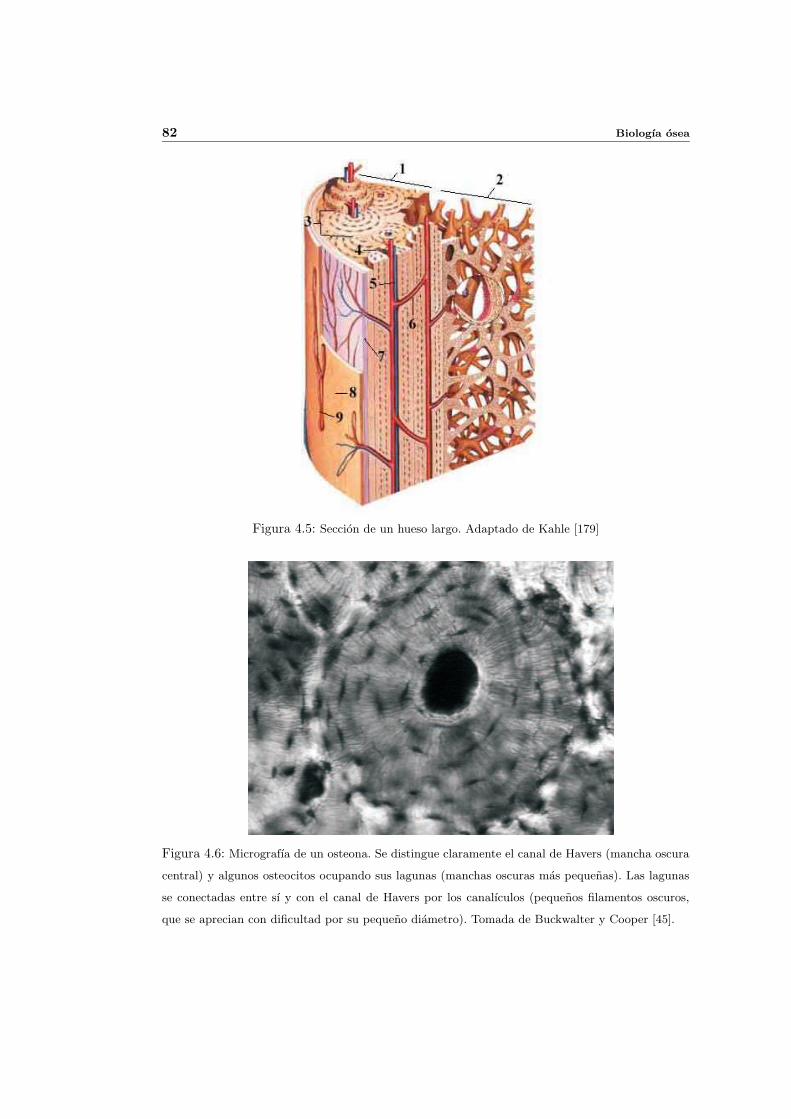
82 Biologıa osea
Figura 4.5: Seccion de un hueso largo. Adaptado de Kahle [179]
Figura 4.6: Micrografıa de un osteona. Se distingue claramente el canal de Havers (mancha oscura
central) y algunos osteocitos ocupando sus lagunas (manchas oscuras mas pequenas). Las lagunas
se conectadas entre sı y con el canal de Havers por los canalıculos (pequenos filamentos oscuros,
que se aprecian con dificultad por su pequeno diametro). Tomada de Buckwalter y Cooper [45].

4.3 Tipos de tejido oseo 83
la lınea de rotura tiende a seguir las lıneas de cementacion sin atravesar la osteona [83], lo
que puede prevenir fallos por fatiga, permitiendo a las celulas oseas reparar el fallo antes de
la fractura completa.
4.3.2. Lamelar y fibroso
Si se examina el hueso cortical y el hueso trabecular mas de cerca, se puede observar que
estan compuestos de dos tipos de tejido:
a) Hueso lamelar, que se forma lentamente y de manera muy organizada. Consiste en
una serie de capas o lamelas, que contienen una matriz anisotropa de cristales de mineral y
fibras de colageno. Cuando las lamelas son cilındricas se tiene una osteona, como se acaba
de ver. Segun Giraud-Guille [124] existen dos tipos de disposicion de las lamelas segun la
orientacion de las fibras de colageno, que discurren paralelas dentro de cada lamela. En la
primera de ellas, las lamelas alternan la orientacion de sus fibras a 90 (ver figura 4.7).
En otras zonas se produce una variacion progresiva de la orientacion de una lamela a otra,
resultando una disposicion helicoidal (ver figura 4.8).
Figura 4.7: Osteona en la que las sucesivas lamelas alternan la direccion de las fibras de colageno
a 90. Tomada de Martin et al. [220].
La presencia de un tipo u otro puede responder a la necesidad de soportar diferentes
estados de cargas, para lo que la disposicion de las fibras de colageno es crucial, dando mas
rigidez al tejido en determinadas direcciones. Ası por ejemplo, la disposicion ortogonal de la
figura 4.7 es bastante adecuada para soportar traccion en direccion longitudinal, mientras que
la disposicion helicoidal serıa mas conveniente si las cargas tienen una orientacion variable.
b) Hueso fibroso o “woven bone”. Se forma de manera rapida y poco organizada, con
los cristales de mineral y las fibras de colageno dispuestas aleatoriamente, lo que hace que

84 Biologıa osea
Figura 4.8: Lamelas sucesivas que cambian la direccion de las fibras de colageno progresivamente
con un aspecto helicoidal. Tomada de Martin et al. [220].
sea menos rıgido y resistente. Para compensar esa falta de rigidez tiene mayor contenido
mineral que el hueso lamelar. En contraposicion al hueso lamelar, en el que la orientacion
de las lamelas determina su caracter anisotropo, el hueso fibroso es practicamente isotropo.
4.3.3. Primario y secundario
El hueso cortical a su vez puede dividirse en primario o secundario segun el estado de
desarrollo en el que se encuentre.
1) El hueso primario es tejido depositado sobre la superficie osea, por ejemplo la del
periostio, durante el perıodo de crecimiento. Se distinguen dos tipos:
a) Hueso lamelar primario o circunferencial, en el que las lamelas son paralelas a la
superficie del hueso. Los vasos sanguıneos se incorporan al tejido rodeados de unas cuantas
lamelas circunferenciales, formando una osteona primaria (ver figura 4.9).
b) Hueso plexiforme. Se forma muy rapidamente con un entramado trabecular de “woven
bone” cuyos huecos se rellenan con hueso lamelar. Los espacios residuales que ocupan los
vasos le dan un aspecto de muro de fabrica [220], como se puede observar en la figura 4.10.
Esta presente en animales con un crecimiento muy rapido, como el ganado vacuno. En el
hombre forma parte del embrion del esqueleto, que va siendo sustituido progresivamente por
hueso mas maduro, y en el callo oseo despues de una fractura. Ası, salvo que exista lesion
osea, no se presenta en el esqueleto humano a partir de los cuatro o cinco anos de edad.

4.3 Tipos de tejido oseo 85
Osteonasprimarias
lamelas paralelas al periostio
vasos sanguıneosperiostio
1
Figura 4.9: Esquema de la estructura de hueso lamelar primario. Tomada de Martin et al. [220].
Figura 4.10: Hueso plexiforme. Tomada de Martin et al. [220].

86 Biologıa osea
2) El hueso secundario resulta de la reabsorcion de tejido existente y su sustitucion
por tejido lamelar, en el proceso de remodelacion osea, que sera descrito con detalle mas
adelante. En el hueso cortical, el tejido viejo es sustituido por osteonas secundarias, como
se observa en el esquema de la figura 4.11. Dichas osteonas secundarias sustituyen bien al
tejido lamelar primario, bien a otras osteonas anteriores, de las que reabsorben una parte y
dejan otra intacta, que se conoce como hueso intersticial. En el hueso trabecular tambien se
sustituye el hueso primario por hueso secundario lamelar, mas rapidamente que en el hueso
compacto, aunque no mediante osteonas, que no cabrıan en una trabecula.
Figura 4.11: Hueso lamelar secundario. Tomada de Martin et al. [220].
4.4. Estructura del hueso
Se pueden distinguir entre tres tipos de hueso segun su forma: largos, planos y cortos.
Se llaman huesos cortos aquellos que no tienen una dimension predominante, como los
huesos del tarso, huesos del carpo, vertebras, etc. Los huesos planos son aquellos que tienen
una dimension mucho mas pequena que las otras dos, como el omoplato y las crestas del
ilion. Normalmente su funcion es protectora y apenas soportan carga, como en el caso del
craneo. Los huesos largos tienen una dimension mucho mayor que las otras dos, como el
femur, la tibia, el humero, los metatarsianos, metacarpianos, las falanges, etc.
Normalmente soportan cargas muy elevadas que pueden ser de distintos tipos: de flexion-
compresion en el caso del femur, flexion-traccion en el caso del humero o flexion predomi-

4.4 Estructura del hueso 87
nantemente en el caso de radio y cubito. La explicacion de por que esas cargas tan altas fue
dada por Borelli en el siglo XVII [34]. El movimiento de los miembros se consigue por la
contraccion de los musculos, que tiran de los huesos. Sin embargo, dicha contraccion es muy
limitada por lo que es necesario amplificarla para conseguir unos movimientos normales.
Para ello, los huesos actuan de palanca, pero no con ganancia mecanica, sino todo lo con-
trario. Ası, para levantar un determinado peso con la mano, el bıceps, que se inserta en el
radio muy cerca del codo, debe ejercer una fuerza unas 9 veces mayor que el peso a levantar.
Estas cargas tan altas condicionan el comportamiento y las propiedades mecanicas del hue-
so, como se vera mas adelante. La estructura interna de los huesos largos esta estrechamente
relacionada con la carga que soportan, mientras que en los huesos planos, como el craneo,
con una mision protectora mas que estructural, la relacion entre carga y estructura interna
no es tan clara.
Todos los huesos estan recubiertos de una membrana dura denominada periostio, excepto
en las regiones proximas a las articulaciones y en las zonas de insercion de tendones y
ligamentos. El periostio tiene dos capas: una externa, densa y fibrosa, con alto contenido
en colageno, y una capa interna vascularizada y con gran actividad celular, que actua como
fuente de osteoblastos.
Los huesos largos tienen una zona tubular de gran espesor denominada diafisis, formada
por hueso cortical, (ver figura 4.12). Esta recubierta por el periostio y separada de la cavidad
medular por una superficie denominada endostio. La diafisis se ensancha en los extremos, a la
vez que disminuye el espesor de la capa de cortical y comienza a aparecer hueso trabecular. A
esta zona del hueso se le denomina metafisis. Despues de esta se encuentra la epıfisis, a la que
se une por una zona cartilaginosa denominada placa epifisaria, cuya osificacion progresiva
produce el crecimiento del hueso en longitud. En el extremo de la epıfisis se encuentra el
cartılago articular, que forma parte de la superficie articular.
La estructura tubular de la diafisis es optima desde el punto de vista resistente, dado
que los esfuerzos que soporta un hueso largo son de flexion principalmente, con una pequena
componente de torsion en algunos casos. Ademas, la direccion de maxima rigidez del tejido
coincide con la direccion longitudinal del hueso. En la epıfisis, sin embargo, el hueso exis-
tente es trabecular sin una direccion preferente clara. En esta zona las cargas estan mas
repartidas, ya que los esfuerzos se transmiten ahı a los huesos vecinos, en una zona mas o
menos amplia. Ademas, esta zona de hueso trabecular permite grandes deformaciones, lo
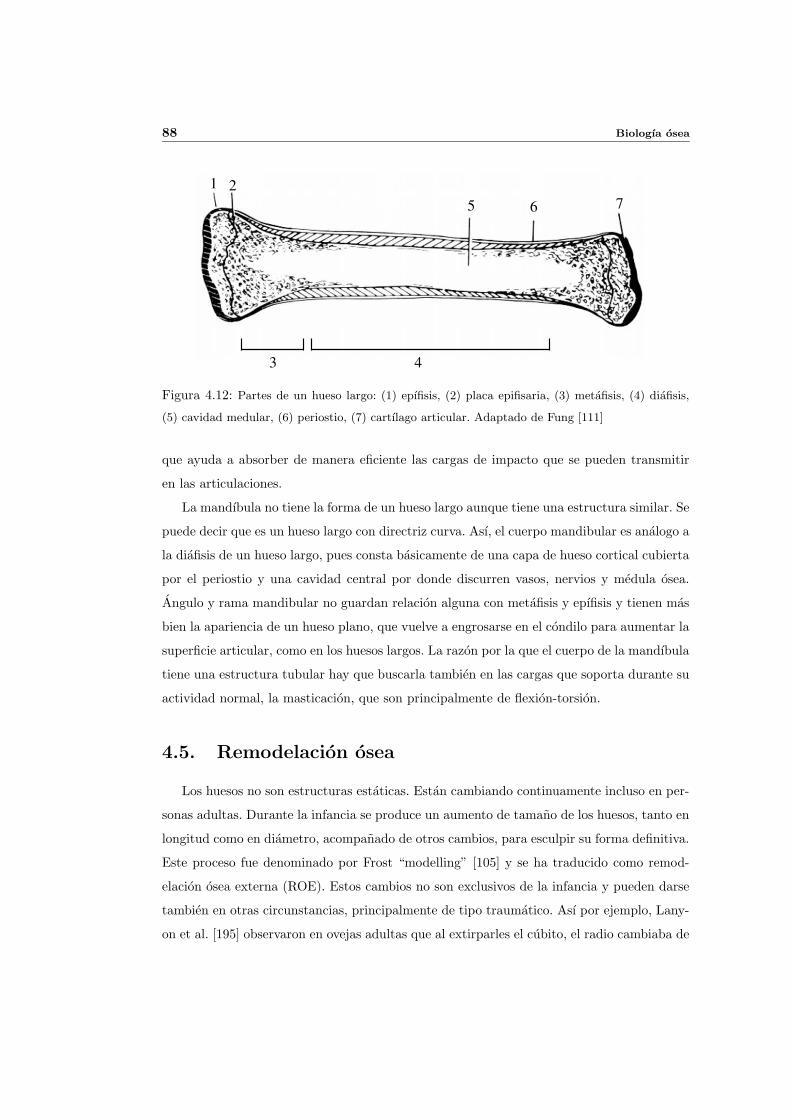
88 Biologıa osea
1 2
3 4
5 6 7
1
Figura 4.12: Partes de un hueso largo: (1) epıfisis, (2) placa epifisaria, (3) metafisis, (4) diafisis,
(5) cavidad medular, (6) periostio, (7) cartılago articular. Adaptado de Fung [111]
que ayuda a absorber de manera eficiente las cargas de impacto que se pueden transmitir
en las articulaciones.
La mandıbula no tiene la forma de un hueso largo aunque tiene una estructura similar. Se
puede decir que es un hueso largo con directriz curva. Ası, el cuerpo mandibular es analogo a
la diafisis de un hueso largo, pues consta basicamente de una capa de hueso cortical cubierta
por el periostio y una cavidad central por donde discurren vasos, nervios y medula osea.
Angulo y rama mandibular no guardan relacion alguna con metafisis y epıfisis y tienen mas
bien la apariencia de un hueso plano, que vuelve a engrosarse en el condilo para aumentar la
superficie articular, como en los huesos largos. La razon por la que el cuerpo de la mandıbula
tiene una estructura tubular hay que buscarla tambien en las cargas que soporta durante su
actividad normal, la masticacion, que son principalmente de flexion-torsion.
4.5. Remodelacion osea
Los huesos no son estructuras estaticas. Estan cambiando continuamente incluso en per-
sonas adultas. Durante la infancia se produce un aumento de tamano de los huesos, tanto en
longitud como en diametro, acompanado de otros cambios, para esculpir su forma definitiva.
Este proceso fue denominado por Frost “modelling” [105] y se ha traducido como remod-
elacion osea externa (ROE). Estos cambios no son exclusivos de la infancia y pueden darse
tambien en otras circunstancias, principalmente de tipo traumatico. Ası por ejemplo, Lany-
on et al. [195] observaron en ovejas adultas que al extirparles el cubito, el radio cambiaba de

4.5 Remodelacion osea 89
forma y se engrosaba en la zona proxima al lugar donde antes estaba el cubito, para compen-
sar en cierta forma su ausencia. Ese cambio de forma conlleva reabsorcion en determinadas
zonas y formacion en otras, que pueden ocurrir de forma independiente.
En una persona adulta que no sufra trauma alguno tambien se producen cambios. El hue-
so esta en continua evolucion, reparando el dano que la actividad diaria pueda producir en el
por fatiga. Tambien se adapta a los cambios que dicha actividad diaria pueda experimentar.
Para ello se retira el hueso antiguo y se forma hueso nuevo, con una actividad coordinada
de osteoclastos y osteoblastos. Frost [105] denomino “remodelling” a este proceso, que se ha
traducido como remodelacion osea interna (ROI).
Las diferencias entre remodelacion osea interna y externa son las siguientes:
a) La ROE requiere la accion de osteoblastos y osteoclastos pero de forma independiente,
mientras que en la ROI esa actividad es secuencial y coordinada como se vera mas adelante.
b) La ROE produce cambios de forma y/o de tamano en el hueso, mientras que la ROI
normalmente no afecta ni a la forma ni al tamano.
c) La velocidad de la ROE decrece con la edad a la vez que los huesos van madurando.
La ROI ocurre durante toda la vida aunque tambien se ve frenada una vez acabado el
crecimiento.
d) La ROE es un proceso continuo y prolongado mientras que la ROI ocurre de manera
episodica, con un principio y un final claro en cada punto de remodelacion.
4.5.1. Remodelacion osea externa
La ROE es necesaria aunque no medie un trauma, dado que el crecimiento longitudinal
de los huesos no produce siempre la forma adecuada para cada individuo. Algunos casos en
los que tiene lugar la ROE son los siguientes:
a) Reduccion del diametro de la metafisis. Durante el crecimiento de algunos huesos, por
osificacion de la lınea epifisaria, es necesario reducir simultaneamente el diametro del hueso
que queda detras, para crear la diafisis (ver figura 4.13a)
b) Aumento del diametro de la diafisis que acompana al crecimiento de los huesos largos.
Ello ocurre por formacion en el periostio y reabsorcion en el endostio (ver figura 4.13b)
c) Modificacion de la curvatura de la diafisis, que tambien es necesaria durante el crecimiento
de huesos largos (ver figura 4.13c)
d) ROE en huesos planos, como por ejemplo el craneo, que aumenta de tamano durante

90 Biologıa osea
Diafisis
Hueso reabsorbidopor osteoclastos
Crecimiento
Osteoblastos a
b c
1
Figura 4.13: Ejemplos de ROE: (a) Reduccion del diametro de la metafisis. (b) Aumento del
diametro de la diafisis. (c) Modificacion de la curvatura de a diafisis. Tomada de Martin et al. [220].
el crecimiento para albergar al cerebro que tambien crece durante la infancia. No solo es
necesaria la formacion osea en las lıneas de sutura, para ir cerrando las fontanelas, sino
tambien en la superficie exterior, acompanada de reabsorcion en la superficie interior para
modificar la curvatura de la placa osea.
4.5.2. Remodelacion osea interna
Este fenomeno es el que se ha implementado en esta tesis y por ello se estudia con algo
mas de profundidad. La ROI tiene lugar durante toda la vida y no solo durante el perıodo
de crecimiento.
Segun Currey [83], el hueso es una estructura optima desde el punto de vista resistente,
porque consigue la maxima rigidez con el mınimo peso. Ese es uno de los objetivos de la ROI:
adaptar la estructura osea para conseguir dicha optimalidad. Como ejemplo de esto, cabe
recordar lo que se dijo en un apartado previo sobre la diafisis de los huesos largos y el cuerpo
de la mandıbula: que tienen una estructura tubular para resistir esfuerzos de flexion-torsion.
Por otra parte, como consecuencia de la actividad diaria, se acumula un cierto dano en el

4.5 Remodelacion osea 91
hueso, en forma de microgrietas, que es necesario reparar para mantener la estructura en
estado optimo. Este es el segundo objetivo de la ROI.
Aparte de esta mision de mantenimiento y optimizacion de la estructura osea, se produce
una remodelacion osea no diferenciada que responde a la necesidad de regular la concen-
tracion de calcio en la sangre. Esta regulacion es un proceso homeostatico en el que el sistema
hormonal tambien juega un papel muy importante. Si la concentracion de calcio en sangre
disminuye, las glandulas paratiroides segregan PTH, una hormona que estimula la actividad
de los osteoclastos, lo que acelera la reabsorcion osea, pasando el calcio de la matriz osea a
la sangre. Por el contrario, si la concentracion de calcio sanguıneo es muy alta las celulas C
de la glandula tiroides segregan calcitonina que inhibe la actividad de los osteoclastos. El
exceso de calcio sanguıneo se acumula en la matriz osea por la mineralizacion del osteoide,
sin que apenas haya transito de calcio en sentido contrario.
Como ya se ha dicho, en la ROI intervienen osteoclastos y osteoblastos, actuando de
forma coordinada y secuencial, en una asociacion de celulas denominada BMU (Basic Mul-
ticellular Unit) [108], que incluye unos 10 osteoclastos y varios cientos de osteoblastos cuando
la BMU alcanza el perıodo de madurez [220].
En la figura 4.14 se observa una micrografıa de una BMU de hueso trabecular. Estas
BMUs actuan siempre sobre la superficie de la matriz osea como se puede apreciar en el
esquema de la figura 4.15.
Las BMUs de hueso cortical tienen una forma diferente y aunque tambien se originan en
la superficie de la matriz osea, la pueden atravesar en su avance posterior. En la figura 4.16
se muestra un esquema de una BMU de hueso cortical. En una determinada zona del hueso
los osteoclastos estan reabsorbiendo tejido viejo, mientras que por detras, los osteoblastos
estan formando nuevo osteoide, a la vez que se forman los vasos que discurren por el canal
de Havers. Mas atras aun, el osteoide se esta mineralizando y algunos osteoblastos quedan
atrapados en la matriz osea, diferenciandose en osteocitos. En la figura 4.17 se muestra un
esquema de la progresion de una BMU de hueso cortical.
Dentro de la BMU la actividad de los osteoclastos precede siempre a la de los osteoblastos
en una secuencia invariable de activacion-reabsorcion-formacion (secuencia ARF) [260], en
la que juegan un papel muy importante las comunicaciones entre celulas. El proceso completo
de la secuencia ARF incluye seis fases, que son: activacion, reabsorcion, inversion, formacion,
mineralizacion, fin de la adaptacion (figura 4.18).
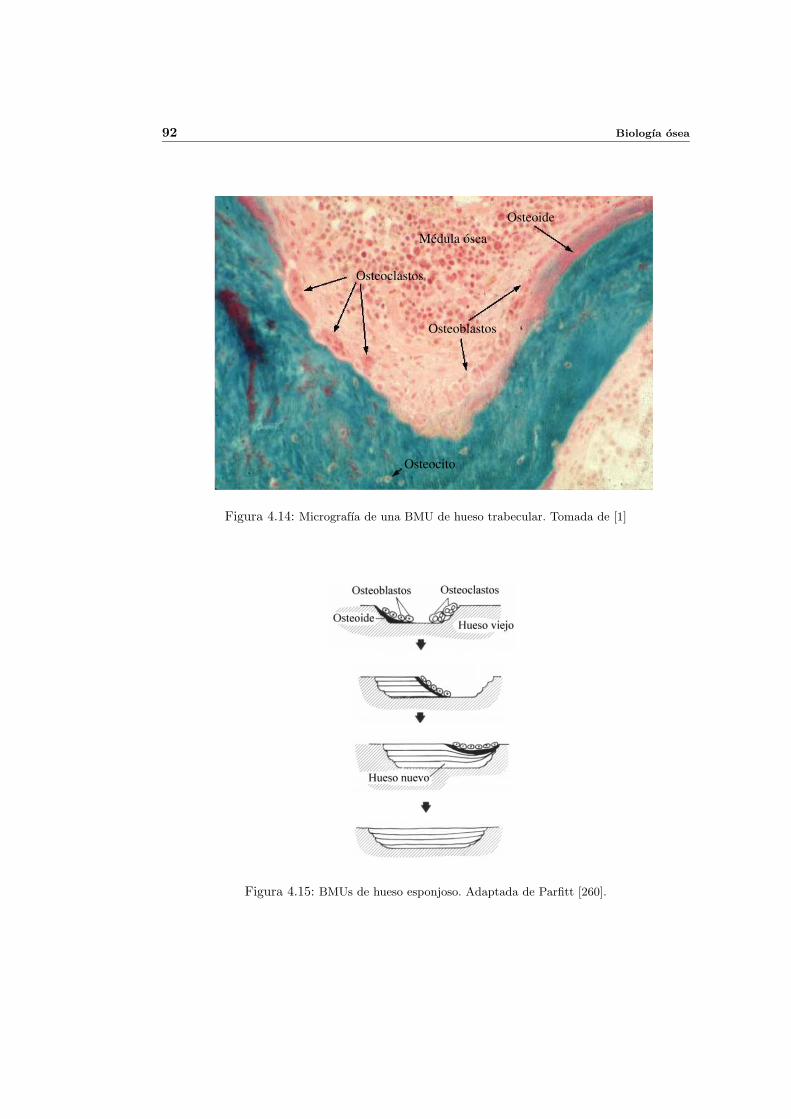
92 Biologıa osea
Osteoclastos
Osteoblastos
Osteocito
Osteoide
Medula osea
1
Figura 4.14: Micrografıa de una BMU de hueso trabecular. Tomada de [1]
1
Figura 4.15: BMUs de hueso esponjoso. Adaptada de Parfitt [260].

4.5 Remodelacion osea 93
OsteoclastosOsteoblastosOsteocitos
Osteoide
Cavidad dereabsorcion
VasosTejido mineralizado
40µm
200µm
4800µm 350µm
v = 40 µmdıa
1
Figura 4.16: Esquema de una BMU de hueso cortical. Se observa en varias secciones la formacion
del canal de Havers. Adaptada de Fridez [103]
Hueso viejo
Hueso nuevo
Osteoide
1
Figura 4.17: Esquema de la progresion de un BMU de hueso cortical. Tomada de Garcıa [115].

94 Biologıa osea
OsteoclastosOsteoblastos
Celulas de borde
1
Figura 4.18: Fases de la actividad de una BMU de hueso trabecular: (0) Activacion, (1) Reabsor-
cion,(2) Inversion, (3) Comienzo de la formacion, (4) Formacion de osteoide, (5) Mineralizacion, (6)
Fin de la adaptacion. Adaptada de Fridez [103].
Activacion
En el entramado oseo se generan ciertas senales bioquımicas que se transmiten a traves
de los osteocitos hasta las celulas de borde. Segun la “teorıa inhibitoria” de Martin [215],
esa senal inhibe la remodelacion, pero si las celulas de borde dejan de sentirla en algun
momento, ponen en marcha los mecanismos necesarios para corregir la anomalıa. La mision
de mantenimiento del hueso que llevan a cabo los osteocitos es desempenada por estos de
una forma muy particular: actuando como sensores de la carga mecanica y transmisores de
la senal hasta las celulas de borde.
Hay que recordar que los osteocitos son osteoblastos que han quedado atrapados dentro
de la matriz osea, ocupando las lagunas y los canalıculos. Estas lagunas y canalıculos se
conectan entre sı, circulando por ellos los nutrientes que provienen del torrente sanguıneo.
En este flujo tiene un papel fundamental la carga mecanica. Dicha carga provoca deforma-
ciones que ensanchan unos canalıculos y estrechan otros, creando una diferencia de presion
que origina el referido flujo [47]. Esta circulacion de fluidos extracelulares solo es apreciable
en los canalıculos, siendo muy pequena en los canales de Havers y en los canales de Volk-
mann, con un diametro mucho mayor (unas 30000 veces). El flujo, laminar, es detectado
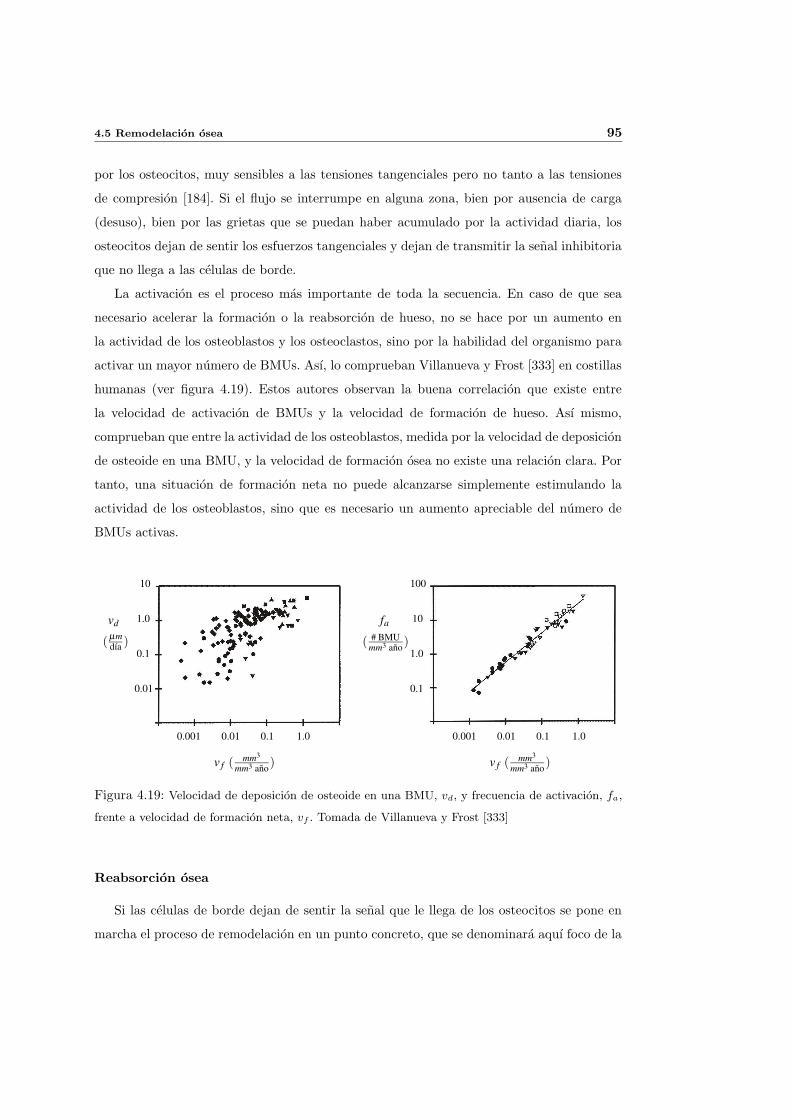
4.5 Remodelacion osea 95
por los osteocitos, muy sensibles a las tensiones tangenciales pero no tanto a las tensiones
de compresion [184]. Si el flujo se interrumpe en alguna zona, bien por ausencia de carga
(desuso), bien por las grietas que se puedan haber acumulado por la actividad diaria, los
osteocitos dejan de sentir los esfuerzos tangenciales y dejan de transmitir la senal inhibitoria
que no llega a las celulas de borde.
La activacion es el proceso mas importante de toda la secuencia. En caso de que sea
necesario acelerar la formacion o la reabsorcion de hueso, no se hace por un aumento en
la actividad de los osteoblastos y los osteoclastos, sino por la habilidad del organismo para
activar un mayor numero de BMUs. Ası, lo comprueban Villanueva y Frost [333] en costillas
humanas (ver figura 4.19). Estos autores observan la buena correlacion que existe entre
la velocidad de activacion de BMUs y la velocidad de formacion de hueso. Ası mismo,
comprueban que entre la actividad de los osteoblastos, medida por la velocidad de deposicion
de osteoide en una BMU, y la velocidad de formacion osea no existe una relacion clara. Por
tanto, una situacion de formacion neta no puede alcanzarse simplemente estimulando la
actividad de los osteoblastos, sino que es necesario un aumento apreciable del numero de
BMUs activas.
v f ( mm3
mm3 ano) v f ( mm3
mm3 ano)
0.001 0.01 0.1 1.0 0.001 0.01 0.1 1.0
0.1
1.0
10
100
0.01
0.1
1.0
10
fa
( # BMUmm3 ano)
vd
(µmdıa )
1
Figura 4.19: Velocidad de deposicion de osteoide en una BMU, vd, y frecuencia de activacion, fa,
frente a velocidad de formacion neta, vf . Tomada de Villanueva y Frost [333]
Reabsorcion osea
Si las celulas de borde dejan de sentir la senal que le llega de los osteocitos se pone en
marcha el proceso de remodelacion en un punto concreto, que se denominara aquı foco de la

96 Biologıa osea
BMU. Las celulas de borde cambian su forma aplanada, se encogen ligeramente y aumentan
su espesor, hacia una forma mas compacta, dejando de cubrir la superficie osea para que
en ella actuen los osteoclastos. Tambien segregan RANK-L que intearcciona con el receptor
RANK de los precursores de osteoclastos, haciendo que estos se diferencien y se fundan
en osteoclastos maduros, multinucleados. Con el RANK-L tambien puede interaccionar la
osteoprotegerina (OPG), sustancia segregada por los osteoblastos y que puede inhibir la
union entre RANK y RANK-L y por tanto la diferenciacion de osteoclastos, por lo que se
esta probando en ensayos clınicos como medida contra la osteoporosis [17, 150].
Los osteoclastos comienzan a reabsorber hueso en el foco de la BMU, formando un
hueco, que se denomina laguna de Howship (ver figura 4.15), en el hueso trabecular y un
tunel de seccion aproximadamente circular en el hueso cortical (ver figuras 4.16 y 4.17). Los
osteoclastos avanzan reabsorbiendo el hueso antiguo y haciendo la laguna o el tunel cada
vez mas grande.
¿A que velocidad? Con un estudio histomorfometrico1 se puede medir la distancia entre
los extremos de las etiquetas de tetraciclina (ver figura 4.20). Dividiendo esa distancia x por
el tiempo transcurrido entre administraciones, TL, se estima la velocidad de avance del frente
de mineralizacion. Si ademas se supone que la forma de la BMU permanece invariable, dicha
velocidad coincide con la de avance de la BMU. Jaworski y Lok [170] midieron esta velocidad
de avance en costillas de perro obteniendo 39 ± 14 µm/dıa. Como puede observarse en la
figura 4.20 el frente de reabsorcion tiene forma practicamente elipsoidal, con un semieje
mayor de 300 µm (en costillas de perro) y un semieje menor (el radio de la osteona) de
100 µm . Eso quiere decir que en un determinado punto de remodelacion se tardan 300/39,
aproximadamente 8 dıas, en abrir por completo la cavidad. A dicho lapso de tiempo se le
denomina perıodo de reabsorcion, TR. La velocidad de erosion de los osteoclastos en direccion
perpendicular a la de avance de la BMU, serıa entonces 100/8 ' 12µm/dıa.
Poco se sabe sobre la forma en que los osteoclastos reabsorben hueso, si lo hacen radial-
mente o avanzando en lınea recta en la direccion de avance de la BMU. Parece menos logico
el avance radial porque requiere la recirculacion de los osteoclastos.
El perıodo de vida de un osteoclasto es un concepto que no esta muy claro y podrıa
durar todo el tiempo que esta activa la BMU. Segun Jaworski et al. [171] los osteoclastos
1Consiste en la administracion de tetraciclina, que se incorpora al hueso durante la mineralizacion y se
ve fluorescente al microscopio cuando se ilumina con luz ultravioleta. De esta forma, se etiqueta el tejido
que se esta mineralizando el dıa en que se administra la tetraciclina.

4.5 Remodelacion osea 97
x
T1 T2 v300µm
100µm
tejidomineralizado osteoide
1
Figura 4.20: Esquema que muestra la forma de medir el avance de la osteona mediante una seccion
histomorfometrica longitudinal. Se administra tetraciclina dos dıas separados un perıodo ∆t, que
producen sendas etiquetas T1 y T2 en el tejido mineralizado. Si la forma de la BMU no cambia, su
velocidad de avance se puede estimar como v =x
∆t.
maduros, renuevan continuamente sus nucleos, fundiendo nucleos nuevos, preosteoclastos, y
eliminando nucleos antiguos, postosteoclastos, que podrıan continuar reabsorbiendo hueso
por detras de los osteoclastos multinucleados. La apoptosis de los osteoclastos sobrevendrıa
entonces al cesar la actividad de la BMU.
Inversion
La transicion entre la actividad de los osteoclastos y los osteoblastos no es instantanea.
En el ser humano dura unos 30 dıas. Durante este perıodo los postosteoclastos, fagocitos
mononucleados, podrıan terminar de reabsorber hueso y preparar la lınea de cementacion
sobre la que deposita el osteoide [22]. Para ello los postosteoclastos son atraıdos por los
residuos de la osteoclasis (fibras de colageno, enzimas lisosomales y glicosaminoglicanos)
hacia la pared de la cavidad.
Formacion osea
Los osteoblastos aparecen sobre la superficie de la cavidad que dejaron los osteoclastos
y comienzan a depositar lamelas de osteoide.
El origen de los osteoblastos es diferente al de los osteoclastos. Mientras que estos ultimos
provienen de celulas hematopoyeticas, los osteoblastos provienen de celulas madre mesen-
quimales [258], lo que descartarıa que los postosteoclastos pudieran diferenciarse en os-
teoblastos una vez separados del osteoclasto.

98 Biologıa osea
Lo precursores de osteoblastos (preosteoblastos) presentes en la zona de inversion son
atraıdos por factores quimiotacticos, tales como porciones de colageno resultantes de la
osteoclasis, hacia la superficie de la cavidad. Segun Roberts et al. [289], estos preosteoblastos
se dividen por mitosis, para mantener su poblacion, y una parte de ellos se diferencian en
osteoblastos, siendo necesaria para esta diferenciacion un cierto nivel de tensiones segun
confirman Barou et al. [23]. Tras la diferenciacion se vuelven a dividir por mitosis para
formar los osteoblastos definitivos, con forma cuboidal, que se adhieren a la superficie de la
cavidad. El proceso completo de diferenciacion dura unas 60 horas. Si la BMU avanza a una
velocidad de unos 40µm/dıa, en esos dos dıas y medio que dura la diferenciacion, el cono de
reabsorcion de la BMU ha avanzado 100 µm. Teniendo en cuenta otros procesos que tienen
lugar antes de la formacion, aparte de los observados por Roberts et al., como la preparacion
de la lınea de cementacion antes comentada, el cono de reabsorcion habra avanzado algo mas,
antes de que comience a depositarse osteoide. Esto estarıa de acuerdo con los tamanos de la
zona de inversion medidos experimentalmente, entre 100− 200 µm.
Jaworski y Hooper [172] demostraron que los osteoblastos no avanzan con la BMU, como
los osteoclastos, sino que permanecen estacionarios en una seccion de la BMU. Una vez
diferenciados ocupan una zona de la cavidad aproximadamente cuadrada de 15 µm de lado
y van alargandose y aplanandose a la vez que forman osteoide, de forma que cuando se
diferencian en osteocitos o celulas de borde abarcan una zona de 300 µm2. Como la BMU
avanza a una velocidad de 40µm/dıa cada dıa es necesario formar tres anillos de osteoblastos.
En el caso de una osteona (BMU de hueso cortical) que tienen un radio de unas 100 µm el
perımetro es aproximadamente 600 µm en el que cabrıan 40 osteoblastos. Por tanto, para
cada BMU es necesario reclutar 120 osteoblastos diarios.
Cabe hacerse en este momento la siguiente pregunta: ¿la capa inicial de osteoblastos
completa la formacion de una seccion o es necesario que haya distintas generaciones de os-
teoblastos que completen el trabajo?. La respuesta no esta del todo clara, aunque la segunda
opcion parece mas factible. Frost [104] describe unas suturas en el osteoide depositado en
BMUs de hueso cortical y sugiere que estas pueden deberse a un receso en la deposicion de
osteoide. Este receso puede obedecer bien a una pausa en la actividad de los osteoblastos o
bien a una renovacion de los mismos, lo que confirmarıa la hipotesis de que son varias las
generaciones de osteoblastos que completan la formacion, segun indica Johnson [176]. Cada
generacion contribuirıa entonces a formar una lamela distinta. Otro hecho que tambien con-

4.5 Remodelacion osea 99
firmarıa esta hipotesis serıa el de los osteoblastos que quedan encerrados en la matriz osea
para diferenciarse en osteocitos. Estos osteocitos serıan osteoblastos que dejan de segregar
osteoide y son enterrados por el tejido generado por la siguiente generacion de osteoblastos.
Se apunta a continuacion otra posible causa por la que los osteocitos quedan atrapados
en la matriz osea, al menos en hueso cortical. Con la formacion osea, se va cerrando el
hueco cilındrico que crearon los osteoclastos y consecuentemente, cada vez es menor la
superficie sobre la que se apoyan los osteoblastos. Ası, es probable que algunos de ellos se
vean enterrados por los osteoblastos vecinos. Esta explicacion, sin embargo, es valida solo
para hueso cortical, ya que en las BMUs de hueso trabecular la superficie donde trabajan
los osteoblastos no disminuye necesariamente con el avance de la formacion.
Otro aspecto a tener en cuenta es que la velocidad de deposicion, vf , decrece con el
tiempo en una determinada seccion. Esta velocidad de deposicion se refiere, en el caso de
hueso cortical, a la velocidad con que disminuye el radio de la cavidad,
vf =dR
dt. (4.9)
Si se supone constante esta velocidad, el tiempo que tardarıa en completarse la formacion
en una seccion, Tf , podrıa expresarse
Tf =Ro −RH
vf(4.10)
donde Ro es el radio de una osteona y RH el radio del canal de Havers.
Mason y Waters [212] conjeturaron que la velocidad de deposicion es directamente pro-
porcional al radio de la cavidad.
vf =dR
dt= −k R, (4.11)
con signo menos porque la deposicion va en sentido contrario al radio, lo que darıa para la
duracion del perıodo de formacion:
Tf = −1
kln(
RHRo
) (4.12)
Con un estudio histomorfometrico de doble etiquetado se pueden medir los radios de las
lamelas etiquetadas con tetraciclina en dos dıas diferentes R1 y R2. Si TL es el tiempo que
transcurrio entre las dos administraciones de tetraciclina, R2 y R1 estaran relacionadas de
la siguiente forma, supuesta una velocidad de deposicion constante
R2 = R1 + vf · TL (4.13)

100 Biologıa osea
y de esta otra, con la hipotesis de Mason y Waters
R2 = R1 e−k TL (4.14)
que se ajusta mejor a los resultados experimentales que la ley de velocidad constante. Con la
ley exponencial la forma del frente de formacion ya no es un cono como lo serıa con velocidad
de deposicion constante (ver figura 4.21)
t
t
e
e
Lineal
Exponencial
1
Figura 4.21: Comparacion entre los modelos de deposicion de osteoide lineal y exponencial y forma
del frente de formacion resultante de cada uno. Tomada de Martin et al. [220].
¿Por que disminuye la velocidad de deposicion con el tiempo? Segun Manson y Waters
[212], la actividad de los osteoblastos se ve favorecida por la lejanıa de los vasos sanguıneos,
algo que no parece muy logico ya que las celulas se nutren de los capilares cercanos. Lo que
ocurre en realidad es que los osteoblastos disponen de menos espacio para diferenciarse de
sus precursores cuando el radio de la cavidad es muy pequeno, mas pequeno aun cuando
el vaso sanguıneo se esta formando. Este problema de espacio dificulta la diferenciacion de
osteoblastos y aunque la necesidad de este tipo de celulas es menor en la ultima fase (la
superficie sobre la que depositan osteoide es menor), la velocidad de deposicion decrece.
La disminucion de espacio disponible para la diferenciacion, es inherente a las BMUs
de hueso cortical y es lo que hace que disminuya la velocidad de formacion de osteoide en
este tipo de BMUs. Esa es la unica causa y no otros factores intrınsecos de los osteoblastos,
ya que en hueso trabecular, en el que no existe este problema de espacio, la velocidad de
deposicion es aproximadamente constante, como comprueban Martin et al. [221].

4.5 Remodelacion osea 101
Otra razon para esta desaceleracion podrıa ser, segun Martin et al. [220], el nivel de
tensiones que existirıa en el entorno del cono de formacion en la ultima fase del proceso,
mas bajo como consecuencia del aumento en la cantidad de material. Hay que recordar que,
segun Roberts et al. [288], para la diferenciacion de osteoblastos es necesario un cierto nivel
de tensiones. No tienen en cuenta Martin y sus colaboradores que en el hueso trabecular
las tensiones tambien deben disminuir como consecuencia del aumento en la cantidad de
material y sin embargo no se observa tal desaceleracion, seguramente porque el nivel de
tensiones es adecuado para la diferenciacion, en cualquier caso.
Mineralizacion
Unos 10 o 20 dıas despues de ser depositado el osteoide comienzan a aparecer en el los
primeros cristales de hidroxiapatita, que desplazan el agua que quedo atrapada entre las
fibras de colageno.
La mineralizacion es muy rapida al principio, lentificandose despues conforme el tejido se
va saturando de mineral. De acuerdo con esto se suelen distinguir dos fases en el proceso de
mineralizacion: una fase primaria muy rapida, que dura unos pocos dıas [262] y en la que se
alcanza aproximadamente el 70 % del contenido maximo de mineral,1 y una fase secundaria
en la que la velocidad de deposicion de mineral decrece con el tiempo de forma exponencial.
Esta fase secundaria se define como el perıodo de tiempo que tarda en alcanzarse el 95 % del
contenido maximo de mineral [262] y dura entre unos 6 meses, segun Parfitt, [262] y varios
anos, segun Frost [107].
Fin de la adaptacion
Los osteoclastos sufren apoptosis y la BMU se detiene. Por detras siguen actuando los
osteoblastos durante cierto tiempo hasta que rellenan de osteoide el hueco dejado por los
osteoclastos. El resultado final es una osteona nueva, o una laguna de Howship, rellena de
hueso nuevo con bajo contenido en mineral y que se ira mineralizando poco a poco, despues
de que cese la actividad de la BMU. Dentro de la matriz osea han quedado atrapados algunos
osteoblastos, que se diferencian en osteocitos, mientras que otros quedan sobre la superficie
osea cuando dejan de depositar osteoide y se convierten en celulas de borde. Ambos tipos
1El contenido maximo de mineral es el que tendrıa un tejido en el que todo el agua atrapada entre el
colageno ha sido sustituida por mineral y corresponde aproximadamente a una fraccion de ceniza α = 0.7.

102 Biologıa osea
de celulas son las encargadas del mantenimiento del nuevo hueso depositado y de comenzar
de nuevo el proceso si la situacion lo requiere.
4.6. Propiedades mecanicas del hueso
4.6.1. Aspectos generales
No se puede generalizar al hablar de las propiedades mecanicas del hueso, dado que dichas
propiedades dependen de que hueso se trate. Y ello es ası porque las propiedades mecanicas
dependen en gran medida de la microestructura osea y esta, como ya se ha indicado, de las
cargas a las que esta sometido el hueso durante su desarrollo. Ası, por ejemplo, el hueso
compacto del cubito y del radio son mas resistentes y rıgidos a traccion que el de la tibia y
el femur. En cambio la resistencia y la rigidez a compresion de estos ultimos son superiores
a las de cubito y radio. En flexion, las propiedades de radio y cubito son de nuevo superiores
a las de tibia y femur. Cada hueso parece tener mejores propiedades frente a las cargas que
normalmente soporta, lo que esta de acuerdo con lo anteriormente dicho sobre la optimalidad
de la estructura osea gracias a la remodelacion.
En el caso de la mandıbula humana, varios autores han medido las propiedades mecanicas
del hueso que la compone. Arendts y Sigolotto [10] extrajeron muestras de dimensiones 7x2x2
mm en la capa de cortical de diferentes mandıbulas y mediante ensayos de compresion
uniaxial determinaron los modulos de Young en direccion axial, Ea, la que recorre el eje del
cuerpo mandibular; en direccion transversal, Et, contenida en una seccion perpendicular al
eje del cuerpo mandibular y tangente al perfil de dicha seccion; y en direccion radial, Er,
perpendicular a las dos anteriores. En la figura 4.22 se muestran sendos esquemas de las
muestras extraıdas para medir Et (izquierda) y Ea (derecha). Las dimensiones de la capa
de cortical impiden extraer probetas de tamano suficientes para medir Er por lo que estos
autores establecieron la relacion Et = 1.2 · Er, de manera arbitraria.
Los resultados que obtuvieron son los siguientes:
16.0GPa < Ea < 18.0GPa
7.0GPa < Et < 9.5GPa
6.0GPa < Er < 8.5GPa
(4.15)
Tambien midieron la resistencia a compresion en las tres mismas direcciones obteniendo los
4.6 Propiedades mecanicas del hueso 103
1
Figura 4.22: Probetas extraıdas por Arendts y Sigolotto [11] para sus ensayos.
siguientes valores medios:
σult a = 200MPa
σult t = 110MPa
σult r = 100MPa
(4.16)
No midieron la resistencia a traccion, sin duda por la dificultad que entrana el manejo de
probetas de dimensiones tan reducidas. Sin embargo, son de esperar valores mas pequenos,
como ocurre en el femur, que presenta una resistencia a traccion de 135 MPa y una resistencia
a compresion de 193 MPa, segun Reilly y Burstein [280].
Schwartz-Dabney et al. [301] midieron los modulos de Young en las tres mismas direc-
ciones, pero solo en la region sinfisaria. Las medidas las realizaron mediante ultrasonidos en
probetas de dimensiones 4x4x2 mm, obteniendo valores algo superiores a los obtenidos por
Arendts y Sigolotto:
Ea = 23GPa
Et = 15GPa
Er = 10GPa
(4.17)
Los anteriores son valores promedio de las probetas y deben ser considerados con cierta
cautela, dado que la caracterıstica principal del hueso es su heterogeneidad. Otro factor a
tener en cuenta es el hecho de que dichas medidas han sido tomadas in vitro, con lo cual, es
posible que se haya perdido parte de la medula osea si ha transcurrido cierto tiempo entre
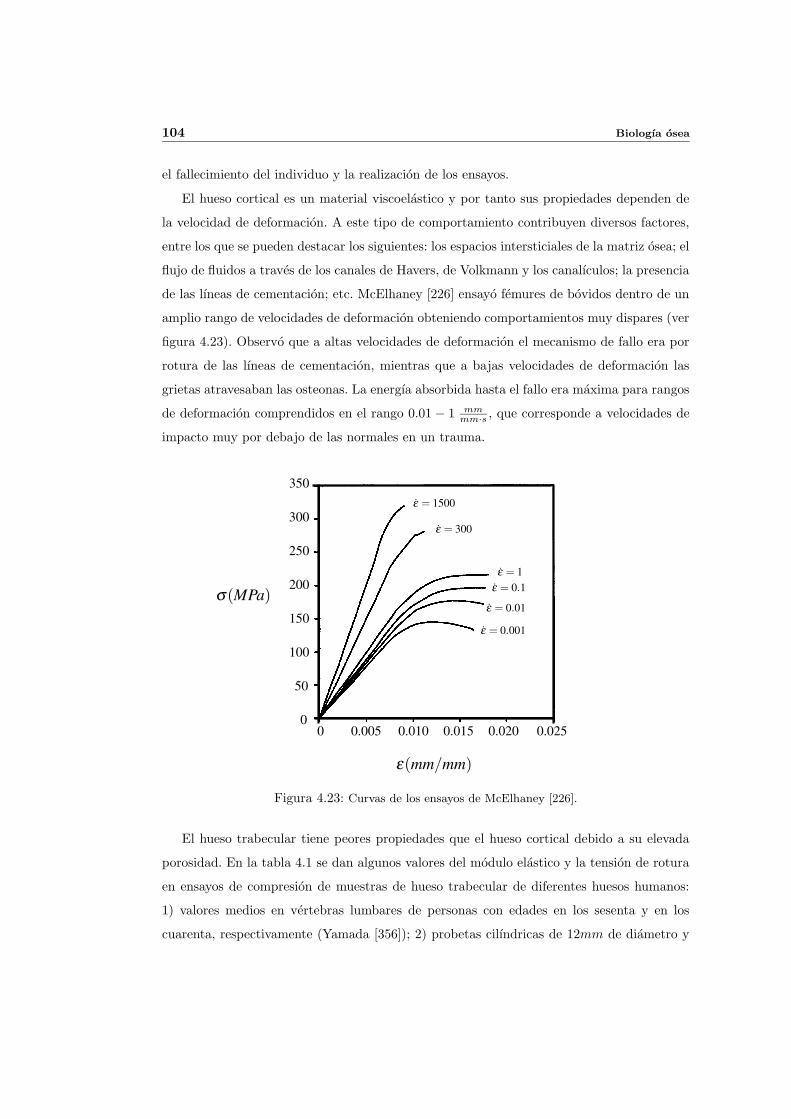
104 Biologıa osea
el fallecimiento del individuo y la realizacion de los ensayos.
El hueso cortical es un material viscoelastico y por tanto sus propiedades dependen de
la velocidad de deformacion. A este tipo de comportamiento contribuyen diversos factores,
entre los que se pueden destacar los siguientes: los espacios intersticiales de la matriz osea; el
flujo de fluidos a traves de los canales de Havers, de Volkmann y los canalıculos; la presencia
de las lıneas de cementacion; etc. McElhaney [226] ensayo femures de bovidos dentro de un
amplio rango de velocidades de deformacion obteniendo comportamientos muy dispares (ver
figura 4.23). Observo que a altas velocidades de deformacion el mecanismo de fallo era por
rotura de las lıneas de cementacion, mientras que a bajas velocidades de deformacion las
grietas atravesaban las osteonas. La energıa absorbida hasta el fallo era maxima para rangos
de deformacion comprendidos en el rango 0.01 − 1 mmmm·s , que corresponde a velocidades de
impacto muy por debajo de las normales en un trauma.
ε(mm/mm)
σ(MPa)
0
50
100
150
200
250
300
350
0 0.005 0.010 0.015 0.020 0.025
ε = 1500
ε = 300
ε = 1ε = 0.1
ε = 0.01
ε = 0.001
1
Figura 4.23: Curvas de los ensayos de McElhaney [226].
El hueso trabecular tiene peores propiedades que el hueso cortical debido a su elevada
porosidad. En la tabla 4.1 se dan algunos valores del modulo elastico y la tension de rotura
en ensayos de compresion de muestras de hueso trabecular de diferentes huesos humanos:
1) valores medios en vertebras lumbares de personas con edades en los sesenta y en los
cuarenta, respectivamente (Yamada [356]); 2) probetas cilındricas de 12mm de diametro y
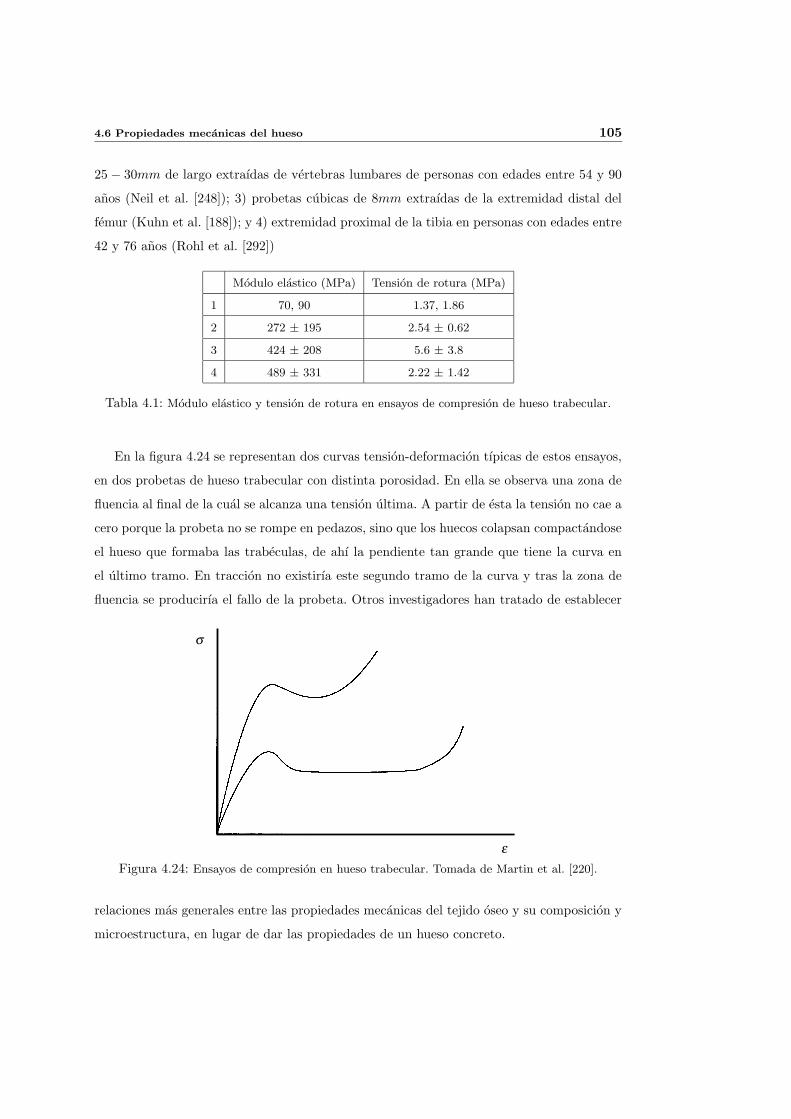
4.6 Propiedades mecanicas del hueso 105
25− 30mm de largo extraıdas de vertebras lumbares de personas con edades entre 54 y 90
anos (Neil et al. [248]); 3) probetas cubicas de 8mm extraıdas de la extremidad distal del
femur (Kuhn et al. [188]); y 4) extremidad proximal de la tibia en personas con edades entre
42 y 76 anos (Rohl et al. [292])
Modulo elastico (MPa) Tension de rotura (MPa)
1 70, 90 1.37, 1.86
2 272 ± 195 2.54 ± 0.62
3 424 ± 208 5.6 ± 3.8
4 489 ± 331 2.22 ± 1.42
Tabla 4.1: Modulo elastico y tension de rotura en ensayos de compresion de hueso trabecular.
En la figura 4.24 se representan dos curvas tension-deformacion tıpicas de estos ensayos,
en dos probetas de hueso trabecular con distinta porosidad. En ella se observa una zona de
fluencia al final de la cual se alcanza una tension ultima. A partir de esta la tension no cae a
cero porque la probeta no se rompe en pedazos, sino que los huecos colapsan compactandose
el hueso que formaba las trabeculas, de ahı la pendiente tan grande que tiene la curva en
el ultimo tramo. En traccion no existirıa este segundo tramo de la curva y tras la zona de
fluencia se producirıa el fallo de la probeta. Otros investigadores han tratado de establecer
ε
σ
1
Figura 4.24: Ensayos de compresion en hueso trabecular. Tomada de Martin et al. [220].
relaciones mas generales entre las propiedades mecanicas del tejido oseo y su composicion y
microestructura, en lugar de dar las propiedades de un hueso concreto.

106 Biologıa osea
Son tres los parametros que tienen mayor influencia en el comportamiento mecanico: la
composicion, o el contenido de mineral para ser mas exactos; la cantidad de material, medida
a traves de la porosidad, de la densidad aparente, de la fraccion volumetrica de hueso, etc.;
y por otro lado, la direccionalidad del entramado oseo.
4.6.2. Influencia del contenido de mineral
En general, el grado de mineralizacion promedio de un hueso normal esta dentro de
un rango estrecho de valores, siendo variable a nivel local, en los puntos de remodelacion
donde se esta mineralizando el nuevo osteoide. Solo en determinadas patologıas esos valores
promedio pueden alejarse de dicho rango. Por ejemplo, la osteomalacia se caracteriza por un
excesivo retraso en la mineralizacion del osteoide, lo que hace que la mineralizacion promedio
sea muy baja. Por contra, la osteopetrosis esta relacionada con una reduccion en la actividad
de los osteoclastos. Esto hace que aumente la densidad osea, y al disminuir la cantidad de
matriz mineralizada que se reabsorbe, que el calcio se acumule en los huesos aumentando el
grado de mineralizacion de manera muy acusada.
Son pequenas las variaciones que experimenta el contenido mineral del hueso, sin embar-
go, su influencia en las propiedades mecanicas es notable. Vose y Kubala [336] analizaron la
resistencia a flexion del hueso y observaron que el aumento de la fraccion de ceniza de 63 %
a 71 % incrementaba la resistencia en 3.7 veces. Entonces, si las propiedades mejoran tanto,
¿por que el hueso tiene un contenido de mineral tan limitado? Currey [81] lo explico dicien-
do que el hueso se vuelve mas fragil con un aumento excesivo de la fraccion de ceniza. Es
mas debil a altas velocidades de deformacion, lo que aumenta el riesgo de fractura frente a
impactos. Este autor sugirio que la fraccion de ceniza optima esta entre 66-67 %.
Schaffler y Burr [299] obtuvieron la siguiente correlacion para el modulo de Young del
hueso cortical en bovidos:
E(MPa) = 89.1 α3.91 (4.18)
Hernandez [144] obtuvo las siguientes correlaciones en humanos, que incluyen tambien
la dependencia con la cantidad de material a traves de la fraccion volumetrica de hueso, vb:
E(MPa) = 84370 v2.58b α2.74 (4.19a)
σult(MPa) = 794.33 v1.92b α2.79 (4.19b)

4.6 Propiedades mecanicas del hueso 107
4.6.3. Influencia de la porosidad
Obviamente, la presencia de poros en una estructura la debilita y eso es lo que ocurre en
el hueso. Existen varias relaciones empıricas entre la porosidad y las propiedades mecanicas,
pero se suelen encontrar relaciones diferentes para hueso cortical y trabecular.
Hay que tener en cuenta, en primer lugar, la forma tan diferente que tienen los poros de
uno y otro tipo de hueso. Pero no solo es diferente la forma sino tambien el volumen total de
poros, siendo los rangos de porosidad muy diferentes en uno y otro caso. Aun ası, si el tejido
fuera el mismo, una sola curva podrıa servir para relacionar el modulo elastico y la porosidad
de ambos tipos de hueso. No sucede ası. El hueso cortical tiene mejores caracterısticas que
tendrıa el hueso trabecular con la misma porosidad [282]. El tejido no es el mismo, como ya
se ha visto en este capıtulo, siendo la estructura de osteonas mejor desde el punto de vista
resistente, por la siguiente razon: una grieta que rompa las lıneas de cementacion en el tejido
trabecular puede exfoliar una capa de hueso, reduciendo significativamente su resistencia y
rigidez (ver figura 4.25). Sin embargo, en el hueso cortical las grietas pueden separar las
lıneas de cementacion e incluso continuar creciendo por el hueso intersticial, pero no suelen
alcanzar la superficie. De esta forma no se compromete excesivamente la rigidez del tejido,
al ser menos probable que se separe un trozo como ocurre en el hueso trabecular. Tambien
es diferente el grado de mineralizacion de uno y otro, siendo menor en el trabecular, lo que
hace que su rigidez y resistencia sean menores, como ya se ha visto. La razon por la que
el contenido mineral es menor en el hueso trabecular es su actividad remodelatoria, mas
intensa que la del hueso cortical, lo que impide al tejido alcanzar un grado de mineralizacion
alto antes de ser reabsorbido.
Schaffler y Burr [299] obtuvieron para el modulo elastico del hueso cortical de bovidos
la siguiente relacion:
E(MPa) = 33.9 (1− p)10.9 (4.20)
donde p es la porosidad y 1− p es la fraccion volumetrica de hueso. Currey [84] obtuvo esta
otra relacion para el hueso cortical de una amplia variedad de especies:
E(MPa) = 23.4 (1− p)5.74 (4.21)
Carter y Hayes [58] encontraron que el modulo elastico es proporcional al cubo de la
densidad aparente en probetas que incluıan hueso cortical y hueso trabecular. Rice et al.
[283] encontraron mas apropiado un exponente de 2 para especımenes formados por hueso

108 Biologıa osea
1
Figura 4.25: Grietas tıpicas en el hueso trabecular (izquierda) y en el cortical (derecha). Tomada
de Martin et al. [220].
trabecular exclusivamente. Beaupre et al. [27] usan las siguientes expresiones del modulo
elastico y el coeficiente de Poisson, obtenidas experimentalmente:
E =
2014 ρ2.5 si ρ ≤ 1.2g/cm3
1763 ρ3.2 si ρ > 1.2g/cm3,(4.22)
ν =
0.2 si ρ ≤ 1.2g/cm3
0.32 si ρ > 1.2g/cm3,(4.23)
en el que se hace la distincion entre hueso trabecular y hueso cortical para una densidad
aparente de ρ = 1.2 g/cm3.
4.6.4. Influencia de la microestructura
La influencia de la microestructura es diferente dependiendo del tipo de hueso.
Hueso cortical
En el hueso cortical la dependencia con la microestructura es debida principalmente a
la orientacion de las osteonas. Estas forman la unidad estructural basica del hueso cortical
y estan separadas del resto del hueso por las lıneas de cementacion, por las que el material

4.6 Propiedades mecanicas del hueso 109
puede sufrir delaminacion. Ademas, la orientacion de las osteonas determina la orientacion
de los poros, de los canales de Havers principalmente, con los que estan alineados. Dado que
lo normal es que las osteonas esten orientadas longitudinalmente en huesos largos, la capa
de tejido cortical presenta un comportamiento transversalmente isotropo, siendo la direccion
longitudinal la de mayor rigidez y resistencia.
Tambien tiene gran influencia sobre las propiedades mecanicas del hueso cortical la
orientacion de las fibras de colageno dentro de las osteonas. Segun Martin y sus colabo-
radores [218, 222], casi tanto como la densidad o el grado de mineralizacion. Cabe entonces
preguntarse si la carga que soporta el hueso tiene alguna influencia en la disposicion de las
fibras de colageno. Portigliatti-Barbos et al. [271, 272] demostraron que ası ocurre efectiva-
mente. La orientacion de las fibras de colageno en la diafisis del femur humano no es aleatoria
y en las zonas que soportan traccion predominan las fibras orientadas longitudinalmente,
mientras que en las sometidas a compresion son mas numerosas las fibras transversales.
El hueso cortical permite elevados niveles de deformacion plastica tanto en traccion como
en compresion. Dicha deformacion plastica es mayor cuando el hueso se carga en la direccion
de las fibras de colageno [50] y mayor en hueso secundario que en hueso primario [281] lo que
sugiere que la fluencia de las lıneas de cementacion y el deslizamiento de unas sobre otras
sean los responsables de la deformacion plastica.
Hueso trabecular
En el hueso trabecular la microestructura esta definida por la forma y orientacion de los
poros. El volumen de los mismos ya esta recogido en la porosidad. Segun Gibson [123] son
tres las microestructuras basicas que existen en el hueso trabecular:
a) Isotropa (figura 4.26a). Propia de regiones con estados de tension triaxial. El modulo
elastico a nivel macroscopico de este tipo de hueso es aproximadamente:
E∗ ≈ ET v2b (4.24)
donde ET es el modulo elastico de las barras que forman la estructura trabecular.
b) Laminar . Tıpica en zonas con estados de tension biaxial (en las direcciones 1 y 2 de
la figura 4.26b). Los modulos de Young en las direcciones del plano son
E∗
1 = E∗
2 ≈ ET vb (4.25)
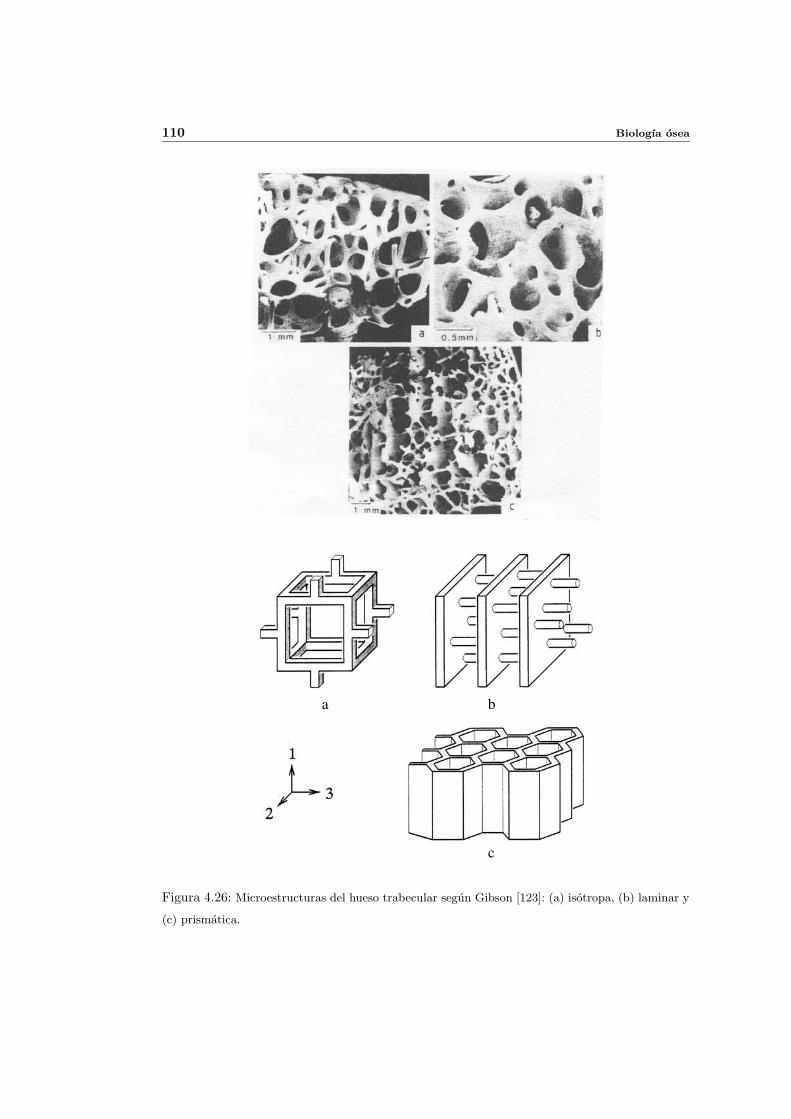
110 Biologıa osea
a b
c
1
Figura 4.26: Microestructuras del hueso trabecular segun Gibson [123]: (a) isotropa, (b) laminar y
(c) prismatica.

4.6 Propiedades mecanicas del hueso 111
y en la direccion perpendicular
E∗
3 ≈ ET v3b (4.26)
c) Prismatica. Presente en zonas con tension uniaxial (en la direccion 1 de la figura
4.26c). En la direccion del eje de los prismas, el modulo elastico es aproximadamente
E∗
1 ≈ ET vb (4.27)
y en las otras dos direcciones
E∗
2 = E∗
3 ≈ 1.5ET v3b (4.28)
en el caso de que la base de los prismas sea un hexagono regular.
Pettermann et al. [269] consideran que pueden existir otras microestructuras en el hueso
trabecular, combinacion de las anteriores. Introducen los parametros de ortotropıa, βi para
expresar de manera general los modulos de Young a nivel macroscopico en la direcciones de
ortotropıa, paralelas a la direcciones principales de tension:
E∗
i ≈ ET vβi
b (4.29)
Con las tensiones principales ordenadas de forma que |σ1| ≥ |σ2| ≥ |σ3|, Pettermann et al.
dan los siguientes valores para los parametros de ortotropıa:
βi = 3− |σi|2
|σ1|+ |σ3|(4.30)
que reproduce las expresiones dadas por Gibson para las tres microestructuras basicas:
|σ1| = |σ2| = |σ3| β1 = β2 = β3 = 2 isotropa
|σ1| = |σ2| 6= 0 |σ3| = 0 β1 = β2 = 1 β3 = 3 laminar
|σ1| 6= 0 |σ2| = |σ3| = 0 β1 = 1 β2 = β3 = 3 prismatica
(4.31)
Pettermann et al. usaron estas leyes de comportamiento en su modelo de remodelacion osea,
que proporciona buenos resultados. Sin embargo, no comprobaron con resultados experimen-
tales la validez de esta ley de comportamiento. Aun ası, es meritorio el intento de englobar
en un unico modelo de comportamiento la amplia variedad de microestructuras que presenta
el hueso trabecular. Es interesante tambien por cuanto dicha ley de comportamiento incluye
una relacion entre microestructura y estado de tension.

112 Biologıa osea
4.6.5. Fabric tensor
A continuacion se presenta una forma mas general de relacionar las propiedades mecanicas
del hueso trabecular con su microestructura. Su validez ha sido comprobada experimental-
mente y ademas es el metodo empleado en esta tesis.
1
Figura 4.27: Esquema de una estructura anisotropa de hueso trabecular y una lınea de muestreo
para la evaluacion de la MIL. Tomada de Martin et al. [220].
Whitehouse y Dyson [344] desarrollaron un metodo para medir la anisotropıa de los
poros de hueso trabecular. Sobre una cuadrıcula como la de la figura 4.27, perteneciente a
una seccion de hueso trabecular, trazaron una serie de lıneas paralelas que interceptan a
las trabeculas. En la figura 4.27 se representa solo una, por claridad. Denominaron longitud
de interseccion promedio (mean intercept length, MIL) a la distancia media entre dos inter-
secciones, medida sobre los poros. Whitehouse y Dyson [344] repitieron estas medidas para
distintas orientaciones de las lıneas, θ, y comprobaron que en dos dimensiones los datos de
MIL se ajustaban bastante bien a una elipse:
1
MIL2(θ)= L11 cos
2θ + L22 sen2θ + 2L12 senθ cosθ (4.32)
donde L11, L22 y L12 son constantes para una determinada microestructura. Harrigan y
Mann [136] extendieron esta conclusion al caso tridimensional e interpretaron el elipsoide
resultante como el elipsoide de Lame de un tensor de segundo orden simetrico y definido
positivo. Este se denominara a partir de ahora tensor MIL y se denotara por L. La ecuacion

4.6 Propiedades mecanicas del hueso 113
anterior se puede escribir entonces en funcion de las componentes del tensor MIL, de la
siguiente forma:1
MIL2(n)= ni Lij nj (4.33)
donde n es el vector unitario en la direccion de la lınea de referencia. Cowin [70] completo el
trabajo relacionando el tensor de elasticidad de un material anisotropo con la densidad
aparente y el “fabric tensor”, H, un tensor definido por este autor para medir la orientacion
de la microestructura osea y relacionado con el tensor MIL a traves de
H = L−1/2 (4.34)
Otros metodos han sido desarrollados con posterioridad para cuantificar la anisotropıa
del entramado trabecular. En el metodo de la orientacion del volumen (VO), desarrollado
por Odgaard et al. [253], se mide en distintos puntos del hueso la longitud de interseccion
mas larga en el entorno de dicho punto (ver figura 4.28a). En el metodo de la distribucion
de volumen radial (star volumen distribution, SVD), desarrollado por Cruz-Orive et al. [79],
se toman al azar un conjunto de puntos dentro del hueso y se determina el promedio de las
longitudes de interseccion para un numero concreto de orientaciones (ver figura 4.28b).
a b
1
Figura 4.28: Esquema del muestreo para la evaluacion de: (a) VO y (b) SVD. Tomada de Garcıa
[115].
Zysset y sus colaboradores [362,363] dan una interpretacion alternativa de la MIL, como
la longitud promedio de las intersecciones entre la lınea de referencia y la interfase de hueso,
en lugar de la interfase de poros. Existe por tanto una relacion entre el tensor MIL medido por

114 Biologıa osea
estos autores, el tensor MIL tradicional y la porosidad. Zysset y Curnier [362] descomponen
el “fabric tensor” de la siguiente forma
H = g I + G (4.35)
donde g es una constante relacionada con la densidad aparente, I es el tensor identidad de
segundo orden y G es un tensor simetrico de segundo orden con traza nula. Estos autores
suponen que la funcion que relaciona el tensor de elasticidad y el tensor H es una funcion
homogenea y por tanto se puede normalizar H obligando a que su traza sea unitaria.
tr(H) = 1 (4.36)
Con ello se consigue desacoplar la influencia de la densidad (o la porosidad) de la influencia
de la anisotropıa en las ecuaciones constitutivas del material, dado que ahora los autovalores
del “fabric tensor” representan las proporciones entre las dimensiones del poro y no las
dimensiones en sı mismas. Zysset y Curnier [362] llegaron con esta hipotesis a unas relaciones
constitutivas generales del tipo:
Ei = E0 ρk1 h2k2
i (4.37a)
Ejνij
=E0
ν0ρk3 hk4i hk4j (4.37b)
Gij = G0 ρk5 hk6i hk6j (4.37c)
Zysset et al. [363] ajustaron las constantes de las ecuaciones anteriores mediante mınimos
cuadrados. Para ello usaron las constates elasticas obtenidas mediante tecnicas de homo-
geneizacion [243] en una serie de muestras de diferentes huesos humanos. Para caracterizar
la anisotropıa de la microestructura, o lo que es lo mismo, los autovalores del “fabric tensor”,
hi, usaron el metodo MIL y el metodo SVD obteniendo las constantes de la tabla 4.2. Los
E0(MPa) E0/ν0(MPa) G0(MPa) k1 k2 k3 k4 k5 k6
MIL 17607 45800 7799 3.2 1.6 2.7 2.4 3.3 1.6
SVD 16956 43104 7513 3.1 1.3 2.6 1.8 3.3 1.3
Tabla 4.2: Valores de las constantes de la ecuacion (4.37) ajustados mediante mınimos
cuadrados.
coeficientes de correlacion lineal de estos ajustes son bastante aceptables: R2 = 0.94 para la

4.6 Propiedades mecanicas del hueso 115
ecuacion 4.37a, R2 = 0.84 para la ecuacion 4.37b y R2 = 0.95 para la ecuacion 4.37c. En esta
tesis se usa el tensor MIL, relacionado como ya se ha dicho con el “fabric tensor”, para rela-
cionar anisotropıa y propiedades mecanicas. Este tensor MIL se normaliza para que tenga
traza unitaria y se usan ecuaciones similares a las (4.37), aunque con exponentes ligeramente
diferentes, e iguales para todas las ecuaciones. En un capıtulo posterior se justificara el por
que de esta eleccion.


Capıtulo 5
Modelos de remodelacion osea
interna
5.1. Introduccion
“Todos los cambios que sufren los huesos en su forma y funcion o bien solo en su funcion,
van seguidos de ciertos cambios concretos en su arquitectura interna e igualmente en su
conformacion externa, de acuerdo a leyes matematicas”. Esta afirmacion de Julius Wolff,
hecha a principios del siglo XX resumıa sus investigaciones, realizadas a finales del siglo
XIX, en las que pretendıa explicar la capacidad del hueso para adaptar su estructura al
estado de cargas al que se encuentra sometido, evidencia a la que habıan llegado otros
cientıficos anteriores a su epoca (Bell [28], Bourgery [35], Ward [338] e incluso Galileo [114]).
La originalidad de Wolff [354] es su interes por representar esa relacion de forma matematica.
Aunque no llego a intentarlo pues no era ni ingeniero ni matematico, sı logro sentar las bases
para hacerlo, no sin cierta controversia entre la comunidad cientıfica de su tiempo con la
que polemizo abiertamente. A pesar de ello, abrio camino a posteriores investigaciones que
culminaron con el primer modelo matematico en 1976. Numerosos trabajos se publicaron
hasta entonces y se siguen publicando, sin proponer modelos matematicos que cuantifiquen
las evidencias experimentales. Por su interes y aplicabilidad a simulaciones computacionales,
como la que se realiza en esta tesis, solo aquellos que sı establecen un modelo matematico,
van a ser revisados aquı.
117

118 Modelos de remodelacion osea interna
5.2. Revision historica de los modelos de remodelacion
osea interna
Antes de pasar a describir los modelos matematicos, se resumen los trabajos de Wolff y
otros autores del siglo XIX, que confirman las observaciones de cientıficos de siglos anteriores
y a su vez sustentan los referidos modelos matematicos. Esta seccion, en la que los modelos se
presentan ordenados cronologicamente, termina con el modelo de remodelacion osea interna
basado en la Mecanica del Dano, desarrollado en la Universidad de Zaragoza por Doblare y
Garcıa [91,115]. A este modelo se dedica el siguiente apartado por la importancia que tiene
en este trabajo: este es el modelo empleado para simular la morfogenesis de la mandıbula,
objeto del capıtulo siguiente.
5.2.1. Primeros conceptos. Ley de Wolff
La idea de que el hueso siente y se adapta a los esfuerzos mecanicos que soporta es
muy antigua y aunque tradicionalmente se atribuye a Wolff [354], este no fue el primero
en darse cuenta. Galileo [114] ya habıa observado las implicaciones mecanicas que tiene
la forma y tamano de los huesos. Bell [28] observo que la estructura del hueso trabecular
esta “relacionada con las fuerzas que actuan sobre el hueso”. Tanto el como Bourgery [35]
reconocıan que la arquitectura del hueso trabecular optimizaba la cantidad de material:
maxima rigidez con la mınima cantidad de material.
Ward [338] comparo la disposicion de las trabeculas en la cabeza del femur con la es-
tructura del soporte de una farola (Fig 5.1). A esa zona de la cabeza del femur se la conoce
como triangulo de Ward. Este descubrimiento abunda en la idea anterior de optimalidad de
la estructura osea: no se dispone material (hueso en este caso) donde no se necesita, al igual
que ocurre en el caso de la farola.
Otro de los conceptos introducidos en esa epoca es la trajectorial theory. Culmann, un
reconocido ingeniero de la segunda mitad del siglo XIX, fue uno de los primeros en darse
cuenta de la importancia de las direcciones principales de tension, hallazgo de Cauchy, hacia
1822, que habıa pasado desapercibido durante decadas, y desarrollo un metodo grafico que
permitıa calcularlas [80]. Habıa estado analizando una columna disenada por Fairbarn y,
segun afirma Roesler [291], no oculto su sorpresa cuando en una visita al laboratorio de un
anatomista contemporaneo suyo, von Meyer, reconocio, en unos dibujos de una seccion de la

5.2 Revision historica de los modelos de remodelacion osea interna 119
Figura 5.1: Comparacion entre la estructura soporte de una farola y la arquitectura trabecular de
la extremidad proximal del femur humano.
extremidad proximal del femur [335], la columna de Fairbarn. En efecto, como puede com-
probarse en la figura 5.2, la coincidencia entre las trayectorias de las direcciones principales
en la columna de Fairbarn y las arcadas de trabeculas en el femur es asombrosa. A pesar de
todo, conviene no olvidar que la columna es una estructura continua y el femur un conjunto
discontinuo de barras y placas de hueso, relleno de medula osea y que en medios discontin-
uos no se puede hablar de direcciones principales de tension. Tampoco se puede establecer
una relacion biunıvoca entre trabeculas y trayectorias de tension, como hace la trajectorial
theory, puesto que hay infinitas trayectorias de tension y solo unas cuantas trabeculas. Este
hecho ha sido pasado por alto durante mucho tiempo como refiere Cowin [72].
A pesar de las evidencias anteriores, solo a partir de los trabajos de Wolff esas ideas
empezaron a ser ampliamente aceptadas. Fue precisamente Wolff quien acuno el termino
trajectorial theory. El pensaba que las trabeculas debıan intersectar siempre en angulo recto:
si las direcciones principales de tension forman 90 grados y las trabeculas siguen la direccion
de estas, es logico que ası ocurra. Por esta razon, critico los dibujos de von Meyer, que
en diversas zonas incluıan trabeculas intersectando de manera oblicua e incluso los rehizo.
Todas sus conclusiones las incluyo en su famoso libro Das Gesetz der Transformation der
Knochen [354] y pronto fueron conocidas como ley de Wolff. Aunque nunca llevo a termino
una formulacion matematica de su ley, es muy posible que tuviera en mente un modelo
120 Modelos de remodelacion osea interna
Figura 5.2: Comparacion entre la arquitectura trabecular del femur y la columna de Fairbarn.
constitutivo continuo para el hueso.
Si bien su papel se considera clave en el desarrollo de la teorıa de la remodelacion osea,
muchas de sus ideas son erroneas, como por ejemplo la teorıa de crecimiento intersticial que
describe la manera en que, segun el, se forma nuevo hueso: las celulas lo producen en el
interior de la matriz osea y el hueso formado empuja al ya existente hacia afuera. Esto es
fısicamente imposible. Mas curioso incluso resulta el hecho de que nunca aceptara la idea
de remodelacion. Maquet y Furlong tradujeron su libro al ingles y Transformation paso a
ser remodeling, cuando el se referıa en realidad a lo que hoy dıa se conoce como modeling.
Segun el, el hueso se adapta a la carga mecanica exterior durante su crecimiento y desarrollo
y tambien durante el proceso de curacion que sigue a un cambio brusco de las condiciones
externas, como por ejemplo una fractura. Sin embargo, despues de que ocurra algo ası, su
comportamiento deja de ser adaptativo. Tambien pensaba que el hueso cortical tenıa la
misma estructura que el trabecular y simplemente era mas denso. Todas estas ideas sin
fundamento, fueron defendidas de forma vehemente por Wolff ante las numerosas crıticas
que recibio y sigue recibiendo [90,325].
Como se refiere en el parrafo anterior, Wolff estaba en contra de uno de los pilares basicos

5.2 Revision historica de los modelos de remodelacion osea interna 121
de la teorıa de la remodelacion: la continua autorregulacion de la estructura osea como res-
puesta a un estımulo mecanico. Bernard (1813-1878) introdujo el concepto de homeostasis
en fisiologıa: los sistemas fisiologicos se autorregulan para mantener constante una deter-
minada variable interna. Roux [293], influenciado por Darwin, establecio en su teorıa de
la adaptacion funcional la hipotesis de que cualquier sistema biologico, no solo los huesos,
posee la habilidad de adaptarse a los cambios que se producen en sus condiciones de vida.
Conviene no confundir adaptacion con evolucion: dichos cambios se pueden producir en dıas
y no es necesario esperar a cambios geneticos a traves de generaciones. Para Roux, una
prueba de que su teorıa era cierta es el principio de optimalidad antes comentado.
En resumen, los cientıficos del siglo XIX habıan legado tres ideas fundamentales para
el desarrollo futuro de este campo, como apunta Roesler [291]: la estructura osea maximiza
su resistencia con el mınimo peso, las trabeculas se orientan en promedio siguiendo las
direcciones principales de tension y a la vez el hueso se autorregula mediante la respuesta
de las celulas oseas a determinados estımulos mecanicos.
5.2.2. Primeros modelos matematicos. Pauwels y Kummer
Pasaron muchos anos en los que estas ideas estuvieron practicamente en el olvido. Coinci-
diendo con la aparicion de los computadores y el creciente interes por los metodos numericos,
los trabajos de Pauwels y Kummer las rescataron con la intencion de formular un modelo
constitutivo de la adaptacion osea.
Basandose en las ideas de Roux, Pauwels [266] cuantifico el estımulo funcional que con-
duce a la atrofia o hipertrofia de hueso, mediante la tension. Propuso la existencia de una
tension optima, que no provoca una variacion neta de la cantidad de hueso (la atrofia y
la hipertrofia se compensan). Cuando el nivel de tension supera este valor se produce una
formacion neta de hueso, mientras que si esta por debajo se produce reabsorcion. Un nivel
de tensiones mucho mayor que el optimo conduce igualmente a la perdida de hueso debido
a los danos que puede producir.
El papel de la deformacion no esta muy claro en este modelo, que lo deja todo en manos
de la tension. Aunque deformacion y tension estan ıntimamente relacionadas, se conoce hoy
en dıa que es realmente la deformacion la que juega el papel primordial en los mecanismos
celulares que provocan la respuesta al estımulo.
Pauwels veıa en su modelo una explicacion de la trajectorial theory. Segun el, las trabecu-
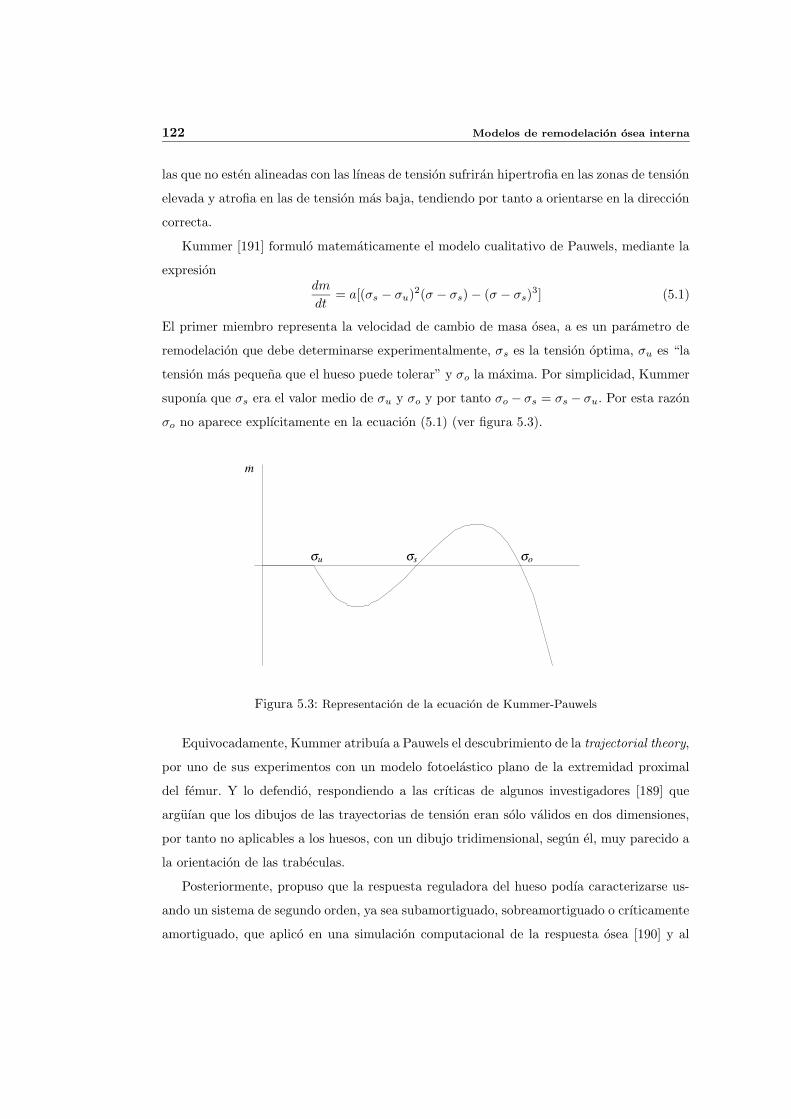
122 Modelos de remodelacion osea interna
las que no esten alineadas con las lıneas de tension sufriran hipertrofia en las zonas de tension
elevada y atrofia en las de tension mas baja, tendiendo por tanto a orientarse en la direccion
correcta.
Kummer [191] formulo matematicamente el modelo cualitativo de Pauwels, mediante la
expresiondm
dt= a[(σs − σu)2(σ − σs)− (σ − σs)3] (5.1)
El primer miembro representa la velocidad de cambio de masa osea, a es un parametro de
remodelacion que debe determinarse experimentalmente, σs es la tension optima, σu es “la
tension mas pequena que el hueso puede tolerar” y σo la maxima. Por simplicidad, Kummer
suponıa que σs era el valor medio de σu y σo y por tanto σo − σs = σs − σu. Por esta razon
σo no aparece explıcitamente en la ecuacion (5.1) (ver figura 5.3).
m
σu σs σo
1
Figura 5.3: Representacion de la ecuacion de Kummer-Pauwels
Equivocadamente, Kummer atribuıa a Pauwels el descubrimiento de la trajectorial theory,
por uno de sus experimentos con un modelo fotoelastico plano de la extremidad proximal
del femur. Y lo defendio, respondiendo a las crıticas de algunos investigadores [189] que
arguıan que los dibujos de las trayectorias de tension eran solo validos en dos dimensiones,
por tanto no aplicables a los huesos, con un dibujo tridimensional, segun el, muy parecido a
la orientacion de las trabeculas.
Posteriormente, propuso que la respuesta reguladora del hueso podıa caracterizarse us-
ando un sistema de segundo orden, ya sea subamortiguado, sobreamortiguado o crıticamente
amortiguado, que aplico en una simulacion computacional de la respuesta osea [190] y al

5.2 Revision historica de los modelos de remodelacion osea interna 123
problema de reabsorcion osea que se presenta en vuelos espaciales, por la condicion de in-
gravidez [192].
La contribucion principal de estos autores fue renovar el interes por el problema de la
remodelacion osea, que a partir de entonces experimento un auge muy importante. Aunque
llegaron a establecer varias relaciones matematicas de la adaptacion mecanica del hueso,
no culminaron sus trabajos con una ley constitutiva continua, que relacionara tensiones y
deformaciones en el hueso, como harıa posteriormente Stephen Cowin [74].
5.2.3. Teorıa de la elasticidad adaptativa de Cowin
En 1976 aparece el primer modelo matematico continuo de remodelacion osea, en un
artıculo de Cowin y Hegedus [74]. Es un modelo termomecanico continuo que considera al
hueso como un material poroelastico con una fase solida, llamada matriz osea, rodeada de
un fluido (medula osea). Ambas fases intercambian masa, como consecuencia de reacciones
quımicas en las que las celulas oseas desempenan un papel fundamental. La velocidad de
dichas reacciones, muy pequena, con tiempos caracterısticos del orden de meses, esta con-
trolada por el nivel de deformaciones, segun postulan estos autores. Como consecuencia del
flujo de masa que se establece entre el fluido y la matriz osea, se produce tambien una trans-
ferencia de energıa, cantidad de movimiento y entropıa, que el modelo cuantifica y en los
que influye igualmente el nivel de tensiones. Como consecuencia del referido flujo de masa,
disminuye o aumenta la porosidad de la matriz osea que ve ası modificado su modulo de
elasticidad.
Plantean, como en cualquier problema elastico, las ecuaciones de equilibrio (5.2), com-
patibilidad (en pequenas deformaciones) (5.3) y comportamiento (5.4):
∇σ + γνb = 0 (5.2)
εij =1
2
(∂ui∂xj
+∂uj∂xi
), (5.3)
σ = (ξ0 + e) C : ε, (5.4)
donde σ y ε son los tensores de tension y de pequenas deformaciones respectivamente, b es
el vector de fuerzas por unidad de volumen, γ y ν son la densidad y la fraccion volumetrica
de matriz osea en el estado deformado, respectivamente, u es el campo de desplazamientos,
ξ0 la fraccion volumetrica de matriz osea en el estado indeformado al comienzo del analisis,
C es el tensor de comportamiento del material y por ultimo, e es la variacion de la fraccion

124 Modelos de remodelacion osea interna
volumetrica de matriz osea en el estado indeformado como consecuencia de la transferencia
de masa que se produce entre matriz osea y fluido extracelular. Las fracciones volumetricas
de matriz osea en los estados deformado e indeformado, ν y ξ, estan relacionadas a traves
de la deformacion volumetrica, que en pequenas deformaciones es igual a trε.
ξ = (ξ0 + e) = ν(1 + tr ε) (5.5)
La velocidad de las reacciones quımicas que determinan esta transferencia de masa de-
penden, segun este modelo, de las deformaciones a las que se encuentra sometido el hueso y
de la fraccion volumetrica de hueso. Ası, la velocidad de variacion de la fraccion volumetrica
la expresan Cowin y Hegedus de la siguiente forma:
e = a(e) + A(e) : ε (5.6)
Esta ecuacion de remodelacion (5.6) surge como un desarrollo en serie de la funcion generica
e(e, ε), que es lo que inicialmente plantean estos autores [74]. En un segundo trabajo [142],
llevan a cabo dicho desarrollo en serie, incluyendo tan solo terminos de orden e, e2, ε y
e ε, despreciando los de orden superior y los terminos en ε2, de acuerdo con la hipotesis de
pequenas deformaciones.
e = c0 + c1e+ c2e2 + A0 : ε+ eA1 : ε (5.7)
Esta aproximacion tiene sentido para valores pequenos de e, lo cual esta justificado en huesos
sanos pero deja de ser cierto en determinadas patologıas de interes, como la osteoporosis. Hay
que indicar que los coeficientes c0, c1 y c2, ası como las matrices A0 y A1 son propiedades
del material que deben obtenerse experimentalmente.
Cowin y Hegedus resolvieron esta ecuacion diferencial para los casos en que el tensor de
deformaciones o el tensor de tensiones se modifica en t = 0 y permanece estacionario de ahı en
adelante. Dicha ecuacion diferencial, ordinaria, de primer orden y no lineal, proporciona la
solucion de un problema homogeneo de remodelacion. Estos autores dieron igualmente las
condiciones para que la solucion de e cuando t tiende a infinito este acotada, correspondi-
ente a una situacion de equilibrio con una porosidad determinada. Si no se cumplen dichas
condiciones e tendera hacia alguno de sus valores extremos 1− ξ0 > e > −ξ0. Firoozbaksh y
Cowin [99], mostraron con posterioridad, resolviendo para ello la ecuacion (5.7) en un caso
no homogeneo, como las heterogeneidades iniciales se van suavizando con el tiempo, tendi-
endo la solucion hacia una situacion homogenea si el campo de tensiones o deformaciones

5.2 Revision historica de los modelos de remodelacion osea interna 125
es uniforme. En el caso de que se parta de una situacion heterogenea y se impongan unas
condiciones de deformacion no uniforme es necesario resolver el problema elastico (ecuaciones
(5.2)-(5.5)) conjuntamente con la ecuacion de remodelacion (5.6).
En otro trabajo, Firoozbaksh y Cowin [100] modificaron el desarrollo en serie (5.7) para
incluir terminos de orden e3 y lo que es mas importante, de orden ε2. Demostraron que
este modelo de remodelacion cubico reproduce la ecuacion de Kummer-Pauwels (5.1), que
tambien es cubica en las tensiones. Ademas, el hecho de retener terminos de orden ε2 en la
ecuacion de remodelacion tiene dos consecuencias muy importantes:
a) Un estado de torsion pura induce a la remodelacion, a diferencia del modelo descrito
por la ecuacion (5.7), lineal en deformaciones.
b) Una carga cıclica con valor medio nulo induce a la remodelacion si se incluyen terminos
de orden ε2, pero no en un modelo lineal en deformaciones.
Hasta aquı lo referido a remodelacion interna. Con respecto a remodelacion externa,
tambien se plantea una ecuacion que relaciona el movimiento de la superficie externa del
hueso (ya sea periostio o endostio) con las componentes del tensor de deformaciones en dicha
superficie:dX
dt= B : (ε− ε0) (5.8)
donde X es la coordenada normal a la superficie (se supone que esta no se mueve en las otras
dos direcciones), ε0 es el tensor de deformaciones de referencia, para el que no se produce
remodelacion y B es un tensor de coeficientes de remodelacion externa, que se determina
experimentalmente, imponiendo que la teorıa reproduzca determinados ensayos clınicos.
En la teorıa precedente anterior, Cowin se cine a lo que ocurre en el hueso cortical. En
1986, da una relacion constitutiva para el hueso esponjoso [71], cuantificando su arquitectura
trabecular a traves del denominado fabric tensor, H. Este, es un tensor de segundo orden
definido positivo cuyas direcciones principales coinciden con las direcciones promedio de las
trabeculas y cuyos valores principales son proporcionales a la cantidad de masa de tejido en
la direccion correspondiente.
Cowin postula que la anisotropıa del hueso esponjoso es debida a su arquitectura trabec-
ular, siendo la matriz osea de la que esta compuesto, intrınsecamente isotropa. Ası, relaciona
el tensor de comportamiento con la fraccion volumetrica de matriz osea ν y con H, estable-
ciendo la ley constitutiva de la siguiente forma:
σ = β11 + β2ε+ β3H + β4H2 + β5(Hε+ εH) + β6(H
2ε+ εH2) (5.9)

126 Modelos de remodelacion osea interna
en la que todos los parametros βi son funciones de e, trH, trH2 y trH3 y algunos pueden
ser funciones de la deformacion, en concreto de tr ε, tr(εH) y tr(εH2), si se mantiene la
dependencia lineal del modelo con la deformacion.
Se define el equilibrio de remodelacion como la ausencia de remodelacion, en dos sentidos:
no hay reabsorcion ni formacion neta de tejido y tampoco existe variacion de la orientacion
de las trabeculas. De esta forma, el equilibrio de remodelacion se caracteriza por una de-
terminada arquitectura trabecular (ν∗, H∗) y un cierto estado de tension-deformacion (σ∗,
ε∗) que se mantiene dentro de un rango. La ley de Wolff determina que, en el equilibrio,
la orientacion de las trabeculas siga las direcciones principales de tension. Esta condicion
se expresa diciendo que los tensores σ∗ y H∗ se encuentran alineados (tienen direcciones
principales coincidentes), o matematicamente:
σ∗H∗ = H∗σ∗ (5.10)
Ademas, debe cumplirse que la direccion en la que hay mayor cantidad de material, sea
tambien la de mayor rigidez. Si los modulos de Young en direcciones principales se ordenan
ası: E1 > E2 > E3, entonces los autovalores del fabric tensor lo estaran: H1 > H2 > H3.
En la situacion de equilibrio tambien se verifica que el tensor de deformacion tambien se
encuentra alineado con los anteriores. Ası pues, la ley de Wolff puede resumirse de la siguiente
forma:
σ∗H∗ = H∗σ∗ ε∗H∗ = H∗ε∗ ε∗σ∗ = σ∗ε∗ (5.11)
Si partiendo de una situacion de equilibrio de remodelacion (ν0,H0,ε0,σ0), se modifica
el estado tensional y se mantiene estacionario (σ∗) se modifica la fraccion volumetrica de
matriz osea y la orientacion de las trabeculas segun las leyes de flujo
H = α11 + α2ε+ α3H + α4H2 + α5(Hε+ εH) + α6(H
2ε+ εH2), (5.12)
e = e(e, trε, trH, trH2, trH3, trεH , trεH2), (5.13)
hasta alcanzar un nuevo estado de equilibrio (ν∗,H∗,ε∗,σ∗) en el que los tres tensores esten
alineados y no se produzca variacion neta de la masa de tejido. En las anteriores ecuaciones
de flujo, las funciones αi dependen de los mismos parametros que las βi.
La gran cantidad de variables que aparecen en el modelo y que deben determinarse ex-
perimentalmente, hacen de su empleo una ardua tarea. De hecho se han resuelto unicamente

5.2 Revision historica de los modelos de remodelacion osea interna 127
problemas muy sencillos de regiones de hueso homogeneas, mediante metodos analıticos [75],
no habiendose implementado aun en un modelo de elementos finitos.
5.2.4. Carter: el concepto de auto-optimizacion
Basandose en las ideas de Pauwels, esbozadas con anterioridad, Carter y sus colabo-
radores introdujeron una formulacion matematica del estımulo funcional que controla la
atrofia o hipertrofia en el hueso trabecular. Debe existir una cierta magnitud, que el hue-
so controle de manera natural, de tal forma que mediante mecanismos biologicos dicha
magnitud, o estımulo, se pueda mantener en un cierto rango, que caracteriza el estado
cuasi-estacionario de remodelacion osea nula.
Pauwels suponıa que la tension tiene un papel fundamental en dicho estımulo y ası lo
entendio Kummer que cuantifico el modelo de Pauwels como ya se ha dicho. En concreto,
dio la respuesta del hueso a dicho estımulo en funcion de la tension local, ecuacion (5.1).
Carter et al. [57] proponen una definicion alternativa del estımulo (S), que tiene en cuenta
el hecho de que el proceso de remodelacion esta influenciado por la historia de carga a la
que se somete a un tejido oseo durante un cierto perıodo de tiempo.
S ∝N∑
i=1
niσmi (5.14)
donde ni es el numero de veces que se realiza a diario la actividad i, N es el numero de
actividades distintas que se realiza habitualmente y σi es una tension efectiva asociada a
dicha actividad y que se define a continuacion. La principal aportacion de este modelo es
precisamente que incluye la influencia del numero de ciclos en el estımulo (ni), ası como
la historia de carga reciente, a traves de la combinacion de los distintos casos de carga
(i = 1, ..., N). La influencia del estado tensional se tiene en cuenta a traves de la mencionada
tension efectiva, que se define de la siguiente forma:
σi =√
2E(ρ)Ui(σi), (5.15)
como funcion del modulo de Young E, que depende de la densidad del hueso, ρ, y de la
energıa de deformacion correspondiente a cada caso de carga, Ui, que depende del tensor de
tensiones local, σi.
Que una carga dinamica es mas efectiva para activar la remodelacion osea que una carga
estatica, es un hecho ampliamente aceptado [196,252,295]. Lanyon y Rubin [196] demostraron

128 Modelos de remodelacion osea interna
que una carga constante aplicada in vivo, por medio de una serie de muelles e inmovilizando
el hueso, provoca reabsorcion en el hueso, como ocurrirıa si no estuviera sometido a ningun
tipo de carga. Sin embargo, una carga moderada repetida periodicamente puede inducir la
remodelacion osea, dependiendo del nivel de tension y del numero de ciclos que ocurra la
carga en el perıodo de estudio; por tanto, dependera de forma indirecta de la velocidad de
aplicacion de la carga. El peso de uno y otro factor esta controlado por el exponente m, que
Whalen y Carter [343] estiman entre 3 y 8, dependiendo del tipo de actividad.
Un sistema biologico de remodelacion osea ha de definir un determinado funcional de
adaptacion, que permita al hueso maximizar su integridad estructural con la menor cantidad
posible de masa osea. De esta forma, la tension actua como una herramienta de optimizacion
que minimiza una determinada funcion objetivo, directamente relacionada con la integridad
estructural [112]. A partir de esta idea, el modelo propone una relacion entre la densidad
aparente y la tension efectiva, de la siguiente forma:
ρ ∝( N∑
i=1
niσmi
)1/2m
(5.16)
Este modelo fue implementando, usando el MEF para el calculo de las tensiones locales,
y usado para predecir la distribucion de densidades en la extremidad proximal del femur.
Se parte de una distribucion homogenea de densidad y se aplican varios casos de carga que
simulan el proceso de caminar.
Carter et al. [59] tambien tratan el problema de la anisotropıa. Establecen que las trabecu-
las tienden a orientarse segun la direccion en que es maxima una cantidad denominada ten-
sion equivalente normal, σn, que tambien tiene en cuenta la contribucion de los distintos
casos de carga:
σn(n) =
[ N∑
i=1
(nint
)(nσi n)m
]1/m, (5.17)
donde nt es el numero total de ciclos aplicados entre los distintos casos de carga y n es un
vector unitario en la direccion considerada.
5.2.5. El modelo isotropo de Huiskes
Huiskes et al. [158] plantean un modelo de remodelacion osea adaptativa que da el papel
preponderante a la deformacion, por medio de la energıa de deformacion U(ε), en lugar de a
la tension, como ocurre en los modelos anteriores. El modelo incorpora igualmente una idea

5.2 Revision historica de los modelos de remodelacion osea interna 129
de Carter [55], la de que el hueso responde con una cierta “pereza” al estımulo mecanico.
Existe una “zona muerta” (de ancho 2s, expresado en tanto por uno de U0), tal que, mientras
la energıa de deformacion se mantenga dentro de ella, la remodelacion no tiene efecto neto,
como pretende mostrar la figura 5.4.
dE
dt,dX
dt
2s
C
C
UU0
Figura 5.4: Remodelacion interna-externa del hueso en funcion de la energıa de deformacion
En remodelacion interna el modelo proporciona la velocidad de variacion del modulo de
Young, E, y en remodelacion externa la velocidad de variacion de la geometrıa, en concreto
de la coordenada en la direccion del vector unitario normal a la superficie, X:
dE
dt=
Ce(U − (1 + s)U0
)si U > (1 + s)U0
0 si (1− s)U0 ≤ U ≤ (1 + s)U0
Ce(U − (1− s)U0
)si U < (1− s)U0
(5.18)
dX
dt=
Cx(U − (1 + s)U0
)si U > (1 + s)U0
0 si (1− s)U0 ≤ U ≤ (1 + s)U0
Cx(U − (1− s)U0
)si U < (1− s)U0
(5.19)
donde Ce y Cx son parametros del modelo que controlan la remodelacion interna y externa
respectivamente mientras que U0 es el valor de equilibrio de la energıa de deformacion.
Esta formulacion ha sido implementada en modelos de elementos finitos y aplicada a
diversos ejemplos: remodelacion externa de una viga en voladizo de material oseo, prediccion
de la distribucion de densidad en la extremidad proximal del femur y prediccion del fenomeno
de “stress shielding” cuando en dicha extremidad se incorpora una protesis. La remodelacion

130 Modelos de remodelacion osea interna
interna tiene el problema de que puede conducir a modulos elasticos negativos si no se
preve esta posibilidad a priori. En remodelacion externa se pueden presentar problemas con
la forma de los elementos si la geometrıa se altera en exceso.
Weinans, colaborador de Huiskes, presenta en un trabajo posterior [341] un modelo de
comportamiento mecanico que tiene como variable independiente la densidad aparente, en
lugar del modulo de elasticidad:
dρ
dt= B
(Uaρ− k)
con 0 < ρ ≤ ρ (5.20)
donde ρ es la densidad aparente, B y k son constantes del modelo y Ua tiene en cuenta la
contribucion a la energıa de deformacion de todos los casos de carga:
Ua =1
N
N∑
i=1
Ui (5.21)
Si esta ley se implementa en un modelo de elementos finitos es necesario conocer como
se relaciona el modulo de Young del hueso con la densidad aparente, que es la variable
independiente del modelo. Para ello se utilizan correlaciones entre ambas variables, obtenidas
experimentalmente.
Hay que indicar que la forma de la ecuacion de remodelacion (5.20) hace que huesos
con valores patologicos de densidad sean muy sensibles a la carga en su respuesta a la
remodelacion.
Este modelo ha sido aplicado principalmente en el diseno de implantes protesicos para la
extremidad proximal del femur [154–156]. Huiskes ha llegado incluso a disenar una protesis
de fijacion de cadera (protesis femoral SHP), que se usa en la actualidad, basandose en
dichos trabajos. Por ultimo, hay que decir que este modelo no ha sido ampliado aun para
intentar reproducir la anisotropıa del hueso, siendo precisamente este caracter isotropo la
principal limitacion.
5.2.6. Modelo de remodelacion osea interna isotropa de Stanford
Este modelo se va a comentar con mas detalle que los anteriores, porque contiene muchas
ideas utilizadas en el modelo anisotropo basado en la mecanica del dano de Doblare y Garcıa,
que sera explicado mas adelante y que ha sido utilizado en algunas simulaciones de esta tesis.
El modelo isotropo de Stanford fue desarrollado a finales de la decada de los 80 y princi-
pios de los 90 del siglo pasado, en dicha universidad, bajo la direccion del Dr. Dennis Carter,

5.2 Revision historica de los modelos de remodelacion osea interna 131
de cuyo modelo, ası como de otros tambien referidos anteriormente, toma algunas ideas im-
portantes. En primer lugar, establece una condicion de remodelacion homeostatica a nivel
local, como sugieren los trabajos de Carter et al. [57], Fyhrie y Carter [112], Frost [109] y
Turner [328]. Esto quiere decir que el tejido oseo dispone de una serie de mecanismos para
autorregularse y mantener dentro de un cierto rango de valores una determinada magnitud
fisiologica, denominada estımulo mecanico. Esos mecanismos tienen como mision primera
modificar la densidad osea y, relacionadas con ella, las propiedades mecanicas, con el objeti-
vo global de homogeneizar el estımulo local y mantenerlo dentro del rango antes comentado.
El estımulo mecanico local, Ψt, tambien denominado estımulo tensional tisular diario, es
una magnitud que esta relacionada con la carga a la que se encuentra sometido el tejido y
que, de forma muy parecida a la que lo hace Carter (ec. (5.14)), tiene en cuenta los distintos
casos de carga que conforman la actividad diaria habitual:
Ψt =
( N∑
i=1
niσmti
)1/m
(5.22)
donde N es, como hasta ahora, el numero de casos de carga, ni es el numero promedio
de ciclos diarios del caso de carga i, σti es la tension efectiva local para el caso de carga
i, parametro escalar que representa la intensidad del estado tensional local en el tejido
mineralizado y m es de nuevo el exponente de la ecuacion (5.14).
Jacobs [166] propone una simplificacion de la historia de carga que tiene en cuenta la
escala de tiempos tan diferente en que ocurren la variacion de las cargas y la respuesta del
hueso. Mientras que las tensiones pueden variar en tiempos del orden de segundos, al cam-
inar por ejemplo, o incluso menos en el caso de un impacto, la respuesta de remodelacion
tiene tiempos caracterısticos del orden de semanas o meses. Por ello, es conveniente sustituir
las tensiones o deformaciones instantaneas por variables cuasi-estaticas que contienen infor-
macion sobre todas las actividades que se producen durante la historia reciente de carga.
En concreto, la informacion que interesa de cada actividad es la tension maxima promedio
que origina y el numero de ciclos que tiene lugar dicha actividad en el perıodo de control en
el que se toma dicho promedio. La duracion de este perıodo no afecta de manera relevante
a la respuesta de la remodelacion, siempre que no sea excesivamente grande ni pequeno.
Por motivos practicos, se suelen agrupar las distintas actividades diarias segun sea su
influencia en la remodelacion. Con esta aproximacion, se resume la historia de carga en una
serie reducida de actividades, cada una de ellas con unas determinadas condiciones de apoyo,

132 Modelos de remodelacion osea interna
de carga y un numero de ciclos que dicha carga se repite diariamente. Ademas, se supone
que el orden de aplicacion de las cargas no es relevante en la respuesta, dada la diferencia de
escala de tiempos ya comentada. La figura 5.5 muestra como se agruparıan las cargas segun
niveles de tension similares.
σ
1
2 2
3
t
Figura 5.5: Evolucion temporal de las tensiones en una actividad generica. Cada una de las activi-
dades con rangos de tensiones similares se agrupan en un caso de carga, como los senalados en la
grafica: 1, 2 y 3.
Despues de agrupar las cargas, Jacobs [166], vio necesario hacer algun tipo de simplifi-
cacion de la expresion (5.22), para que el algoritmo pudiera ser implementado de manera
sencilla. Aunque se puede observar en dicha expresion que todos los casos de carga con-
tribuyen de manera simultanea al evaluar el estımulo diario, Jacobs, los aplica de manera
secuencial. Supongase, por un momento, que existen tres casos de carga como los de la figura
5.5. Despues de promediar las tensiones y contabilizar el numero de ciclos correspondientes
a cada actividad en un tiempo de muestreo suficientemente amplio, se supone que todos los
dıas se repite la misma secuencia de ciclos. El histograma de actividades quedara entonces
como muestra la figura 5.6. La simplificacion propuesta por Jacobs modifica dicho histogra-
ma, aplicando cada dıa una sola actividad, secuencialmente y repitiendo alguna de ellas,
si es necesario, para que la proporcion entre los numeros de ciclos de cada actividad y el
numero de ciclos total sea igual a la del histograma original. El histograma despues de la
agrupacion de cargas quedarıa como en la figura 5.7, repitiendose la secuencia representada
cada cuatro dıas. De esa forma, las actividades 1 y 3 ocurren el mismo numero de veces y
la mitad de veces que la actividad 2, al igual que en el histograma original de la figura 5.6.
5.2 Revision historica de los modelos de remodelacion osea interna 133
1 2 3 4 5 6 7 8
4
8
12
16
Actividad 1
Actividad 2
Actividad 3
dıas
no
cicl
osdia
rios
Figura 5.6: Distribucion del numero de ciclos correspondiente a cada caso de carga a lo largo del
tiempo, antes de realizar la simplificacion propuesta por Jacobs.
1 2 3 4 5 6 7 8
4
8
12
16
Actividad 1
Actividad 2
Actividad 3
dıas
no
cicl
osdia
rios
Figura 5.7: Distribucion del numero de ciclos correspondiente a cada caso de carga a lo largo del
tiempo, despues de realizar la simplificacion.

134 Modelos de remodelacion osea interna
Con esta simplificacion, el numero de ciclos que se repite cada actividad es el mismo, 16
en el ejemplo de las figuras anteriores. Si en la simulacion la unidad de tiempo es un dıa, ya
no es necesario superponer diariamente los casos de carga para calcular el estımulo mecanico
segun la ecuacion (5.22). Ahora se puede aplicar cada dıa una sola actividad, que produce
un estımulo:
Ψt = n1/mc σt donde nc =
N∑
i=1
ni, (5.23)
y repetir esa actividad varios dıas consecutivos si es necesario, como lo es con la actividad
2 del ejemplo anterior. Jacobs comprobo que la diferencia entre la aplicacion simultanea y
la aplicacion secuencial de las cargas no es significativa, mientras el numero de dıas que
dure una secuencia no sea excesivo. Sin embargo el coste computacional sı disminuye de
forma muy favorable con esta aproximacion. Que la secuencia de agrupamiento no resulte
muy larga dependera del numero de actividades que se tengan en cuenta. En un ejemplo
tan sencillo como el anterior, con cuatro dıas basta, pero si el numero de actividades es
mas amplio, es conveniente hacer un nuevo agrupamiento para reducirlo en la medida de lo
posible.
Otra hipotesis de este modelo es la de suponer que la matriz osea esta completamente
mineralizada, con una densidad local igual en todos los puntos, ρ. De esta forma, la densidad
aparente del tejido oseo, ρ, definida como la masa de matriz osea contenida en la unidad de
volumen de tejido, depende tan solo de la porosidad local, n, y no del grado de mineralizacion
de la matriz osea, que se supone que no varıa en este modelo.
n = 1− VmVT
= 1− ρ
ρ(5.24)
Segun Martin [213] y el propio Jacobs [166] el hueso cortical presenta una porosidad
mınima de 0.05. Jacobs asigna ademas al hueso cortical de mınima porosidad una densidad
aparente de 1.92 g/cm3, de lo que se deduce facilmente el valor asignado a la densidad de la
matriz osea, ρ = 2.02 g/cm3.
Para poder tratar al hueso como un medio continuo, es necesario pasar de la escala
microscopica a nivel de tejido, en el que el material es heterogeneo, al nivel continuo. Esto
se hace de forma indirecta, relacionando la resistencia ultima del hueso trabecular a nivel
continuo, σult, con la resistencia ultima del tejido oseo trabecular, σultt , que se supone
constante. Gibson [123] establece que ambas magnitudes estan relacionadas a traves de la
densidad aparente por medio de alguna funcion R(ρ).

5.2 Revision historica de los modelos de remodelacion osea interna 135
σult(ρ) = R(ρ)σultt (5.25)
La evidencia experimental [58] muestra que esta funcion debe ser proporcional al cuadra-
do de la densidad aparente. Comoquiera que la resistencia ultima a nivel continuo y a nivel
tisular deben coincidir en el caso de porosidad nula, la funcion debe ser tal que R(ρ) = 1.
De esta forma la ecuacion (5.25) se puede reescribir de la siguiente forma [26]:
σult(ρ) =(ρρ
)2
σultt (5.26)
Esta relacion se debe cumplir no solo para la resistencia ultima sino para cualquier valor
de tension, y en particular para la tension efectiva. Ası, sustituyendo en la ecuacion (5.22)
la tension efectiva a nivel tisular o local por la tension efectiva a nivel continuo y operando,
se puede obtener el estımulo tensional diario continuo
Ψ =
( N∑
i=1
niσmi
)1/m
(5.27)
que se relaciona con el estımulo tensional diario tisular de la misma forma que las tensiones
Ψt =( ρρ
)2
Ψ (5.28)
y que es posible simplificar de la misma manera que se hizo en la ecuacion (5.23)
Ψ = n1/mc σ (5.29)
Si es posible de alguna manera determinar la distribucion de tensiones y deformaciones
a nivel continuo en un determinado hueso, por ejemplo por medio del MEF, con la ecuacion
(5.15) se puede determinar la tension efectiva continua y usando las ecuaciones (5.28) y
(5.29) obtener el estımulo tensional tisular diario, Ψt. Una vez conocido este, el siguiente
paso consiste en determinar cuan lejos esta el tejido de cumplir el equilibrio homeostatico.
Para ello se define el error de remodelacion, e, como la diferencia entre el estımulo tensional
tisular diario y un valor de referencia que se considera de equilibrio, Ψ∗
t .
e = Ψt −Ψ∗
t (5.30)
Si este error de remodelacion no es nulo o esta fuera de un determinado rango, se produce
una respuesta del hueso que se traduce en una variacion de la densidad aparente a nivel local.
Sin embargo, el modelo no relaciona de forma directa ambas variables, sino a traves de la
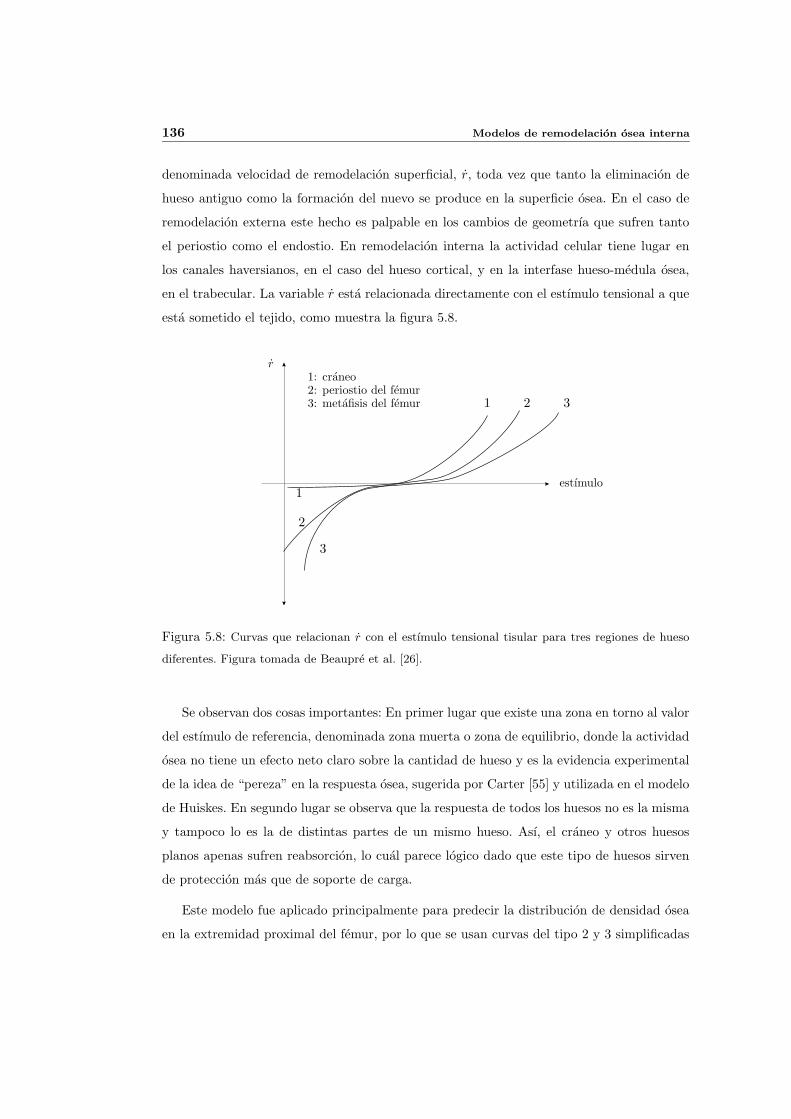
136 Modelos de remodelacion osea interna
denominada velocidad de remodelacion superficial, r, toda vez que tanto la eliminacion de
hueso antiguo como la formacion del nuevo se produce en la superficie osea. En el caso de
remodelacion externa este hecho es palpable en los cambios de geometrıa que sufren tanto
el periostio como el endostio. En remodelacion interna la actividad celular tiene lugar en
los canales haversianos, en el caso del hueso cortical, y en la interfase hueso-medula osea,
en el trabecular. La variable r esta relacionada directamente con el estımulo tensional a que
esta sometido el tejido, como muestra la figura 5.8.
r1: craneo2: periostio del femur3: metafisis del femur
estımulo1
1
2
2
3
3
Figura 5.8: Curvas que relacionan r con el estımulo tensional tisular para tres regiones de hueso
diferentes. Figura tomada de Beaupre et al. [26].
Se observan dos cosas importantes: En primer lugar que existe una zona en torno al valor
del estımulo de referencia, denominada zona muerta o zona de equilibrio, donde la actividad
osea no tiene un efecto neto claro sobre la cantidad de hueso y es la evidencia experimental
de la idea de “pereza” en la respuesta osea, sugerida por Carter [55] y utilizada en el modelo
de Huiskes. En segundo lugar se observa que la respuesta de todos los huesos no es la misma
y tampoco lo es la de distintas partes de un mismo hueso. Ası, el craneo y otros huesos
planos apenas sufren reabsorcion, lo cual parece logico dado que este tipo de huesos sirven
de proteccion mas que de soporte de carga.
Este modelo fue aplicado principalmente para predecir la distribucion de densidad osea
en la extremidad proximal del femur, por lo que se usan curvas del tipo 2 y 3 simplificadas

5.2 Revision historica de los modelos de remodelacion osea interna 137
como en la figura 5.9 y que se pueden expresar:
r =
c1(e+ w) si e < −w0 si −w ≤ e ≤ wc2(e− w) si e > w
(5.31)
r(µm/dıa)
w w
c1
c2
e
Figura 5.9: Ley de remodelacion usada por Jacobs [166].
Algunos autores han propuesto variaciones a esta ley de remodelacion [269,315]. Una de
ellas es suponer, parece logico, que el hueso no puede crecer o ser reabsorbido de forma ilim-
itada. La ley de remodelacion quedarıa entonces como muestra la figura 5.10. Sin embargo,
dado que en situaciones normales el estımulo mecanico se mantiene relativamente proximo
al valor de equilibrio, dicha modificacion no introduce cambios significativos.
Como ya se ha dicho, el hueso aparece y desaparece en la superficie osea y ası se ha
introducido la variable velocidad de remodelacion superficial, que se define como el volumen
de hueso generado o eliminado por unidad de tiempo y por unidad de superficie disponible
para la remodelacion. Martin [213] definio la superficie especıfica, Sv, como la cantidad de
superficie disponible para la remodelacion por unidad de volumen de hueso y la relaciono con
la porosidad, encontrando que un polinomio de quinto grado (ecuacion (5.32)) se ajusta a
huesos de distintas zonas del cuerpo, de diferentes edades y estados de salud. En la figura 5.11
se representa esta correlacion pero frente a la densidad aparente en lugar de la porosidad.
Sv = 32.3n− 93.9n2 + 134n3 − 101n4 + 28.8n5 (5.32)
138 Modelos de remodelacion osea interna
r(µm/dıa)
w w
c1
c2e
rmax
rmin
Figura 5.10: Ley de remodelacion modificada.
1
2
3
4
Sv(mm
2/m
m3)
ρ(g/cm3)
Figura 5.11: Superficie especıfica en funcion de la densidad aparente.

5.2 Revision historica de los modelos de remodelacion osea interna 139
Se recuerda que en este modelo el hueso se encuentra completamente mineralizado y ası lo
estara pues, tanto el hueso que se elimina como el nuevo que se forma sobre la superficie osea.
De esta forma la variacion de la densidad aparente viene dada por la siguiente expresion:
ρ = k r Sv ρ, (5.33)
en la que todas las variables han sido ya definidas a excepcion de k, que es el porcentaje
de superficie disponible que se encuentra activa para la remodelacion y que normalmente se
toma igual a la unidad.
Para obtener la densidad aparente en el siguiente paso de carga, se integra la ecuacion
(5.33) mediante un algoritmo de integracion de Euler explıcito,
ρ(t+ ∆t) = ρ(t) + ρ(t)∆t, (5.34)
y finalmente se actualizan las propiedades elasticas en cada punto del dominio solido (en
la malla de elementos finitos se hace en los puntos de integracion de Gauss) segun las
expresiones experimentales obtenidas por Beaupre et al. [27, 166] y teniendo en cuenta que
el hueso se supone un material isotropo:
C =E
1 + ν
( ν
1− 2ν1⊗ 1 + I
), (5.35)
E =
2014ρ2.5 si ρ ≤ 1.2g/cm3
1763ρ3.2 si ρ > 1.2g/cm3,(5.36)
ν =
0.2 si ρ ≤ 1.2g/cm3
0.32 si ρ > 1.2g/cm3,(5.37)
donde 1 es el tensor identidad de segundo orden y I es el tensor identidad simetrico de
cuarto orden (Iijkl = 12 (δikδjl + δilδjk)). Con estas expresiones se completa el algoritmo y
se puede resolver el problema elastico con las propiedades actualizadas, por ejemplo con el
MEF, como ya se ha comentado.
Un problema que presenta este algoritmo cuando se aplica a una malla de elementos fini-
tos es el denominado “checkerboard”: se obtienen distribuciones de densidad con un tipo de
discontinuidad muy particular, que consiste en la alternancia de elementos de densidad maxi-
ma y elementos de densidad nula. Este problema se resuelve mediante tecnicas de proyeccion
(ver por ejemplo [359]), mediante las cuales la densidad o cualquier otra magnitud derivada
de las variables nodales, como por ejemplo las tensiones, que se evaluan normalmente en

140 Modelos de remodelacion osea interna
los puntos de integracion de Gauss, se extrapola a los nodos asegurando de esta forma la
continuidad interelemental, que de otro modo no esta garantizada.
Este modelo fue implementado usando el MEF para predecir la distribucion de densidad
osea en la extremidad proximal del femur con buenos resultados y una convergencia mas
que aceptable. Sin embargo, su caracter isotropo limita mucho su uso para el fin requerido
en este trabajo.
Por ultimo, conviene hacer un breve comentario sobre la estabilidad de este algoritmo.
Anteriormente se ha comentado que el metodo numerico usado para integrar la ecuacion
diferencial que gobierna la evolucion de la densidad es el metodo de Euler explıcito (ecuacion
(5.34)). La utilizacion de este esta justifica por la lentitud del proceso de remodelacion
comentada anteriormente y conduce a buenos resultados siempre que el paso de integracion
no sea excesivamente grande. Levenston [201] ha analizado la estabilidad de este problema
concreto obteniendo un paso de integracion crıtico:
∆tc =ρ∗
θΨ∗Sv(ρ∗)ρ(5.38)
donde ρ∗ es la densidad aparente de equilibrio y θ es la pendiente de la curva de remodelacion
(el analisis de estabilidad se desarrolla para una ley de remodelacion lineal).
5.3. Modelos de remodelacion osea anisotropa
Por su importancia en esta tesis, en la que precisamente se propone un nuevo modelo de
remodelacion osea anisotropa, la revision que se viene desarrollando se interrumpe aquı, para
dedicar un apartado completo a los modelos anisotropos. Ya se hablo de uno de ellos cuando
se menciono la teorıa de la elasticidad adaptativa de Cowin. Sin embargo, ese intento de
extender la teorıa de la elasticidad adaptativa al caso anisotropo no es viable desde el punto
de vista practico, como ya se comento, por el gran numero de parametros que involucra.
Como es bien sabido, el proceso de remodelacion osea produce no solo cambios en la den-
sidad aparente del tejido sino tambien reorientacion de la estructura osea: de las trabeculas
en el hueso esponjoso o de las fibras de tejido en el hueso compacto. Una vez que se ha
podido estimar la distribucion de densidad con los modelos isotropos, se pretende conocer
ahora la evolucion de las propiedades elasticas del hueso, supuesto un material anisotropo.
Para caracterizar el hueso como material isotropo se ha relacionado el modulo de Young con
la densidad aparente y se ha supuesto el coeficiente de Poisson constante. Solo es necesario

5.3 Modelos de remodelacion osea anisotropa 141
por tanto una variable independiente, el modulo de Young (como en el modelo de Huiskes) o
la densidad aparente (como en el modelo isotropo de Stanford). Ahora es necesario anadir a
las variables independientes las constantes elasticas, directamente, o definir alguna variable
relacionada con la orientacion de los tejidos (como el fabric tensor) y relacionar el tensor de
comportamiento con esta variable y la densidad aparente.
La mayorıa de los modelos de remodelacion osea estan basados en dos ideas: la opti-
mizacion global de la estructura y/o la regulacion local a nivel celular o tisular.
5.3.1. Modelo de remodelacion osea anisotropa de Stanford basado
en la energıa
Fundamento teorico
La idea principal de este modelo consiste en suponer que la velocidad de disipacion de
energıa mecanica es una medida de la eficiencia de la respuesta osea. Dicha velocidad se define
como la diferencia entre la potencia desarrollada por las fuerzas externas y la velocidad de
variacion de la energıa interna total (suma de energıa interna y cinetica):
Ξ =
(∫
Γ
t · v dΓ +
∫
Ω
b · v dV)− d
dt
(∫
Ω
(χ+K)dV
)(5.39)
donde t es el vector de fuerzas en el contorno Γ, b el de fuerzas de volumen en el dominio Ω,
v es el vector velocidad en cada punto, debida tanto a la deformacion como a la respuesta
adaptativa, χ es la densidad de energıa interna y K = 12ρ|v|2 es la densidad de energıa
cinetica.
La disipacion de energıa total coincide con la disipacion de energıa mecanica en materiales
inertes. Sin embargo, los materiales vivos no son sistemas termodinamicamente cerrados,
pues reciben un aporte metabolico externo que les ayuda a desarrollar su actividad. Ası, la
velocidad de disipacion total, D, se descompone de la siguiente forma:
D = B + Ξ (5.40)
donde B es la energıa disipada en los procesos metabolicos por unidad de tiempo. La disi-
pacion de energıa total es estrictamente positiva tanto en materiales inertes como en ma-
teriales vivos. Sin embargo, el termino de disipacion metabolica los diferencia de manera
fundamental. Dicho termino hace que en materiales vivos la disipacion de energıa mecanica

142 Modelos de remodelacion osea interna
no tenga por que ser estrictamente positiva como lo es en materiales inertes. En efecto,
para el tejido oseo es positiva en reabsorcion y negativa en formacion. En reabsorcion, el
hueso disminuye su rigidez, comportandose de este modo como los materiales inertes. Sin
embargo, en formacion aumenta la rigidez, algo que no ocurre en un material inerte, que no
tiene posibilidad de reparar el dano. Para que se produzca formacion osea y la consecuente
reparacion de dano es estrictamente necesario que haya un aporte energetico externo de tipo
metabolico.
El primer parentesis de la ecuacion (5.39) es la potencia desarrollada por las fuerzas
externas, Pext, que se puede expresar como la suma de la potencia generada por el campo
de tensiones mas la velocidad de cambio de la energıa cinetica:
Pext =
∫
Ω
σ : ε dV +d
dt
∫
Ω
1
2ρ|v|2dV (5.41)
Por tanto, la velocidad de disipacion mecanica se expresa como
Ξ =
∫
Ω
(σ : ε− χ)dV (5.42)
Si el comportamiento fuera puramente elastico, la energıa interna dependerıa exclusiva-
mente del campo de deformaciones y la matriz de comportamiento, y por tanto, el integrando
de (5.42) serıa identicamente cero. Sin embargo, no lo es cuando la respuesta del material
es adaptativa. En el caso concreto del hueso se supone que la energıa interna es tambien
funcion de la densidad aparente:
χ(ε,C, ρ) =1
2ε : C : ε+ Θ(ρ) (5.43)
donde Θ(ρ) es la parte de la energıa interna asociada a los cambios de densidad y C es
el tensor de rigidez del material. Derivando (5.43) y sustituyendo en (5.42) se obtiene la
expresion definitiva de la velocidad de disipacion mecanica o eficiencia de la remodelacion:
Ξ =
∫
Ω
(− 1
2ε : C : ε− dΘ
dρρ)dV (5.44)
Hay que hacer notar que el termino −ε : C : ε es positivo en caso de reabsorcion, para el
que el tensor C es definido negativo y al contrario en el caso de formacion.
Las condiciones que han de cumplirse para que exista remodelacion son condiciones de
caracter local, que en general dependen de la densidad aparente, del tensor de compor-
tamiento y del campo de deformaciones:
fj(ε,C, ρ) ≥ 0 se produce remodelacion
fj(ε,C, ρ) < 0 no se produce remodelacion(5.45)

5.3 Modelos de remodelacion osea anisotropa 143
donde el subındice j abarca todos los criterios de remodelacion empleados. Este serıa el caso
de remodelacion independiente de la velocidad. En el caso de remodelacion dependiente
de la velocidad se anadirıa la condicion fj > 0 para que pudiera producirse remodelacion.
Las condiciones (5.45) son parecidas a las utilizadas en Mecanica del Dano, si bien allı la
condicion fj > 0 es imposible, aumentando el dano si se cumple fj = 0 y permaneciendo
invariable si fj < 0.
Jacobs [166] establecio que la remodelacion osea se produce de manera tal que la efi-
ciencia Ξ es maxima, condicion equivalente al principio de maxima disipacion mecanica.
Sin embargo, de esta forma se llega a expresiones contradictorias y sin sentido fısico alguno
como se vera mas adelante. Garcıa sugirio en su tesis [115] que los materiales adaptativos
cumplen el principio de mınima disipacion, tanto en reabsorcion como en formacion; justo al
contrario que lo supuesto en la Mecanica del Dano aplicada a materiales inertes (principio
de maxima disipacion [307]). En formacion, donde la disipacion mecanica es negativa, se
maximiza su valor absoluto, conservando el paralelismo con la teorıa general de la Mecanica
del Dano. La diferencia de comportamiento en reabsorcion la explica diciendo que mientras
en materiales inertes la disipacion mecanica se produce por la aparicion de microfisuras en
la zona mas solicitada, en materiales adaptativos, en cambio, la reabsorcion se produce en
la zona menos solicitada.
Formulacion del modelo
A continuacion se sigue el desarrollo hecho por Jacobs en su tesis [166]. Este desarrollo se
hace para un criterio de remodelacion generico. Posteriormente se particularizaran las ecua-
ciones para los criterios de remodelacion del modelo de Stanford y finalmente se comentaran
las incongruencias que presenta el modelo, que ya se han advertido en el parrafo anterior.
La idea basica del modelo es que la remodelacion osea maximiza la velocidad de disi-
pacion mecanica cumpliendo los criterios de remodelacion. Este problema fısico se puede
resolver matematicamente como un problema de optimizacion, de Ξ, con restricciones, las
que imponen los criterios de remodelacion, fj ≤ 0. Se hace mediante los multiplicadores de
Lagrange, con un funcional aumentado de la siguiente forma:
L = −Ξ +n∑
j=1
∫
Ω
γjfj(ε,C, ρ)dV (5.46)

144 Modelos de remodelacion osea interna
L =
∫
Ω
[1
2ε : C : ε+ Θ′ρ+
n∑
j=1
γjfj(ε,C, ρ)
]dV (5.47)
donde γj son los multiplicadores de Lagrange para cada criterio de remodelacion. La condi-
cion estacionaria requiere:δL(ε,C, ρ, δε, δρ) = 0 (5.48)
lo que implica:∫
Ω
[( n∑
j=1
(γj∂fj∂ε
)+ C : ε
): δε+
( n∑
j=1
(γj∂fj∂ρ
)+ Θ′′ρ
)δρ
]dV = 0 (5.49)
Como las variaciones de deformacion y de densidad pueden tomar cualquier valor en
cualquier punto del dominio, para que se cumpla la ecuacion anterior, deben anularse cada
uno de los parentesis por separado. En ese caso:
C : ε = −n∑
j=1
γj∂fj∂ε
y Θ′′ρ = −n∑
j=1
γj∂fj∂ρ
, (5.50)
que junto a las condiciones de Kuhn-Tucker cierran el problema de optimizacion:
γj ≥ 0 , fj ≤ 0 , γjfj = 0 (5.51)
El siguiente paso consiste en formular el criterio de remodelacion separando la depen-
dencia de la deformacion y de la densidad:
fj(ε,C, ρ) = φj(ε,C)−Rj(ρ) (5.52)
y suponiendo que la funcion φj es homogenea de grado uno en la deformacion, es decir:
φj(αε,C) = αφj(ε,C) ⇔ φj(ε,C) =∂φj(ε,C)
∂ε: ε (5.53)
Esta condicion es satisfecha tanto por el modelo isotropo de Stanford [166] como por el
de Huiskes [158]. Con estas hipotesis y operando en la primera ecuacion de (5.50), Jacobs
llega a la siguiente expresion de C (ver Anexo A):
C =n∑
j=1
( γjφj
∂φj∂ε⊗ ∂φj
∂ε
)(5.54)
Esta expresion conduce a tensores de comportamiento simetricos y definido positivos. Se
puede observar que el tensor velocidad de cambio de rigidez es un tensor de cuarto orden
que depende exclusivamente del criterio de remodelacion. El escalar γj se calcula forzan-
do el cumplimiento de las condiciones de Kuhn-Tucker (5.51) en el caso de remodelacion
independiente de la velocidad. En el caso de remodelacion dependiente de la velocidad se

5.3 Modelos de remodelacion osea anisotropa 145
sustituyen dichas condiciones por una ecuacion constitutiva para γj en funcion de ρ, proceso
conocido como “regularizacion viscosa” [306]. Esta ecuacion constitutiva se obtiene, en la
practica, imponiendo que la formulacion anisotropa reproduzca el modelo isotropo cuando
un material inicialmente isotropo se encuentre sometido a un estado de tension hidrostatico.
Particularizacion para el criterio de remodelacion de Stanford
La ecuacion (5.54) se particulariza usando el criterio de remodelacion propuesto por
Beaupre [26] (ver ecuacion (5.31)). En este las funciones fa y fr, que definen respectivamente
los criterios de formacion y reabsorcion, se pueden escribir:
fr(ε,C, ρ) = (−e− w)ρ2−β/2
fa(ε,C, ρ) = (e− w)ρ2−β/2(5.55)
donde β es el exponente de la ecuacion (5.36) y e es el error de remodelacion, que se puede
reescribir recordando la ecuaciones (5.28), (5.29) y (5.30)(ver Anexo A):
e = K
√ε : C : ε
ρ2−β/2−Ψ∗
t donde K = ρ2n1/m√B (5.56)
en la que B es el coeficiente de la ecuacion (5.36) y todas las demas constantes ya se
introdujeron cuando se hablo del modelo isotropo de Stanford. Recordando la ecuacion
(5.52), en la anterior se pueden identificar las funciones φa, φr, Ra y Rr:
φr(ε,C) = −K√ε : C : ε
φa(ε,C) = K√ε : C : ε
Rr(ρ) = (w −Ψ∗
t )ρ2−β/2
Ra(ρ) = (w + Ψ∗
t )ρ2−β/2
(5.57)
Diferenciando estas funciones y recordando la expresion (5.54) se obtiene la evolucion
del tensor C:
C =
−γrσ ⊗ σσ : ε
si fr ≥ 0 y fa < 0
0 si fa < 0 y fr < 0
γaσ ⊗ σσ : ε
si fa ≥ 0 y fr < 0
(5.58)
Jacobs calcula γr y γa imponiendo que la ley anisotropa reproduzca el modelo isotropo de
Stanford cuando se aplica a un material isotropo sometido a un estado tensional hidrostatico.
De esta forma llega a la siguiente expresion (ver Anexo A):
C =βρ
ρ
σ ⊗ σσ : ε
(5.59)

146 Modelos de remodelacion osea interna
Para la velocidad de variacion de la densidad se toma la usada en el modelo isotropo de
Stanford:
ρ =
−k crfr(ε,C, ρ)
ρ2−β/2Sv(ρ) ρ si fr ≥ 0 y fa < 0
0 si fa < 0 y fr < 0
k cafa(ε,C, ρ)
ρ2−β/2Sv(ρ) ρ si fa ≥ 0 y fr < 0
(5.60)
Si se implementa este modelo numericamente, por ejemplo mediante el MEF, es necesario
hacerlo de forma incremental integrando la evolucion de la densidad aparente y del tensor
de comportamiento mediante un esquema de integracion explıcito de Euler, por ejemplo:
ρ(t+ ∆t) = ρ(t) + ∆t ρ(t)
C(t+ ∆t) = C(t) + ∆tβρ
ρ
σ ⊗ σσ : ε
(5.61)
Pueden aparecer problemas en el caso de reabsorcion cuando ρ < 0, ya que es posible
entonces que C pierda su caracter definido positivo si el incremento de tiempo excesivamente
grande. Esto se evita planteando el algoritmo en funcion del tensor de flexibilidad, D, que
cumple:
D : C = I (5.62)
Derivando (5.62) con respecto al tiempo se obtiene:
D = −D : C : D (5.63)
y sustituyendo (5.59) en la anterior, queda finalmente:
D = −βρρ
ε⊗ εσ : ε
(5.64)
Aparte de pequenos errores que se han advertido en la formulacion y que se comentan en
el Anexo A, hay que indicar que el modelo no es del todo consistente con la optimizacion en la
que se basa. La expresion que permite calcular ρ, (5.60), es identica a la del modelo isotropo
de Stanford. Para su obtencion no se ha tenido en cuenta el criterio de optimalidad, (5.50),
que ademas no se puede cumplir con tal definicion de ρ. Si se identificaran las ecuaciones
(5.50) y (5.60), Θ no podrıa ser funcion exclusiva de ρ, como se ha supuesto, ya que los
criterios de remodelacion, fi(ε,C, ρ), dependen de la deformacion, la densidad y el material.
Este modelo fue implementado usando el MEF para predecir la distribucion de densidad
osea y la anisotropıa en la extremidad proximal del femur [166]. Los resultados de densidad

5.3 Modelos de remodelacion osea anisotropa 147
son tan buenos como lo pueden ser los obtenidos con el modelo isotropo de Stanford, ya que
la ley de flujo de la densidad es la misma que la allı utilizada. En cuanto a las propiedades
elasticas y el grado de anisotropıa no se puede decir lo mismo, ya que los resultados no
concuerdan con la realidad. Esto era esperable, por otra parte, a la vista de las incongruencias
que el modelo presenta. En concreto, las capas de hueso cortical de la diafisis tienen una
rigidez menor de lo esperado y un mayor grado de anisotropıa del medido experimentalmente.
5.3.2. Modelo de remodelacion osea anisotropa de Stanford basado
en las tensiones
En respuesta a la falta de concordancia entre los resultados que proporciona el modelo
anterior y los experimentales, Jacobs [166] propuso un modelo alternativo basado en la
suposicion de que el hueso responde adaptandose a las direcciones principales de tension,
mas que para conseguir una estructura globalmente optima.
Como ya se ha dicho, Wolff [354], sugirio a finales del siglo XIX que las trabeculas se
alinean con las direcciones principales de tension y como estas son perpendiculares entre sı las
trabeculas tambien han de serlo. Esta controvertida idea, refutada por numerosos autores
[139,266,335] e incluso por estudios estereologicos [19], es erronea desde el momento en que
considera que solo existe una situacion de carga posible y por tanto un unico sistema de
direcciones principales. Matizando esta idea, Carter et al. [59] propusieron que las trabeculas
se orientan en la direccion que hace maxima la tension equivalente normal (ver ec. (5.17)),
parametro que tiene en cuenta todos los casos de carga que soporta el hueso. Basado en
esta idea, Jacobs desarrollo en su tesis [166] un modelo de comportamiento que se describe
a continuacion.
El desarrollo del modelo parte de la division del tensor C en dos, uno isotropo y otro
anisotropo.
C = Ciso + Caniso (5.65)
El primero de ellos se obtiene de forma sencilla (ver Anexo A, ecuacion (A.16b))
Ciso =( ν
1− 2ν1⊗ 1 + I
) E
1 + ν
β ρ(σiso, ρ)
ρ(5.66)
En este modelo, la variacion de la densidad aparente, ρ, se calcula de la misma forma que
se hace en el modelo isotropo de Stanford pero usando una tension efectiva isotropa σiso,
definida segun la ecuacion (5.15) que, para un estado tensional pseudo esferico, se obtiene

148 Modelos de remodelacion osea interna
de las expresiones:
p =|σI |+ |σII |+ |σIII |
3(5.67)
σiso = p 1 (5.68)
σiso(p) =√Bρβ(σiso : D : σiso) (5.69)
donde σI , σII y σIII son las tensiones principales. La perturbacion anisotropa, Caniso, tiene
en cuenta cada direccion principal por separado: si una tension principal es mayor que la
pseudo presion hidrostatica, p, la correspondiente componente del tensor de rigidez aumenta
y viceversa. La velocidad con que aumenta o disminuye es para todas las direcciones la misma
y no depende de la diferencia entre la tension principal y la pseudo presion hidrostatica, la
gobierna un unico escalar, γ. En ejes principales y en notacion compacta, queda:
Caniso = γ
sign(|σI | − p) 0 0
0 sign(|σII | − p) 0 0
0 0 sign(|σIII | − p)0 0
(5.70)
que se expresa de la siguiente forma en el sistema de coordenadas global:
Caniso = γ
III∑
i=I
sign(|σi| − p)(vi ⊗ vi ⊗ vi ⊗ vi) (5.71)
Los cambios adaptativos que predice esta aproximacion ocurren en direcciones perpen-
diculares entre sı de tal modo que un material inicialmente isotropo evolucionara hacia un
comportamiento ortotropo con ejes de ortotropıa coincidentes con las direcciones principales
de tension. Sin embargo, esta simetrıa desaparecera en cuanto se aplique una carga distinta
a la primera y el hueso acabara, por lo general, teniendo un comportamiento anisotropo
como algunos autores [19,59,335] sugieren que ocurre en realidad.
Conviene no confundir el parametro p con la tension hidrostatica, que se obtendrıa sin
tomar el valor absoluto de las tensiones principales en la expresion (5.67). Jacobs usa p,
segun el, porque el “hueso responde de la misma forma a tensiones de traccion o de compre-
sion, haciendo que el signo de las tensiones principales sea irrelevante”. Ası, el parametro p
representa una media de las tensiones principales sin tener en cuenta el signo de estas. Si una
determinada tension principal es mayor en valor absoluto que esta media, el hueso responde
aumentando la rigidez en esa direccion y disminuyendola si es menor que la media en valor
absoluto. Tomar el valor absoluto de las tensiones principales para compararlas con p en la

5.3 Modelos de remodelacion osea anisotropa 149
expresion (5.70)1 es consecuente con la idea de que el signo de las tensiones es irrelevante
en la respuesta adaptativa. De otra forma, una tension de compresion siempre producirıa
reabsorcion.
Calculo del parametro γ
Ahora no cabe hacer como en el modelo basado en la energıa para determinar γ: iden-
tificar el modelo anisotropo con el isotropo de Stanford cuando se aplica a un material
inicialmente isotropo sometido a un estado hidrostatico de tension. En ese caso (|σi| − p) y
por tanto Caniso es nulo. En su lugar, el problema empleado para la calibracion del modelo es
un estado de tension uniaxial que, sin perdida de generalidad, se puede considerar actuando
segun la direccion x. Es evidente que no es posible identificar por completo ambos modelos
con un solo parametro de ajuste. Ası, la calibracion se limita a identificar la evolucion de
uno solo de los elementos del tensor de comportamiento, C1111.
C1111 = Ciso1111 + Caniso1111 (5.72)
En el modelo isotropo de Stanford (que supone ν constante) la evolucion de este termino
es:
C1111 = 2µ+ λ = E1− ν
(1 + ν)(1− 2ν)=βE
ρρ(σ, ρ)
1− ν(1 + ν)(1− 2ν)
(5.73)
con µ y λ las constantes de Lame y ρ(σ, ρ) definida en la ecuacion (5.33).
En el caso de tension uniaxial el tensor de tensiones es:
σ =
σ 0 0
0 0 0
0 0 0
(5.74)
y la tension efectiva σ = σ, con lo cual ya se puede completar la ecuacion (5.73). Por otro
lado, en el caso de tension uniaxial p =σ
3y es facil comprobar que:
σiso =σ
3
√Bρβ
κ(5.75)
Ciso1111 =βE
ρρ(σiso, ρ)
1− ν(1 + ν)(1− 2ν)
(5.76)
1En su tesis [166] Jacobs no usa el valor absoluto en la ecuacion (5.70). Probablemente sea una errata ya
que en la programacion del modelo sı lo hace

150 Modelos de remodelacion osea interna
Caniso = γ
1 0 0
0 −1 0 0
0 0 −1
0 0
(5.77)
Sumando (5.76) y el termino correspondiente de (5.77) y sustituyendo en la ecuacion
(5.72) se puede despejar γ. Jacobs llega a la siguiente expresion:
γ = βBρβ−1
[ρ(σ, ρ)− ρ(σ
3
√Bρβ
κ, ρ)
](5.78)
con la que queda completa la formulacion.
Jacobs aplico tanto este modelo como el descrito en el apartado anterior, basado en la
energıa, a la simulacion de la morfogenesis de la extremidad proximal del femur. El modelo
basado en las tensiones proporciona mejores resultados a pesar de que no es mecanicamente
consistente, como se vera a continuacion.
Discusion sobre la formulacion
En primer lugar hay que destacar que el proceso de calibracion que permite calcular γ
es totalmente arbitrario y no obedece a ningun tipo de mecanismo biologico. No se puede
decir que un estado de tension uniaxial sea muy frecuente en huesos. Aunque lo fuera, la
expresion a la que llega con dicho proceso de calibracion, (5.78) no es correcta, al menos
si se siguen los pasos que el propio Jacobs describe. En efecto, sumando (5.76) y (5.77) y
sustituyendo en (5.72) se obtiene un γ distinto:
γ =1− ν
(1 + ν)(1− 2ν)βBρβ−1
[ρ(σ, ρ)− ρ(σ
3
√Bρβ
κ, ρ)
](5.79)
La diferencia esta en el primer factor de la ecuacion anterior. Este factor esta en torno a
1.4 para los valores de ν que se usan normalmente en el hueso. En caso de usar la ecuacion
(5.79) en lugar de la (5.80) cabrıa esperar diferencias apreciables con respecto a los resultados
dados por Jacobs, tanto en la velocidad de convergencia como en el nivel de anisotropıa.
Por otro lado, la base de la formulacion no es consistente desde el punto de vista mecanico,
ya que la perturbacion anisotropa propuesta modifica solo parte de las componentes del
tensor de comportamiento. Dicha modificacion no se corresponde con ningun tipo de simetrıa
de material conocida y tampoco con un caso de anisotropıa propiamente dicha. Como explica

5.3 Modelos de remodelacion osea anisotropa 151
Garcıa [115], ante un estado de tension biaxial:
σ =
σ 0 0
0 σ 0
0 0 0
(5.80)
lo logico es que el material evolucione hacia un comportamiento transversalmente isotropo.
El tensor de rigidez de un material de este tipo es como sigue:
Caniso = γ
C11 C12 C13 0 0 0
C12 C11 C13 0 0 0
C13 C13 C33 0 0 0
0 0 0 C44 0 0
0 0 0 0 C44 0
0 0 0 0 0 C11 − C12
(5.81)
Sin embargo, conseguir esto es imposible con el modelo propuesto que, para el caso de tension
biaxial de la ecuacion (5.80), darıa una perturbacion anisotropa igual a:
Caniso = γ
1 0 0
0 1 0 0
0 0 −1
0 0
(5.82)
Se observa que con esta evolucion de la rigidez es imposible diferenciar C66 de C44 y C55.
La gran limitacion de este modelo es la de asignar a todas las direcciones la misma
velocidad de evolucion de la rigidez, con independencia de la magnitud de las tensiones en
cada direccion. Usar un solo parametro de calibracion trae como consecuencia que dicho
modelo solo funcione apropiadamente en determinadas circunstancias.
5.3.3. Modelo de remodelacion osea anisotropa basado en la Mecanica
del dano
Fundamentos de la Mecanica del Dano
La iniciacion y crecimiento de microgrietas en materiales fragiles, de microcavidades en
materiales ductiles y de huecos en materiales porosos ha sido estudiado por medio de analisis
microestructural [225, 284] y por medio de la Mecanica del Dano, [178, 199, 307, 308], que

152 Modelos de remodelacion osea interna
aborda el problema desde un punto de vista macroscopico, tıpico de la Mecanica de Medios
Continuos. Se define para ello un parametro de dano, d, que cuantifica en cierta forma la
densidad efectiva de grietas y que varıa entre d = 0 (estado no danado) y d = 1 (estado de
ruptura local).
Basado en el principio de deformacion equivalente [199], se introduce la tension efectiva,
(σ), como aquella a la que hay que someter al modelo no danado para tener la misma
deformacion que en el modelo con dano (figura 5.12)
1 1
1+ ε 1+ ε
σ σ
1
Figura 5.12: Representacion esquematica del concepto de tension efectiva
Esta definicion se puede escribir de forma matematica como sigue:
σ =σ
1− d (5.83)
y aplicando el principio de deformacion equivalente se tiene de forma inmediata el tensor de
comportamiento del material danado, C, en funcion del correspondiente al material virgen,
C0,
C = (1− d)C0 (5.84)
Para completar la formulacion es necesario definir la variable que hace que el dano se
modifique (estımulo), el umbral que hay que superar para se produzca dicha modificacion
(criterio de dano), la evolucion del parametro de dano (ley de flujo) y del resto de parametros
que aparecen en el criterio de dano (ley de endurecimiento). Estas leyes deben ser conse-
cuentes con las leyes de la Termodinamica. Ası, el Segundo Principio establece que en todo

5.3 Modelos de remodelacion osea anisotropa 153
proceso termodinamico se debe cumplir:
D = −ψ + σ : ε ≥ 0 (5.85)
donde D es la velocidad de disipacion y ψ es la variacion de la energıa libre que, para un
material con comportamiento elastico, viene dada por:
ψ =1
2ε : C : ε (5.86)
con lo cual
D = −1
2ε : C : ε (5.87)
Siguiendo la nomenclatura de Simo y Ju [307] se denota por τ 0 el estımulo o causa
intrınseca del dano. El criterio de dano define los valores del estımulo que no producen dano.
Son aquellos que cumplen que una cierta funcion, g se mantiene no positiva. El criterio de
dano se suele escribir separando la influencia del dano y del estımulo, a traves de una funcion
auxiliar G, de la siguiente forma:
g(τ0, d) ≡ G(τ0)− d ≤ 0 (5.88)
El estımulo τ0 suele ser funcion de la variable termodinamica asociada al dano, que en
materiales isotropos y elasticos es ψ0, la densidad de energıa libre asociada a la deformacion
real y a la tension efectiva:
ψ0 =1
2ε : C0 : ε (5.89)
Aplicando el principio de maxima disipacion con la restriccion impuesta por (5.88) se
obtiene:
d = µ∂G
∂ψ0(5.90)
donde µ es el parametro de consistencia del dano que se obtiene aplicando las condiciones
de Kuhn-Tucker:
µ ≥ 0 g(τ0, d) ≤ 0 µ g(τ0, d) = 0 (5.91)
La extension al caso anisotropo que hicieron Cordebois y Sideroff [69] considera el dano
como un tensor, d y define la tension efectiva de manera analoga a (5.83) pero de forma que
conserva la simetrıa:
σ = (I− d)−1/2 σ (I− d)−1/2 (5.92)

154 Modelos de remodelacion osea interna
La ecuacion constitutiva del dano que dieron estos autores se basa en una identificacion
energetica: la tension efectiva es tal que aplicada a un material sin dano produce la misma
energıa de deformacion que tiene el material danado (d) sujeto a la tension real, σ.
U(σ,d) = U(σ, 0) (5.93)
Formulacion teorica
Utilizando ideas de la teorıa de dano anisotropo de Cordebois y Sideroff [69] y el “fabric
tensor” como medida de la anisotropıa osea, Doblare y Garcıa [91] plantean un nuevo modelo
de dano-reparacion aplicable a la remodelacion osea interna, que podrıa coynsiderarse, en
parte, una extension del modelo isotropo de Stanford al caso anisotropo.
El dano se interpreta como una medida del volumen de huecos en el interior del tejido
oseo y de la direccionalidad de los mismos, a traves del “fabric tensor”. De acuerdo con
esto, se define el material virgen como aquel con porosidad nula y en este caso se considera
isotropo. Eso conlleva que este modelo considere asimismo isotropo el tejido oseo cortical
ideal sin porosidad.
Existen dos diferencias fundamentales con la teorıa de dano clasica. La primera es que
en esta siempre se produce aumento del dano, mientras que aquı, en el proceso de formacion
osea, la porosidad puede disminuir. Lo hace como consecuencia del aporte energetico puesto
en juego por los procesos metabolicos que tienen lugar en un organismo vivo y que no
estan presentes en los materiales inertes que usualmente trata la teorıa clasica de dano. La
segunda diferencia es que en este modelo el aumento del dano-porosidad, reabsorcion osea, se
produce en las regiones con un nivel bajo de tensiones mientras que el aumento del dano en
la teorıa clasica siempre se produce por un nivel elevado de tensiones. Estas peculiaridades
llevan a que el tejido oseo cumpla, con los postulados utilizados en este modelo, el principio
de mınima disipacion mecanica, es decir, la respuesta del hueso al estımulo mecanico es
tal que minimiza la energıa mecanica disipada, condicion contraria al principio de maxima
disipacion, que cumplen los materiales inertes en la Mecanica de Dano [307].
Para el caso de reabsorcion, se define el tensor de dano, o tensor de porosidad, mediante
la expresion
d = 1−H2 = 1−(ρ
ρ
)β/2√A H (5.94)
donde ρ es la densidad aparente, ρ la densidad maxima del hueso cortical, β el exponente
de la densidad aparente en la ecuacion (5.36), H es el “fabric tensor” normalizado de forma

5.3 Modelos de remodelacion osea anisotropa 155
que det(H) = 1 y A un parametro que se introduce para asegurar que en el caso isotropo la
formulacion reproduzca el modelo isotropo de Stanford. Su valor es:
A =B(ρ)
Bρβ(ρ)−β (5.95)
donde B es el coeficiente de la ecuacion (5.36) y las variables con “gorro” hacen referencia
a los valores correspondientes al hueso cortical de maxima densidad.
Sustituyendo (5.94) en la ecuacion (5.92), que proporciona la tension efectiva y traba-
jando con el tensor H, se llega a:
σ = H−1 σH−1, (5.96)
Esta expresion junto con la ecuacion constitutiva del dano (5.92) da lugar a un tensor de
flexibilidad:
D = H−2 DH−2, (5.97)
cuyas direcciones de ortotropıa coinciden con los ejes principales del tensor H, o del tensor de
dano d, del que es paralelo. Con la anterior ecuacion, se puede calcular el tensor de compor-
tamiento a partir de los autovectores y autovalores del tensor H. Las propiedades elasticas
en ejes de ortotropıa vienen dadas por expresiones como las que siguen, que corresponden a
la direccion principal de dano i:
1
Ei=
1
E
1
h4i
− νijEi
= − νE
1
h2i h
2j
1
2Gij=
1 + ν
E
1
h2i h
2j
(5.98)
donde E y ν son respectivamente el modulo de Young y el coeficiente de Poisson correspon-
diente al hueso con porosidad nula y hi son los autovalores del tensor H.
En reabsorcion se usa el tensor de dano, tal como se ha indicado hasta ahora, mientras
que para formacion se define un tensor analogo al de dano pero asociado a la reparacion,
R = H2. La formulacion es identica para uno y otro caso.
Una vez establecida la ley de comportamiento del hueso, el siguiente paso es definir como
varıa dicho comportamiento con la carga. Esto equivale a preguntarse como se modifica H
con el estado de tensiones. En primer lugar, se define el estımulo de remodelacion, Y, de
forma analoga a como se hace en plasticidad, como la variable termodinamica asociada al
dano o a la reparacion, segun se trate de reabsorcion o formacion. En la practica no es

156 Modelos de remodelacion osea interna
necesario hacer tal distincion y se define como la variable termodinamica asociada al tensor
H que caracteriza tanto al dano como a la reparacion. Sı es necesario definir la variable
mecanica independiente del proceso: deformacion o tension. En formacion, la variable basica
que rige el proceso es la tension, en la lınea de lo indicado por Wolff, que postulo que los
ejes principales de ortotropıa se orientan segun las direcciones principales de tension. Por el
contrario, en reabsorcion se utiliza la deformacion como variable basica independiente. Con
ello, se tiene, para el caso de reabsorcion:
Y =∂ψ(H, ε)
∂H(5.99)
Garcıa mostro [115] como derivar en la ecuacion anterior para llegar a la siguiente expresion:
Y = 2[2G sym
[(HεH) (Hε)
]+ λ tr(H2ε) sym(Hε)
](5.100)
siendo G y λ las constantes de Lame correspondientes al hueso cortical de porosidad nula.
Para cuantificar la importancia del nivel de anisotropıa en el estımulo mecanico se define
un nuevo tensor, J, definido mediante la expresion
J =(1− ω)
3tr(Y)1 + ω dev(Y) =
(1− 2ω)
3tr(Y)1 + ωY (5.101)
El factor de anisotropıa, ω, es el que pesa la importancia del nivel de anisotropıa del estımulo
en el modelo. Es tal que ω ∈ [0, 1], de manera que ω = 0 conduce a un modelo que depende
unicamente de la componente isotropa del estımulo. De esa forma la velocidad de variacion
de las componentes del tensor de comportamiento son independientes de la direccion. El
otro extremo, ω = 1, produce el mayor grado de anisotropıa posible con este modelo.
En funcion de este nuevo estımulo se plantean los criterios de remodelacion, que deter-
minan las condiciones bajo las cuales se activa los mecanismos de formacion y reabsorcion
osea. Se definen las funciones gr y gf de la siguiente manera:
gr =
√2(1− ω)
n1/m√B ρ2−β/8 ρβ/8 A1/8 271/4
(J−1 : J−1
)1/4 − 1
ρ2−β/2(Ψ∗
t − w)
gf =n1/m
√B ρ2−β/8 ρβ/8 A1/8 31/4
√2(1− ω)
(J : J)1/4 − ρ2−β/2(Ψ∗
t + w)
(5.102)
con una forma muy particular y que incluyen un buen numero de constantes. La presencia
de todas esas constantes asegura que al aplicar el algoritmo a un caso isotropo se reproduce

5.3 Modelos de remodelacion osea anisotropa 157
el modelo isotropo de Stanford. Los criterios de remodelacion se definen:
gf (J,Ψ∗
t , w) ≤ 0 gr(J,Ψ∗
t , w) > 0 reabsorcion
gr(J,Ψ∗
t , w) ≤ 0 gf (J,Ψ∗
t , w) > 0 formacion
gr(J,Ψ∗
t , w) ≤ 0 gf (J,Ψ∗
t , w) ≤ 0 zona muerta
(5.103)
La ley de evolucion de la porosidad se define mediante la expresion
H = µr∂gr
∂Yreabsorcion
H = µf∂gf
∂Yformacion
(5.104)
debiendo cumplirse la condicion de consistencia (condiciones de Kuhn-Tucker):
µr, µf ≥ 0 gr, gf ≤ 0 µrgr, µfgf = 0 (5.105)
La forma de calcular los parametros µr y µf consiste en forzar que se cumpla la condicion
establecida anteriormente de que det(H) = 1. Los valores ası obtenidos cumplen la condicion
de consistencia, como demuestra Garcıa [115]. Despues de una serie de calculos se llega a la
siguiente expresion de H:
H =3β k rSv
4 tr (H−1J−3ω)
ρ
ρJ−3 ω reabsorcion
H =3β k rSv
4 tr (H−1Jω)
ρ
ρJ ω formacion
(5.106)
en la que se ha introducido el tensor ω para simplificar la formulacion
ω =(1− 2ω)
31⊗ 1 + ω I (5.107)
En la expresion anterior tambien se ha sustituido la derivada temporal de la densidad
aparente, que surge de la derivacion de H, por la expresion utilizada en el modelo isotropo
de Stanford, (5.33). Esto implica que en un material isotropo si se utiliza un criterio de dano
isotropo, (ω = 0), se reproduce el modelo isotropo de Stanford.
Cabe destacar una propiedad de este modelo: si el tensor de porosidad, o de forma
equivalente el “fabric tensor”, esta alineado con el tensor de tensiones (en ese caso tambien
estara alineado con el tensor de deformaciones), se llega a una situacion de equilibrio direc-
cional. Los autovectores del tensor de porosidad no varıan y por tanto tampoco lo hacen
los ejes de ortotropıa del material, tal como ocurre en realidad, segun indica Cowin [75] y
Odgaard et al. [254] a partir de datos experimentales.

158 Modelos de remodelacion osea interna
Es preciso comentar que el valor de r en la ecuacion (5.106) se obtiene a partir del criterio
que se encuentre activo en ese momento:
r =
−crgr
ρ2−β/2si gr ≥ 0 y gf < 0
0 si gr < 0 y gf < 0
cfgf
ρ2−β/2si gr < 0 y gf ≥ 0
(5.108)
Esta respuesta remodelatoria es equivalente a la representada en la figura 5.9, corre-
spondiente al modelo isotropo de Stanford (ver ecuacion (5.31)) y presenta una limitacion
importante, que se comenta a continuacion. Segun esa ecuacion, un estımulo mecanico por
encima del de equilibrio produce formacion osea y ademas a una velocidad creciente si el
estımulo crece de manera indefinida. Esto implica que cargas muy elevadas producen for-
macion de hueso de manera indefinida, si acaso limitada por el maximo que la densidad
osea puede alcanzar. Como consecuencia del proceso de formacion, la rigidez crecera rapi-
damente. En ningun caso se contempla la posibilidad de que el hueso pueda danarse como
consecuencia de un estado tensional demasiado alto, como sugieren gran cantidad de au-
tores [49,175,198,265,360].
El modelo anisotropo basado en la Mecanica del Dano tampoco distingue el compor-
tamiento del hueso en traccion del de compresion. Esto se puede comprobar en la ecuacion
que define el estımulo, (5.100), cuadratrica en las deformaciones y por tanto independiente
del signo de estas.
Las dos limitaciones comentadas, que no se contempla la acumulacion de dano por sobre-
carga y que no se distingue entre traccion y compresion, estan relacionadas entre sı. Algunos
autores [117, 265] achacan la diferente respuesta del hueso frente a traccion y compresion a
la distinta forma en que se acumula dano a nivel microscopico en uno y otro caso.
Este modelo fue utilizado por Doblare y Garcıa [91,115] para reproducir la distribucion
de densidad y propiedades elasticas del tejido oseo en el femur humano. Partieron de una
situacion inicial arbitraria, densidad aparente uniforme de 0.5 g/cm3 y hueso isotropo y
aplicando las cargas normales al caminar obtuvieron la distribucion de densidad aparente que
se muestra en la figura 5.13. Las propiedades elasticas obtenidas (ver figura 5.13 igualmente)
presentan un grado de anisotropıa mas parecido a la realidad, que los obtenidos con los otros
modelos anisotropos presentados aquı, como demuestra Garcıa [115].
Este modelo ha sido utilizado para simular la morfogenesis de la mandıbula, de la misma
forma planteada por Doblare y Garcıa para el femur. Los resultados de esta simulacion se

5.4 Otros modelos posteriores 159
a b
2 GPa
Figura 5.13: Resultados obtenidos en el femur con el modelo de remodelacion basado en la Mecanica
del Dano: (a) densidad en g/cm3, (b) propiedades elasticas en GPa.
comentaran en el capıtulo siguiente.
5.4. Otros modelos posteriores
A continuacion se presentan dos modelos de remodelacion osea interna desarrollados con
posterioridad al modelo basado en la Mecanica del Dano. El primero de ellos, el Modelo de
Bagge, usa tecnicas de optimizacion en su desarrollo y no es anisotropo, puesto que considera
que el tejido oseo trabecular tiene simetrıa cubica. El segundo, Modelo de Tsubota-Adachi-
Tomita, consigue simular la anisotropıa del hueso por medio de su arquitectura trabecular,
que dicho modelo es capaz de evaluar a nivel microscopico. Para obtener dicha arquitectura
supone que el fin de la remodelacion es conseguir la homogeneizacion de las tensiones, que,
en definitiva, equivale a conseguir el mejor aprovechamiento posible del material.
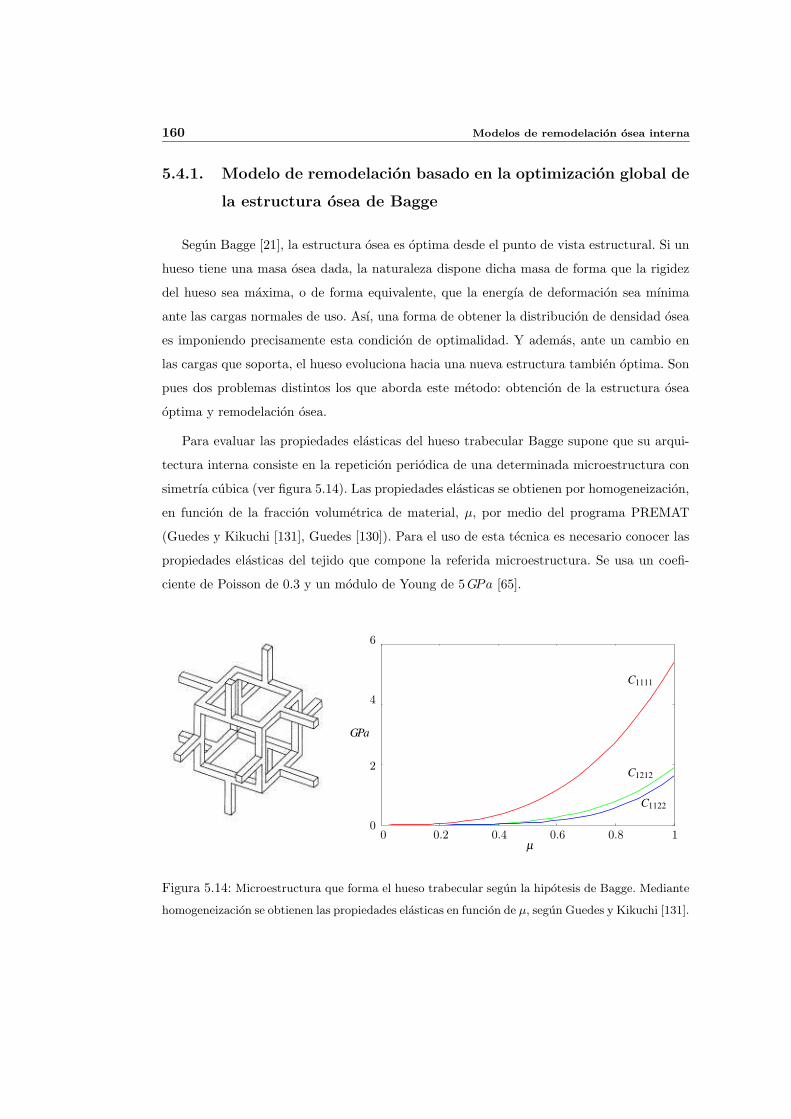
160 Modelos de remodelacion osea interna
5.4.1. Modelo de remodelacion basado en la optimizacion global de
la estructura osea de Bagge
Segun Bagge [21], la estructura osea es optima desde el punto de vista estructural. Si un
hueso tiene una masa osea dada, la naturaleza dispone dicha masa de forma que la rigidez
del hueso sea maxima, o de forma equivalente, que la energıa de deformacion sea mınima
ante las cargas normales de uso. Ası, una forma de obtener la distribucion de densidad osea
es imponiendo precisamente esta condicion de optimalidad. Y ademas, ante un cambio en
las cargas que soporta, el hueso evoluciona hacia una nueva estructura tambien optima. Son
pues dos problemas distintos los que aborda este metodo: obtencion de la estructura osea
optima y remodelacion osea.
Para evaluar las propiedades elasticas del hueso trabecular Bagge supone que su arqui-
tectura interna consiste en la repeticion periodica de una determinada microestructura con
simetrıa cubica (ver figura 5.14). Las propiedades elasticas se obtienen por homogeneizacion,
en funcion de la fraccion volumetrica de material, µ, por medio del programa PREMAT
(Guedes y Kikuchi [131], Guedes [130]). Para el uso de esta tecnica es necesario conocer las
propiedades elasticas del tejido que compone la referida microestructura. Se usa un coefi-
ciente de Poisson de 0.3 y un modulo de Young de 5GPa [65].
00
2
4
6
0.2 0.4 0.6 0.8 1
µ
GPa
C1111
C1122
C1212
1
Figura 5.14: Microestructura que forma el hueso trabecular segun la hipotesis de Bagge. Mediante
homogeneizacion se obtienen las propiedades elasticas en funcion de µ, segun Guedes y Kikuchi [131].

5.4 Otros modelos posteriores 161
Estructura osea optima
Si un hueso, de volumen V , esta sometido diariamente a un conjunto de Nl estados de
carga, la disposicion del tejido oseo sera aquella que consigue la maxima rigidez o la menor
energıa de deformacion asociada a esos estados de carga, con el volumen dado, V . Para
cumplir esta hipotesis, la estructura osea debe ser la solucion del siguiente problema de
optimizacion:
min
Nl∑
j=1
wj Uj
sujeto a
Ne∑
i=1
µiVsi = V ,
0 < µmin ≤ µi ≤ 1, i = 1, ..., Ne
(5.109)
que Bagge resuelve con ayuda del MEF. En la expresion anterior Uj es la energıa de defor-
macion para el caso de carga j, wj es un factor de peso asociado a dicha carga, Ne es el
numero de elementos de la malla de elementos finitos, V si es el volumen del elemento i y µi
es su fraccion volumetrica de hueso, que debe estar siempre por encima de un valor mınimo
µmin.
La solucion del problema anterior se obtiene aplicando el metodo de los multiplicadores
de Lagrange (ver Bendsøe [30] para referencias y Bagge [20] para el desarrollo). Las condi-
ciones de optimalidad se usan en sucesivas iteraciones, para la actualizacion de la fraccion
volumetrica de hueso de cada elemento. Ası, la fraccion volumetrica del elemento i en la
iteracion k + 1 sera:
µk+1i = Bηi µ
ki , i = 1, ..., Ne (5.110)
donde η ≤ 1 es un factor de amortiguamiento que se usa para limitar el cambio de volumen
del elemento en cada iteracion. Los parametros Bi vienen dados por:
Bi =1
λ V si
Nl∑
j=1
wj∂Uj∂µi
, i = 1, ..., Ne (5.111)
donde λ es el multiplicador de Lagrange de la restriccion de volumen y la sensibilidad de la
energıa de deformacion al cambio de volumen se evalua de la siguiente forma:
∂Uj∂µi
=
(∫
Vi
1
2εT :
∂[C(µ)]Hi∂µi
: ε dVi
)
j
, i = 1, ..., Ne, j = 1, ..., Nl (5.112)
La derivada parcial del integrando se obtiene a partir de la grafica de la figura 5.14.

162 Modelos de remodelacion osea interna
Como el material posee simetrıa cubica, es necesario establecer la orientacion del material
que proporciona la mınima energıa de deformacion. Existe solucion analıtica a este problema
para el caso bidimensional de un unico caso de carga. Esta solucion consiste en alinear los
ejes del material con las direcciones principales de tension. Para el problema general de un
hueso 3D sometido a una combinacion de cargas no existe solucion analıtica. Bagge propone
alinear los ejes del material con los autovalores de un tensor de tensiones promedio que tenga
en cuenta todos los casos de carga pesados con los factores wj , como en la ecuacion (5.109),
lo que esta de acuerdo con la trajectorial theory de la que se hablo al principio del capıtulo.
Bagge [21] aplico esta tecnica a la extremidad proximal del femur y obtuvo los resultados
mostrados en la figura 5.15. Hay que indicar que V es una propiedad de cada hueso y de
cada individuo, desconocida a priori y que esta relacionada con la fraccion volumetrica global
del hueso en cuestion. Bagge probo con valores de volumen total de tejido de 25 %, 50 %
y 75 % respecto del volumen total del femur. Los resultados muestran que la distribucion
correspondiente al 50 % se asemeja a la realidad bastante mejor que las otras dos. Esta es, por
tanto, la que toma como referencia y la que le permite estimar para el femur una fraccion
volumetrica del 50 %. Los problemas de “checkerboard” que presenta la implementacion
del problema con el MEF fueron resueltos mediante un metodo de filtrado sugerido por
Sigmund [305].
Remodelacion osea
Para simular una situacion de remodelacion es necesario partir de una distribucion de
hueso realista. Bagge [21] tomo como situacion de partida la estructura osea optima cor-
respondiente a un volumen de tejido 50 % del volumen total del hueso y aplico la misma
combinacion de casos de carga con la que obtuvo dicha estructura pero variando la magnitud
de todas las cargas en proporcion.
Ante un cambio de actividad el hueso modifica su masa evolucionando hacia una nueva
estructura, que es optima para la nuevas cargas. Por ello, la ecuacion (5.110) sirve tambien
como esquema de actualizacion de la fraccion volumetrica de hueso en remodelacion.
µt+∆ti = (Bti )
ηti µti =
(1
λ V si
∂U t
∂µti
)ηti
µti, i = 1, ..., Ne (5.113)
donde se han cambiado los superındices que hacıan referencia al numero de iteracion por
otros que se refieren al tiempo transcurrido desde que comenzo la remodelacion. El multipli-
cador de Lagrange correspondiente al optimo de la estructura de partida (50 % de fraccion
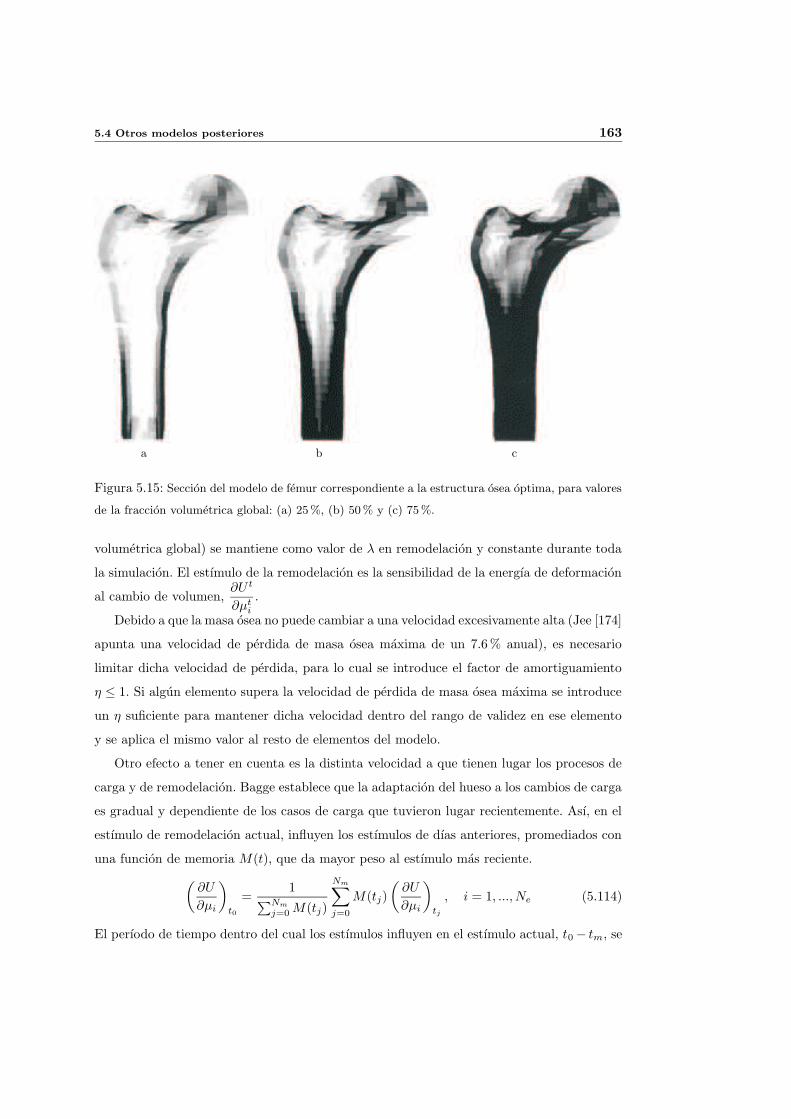
5.4 Otros modelos posteriores 163
a b c
Figura 5.15: Seccion del modelo de femur correspondiente a la estructura osea optima, para valores
de la fraccion volumetrica global: (a) 25%, (b) 50% y (c) 75%.
volumetrica global) se mantiene como valor de λ en remodelacion y constante durante toda
la simulacion. El estımulo de la remodelacion es la sensibilidad de la energıa de deformacion
al cambio de volumen,∂U t
∂µti.
Debido a que la masa osea no puede cambiar a una velocidad excesivamente alta (Jee [174]
apunta una velocidad de perdida de masa osea maxima de un 7.6 % anual), es necesario
limitar dicha velocidad de perdida, para lo cual se introduce el factor de amortiguamiento
η ≤ 1. Si algun elemento supera la velocidad de perdida de masa osea maxima se introduce
un η suficiente para mantener dicha velocidad dentro del rango de validez en ese elemento
y se aplica el mismo valor al resto de elementos del modelo.
Otro efecto a tener en cuenta es la distinta velocidad a que tienen lugar los procesos de
carga y de remodelacion. Bagge establece que la adaptacion del hueso a los cambios de carga
es gradual y dependiente de los casos de carga que tuvieron lugar recientemente. Ası, en el
estımulo de remodelacion actual, influyen los estımulos de dıas anteriores, promediados con
una funcion de memoria M(t), que da mayor peso al estımulo mas reciente.
(∂U
∂µi
)
t0
=1
∑Nm
j=0M(tj)
Nm∑
j=0
M(tj)
(∂U
∂µi
)
tj
, i = 1, ..., Ne (5.114)
El perıodo de tiempo dentro del cual los estımulos influyen en el estımulo actual, t0− tm, se

164 Modelos de remodelacion osea interna
divide en Nm pasos de tiempo, siendo t0 el instante actual y tm una constante del modelo,
que se tomo igual a 60 dıas.
La orientacion del material se mantiene alineada con el tensor de tensiones promedio,
antes comentado. Dado que en la simulacion de la remodelacion unicamente se modifica
la magnitud de las diferentes cargas y se mantiene la proporcion entre ellas, la estructura
apenas sufre cambios de orientacion.
Bagge [21] simulo tres casos, en los que se partıa de la estructura osea optima para
soportar las cargas de referencia, P , y un volumen de tejido 50 %. En la simulacion S1
alternaba 10 dıas con una carga mitad de la de referencia, P/2, y 10 dıas con una carga
doble de la referencia, 2P . En la grafica 5.16 se observa como crece la fraccion volumetrica
de hueso hasta aproximadamente el 59 %, alcanzandose la convergencia tras unos 6000 dıas.
Es por tanto mayor el efecto de la formacion producida por las cargas 2P que el de la
reabsorcion producida por P/2.
En la simulacion S2 se aplica P/2 durante 3000 dıas seguidos y posteriormente se apli-
ca 2P hasta alcanzar la convergencia, para comparar el efecto que tiene alternar estados
prolongados de desuso con estados prolongados de actividad intensa. Primero disminuye el
volumen de hueso y con el aumento de la carga se produce un aumento brusco de la masa
osea, superando incluso el valor obtenido en la simulacion S1. Es decir, aplicar las mismas
cargas pero repartidas de manera diferente produce resultados de masa osea diferentes.
Para investigar la irreversibilidad del proceso, Bagge [21] simulo un tercer caso, S3, en
el que partiendo de la situacion de equilibrio obtenida al final de S1, se volvıan a aplicar
las cargas de referencia, P , hasta alcanzar un nuevo estado de equilibrio. Una vez alcanzado
este la fraccion volumetrica obtenida es del 50.2 %, ligeramente superior al valor de partida
(50 %), pero tan parecida que Bagge concluye que el proceso de remodelacion osea no es
irreversible, como sugieren Weinans y Prendergast [342].
Algunas limitaciones de este modelo merecen un comentario aparte. En primer lugar hay
que senalar que la simetrıa cubica que se impone al material es demasiado restrictiva. La
disposicion de tejidos en el hueso cortical hace que este tenga, aproximadamente, isotropıa
transversal, mientras que el hueso trabecular es en general anisotropo, aunque se suele
aceptar como ortotropo, siendo muy variable el grado de ortotropıa. Sin duda esta es una
limitacion que influye de manera importante en los resultados. Ası, la diafisis del femur, que
tiene una rigidez en direccion longitudinal mucho mayor que en las otras dos direcciones,
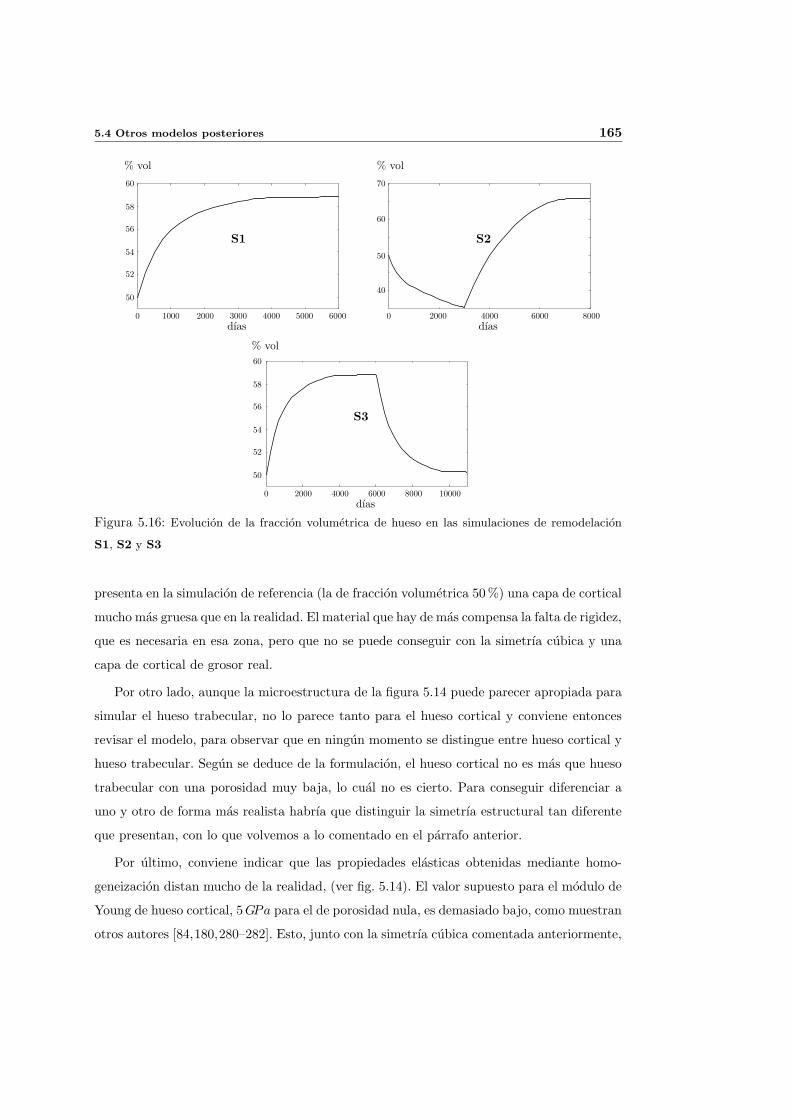
5.4 Otros modelos posteriores 165
dıas dıas
dıas0 2000 4000 6000 8000 10000
0 1000 2000 3000 4000 5000 6000 0 2000 4000 6000 8000
50
52
54
56
58
60
50
52
54
56
58
60
40
50
60
70
% vol % vol
% vol
S1 S2
S3
1
Figura 5.16: Evolucion de la fraccion volumetrica de hueso en las simulaciones de remodelacion
S1, S2 y S3
presenta en la simulacion de referencia (la de fraccion volumetrica 50 %) una capa de cortical
mucho mas gruesa que en la realidad. El material que hay de mas compensa la falta de rigidez,
que es necesaria en esa zona, pero que no se puede conseguir con la simetrıa cubica y una
capa de cortical de grosor real.
Por otro lado, aunque la microestructura de la figura 5.14 puede parecer apropiada para
simular el hueso trabecular, no lo parece tanto para el hueso cortical y conviene entonces
revisar el modelo, para observar que en ningun momento se distingue entre hueso cortical y
hueso trabecular. Segun se deduce de la formulacion, el hueso cortical no es mas que hueso
trabecular con una porosidad muy baja, lo cual no es cierto. Para conseguir diferenciar a
uno y otro de forma mas realista habrıa que distinguir la simetrıa estructural tan diferente
que presentan, con lo que volvemos a lo comentado en el parrafo anterior.
Por ultimo, conviene indicar que las propiedades elasticas obtenidas mediante homo-
geneizacion distan mucho de la realidad, (ver fig. 5.14). El valor supuesto para el modulo de
Young de hueso cortical, 5GPa para el de porosidad nula, es demasiado bajo, como muestran
otros autores [84,180,280–282]. Esto, junto con la simetrıa cubica comentada anteriormente,

166 Modelos de remodelacion osea interna
contribuye a hacer la capa de hueso de cortical de la diafisis mas gruesa que en la realidad.
No obstante las objeciones, la idea de buscar la estructura osea optima es interesante,
y concuerda con las sugerencias de algunos autores, como Currey [83], de que la naturaleza
dispone el hueso de forma que mejor se aprovecha el material para cumplir su funcion
estructural.
5.4.2. Modelo de las tensiones locales uniformes de Tsubota, Adachi
y Tomita
Este modelo se centra en buscar las relaciones entre el estımulo mecanico que recibe
el hueso y los cambios estructurales que sufre a nivel microscopico. El objetivo de estos
cambios estructurales es, segun Tsubota et al. [327], conseguir una distribucion uniforme
de tensiones. Ası, un gradiente de deformaciones promueve la remodelacion produciendo el
flujo de fluidos vitales necesario para activar las celulas oseas. Para comprobar la validez de
la hipotesis anterior estos autores construyeron un modelo de EF 2D de un femur humano,
en el que la geometrıa de las trabeculas se modela con elementos de tipo pıxel, es decir, que
tienen un area de hueso 0 o 1.
Como se ha dicho, la hipotesis fundamental del modelo establece que la remodelacion
osea se produce para conseguir que las tensiones o las deformaciones sean uniformes en el
hueso. La falta de uniformidad, que es la causa de la remodelacion, se mide por el parametro
Γ, que se define como
Γ(xc) = ln
(σcσd,
)(5.115)
donde σc es la tension en el punto xc y σd es una tension representativa del entorno del
punto xc, que viene dada por
σd =
∫
S
w(l)σrdS∫
S
w(l)dS
, (5.116)
donde σr representa la tension del punto xr perteneciente a la superficie trabecular S, del
entorno de xc. La funcion de peso w, que se usa en el promedio ponderado de la ecuacion
(5.116), depende de la distancia l entre xr y xc. Dicho promedio se realiza en un entorno
de tamano lL (ver figura 5.17) alrededor del punto xc, siendo lL un parametro del modelo,
que mide la distancia a la que las celulas oseas pueden sentir el estımulo mecanico y que
Tsubota et al. tomaron igual a 1mm [326].

5.4 Otros modelos posteriores 167
formacion
reabsorcion
trabecula
medula
Figura 5.17: (a) Esquema de una trabecula y el entorno alrededor de un punto xc en el que se
evalua el parametro Γ. (b) Velocidad de remodelacion M en funcion de la falta de uniformidad, Γ.
La falta de uniformidad, Γ, determina la velocidad con que se produce la remodelacion,
M . Esta se entiende como una velocidad de movimiento de la superficie exterior de la
trabecula que en el modelo de pıxeles se traduce en la velocidad a la que aparecen junto a
la superficie pıxeles de hueso nuevos. En la figura 5.17 se observa que en los puntos donde
la tension sea inferior a la tension promedio del entorno, esto es, que tengan Γ < 0, se
produce reabsorcion, M < 0, mientras que si la tension es mayor que la tension promedio,
Γ > 0, se produce formacion, M > 0. Las constantes Γu y Γl son parametros del modelo
que determinan el ancho de la zona muerta, el rango de valores de Γ con poca probabilidad
de remodelacion. Tsubota et al. tomaron Γu = 1.0 y Γl = −2.0 [326].
Este modelo de remodelacion fue aplicado por Tsubota et al. [327] a la extremidad prox-
imal del femur, con el referido modelo de EF 2D de pıxeles, en deformacion plana. Partiendo
de una situacion practicamente isotropa, con las trabeculas dejando huecos elıpticos de di-
mensiones H1 = 714 µm y H2 = 713 µm aplicaron tres casos de carga: abduccion, aduccion
y apoyo sobre una sola pierna. Usaron la tension de von Mises en las ecuaciones (5.115)
y (5.116) y definieron un parametro de no uniformidad para cargas multiples, Γmlt, que
promedia los estados de tension segun su importancia por numero de ciclos:
Γmlt =
3∑
i=1
wiΓi con wi =ni
n1 + n2 + n3(5.117)
donde ni es el numero de ciclos diarios que se repite el caso de carga i. La funcion de peso
w(l) que tomaron fue lineal y decreciente con la distancia
w(l) =
1− l/lL si 0 < l < lL
0 si lL ≤ l(5.118)

168 Modelos de remodelacion osea interna
Figura 5.18: Esquema de la simulacion de carga multiple: (a) Modelo de EF. (b) Estructura
trabecular inicial y elipse del “fabric tensor”. (c) Cargas y condiciones de contorno segun Beaupre et
al. [27]. Figura tomada de Tsubota et al. [327].
La relacion Γ− M de la figura 5.17 se trata numericamente como una funcion de prob-
abilidad. Si M es alto, proximo a Mmax, es probable que aparezca un pıxel nuevo junto al
punto en cuestion. Por el contrario si M esta proximo a Mmin, es probable que el pıxel de
dicho punto desaparezca.
Este proceso, por el cual desaparecen elementos y aparecen otros nuevos se repitio hasta
la convergencia, estimada como el paso a partir del que el numero de pıxeles que cambian
de estado es menor que el 2 % del area trabecular. Eso ocurrio tras 16 pasos de carga y la
distribucion osea quedo como se aprecia en la figura 5.19. Los resultados no son muy realistas
como reconocen los propios autores y las poros obtenidos son excesivamente grandes. En su
opinion es debido a que ya en la situacion de partida son demasiado grandes. Esto pone
de manifiesto una limitacion del modelo: no considera la idea de estımulo mecanico como
medida fisiologica de las tensiones. Ası, el numero de ciclos no influye en la remodelacion mas
que a traves de la proporcion de la ecuacion (5.117). Ni con desuso se produce reabsorcion,
ni con una actividad anormalmente alta se produce hipertrofia, si la proporcion de las cargas
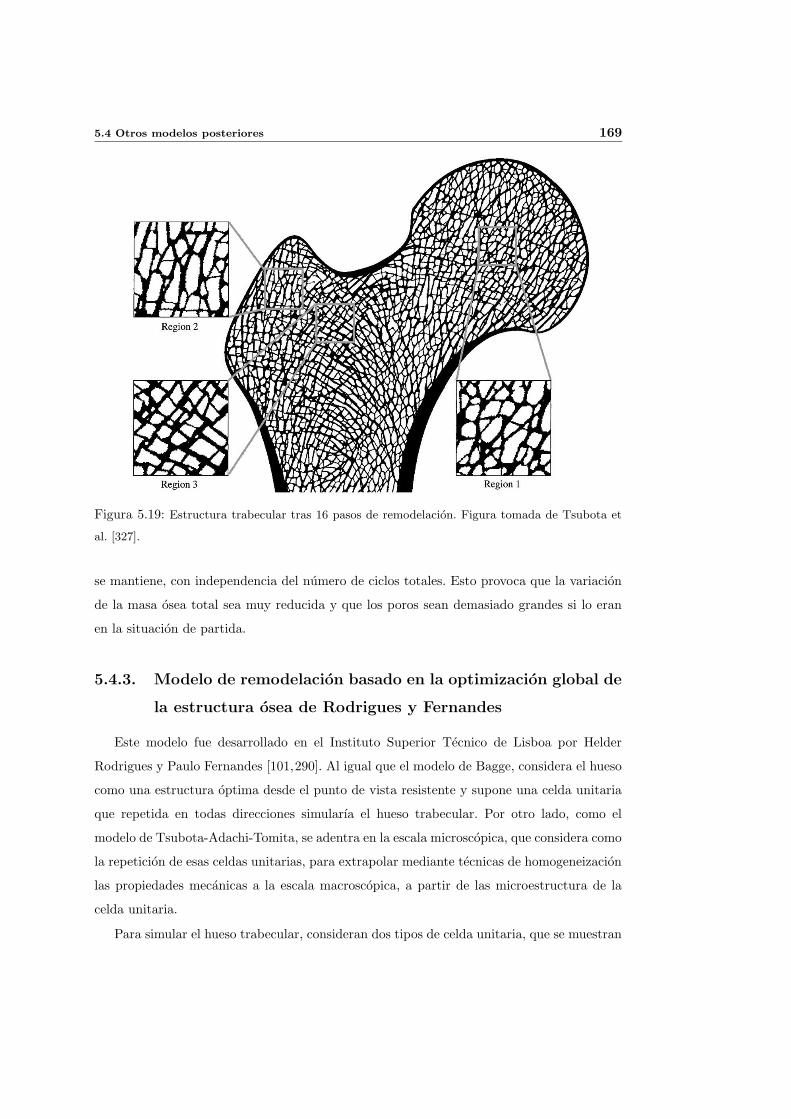
5.4 Otros modelos posteriores 169
Figura 5.19: Estructura trabecular tras 16 pasos de remodelacion. Figura tomada de Tsubota et
al. [327].
se mantiene, con independencia del numero de ciclos totales. Esto provoca que la variacion
de la masa osea total sea muy reducida y que los poros sean demasiado grandes si lo eran
en la situacion de partida.
5.4.3. Modelo de remodelacion basado en la optimizacion global de
la estructura osea de Rodrigues y Fernandes
Este modelo fue desarrollado en el Instituto Superior Tecnico de Lisboa por Helder
Rodrigues y Paulo Fernandes [101,290]. Al igual que el modelo de Bagge, considera el hueso
como una estructura optima desde el punto de vista resistente y supone una celda unitaria
que repetida en todas direcciones simularıa el hueso trabecular. Por otro lado, como el
modelo de Tsubota-Adachi-Tomita, se adentra en la escala microscopica, que considera como
la repeticion de esas celdas unitarias, para extrapolar mediante tecnicas de homogeneizacion
las propiedades mecanicas a la escala macroscopica, a partir de las microestructura de la
celda unitaria.
Para simular el hueso trabecular, consideran dos tipos de celda unitaria, que se muestran

170 Modelos de remodelacion osea interna
en la figura 5.20. La celda a es de tipo cerrado (solo se muestra 1/8 en la figura 5.20a) y en
ella la densidad relativa viene dada por las dimensiones del poro, como µ = 1−a1 a2 a3. Esta
geometrıa es similar a la supuesta en el modelo desarrollado en esta tesis, como se vera mas
adelante en el capıtulo 7. En otro trabajo [101], Rodrigues y sus colaboradores han utilizado
una celda de tipo abierto (figura 5.20b), de densidad relativa µ = 1− b1 b2 − b1 b3 − b2 b3 +
b1 b2 b3.
Figura 5.20: Celdas unitarias usadas en este modelo para modelar el hueso trabecular. (a) Celda
cerrada. (b) Celda abierta. Tomado de Folgado et al. [101].
La densidad relativa, µ, representa el volumen que ocupa la matriz osea dentro de la celda
unitaria. Esta matriz osea se considera homogenea y con propiedades mecanicas identicas
a las del hueso cortical. Ası, dicho hueso cortical vendrıa dado por µ = 1, mientras que
el hueso trabecular es aquel que tiene 0 < µ < 1 [101]. Para evaluar las propiedades de
dicho hueso se utilizan tecnicas de homogeneizacion [298], que se basan en la existencia de
dos escalas geometricas: la escala macro, de dimension caracterıstica la del hueso (D), y la
escala micro, de dimension caracterıstica la de las trabeculas (d). Por medio del factor de
escala ε = d/D, es posible expresar el campo de desplazamientos en funcion de una variable
de posicion macroscopica, x, y otra microscopica, y, definida como y = x/ε. El campo de
desplazamientos se expresa por medio de una expansion asintotica en el parametro ε,
uε(x,y) = u0(x) + ε u1(x,y) + ε2 u2(x,y) + . . . (5.119)

5.4 Otros modelos posteriores 171
Imponiendo el equilibrio de este campo de desplazamientos y tomando el lımite ε → 0,
se encuentra que u0(x), el campo de desplazamientos homogeneizado, es la solucion de un
problema de equilibrio en el que ha sustituido el material formado por celdas unitarias por
un material homogeneizado.
Si el material que forma la celda unitaria es homogeneo (como se supone aquı con el
hueso cortical), las propiedades homogeneizadas del material vienen dadas por:
EHijkl(µ) = Eijkl µ−1
|Y |
∫
U
Eijpm∂χklp∂ym
dY (5.120)
donde U es el subdominio ocupado por el material dentro de la celda unitaria y χkl son un
conjunto de soluciones periodicas del problema de equilibrio
∫
U
Eijpm∂χklp∂ym
∂vi(y)
∂yjdY =
∫
U
Eijkl∂vi(y)
∂yjdY, ∀v Y − Periodica (5.121)
El metodo de homogeneizacion es aplicable a materiales con una microestructura local-
mente periodica, que no es el caso general del tejido oseo. Sin embargo, la aproximacion
que supone aplicar este metodo al hueso queda restringida al nivel microscopico (de la co-
ordenada y), ya que a nivel macroscopico (de la coordenada x) no se impone ningun tipo
de restriccion al material. El metodo de homogeneizacion puede verse con mas detalle en
Sanchez-Palencia [298].
Ademas de depender de las dimensiones del poro, el tensor de comportamiento depende
de la orientacion de las celdas unitarias, θ, a traves de
(EHijkl)θ = RimRjnRkpRlqEHmnpq , (5.122)
donde R es el tensor de giro asociado a la rotacion θ.
Partiendo de estas relaciones entre la microestructura y el tensor de comportamiento,
Rodrigues y Fernandes desarrollan un modelo de remodelacion que se basa en la optimalidad
de la estructura osea desde el punto de vista resistente [290]. Segun este criterio de opti-
malidad, la estructura osea se adapta al entorno mecanico externo de forma que consigue la
maxima rigidez con la mınima cantidad de hueso. Ası, si un hueso esta sometido a una serie
de fuerzas fp, que produce unos desplazamientos up, la estructura osea sera la solucion del
siguiente problema de optimizacion:

172 Modelos de remodelacion osea interna
minai,θ
(NC∑
p=1
αp fp · up + β
∫
Ω
µ(a)dΩ
)
sujeto a K(a, θ)up = fp(5.123)
La condicion impuesta es la de equilibrio a nivel global y las dimensiones del poro, obvia-
mente, deben cumplir 0 ≤ ai ≤ 1.1 El primer termino de la funcion objetivo es un promedio
ponderado de la energıa de deformacion de la estructura en los p casos de carga, donde NC
es el numero de casos de carga y αp son los pesos correspondientes a cada uno de ellos, que
cumplen que∑NCp=1 α
p = 1. El segundo termino controla la cantidad de hueso que se utiliza
para optimizar la rigidez de la estructura y puede variar, dado que el hueso se considera un
sistema abierto. La respuesta del hueso es distinta en cada individuo, aunque se mantenga
constante la carga. Para tener esto en cuenta se introduce el parametro β, que puede en-
globar diversos aspectos que influyen en la respuesta, como la edad, el estado de salud, los
trastornos hormonales, etc. Ası, cuanto mayor es β, menor es la cantidad de hueso disponible
para conseguir una cierta rigidez. Con valores de β muy elevados Folgado et al. [101] estu-
dian la incidencia de la osteoporosis en la distribucion y orientacion del material oseo. El
problema de optimizacion se resuelve mediante multiplicadores de Lagrange, resultando las
siguientes condiciones de optimalidad
NC∑
p=1
(αp up
∂K(a, θ)
∂aiup)− β
∫
Ω
∂µ(a)
∂aidΩ = 0 (5.124)
NC∑
p=1
(αp up
∂K(a, θ)
∂θup)
= 0 (5.125)
Estas ecuaciones se pueden interpretar como unas ecuaciones de remodelacion, de forma
que su cumplimiento indica que se ha alcanzado la situacion de equilibrio de remodelacion
(no se produce mas remodelacion). Si no se cumplen, es necesario modificar localmente la
orientacion de las celdas y las dimensiones de los poros, hasta que se cumpla. Estos autores
han aplican el modelo a un femur humano intacto y a otro con un implante de cadera [290].
Asimismo, han analizado una vertebra humana [101], estudiando la influencia que tiene la
1En todas estas ecuaciones se han sustituido las dimensiones de la celda unitaria por las de la celda
cerrada, ai, sin perdida de generalidad. En caso de utilizar la celda abierta no hay mas que cambiar ai por
bi.

5.4 Otros modelos posteriores 173
osteoporosis en su estructura. En la figura 5.20a se muestra el modelo de elementos finitos de
1/4 de vertebra. La vertebra se aproxima por un hiperboloide de seccion circular sometido a
una presion uniforme en las superficies inferior y superior. Tras la remodelacion, la estructura
que optimiza la rigidez con la mınima cantidad de material es la que se muestra en la figura
5.20b, con la orientacion mostrada en la figura 5.20c.
a b c
1
Figura 5.21: (a) Modelo de elementos finitos de un 1/4 de vertebra humana. (b) Resultados de
densidad relativa en el equilibrio de remodelacion. (c) Resultados de orientacion de las celdas en el
equilibrio de remodelacion. Tomado de Folgado et al. [101].
Se observa que la situacion de equilibrio de remodelacion esta caracterizada por una
zona central de hueso esponjoso y hueso cortical en la periferia. La direccion de mayor
rigidez coincide con la direccion de carga. La conclusion a la que llegan en su estudio de la
influencia de la osteoporosis es que cuanto menor es la cantidad de hueso, mayor es el grado
de ortotropıa. Ası, con la osteoporosis disminuye la rigidez del hueso, pero lo hace menos en
la direccion de carga que en las otras dos.


Capıtulo 6
Obtencion de las propiedades
mecanicas de la mandıbula
humana mediante simulacion
numerica. Resultados I.
6.1. Introduccion
En este capıtulo se muestran los resultados obtenidos en la simulacion de la morfogenesis
de la mandıbula. No es una morfogenesis en sentido estricto, ya que la forma de la mandıbu-
la es un dato de partida (no se usa modelo de remodelacion osea externa alguno) y tan
solo se pretende estimar la distribucion de densidad y la orientacion de la microestructura
osea, parametros ambos que influyen en las propiedades elasticas. Hay que recordar que la
morfogenesis es la primera fase del analisis y sirve de punto de partida para la segunda fase.
En esta se analiza el implante dental y sus resultados se veran en un capıtulo posterior. Los
resultados de propiedades elasticas obtenidos aquı se usan en el modelo de elementos finitos
empleado en la segunda fase.
La situacion de partida de la morfogenesis es completamente irreal: distribucion uniforme
del tejido, con una densidad inicial de 0.5 g/cm3, correspondiente a un hueso trabecular muy
175

176 Obtencion de las propiedades mecanicas de una mandıbula humana
ligero, y simetrıa isotropa. A partir de esta situacion y aplicando repetidamente las cargas
de masticacion descritas en los capıtulos 2 y 3, cambia la distribucion y la orientacion de
los tejidos, segun el modelo de remodelacion osea interna de Doblare y Garcıa [91, 115],
alcanzandose tras un cierto tiempo una situacion de equilibrio.
En primer lugar se mostraran las distribuciones de tension que los diferentes casos de
carga considerados en el proceso masticatorio originan en el modelo inicial de la mandıbula
(isotropo y con densidad uniforme 0.5 g/cm3). A la vista de esos resultados se justificaran
las distribuciones de densidad y propiedades elasticas obtenidas en el apartado siguiente,
dedicado a la simulacion de referencia. Dicha simulacion esta realizada con los parametros
que mejor ajustan los resultados a la realidad y sera la utilizada para la segunda fase del
analisis. Por esta razon, la simulacion de referencia se analiza con especial detalle: ademas de
dar la distribucion de densidad y propiedades elasticas, se comprueba que efectivamente se
ha alcanzado la situacion de equilibrio, en particular en los desplazamientos, que se usan para
esa segunda fase; se comparan las tensiones en la situacion de equilibrio con las representadas
anteriormente para la situacion inicial; y se analizan las fuerzas de apriete con los dientes,
comparandolas con algunos estudios experimentales.
Por ultimo, se realiza un analisis de sensibilidad de diferentes parametros del modelo
de remodelacion utilizado, ası como del patron de masticacion, partiendo de la simulacion
de referencia. Se pretende conocer ası la influencia de dichos parametros en la respuesta
remodelatoria y justificar los valores utilizados en la simulacion de referencia.
6.2. Distribucion de tensiones inicial
A continuacion, las figuras 6.1 a 6.4 muestran la distribucion de tensiones de von Mises
que provocan los diferentes casos de carga en el modelo de partida. El modelo no es com-
pletamente simetrico ya que en el lado izquierdo no se ha dispuesto el tercer molar. Sin
embargo, esto no tiene gran influencia por la similitud entre las propiedades elasticas del
hueso cortical y la dentina que forma el diente. Ası, los resultados de masticaciones simetri-
cas respecto al plano sinfisario practicamente coinciden. Por esta razon, solo se presentan
aquı los casos de carga siguientes: mordida incisiva, mordida canina con el lado derecho y
masticaciones con los molares del lado derecho, primero y segundo.
En todos los casos las tensiones mas altas estan concentradas cerca de los apoyos, condilos
y dientes con los que se muerde o mastica. Las tensiones en los dientes son especialmente
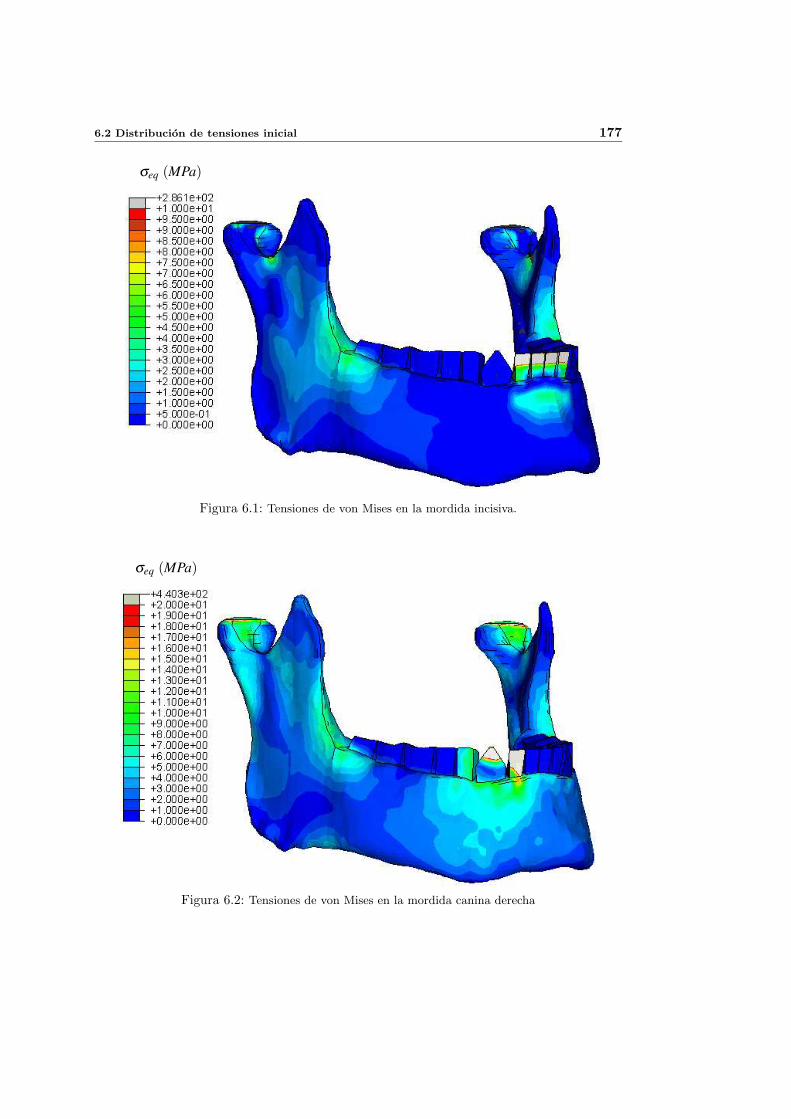
6.2 Distribucion de tensiones inicial 177
σeq (MPa)
1
Figura 6.1: Tensiones de von Mises en la mordida incisiva.
σeq (MPa)
1
Figura 6.2: Tensiones de von Mises en la mordida canina derecha
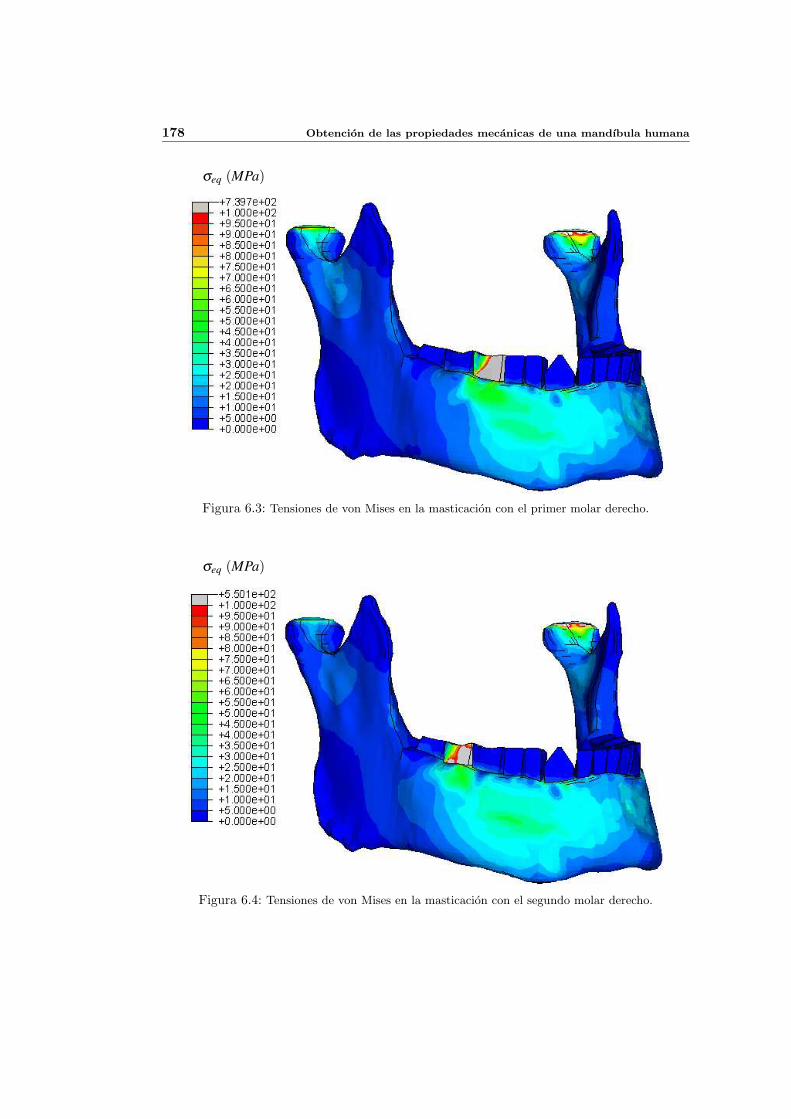
178 Obtencion de las propiedades mecanicas de una mandıbula humana
σeq (MPa)
1
Figura 6.3: Tensiones de von Mises en la masticacion con el primer molar derecho.
σeq (MPa)
1
Figura 6.4: Tensiones de von Mises en la masticacion con el segundo molar derecho.

6.2 Distribucion de tensiones inicial 179
altas, incluso, estan fuera del rango representado, debido a que los dientes son mucho mas
rıgidos que el hueso circundante, especialmente al principio del analisis, cuando el hueso
tiene una densidad tan baja.
Las tensiones en las mordidas, tanto incisiva como canina, son mucho menores que en las
masticaciones. Para encontrar una explicacion a esto hay que recurrir a las fuerzas ejercidas
por los dientes en cada caso, que se analizan mas adelante, para comprobar que en las
mordidas son mucho menores que en las masticaciones. Esto puede indicar que los datos
de actividad muscular corresponden en estos casos a unas mordidas suaves. Otras mordidas
de mayor intensidad, que suelen denominarse aprietes, podrıan haberse incluido, pero no
se ha hecho, ya que no son muy habituales. Las mordidas tambien se producen con mucha
menor frecuencia que las masticaciones, por lo que su influencia sobre la remodelacion osea
sera pequena.
En la figura 6.5 se muestra la distribucion de las componentes del tensor de tensiones en
el caso de carga MI1. Se muestra una seccion perpendicular al eje del cuerpo mandibular,
proxima al primer molar derecho. Puede comprobarse que los valores de σx son mucho
mayores que los de σy y σz, lo que indica que hay un predominio de la flexion. Tambien
puede comprobarse que la lınea neutra forma unos 60 respecto al eje Y , y que en la zona
alveolar, las tensiones estan afectadas de manera importante por la presencia del diente,
mucho mas rıgido. Se observa en la distribucion de σy como el diente comprime el hueso
alveolar del lado bucal, a la izquierda, mientras que en el lado lingual las tensiones son muy
pequenas. Tambien se puede apreciar una pequena torsion de la seccion: los valores de σxz
tienen distinto signo en las caras lingual y bucal, y provocan un giro dextrogiro de la seccion,
si se observa la mandıbula de frente. La distribucion de las tensiones en las mordidas incisiva
y canina tiene una forma similar, aunque con valores mucho mas pequenos, como ya se ha
comentado.
A la vista de estos resultados se puede concluir que la accion de los musculos elevadores
producen flexion y torsion en la mandıbula. La torsion se produce no solo por ser curva la
directriz de la misma sino tambien por el hecho de que las condiciones de carga y de apoyo
no son simetricas, excepto en la mordida incisiva. La estructura optima que deberıa tener
la mandıbula, para resistir estas cargas con el mınimo peso, es una estructura tubular. Esta
es precisamente la que tiene, como se vera en el siguiente apartado.
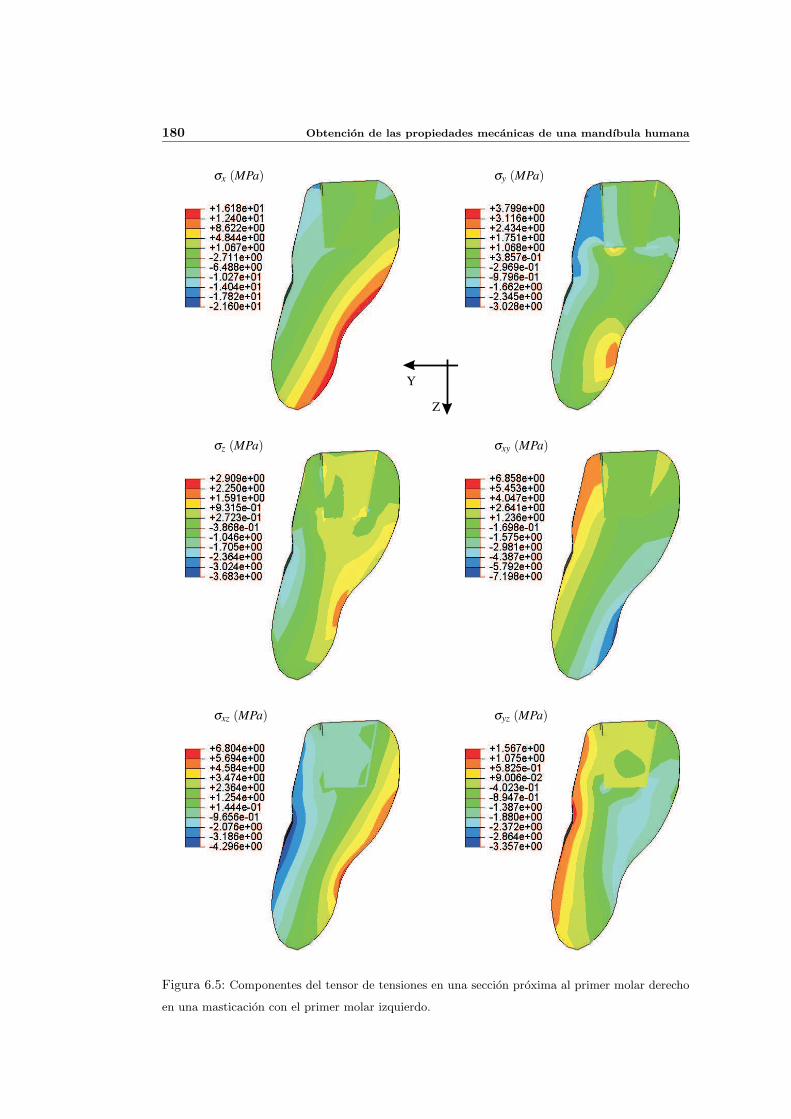
180 Obtencion de las propiedades mecanicas de una mandıbula humana
Y
Z
σx (MPa) σy (MPa)
σz (MPa) σxy (MPa)
σxz (MPa) σyz (MPa)
1Figura 6.5: Componentes del tensor de tensiones en una seccion proxima al primer molar derecho
en una masticacion con el primer molar izquierdo.

6.3 Distribucion de densidad osea y propiedades elasticas 181
6.3. Distribucion de densidad osea y propiedades elasti-
cas
6.3.1. Introduccion y resultados experimentales de referencia
El modelo matematico de remodelacion osea interna desarrollado por Doblare y Garcıa
[91, 115], y descrito en el capıtulo 5, tiene algunos parametros cuyos valores han sido
obtenidos experimentalmente y otros que estos autores han estimado, basandose en los re-
sultados numericos que proporciona el modelo y eligiendo aquellos que producen resultados
mas ajustados a la realidad:
1) El parametro m, que aparece en las ecuaciones (5.14), (5.22) y (5.102), tiene en cuenta
la influencia del numero de ciclos en la respuesta remodelatoria, fue estimado por Whalen
et al. [343] para el calcaneo. Segun estos autores, los valores mas adecuados estan entre 3 y
8. Posteriormente, otros autores [27, 91, 166] han usado m = 4 y ese sera el valor adoptado
aquı.
2) En la ecuacion (6.3) se introdujo el parametro ω para ponderar la influencia de la
componente desviadora del estımulo, esto es, la anisotropıa de la carga, en la respuesta
remodelatoria. Doblare y Garcia aplicaron este modelo al femur humano usando ω = 0.1.
Las cargas en el femur son principalmente de flexion, mientras que en la mandıbula, la
torsion que producen las masticaciones unilaterales tambien es importante, como se acaba
de comprobar. En flexion-torsion la componente desviadora del tensor de tensiones es mayor
que en flexion pura, y por tanto, cabe esperar que valores mas altos de ω proporcionen
mejores resultados en la mandıbula. Se han analizado tres casos: ω = 0.18 , 0.24 y 0.3.
3) El siguiente parametro analizado es el estımulo de referencia, Ψ∗
t , que aparece en el
criterio de remodelacion, ecuacion (5.103) y esta tomado del modelo isotropo de Stanford
(ecuacion (5.30)). Este parametro junto con w definen la zona muerta de remodelacion:
el rango de valores del estımulo para los que no hay respuesta remodelatoria. Beaupre et
al. [27] usaron 50MPa/dıa para el femur humano y el mismo valor adoptaron posteriormente
Doblare y Garcıa [91,115]. Sin embargo, hay que tener en cuenta que el femur soporta cargas
mucho mayores que las de masticacion. Tambien es mayor el numero de pasos que el numero
de masticaciones diarias y en consecuencia, el estımulo que soporta el femur sera mayor
que el soportado por la mandıbula. El estımulo de referencia o estımulo de equilibrio, se
adapta de alguna forma al estımulo externo aplicado como establece Turner en el principio

182 Obtencion de las propiedades mecanicas de una mandıbula humana
de acomodacion celular [329]. Si esta adaptacion se produce durante el crecimiento del hueso,
el estımulo de referencia, Ψ∗
t , en un adulto debe ser algo menor en la mandıbula que en el
femur. Se han adoptado tres valores para su comparacion: 50, 25 y 10MPa/dıa. Para el
semiancho de la zona muerta se ha adoptado el mismo criterio que usan Doblare y Garcıa,
esto es, w = 0.25Ψ∗
t
4) La curva de remodelacion usada es del tipo representado en la figura 5.9, siendo las
pendientes c1 = c2 = 0.02µm/dıa, igual que las utilizadas para el femur.
5) El numero de ciclos diarios tambien es menor, como ya se ha dicho. En el femur se
reparten 10000 ciclos entre tres tipos de actividades. Para la mandıbula se supondran 500
ciclos diarios repartidos entre todos los casos de carga, cualquiera que sea la secuencia de
masticacion. Se hace la simplificacion introducida por Jacobs [166], comentada en un capıtulo
precedente, que consiste en aplicar un unico caso de carga por dıa. Los dıas sucesivos ocurren
distintos casos de carga, de forma que se aplican todos los casos que incluye la secuencia de
masticacion y en la misma proporcion establecida en ella.
6) Las propiedades elasticas se relacionan con el “fabric tensor” por medio de la ecuacion
(5.98), pero tambien influye en el modelo de remodelacion la relacion existente entre el modu-
lo de Young y la densidad aparente. Doblare y Garcıa usaron para el femur la correlacion
obtenida por Beaupre experimentalmente, (5.36). En esta tesis se ha usado otra mas actual,
obtenida por Hernandez de forma experimental [144]. La correlacion de Beaupre fue utiliza-
da en un trabajo previo, durante el desarrollo de esta tesis [223], proporcionando resultados
con una peor aproximacion que los mostrados a continuacion. En realidad Hernandez rela-
ciono el modulo de Young con la fraccion volumetrica de hueso y la fraccion de ceniza, pero
esta ultima, que varıa como consecuencia del proceso de mineralizacion, no se considera una
variable de este modelo de remodelacion y se supondra constante e igual a 0.65 [220]. Por
tanto, en ese caso, la correlacion se puede escribir:
E = 3204ρ2.58 (6.1)
donde el modulo de Young esta dado en MPa y ρ representa la densidad aparente.
6.3.2. Simulacion de referencia
La simulacion que mas se aproxima a las medidas experimentales se ha denominado
simulacion de referencia y es aquella en la que se utilizan los siguientes parametros:

6.3 Distribucion de densidad osea y propiedades elasticas 183
Simulacion de referencia
m = 4
ω = 0.24
Ψ∗
t = 25MPa/dıa
w = 0.25Ψ∗
c1 = c2 = 0.02µm/dıa
nc = 500 ciclos diarios
Correlacion de Hernandez [144]
Tabla 6.1: Parametros del modelo de remodelacion utilizados en la simulacion de referencia.
La secuencia de masticacion usada en esta simulacion es la que se denomino S2, que com-
prende cuatro casos de carga, MD1-MI1-MD2-MI2, repetidos en este orden hasta alcanzar
el equilibrio de remodelacion. Si se suponen 500 ciclos de masticacion diarios, la aplicacion
de esta secuencia implica que en cada dıa se producen 125 ciclos correspondientes a cada
tipo de carga. Con la agrupacion propuesta por Jacobs (figura 5.7) se aplican 500 ciclos de
carga MD1 el primer dıa, 500 ciclos de carga MI1 el dıa siguiente y ası sucesivamente hasta
completar la secuencia, para empezar otra secuencia el cuarto dıa. El hecho de aplicar 500
ciclos de un tipo de carga no significa que haya que resolver 500 problemas elasticos para
simular un dıa. Puesto que se supone que la remodelacion se produce al final del dıa, se
resuelve un solo problema elastico para ese tipo de carga y en el algoritmo de remodelacion
se usa nc = 500 en el calculo del estımulo diario (ecuacion (5.23)).
Convergencia de los resultados
Para comprobar que se ha alcanzado el equilibrio de remodelacion, al menos globalmente,
se definio la variable conv,
conv =
∫v∆ρ dV∫vρ dV
(6.2)
que mide la variacion de la masa de la mandıbula en cada paso de carga, respecto a la
masa total. El analisis se detuvo cuando conv < 0.00015, lo que ocurrio tras 624 dıas de
masticacion. En la figura 6.6 se muestra la evolucion del parametro conv en la simulacion
de referencia.
La variacion temporal de masa osea tras 624 dıas es muy pequena y se puede considerar
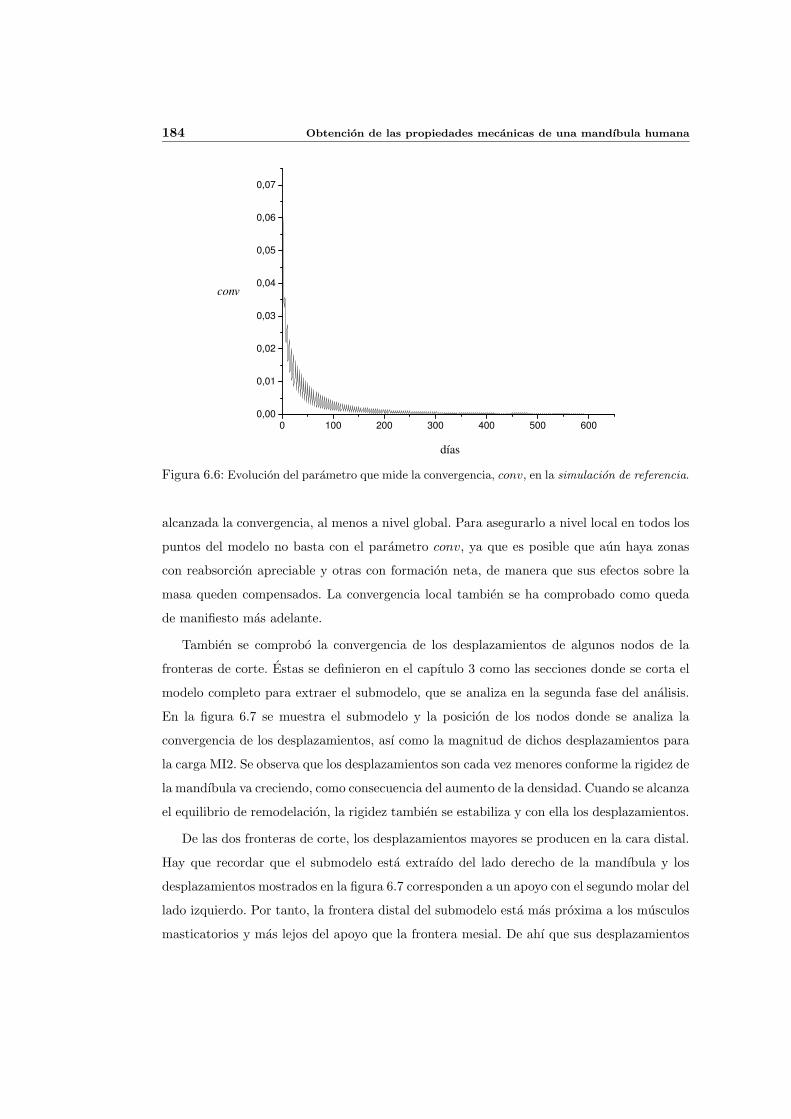
184 Obtencion de las propiedades mecanicas de una mandıbula humana
0 100 200 300 400 500 6000,00
0,01
0,02
0,03
0,04
0,05
0,06
0,07
dıas
conv
1
Figura 6.6: Evolucion del parametro que mide la convergencia, conv, en la simulacion de referencia.
alcanzada la convergencia, al menos a nivel global. Para asegurarlo a nivel local en todos los
puntos del modelo no basta con el parametro conv, ya que es posible que aun haya zonas
con reabsorcion apreciable y otras con formacion neta, de manera que sus efectos sobre la
masa queden compensados. La convergencia local tambien se ha comprobado como queda
de manifiesto mas adelante.
Tambien se comprobo la convergencia de los desplazamientos de algunos nodos de la
fronteras de corte. Estas se definieron en el capıtulo 3 como las secciones donde se corta el
modelo completo para extraer el submodelo, que se analiza en la segunda fase del analisis.
En la figura 6.7 se muestra el submodelo y la posicion de los nodos donde se analiza la
convergencia de los desplazamientos, ası como la magnitud de dichos desplazamientos para
la carga MI2. Se observa que los desplazamientos son cada vez menores conforme la rigidez de
la mandıbula va creciendo, como consecuencia del aumento de la densidad. Cuando se alcanza
el equilibrio de remodelacion, la rigidez tambien se estabiliza y con ella los desplazamientos.
De las dos fronteras de corte, los desplazamientos mayores se producen en la cara distal.
Hay que recordar que el submodelo esta extraıdo del lado derecho de la mandıbula y los
desplazamientos mostrados en la figura 6.7 corresponden a un apoyo con el segundo molar del
lado izquierdo. Por tanto, la frontera distal del submodelo esta mas proxima a los musculos
masticatorios y mas lejos del apoyo que la frontera mesial. De ahı que sus desplazamientos
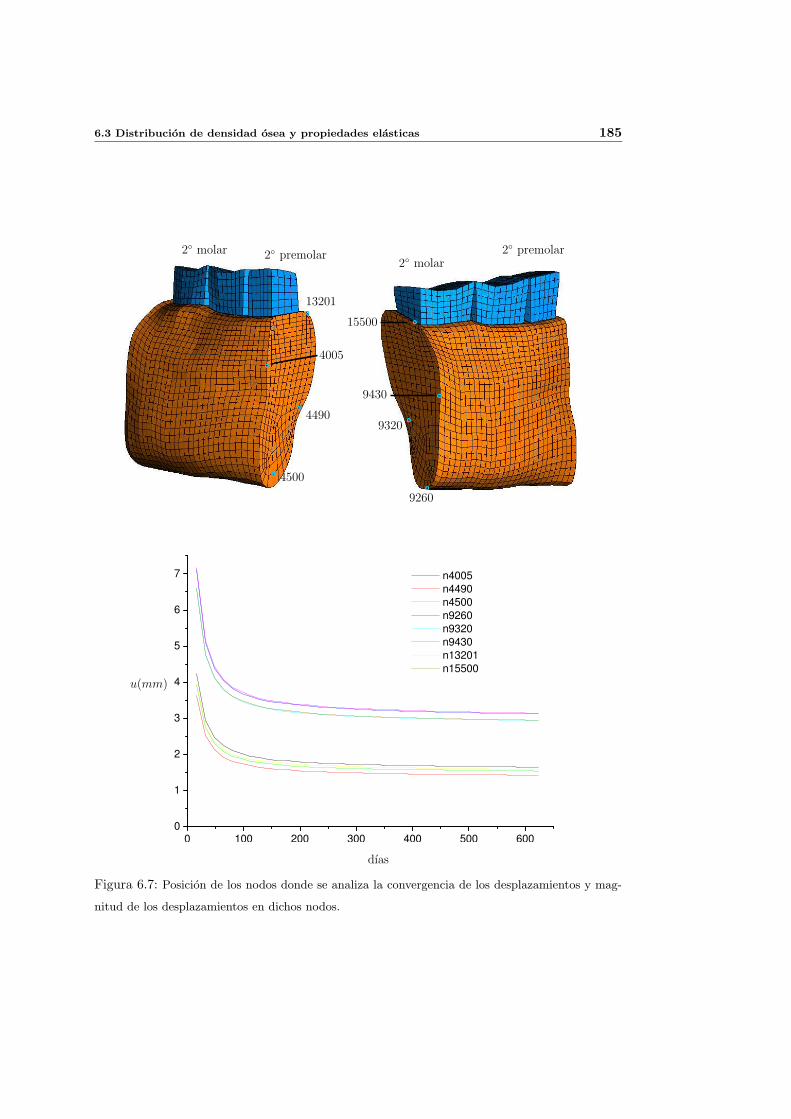
6.3 Distribucion de densidad osea y propiedades elasticas 185
4005
4500
4490
13201
9260
9320
9430
15500
2 premolar2 molar2 molar
2 premolar
0 100 200 300 400 500 6000
1
2
3
4
5
6
7
n4005 n4490 n4500 n9260 n9320 n9430 n13201 n15500
dıas
u(mm)
1
Figura 6.7: Posicion de los nodos donde se analiza la convergencia de los desplazamientos y mag-
nitud de los desplazamientos en dichos nodos.

186 Obtencion de las propiedades mecanicas de una mandıbula humana
sean mayores.
Los incrementos de desplazamientos entre pasos de carga consecutivos estan en torno al
0.1 %, en el dıa 624, en el que se detiene el analisis, por lo que se puede suponer alcanzada la
convergencia de la solucion. Aunque la convergencia local de la densidad aparente y de las
propiedades elasticas no se haya comprobado en todos los puntos de la mandıbula, el hecho
de que los desplazamientos se hayan estabilizado en las fronteras de corte es importante. Este
hecho permite usar dichos desplazamientos como condiciones de contorno en el submodelo,
si se tiene en cuenta que la solucion ası obtenida puede no ser valida localmente, cerca de las
fronteras de corte, pero sı en la zona donde se colocara el implante, que esta suficientemente
alejada de ellas.
Densidad y propiedades elasticas obtenidas con la simulacion numerica. Com-
paracion con resultados experimentales
La distribucion de densidad osea tras 624 dıas de masticacion se muestra en la figura 6.8.
ρ(g/cm3)
1
Figura 6.8: Distribucion de densidad osea en la simulacion de referencia.
Se puede comprobar que practicamente toda la mandıbula esta cubierta por una capa de
hueso cortical muy compacto. De hecho, se alcanza la maxima densidad aparente que permite
el modelo, 1.92 g/cm3, valor tomado de Beaupre et al. [27], excepto en dos zonas concretas:
la apofisis coronoide y la tuberosidad pterigomaseterica, en el angulo de la mandıbula. En
cualquier caso, en estas zonas se tiene un hueso cortical mas ligero o un trabecular compacto
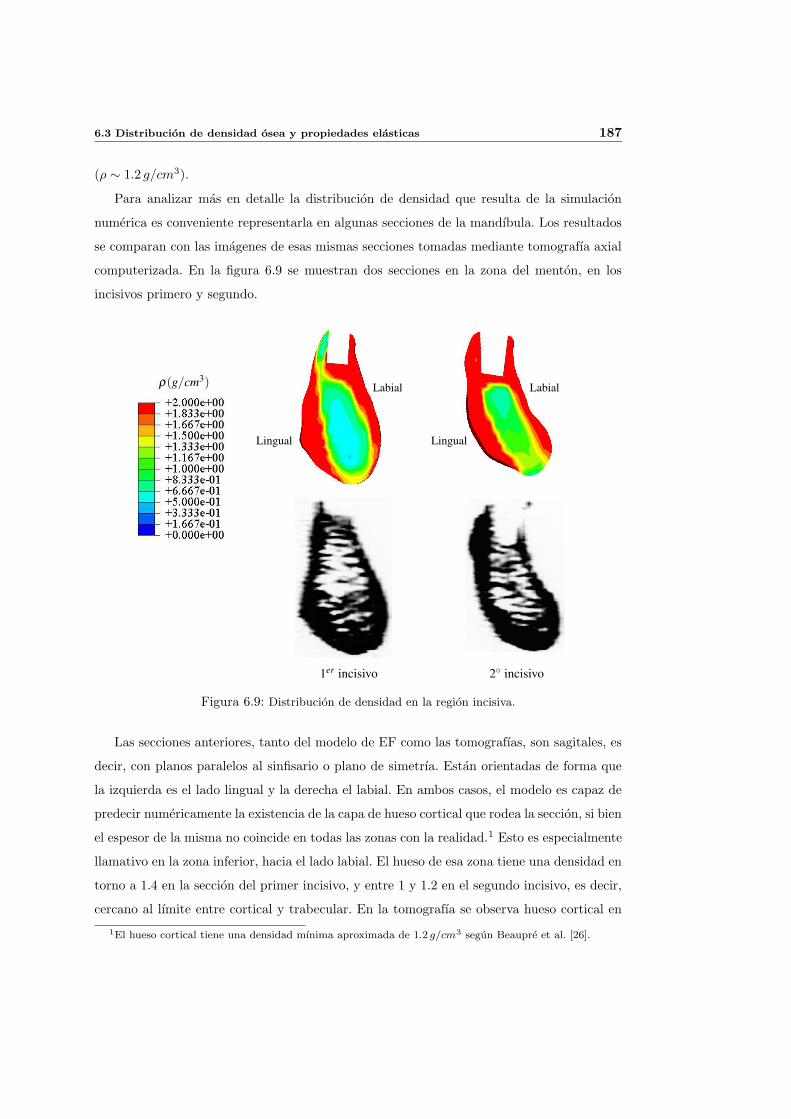
6.3 Distribucion de densidad osea y propiedades elasticas 187
(ρ ∼ 1.2 g/cm3).
Para analizar mas en detalle la distribucion de densidad que resulta de la simulacion
numerica es conveniente representarla en algunas secciones de la mandıbula. Los resultados
se comparan con las imagenes de esas mismas secciones tomadas mediante tomografıa axial
computerizada. En la figura 6.9 se muestran dos secciones en la zona del menton, en los
incisivos primero y segundo.
Lingual
Labial
Lingual
Labialρ(g/cm3)
1er incisivo 2 incisivo
1
Figura 6.9: Distribucion de densidad en la region incisiva.
Las secciones anteriores, tanto del modelo de EF como las tomografıas, son sagitales, es
decir, con planos paralelos al sinfisario o plano de simetrıa. Estan orientadas de forma que
la izquierda es el lado lingual y la derecha el labial. En ambos casos, el modelo es capaz de
predecir numericamente la existencia de la capa de hueso cortical que rodea la seccion, si bien
el espesor de la misma no coincide en todas las zonas con la realidad.1 Esto es especialmente
llamativo en la zona inferior, hacia el lado labial. El hueso de esa zona tiene una densidad en
torno a 1.4 en la seccion del primer incisivo, y entre 1 y 1.2 en el segundo incisivo, es decir,
cercano al lımite entre cortical y trabecular. En la tomografıa se observa hueso cortical en
1El hueso cortical tiene una densidad mınima aproximada de 1.2 g/cm3 segun Beaupre et al. [26].

188 Obtencion de las propiedades mecanicas de una mandıbula humana
esta zona, aunque parece que de baja densidad por el poro que se observa en la seccion del
segundo incisivo.
El software del tomografo TAC permitio medir la densidad en esa zona mediante la escala
de Hounsfield. Este metodo de medicion determina, dentro de una escala de grises, el nivel
de oscuridad de los pıxeles de una determinada ventana de muestra, en la que se toma la
media. La escala se calibra asignando un valor de -1000 al aire y 0 al agua, correspondiendole
al hueso compacto valores superiores a +1000. En la figura 6.10 muestra la tomografıa y la
posicion de las ventanas donde se midio la densidad. En la region medular, (1), la densidad
Hounsfield es 288; en la ventana (2), inferior lingual, 1775; en (3), inferior, 1633; y por
ultimo, en la ventana inferior labial, (4), 1932.
Lingual Labial
1
2
3
4
1
Figura 6.10: Distribucion de densidad en la region incisiva y posicion de las unidades o ventanas
de Hounsfield.
Se deduce, por tanto, que la parte inferior es la zona de menor densidad de la capa
cortical, lo que esta de acuerdo con los resultados numericos obtenidos, si bien la diferencia
no es tan grande como estos predicen. Hay que decir que en esta zona la convergencia de
los resultados es muy lenta. Ya se ha comentado que la convergencia global se ha alcanzado,
pero esto no asegura la convergencia local en todos los puntos y esta es precisamente una
de las zonas donde no se ha alcanzado dicha convergencia, al detener el analisis tras 624
dıas de masticacion. La tendencia de la densidad en esta zona es seguir aumentando, muy
poco a poco. La lentitud de esta convergencia es debida a que el estımulo mecanico esta muy
proximo a la zona muerta y esto se debe, a su vez, a que las tensiones son relativamente

6.3 Distribucion de densidad osea y propiedades elasticas 189
pequenas en esta zona, en los casos que contempla la secuencia S2, masticaciones con los
molares unicamente. Al incluir mordidas con los caninos e incisivos se carga mas esta zona
como se vera mas adelante, cuando se estudie la influencia del patron de masticacion.
En la parte superior de las dos secciones anteriores, la zona del proceso alveolar, los
resultados tambien difieren un tanto de la realidad. Hay que decir, en primer lugar, que
el proceso alveolar, que en esta zona es muy delgado, no se ha conservado ıntegro en la
mandıbula y no aparece por tanto en las tomografıas. Por otro lado, hay que tener en
cuenta que la geometrıa de los dientes y del proceso alveolar ha sido aproximada, que no
medida, a partir de las medidas del hueso basal, dada la dificultad de medir los alveolos
con la maquina de medicion por coordenadas. Esta discrepancia de los resultados en la zona
superior es una tonica general de todas las secciones.
Las siguientes tomografıas, mostradas en las figuras 6.11 y 6.12, son secciones con un
plano coronal. Sin embargo, las secciones del modelo de EF que aparecen en dichas figuras,
son perpendiculares al eje del cuerpo mandibular, porque ası son las rebanadas de elementos.
Por tanto, las secciones que se comparan no son exactamente las mismas y la relacion entre
sus anchos es el coseno del angulo formado por ambos planos. Como este angulo es pequeno,
aproximadamente 8, las diferencias no son muy importantes. En la figura 6.11 se muestran
dos secciones en la region premolar, concretamente en el segundo premolar y en la zona
mesial del primer molar.
En ambas secciones se observa una capa de cortical bastante gruesa, tanto en la tomo-
grafıa como en el modelo de EF. En la seccion premolar se observan algunas coincidencias
resenables. En primer lugar, la zona densa debajo del diente no esta presente en la tomo-
grafıa pero en su lugar esta el hueco que aquel ocupaba. El premolar se ha modelado mas
corto de lo que es en realidad y en la zona de la raız, la ausencia de un material con una
rigidez elevada como es la del diente, se compensa poniendo un tejido mas denso. En el
tercio superior lingual se observa una zona de baja densidad entre la capa de cortical y el
diente y rodeando a este una capa muy delgada de cortical.
En la seccion del primer molar destaca la ausencia del diente en la tomografıa. Esta
vez la perdida se produjo con anterioridad a la muerte. Esto se observa en la mandıbula a
simple vista, ya que por remodelacion osea se ha reabsorbido el proceso alveolar y el alveolo
se ha rellenado de hueso. En la tomografıa se observa un hueso trabecular ligero en el lugar
del hueco y una fina capa de cortical a modo de tapa. Llama la atencion que esta capa
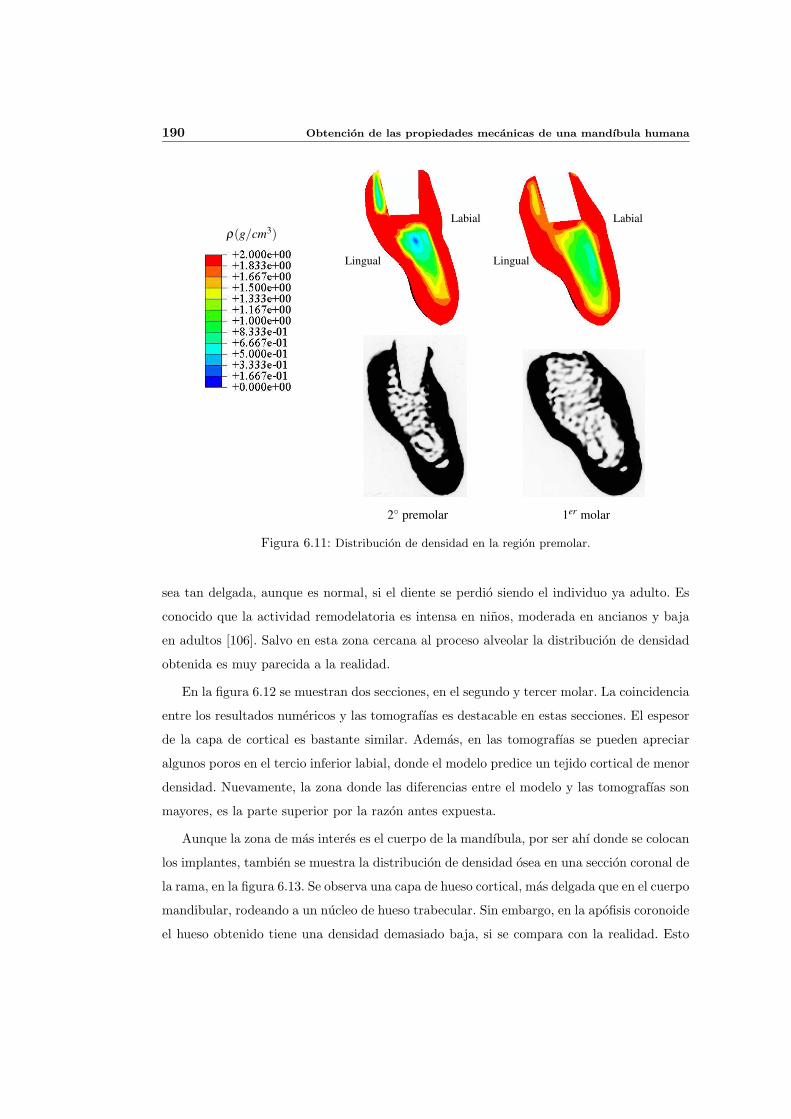
190 Obtencion de las propiedades mecanicas de una mandıbula humana
Lingual
Labial
Lingual
Labialρ(g/cm3)
2 premolar 1er molar
1
Figura 6.11: Distribucion de densidad en la region premolar.
sea tan delgada, aunque es normal, si el diente se perdio siendo el individuo ya adulto. Es
conocido que la actividad remodelatoria es intensa en ninos, moderada en ancianos y baja
en adultos [106]. Salvo en esta zona cercana al proceso alveolar la distribucion de densidad
obtenida es muy parecida a la realidad.
En la figura 6.12 se muestran dos secciones, en el segundo y tercer molar. La coincidencia
entre los resultados numericos y las tomografıas es destacable en estas secciones. El espesor
de la capa de cortical es bastante similar. Ademas, en las tomografıas se pueden apreciar
algunos poros en el tercio inferior labial, donde el modelo predice un tejido cortical de menor
densidad. Nuevamente, la zona donde las diferencias entre el modelo y las tomografıas son
mayores, es la parte superior por la razon antes expuesta.
Aunque la zona de mas interes es el cuerpo de la mandıbula, por ser ahı donde se colocan
los implantes, tambien se muestra la distribucion de densidad osea en una seccion coronal de
la rama, en la figura 6.13. Se observa una capa de hueso cortical, mas delgada que en el cuerpo
mandibular, rodeando a un nucleo de hueso trabecular. Sin embargo, en la apofisis coronoide
el hueso obtenido tiene una densidad demasiado baja, si se compara con la realidad. Esto
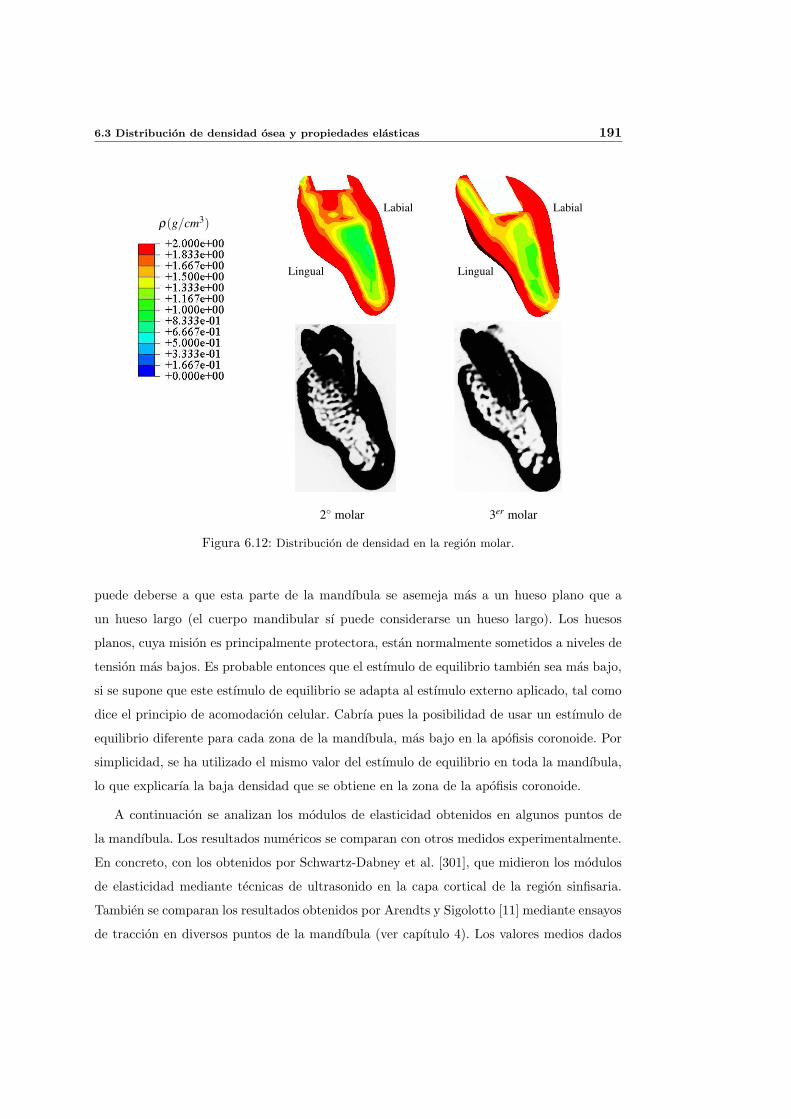
6.3 Distribucion de densidad osea y propiedades elasticas 191
ρ(g/cm3)
2 molar 3er molar
Lingual
Labial
Lingual
Labial
1
Figura 6.12: Distribucion de densidad en la region molar.
puede deberse a que esta parte de la mandıbula se asemeja mas a un hueso plano que a
un hueso largo (el cuerpo mandibular sı puede considerarse un hueso largo). Los huesos
planos, cuya mision es principalmente protectora, estan normalmente sometidos a niveles de
tension mas bajos. Es probable entonces que el estımulo de equilibrio tambien sea mas bajo,
si se supone que este estımulo de equilibrio se adapta al estımulo externo aplicado, tal como
dice el principio de acomodacion celular. Cabrıa pues la posibilidad de usar un estımulo de
equilibrio diferente para cada zona de la mandıbula, mas bajo en la apofisis coronoide. Por
simplicidad, se ha utilizado el mismo valor del estımulo de equilibrio en toda la mandıbula,
lo que explicarıa la baja densidad que se obtiene en la zona de la apofisis coronoide.
A continuacion se analizan los modulos de elasticidad obtenidos en algunos puntos de
la mandıbula. Los resultados numericos se comparan con otros medidos experimentalmente.
En concreto, con los obtenidos por Schwartz-Dabney et al. [301], que midieron los modulos
de elasticidad mediante tecnicas de ultrasonido en la capa cortical de la region sinfisaria.
Tambien se comparan los resultados obtenidos por Arendts y Sigolotto [11] mediante ensayos
de traccion en diversos puntos de la mandıbula (ver capıtulo 4). Los valores medios dados
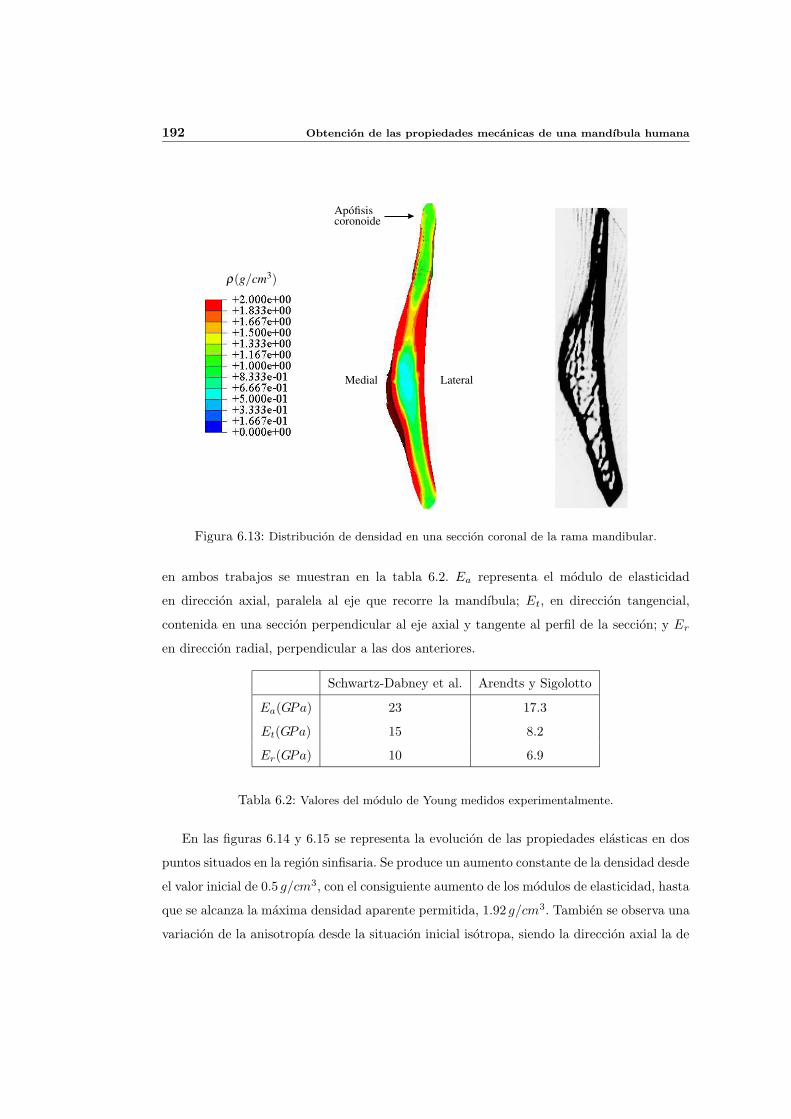
192 Obtencion de las propiedades mecanicas de una mandıbula humana
ρ(g/cm3)
Medial Lateral
Apofisiscoronoide
1
Figura 6.13: Distribucion de densidad en una seccion coronal de la rama mandibular.
en ambos trabajos se muestran en la tabla 6.2. Ea representa el modulo de elasticidad
en direccion axial, paralela al eje que recorre la mandıbula; Et, en direccion tangencial,
contenida en una seccion perpendicular al eje axial y tangente al perfil de la seccion; y Er
en direccion radial, perpendicular a las dos anteriores.
Schwartz-Dabney et al. Arendts y Sigolotto
Ea(GPa) 23 17.3
Et(GPa) 15 8.2
Er(GPa) 10 6.9
Tabla 6.2: Valores del modulo de Young medidos experimentalmente.
En las figuras 6.14 y 6.15 se representa la evolucion de las propiedades elasticas en dos
puntos situados en la region sinfisaria. Se produce un aumento constante de la densidad desde
el valor inicial de 0.5 g/cm3, con el consiguiente aumento de los modulos de elasticidad, hasta
que se alcanza la maxima densidad aparente permitida, 1.92 g/cm3. Tambien se observa una
variacion de la anisotropıa desde la situacion inicial isotropa, siendo la direccion axial la de
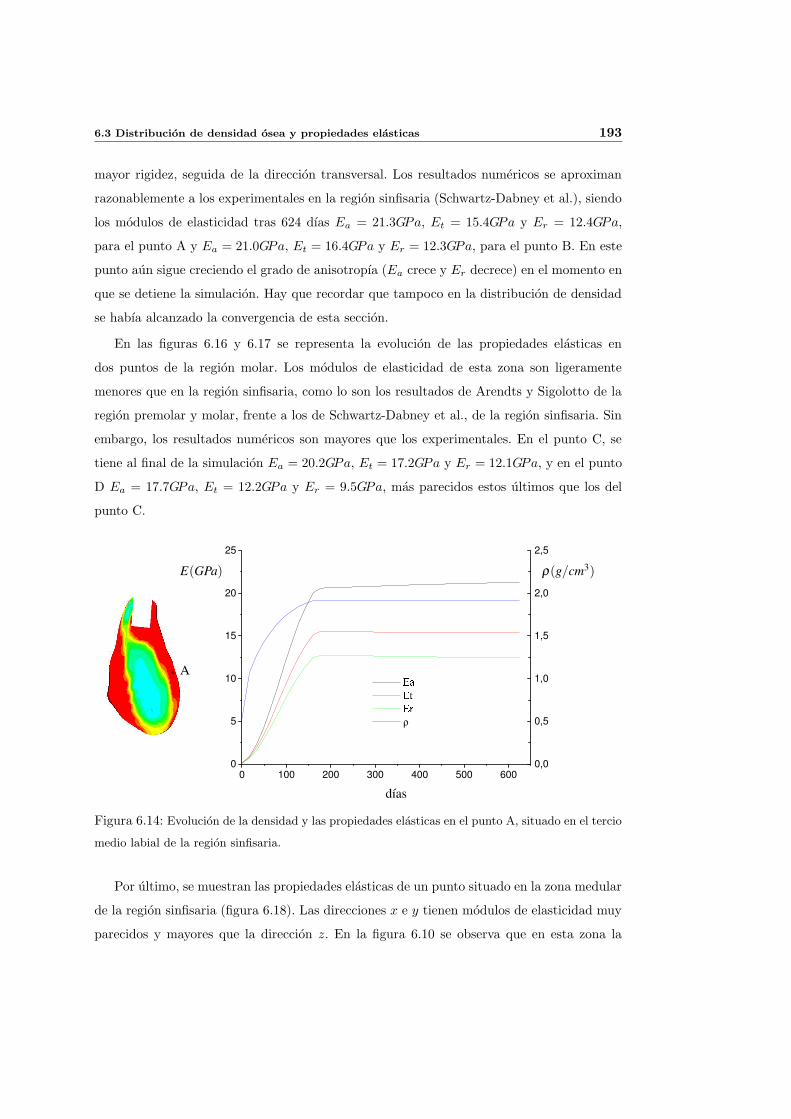
6.3 Distribucion de densidad osea y propiedades elasticas 193
mayor rigidez, seguida de la direccion transversal. Los resultados numericos se aproximan
razonablemente a los experimentales en la region sinfisaria (Schwartz-Dabney et al.), siendo
los modulos de elasticidad tras 624 dıas Ea = 21.3GPa, Et = 15.4GPa y Er = 12.4GPa,
para el punto A y Ea = 21.0GPa, Et = 16.4GPa y Er = 12.3GPa, para el punto B. En este
punto aun sigue creciendo el grado de anisotropıa (Ea crece y Er decrece) en el momento en
que se detiene la simulacion. Hay que recordar que tampoco en la distribucion de densidad
se habıa alcanzado la convergencia de esta seccion.
En las figuras 6.16 y 6.17 se representa la evolucion de las propiedades elasticas en
dos puntos de la region molar. Los modulos de elasticidad de esta zona son ligeramente
menores que en la region sinfisaria, como lo son los resultados de Arendts y Sigolotto de la
region premolar y molar, frente a los de Schwartz-Dabney et al., de la region sinfisaria. Sin
embargo, los resultados numericos son mayores que los experimentales. En el punto C, se
tiene al final de la simulacion Ea = 20.2GPa, Et = 17.2GPa y Er = 12.1GPa, y en el punto
D Ea = 17.7GPa, Et = 12.2GPa y Er = 9.5GPa, mas parecidos estos ultimos que los del
punto C.
0 100 200 300 400 500 6000
5
10
15
20
25
ρ
0,0
0,5
1,0
1,5
2,0
2,5
A
dıas
E(GPa) ρ(g/cm3)
1
Figura 6.14: Evolucion de la densidad y las propiedades elasticas en el punto A, situado en el tercio
medio labial de la region sinfisaria.
Por ultimo, se muestran las propiedades elasticas de un punto situado en la zona medular
de la region sinfisaria (figura 6.18). Las direcciones x e y tienen modulos de elasticidad muy
parecidos y mayores que la direccion z. En la figura 6.10 se observa que en esta zona la
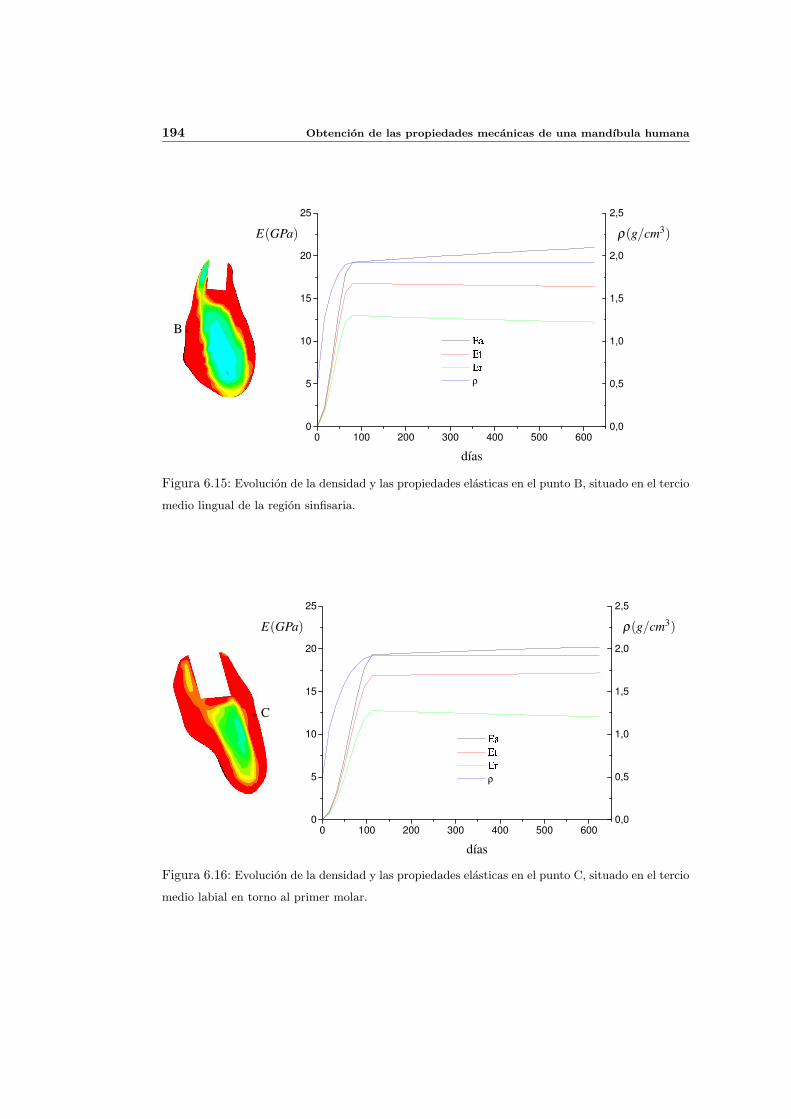
194 Obtencion de las propiedades mecanicas de una mandıbula humana
0 100 200 300 400 500 6000
5
10
15
20
25
ρ
0,0
0,5
1,0
1,5
2,0
2,5
B
dıas
E(GPa) ρ(g/cm3)
1
Figura 6.15: Evolucion de la densidad y las propiedades elasticas en el punto B, situado en el tercio
medio lingual de la region sinfisaria.
0 100 200 300 400 500 6000
5
10
15
20
25
ρ
0,0
0,5
1,0
1,5
2,0
2,5
C
dıas
E(GPa) ρ(g/cm3)
1
Figura 6.16: Evolucion de la densidad y las propiedades elasticas en el punto C, situado en el tercio
medio labial en torno al primer molar.
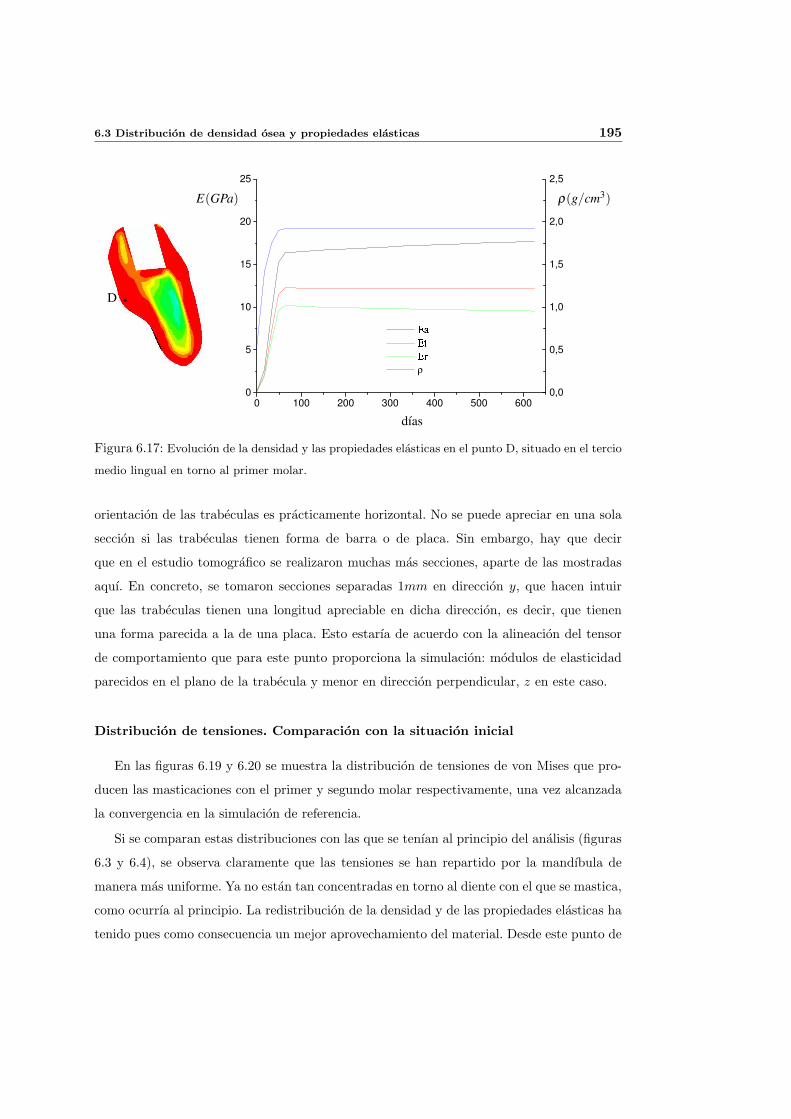
6.3 Distribucion de densidad osea y propiedades elasticas 195
0 100 200 300 400 500 6000
5
10
15
20
25
ρ
0,0
0,5
1,0
1,5
2,0
2,5
D
dıas
E(GPa) ρ(g/cm3)
1
Figura 6.17: Evolucion de la densidad y las propiedades elasticas en el punto D, situado en el tercio
medio lingual en torno al primer molar.
orientacion de las trabeculas es practicamente horizontal. No se puede apreciar en una sola
seccion si las trabeculas tienen forma de barra o de placa. Sin embargo, hay que decir
que en el estudio tomografico se realizaron muchas mas secciones, aparte de las mostradas
aquı. En concreto, se tomaron secciones separadas 1mm en direccion y, que hacen intuir
que las trabeculas tienen una longitud apreciable en dicha direccion, es decir, que tienen
una forma parecida a la de una placa. Esto estarıa de acuerdo con la alineacion del tensor
de comportamiento que para este punto proporciona la simulacion: modulos de elasticidad
parecidos en el plano de la trabecula y menor en direccion perpendicular, z en este caso.
Distribucion de tensiones. Comparacion con la situacion inicial
En las figuras 6.19 y 6.20 se muestra la distribucion de tensiones de von Mises que pro-
ducen las masticaciones con el primer y segundo molar respectivamente, una vez alcanzada
la convergencia en la simulacion de referencia.
Si se comparan estas distribuciones con las que se tenıan al principio del analisis (figuras
6.3 y 6.4), se observa claramente que las tensiones se han repartido por la mandıbula de
manera mas uniforme. Ya no estan tan concentradas en torno al diente con el que se mastica,
como ocurrıa al principio. La redistribucion de la densidad y de las propiedades elasticas ha
tenido pues como consecuencia un mejor aprovechamiento del material. Desde este punto de
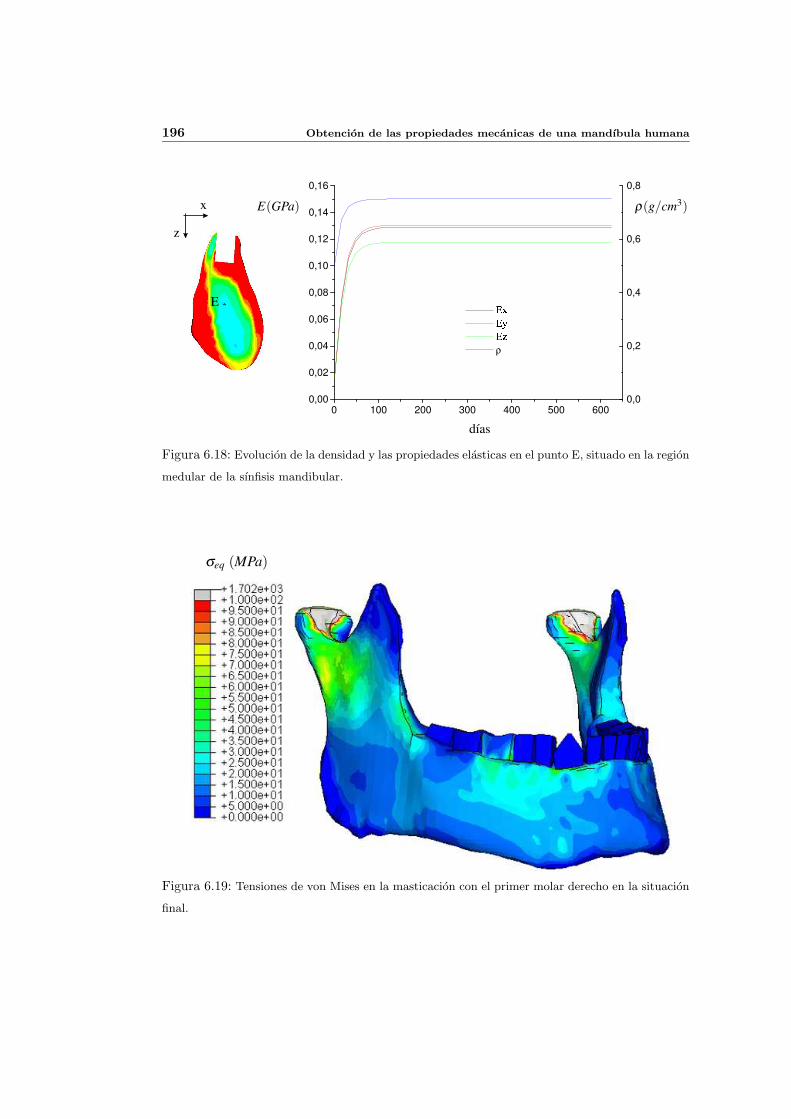
196 Obtencion de las propiedades mecanicas de una mandıbula humana
0 100 200 300 400 500 6000,00
0,02
0,04
0,06
0,08
0,10
0,12
0,14
0,16
ρ
0,0
0,2
0,4
0,6
0,8
E
dıas
E(GPa) ρ(g/cm3)x
z
1
Figura 6.18: Evolucion de la densidad y las propiedades elasticas en el punto E, situado en la region
medular de la sınfisis mandibular.
σeq (MPa)
1
Figura 6.19: Tensiones de von Mises en la masticacion con el primer molar derecho en la situacion
final.
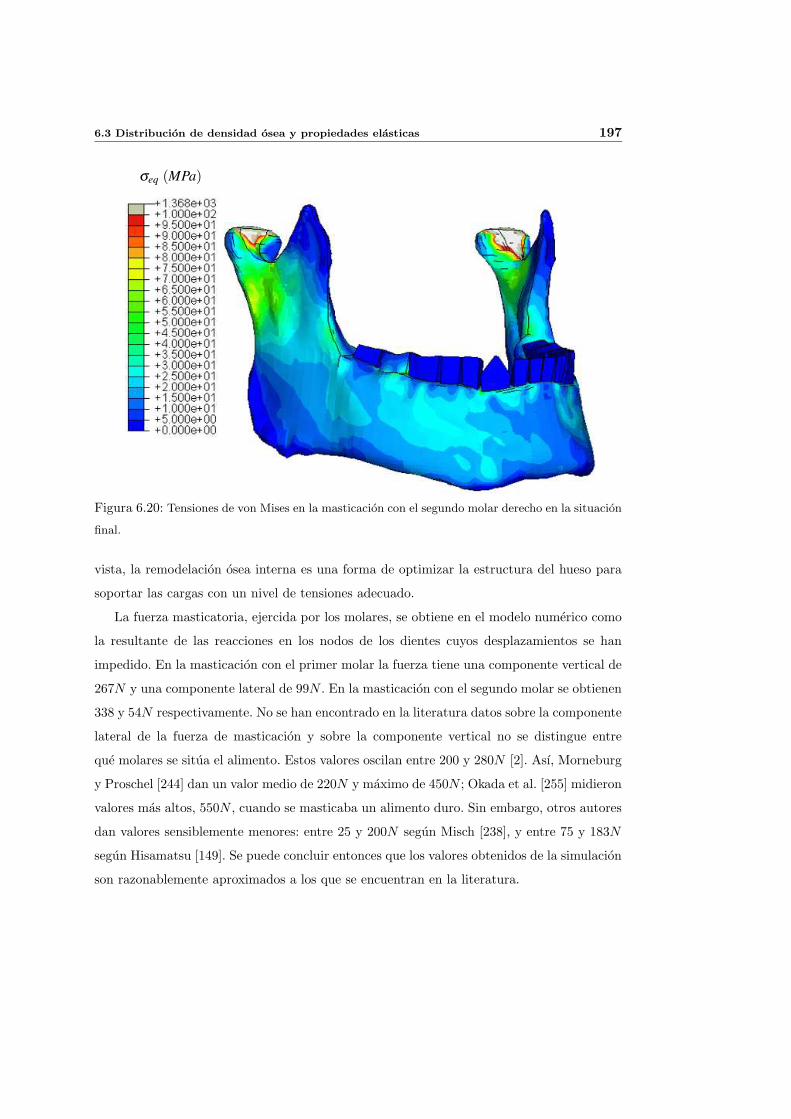
6.3 Distribucion de densidad osea y propiedades elasticas 197
σeq (MPa)
1
Figura 6.20: Tensiones de von Mises en la masticacion con el segundo molar derecho en la situacion
final.
vista, la remodelacion osea interna es una forma de optimizar la estructura del hueso para
soportar las cargas con un nivel de tensiones adecuado.
La fuerza masticatoria, ejercida por los molares, se obtiene en el modelo numerico como
la resultante de las reacciones en los nodos de los dientes cuyos desplazamientos se han
impedido. En la masticacion con el primer molar la fuerza tiene una componente vertical de
267N y una componente lateral de 99N . En la masticacion con el segundo molar se obtienen
338 y 54N respectivamente. No se han encontrado en la literatura datos sobre la componente
lateral de la fuerza de masticacion y sobre la componente vertical no se distingue entre
que molares se situa el alimento. Estos valores oscilan entre 200 y 280N [2]. Ası, Morneburg
y Proschel [244] dan un valor medio de 220N y maximo de 450N ; Okada et al. [255] midieron
valores mas altos, 550N , cuando se masticaba un alimento duro. Sin embargo, otros autores
dan valores sensiblemente menores: entre 25 y 200N segun Misch [238], y entre 75 y 183N
segun Hisamatsu [149]. Se puede concluir entonces que los valores obtenidos de la simulacion
son razonablemente aproximados a los que se encuentran en la literatura.

198 Obtencion de las propiedades mecanicas de una mandıbula humana
6.3.3. Influencia del patron de masticacion
Para estudiar la influencia de los habitos masticatorios se compara la simulacion de
referencia, en la que se usa la secuencia de masticacion S2 (MD1-MI1-MD2-MI2), (MD1-
MI1-MD2-MI2),. . . , con otras simulaciones en las que se usan las secuencias S1, que incluye
mordidas con los incisivos y los caninos (I-CD-CI-MD1-MI1-MD2-MI2-MD1-MI1-MD2-MI2-
MD1-MI1-MD2-MI2), (I-CD-CI-MD1-. . . ),. . . , y S3, que solo comprende masticaciones uni-
laterales con el lado derecho (MD1-MD2), (MD1-MD2), . . . . En todos los casos los paramet-
ros del modelo son los mismos que en la simulacion de referencia (ver tabla 6.1). Con la
secuencia S1 se detuvo el analisis despues de 624 dıas de actividad, como en la simulacion
de referencia y con la secuencia S3 tras 480 dıas, cuando se alcanza la convergencia.
En la figura 6.21 se muestra la distribucion de densidad aparente en un par de secciones,
obtenidas con las tres secuencias de masticacion. En general, la secuencia S1 produce resul-
tados muy parecidos a los de la simulacion de referencia, que usa la secuencia S2, aunque
con densidades ligeramente menores. En la region incisiva se observan leves diferencias: con
la secuencia S1 no se cierra la capa de cortical en la zona inferior, que sı lo hace con la
secuencia S2, aunque con un hueso cortical de densidad media; la capa de cortical obtenida
con la secuencia S1 es ligeramente mas fina; por ultimo, el hueso trabecular del nucleo de
la seccion es mas poroso si se utiliza la secuencia S1. De hecho, se puede apreciar que ha
habido incluso reabsorcion, ya que la densidad final es menor que 0.5 g/cm3, la densidad
de partida. Esto no ocurre con la secuencia S2. Con esta secuencia el hueso de esta zona
debe estar en zona muerta de remodelacion o proximo a ella, ya que no ha modificado su
densidad.
En la seccion molar tambien existen algunas diferencias en los resultados obtenidos con
ambas secuencias, principalmente en el tercio superior, en el que la densidad obtenida con
S1 es bastante menor. Tambien lo es el espesor de la capa cortical, aunque en este caso las
diferencias son menores. Finalmente, destaca en el caso de S1 la presencia de una zona de
hueso trabecular muy poroso, que con S2 no aparece.
Las diferencias entre los resultados de ambas secuencias, que pueden resumirse diciendo
que S1 conduce a densidades menores que S2, pueden explicarse a la vista de las tensiones
que producen las mordidas incisiva y canina, que son los unicos casos de carga en los que
se diferencian una y otra secuencia. En las figuras 6.22 y 6.23 se muestra la distribucion
de tensiones de von Mises en ambas mordidas una vez alcanzada la convergencia de las
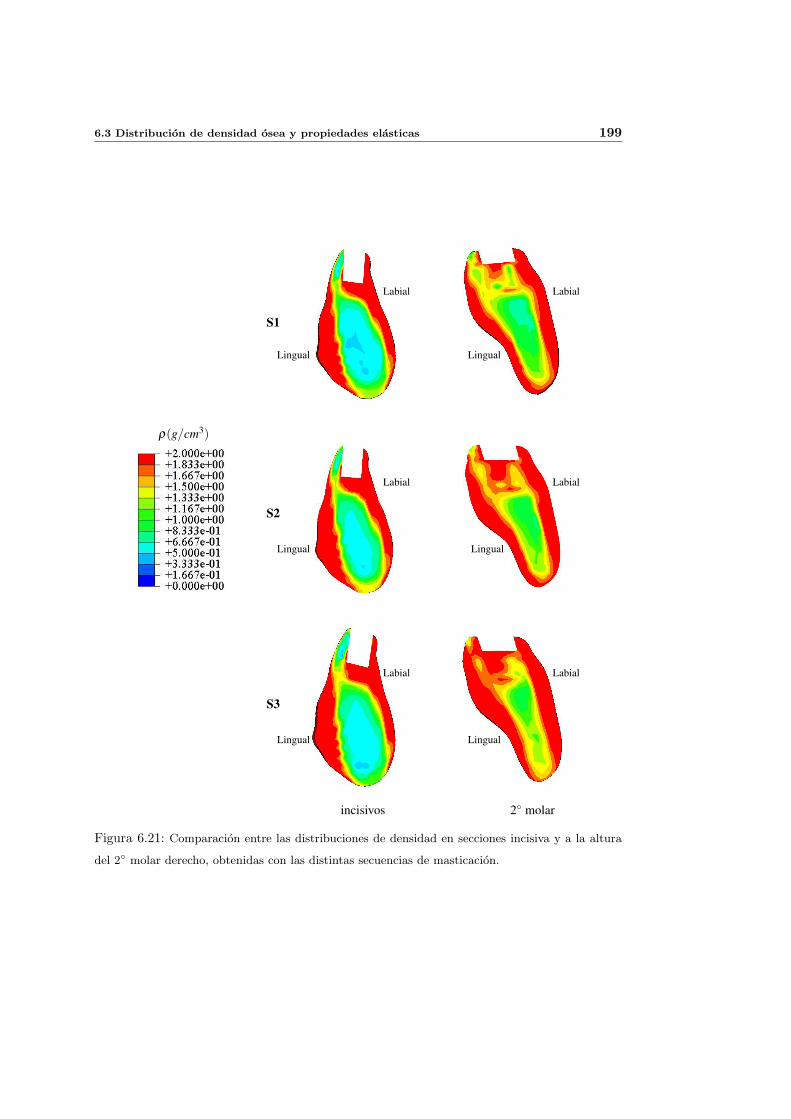
6.3 Distribucion de densidad osea y propiedades elasticas 199
ρ(g/cm3)
Lingual
Labial
Lingual
Labial
Lingual
Labial
Lingual
Labial
Lingual
Labial
Lingual
Labial
incisivos 2 molar
S1
S2
S3
1
Figura 6.21: Comparacion entre las distribuciones de densidad en secciones incisiva y a la altura
del 2 molar derecho, obtenidas con las distintas secuencias de masticacion.
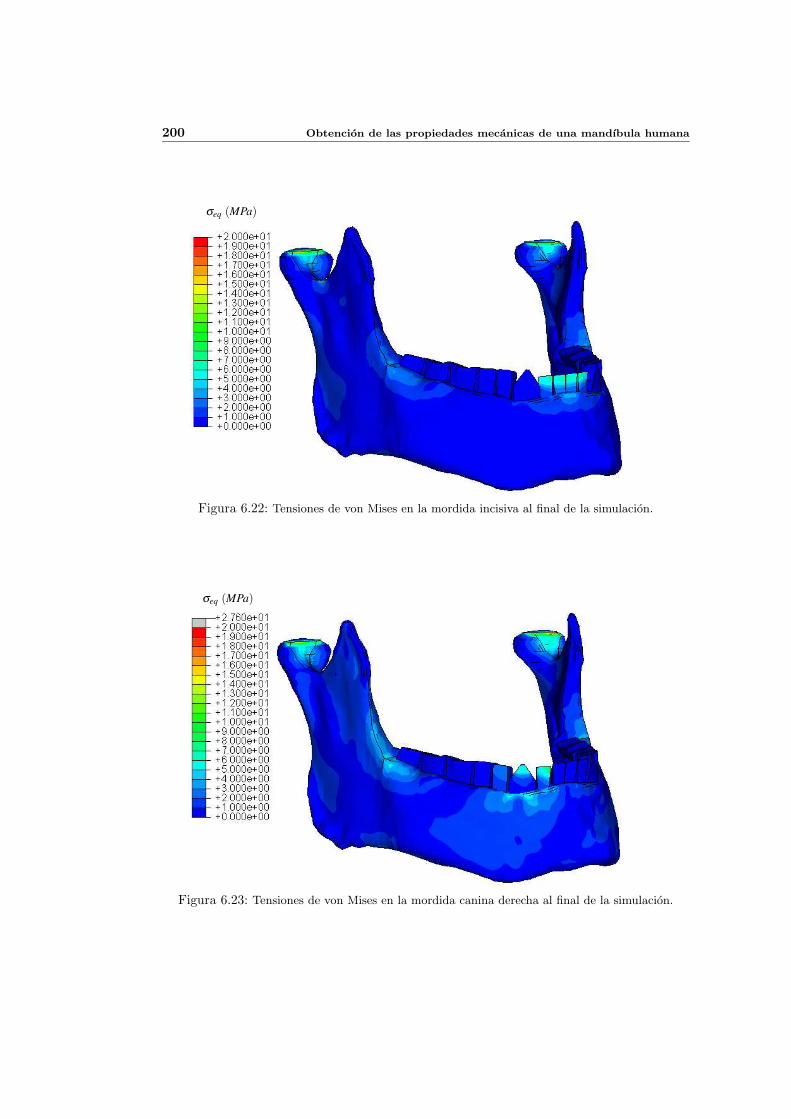
200 Obtencion de las propiedades mecanicas de una mandıbula humana
σeq (MPa)
1
Figura 6.22: Tensiones de von Mises en la mordida incisiva al final de la simulacion.
σeq (MPa)
1
Figura 6.23: Tensiones de von Mises en la mordida canina derecha al final de la simulacion.

6.3 Distribucion de densidad osea y propiedades elasticas 201
propiedades elasticas. Se observa nuevamente que las tensiones se uniformizan a nivel global
tras la remodelacion: con la redistribucion de la rigidez que esta conlleva bajan las tensiones
maximas y suben las tensiones mınimas. Pero es fundamental observar que las tensiones
siguen siendo muy pequenas. Este nivel tan bajo de tensiones no favorece la formacion
osea, mas bien al contrario. Ademas, al incluir los ciclos de mordida hay menos ciclos de
masticacion, que sı producen tensiones elevadas, con lo que el estımulo mecanico diario
desciende y con el la densidad aparente.
Las fuerzas verticales de mordida obtenidas en la simulacion son 150N en la mordida
incisiva y 192N en la mordida canina. Los datos encontrados en la literatura sobre fuerzas de
mordida son algo mayores. Para mordidas con los molares oscilan entre 220 y 2650N segun
Craig [77] o entre 100 y 730N , segun Helkimo [143]. Los datos anteriores no distinguen
con que dientes se produce la mordida y hay que tener en cuenta que con los molares se
aprieta mas fuerte que con los incisivos. Arnold [12] establece una relacion de fuerzas 4:2:1
si se muerde con molares, premolares o incisivos respectivamente. Aun ası, los resultados
numericos estan muy por debajo, lo que hace pensar que los datos de actividad muscular
usados aquı, y que se tomaron de la literatura, corresponden a mordidas moderadas, que por
otro lado son mas habituales. Se han encontrado datos de actividad muscular en aprietes
mas intensos, pero no se han incluido en la secuencia de masticacion por considerarlos casos
muy particulares y poco frecuentes en situaciones normales.
Mas diferencias pueden encontrarse entre las secuencias S2 y S3. En la seccion incisiva
no se cierra la capa de cortical en el tercio inferior y en la region molar la distribucion de
densidad es ligeramente diferente: mayor en la parte superior y con una capa de cortical mas
gruesa en la parte inferior, aunque de menor densidad.
Si estas diferencias son notables, mas lo son en el lado izquierdo, con el que no se mastica.
Las secuencias S1 y S2 son simetricas y por tanto tambien lo es la distribucion de densidad.
En la secuencia S3, sin embargo, solo se mastica con el lado derecho y eso se nota en el lado
izquierdo, que ha recibido un estımulo menor y tiene menor densidad, bastante menor como
se puede apreciar en la figura 6.24.
6.3.4. Influencia del parametro de anisotropıa ω
Hay que recordar que ω ponderaba la influencia de la componente desviadora de las
cargas (o del estımulo Y) en la remodelacion. Cuanto mayor es ω, mayor es la influencia de

202 Obtencion de las propiedades mecanicas de una mandıbula humana
ρ(g/cm3)
1
Figura 6.24: Distribucion de densidad osea con la secuencia S3.
dicha componente desviadora.
J =(1− ω)
3tr(Y)1 + ω dev(Y) =
(1− 2ω)
3tr(Y)1 + ωY (6.3)
Este parametro hay que elegirlo con cuidado. Supongase, por ejemplo, que se tiene un estımu-
lo Y elevado, en el que predomina la componente desviadora. Si se usa un valor pequeno de
ω, el estımulo ponderado J sera pequeno y la respuesta remodelatoria probablemente sea
reabsorcion, cuando el estımulo es alto. Parece claro que en ese caso hay que elegir un valor
de ω elevado y, por contra, un valor pequeno ante un estado de cargas cuasi-hidrostatico.
El analisis de sensibilidad del parametro ω se ha realizado usando para los demas
parametros el valor que tienen en la simulacion de referencia y la secuencia de masti-
cacion S2. Los valores de ω analizados son 0.18, 0.24 y 0.3. El valor utilizado por Garcıa y
Doblare [91, 115] en el femur es sensiblemente menor: 0.1.
En la figura 6.25 se muestra la distribucion de densidad en las secciones incisiva y a
la altura del segundo molar, para los tres valores de ω utilizados. La densidad es bastante
parecida en los tres casos, siendo mayor cuanto menor es ω. Con un valor de ω demasiado
grande se esta dando mas importancia de la que tiene a la componente desviadora del
estımulo y eso es precisamente lo que ocurre en la seccion incisiva, donde se dan las diferencias
mas notables. En esta region la componente desviadora del estımulo es menor que en la zona
molar por ser menor la torsion.
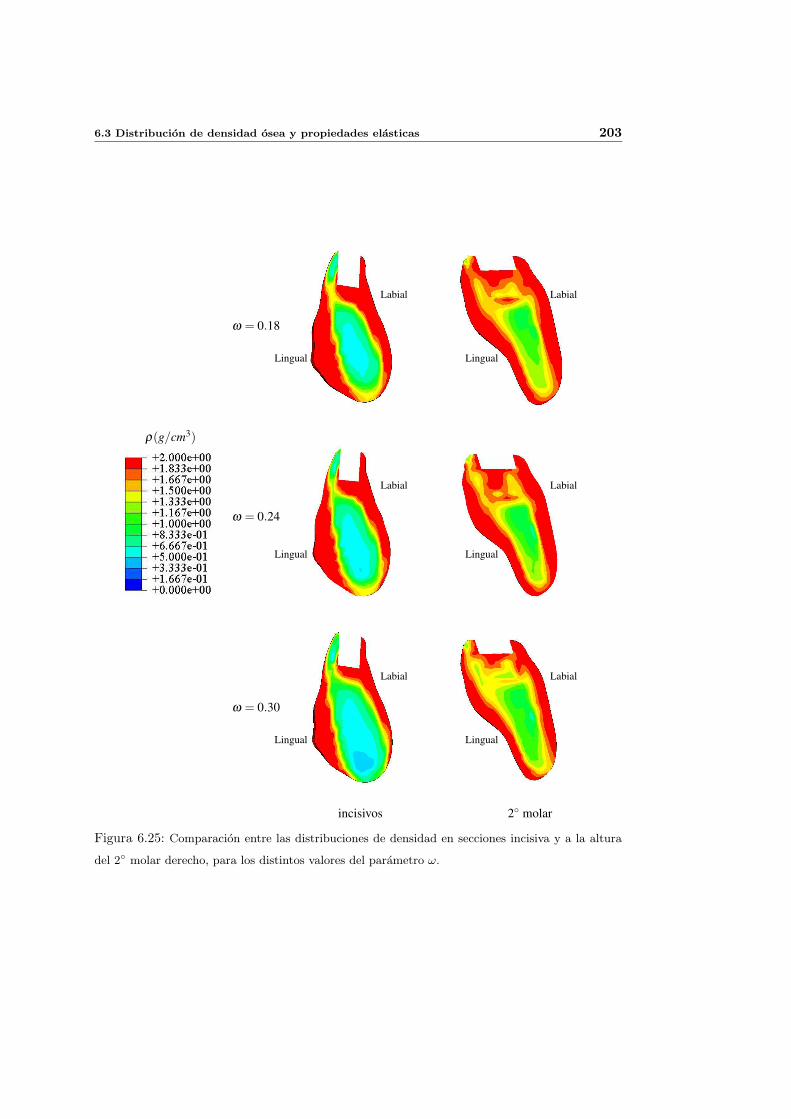
6.3 Distribucion de densidad osea y propiedades elasticas 203
ρ(g/cm3)
Lingual
Labial
Lingual
Labial
Lingual
Labial
Lingual
Labial
Lingual
Labial
Lingual
Labial
incisivos 2 molar
ω = 0.18
ω = 0.24
ω = 0.30
1
Figura 6.25: Comparacion entre las distribuciones de densidad en secciones incisiva y a la altura
del 2 molar derecho, para los distintos valores del parametro ω.

204 Obtencion de las propiedades mecanicas de una mandıbula humana
En cuanto a los modulos de elasticidad, la diferencia es mucho mayor, como se puede
observar en las figuras 6.26 y 6.27. Al aumentar ω aumenta el grado de anisotropıa de manera
notable. El modulo de Young intermedio, Et, apenas varıa, pero Ea aumenta y Er disminuye
en ambos casos. Si se comparan los resultados anteriores y los obtenidos en la simulacion de
referencia (figuras 6.14 y 6.16), con los resultados experimentales de la tabla 6.2, se observa
que el valor adoptado en dicha simulacion de referencia es el que conduce a resultados mas
ajustados a la realidad.
6.3.5. Influencia del estımulo de equilibrio
El estımulo de equilibrio, en torno al cual se halla la zona muerta, determina los valores
del estımulo que no producen ni formacion ni reabsorcion. Un estımulo de equilibrio muy alto
hace que sea necesario un estımulo elevado para que se produzca formacion y, por contra,
que con niveles normales de estımulo se produzca reabsorcion. Ello conduce a una densidad
osea muy baja. El otro caso extremo es que se utilice un estımulo de equilibrio demasiado
bajo. Con ello se esta favoreciendo la formacion osea y se obtiene una densidad excesiva. El
estımulo mas adecuado sera aquel que produzca una distribucion de densidad mas parecida
a la realidad.
Los tres valores de Ψ∗ analizados son 50, 25 y 10MPa/dıa. El semiancho de la zona
muerta es en todos los casos igual al 25 % del valor del estımulo de equilibrio. Los demas
parametros se mantienen constantes, en los valores usados para la simulacion de referencia.
En la figura 6.28 se compara la distribucion de densidad osea en sendas secciones incisiva
y a la altura del segundo molar derecho, en las simulaciones con los tres valores del estımulo
de equilibrio. Se observa que el valor utilizado por Doblare y Garcıa [91,115] para el femur,
50MPa/dıa, es demasiado alto para la mandıbula. En las dos secciones mostradas, la capa
de cortical es demasiado fina e incluso no se llega a cerrar por completo en la parte inferior.
El valor 10MPa/dıa, por el contrario, es demasiado bajo y conduce a una densidad excesiva.
Las propiedades elasticas del hueso resultaron muy similares en los tres casos. Por esa
razon y porque el caso Ψ∗ = 50MPa/dıa es en realidad la simulacion de referencia (descrita
de manera exhaustiva en la seccion 6.3.2), las propiedades elasticas no se muestran aquı. En
los tres casos se ha usado el mismo valor de ω, que es el parametro que tiene mas influencia
en las propiedades elasticas, y esa es la razon de dicha similitud, al menos en lo que a
propiedades elasticas se refiere.
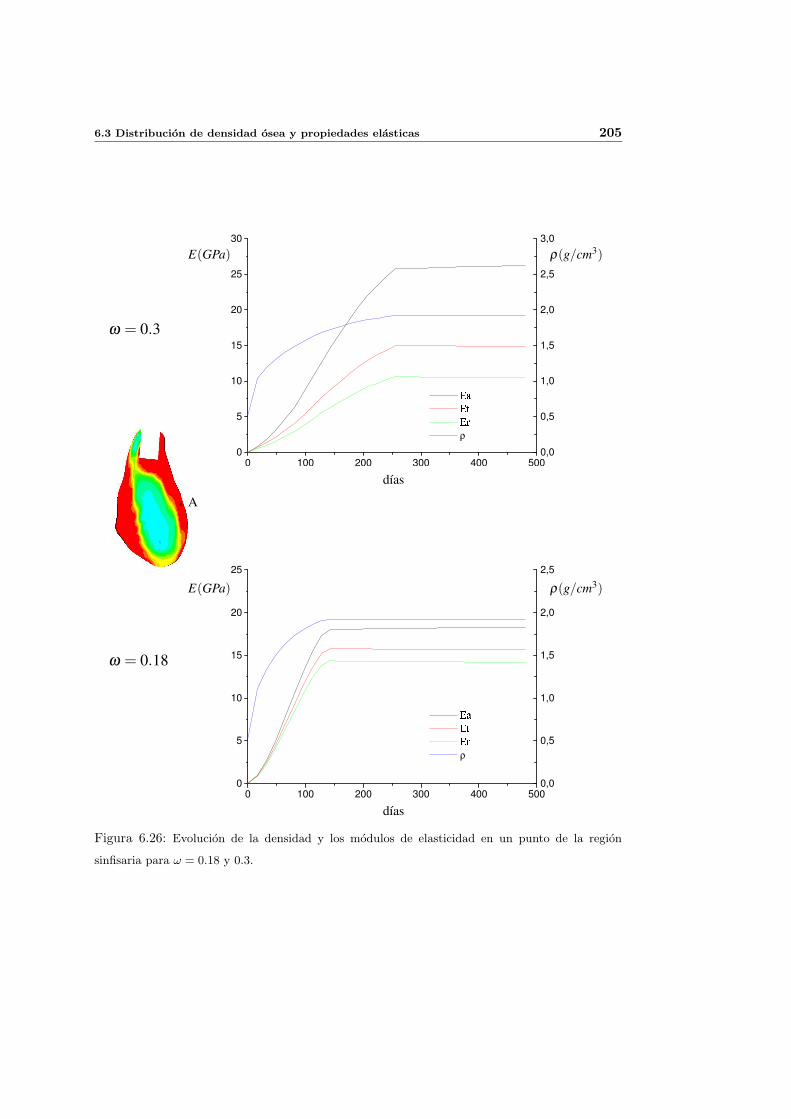
6.3 Distribucion de densidad osea y propiedades elasticas 205
0 100 200 300 400 5000
5
10
15
20
25
ρ
0,0
0,5
1,0
1,5
2,0
2,5
0 100 200 300 400 5000
5
10
15
20
25
30
ρ
0,0
0,5
1,0
1,5
2,0
2,5
3,0
A
dıas
E(GPa) ρ(g/cm3)
dıas
E(GPa) ρ(g/cm3)
ω = 0.3
ω = 0.18
1
Figura 6.26: Evolucion de la densidad y los modulos de elasticidad en un punto de la region
sinfisaria para ω = 0.18 y 0.3.
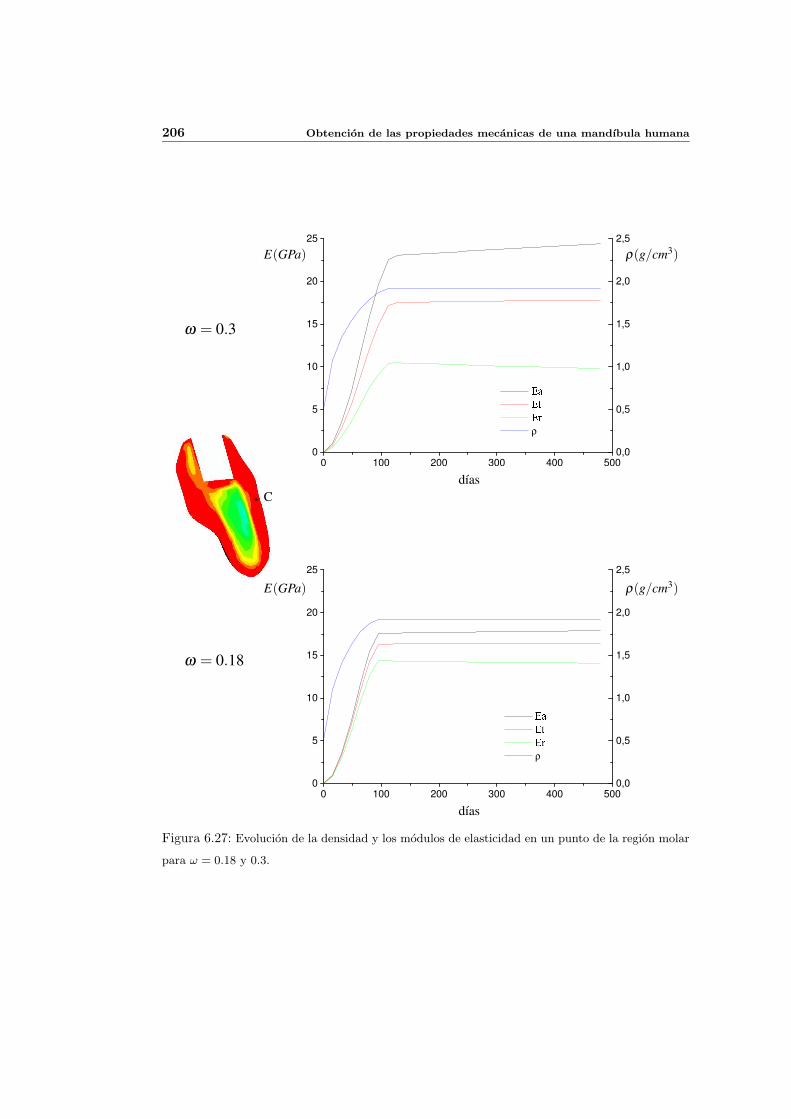
206 Obtencion de las propiedades mecanicas de una mandıbula humana
0 100 200 300 400 5000
5
10
15
20
25
! " #ρ
0,0
0,5
1,0
1,5
2,0
2,5
0 100 200 300 400 5000
5
10
15
20
25
$%$&$'ρ
0,0
0,5
1,0
1,5
2,0
2,5
C
dıas
E(GPa) ρ(g/cm3)
dıas
E(GPa) ρ(g/cm3)
ω = 0.3
ω = 0.18
1
Figura 6.27: Evolucion de la densidad y los modulos de elasticidad en un punto de la region molar
para ω = 0.18 y 0.3.
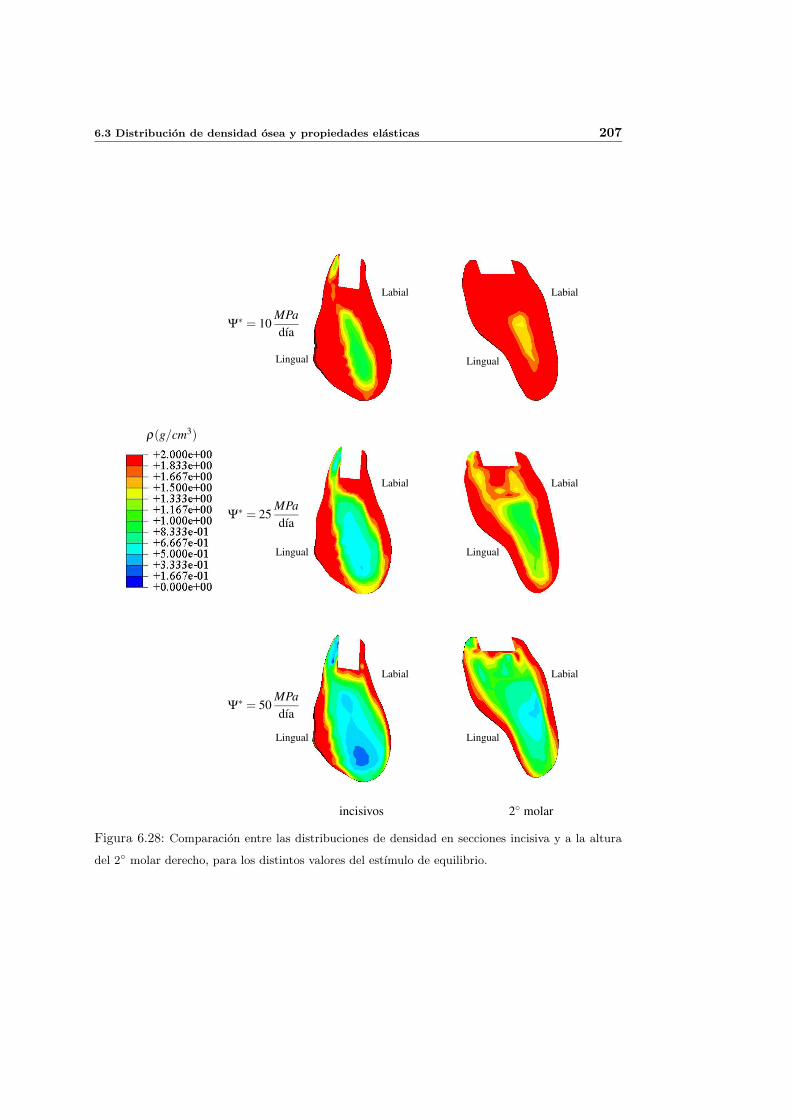
6.3 Distribucion de densidad osea y propiedades elasticas 207
ρ(g/cm3)
Lingual
Labial
Lingual
Labial
Lingual
Labial
Lingual
Labial
Lingual
Labial
Lingual
Labial
incisivos 2 molar
Ψ∗ = 10MPadıa
Ψ∗ = 25MPadıa
Ψ∗ = 50MPadıa
1
Figura 6.28: Comparacion entre las distribuciones de densidad en secciones incisiva y a la altura
del 2 molar derecho, para los distintos valores del estımulo de equilibrio.

208 Obtencion de las propiedades mecanicas de una mandıbula humana
6.3.6. Discusion de los resultados
La comparacion entre los resultados obtenidos con la simulacion numerica y las tomo-
grafıas realizadas a la mandıbula, corroboran que el modelo de remodelacion osea desarrol-
lado por Doblare y Garcıa es capaz de aproximar de manera razonable la distribucion de
densidad osea de la mandıbula y no solo del femur, como comprobaron estos autores. No
solo de la densidad, sino tambien de las propiedades elasticas, como se puede concluir de la
comparacion con los resultados encontrados en la literatura.
Para adaptar el modelo a la mandıbula, solo es necesario usar unos valores de los paramet-
ros del modelo distintos a los usados por Doblare y Garcıa para el femur. El estımulo de
equilibrio, Ψ∗, tiene gran influencia en la densidad, o lo que es lo mismo, en el espesor de la
capa de cortical, pero no ası en las propiedades elasticas. Sobre estas tiene mas influencia el
parametro ω, que determina el grado de anisotropıa del tejido y que, sin embargo, afecta de
manera muy leve a la densidad osea.
El valor de Ψ∗ = 50MPa/dıa usado para el femur es excesivo para la mandıbula, para la
que parece mas adecuado usar 10MPa/dıa. Hay que tener en cuenta que el femur esta someti-
do a cargas mas altas que las de masticacion. Igualmente, el valor de ω = 0.1 usado para
el femur, no resulta adecuado para la mandıbula, que necesita un valor considerablemente
mas alto (0.24 parece el mas adecuado) para reproducir la anisotropıa del tejido real. Ello
es debido a que las cargas de masticacion producen una torsion apreciable en la mandıbula,
que hace que la componente desviadora del estımulo sea alta, mayor que la existente en el
femur al caminar.
Se ha comprobado la convergencia de la densidad a nivel global, es decir, la convergencia
de la masa total de hueso. Esto no asegura la convergencia a nivel local. De hecho, existen
zonas de la mandıbula que siguen experimentando cambios en la densidad, especialmente
en la region mentoniana. Tampoco se puede asegurar que se haya alcanzado la convergencia
de las propiedades elasticas. En las graficas de la evolucion de las propiedades elasticas se
observa una tendencia convergente, aunque en algunos puntos dicha convergencia aun no se
haya alcanzado. A pesar de todo ello, se ha interrumpido la simulacion en un cierto punto.
En concreto, la simulacion de referencia, tras 624 dıas de masticacion. La primera razon
para ello es puramente practica: el tiempo de computacion que requiere el analisis de la
mandıbula completa es muy elevado. En segundo lugar, hay que tener en cuenta que el
objetivo perseguido en esta primera fase del analisis es establecer una base para el analisis

6.3 Distribucion de densidad osea y propiedades elasticas 209
del submodelo, en el que se ha incluido el implante. Es posible que las propiedades elasticas
del hueso incluido en dicho submodelo no hayan alcanzado la convergencia. Sin embargo,
sı ha quedado demostrado, que en el momento de interrumpir la simulacion, han convergido
los desplazamientos en la frontera de corte. Estos desplazamientos son los que se utilizan
en el analisis del submodelo y por tanto, se puede asegurar que dicho analisis no va a estar
afectado, de forma apreciable, por los cambios que sufran los elementos fuera del submodelo.
Ademas, dado que se continua la simulacion tras la inclusion del implante, los elementos
de hueso que permanecen en el submodelo pueden seguir cambiando sus propiedades hasta
alcanzar la convergencia.
Teniendo en cuenta lo anterior, es aceptable el uso de las propiedades elasticas de los
elementos del submodelo y de los desplazamientos de la frontera de corte como punto de
partida de la simulacion del submodelo. Ademas, es necesario suponer que los desplazamien-
tos en la frontera de corte no se ven afectados por la sustitucion del diente natural por un
implante. Esto no es cierto, pero, el no aceptar esta suposicion implica simular el implante
con la mandıbula completa, lo que resulta excesivamente costoso con los medios disponibles.


Capıtulo 7
Modelos mecanobiologicos
7.1. Introduccion
Los modelos descritos en el capıtulo 4 se suelen denominar fenomenologicos. Este califica-
tivo responde a una caracterıstica fundamental de los mismos: no consideran la naturaleza
intrınseca de los fenomenos biologicos que intervienen en la remodelacion osea. Su interes
se centra, por el contrario, en las consecuencias mecanicas que esos fenomenos tienen. Su
proposito, por tanto, es describir de forma matematica la relacion causa-efecto, o carga-
remodelacion si se quiere, de estos fenomenos, sin entrar en detalle en los procesos biologicos
involucrados.
En contraposicion a estos modelos, hace unos anos comenzaron a desarrollarse otros,
mas conocidos como mecanobiologicos, que intentan caracterizar matematicamente lo que
ocurre a nivel biologico, para poder explicar la remodelacion osea. Esta profundizacion en el
caracter biologico esta motivada por los resultados poco satisfactorios, que en determinadas
situaciones, proporcionan los modelos fenomenologicos.
Para aclarar la diferencia entre unos y otros puede servir un sencillo ejemplo de la In-
genierıa de Materiales, referente a la resistencia a fatiga de aceros templados. Un modelo
fenomenologico intentarıa relacionar, sin mas, el lımite de fatiga con la velocidad de tem-
ple mediante una ecuacion, que en todo caso podrıa ser corroborada experimentalmente.
Un modelo mecanico intentarıa ir mas alla en la explicacion del fenomeno. Relacionarıa,
en primer lugar, la velocidad de temple con el tamano de grano. A partir de este hecho,
explicarıa la relacion entre la resistencia a fatiga y la velocidad de temple, indicando que
211

212 Modelos mecanobiologicos
la resistencia a fatiga crece al disminuir el tamano de grano. Esto es ası porque los lımites
de grano actuan como barrera al progreso de las grietas. La conclusion final es que a mayor
velocidad de temple menor es el tamano de grano y, por tanto, mayor la resistencia a fatiga.
Mientras que el modelo fenomenologico se limita a ajustar una ecuacion entre las dos mag-
nitudes, el modelo mecanıstico, intenta explicar por que se produce esa relacion entre ellas
y si es posible cuantificar los fenomenos a ese nivel.
Mientras un modelo fenomenologico predice que el desuso conlleva perdida de masa osea;
una actividad por encima de lo habitual produce un aumento de masa osea, hasta un cierto
lımite, y un nivel de actividad normal mantiene el hueso dentro de unos parametros salud-
ables, el modelo mecanobiologico se pregunta por que ocurre ası y cuales son los mecanismos
biologicos que entran en juego, fundamentalmente la actividad de las BMUs, artıfices de la
remodelacion osea. Para ello investigan fenomenos tales como la generacion de BMUs, proce-
so de mineralizacion del tejido oseo, aparicion de microgrietas y reparacion de las mismas por
medio de la remodelacion, influencia de la presencia de estas microgrietas en la generacion de
nuevas BMUs, influencia de diversos parametros metabolicos, de la edad, de enfermedades
y tratamientos farmacologicos, etc.
7.2. Revision de algunos modelos de remodelacion mecanobi-
ologicos
7.2.1. Modelo de adaptacion osea de hueso esponjoso de Huiskes
et al.
En un tejido vivo como el hueso son las celulas las responsables de su mantenimiento y
de todos los cambios que en el se producen. Esa idea es la base del modelo desarrollado por
Huiskes et al. [157], uno de los primeros en cuantificar la actividad de las celulas oseas en la
remodelacion. Lo aplican unicamente a hueso trabecular, considerando que la remodelacion
ocurre en la superficie libre de los poros.
Los osteocitos tienen un papel fundamental en la transmision de senales dentro de la ma-
triz osea [194,296]. Estas celulas serıan capaces de “sentir” la deformacion y generar senales
consecuentemente. Huiskes et al. [157] las llegan a denominar incluso “galgas celulares”.
Dos procesos distintos como son el de remodelacion osea externa (bone modelling o

7.2 Revision de algunos modelos de remodelacion mecanobiologicos 213
external bone remodelling) y la remodelacion osea interna (bone remodelling o internal bone
remodelling) podrıan tener el mismo origen, el elevado nivel de tension-deformacion que
sentirıan los osteocitos. En el primer caso serıa producido por cargas externas elevadas y
conducirıa a un aumento de la masa osea. En la remodelacion, la formacion se producirıa
para rellenar el hueco que previamente han reabsorbido los osteoclastos. Este hueco actua
como concentrador de tensiones, elevando el nivel de deformaciones y por tanto reclutando
osteoblastos que rellenen el hueco. En cualquier caso los osteoblastos aparecen allı donde las
deformaciones son elevadas. El proceso de remodelacion propuesto parte de las siguientes
hipotesis:
1) La variable mecanica que controla el proceso es la velocidad de variacion de la densidad
de energıa de deformacion. En esta influyen la frecuencia y la amplitud de las cargas [330],
mientras que el numero de ciclos no es relevante, siendo suficiente con unos pocos ciclos para
el mantenimiento de la masa osea [296].
2) Los osteocitos reaccionan ante la carga produciendo una senal bioquımica que sirve
de estımulo para el reclutamiento de osteoblastos y que es proporcional a la velocidad de
variacion de la densidad de energıa de deformacion.
3) Dicha senal se transmite a traves de la red de osteocitos, amortiguandose con la
distancia recorrida, hasta que se recibe en la superficie osea, que es donde se produce el
reclutamiento de osteoblastos.
La intensidad de la senal, P (x, t), varıa con el tiempo y con la posicion dentro de la
superficie osea, x, y se expresa:
P (x, t) =
n∑
i=1
fi(x)µiRti(t) (7.1)
que como se observa es proporcional a la velocidad de variacion de la densidad de energıa
de deformacion, Rti, en la posicion ocupada por el osteocito i; a la sensibilidad mecanica
del osteocito, µi; a una funcion exponencial de amortiguamiento de la senal bioquımica, fi,
que depende de la distancia del osteocito i a la posicion de la superficie donde se reciba
esa senal, x. Ademas, la senal es dependiente tambien del numero de osteocitos proximos a
dicha posicion, n.
4) La reabsorcion ocurre de manera aleatoria segun dicte una funcion de densidad de
probabilidad p(x, t), que depende del mecanismo que active la reabsorcion. Segun una
primera hipotesis, la reabsorcion se producirıa como respuesta al dano microscopico acu-
mulado, para retirar el hueso danado y de esa forma mantener dicho dano dentro de unos

214 Modelos mecanobiologicos
lımites. Ese dano es consustancial con la carga y se distribuye de manera uniforme. Ocurre
ası, dado que las tensiones tambien se reparten de manera mas o menos uniforme, suponien-
do que la estructura osea es optima desde el punto de vista resistente. Por ello, la funcion
de densidad de probabilidad se supone uniforme en esta hipotesis.
Una segunda hipotesis establece que la reabsorcion ocurre por desuso. Esta ausencia
de carga serıa convertida en senal por los osteocitos y celulas de borde, que segregarıan
citokinas, las sustancias que activan los osteoclastos [61]. Es probable que ambas causas,
desuso y dano, concurran para activar la reabsorcion. De hecho, el dano produce un efecto
similar al desuso al romper la conectividad de la matriz osea. La funcion de densidad de
probabilidad es:
Hipotesis I p(x, t) = 10% uniformemente distribuida
Hipotesis II p(x, t) = c[a− P (x, t)] si P < a
p(x, t) = 0 si P ≥ a
(7.2)
donde c y a son constantes del modelo.
El cambio de densidad osea relativa, m (1.0 para hueso completamente mineralizado) se
expresa como:dm
dt= τ [P (x, t)− ktr]− roc si P (x, t) > ktr
dm
dt= −roc si P (x, t) ≤ ktr
(7.3)
donde roc es la cantidad diaria de hueso reabsorbido por los osteoclastos por unidad de
volumen, ktr es el valor mınimo de la senal necesaria para que se activen los osteoblastos y
τ es una constante de proporcionalidad. El parametro roc entra en juego con la probabilidad
dada por la ecuacion (7.2), mediante un generador de Monte Carlo.
Los autores aplicaron estas ideas a un modelo microscopico bidimensional de hueso, de
dimensiones 2 x 2 mm, sujeto a carga cıclica (2MPa a 1Hz). En la figura 7.1 se pueden
observar los resultados de las simulaciones. En color blanco se representa la medula y en
color negro el hueso. Encima de los cuadros aparecen los dıas de simulacion requeridos para
alcanzar la estructura correspondiente. Partiendo de dos estructuras trabeculares distintas,
A y B, llegaron a la misma estructura final, lo que demuestra la estabilidad de la formulacion.
Dicha estructura final corresponde a un estado homeostatico y esta alineada con la carga, lo
que concuerda con numerosas evidencias experimentales ya referidas en esta tesis. Partiendo
de dicha estructura homeostatica, en C se simula un cambio en la orientacion de la carga. Se

7.2 Revision de algunos modelos de remodelacion mecanobiologicos 215
observa como la estructura osea se adapta ligeramente para tratar de alinearse con la carga.
Por ultimo, en la simulacion D se parte de la situacion de equilibrio de las simulaciones A
y B y se aplica una estado de deformacion constante, que produce reabsorcion por desuso
segun la hipotesis II, siendo a = 1.6 y c = 12.5.
A
B C D
100 300 500 2000
500100030003000
Homeostasis
Homeostasis Homeostasis Homeostasis
Figura 7.1: Resultados de la simulacion. Adaptado de Huiskes et al. [157].
Las dos hipotesis hechas acerca de la reabsorcion conducen a los mismos resultados
pero con velocidades diferentes. Con la hipotesis II se alcanza mucho antes el equilibrio
homeostatico, ya que la reabsorcion ayuda a “esculpir” la forma externa del hueso, retirando
material de donde no es necesario por no estar cargado.
Frente a un aumento del nivel de carga una vez alcanzado el equilibrio, el modelo simula
bien el incremento de masa osea que se deriva. Tambien se simula con resultados razonables
la perdida de masa osea en individuos que bajan el nivel de actividad. Sin embargo, algo
que no comprobaron los autores es que en una situacion de desuso total, el modelo predice
la perdida total del hueso a largo plazo, algo que no ocurre en la realidad. A esta paradoja
la denominaron Fyhrie y Schaffler la “disuse fallacy” [113].
La necesidad de conocer la energıa de deformacion local en cada punto del modelo com-
plica enormemente su aplicabilidad ya que requiere el uso de modelos microscopicos, lo que lo
hace muy costoso desde el punto de vista computacional e inviable, con los medios actuales,
para simular huesos completos. De hecho, el modelo propuesto es bidimensional.
Se echa de menos una atencion mas exhaustiva al dano y sus consecuencias. Se supone

216 Modelos mecanobiologicos
que es una de las razones para la activacion de osteoclastos, pero no se estima su valor
en ningun momento. La probabilidad de que ocurra tal activacion es un valor constante e
independiente del nivel de dano real que acumule en el hueso, como consecuencia de que
haya mas o menos carga y mas o menos reparacion por remodelacion.
Con todo, la principal limitacion del modelo deriva de considerar la accion de los osteo-
clastos y los osteoblastos de forma conjunta (ecuacion (7.3)). No tiene en cuenta que ambos
procesos ocurren secuencialmente, primero la reabsorcion y despues la formacion (secuen-
cia ARF), ni que ese orden viene dado por la existencia de BMUs, que estos autores no
consideran.
7.2.2. Modelo mecanobiologico de BMUs de Hernandez
Este modelo fue desarrollado en la Universidad de Stanford por Hernandez, discıpulo del
profesor Carter [144]. En el sı se consideran las BMUs y la secuencia ARF, a diferencia del
modelo anterior. Ademas Hernandez lo aplica tanto a hueso trabecular como a cortical.
Partiendo del algoritmo de remodelacion isotropo de Stanford (ver capıtulo 5), Hernandez
propone algunas modificaciones para buscar sentido biologico a las expresiones que allı se
utilizan de forma puramente fenomenologica. En concreto, pretende sustituir la ecuacion de
remodelacion
ρ = krSvρ, (7.4)
por otra con mayor sentido biologico. Como se vio en el capıtulo 5, esta ecuacion proporciona
la velocidad de variacion de la densidad aparente, en funcion del volumen de matriz osea
que se deposita en la unidad de superficie por unidad de tiempo, r, de la densidad del tejido
depositado ρ, de la superficie que hay disponible para la remodelacion, Sv, y de un factor
biologico, k, que determina que proporcion de la superficie disponible es adecuada para
la remodelacion. Hernandez investiga mas en profundidad el fundamento biologico de esta
ecuacion y asigna un papel esencial en ella a la BMU.
Hernandez considera ademas que la densidad aparente no basta como variable inde-
pendiente para describir el tejido oseo y considera tres variables en su lugar: volumen de
poros, volumen de osteoide y volumen mineralizado. La conversion de un volumen en otro
es consecuencia de los procesos biologicos que tienen lugar en el tejido.
En el proceso de remodelacion osea, un determinado volumen de hueso mineralizado se
convierte en volumen de poros en la fase de reabsorcion y en la fase de formacion el hueco

7.2 Revision de algunos modelos de remodelacion mecanobiologicos 217
que deja la BMU se rellena de osteoide. Hernandez aproximo la forma del hueco primero
reabsorbido y despues formado por las BMUS. En el caso de las BMUs de hueso cortical
u osteonas, lo aproximo por un cilindro hueco de seccion circular de diametro interior dh y
diametro exterior do (figura 7.2). En el caso de las BMUs de hueso trabecular, denominadas
hemiosteonas por Parfitt [262], lo hizo por medio cilindro de seccion elıptica de semiejes de
y dBMU .
dBMU
de
1
dh
do
1
Figura 7.2: Forma simplificada del volumen remodelado por una BMU, en hueso trabecular (izquier-
da) y en hueso cortical (derecha).
La velocidad con que se remodela el hueso depende de la cantidad de BMUs que hay
activas en cada instante y de la duracion de los distintos perıodos de la secuencia ARF: TR,
perıodo de reabsorcion, TI , perıodo de inversion y TF , perıodo de formacion.
Para evaluar la poblacion de BMUs es necesario conocer la frecuencia de activacion
de BMUs. Esta depende de diversos factores metabolicos y mecanobiologicos, ası como de
la superficie especıfica de poros, Sv, cuya dependencia con la porosidad esta dada por la
correlacion de Martin [213].
Sv = 32.3n− 93.9n2 + 134n3 − 101n4 + 28.8n5 (7.5)
El modelo se aplica a un elemento diferencial en el que se suponen constantes los estımulos
metabolicos y mecanicos. De esta manera, se obvia la influencia del estado tensional tanto
en la activacion de BMUs como en su posterior actividad. Se trata pues de un modelo
puramente biologico, en el que la actividad de las BMUs es la unica responsable del cambio
de porosidad, siendo la velocidad de activacion de BMUs, NBMU , funcion de de Sv y de la

218 Modelos mecanobiologicos
frecuencia de activacion for, que se supone constante.
NBMU = Sv · for (7.6)
En la figura 7.3 se muestra el algoritmo del modelo de Hernandez.
BMUs en apoptosis
BMUs activadas, NBMU
BMUs progresando, NBMUPerıodo
reabsorcionPerıodo
inversionPerıodo
formacion
Numero de BMUsen reabsorcion
Numero de BMUsen formacion
Volumen de huesoreabsorbido
Volumen de huesoformado
Nueva porosidad
Superficie especıficade poros, Sv
for constante. No considera
influencia del estado mecanico
1
Figura 7.3: Algoritmo del modelo biologico de Hernandez [144].
El otro proceso biologico investigado por Hernandez es el de mineralizacion osea. En
este proceso, el osteoide se convierte en hueso mineralizado. El osteoide recien formado por
los osteoblastos esta compuesto principalmente de colageno y agua. Con el paso del tiempo
este osteoide se mineraliza y parte del agua es sustituida por cristales de hidroxiapatita. La
mineralizacion es muy rapida al principio, lentificandose despues conforme el tejido se va
saturando de mineral. De acuerdo con esto se suelen distinguir dos fases en el proceso de
mineralizacion: una fase primaria muy rapida, que dura unos pocos dıas [262] y en la que se
alcanza aproximadamente el 70 % del contenido maximo de mineral, y una fase secundaria
que puede durar anos y en la que la velocidad de deposicion de mineral decrece de forma
exponencial [262] (ver figura 7.4).
Partiendo de un volumen de hueso con unos valores de contenido en mineral (fraccion de
ceniza) y fraccion volumetrica de hueso determinados, y en el que no existen BMUs activas,
se analiza la evolucion temporal de los dos parametros anteriores cuando se empiezan a
activar BMUs hasta la convergencia.

7.2 Revision de algunos modelos de remodelacion mecanobiologicos 219
2 4 6
α0
αmax
t (anos)
fraccion deceniza ←− fase primaria
↑fase secundaria
Figura 7.4: Evolucion de la fraccion de ceniza propuesta por Hernandez. La fase primaria ocurre
de manera instantanea y en ella se alcanza el 70% del contenido de mineral maximo (αmax = 0.7).
La fase secundaria dura aproximadamente 6 anos y en ella el tejido se mineraliza del 70% a 95 %
del contenido maximo.
7.2.3. Modelo de BMUs con activacion por desuso y dano de
Hazelwood
Este discıpulo de Martin sı da un papel fundamental al estado tensional en la actividad
de las BMUs, a diferencia de Hernandez. De hecho, estas se activan por dos razones segun
Hazelwood et al. [140]: el desuso y el dano, o lo que es lo mismo, por tensiones demasiado
bajas o demasiado elevadas, respectivamente. En funcion de las tensiones locales, que se
pueden evaluar por medio del MEF, estima la frecuencia de activacion de BMUs y con
ella su poblacion. El efecto de las BMUs que se encuentren activas es doble: por un lado
modifican la porosidad y por otro reducen el dano que se pueda producir por fatiga ante un
estado de tensiones elevado. A continuacion se describe el modelo con mas detalle y se dan
las ecuaciones que cuantifican los conceptos recien introducidos.
La actividad de las BMUs produce cambios en la porosidad, que tienen en cuenta el
numero de BMUs que se encuentran en fase de reabsorcion, NR, y en fase de formacion, NF :
p = QRNR −QFNF (7.7)
donde QR = A/TR y QF = A/TF son respectivamente la velocidad de reabsorcion y de

220 Modelos mecanobiologicos
formacion, A es la seccion de la BMU,1 mientras que TR y TF son los perıodos (numero
de dıas) de reabsorcion y de formacion, que junto al perıodo de inversion TI completan la
secuencia ARF. De una serie de estudios histomorfometricos [32, 40, 41, 97, 107, 314, 316] se
toman los siguientes valores TR = 24, TF = 64 y TI = 8 dıas. La poblacion de osteonas NR
y NF se obtiene por integracion del historial reciente de la frecuencia de activacion, fa:
NR(t) =
∫ t
t−TR
fa(t′)dt′ (7.8a)
NF (t) =
∫ t−TR−TI
t−TR−TI−TF
fa(t′)dt′ (7.8b)
La frecuencia de activacion fa definida como el numero de BMUs que se activan por unidad
de area y por unidad de tiempo depende, como se dijo al principio, del nivel de dano y del
desuso:
fa = Sv(fa(desuso) + fa(dano)) (7.9)
ası como de la superficie especıfica de poros [213], como en tantos otros modelos. Los
parametros fa(desuso) y fa(dano) representan la contribucion a la frecuencia de activacion
de del desuso y del dano respectivamente.
El dano, d(mm/mm2), se define en este modelo como la longitud total de grietas que se
puede contabilizar en una seccion de hueso de area unitaria. El dano aumenta por fatiga y
disminuye como consecuencia de la remodelacion (disminuir el dano es precisamente uno de
los objetivos de la remodelacion [31,49,241]):
d = dF − dR (7.10)
El aumento de dano por fatiga, dF , se supone proporcional a un estımulo mecanico analogo
al definido por Carter (ver ecuacion (5.22))
dF = kDψ = kD
N∑
i=1
niεmi (7.11)
De hecho toma el mismo exponente m = 4 que obtienen los colaboradores de Carter [27,343],
pero no eleva el sumatorio a 1/m como hacen ellos.
Si se supone que las BMUs y el dano estan distribuidos uniformemente en el hueso, la
velocidad de disminucion de dano serıa d faA, ya que las BMUs en su avance se encontrarıan
1Parfitt [260] determino el area de una osteona A = 2.84 · 10−2mm2 y Hazelwood et al [140] toman el
mismo valor para hueso trabecular y cortical, lo que parece una simplificacion aparentemente innecesaria y
de todo punto incongruente, como muestra Hernandez [144].

7.2 Revision de algunos modelos de remodelacion mecanobiologicos 221
aleatoriamente con el tejido danado. Sin embargo, Hazelwood et al. [140] tomaron:
dR = d faAFs (7.12)
en la que introdujeron el factor Fs = 5 para tener en cuenta que la eficacia de la reparacion
del dano es mejor que la reparacion uniforme, ya que las BMUs se activan para reparar el
dano. No ocurre que lo encuentren aleatoriamente y entonces lo reparen, sino que se activan
y lo reparan principalmente donde mas dano se concentra.
La simulacion parte de un estado de equilibrio en el que se cumple dF = dR. Esto permite
estimar el parametro kD de la ecuacion (7.11)
kD =d0 fa0AFs
ψ0(7.13)
en el que el subındice 0 indica estado inicial o de equilibrio. Usando una longitud promedio
de grietas de 0.088mm [48] y la densidad promedio de grietas de un adulto de 40 anos
de los estudios de Schaffler et al. [300] se tiene d0 = 0.0366 mmmm2 . Hazelwood et al. toman
una frecuencia de activacion inicial fa0 = 0.00670 BMUsmm2 dıa basandose en diversos estudios
[32, 40, 41, 107, 316]. El estımulo de equilibrio lo estiman tomando n = 3000 (individuo que
camina diariamente 4.5 km a razon de 0.75 m por paso) y una deformacion de 500 µε,
que Beaupre et al. [27] consideran como deformacion de referencia para el hueso cortical, lo
que proporciona ψ0 = 1.875 · 10−10 µε·ciclosdıa . Sustituyendo en la ecuacion (7.13) se obtiene
kD = 1.85 · 105 mmmm2 .
El desuso se produce cuando ψ < ψ0, en cuyo caso se activan BMUs que reabsorben mas
hueso del que indica la ecuacion (7.7). Concretamente, se sustituye el area reabsorbida por
0.5A[1 + ψψ0
]. La frecuencia de activacion debida al dano se supone una funcion sigmoidal,
como la mayorıa de las respuestas a tratamientos farmacologicos.
fa(desuso) =fa(max)
1 + ekb(ψ−kc)si ψ < ψ0 (7.14)
donde kb = 6.5 · 1010 ciclosdıa y kc = ψ0
2 son constantes del modelo.
La frecuencia de activacion debida al dano es similar a la usada por Martin [214]
fa(dano) =fa(max) fa0
fa0 + (fa(max) − fa0)e[kr fa(max)
d−d0d0
] (7.15)
con la constante kr = −1.6 y la frecuencia de activacion maxima fa(max) = 0.50 BMUsmm2 dıa ,
que se toma considerable e intencionadamente mas alta que la de 0.14 BMUsmm2 dıa medida por
Frost [107] y que considera muy por debajo de un valor fisiologico normal.

222 Modelos mecanobiologicos
Las anteriores forman un conjunto de ecuaciones diferenciales que permiten calcular los
cambios que sufre la porosidad en funcion de las deformaciones a que esta sometido el hueso.
Para conocer dichas deformaciones es necesario resolver un problema elastico. El hueso se
supone un material elastico lineal pero con el modulo de Young variando de forma no lineal
con la porosidad, que tambien varıa de forma no lineal durante el proceso de remodelacion.
Para relacionar el modulo de Young y la porosidad se usa la siguiente correlacion:
E(MPa) =8.83 · 105 p6 − 2.99 · 109 p5 + 3.39 · 106 p4 − 2.64 · 106 p3 + 9.08 · 105 p2−
1.68 · 105 p+ 2.37 · 104(7.16)
ajustando un polinomio de grado 6 a los resultados experimentales de Currey [84] y Rho
et al. [282] que dan resultados de E frente a la densidad aparente, ρ, por lo que hay que
suponer una relacion p − ρ, lineal y de forma que para p = 0, ρ = 2.0 g/cm3 y para p = 1,
ρ = 0.
Modulo de Young, E Cargas y geometrıa
Deformaciones
Aumento de danopor fatiga, dF
Dano, dReparacion de
dano, dR
Frecuencia deactivacion, fa
Superficie especıficade poros, Sv
Porosidad , p
MEFdesuso
1
Figura 7.5: Algoritmo del modelo de Hazelwood. Traducido de [140]
El caso de carga simulado por Hazelwood et al. [140] es muy simple, compresion pura en
2-D, lo que constituye la principal limitacion de este modelo. Sin embargo, es pionero en usar

7.2 Revision de algunos modelos de remodelacion mecanobiologicos 223
algunas ideas y ecuaciones que luego son profusamente utilizadas en modelos posteriores.
7.2.4. Modelo de fallo y reparacion probabilista de Taylor
David Taylor y sus colaboradores han desarrollado una intensa labor de investigacion
sobre el dano en el tejido oseo, que ha dado lugar a numerosas publicaciones [319–322]. Han
aplicado la Mecanica de la Fractura Elastica Lineal para analizar el crecimiento de grietas
en propagacion y las condiciones necesarias para que la grieta se detenga en las barreras
que suponen las lıneas de cementacion entre osteonas [321]. En este mismo trabajo [321]
investiga sobre las tensiones y desplazamientos en la grieta y la influencia que pueden tener
estas tensiones y desplazamientos en los osteocitos proximos. Un nivel elevado de tensiones,
sobre todo tangenciales, puede conducir a la apoptosis (una modalidad de muerte celular)
de los mismos o a la interrupcion de las senales bioquımicas que transmiten y que podrıa ser
la causa de que se activara el proceso de remodelacion para reparar el dano. Ese proceso de
reparacion consiste en la renovacion del tejido danado y lo lleva a cabo la BMU que avanza
“cosiendo” las grietas. La forma en que varıa el factor de intensidad de tensiones con el
avance de la BMU ha sido objeto de otro de sus estudios [322].
En el modelo presentado a continuacion, Taylor parte de un modelo de dano probabilista
que predice la posibilidad de fallo por fatiga que puede sufrir un hueso sometido a carga
cıclica [319]. Para ello utiliza una funcion de densidad de probabilidad de Weibull, tıpico del
analisis de fiabilidad de materiales. Segun esta distribucion, la probabilidad de fallo de un
volumen de material Vs, sometido a una tension σ es:
Pf = 1− exp[− VsVso
( σσ∗
)α](7.17)
en la que Vso es un volumen de referencia, σ∗ es una constante relacionada con la resistencia
del material y α otra, relacionada con el grado de dispersion de la distribucion aleatoria.
Si aumenta σ, aumenta la probabilidad de fallo. Tambien lo hace si aumenta Vs, pues se
supone que el fallo comenzara en el lugar mas debil de la probeta y al aumentar su tamano
hay mayor probabilidad de encontrar zonas debiles.
Si el fallo considerado es por fatiga, se sustituye σ por el rango de tensiones ∆σ
Pf = 1− exp[− VsVso
(∆σ
∆σ∗
)α](7.18)
que se suele relacionar con el numero de ciclos hasta el fallo, nf , por expresiones del tipo:
nf =C
∆σn(7.19)
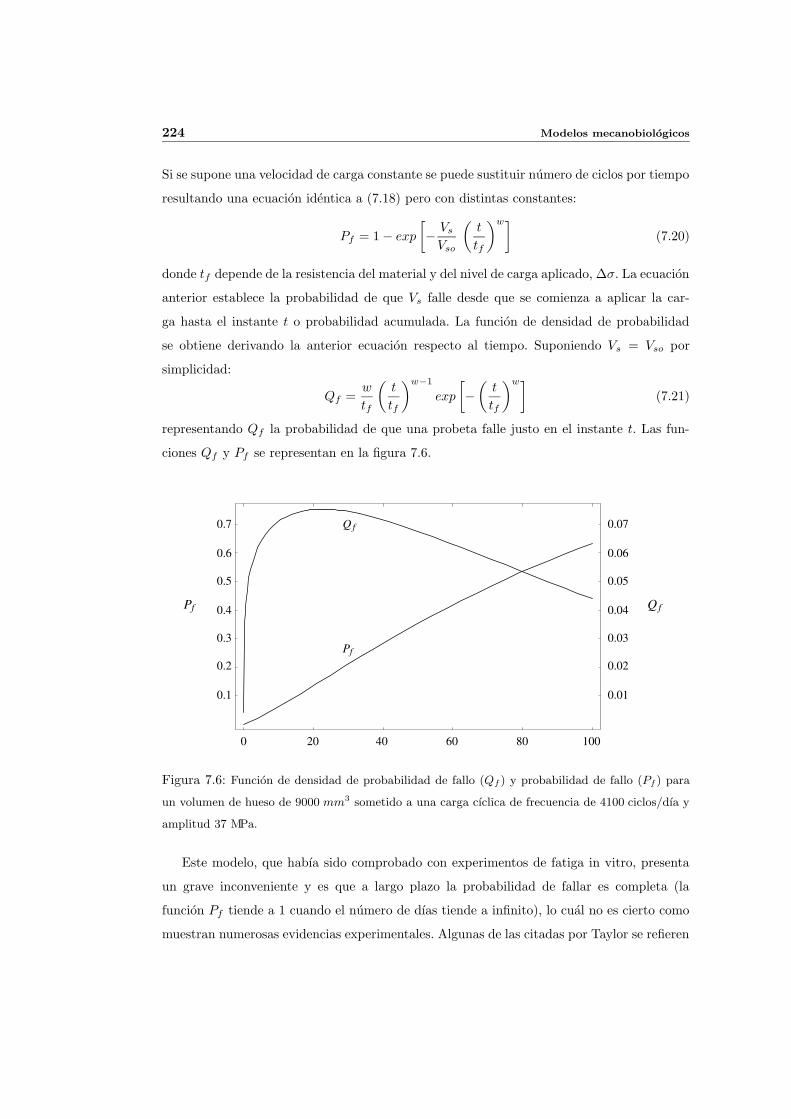
224 Modelos mecanobiologicos
Si se supone una velocidad de carga constante se puede sustituir numero de ciclos por tiempo
resultando una ecuacion identica a (7.18) pero con distintas constantes:
Pf = 1− exp[− VsVso
(t
tf
)w](7.20)
donde tf depende de la resistencia del material y del nivel de carga aplicado, ∆σ. La ecuacion
anterior establece la probabilidad de que Vs falle desde que se comienza a aplicar la car-
ga hasta el instante t o probabilidad acumulada. La funcion de densidad de probabilidad
se obtiene derivando la anterior ecuacion respecto al tiempo. Suponiendo Vs = Vso por
simplicidad:
Qf =w
tf
(t
tf
)w−1
exp
[−(t
tf
)w](7.21)
representando Qf la probabilidad de que una probeta falle justo en el instante t. Las fun-
ciones Qf y Pf se representan en la figura 7.6.
0 20 40 60 80 100
0.1
0.2
0.3
0.4
0.5
0.6
0.7
0.01
0.02
0.03
0.04
0.05
0.06
0.07
Pf
Q f
Pf Q f
1
Figura 7.6: Funcion de densidad de probabilidad de fallo (Qf ) y probabilidad de fallo (Pf ) para
un volumen de hueso de 9000 mm3 sometido a una carga cıclica de frecuencia de 4100 ciclos/dıa y
amplitud 37 MPa.
Este modelo, que habıa sido comprobado con experimentos de fatiga in vitro, presenta
un grave inconveniente y es que a largo plazo la probabilidad de fallar es completa (la
funcion Pf tiende a 1 cuando el numero de dıas tiende a infinito), lo cual no es cierto como
muestran numerosas evidencias experimentales. Algunas de las citadas por Taylor se refieren

7.2 Revision de algunos modelos de remodelacion mecanobiologicos 225
a deportistas y militares de reemplazo que sufren un notable incremento de su actividad fısica
en un momento determinado de su vida [224,235,257] y que padecen problemas oseos durante
la primera fase de la nueva actividad, disminuyendo posteriormente. Es mas que probable
que la causa de esa estabilizacion sea la reparacion del dano acumulado tras la sobrecarga.
Para contemplar esta posibilidad Taylor lleva a cabo una revision del modelo [320] que
incluye tanto la posibilidad de reparacion del dano por la accion de las BMUs, como el
aumento de la masa osea, factores ambos que redundan en un aumento de la resistencia de
la probeta. La mencionada revision del modelo propone que la reparacion de una grieta se
produce cuando una BMU atraviesa el plano de grieta disminuyendo su tamano, con lo que
baja drasticamente la probabilidad de fallo. Si la grieta es mas pequena que el diametro de
la BMU puede ocurrir que la grieta se repare por completo. Por el contrario, si la grieta
es mucho mayor que la BMU el efecto de este puede ser inutil y crecer la grieta hasta el
fallo antes de que otra BMU pueda reparar el resto de la grieta. La probabilidad de que una
BMU repare la grieta sigue igualmente una distribucion de Weibull, si bien independiente
del volumen considerado, porque en este caso no aumenta la posibilidad de encontrar puntos
de reparacion preferente.
Pr = 1− exp[−(t
tr
)v](7.22)
Qr =v
tr
(t
tr
)v−1
exp
[−(t
tr
)v](7.23)
Para Taylor no hay evidencia de que la velocidad de avance de la BMU dependa de la carga.
Segun el, la frecuencia de activacion de BMUs tambien es independiente de la carga, lo cual
es mas discutible, como se vera mas adelante. Supuesto cierto lo anterior, la probabilidad
de reparacion serıa independiente de la carga aplicada. No ası la probabilidad de fallo que
depende de la carga a traves de tf .
La reparacion y el fallo se suponen dos sucesos independientes. Ası, si Qf representa la
proporcion de probetas que fallan en el instante t, una parte de esas probetas es susceptible
de haber sido reparada, concretamente Pr, con lo cual, la nueva funcion de densidad de
probabilidad de fallo sera
Qfr = Qf (1− Pr) (7.24)
pudiendose obtener la probabilidad acumulada por integracion:
Pfr =
∫ t
0
Qfr(τ) dτ (7.25)
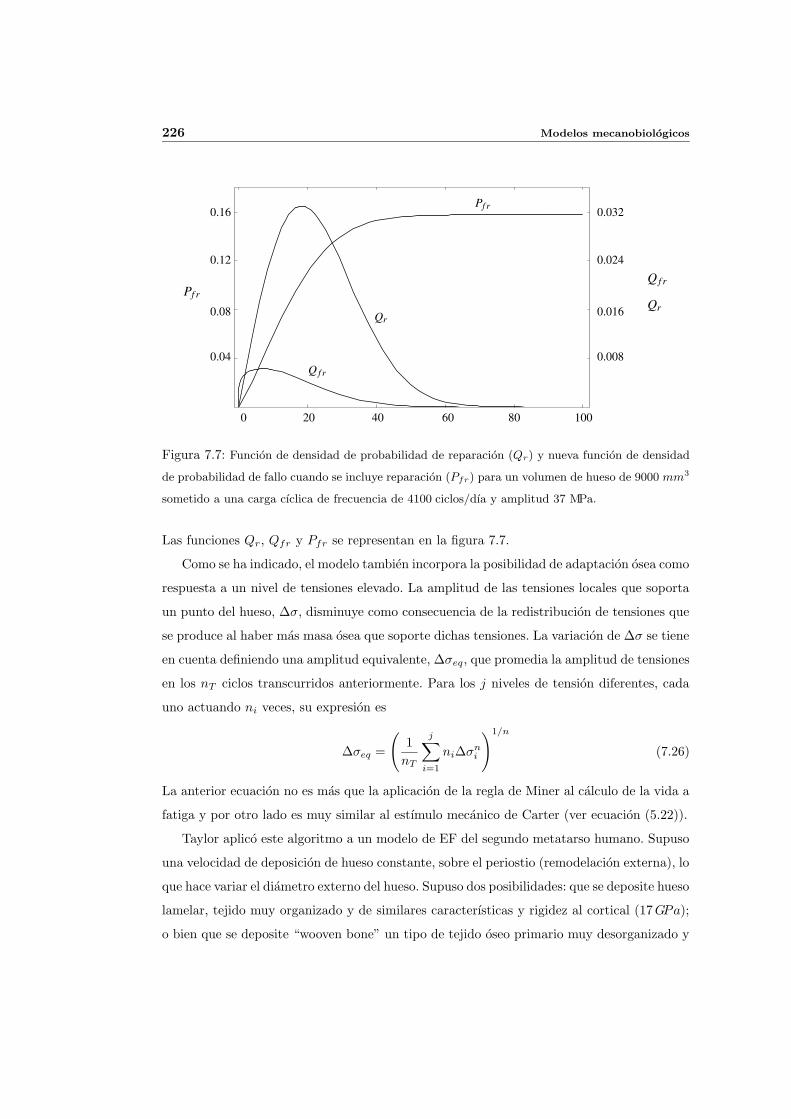
226 Modelos mecanobiologicos
0 20 40 60 80 100
0.04
0.08
0.12
0.16
0.008
0.016
0.024
0.032Pf r
Q f r
Qr
Pf r
Q f r
Qr
1
Figura 7.7: Funcion de densidad de probabilidad de reparacion (Qr) y nueva funcion de densidad
de probabilidad de fallo cuando se incluye reparacion (Pfr) para un volumen de hueso de 9000 mm3
sometido a una carga cıclica de frecuencia de 4100 ciclos/dıa y amplitud 37 MPa.
Las funciones Qr, Qfr y Pfr se representan en la figura 7.7.
Como se ha indicado, el modelo tambien incorpora la posibilidad de adaptacion osea como
respuesta a un nivel de tensiones elevado. La amplitud de las tensiones locales que soporta
un punto del hueso, ∆σ, disminuye como consecuencia de la redistribucion de tensiones que
se produce al haber mas masa osea que soporte dichas tensiones. La variacion de ∆σ se tiene
en cuenta definiendo una amplitud equivalente, ∆σeq, que promedia la amplitud de tensiones
en los nT ciclos transcurridos anteriormente. Para los j niveles de tension diferentes, cada
uno actuando ni veces, su expresion es
∆σeq =
(1
nT
j∑
i=1
ni∆σni
)1/n
(7.26)
La anterior ecuacion no es mas que la aplicacion de la regla de Miner al calculo de la vida a
fatiga y por otro lado es muy similar al estımulo mecanico de Carter (ver ecuacion (5.22)).
Taylor aplico este algoritmo a un modelo de EF del segundo metatarso humano. Supuso
una velocidad de deposicion de hueso constante, sobre el periostio (remodelacion externa), lo
que hace variar el diametro externo del hueso. Supuso dos posibilidades: que se deposite hueso
lamelar, tejido muy organizado y de similares caracterısticas y rigidez al cortical (17GPa);
o bien que se deposite “wooven bone” un tipo de tejido oseo primario muy desorganizado y

7.3 Modelo isotropo de remodelacion osea basado en la actividad de las BMUs 227
con peores propiedades (4.6GPa). El primero suele formarse mas lentamente que el segundo
que es caracterıstico de situaciones traumaticas, como las fracturas. En la simulacion se
suponen unas velocidades de deposicion de 4µm/dıa y 40µm/dıa respectivamente.
Con el modelo mecanico de elementos finitos es posible determinar ∆σeq, asociada a
la cual se puede definir una vida a fatiga equivalente. Con ella se calcula tf , que permite
calcular Qf con la ecuacion (7.21). Esta probabilidad se corrige segun (7.24) con Pr que es
independiente de la tension, para obtener Pfr, en realidad Pfra, pues se incluye el efecto del
hueso anadido.
En la simulacion realizada, la probabilidad de fallo aumenta con el tiempo hasta los 40
dıas, momento a partir del cual se mantiene constante, como se observa en la figura 7.7. Eso
es lo mismo que decir que la funcion de densidad de probabilidad de fallo es nula a partir de
40 dıas. Pasado ese perıodo el individuo ha podido reparar el dano acumulado inicialmente
sin que se produzca la fractura [224, 235, 257]. Se comprueba tambien que el efecto de la
reparacion es importante pero no lo es tanto el de la formacion osea, que apenas contribuye
a disminuir la probabilidad de fallo. Antes al contrario, la probabilidad de fallo aumenta si
lo que se deposita es “wooven bone”, pues las grietas crecen sobre este mas rapidamente que
sobre el hueso original.
7.3. Modelo isotropo de remodelacion osea basado en
la actividad de las BMUs
7.3.1. Introduccion
Desde el modelo de Huiskes, que comienza a abordar la necesidad de investigar el papel
de las celulas oseas en el proceso mecanobiologico de remodelacion, hasta el modelo de
Taylor, se han visto algunos conceptos interesantes. Conceptos que se aplican en el modelo
desarrollado en esta tesis y en el modelo isotropo en el que se basa, y que se expone en
el presente apartado. Hernandez [144] introdujo el concepto de BMU y secuencia ARF,
que Huiskes [157] no habıa tenido en cuenta. Hazelwood et al. [140] tambien consideraron
la influencia de las BMUs en los cambios de porosidad, pero no solo desde el punto de
vista biologico, como hizo Hernandez, que no incluyo el efecto que la carga tiene sobre la
activacion de las BMUs. Hazelwood et al. sı lo incluyeron, ademas del efecto que tambien

228 Modelos mecanobiologicos
tiene el microdano. Taylor se centro precisamente en este como la principal causa de la
activacion de las BMUs en casos de sobrecarga. No considero los casos de desuso. Huiskes
tambien tuvo en cuenta en su modelo la influencia del dano en la proliferacion de BMUs,
aunque de una forma algo superficial.
Con todo, ninguno de los modelos anteriores considera en sus simulaciones que el mi-
crodano cambia las propiedades elasticas del hueso, que no solo dependen de la porosidad.
Esto ultimo sı se considera en el modelo descrito a continuacion, que fue propuesto por
Garcıa-Aznar et al. [117]. Ademas, contempla el hecho observado experimentalmente de que
una situacion de desuso prolongado no produce la perdida total del hueso, sino que se al-
canza antes una situacion de equilibrio. Esto no lo hacen los otros modelos, que predirıan la
perdida indefinida de hueso, por suponer constante el estımulo mecanico de equilibrio.
Antes de entrar a describir el modelo conviene recordar algunos parametros definidos en
el capıtulo 4 acerca de la composicion volumetrica del tejido oseo. En una muestra de hueso
de volumen Vt existen huecos o poros, que ocupan un volumen Vv y tejido que ocupa el Vb
restante. Esta matriz de tejido oseo tiene una cierta cantidad de tejido danado Vd, que se
supone uniformemente repartido dentro del volumen de tejido. El resto es tejido intacto, que
comprende un componente inorganico Vm, principalmente hidroxiapatita, y un componente
organico Vo, colageno en su mayorıa.
Vt = Vb + Vv = Vm + Vo + Vd + Vv (7.27)
La fraccion volumetrica de hueso, vb, se define como:
vb =VbVt
(7.28)
Aparte de vb, las variables independientes del modelo isotropo de BMUs son el grado de
mineralizacion y el nivel de dano. Para medir la primera se utiliza la fraccion de ceniza, α,
definida en el capıtulo 4 como
α =mm
mm +mo(7.29)
El nivel de dano puede entenderse de dos formas. En primer lugar como una medida de
la degradacion de las propiedades mecanicas, fundamentalmente la rigidez, o bien como una
medida de la densidad de microgrietas dentro del material. Para la primera interpretacion
se define la variable degradacion, d, definida como:
d = 1− E
E0(7.30)

7.3 Modelo isotropo de remodelacion osea basado en la actividad de las BMUs 229
en la que E es el modulo de Young del material danado y E0 el del material virgen [49, 85,
175,265,360]. Esta es la forma usual de definir el dano en la Mecanica del Dano [200,307].
Para evaluar la densidad de microgrietas se define la variable h, densidad de dano, como:
h =VdVt
(7.31)
La relacion entre h y d puede depender de muchos factores como el estado de carga,
la porosidad, el tipo de hueso, etc. Actualmente no existe una correlacion experimental de
caracter general entre ambas magnitudes, tan solo correlaciones para casos particulares,
como el desarrollado por Burr et al. [49]. Estos autores observaron una relacion lineal entre
la perdida de rigidez y el area de grietas en ensayos de flexion realizados a femures de perro
inicialmente sin dano. Consecuentemente, en este modelo isotropo de BMUs se supone una
relacion lineal, del tipo:
h = k d (7.32)
donde k es una constante a determinar experimentalmente.
Es equivalente tener como variable independiente a la fraccion volumetrica de hueso, que
a la densidad aparente, como ocurre en la mayorıa de modelos anteriores, ya que ambas
variables estan relacionadas:
ρ = ρt (vb − h) (7.33)
donde ρt es la densidad del tejido oseo, que depende del grado de mineralizacion. Hernandez
et al. [144, 145] encontraron una relacion lineal entre ρt y α que reproduce con razonable
aproximacion los resultados experimentales
ρt(g/cm3) = 1.41 + 1.29α (7.34)
Estos autores comprobaron experimentalmente que las propiedades elasticas del hueso pre-
sentan una mejor correlacion con la fraccion volumetrica del hueso y la fraccion de ceniza, que
con la densidad aparente. Esa es la razon por la que se elige vb como variable independiente
en lugar de ρ. La correlacion que encontraron entre E y vb es la siguiente:
E(MPa) = 84370 v2.58b α2.74 (7.35)
que se usa en este modelo como alternativa a la ecuacion (5.36) dada por Beaupre [26] y
empleada en modelos anteriores. El coeficiente de Poisson se considera constante (ν = 0.3),
hipotesis bastante razonable ya que su variacion es poco importante [220].

230 Modelos mecanobiologicos
Una vez definida la variable fundamental del modelo, vb, y su relacion con las propiedades
elasticas, se describe a continuacion como se modifica dicha variable por el proceso de re-
modelacion osea.
7.3.2. Descripcion matematica de la actividad de las BMUs
Como ya se indico en el capıtulo 4, las BMUs (basic multicellular units) son asocia-
ciones de celulas, osteoclastos y osteoblastos que trabajan de forma conjunta reabsorbiendo
el hueso antiguo y generando tejido nuevo. Ambos procesos tienen lugar continuamente, in-
cluso en una situacion de equilibrio de remodelacion. La velocidad de cambio de la fraccion
volumetrica de hueso, vb, vendra dada por el balance de materia siguiente:
vb = vF − vR (7.36)
donde vF y vR son, respectivamente, las velocidades de cambio de volumen formado y re-
absorbido por unidad de volumen. En la situacion de equilibrio de remodelacion ambas son
iguales y por tanto la cantidad de tejido no se modifica. Aunque no cambie la cantidad, el
tejido sı se modifica, se regenera. El objetivo de esta regeneracion es disminuir el nivel de
dano y regular el contenido de mineral.
Estos cambios de volumen de tejido dependen del numero de BMUs por unidad de
volumen que se encuentren activas en cada instante, parametro que se denomina densidad
de BMUs, NBMU . Una vez activada, la BMU recorre una distancia LBMU = σL vBMU . La
variable σL representa la vida de una BMU, es decir, el tiempo que permanecen activos
los primeros osteoclastos generados; mientras que vBMU es la velocidad de progresion de la
BMU. Tanto vF como vR pueden expresarse de forma generica como:
v(t′) =
∫ LBMU
0
(∫ t′
t′−σL
NBMU(t′′)dt′′
)A(x)dx (7.37)
donde x es la direccion de progreso de la BMU, A(x) es la velocidad de variacion de volumen
por unidad de longitud en la posicion x debida a una BMU y el factor entre parentesis
proporciona el numero de BMUs que se encuentran activas en el instante t′.
A(x) es una variable que mide la velocidad a la que se reabsorbe o forma hueso en la
posicion x de la BMU. Toma valores distintos en las zonas de reabsorcion y formacion y se
supone, por simplicidad, que dentro de cada zona toma un valor constante, independiente

7.3 Modelo isotropo de remodelacion osea basado en la actividad de las BMUs 231
de la variable x:
A(x) =
ABMU
TR
fc en la zona de reabsorcion
0 en la zona de inversion
ABMU
TF
fb en la zona de formacion
(7.38)
en la que TR y TF representan el tiempo que dura, en una seccion, la fase de reabsorcion y
formacion, respectivamente. Los parametros fb y fc son dos variables con rango de variacion
[0, 1] que representan el nivel de actividad de osteoblastos y osteoclastos, respectivamente.
Por ultimo, ABMU es el area de la seccion de una BMU. Si se aproxima la geometrıa de las
BMUs de la misma forma que lo hace Hernandez (ver figura 7.2):
ABMU =
(d2o
4− d2
h
4
)π en hueso cortical
π
4de dBMU en hueso trabecular
(7.39)
Combinando las ecuaciones (7.37), (7.38) y (7.39) y haciendo en la integral el cambio de
variables dx = vBMUdt:
vF (t) =
∫ t−TR−TI
t−TR−TI−TF
(∫ t′
t′−σL
NBMU(t′′)dt′′
)ABMU
TF
fb(t′) vBMUdt
′ (7.40a)
vR(t) =
∫ t
t−TR
(∫ t′
t′−σL
NBMU(t′′)dt′′
)ABMU
TR
fc(t′) vBMUdt
′ (7.40b)
donde TI es el tiempo que dura la fase de inversion de la secuencia ARF. Las ecuaciones
(7.40) describen los cambios de la fraccion volumetrica de hueso en el instante t en funcion
de los cambios de densidad de BMUs ocurridos recientemente.
Otro factor que controla la cantidad de hueso remodelado es el balance de la actividad de
osteoclastos y osteoblastos, fbb = (fb/fc), conocido como balance local de hueso (focal bone
balance) [144]. Dicho parametro depende del estımulo mecanico ξ [91, 157, 167, 294] y de la
edad [56,220]. Entre fbb y ξ se establece una relacion lineal a trozos (figura 7.8), analoga a la
existente entre la velocidad de remodelacion y el estımulo en el modelo isotropo de Stanford
(recordar figura 5.10). La dependencia con la edad es difıcil de establecer. Lo que proponen
Garcıa-Aznar [117] es considerar que el hueso de una persona adulta tiene un balance local
de hueso neutro, es decir, fbb,max = fbb,min = 1.0, mientras que en un hueso en crecimiento
o en un hueso viejo fbb,max > 1 y fbb,min < 1 pudiendo producirse cambios significativos en
la masa osea.
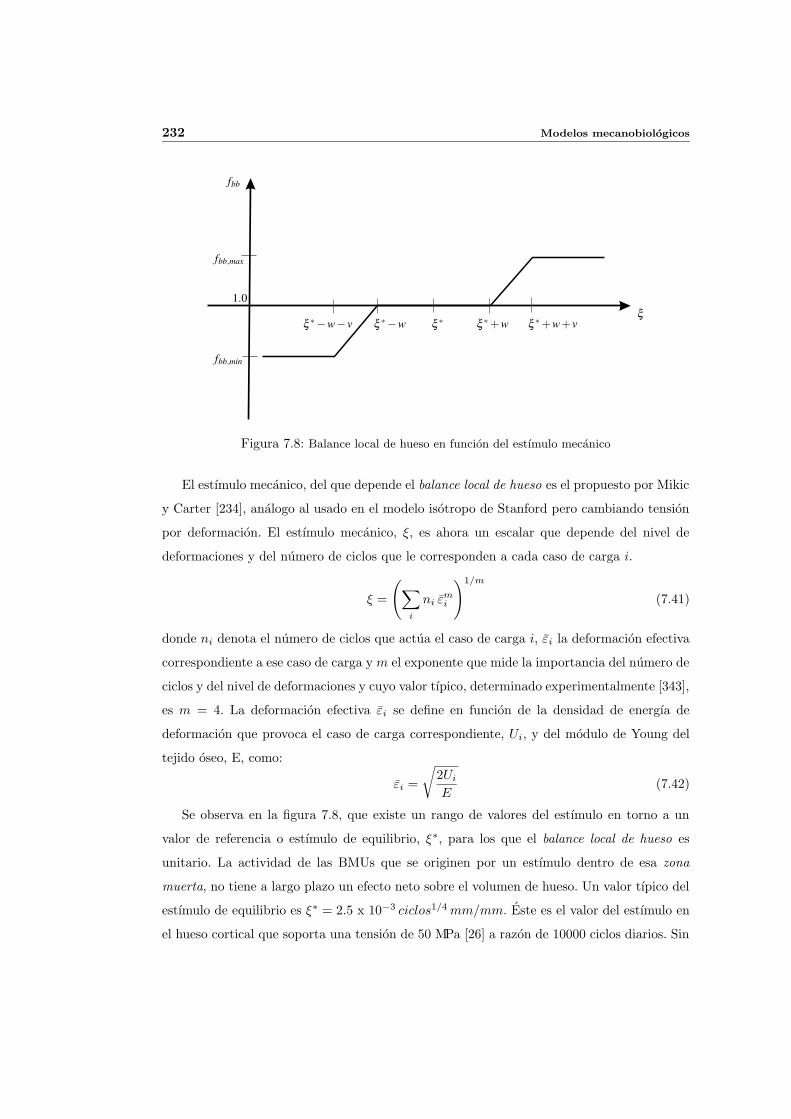
232 Modelos mecanobiologicos
ξξ ∗ ξ ∗
+w ξ ∗+w+ vξ ∗
−wξ ∗−w− v
1.0
fbb
fbb,max
fbb,min
1
Figura 7.8: Balance local de hueso en funcion del estımulo mecanico
El estımulo mecanico, del que depende el balance local de hueso es el propuesto por Mikic
y Carter [234], analogo al usado en el modelo isotropo de Stanford pero cambiando tension
por deformacion. El estımulo mecanico, ξ, es ahora un escalar que depende del nivel de
deformaciones y del numero de ciclos que le corresponden a cada caso de carga i.
ξ =
(∑
i
ni εmi
)1/m
(7.41)
donde ni denota el numero de ciclos que actua el caso de carga i, εi la deformacion efectiva
correspondiente a ese caso de carga y m el exponente que mide la importancia del numero de
ciclos y del nivel de deformaciones y cuyo valor tıpico, determinado experimentalmente [343],
es m = 4. La deformacion efectiva εi se define en funcion de la densidad de energıa de
deformacion que provoca el caso de carga correspondiente, Ui, y del modulo de Young del
tejido oseo, E, como:
εi =
√2UiE
(7.42)
Se observa en la figura 7.8, que existe un rango de valores del estımulo en torno a un
valor de referencia o estımulo de equilibrio, ξ∗, para los que el balance local de hueso es
unitario. La actividad de las BMUs que se originen por un estımulo dentro de esa zona
muerta, no tiene a largo plazo un efecto neto sobre el volumen de hueso. Un valor tıpico del
estımulo de equilibrio es ξ∗ = 2.5 x 10−3 ciclos1/4mm/mm. Este es el valor del estımulo en
el hueso cortical que soporta una tension de 50 MPa [26] a razon de 10000 ciclos diarios. Sin

7.3 Modelo isotropo de remodelacion osea basado en la actividad de las BMUs 233
embargo, este valor parece ser aplicable solo a huesos cuya mision sea la de soportar carga,
como el femur. Huesos menos cargados, como el craneo y otros huesos planos, cuya mision
es principalmente de proteccion, no soportan valores tan altos.
La diferencia de comportamiento entre huesos largos y huesos planos se obtiene en el
modelo considerando que el valor de ξ∗ no es una constante caracterıstica de un hueso
sino que varıa con el tiempo, adaptandose al valor del estımulo externo aplicado, segun el
principio de acomodacion celular propuesto por Turner [329].
Esta teorıa corrige, o mas bien matiza, la teorıa del “mecanostato” propuesta por Frost
[109]. Segun esta el hueso dispone de un mecanismo, que el denomino mecanostato, que
regula la densidad osea de la misma forma que un termostato regula la temperatura de una
habitacion: la velocidad de cambio de la temperatura es proporcional a la diferencia entre la
temperatura actual, T , y la temperatura de control del termostato, To. De la misma forma,
la velocidad de cambio de densidad osea, o velocidad de remodelacion, es proporcional a la
diferencia entre la deformacion que soporta el hueso y una deformacion de referencia, εo,
que es la misma para cualquier tipo de hueso.
termostatodT
dt= −k1 (T − To) (7.43a)
mecanostatodρ
dt= −k2 (ε(ρ)− ε∗o) (7.43b)
Esta ley de remodelacion se usa en gran cantidad de modelos, por ejemplo el modelo
isotropo de Stanford. Con dicha ley, una deformacion o un estımulo demasiado bajo, co-
mo el que sufren los huesos planos, provocarıa reabsorcion de forma indefinida hasta que
desapareciera todo el hueso, algo que no ocurre en la realidad.
Turner, con el mencionado principio de acomodacion celular [329], modifica la ley del
mecanostato diciendo que la deformacion de referencia, εo, no es una constante del tejido
oseo, sino que puede variar para adaptarse a la deformacion que normalmente soporta el
tejido. En otras palabras, el tejido se acomoda a la deformacion que soporta. Ası, la defor-
macion de referencia tiende hacia la deformacion normal de uso siguiendo una ley identica
a las ecuaciones (7.43):dεodt
= ϕ (ε− εo) (7.44)
En este modelo isotropo de BMUs, dicha adaptacion se expresa en terminos de estımulo,
en lugar de deformaciones, siendo la ecuacion diferencial totalmente analoga,
dξ∗
dt= ϕ (ξ − ξ∗) (7.45)

234 Modelos mecanobiologicos
donde ϕ es el parametro que controla la velocidad de adaptacion y que toma en el modelo
un valor de 0.002 dıas−1.
De esta manera, una deformacion o un estımulo demasiado bajo, como el que sufren los
huesos planos, no produce reabsorcion de manera indefinida, sino solo hasta que el estımulo
de equilibrio se adapte a ese valor tan bajo del estımulo exterior. Esto podrıa explicar la
forma de la figura 5.8, que en huesos poco cargados, como el craneo, evita que se pierda
masa osea por desuso. En este sentido, Frost [110] afirma que nunca se producen un estado
de deformacion nula, ya que, incluso en un caso de completo desuso, la presion sistolica hace
fluir la sangre por los capilares del hueso cortical lo que provoca una pequena deformacion.
Bien es verdad que esta deformacion es muy pequena, como argumenta Tuner en el mismo
trabajo [110], donde dice que debe estar en torno a 1.1µε, para una presion sistolica normal.
7.3.3. Balance de masa osea
A continuacion se analiza como varıa la densidad aparente del tejido oseo como conse-
cuencia de los procesos biologicos y mecanicos que ocurren en el hueso y que tiene en cuenta
este modelo.
Derivando con respecto al tiempo la expresion (7.33) se obtiene:
∂ρ
∂t=∂ρt∂t
(vb − h) + ρt∂vb∂t− ρt
∂h
∂t= Πb (7.46)
Cada termino de la expresion anterior determina como evoluciona la masa osea. El primer
termino describe como se mineraliza el osteoide, el segundo representa la evolucion de la
masa osea por reabsorcion y formacion por parte de las BMUs y el tercero representa el
efecto que sobre la masa osea tiene la evolucion del dano.
Crecimiento del dano
Las cargas cıclicas a las que esta sometido el hueso producen una acumulacion de dano
microestructural que da lugar a la formacion y crecimiento de grietas. En este proceso no
hay aportacion de hueso, Πb, ni varıa la densidad del tejido,∂ρt∂t
= 0, con lo que del balance
de masa de la ecuacion (7.46) se deduce que:
∂h
∂t=∂vb∂t
(7.47)
Esta ecuacion establece que el aumento del dano por acumulacion de microgrietas revierte
en un aumento de la cantidad de tejido. Conviene recordar que el hueso danado forma parte

7.3 Modelo isotropo de remodelacion osea basado en la actividad de las BMUs 235
del tejido oseo y no de los huecos (ecuacion (7.27)). La ecuacion (7.47) indica que al abrirse
las grietas, el volumen asociado expande el tejido completo, disminuyendo la proporcion de
volumen ocupada por los poros.
A continuacion se tiene en cuenta el comportamiento a fatiga del hueso para relacionar el
dano acumulado con las cargas a las que esta sometido el tejido. El comportamiento a fatiga
del hueso ha sido estudiado de manera exhaustiva por diversos autores [49,175,198,265,361].
El dano acumulado por fatiga se suele medir a traves de la degradacion del modulo elastico,
para la que se ha definido la variable d. La variacion de este parametro se puede relacionar
con la variacion del volumen danado, h, usando la ecuacion (7.32), y ası:
∂h
∂t= k
∂d
∂t(7.48)
Lemaitre y Chaboche expresaron el crecimiento de dano por fatiga de forma generica
[200]. Usaron expresiones del tipo∂d
∂n= G(d, ε) (7.49)
donde n es el numero de ciclos. Por otro lado, la vida a fatiga, nF , se suele aproximar por
expresiones del tipo:
nF = Cεδ (7.50)
En las dos ecuaciones anteriores es necesario distinguir entre traccion y compresion,
dado que cada uno de estos casos produce una evolucion del dano diferente. Garcıa-Aznar
et al. [117] utilizan las siguientes leyes de evolucion del dano para uno y otro caso:
∂dc∂n
= Gc(dc, ε) =C1
γ1eγ1dc εδ1 en compresion (7.51a)
∂dt∂n
= Gt(dt, ε) =C2
C3γ2(1− dt)1−γ2e−C3(1−dt)
γ2εδ2 en traccion (7.51b)
donde dc y dt son los parametros de degradacion del modulo de Young en compresion
y traccion, respectivamente; N es el numero de ciclos y C1, C2, C3, γ1, γ2, δ1 y δ2 son
parametros del modelo. La forma particular de las funciones anteriores fue elegida ex profeso
por Garcıa-Aznar et al. para ajustar los resultados experimentales obtenidos por Pattin et
al. [265].
La integracion de las ecuaciones (7.51) conduce a las siguientes expresiones:
dc = − 1
γ1[ln(1− C1ε
δ1n)] (7.52a)
dt = 1− γ2
√1
C3ln(eC3 − C2εδ2n) (7.52b)
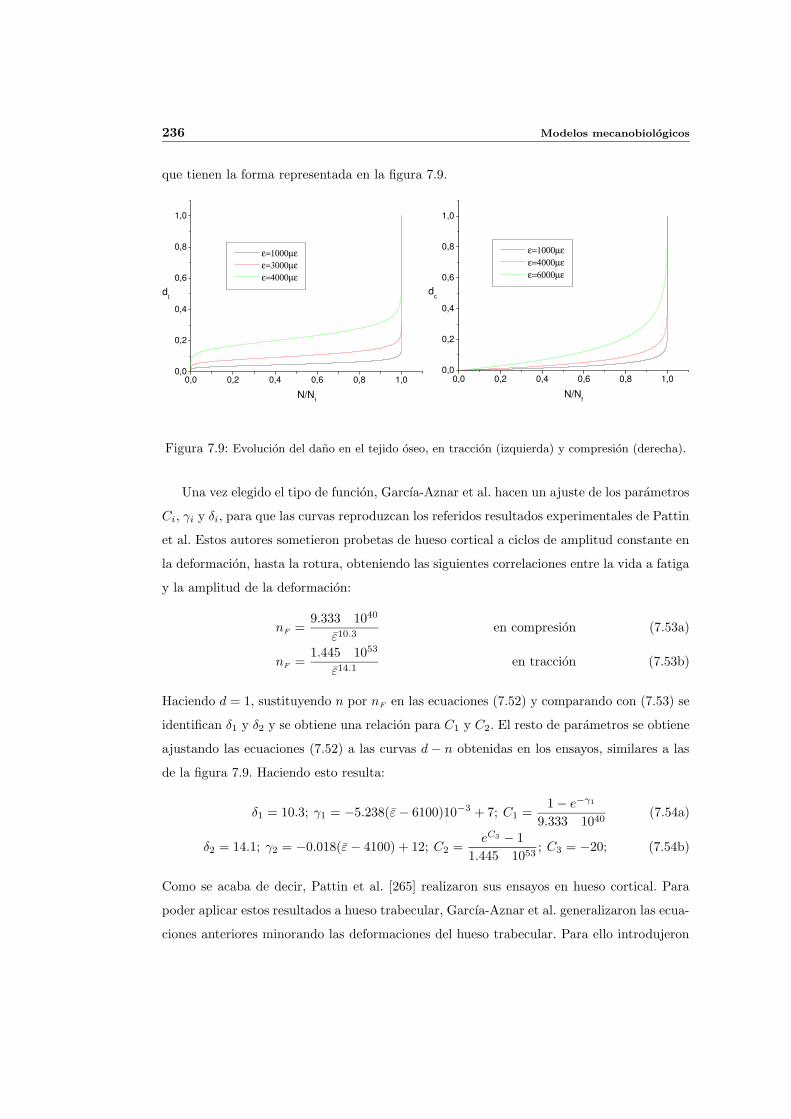
236 Modelos mecanobiologicos
que tienen la forma representada en la figura 7.9.
0,0 0,2 0,4 0,6 0,8 1,00,0
0,2
0,4
0,6
0,8
1,0
dt
N/Nf
ε=1000µε ε=3000µε ε=4000µε
0,0 0,2 0,4 0,6 0,8 1,00,0
0,2
0,4
0,6
0,8
1,0
dc
N/Nf
ε=1000µε ε=4000µε ε=6000µε
Figura 7.9: Evolucion del dano en el tejido oseo, en traccion (izquierda) y compresion (derecha).
Una vez elegido el tipo de funcion, Garcıa-Aznar et al. hacen un ajuste de los parametros
Ci, γi y δi, para que las curvas reproduzcan los referidos resultados experimentales de Pattin
et al. Estos autores sometieron probetas de hueso cortical a ciclos de amplitud constante en
la deformacion, hasta la rotura, obteniendo las siguientes correlaciones entre la vida a fatiga
y la amplitud de la deformacion:
nF =9.333 1040
ε10.3en compresion (7.53a)
nF =1.445 1053
ε14.1en traccion (7.53b)
Haciendo d = 1, sustituyendo n por nF en las ecuaciones (7.52) y comparando con (7.53) se
identifican δ1 y δ2 y se obtiene una relacion para C1 y C2. El resto de parametros se obtiene
ajustando las ecuaciones (7.52) a las curvas d − n obtenidas en los ensayos, similares a las
de la figura 7.9. Haciendo esto resulta:
δ1 = 10.3; γ1 = −5.238(ε− 6100)10−3 + 7; C1 =1− e−γ1
9.333 1040(7.54a)
δ2 = 14.1; γ2 = −0.018(ε− 4100) + 12; C2 =eC3 − 1
1.445 1053; C3 = −20; (7.54b)
Como se acaba de decir, Pattin et al. [265] realizaron sus ensayos en hueso cortical. Para
poder aplicar estos resultados a hueso trabecular, Garcıa-Aznar et al. generalizaron las ecua-
ciones anteriores minorando las deformaciones del hueso trabecular. Para ello introdujeron

7.3 Modelo isotropo de remodelacion osea basado en la actividad de las BMUs 237
un modulo de Young de referencia, E∗, y sustituyeron ε por
(E
E∗ε
)en las ecuaciones anteri-
ores, donde E es el modulo de Young del hueso considerado. Para el hueso cortical el cocienteE
E∗es la unidad y no hay minoracion, puesto que no es necesario hacer la adaptacion de los
resultados experimentales. Para el hueso trabecular se toma E∗ = E(vb = 0.95; α = 0.69)
segun la ecuacion (7.35).
Reabsorcion osea y reparacion del dano
La reabsorcion es la forma que tiene el hueso de reparar el tejido danado. Como se
vera mas adelante, cuando el dano acumulado en el hueso es muy alto se origina un gran
numero de BMUs, que en primer lugar retiran el hueso danado y posteriormente depositan
hueso nuevo, intacto. Con ello se consigue mantener el nivel de dano dentro de unos niveles
aceptables. Lo mismo ocurre con la concentracion de calcio, ya que el hueso antiguo, reab-
sorbido por la BMU, tiene mayor concentracion de mineral que el nuevo hueso depositado.
En este modelo se hace la suposicion de que el volumen danado, Vd, esta uniformemente
distribuido por todo el volumen de tejido, Vb. De esta forma, Vd y Vb disminuyen en la misma
proporcion durante el proceso de reabsorcion, es decir:
1
Vd
∂Vd∂t
=1
Vb
∂Vb∂t
(7.55)
Recordando que h =VdVt
y teniendo en cuenta que Vt no varıa en el proceso de reabsorcion:
∂h
∂t=
1
Vt
∂Vd∂t
=1
Vt
∂Vb∂t
VdVb
= −vR
h
vb, (7.56)
Como durante el proceso de reabsorcion no varıa la densidad del tejido oseo (∂ρt∂t
= 0), de
(7.46) se deduce que el hueso eliminado sera
Πb = −ρtvR
(1− h
vb
)(7.57)
Formacion osea y mineralizacion
El nuevo tejido formado por los osteoblastos, llamado osteoide, esta compuesto princi-
palmente de colageno y agua, como ya se indico en el capıtulo 4. Con el paso del tiempo
este osteoide se mineraliza y parte del agua es sustituida por hidroxiapatita. Tambien se ha
indicado que este proceso de mineralizacion suele dividirse en dos fases: la fase primaria,
muy rapida, dura unos pocos dıas [260], y en la que se alcanza aproximadamente el 70 % del

238 Modelos mecanobiologicos
contenido maximo de mineral y la fase secundaria que puede durar anos. En este modelo
se supone que la fase primaria ocurre instantaneamente, una vez formado el nuevo tejido, y
que en la fase secundaria la velocidad de mineralizacion decrece exponencialmente
α(t) = αmax + (α0 − αmax) e−κt (7.58)
donde α0 es la fraccion de ceniza al final de la fase primaria, αmax es su valor maximo y κ
es una constante que determina la velocidad del proceso y para el que se toma un valor de
6 anos−1.
En el modelo se define un promedio de la fraccion de ceniza en el volumen de control.
Este promedio tiene en cuenta los diferentes procesos que afectan al contenido de mineral
del tejido contenido en dicho volumen:
α(t) =(vb,0 − h0)α(t) +
∫ t0
(vF (τ)− h(τ)
)α(t− τ) dτ −
∫ t0
(vR(τ)− h(τ)
)α(τ) dτ
vb(t)− h(t),
(7.59)
Los diferentes terminos de esta ecuacion representan lo siguiente:
a) El primer termino es el contenido mineral que aporta el tejido que habıa inicialmente
vb,0,
b) La integral de convolucion del segundo termino representa el contenido de mineral
que aporta el tejido formado desde el instante inicial hasta ahora.
c) El tercer termino es una aproximacion de la cantidad de mineral que se retira con la
reabsorcion. Como no se conoce de manera exacta que cantidad de mineral tiene el tejido
reabsorbido, se supone que su fraccion de ceniza es igual al promedio en ese instante.
Los parametros h0 y h aparecen restando en la ecuacion (7.59) porque se supone que la
parte danada de tejido no se mineraliza. Para reducir el coste computacional que requiere
la implementacion de la expresion anterior, Garcıa-Aznar et al. [117] aproximan el valor de
la fraccion de ceniza en el instante t+ ∆t por:
α(t+ ∆t) =α(t)((vb(t)− h(t))e−κ∆t −∆vR) + ∆vFα0
vb(t+ ∆t)− h(t+ ∆t)
+αmax(1− e−κ∆t)(vb(t)− h(t)) + ∆h(α(t)− α0)
vb(t+ ∆t)− h(t+ ∆t)
+(1− e−κ∆t)
∫ t0(vR − h)(αmax − α(τ))dτ
vb(t+ ∆t)− h(t+ ∆t)
(7.60)
donde ∆vF y ∆vR son las cantidades de tejido formado y reabsorbido en el intervalo [t, t+∆t].

7.3 Modelo isotropo de remodelacion osea basado en la actividad de las BMUs 239
Durante el proceso de mineralizacion cambia la densidad del tejido (ver ecuacion (7.46))
y como consecuencia varıa la masa osea en una cantidad
Πb ∆t = [vb(t+ ∆t)− h(t+ ∆t)] ρt(t+ ∆t)− [vb(t)− h(t)] ρt(t) (7.61)
7.3.4. Activacion de BMUs
Para completar el modelo es necesario establecer como aparecen las BMUs y en que can-
tidad, una vez que se conoce el efecto que tiene una BMU sobre el volumen de tejido formado
o reabsorbido. En el capıtulo 4 se comentaron los mecanismos biologicos que se ponen en
marcha para la activacion de las BMUs. A continuacion se relaciona de forma matematica
la cantidad de BMUs que se activan en cada instante con los otros parametros del modelo,
en concreto, con el dano y el estımulo.
Las BMUs estan continuamente originandose y desapareciendo, siendo variable, en gen-
eral, el numero de BMUs activas en cada instante. Las BMUs cesan en su actividad de
manera automatica cuando superan su tiempo de vida, σL, en un fenomeno conocido co-
mo apoptosis. El proceso de activacion, sin embargo, no ocurre automaticamente sino que
esta controlado por diversos factores, que se analizan a continuacion. La velocidad a la que
se generan BMUs se expresa en este modelo de la siguiente forma:
∂NBMU
∂t
(NBMUS
mm3dıa
)= forSv (7.62)
siendo proporcional a la frecuencia de activacion, for, y a la superficie especıfica disponible
para tal activacion, Sv, parametro utilizado tambien en el modelo isotropo de Stanford. La
frecuencia de activacion depende de ciertos factores metabolicos y otros de tipo mecanico. Se
sabe de estudios experimentales que el desuso produce reabsorcion [173,294,318], y que una
carga excesiva tambien induce la remodelacion para reparar el dano que dicha sobrecarga
provoca [216,217,241,332].
Este modelo sigue la teorıa inhibitoria propuesta por Martin [215]. Esta establece que
las celulas de borde tienen una tendencia natural a activar BMUs, tendencia que se inhibe
solo cuando reciben una senal por parte del entramado de osteocitos. Ası, la frecuencia de
activacion viene dada por
for = fbio(1− s) (7.63)
donde fbio es un parametro que engloba a los factores biologicos que influyen en la activacion
de BMUs y s es el valor de la senal inhibitoria, normalizado entre 0 y 1. Esta senal se genera
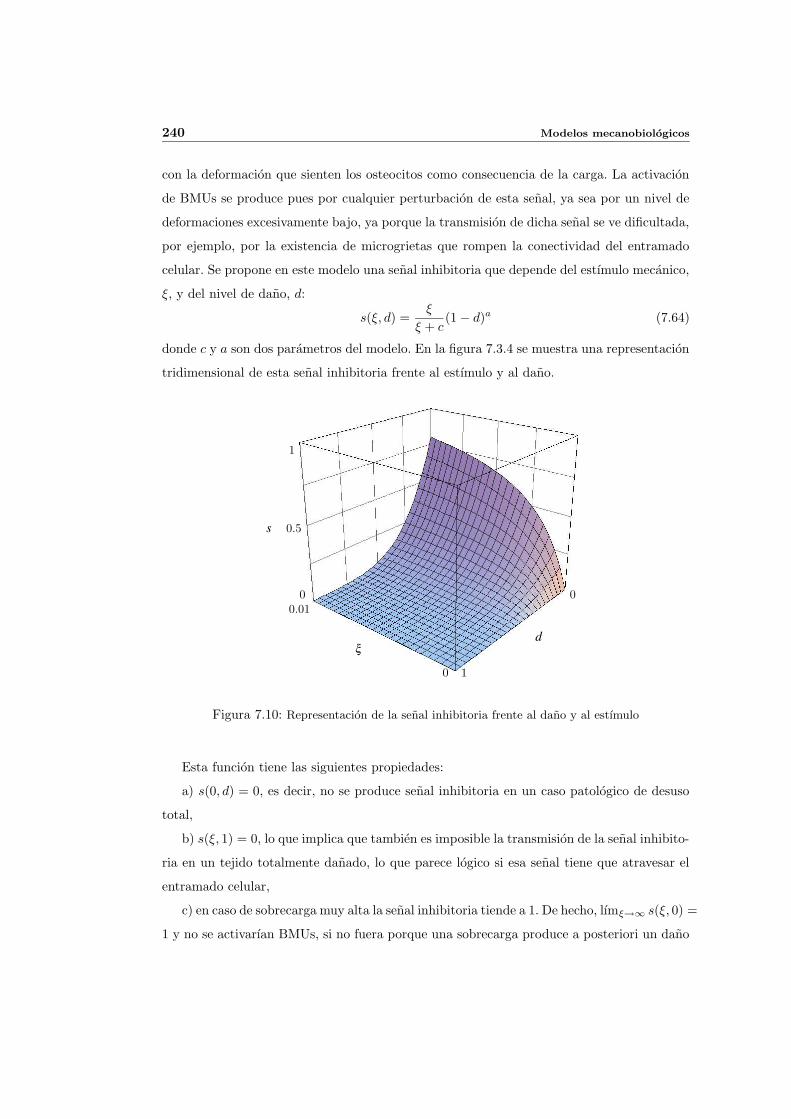
240 Modelos mecanobiologicos
con la deformacion que sienten los osteocitos como consecuencia de la carga. La activacion
de BMUs se produce pues por cualquier perturbacion de esta senal, ya sea por un nivel de
deformaciones excesivamente bajo, ya porque la transmision de dicha senal se ve dificultada,
por ejemplo, por la existencia de microgrietas que rompen la conectividad del entramado
celular. Se propone en este modelo una senal inhibitoria que depende del estımulo mecanico,
ξ, y del nivel de dano, d:
s(ξ, d) =ξ
ξ + c(1− d)a (7.64)
donde c y a son dos parametros del modelo. En la figura 7.3.4 se muestra una representacion
tridimensional de esta senal inhibitoria frente al estımulo y al dano.
0
0
0
0.5
1
1
0.01
dξ
s
1
Figura 7.10: Representacion de la senal inhibitoria frente al dano y al estımulo
Esta funcion tiene las siguientes propiedades:
a) s(0, d) = 0, es decir, no se produce senal inhibitoria en un caso patologico de desuso
total,
b) s(ξ, 1) = 0, lo que implica que tambien es imposible la transmision de la senal inhibito-
ria en un tejido totalmente danado, lo que parece logico si esa senal tiene que atravesar el
entramado celular,
c) en caso de sobrecarga muy alta la senal inhibitoria tiende a 1. De hecho, lımξ→∞ s(ξ, 0) =
1 y no se activarıan BMUs, si no fuera porque una sobrecarga produce a posteriori un dano
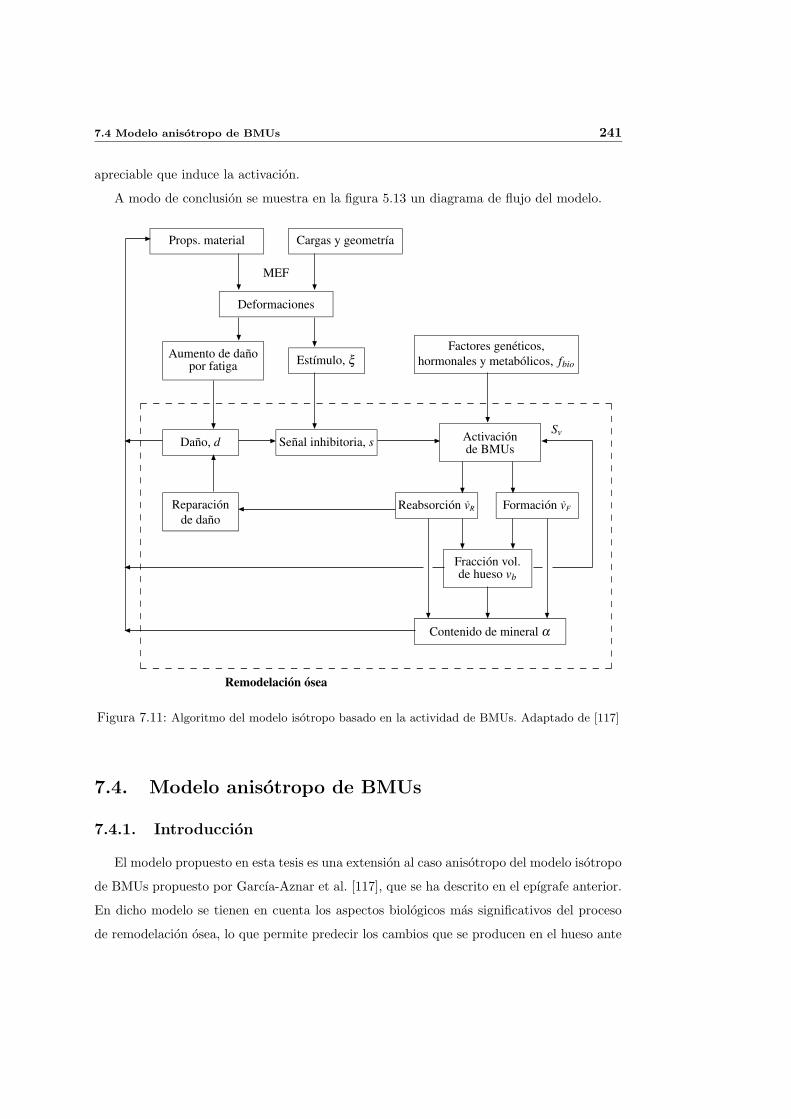
7.4 Modelo anisotropo de BMUs 241
apreciable que induce la activacion.
A modo de conclusion se muestra en la figura 5.13 un diagrama de flujo del modelo.
Props. material Cargas y geometrıa
Deformaciones
Estımulo, ξFactores geneticos,
hormonales y metabolicos, fbioAumento de dano
por fatiga
Dano, d
Reparacionde dano
Senal inhibitoria, s Activacionde BMUs
Reabsorcion vR Formacion vF
Fraccion vol.de hueso vb
Contenido de mineral α
MEF
Sv
Remodelacion osea
1
Figura 7.11: Algoritmo del modelo isotropo basado en la actividad de BMUs. Adaptado de [117]
7.4. Modelo anisotropo de BMUs
7.4.1. Introduccion
El modelo propuesto en esta tesis es una extension al caso anisotropo del modelo isotropo
de BMUs propuesto por Garcıa-Aznar et al. [117], que se ha descrito en el epıgrafe anterior.
En dicho modelo se tienen en cuenta los aspectos biologicos mas significativos del proceso
de remodelacion osea, lo que permite predecir los cambios que se producen en el hueso ante

242 Modelos mecanobiologicos
un cambio de actividad. Sin embargo, solo da informacion de lo que le ocurre a la densidad
y no a la direccionalidad del entramado oseo. Con el presente modelo se pretende salvar esa
limitacion y predecir tanto los cambios de densidad de masa osea como los de arquitectura
tisular. Un aspecto destacado del mismo es que distingue la forma tan diferente de actuar
que tienen las BMUs de hueso cortical y hueso trabecular.
7.4.2. Extension al caso anisotropo de las variables basicas
Las variables del modelo anterior son basicamente las mismas que se utilizan aquı, modif-
icandose en algunos casos su sentido fısico, para que adquieran significado en un modelo
anisotropo como este. Unicamente la fraccion de ceniza, α, y la fraccion volumetrica de hue-
so, vb, tienen la misma definicion que en el modelo isotropo, como magnitudes escalares que
son. El resto de variables que se usaron allı y otras nuevas que se definen especıficamente
para este modelo son magnitudes tensoriales, cuyo sentido fısico se describe a continuacion.
Estas variables son: el tensor MIL, el tensor de deformacion acumulada, el estımulo mecanico
y el estımulo de equilibrio, el dano y la velocidad de activacion de BMUs.
Porosidad y tensor MIL
Quiza la variable mas importante del modelo sea el tensor MIL, que se denominara L,
pues con el va a quedar definida la direccionalidad del tejido oseo, ya sea cortical o trabecular.
Mientras vb establece que cantidad de tejido hay en cada punto del dominio, L, describe como
se reparte ese tejido en el espacio. Este tensor proporciona una medida de la anisotropıa e
influye de manera fundamental en las propiedades elasticas del hueso.
Sobre el tensor MIL ya se hablo en el capıtulo 4. Conviene recordar, sin embargo, algunos
aspectos sobre el. En esta tesis se usa la definicion del tensor MIL (mean interception length)
basada en la definicion dada originalmente por Whitehouse y Dyson [344]. Estos autores
definen el MIL como la longitud promedio de los poros en una determinada direccion. La
definicion usada en esta tesis considera un poro promedio contenido en la unidad de volumen:
Definicion 1 El tensor MIL, L, se define como aquel cuya proyeccion en una determinada
direccion, L(n) = nTLn, representa la longitud del poro promedio contenido en la unidad
de volumen, medida en dicha direccion n.
Cowin fue el primero en asociar dimensiones promedio de los poros con un tensor [70],

7.4 Modelo anisotropo de BMUs 243
en dos dimensiones. Harrigan y Mann [136] extendieron esta relacion a tres dimensiones
y observaron que la longitud de intercepcion promedio para distintas direcciones en el es-
pacio, se ajustaba razonablemente bien a un elipsoide. Este elipsoide se podıa interpretar
matematicamente como el elipsoide de un tensor, el tensor MIL, y fısicamente como un poro
promedio, representativo de la estructura trabecular. El modelo se simplifica enormemente
si se supone que los poros tienen todos la misma geometrıa aunque con distintas dimensiones
y esta geometrıa elipsoidal parece en principio la mas adecuada. La simplificacion no solo es
ventajosa, ademas es necesaria. No hay que olvidar que se esta haciendo una aproximacion
macroscopica de lo que ocurre a nivel microscopico. Para tener en cuenta la forma exacta del
poro, muy variable por otro lado, serıa obligado hacer un modelo microscopico del tejido, lo
que se escapa de los objetivos de esta tesis. Con vistas a facilitar aun mas el desarrollo del
modelo se hace la siguiente suposicion acerca de la geometrıa de los poros.
Hipotesis 1 El poro promedio contenido en la unidad de volumen se supone ortoedrico, de
forma que la porosidad y tensor MIL estan relacionados por la siguiente expresion:
p = det(L) (7.65)
Con esta hipotesis los autovectores del tensor L son las normales a las caras del ortoedro y
los autovalores son las dimensiones de dicho ortoedro, lo que justifica la ecuacion (7.65).
Esta hipotesis no contradice la equivalencia entre poro promedio y elipsoide, por la
siguiente razon. La geometrıa supuesta tiene caracter local, ya que esta asociada al tensor
MIL en un punto o, si se quiere, en un elemento. Sin embargo, las componentes del tensor
MIL se miden experimentalmente en una muestra de tamano macroscopico. Si la escala del
modelo es suficientemente pequena como para que en la citada muestra macroscopica haya un
buen numero de elementos, la intercepcion promedio seguirıa aproximandose razonablemente
bien a un elipsoide, aunque los poros se representaran localmente como ortoedros.
En la teorıa que sigue se estudiara como la actividad de las BMUs modifica la longi-
tud promedio de los poros, es decir, como modifica el tensor L. Al hacerlo varıa tanto la
porosidad, segun la ecuacion (7.65) como la anisotropıa, factores ambos que afectan a las
propiedades elasticas del hueso. Para desacoplar la influencia de estos factores en el tensor
de comportamiento se normaliza el tensor L, definiendose un nuevo tensor M, o tensor MIL
normalizado.

244 Modelos mecanobiologicos
Definicion 2 El tensor M es un tensor paralelo al tensor L, con autovalores proporcionales
a los autovalores de este pero de traza unitaria:
M =1
tr(L)L (7.66)
De esta forma, el nuevo tensor M, solo tiene informacion acerca de la anisotropıa. La
normalizacion anterior es la usada por diversos autores [362, 363] para tensores “fabric”,
analogos al definido aquı. Estos autores relacionan el tensor “fabric” con el tensor de com-
portamiento, pero solo una vez que el tensor se ha normalizado con la traza. Las propiedades
elasticas dependen por un lado de la cantidad de tejido, a traves de la porosidad, y por otro
de la anisotropıa, a traves del tensor “fabric” normalizado, quedando ası desacoplada la
influencia de ambos factores en el tensor de comportamiento.
La normalizacion anterior obliga al uso simultaneo de ambos tensores, L y M, en la
formulacion. Su uso, que se describira en detalle mas adelante, se puede resumir de la
siguiente forma: la actividad de las BMUs afecta directamente al tensor L. Esto es evidente,
ya que formacion y reabsorcion son fenomenos que modifican la longitud de los poros y por
tanto las componentes del tensor L. El determinante de este proporciona la cantidad de
tejido, pero es necesario normalizarlo con traza unitaria, es decir, evaluar M, para conocer
completamente el tensor de comportamiento. La velocidad de variacion de la porosidad se
puede obtener a partir de la velocidad de variacion de L sin mas que derivar respecto al
tiempo la expresion (7.65):
p = p tr(L−1L) (7.67)
Activacion de BMUs. Tensor de estımulo
Otra variable esencial para entender el modelo es el numero de BMUs que se encuen-
tran activas en un determinado instante. Esta era una magnitud escalar en el modelo
isotropo, NBMU , pero ahora, es necesario darle un caracter direccional y distinguir la di-
reccion de avance de las BMUs, que tiene una influencia fundamental en la modificacion de
la anisotropıa.
Definicion 3 Se define el tensor N como aquel cuya proyeccion en una determinada di-
reccion, nT Nn, proporciona el numero de BMUs activas que avanzan en dicha direccion n
por unidad de volumen.

7.4 Modelo anisotropo de BMUs 245
Tambien es interesante conocer el numero de BMUs por unidad de tiempo que se activan
en un determinado instante avanzando en cada direccion. Para ello se define el tensor, N.
Definicion 4 Se define el tensor N como aquel cuya proyeccion en una determinada direc-
cion, nT N n, proporciona el numero de BMUs que se activan por unidad de tiempo y por
unidad de volumen, avanzando en dicha direccion n.
Hipotesis 2 (Orientacion de N para un solo estado de carga) Si un hueso esta someti-
do a un estado de carga determinado, que produce un estado de deformacion local con direc-
ciones principales ei, las BMUs que en un instante dado se activan en dicho punto, avanzan
segun dichas direcciones principales de deformacion y continuan avanzando en esas direc-
ciones hasta que cesa su actividad por apoptosis. Esto equivale a decir que el tensor N es
paralelo al tensor de deformaciones en el instante en que se activan las BMUs. Se puede
escribir entonces:
N(t) =
3∑
i=1
Ni(t) ei(t)⊗ ei(t) (7.68)
donde Ni(t) representa el numero de BMUs por unidad de volumen que se activan en el
instante t avanzando en direccion i y ei(t) es la direccion principal de deformacion i en el
instante t.
Cuando un hueso esta sometido a varios estados de carga durante la actividad normal del
individuo, la anterior hipotesis no es aplicable, puesto que cada caso de carga tendra aso-
ciadas direcciones principales distintas. Hay que tener en cuenta que el tiempo medio de
variacion de las deformaciones durante la actividad normal es mucho menor que el tiempo
que tardan en activarse las BMUs. Por tanto, dichas BMUs no pueden seguir las direcciones
principales de deformacion de manera instantanea. Es necesario establecer una hipotesis mas
general para el caso en el que el hueso soporte diariamente mas de un estado de carga. Se
supondra en ese caso que el hueso recopila informacion de todo lo que ocurre diariamente y
las BMUs se activan en funcion de esa historia diaria mas reciente. Esta hipotesis requiere la
definicion de una deformacion acumulada, analoga al estımulo mecanico definido por Carter
et al. [57]. Este estımulo mecanico ya se uso en el modelo isotropo de BMUs (ver ecuaciones
(5.14) y (7.41)) y tiene en cuenta las distintas actividades que soporta el hueso ası como el
numero de ciclos de cada actividad.
Definicion 5 El tensor de deformacion acumulada se define como la superposicion ponder-
ada de los diferentes estados de deformacion que producen las actividades diarias a las que

246 Modelos mecanobiologicos
se encuentra sometido el hueso.
ε =
N∑
j=1
nj εmj
1/m
=
N∑
j=1
3∑
i=1
(nj εmij ) eij(t)⊗ eij(t)
1/m
(7.69)
donde nj es el numero de ciclos correspondiente al caso de carga j, N es el numero de
actividades distintas que soporta el hueso, m es el exponente introducido por Carter et al.
[57] (se suele tomar el valor m = 4) y εij y eij son respectivamente las deformaciones y
direcciones principales de deformacion i correspondiente al caso de carga j.
Es conveniente hacer la descomposicion espectral del tensor anterior:
ε =3∑
i=1
εi ei ⊗ ei (7.70)
donde ei son las direcciones principales del tensor de deformacion acumulada y εi son sus
autovalores. Una vez conocidos los autovectores del tensor deformacion acumulada, ya es
posible hacer la hipotesis general sobre la orientacion del tensor N.
Hipotesis 3 (Orientacion de N para varios estados de carga) En un hueso someti-
do diariamente a una combinacion de actividades que produce un tensor de deformacion
acumulada ε, se activan BMUs que avanzan segun las direcciones principales de dicho ten-
sor y continuan avanzando en esas direcciones hasta que cesa su actividad por apoptosis. El
tensor N es en este caso paralelo al tensor de deformacion acumulada.
N(t) =
3∑
i=1
Ni(t) ei(t)⊗ ei(t) (7.71)
Puede observarse que la hipotesis 2 es un caso particular de la hipotesis general 3 para
una sola actividad. En ese caso concreto el tensor deformacion acumulada es paralelo al
tensor de deformacion correspondiente a dicha actividad.
El concepto de estımulo mecanico no ha aparecido aun entre las magnitudes definidas.
De hecho, ε es una magnitud analoga a la que en el modelo isotropo se define como estımulo.
Sin embargo, aquı se le ha dado el nombre de deformacion acumulada y se reserva el estımulo
para una magnitud que tenga en cuenta no la deformacion, sino la energıa de deformacion.
Se elige la energıa de deformacion como medida del estımulo mecanico para distinguir entre
situaciones que tienen la misma deformacion pero que son completamente diferentes. Ası,
un hueso intacto sometido a tensiones elevadas tendra deformaciones tambien elevadas.

7.4 Modelo anisotropo de BMUs 247
Igualmente altas pueden ser las deformaciones que sufra un hueso muy danado (por tanto
muy flexible) aunque en este caso las tensiones no tienen que ser necesariamente altas. Si
se usa la deformacion como medida del estımulo mecanico, ambos casos pueden tener el
mismo estımulo, aunque el comportamiento sin duda debe ser diferente en uno y otro hueso.
Ese problema se solventa utilizando la energıa de deformacion absorbida por el tejido. En
el hueso intacto dicha energıa sera alta si las deformaciones lo son, mientras que un hueso
danado apenas podra absorber energıa de deformacion y su estımulo mecanico sera pequeno.
Definicion 6 Se define el tensor estımulo mecanico como un tensor paralelo al tensor de
deformacion acumulada, cuyos autovalores representan una energıa de deformacion asociada
a la direccion principal correspondiente de este ultimo tensor.
ξ =
3∑
i=1
ξi ei ⊗ ei (7.72)
Estos autovalores, ξi, se expresan como
ξi =1
2σpi εi =
1
2(eTi σ ei) εi (7.73)
donde σpi es la proyeccion del tensor de tensiones en la direccion principal de deformacion
acumulada ei y el autovalor ξi es la energıa de deformacion asociada a dicha direccion,
tomando como deformacion la acumulada, εi.
Queda aun por establecer el valor de Ni en las ecuaciones anteriores para que estas
adquieran sentido. El numero de BMUs que se activan en un instante determinado, depen-
dera de la superficie disponible para ello y de una frecuencia de activacion, al igual que en
el modelo isotropo. En el modelo propuesto en esta tesis tambien se sigue la teorıa inhibito-
ria de Martin [215] para definir la frecuencia de activacion. Sin embargo, hay que darle un
caracter direccional a la senal inhibitoria, como se ha hecho con el resto de variables. Eso
se hara inmediatamente, pero antes, conviene introducir el tensor de dano aunque sea de
forma breve, ya que aparecera en la expresion de la senal inhibitoria.
El tensor de dano considerado en este modelo, d, es analogo a la degradacion, d, del
modelo isotropo. La proyeccion del tensor de dano en una determinada direccion esta rela-
cionada con la perdida de rigidez del material en dicha direccion. En apartados posteriores
se vera como es dicha relacion y como se modifica el dano: por fatiga y como consecuencia
del proceso de remodelacion. El tensor de dano esta relacionado con el volumen de teji-
do danado, h, como ya se discutio en el modelo isotropo, a traves de la constante k de la

248 Modelos mecanobiologicos
ecuacion (7.32). Ahora, sin embargo, se ha de tener en cuenta la degradacion en todas las
direcciones, por medio de la traza del tensor d. Ası:
h =k
3tr(d) (7.74)
Si el diferencial de volumen en cuestion esta completamente danado, d sera el tensor unidad
de segundo orden, su traza sera 3 y la ecuacion anterior reproduce la (7.32) del modelo
isotropo.
Frecuencia de activacion. Reparto direccional de las BMUs
Para cada direccion de avance de BMUs se va definir una frecuencia de activacion cuyo
calculo se hace en dos partes. En primer lugar se estima la frecuencia de activacion total o
frecuencia de activacion isotropa,1 sin distinguir direccion de avance. En segundo lugar se
reparten esas BMUs segun las direcciones en las que progresan.
Definicion 7 Se define la frecuencia de activacion isotropa for iso como el numero de BMUs
que se activan por unidad de tiempo y por unidad de superficie disponible para la activacion.
Se calcula mediante la siguiente expresion:
for iso = fbio
(1− ξiso
ξiso + c(1− diso)a
)(7.75)
donde c y a son constantes del modelo; fbio es un parametro que tiene en cuenta diversos
factores biologicos que influyen en la activacion de BMUs, como por ejemplo la edad; la
variable diso es la componente octaedrica del tensor de dano (diso =tr(d)
3), que es una
medida del nivel de dano global y el estımulo ξiso =tr(ξ)
3es la componente octaedrica del
tensor de estımulo.
Segun la expresion anterior, la cantidad de BMUs que se activen dependera de la energıa
de deformacion total que este soportando el hueso y del nivel de dano global, sin distinguir
la direccion de este dano. Ası, una energıa de deformacion alta no induce la activacion
de muchas BMUs aunque haya alguna direccion en la que la energıa de deformacion sea
pequena. Este caso se podrıa interpretar, desde el punto de vista biologico, diciendo que los
osteocitos sienten la deformacion, al menos en alguna direccion, y por tanto no dejan de
enviar la senal que inhibe la activacion de BMUs. Si la energıa de deformacion es pequena,
1Se denomina ası porque la expresion que la define es identica a la del modelo isotropo

7.4 Modelo anisotropo de BMUs 249
entonces sı dejan de sentir la deformacion y dejan de transmitir la senal inhibitoria a las
celulas de borde, que ponen en marcha los mecanismos de activacion. En cuanto al dano,
hay que pensar que las grietas son un obstaculo para la transmision de la senal inhibitoria,
pero es necesario que el dano sea generalizado para que esa senal se vea verdaderamente
obstaculizada. Supongase un caso en el que el dano es elevado en una direccion pero muy
pequeno en las otras dos. La senal puede sortear el dano en la direccion en la que se acumulan
las grietas y llegar a las celulas de borde aprovechando la conexion entre otros osteocitos.
La senal que reciben las celulas de borde sera mas debil, pero algo de senal recibiran. No
dejaran de recibir senal inhibitoria hasta que el dano sea generalizado (elevado en todas
direcciones) y la senal no encuentre un camino posible para alcanzar la superficie del tejido.
En ese caso de dano generalizado sı se activaran muchas BMUs como dice la ecuacion (7.75).
Una vez que se conoce el numero total de BMUs que se activan por unidad de tiempo
y por unidad de superficie especıfica, se hace un reparto de las BMUs segun su direccion
de avance. Para ello se define primero una senal inhibitoria direccional y despues el tensor
frecuencia de activacion.
Para cada direccion principal de estımulo i,1 habra una senal inhibitoria, si, que depende
del estımulo y del dano asociados a esa direccion. La funcion que relaciona estımulo y dano
con la senal inhibitoria es identica a la usada en el caso isotropo (ver ecuacion (7.110)).
Definicion 8 El tensor senal inhibitoria, s, es un tensor paralelo al tensor de estımulo y
de autovalores,
si(ξi, dei) =
ξiξi + c
(1− dei)a (7.76)
donde ξi es el autovalor del tensor de estımulo correspondiente a la direccion i y dei= eTi d ei
es la proyeccion del tensor de dano en dicha direccion. Las constantes c y a son las mismas
de la ecuacion 7.75 e iguales a las del modelo isotropo.
Definicion 9 El tensor frecuencia de activacion for es un tensor paralelo al tensor de
estımulo:
for =
3∑
i=1
for i ei ⊗ ei (7.77)
1En lo que sigue se hara referencia a las direcciones principales de deformacion acumulada como direc-
ciones principales de estımulo. En realidad son las mismas ya que ambos tensores son paralelos. Se hace
ası por brevedad y por tener mas sentido fısico asociar la actividad de las BMUs a un estımulo que a una
mera deformacion.

250 Modelos mecanobiologicos
cuyos autovalores dependen de la senal inhibitoria en la direccion correspondiente, si, y de
la frecuencia de activacion isotropa, for iso,
for i = for iso1− si
3∑
i=1
(1− si)(7.78)
La interpretacion del tensor for es basica para comprender el modelo. Que en una de-
terminada direccion la senal inhibitoria, si, sea alta (muy proxima a 1), quiere decir que
de todas las BMUs que se activen, muy pocas avanzaran en esa direccion. Eso es lo que
ocurre cuando la deformacion en dicha direccion es alta y el dano pequeno. Sin embargo,
cuando haya desuso o cuando el dano en esa direccion sea grande, se interrumpira la senal
inhibitoria, se hara mas proxima a 0, y aumentara la proporcion de BMUs avanzando en esa
direccion. Si el motivo de esta activacion fue el desuso, las BMUs reabsorberan hueso para
hacer menos rıgida esa direccion, ya que no es necesario tanto material, a la vista del nivel de
deformaciones que esta soportando. Por contra, si la activacion se debio al nivel de dano en
esa direccion, las BMUs tendran como mision principal reparar dicho dano. Piensese en una
grieta plana dentro de la matriz osea. En las proximidades de la grieta el dano es unitario
en direccion perpendicular al plano de grieta, entendiendo el dano como una medida de la
degradacion de la seccion resistente. En efecto, la energıa de deformacion que el material
absorbe en direccion perpendicular al plano de grieta, es nula en la cara de la misma. Pues
bien, las expresiones (7.76) y (7.78) establecen que una parte importante de las BMUs que se
activen lo haran avanzando en direccion perpendicular al plano de la grieta. En cierto modo,
esas BMUs van a “coser” la grieta, permitiendo que de nuevo se pueda transmitir energıa
de deformacion de un lado a otro de la misma. En cualquier direccion paralela al plano de
la grieta y si no hay mas grieta que esa, no es necesaria la activacion de BMUs, si no es
por desuso, ya que la transmision de esa senal es perfectamente posible en esas direcciones.
Ası lo establece la ecuacion (7.76), dado que el dano en las direcciones paralelas al plano de
la grieta es nulo.
Se ha comentado que ademas de los factores puramente mecanicos, como el estımulo y el
nivel de dano, existen otros factores que influyen en la activacion de las BMUs. Los factores
biologicos, que son muchos y de muy diversa ındole, se han tenido en cuenta mediante un
unico parametro escalar, fbio, por la dificultad que entrana su cuantificacion. Tambien afecta
a la cantidad de BMUs activadas la superficie disponible para su activacion. Como ya se

7.4 Modelo anisotropo de BMUs 251
ha dicho, las BMUs se originan sobre la superficie libre, tanto del hueso cortical como del
trabecular. Falta, pues, contar con la superficie especıfica de poros, Sv, para completar la
evaluacion del numero de BMUs que se activan por unidad de tiempo. Dado que en este
modelo su influencia sobre N es distinta para hueso cortical y trabecular, se va a analizar
por separado en los dos siguientes epıgrafes.
7.4.3. Actividad de las hemiosteonas. Variacion del tensor MIL en
el hueso trabecular
Se describe ahora como la actividad de una BMU, bien en la fase de reabsorcion, bien
en la de formacion, modifica el tensor L. Existen dos grandes diferencias entre las BMUs de
hueso cortical u osteonas y las de hueso trabecular, tambien llamadas hemiosteonas: por un
lado la geometrıa (ver figura 7.2) y por otro la manera en que avanzan: la osteona atraviesa
el tejido cortical, mientras que la hemiosteona avanza sobre la superficie libre de los poros
del hueso trabecular. Por esta razon se van a tratar por separado.
Activacion de hemiosteonas. Orientacion de la superficie libre de poros
En su tesis doctoral [144], Hernandez, aproxima la forma de las hemiosteonas por medio
cilindro de seccion elıptica (ver figura 7.2). Estas BMUs se originan en la superficie libre
de poros y avanzan por ella hasta la apoptosis. De acuerdo con la hipotesis 3, la BMU
mantiene su direccion de avance durante el perıodo que dura su actividad y por tanto el
numero de BMUs que se activan avanzando en una determinada direccion dependera tambien
de si disponen de mucha o poca superficie para su posterior avance. Ası, el parametro de
superficie especıfica, Sv, debe tener en consideracion, de alguna forma, la arquitectura del
entramado oseo y no solo la relacion entre superficie de poros y volumen de tejido. El
numero de BMUs que se generan por unidad de tiempo y unidad de volumen avanzando en
la direccion principal de estımulo ei se expresa como
Ni =3
2for i Sv i (7.79)
donde Sv i representa la fraccion de la superficie especıfica sobre la que las BMUs pueden
avanzar en direccion ei. El factor 3/2 aparece para asegurar que si el modelo propuesto se
aplica a una situacion isotropa se reproduce el modelo de remodelacion isotropo, como se
comprobara en la demostracion 1 de la pagina 255.

252 Modelos mecanobiologicos
Para calcular Sv i supongase en primer lugar un poro ideal ortoedrico contenido en la
unidad de volumen, como el representado en la figura 7.12. La superficie de dicho poro es
igual, por definicion, a la superficie especıfica de poros y para esta geometrıa en concreto
vale
Sv = 2(li · lj + li · lk + lj · lk) = 2 I2(L) (7.80)
donde li son las dimensiones del poro, es decir, los autovalores del tensor MIL. De acuerdo
con esto y para esta geometrıa sencilla, Sv es dos veces el segundo invariante del tensor L,
que se denominara I2(L).
Sobre una cara cualquiera de ese ortoedro se pueden activar BMUs avanzando en todas
las direcciones ortogonales a la normal a dicha cara, ya que las hemiosteonas avanzan sobre
la superficie de los poros, como se esquematiza en la figura 7.12. Las BMUs senaladas con A
avanzan en direcciones contenidas en la cara sobre la que se han activado. Sin embargo, la
BMU B no puede avanzar sobre esa cara porque atravesarıa la matriz osea. Para generalizar
el concepto, considerese una BMU que avanza en direccion i sobre el poro de la figura. Dicha
BMU podra hacerlo en las caras de normales j y k y dispondra para su activacion de una
fraccion de la superficie disponible de poros que se denominara Sv i, de valor:
Sv i = Svli lj + li lk
li lj + li lk + lj lki, j, k = 1, 2, 3 i 6= j 6= k 6= i (7.81)
En la ecuacion anterior se podrıan usar indistintamente los autovalores del tensor L
que representan las dimensiones reales del poro o los del tensor MIL normalizado M que
representan proporciones entre longitudes, por tratarse de una cociente adimensional. En
ese caso
Sv i = Svmimj +mimk
mimj +mimk +mjmk=
SvI2(M)
mi(1−mi) (7.82)
donde mi son los autovalores del tensor M. Para la segunda igualdad se ha expresado el
numerador del segundo miembro en funcion de la traza, que en el tensor M es unitaria.
Es necesario generalizar esta formulacion para cubrir el caso general en el que la BMU
no avance paralela a alguna de las caras del poro, poro que ademas puede tener una for-
ma cualquiera. En el proceso de generalizacion siguiente se consigue ademas una notacion
tensorial compacta para la superficie especıfica. Para ello se define un tensor Sv que tiene
en cuenta tanto la superficie especıfica disponible para la activacion de las BMU, como su
reparto direccional.

7.4 Modelo anisotropo de BMUs 253
i
j
k
A
A
A
A
B
Figura 7.12: Poro ideal ortoedrico y BMUs avanzando sobre una cara del poro.
Definicion 10 El tensor superficie especıfica trabecular Sv es un tensor paralelo al tensor
MIL cuyos autovalores son los parametros Sv i calculados mediante (7.82). Esto es:
Sv =Sv
I2(M)
3∑
i=1
mi(1−mi) mi ⊗mi (7.83)
La interpretacion de este tensor es importante para entender la ecuacion (7.79). Supongase
un hueso con poros muy alargados y estrechos, como el de la figura 7.13. En este tejido a la
direccion de ortotropıa 2 le corresponde un autovalor m2 muy pequeno. Puede imaginarse
una situacion lımite en la que ese autovalor fuera nulo. En ese caso serıa imposible que se
activaran BMUs avanzando en direccion 2, porque no hay superficie libre de poros disponible
para que progresen sobre ella. Eso es precisamente lo que establece la ecuacion (7.83). En
efecto, lımm2→0 Sv 2 = 0, siendo los otros dos autovalores finitos. Ademas, como en ese caso
lımite m1 +m3 → 1 es inmediato colegir que en el lımite Sv 1 = Sv 3 = Sv, es decir, toda la
superficie del poro esta disponible para el avance de las BMUs en direcciones 1 y 3.
Para el poro ortoedrico ideal considerado hasta ahora las direcciones de avance de las
BMUs estan muy limitadas, por la orientacion de las caras del poro. Sin embargo, la realidad

254 Modelos mecanobiologicos
1
2
Figura 7.13: Poro tıpico de una situacion con elevado grado de anisotropıa: m1 m2
es otra y la geometrıa de los poros sera, por lo general, bien distinta y muy variable. De todos
modos, la definicion del tensor Sv puede generalizarse a cualquier geometrıa de poro, en la
que las BMUs puedan avanzar, a priori, en direcciones cualesquiera. Esta generalizacion se
hara con la proyeccion de Sv en una direccion determinada, que puede interpretarse como la
superficie especıfica de que disponen para activarse las BMUs que avanzan en dicha direccion.
Ası, la ecuacion (7.79) puede reescribirse de la siguiente forma:
Ni =3
2(eTi forei)(e
Ti Svei) (7.84)
y el tensor N, recordando la ecuacion (7.68) y escribiendola en forma compacta:
N =3
2
3∑
i=1
(ei ⊗ ei : for ⊗ Sv : ei ⊗ ei) ei ⊗ ei (7.85)
Como se vera a continuacion, la contribucion al cambio de anisotropıa no es igual para
todas las BMUs que avanzan en una determinada direccion. Si se establece un triedro de
referencia con las direcciones principales de estımulo i, j, k; de entre las Ni BMUs que se
originan avanzando en direccion i, se va a distinguir entre aquellas que aumentan el MIL en
direccion j, que se denominaran Nij , de las que lo hacen en direccion k, Nik, de tal manera
que
Ni = Nij + Nik, (7.86)
no siendo posible que las BMUs que avancen en direccion i aumenten la longitud de poros
en esa misma direccion (Nii = 0), ya que no pueden progresar sobre superficies normales a
i.

7.4 Modelo anisotropo de BMUs 255
El anterior reparto dependera obviamente de la orientacion de la superficie libre de poros.
Por ejemplo, en el poro ortoedrico de la figura 7.12, se puede observar que las BMUs que
avanzan sobre la cara de normal k, modifican la longitud promedio de poros en direccion k.
Si Sv ij designa la superficie disponible para el avance de las Nij , la distincion entre Nij y
Nik es consecuencia del reparto de la superficie de activacion,
Sv i = Sv ij + Sv ik, (7.87)
en la que Sv ij mide que fraccion de Sv i es normal a j. En un poro ortoedrico hay 3 ori-
entaciones unicamente, pero, por lo general, la orientacion de la superficie del poro sera una
funcion continua. En ese caso, Sv ij mide que fraccion de Sv i tiene una normal de orientacion
proxima a j.1 La cantidad de superficie normal a una direccion es proporcional a la longitud
promedio de poros en las otras dos direcciones. Ası:
Sv ij =Sv
I2(M)mimk i, j, k = 1, 2, 3 i 6= j 6= k 6= i (7.88)
y el reparto de la ecuacion (7.87) reproduce (7.82).
Por ultimo, queda demostrar que si el modelo propuesto se aplica a un material isotropo
sometido a un estado de carga hidrostatico, el numero de BMUs que se activan es el mismo
que establece el modelo isotropo.
Demostracion 1 En efecto, en un material isotropo Sv i =2
3Sv segun la expresion (7.81).
Si ademas se carga con un estado de deformacion hidrostatico si = s ∀ i. Entonces, el
numero total de BMUs que se activan por unidad de tiempo sera
N = tr(N) =3
2
3∑
i=1
for i Sv i =3
2
3∑
i=1
for iso1− si
3∑
i=1
1− siSv i = for iso Sv
que coincide con lo que establece el modelo isotropo de BMUs.
Avance de la hemiosteona. Variacion del tensor L
A continuacion se establece de forma matematica como cambia la porosidad y la anisotropıa
del hueso con el avance de las hemiosteonas. Supongase que entre t′ y t′ + dt′ se activan
Nij(t′) dt′ BMUs por unidad de volumen, avanzando en direccion i y aumentando la longitud
1Si nj designa a la direccion j y n a la normal a la superficie en un punto, la condicion podrıa ser
n · nj ≥ cos(45)

256 Modelos mecanobiologicos
de los poros en direccion j, segun la frecuencia de activacion y la superficie disponible, tal
como se ha descrito anteriormente
Nij(t′) = for i(t
′) Sv ij(t′) (7.89)
En un instante posterior, t, esas BMUs tendran algunos osteoclastos activos si se cumple que
t ∈ [t′, t′ + σL + TR]. En ese caso, el volumen de hueso reabsorbido por unidad de volumen
y por unidad de tiempo en el instante t por dichas BMUs vendra dado por:
vR ij(t, t′) = Nij(t
′)AR(t′) vBMU fci(t, t′) dt′ (7.90)
donde fci(t, t′) es una variable que mide la actividad de los osteoclastos y que depende del
instante t′ en que se origino la BMU y del instante actual, t, en el que los osteoclastos de esa
BMU estan reabsorbiendo tejido. La variable AR(t′) mide la cantidad de tejido reabsorbido
en una determinada fase del perıodo de reabsorcion. En concreto, AR(t′) vBMU , proporciona
el volumen de tejido reabsorbido por unidad de tiempo por cada BMU, en el instante t′.
La variable AR se ha introducido porque la BMU no reabsorbe hueso siempre a la misma
velocidad. En efecto, al principio de la fase de reabsorcion la cantidad de tejido eliminado
sera menor mientras se siguen fusionando nucleos de preosteoclastos para formar osteoclastos
maduros. Cuando el numero de osteoclastos de la BMU se estabiliza, tambien lo hace la ve-
locidad de reabsorcion. En la etapa final de este proceso de reabsorcion, cuando los primeros
osteoclastos sufren apoptosis, la velocidad vuelve a decrecer, haciendose nula cuando no
quedan osteoclastos activos. Los perıodos en los que cambia la velocidad de reabsorcion son
breves comparados con la vida total de la BMU y, sobre todo, comparados con el tiempo
que tardan en ser apreciables los cambios en el hueso por efecto de la remodelacion. Por
ello, la influencia de la variacion de la velocidad de reabsorcion con el tiempo es pequena,
por lo general. Sin embargo, sı puede tenerla en algunas situaciones, en las que la poblacion
de BMUs sufra un aumento muy brusco.
Para tener en cuenta dicho efecto se puede usar la siguiente aproximacion de AR:
AR(t′) =
ABMU
TR
t′ si t′ ≤ TR
ABMU si TR < t′ ≤ σL
ABMU
TR
(σL + TR − t′) si σL < t′ ≤ TR + σL
(7.91)

7.4 Modelo anisotropo de BMUs 257
donde ABMU es la seccion de la BMU. En el caso de hueso trabecular la BMU se supone
semielıptica, como hace Parfitt [262] (ver figura 7.2), con lo cual ABMU =π
4de dBMU .
La figura 7.14 muestra un esquema que ayuda a comprender esta expresion. En ella se
simplifica el progreso de una BMU, mostrando cinco instantes del proceso de reabsorcion. La
formacion no se ha representado, por claridad. Se supone que el tejido se reabsorbe en bandas
como las representadas en gris oscuro, de volumen ARvBMUdt′, y que la velocidad de avance
de la BMU, vBMU , es constante. En a aun no se han diferenciado todos los osteoclastos y la
velocidad de reabsorcion aumenta linealmente. TR es el perıodo que tarda en completarse la
reabsorcion en una determinada seccion. En ese instante, b, se supone que la velocidad de
reabsorcion se estabiliza y es constante durante la fase c. En d, cuando ha transcurrido el
perıodo de tiempo σL, que se denomina vida de la BMU, los primeros osteoclastos que se
activaron sufren apoptosis y a partir de aquı, durante la fase e, la velocidad de reabsorcion,
decrece al disminuir el numero de osteoclastos activos. Decrece linealmente con el tiempo,
hasta el instante σL + TR cuando todos los osteoclastos han sufrido apoptosis, en el que se
hace cero.
Tambien se puede utilizar un valor de AR promedio, constante durante el perıodo que
dura la fase de reabsorcion,
AR =1
TR + σL
∫ TR+σL
0
AR(t′) dt′ =ABMU σL
TR + σL
(7.92)
En esta tesis se ha adoptado esta ultima opcion por simplicidad. El uso de una u otra
opcion afecta unicamente al perıodo transitorio que tiene lugar tras un cambio en la actividad
que soporta el hueso y, solo si ese cambio provoca una variacion muy brusca en la poblacion
de BMUs. Aun ası, las diferencias son pequenas y se van atenuando a medio plazo, al llegar
al equilibrio de remodelacion.
Hasta aquı lo que se refiere a la fase de reabsorcion. En la fase de formacion, los os-
teoblastos rellenan de osteoide el hueco dejado anteriormente por los osteoclastos. Las
BMUs activadas en el instante t′ se encuentran en fase de formacion en el instante t si
t ∈ [t′ + TR + TI , t′ + σL + TR + TI + TF ]. En ese caso formaran un volumen de hueso por
unidad de volumen y por unidad de tiempo:
vF ij(t, t′) = Nij(t
′)AF (t′) vBMU fbi(t, t′) dt′ (7.93)

258 Modelos mecanobiologicos
vBMU
a
t ′< TR
b
t ′= TR
c
TR < t ′< σL
d
t ′= σL
e
σL < t ′< σL +TR
Hueco Hueso Hueso en reabsorcion
1
Figura 7.14: Progreso de la reabsorcion en una BMU sobre la superficie de una trabecula de hueso
esponjoso.

7.4 Modelo anisotropo de BMUs 259
donde AF (t′) es una variable analoga a AR(t′) para el perıodo de formacion y viene dada
por
AF (t′) =
ABMU
TF
(t′ − TR − TI) si TR + TI < t′ ≤ TR + TI + TF
ABMU si TR + TI + TF < t′ ≤ TR + TI + σL
ABMU
TF
(TR + TI + TF + σL − t′) si TR + TI + σL < t′ ≤ TR + TI + TF + σL
(7.94)
Igualmente, fbi(t, t′) es analoga a fci(t, t
′) pero ahora mide la actividad de los osteoblastos
en las BMUs que avanzan en direccion i. La razon entre las dos, fbbi = fbi/fci, denominada
balance local de hueso, esta relacionada con el nivel de estımulo y con la edad. Aquı se
supondra el mismo tipo de relacion estımulo-balance local de hueso que en el caso isotropo
(ver figura 7.8), pero ahora teniendo en cuenta la direccion correspondiente. Su papel en el
modelo se vera en detalle en un apartado posterior.
Se puede utilizar la expresion (7.94) o bien un valor promedio constante como se hizo en
reabsorcion. De hecho, esto ultimo es lo que se hace en esta tesis, usando
AF =ABMU σL
TF + σL
(7.95)
Una vez analizados todos los factores presentes en la ecuaciones (7.90) y (7.93), es posible
obtener la variacion del tensor MIL, L. El aumento de la longitud media de poros que la
reabsorcion produce en la direccion principal de estımulo i y que se denominara dlR i, provoca
un cambio en el tensor L:
dL = dlR i ei ⊗ ei (7.96)
y es debida a BMUs que avanzan en direcciones principales de estımulo j y k, en concreto
a Nji y Nki. Estas BMUs en su avance provocan un cambio en la porosidad dado por la
ecuacion (7.90):
dp = (vR ji + vR ki)dt =(Nji(t
′)fcj(t, t′)dt′ + Nki(t
′)fck(t, t′)dt′
)AR(t′) vBMU dt (7.97)
Recordando la ecuacion (7.67) es posible relacionar dp con dL:
dp = p tr(L−1 ei ⊗ ei)dlR i (7.98)
de la que se puede despejar dlR i:
dlR idt
=
(Nji(t
′)fcj(t, t′)dt′ + Nki(t
′)fck(t, t′)dt′
)AR(t′) vBMU
p tr(L−1 ei ⊗ ei)(7.99)

260 Modelos mecanobiologicos
Hasta ahora solo se ha tenido en consideracion la contribucion de las BMUs generadas
entre t′ y t′ + dt′. Para incluir la contribucion de las BMUs que se originaron previamente
y que aun estan activas, se integran las expresiones anteriores respecto a t’. Evaluando el
crecimiento de los poros en las tres direcciones principales de estımulo del instante t’ y
usando la descomposicion espectral, el tensor LR se puede expresar de la siguiente forma:
LR(t) =
3∑
i=1
∫ t
t−σL−TR
dlR i
dt dt′ei(t
′)⊗ ei(t′) dt′ (7.100)
Las dimensiones de los poros se modifican de forma analoga en formacion:
dlF idt
=
(Nji(t
′)fbj(t, t′)dt′ + Nki(t
′)fbk(t, t′)dt′
)AF (t′) vBMU
p tr(L−1 ei ⊗ ei)(7.101)
LF (t) =
3∑
i=1
∫ t−TR−TI
t−σL−TR−TI−TF
dlF i
dt dt′ei(t
′)⊗ ei(t′) dt′ (7.102)
Finalmente, se superponen los efectos de formacion y la reabsorcion:
L = LR − LF (7.103)
y se actualiza el tensor L integrando L, por ejemplo, con un esquema de integracion de
Euler explıcito:
L(t+ ∆t) = L(t) + L(t)∆t (7.104)
que se normaliza dividiendo por su traza, para obtener el tensor M, del que dependen las
propiedades elasticas del tejido oseo, en la forma que se estudiara mas adelante.
7.4.4. Actividad de las osteonas. Variacion del tensor MIL en el
hueso cortical
La principal diferencia entre las hemiosteonas y las osteonas, es que estas ultimas no
avanzan sobre la superficie de los poros como la hemiosteonas, sino que atraviesan el tejido
de la misma forma en que lo harıa una tuneladora. De ahı que su forma sea un cilindro
completo, de seccion aproximadamente circular. Hay que aclarar que aunque no avancen
sobre la superficie de los poros sı se originan en ella, por lo que la superficie especıfica, Sv,
tambien tiene un papel importante en el numero de BMUs que se activan por unidad de
tiempo, aunque esta influencia sea menor y no tenga el caracter direccional que tiene en

7.4 Modelo anisotropo de BMUs 261
el hueso trabecular. Se mantiene la definicion de Ni(t′), como el numero de BMUs que se
activan entre t′ y t′ + dt′ avanzando en la direccion principal de estımulo i. Ası:
Ni = for i Sv (7.105)
donde for i tiene el mismo significado que en hueso trabecular. Es inmediato demostrar que
en un tejido cortical sometido a un estado de carga hidrostatico, el numero de BMUs que
se activan por unidad de tiempo coincide con el que establece el modelo isotropo
Demostracion 2 En efecto, no hay que exigir ningun tipo de simetrıa al material. Si la
carga es hidrostatica si = s ∀ i y el numero total de BMUs que se activan por unidad de
tiempo es
N = tr(N) =
3∑
i=1
for i Sv = Sv
3∑
i=1
for iso1− si
3∑
i=1
1− si= for iso Sv
que coincide con lo que establece el modelo isotropo de BMUs.
En la figura 7.15 se esquematiza el progreso de una BMU de hueso cortical. Se observa
que el avance de la BMU en una determinada direccion provoca la variacion de la longitud
media de poros en las direcciones perpendiculares, manteniendo de esta forma la isotropıa
transversal, simetrıa caracterıstica del hueso cortical. Ası, una BMU avanzando en la direc-
cion principal de estımulo i y en fase de reabsorcion provoca una variacion del tensor L dada
por:
dL = dl (ej ⊗ ej + ek ⊗ ek) (7.106)
Xi
j
ki
j
k
vBMU
1
Figura 7.15: Progreso de la BMU en hueso cortical.

262 Modelos mecanobiologicos
De la misma forma que se ha relacionado dL con el cambio de porosidad que experimenta
el hueso trabecular durante el avance de la BMU, se puede hacer en el caso de hueso cortical,
por medio de la ecuacion (7.67):
dp = p tr(L−1(ej ⊗ ej + ek ⊗ ek)
)dl (7.107)
Se supondra que cada BMU reabsorbe y forma un volumen VBMU =π
4LBMU (d2
O − d2H)
a una velocidad constante durante el tiempo que dura cada proceso, TR + σL y TF + σL
para reabsorcion y formacion, respectivamente. La variable introducida LBMU = vBMU σL es
la longitud recorrida por la BMU en su progreso. Tambien cabe la posibilidad de suponer
una ley de variacion de la velocidad de reabsorcion (o formacion) como la que proporciona
el uso de la ecuacion (7.91) (o (7.94)). Sin embargo, se ha optado por suponer un valor
promedio, constante, como se hizo en el hueso trabecular. En reabsorcion y formacion la
velocidad de variacion de la porosidad debida a las BMUs activadas entre t′ y t′ + dt′,
sera respectivamente:
vR = p =VBMU Ni fciTR + σL
dt′ (7.108a)
vF = −p =VBMU Ni fbiTF + σL
dt′ (7.108b)
Teniendo en cuenta (7.108a) y (7.108b) se puede obtenerdljdt
=dlkdt
=dl
dtdebida a todas
las BMUs que avanzan en direccion i y se encuentran activas en el instante t, integrando en
la variable t′ que mide el origen de tiempos de la activacion.
dlRdt
=VBMU
p (TR + σL)
∫ t
t−σL−TR
Ni(t′) fci(t, t
′)
tr[L−1
(ej(t′)⊗ ej(t′) + ek(t′)⊗ ek(t′)
)] dt′ (7.109a)
dlFdt
=VBMU
p (TF + σL)
∫ t−TR−TI
t−σL−TR−TI−TF
Ni(t′) fbi(t, t
′)
tr[L−1
(ej(t′)⊗ ej(t′) + ek(t′)⊗ ek(t′)
)] dt′ (7.109b)
En el caso general en el que existan BMUs avanzando en las tres direcciones principales
de estımulo:
LR =VBMU
p (TR + σL)
3∑
i=1
∫ t
t−σL−TR
Ni(t′) fci(t, t
′)(ej ⊗ ej + ek ⊗ ek
)
tr[L−1
(ej ⊗ ej + ek ⊗ ek
)] dt′ (7.110a)
LF =VBMU
p (TF + σL)
3∑
i=1
∫ t−TR−TI
t−σL−TR−TI−TF
Ni(t′) fbi(t, t
′)(ej ⊗ ej + ek ⊗ ek
)
tr[L−1
(ej ⊗ ej + ek ⊗ ek
)] dt′ (7.110b)
que junto con las ecuaciones (7.103) y (7.104) permite actualizar el tensor L(t+ ∆t).

7.4 Modelo anisotropo de BMUs 263
7.4.5. Balance local direccional de hueso y principio de acomodacion
celular
Para completar el analisis de la actividad de las BMUs es necesario conocer en cada
momento que cantidad de tejido estan reabsorbiendo los osteoclastos y que cantidad estan
formando los osteoblastos. En concreto, es necesario cononcer cual es el balance entre ambas
cantidades, para lo que se define el llamado balance local direccional de hueso.
Principio de acomodacion celular. Error de remodelacion
Antes de describir el balance local direccional de hueso se extendera al caso anisotropo el
principio de acomodacion celular, del que ya se ha hablado anteriormente. Se hace en este
momento ya que tiene una influencia fundamental en el balance local direccional de hueso.
El principio de acomodacion celular, introducido por Turner [329], establece que los sis-
temas biologicos que responden a senales externas tienden a hacerlo de forma cada vez mas
atenuada, conforme la “fuerza” que les lleva a responder se acomoda a la senal externa. En
el caso de la remodelacion osea, la “fuerza” que provoca la respuesta del hueso es el estımulo
mecanico. Ante un cambio de este estımulo mecanico el sistema responde haciendo que las
BMUs formen mas o menos hueso. Pero si ese estımulo mecanico se mantiene constante,
el sistema se acomoda, de manera que pasado un cierto tiempo las BMUs forman el mis-
mo volumen de tejido que reabsorben, alcanzandose una situacion de equilibrio (equilibrio
homeostatico).
Se define ası un tensor estımulo de equilibrio, ξ∗, que tiende hacia el tensor de estımulo
externo siguiendo la ecuacion del mecanostato
dξ∗
dt= ϕ (ξ(t)− ξ∗) (7.111)
Esto quiere decir que la acomodacion no solo ocurre en magnitud (de los autovalores)
sino que tambien es direccional (de los autovectores). Ası, si se produce un cambio en la
magnitud del estımulo, las BMUs van a responder a ese cambio formando mas o menos
hueso, pero esa respuesta se va a atenuar con el tiempo por la acomodacion en magnitud del
estımulo. Si lo que se produce es un cambio en la orientacion del estımulo la respuesta de las
BMUs va a ser direccional, distinta segun la direccion en la que avanzan. Pero nuevamente,
tras un cierto tiempo, la respuesta se va a hacer uniforme, por la acomodacion direccional
del estımulo.

264 Modelos mecanobiologicos
Observese igualmente la dependencia del tiempo que tiene el tensor estımulo externo en
la ecuacion (7.111), que corresponde a la situacion mas general. Esto hace que para integrar
dicha ecuacion sea necesario recurrir a la integracion numerica, para lo que se ha empleado
el metodo de Euler explıcito.
Definicion 11 El tensor error de remodelacion, ψ, se define como la diferencia entre el
tensor de estımulo y el tensor de estımulo de equilibrio y mide lo lejos que esta el sistema
de alcanzar el equilibrio homeostatico.
ψ(t) = ξ(t)− ξ∗(t) (7.112)
Para evaluar la cercanıa del equilibrio homeostatico se utilizara, mas concretamente, un
escalar, el modulo del tensor error de remodelacion, ψ, definido de la siguiente forma
ψ = |√ψ : ψ| (7.113)
Segun el principio de acomodacion celular, el tensor error de remodelacion tiende con el
tiempo al tensor nulo de segundo orden, y su modulo a 0, lo que caracteriza a un estado de
equilibrio homeostatico. De forma analoga a lo que hacen Beaupre et al. [26,27], el balance
entre formacion y reabsorcion, el balance local direccional de hueso en definitiva, se va a
definir en funcion de este error de remodelacion.
Papel del balance local direccional de hueso
En las ecuaciones (7.99), (7.101) y (7.110), la actividad de osteoclastos y osteoblastos
esta cuantificada con las variables fci y fbi respectivamente. Se pueden definir sendos ten-
sores, f c y f b para medir dichas actividades, paralelos al tensor de estımulo y con autovalores
fci y fbi. A la relacion fbb i = fbi/fci se le denomina balance local direccional de hueso y
analogamente puede definirse un tensor f bb, tambien paralelo al tensor de estımulo y de
autovalores fbb i.
Puede llamar la atencion que en las citadas ecuaciones, fbi y fci dependan de t y de t′
cuando el resto de variables solo depende de t′. Esto obedece a dos motivos:
a) La BMU responde al estımulo que recibe el hueso en este preciso instante, t, que, en
general, no ha de coincidir con el que recibıa en t′, cuando la BMU se activo. De esta forma,
si el hueso esta soportando ahora deformaciones mas elevadas que entonces, los osteoblastos

7.4 Modelo anisotropo de BMUs 265
formaran mas hueso del que quitaron los osteoclastos en la fase de reabsorcion y el tejido se
adaptara mas rapidamente a un cambio de actividad.
b) Por otro lado, dicha dependencia de t y t′ responde a la necesidad de matizar la
hipotesis fundamental del modelo: que las BMUs avanzan en las direcciones principales de
estımulo del instante en el que se originan. A continuacion se discute esa hipotesis y la forma
en que se matiza, introduciendo la dependencia de t en el balance local direccional de hueso.
Las BMUs que se activan en el instante t′ avanzan segun las direcciones principales de
estımulo de ese instante, ei(t′), y lo siguen haciendo ası hasta la apoptosis. Sin embargo, es
posible que en la realidad el progreso de una BMU sufra cambios de orientacion. En el hueso
trabecular, estos pueden ser debidos a la orientacion de la superficie sobre la que progresa
la BMU. Ası, es obvio que si la superficie libre de los poros no es plana y se ha supuesto
que la BMU progresa sobre la superficie, dicha BMU debe girar para mantenerse sobre ella.
Sin embargo, esto carece de sentido con la hipotesis de poro ortoedrico hecha aquı. Por el
contrario, sı es posible que el mismo proceso de remodelacion, al modificar la anisotropıa
haga girar el ortoedro, lo que tambien obligarıa a la BMU a girar para mantenerse sobre la
superficie.
En el hueso cortical, en cambio, las BMUs pueden atravesar el tejido y el cambio de
orientacion, de ocurrir, se debera a otros motivos. Uno de ellos puede ser la reorientacion
de las direcciones principales de estımulo. En tal caso, el modelo aquı propuesto no permite
que las BMUs previamente activadas giren para seguir las nuevas direcciones principales
de estımulo. Lo que sı ocurrira es que las nuevas BMUs progresaran en las nuevas direc-
ciones. De todos modos, las consecuencias de esta limitacion no son importantes, ya que se
esta hablando de giros inapreciables. En efecto, esos giros son muy pequenos por la siguiente
razon: las direcciones principales de estımulo se mantienen aproximadamente inalteradas,
salvo en casos excepcionales como cambios drasticos en la actividad del individuo o de tipo
traumatico. En cualquier caso, la adaptacion de la estructura osea y, consecuentemente, el
cambio de las direcciones principales de estımulo, se produce lentamente. Por tanto, antes
de que cambie de forma apreciable, la BMU ha sufrido apoptosis y en su lugar, otras BMUs
se habran originado, ya en las nuevas direcciones principales de estımulo.
Por tanto, la hipotesis de partida del modelo: que la BMU, una vez activada, no cambia
su direccion de avance ante un cambio en las direcciones principales de estımulo, puede
considerarse suficientemente valida para simular la mayorıa de las situaciones. Sin embargo,

266 Modelos mecanobiologicos
si los cambios de estımulo son muy bruscos, se puede conseguir que influyan en la respuesta
del modelo, aunque las BMUs no giren. Esto se consigue dando mas importancia a las BMUs
que avanzan en direcciones proximas a las actuales direcciones principales de estımulo. Para
ello se hace la siguiente proyeccion del tensor error de remodelacion
Definicion 12 Se define el error de remodelacion proyectado, ψi(t, t′), como la proyeccion
del tensor error de remodelacion actual ψ(t) en cada direccion principal de estımulo ei(t′)
en la que avanzan una serie de BMUs.
ψi(t, t′) = ei(t
′)Tψ(t)ei(t′) (7.114)
Uno de los objetivos de esta proyeccion es permitir que cambie el balance formacion-
reabsorcion que tienen las BMUs que ya avanzan en una determinada direccion, cuando
cambia repentinamente el estımulo. Supongase, por ejemplo, que en un instante determi-
nado un hueso deja de soportar carga en direccion x. Como consecuencia del desuso se
activaran muchas BMUs avanzando en esa direccion y su efecto neto sera de reabsorcion. Si
por algun motivo cambia la actividad del hueso y la carga en direccion x pasa a ser alta,
las BMUs que siguen avanzando en esa direccion, aunque se activaran por el desuso, ahora
van a formar gran cantidad de tejido, como respuesta al elevado nivel de estımulo actual en
esa direccion. Todo ello gracias a que la proyeccion del tensor error de remodelacion en la
direccion original de avance ha cambiado de signo.
Pero con la anterior proyeccion tambien se consigue dar mas relevancia a aquellas BMUs
que avanzan paralelas al tensor error de remodelacion actual, puesto que ellas son precisa-
mente las que tienen mayor protagonismo en la correccion de dicho error de remodelacion.
Sin llegar a cambiar sus trayectorias, el efecto de las BMUs sı estara girando, en cierta forma,
con el cambio de las direcciones principales de estımulo.
Zona muerta de remodelacion. Relacion error proyectado-balance local direc-
cional de hueso
Como ya se ha dicho, el balance local direccional de hueso va a depender del error de
remodelacion, en la lınea de lo establecido por muchos autores [26, 158, 166, 167]. Esa de-
pendencia se va a dividir en dos partes. Por un lado se define la zona muerta de remod-
elacion, donde el balance local direccional de hueso es neutro, y en segundo lugar se describe
que ocurre cuando el error de remodelacion esta fuera de la zona muerta.

7.4 Modelo anisotropo de BMUs 267
Definicion 13 Se define la zona muerta de remodelacion como el rango de valores de ψ
para el que existe equilibrio homeostatico de remodelacion local, es decir, en zona muerta los
osteoblastos forman la misma cantidad de tejido que reabsorben los osteoclastos (fbb i = 1).
Si no se esta dentro de la zona muerta de remodelacion el balance local direccional de hueso
sera mayor o menor que uno (de formacion neta o reabsorcion neta), en funcion del recien
definido error de remodelacion proyectado. Dicha funcion es lineal a trozos y se representa
en la figura 7.16. En resumen, la relacion entre el error de remodelacion y el balance local
direccional de hueso se puede expresar de la siguiente forma:
Si 0 ≤ ψ ≤ w ⇒ fbb i = 1 ∀ i
Si ψ > w ⇒ fbb i =
fbbmin si ψi(t, t′) ≤ −v
1 +1− fbbmin
vψi(t, t
′) si − v < ψi(t, t′) ≤ 0
1 +fbbmax − 1
vψi(t, t
′) si 0 < ψi(t, t′) ≤ v
fbbmax si v < ψi(t, t′)
(7.115)
donde w es el semiancho de la zona muerta y v representa el ancho de las zonas de transicion
lineal. Se mantiene el nombre de semiancho para w, por analogıa con el caso isotropo, pero
serıa mas exacto llamarlo ancho de la zona muerta por el caracter positivo de ψ, razon por
la cual no esta definido el valor de fbb i en la region ψ < 0.
La actividad de osteoclastos se supone unitaria, fc i = 1, con lo que el balance local
direccional de hueso se identifica con la actividad de osteoblastos. Esto quiere decir que los
osteoclastos reabsorben siempre el mismo volumen de hueso y los osteoblastos forman mas
o menos osteoide en funcion del error de remodelacion.
Para entender como afecta la anterior relacion ψi−fbb i al grado de anisotropıa, supongase
que se parte de una situacion isotropa y se aplica una carga cuasi-uniaxial. Supongase
tambien que el estımulo ξ1 es alto, por encima del de equilibrio, y que los estımulos en las
otras dos direcciones estan cerca de los correspondientes estımulos de equilibrio y el error
de remodelacion, por tanto, en la zona muerta. En esta situacion se pueden extraer las
siguientes conclusiones:
a) Si ψ > w y ψ1 > 0 entonces fbb1 > 1 y en las BMUs que avanzan en direccion
1 predomina la formacion frente a la reabsorcion. En las otras direcciones el balance local
direccional de hueso sera proximo al neutro y las BMUs que avanzan segun ellas forman
practicamente la misma cantidad de hueso que antes han retirado.

268 Modelos mecanobiologicos
ψiv−v
1.0
fbbi =
fbi
fc i
fbb,max
fbb,min
1
Figura 7.16: Balance local de hueso en funcion del error de remodelacion proyectado en la direccion
ei de avance de la BMU.
b) Segun las ecuaciones (7.99), (7.101) (de hueso trabecular) y (7.110) (de hueso cortical),
el efecto de las BMUs que avanzan en direccion 1 es el de disminuir la longitud promedio
de los poros en direcciones 2 y 3, perpendiculares a la de avance. Mientras tanto, las otras
BMUs no tienen efecto, a largo plazo, sobre el tensor MIL.
c) Como consecuencia de lo anterior aumenta la rigidez en direccion 1, ya que aumenta
la cantidad de material presente en las secciones normales a 1.
La conclusion es que un estado de tension uniaxial modifica la anisotropıa del hueso, ha-
ciendo que la direccion de carga se convierta en la de mayor rigidez. En definitiva, orientando
la rigidez con la carga y aprovechando el material de manera optima.
7.4.6. Evolucion del dano en el caso anisotropo
La acumulacion de dano microestructural en el tejido oseo es un proceso que tambien se
considera en el modelo anisotropo, ya que tiene un papel fundamental en la remodelacion
osea.
Como consecuencia de un estado de tensiones elevado puede aumentar el dano del tejido
oseo a nivel local. Este dano activa la remodelacion [31, 216, 217, 241, 279, 332], porque de
alguna manera es percibido por los osteocitos. Expresado como lo harıa la teorıa inhibitoria,

7.4 Modelo anisotropo de BMUs 269
se podrıa decir que el dano interrumpe la transmision de la senal inhibitoria a traves de la
red de osteocitos. En cuanto las celulas de borde dejan de sentir esta senal se activan las
BMUs, que primero reabsorben el tejido danado para despues sustituirlo por tejido intacto
en la fase de formacion, con lo que el dano quedarıa reparado. Este continuo proceso de
aparicion y reparacion de dano puede alcanzar una situacion de equilibrio si el dano no
aumenta excesivamente rapido y da tiempo a ser reparado por remodelacion, pero si el dano
se acumula muy rapidamente, puede producir una fractura por fatiga.
Como ya se dijo, el tensor de dano mide la degradacion del material. En concreto, su
proyeccion en una determinada direccion mide la perdida de rigidez en dicha direccion. Si
se mide el dano en una direccion por medio del volumen de tejido que contiene grietas
perpendiculares a esa direccion, es obvio que el volumen danado y la perdida de rigidez
direccionales estan relacionados entre sı, como lo estaban h y d en el caso isotropo (ver
ecuacion (7.32)).
La variacion del tensor de dano, ∆d, se evalua considerando los procesos de acumulacion
y reparacion de dano de forma separada, mediante los respectivos tensores ∆dA y ∆dR.
Para la acumulacion de dano se usan las mismas ecuaciones que usaron Garcıa-Aznar et
al. [117] en el modelo isotropo. Sin embargo, hay una diferencia fundamental y es que se
supone que solo las deformaciones de traccion producen dano y que si un trozo de hueso
esta sometido a compresion, acumula dano por las deformaciones de traccion que tiene por
efecto Poisson.
Hipotesis 4 El tensor de acumulacion de dano por fatiga, ∆dA es paralelo al tensor de
deformaciones. Ademas, se supone que para un estado de carga concreto, el dano aumenta
unica y exclusivamente en la direccion principal de maxima deformacion y si esa maxima
deformacion es positiva (de traccion). Si se denota por emax a esta direccion principal, ∆dA
se puede expresar del siguiente modo:
∆dA = ∆dA emax ⊗ emax (7.116)
donde ∆dA, es el incremento de dano en la direccion emax y se evalua usando las mismas
expresiones del caso isotropo.
Para calcular el incremento del dano no se utilizan las derivadas∂d
∂ndadas en las ecua-
ciones (7.51), como se hace en el modelo isotropo, por los problemas numericos que presentan.
Concretamente, estos problemas numericos aparecen solo en traccion, debido a la pendiente

270 Modelos mecanobiologicos
tan elevada que tiene la curva d − N para bajos numeros de ciclos (ver figura 7.9). Como
alternativa se utilizan directamente las leyes de evolucion del dano con el numero de cic-
los, presentadas en las ecuaciones (7.52). La ley de dano frente a deformacion y numero de
ciclos del caso isotropo se usa en el caso anisotropo, sin mas que sustituir la deformacion
equivalente, ε, de la ecuacion (7.47) correspondiente a traccion, por la deformacion principal
maxima en valor absoluto, εmax:
d = 1− γ2
√1
C3ln(eC3 − C2 ε
δ2max n) (7.117)
La interpretacion de esta ecuacion es la siguiente: un estado de deformacion cualquiera
provoca la iniciacion y el crecimiento de grietas perpendiculares a la direccion en la que
son mayores las deformaciones en valor absoluto (modo I). El efecto inmediato de esta
acumulacion de dano es la disminucion de la rigidez del hueso en direccion perpendicular a
las grietas, en la direccion predominante de carga. Precisamente, esa disminucion de rigidez
es lo que mide d, que no es mas que la proyeccion del tensor de dano en la direccion principal
de deformacion emax.
Para la vida a fatiga tambien se sustituye ε por εmax en las ecuaciones (7.53):
nF =1.445 · 1053
ε14.1max
(7.118)
Se utilizan las mismas expresiones para hueso cortical y para hueso trabecular. Esto es,
no se incluye el factor corrector de las deformaciones,E
E∗, como proponen Garcıa-Aznar
et al. [117] en el modelo isotropo de BMUs para distinguir uno y otro. Se recuerda que
dicho factor es la unidad en hueso cortical y en hueso trabecular, E representa modulo de
Young del tejido y E∗ un modulo de Young de referencia, del tejido cortical de mınima
porosidad. Estos autores proponen estas leyes de evolucion para predecir la acumulacion
de dano observada experimentalmente en los ensayos realizados por Pattin et al. [265] en
hueso cortical. Debido a la falta de datos experimentales en hueso esponjoso, dichos autores
propusieron la incorporacion de este factor corrector. La razon por la que no se incluye
en este trabajo es la siguiente: dicho termino se utilizaba para minorar las deformaciones
del hueso trabecular, muy grandes por lo general. Al multiplicar las deformaciones por el
modulo de Young lo que se esta haciendo en realidad es basar el calculo del dano en las
tensiones y no en las deformaciones. Sin embargo, aunque las tensiones macroscopicas que
soporta el tejido trabecular son pequenas no ocurre ası con las tensiones locales, a nivel de
las trabeculas. De hecho,E
E∗es proporcional a ρ2.58, segun la correlacion de Hernandez [144]

7.4 Modelo anisotropo de BMUs 271
y la relacion entre tensiones macroscopicas y tensiones locales es inversamente proporcional
a ρ2, segun Beaupre et al. [26], por lo que ambos efectos practicamente se compensan.
En cualquier caso, el hecho de que los resultados experimentales que sustentan estas
ecuaciones se obtuvieron en hueso cortical (por Pattin et al. [265]) hace que los resultados
obtenidos en hueso trabecular se deban tomar con cierta precaucion. En el capıtulo de resul-
tados se comenta uno de los problemas que han surgido, precisamente en la aplicacion del
modelo a un hueso trabecular. El dano simulado es excesivo y carente de sentido fisiologico.
Hay que tener en cuenta que los mecanismos de acumulacion de dano son completamente
distintos en hueso cortical y trabecular (ver figura 4.25), porque su microestructura es com-
pletamente distinta y la acumulacion de dano depende enormemente de la microestructura.
Una vez hecha esta salvedad se sigue el desarrollo del modelo de crecimiento de dano
describiendo la forma en que se evalua ∆dA mediante las expresiones (7.117), sin necesidad
de derivarlas. Hay que tener en cuenta que los experimentos en los que se basan estas
expresiones fueron realizados con una deformacion constante [265], algo que no ocurre en
la practica en un hueso real. Sin embargo, es posible responder a la siguiente pregunta:
¿que dano acumulara un material que ya posee un cierto dano d en la direccion emax, si se
somete a n ciclos de deformacion constante εmax? La respuesta es precisamente lo que se
esta buscando, ∆dA.
Previamente, mediante la expresion (7.117), se puede determinar cuantos ciclos de defor-
macion constante εmax hubieran sido necesarios para acumular el nivel de dano que tiene el
material actualmente, d, aunque ese dano haya sido acumulado, incluso reparado, siguiendo
otra evolucion. Ese numero de ciclos se denominara n y se puede determinar sustituyendo el
dano actual d y la deformacion εmax en la ecuacion (7.117) y despejando n. El nuevo dano
tras aplicar los n ciclos planeados sera entonces:
d+ ∆dA = 1− γ2
√1
C3ln(eC3 − C2 ε
δ2max (n+ n)) (7.119)
de donde se despeja inmediatamente ∆dA. Sustituyendo este valor en la ecuacion (7.116) se
obtiene el tensor de acumulacion de dano, ∆dA.
El proceso de remodelacion osea hace que disminuya el volumen de hueso danado, que es
reabsorbido y sustituido por tejido intacto. Se supondra que el volumen danado Vd esta uni-
formemente repartido en el volumen de tejido Vb y que el tensor de dano tambien se reparte
de manera uniforme en Vd. Se supondra igualmente que el dano disminuye en la misma

272 Modelos mecanobiologicos
proporcion en todas las direcciones y que se repara mas dano cuanto mas dano haya, lo cual
parece logico. Ası:
∆dR =∆ vR
vbd (7.120)
Por ultimo, la velocidad de variacion del dano se obtiene teniendo en cuenta el aumento
por fatiga y la disminucion por reparacion:
∆d = ∆dA −∆dR (7.121)
Y el tensor de dano se actualiza de manera sencilla:
d(t+ ∆t) = d(t) + ∆d(t) (7.122)
En funcion del dano d se establece la condicion de rotura local del material. Esta puede
definirse como aquella situacion en la que un diferencial de volumen no puede absorber mas
energıa de deformacion. En el caso isotropo, esto ocurre cuando el parametro de dano, d,
alcanza el valor unidad, lo que equivale a decir que su rigidez es nula (ver ecuacion (7.30))
y por tanto, tambien lo es su capacidad de absorber energıa de deformacion.
Ahora, esta condicion hay que matizarla por el caracter anisotropo del dano. Cuando un
autovalor del tensor d alcanza el valor unidad, es nula la capacidad de absorber energıa de
deformacion en la direccion principal de dano correspondiente. Sin embargo, no tiene por
que serlo en las otras direcciones. Para entender esto conviene recordar la interpretacion del
dano hecha con anterioridad. Imagınese una sola grieta plana macroscopica en el seno de un
material. El dano en la cara de la grieta es unidad en la direccion perpendicular al plano de
grieta y nulo en las otras direcciones. Ası, si el material se carga en la direccion perpendicular
al plano de grieta, las tensiones en la cara de la grieta son nulas y la region donde la energıa
de deformacion alta rodea la grieta. Sin embargo, si se carga en direccion paralela al plano
de la grieta, el material se comporta igual que si estuviera intacto. De hecho lo esta en las
otras dos direcciones, con la interpretacion hecha aquı del tensor de dano.
Como conclusion, se puede decir que para que un punto este totalmente danado, el tensor
de dano debe ser igual al tensor unidad de segundo orden. Entonces, su rigidez sera nula en
cualquier direccion y tambien lo sera, por tanto, la energıa de deformacion absorbida por
dicho punto. La condicion de rotura local expresada en terminos del parametro h es segun
la ecuacion (7.74), h = k, identica condicion a la del caso isotropo. Por tanto, la constante
k se puede interpretar como la fraccion de tejido danado que conduce a la rotura local. Este

7.4 Modelo anisotropo de BMUs 273
valor no se puede superar ya que en el algoritmo se limitan los autovalores del tensor de
dano, para que no superen la unidad.
7.4.7. Propiedades elasticas del hueso
El principal objetivo de este modelo de remodelacion osea es establecer como cambian las
propiedades elasticas del hueso durante la actividad que desarrolla y para ello es necesario
conocer como varıan los factores de los que dependen dichas propiedades.
Las propiedades elasticas de un trozo de hueso dependen de la distribucion de poros, que
en este modelo se mide por medio del tensor L; del nivel dano, a traves del tensor d y de las
propiedades elasticas del tejido oseo. Como varıan los dos primeros durante la actividad del
hueso ha sido objeto de los epıgrafes anteriores. Queda por establecer como es la dependencia
entre esas variables y las propiedades del trozo de hueso. Pero tambien resta aun por definir
como son las propiedades elasticas del tejido oseo que compone el trozo de hueso. Estas
dependen principalmente de la porosidad y de la composicion de dicho tejido y en concreto
de la fraccion de ceniza, α, que varıa a lo largo del proceso de mineralizacion. Para relacionar
porosidad y fraccion de ceniza con el modulo de Young del tejido se utiliza, como en el caso
isotropo, la correlacion obtenida experimentalmente por Hernandez et al. [144,145]:
E(MPa) = 84370 v2.58b α2.74 (7.123)
El proceso de mineralizacion ya fue descrito en el modelo isotropo y aquı se supon-
dra identica evolucion para el contenido mineral del tejido. La fraccion de ceniza promedio
en el instante t+ ∆t del volumen diferencial se puede aproximar por la ecuacion (7.60), en
funcion de la fraccion de ceniza promedio en el instante t y las cantidades de hueso formado
y reabsorbido en el intervalo ∆t
α(t+ ∆t) =α(t)((vb(t)− h(t))e−κ∆t −∆vR) + ∆vFα0
vb(t+ ∆t)− h(t+ ∆t)
+αmax(1− e−κ∆t)(vb(t)− h(t)) + ∆h(α(τ)− α0)
vb(t+ ∆t)− h(t+ ∆t)
+(1− e−κ∆t)
∫ t0(vR − h)(αmax − α(τ))dτ
vb(t+ ∆t)− h(t+ ∆t)
(7.124)
La unica diferencia entre el caso isotropo y el anisotropo estriba en la distinta forma que
ambos modelos tienen de evaluar: ∆vR, ∆vF y h(t).
A continuacion se incorpora el efecto de la porosidad. La influencia de la cantidad de
material y de la orientacion de los poros se tiene en cuenta de forma independiente. De esta

274 Modelos mecanobiologicos
forma lo hicieron Zysset et al. [363], como ya se comento al hablar del “fabric tensor” (ver
capıtulo 4). Estos autores relacionaron las propiedades elasticas del hueso con la densidad
aparente y las componentes del tensor MIL, normalizado con traza unitaria. Obtuvieron
experimentalmente las siguientes correlaciones para las constantes elasticas:
Ei = 17607 ρ3.2m3.2i
Eiνij
= 45800 ρ2.7m2.4i m2.4
j
Gij = 7799 ρ3.3m1.6i m1.6
j ,
(7.125)
expresiones muy similares a las ecuaciones (5.98) del modelo anisotropo basado en la Mecanica
del Dano y en las que se observa claramente que esta desacoplada la dependencia de la
porosidad y de la anisotropıa.
Ya se ha visto que la dependencia de las propiedades elasticas con la porosidad se tiene en
cuenta de manera indirecta, por medio de la fraccion volumetrica de hueso, como establece
la ecuacion (7.123). La dependencia con la orientacion del entramado oseo se establece
con el tensor MIL normalizado de traza unitaria, como hacen Zysset et al. [362, 363]. Sin
embargo, hay que observar que los exponentes a los que se elevan sus autovalores, mi, en
la ecuacion (7.125), son ligeramente diferentes a los del modelo basado en la Mecanica del
Dano (ecuacion (5.98)). Estos ultimos se obtienen de forma puramente teorica, mientras
que los exponentes de la ecuacion (7.125) se han obtenido mediante el ajuste por mınimos
cuadrados de resultados experimentales. Parece, en principio, mas apropiado usar estos
ultimos. Sin embargo hay que tener en cuenta que la interpretacion del tensor MIL que se
ha hecho en este trabajo es distinta de la que hacen Zysset et al. [363]. Mientras que en
esta tesis el MIL representa la longitud promedio de poros, para Zysset et al. representa
la longitud promedio de las trabeculas. Ambas estan relacionadas y es posible encontrar
una correlacion experimental con esa otra interpretacion de mi, aunque evidentemente con
distintos exponentes y constantes. Por esa razon y dado que la diferencia entre los exponentes
de la ecuacion (7.125) y los del modelo teorico basado en la Mecanica del Dano, no es muy
importante, se ha optado por mantener los teoricos. Ası, las componentes del tensor de
flexibilidad de un hueso con cierta porosidad pero sin dano seran:
Diiii =1
Ei=
1
84370 v2.58b α2.74
1
(3mi)4
Diijj = −νijEi
= − ν
84370 v2.58b α2.74
1
(3mi)2 (3mj)2
Dijij =1
2Gij=
1 + ν
2 · 84370 v2.58b α2.74
1
(3mi)2 (3mj)2
(7.126)

7.4 Modelo anisotropo de BMUs 275
donde ν es el coeficiente de Poisson del tejido oseo, que se supone igual a 0.3 e independiente
de la porosidad [220]. Es necesario aclarar que las propiedades anteriores estan dadas en ejes
de ortotropıa y que estas coinciden con los ejes principales del tensor MIL en el hueso sin
dano microestructural.
Las ecuaciones (7.126) reproducen la correlacion de Hernandez el at. (ecuacion (7.123)),
si se tiene en cuenta que el tensor MIL esta normalizado con traza unitaria y se particulariza
para un material isotropo, en cuyo caso mi = 1/3 ∀i.A continuacion se incorpora efecto del dano microestructural al tensor de comportamien-
to. Para tenerlo en cuenta se sigue la teorıa de dano anisotropo de Cordebois y Sideroff [69],
de la misma forma que hacen Doblare y Garcıa [91] en el modelo de remodelacion osea
anisotropo basado en la Mecanica del Dano (ver apartado 5.3.3), con la diferencia de que
allı la variable de dano medıa la porosidad y no el dano microestructural. Se define en primer
lugar la tension efectiva σ en funcion del tensor de dano y la tension real σ, de tal forma
que conserva la simetrıa,
σ = (1− d)−1/2 σ (1− d)−1/2 (7.127)
La ecuacion constitutiva del dano establece que la tension efectiva es aquella que aplicada
al material sin dano, darıa lugar a una energıa de deformacion igual a la que existe realmente
en el material danado, sujeto a las tensiones reales. Esto es:
U(σ,d) = U(σ, 0) (7.128)
Si se supone un comportamiento elastico lineal, la ecuacion anterior conduce al siguiente
tensor de flexibilidad del material poroso y danado, D:
D = R−1 R−1 DR−1 R−1 (7.129)
donde D es el tensor de flexibilidad del material poroso pero sin dano, calculado por medio
de las ecuaciones (7.126) y R = (1− d)1/2.
Demostracion 3 Si el comportamiento es elastico lineal, el primer miembro de la ecuacion
(7.128) se puede escribir
U(σ,d) =1
2σ : D : σ, (7.130)
y el segundo,
U(σ, 0) =1
2σ : D : σ =
1
2(R−1σR−1) : D : (R−1σR−1) (7.131)

276 Modelos mecanobiologicos
1 1
1+ ε 1+ ε
σ σ
σ = D : ε σ = D : ε
1
Figura 7.17: Representacion esquematica del concepto de tension efectiva
donde se ha usado la ecuacion (7.127) para expresar σ y la definicion que se acaba de hacer
de R. Igualando ambas y expresandolas en notacion de ındices, se tiene:
1
2σij Dijkl σkl =
1
2(R−1
mr σrsR−1sn ) Dmnpq (R−1
pt σtuR−1uq ) (7.132)
En un material elastico, el tensor de flexibilidad se puede obtener derivando la energıa de
deformacion respecto al tensor de tensiones dos veces, Dijkl =∂U
∂σij∂σkl. Si se tiene en
cuenta que R es simetrico (porque lo es el tensor de dano d, tal como se definio) y se deriva
la ecuacion anterior, se obtiene, tras una serie de operaciones elementales:
Dijkl = R−1im R−1
jn Dmnpq R−1pk R
−1ql (7.133)
expresion que escrita en forma compacta coincide con la ecuacion (7.129).
Con la expresion anterior se tiene en cuenta la influencia del dano en las propiedades
elasticas del material. Sustituyendo (7.126) en (7.129) se obtienen las constantes elasticas
del hueso real, teniendo en cuenta dano y porosidad.
Hay que observar de las ecuaciones (7.126) y (7.130) que el hueso es anisotropo, en el caso
mas general. Serıa ortotropo si no hubiera dano, en cuyo caso, las direcciones de ortotropıa
coincidirıan con las direcciones principales del tensor MIL. Sin embargo este no sera paralelo

7.4 Modelo anisotropo de BMUs 277
al tensor de dano, por lo general, y no se conserva la ortotropıa, mas que en aquellos casos
en que el dano se mantenga en valores muy pequenos o sea paralelo al tensor MIL. Y eso
es precisamente lo que tiende a ocurrir a largo plazo. En efecto, el dano esta alineado con
la deformacion, como se ve en la ecuacion (7.116) y la estructura osea tiende a alinearse
con ella a largo plazo. En la figura 7.18 se muestra un diagrama de flujo del modelo de
remodelacion propuesto en esta tesis.
Props. material Cargas y geometrıa
MEF
DeformacionesTensiones
εmax Estımulo, ξ
ξiso
Senalinhibitoria, si
fbio
Est. equilibrio, ξ ∗
Balance localde hueso
fbb
Aumento de danopor fatiga
Dano, d BMUs activadasdiso
Sv
Reparto deBMUs, Ni
Reparacionde dano
LR
LF L = LR − LF
vR vF
Fraccion vol.de hueso vb
Contenido de mineral α
L(t +∆t) = L(t)+ L∆t
Hueso trabecular
M =L
tr(L)
1
Figura 7.18: Diagrama de flujo del modelo de remodelacion propuesto en esta tesis.

278 Modelos mecanobiologicos
7.5. Aplicacion del modelo propuesto a algunos casos
sencillos. Analisis de sensibilidad de los parametros
En esta seccion se aplicara el modelo desarrollado en esta tesis a algunos casos sencillos
para analizar el efecto que tienen las hipotesis que se han hecho y los parametros que se
han introducido. Este analisis se hara con la finalidad de que el modelo reproduzca numeri-
camente algunos de los cambios que sufre el tejido oseo bajo determinadas circunstancias,
como pueden ser el desuso, una sobrecarga que produzca un aumento considerable del dano,
un cambio en la orientacion de las cargas, etc.
Para analizar la respuesta del modelo se utiliza un unico elemento con condiciones de
contorno que simulen estados de deformacion constante o tension constante segun el caso.
En todas las simulaciones se parte de una situacion de equilibrio de remodelacion. Esto
significa que el estımulo externo aplicado es igual al estımulo de equilibrio y que desde hace
un tiempo suficiente no se producen cambios en la frecuencia de activacion de BMUs, con
lo que la poblacion de BMUs permanece estable.
Se han llevado a cabo tres tipos de simulaciones (ver tabla 7.1).
Simulacion Estado inicial Estado de cargas Estado final
Isotropa isotropo isotropo isotropo
Anisotropa isotropo anisotropo A anisotropo 1
Cambio de anisotropıa anisotropo 1 anisotropo B anisotropo 2
Tabla 7.1: Resumen de las simulaciones realizadas en este apartado.
En las simulaciones isotropas se parte de una situacion inicial isotropa en equilibrio
de remodelacion y se aplica una deformacion (o tension en otros casos) hidrostatica, que
hara que el material mantenga la isotropıa al remodelar. Estos casos no se diferencian en
nada del caso isotropo y sus resultados se comparan con los que proporciona el modelo
isotropo de BMUs.
Que la situacion inicial sea isotropa significa que tanto la porosidad, como la actividad
de las BMUs, como el dano, son isotropos. El dano, en concreto, se supone nulo inicialmente.
Como ademas se parte de una situacion de equilibrio de remodelacion, la situacion de partida
isotropa sera la que indica la tabla 7.2:

7.5 Aplicacion del modelo propuesto a algunos casos sencillos 279
Situacion inicial isotropa
d = 0
α = α0
ρ = ρ0
vb0 =ρ0
ρt=
ρ0
1.41 + 1.29 α0
L0 = v1/3
b0 I
ξ0 = ξ∗0
Ni0 =1
3fbio (1 −
ξ0
ξ0 + c) Sv(ρ0) ∀ i
Tabla 7.2: Variables del modelo de remodelacion en la situacion inicial isotropa.
En las simulaciones anisotropas se parte de una situacion inicial isotropa y se aplica
un estado de deformacion con una componente desviadora no nula. El hueso modifica su
estructura interna hasta alcanzar un nuevo equilibrio de remodelacion con un cierto grado
de anisotropıa.
A partir de este ultimo estado se cambia la orientacion de las cargas en las simulaciones
denominadas “cambio de anisotropıa”. Se produce una redistribucion de los tejidos y se
alcanza un nuevo equilibrio de remodelacion con una orientacion diferente del material.
7.5.1. Simulacion isotropa
Se han simulado cuatro casos: desuso, sobrecarga moderada, sobrecarga muy alta, que
produce rotura inmediatamente, y sobrecarga alta, que produce una rotura a largo plazo. En
cada uno de estos casos hay parametros que influyen mas que otros. Los que tienen mayor
influencia se hacen variar en el analisis de sensibilidad que se realiza a continuacion.
Desuso. Influencia de fbio, fbb,min, ϕ y de la duracion de los perıodos de la se-
cuencia ARF.
Se simula en este caso un completo desuso, en el que la deformacion es nula en todas las
direcciones. Esta situacion hace que la senal inhibitoria sea nula, cualquiera que sea el dano,
(ver ecuacion (7.76)), aunque en este caso concreto de deformacion nula el dano se mantiene
1Este valor implica que el perıodo de mineralizacion secundaria dura 6 anos. Se considera este perıodo
el que transcurre desde el final de la mineralizacion primaria, cuando se tiene αini, hasta que se alcanza el
95 % del maximo contenido mineral, αmax.

280 Modelos mecanobiologicos
Parametro Valor nominal Otros valores usados
N Numero de ciclos 500 -
m Exponente de Carter 4 a -
k Razon volumen danado - degradacion 0.000333 b -
c Parametro de activacion por estımulo 0.0025 b -
a Exponente de activacion por dano 10 b 0, 40
fbio Factor biologico de activacion 0.03BMUs
mm2 dıab 0.05, 0.1
fbb,min balance local direccional de hueso mınimo 0.9 0.7, 1.0
fbb,max balance local direccional de hueso maximo 1.1 1.0, 1.2
ξ∗0
Estımulo de equilibrio inicial 0.0025 b -
v Ancho de la zona de transicion 0.2 ξ∗0
b -
w Semiancho de la zona muerta 0.1 ξ∗0
-
ϕ Exponente de acomodacion del estımulo 0.004 0.002
TR Duracion del perıodo de reabsorcion 24 dıas c 60 dıas d
TI Duracion del perıodo de inversion 8 dıas c 56 dıas d
TF Duracion del perıodo de formacion 64 dıas c 175 dıas d
vBMU Velocidad de avance de la BMU 40 µm/dıa e 10 µm/dıa f
σL Vida de la BMU 100 dıas f -
dO Diametro de la osteona 0.076 mm b -
dH Diametro del canal haversiano 0.0145 mm b -
dE Profundidad de erosion de la hemiosteona 0.0491 mm b -
dE Ancho de la hemiosteona 0.152 mm b -
αini Fraccion de ceniza inicial 0.6 b -
α0 Fraccion de ceniza mınima 0.45 g -
αmax Fraccion de ceniza maxima 0.7 g -
κ Exponente de velocidad de mineralizacion 0.0009 g 1 -
Tabla 7.3: Valores nominales de los distintos parametros del modelo y valores alternativos analiza-
dos para estudiar su influencia. a Whalen et al. [343], b Garcıa-Aznar [117], c Hazelwood et al. [140],
d Eriksen et al. [98], e Jaworski y Lok [170], f Parfitt et al. [263], g Hernandez [144].

7.5 Aplicacion del modelo propuesto a algunos casos sencillos 281
nulo. Eso hace que se active el mayor numero posible de BMUs, que van a reabsorber
un tejido que no esta recibiendo carga alguna. El numero de BMUs que se activen va a
depender de la densidad, a traves de Sv, pero principalmente de fbio. La cantidad de hueso
reabsorbido por estas BMUs va a depender de fbb,min, que controlara la velocidad a la que
se va perdiendo hueso. Esta reabsorcion no se produce de manera ilimitada, puesto que la
adaptacion de la energıa de deformacion de referencia a la energıa de deformacion aplicada,
hara que, tras un cierto perıodo, el volumen de hueso deje de disminuir. Esto significa que
perıodos prolongados, o incluso permanentes, de inmovilidad no conducen a la perdida total
de masa osea, aunque sı a una perdida severa. En ello influye de manera fundamental la
velocidad a la que se produce dicha adaptacion de la energıa de deformacion de equilibrio,
es decir del parametro ϕ de la ecuacion (7.111). La influencia de todos estos parametros,
fbio, fbb,min y ϕ, se analiza en este apartado. Los valores nominales que se han tomado son
los siguientes: ϕ = 0.004, fbb,min = 0.9 y fbio = 0.03 BMUsmm2 dıa . Para estudiar la influencia de
cada parametro se prueban distintos valores, que se pueden consultar en la tabla 7.3, ceteris
paribus.
En la figura 7.19 se representa la evolucion de vb para diferentes valores de fbio y tres
tipos de tejido: cortical de alta densidad (ρ0 = 2.0 g/cm3), trabecular de densidad intermedia
(ρ0 = 1.0 g/cm3) y de baja densidad (ρ0 = 0.5 g/cm3), esto es, un hueso osteoporotico. La
primera consecuencia significativa que hay que destacar de esta figura es que el desuso
produce reabsorcion osea, algo sobradamente conocido. Ademas, se puede observar que la
reabsorcion mas acusada se produce en el hueso cortical y en el trabecular de densidad media.
Es especialmente llamativa en el hueso cortical para el caso fbio = 0.1, pero este valor de fbio
no tiene mucho sentido fisiologico como se vera enseguida. Para el hueso osteoporotico la
reabsorcion osea no es tan acusada, lo cual tiene sentido fisiologico. En efecto, en las personas
con osteoporosis, generalmente de avanzada edad, son usuales los estados de desuso, o al
menos de cargas pequenas, y se sabe de estudios clınicos que estas personas no pierden por
completo la poca masa osea que les queda. La razon por la que el modelo predice menos
reabsorcion en hueso osteoporotico, se encuentra en las ecuaciones, en concreto, en la relacion
de proporcionalidad que se ha establecido entre la frecuencia de activacion de BMUs, for y
la superficie especıfica de poros, Sv. El maximo de esta funcion, que depende exclusivamente
de la porosidad, se produce para el hueso trabecular de densidad media y se mantiene en
valores altos para el hueso cortical, excepto para un hueso muy compacto (ver figura 5.11).
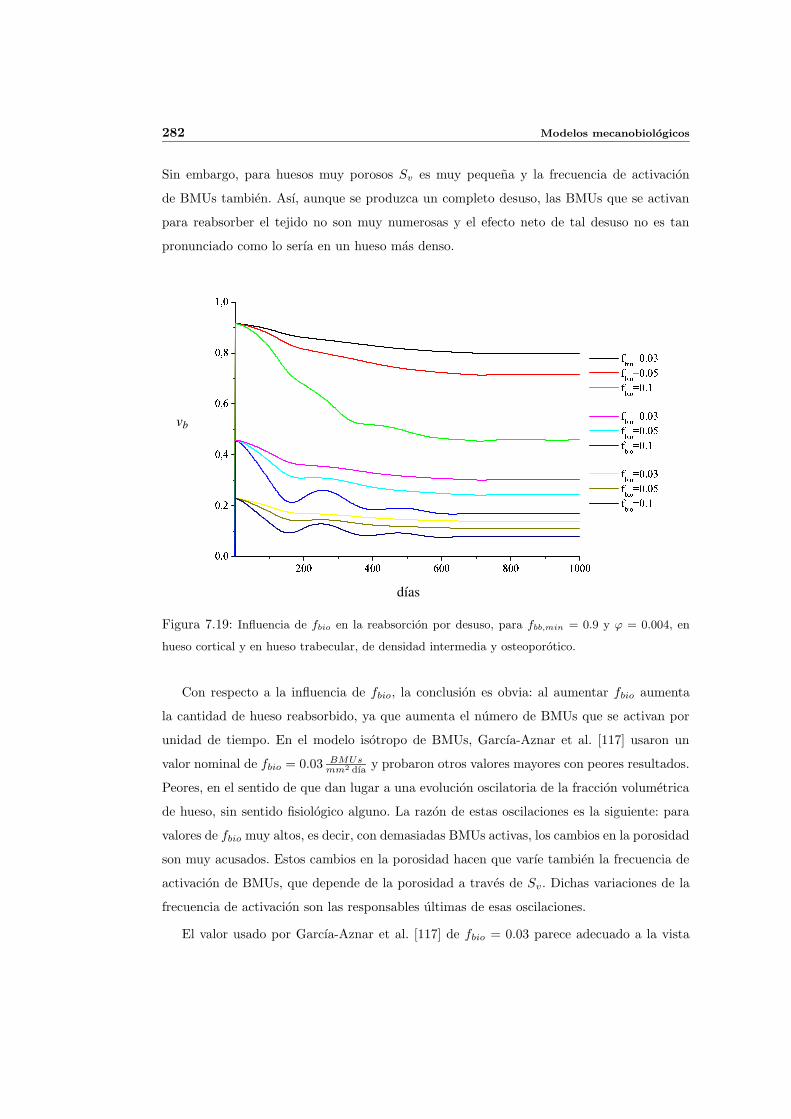
282 Modelos mecanobiologicos
Sin embargo, para huesos muy porosos Sv es muy pequena y la frecuencia de activacion
de BMUs tambien. Ası, aunque se produzca un completo desuso, las BMUs que se activan
para reabsorber el tejido no son muy numerosas y el efecto neto de tal desuso no es tan
pronunciado como lo serıa en un hueso mas denso.
(*)+) ,*)+) -+)+) ./)+) 01)+)2))+3 )
)+3 (
)+3 ,
)+3 -
)+3 .
013 )
4 5 6 7 8 ):9 )+;4 5 6 7 8 ):9 )2<4 5 6 7 8 ):9 0
4 5 6 7 8 ):9 )+;4 5 6 7 8 ):9 )2<4 5 6 7 8 ):9 0
4 5 6 7 8 ):9 )+;4 5 6 7 8 ):9 )2<4 5 6 7 8 ):9 0
dıas
vb
1
Figura 7.19: Influencia de fbio en la reabsorcion por desuso, para fbb,min = 0.9 y ϕ = 0.004, en
hueso cortical y en hueso trabecular, de densidad intermedia y osteoporotico.
Con respecto a la influencia de fbio, la conclusion es obvia: al aumentar fbio aumenta
la cantidad de hueso reabsorbido, ya que aumenta el numero de BMUs que se activan por
unidad de tiempo. En el modelo isotropo de BMUs, Garcıa-Aznar et al. [117] usaron un
valor nominal de fbio = 0.03 BMUsmm2 dıa y probaron otros valores mayores con peores resultados.
Peores, en el sentido de que dan lugar a una evolucion oscilatoria de la fraccion volumetrica
de hueso, sin sentido fisiologico alguno. La razon de estas oscilaciones es la siguiente: para
valores de fbio muy altos, es decir, con demasiadas BMUs activas, los cambios en la porosidad
son muy acusados. Estos cambios en la porosidad hacen que varıe tambien la frecuencia de
activacion de BMUs, que depende de la porosidad a traves de Sv. Dichas variaciones de la
frecuencia de activacion son las responsables ultimas de esas oscilaciones.
El valor usado por Garcıa-Aznar et al. [117] de fbio = 0.03 parece adecuado a la vista

7.5 Aplicacion del modelo propuesto a algunos casos sencillos 283
de los resultados. El valor fbio = 0.05 produce mucha reabsorcion y quiza sea mas adecuado
para jovenes, con mayor frecuencia de activacion de BMUs, como indica Frost [107]. Por
ultimo, el valor fbio = 0.1 resulta excesivo por lo que se acaba de comentar: las oscilaciones
que produce, carentes por completo de sentido fisiologico.
En relacion con esto, hay que decir que Hazelwood et al. [140] suponen en su modelo
de remodelacion, descrito en este capıtulo, una frecuencia maxima de activacion de BMUs
bastante mayor, en concreto, 0.5 BMUsmm2 dıa . En el modelo desarrollado en esta tesis, la frecuencia
maxima de activacion se alcanza cuando la senal inhibitoria es nula y en ese caso vale fbio ·Sv.El valor maximo de Sv esta en torno a 4, correspondiente a un hueso trabecular denso y en
tal caso, para alcanzar el valor utilizado por Hazelwood et al. habrıa que suponer un valor de
fbio > 1, lo que conduce a resultados con poco sentido fisiologico, como ya se ha visto. Mas
fiable parece el valor de la frecuencia maxima de activacion, 0.14BMUs/mm2/dıa, medida
experimentalmente por Frost con tecnicas de histomorfometrıa en costillas humanas [107].
Este valor corresponde a valores de fbio en torno a 0.04, muy proximo a los dos propuestos
aquı: 0.03 para adultos y 0.05 para adolescentes.
A continuacion se analiza la influencia del parametro fbb,min. En la figura 7.20 se mues-
tra la evolucion de vb para tres valores de fbb,min, 1.0, 0.9 y 0.7. En ella se observa que
al disminuir fbb,min aumenta la reabsorcion. Hay que recordar que este factor minora la
actividad de los osteoblastos. Ası, en ausencia de energıa de deformacion, en cuyo caso
fbb = fbb,min, los osteoblastos encuentran mas dificultad para diferenciarse de sus celulas
precursoras. Habra pocos osteoblastos y por tanto la cantidad de nuevo osteoide formado
sera pequena. Por lo general sera menor que la cantidad de tejido reabsorbido por los os-
teoclastos y por tanto la porosidad aumentara. Cuanto menor es el valor de fbb,min, menor
es el numero de osteoblastos que aparecen, menor la cantidad de osteoide formado y, por
tanto, mayor el aumento de la porosidad o la disminucion de la cantidad de hueso, como se
quiera.
Resulta llamativo que con fbb,min = 1.0 haya reabsorcion, pero su explicacion es sencilla.
En definitiva, fbb,min = 1.0 significa que aunque la actividad de los osteoblastos este en su
valor mınimo, estos forman la misma cantidad de hueso que los osteoclastos reabsorbieron.
Parece logico pensar entonces, que no varıe la cantidad de hueso. Sin embargo, hay que tener
en cuenta que el desuso provoca tambien un aumento de la poblacion de BMUs (recuerdese la
expresion de la frecuencia de activacion (7.75)). Las nuevas BMUs, activadas por el desuso,
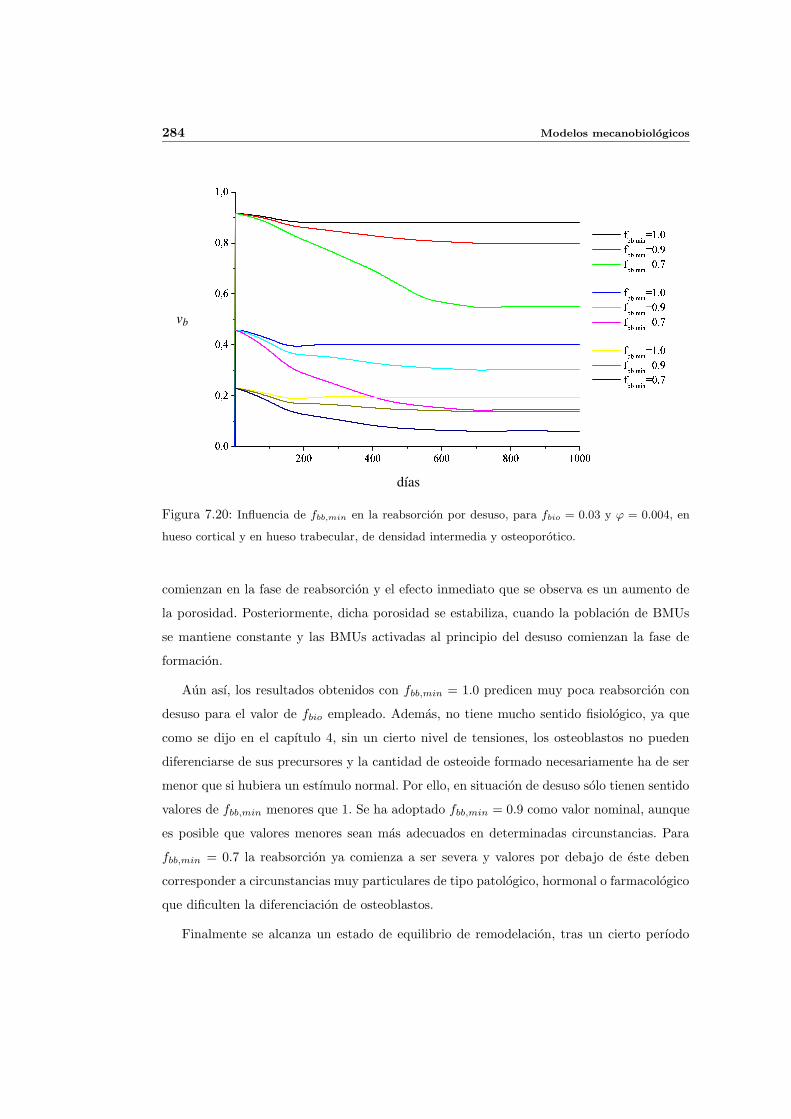
284 Modelos mecanobiologicos
=*>+> ?*>+> @+>+> A/>+> B1>+>2>>+C >
>+C =
>+C ?
>+C @
>+C A
B1C >
D E EGFIH J K BML >D E EGFIH J K >*L ND E EGFIH J K >*L O
D E EGFIH J K BML >D E EGFIH J K >*L ND E EGFIH J K >*L O
D E EGFIH J K BML >D E EGFIH J K >*L ND E EGFIH J K >*L O
dıas
vb
1
Figura 7.20: Influencia de fbb,min en la reabsorcion por desuso, para fbio = 0.03 y ϕ = 0.004, en
hueso cortical y en hueso trabecular, de densidad intermedia y osteoporotico.
comienzan en la fase de reabsorcion y el efecto inmediato que se observa es un aumento de
la porosidad. Posteriormente, dicha porosidad se estabiliza, cuando la poblacion de BMUs
se mantiene constante y las BMUs activadas al principio del desuso comienzan la fase de
formacion.
Aun ası, los resultados obtenidos con fbb,min = 1.0 predicen muy poca reabsorcion con
desuso para el valor de fbio empleado. Ademas, no tiene mucho sentido fisiologico, ya que
como se dijo en el capıtulo 4, sin un cierto nivel de tensiones, los osteoblastos no pueden
diferenciarse de sus precursores y la cantidad de osteoide formado necesariamente ha de ser
menor que si hubiera un estımulo normal. Por ello, en situacion de desuso solo tienen sentido
valores de fbb,min menores que 1. Se ha adoptado fbb,min = 0.9 como valor nominal, aunque
es posible que valores menores sean mas adecuados en determinadas circunstancias. Para
fbb,min = 0.7 la reabsorcion ya comienza a ser severa y valores por debajo de este deben
corresponder a circunstancias muy particulares de tipo patologico, hormonal o farmacologico
que dificulten la diferenciacion de osteoblastos.
Finalmente se alcanza un estado de equilibrio de remodelacion, tras un cierto perıodo
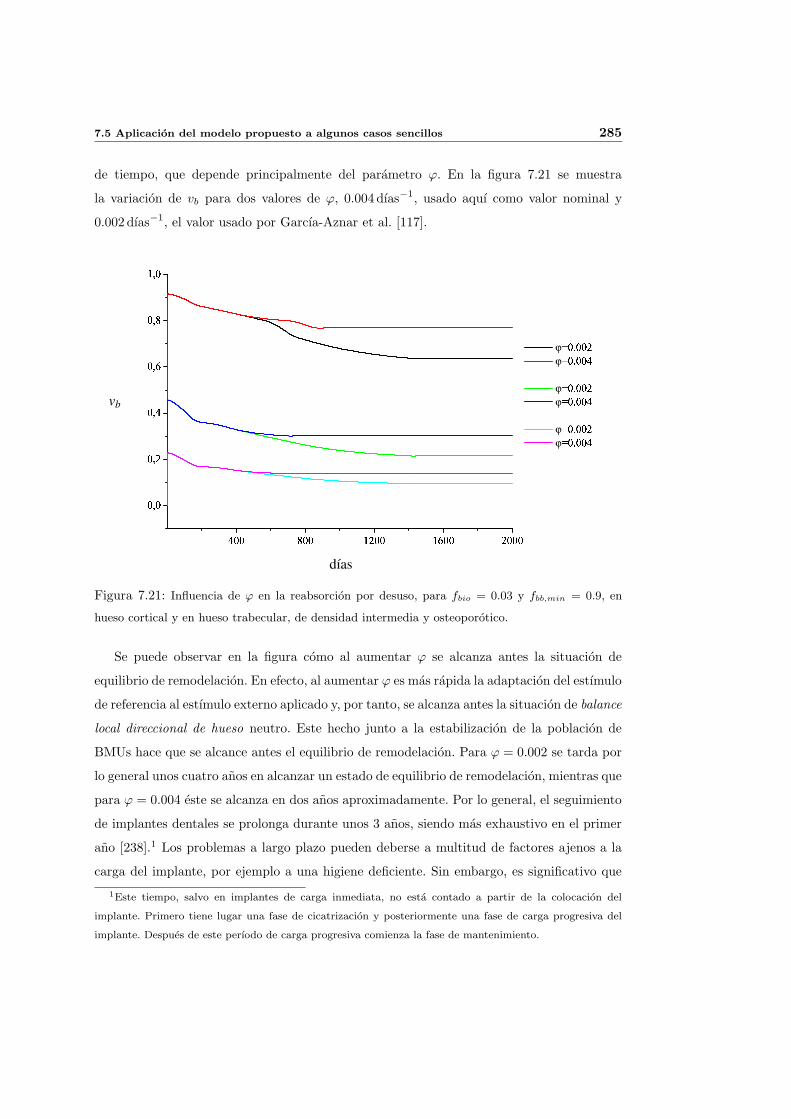
7.5 Aplicacion del modelo propuesto a algunos casos sencillos 285
de tiempo, que depende principalmente del parametro ϕ. En la figura 7.21 se muestra
la variacion de vb para dos valores de ϕ, 0.004 dıas−1, usado aquı como valor nominal y
0.002 dıas−1, el valor usado por Garcıa-Aznar et al. [117].
P*Q+Q R+Q+Q S1T2Q+Q S1U+Q2Q T2Q+Q2Q
Q+V Q
Q+V T
Q+V P
Q+V U
Q+V R
S1V Q
ϕ W Q*X Q2Q/Tϕ W Q*X Q2Q/P
ϕ W Q*X Q2Q/Tϕ W Q*X Q2Q/P
ϕ W Q*X Q2Q/Tϕ W Q*X Q2Q/P
dıas
vb
1
Figura 7.21: Influencia de ϕ en la reabsorcion por desuso, para fbio = 0.03 y fbb,min = 0.9, en
hueso cortical y en hueso trabecular, de densidad intermedia y osteoporotico.
Se puede observar en la figura como al aumentar ϕ se alcanza antes la situacion de
equilibrio de remodelacion. En efecto, al aumentar ϕ es mas rapida la adaptacion del estımulo
de referencia al estımulo externo aplicado y, por tanto, se alcanza antes la situacion de balance
local direccional de hueso neutro. Este hecho junto a la estabilizacion de la poblacion de
BMUs hace que se alcance antes el equilibrio de remodelacion. Para ϕ = 0.002 se tarda por
lo general unos cuatro anos en alcanzar un estado de equilibrio de remodelacion, mientras que
para ϕ = 0.004 este se alcanza en dos anos aproximadamente. Por lo general, el seguimiento
de implantes dentales se prolonga durante unos 3 anos, siendo mas exhaustivo en el primer
ano [238].1 Los problemas a largo plazo pueden deberse a multitud de factores ajenos a la
carga del implante, por ejemplo a una higiene deficiente. Sin embargo, es significativo que
1Este tiempo, salvo en implantes de carga inmediata, no esta contado a partir de la colocacion del
implante. Primero tiene lugar una fase de cicatrizacion y posteriormente una fase de carga progresiva del
implante. Despues de este perıodo de carga progresiva comienza la fase de mantenimiento.
286 Modelos mecanobiologicos
a los seis meses se realice una radiografıa de control. Esto hace indicar que los cambios que
se puedan producir en el hueso periimplantario ocurren, principalmente, dentro de esos seis
primeros meses, de manera que con esa radiografıa se puede detectar si esta teniendo lugar
un problema oseo. En la tabla 7.4 se muestran los resultados de vb en las cuatro simulaciones
de la figura 7.21, tras 6 meses y al final de la simulacion, tras 2000 dıas, una vez alcanzado
el equilibrio de remodelacion.
vb t=0 vb t=180 vb t=2000 (∆vb)t=180
t=0 (∆vb)t=2000
t=0
(∆vb)t=180
t=0
(∆vb)t=2000
t=0
cortical ϕ = 0.002 0.916 0.865 0.638 0.051 0.278 18 %
trabecular ϕ = 0.002 0.550 0.363 0.216 0.187 0.334 56 %
osteop. ϕ = 0.002 0.229 0.170 0.096 0.059 0.133 44 %
cortical ϕ = 0.004 0.916 0.865 0.770 0.051 0.146 35 %
trabecular ϕ = 0.004 0.550 0.363 0.302 0.187 0.248 75 %
osteop. ϕ = 0.004 0.229 0.170 0.139 0.059 0.090 66 %
Tabla 7.4: Fraccion volumetrica de hueso en el caso de desuso para ϕ = 0.002 y ϕ = 0.004, a los seis
meses y en el equilibrio de remodelacion. Fraccion volumetrica de hueso reabsorbido y porcentaje
de hueso reabsorbido en los seis primeros meses respecto al total reabsorbido.
De la tabla 7.4 se concluye que: para ϕ = 0.004 los cambios de vb que se producen tras
los seis primeros meses pueden servir para dar una idea aproximada de lo va a ocurrir a
largo plazo, excepto en hueso cortical, en el que sigue habiendo cambios significativos mas
alla de los seis meses (segun este modelo y para ϕ = 0.004). En cambio, para ϕ = 0.002 no
ocurre ası y los cambios posteriores a esos seis primeros meses siguen siendo importantes.
Suponiendo que esta fundamentada la toma de radiografıas a los seis meses, por ser repre-
sentativa de lo que ocurre a largo plazo, no serıa congruente usar ϕ = 0.002. Por esta razon
se toma ϕ = 0.004 como valor nominal.
Antes de pasar a otro caso de carga, se analiza la influencia que tiene en el modelo la
duracion de las distintas fases de la secuencia ARF. Los valores usados por Garcıa-Aznar et
al. [117] son muy elevados. En concreto, usan TR = 60, TI = 56 y TF = 175 dıas, tomados
de un estudio experimental en mujeres postmenopausicas con osteoporosis [98], que es un
caso demasiado especıfico. Se han usado aquı los valores que emplean Hazelwood et al. [140]
en su modelo, que estan basados en una serie mas amplia de estudios histomorfometricos
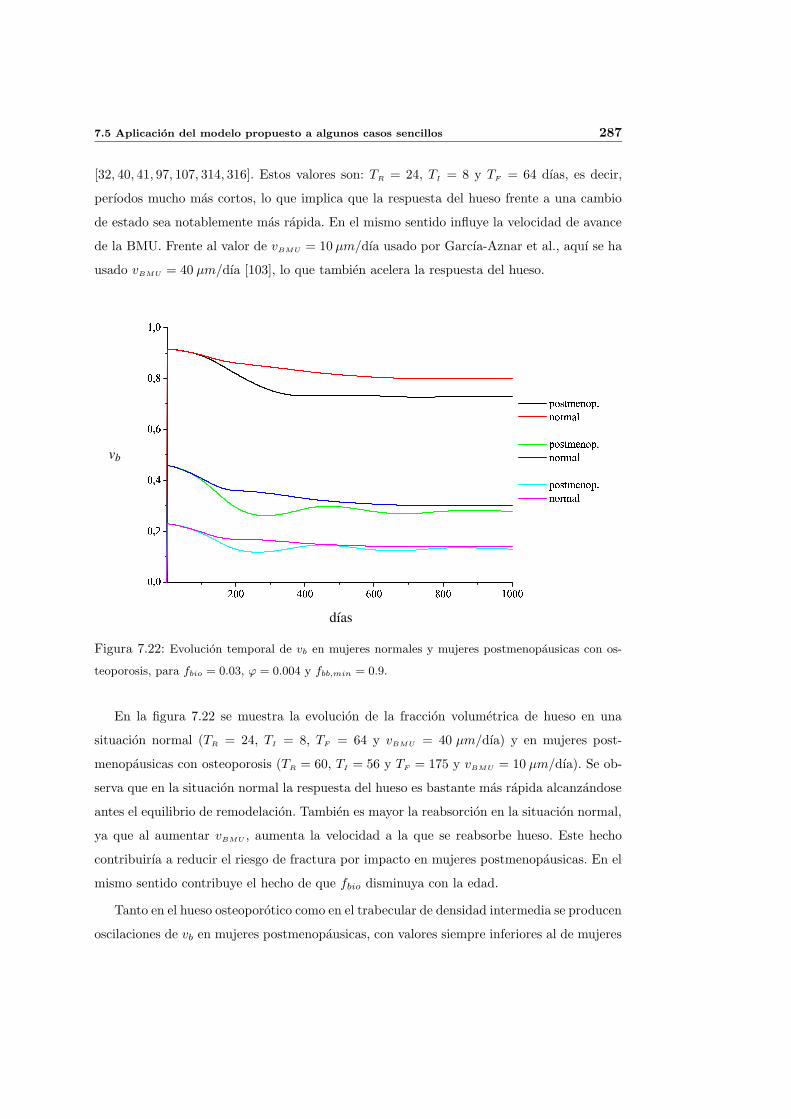
7.5 Aplicacion del modelo propuesto a algunos casos sencillos 287
[32, 40, 41, 97, 107, 314, 316]. Estos valores son: TR = 24, TI = 8 y TF = 64 dıas, es decir,
perıodos mucho mas cortos, lo que implica que la respuesta del hueso frente a una cambio
de estado sea notablemente mas rapida. En el mismo sentido influye la velocidad de avance
de la BMU. Frente al valor de vBMU = 10 µm/dıa usado por Garcıa-Aznar et al., aquı se ha
usado vBMU = 40 µm/dıa [103], lo que tambien acelera la respuesta del hueso.
Y*Z+Z [*Z+Z \+Z+Z ]/Z+Z ^1Z+Z2ZZ+_ Z
Z+_ Y
Z+_ [
Z+_ \
Z+_ ]
^1_ Z
`*a2bdc egf1h2a/`jih*aGk emlMn
`*a2bdc egf1h2a/`jih*aGk emlMn
`*a2bdc egf1h2a/`jih*aGk emlMn
dıas
vb
1
Figura 7.22: Evolucion temporal de vb en mujeres normales y mujeres postmenopausicas con os-
teoporosis, para fbio = 0.03, ϕ = 0.004 y fbb,min = 0.9.
En la figura 7.22 se muestra la evolucion de la fraccion volumetrica de hueso en una
situacion normal (TR = 24, TI = 8, TF = 64 y vBMU = 40 µm/dıa) y en mujeres post-
menopausicas con osteoporosis (TR = 60, TI = 56 y TF = 175 y vBMU = 10 µm/dıa). Se ob-
serva que en la situacion normal la respuesta del hueso es bastante mas rapida alcanzandose
antes el equilibrio de remodelacion. Tambien es mayor la reabsorcion en la situacion normal,
ya que al aumentar vBMU , aumenta la velocidad a la que se reabsorbe hueso. Este hecho
contribuirıa a reducir el riesgo de fractura por impacto en mujeres postmenopausicas. En el
mismo sentido contribuye el hecho de que fbio disminuya con la edad.
Tanto en el hueso osteoporotico como en el trabecular de densidad intermedia se producen
oscilaciones de vb en mujeres postmenopausicas, con valores siempre inferiores al de mujeres

288 Modelos mecanobiologicos
normales. Sin embargo, si se parte de un hueso osteoporotico apenas hay diferencias a largo
plazo. Esto querrıa decir que un estado de desuso tras la menopausia puede afectar al hueso
trabecular normal y convertirlo en osteoporotico, pero si ya se padece osteoporosis el desuso
no afecta de manera importante. Se podrıa decir que se ha llegado a una situacion lımite,
muy peligrosa para la integridad del hueso.
Sobrecarga moderada. Influencia de fbb,max
En este caso se simula un caso de sobrecarga moderada hidrostatica, sobre huesos de
tres densidades distintas: cortical de densidad intermedia (ρ0 = 1.7 g/cm3), trabecular de
densidad intermedia (ρ0 = 1.0 g/cm3) y trabecular de densidad muy baja (ρ0 = 0.5 g/cm3).
En el hueso cortical se ha aplicado una tension hidrostatica constante de 20MPa, que
produce inicialmente una deformacion de 734µε. Para los otros dos tipos de hueso, que
tienen una rigidez diferente, se ha aplicado igualmente una tension hidrostatica constante,
tal que produce la misma deformacion inicial: 734µε. En el hueso trabecular de densidad
intermedia la tension aplicada es 5MPa y en el osteoporotico 0.85MPa.
Como consecuencia de esta sobrecarga se produce un aumento de la fraccion volumetrica
de hueso, debido por un lado al descenso de la poblacion de BMUs y por otro a que las
que hay activas forman mas hueso del que reabsorben. El factor mas importante del que
depende la cantidad neta de hueso formado es el valor maximo del balance local direccional
de hueso, fbb,max. Que este parametro sea mayor que la unidad significa que los osteoblastos
forman mas hueso del que reabsorbieron los osteoclastos. Cuanto mayor es fbb,max, mayor
es la cantidad de hueso formado por los osteoblastos, siendo constante la cantidad de hueso
reabsorbido por los osteoclastos, que es igual a ABMU ·LBMU . En la figura 7.23 se analiza la
influencia de fbb,max en la fraccion volumetrica de hueso.
Se observa que la formacion neta de hueso es ligeramente mas acusada en el hueso
trabecular de densidad intermedia, de nuevo porque la activacion de BMUs se ve favorecida
en este tipo de hueso por la elevada superficie especıfica disponible, Sv. Para fbb,max = 1.0
(balance local direccional de hueso neutro) aumenta la cantidad de tejido, unicamente por
la disminucion de la poblacion de BMUs. Sin embargo, ese aumento es pequeno, ya que los
osteoblastos no forman mas que la misma cantidad de hueso reabsorbida por los osteoclastos.
Para fbb,max = 1.2 la formacion neta es excesiva y por esa razon se ha tomado como valor
nominal fbb,max = 1.1. En hueso cortical muy compacto se limita la cantidad de osteoide
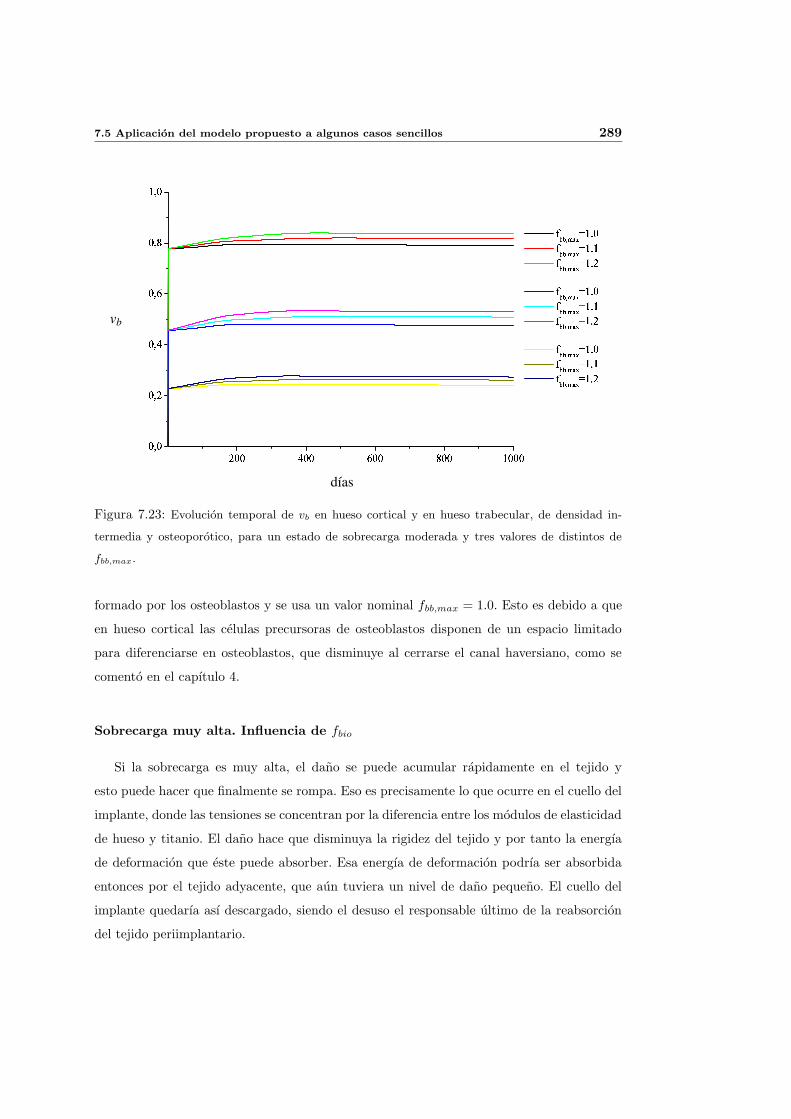
7.5 Aplicacion del modelo propuesto a algunos casos sencillos 289
o*p+p q*p+p r+p+p s/p+p t1p+p2pp+u p
p+u o
p+u q
p+u r
p+u s
t1u p
v w wx yIz |
tM pv w wx yIz |
tM tv w wx yIz |
tM o
v w wx yIz |
tM pv w wx yIz |
tM tv w wx yIz |
tM o
v w wx yIz |
tM pv w wx yIz |
tM tv w wx yIz |
tM o
dıas
vb
1
Figura 7.23: Evolucion temporal de vb en hueso cortical y en hueso trabecular, de densidad in-
termedia y osteoporotico, para un estado de sobrecarga moderada y tres valores de distintos de
fbb,max.
formado por los osteoblastos y se usa un valor nominal fbb,max = 1.0. Esto es debido a que
en hueso cortical las celulas precursoras de osteoblastos disponen de un espacio limitado
para diferenciarse en osteoblastos, que disminuye al cerrarse el canal haversiano, como se
comento en el capıtulo 4.
Sobrecarga muy alta. Influencia de fbio
Si la sobrecarga es muy alta, el dano se puede acumular rapidamente en el tejido y
esto puede hacer que finalmente se rompa. Eso es precisamente lo que ocurre en el cuello del
implante, donde las tensiones se concentran por la diferencia entre los modulos de elasticidad
de hueso y titanio. El dano hace que disminuya la rigidez del tejido y por tanto la energıa
de deformacion que este puede absorber. Esa energıa de deformacion podrıa ser absorbida
entonces por el tejido adyacente, que aun tuviera un nivel de dano pequeno. El cuello del
implante quedarıa ası descargado, siendo el desuso el responsable ultimo de la reabsorcion
del tejido periimplantario.
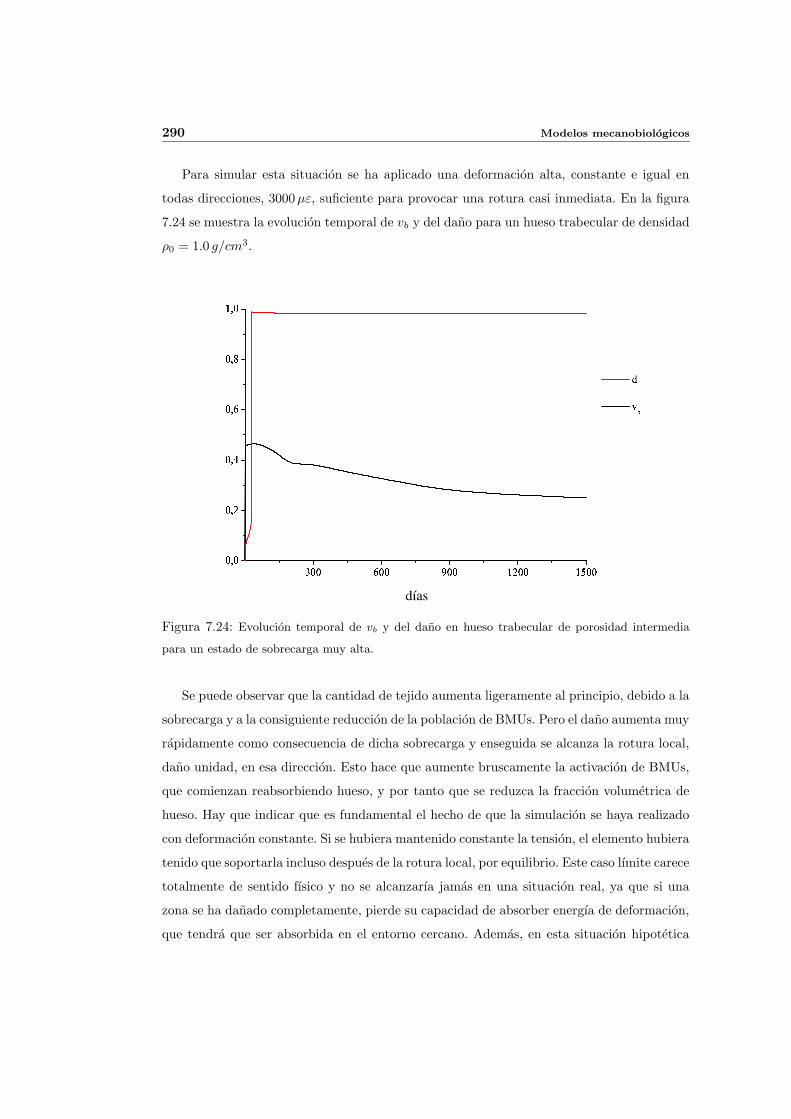
290 Modelos mecanobiologicos
Para simular esta situacion se ha aplicado una deformacion alta, constante e igual en
todas direcciones, 3000µε, suficiente para provocar una rotura casi inmediata. En la figura
7.24 se muestra la evolucion temporal de vb y del dano para un hueso trabecular de densidad
ρ0 = 1.0 g/cm3.
~++ ++ ++ 2+ I/++
+
+
+
+
1
/
dıas
1
Figura 7.24: Evolucion temporal de vb y del dano en hueso trabecular de porosidad intermedia
para un estado de sobrecarga muy alta.
Se puede observar que la cantidad de tejido aumenta ligeramente al principio, debido a la
sobrecarga y a la consiguiente reduccion de la poblacion de BMUs. Pero el dano aumenta muy
rapidamente como consecuencia de dicha sobrecarga y enseguida se alcanza la rotura local,
dano unidad, en esa direccion. Esto hace que aumente bruscamente la activacion de BMUs,
que comienzan reabsorbiendo hueso, y por tanto que se reduzca la fraccion volumetrica de
hueso. Hay que indicar que es fundamental el hecho de que la simulacion se haya realizado
con deformacion constante. Si se hubiera mantenido constante la tension, el elemento hubiera
tenido que soportarla incluso despues de la rotura local, por equilibrio. Este caso lımite carece
totalmente de sentido fısico y no se alcanzarıa jamas en una situacion real, ya que si una
zona se ha danado completamente, pierde su capacidad de absorber energıa de deformacion,
que tendra que ser absorbida en el entorno cercano. Ademas, en esta situacion hipotetica
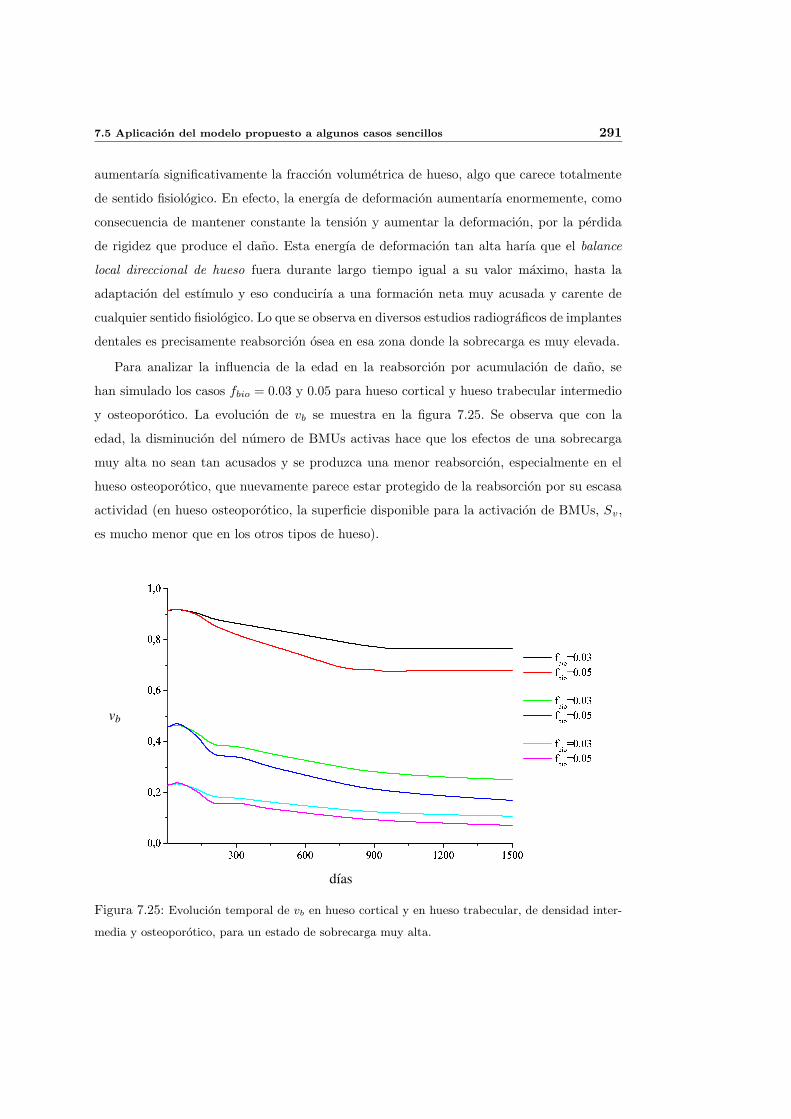
7.5 Aplicacion del modelo propuesto a algunos casos sencillos 291
aumentarıa significativamente la fraccion volumetrica de hueso, algo que carece totalmente
de sentido fisiologico. En efecto, la energıa de deformacion aumentarıa enormemente, como
consecuencia de mantener constante la tension y aumentar la deformacion, por la perdida
de rigidez que produce el dano. Esta energıa de deformacion tan alta harıa que el balance
local direccional de hueso fuera durante largo tiempo igual a su valor maximo, hasta la
adaptacion del estımulo y eso conducirıa a una formacion neta muy acusada y carente de
cualquier sentido fisiologico. Lo que se observa en diversos estudios radiograficos de implantes
dentales es precisamente reabsorcion osea en esa zona donde la sobrecarga es muy elevada.
Para analizar la influencia de la edad en la reabsorcion por acumulacion de dano, se
han simulado los casos fbio = 0.03 y 0.05 para hueso cortical y hueso trabecular intermedio
y osteoporotico. La evolucion de vb se muestra en la figura 7.25. Se observa que con la
edad, la disminucion del numero de BMUs activas hace que los efectos de una sobrecarga
muy alta no sean tan acusados y se produzca una menor reabsorcion, especialmente en el
hueso osteoporotico, que nuevamente parece estar protegido de la reabsorcion por su escasa
actividad (en hueso osteoporotico, la superficie disponible para la activacion de BMUs, Sv,
es mucho menor que en los otros tipos de hueso).
2+ 2+ ++ 22 I/2+
+
+
+
+
1
* + * 2
* + * 2
* + * 2
dıas
vb
1
Figura 7.25: Evolucion temporal de vb en hueso cortical y en hueso trabecular, de densidad inter-
media y osteoporotico, para un estado de sobrecarga muy alta.

292 Modelos mecanobiologicos
Sobrecarga alta. Influencia de fbio y del exponente de activacion por dano, a
En este apartado se simula una sobrecarga alta, pero no tan alta como para producir
una rotura inmediata. En concreto se aplica una deformacion constante e igual en todas
direcciones, de valor 2500µε. El dano aumenta progresivamente, aunque de forma mas lenta
que en el apartado anterior. Este dano moderado puede inducir la activacion de muchas
BMUs, dependiendo del valor del parametro a en la expresion de la senal inhibitoria, ecuacion
(7.76).
si(ξi, di) =ξi
ξi + c(1− dei
)a (7.76)
En la figura 7.26 se muestra la variacion de vb y el dano con esta sobrecarga en hueso
cortical, para el valor nominal de a = 10. Se observa al principio una formacion neta de hueso
apreciable, mientras el dano se acumula de forma progresiva. El dano acumulado crece mas
rapido de lo que da tiempo a repararse por remodelacion y finalmente se produce la rotura
tras unos 440 dıas de sobrecarga, aproximadamente. El fenomeno anterior es conocido en
atletas con sobrecarga de entrenamiento, que puede producir una acumulacion de dano a
nivel local, que conduzca finalmente a la rotura del hueso.
A continuacion se analiza la influencia del parametro a en este fenomeno. Dicho parametro
no tiene ninguna influencia a partir del momento de la rotura, cuando el dano se mantiene
igual a la unidad sin posibilidad de reparacion (ver ecuacion (7.76)). Pero hasta ese mo-
mento sı la tiene, de tal suerte que cuanto mayor es a, mayor es la influencia del dano en
la activacion de BMUs. Se puede decir que de las dos misiones que este modelo asigna a la
BMU, que se ven plasmadas en la ecuacion (7.76) y que son: retirar hueso de donde no es
necesario (estımulo pequeno) y reparar el dano microestructural; esta ultima adquiere mas
importancia al aumentar a. En las figuras 7.27 y 7.28 se muestra la evolucion de vb y del
dano hasta el momento de la rotura para a = 0 y a = 40 respectivamente, para compararlas
con el caso nominal, a = 10, de la figura 7.26.
El uso de a = 0 equivale a decir que la senal inhibitoria es independiente del dano,
es decir, que las BMUs no se activan al aparecer dano microestructural y por tanto, su
objetivo no es repararlo. Muchos autores han establecido que esto no es cierto, mas bien al
contrario, la reparacion del dano microestructural es uno de los principales objetivos de la
remodelacion osea y causa de la activacion de BMUs [92,117,140,274,279]. Sin embargo, si
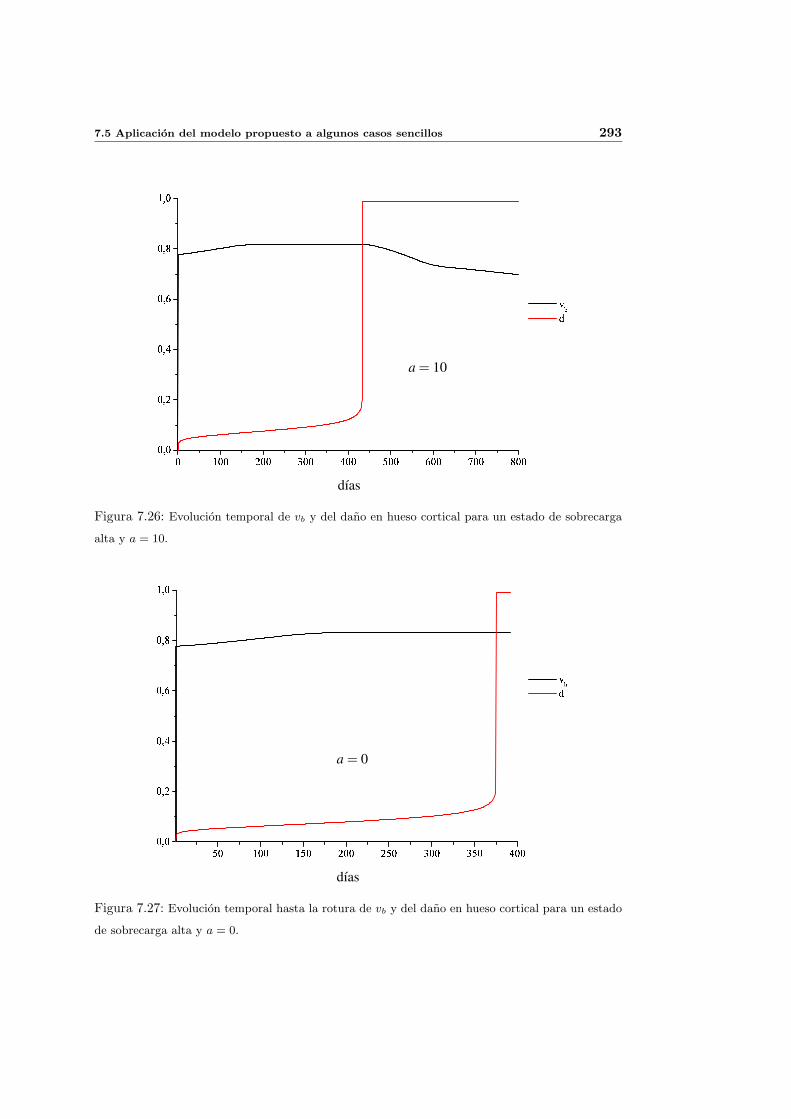
7.5 Aplicacion del modelo propuesto a algunos casos sencillos 293
1+ 2+ ++ 2+ ¡/+ ¢++ £++ ¤/++¥
+¥
+¥
+¥ ¢
+¥ ¤
1¥
¦/§¨
dıas
a = 10
1
Figura 7.26: Evolucion temporal de vb y del dano en hueso cortical para un estado de sobrecarga
alta y a = 10.
©/ª «1ª+ª «I©/ª ¬2ª+ª ¬*©/ª +ª+ª 2©/ª ®2ª+ªª+¯ ª
ª+¯ ¬
ª+¯ ®
ª+¯ °
ª+¯ ±
«1¯ ª
²/³´
dıas
a = 0
1
Figura 7.27: Evolucion temporal hasta la rotura de vb y del dano en hueso cortical para un estado
de sobrecarga alta y a = 0.
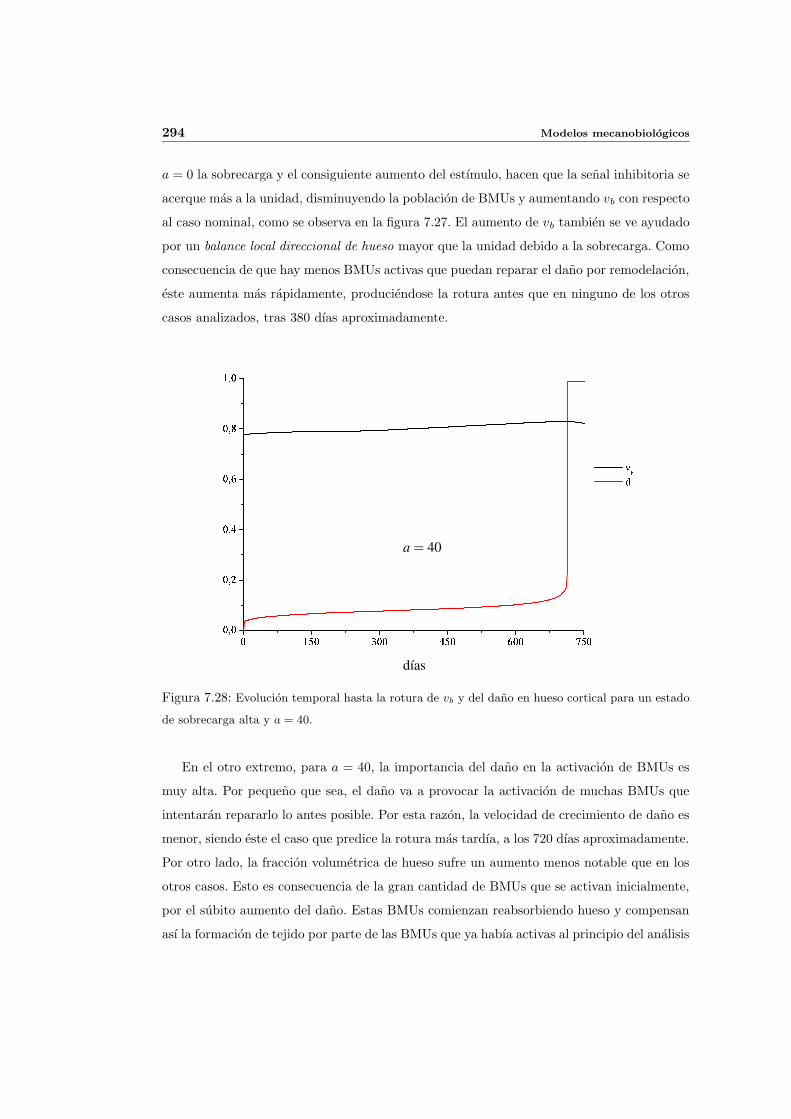
294 Modelos mecanobiologicos
a = 0 la sobrecarga y el consiguiente aumento del estımulo, hacen que la senal inhibitoria se
acerque mas a la unidad, disminuyendo la poblacion de BMUs y aumentando vb con respecto
al caso nominal, como se observa en la figura 7.27. El aumento de vb tambien se ve ayudado
por un balance local direccional de hueso mayor que la unidad debido a la sobrecarga. Como
consecuencia de que hay menos BMUs activas que puedan reparar el dano por remodelacion,
este aumenta mas rapidamente, produciendose la rotura antes que en ninguno de los otros
casos analizados, tras 380 dıas aproximadamente.
µ ¶I·/µ ¸+µ+µ ¹*·/µ º+µ+µ »2·/µµ+¼ µ
µ+¼ ½
µ+¼ ¹
µ+¼ º
µ+¼ ¾
¶1¼ µ
¿/ÀÁ
dıas
a = 40
1
Figura 7.28: Evolucion temporal hasta la rotura de vb y del dano en hueso cortical para un estado
de sobrecarga alta y a = 40.
En el otro extremo, para a = 40, la importancia del dano en la activacion de BMUs es
muy alta. Por pequeno que sea, el dano va a provocar la activacion de muchas BMUs que
intentaran repararlo lo antes posible. Por esta razon, la velocidad de crecimiento de dano es
menor, siendo este el caso que predice la rotura mas tardıa, a los 720 dıas aproximadamente.
Por otro lado, la fraccion volumetrica de hueso sufre un aumento menos notable que en los
otros casos. Esto es consecuencia de la gran cantidad de BMUs que se activan inicialmente,
por el subito aumento del dano. Estas BMUs comienzan reabsorbiendo hueso y compensan
ası la formacion de tejido por parte de las BMUs que ya habıa activas al principio del analisis

7.5 Aplicacion del modelo propuesto a algunos casos sencillos 295
y que ahora estan en fase de formacion, con un balance local direccional de hueso mayor que
la unidad, por el elevado estımulo. Cuanto mayor es el numero de BMUs activadas por la
aparicion de dano, menor es la formacion neta por sobrecarga.
7.5.2. Simulacion anisotropa
En las simulaciones que se presentan a continuacion se parte de una situacion inicial
isotropa y se aplica un caso de carga no hidrostatico, que modifica a largo plazo la anisotropıa
del material. El termino anisotropo esta usado en sentido general. En los casos concretos
aquı analizados, el material evoluciona hasta tener una simetrıa transversalmente isotropa
y ortotropa, segun el caso.
En primer lugar se simula un estado de traccion uniaxial, que hace que el tejido adquiera
una simetrıa transversalmente isotropa, siendo la direccion de traccion la direccion prefer-
ente, es decir, la direccion perpendicular al plano de isotropıa. Posteriormente, se analiza un
estado de compresion uniaxial. Por ultimo, se simula un estado de tensiones arbitrario, que
tiende a alinear el material con las direcciones principales de deformacion.1
Traccion uniaxial
El caso de carga propuesto es el siguiente:
a) σy = 20MPa y σx = σz = 0 para un tejido isotropo de densidad inicial ρ0 = 1.7 g/cm3,
correspondiente a un hueso cortical de densidad intermedia.
b) σy = 4MPa y σx = σz = 0 para un tejido isotropo de densidad inicial ρ0 = 1.0 g/cm3,
correspondiente a un hueso trabecular de densidad intermedia.
En las figuras 7.29 y 7.30 se muestra la evolucion de la fraccion volumetrica de hueso y de
los autovalores del tensor MIL. Las direcciones principales de dicho tensor coinciden con las
de la carga: la direccion principal y es la direccion de carga y las otras son cualesquiera dos
direcciones ortogonales contenidas en el plano de isotropıa. Se recuerda que los autovalores
del tensor MIL representan la longitud en la direccion principal correspondiente de un poro
promedio contenido en la unidad de volumen.
1A largo plazo, cuando se alcance el equilibrio de remodelacion, las direcciones principales de deformacion
coincidiran en cualquier caso con las de tension. En el caso concreto de que se parta de una situacion inicial
isotropa, el alineamiento entre tensiones y deformaciones ocurre desde el principio.
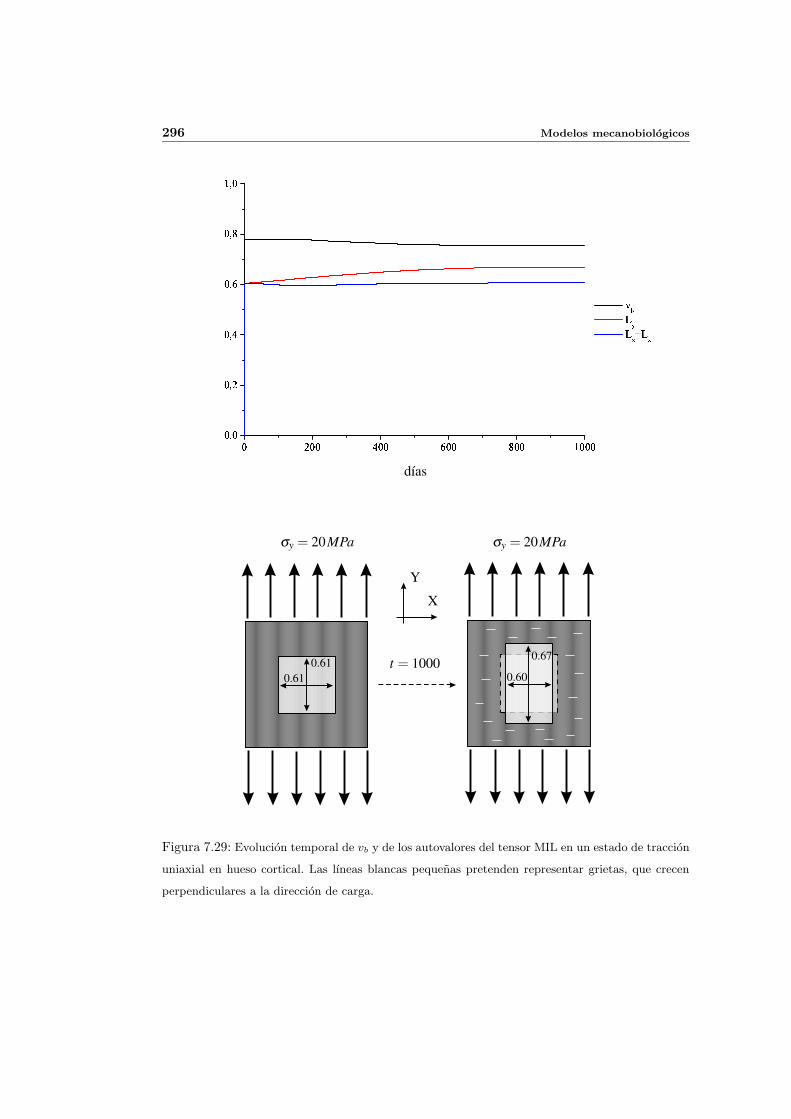
296 Modelos mecanobiologicos
 Ã2Â+ Ä2Â+ Å+Â+ Æ/Â+ Ç1Â+Â+ÂÂ/È Â
Â/È Ã
Â/È Ä
Â/È Å
Â/È Æ
ÇÈ Â
É/ÊËÌËÍ ÎÏËÐ
dıas
X
Y
t = 1000
σy = 20MPa σy = 20MPa
0.610.61
0.60
0.67
1
Figura 7.29: Evolucion temporal de vb y de los autovalores del tensor MIL en un estado de traccion
uniaxial en hueso cortical. Las lıneas blancas pequenas pretenden representar grietas, que crecen
perpendiculares a la direccion de carga.
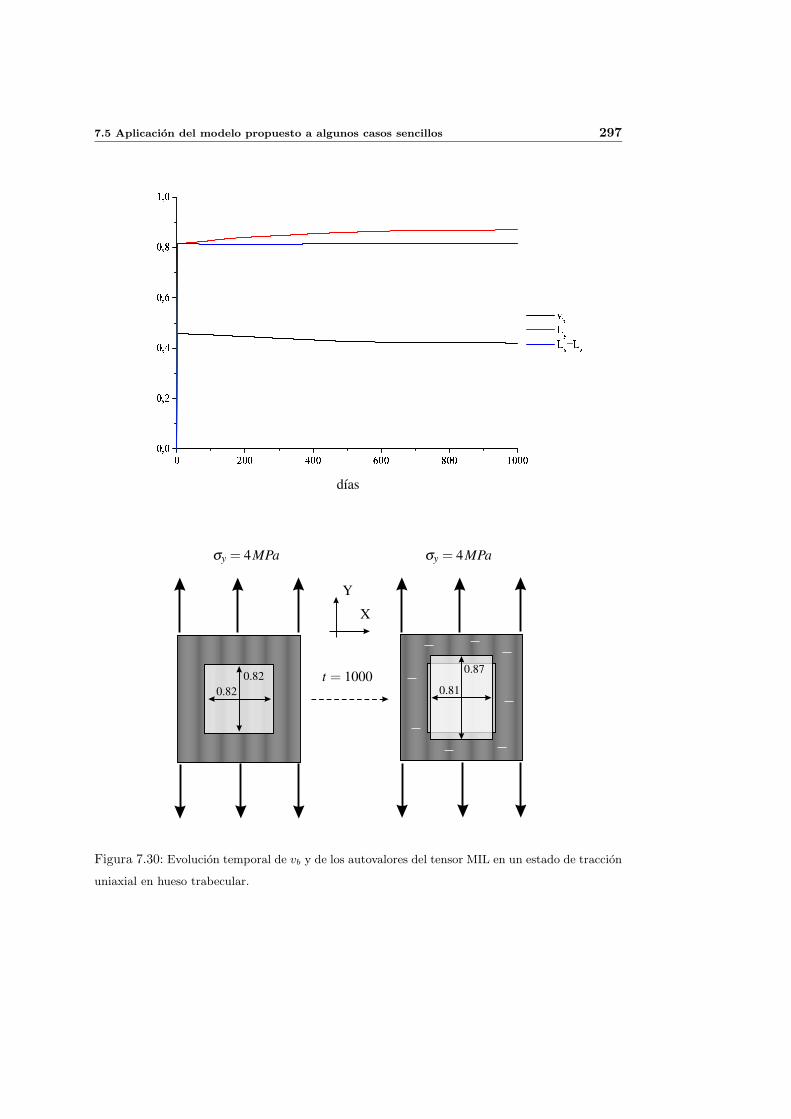
7.5 Aplicacion del modelo propuesto a algunos casos sencillos 297
Ñ Ò2Ñ+Ñ Ó2Ñ+Ñ Ô+Ñ+Ñ Õ/Ñ+Ñ Ö1Ñ+Ñ+ÑÑ/× Ñ
Ñ/× Ò
Ñ/× Ó
Ñ/× Ô
Ñ/× Õ
Ö× Ñ
Ø/ÙÚÛÚÜ ÝÏÚÞ
dıas
X
Y
t = 1000
σy = 4MPa σy = 4MPa
0.820.82
0.81
0.87
1
Figura 7.30: Evolucion temporal de vb y de los autovalores del tensor MIL en un estado de traccion
uniaxial en hueso trabecular.

298 Modelos mecanobiologicos
En estas figuras se observa como las longitudes de los poros van cambiando, permanecien-
do iguales en todo momento las longitudes en direccion z y x. Puesto que no hay distincion
entre ellas en lo que a cargas se refiere, tampoco la va a haber en las longitudes de poro,
en la anisotropıa o en las propiedades elasticas. Se alcanza, efectivamente, una situacion
de isotropıa transversal, muy tıpica en huesos que estan cargados preferentemente en una
direccion.
En la direccion de carga se alargan los poros, mientras que en las direcciones perpendicu-
lares disminuyen su longitud ligeramente, como se esquematiza en las figuras 7.29 y 7.30. De
esa forma aumenta la seccion resistente de material en la direccion en la que dicho material
se hace necesario: aumenta la rigidez en la direccion de la carga y disminuye en las otras dos,
en las que hay desuso. Igualmente, se observa que la fraccion volumetrica de hueso disminuye
al principio para recuperarse a largo plazo. Esto es debido a que inicialmente se activa un
buen numero de BMUs por el aumento brusco del dano en la direccion de la carga. Hay
que pensar que se parte de una situacion de dano nulo y que una carga de 4MPa produce
deformaciones considerables en un hueso trabecular como este (el dano en la direccion de la
carga sube rapidamente a dy = 0.026, que es un valor considerable). Ademas, la energıa de
deformacion es nula en las direcciones en las que no hay carga y por tanto la senal inhibitoria
tambien es nula, con lo que gran parte de las BMUs que se activan lo hacen avanzando en
estas direcciones, segun el reparto de la ecuacion (7.78). Como esas BMUs tienen un balance
local direccional de hueso menor que la unidad, su efecto es reducir vb. Pero una vez que el
estımulo de equilibrio se adapta al estımulo externo en las direcciones z y x, el balance local
direccional de hueso de esas BMUs se hace neutro. Ademas, por entonces, las BMUs que se
activaron al principio del analisis se encuentran en la fase de formacion y dado que en la
direccion de carga el estımulo es alto, su efecto neto es el aumento de vb. Cuando se llega
al equilibrio de remodelacion vb incluso supera, levemente eso sı, la fraccion volumetrica de
hueso inicial.
Resistencia a fatiga en compresion y en traccion uniaxial
Todos los casos simulados hasta ahora son de traccion. Se ha simulado tambien un caso
de compresion para comparar el distinto comportamiento exhibido por el tejido oseo en
uno y otro caso. Se recuerda que, con la ley de dano propuesta, solo se acumula dano
con deformaciones de traccion y por tanto, si se acepta dicha hipotesis, en un ensayo de

7.5 Aplicacion del modelo propuesto a algunos casos sencillos 299
compresion solo podrıa aumentar el dano por efecto Poisson. Esto hara que la resistencia a
fatiga en compresion sea bastante mayor.
Un tejido isotropo de densidad inicial ρ0 = 1.7 g/cm3 y dano nulo se ha sometido a una
carga uniaxial cıclica de amplitud constante σ = 26MPa y un coeficiente de asimetrıa R = 0,
en traccion, y R = −∞, en compresion. En las figuras 7.31 y 7.32 se muestra la evolucion
del dano y vb en traccion y en compresion respectivamente. El fallo en traccion se produce
casi inmediatamente, a los 100 dıas, es decir, tras 50000 ciclos, a razon de 500 ciclos por
dıa. En compresion, sin embargo, no se produce fallo. El dano se acumula muy lentamente
y la reabsorcion del tejido danado y su sustitucion por un nuevo tejido, frena el aumento
del dano, de forma que a partir de un cierto instante deja de crecer. Se puede concluir, si se
aceptan las hipotesis de este trabajo, que un estado de traccion es mas peligroso para el tejido
oseo desde el punto de vista resistente, ya que el dano se acumula muy rapidamente y si la
carga es suficientemente alta, no da tiempo a que se elimine dicho dano por remodelacion.
Si la compresion es mas alta tambien puede ocurrir que el dano se acumule mas rapi-
damente de lo que se elimina por remodelacion, pudiendo llevar al material a la rotura por
fatiga. Se ha simulado el mismo ensayo anterior pero con una carga superior, de amplitud
σ = 88MPa, produciendose el fallo tras 430 dıas, es decir, tras 215000 ciclos. A pesar de que
tambien se puede producir el fallo por compresion, evidentemente, la resistencia a fatiga en
compresion es mucho mayor que en traccion. Para esta misma amplitud de carga en traccion,
el fallo se produce de manera inmediata, ya que ese valor de tension esta por encima de la
resistencia a traccion para un hueso de esa densidad.
Estado arbitrario de carga
Se simula en este apartado un estado arbitrario de cargas partiendo de una situacion
inicial isotropa, con densidad inicial ρ0 = 1.7 g/cm3 y sin dano. El tejido evoluciona modif-
icando su anisotropıa, la longitud de sus poros, para adaptarse a la carga externa aplicada.
En concreto, alineara sus poros con las direcciones principales de tension y hara que su
rigidez sea maxima en la direccion mas solicitada, como se vera a continuacion.
El estado de cargas simulado es σx = 18MPa, σy = 0.04MPa y σz = −8MPa. En la
figura 7.33 se muestra la evolucion de la fraccion volumetrica de hueso y de los autovalores
del tensor MIL.
Se puede observar que una vez alcanzado el equilibrio de remodelacion, la direccion x, a
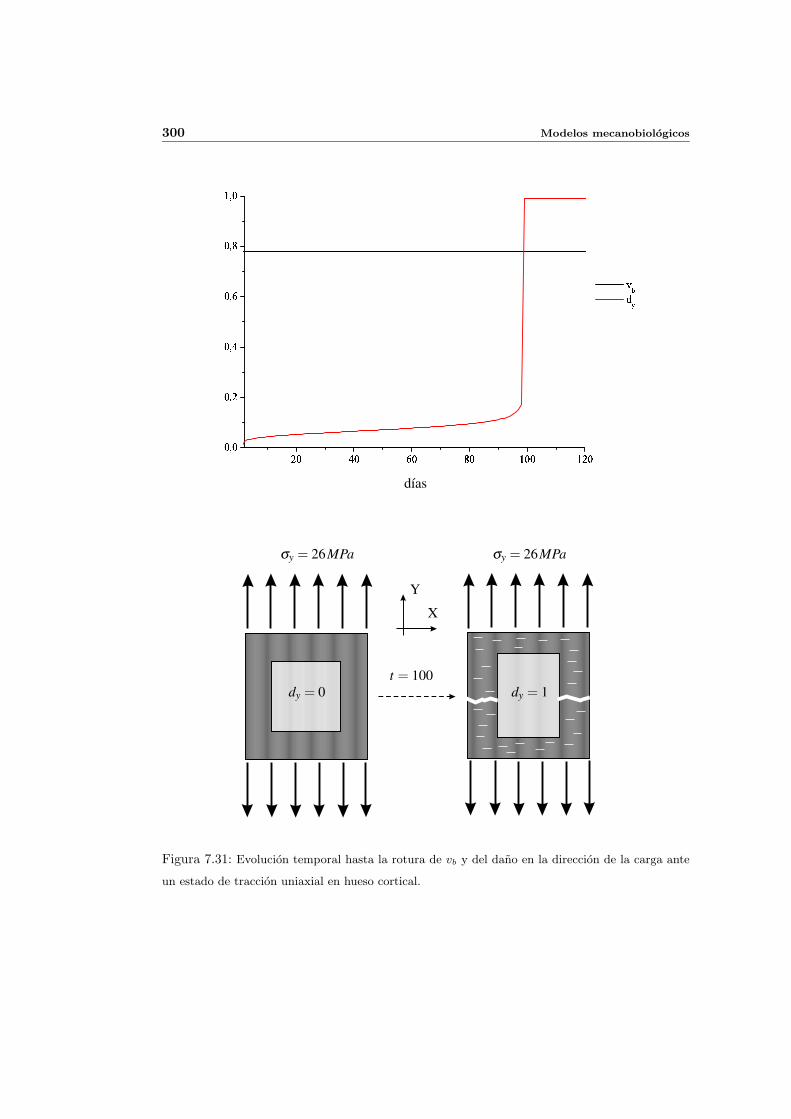
300 Modelos mecanobiologicos
ß2à á2à â+à ã/à ä1à+à äß2àà/å à
à/å ß
à/å á
à/å â
à/å ã
äå à
æ/çèMé
dıas
X
Y
t = 100
σy = 26MPa σy = 26MPa
dy = 0 dy = 1
1
Figura 7.31: Evolucion temporal hasta la rotura de vb y del dano en la direccion de la carga ante
un estado de traccion uniaxial en hueso cortical.
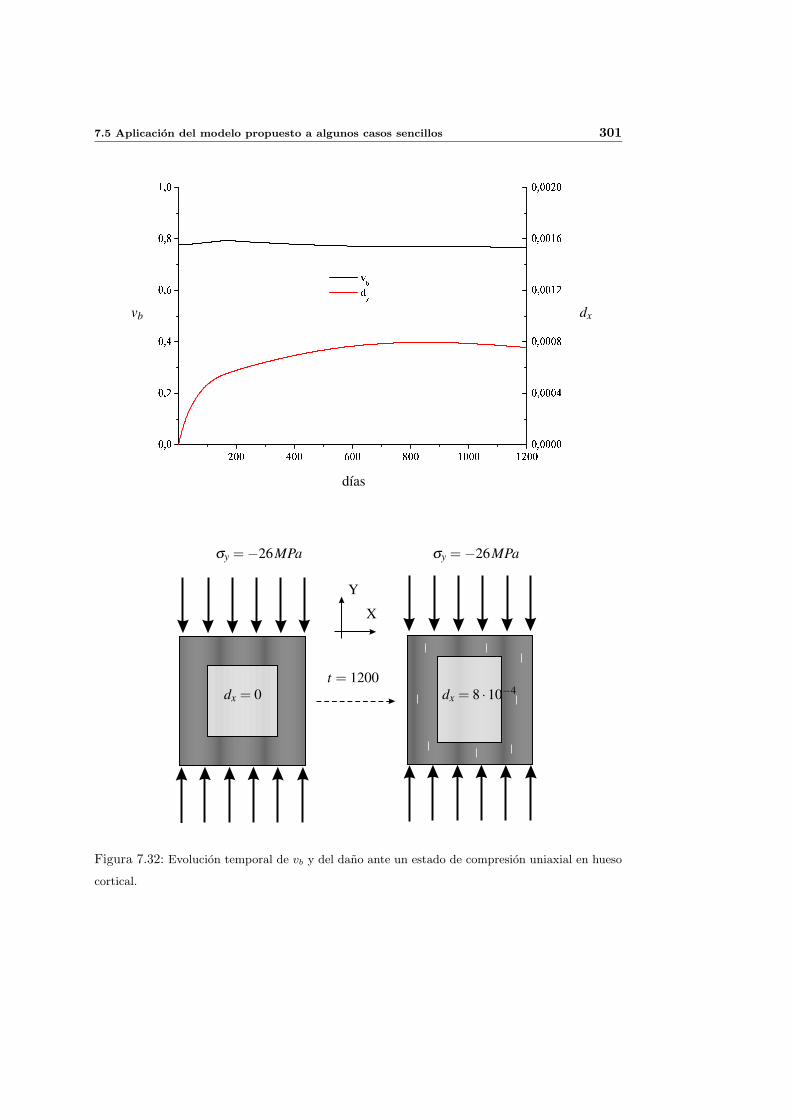
7.5 Aplicacion del modelo propuesto a algunos casos sencillos 301
ê*ë2ë ì*ë2ë í+ë2ë î+ë2ë ï1ë2ë2ë ï1ê*ë+ëë+ð ë
ë+ð ê
ë+ð ì
ë+ð í
ë+ð î
ï1ð ë
ñ+òóGô
ë+ð ë+ë2ë2ë
ë+ð ë+ë2ë+ì
ë+ð ë+ë2ë*î
ë+ð ë+ëï1ê
ë+ð ë+ëïIí
ë+ð ë+ë+ê*ë
dıas
X
Y
t = 1200
σy = −26MPa σy = −26MPa
dx = 0 dx = 8 ·10−4
vb dx
1
Figura 7.32: Evolucion temporal de vb y del dano ante un estado de compresion uniaxial en hueso
cortical.
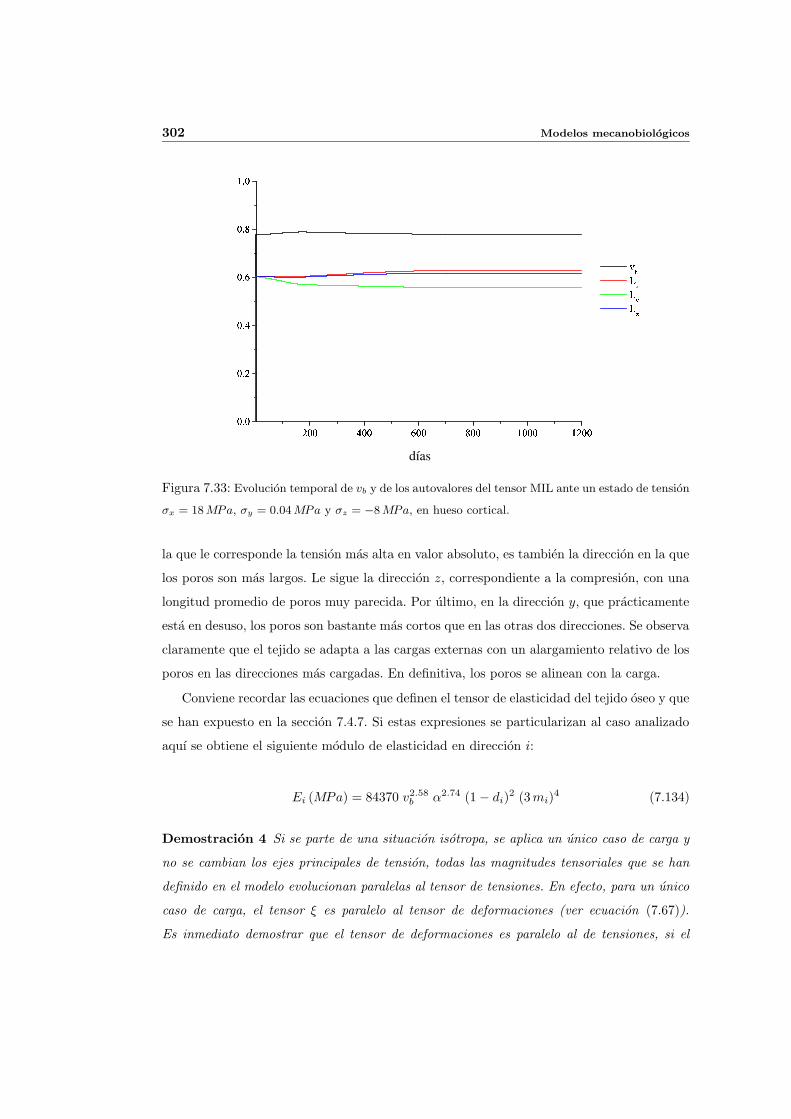
302 Modelos mecanobiologicos
õ/öGö ÷+öGö øGö/ö ùGöGö úûö/öGö úûõ/ö/öö/ü ö
ö/ü õ
ö/ü ÷
ö/ü ø
ö/ü ù
úü ö
ýGþÿÿ ÿ
dıas
1
Figura 7.33: Evolucion temporal de vb y de los autovalores del tensor MIL ante un estado de tension
σx = 18 MPa, σy = 0.04 MPa y σz = −8 MPa, en hueso cortical.
la que le corresponde la tension mas alta en valor absoluto, es tambien la direccion en la que
los poros son mas largos. Le sigue la direccion z, correspondiente a la compresion, con una
longitud promedio de poros muy parecida. Por ultimo, en la direccion y, que practicamente
esta en desuso, los poros son bastante mas cortos que en las otras dos direcciones. Se observa
claramente que el tejido se adapta a las cargas externas con un alargamiento relativo de los
poros en las direcciones mas cargadas. En definitiva, los poros se alinean con la carga.
Conviene recordar las ecuaciones que definen el tensor de elasticidad del tejido oseo y que
se han expuesto en la seccion 7.4.7. Si estas expresiones se particularizan al caso analizado
aquı se obtiene el siguiente modulo de elasticidad en direccion i:
Ei (MPa) = 84370 v2.58b α2.74 (1− di)2 (3mi)
4 (7.134)
Demostracion 4 Si se parte de una situacion isotropa, se aplica un unico caso de carga y
no se cambian los ejes principales de tension, todas las magnitudes tensoriales que se han
definido en el modelo evolucionan paralelas al tensor de tensiones. En efecto, para un unico
caso de carga, el tensor ξ es paralelo al tensor de deformaciones (ver ecuacion (7.67)).
Es inmediato demostrar que el tensor de deformaciones es paralelo al de tensiones, si el

7.5 Aplicacion del modelo propuesto a algunos casos sencillos 303
tensor de comportamiento inicial es isotropo. El tensor acumulacion de dano, paralelo al de
deformaciones (ver ecuacion (7.116)), tambien sera paralelo al de tensiones. Como se parte
de una situacion de dano nulo y el tensor de reparacion de dano es paralelo al tensor de dano
(ver ecuacion (7.120)), dicho tensor de dano evoluciona paralelo al tensor de tensiones. La
evolucion del tensor MIL sigue al tensor de deformaciones (ver ecuaciones (7.100)-(7.103) y
(7.110)) y por tanto al de tensiones. Finalmente, como el tensor de comportamiento depende
del tensor de dano y del tensor MIL (ver ecuaciones (7.126) y (7.129)), las direcciones de
ortotropıa se mantendran paralelas al tensor de tensiones y consecuentemente, todas las
magnitudes tensoriales anteriores.
La direccion de mayor rigidez sera aquella con mayor autovalor mi, es decir, aquella en
la que los poros son mas largos. En la figura 7.34 se muestra la evolucion de los modulos de
elasticidad en las direcciones x,y,z, principales de tension, que coinciden con las direcciones
de ortotropıa. Se observa que la direccion x, la de traccion, es finalmente la de mayor
rigidez, aunque al principio comienza siendo la de menor rigidez, debido al dano en esta
direccion, que crece muy rapidamente por la carga de traccion. Una vez que el dano es
reparado por remodelacion, se recupera la rigidez. A la direccion x le sigue la z, que tiene
una rigidez similar; como la carga, que en valor absoluto tambien es del mismo orden. La
rigidez en direccion y es bastante menor, algo logico puesto que en esa direccion apenas se
esta cargando el hueso y por tanto no es necesaria dicha rigidez.
El crecimiento generalizado de la rigidez, a pesar de que se ha alcanzado un estado de
equilibrio, en lo que a anisotropıa de los poros y a fraccion volumetrica de hueso se refiere,
obedece al aumento del contenido en mineral que se produce al final de la simulacion.
En la figura 7.35 se muestra la evolucion de la fraccion de ceniza promedio. El descenso
que sufre el contenido mineral al principio, debido a la reabsorcion inicial de un tejido
altamente mineralizado, afecta a las propiedades elasticas, pero hay otros factores igualmente
importantes que tambien influyen, como el cambio de porosidad y de orientacion de la
microestructura. Sin embargo, al final del analisis, cuando porosidad y microestructura se
han estabilizado, lo unico que cambia es la fraccion de ceniza, que aumenta y es la responsable
de la variacion de las propiedades elasticas.
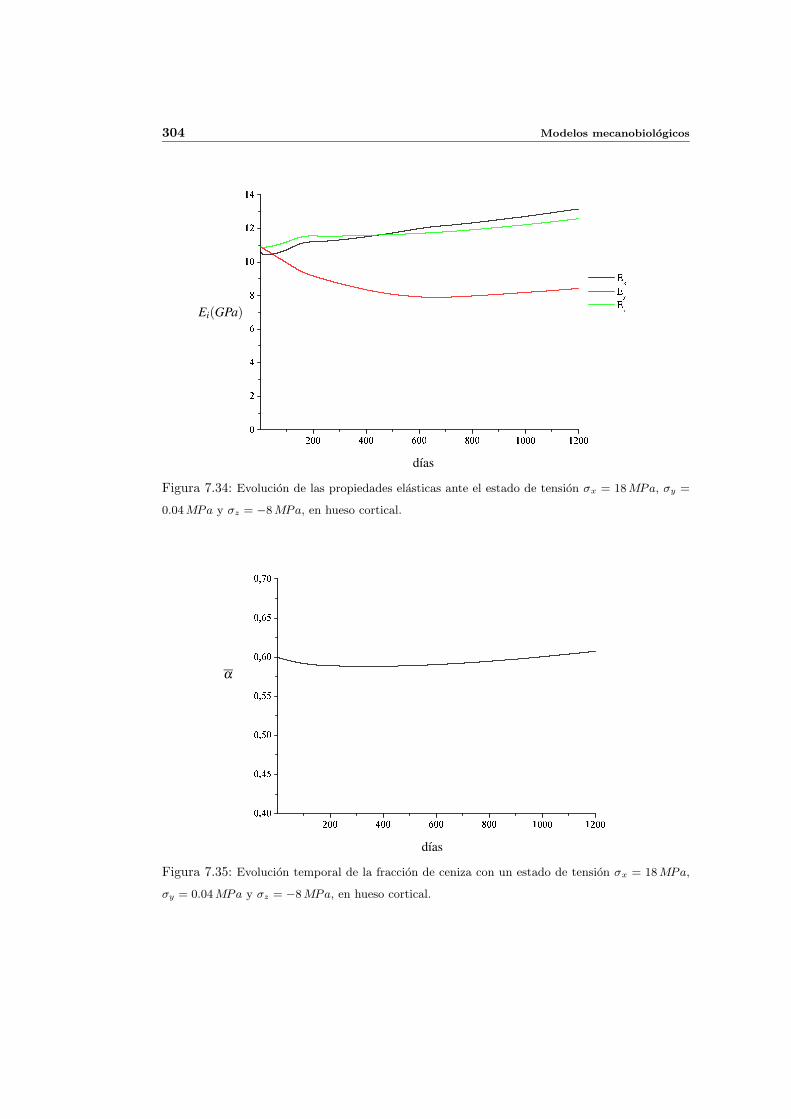
304 Modelos mecanobiologicos
dıas
Ei(GPa)
1
Figura 7.34: Evolucion de las propiedades elasticas ante el estado de tension σx = 18 MPa, σy =
0.04 MPa y σz = −8 MPa, en hueso cortical.
dıas
α
1
Figura 7.35: Evolucion temporal de la fraccion de ceniza con un estado de tension σx = 18 MPa,
σy = 0.04 MPa y σz = −8 MPa, en hueso cortical.

7.5 Aplicacion del modelo propuesto a algunos casos sencillos 305
7.5.3. Cambio de anisotropıa
En este apartado se va a ilustrar como el modelo de remodelacion propuesto en esta tesis
es capaz de predecir la adaptacion que experimenta el tejido ante cambios en el estado de
cargas externo. Para ello se parte de una situacion no isotropa, en equilibrio de remodelacion
y se aplica un estado de cargas que modifique la orientacion de la microestructura.
Cambio de la anisotropıa sin giro de la estructura
En esta simulacion se parte del estado de equilibrio obtenido en el apartado 7.5.2 para
un tejido de densidad inicial ρ0 = 1.0 g/cm3 e isotropo, sometido a un estado de traccion
uniaxial: σy = 4MPa y σx = σz = 0. Tras 1200 dıas aplicando este estado de carga (tiempo
suficiente para que se alcance el equilibrio de remodelacion) se cambia la direccion de carga:
σx = 4MPa y σy = σz = 0, que se aplica durante otros 1200 dıas, hasta que se alcanza un
nuevo equilibrio de remodelacion.
En la figura 7.36 se muestra la evolucion de vb y de los autovalores del tensor MIL desde el
principio del analisis, cuando el tejido es isotropo. Al analizar el caso de carga uniaxial se vio
como la direccion de carga se convertıa en la de mayor rigidez, es decir, la direccion en la que
los poros son mas largos. Al cambiar la orientacion de la carga se produce una reorientacion
de los poros, de forma que la nueva direccion de carga pasa a ser la de mayor rigidez. Los
poros se van acortando en direccion y para que aumente la seccion que resiste la carga σx.
Mientras tanto, se van alargando en direccion x, porque en las direcciones perpendiculares
ya no hay carga y no es necesaria una seccion resistente demasiado grande. En el equilibrio
de remodelacion, las longitudes promedio de los poros en direcciones x e y practicamente se
han intercambiado con el cambio de direccion de la carga.
Conviene observar que el tiempo que tarda en alcanzarse el nuevo equilibrio de remod-
elacion es ligeramente mayor en la segunda parte. Mientras que para pasar de la situacion
isotropa al alineamiento con la carga σy se tarda unos 700 dıas, para reorientar la arquitec-
tura trabecular se tarda cerca de 900 dıas. Este resultado es logico, ya que parece una tarea
complicada cambiar de forma tan radical la anisotropıa del entramado oseo. En la practica
no es muy usual encontrar cambios de orientacion tan pronunciados salvo en casos de tipo
traumatico.
Otra peculiaridad de la segunda parte de la simulacion es el hecho de que el cambio de
anisotropıa este acompanado de una leve reabsorcion osea (aunque justo despues de cambiar
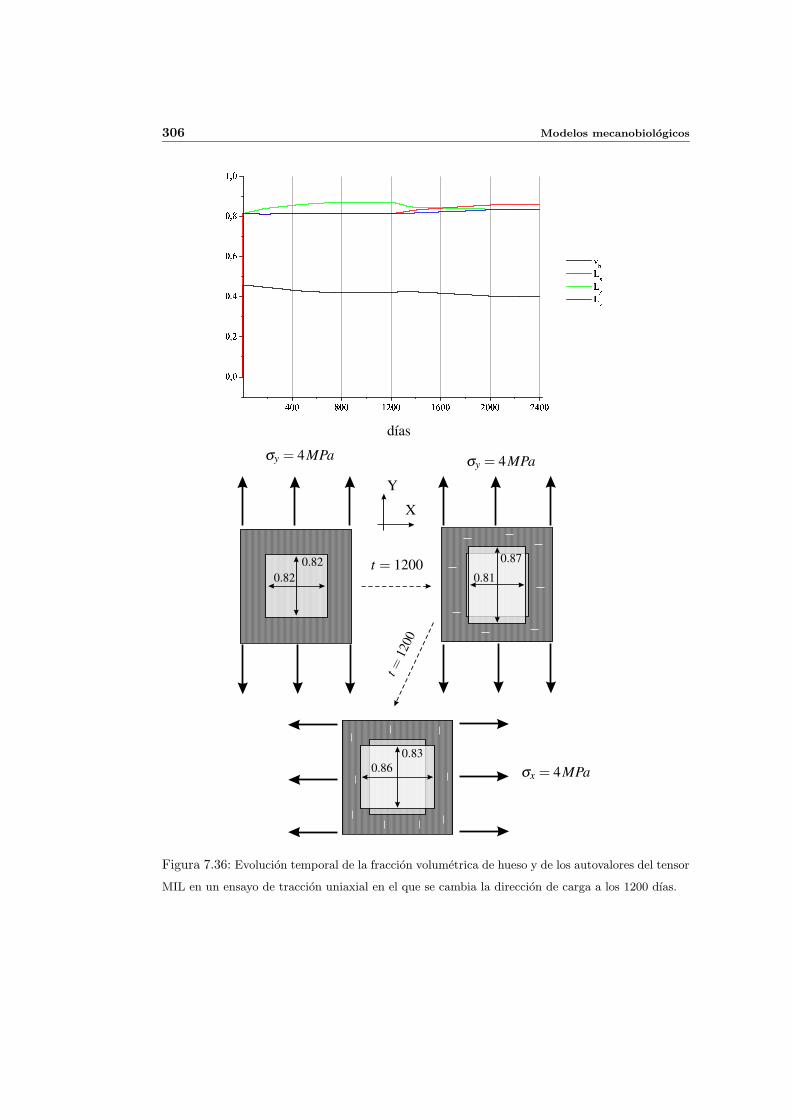
306 Modelos mecanobiologicos
! "! #%$ ! #%& $ $
'
' $
'
' &
' "
#'
(!)*,+*.-*,/
dıas
X
Y
t = 1200
σy = 4MPa σy = 4MPa
0.820.82
0.81
0.87
0.860.83
σx = 4MPa
t =12
00
1
Figura 7.36: Evolucion temporal de la fraccion volumetrica de hueso y de los autovalores del tensor
MIL en un ensayo de traccion uniaxial en el que se cambia la direccion de carga a los 1200 dıas.

7.5 Aplicacion del modelo propuesto a algunos casos sencillos 307
la carga se produce formacion, a largo plazo disminuye vb). Esta reabsorcion afecta incluso
a la longitud de los poros en direccion z (no mostrada en la figura), que aumenta, no porque
haya carga en esa direccion, sino porque hay reabsorcion neta. Ya se comento en el apartado
7.5.2 la evolucion de vb en la primera fase de la simulacion y como el aumento inicial del
dano y el desuso en direcciones x y z produce una activacion fuerte de BMUs y reabsorcion
inicial, que posteriormente se frena por el estımulo moderadamente alto que existe en la
direccion de carga.
En cambio, al girar la carga, la poblacion de BMUs no experimenta un aumento tan
brusco como al principio del analisis. Y es que el dano no varıa tan rapidamente como
entonces, ya que ahora se parte de una situacion con dano. Y sin embargo, se produce
reabsorcion a largo plazo, pero es debido a otro motivo: que se ha pasado a un estado de
desuso en la direccion y. Esta direccion se ha acomodado durante la primera fase del analisis
a un estımulo alto, incrementando su rigidez, pero de repente, baja a cero el estımulo externo
en esa direccion. Parece evidente que el desuso debe tener un efecto mas acusado cuanto
mayor sea el estımulo al que se ha “acostumbrado” el hueso y eso es lo que le ocurre a la
muestra analizada aquı, que el desuso en direccion y tiene un efecto muy importante porque
esa es una direccion de rigidez elevada. Por un razonamiento similar parece logico pensar
que el hecho de cargar una direccion que antes no estaba cargada puede producir formacion
neta, que compensarıa la reabsorcion anterior y harıa que vb no variara mucho. Pero cargar
una direccion que antes no estaba cargada no tiene tanta influencia en la remodelacion como
el desuso, porque, aunque el efecto de esas BMUs es efectivamente el de formar tejido, su
numero es mas reducido que en la direccion en la que hay desuso (la senal inhibitoria es nula
con desuso).
Cambio de la anisotropıa con giro de la estructura
Para comprobar que la estructura osea se alinea con las cargas (en la hipotesis de este
modelo, con las direcciones principales de estımulo), se ha simulado un caso de carga no
hidrostatico que cambia de direccion en un momento determinado.
Conviene tener en cuenta un aspecto importante del modelo relacionado con la hipotesis
de poro ortoedrico. Los poros de la matriz osea se han considerado ortoedricos para evaluar
la superficie sobre la que pueden avanzar las BMUs. Las dimensiones de este poro coinciden
con los autovalores del tensor MIL. Ası, en una situacion isotropa los tres autovalores son

308 Modelos mecanobiologicos
iguales y el poro promedio es un hexaedro. A priori, para que el comportamiento fuera
isotropo los poros deberıan ser esfericos, pero se supone que los poros estan aleatoriamente
distribuidos (hay que recordar que el MIL se mide haciendo un promedio), con lo que el
comportamiento es muy similar al isotropo.
En la situacion inicial isotropa se supone que las caras de los hexaedros son paralelas
a las direcciones XY Z globales y que las BMUs que hay activas antes del comienzo de la
simulacion avanzan paralelas a esas direcciones. Un hueso inicialmente isotropo de densidad
ρ0 = 1.4 g/cm3, se va a someter a un estado de tension constante en el que las direcciones
principales de tension coinciden con las direcciones XY Z globales, siendo σy = 10MPa,
σx = σz = 8MPa. Posteriormente, tras 1000 dıas aplicando cıclicamente el estado de
tensiones anterior, se hacen girar la direcciones principales de tension 30 alrededor del eje
global Z, manteniendose el valor de las tensiones principales. El nuevo tensor de tension en
ejes globales es:
σ =
8.5 0.866 0
0.866 9.5 0
0 0 8
MPa (7.135)
y se pretende simular como los poros tienden a alinearse con las nuevas direcciones principales
de tension, como han hecho otros autores [157,327]. Para ello se aplica el estado de tension
dado por (7.135) 1000 dıas mas.
En la figura 7.37 se muestra la evolucion de la fraccion volumetrica de hueso y de las
componentes del tensor MIL en las dos partes de la simulacion. La primera, la que lleva al
material del estado isotropo inicial al de isotropıa transversal y la segunda, en la que se giran
las direcciones de carga. Se observa una ligera formacion neta, al final de la primera parte.
Los poros se alargan en direccion y, pero se acortan en direcciones x y z. Tras los primeros
1000 dıas de simulacion el grado de anisotropıa es muy pequeno, dado que las tensiones
principales son muy parecidas entre sı. El tensor MIL al final de esta primera parte de la
simulacion es:
L1000 =
0.697 0 0
0 0.728 0
0 0 0.697
(7.136)
La direccion de maxima rigidez tras los 1000 primeros dıas es la direccion Y , en la que
se aplica la maxima tension principal. De repente, la tension principal maxima se empieza
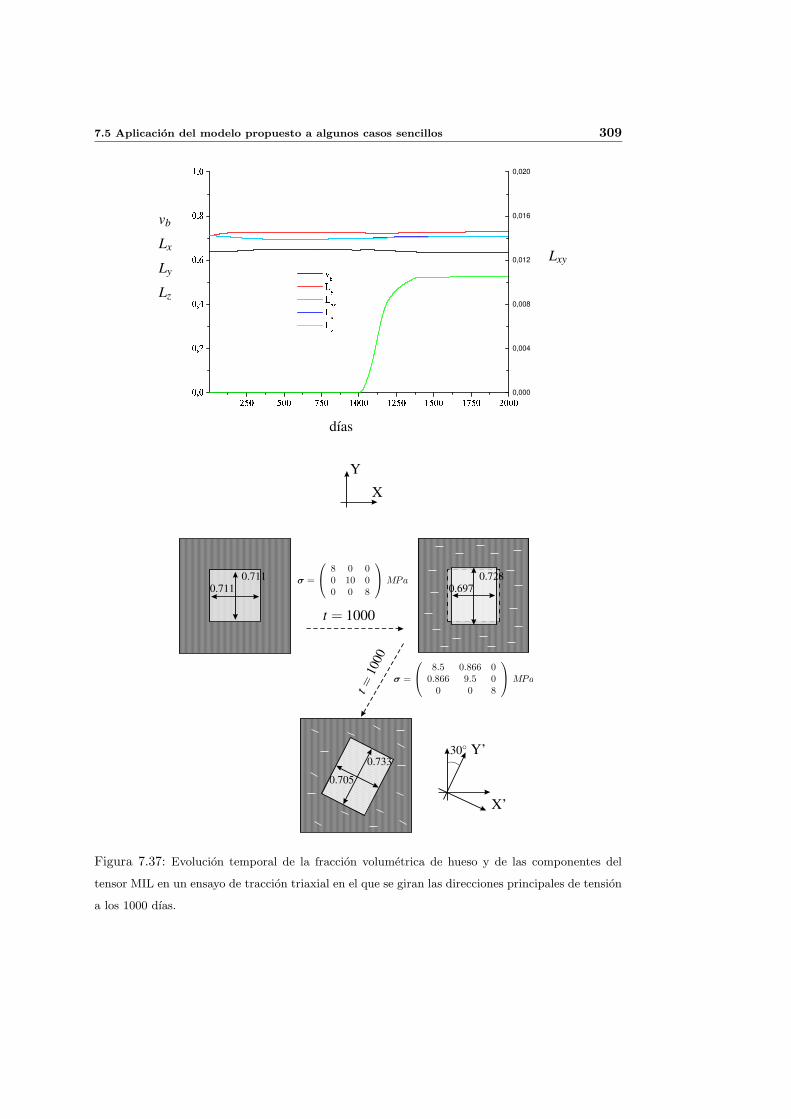
7.5 Aplicacion del modelo propuesto a algunos casos sencillos 309
01!2 1!22 3142 522265701!2851422653142902222: 2
2: 0
2: ;
2: <
2: =
5: 2
>!?@[email protected] [email protected]@.D
0,000
0,004
0,008
0,012
0,016
0,020
σ =
8 0 00 10 00 0 8
MPa (1)
1
σ =
8.5 0.866 00.866 9.5 0
0 0 8
MPa (1)
1
dıas
vb
Lx
Ly
Lz
Lxy
X
Y
X’
Y’30
t = 1000
0.7110.711
0.6970.728
0.705
0.733
t =10
00
1
Figura 7.37: Evolucion temporal de la fraccion volumetrica de hueso y de las componentes del
tensor MIL en un ensayo de traccion triaxial en el que se giran las direcciones principales de tension
a los 1000 dıas.

310 Modelos mecanobiologicos
a aplicar en una direccion distinta, con una menor rigidez, razon por la que aumenta la
deformacion principal maxima. Esto tiene como consecuencia un ligero aumento del dano
tras el giro de la carga, y esto, un aumento de la poblacion de BMUs. Ese aumento en la
poblacion de BMUs se traduce en una disminucion de la cantidad de hueso, aunque muy leve
como se puede apreciar en la figura 7.37. Pero esto es accesorio, lo realmente importante es
que las BMUs que se activan tras el giro de la carga deben conseguir la remodelacion de la
estructura osea, necesaria para que los poros se alineen con la nueva carga y la rigidez sea
maxima en la nueva direccion de maxima carga, Y ′. Las BMUs generadas con anterioridad
al giro de la carga tambien ayudan en esta mision, segun el modelo propuesto aquı. Para
ello se definio el estımulo proyectado y en funcion de el el balance local direccional de hueso.
Ası, por ejemplo, una BMU avanzando en direccion X hacıa crecer el poro en direccion Y
antes del giro de la carga. Tras el giro, la proyeccion del nuevo estımulo en la direccion de
avance de esa BMU, hace que el papel de esta cambie. Si antes su balance local direccional
de hueso era menor que la unidad (ası hacıa crecer Ly), ahora, con la proyeccion, es mayor
que la unidad, contribuyendo a cerrar el poro en esa direccion (Ly disminuye).
Tras los 1000 dıas de la segunda fase de carga, el grado de anisotropıa sigue siendo
pequeno, pero se observa un giro del tensor MIL, que finalmente queda
L2000 =
0.710 0.0106 0
0.0106 0.729 0
0 0 0.707
(7.137)
cuyos autovalores son L1 = 0.705, L2 = 0.733 y L3 = 0.707 y cuyos autovectores estan
practicamente alineados con las direcciones principales de tension, una vez alcanzado el
equilibrio de remodelacion. La direccion Z es principal de deformacion en las dos partes de
la simulacion y tambien autovector del tensor MIL. Al girar la direcciones principales de
deformacion se produce un giro de los poros alrededor del eje Z. Como ya se ha dicho, este
giro es una accion combinada de las BMUs que ya habıa activas y las que se han activado
tras el giro. Ası, las BMUs que antes del giro avanzaban en direccion Y , ahora agrandan los
poros en direccion X. Las BMUS que antes avanzaban en direccion X, ahora acortan los
poros en direccion Y y las nuevas BMUs, que se activan avanzando en direcciones X ′ e Y ′,
tratan de alinearlos con dichas direcciones.
Efectivamente, las BMUs intentan alinear los poros con la carga, pero no lo consiguen del

7.5 Aplicacion del modelo propuesto a algunos casos sencillos 311
todo: la direccion principal X ′′ del tensor MIL forma 24 con la direccion global X, a pesar
de que la carga ha girado 30. En esto tiene mucho que ver la zona muerta de remodelacion,
que se definio en funcion del modulo del tensor ψ. Este tensor error de remodelacion mide
lo cerca que esta el estımulo de equilibrio del estımulo externo. Evidentemente, al girar la
carga, gira el tensor de estımulo y el tensor de estımulo de equilibrio se intenta adaptar a
este. Cuando la diferencia (ψ) entre ambos tensores es pequena se entra en zona muerta
de remodelacion y el balance local direccional de hueso se hace neutro, disminuyendo la
capacidad de las BMUs de reorientar la estructura osea. La diferencia es pequena, solo
6, y se puede considerar que practicamente se ha alcanzado el equilibrio direccional. Sin
embargo, hay que recordar que el grado de anisotropıa de partida de la segunda fase era
bastante pequeno en este caso concreto. ¿Que ocurre si el grado de anisotropıa es mayor y
la reorientacion de la estructura requiere de un mayor “esfuerzo” por parte de las BMUs?
Para analizar que ocurre se ha realizado otra simulacion con cargas que conducen a un
mayor grado de anisotropıa. En concreto, σy = 8, σx = −1 y σz = −4MPa. Nuevamente
se parte de un hueso inicialmente isotropo de densidad ρ0 = 1.4 g/cm3, se aplican 1000 dıas
de carga, suficientes para alcanzar el equilibrio de remodelacion y se giran las cargas 30
alrededor del eje Z, aplicando 1000 dıas mas con esta nueva carga. Al final de la primera
fase el tensor MIL tiene unos autovalores Lx = 0.679, Ly = 0.760 y Lz = 0.725, con un
grado de anisotropıa superior al del caso de carga anterior. Tras el giro de la carga, el tensor
MIL gira con ella, pero solo lo hace 19.3, bastante mas lejos de los 30 que antes. Es decir,
cuanto mayor es el grado de anisotropıa, mas difıcil resulta lograr la alineacion entre poros
y carga. La razon de esto es que el error de remodelacion tiende a cero segun el principio de
acomodacion celular, con lo que, pasado un tiempo despues de un cambio de carga se alcanza
la zona muerta y se frena la adaptacion del hueso. Si la remodelacion es muy costosa, como
ocurre cuando el grado de anisotropıa es grande, la zona muerta puede alcanzarse antes de
que se logre la alineacion completa entre poros y carga.
Se ha estudiado la influencia que tienen diversos parametros del modelo en este caso de
carga y en concreto, en el angulo girado por el tensor MIL. En la tabla 7.5 se muestra un
resumen de los resultados obtenidos en este analisis de sensibilidad. El caso A corresponde
a los valores nominales que se mostraron en la tabla 7.3. En el resto de casos se modifica el
valor de uno de los parametros (el que se indica en cada caso), manteniendo constante los
demas. Se muestran los autovalores X e Y del tensor MIL en el equilibrio de remodelacion
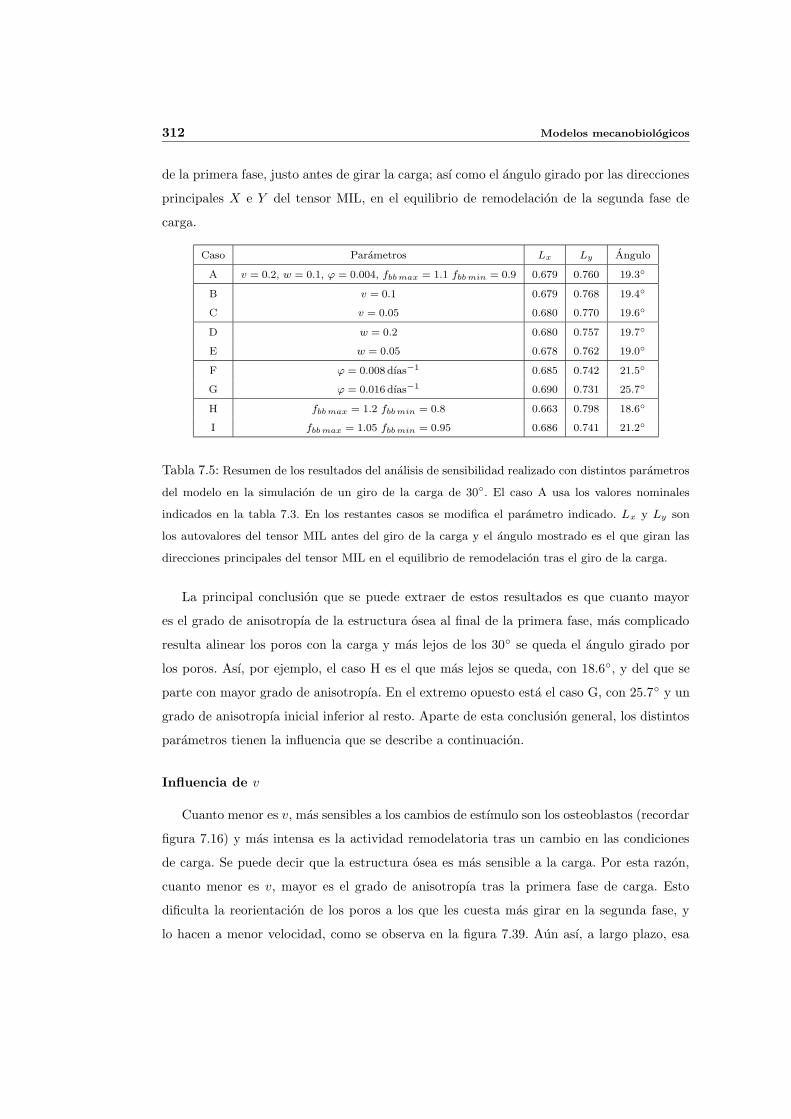
312 Modelos mecanobiologicos
de la primera fase, justo antes de girar la carga; ası como el angulo girado por las direcciones
principales X e Y del tensor MIL, en el equilibrio de remodelacion de la segunda fase de
carga.
Caso Parametros Lx Ly Angulo
A v = 0.2, w = 0.1, ϕ = 0.004, fbb max = 1.1 fbb min = 0.9 0.679 0.760 19.3
B v = 0.1 0.679 0.768 19.4
C v = 0.05 0.680 0.770 19.6
D w = 0.2 0.680 0.757 19.7
E w = 0.05 0.678 0.762 19.0
F ϕ = 0.008 dıas−1 0.685 0.742 21.5
G ϕ = 0.016 dıas−1 0.690 0.731 25.7
H fbb max = 1.2 fbb min = 0.8 0.663 0.798 18.6
I fbb max = 1.05 fbb min = 0.95 0.686 0.741 21.2
Tabla 7.5: Resumen de los resultados del analisis de sensibilidad realizado con distintos parametros
del modelo en la simulacion de un giro de la carga de 30. El caso A usa los valores nominales
indicados en la tabla 7.3. En los restantes casos se modifica el parametro indicado. Lx y Ly son
los autovalores del tensor MIL antes del giro de la carga y el angulo mostrado es el que giran las
direcciones principales del tensor MIL en el equilibrio de remodelacion tras el giro de la carga.
La principal conclusion que se puede extraer de estos resultados es que cuanto mayor
es el grado de anisotropıa de la estructura osea al final de la primera fase, mas complicado
resulta alinear los poros con la carga y mas lejos de los 30 se queda el angulo girado por
los poros. Ası, por ejemplo, el caso H es el que mas lejos se queda, con 18.6, y del que se
parte con mayor grado de anisotropıa. En el extremo opuesto esta el caso G, con 25.7 y un
grado de anisotropıa inicial inferior al resto. Aparte de esta conclusion general, los distintos
parametros tienen la influencia que se describe a continuacion.
Influencia de v
Cuanto menor es v, mas sensibles a los cambios de estımulo son los osteoblastos (recordar
figura 7.16) y mas intensa es la actividad remodelatoria tras un cambio en las condiciones
de carga. Se puede decir que la estructura osea es mas sensible a la carga. Por esta razon,
cuanto menor es v, mayor es el grado de anisotropıa tras la primera fase de carga. Esto
dificulta la reorientacion de los poros a los que les cuesta mas girar en la segunda fase, y
lo hacen a menor velocidad, como se observa en la figura 7.39. Aun ası, a largo plazo, esa
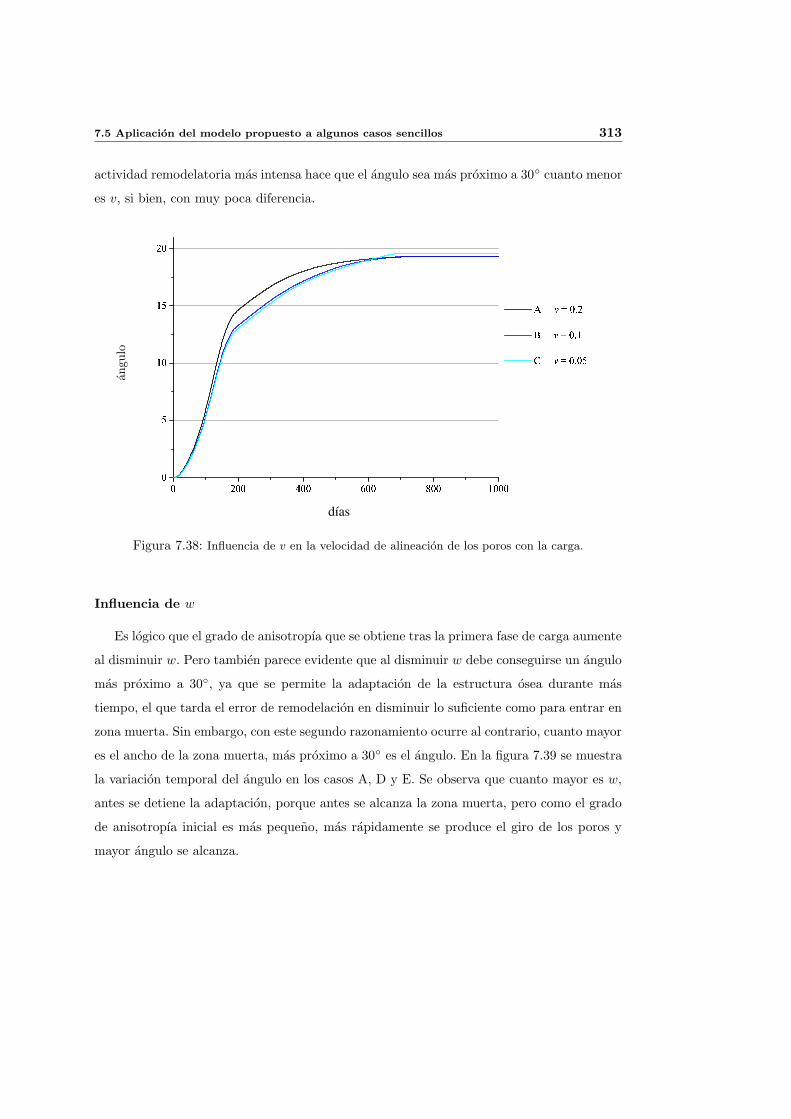
7.5 Aplicacion del modelo propuesto a algunos casos sencillos 313
actividad remodelatoria mas intensa hace que el angulo sea mas proximo a 30 cuanto menor
es v, si bien, con muy poca diferencia.
E FEE GEE HEE IEE J4EEEE
K
JE
J K
FE
LNMPOQESR F
T MPOQEBR J
UVMPOQEBR E K
dıas
angulo
1
Figura 7.38: Influencia de v en la velocidad de alineacion de los poros con la carga.
Influencia de w
Es logico que el grado de anisotropıa que se obtiene tras la primera fase de carga aumente
al disminuir w. Pero tambien parece evidente que al disminuir w debe conseguirse un angulo
mas proximo a 30, ya que se permite la adaptacion de la estructura osea durante mas
tiempo, el que tarda el error de remodelacion en disminuir lo suficiente como para entrar en
zona muerta. Sin embargo, con este segundo razonamiento ocurre al contrario, cuanto mayor
es el ancho de la zona muerta, mas proximo a 30 es el angulo. En la figura 7.39 se muestra
la variacion temporal del angulo en los casos A, D y E. Se observa que cuanto mayor es w,
antes se detiene la adaptacion, porque antes se alcanza la zona muerta, pero como el grado
de anisotropıa inicial es mas pequeno, mas rapidamente se produce el giro de los poros y
mayor angulo se alcanza.
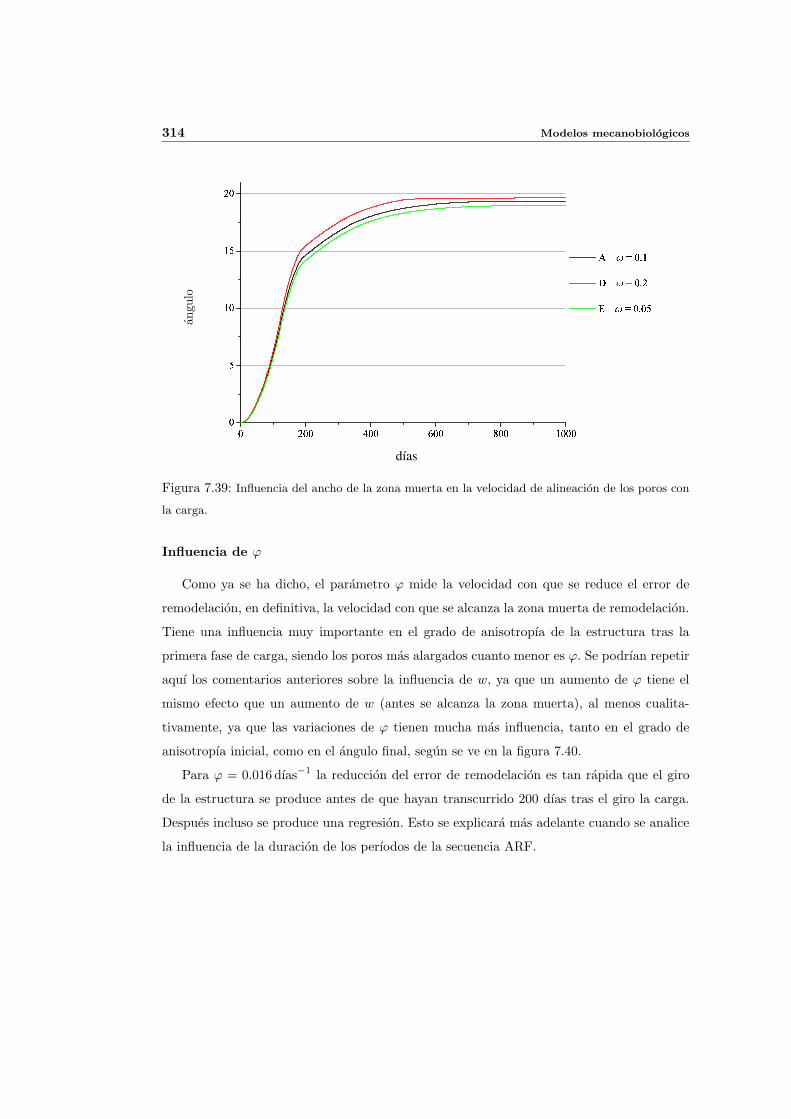
314 Modelos mecanobiologicos
W XWW YWW ZWW [WW \4WWWW
]
\W
\ ]
XW
^`_baQWSc \
d _baQWSc X
e _fagWSc W ]
dıas
angulo
1
Figura 7.39: Influencia del ancho de la zona muerta en la velocidad de alineacion de los poros con
la carga.
Influencia de ϕ
Como ya se ha dicho, el parametro ϕ mide la velocidad con que se reduce el error de
remodelacion, en definitiva, la velocidad con que se alcanza la zona muerta de remodelacion.
Tiene una influencia muy importante en el grado de anisotropıa de la estructura tras la
primera fase de carga, siendo los poros mas alargados cuanto menor es ϕ. Se podrıan repetir
aquı los comentarios anteriores sobre la influencia de w, ya que un aumento de ϕ tiene el
mismo efecto que un aumento de w (antes se alcanza la zona muerta), al menos cualita-
tivamente, ya que las variaciones de ϕ tienen mucha mas influencia, tanto en el grado de
anisotropıa inicial, como en el angulo final, segun se ve en la figura 7.40.
Para ϕ = 0.016 dıas−1 la reduccion del error de remodelacion es tan rapida que el giro
de la estructura se produce antes de que hayan transcurrido 200 dıas tras el giro la carga.
Despues incluso se produce una regresion. Esto se explicara mas adelante cuando se analice
la influencia de la duracion de los perıodos de la secuencia ARF.
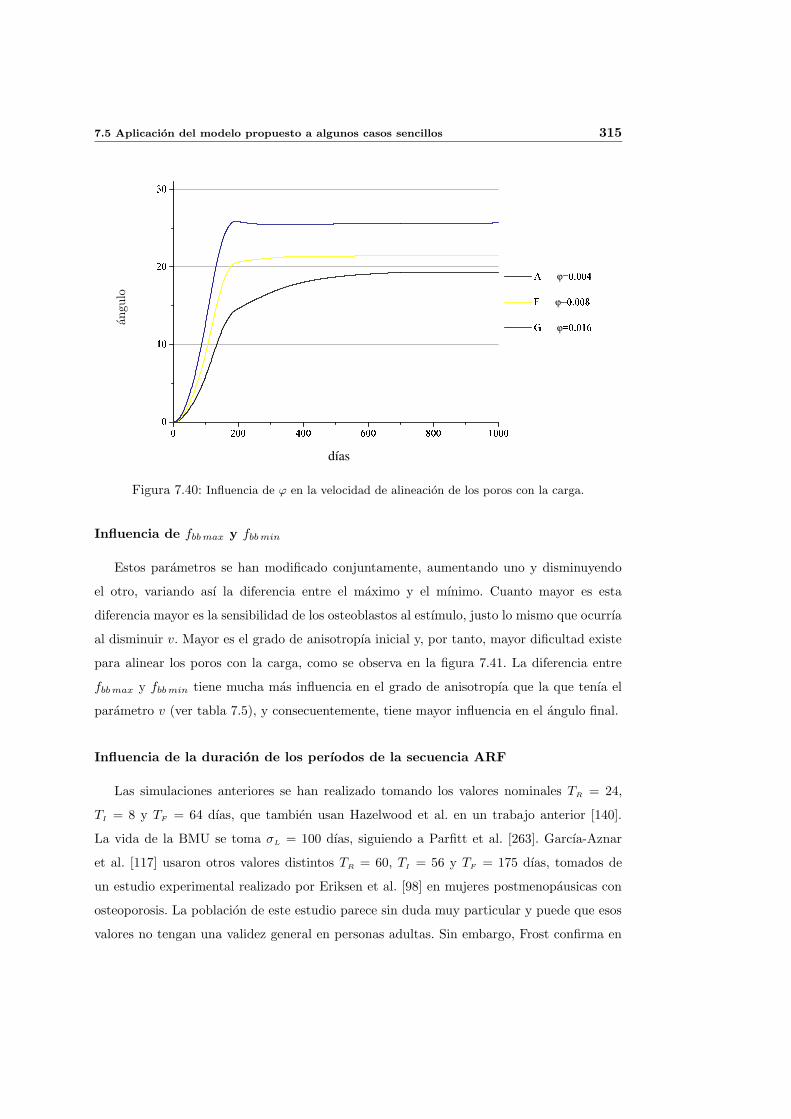
7.5 Aplicacion del modelo propuesto a algunos casos sencillos 315
h ihh jhh khh lhh m4hhhh
mh
ih
nh
oϕp hSq hhj
rϕp hSq hhSl
sϕp hSq h,mk
dıas
angulo
1
Figura 7.40: Influencia de ϕ en la velocidad de alineacion de los poros con la carga.
Influencia de fbbmax y fbbmin
Estos parametros se han modificado conjuntamente, aumentando uno y disminuyendo
el otro, variando ası la diferencia entre el maximo y el mınimo. Cuanto mayor es esta
diferencia mayor es la sensibilidad de los osteoblastos al estımulo, justo lo mismo que ocurrıa
al disminuir v. Mayor es el grado de anisotropıa inicial y, por tanto, mayor dificultad existe
para alinear los poros con la carga, como se observa en la figura 7.41. La diferencia entre
fbbmax y fbbmin tiene mucha mas influencia en el grado de anisotropıa que la que tenıa el
parametro v (ver tabla 7.5), y consecuentemente, tiene mayor influencia en el angulo final.
Influencia de la duracion de los perıodos de la secuencia ARF
Las simulaciones anteriores se han realizado tomando los valores nominales TR = 24,
TI = 8 y TF = 64 dıas, que tambien usan Hazelwood et al. en un trabajo anterior [140].
La vida de la BMU se toma σL = 100 dıas, siguiendo a Parfitt et al. [263]. Garcıa-Aznar
et al. [117] usaron otros valores distintos TR = 60, TI = 56 y TF = 175 dıas, tomados de
un estudio experimental realizado por Eriksen et al. [98] en mujeres postmenopausicas con
osteoporosis. La poblacion de este estudio parece sin duda muy particular y puede que esos
valores no tengan una validez general en personas adultas. Sin embargo, Frost confirma en
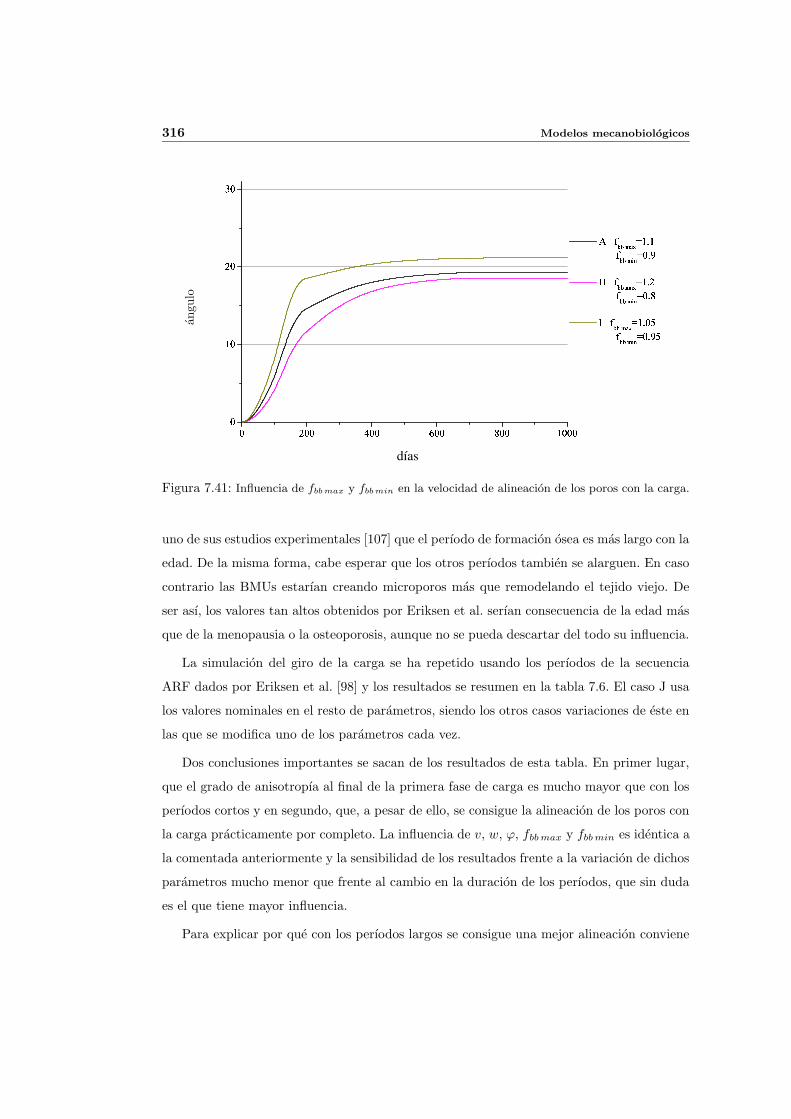
316 Modelos mecanobiologicos
t utt vtt wtt xtt y4tttt
yt
ut
zt
| ~ ~! Py! y| ~ ~!! tS
| ~ ~! Py! u| ~ ~!! tS x
| ~ ~! Py! tS| ~ ~!! tS
dıas
angulo
1
Figura 7.41: Influencia de fbb max y fbb min en la velocidad de alineacion de los poros con la carga.
uno de sus estudios experimentales [107] que el perıodo de formacion osea es mas largo con la
edad. De la misma forma, cabe esperar que los otros perıodos tambien se alarguen. En caso
contrario las BMUs estarıan creando microporos mas que remodelando el tejido viejo. De
ser ası, los valores tan altos obtenidos por Eriksen et al. serıan consecuencia de la edad mas
que de la menopausia o la osteoporosis, aunque no se pueda descartar del todo su influencia.
La simulacion del giro de la carga se ha repetido usando los perıodos de la secuencia
ARF dados por Eriksen et al. [98] y los resultados se resumen en la tabla 7.6. El caso J usa
los valores nominales en el resto de parametros, siendo los otros casos variaciones de este en
las que se modifica uno de los parametros cada vez.
Dos conclusiones importantes se sacan de los resultados de esta tabla. En primer lugar,
que el grado de anisotropıa al final de la primera fase de carga es mucho mayor que con los
perıodos cortos y en segundo, que, a pesar de ello, se consigue la alineacion de los poros con
la carga practicamente por completo. La influencia de v, w, ϕ, fbbmax y fbbmin es identica a
la comentada anteriormente y la sensibilidad de los resultados frente a la variacion de dichos
parametros mucho menor que frente al cambio en la duracion de los perıodos, que sin duda
es el que tiene mayor influencia.
Para explicar por que con los perıodos largos se consigue una mejor alineacion conviene
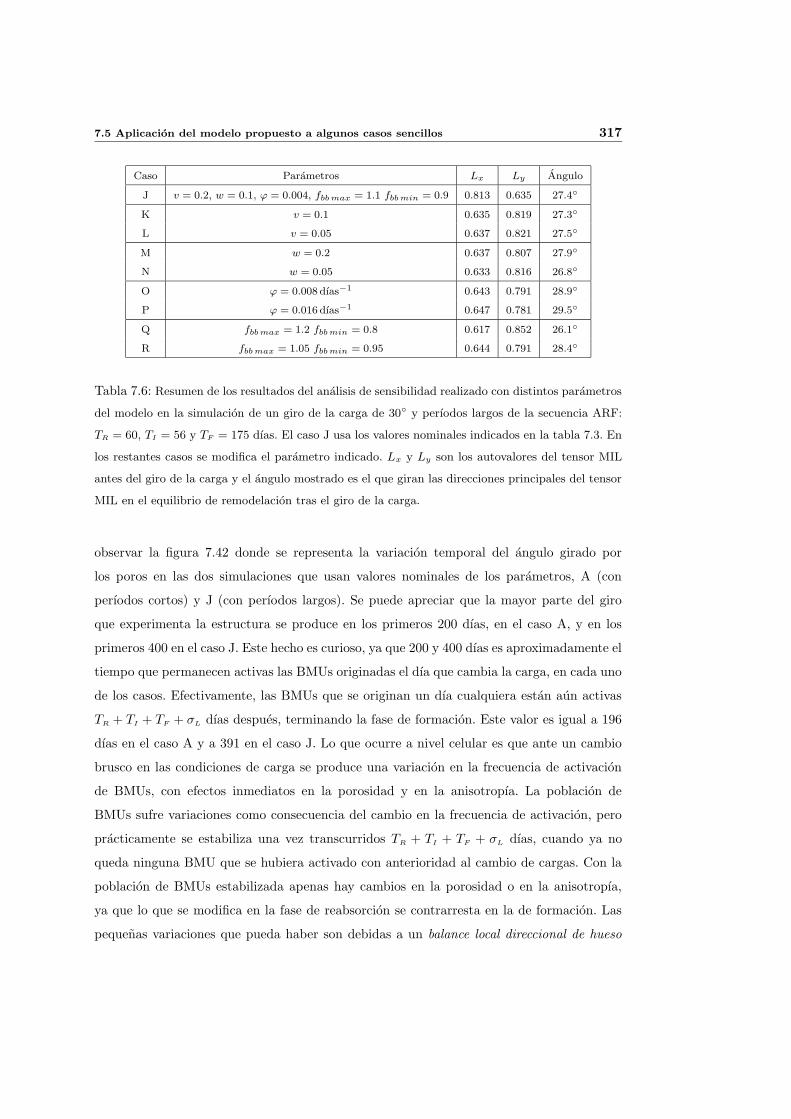
7.5 Aplicacion del modelo propuesto a algunos casos sencillos 317
Caso Parametros Lx Ly Angulo
J v = 0.2, w = 0.1, ϕ = 0.004, fbb max = 1.1 fbb min = 0.9 0.813 0.635 27.4
K v = 0.1 0.635 0.819 27.3
L v = 0.05 0.637 0.821 27.5
M w = 0.2 0.637 0.807 27.9
N w = 0.05 0.633 0.816 26.8
O ϕ = 0.008 dıas−1 0.643 0.791 28.9
P ϕ = 0.016 dıas−1 0.647 0.781 29.5
Q fbb max = 1.2 fbb min = 0.8 0.617 0.852 26.1
R fbb max = 1.05 fbb min = 0.95 0.644 0.791 28.4
Tabla 7.6: Resumen de los resultados del analisis de sensibilidad realizado con distintos parametros
del modelo en la simulacion de un giro de la carga de 30 y perıodos largos de la secuencia ARF:
TR = 60, TI = 56 y TF = 175 dıas. El caso J usa los valores nominales indicados en la tabla 7.3. En
los restantes casos se modifica el parametro indicado. Lx y Ly son los autovalores del tensor MIL
antes del giro de la carga y el angulo mostrado es el que giran las direcciones principales del tensor
MIL en el equilibrio de remodelacion tras el giro de la carga.
observar la figura 7.42 donde se representa la variacion temporal del angulo girado por
los poros en las dos simulaciones que usan valores nominales de los parametros, A (con
perıodos cortos) y J (con perıodos largos). Se puede apreciar que la mayor parte del giro
que experimenta la estructura se produce en los primeros 200 dıas, en el caso A, y en los
primeros 400 en el caso J. Este hecho es curioso, ya que 200 y 400 dıas es aproximadamente el
tiempo que permanecen activas las BMUs originadas el dıa que cambia la carga, en cada uno
de los casos. Efectivamente, las BMUs que se originan un dıa cualquiera estan aun activas
TR + TI + TF + σL dıas despues, terminando la fase de formacion. Este valor es igual a 196
dıas en el caso A y a 391 en el caso J. Lo que ocurre a nivel celular es que ante un cambio
brusco en las condiciones de carga se produce una variacion en la frecuencia de activacion
de BMUs, con efectos inmediatos en la porosidad y en la anisotropıa. La poblacion de
BMUs sufre variaciones como consecuencia del cambio en la frecuencia de activacion, pero
practicamente se estabiliza una vez transcurridos TR + TI + TF + σL dıas, cuando ya no
queda ninguna BMU que se hubiera activado con anterioridad al cambio de cargas. Con la
poblacion de BMUs estabilizada apenas hay cambios en la porosidad o en la anisotropıa,
ya que lo que se modifica en la fase de reabsorcion se contrarresta en la de formacion. Las
pequenas variaciones que pueda haber son debidas a un balance local direccional de hueso
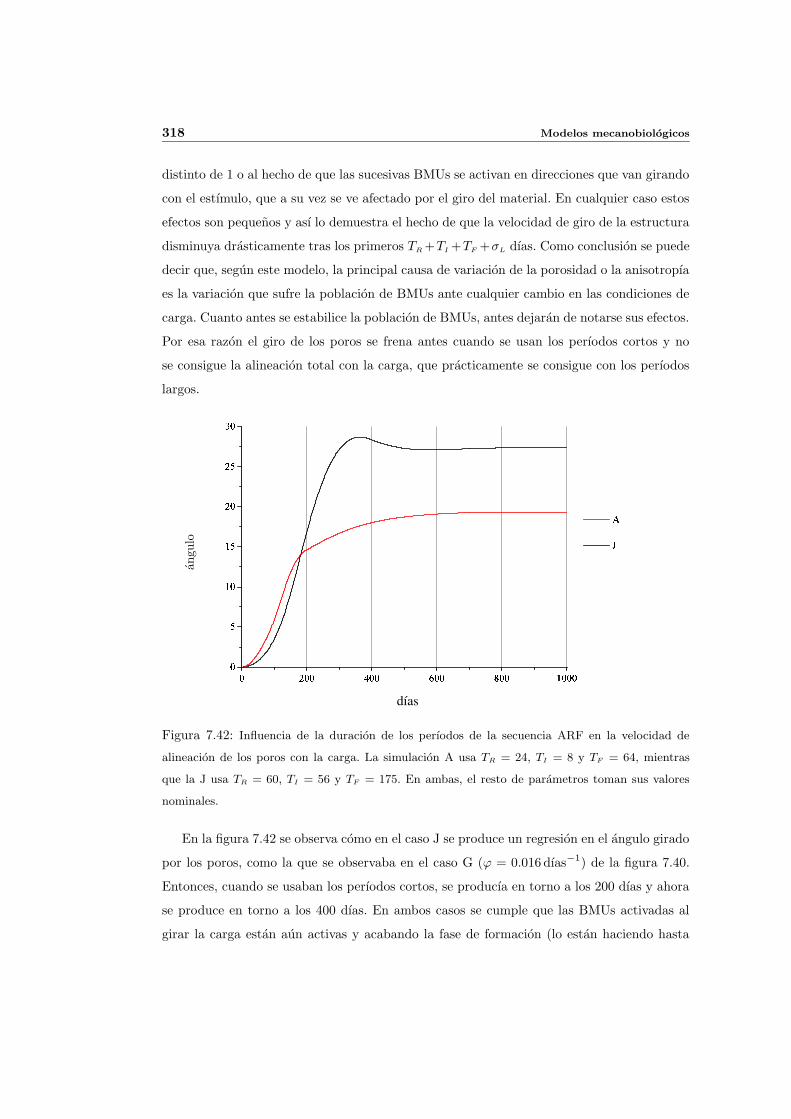
318 Modelos mecanobiologicos
distinto de 1 o al hecho de que las sucesivas BMUs se activan en direcciones que van girando
con el estımulo, que a su vez se ve afectado por el giro del material. En cualquier caso estos
efectos son pequenos y ası lo demuestra el hecho de que la velocidad de giro de la estructura
disminuya drasticamente tras los primeros TR +TI +TF +σL dıas. Como conclusion se puede
decir que, segun este modelo, la principal causa de variacion de la porosidad o la anisotropıa
es la variacion que sufre la poblacion de BMUs ante cualquier cambio en las condiciones de
carga. Cuanto antes se estabilice la poblacion de BMUs, antes dejaran de notarse sus efectos.
Por esa razon el giro de los poros se frena antes cuando se usan los perıodos cortos y no
se consigue la alineacion total con la carga, que practicamente se consigue con los perıodos
largos.
4
dıas
angulo
1
Figura 7.42: Influencia de la duracion de los perıodos de la secuencia ARF en la velocidad de
alineacion de los poros con la carga. La simulacion A usa TR = 24, TI = 8 y TF = 64, mientras
que la J usa TR = 60, TI = 56 y TF = 175. En ambas, el resto de parametros toman sus valores
nominales.
En la figura 7.42 se observa como en el caso J se produce un regresion en el angulo girado
por los poros, como la que se observaba en el caso G (ϕ = 0.016 dıas−1) de la figura 7.40.
Entonces, cuando se usaban los perıodos cortos, se producıa en torno a los 200 dıas y ahora
se produce en torno a los 400 dıas. En ambos casos se cumple que las BMUs activadas al
girar la carga estan aun activas y acabando la fase de formacion (lo estan haciendo hasta

7.5 Aplicacion del modelo propuesto a algunos casos sencillos 319
TR +TF +TI +σL dıas despues), en el momento en el que se produce dicha regresion. Se acaba
de comentar que los cambios en el hueso son ya menos importantes una vez transcurridos esos
TR+TF +TI +σL dıas, cuando se estabiliza la poblacion de BMUs. Pero esos cambios tambien
pueden frenarse por otro motivo: que las BMUs entren en zona muerta de remodelacion. En
ese caso el balance local direccional de hueso se hace neutro (se iguala a 1) y las BMUs en
fase de formacion no influyen tanto en la anisotropıa. Eso es lo que ocurre en ambos casos,
que concurren las dos razones anteriores para que se frene la velocidad de giro de los poros:
se estabiliza la poblacion de BMUs y ademas estas entran en zona muerta. En el caso J,
400 dıas son suficientes para que el estımulo de equilibrio se adapte al estımulo externo y
se entre en zona muerta, y en el caso G, 200 dıas tambien lo son, porque el valor de ϕ es
muy alto y la adaptacion se produce muy rapidamente. La concurrencia de las dos razones
comentadas hace que la velocidad de giro de los poros no solo se frene, sino que incluso se
produzca una regresion en el giro, como la que se observa en el caso J.
Como conclusion a este apartado se puede hablar de la influencia de la edad en los
cambios de orientacion de la estructura, si se toma como cierta la observacion de Frost [107]
de que los perıodos de la secuencia ARF se hacen mas largos con la edad. En tal caso los
resultados de este modelo predirıan que en personas mayores es mas facil que un cambio en la
forma de cargar el hueso afecte a la arquitectura osea y, por tanto, que sean mas propensas
a sufrir deformacion en los huesos, como ocurre en la realidad. En cambio, en personas
jovenes esos cambios de actividad no afectan tanto a la estructura osea interna. Pero este
razonamiento no esta completo, ya que no se ha dicho nada sobre como influye la edad en
la remodelacion osea externa, que es el proceso que realmente determina la forma de los
huesos. Lo que se sugiere aquı es que si ese cambio de forma se produce, por remodelacion
externa, la estructura interna puede alinearse mas facilmente con las cargas en personas
ancianas que en jovenes, al menos con las hipotesis propuestas en este modelo.


Capıtulo 8
Analisis de implantes dentales
con el modelo de ROI
propuesto. Resultados II
8.1. Introduccion
En el anterior capıtulo se ha desarrollado un nuevo modelo de remodelacion osea interna
anisotropo de tipo mecanobiologico, que analiza con cierto detalle alguno de los procesos
biologicos de relevancia en la remodelacion osea. Este modelo presenta algunas ventajas con
respecto a los fenomenologicos, y en concreto, con respecto al modelo anisotropo basado
en la Mecanica del Dano, que se utilizo para simular la morfogenesis de la mandıbula. En
primer lugar, el nuevo modelo permite cuantificar de forma mas aproximada la velocidad
de remodelacion real del tejido oseo. Pero sobre todo, el nuevo modelo salva una limitacion
importante que tiene el modelo basado en la Mecanica del Dano. Resulta curioso, porque
esta limitacion es, precisamente, que no tiene en cuenta el dano microestructural real que se
acumula en el hueso por la carga. En el modelo fenomenologico, el dano es en realidad una
medida de la porosidad, pero no del dano microestructural. Y este dano se supone que es
precisamente uno de los factores reguladores de la activacion de BMUs y consecuentemente
de la remodelacion osea, de ahı la importancia de tenerlo en cuenta. Es mas, el dano es
especialmente importante cuando se coloca un implante, pues se va a acumular en el tejido
321

322 Analisis de implantes dentales
periimplantario de manera inevitable. Esa es la principal diferencia entre el modelo usado
para simular la morfogenesis y el nuevo modelo desarrollado en esta tesis: la respuesta ante
una sobrecarga muy alta. Si el modelo basado en la Mecanica del Dano predice en ese caso
un aumento importante de la cantidad de tejido, este ultimo, predice algo mas parecido a lo
que ocurre en realidad, que el hueso se pierde a largo plazo, en el intento de reparar un dano
tan elevado. Estas dos caracterısticas hacen muy apropiado el uso del nuevo modelo en la
simulacion del comportamiento del tejido oseo tras la colocacion de un implante dental: los
cambios que sufra el hueso se pueden seguir en tiempo real (al menos de forma cualitativa)
y va a ser posible simular la reabsorcion crestal debida a la sobrecarga que sufre el cuello
del implante.
Por otro lado, conviene aclarar que el modelo usado en la morfogenesis es perfectamente
valido para el objetivo que allı se perseguıa, ya que no habıa sobrecargas como las que
ocurren con un implante dental, sino masticaciones normales con dientes naturales, y no
existıa interes alguno en simular el proceso de morfogenesis en tiempo real, sino simplemente
obtener la distribucion de densidad y propiedades elasticas en la mandıbula de manera
sencilla.
El elevado coste computacional del modelo propuesto aquı ha hecho necesarias dos sim-
plificaciones. Por un lado, se ha limitado su uso al hueso periimplantario, dejando que el
resto del hueso se siga comportando segun el modelo basado en la Mecanica del Dano. Esto
esta justificado puesto que se supone que solo el hueso que rodea al implante va a sufrir cam-
bios considerables en el estado de tension-deformacion. Por otro lado, ha sido necesaria la
extraccion de un submodelo en el modelo completo de la mandıbula, tal como se describio en
el capıtulo 3. La malla de EF del submodelo se muestra en la figura 8.1.
La parte de hueso mas alejada del implante, la que se sigue comportando segun el modelo
basado en la Mecanica del Dano, tiene las propiedades elasticas que con dicho modelo se
han simulado en la primera fase del analisis (ver capıtulo 6). Para definir las constantes
elasticas del hueso periimplantario en la situacion inicial ha sido necesario suponer algunas
propiedades del mismo. En concreto, se ha supuesto una densidad inicial y una distribucion
uniforme e isotropa de los tejidos. Las razones por las que se hacen estas suposiciones ya se
explicaron en el capıtulo 3, pero se recuerdan aquı brevemente. Lo que ocurre tras la perdida
de un diente natural es que el hueco dejado por este se rellena de hueso, normalmente de
tipo trabecular de baja densidad, mediante un proceso de remodelacion osea externa que

8.1 Introduccion 323
a b
2 premolarimplante
2 molar
1
Figura 8.1: (a) Malla de elementos finitos del submodelo. (b) Detalle del hueso periimplantario y
el implante.
no se ha simulado aquı. Pero, este hueso sufre otra alteracion que tampoco se ha simulado,
la que le infiere el proceso quirurgico de implantacion. Por evitar estas simulaciones, que
complicarıan el estudio sobremanera, y por la falta de datos acerca de las propiedades del
tejido periimplantario previo a la entrada en carga, que es el unico proceso que se simula
aquı, se ha optado por la simplificacion de adoptar un valor inicial uniforme de la densidad
y las propiedades mecanicas.
Como tambien se dijo en el capıtulo 3, se van a analizar dos tipos de implante. El
primer tipo esta completamente osteointegrado en toda su superficie. Esta es la situacion
que se da en los implantes con recubrimiento de spray de plasma de titanio, que tienen una
superficie rugosa que facilita la osteointegracion. A falta de un mejor conocimiento sobre
este proceso, la osteointegracion se supondra perfecta en toda la superficie, lo que quiere
decir que el implante y el hueso circundante comparten los mismos nodos en la interfase y
ademas que dicha union no se va a perder. El segundo tipo de implante esta parcialmente
osteointegrado ya que en la zona del cuello se supone que se ha perdido la osteointegracion.
Esta situacion es la que se produce en implantes como el IMZ que tienen el recubrimiento

324 Analisis de implantes dentales
de spray de plasma de titanio excepto en el cuello del implante, donde presenta una zona
lisa cuya osteointegracion es mas debil. Se supondra que la osteointegracion en esta zona se
pierde inmediatamente cuando haya la mas mınima carga sobre el implante, rompiendose
la interfase y permitiendo entonces el contacto entre las superficies de implante y hueso. La
forma de transmitirse las cargas es completamente distinta en uno y otro caso y por tanto
tambien lo sera la remodelacion. Su efecto se va a comparar aquı.
Tambien se compara el efecto que tiene no cargar el implante durante un cierto perıodo
despues de conseguida la osteointegracion. Para ello se definio una secuencia de masticacion
S5, unilateral, con las cargas (MI1-MI2)-(MI1-MI2)-. . . Esta se compara con una secuencia
de masticacion unilateral alternante, S4, con la sucesion de cargas (MI1-MI2-MImp-MD2)-
(MI1-MI2-MImp-MD2)-. . .
Por ultimo se compara el efecto de la edad del paciente, modificando el parametro del
modelo fbio, que controla la frecuencia de activacion de BMUs y que es mayor en jovenes.
Antes de pasar a los resultados conviene hacer una aclaracion sobre la forma en que se
aplican las cargas de la secuencia de masticacion. Considerese, por ejemplo, la secuencia S4
y supongase, como se hace aquı, que diariamente se producen 500 ciclos de carga (125 ciclos
para cada tipo de carga). En la simulacion se comienza aplicando 125 ciclos de la carga
MI1, se actualiza el dano y se calcula el estımulo producido por los 125 ciclos de esa carga.
A continuacion se aplican 125 ciclos de la carga MI2, se actualiza el dano y se acumula el
estımulo segun la superposicion de Carter et al. [57] (ver ecuacion (7.69)). Las dos cargas
restantes de la secuencia de masticacion se aplican a continuacion, actualizando el dano
despues de cada una de ellas y solo al final de la secuencia, es decir, “al final del dıa”, se
aplica el algoritmo de remodelacion y se actualizan el resto de variables. En resumen, el
dano se actualiza despues de cada carga, pero la densidad y el tensor MIL se actualizan al
final de la secuencia, “al final del dıa”.
El hecho de aplicar 125 ciclos de carga no implica resolver un problema dinamico. Segun la
aproximacion hecha por Beaupre et al. [27], se pueden estimar las tensiones o deformaciones
maximas (aquı se hace resolviendo un unico problema estatico) y considerar para el calculo
del estımulo un numero de ciclos nc = 125. El calculo del dano tambien se simplifica usando
nc = 125 en las curvas de fatiga, y las deformaciones dadas por la resolucion del problema
correspondiente a un solo ciclo de carga. Esta limitacion que tiene el modelo es importante
ya que, ası como en la remodelacion osea se supone que solo influye el nivel de tensiones

8.2 Implantes completamente osteointegrados 325
(o deformaciones) y el numero de ciclos diarios (al menos eso suponen Beaupre et al. [27]),
con el dano microestructural esa hipotesis deja de tener sentido y el orden de aplicacion de
las cargas puede influir en el resultado final. Sin embargo, eso no se esta teniendo en cuenta
aquı, con la agrupacion de cargas antes comentada, por el elevado coste computacional que
tendrıa hacerlo.
8.2. Implantes completamente osteointegrados
Los parametros del modelo de remodelacion utilizados en esta simulacion se muestran
en la tabla 8.1. Para la densidad inicial se han elegido valores de 2.0 y 1.7 g/cm3, corre-
spondientes a un hueso cortical denso y a un cortical ligero, respectivamente. El modulo de
elasticidad del titanio es E = 110GPa y su coeficiente de Poisson ν = 0.35. Mientras tanto,
el modulo de elasticidad del hueso en la situacion inicial (sin dano, isotropo y con una frac-
cion de ceniza 0.6) es 16.5 y 10.9GPa, para las densidades 2.0 y 1.7 g/cm3 respectivamente.
En cualquiera de los casos, muy diferente al modulo de elasticidad del titanio, un hecho
crucial a la hora de analizar las tensiones que aparecen en la interfaz hueso - titanio. La
diferencia de modulos de elasticidad y los problemas que se derivan de esta discontinuidad
de propiedades elasticas en la interfaz son mas importantes cuanto menor es la densidad
inicial adoptada para el tejido periimplantario.
Volviendo a los valores de la densidad inicial supuesta para dicho tejido periimplantario,
hay que llamar la atencion sobre las diferencias entre los valores supuestos aquı y los valores
reales. De la observacion directa del hueso que rellena el hueco dejado por el primer molar (ver
3.10), no se puede saber su densidad. Solo se puede decir que es alta, aunque su estructura
no es la de un hueso cortical, mas bien parece un hueso trabecular denso. En la figura 6.11
se observa una tomografıa tomada de la mandıbula real a la altura del segundo premolar,
diente que el individuo habıa perdido hacıa algun tiempo. Del analisis de esta sı se puede
conocer mejor la forma en que se rellena el hueco dejado por el diente: una capa superior
de tejido denso aunque con estructura trabecular, como ya se ha dicho, y debajo de ella un
hueso bastante mas poroso. En principio, nada tiene que ver la densidad supuesta aquı con
la densidad real. Por ello, durante este trabajo tambien se ha simulado un caso con una
densidad inicial menor que las anteriores, esto es, mas parecida a la real. Sin embargo, sus
1En las BMUs de tejido cortical, el volumen maximo de tejido depositado por los osteoblastos esta limitado
por razones de espacio. Por esa razon se ha tomado fbb,max = 1.0.

326 Analisis de implantes dentales
Parametro Valor nominal
N Numero de ciclos 500
m Exponente de Carter 4 a
k Razon volumen danado - degradacion 0.000333 b
c Parametro de activacion por estımulo 0.0025
a Exponente de activacion por dano 10 b
fbio Factor biologico de activacion 0.03BMUs
mm2 dıab
fbb,min Balance local de hueso mınimo 0.8
fbb,max Balance local de hueso maximo 1.0 1
ξ∗0
Estımulo de equilibrio inicial 0.0025
v Ancho de la zona de transicion 0.2ξ∗0
b
w Semiancho de la zona muerta 0.1ξ∗0
ϕ Exponente de acomodacion del estımulo 0.004
TR Duracion del perıodo de reabsorcion 24 dıas c
TI Duracion del perıodo de inversion 8 dıas c
TF Duracion del perıodo de formacion 64 dıas c
vBMU Velocidad de avance de la BMU 40 µm/dıa d
σL Vida de la BMU 100 dıas e
dO Diametro de la osteona 0.076 mm b
dH Diametro del canal haversiano 0.0145 mm b
dE Profundidad de erosion de la hemiosteona 0.0491 mm b
dE Ancho de la hemiosteona 0.152 mm b
α0 Fraccion de ceniza inicial 0.6 b
αmin Fraccion de ceniza tras la mineralizacion primaria 0.45 f
αmax Fraccion de ceniza maxima 0.7 f
κ Exponente de velocidad de mineralizacion 0.0009 f
ρ0 Densidad inicial 2.0, 1.7
di Nivel de dano inicial 0 ∀i
Li Componentes del tensor MIL Iguales ∀i
Tabla 8.1: Valores de los distintos parametros del modelo usados en la simulacion del implante. a
Whalen et al. [343], b Garcıa-Aznar [117], c Hazelwood et al. [140], d Jaworski y Lok [170], e Parfitt
et al. [263],f Hernandez [144].

8.2 Implantes completamente osteointegrados 327
resultados no se muestran, porque los problemas que presenta esta simulacion hacen que
dichos resultados no sean validos. Esos problemas estan relacionados con el dano que se
acumula en huesos muy porosos y que alcanza valores muy elevados y carentes de sentido
fisiologico. La razon de que ese dano no sea representativo reside en el hecho de que el
modelo de dano usado en esta tesis esta calibrado para hueso cortical pero no para hueso
trabecular. Al hablar del dano en el capıtulo 7 se comento que las ecuaciones en las que
se basa el modelo se habıan obtenido ajustando los ensayos de fatiga realizados por Pattin
et al. [265] en hueso cortical. Para hueso trabecular se usaron las mismas ecuaciones por
falta de datos, pero es evidente que el comportamiento del hueso trabecular en fatiga debe
ser completamente distinto, porque la estructura interna tambien lo es y los mecanismos de
acumulacion de dano dependen enormemente de la estructura. Esta limitacion del modelo es
importante, pues impide simular con ciertas garantıas un caso mas realista, en el que se parta
de un tejido periimplantario con una densidad inicial correspondiente a hueso trabecular.
Esto quiere decir que los resultados deben analizarse desde un punto de vista cualitativo, en
tanto no se mejore el modelo de crecimiento de dano en hueso trabecular.
8.2.1. Secuencia S4. Densidad inicial 2.0g/cm3 en el hueso periim-
plantario
Para la secuencia de masticacion S4 (MI1-MI2-MImp-MD2)- (MI1-MI2-MImp-MD2),. . . ,
se suponen 500 ciclos de masticacion diarios, es decir, 125 ciclos de cada carga. En la figura
8.2 se muestra la distribucion de tensiones en el hueso periimplantario para los cuatro casos
de carga simulados. Dichas distribuciones de tension corresponden al inicio de la simulacion,
cuando el dano acumulado es aun pequeno y no tiene mucha influencia en las propiedades
elasticas. Estas distribuciones de tension van a cambiar como consecuencia de la acumulacion
de dano en ciertas zonas y tambien con el cambio de la anisotropıa del tejido.
Se observa claramente que las tensiones son mucho mayores en el borde libre de la
interfase titanio - hueso (que se suele denominar cuello del implante), especialmente en el
caso MImp. Esto es debido a lo comentado anteriormente sobre los modulos de elasticidad
tan diferentes de uno y otro material. La distribucion de tensiones y deformaciones que se
obtendrıan analıticamente serıan, por lo general, singulares en la interfase. Que haya o no
singularidad depende de los angulos que forman los dos materiales que concurren en el borde
libre, de los modulos de elasticidad de dichos materiales y del tipo de union que haya entre
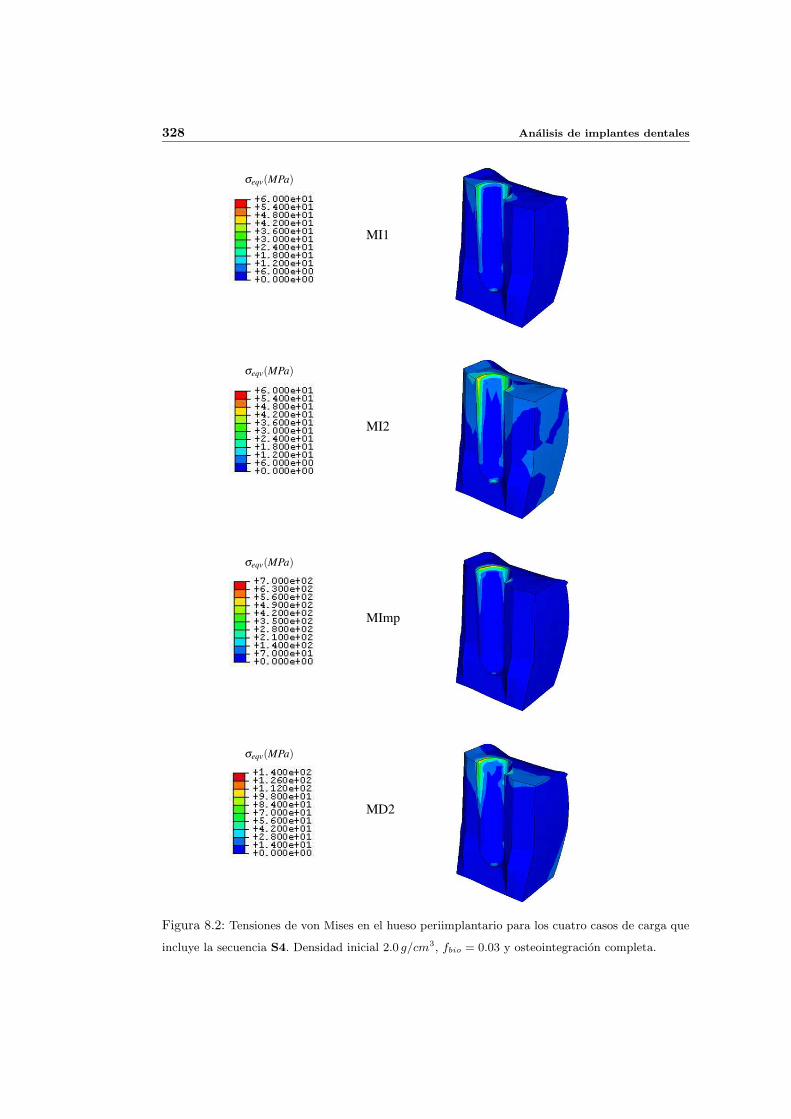
328 Analisis de implantes dentales
σeqv(MPa)
σeqv(MPa)
σeqv(MPa)
σeqv(MPa)
MI1
MI2
MImp
MD2
1
Figura 8.2: Tensiones de von Mises en el hueso periimplantario para los cuatro casos de carga que
incluye la secuencia S4. Densidad inicial 2.0 g/cm3, fbio = 0.03 y osteointegracion completa.

8.2 Implantes completamente osteointegrados 329
ellos [33, 352]. Mediante el metodo de los elementos finitos, esta singularidad no puede ser
simulada con elementos isoparametricos de interpolacion lineal como los que se usan aquı,
lo que hace que la solucion numerica de tensiones sea erronea cerca de la singularidad. Se ha
comprobado en los resultados que el error se encuentra muy localizado en los dos elementos
mas cercanos a la interfase, tanto de un lado como de otro. En esos elementos se observa
una abrupta oscilacion de la solucion de tensiones, mientras que lo esperable hubiera sido
que las tensiones crecieran monotonamente al aproximarnos a la singularidad. A pesar de
no reproducir la singularidad, el nivel de tensiones en esos dos elemento es alto, en cualquier
caso.
De entre los tres casos de carga, se observa que las mayores tensiones se dan en el
caso MImp, seguido de MD2, MI2 y por ultimo MI1 (hay que observar que la escala de
colores es diferente). Ademas, en el caso de cargas MImp, las tensiones estan especialmente
concentradas alrededor del cuello del implante. Tambien se observa que, en en todos los
casos, las tensiones en el apice del implante son muy pequenas. En las cargas MD2, MI1 y
MI2 es logico dado que la carga de la mandıbula es principalmente de flexion y esta zona
esta mas cerca de la lınea neutra. Pero lo curioso es que tambien sucede ası en el caso de
carga MImp. En este caso, parte de la carga se transmite en el apice por compresion, pero
la mayor parte de ella se transmite a lo largo de la interfase y principalmente en el cuello,
debido al elevado contraste entre la rigidez del hueso y del titanio, que se ha comentado
antes. Este hecho hace del implante un dispositivo mucho menos eficiente que el diente
natural. Este ultimo dispone del ligamento periodontal para repartir las tensiones por toda
la superficie de contacto con el hueso, de manera mas o menos uniforme. Sin embargo, el
implante carga en exceso la zona del cuello y deja practicamente en desuso el hueso apical,
lo que puede producir reabsorcion en ambos casos, ya que la mejor forma de mantener el
hueso en optimas condiciones es someterlo a un estımulo moderado.
Evolucion del dano
El hecho de que las tensiones sean especialmente elevadas en el cuello del implante
hace que en esta zona el dano comience a acumularse inmediatamente y de manera muy
notable. Conforme los elementos del hueso situados en la interfase se van danando, dejan de
absorber energıa de deformacion, que tiene que ser absorbida por los elementos vecinos. Era
de esperar que al ocurrir esto, aumentaran las tensiones soportadas por dichos elementos

330 Analisis de implantes dentales
vecinos y consecuentemente el dano. Esto harıa que el dano se fuera propagando a lo largo del
hueso, siempre que el proceso de remodelacion osea no fuera capaz de repararlo rapidamente,
al menos tan rapidamente como crece por fatiga. Sin embargo, no ocurre ası y el dano se
mantiene muy localizado en torno a la interfase. No da tiempo a que por remodelacion se
repare el dano acumulado en esta interfase, al menos este no; pero tampoco se extiende
mucho mas alla, como se puede observar en la figura 8.3. En la interfase, como consecuencia
de la singularidad de tensiones, la acumulacion de dano por fatiga es muy rapida, mas que
la reparacion y por ello el dano permanece estable en niveles muy elevados. Pero lejos de
la interfase los niveles de deformacion son moderados y la remodelacion osea es capaz de
reparar el dano y mantenerlo dentro de unos niveles fisiologicos normales. En la figura 8.3
se muestra la distribucion de las componentes del tensor de dano en direcciones longitudinal
(x), transversal (y) y vertical (z).
Lo primero que llama la atencion es que haya una buena parte del modelo en la que el
dano esta fuera del rango de variacion permitido [0, 1]. Este hecho merece, que duda cabe, una
explicacion. El tensor de dano se interpreta en el modelo como una medida de la degradacion
de las propiedades elasticas. Ası, un dano nulo significa que el tejido mantiene intactas sus
propiedades y un dano unidad, que su rigidez es nula.1 Para mantener esta interpretacion
y la coherencia de los resultados, se impone en el algoritmo que los autovalores del tensor
de dano esten entre 0 y 1. Esa restriccion se impone en los puntos de integracion de Gauss
del modelo de elementos finitos, que es donde se aplica el algoritmo de remodelacion y se
actualizan las propiedades elasticas. Los mostrados en todas las figuras de este capıtulo
son resultados nodales, obtenidos extrapolando a cada nodo los resultados de los puntos
de Gauss vecinos. Esta extrapolacion puede hacer que los resultados nodales esten fuera
de rango, pero ello no constituye problema alguno, ya que donde se tiene que cumplir la
restriccion es en los puntos de integracion, que es donde se evaluan las propiedades, y ahı el
dano se mantiene obligatoriamente entre 0 y 1.
En la figura 8.3 se puede observar lo comentado anteriormente: que la zona con dano
elevado esta muy localizada en torno al implante y ademas apenas se extiende con el tiempo.
El mayor dano se produce en direccion x, en la que las tensiones son mayores debido a la
flexion de la mandıbula, aunque las diferencias con el dano en las otras direcciones no es muy
importante. Tambien se observa que en la zona apical, en la base del implante, hay un dano
1Al ser el dano direccional, la rigidez se refiere a la direccion correspondiente
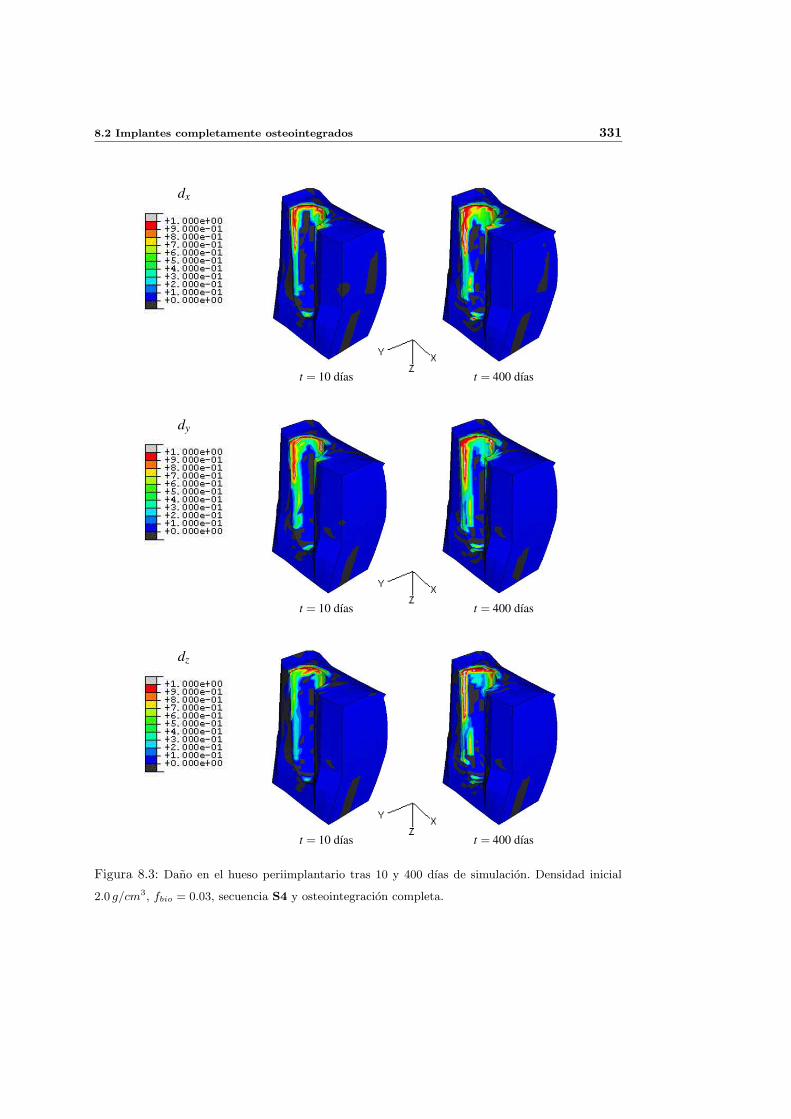
8.2 Implantes completamente osteointegrados 331
dx
t = 10 dıas t = 400 dıas
dy
t = 10 dıas t = 400 dıas
dz
t = 10 dıas t = 400 dıas
1
Figura 8.3: Dano en el hueso periimplantario tras 10 y 400 dıas de simulacion. Densidad inicial
2.0 g/cm3, fbio = 0.03, secuencia S4 y osteointegracion completa.

332 Analisis de implantes dentales
muy elevado. Se ha podido comprobar que el dano en esta zona aumenta principalmente
por la aplicacion del caso de carga MImp. En esta carga, al subir la mandıbula por la
accion de los musculos masticatorios, el implante, que tiene los desplazamientos verticales
impedidos, se clava en el hueso que tiene justo debajo. La compresion en direccion z produce
en las otras dos direcciones deformaciones de traccion, que son las responsables del dano.
De hecho, se puede observar que en esta zona el dano en direccion z es menor que en las
otras dos direcciones, ya que con la hipotesis de crecimiento del dano adoptada en esta tesis,
las deformaciones de compresion no producen dano. El dano en direccion z de esta zona se
debe a otros casos de carga que produzcan tracciones, que no al MImp. Como esos casos de
cargas producen tensiones mas pequenas, el dano tambien es mas pequeno.
En la figura 8.4 se vuelve a representar el dano en las tres direcciones, pero ahora en un
rango mas reducido, entre 0 y 0.02. El maximo de ese rango es un valor aun elevado, pero mas
cerca del valor normal que el hueso puede tener como consecuencia de su actividad. Aparte de
la zona fuera de rango, que se circunscribe al hueso que rodea al implante, se pueden observar
dos zonas de dano elevado, a uno y otro lado del implante, en direccion mesio-distal. Este
dano es debido principalmente a la flexion de la mandıbula durante la masticacion, hecho
que se ve agravado por la existencia de un dano elevado en la region del cuello del implante,
que hace que sea menor la cantidad de hueso con capacidad suficiente para absorber energıa
de deformacion. Este dano, ciertamente elevado, producira reabsorcion en esta zona, como
se vera en el apartado siguiente donde se analiza la evolucion de la porosidad.
Se observa tambien en la figura 8.4 que el dano en direccion z es muy pequeno, salvo
cerca del implante, donde esta fuera del rango representado y es muy proximo a 1 como se
puede apreciar en la figura 8.3. Este dano dz tan localizado se acumula casi exclusivamente
por la carga MImp que hace que se clave el implante en el hueso periimplantario.
Como ya se ha dicho, el dano se acumula en la interfase entre el hueso y el implante,
como consecuencia de la singularidad de tensiones. El dano crece muy rapidamente en los
primeros dıas y luego se estabiliza, como tambien quedo patente en la figura 8.3. Sin embargo,
en su evolucion durante los primeros dıas (ver figura 8.5) se observa como el dano crece a
partir del cuello del implante hacia abajo, siempre a lo largo de la interfase hueso-implante.
Esa propagacion tan rapida sucede por la siguiente razon: en cualquier punto de la interfase
donde se tenga un elemento muy danado junto a otro no danado existe una alta concentracion
de tensiones. En efecto, la rigidez de un elemento altamente danado es muy baja, ası, en
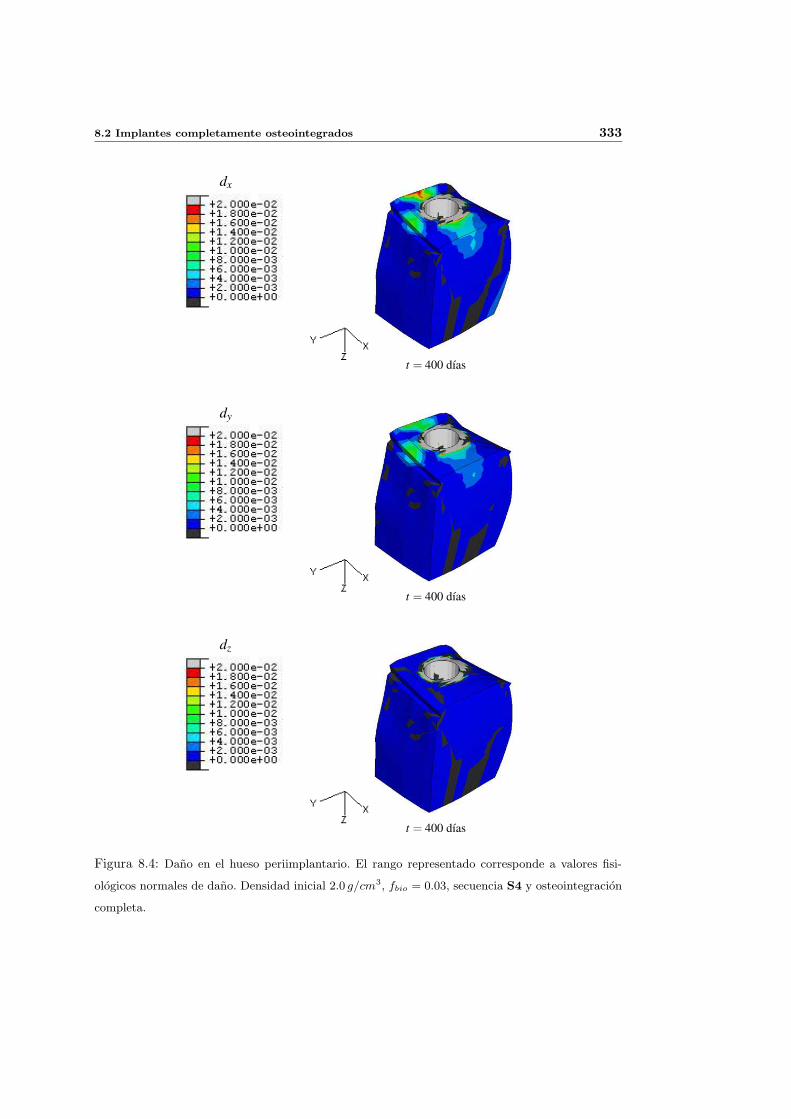
8.2 Implantes completamente osteointegrados 333
dx
t = 400 dıas
dy
t = 400 dıas
dz
t = 400 dıas
1
Figura 8.4: Dano en el hueso periimplantario. El rango representado corresponde a valores fisi-
ologicos normales de dano. Densidad inicial 2.0 g/cm3, fbio = 0.03, secuencia S4 y osteointegracion
completa.
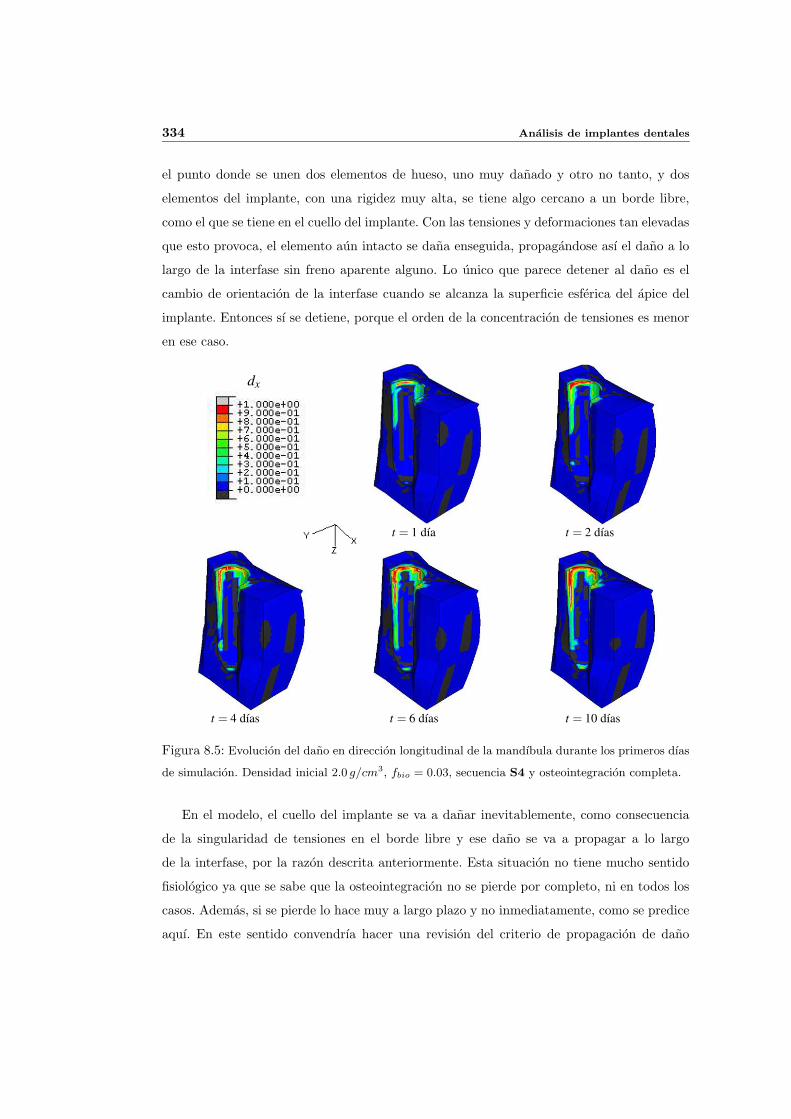
334 Analisis de implantes dentales
el punto donde se unen dos elementos de hueso, uno muy danado y otro no tanto, y dos
elementos del implante, con una rigidez muy alta, se tiene algo cercano a un borde libre,
como el que se tiene en el cuello del implante. Con las tensiones y deformaciones tan elevadas
que esto provoca, el elemento aun intacto se dana enseguida, propagandose ası el dano a lo
largo de la interfase sin freno aparente alguno. Lo unico que parece detener al dano es el
cambio de orientacion de la interfase cuando se alcanza la superficie esferica del apice del
implante. Entonces sı se detiene, porque el orden de la concentracion de tensiones es menor
en ese caso.
dx
t = 1 dıa t = 2 dıas
t = 4 dıas t = 6 dıas t = 10 dıas
1
Figura 8.5: Evolucion del dano en direccion longitudinal de la mandıbula durante los primeros dıas
de simulacion. Densidad inicial 2.0 g/cm3, fbio = 0.03, secuencia S4 y osteointegracion completa.
En el modelo, el cuello del implante se va a danar inevitablemente, como consecuencia
de la singularidad de tensiones en el borde libre y ese dano se va a propagar a lo largo
de la interfase, por la razon descrita anteriormente. Esta situacion no tiene mucho sentido
fisiologico ya que se sabe que la osteointegracion no se pierde por completo, ni en todos los
casos. Ademas, si se pierde lo hace muy a largo plazo y no inmediatamente, como se predice
aquı. En este sentido convendrıa hacer una revision del criterio de propagacion de dano

8.2 Implantes completamente osteointegrados 335
usado en esta tesis, que permitiera introducir algun tipo de barrera a dicha propagacion.
Una posible forma de solventar el problema serıa definir un criterio energetico de crecimiento
de dano a nivel global, porque esta claro que las deformaciones locales no sirven para estimar
el crecimiento del dano, al menos cuando existen altas concentraciones de tension, en las que
el dano estimado mediante las deformaciones locales no deja de propagarse como se acaba
de ver.
Otra limitacion del modelo es que no considera que la osteointegracion se puede perder.
Cuando en el parrafo anterior se ha hablado de perdida de osteointegracion, se hacıa ref-
erencia en realidad a un dano de la interfase, pero no a una falta de union entre hueso e
implante. Como ya se dijo en el capıtulo 3, se supone que la osteointegracion, simulada como
una union perfecta entre los elementos de hueso e implante, que comparten los nodos de la
interfase, no se pierde durante el analisis. Los elementos de la interfase siguen compartiendo
esos nodos aunque el hueso se dane por completo. Esto que, sin duda, influye en la propa-
gacion del dano a lo largo de la interfase, que como se ha comentado parece incongruente
con la realidad, se podrıa resolver utilizando un modelo de evolucion de la union como el
que han propuesto Moreo et al. [270] en protesis de cadera cementadas.
Para evaluar de forma global la cantidad de tejido danado, se ha definido una fraccion
de volumen danado promedio en el hueso periimplantario, segun la expresion
h =
∫Vperi
h dVT∫Vperi
dVT(8.1)
donde h = Vd/Vt es la fraccion de tejido danado sobre el volumen total. En la figura 8.6
se representa la evolucion de h en esta simulacion. Se observa como el dano crece muy
rapidamente al principio para estabilizarse despues. Aunque aun esta creciendo cuando se
interrumpe la simulacion, el crecimiento no es muy acusado, como tampoco lo es el dano
promedio. De hecho, si se recuerda la relacion existente entre h y el tensor de dano (h =
k tr(d/3) siendo la constante k = 0.000333), el promedio de la traza del tensor de dano
esta en torno a 0.06, valor que se puede considerar dentro de la normalidad.
Evolucion de la fraccion volumetrica de hueso
Segun la correlacion dada por Hernandez [145] entre la densidad del tejido oseo y la
fraccion de ceniza,
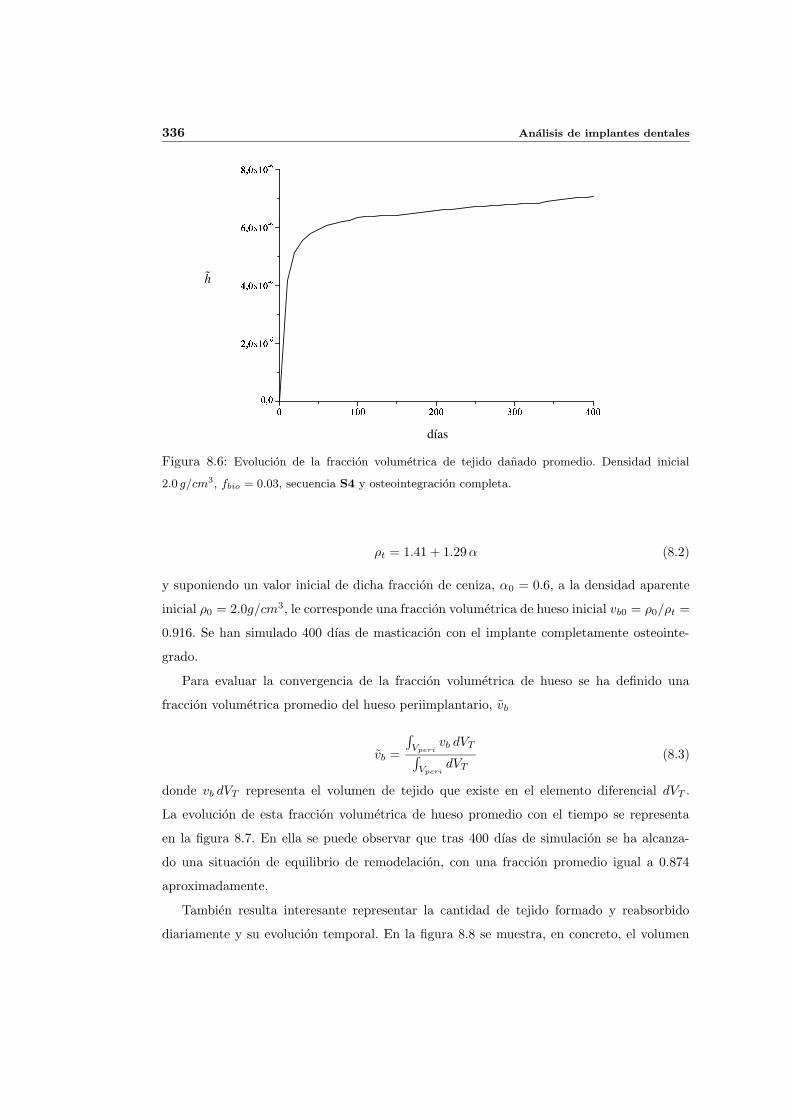
336 Analisis de implantes dentales
.
.
.
.
dıas
h
1
Figura 8.6: Evolucion de la fraccion volumetrica de tejido danado promedio. Densidad inicial
2.0 g/cm3, fbio = 0.03, secuencia S4 y osteointegracion completa.
ρt = 1.41 + 1.29α (8.2)
y suponiendo un valor inicial de dicha fraccion de ceniza, α0 = 0.6, a la densidad aparente
inicial ρ0 = 2.0g/cm3, le corresponde una fraccion volumetrica de hueso inicial vb0 = ρ0/ρt =
0.916. Se han simulado 400 dıas de masticacion con el implante completamente osteointe-
grado.
Para evaluar la convergencia de la fraccion volumetrica de hueso se ha definido una
fraccion volumetrica promedio del hueso periimplantario, vb
vb =
∫Vperi
vb dVT∫Vperi
dVT(8.3)
donde vb dVT representa el volumen de tejido que existe en el elemento diferencial dVT .
La evolucion de esta fraccion volumetrica de hueso promedio con el tiempo se representa
en la figura 8.7. En ella se puede observar que tras 400 dıas de simulacion se ha alcanza-
do una situacion de equilibrio de remodelacion, con una fraccion promedio igual a 0.874
aproximadamente.
Tambien resulta interesante representar la cantidad de tejido formado y reabsorbido
diariamente y su evolucion temporal. En la figura 8.8 se muestra, en concreto, el volumen
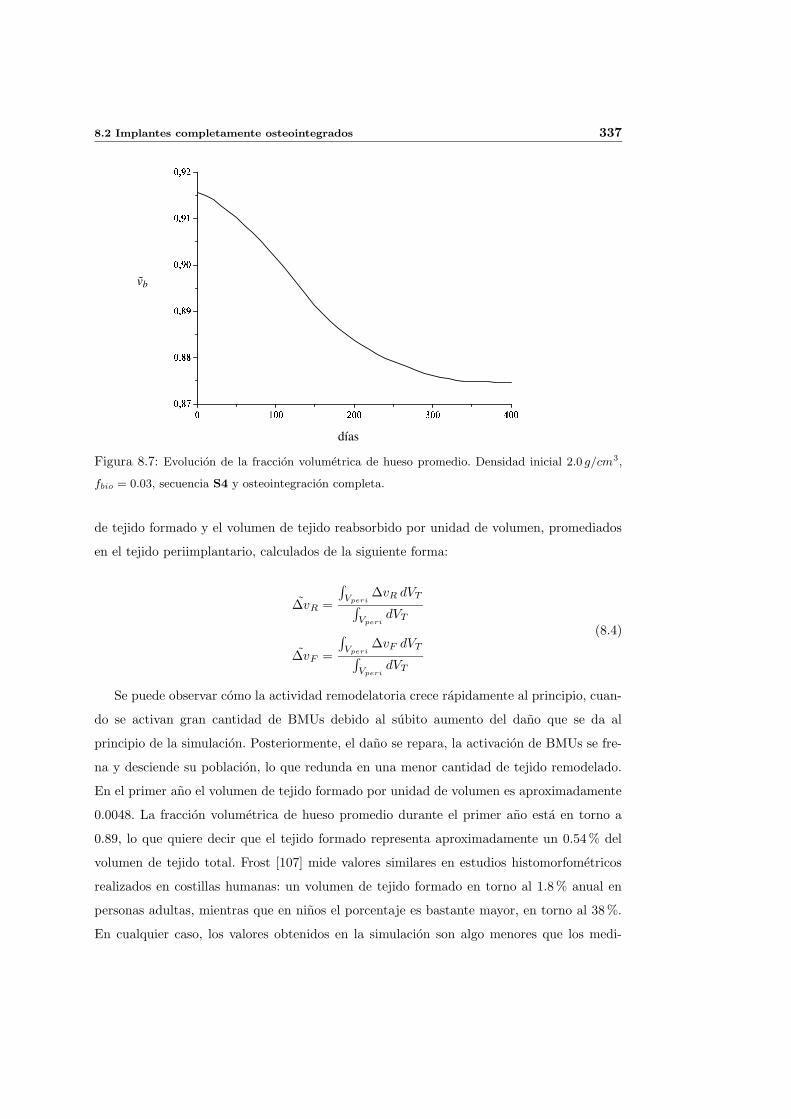
8.2 Implantes completamente osteointegrados 337
¡ ¢ £¤ ¥¦
¤ ¥¥
¤ ¥§
¤ §
¤ §B
¤ §¡
dıas
vb
1
Figura 8.7: Evolucion de la fraccion volumetrica de hueso promedio. Densidad inicial 2.0 g/cm3,
fbio = 0.03, secuencia S4 y osteointegracion completa.
de tejido formado y el volumen de tejido reabsorbido por unidad de volumen, promediados
en el tejido periimplantario, calculados de la siguiente forma:
∆vR =
∫Vperi
∆vR dVT∫Vperi
dVT
∆vF =
∫Vperi
∆vF dVT∫Vperi
dVT
(8.4)
Se puede observar como la actividad remodelatoria crece rapidamente al principio, cuan-
do se activan gran cantidad de BMUs debido al subito aumento del dano que se da al
principio de la simulacion. Posteriormente, el dano se repara, la activacion de BMUs se fre-
na y desciende su poblacion, lo que redunda en una menor cantidad de tejido remodelado.
En el primer ano el volumen de tejido formado por unidad de volumen es aproximadamente
0.0048. La fraccion volumetrica de hueso promedio durante el primer ano esta en torno a
0.89, lo que quiere decir que el tejido formado representa aproximadamente un 0.54 % del
volumen de tejido total. Frost [107] mide valores similares en estudios histomorfometricos
realizados en costillas humanas: un volumen de tejido formado en torno al 1.8 % anual en
personas adultas, mientras que en ninos el porcentaje es bastante mayor, en torno al 38 %.
En cualquier caso, los valores obtenidos en la simulacion son algo menores que los medi-
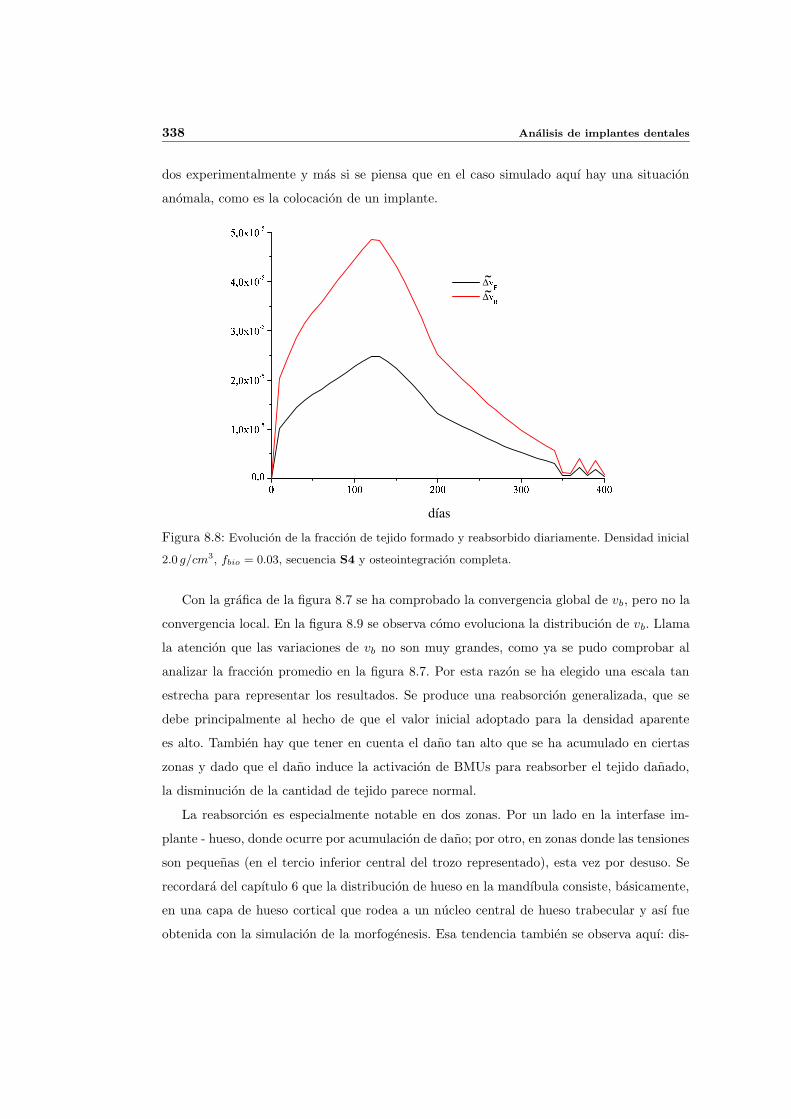
338 Analisis de implantes dentales
dos experimentalmente y mas si se piensa que en el caso simulado aquı hay una situacion
anomala, como es la colocacion de un implante.
¨ ©4¨S¨ ªS¨S¨ «¨S¨ ¬S¨S¨¨ ¨
©4 ¨®©4¨¯ °
ªS ¨®©4¨¯ °
« ¨®©4¨¯ °
¬S ¨®©4¨¯ °
± ¨®©4¨¯ °
∆²³∆²´
dıas
˜˜
1
Figura 8.8: Evolucion de la fraccion de tejido formado y reabsorbido diariamente. Densidad inicial
2.0 g/cm3, fbio = 0.03, secuencia S4 y osteointegracion completa.
Con la grafica de la figura 8.7 se ha comprobado la convergencia global de vb, pero no la
convergencia local. En la figura 8.9 se observa como evoluciona la distribucion de vb. Llama
la atencion que las variaciones de vb no son muy grandes, como ya se pudo comprobar al
analizar la fraccion promedio en la figura 8.7. Por esta razon se ha elegido una escala tan
estrecha para representar los resultados. Se produce una reabsorcion generalizada, que se
debe principalmente al hecho de que el valor inicial adoptado para la densidad aparente
es alto. Tambien hay que tener en cuenta el dano tan alto que se ha acumulado en ciertas
zonas y dado que el dano induce la activacion de BMUs para reabsorber el tejido danado,
la disminucion de la cantidad de tejido parece normal.
La reabsorcion es especialmente notable en dos zonas. Por un lado en la interfase im-
plante - hueso, donde ocurre por acumulacion de dano; por otro, en zonas donde las tensiones
son pequenas (en el tercio inferior central del trozo representado), esta vez por desuso. Se
recordara del capıtulo 6 que la distribucion de hueso en la mandıbula consiste, basicamente,
en una capa de hueso cortical que rodea a un nucleo central de hueso trabecular y ası fue
obtenida con la simulacion de la morfogenesis. Esa tendencia tambien se observa aquı: dis-

8.2 Implantes completamente osteointegrados 339
minuye la densidad del hueso en la parte central, que apenas esta siendo util desde el punto
de vista resistente. El hueso que rodea al apice del implante aumenta su porosidad como se
puede comprobar en la figura 8.10.
El objetivo perseguido con el nuevo modelo de remodelacion es simular la reabsorcion
osea que se observa en algunos casos clınicos. En este sentido, las figuras 8.9 y 8.10 parecen
confirmar que, efectivamente, la acumulacion de dano microestructural en esa zona hace
que se reabsorba hueso localmente. La reabsorcion por dano es mas acusada, evidentemente,
allı donde el dano acumulado es mayor, es decir en la interfase. En la figura 8.10 se observa
que, una vez alcanzado el equilibrio de remodelacion, la zona adyacente al implante presenta
una porosidad mas alta que en el resto. El implante queda practicamente rodeado de una
capa de elementos con un dano muy elevado y por tanto muy flexibles. Esta capa se hace
ligeramente mas gruesa en la zona del cuello, especialmente en el lado mesial (derecho en
las secciones de la figura 8.10) y en la zona inferior distal (izquierdo) donde predominan las
tracciones.
Se ha analizado con algo mas de detalle la evolucion de la fraccion volumetrica de hueso,
vb, en algunos puntos de la interfase. Los resultados se muestran en la figura 8.11 y se puede
comprobar que, al menos en esos puntos, se ha alcanzado la convergencia local de vb. Todos
los puntos representados en la figura sufren reabsorcion pero la distribucion por zonas no
sigue un patron definido, como tambien se puede apreciar en la figura 8.10, en la que se
observa una gran variacion de vb por toda la interfase. Se puede destacar, quiza, el apice,
la region inferior mesial y medio labial, por tener un vb elevado. El resultado de las dos
primeras se pueden corroborar en la figura 8.10. Estas zonas reciben cargas de compresion
principalmente, por lo que no aumenta el dano, con el modelo de crecimiento de dano
propuesto aquı. Sin embargo, el estımulo mecanico sı es elevado y eso contribuye a mantener
una densidad osea alta. Tambien destaca la zona distal y especialmente la region superior, por
todo lo contrario, una vb menor que en el resto. Esto es debido al dano tan alto que acumula
la interfase de este lado. Se observa en la figura 8.5 que el dano comienza a acumularse en el
borde superior distal y a partir de ahı se propaga en direccion circunferencial y hacia abajo.
En esta direccion de manera muy notable, ya que consigue danar todo el borde distal en
pocos dıas. Este dano tan elevado es el responsable de la activacion de muchas BMUs para
intentar repararlo y su efecto se nota claramente en el descenso de vb.
La anterior conclusion sobre la region distal es valida solo en la interfase, ya que, como se
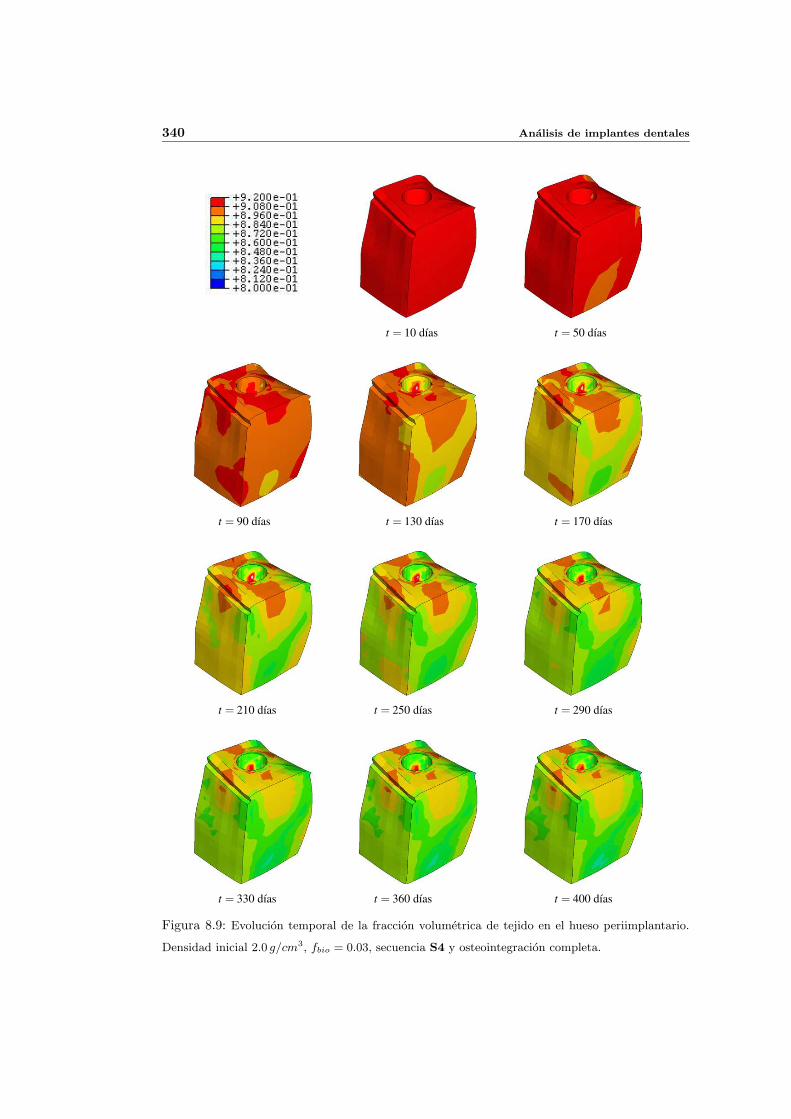
340 Analisis de implantes dentales
t = 10 dıas t = 50 dıas
t = 90 dıas t = 130 dıas t = 170 dıas
t = 210 dıas t = 250 dıas t = 290 dıas
t = 330 dıas t = 360 dıas t = 400 dıas
1
Figura 8.9: Evolucion temporal de la fraccion volumetrica de tejido en el hueso periimplantario.
Densidad inicial 2.0 g/cm3, fbio = 0.03, secuencia S4 y osteointegracion completa.
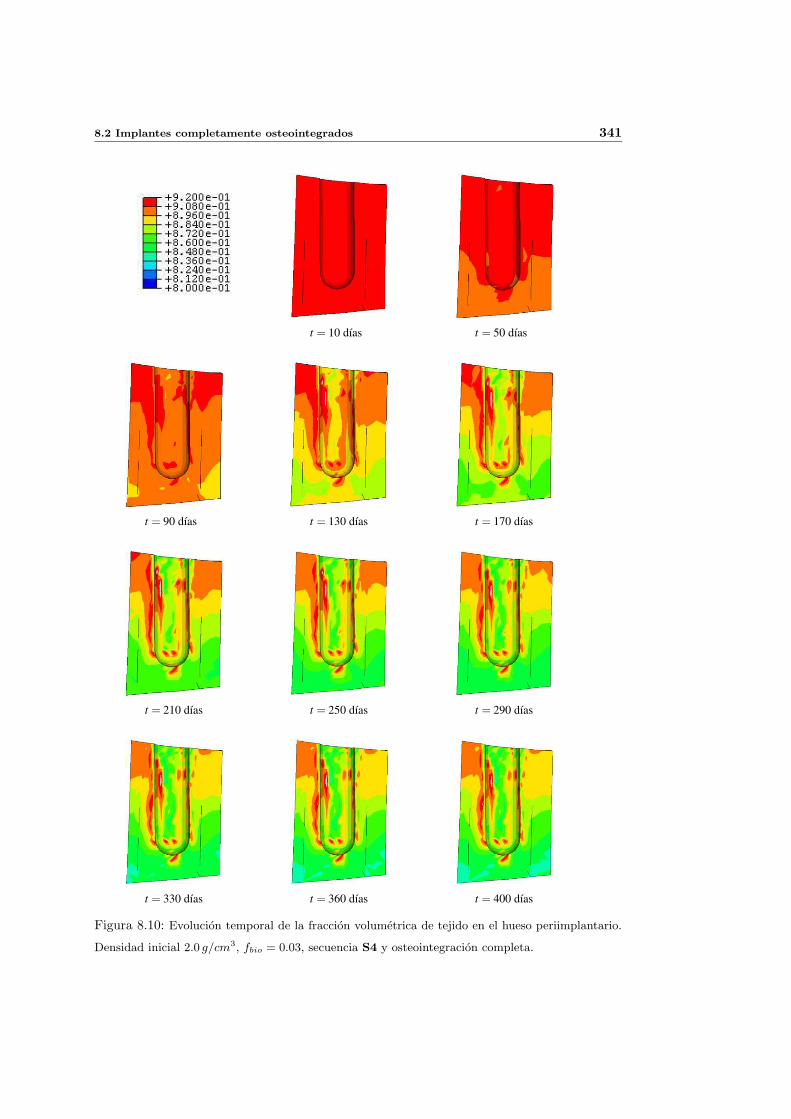
8.2 Implantes completamente osteointegrados 341
t = 10 dıas t = 50 dıas
t = 90 dıas t = 130 dıas t = 170 dıas
t = 210 dıas t = 250 dıas t = 290 dıas
t = 330 dıas t = 360 dıas t = 400 dıas
1
Figura 8.10: Evolucion temporal de la fraccion volumetrica de tejido en el hueso periimplantario.
Densidad inicial 2.0 g/cm3, fbio = 0.03, secuencia S4 y osteointegracion completa.
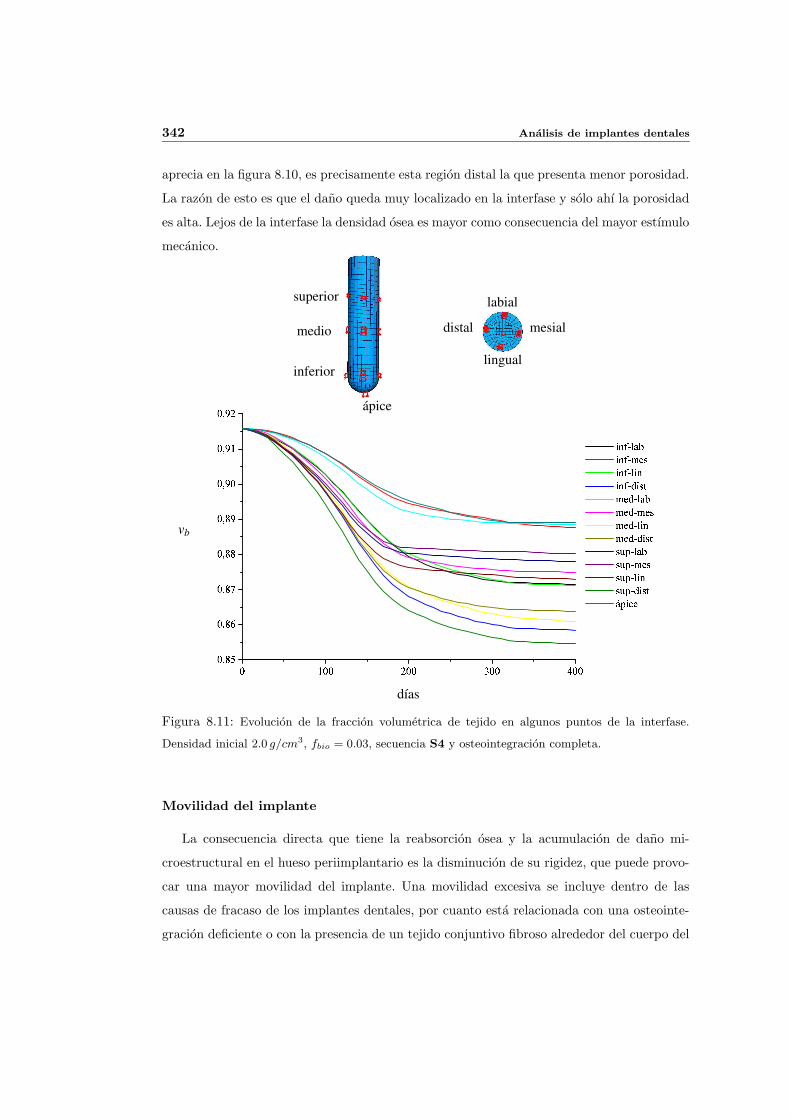
342 Analisis de implantes dentales
aprecia en la figura 8.10, es precisamente esta region distal la que presenta menor porosidad.
La razon de esto es que el dano queda muy localizado en la interfase y solo ahı la porosidad
es alta. Lejos de la interfase la densidad osea es mayor como consecuencia del mayor estımulo
mecanico.
µ ¶4µSµ ·SµSµ ¸µSµ ¹SµSµµº »¼
µº »½
µº »¾
µº »»
µº »¿
µº ¿µ
µº ¿¶
µº ¿·
À ÁBÂ Ã Ä ÅÆÀ ÁBÂ Ã ÇÉÈÊÀ ÁBÂ Ã Ä À ÁÀ ÁBÂ Ã ËÀ Ê ÌÇÉÈËÃ Ä ÅÆÇÉÈËÃ ÇÍÈÊÇÉÈËÃ Ä À ÁÇÉÈËÃ ËÀ Ê ÌÊ ÎSÏBÃ Ä Å4ÆÊ ÎSÏBÃ ÇÍÈÊÊ ÎSÏBÃ Ä À ÁÊ ÎSÏBÃ ËÀ Ê ÌÐ ÏBÀ Ñ4È
distal mesial
lingual
labialsuperior
medio
inferior
apice
dıas
vb
1
Figura 8.11: Evolucion de la fraccion volumetrica de tejido en algunos puntos de la interfase.
Densidad inicial 2.0 g/cm3, fbio = 0.03, secuencia S4 y osteointegracion completa.
Movilidad del implante
La consecuencia directa que tiene la reabsorcion osea y la acumulacion de dano mi-
croestructural en el hueso periimplantario es la disminucion de su rigidez, que puede provo-
car una mayor movilidad del implante. Una movilidad excesiva se incluye dentro de las
causas de fracaso de los implantes dentales, por cuanto esta relacionada con una osteointe-
gracion deficiente o con la presencia de un tejido conjuntivo fibroso alrededor del cuerpo del

8.2 Implantes completamente osteointegrados 343
implante [312]. Ademas, la posibilidad de recuperar la osteointegracion es tanto mas remota
cuanto mayor es la movilidad, por lo que la perdida del implante puede ser irreversible. Segun
Misch [238], los implantes que tienen menos de 0.5 mm de desplazamiento horizontal pueden
recuperar la rigidez de fijacion y la movilidad cero. En caso contrario, la recuperacion de la
osteointegracion es mas improbable. Este autor establece que un implante que tenga mas de
un 1 mm de movilidad horizontal o cualquier grado de movilidad vertical no debe entrar en
carga, para evitar la perdida continuada de tejido oseo y el deterioro del emplazamiento del
implante.
Para medir la movilidad se puede utilizar un dispositivo denominado Periotest r©(Siemens)
que mide el tiempo que dura el impacto entre una varilla metalica y el implante; tiempo que
se correlaciona con la rigidez de la fijacion, es decir, con la movilidad. En esta tesis no se ha
simulado este ensayo de impacto. En cualquier caso, el ensayo esta pensado para evitar tener
que medir la movilidad en carga, durante la masticacion, que es difıcil de medir a efectos
practicos, pero que en realidad es lo que importa. Aquı se pretende evaluar precisamente eso:
medir los desplazamientos que tienen lugar durante la masticacion. En concreto durante la
masticacion con el implante, que es el caso de carga mas comprometido. Segun Misch [238],
la movilidad mas peligrosa es la vertical y por ello se analizan los desplazamientos en esta
direccion. Misch se refiere en realidad a la movilidad segun el eje del implante, que no tiene
por que ser vertical obligatoriamente. En la figura 8.12 se muestran estos desplazamien-
tos, en la direccion axial del implante, la representada como y, para el ultimo dıa de carga
simulado, es decir, una vez asegurada la convergencia.
Hay que hacer notar que la movilidad relativa entre cada una de las partes de la interfase
hueso-implante es nula por definicion, ya que se ha supuesto osteointegracion perfecta. Pero
mas alla de lo que ocurre en la interfase, los resultados son interesantes, en tanto que el
dano y la perdida de rigidez de la capa de hueso en contacto con el implante afecta a
los desplazamientos del hueso un poco mas alejado de aquel. De este modo, aunque los
desplazamientos de los nodos de la interfase sean los mismos en el implante y en el hueso,
los desplazamientos de una capa un poco mas alejada, sı pueden servir para estimar la
movilidad global del implante. Ası, en la figura 8.12 se observa una cierta diferencia entre
los desplazamientos del implante y los del hueso en el lado distal, siendo algo menor esa
diferencia en el lado mesial. Los desplazamientos relativos entre el hueso y el implante son
los que dan una idea de la movilidad. En el lado distal estan en torno a 30µm, a la altura del

344 Analisis de implantes dentales
uy(mm)
distalmesial
x
y
P
1
Figura 8.12: Desplazamientos en la direccion axial del implante en el caso de carga de masticacion
con el implante. Densidad inicial 2.0 g/cm3, fbio = 0.03, secuencia S4 y osteointegracion completa.
Se muestra solo la zona crestal del implante y el hueso periimplantario.
cuello, mientras que en el lado mesial son del orden de 10µm. Estos valores estan muy por
debajo de 1 mm, valor establecido como lımite para la movilidad horizontal por Misch [238].
Bien es cierto que este autor no permite movilidad vertical alguna, pero tambien lo es que
el valor obtenido para la movilidad vertical es dos ordenes inferior al lımite de la movilidad
horizontal y por tanto, se puede considerar despreciable.
Es resenable que la movilidad sea maxima en el lado distal, que como se ha dicho antes
es el lado que presenta una mejor integridad. Sin embargo, tambien se ha dicho que en es-
ta zona distal el dano y la reabsorcion estan muy localizados, en los elementos adyacentes
al implante. Eso es precisamente lo que se observa en los desplazamientos, que sufren una
variacion importante, pero en una zona relativamente estrecha, donde el dano microstruc-
tural acumulado es muy alto.
Tambien resulta interesante conocer la evolucion temporal de los desplazamientos rel-
ativos entre el implante y el hueso periimplantario. En particular, la carga en la que se
mastica con el implante es la que produce mayores desplazamientos relativos y sera la que se
analice aquı. Dado que con dicha carga se impiden los desplazamientos axiales (en direccion
y) de la cabeza del implante, los desplazamientos del hueso periimplantario dan una idea
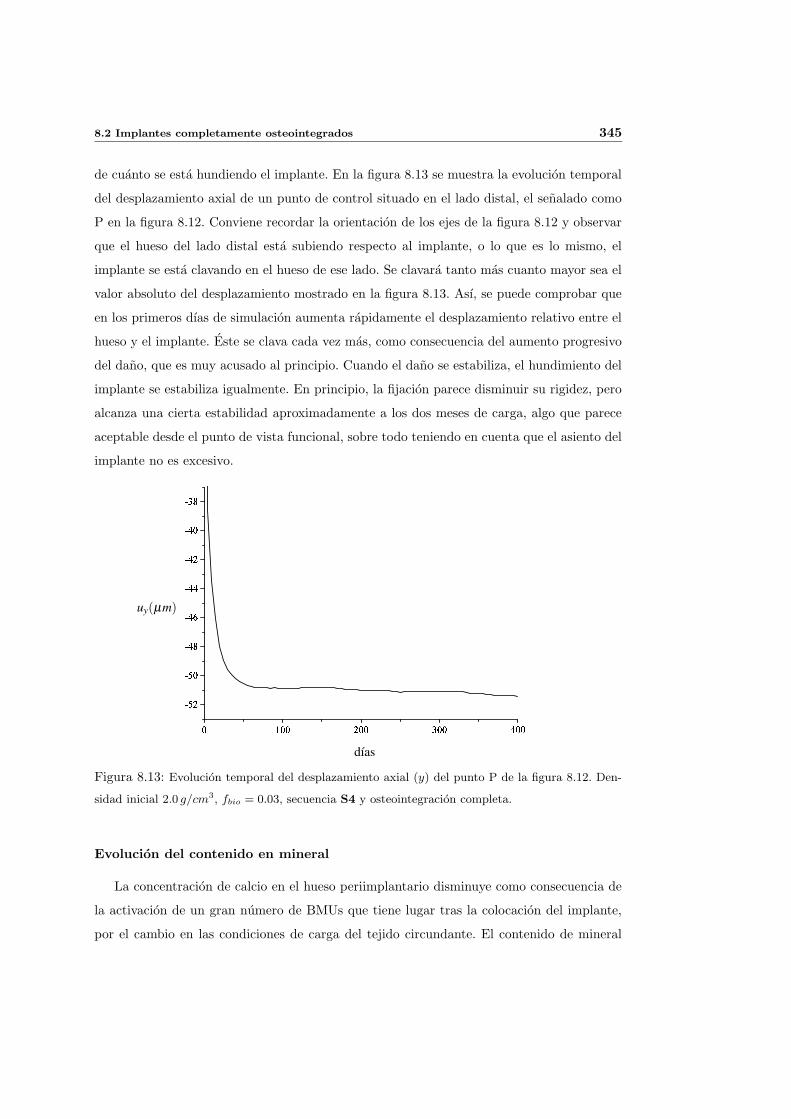
8.2 Implantes completamente osteointegrados 345
de cuanto se esta hundiendo el implante. En la figura 8.13 se muestra la evolucion temporal
del desplazamiento axial de un punto de control situado en el lado distal, el senalado como
P en la figura 8.12. Conviene recordar la orientacion de los ejes de la figura 8.12 y observar
que el hueso del lado distal esta subiendo respecto al implante, o lo que es lo mismo, el
implante se esta clavando en el hueso de ese lado. Se clavara tanto mas cuanto mayor sea el
valor absoluto del desplazamiento mostrado en la figura 8.13. Ası, se puede comprobar que
en los primeros dıas de simulacion aumenta rapidamente el desplazamiento relativo entre el
hueso y el implante. Este se clava cada vez mas, como consecuencia del aumento progresivo
del dano, que es muy acusado al principio. Cuando el dano se estabiliza, el hundimiento del
implante se estabiliza igualmente. En principio, la fijacion parece disminuir su rigidez, pero
alcanza una cierta estabilidad aproximadamente a los dos meses de carga, algo que parece
aceptable desde el punto de vista funcional, sobre todo teniendo en cuenta que el asiento del
implante no es excesivo.
Ò Ó4ÒÒ ÔÒÒ ÕÒÒ ÖÒÒ× Ø Ô
× Ø Ò
× ÖÙ
× ÖÚ
× ÖÖ
× ÖÔ
× ÖÒ
× ÕÙ
dıas
uy(µm)
1
Figura 8.13: Evolucion temporal del desplazamiento axial (y) del punto P de la figura 8.12. Den-
sidad inicial 2.0 g/cm3, fbio = 0.03, secuencia S4 y osteointegracion completa.
Evolucion del contenido en mineral
La concentracion de calcio en el hueso periimplantario disminuye como consecuencia de
la activacion de un gran numero de BMUs que tiene lugar tras la colocacion del implante,
por el cambio en las condiciones de carga del tejido circundante. El contenido de mineral
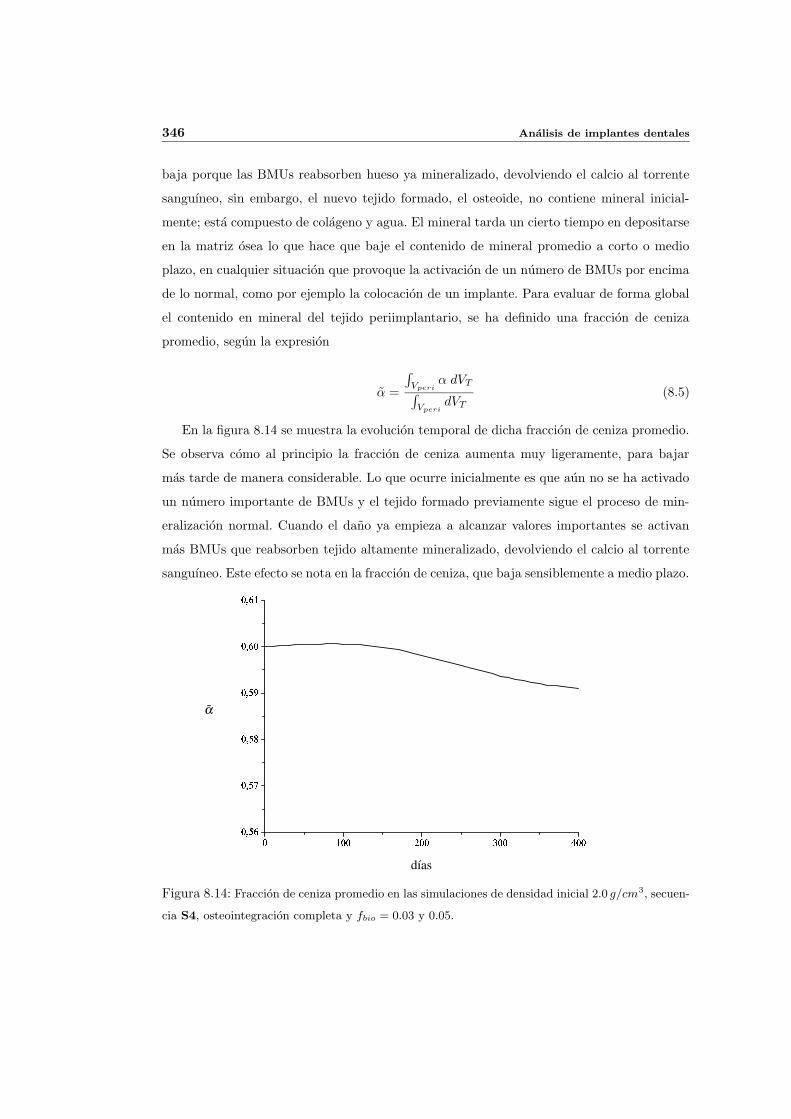
346 Analisis de implantes dentales
baja porque las BMUs reabsorben hueso ya mineralizado, devolviendo el calcio al torrente
sanguıneo, sin embargo, el nuevo tejido formado, el osteoide, no contiene mineral inicial-
mente; esta compuesto de colageno y agua. El mineral tarda un cierto tiempo en depositarse
en la matriz osea lo que hace que baje el contenido de mineral promedio a corto o medio
plazo, en cualquier situacion que provoque la activacion de un numero de BMUs por encima
de lo normal, como por ejemplo la colocacion de un implante. Para evaluar de forma global
el contenido en mineral del tejido periimplantario, se ha definido una fraccion de ceniza
promedio, segun la expresion
α =
∫Vperi
α dVT∫Vperi
dVT(8.5)
En la figura 8.14 se muestra la evolucion temporal de dicha fraccion de ceniza promedio.
Se observa como al principio la fraccion de ceniza aumenta muy ligeramente, para bajar
mas tarde de manera considerable. Lo que ocurre inicialmente es que aun no se ha activado
un numero importante de BMUs y el tejido formado previamente sigue el proceso de min-
eralizacion normal. Cuando el dano ya empieza a alcanzar valores importantes se activan
mas BMUs que reabsorben tejido altamente mineralizado, devolviendo el calcio al torrente
sanguıneo. Este efecto se nota en la fraccion de ceniza, que baja sensiblemente a medio plazo.
Û ÜÛÛ ÝÛÛ ÞÛÛ ßÛÛÛà áâ
Ûà áã
Ûà áä
Ûà áå
Ûà âÛ
Ûà âBÜ
dıas
α
1
Figura 8.14: Fraccion de ceniza promedio en las simulaciones de densidad inicial 2.0 g/cm3, secuen-
cia S4, osteointegracion completa y fbio = 0.03 y 0.05.

8.2 Implantes completamente osteointegrados 347
Evolucion de las propiedades elasticas
A continuacion se muestra la evolucion temporal de las propiedades elasticas en algunos
puntos del hueso periimplantario.
En la figura 8.15 se muestran los modulos de Young en direccion longitudinal (x),
transversal (y) y vertical (z), en puntos muy proximos al implante, concretamente en la
segunda capa. Aunque la porosidad no es demasiado baja porque la reabsorcion no ha sido
excesiva, los valores del modulo de Young son relativamente pequenos. Hay que tener en
cuenta que el dano acumulado en esa zona es bastante alto y eso hace que disminuya la
rigidez sustancialmente. Especialmente alto es el dano en D, punto situado en la cara medi-
al, de ahı que las propiedades sean sensiblemente peores a las del resto de puntos. La cara
labial no se distingue por tener un dano especialmente elevado y, sin embargo, el punto A
tiene una rigidez muy pequena, pero es debido a que este punto esta mas cerca del implante
que los demas.
Las variaciones que sufre la rigidez del elemento B son debidas a las variaciones del dano,
que en ese elemento son algo bruscas. Esas variaciones bruscas se deben a que en esa zona
el dano no es demasiado alto y en parte se repara por remodelacion, a diferencia de las
otras zonas representadas, en las que el aumento del dano por la carga es mas rapido que la
reparacion por remodelacion y el dano se mantiene en valores elevados.
Dado que en la cercanıa del implante la rigidez esta muy influenciada por el dano, la
direccion de menor rigidez coincide con la de mayor dano. Ası, en los puntos A y D, el dano
es mayor en direccion normal a la superficie del implante (y para el punto A y x para el
punto D) y en esas direcciones se tiene la menor rigidez. La razon de que esas direcciones
sean las de mayor dano reside en el hecho de que en esa zona el implante esta tirando del
hueso (hay que recordar que se considera osteointegracion perfecta). La traccion a la que
se encuentra sometido el hueso es la causante de un dano tan alto. En lo puntos B y C,
en cambio, el implante esta comprimiendo el hueso en direccion normal a la superficie. La
compresion no provoca dano, con la suposicion hecha aquı, pero provoca alargamientos en
direccion z por efecto Poisson, que hace que aumente el dano en esa direccion.
En la figura 8.16 se muestran los modulos de Young en puntos cercanos al implante, pero
algo mas alejados que los anteriores. Las rigideces son sensiblemente mayores al alejarse del
implante, por la disminucion del dano, pero este aun es considerable en la zona mesial. Ası,
el punto H, como antes el D, es el de menor rigidez de todos los de esta capa.
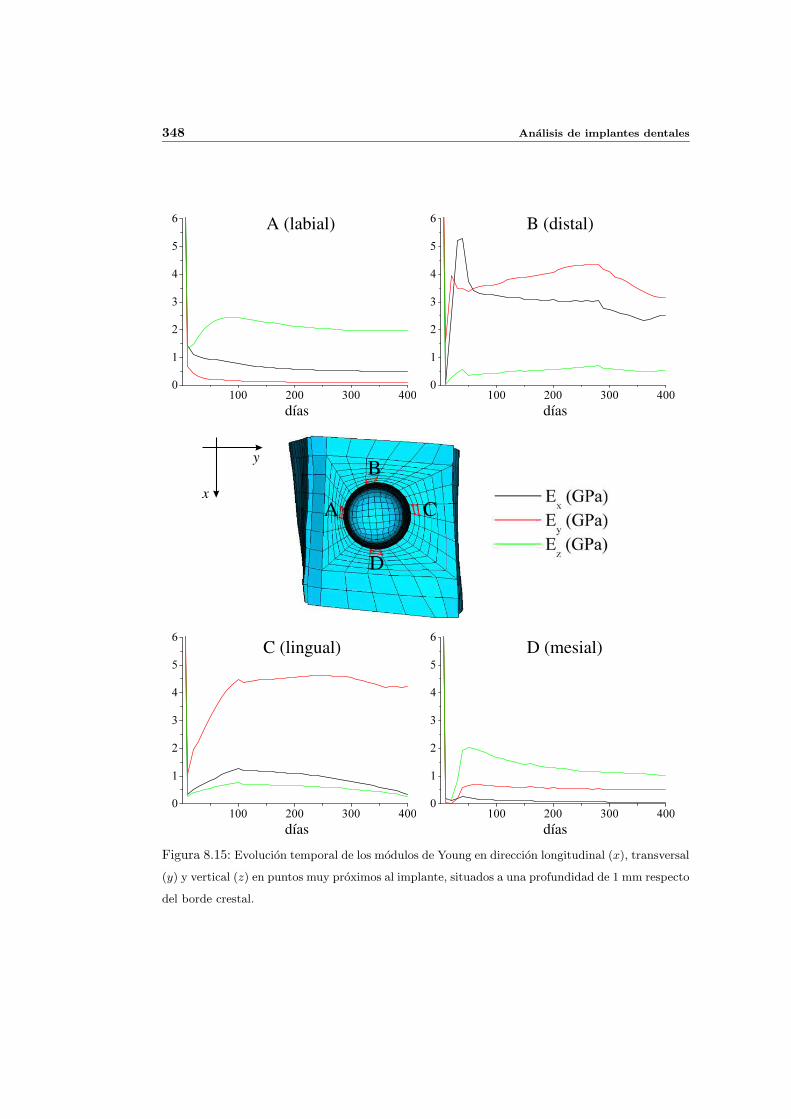
348 Analisis de implantes dentales
A
B
C
D
1
x
y
100 200 300 4000
1
2
3
4
5
6
dıas
100 200 300 4000
1
2
3
4
5
6
dıas
100 200 300 4000
1
2
3
4
5
6
dıas
100 200 300 4000
1
2
3
4
5
6
dıas
A (labial) B (distal)
C (lingual) D (mesial)
1
Figura 8.15: Evolucion temporal de los modulos de Young en direccion longitudinal (x), transversal
(y) y vertical (z) en puntos muy proximos al implante, situados a una profundidad de 1 mm respecto
del borde crestal.

8.2 Implantes completamente osteointegrados 349
En el resto de puntos la rigidez es muy parecida, entre 14 y 17GPa, y el grado de
anisotropıa relativamente pequeno. Se puede decir que no hay direcciones preferentes, como
sucede en la mandıbula sana, donde la direccion de mayor rigidez es la longitudinal, paralela
al eje del cuerpo mandibular. La razon es que las condiciones de carga han cambiado con la
inclusion del implante y la acumulacion de dano, y la consecuente redistribucion de tensiones
afecta a la rigidez del tejido circundante.
El dano tambien influye en el hecho de que el grado de anisotropıa no sea muy elevado.
En efecto, la direccion de carga preferente es la que tiende a hacerse mas rıgida con la
remodelacion, pero como tambien sera la que acumule mas dano, ese incremento de rigidez
se vera mermado. Esto se observa claramente en el punto F. Por la flexion de la mandıbula,
en ese punto son mas altas las tensiones σx y la tendencia deberıa ser aumentar la rigidez
en esa direccion. Pero en su lugar, lo que ocurre es que se acumula muy rapidamente el
dano y por eso cae subitamente Ex al principio del analisis. Cuando ese dano se repara
paulatinamente por remodelacion la rigidez se recupera y crece poco a poco.
Discusion de los resultados
Algunos parametros que se usan para evaluar el exito del implante desde el punto de
vista clınico son la sensibilidad local, la existencia de placa, la movilidad, el sangrado y
la perdida de hueso crestal entre otros [24]. Entre los anteriores quiza los mas importantes
sean la movilidad y la cantidad de hueso reabsorbido, que es precisamente lo que se pretende
simular en este analisis. Para comprobar si existe reabsorcion crestal clınicamente se realiza
un seguimiento radiografico despues de la colocacion del implante. Sobre las radiografıas se
traza una lınea de referencia siguiendo la cresta alveolar (ver figura 8.17). Todo lo que haya
por debajo de la lınea de referencia se considera defecto, que se mide bien por la superficie
que ocupa (figura 8.17 izquierda) o bien su altura (figura de la derecha).
Existen numerosos estudios clınicos publicados sobre la supervivencia de implantes IMZ.
Willer et al. [349], por ejemplo, han analizado 1041 implantes de los cuales 136 fallaron
a largo plazo, 35 fallaron a medio plazo, despues del perıodo de curacion y 11 fallaron
durante el perıodo de curacion. El resto no fallaron, lo que representa una buena proporcion
de exito. Willer et al. se limitan a diferenciar los implantes como exitosos o fallidos (si
es necesario retirarlos), sin distinguir la razon del fracaso. Batenburg et al. [25] sı tienen
en cuenta las razones que pueden llevar al fracaso del implante. En concreto, en los dos
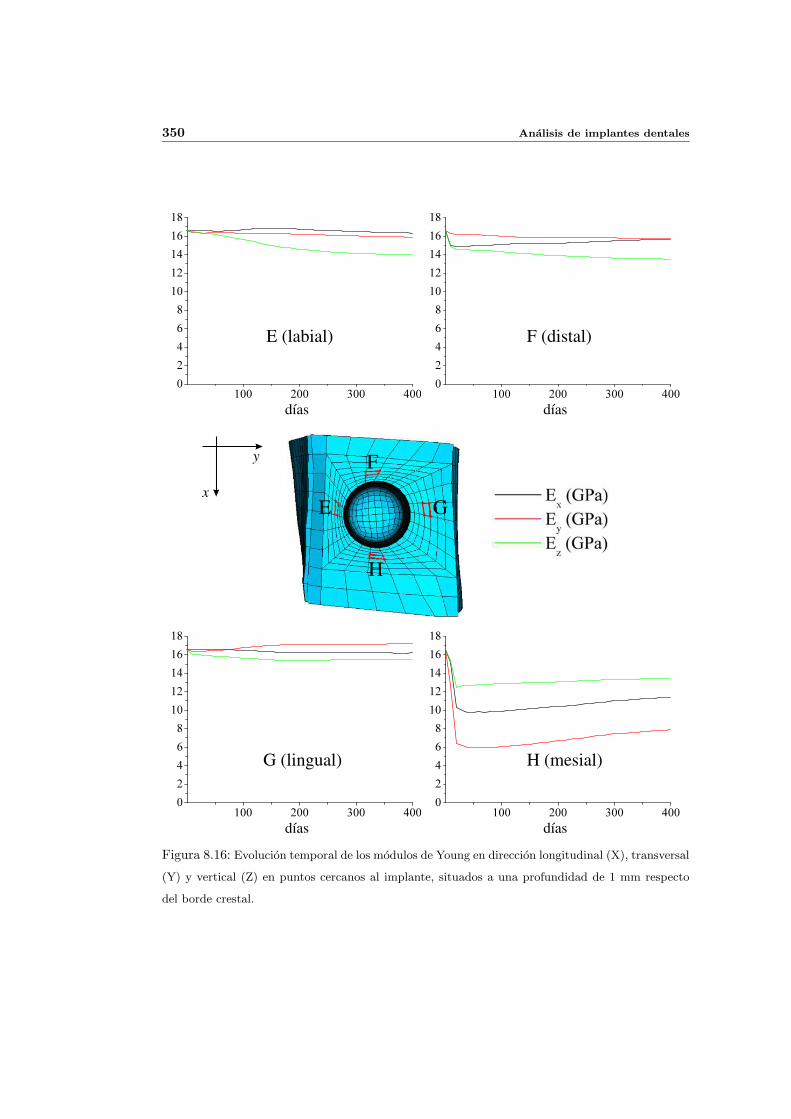
350 Analisis de implantes dentales
E
F
G
H
1
x
y
100 200 300 400024681012141618
dıas
100 200 300 400024681012141618
dıas
100 200 300 400024681012141618
dıas
100 200 300 400024681012141618
dıas
E (labial) F (distal)
G (lingual) H (mesial)
1
Figura 8.16: Evolucion temporal de los modulos de Young en direccion longitudinal (X), transversal
(Y) y vertical (Z) en puntos cercanos al implante, situados a una profundidad de 1 mm respecto
del borde crestal.

8.2 Implantes completamente osteointegrados 351
1
Figura 8.17: Formas de medir la perdida osea crestal: Superficie ocupada por el defecto (Izq.) o
altura del mismo (Dcha.). Tomado de [24].
primeros anos se ha producido reabsorcion crestal en 14 de los 57 implantes analizados por
estos autores, aunque ello no quiere decir que los 14 hayan fallado. Heydenrijk et al. [146]
son mas precisos y distinguen entre reabsorcion leve o severa. Ası, en el primer ano, de
37 implantes IMZ analizados, 11 no exhiben reabsorcion osea, 14 de ellos tienen menos
de 1 mm de reabsorcion y 12 presentan una altura de defecto mayor que 1 mm. Como se
ve la proporcion de exito es relativamente buena por lo general y aunque la reabsorcion
osea crestal puede ser un problema que hay que intentar evitar, no es una situacion que
se de indefectiblemente. En la figura 8.18 se muestra un par de radiografıas de implantes
IMZ. En el A no hay perdida osea crestal y la bajada de la cresta alveolar que se observa
se debe probablemente a la reabsorcion asociada a la perdida del diente natural, que ocurre
con anterioridad a la colocacion del implante. En la radiografıa B, en cambio, sı se puede
observar claramente que existe perdida osea. No solo baja la cresta alveolar por la perdida
del diente, sino que ademas, junto al implante se ha formado un hueco que se extiende hacia
abajo dejando al descubierto parte de la superficie del implante. Desde luego, la forma que
presenta este hueco de reabsorcion es muy parecida a la que tiene en el modelo de elementos
finitos la zona de alta porosidad que se observa en el cuello del implante (ver figura 8.10).
Si se compara la radiografıa 8.18 con los resultados de la simulacion (figura 8.10), se puede
observar cierta similitud, ya que en el cuello se predice una reabsorcion importante y en los
implantes que fallan en la realidad es ahı donde comienza a perderse hueso, como se observa
en la radiografıa. De todos modos, hay que tener en cuenta que la simulacion tampoco
predice una reabsorcion demasiado severa (la porosidad final en esa zona no es demasiado

352 Analisis de implantes dentales
1
Figura 8.18: Radiografıa de sendos implantes IMZ: (A) sin perdida osea crestal y (B) con perdida
osea. Tomado de [25].
alta), algo que esta de acuerdo con los estudios clınicos, que no establecen que haya un fallo
generalizado de los implantes IMZ por reabsorcion crestal. Eso sı, de existir la reabsorcion,
suele aparecer en una zona muy proxima a la predicha por la simulacion. En cualquier
caso, para predecir la reabsorcion crestal de manera precisa hubiera sido necesario simular
el proceso de remodelacion osea externa, que no se hace en esta tesis. Con la prediccion de
un aumento de la porosidad solo se esta comprobando que el hueso de esa zona tiende a
desaparecer, aumentando la probabilidad de que haya remodelacion osea externa.
El hecho de que la reabsorcion simulada sea mas notable en uno de los lados (el mesial
en este caso) se debe a la posicion del implante dentro de la mandıbula (ocupa el lugar
del primer molar). En el lado distal del implante la flexion es menor que en el lado mesial,
de ahı que el dano acumulado y la reabsorcion tambien sea menor. Si el implante estuviera
localizado en la region premolar o incisiva las tensiones serıan mas uniformes y la reabsorcion
mas parecida a uno y otro lado.
Uno de los resultados mas llamativos de la simulacion es, que aunque se suponga una
osteointegracion inicial perfecta, alrededor del implante acaba quedando una capa de hueso
danado y algo mas poroso, por lo tanto mas flexible. En este sentido, resultan clarificadores
algunos estudios clınicos e histologicos sobre la osteointegracion de implantes. Albrektsson
et al. [8] aseguran que incluso en implantes osteointegrados adecuadamente existe una capa
de tejido blando, muy delgada eso sı (1µm), adyacente a la superficie del implante. Cuando
la osteointegracion no es tan buena puede existir cierta movilidad y laxitud de la fijacion,
siendo la capa de tejido blando mas gruesa en este caso, entre 200 y 500µm [43,44,52,95]. La

8.2 Implantes completamente osteointegrados 353
capa de tejido danado y poroso que resulta en el modelo podrıa estar simulando este tejido
blando adyacente al implante. El espesor de esa capa es variable a lo largo de la interfase
y oscila entre 20 y 200µm, sin contar la zona del cuello del implante, donde el dano y la
porosidad del hueso es sensiblemente mayor y el espesor de la zona danada tambien. Como
se ve, los espesores son muy similares a los medidos en estudios clınicos. Hay que matizar
este resultado puesto que esta capa de tejido blando no es continua, ni rodea por completo
al implante, ya que, como se ve en las figuras 8.3 y 8.10, no toda la interfase esta danada,
ni tiene una porosidad elevada. Existen algunas zonas donde el nivel de dano es moderado
y apenas se ha producido reabsorcion, lo que quiere decir que en ellas la osteointegracion se
mantiene en buenas condiciones. Existe como se ve una gran variabilidad de los resultados
a lo largo de la interfase. Numerosos estudios histomorfometricos [64,132,163,177,297,337]
han confirmado que efectivamente el grado de osteointegracion es muy variable a lo largo de
la interfase y que el porcentaje de hueso en contacto directo con el implante, que se suele
denominar BIC (Bone Implant Contact), depende de muchos factores, pero rara vez supera
el 70 %.
Un aspecto relacionado con el grado de osteointegracion y la presencia de la capa de
tejido flexible mencionada antes, es la movilidad que pueda causar al implante durante la
masticacion. Esta movilidad se ha analizado en el caso de carga mas comprometido, que
es la masticacion con el implante. En este caso, la movilidad vertical es bastante pequena,
del orden de 30µm en el lado distal, que se puede considerar despreciable si se compara
con los valores lımites de seguridad que establece Misch [238] para la movilidad (1mm para
la horizontal). Hay que tener en cuenta que la movilidad horizontal es admisible, pero la
vertical, que es la que se ha medido aquı, no lo es. Sin embargo, tambien es cierto que la
movilidad se ha medido en unas condiciones mas desfavorables (en carga), que aquellas en
las que se mide clınicamente (con un pequeno impacto, mediante el Periotest r©). Se puede
concluir entonces que los resultados de movilidad del implante estan dentro de los valores
admisibles, como el resto de parametros que determinan el exito del implante.
8.2.2. Secuencia S4. Densidad inicial 1.7g/cm3 en el hueso periim-
plantario
La densidad inicial supuesta para el hueso periimplantario en al apartado anterior,
2.0 g/cm3, quiza sea un valor demasiado elevado para el hueso que ocupa el hueco deja-

354 Analisis de implantes dentales
do por el diente natural. Tampoco se puede saber con exactitud la distribucion de densidad
y por eso se han probado otros valores de la densidad inicial. En este apartado se analiza
un valor ligeramente inferior: 1.7 g/cm3.
Evolucion del dano
En la figura 8.19 se muestran tres componentes del tensor de dano tras 10 y 400 dıas de
simulacion. Se puede observar que ahora el dano sı aumenta sensiblemente con el tiempo, a
diferencia de lo que ocurrıa en el caso anterior. Para ρ0 = 2.0 g/cm3, el dano se acumulaba
especialmente durante los primeros dıas, pero luego se mantenıa relativamente estable. Ahora
sı crece y alcanza valores mas altos que los obtenidos con ρ0 = 2.0 g/cm3. Los resultados de
esta simulacion y la anterior se comparan en la figura 8.20, donde se observa una sensible
diferencia entre ambas situaciones.
El dano sigue siendo mas alto en direccion x debido a la flexion de la mandıbula, que
esta sometida a un momento flector en direccion y. Es especialmente alto en la zona apical del
lado distal (derecha) y dentro de la zona crestal es mas alto en el lado mesial. Precisamente,
en esas dos zonas, el hueso esta sometido a traccion, como consecuencia de la rigidez tan
diferente que tienen hueso e implante y de que la osteointegracion se mantiene ıntegra, por
hipotesis.
Que el dano sea mayor en el caso de ρ0 = 1.7 g/cm3 es logico, dado que las deformaciones
son mayores, por ser el hueso periimplantario mas flexible. Ademas, la singularidad de ten-
siones que existe en el borde libre de la interfase hueso-implante es mas severa, por ser mayor
el contraste de rigidez entre uno y otro material y eso contribuye a elevar las tensiones y
las deformaciones, que son finalmente las que determinan el crecimiento del dano, con la ley
de dano supuesta. En la figura 8.21 se comprueba esto en la fraccion volumetrica de tejido
danado promedio, que en el caso de ρ0 = 1.7 g/cm3 es casi tres veces superior a la del caso
ρ0 = 2.0 g/cm3.
Evolucion de la fraccion volumetrica de hueso
La fraccion volumetrica de hueso inicial es vb0 = 0.778 para la densidad ρ0 = 1.7 g/cm3
y la fraccion de ceniza inicial supuesta α0 = 0.6. Para comprobar la convergencia, se rep-
resenta en la figura 8.22 la evolucion temporal de la fraccion volumetrica promedio, vb, en
el hueso periimplantario. Efectivamente, tras 400 dıas de simulacion se alcanza un equilib-
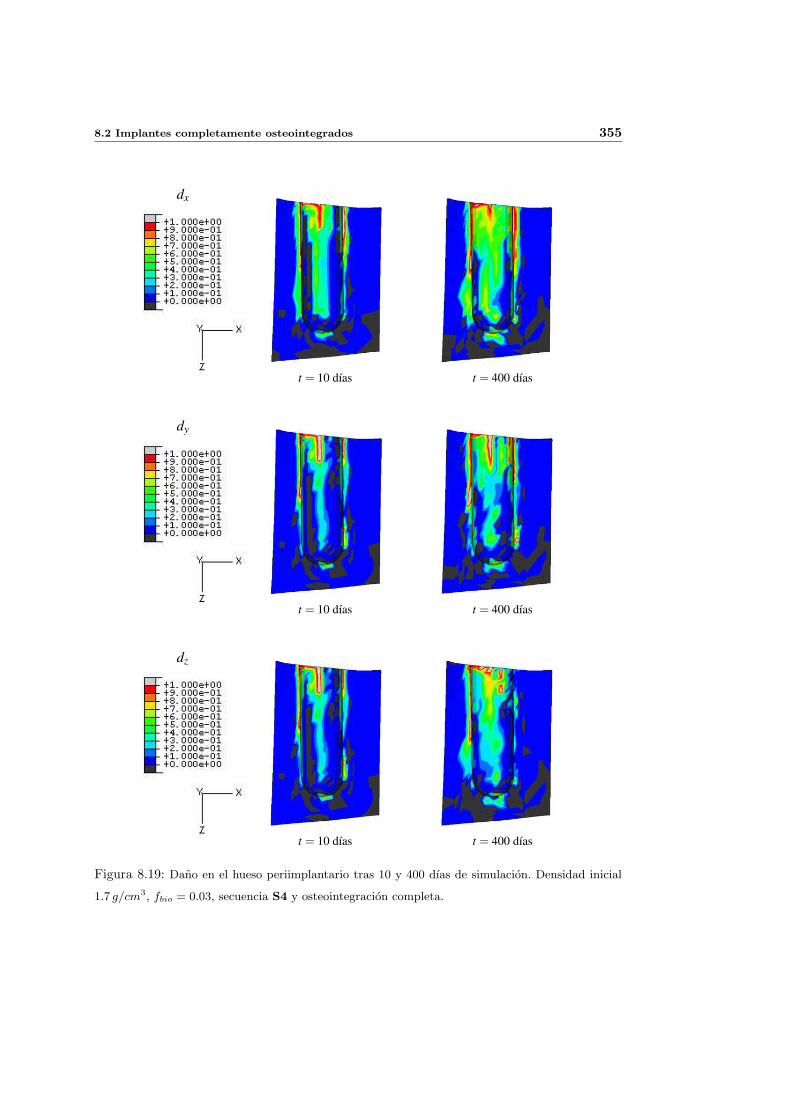
8.2 Implantes completamente osteointegrados 355
dx
t = 10 dıas t = 400 dıas
dy
t = 10 dıas t = 400 dıas
dz
t = 10 dıas t = 400 dıas
1
Figura 8.19: Dano en el hueso periimplantario tras 10 y 400 dıas de simulacion. Densidad inicial
1.7 g/cm3, fbio = 0.03, secuencia S4 y osteointegracion completa.
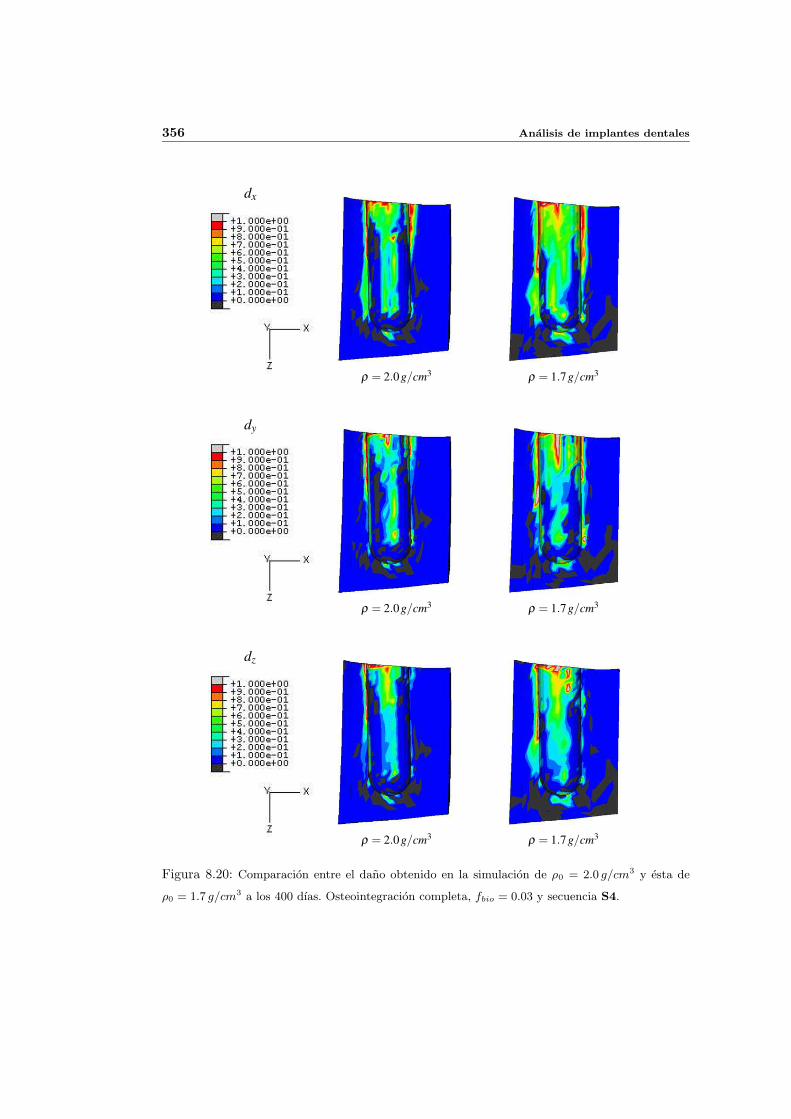
356 Analisis de implantes dentales
dx
ρ = 2.0g/cm3 ρ = 1.7g/cm3
dy
ρ = 2.0g/cm3 ρ = 1.7g/cm3
dz
ρ = 2.0g/cm3 ρ = 1.7g/cm3
1
Figura 8.20: Comparacion entre el dano obtenido en la simulacion de ρ0 = 2.0 g/cm3 y esta de
ρ0 = 1.7 g/cm3 a los 400 dıas. Osteointegracion completa, fbio = 0.03 y secuencia S4.
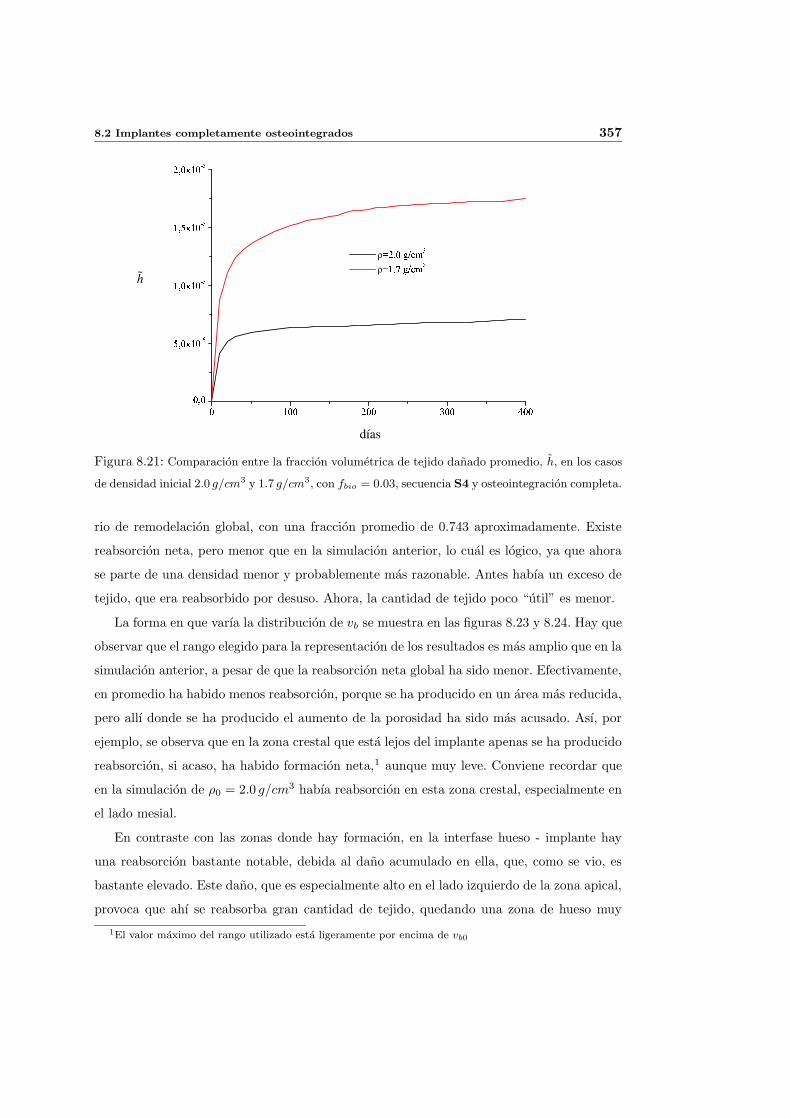
8.2 Implantes completamente osteointegrados 357
æ çææ èææ éææ êæææë æ
ì ë æí.çæî ï
ç4ë æí.çæî ð
ç4ë ì í.çæî ð
èë æí.çæ î ð
ρñ èSò æôó!õ ö÷ùøρñ ç!ò úôó!õ ö÷ ø
dıas
h
1
Figura 8.21: Comparacion entre la fraccion volumetrica de tejido danado promedio, h, en los casos
de densidad inicial 2.0 g/cm3 y 1.7 g/cm3, con fbio = 0.03, secuencia S4 y osteointegracion completa.
rio de remodelacion global, con una fraccion promedio de 0.743 aproximadamente. Existe
reabsorcion neta, pero menor que en la simulacion anterior, lo cual es logico, ya que ahora
se parte de una densidad menor y probablemente mas razonable. Antes habıa un exceso de
tejido, que era reabsorbido por desuso. Ahora, la cantidad de tejido poco “util” es menor.
La forma en que varıa la distribucion de vb se muestra en las figuras 8.23 y 8.24. Hay que
observar que el rango elegido para la representacion de los resultados es mas amplio que en la
simulacion anterior, a pesar de que la reabsorcion neta global ha sido menor. Efectivamente,
en promedio ha habido menos reabsorcion, porque se ha producido en un area mas reducida,
pero allı donde se ha producido el aumento de la porosidad ha sido mas acusado. Ası, por
ejemplo, se observa que en la zona crestal que esta lejos del implante apenas se ha producido
reabsorcion, si acaso, ha habido formacion neta,1 aunque muy leve. Conviene recordar que
en la simulacion de ρ0 = 2.0 g/cm3 habıa reabsorcion en esta zona crestal, especialmente en
el lado mesial.
En contraste con las zonas donde hay formacion, en la interfase hueso - implante hay
una reabsorcion bastante notable, debida al dano acumulado en ella, que, como se vio, es
bastante elevado. Este dano, que es especialmente alto en el lado izquierdo de la zona apical,
provoca que ahı se reabsorba gran cantidad de tejido, quedando una zona de hueso muy
1El valor maximo del rango utilizado esta ligeramente por encima de vb0
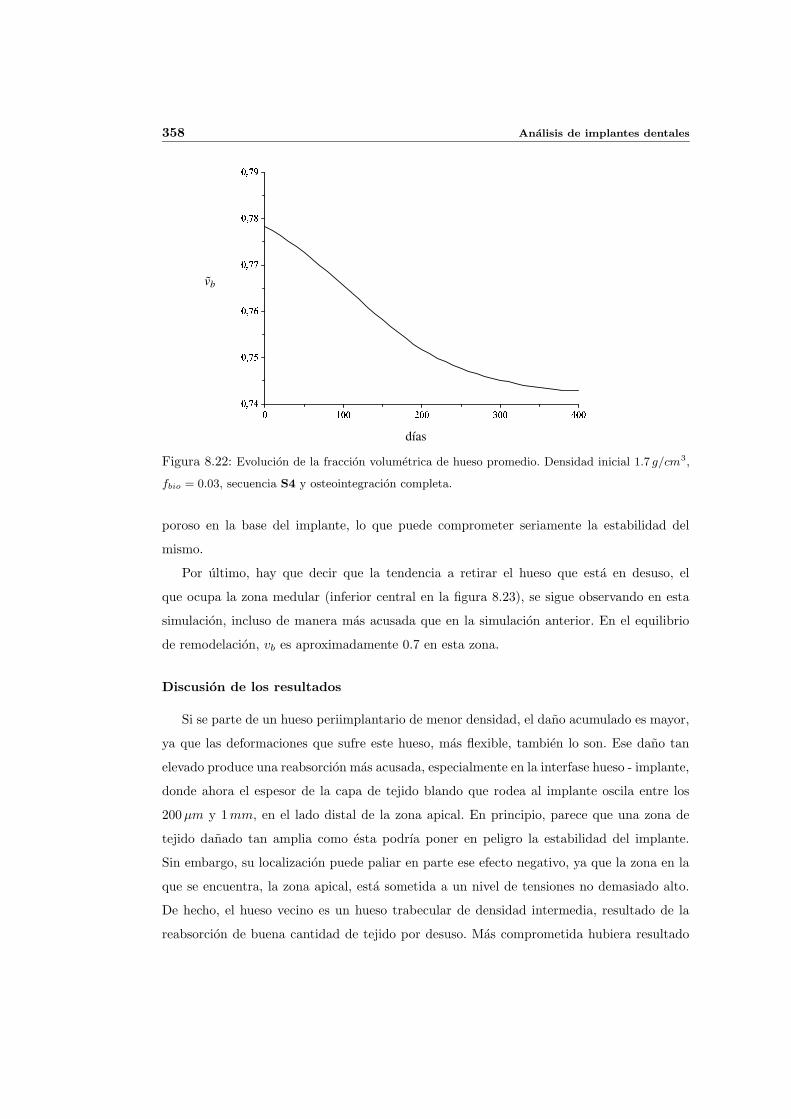
358 Analisis de implantes dentales
û üûû ýûû þûû ÿûûû ÿ
û
û
û
û
û
dıas
vb
1
Figura 8.22: Evolucion de la fraccion volumetrica de hueso promedio. Densidad inicial 1.7 g/cm3,
fbio = 0.03, secuencia S4 y osteointegracion completa.
poroso en la base del implante, lo que puede comprometer seriamente la estabilidad del
mismo.
Por ultimo, hay que decir que la tendencia a retirar el hueso que esta en desuso, el
que ocupa la zona medular (inferior central en la figura 8.23), se sigue observando en esta
simulacion, incluso de manera mas acusada que en la simulacion anterior. En el equilibrio
de remodelacion, vb es aproximadamente 0.7 en esta zona.
Discusion de los resultados
Si se parte de un hueso periimplantario de menor densidad, el dano acumulado es mayor,
ya que las deformaciones que sufre este hueso, mas flexible, tambien lo son. Ese dano tan
elevado produce una reabsorcion mas acusada, especialmente en la interfase hueso - implante,
donde ahora el espesor de la capa de tejido blando que rodea al implante oscila entre los
200µm y 1mm, en el lado distal de la zona apical. En principio, parece que una zona de
tejido danado tan amplia como esta podrıa poner en peligro la estabilidad del implante.
Sin embargo, su localizacion puede paliar en parte ese efecto negativo, ya que la zona en la
que se encuentra, la zona apical, esta sometida a un nivel de tensiones no demasiado alto.
De hecho, el hueso vecino es un hueso trabecular de densidad intermedia, resultado de la
reabsorcion de buena cantidad de tejido por desuso. Mas comprometida hubiera resultado
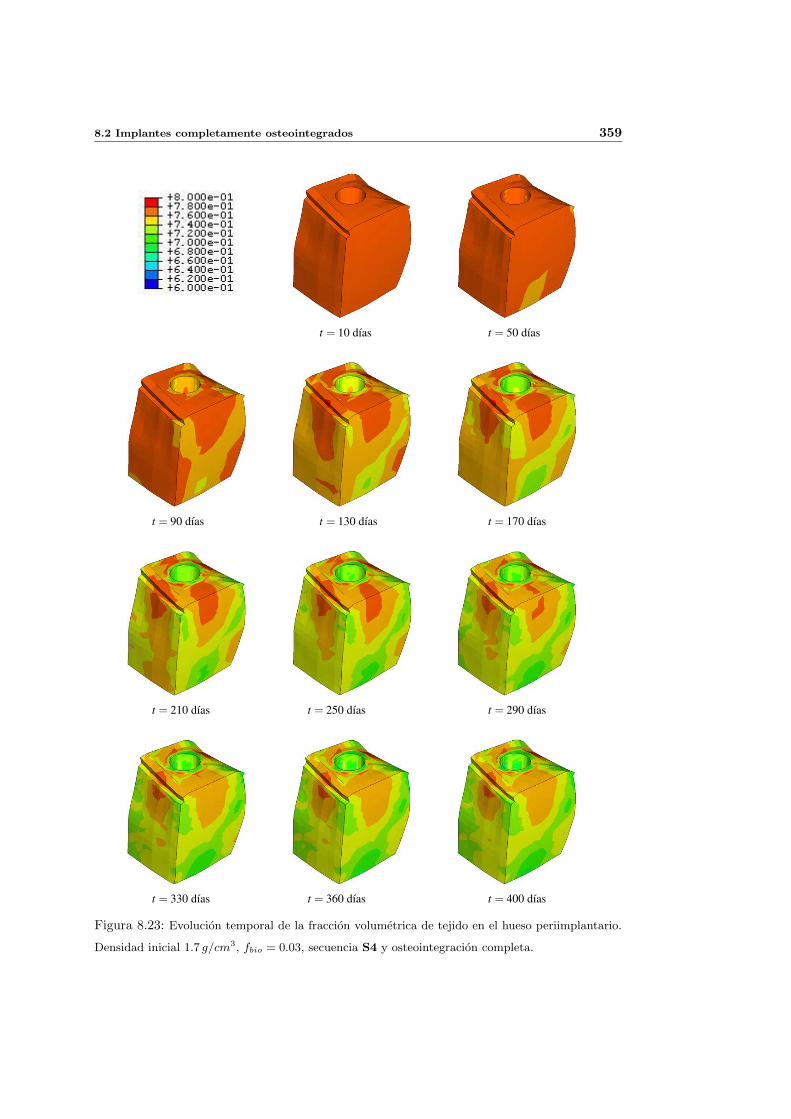
8.2 Implantes completamente osteointegrados 359
t = 10 dıas t = 50 dıas
t = 90 dıas t = 130 dıas t = 170 dıas
t = 210 dıas t = 250 dıas t = 290 dıas
t = 330 dıas t = 360 dıas t = 400 dıas
1
Figura 8.23: Evolucion temporal de la fraccion volumetrica de tejido en el hueso periimplantario.
Densidad inicial 1.7 g/cm3, fbio = 0.03, secuencia S4 y osteointegracion completa.
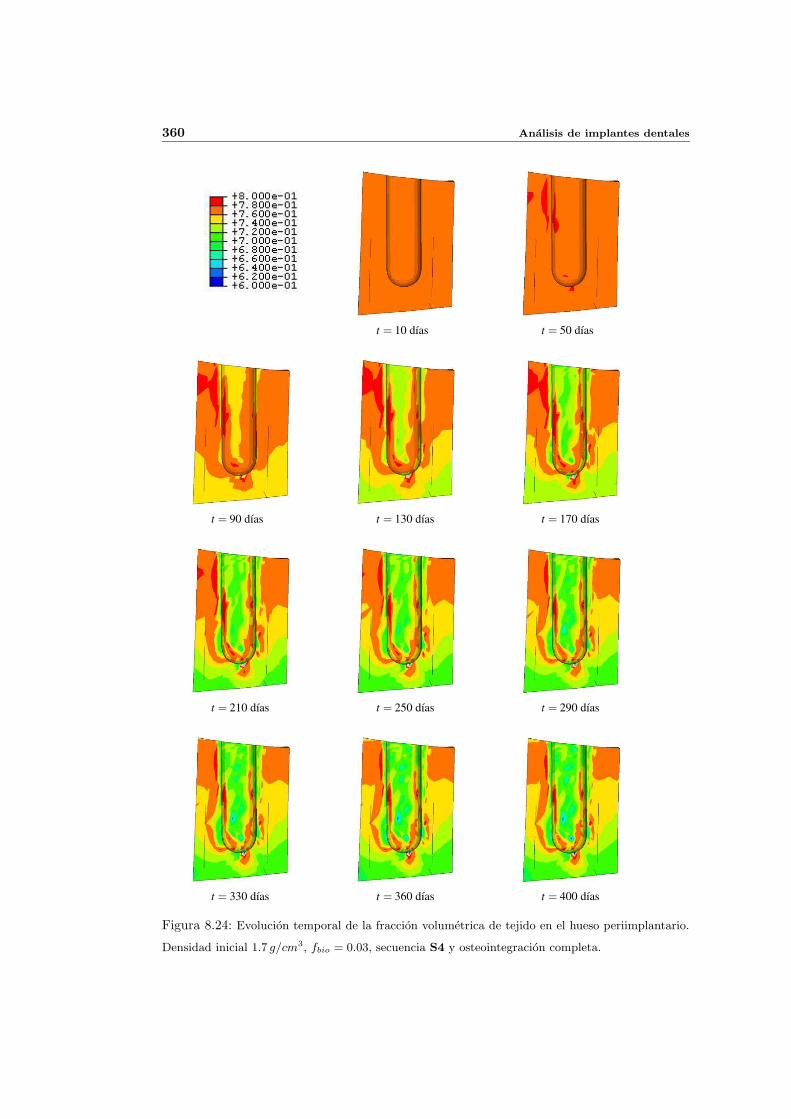
360 Analisis de implantes dentales
t = 10 dıas t = 50 dıas
t = 90 dıas t = 130 dıas t = 170 dıas
t = 210 dıas t = 250 dıas t = 290 dıas
t = 330 dıas t = 360 dıas t = 400 dıas
1
Figura 8.24: Evolucion temporal de la fraccion volumetrica de tejido en el hueso periimplantario.
Densidad inicial 1.7 g/cm3, fbio = 0.03, secuencia S4 y osteointegracion completa.

8.2 Implantes completamente osteointegrados 361
la situacion de haberse obtenido un dano muy elevado en la zona crestal, en la que sı es
necesario un soporte oseo firme para soportar las cargas de masticacion. En esta zona, el
dano y la reabsorcion son mayores que en el caso ρ0 = 2.0 g/cm3, pero las diferencias no
son dramaticas y se puede concluir que el riesgo de perdida del implante por reabsorcion es
mayor si el hueso de partida es de peor calidad, pero tampoco es excesivo.
La reabsorcion neta es menor en este caso de ρ0 = 1.7 g/cm3, segun se deduce de la
evolucion de vb, ya que se produce reabsorcion en una region mas reducida y en algunas
zonas se produce incluso formacion. Ası, por ejemplo, en la zona crestal disminuye vb, si se
parte de ρ0 = 2.0 g/cm3, y aumenta si se comienza con ρ0 = 1.7 g/cm3. Es decir, el modelo
tiende a alcanzar un valor intermedio de vb en esa zona, independientemente del valor inicial
de vb supuesto.
Ası pues, la reabsorcion neta es menor debido a que el valor ρ0 = 1.7 g/cm3 debe estar
mas cerca de la densidad promedio del hueso periimplantario que el valor ρ0 = 2.0 g/cm3 de
la simulacion anterior. Pero tambien hay que decir, que allı donde se produce reabsorcion,
esta es mas acusada. La razon para esto no hay que buscarla solo en el hecho de que el dano
sea mayor, que lo es, sino tambien en el hecho de que el numero de BMUs activadas por
unidad de tiempo es mayor en un hueso de densidad intermedia que un hueso muy compacto,
que dispone de poca superficie especıfica de poros, Sv (ver figura 5.11), para la activacion
de BMUs. La actividad remodelatoria en este hueso de densidad intermedia es mas intensa
y de ahı que la diferencia de porosidad entre unas zonas y otras sea mas grande.
8.2.3. Secuencia S5. Densidad inicial 2.0g/cm3 en el hueso periim-
plantario
A continuacion se analiza lo que ocurre si se alteran los habitos masticatorios y no se
mastica con el lado del implante, ya sea por prescripcion medica, por falta de confianza del
paciente o por cualquier otra razon. Como ya se indico, no se pretende simular el perıodo
de curacion tras la implantacion, en el que es habitual no masticar con el lado del implante,
ya que el proceso mecanobiologico de osteointegracion, que tiene lugar durante este perıodo,
no ha sido simulado en esta tesis. Tan solo se pretende simular que ocurrirıa si ya se ha
alcanzado la osteointegracion pero aun no se mastica con ese lado y si es conveniente o no
un perıodo de curacion excesivamente largo, antes de colocar la protesis y que sea funcional.
La densidad inicial supuesta para el hueso periimplantario es ρ0 = 2.0 g/cm3 y se aplica

362 Analisis de implantes dentales
una secuencia de masticacion unilateral con el lado izquierdo S5: (MI1-MI2)-(MI1-MI2)-
(MI1-MI2)-. . . con 500 ciclos diarios, 250 para cada carga, MI1 y MI2.
Al principio de este capıtulo, cuando se analizo la distribucion de tensiones que producıan
los distintos casos de carga sobre el hueso periimplantario se vio que las masticaciones con
el lado izquierdo producıan unas tensiones mucho mas reducidas en el entorno del implante
que las masticaciones con el lado derecho. Este hecho va a condicionar todos los resultados
obtenidos con esta simulacion ya que el hecho de que el hueso este sometido a un estımulo
menor hara que el dano acumulado sea menor, y la reabsorcion por dano tambien, pero por
contra inducira la reabsorcion por desuso.
Evolucion del dano
En la figura 8.25 se muestra la distribucion de tres componentes del tensor de dano a
los 30 y a los 500 dıas de simulacion. Se ha mostrado el dano a los 30 dıas en lugar de
a los 10 dıas, como se venıa haciendo hasta ahora, porque el dano a los 10 dıas es aun
muy pequeno. Y es que el dano en esta simulacion no solo es menor, sino que ademas se
propaga de manera mas gradual. De hecho, a los 30 dıas de simulacion, la zona danada aun
es pequena en comparacion con la extension que alcanza al final de la simulacion, cuando
tampoco es muy extensa, de todos modos.
Es interesante analizar el nivel de dano en los elementos de la interfase hueso - implante,
dado que en el perıodo de curacion, en el que se consigue la osteointegracion, no se suele
masticar con este lado de la mandıbula, tal como se hace en esta simulacion. De esta forma
se puede comprobar si el hecho de no masticar con este lado puede ayudar a una mejor con-
solidacion del hueso que rodea al implante. Los resultados indican que el dano que acumula
la interfase es sensiblemente menor que en la simulacion de la secuencia S4, aunque algo de
dano existe. No se produce en toda la superficie de la interfase y esta muy localizado, como
en todos los casos anteriores, en el cuello del implante, donde es similar al de la secuencia
S4. Aparte de la zona crestal, el resto de la interfase tiene un nivel de dano normal y el
espesor de la capa de tejido danado que rodea al implante tiene en este caso un espesor
mucho mas reducido, de unas decenas de micras.
En la figura 8.26 se compara la fraccion volumetrica de tejido danado promedio con las
secuencias S4 y S5, comprobandose que con esta ultima el dano es ligeramente menor.
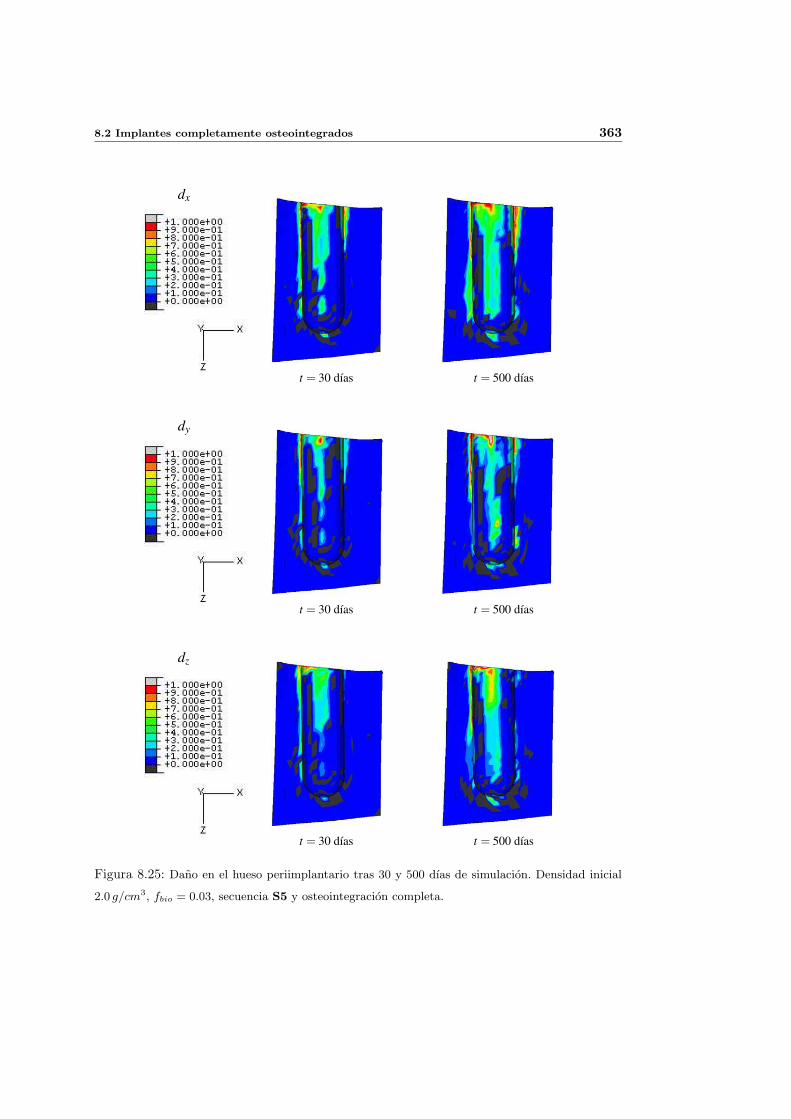
8.2 Implantes completamente osteointegrados 363
dx
t = 30 dıas t = 500 dıas
dy
t = 30 dıas t = 500 dıas
dz
t = 30 dıas t = 500 dıas
1
Figura 8.25: Dano en el hueso periimplantario tras 30 y 500 dıas de simulacion. Densidad inicial
2.0 g/cm3, fbio = 0.03, secuencia S5 y osteointegracion completa.
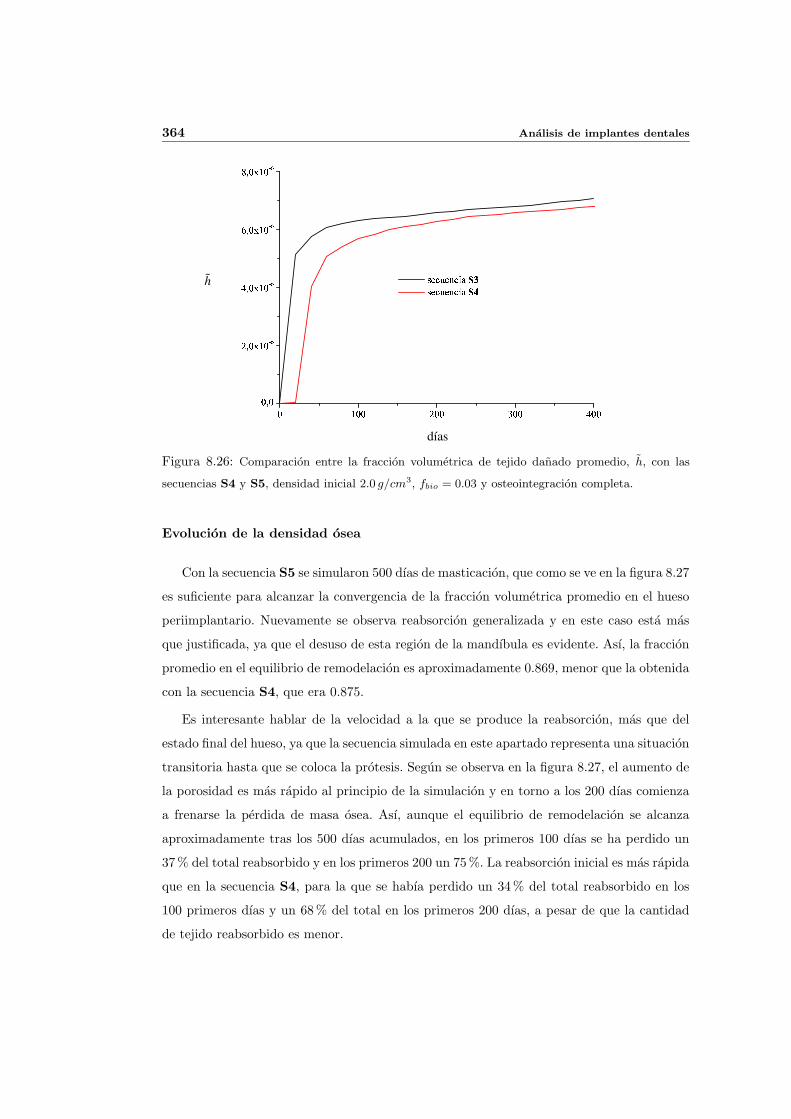
364 Analisis de implantes dentales
"!# "!$
dıas
h
1
Figura 8.26: Comparacion entre la fraccion volumetrica de tejido danado promedio, h, con las
secuencias S4 y S5, densidad inicial 2.0 g/cm3, fbio = 0.03 y osteointegracion completa.
Evolucion de la densidad osea
Con la secuencia S5 se simularon 500 dıas de masticacion, que como se ve en la figura 8.27
es suficiente para alcanzar la convergencia de la fraccion volumetrica promedio en el hueso
periimplantario. Nuevamente se observa reabsorcion generalizada y en este caso esta mas
que justificada, ya que el desuso de esta region de la mandıbula es evidente. Ası, la fraccion
promedio en el equilibrio de remodelacion es aproximadamente 0.869, menor que la obtenida
con la secuencia S4, que era 0.875.
Es interesante hablar de la velocidad a la que se produce la reabsorcion, mas que del
estado final del hueso, ya que la secuencia simulada en este apartado representa una situacion
transitoria hasta que se coloca la protesis. Segun se observa en la figura 8.27, el aumento de
la porosidad es mas rapido al principio de la simulacion y en torno a los 200 dıas comienza
a frenarse la perdida de masa osea. Ası, aunque el equilibrio de remodelacion se alcanza
aproximadamente tras los 500 dıas acumulados, en los primeros 100 dıas se ha perdido un
37 % del total reabsorbido y en los primeros 200 un 75 %. La reabsorcion inicial es mas rapida
que en la secuencia S4, para la que se habıa perdido un 34 % del total reabsorbido en los
100 primeros dıas y un 68 % del total en los primeros 200 dıas, a pesar de que la cantidad
de tejido reabsorbido es menor.
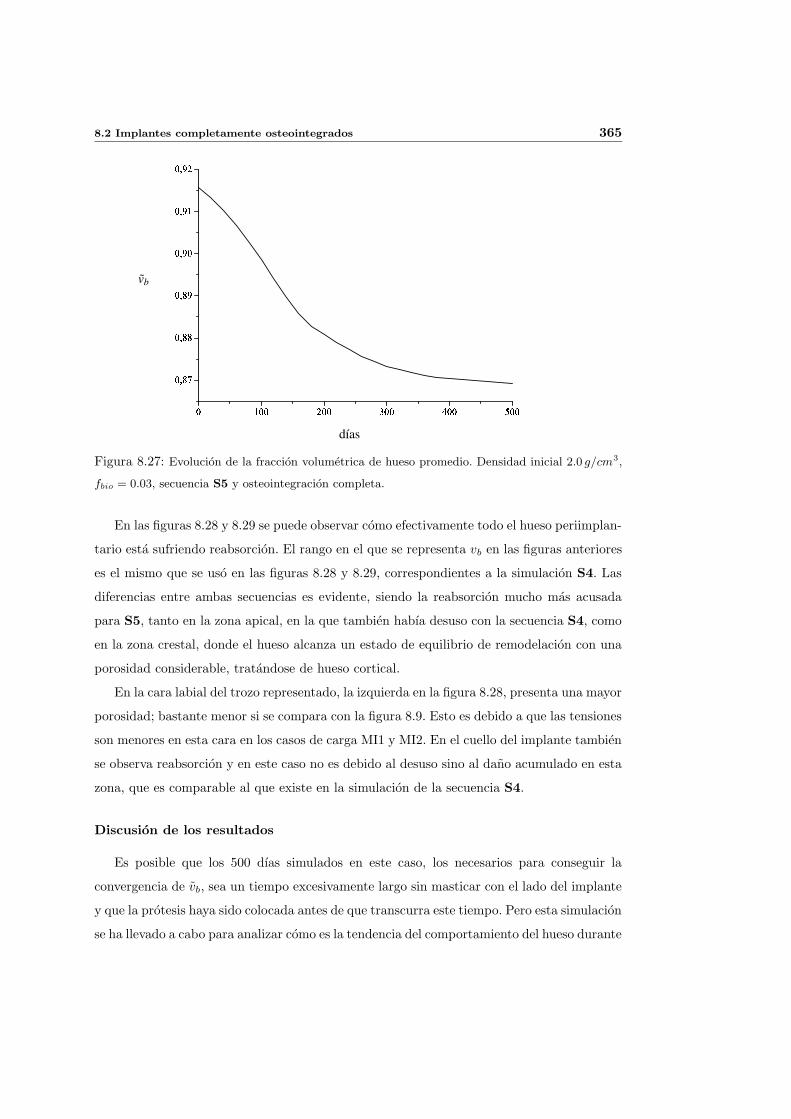
8.2 Implantes completamente osteointegrados 365
% &%% '%% (%% )%% *%%
%+ ,-
%+ ,,
%+ ,.
%+ .%
%+ ./&
%+ .'
dıas
vb
1
Figura 8.27: Evolucion de la fraccion volumetrica de hueso promedio. Densidad inicial 2.0 g/cm3,
fbio = 0.03, secuencia S5 y osteointegracion completa.
En las figuras 8.28 y 8.29 se puede observar como efectivamente todo el hueso periimplan-
tario esta sufriendo reabsorcion. El rango en el que se representa vb en las figuras anteriores
es el mismo que se uso en las figuras 8.28 y 8.29, correspondientes a la simulacion S4. Las
diferencias entre ambas secuencias es evidente, siendo la reabsorcion mucho mas acusada
para S5, tanto en la zona apical, en la que tambien habıa desuso con la secuencia S4, como
en la zona crestal, donde el hueso alcanza un estado de equilibrio de remodelacion con una
porosidad considerable, tratandose de hueso cortical.
En la cara labial del trozo representado, la izquierda en la figura 8.28, presenta una mayor
porosidad; bastante menor si se compara con la figura 8.9. Esto es debido a que las tensiones
son menores en esta cara en los casos de carga MI1 y MI2. En el cuello del implante tambien
se observa reabsorcion y en este caso no es debido al desuso sino al dano acumulado en esta
zona, que es comparable al que existe en la simulacion de la secuencia S4.
Discusion de los resultados
Es posible que los 500 dıas simulados en este caso, los necesarios para conseguir la
convergencia de vb, sea un tiempo excesivamente largo sin masticar con el lado del implante
y que la protesis haya sido colocada antes de que transcurra este tiempo. Pero esta simulacion
se ha llevado a cabo para analizar como es la tendencia del comportamiento del hueso durante
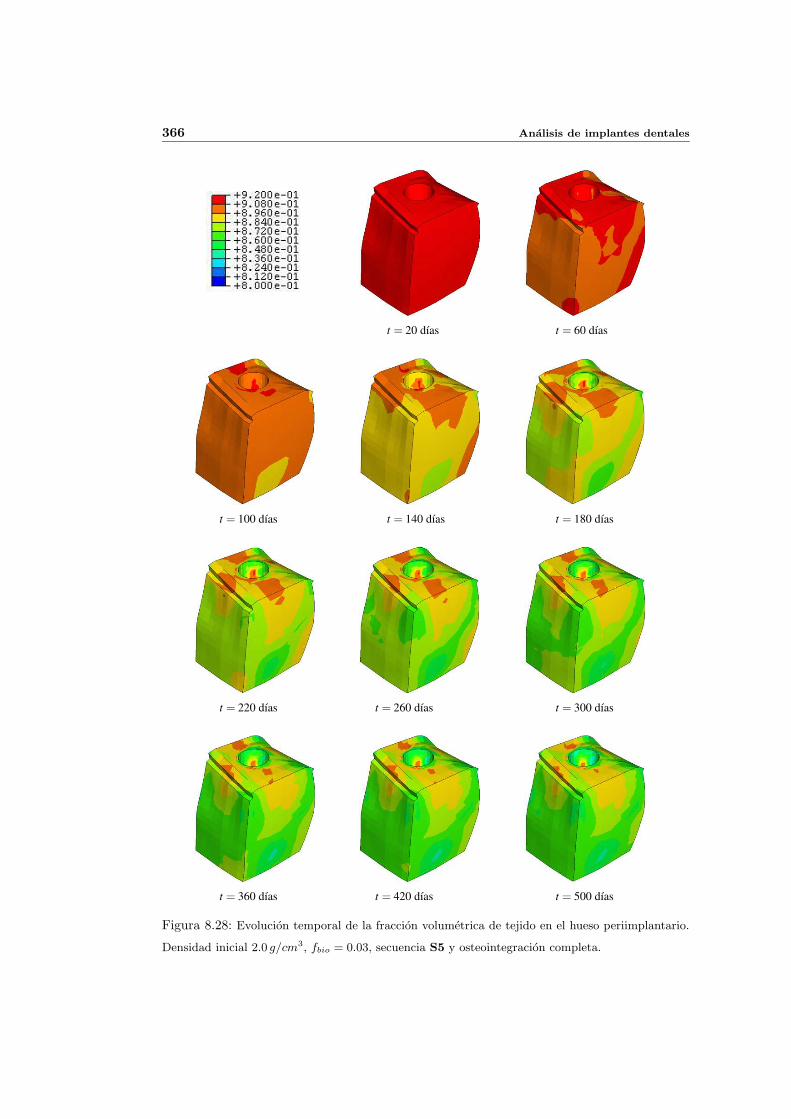
366 Analisis de implantes dentales
t = 20 dıas t = 60 dıas
t = 100 dıas t = 140 dıas t = 180 dıas
t = 220 dıas t = 260 dıas t = 300 dıas
t = 360 dıas t = 420 dıas t = 500 dıas
1
Figura 8.28: Evolucion temporal de la fraccion volumetrica de tejido en el hueso periimplantario.
Densidad inicial 2.0 g/cm3, fbio = 0.03, secuencia S5 y osteointegracion completa.
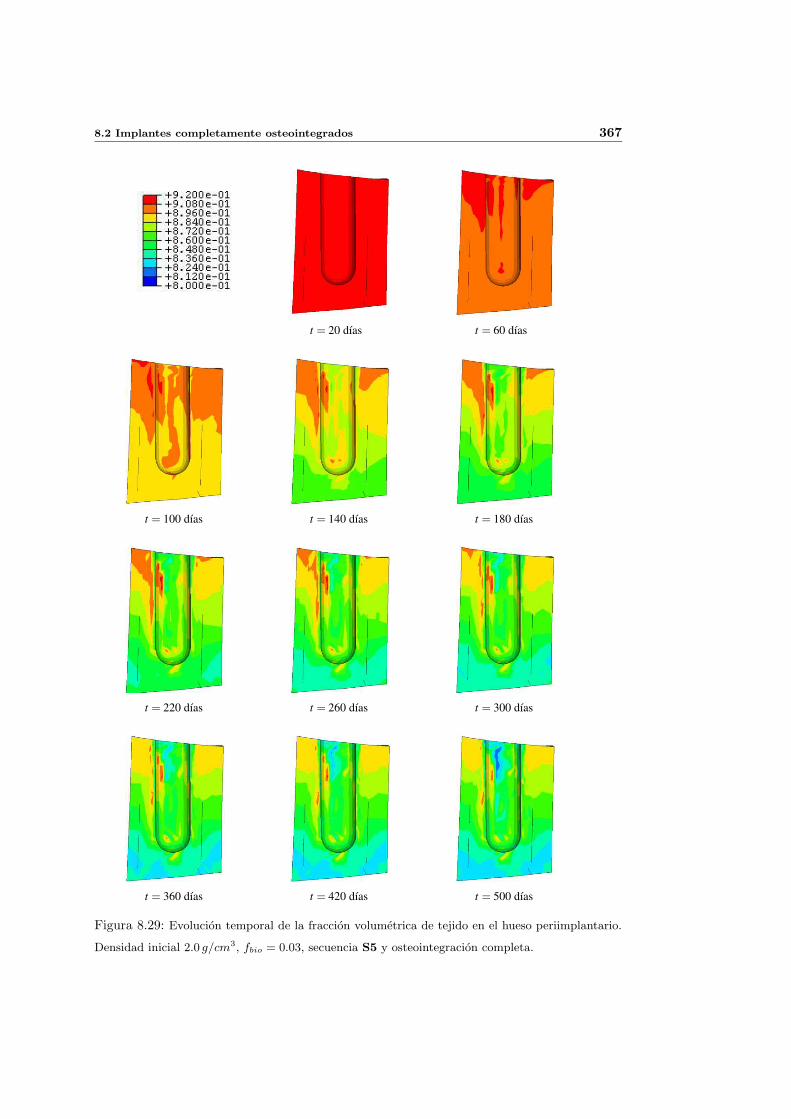
8.2 Implantes completamente osteointegrados 367
t = 20 dıas t = 60 dıas
t = 100 dıas t = 140 dıas t = 180 dıas
t = 220 dıas t = 260 dıas t = 300 dıas
t = 360 dıas t = 420 dıas t = 500 dıas
1
Figura 8.29: Evolucion temporal de la fraccion volumetrica de tejido en el hueso periimplantario.
Densidad inicial 2.0 g/cm3, fbio = 0.03, secuencia S5 y osteointegracion completa.

368 Analisis de implantes dentales
este perıodo, mas que para determinar como queda a largo plazo, ya que hablar de largo
plazo en estas condiciones no tiene mucho sentido.
Pues bien, si no se mastica con el lado del implante se produce una reabsorcion gen-
eralizada del hueso periimplantario provocada por el desuso. Ademas, esta reabsorcion es
especialmente rapida al principio de la simulacion, con lo cual no conviene alargar demasiado
el perıodo de curacion antes de colocar la protesis, ya que se corre el riesgo de perder masa
osea por desuso. Es mas, esta inactividad tampoco asegura que no se acumule dano en la
interfase hueso - implante. En efecto, el dano que se acumula en este caso es inevitable, ya
que aunque se mastique con el lado contrario a donde se situa el implante, la mandıbula
esta sometida a flexion y esas deformaciones, aumentadas por la singularidad en el borde
libre, hacen que la zona danada en torno al cuello del implante sea del mismo tamano aprox-
imadamente que con la secuencia S4, que incluye masticaciones con el lado del implante. En
el resto de la interfase sı es notoria la diferencia con S4 y la zona danada resultante tiene
menor espesor.
Si las tensiones y deformaciones son menores en las cargas MI1 y MI2, resulta extrano
que en el cuello del implante el nivel de dano sea similar al que habıa con la secuencia S4,
con deformaciones mucho mayores. La razon de esto es que la porosidad es mayor con la
secuencia S5. Esto hace que el contraste entre la rigidez del hueso y el titanio sea mayor y
por tanto, el orden de la singularidad mas elevado em el borde libre.
8.3. Influencia de la edad
En este apartado se va a analizar la influencia de la edad en el comportamiento del
hueso periimplantario. Dicha influencia se va a analizar a traves del parametro fbio, que,
como ya se ha dicho, disminuye con la edad. No tiene sentido estudiar el comportamiento
de implantes en ninos, que solo mudan la dentadura decıdua (vulgarmente conocida como
dentadura de leche) por la dentadura adulta. Mas normal es el uso de implantes dentales en
ancianos. Incluso en algunos casos, normalmente de tipo traumatico, hay jovenes a los que se
les colocan implantes dentales. Se analizara un implante colocado a una persona joven, que
tiene una frecuencia de activacion de BMUs mayor que la de un adulto, pudiendo extraerse
conclusiones contrarias en el caso de un anciano, con fbio menores. Si para adultos se supuso
fbio = 0.03 BMUsmm2 dıa , para jovenes se usara un valor ligeramente superior, fbio = 0.05 BMUs
mm2 dıa .
La actividad remodelatoria es mas intensa en jovenes, el numero de BMUs activadas es

8.3 Influencia de la edad 369
mayor, y por tanto, los efectos que tiene la insercion del implante se van a ver magnificados
por este hecho, como se vera a continuacion. La densidad inicial supuesta para el hueso
periimplantario es ρ0 = 2.0 g/cm3 y la secuencia simulada es S4.
Evolucion del dano
En la figura 8.30 se muestra la distribucion de tres componentes del tensor de dano a los
10 y a los 400 dıas de simulacion. Como en las otras simulaciones que usan la secuencia S4,
el dano crece muy rapidamente al principio (practicamente en los primeros dıas), para luego
frenarse en su propagacion, gracias al efecto de las BMUs que se activan, principalmente,
para reparar ese dano.
Dado que ahora, con fbio = 0.05, el numero de BMUs que se activan por unidad de
tiempo es mas alto, hay mas BMUs activas en cada instante para el mismo nivel de dano
y, por tanto, la reparacion del mismo es mas efectiva. Este hecho se puede comprobar en la
figura 8.31, en la que se compara el dano al final de esta simulacion y al final de la simulacion
que utiliza fbio = 0.03. La distribucion del dano es muy parecida en ambas, siendo el dano
mas alto el que se produce en direccion x, debido a la flexion, y la zona mas danada la
del cuello del implante. Sin embargo, debido al mayor numero de BMUs activas, el area
danada no es tan amplia para fbio = 0.05, si bien, las diferencias son muy pequenas. Hay
que tener en cuenta que los valores fisiologicos normales del dano estan bastante por debajo
de 0.1, ocupando la zona azul. El resto tiene valores de dano patologicos muy difıciles de
reparar. Y es que en aquellas zonas donde las tensiones son demasiado altas, como el cuello
del implante, el dano no da tiempo a repararse aunque haya muchas BMUs activas, y la
zona queda danada por completo; en cualquier caso, fbio = 0.03 o fbio = 0.05.
Evolucion de la densidad osea
En este caso se han simulado 400 dıas de masticacion, que como se ve en la figura 8.32 son
suficientes para alcanzar la convergencia de la fraccion volumetrica promedio, vb, es decir,
un estado de equilibrio de remodelacion global en el hueso periimplantario. El valor final
de vb es 0.844, lo que quiere decir que se produce una reabsorcion notable. Esto es normal
ya que la actividad remodelatoria es mas intensa ahora que el numero de BMUs activas es
mayor.
En las figuras 8.33 y 8.34 se muestra la evolucion local de la fraccion volumetrica de
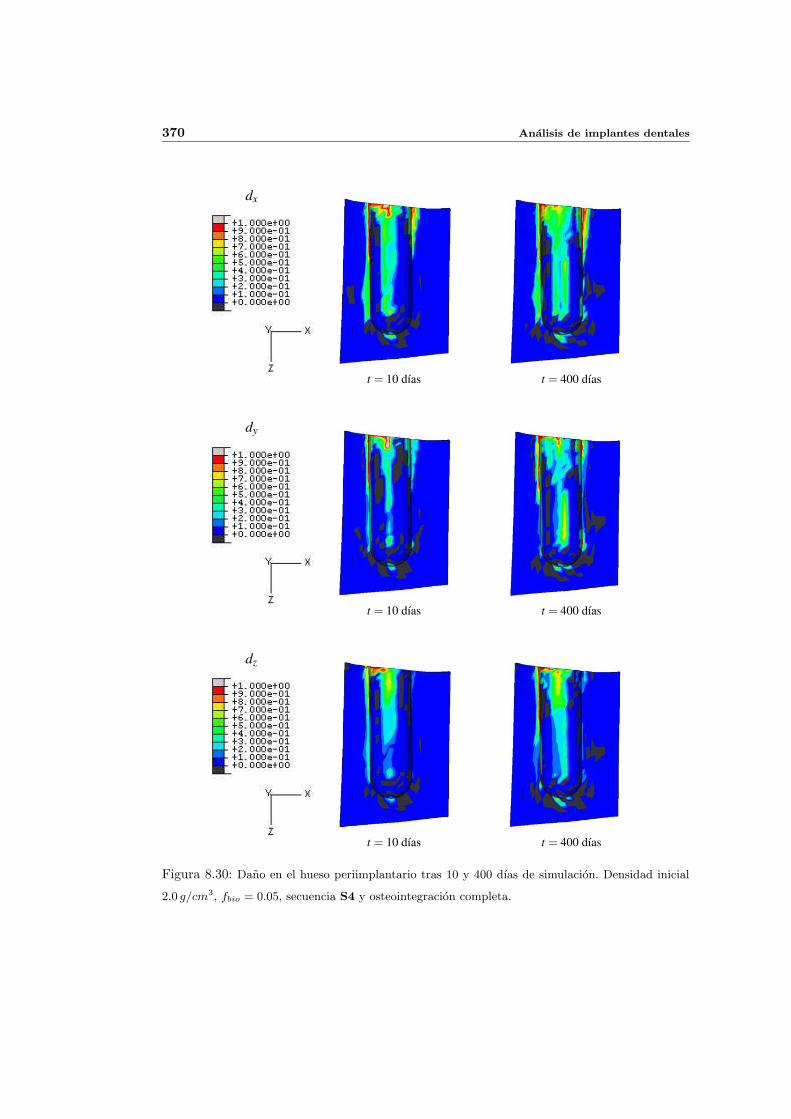
370 Analisis de implantes dentales
dx
t = 10 dıas t = 400 dıas
dy
t = 10 dıas t = 400 dıas
dz
t = 10 dıas t = 400 dıas
1
Figura 8.30: Dano en el hueso periimplantario tras 10 y 400 dıas de simulacion. Densidad inicial
2.0 g/cm3, fbio = 0.05, secuencia S4 y osteointegracion completa.
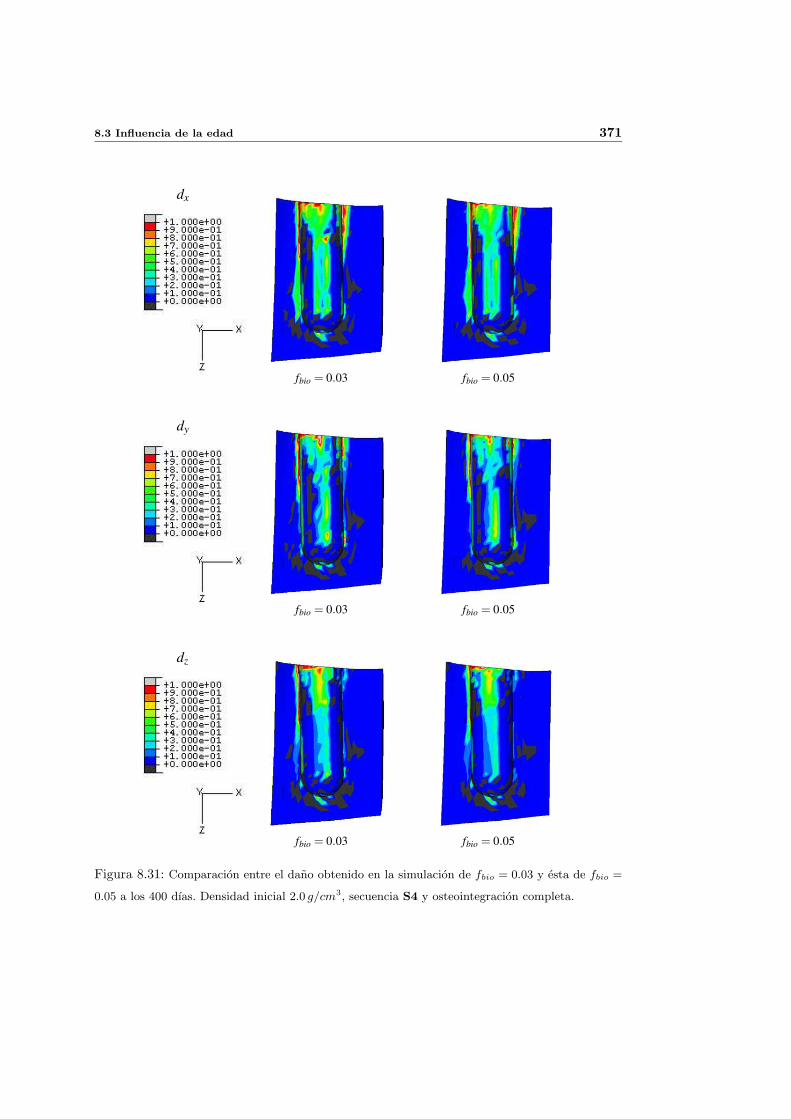
8.3 Influencia de la edad 371
dx
fbio = 0.03 fbio = 0.05
dy
fbio = 0.03 fbio = 0.05
dz
fbio = 0.03 fbio = 0.05
1
Figura 8.31: Comparacion entre el dano obtenido en la simulacion de fbio = 0.03 y esta de fbio =
0.05 a los 400 dıas. Densidad inicial 2.0 g/cm3, secuencia S4 y osteointegracion completa.
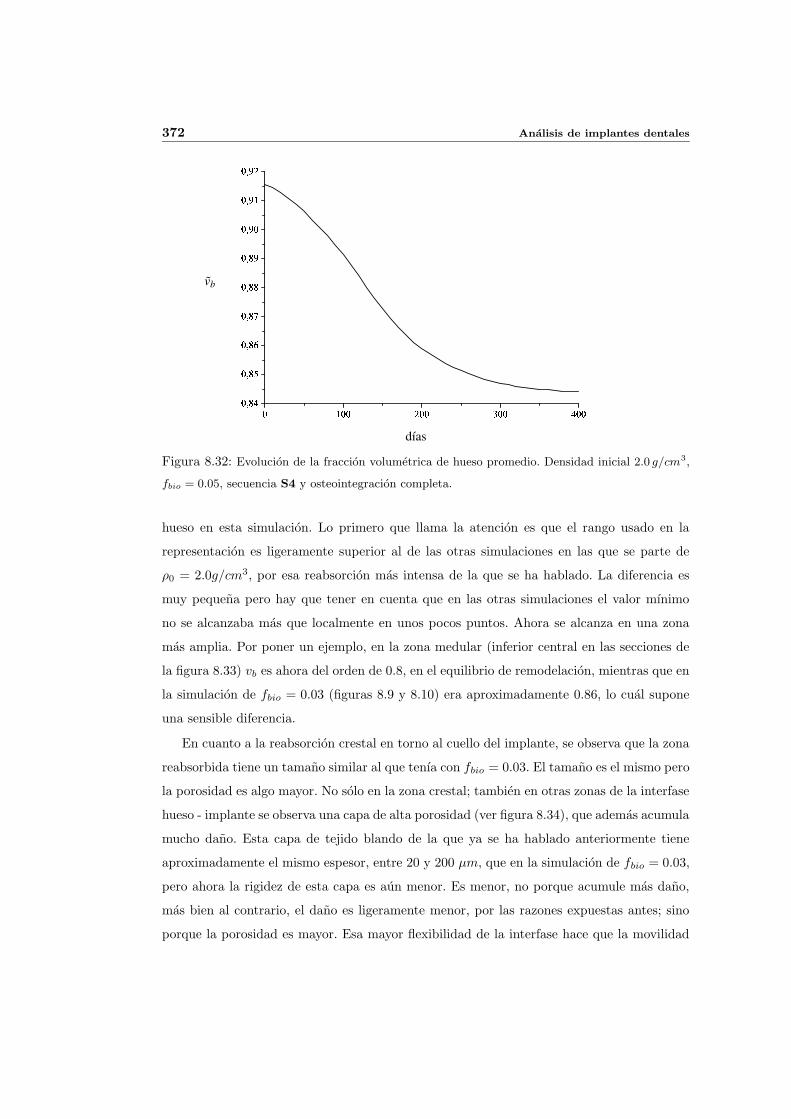
372 Analisis de implantes dentales
0 100 200 300 40005 64
05 67
05 68
05 69
05 66
05 6:
05 :0
05 :/1
05 :2
dıas
vb
1
Figura 8.32: Evolucion de la fraccion volumetrica de hueso promedio. Densidad inicial 2.0 g/cm3,
fbio = 0.05, secuencia S4 y osteointegracion completa.
hueso en esta simulacion. Lo primero que llama la atencion es que el rango usado en la
representacion es ligeramente superior al de las otras simulaciones en las que se parte de
ρ0 = 2.0g/cm3, por esa reabsorcion mas intensa de la que se ha hablado. La diferencia es
muy pequena pero hay que tener en cuenta que en las otras simulaciones el valor mınimo
no se alcanzaba mas que localmente en unos pocos puntos. Ahora se alcanza en una zona
mas amplia. Por poner un ejemplo, en la zona medular (inferior central en las secciones de
la figura 8.33) vb es ahora del orden de 0.8, en el equilibrio de remodelacion, mientras que en
la simulacion de fbio = 0.03 (figuras 8.9 y 8.10) era aproximadamente 0.86, lo cual supone
una sensible diferencia.
En cuanto a la reabsorcion crestal en torno al cuello del implante, se observa que la zona
reabsorbida tiene un tamano similar al que tenıa con fbio = 0.03. El tamano es el mismo pero
la porosidad es algo mayor. No solo en la zona crestal; tambien en otras zonas de la interfase
hueso - implante se observa una capa de alta porosidad (ver figura 8.34), que ademas acumula
mucho dano. Esta capa de tejido blando de la que ya se ha hablado anteriormente tiene
aproximadamente el mismo espesor, entre 20 y 200 µm, que en la simulacion de fbio = 0.03,
pero ahora la rigidez de esta capa es aun menor. Es menor, no porque acumule mas dano,
mas bien al contrario, el dano es ligeramente menor, por las razones expuestas antes; sino
porque la porosidad es mayor. Esa mayor flexibilidad de la interfase hace que la movilidad
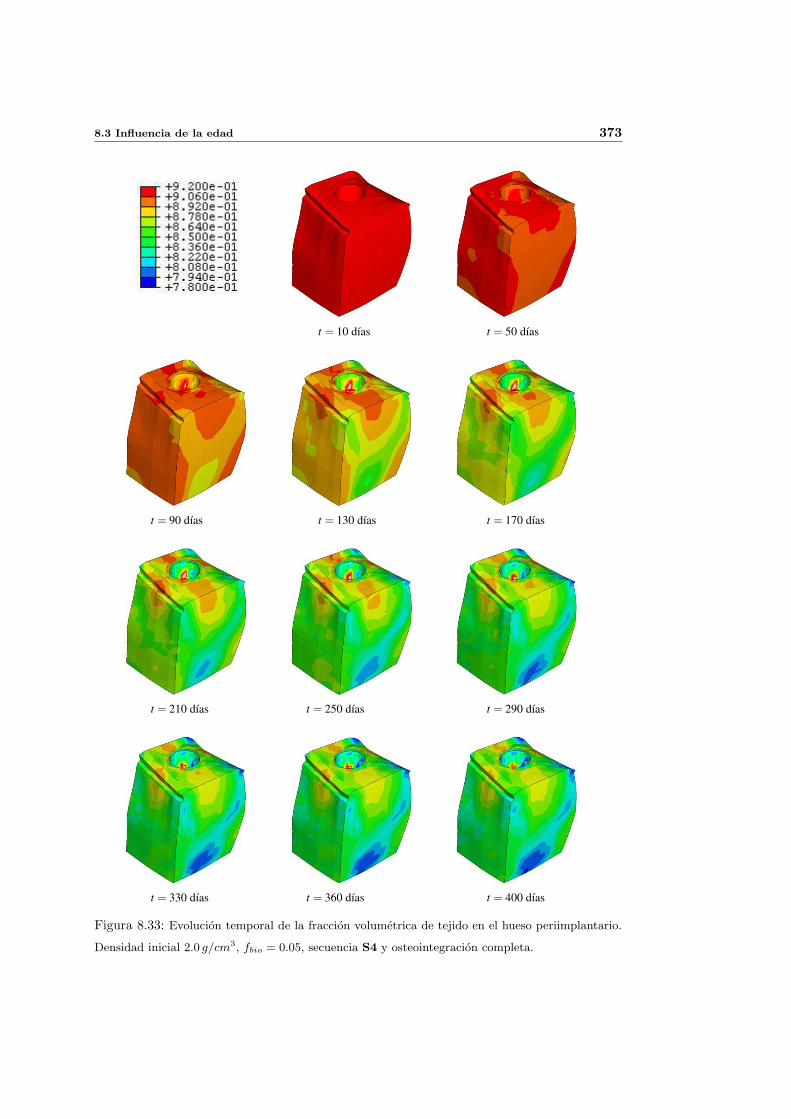
8.3 Influencia de la edad 373
t = 10 dıas t = 50 dıas
t = 90 dıas t = 130 dıas t = 170 dıas
t = 210 dıas t = 250 dıas t = 290 dıas
t = 330 dıas t = 360 dıas t = 400 dıas
1
Figura 8.33: Evolucion temporal de la fraccion volumetrica de tejido en el hueso periimplantario.
Densidad inicial 2.0 g/cm3, fbio = 0.05, secuencia S4 y osteointegracion completa.
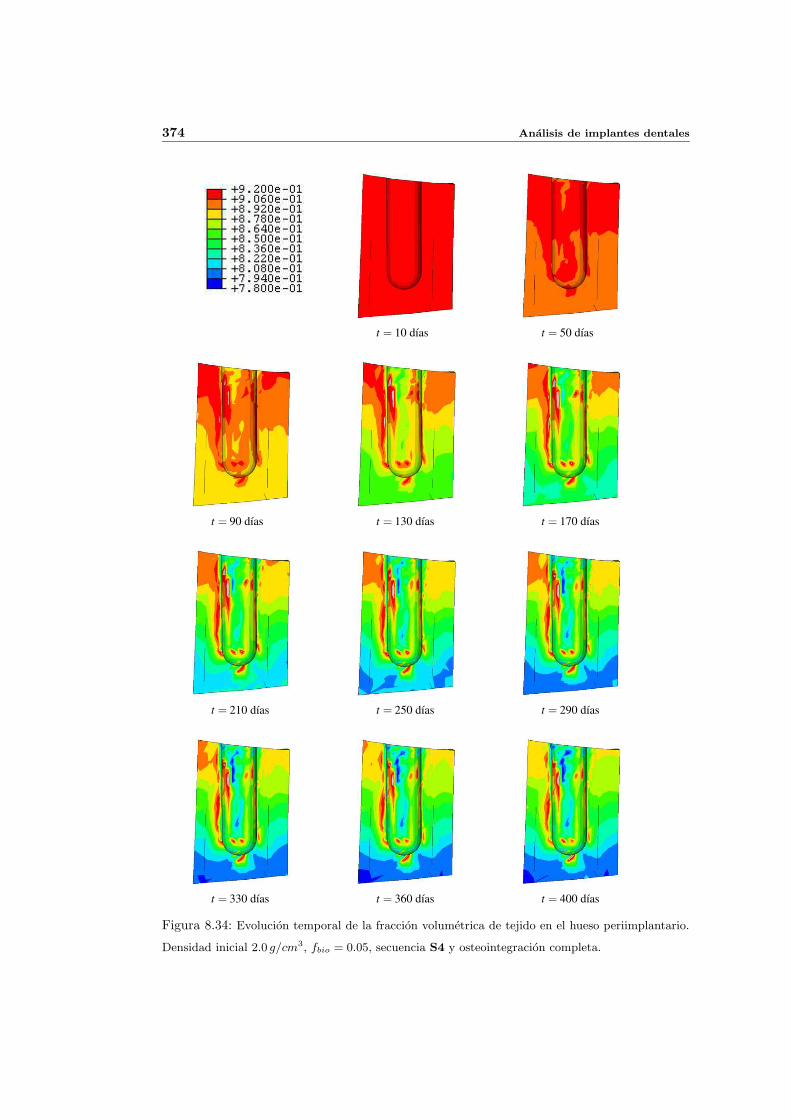
374 Analisis de implantes dentales
t = 10 dıas t = 50 dıas
t = 90 dıas t = 130 dıas t = 170 dıas
t = 210 dıas t = 250 dıas t = 290 dıas
t = 330 dıas t = 360 dıas t = 400 dıas
1
Figura 8.34: Evolucion temporal de la fraccion volumetrica de tejido en el hueso periimplantario.
Densidad inicial 2.0 g/cm3, fbio = 0.05, secuencia S4 y osteointegracion completa.

8.3 Influencia de la edad 375
del implante sea mayor, lo que puede comprometer su estabilidad y consecuentemente su
exito.
Discusion de los resultados
Con la edad disminuye la actividad remodelatoria de las BMUs, que se activan con menor
frecuencia. Esto tiene importantes consecuencias cuando se coloca un implante dental, que
provoca cambios en la forma de transferir las cargas de masticacion al hueso periimplantario.
Estos cambios provocan la activacion de BMUs, que en personas jovenes son mas numerosas
y esta mayor presencia de BMUs hace que aumente la porosidad de forma bastante notable.
Por lo general, el dano es mayor en ancianos. Crece exponencialmente con la edad,
especialmente en mujeres, como afirman Schaffler et al. [300] y Mori et al. [242]. Este hecho
estarıa de acuerdo con los resultados, que para fbio = 0.03, es decir, en adultos, predicen
un dano ligeramente menor al obtenido con fbio = 0.05, en jovenes. Si esa tendencia se
mantiene, en ancianos, con un valor de fbio aun menor que 0.03, el dano serıa mayor que en
adultos de mediana edad, por el escaso numero de BMUs que hay activas para repararlo.
En la figura 8.35 se analiza la influencia de la edad sobre la fraccion volumetrica de
tejido danado promedio, h, observandose diferencias muy pequenas entre un caso y otro.
Para fbio = 0.05 la reparacion del dano es mas efectiva y la cantidad de tejido danado es
menor, pero solo ligeramente. Hay que tener en cuenta que en el promedio que representa
h tiene un peso importante el tejido adyacente al implante, que esta muy danado. Como
el dano de esa zona no da tiempo a repararse antes de que aumente de nuevo por fatiga,
aunque sea mayor la poblacion de BMUs, las diferencias entre ambos casos son pequenas.
Estas diferencias se deben mas a la contribucion sobre el promedio de otras zonas del hueso
periimplantario, con un dano moderado, donde sı se nota la reparacion por remodelacion y
el hecho de que la poblacion de BMUs sea mas numerosa. Tambien hay que tener en cuenta
que la variable h mide la proporcion de volumen de tejido danado respecto al volumen total
y no respecto al volumen de tejido, que en jovenes es menor, como se ha visto antes. Si se
hiciera la proporcion respecto al volumen de tejido las diferencias serıan un poco mayores,
lo que quiere decir que la cantidad de tejido periimplantario es menor pero conserva una
mejor integridad.
Otro aspecto importante que hay que destacar es el bajo contenido en mineral que tiene el
tejido periimplantario en promedio. Ello es debido al gran numero de BMUs que se activan,
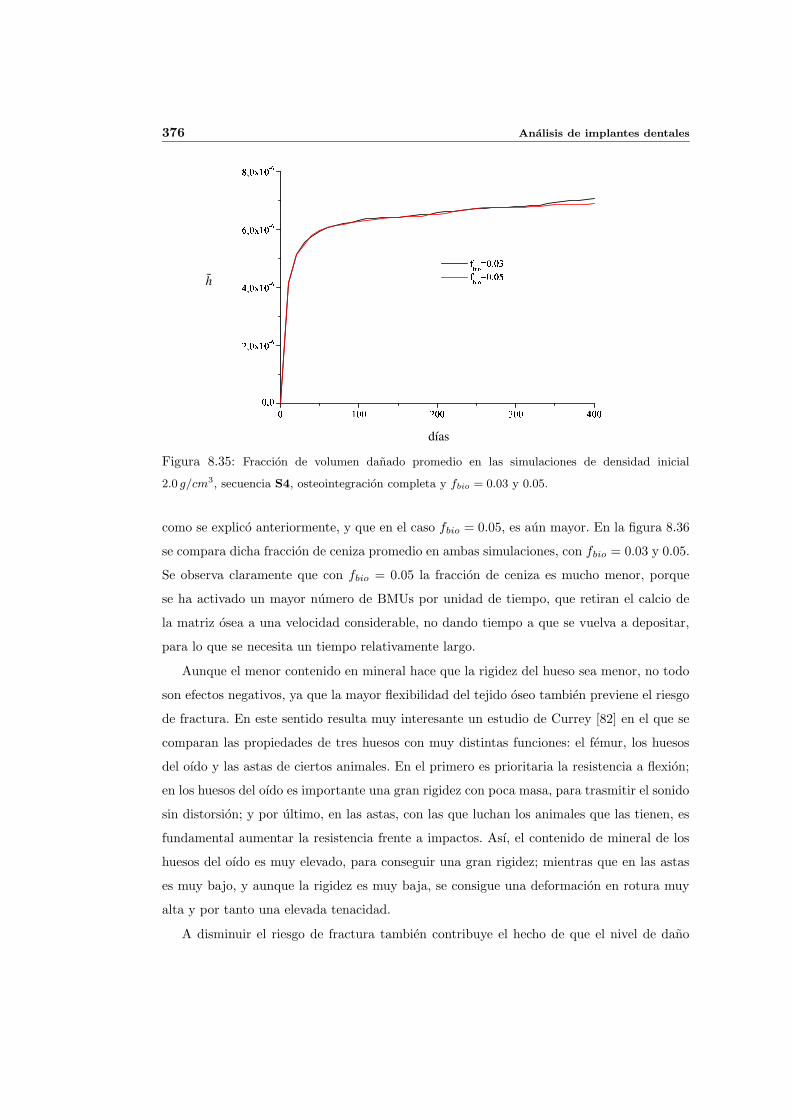
376 Analisis de implantes dentales
; <;; =;; >;; ?;;;@ ;
=@ ;AB<;C D
?@ ;AB<;C D
E@ ;AB<;C D
F@ ;AB<; C D
G H I J K ;L ;>G H I J K ;L ;M
dıas
h
1
Figura 8.35: Fraccion de volumen danado promedio en las simulaciones de densidad inicial
2.0 g/cm3, secuencia S4, osteointegracion completa y fbio = 0.03 y 0.05.
como se explico anteriormente, y que en el caso fbio = 0.05, es aun mayor. En la figura 8.36
se compara dicha fraccion de ceniza promedio en ambas simulaciones, con fbio = 0.03 y 0.05.
Se observa claramente que con fbio = 0.05 la fraccion de ceniza es mucho menor, porque
se ha activado un mayor numero de BMUs por unidad de tiempo, que retiran el calcio de
la matriz osea a una velocidad considerable, no dando tiempo a que se vuelva a depositar,
para lo que se necesita un tiempo relativamente largo.
Aunque el menor contenido en mineral hace que la rigidez del hueso sea menor, no todo
son efectos negativos, ya que la mayor flexibilidad del tejido oseo tambien previene el riesgo
de fractura. En este sentido resulta muy interesante un estudio de Currey [82] en el que se
comparan las propiedades de tres huesos con muy distintas funciones: el femur, los huesos
del oıdo y las astas de ciertos animales. En el primero es prioritaria la resistencia a flexion;
en los huesos del oıdo es importante una gran rigidez con poca masa, para trasmitir el sonido
sin distorsion; y por ultimo, en las astas, con las que luchan los animales que las tienen, es
fundamental aumentar la resistencia frente a impactos. Ası, el contenido de mineral de los
huesos del oıdo es muy elevado, para conseguir una gran rigidez; mientras que en las astas
es muy bajo, y aunque la rigidez es muy baja, se consigue una deformacion en rotura muy
alta y por tanto una elevada tenacidad.
A disminuir el riesgo de fractura tambien contribuye el hecho de que el nivel de dano
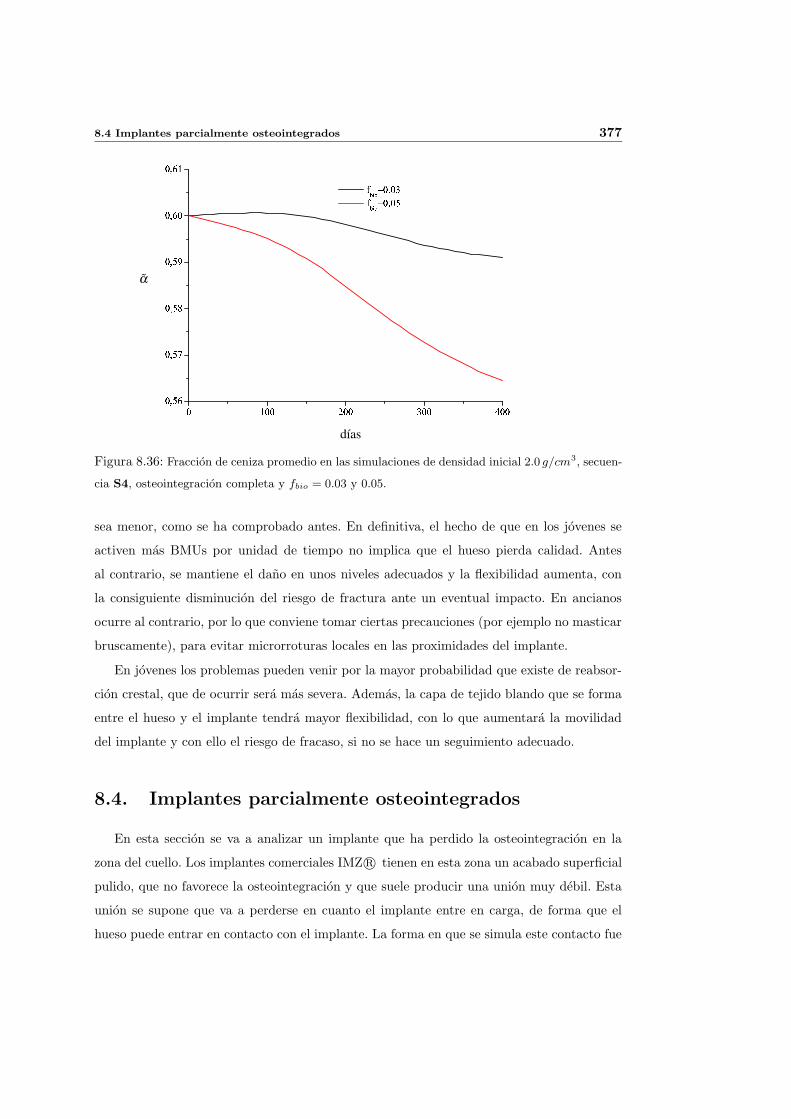
8.4 Implantes parcialmente osteointegrados 377
N ONN PNN QNN RNNNS TU
NS TV
NS TW
NS TX
NS UN
NS U/OY Z [ \ ] N_^ NQY Z [ \ ] N_^ NT
dıas
α
1
Figura 8.36: Fraccion de ceniza promedio en las simulaciones de densidad inicial 2.0 g/cm3, secuen-
cia S4, osteointegracion completa y fbio = 0.03 y 0.05.
sea menor, como se ha comprobado antes. En definitiva, el hecho de que en los jovenes se
activen mas BMUs por unidad de tiempo no implica que el hueso pierda calidad. Antes
al contrario, se mantiene el dano en unos niveles adecuados y la flexibilidad aumenta, con
la consiguiente disminucion del riesgo de fractura ante un eventual impacto. En ancianos
ocurre al contrario, por lo que conviene tomar ciertas precauciones (por ejemplo no masticar
bruscamente), para evitar microrroturas locales en las proximidades del implante.
En jovenes los problemas pueden venir por la mayor probabilidad que existe de reabsor-
cion crestal, que de ocurrir sera mas severa. Ademas, la capa de tejido blando que se forma
entre el hueso y el implante tendra mayor flexibilidad, con lo que aumentara la movilidad
del implante y con ello el riesgo de fracaso, si no se hace un seguimiento adecuado.
8.4. Implantes parcialmente osteointegrados
En esta seccion se va a analizar un implante que ha perdido la osteointegracion en la
zona del cuello. Los implantes comerciales IMZ r© tienen en esta zona un acabado superficial
pulido, que no favorece la osteointegracion y que suele producir una union muy debil. Esta
union se supone que va a perderse en cuanto el implante entre en carga, de forma que el
hueso puede entrar en contacto con el implante. La forma en que se simula este contacto fue

378 Analisis de implantes dentales
descrita en el capıtulo 3 y la zona en que se produce se extiende hasta una cierta profundidad,
donde la superficie del implante vuelve a ser rugosa y la union es mas fuerte. En esta zona
rugosa se supone que la osteointegracion no se va a perder durante toda la simulacion. La
densidad inicial supuesta para el hueso periimplantario es ρ0 = 2.0 g/cm3 y la secuencia
simulada es S4. El resto de parametros del modelo tienen los valores nominales de la tabla
8.1.
Con las nuevas condiciones de contacto en la union hueso - implante, cambia la distribu-
cion de tensiones con respecto a las simulaciones anteriores. Las nuevas tensiones de von
Mises para los cuatro casos de cargas de la secuencia S4 se muestran en la figura 8.37. Se
puede observar que ahora la singularidad de tensiones no se produce en el borde libre de la
interfase, sino en el punto donde comienza la osteointegracion perfecta, que es en realidad
el borde de una grieta. La zona crestal que queda por encima del borde de grieta se ha
descargado ligeramente respecto a la situacion anterior, aunque las tensiones siguen siendo
altas, ya no lo son tanto. La zona que entra en contacto con el implante como consecuencia
de la deformacion de la mandıbula soporta altas tensiones de compresion, pero hay una
parte del hueso que se separa del implante y ya no soporta tracciones como antes, cuando
la osteointegracion hacıa que el implante tirara de ella. El contacto esta redistribuyendo las
tensiones, que ahora tienen un nivel moderado en una zona mas amplia y un valor maximo
mas reducido. Para la carga MImp las tensiones maximas estan en torno a 350MPa mientras
que en el caso de osteointegracion completa una buena zona del cuello del implante soporta-
ba tensiones de von Mises por encima de 450MPa. Esto tiene un efecto muy positivo sobre
el hueso que para un buen mantenimiento necesita un nivel de estımulo adecuado y evitar
tanto el desuso, como una sobrecarga muy alta que provoque dano. Tambien se extiende ha-
cia abajo la zona de hueso que soporta un nivel de tensiones moderado, como consecuencia
de la referida redistribucion de las tensiones. En general, se puede decir que las tensiones
estan mas repartidas cuando se pierde la osteointegracion en parte del cuello del implante y
eso va a tener consecuencias positivas muy importantes, como se vera a continuacion.
Evolucion del dano
El dano que acumula el hueso, ahora que se tiene osteointegracion parcial, es bastante
mas pequeno que en el caso de osteointegracion completa, como puede observarse en la figura
8.38. Este resultado es consecuencia directa de lo que se acaba de decir sobre las tensiones:
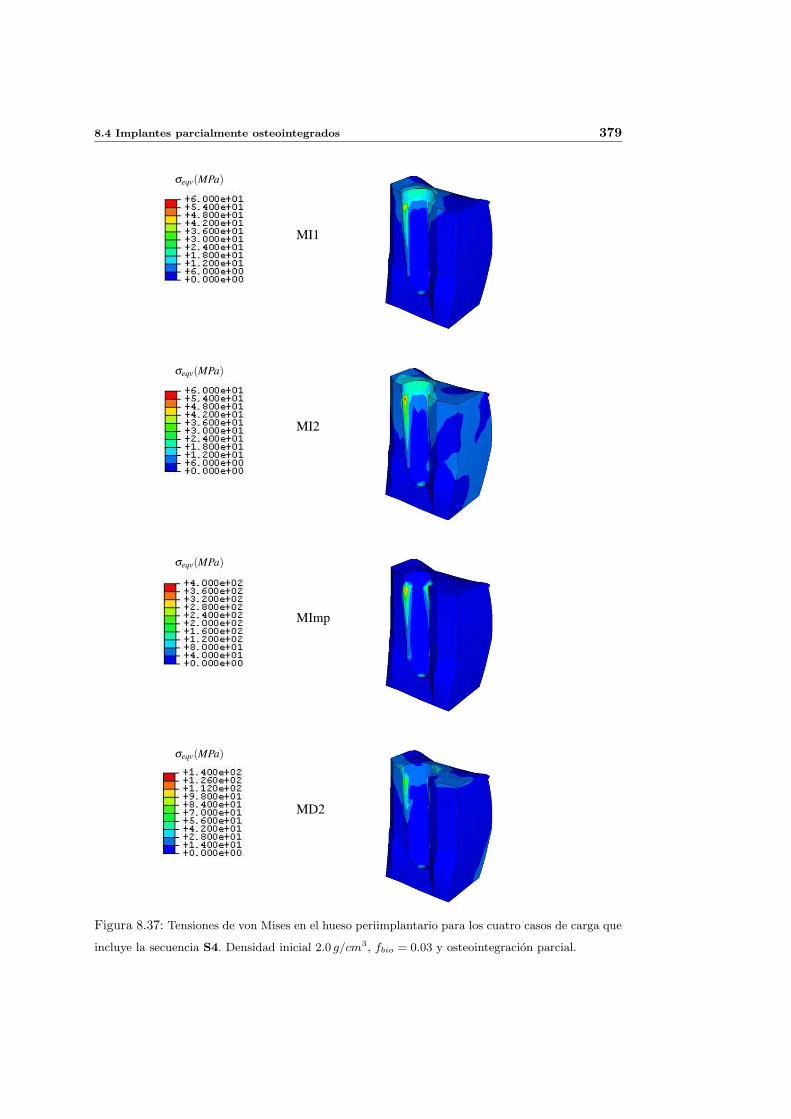
8.4 Implantes parcialmente osteointegrados 379
σeqv(MPa)
σeqv(MPa)
σeqv(MPa)
σeqv(MPa)
MI1
MI2
MImp
MD2
1
Figura 8.37: Tensiones de von Mises en el hueso periimplantario para los cuatro casos de carga que
incluye la secuencia S4. Densidad inicial 2.0 g/cm3, fbio = 0.03 y osteointegracion parcial.

380 Analisis de implantes dentales
al estar mas repartidas, las tensiones maximas son menores. Ademas cambia la distribucion
del dano. Ahora, la zona que acumula mas dano esta por debajo del borde de grieta. La zona
crestal que ha perdido la osteointegracion tiene un dano muy pequeno, porque las tensiones
son moderadas y principalmente de compresion. El dano mas alto se produce en direccion
x, muy concentrado en los bordes mesial y labial, y mas repartido en los bordes distal y
lingual, donde ocupa un area mas extensa.
La evolucion del dano es muy parecida a la de las otras simulaciones en las que se parte
de una densidad de 2.0 g/cm3: el dano se acumula principalmente en los primeros dıas de
simulacion y a partir de entonces se propaga muy lentamente, siendo pequena la diferencia
entre el estado a los 10 y a los 400 dıas de simulacion.
En el borde libre del cuello del implante tambien se observa una zona con dano elevado
situada en posicion mesio-lingual. Existe otra zona opuesta a esta, en posicion disto-labial,
que igualmente tiene un dano considerable. El dano tambien se acumula en esta zona porque
aunque no haya union (por la falta de osteointegracion) sı hay contacto y sigue habiendo una
singularidad de tensiones en este borde libre, aunque con un orden de singularidad menor,
por las condiciones de contorno de contacto con friccion.
Comparando la fraccion volumetrica de tejido danado promedio, h, en los casos de os-
teointegracion completa y parcial, como se hace en la figura 8.39, se comprueba que el dano
es menor en este ultimo caso (h es aproximadamente la mitad). El nivel de dano es similar
en casi todo el hueso periimplantario, excepto en la zona mas proxima al implante, que es
precisamente la que tiene mas peso en el promedio y la que hace las diferencias entre las
curvas de la figura 8.39.
Evolucion de la densidad osea
La figura 8.40 muestra la evolucion de la fraccion volumetrica de hueso promedio en la
region periimplantaria. El valor final, tras 400 dıas de simulacion, en los que se alcanza el
equilibrio de remodelacion, es 0.873, valor muy parecido al que se obtuvo con osteointegracion
completa.
La evolucion local de vb se muestra en las figuras 8.41 y 8.42. En ellas se comprueba la
tendencia, evidenciada en otras simulaciones, de reabsorcion osea en la zona medular y en
la zona del cuello del implante, aunque en este ultimo caso muy leve. De hecho, incluso se
ha tenido que aumentar el extremo superior del rango en el que se representa vb, porque hay
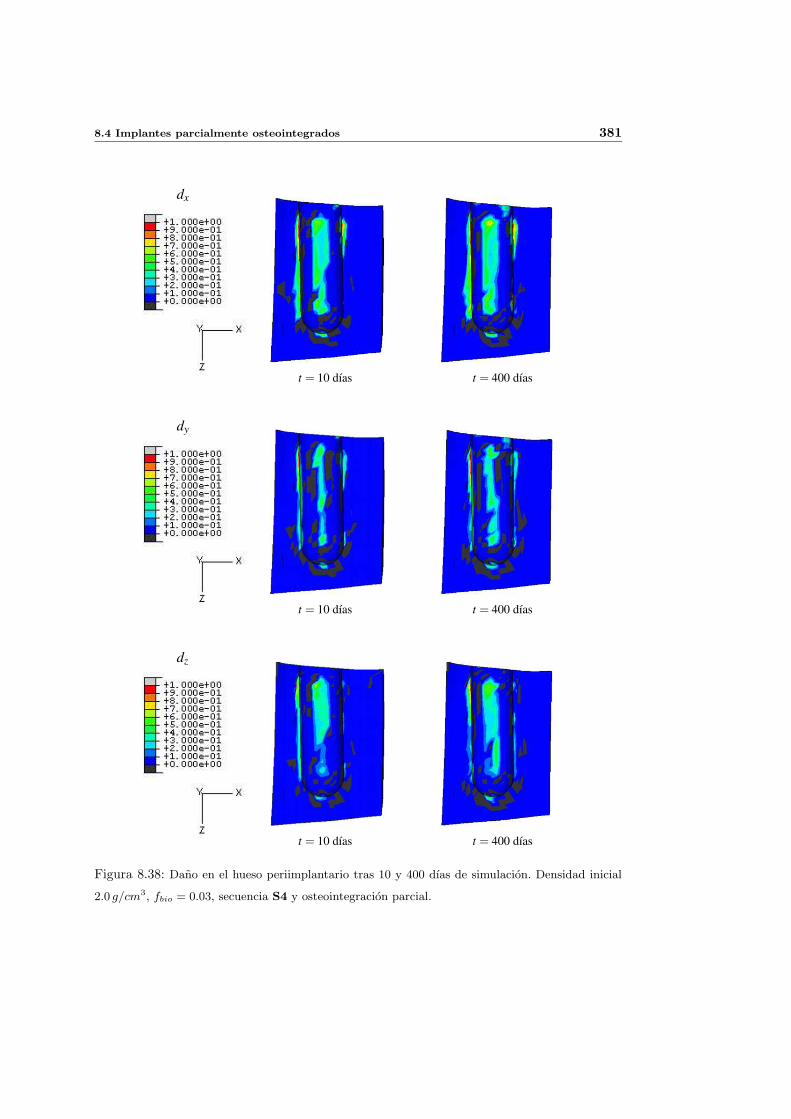
8.4 Implantes parcialmente osteointegrados 381
dx
t = 10 dıas t = 400 dıas
dy
t = 10 dıas t = 400 dıas
dz
t = 10 dıas t = 400 dıas
1
Figura 8.38: Dano en el hueso periimplantario tras 10 y 400 dıas de simulacion. Densidad inicial
2.0 g/cm3, fbio = 0.03, secuencia S4 y osteointegracion parcial.
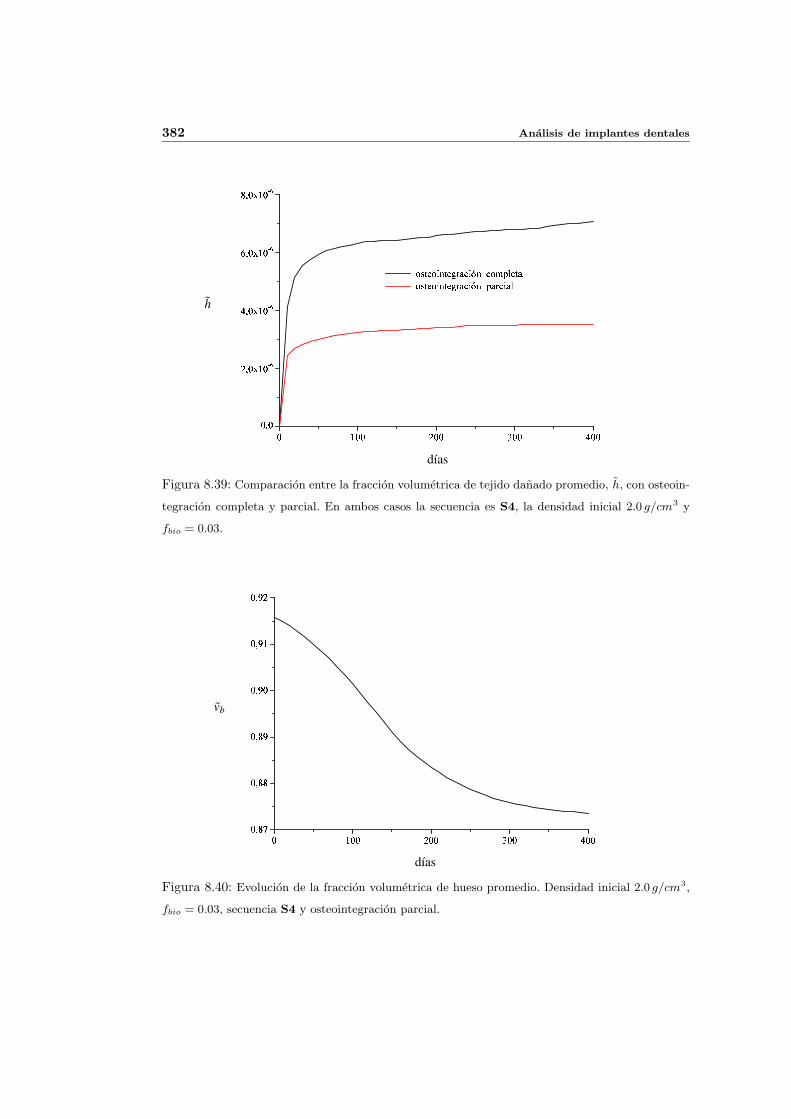
382 Analisis de implantes dentales
` a`` b`` c`` d```e `
be `fBa`g h
de `fBa`g h
ie `fBa`g h
je `fBa` g h
kl m nko pm nqr sto upvtkwyxz nm skl m nko pm nqr sto upxsr to sz
dıas
h
1
Figura 8.39: Comparacion entre la fraccion volumetrica de tejido danado promedio, h, con osteoin-
tegracion completa y parcial. En ambos casos la secuencia es S4, la densidad inicial 2.0 g/cm3 y
fbio = 0.03.
| || ~|| || |||
|
|
| |
| /
| ~
dıas
vb
1
Figura 8.40: Evolucion de la fraccion volumetrica de hueso promedio. Densidad inicial 2.0 g/cm3,
fbio = 0.03, secuencia S4 y osteointegracion parcial.

8.4 Implantes parcialmente osteointegrados 383
zonas del cuello del implante donde se produce incluso formacion neta. Este valor maximo
es 0.932 y se recuerda que vb0 = 0.916.
En el borde distal del cuello del implante, izquierdo en las secciones de la figura 8.42, se
produce reabsorcion como consecuencia del dano acumulado (ver figura 8.38), aunque esa
reabsorcion es muy pequena, como lo es el dano. Solo existe otra zona en la que se produce
reabsorcion crestal y es la disto-lingual, aunque apenas es perceptible. El mantenimiento del
hueso a nivel crestal es aceptable por el nivel de tensiones que soporta, bastante uniformes
y de nivel moderado. En el resto de la interfase la integridad del hueso circundante parece
que tampoco esta comprometida. Se observan distintas zonas donde se produce reabsorcion,
por ejemplo, en el borde lingual, donde el dano estaba muy extendido, aunque no era muy
elevado, vb ronda el valor 0.86 y solo en algunos puntos esta ligeramente por debajo de
0.85. Es decir, en la interfase apenas se produce reabsorcion mientras el dano este en niveles
moderados.
En la zona medular, en cambio, la reabsorcion es mas acusada, siendo notable la difer-
encia con la zona crestal. Tambien hay ligeras diferencias con las anteriores simulaciones.
Efectivamente, en el equilibrio de remodelacion vb esta en torno a 0.83 en buena parte de
esta region, mientras que en la simulacion del implante con osteointegracion completa pocos
eran los puntos en los que vb bajaba de 0.84. Teniendo en cuenta lo poco que varıa vb en
estas simulaciones, esta diferencia, aparentemente pequena, no es para nada despreciable.
Dado que la distribucion de densidades ha cambiado por completo con respecto a los
casos de osteointegracion completa, resulta interesante analizar la distribucion de vb a lo
largo de la interfase. En la figura 8.43 se muestra la evolucion de vb en algunos puntos
de ella. Lo mas destacado de esta grafica es que los puntos de la zona superior tienen una
porosidad por debajo del resto. La reabsorcion en estos puntos es inapreciable e incluso en la
zona superior lingual hay formacion neta. Los puntos de la zona superior estan enfrentados
a la region pulida del implante, donde se ha perdido la osteointegracion y el dano esta muy
por debajo de los valores que tiene en los implantes con osteointegracion completa, como se
acaba de ver. Esto explica que la reabsorcion sea mucho menor. Como ademas, en el cuello
del implante el estımulo mecanico es suficientemente alto por la compresion, se forma incluso
mas tejido del que se supone inicialmente.
Exceptuando la zona superior, la region del apice es la que sufre menor reabsorcion del
resto de la interfase, algo parecido a lo que ocurrıa en el caso de osteointegracion completa,
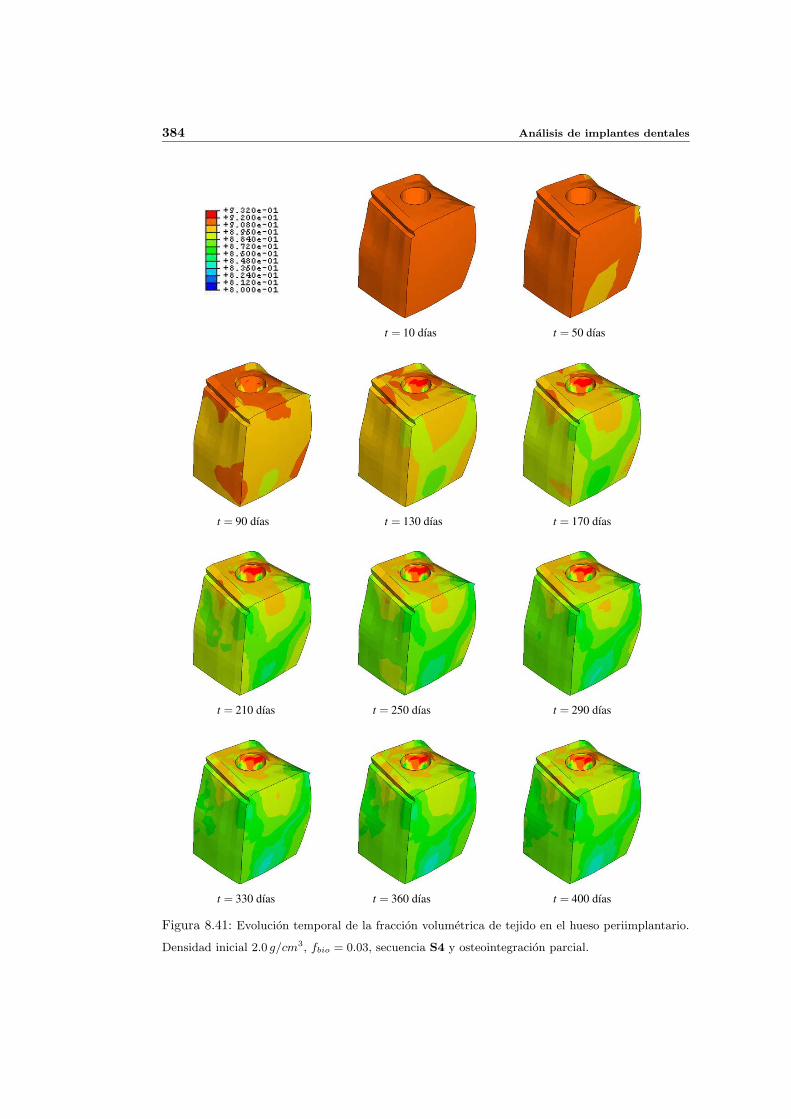
384 Analisis de implantes dentales
t = 10 dıas t = 50 dıas
t = 90 dıas t = 130 dıas t = 170 dıas
t = 210 dıas t = 250 dıas t = 290 dıas
t = 330 dıas t = 360 dıas t = 400 dıas
1
Figura 8.41: Evolucion temporal de la fraccion volumetrica de tejido en el hueso periimplantario.
Densidad inicial 2.0 g/cm3, fbio = 0.03, secuencia S4 y osteointegracion parcial.
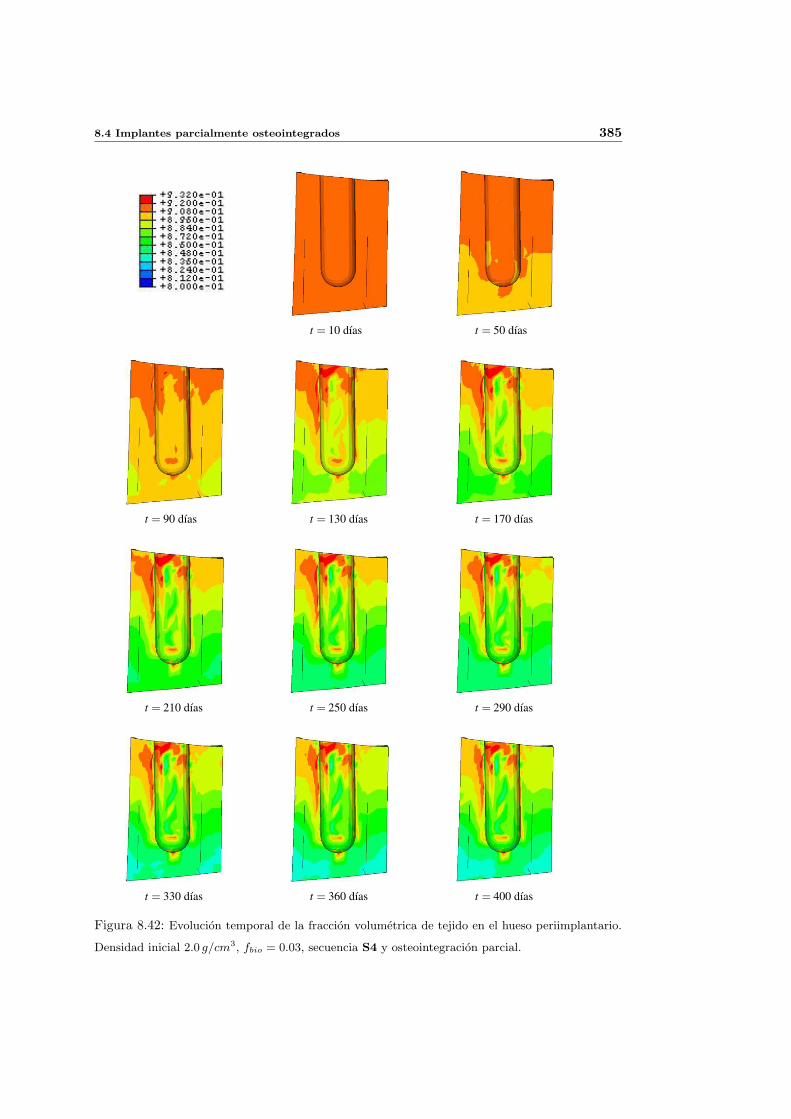
8.4 Implantes parcialmente osteointegrados 385
t = 10 dıas t = 50 dıas
t = 90 dıas t = 130 dıas t = 170 dıas
t = 210 dıas t = 250 dıas t = 290 dıas
t = 330 dıas t = 360 dıas t = 400 dıas
1
Figura 8.42: Evolucion temporal de la fraccion volumetrica de tejido en el hueso periimplantario.
Densidad inicial 2.0 g/cm3, fbio = 0.03, secuencia S4 y osteointegracion parcial.
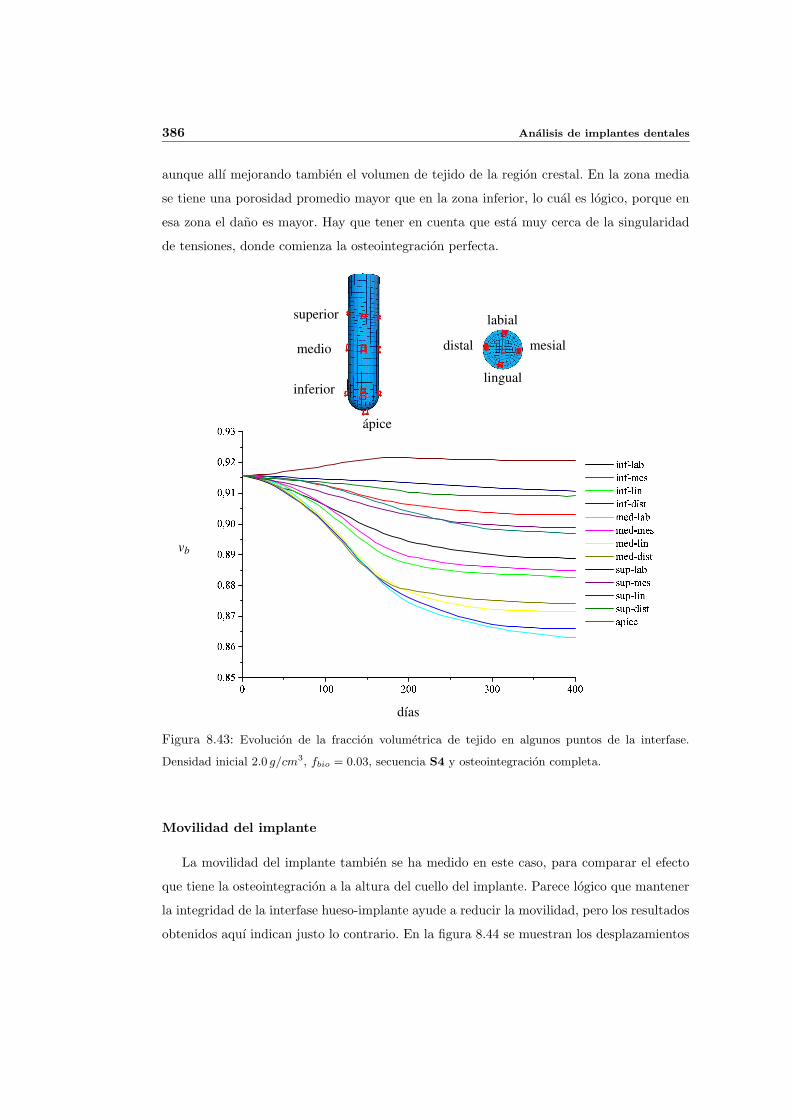
386 Analisis de implantes dentales
aunque allı mejorando tambien el volumen de tejido de la region crestal. En la zona media
se tiene una porosidad promedio mayor que en la zona inferior, lo cual es logico, porque en
esa zona el dano es mayor. Hay que tener en cuenta que esta muy cerca de la singularidad
de tensiones, donde comienza la osteointegracion perfecta.
_ __ _ __
/ / / / _ / _ / _ / _ / / ¡
distal mesial
lingual
labialsuperior
medio
inferior
apice
dıas
vb
1
Figura 8.43: Evolucion de la fraccion volumetrica de tejido en algunos puntos de la interfase.
Densidad inicial 2.0 g/cm3, fbio = 0.03, secuencia S4 y osteointegracion completa.
Movilidad del implante
La movilidad del implante tambien se ha medido en este caso, para comparar el efecto
que tiene la osteointegracion a la altura del cuello del implante. Parece logico que mantener
la integridad de la interfase hueso-implante ayude a reducir la movilidad, pero los resultados
obtenidos aquı indican justo lo contrario. En la figura 8.44 se muestran los desplazamientos
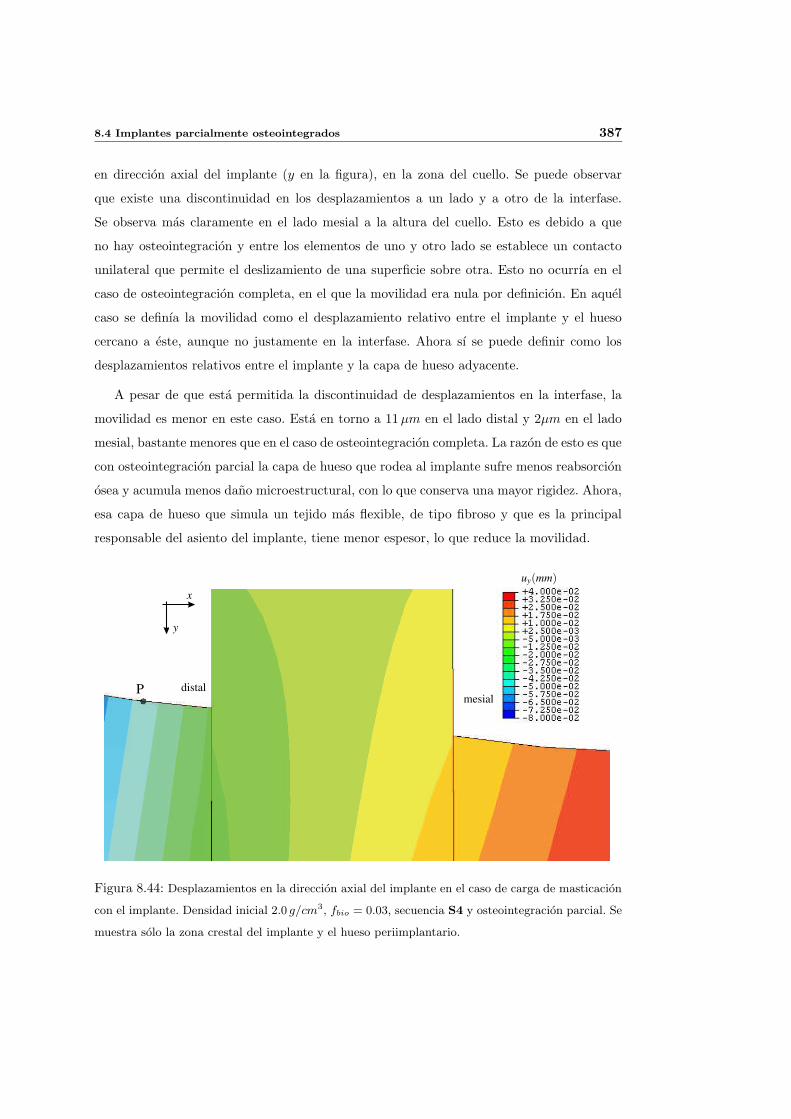
8.4 Implantes parcialmente osteointegrados 387
en direccion axial del implante (y en la figura), en la zona del cuello. Se puede observar
que existe una discontinuidad en los desplazamientos a un lado y a otro de la interfase.
Se observa mas claramente en el lado mesial a la altura del cuello. Esto es debido a que
no hay osteointegracion y entre los elementos de uno y otro lado se establece un contacto
unilateral que permite el deslizamiento de una superficie sobre otra. Esto no ocurrıa en el
caso de osteointegracion completa, en el que la movilidad era nula por definicion. En aquel
caso se definıa la movilidad como el desplazamiento relativo entre el implante y el hueso
cercano a este, aunque no justamente en la interfase. Ahora sı se puede definir como los
desplazamientos relativos entre el implante y la capa de hueso adyacente.
A pesar de que esta permitida la discontinuidad de desplazamientos en la interfase, la
movilidad es menor en este caso. Esta en torno a 11µm en el lado distal y 2µm en el lado
mesial, bastante menores que en el caso de osteointegracion completa. La razon de esto es que
con osteointegracion parcial la capa de hueso que rodea al implante sufre menos reabsorcion
osea y acumula menos dano microestructural, con lo que conserva una mayor rigidez. Ahora,
esa capa de hueso que simula un tejido mas flexible, de tipo fibroso y que es la principal
responsable del asiento del implante, tiene menor espesor, lo que reduce la movilidad.
uy(mm)
distalmesial
x
y
P
1
Figura 8.44: Desplazamientos en la direccion axial del implante en el caso de carga de masticacion
con el implante. Densidad inicial 2.0 g/cm3, fbio = 0.03, secuencia S4 y osteointegracion parcial. Se
muestra solo la zona crestal del implante y el hueso periimplantario.

388 Analisis de implantes dentales
Si se compara la figura 8.44 con la 8.12 se ve que el gradiente de desplazamientos, es
decir, la deformacion, es menor en el primer caso, con osteointegracion parcial. Esto confirma
que la rigidez del hueso periimplantario, sobre todo en el cuello del implante, se conserva
mejor si la osteointegracion se pierde localmente en la zona crestal.
En la figura 8.45 se muestra la evolucion del desplazamiento axial (en direccion y) del
punto P de la figura 8.44, en las masticaciones con el implante. Este desplazamiento da una
idea de cuanto se hunde el implante dentro del hueso periimplantario, ya que en ese caso
de carga, el implante tiene impedido los desplazamientos axiales en su cara superior. En
la misma figura se representa el desplazamiento axial de ese punto en la simulacion de os-
teointegracion completa, para su comparacion. Se observa que en el caso de osteointegracion
parcial tambien aumenta con el tiempo el desplazamiento en valor absoluto, es decir, que el
implante se hunde cada vez mas en el hueso circundante; pero sensiblemente menos que en
el caso de osteointegracion completa, ya que en este ultimo caso el dano acumulado por el
hueso periimplantario es mayor, como ha quedado patente en las simulaciones. Sin embargo,
al inicio de la simulacion de osteointegracion parcial, el hundimiento del implante es mayor;
algo logico, dado que la sujecion inicial del implante es menor cuando no hay osteointe-
gracion a la altura del cuello, mientras que con osteointegracion completa, la union entre el
hueso y el implante limita el hundimiento de este ultimo, al menos mientras el hueso de la
interfase mantiene su rigidez. Cuando pasa el tiempo esa rigidez se pierde y el hundimiento
es cada vez mayor, superando ampliamente al que tiene lugar con osteointegracion parcial.
Discusion de los resultados
En general, el hueso de la zona crestal acumula menos dano microestructural y sufre
menos reabsorcion, es decir, tiene mayor rigidez, en el caso de osteointegracion parcial y ello
trae diversas consecuencias. Por ejemplo, que la region medular sufra mas reabsorcion que
con osteointegracion completa. La mayor rigidez del hueso crestal le permite absorber mas
energıa de deformacion durante la masticacion, quedando la zona medular mas descargada,
con la consiguiente reabsorcion por desuso. Por el contrario, con osteointegracion completa,
el dano que acumula la zona crestal y la reabsorcion asociada a ese dano, hacen que la
rigidez de esta zona decaiga mucho, disminuyendo la capacidad de absorber energıa de
deformacion, que tiene que repartirse a otras regiones, entre ellas la region medular. Esta
region debe soportar ahora mas carga de la que soportarıa si la osteointegracion fuera parcial,
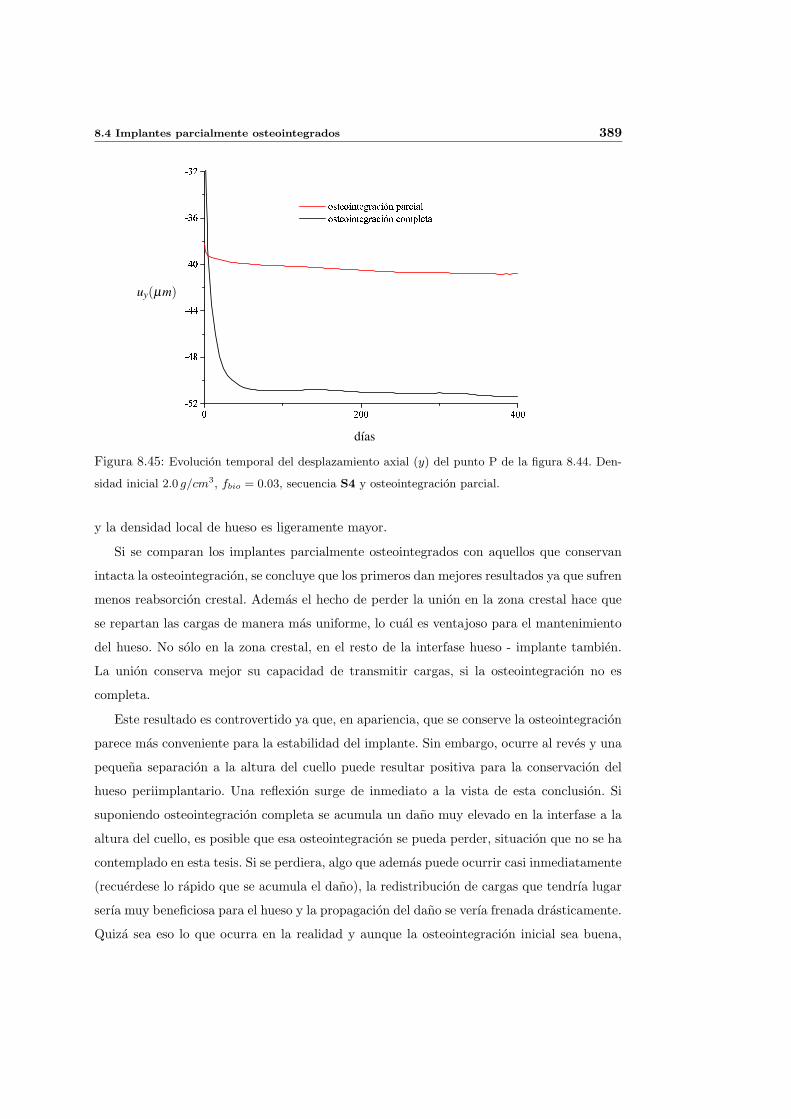
8.4 Implantes parcialmente osteointegrados 389
¢ £¢¢ ¤¢¢¥ ¦ £
¥ ¤§
¥ ¤¤
¥ ¤¢
¥ ¨©
¥ ¨ £
ª« ¬ ª® ¯¬ °± ²³® ´¯¶µ_²·± ³® ²¸ª« ¬ ª® ¯¬ °± ²³® ´¯¹³ªº¶µ_¸ ¬ ²
dıas
uy(µm)
1
Figura 8.45: Evolucion temporal del desplazamiento axial (y) del punto P de la figura 8.44. Den-
sidad inicial 2.0 g/cm3, fbio = 0.03, secuencia S4 y osteointegracion parcial.
y la densidad local de hueso es ligeramente mayor.
Si se comparan los implantes parcialmente osteointegrados con aquellos que conservan
intacta la osteointegracion, se concluye que los primeros dan mejores resultados ya que sufren
menos reabsorcion crestal. Ademas el hecho de perder la union en la zona crestal hace que
se repartan las cargas de manera mas uniforme, lo cual es ventajoso para el mantenimiento
del hueso. No solo en la zona crestal, en el resto de la interfase hueso - implante tambien.
La union conserva mejor su capacidad de transmitir cargas, si la osteointegracion no es
completa.
Este resultado es controvertido ya que, en apariencia, que se conserve la osteointegracion
parece mas conveniente para la estabilidad del implante. Sin embargo, ocurre al reves y una
pequena separacion a la altura del cuello puede resultar positiva para la conservacion del
hueso periimplantario. Una reflexion surge de inmediato a la vista de esta conclusion. Si
suponiendo osteointegracion completa se acumula un dano muy elevado en la interfase a la
altura del cuello, es posible que esa osteointegracion se pueda perder, situacion que no se ha
contemplado en esta tesis. Si se perdiera, algo que ademas puede ocurrir casi inmediatamente
(recuerdese lo rapido que se acumula el dano), la redistribucion de cargas que tendrıa lugar
serıa muy beneficiosa para el hueso y la propagacion del dano se verıa frenada drasticamente.
Quiza sea eso lo que ocurra en la realidad y aunque la osteointegracion inicial sea buena,

390 Analisis de implantes dentales
propiciada por un acabado superficial rugoso del implante y un perıodo de curacion efectivo,
en cuanto entre en carga, es posible que parte de esa union se rompa, mejorando ası la
transferencia de cargas y con ello la estabilidad del implante. No se puede comprobar si esto
ocurre efectivamente ası, sin considerar en la simulacion numerica la posibilidad de que la
osteointegracion del implante se pierda, cosa que no se ha estudiado en esta tesis y que serıa
conveniente estudiar en un futuro.
8.5. Resumen de los resultados
Hay que advertir en primer lugar, que los resultados deben ser analizados cualitativa-
mente. En ese sentido sı pueden aportar cierta luz sobre lo que le ocurre al hueso periim-
plantario cuando se coloca una protesis dental implantosoportada en la mandıbula. Pero de
ahı a que se pueda confiar en los resultados numericos obtenidos en estas simulaciones hay
un gran trecho, ya que son muchos los factores que pueden influir en ellos y que no se han
tenido en cuenta en esta tesis.
Aunque el objetivo perseguido en esta tesis es simular la reabsorcion crestal alrededor
del implante, es interesante observar lo que ocurre con la distribucion de porosidad lejos del
implante. La tendencia mostrada por las simulaciones es la de reabsorber hueso en la region
medular, donde el estımulo mecanico es pequeno, y mantener una densidad osea alta en la
region cortical, que soporta un estımulo mecanico elevado.
Se ha comprobado en las simulaciones que la porosidad inicial supuesta para el hueso
periimplantario influye de forma importante en los resultados, por ello, la eleccion de un
valor aproximado a la realidad es de suma importancia y el hecho de suponer uno arbitrario
es una de las principales limitaciones de este trabajo. En primer lugar, si se quiere que la
distribucion de densidad sea parecida a la realidad, no se puede suponer un valor uniforme
para el tejido que rodea al implante; es necesario conocer la distribucion de densidad osea, al
menos de forma aproximada, bien mediante tomografıas, bien de forma numerica, simulando
los procesos biologicos regenerativos que tienen lugar hasta que el implante entra en carga,
como por ejemplo, el crecimiento y maduracion del callo oseo. De todas formas, hay que
incidir en el hecho de que solo es necesario conocerla de forma aproximada, puesto que esa
distribucion de densidad se va a ir modificando durante la propia simulacion, acercandose a
la distribucion real, si el valor inicial no esta muy alejado de la realidad.
La secuencia de masticacion tambien tiene una gran influencia en los resultados como se

8.5 Resumen de los resultados 391
ha podido comprobar. La conclusion que se puede sacar sobre este aspecto es que el hecho
de no masticar con el lado del implante no asegura que la interfase hueso - implante quede
sin danar, ya que las masticaciones con el lado contrario tambien hacen que se deforme la
mandıbula y el hueso periimplantario. Tambien es cierto que el dano acumulado es menor y
eso podrıa justificar en parte la prescripcion de un perıodo de curacion tras la colocacion del
implante. Sin embargo, se ha comprobado aquı que un perıodo de curacion demasiado largo
hasta la colocacion de la protesis no hace sino empeorar la calidad del hueso cercano, que
pierde masa osea por desuso. Por esta razon se vienen desarrollando ultimamente tecnicas
de implantacion con carga inmediata, en la que el proceso quirurgico se hace coincidir con
el protesico, en la misma sesion, comprobandose que el porcentaje de exito esta tecnica es
similar al que se tiene con protocolos de dos fases [62,193].
La edad del paciente tambien tiene cierta influencia en la posibilidad de fallo del implante.
Dado que en jovenes la actividad remodelatoria es mas intensa, los efectos que se observan en
el hueso periimplantario se ven magnificados y entre ellos la reabsorcion crestal y la perdida
de rigidez de la union. Aun ası las diferencias no son muy importantes y ademas, el nivel de
dano es menor como consecuencia de esa mayor actividad remodelatoria. En la posibilidad
de exito de la implantacion en ancianos influyen muchos otros factores de tipo biologico y
metabolico que no se han tenido en cuenta aquı y por tanto extraer una conclusion definitiva
es algo osado.
Quiza el resultado mas llamativo sea el obtenido en la ultima simulacion: que una perdida
moderada de osteointegracion a nivel crestal puede ayudar a mejorar la estabilidad del im-
plante. Como se ha visto, a partir de que se pierda parte de la union en el cuello del implante,
las tensiones se distribuyen de manera mas uniforme, el dano progresa mas lentamente y la
densidad del hueso periimplantario se mantiene en un nivel bastante alto. Esto querrıa decir
que la perdida moderada de osteointegracion es un mecanismo de proteccion para el propio
implante. De todos modos, esta conclusion hay que tomarla con cautela dado que existen
muchos aspectos que no han sido analizados aquı y entre ellos, uno de los mas importantes,
es precisamente la perdida de osteointegracion. Este proceso, que se ha simplificado enorme-
mente, se produce de forma progresiva y compleja, involucrando muchos procesos biologicos
y mecanicos que es necesario estudiar con mas detalle, porque su influencia en los resultados
puede ser fundamental.


Capıtulo 9
Conclusiones y desarrollos
futuros
9.1. Resumen del trabajo realizado
El objetivo principal de esta tesis ha sido ahondar en el conocimiento del proceso de
adaptacion del hueso de la mandıbula en su estado sano y cuando se coloca una protesis
dental implantosoportada.
Aparte de este objetivo general, en esta tesis se han conseguido tres objetivos particulares.
En primer lugar, se ha construido un modelo de elementos finitos de una mandıbula humana,
sobre el que se ha aplicado un modelo de remodelacion osea interno desarrollado previamente
y que ha permitido estimar la distribucion de densidad osea y propiedades elasticas del hueso
en toda la mandıbula, en funcion de la carga de masticacion. Con ello se ha conseguido
completar el modelo de elementos finitos, en el que no se conocıan a priori las propiedades
elasticas. Este modelo se puede utilizar para otros propositos, como por ejemplo, el analisis
de implantes dentales, que ha sido objeto del capıtulo 8 de esta tesis. Los parametros del
modelo de remodelacion osea utilizado en esta primera parte han sido adaptados para que
puedan reproducir el comportamiento del hueso maxilar, ya que el modelo fue aplicado
originalmente a la extremidad proximal del femur y los valores de sus parametros estaban
ajustados al comportamiento de ese hueso. Los resultados de densidad obtenidos con las
simulaciones se han podido comparar directamente con los datos extraıdos de la propia
393

394 Conclusiones y desarrollos futuros
mandıbula humana, que se midio para obtener el modelo de elementos finitos. De ella se
tomaron una serie de tomografıas que sirvieron de patron para los resultados numericos,
consiguiendo finalmente una buena concordancia entre ambos resultados.
En segundo lugar se ha desarrollado un nuevo modelo de remodelacion osea interna
anisotropo, basado en la actividad de las BMUs. Este modelo mecanobiologico relaciona
directamente el estado de carga mecanica soportado por el tejido con los procesos biologicos
involucrados en la remodelacion osea. Considera las variaciones locales del tamano y forma
de los poros, aunque tratados desde un punto de vista macroscopico, pero que permite
establecer las propiedades elasticas anisotropas del tejido oseo cortical y trabecular, que
se tratan de forma separada. El tiempo es una variable de este modelo, que se incluye
en la formulacion para controlar la velocidad a la que se producen los diferentes procesos
biologicos. Esto permite seguir los cambios que sufre el hueso en tiempo real. Entre estos
cambios se ha simulado una situacion de desuso que provoca reabsorcion; una sobrecarga
moderada, con la consiguiente formacion; una sobrecarga alta, que produce fractura; un
cambio en la magnitud de las cargas, distinta para cada direccion, que provoca cambios
en la anisotropıa del entramado oseo; finalmente, se ha simulado un giro en las direcciones
principales de deformacion, que provoca una reorientacion de los poros, para alinearse con
las cargas.
Por ultimo, se ha aplicado el nuevo modelo de remodelacion osea al estudio de un implante
osteointegrado en la mandıbula, a la altura del primer molar derecho. De esta forma se ha
podido analizar la alteracion que sufre el tejido periimplantario como consecuencia de los
cambios en la forma de transferir las cargas de masticacion. Se han comparado diversos
parametros, como la edad, los habitos masticatorios, la porosidad del hueso previa a la
implantacion y el acabado superficial de la protesis, que influye en el tipo de union que se
establece entre implante y hueso circundante.
9.2. Conclusiones
De caracter general
La Bioingenierıa en general y la Biomecanica en particular, son disciplinas con un futuro
prometedor. Buena prueba de ello son la cantidad de grupos de investigacion que en los
ultimos tiempos le estan prestando su atencion. Uno de los atractivos de esta ciencia reside

9.2 Conclusiones 395
en el estado puber, si se me permite la expresion, en el que se encuentra, lo que hace que
se abran nuevas lıneas de investigacion continuamente, pues aun es largo el camino por
recorrer. Largo y tortuoso, que duda cabe, pues el objetivo final es bien ambicioso: intentar
comprender, y aun mas, reproducir, los fenomenos que gobiernan la vida de la criatura
mas compleja de la Creacion (o mas avanzada de la Evolucion), el Hombre. Una cita de
Alarcon resume de manera precisa esta idea: “La intervencion en el cuerpo humano [. . . ] es
impresionante. El cuerpo humano es la maquina definitiva. Ahora cuando hay un problema
de corazon se dice que se hace fontanerıa, pues imagınese lo que puede hacer la ingenierıa.
Creo que el cuerpo humano es el desafıo total para los ingenieros” [7].
Algunos grupos de investigacion estan reconduciendo sus pasos hacia este nuevo campo
de la ciencia y otros estan aprovechando su experiencia en otros campos de la Ingenierıa
que son de mucha utilidad. Entre ellos, uno que considero fundamental es el de los Metodos
Numericos, que con el desarrollo de la informatica se ha convertido de manera natural en
pieza clave para cualquier estudio avanzado en el campo de la Bioingenierıa. En Biomecanica,
el Metodo de los Elementos Finitos es cada vez mas utilizado para simular procesos biologicos
en los que la Mecanica juega un papel relevante [273]. Y no solo la Mecanica Clasica o
la Teorıa de la Elasticidad, ya que al descender a nivel celular el acoplamiento con otras
disciplinas como la Mecanica de Fluidos, el Electromagnetismo, la Bioquımica, etc. surge
inevitablemente. Lo anterior pone de manifiesto la necesidad de colaborar con otros grupos
de investigacion, que requiere la dedicacion a este campo. Atras deben quedar los modelos
fenomenologicos, de los que se ha hablado en esta tesis, que pretenden simular procesos
biologicos desde un punto de vista estrictamente mecanico. Se ha demostrado la necesidad
de iluminar esa caja negra que representa un modelo fenomenologico, bajar al nivel celular y
tener en cuenta tambien los procesos biologicos que tienen lugar, y sobre todo, relacionarlos
con la Mecanica, pues ambas disciplinas estan necesariamente relacionadas. Solo ası se puede
alcanzar una comprension exhaustiva de los procesos que se quieren simular.
Sobre la simulacion del proceso de masticacion
La distribucion de densidad de una mandıbula humana ha sido obtenida utilizando un
modelo de elementos finitos y simulando con el el proceso de masticacion. De paso se ha
obtenido la distribucion de propiedades elasticas considerando el hueso como un material
anisotropo. Todo ello partiendo de una situacion inicial tan irreal como lo es una distribucion

396 Conclusiones y desarrollos futuros
uniforme de densidad 0.5 g/cm3 e isotropa. Este hecho parece confirmar que la distribucion
de densidad y de propiedades elasticas esta ıntimamente relacionada con la actividad diaria
y que para resistir las cargas normales de uso, la disposicion optima del material, aquella en
la que se aprovecha mejor, es decir, resiste las cargas con el mınimo peso, coincide con la dis-
tribucion real. Se han comparado los resultados numericos de densidad osea con tomografıas
extraıdas de la propia mandıbula, siendo la concordancia entre ambos resultados bastante
buena. Tambien se han comparado los resultados de propiedades elasticas y la similitud tam-
bien es aceptable, a pesar de que los estudios experimentales tomados como patron tienen
validez limitada, bien por el metodo de medida empleado, bien porque proporcionan datos
solo para una region de la mandıbula. Por otro lado, por falta de medios tecnicos no ha
sido posible medir las propiedades elasticas de la mandıbula en cuestion. Este hubiera sido
sin duda el mejor patron de comparacion, ya que estas propiedades elasticas pueden variar
enormemente de un individuo a otro y dependen entre otras cosas de las dimensiones de la
propia mandıbula, que determinan las cargas que esta soporta.
Se han hecho algunas simplificaciones en el modelo como, por ejemplo, la geometrıa
de los dientes, que no fue posible medir en la mandıbula real por faltar gran cantidad de
ellos. A los alveolos no se pudo acceder con la maquina de medicion por coordenadas y su
geometrıa tambien es aproximada. El ligamento periodontal se ha simulado de manera muy
simplificada, como un material isotropo lineal elastico, con una rigidez muy pequena. Como
se ve todas las simplificaciones estan hechas en el entorno de los dientes y precisamente es
en esta zona donde se encuentran las principales diferencias entre los resultados numericos
y las tomografıas. Se hace necesario, por tanto, simular con mas detalle esta zona y sobre
todo el comportamiento del ligamento, si se quiere conseguir una distribucion de densidad
osea mas aproximada en esta region.
El modelo de remodelacion osea basado en la Mecanica del Dano desarrollado por Garcıa
y Doblare [91,115] y otros modelos de remodelacion osea interna han sido tradicionalmente
aplicados al femur y otros huesos largos. Esto es ası porque estos modelos establecen una
relacion directa entre cargas y remodelacion, y los huesos largos estan sometidos a cargas
elevadas, en contraposicion a los huesos planos que tienen una funcion protectora, como el
craneo, y apenas soportan carga. En huesos planos la relacion entre distribucion de masa
osea y estımulo mecanico debe diferir de la que existe en los huesos largos y dicha relacion no
ha sido suficientemente investigada todavıa. En esta tesis el modelo de remodelacion se ha

9.2 Conclusiones 397
aplicado al maxilar inferior, que por geometrıa no se puede clasificar como un hueso largo,
pero que por comportamiento se ha demostrado que sı lo es, o al menos tiene el mismo tipo
de relacion carga - remodelacion que los huesos largos. Si se quiere, la mandıbula se podrıa
ver como un hueso largo de directriz curva.
El estımulo mecanico al que esta sometido la mandıbula es menor que el que sufre
el femur. No solo son menores las cargas sino que tambien es menor el numero de ciclos
en uno y otro caso. La respuesta remodelatoria depende del estımulo mecanico y de las
diferencias entre este estımulo y un valor de referencia, que por pura logica debe ser distinto
para mandıbula y femur, para que la respuesta remodelatoria sea acorde con el estımulo
soportado. Se han probado tres valores distintos del estımulo mecanico de referencia en la
mandıbula, siendo aquel que proporciona mejores resultados igual a la mitad del valor usado
por Garcıa y Doblare en el femur [91,115].
Para obtener con suficiente aproximacion la distribucion de densidad osea es primordial
ajustar el estımulo mecanico de referencia, pero en las propiedades elasticas el parametro que
mayor influencia tiene es el factor de anisotropıa, que pondera la influencia de la componente
desviadora del estımulo en la remodelacion. Con este parametro tambien ha sido necesario
un reajuste con respecto al valor usado en el femur por Garcıa y Doblare [91, 115]. En el
caso de la mandıbula es necesario usar un valor mas alto del factor de anisotropıa, dado
que la componente desviadora de las cargas, y por tanto del estımulo, es mas importante
que en el femur. Hay que tener en cuenta que a la flexion que sufre la mandıbula por la
accion muscular y el apoyo en los dientes hay que anadirle una torsion importante cuando
la masticacion es unilateral.
La secuencia de masticacion tambien tiene una notable influencia en los resultados, siendo
de enorme importancia masticar con ambos lados para mantener una buena calidad de hueso.
En el caso de masticacion unilateral, el lado con el que no se mastica recibe un estımulo
mecanico menor que hace que la densidad de hueso sea muy baja localmente.
Sobre el modelo de remodelacion propuesto
El nuevo modelo de remodelacion propuesto en esta tesis es de tipo mecanobiologico, en
cuanto que tiene en cuenta muchos procesos biologicos que intervienen en la remodelacion
osea interna. Estos procesos son:
1) Activacion de BMUs. Aunque aun no se conoce a ciencia cierta como ocurre, los os-

398 Conclusiones y desarrollos futuros
teocitos son capaces de sentir de alguna manera la deformacion cıclica que sufre la estructura
osea como consecuencia de la actividad diaria. Esta deformacion parece transmitirse de un
osteocito a otro como una senal, que finalmente llega a las celulas de borde que ocupan la
superficie libre del hueso. Si esa senal se interrumpe por algun motivo, las celulas de bor-
de, que dejan de sentirla, segregan sustancias que activan el reclutamiento de osteoclastos,
comenzando de esta forma la actividad de las BMUs. Esa senal se puede interrumpir, bi-
en porque no hay deformacion, bien porque la carga ha hecho crecer grietas que rompen
la conexion entre osteocitos. Ambos casos son contemplados con este modelo, que evalua
la frecuencia de activacion en funcion del estımulo y el nivel de dano. Tambien influye la
superficie disponible para la activacion, que es funcion de la porosidad. Otros factores de
tipo metabolico, patologico, hormonal, etc., pueden influir en la activacion de BMUs, pero
no han sido estudiados en detalle. Se engloban en un unico factor, fbio, que tan solo se ha
modificado para tener en cuenta la edad, factor que se sabe que influye haciendo menos
numerosa la poblacion de BMUs [107].
2) Avance de la BMU. Una vez activados, los primeros osteoclastos comienzan a avanzar
en una determinada direccion, que se ha supuesto coincidente con las direcciones principales
de deformacion. De esta forma las BMUs pueden cumplir las dos misiones principales que se
le han supuesto en este modelo: reabsorber hueso si no esta siendo util y no se deforma en una
determinada direccion, o repararlo si las deformaciones en esa direccion lo han danado en
exceso (las tensiones de traccion producen grietas y las BMUs van “cosiendo” esas grietas).
La forma de avance de las BMUs es distinta en el tejido cortical, en el que pueden atravesar
la matriz osea, y en el tejido trabecular donde avanzan siempre sobre la superficie de hueso.
La orientacion de los poros limita por tanto la accion de las BMUs sobre el hueso trabecular
y la cantidad de de BMUs que se pueden activar avanzando en cada direccion. Esta diferencia
tambien ha sido considerada en el nuevo modelo.
3) Fases de la secuencia ARF. Por cada lugar por el que avanza hasta la apoptosis, la BMU
pasa por tres fases: reabsorcion, inversion y formacion. Todas ellas se simulan en el modelo.
La cantidad de hueso eliminado en la fase de reabsorcion se supone que es siempre la misma,
pero en el proceso de diferenciacion de osteoblastos interviene de manera fundamental la
energıa de deformacion. Ası, cuanto mayor sea esta, mayor es el numero de osteoblastos que
se diferencian de sus precursores y por tanto, mayor la cantidad de osteoide formado. Este
efecto se ha tenido en cuenta en el modelo mediante el balance local direccional de hueso, que

9.2 Conclusiones 399
da el volumen de osteoide depositado, en relacion al reabsorbido, en funcion de la energıa
de deformacion local asociada a la direccion de avance de la BMU.
4) Proceso de mineralizacion. El osteoide recien formado esta compuesto de colageno
y agua, pero no tiene mineral alguno. Este se deposita en la matriz osea en un proceso
que comienza relativamente rapido y posteriormente se ralentiza conforme la matriz osea
se satura de mineral. Este proceso de mineralizacion tambien se ha simulado a lo largo del
tiempo, distinguiendo dos perıodos: uno de mineralizacion primaria, casi instantaneo, y otro
de mineralizacion secundaria, en el que la velocidad de deposicion de mineral decae de forma
exponencial.
5) Proceso de acomodacion celular. Otro fenomeno que se observa en la respuesta re-
modelatoria a un estımulo externo es que si dicho estımulo se mantiene constante de manera
prolongada, el sistema acaba por acomodarse a dicho estımulo de forma que cada vez re-
sponde al estımulo con menor intensidad. Ası, perıodos prolongados de desuso no producen
una perdida total de masa osea, como se puede comprobar en astronautas o en personas
minusvalidas. Este proceso de acomodacion celular se ha introducido en el modelo haciendo
que el estımulo mecanico de referencia tienda hacia el estımulo mecanico con una velocidad
proporcional a la diferencia entre ambos.
6) Acumulacion de dano microestructural. Junto a los fenomenos biologicos anteriores,
tambien se ha simulado el proceso mecanico de acumulacion de dano por deformacion.
El dano en este modelo tiene dos interpretaciones. Por un lado se entiende como la frac-
cion volumetrica de tejido agrietado y por otro como una medida de la degradacion de las
propiedades elasticas de material. De esta forma es como lo entienden Pattin et al. [265]
en sus ensayos de fatiga en hueso cortical. Estos ensayos han sido ajustados mediante ecua-
ciones para dar una ley de crecimiento del dano que usar en el modelo numerico. Pero estas
mismas ecuaciones se han usado tambien en hueso trabecular, a pesar de que no se ha podido
comprobar experimentalmente la validez de esta generalizacion.
Las propiedades elasticas del hueso dependen de muchos factores, algunos de los cuales
se han tenido en cuenta en el modelo propuesto. El proceso de remodelacion modifica las
dimensiones de los poros en cada direccion, pudiendose expresar el promedio local de es-
tas dimensiones de poro mediante un tensor, que ha sido relacionado con las propiedades
elasticas mediante diversos estudios experimentales. Una relacion, del mismo tipo que las
obtenidas experimentalmente, se utiliza en el modelo propuesto aquı. En el tensor de com-

400 Conclusiones y desarrollos futuros
portamiento influye no solo la forma de los poros, sino tambien su volumen, a traves de la
porosidad y el contenido de mineral de la matriz osea. La relacion usada en ambos casos se
ha encontrado en la literatura, concretamente de los estudios experimentales de Hernandez
et al. [145]. Por ultimo, se incluye la influencia del dano de acuerdo con la teorıa de dano
anisotropo de Cordebois y Sideroff [69].
Hasta aquı se ha expuesto un pequeno resumen de los aspectos biologicos y mecanicos
contemplados y relacionados por este modelo, que en buena logica se ha denominado mecanobi-
ologico. A continuacion se resumen algunas de sus caracterısticas, comprobadas con su uso
en las diversas simulaciones llevadas a cabo.
a) El tiempo aparece explıcitamente en la formulacion del modelo y ello permite estimar
la velocidad a la que se produce la adaptacion del tejido oseo frente a un cambio en la
situacion de carga.
b) En relacion al tiempo, y en concreto a la duracion de los perıodos de reabsorcion,
inversion y formacion de la secuencia ARF, llama la atencion la disparidad de valores en-
contrados en la literatura. Garcıa-Aznar et al. [117] usaron, en el modelo isotropo que sirve
de base al desarrollado aquı, unos valores muy particulares, obtenidos en estudios histomor-
fometricos de mujeres postmenopausicas con osteoporosis. Segun estos estudios las fases de
la secuencia ARF ocurren muy lentamente, incluso las BMUs avanzan mas lentamente, en
estas mujeres que en personas sanas sin trastornos hormonales. La reabsorcion por desuso
que predice el modelo con los perıodos mas largos y el avance mas lento es menor, lo que
podrıa prevenir el riesgo de fractura en estas mujeres. En el mismo sentido ayuda el hecho
de que con la edad disminuya la frecuencia de activacion de BMUs, fbio.
Por otro lado, puede que los perıodos mas largos de la secuencia ARF sean solo debidos
a la edad y no a trastornos hormonales. A esa conclusion es a la que llega Frost con el
perıodo de formacion [107], conclusion que se puede extender a los otros perıodos, porque
es de suponer que la duracion de todos ellos varıe en consonancia.
c) Este modelo presenta una diferencia fundamental con la mayorıa de modelos fenomenologi-
cos anteriores. Estos predicen, en resumidas cuentas, aunque sea una forma excesivamente
simplificada de decirlo, que cargas altas producen formacion neta mientras que cargas bajas
producen reabsorcion neta. Sin embargo hay casos en los que una sobrecarga alta continuada
puede producir reabsorcion y conducir finalmente a una fractura por fatiga. Este fenomeno
se ha podido simular con este modelo, en el que se incluye el dano que una sobrecarga puede

9.2 Conclusiones 401
producir, con la consecuente reabsorcion si ese dano es elevado. Por tanto, hay concordancia
con modelos previos en que el desuso produce reabsorcion, pero tambien una sobrecarga
prolongada puede producir un efecto similar, como consecuencia de la acumulacion de dano.
d) La fractura del hueso por fatiga es un fenomeno que a veces ocurre en deportistas
sometidos a sobrecarga de entrenamiento. Se han simulado ensayos de fatiga en traccion
y en compresion uniaxial, obteniendose resultados muy dispares en uno y otro caso. La
resistencia a traccion es mucho menor que a compresion, ya que, por hipotesis, el dano crece
en la direccion de maxima deformacion principal y solo si esa deformacion es de traccion.
Ası, en un caso de compresion uniaxial, el modelo supone que el dano crece unicamente por
efecto Poisson y, por tanto, mucho mas lentamente.
e) Adaptacion del estımulo de equilibrio. El proceso de acomodacion celular descrito
mas arriba, tambien se conoce como teorıa del mecanostato [109,329], por analogıa entre el
comportamiento del hueso y el de un termostato, que controle, por ejemplo, la temperatura
de un recinto, haciendola converger exponencialmente hacia la temperatura de control. Ante
cualquier cambio de la temperatura de control, el termostato actua en consecuencia, con
una nueva adaptacion exponencial de la temperatura.1 Esta situacion llevada al caso del
hueso implica que cualquier cambio en las condiciones de carga tiene que verse reflejada en
una nueva adaptacion del estımulo de equilibrio. La adaptacion es tal que la variacion del
estımulo de equilibrio es proporcional a la diferencia entre el estımulo de equilibrio actual y
el estımulo externo aplicado.
f) El modelo de remodelacion propuesto en esta tesis esta basado en el modelo mecanobi-
ologico basado en la actividad de BMUs, desarrollado por Garcıa-Aznar et al. [117], que
proporciona una evolucion isotropa de las propiedades mecanicas del hueso. De hecho este
modelo es una extension al caso anisotropo. En los casos de carga hidrostatica que se han
simulado la respuesta obtenida es similar a la del modelo isotropo, como es logico.
g) Tambien se han simulado estados de carga no hidrostaticos para estimar las variaciones
de la anisotropıa del tejido oseo en tiempo real. Se ha comprobado que la direccion de
ortotropıa que adquiere mayor rigidez es aquella que soporta mayor energıa de deformacion.
Esa es la forma en la que mejor se aprovecha el material y por tanto la disposicion resistente
de masa mınima.
h) Asimismo, se ha comprobado que ante un cambio en la orientacion de la carga, los
1La respuesta es exponencial en cuanto que el tiempo aparece en las ecuaciones como exponente, siempre
que se mantenga constante la temperatura de control.

402 Conclusiones y desarrollos futuros
poros tienden a alinearse con las direcciones principales de tension, como predice la ley
de Wolff [354] y se viene investigando insistentemente desde entonces. Sin embargo, esta
alineacion no se consigue completar en un caso general, siendo mas difıcil cuanto mayor es el
grado de anisotropıa. Esto es logico, porque para cambiar tanto la anisotropıa es necesario
una intensa actividad remodelatoria mantenida durante bastante tiempo y eso no siempre
es posible, ya que mientras la estructura se esta reorientando, el estımulo de equilibrio se
adapta al nuevo estımulo externo, en magnitud y en direccion, y el balance local direccional
de hueso puede entrar entonces en zona muerta, frenandose la adaptacion. A partir de ese
momento ya no hay cambios apreciables en el tensor MIL, si no se produce una activacion
intensa de BMUs.
Lo que sugiere este resultado de que los poros no se alineen por completo con la carga,
es que en el modelo propuesto puede existir una zona muerta para la reorientacion de la
estructura, al igual que existe una zona muerta de remodelacion para el estımulo mecanico.
En esta ultima no se produce ni formacion ni reabsorcion neta y cuando el estımulo entra en
ella se frena el cambio de porosidad. De forma parecida, cuando se entra en la zona muerta
de reorientacion, los poros dejan de girar en su intento por alinearse con la carga y por tanto
se alcanza un equilibrio direccional.
La falta de datos ha impedido validar estos resultados, por lo que resulta muy conve-
niente seguir investigando en este sentido, para comprobar si efectivamente existe esta zona
muerta de reorientacion o por el contrario se consigue la alineacion total entre la carga y
la estructura. Al menos hasta donde llega el conocimiento del autor, no se han realizado
estudios in vivo que cuantifiquen el giro de la estructura osea ante un giro de la carga y
serıa fundamental hacerlo para validar los resultados obtenidos aquı.
En cualquier caso, conviene recordar la controversia que ha promovido siempre la ley de
Wolff, que es precisamente la que establece la alineacion de las trabeculas con las direcciones
principales de tension. Se ha comprobado en algunos huesos, por ejemplo el femur, que dicha
ley no es del todo cierta. No lo puede ser porque el hueso soporta, en general, diferentes
estados de carga, cada uno con unas direcciones principales de tension diferentes y por tanto
las trabeculas no se pueden alinear con todas a la vez. Si acaso, se podrıan alinear con
las direcciones principales de un tensor que fuera combinacion de los tensores de tension
correspondiente a las cargas mas habituales. Algo parecido es lo que se hace aquı cuando se
establece como hipotesis que las BMUs se activan en las direcciones principales del tensor

9.2 Conclusiones 403
de estımulo, tensor que es una combinacion lineal de los diferentes estımulos.
Sobre la simulacion del modelo implantado
En cuanto a la aplicacion del nuevo modelo de remodelacion osea a la simulacion de
implantes dentales osteointegrados, se pueden sacar las siguientes conclusiones:
a) El modelo propuesto en esta tesis permite simular los procesos biologicos en tiempo
real y en el caso de sobrecargas altas, como las que ocurren en el cuello del implante,
predice reabsorcion osea por acumulacion de dano, no formacion como otros modelos de
remodelacion anteriores. Esto hace que sea muy util para el estudio de la viabilidad a largo
plazo de un implante dental, ya que permite simular la evolucion temporal de una posible
perdida osea a nivel crestal, que es uno de los problemas mas graves que pueden surgir en
este tipo de implantes.
b) La implementacion de un modelo de dano con elementos finitos requiere de un especial
cuidado, ya que el dano sera especialmente alto en concentradores y en puntos singulares de
tension. A veces estos puntos aparecen por un mallado excesivamente grosero, pero otras ve-
ces son consecuencias inevitables de la discretizacion fısica del problema, como por ejemplo:
en cargas puntuales, en las regiones del modelo con desplazamientos impuestos, en las es-
quinas entrantes, en los bordes libres de la interfase entre dos materiales, etc. Cuando solo se
precisa del conocimiento de la solucion elastica se suelen descartar estas zonas, porque ahı la
solucion elastica numerica no es valida. Si la malla es adecuada, lo que ocurre localmente en
estos puntos singulares no afecta a la solucion global. Si se desea conocer la solucion local, se
puede refinar la malla, en el caso de concentradores, o usar elementos finitos singulares si se
trata de una singularidad. Sin embargo, cuando se analiza el dano hay un problema anadido
y es que el dano se puede propagar, afectando a elementos contiguos al punto singular, que
originalmente estaban lejos de la influencia de la singularidad. Esta propagacion se debe a
lo siguiente: la degradacion de las propiedades elasticas de un elemento danado hace que en
la union entre ese elemento y el contiguo haya una discontinuidad de las propiedades y esta
discontinuidad puede originar una nueva singularidad de tensiones, que dane al elemento
contiguo, hasta ahora intacto.
Esta propagacion del dano en el tejido oseo es un proceso que es necesario investigar mas
en profundidad y que conviene tener en cuenta al resolver problemas con singularidades de
tipo numerico, como cargas puntuales, regiones con desplazamientos impuestos, etc., en los

404 Conclusiones y desarrollos futuros
que no deberıa afectar a la solucion originando un dano espurio. Una solucion para estos
casos puede ser no considerar el dano en los elementos proximos a la singularidad. Tambien
se podrıa definir un criterio de propagacion no local de tipo energetico, que evite un eventual
crecimiento del dano solo porque haya unas tensiones altas muy localizadas.
c) La propagacion del dano se observa en los resultados del implante, que en la interfase
con el hueso acumula un dano muy elevado que se inicia en el borde libre (en el cuello del
implante) y progresa hacia el apice sin freno aparente. Este dano ademas produce reabsorcion
osea y da como resultado final una capa de tejido de gran flexibilidad rodeando al implante.
Este resultado se corresponde con un deterioro notable de la osteointegracion y podrıa estar
simulando lo que ocurre en la realidad: que existe una capa de tejido fibroso rodeando el
implante. Una vez que el implante entre en carga, es imposible que el tejido adyacente al
implante quede intacto si la union entre implante y hueso se produce de manera directa, por
la diferencia de modulos elasticos de uno y otro material.1 Con todo, el espesor de la capa
de tejido fibroso es bastante menor que la de tejido blando obtenida numericamente.
En los resultados numericos, el espesor de la capa danada practicamente coincide con el
espesor de los elementos de la interfase, ya que los elementos adyacentes apenas acumulan
dano. Esto es debido a que, hasta cierto punto, el espesor de la zona danada depende del
tamano de la malla. En efecto, el dano se localiza en torno a la singularidad. De esta forma,
si el elemento de la interfase engloba a la region afectada por dicha singularidad, todo el
aparecera muy danado, aunque esa singularidad este mucho mas localizada. Si se refina la
malla, el elemento de la interfase sigue estando afectado por la singularidad, es decir, muy
danado, pero los elementos adyacentes tienen un nivel de tensiones y de dano inferior. Ası,
refinando localmente la malla se podrıa ajustar con mas precision el espesor de esta capa
danada, que simula el tejido fibroso que recubre el implante.
d) De todas maneras, el proceso de osteointegracion es mucho mas complejo y requiere
un estudio mas detallado. No solo habrıa que mejorar la consideracion del dano en esta zona,
sino tambien como las tensiones y las propiedades elasticas podrıan afectar al tipo de union
entre tejido e implante. Aquı se esta suponiendo que dicha union es perfecta o no existe,
pero no se permite un estado intermedio, ni siquiera pasar de uno a otro. El hecho de que
el dano se acumule en la interfase hace pensar que quiza sea conveniente usar un modelo de
degradacion de la union.
1Diferente es lo que ocurre durante el proceso de curacion, mientras se produce la osteointegracion, porque
el implante no se carga directamente y las tensiones son muy bajas.

9.2 Conclusiones 405
De hecho, se ha comprobado que si la osteointegracion se pierde en el cuello del implante,
mejoran sensiblemente las condiciones de carga de esta zona. Efectivamente, en el modelo
parcialmente osteointegrado, el hueso que rodea el cuello del implante mantiene una mejor
integridad: acumula menos dano y mantiene una densidad mayor. En definitiva tiene may-
or rigidez. Esto hace tambien que la movilidad del implante sea menor que en el caso de
osteointegracion completa. Este resultado sugiere que la situacion de osteointegracion com-
pleta puede no estar representando lo que ocurre en la realidad. Ası, la union perfecta entre
el hueso y el implante no serıa posible en toda la interfase, porque ello danarıa en exceso el
hueso adyacente y un hueso muy danado no podrıa soportar las tracciones que le transmite
el implante por la union. De esta manera, si la capa de hueso en contacto con el implante
se debilita, lo mas probable es que parte de la union se pierda y a partir de ese momento
se relajen la tensiones en toda la zona crestal. Esto no es mas que una hipotesis, que habrıa
que comprobar usando un modelo de interfaz para la osteointegracion.
e) Las propiedades iniciales del hueso periimplantario fueron supuestas. En concreto, se
supuso una densidad inicial uniforme y que el comportamiento es isotropo. Se hizo ası por
no disponer de datos al respecto. En esta region, tan poca justificacion tiene extrapolar los
valores obtenidos en la morfogenesis de la mandıbula, como suponer unos valores iniciales,
puesto que el relleno del hueso dejado por el diente, el proceso quirurgico y el perıodo de
curacion posterior van a modificar, sin duda, las propiedades de dicho hueso periimplan-
tario. Uno de los fenomenos que no se tiene en cuenta en este modelo es el crecimiento,
consolidacion y posterior reabsorcion del callo oseo, que puede tener tanta influencia en los
resultados finales como el hecho de suponer una distribucion u otra de densidad para el
hueso periimplantario.
Con todo, la influencia de la densidad inicial es importante y se ha comprobado que
cuanto menor es esta densidad, mayor es la reabsorcion y el dano acumulado en el hueso
periimplantario. Ademas, con un hueso soporte de baja calidad, el dano no solo se acumula
en el cuello del implante, sino que tambien es importante en el apice. Por ello, partir de
una densidad proxima a la real adquiere una gran importancia para conseguir resultados
fiables y conviene analizar con mucho detalle todos los procesos que puedan influir en la
distribucion de densidad osea previa a la entrada en carga del implante. Pero ademas, partir
de una densidad mas parecida a la real implica que sea mas baja que la aquı simulada,
y eso tiene un grave inconveniente y es que el modelo de dano utilizado fue ajustado a

406 Conclusiones y desarrollos futuros
curvas experimentales en hueso cortical, pero su validez no se ha comprobado para hueso
trabecular. De hecho, su utilizacion en huesos muy porosos produce un dano muy elevado
con poco sentido fisiologico.
f) La secuencia de masticacion tiene bastante influencia en los resultados. No solo in-
fluyen que cargas se aplican y con que frecuencia cada una, sino tambien el orden en que se
aplican, como ocurre en la realidad. El orden de las cargas no puede influir directamente en
la respuesta remodelatoria, puesto que el tiempo caracterıstico de esta respuesta es mucho
mayor que el tiempo caracterıstico de variacion de las cargas. Pero sı puede influir indirecta-
mente, ya que en determinadas condiciones el dano acumulado por la estructura osea sı que
puede depender del orden en que se aplican esas cargas. En cualquier caso, la diferencia
debe ser pequena, ya que el dano crece preferentemente con una de las cargas, precisamente
cuando se mastica con la pieza implantada, siendo muy pequeno el aumento del dano debido
a las otras cargas.
Por otro lado, depende del patron de masticacion y esto sı que se ha comprobado. Junto a
la masticacion unilateral alternante, se ha simulado un caso de masticacion unilateral, en la
que no se mastica con el lado donde esta el implante. Aunque no se cargue directamente ese
lado, las deformaciones que sufre la mandıbula por la masticacion con el otro lado, tambien
hacen que se acumule dano en el hueso periimplantario. Ademas, el desuso de este lado del
implante esta provocando una perdida osea generalizada, que puede poner en grave riesgo
la estabilidad del implante. Por tanto, el perıodo de curacion que se prescribe despues de
la operacion debe limitarse al tiempo estrictamente necesario para la consolidacion del callo
oseo. Nada se consigue alargando en exceso dicho perıodo, ya que no esta evitando que se
acumule dano y por el contrario induce la reabsorcion en el lado del implante.
g) La influencia de la edad se ha analizado a traves de uno de los parametros en los
que influye la edad: la frecuencia de activacion de BMUs, que disminuye con la edad. La
actividad remodelatoria es especialmente intensa en jovenes, en cuyo caso la reabsorcion
crestal es mas probable, pero tambien ocurre que el dano acumulado es ligeramente menor,
porque se repara mas rapidamente. Seguramente, hay muchos otros factores biologicos en
los que influye la edad y que no se han tenido en cuenta aquı. Uno de ellos, la duracion de
los perıodos de la secuencia ARF, se analizo en un caso de desuso, en el capıtulo 7, pero no
en la simulacion del implante. Con la edad estos perıodos son mas largos y, en general, la
actividad de las BMUs es mas lenta, lo que, por ejemplo, previene una excesiva reabsorcion

9.3 Aportaciones originales 407
por desuso. Esto es muy importante en personas de cierta edad, en las que el logico descenso
de la actividad diaria podrıa desembocar en una excesiva perdida de masa osea si la actividad
de las BMUs se mantuviese en los mismos parametros que tiene en personas jovenes.
9.3. Aportaciones originales
Las aportaciones originales mas importantes de esta tesis son las siguientes:
a) Aplicacion de un modelo de remodelacion osea interna a una mandıbula humana.
Tradicionalmente, estos modelos se han venido usando para obtener numericamente la dis-
tribucion de densidad osea en la extremidad proximal del femur. Hasta donde llega el
conocimiento de su autor, esta tesis es el primer trabajo en el que se usa para obtener
la distribucion de densidad de una mandıbula humana. Ademas, se obtiene la distribucion
de propiedades elasticas del hueso, supuesto como un material anisotropo.
b) Analisis de la influencia que tiene un patron de masticacion unilateral sobre la densidad
osea de la mandıbula. Comparacion con un patron de masticacion unilateral alternante.
c) Extension de un modelo de remodelacion osea interna isotropo desarrollado anterior-
mente, al caso anisotropo. El modelo resultante, al igual que el modelo en el que esta basado,
es de tipo mecanobiologico y esta basado en la actividad de las BMUs. Aplicacion de dicho
modelo a casos sencillos de carga: desuso, sobrecarga que produce rotura por fatiga, estado
arbitrario de carga y cambio de orientacion de la carga.
d) Aplicacion del nuevo modelo de remodelacion al analisis de un implante dental os-
teointegrado. Simulacion de la perdida de hueso crestal a largo plazo.
e) Empleo de tecnicas de submodelado para el analisis de una rebanada de la mandıbula.
Los modelos de rebanada tradicionalmente empleados en el analisis de implantes dentales
aplican condiciones de carga y/o desplazamientos muy simples en los extremos. Aquı se han
usado los resultados obtenidos en el modelo de mandıbula completa para aplicar las fuerzas
en los extremos, pero tambien para establecer las propiedades elasticas y la densidad inicial
de los elementos de la rebanada.
f) Analisis de la influencia de diversos factores en la reabsorcion crestal alrededor del
implante: edad del paciente, calidad del hueso previa a la puesta en carga y prescripcion de
un perıodo de curacion, durante el que no se mastique con el lado del implante.

408 Conclusiones y desarrollos futuros
9.4. Desarrollos futuros
Muchas son las vıas que quedan abiertas para un futuro desarrollo en esta lınea de
investigacion. Entre ellas, las mas importantes, en opinion del autor, son:
En relacion a la obtencion numerica de las propiedades mecanicas de la mandıbu-
la
a) Simulacion mas precisa del proceso de masticacion. Es necesario mejorar las condi-
ciones de apoyo, tanto en el condilo como en los dientes. En el condilo se debe usar un
modelo de contacto entre la superficie articular del mismo y el disco articular. Ademas, si
se incluye el disco articular, es necesario incluir un modelo de comportamiento adecuado
del mismo. Un modelo porohiperelastico fibrado del disco articular ha sido desarrollado por
Perez-del Palomar y Doblare [87, 88] y puede ser mas adecuado para este fin.
b) Mejora de la malla en el entorno de los dientes. La geometrıa de los dientes, y lo que
es mas importante, la geometrıa de los alveolos, es solo aproximada. Tambien conviene usar
un modelo de comportamiento especıfico del ligamento periodontal, que se ha simplificado
como elastico lineal. Es importante la mejora en esta zona porque es aquı donde se han
obtenido los peores resultados de densidad.
c) Consideracion de la remodelacion osea externa para la simulacion del proceso de
reabsorcion alveolar y posterior cierre del alveolo que tiene lugar tras la perdida de un
diente.
d) Unificacion de fuentes de las que se toman los datos del modelo. En este trabajo se
ha utilizado una mandıbula concreta, pero los datos de fuerzas masticatorias y patrones
de masticacion se han tomado de la literatura. Los resultados de densidad que sirven de
patron para la validacion de los resultados numericos sı pertenecen a la mandıbula medida,
pero los resultados de propiedades mecanicas estan tomados, nuevamente, de la literatura.
Serıa conveniente repetir este estudio en un individuo vivo, del que extraer todos los datos:
la geometrıa y la densidad osea mediante tomografıa, las fuerzas masticatorias mediante
electromiografıa, el patron de masticacion mediante observacion y las propiedades elasticas
con tecnicas de ultrasonido, por ejemplo.
e) Aplicacion al estudio del desarrollo de la arcada dentaria humana y a la Paleoantropologıa.
En la actualidad, los investigadores que estudian el proceso de aparicion de los dientes per-
manentes en humanos centran sus estudios en la cronologıa de dicho desarrollo; principal-

9.4 Desarrollos futuros 409
mente con la toma de datos de los tiempos medios de emergencia de dientes permanentes
en diferentes poblaciones del humano moderno [205]. Sin embargo, los diferentes patrones
de aparicion de los dientes han sido muy poco estudiados en la bibliografıa. Un mejor en-
tendimiento de la variabilidad de estos patrones es fundamental para el estudio del desarrollo
dental, pudiendo a su vez servir como base de trabajo para el analisis de los cambios mor-
fologicos sufridos por la especie humana durante su evolucion. Para dientes permanentes,
existen algunos datos que muestran diferencias en la secuencia de emergencia, habiendose
obtenido dichos datos mediante una simple comparacion entre pares de dientes [205]. Tam-
bien se conoce la variabilidad en las secuencias del desarrollo dental en ninos [309], pero,
este problema no ha sido nunca analizado cientıficamente, debido a la ausencia de una
metodologıa adecuada [36]. Poco se conoce en la actualidad acerca de los mecanismos que
definen el patron de aparicion de los dientes. Se cree que puede existir un acoplamiento entre
diferentes factores: cargas mecanicas, configuracion de la mandıbula (propiedades mecanicas
y geometrıa), sexo, edad y otros. La aplicacion de los metodos empleados en esta tesis puede
ayudar a comprender mejor el efecto de algunos de estos factores.
En relacion al modelo de remodelacion anisotropo propuesto
a) Es necesario hacer un ajuste mas preciso de la velocidad de los procesos biologicos
que intervienen en la remodelacion y de la dependencia temporal de ciertas variables del
modelo, ya que algunas tienen una gran influencia en la respuesta osea y han sido supuestas
sin ninguna base experimental.
b) Modificar la teorıa de remodelacion osea para hueso cortical. Hay dos aspectos del
modelo que es necesario revisar para aplicarlo a hueso cortical:
b1) Por un lado, la consideracion de poros ortoedricos no tiene mucho sentido en hueso
cortical, donde la porosidad es debida principalmente a los canales haversianos y los canales
de Volkmann, de geometrıa bien distinta a la de un ortoedro. Es probable que las propiedades
elasticas de este tipo de tejido no esten bien definidas mediante el tensor MIL en la forma
adoptada aquı. En ese caso serıa necesario buscar una relacion mas precisa entre porosidad,
direccion de las osteonas (por tanto de los canales) y propiedades elasticas.
Ademas, en el tejido cortical, la anisotropıa esta fuertemente influenciada por la direccion
que tienen las fibras de colageno dentro de las lamelas que forman la osteona [13–16,280,281]
y eso no se ha tenido en las relaciones de comportamiento usadas aquı.

410 Conclusiones y desarrollos futuros
b2) Por otro lado, al evaluar el volumen de hueso reabsorbido por una osteona, se ha
supuesto que dicha osteona, hace un tunel en la matriz osea y reabsorbe un volumen de
tejido igual al volumen de un cilindro de radio el de la osteona, sin tener en cuenta que esa
osteona se puede solapar con una anterior y encontrarse en su avance, no solo matriz osea,
sino tambien poros. El volumen de hueso reabsorbido dependerıa entonces de la porosidad.
c) Tambien debe ser mejorada la teorıa para hueso trabecular. En concreto, debe investi-
garse la acumulacion de dano en este tipo de tejido, que probablemente involucre mecanismos
diferentes a los que tienen lugar en el hueso cortical (ver figura 4.25).
d) Estudiar la influencia de la frecuencia de aplicacion de las cargas. Segun Srinivasan
et al. [310] una carga de baja intensidad puede inducir formacion osea si entre cada ciclo de
carga se prescribe un perıodo de inactividad. Para tratar de explicar este hecho, Srinivasan y
sus colaboradores han desarrollado en un estudio reciente [18] un modelo basado en agentes
(agent based model, ABM) para simular la actividad de los osteocitos.
e) Simulacion multiescala de la evolucion de las BMUs. El efecto de las BMUs esta muy
localizado. Algunos autores han propuesto modelos microscopicos que tratan de predecir
la probabilidad de que en un punto concreto del entramado oseo tenga lugar el proceso
de remodelacion [157, 326, 327]. Pero no solo en la activacion de las BMUs es conveniente
bajar a la escala microscopica, ya que el posterior avance de dichas BMUs tambien afecta
localmente a las propiedades mecanicas.
f) Debe investigarse en profundidad el alineamiento de los poros con la carga, desde el
punto de vista experimental, para ver si efectivamente existe una zona muerta dentro de la
cual los poros dejan de girar para adaptarse a la carga o si por el contrario se produce una
alineacion completa. En ese caso, habrıa que conocer bajo que circunstancias se produce y si
tiene algun efecto la edad, como se sugiere en esta tesis, u otros parametros no considerados
aquı.
g) Modelizacion de la compleja red de osteocitos, lagunas y canalıculos para el caso de
tejido cortical. Se cree que el flujo de fluidos a traves de la red de canalıculos del hueso cortical
es el principal estımulo que controla la respuesta de dicho tejido (remodelacion osea) [73].
Por ello, el desarrollo de modelos que permitan estimar de alguna forma los parametros de
dicho flujo es muy importante, ya que serıa de gran utilidad para la elaboracion de modelos
de remodelacion osea mas realistas, que permitan mejorar los actuales, fundamentalmente
basados en la utilizacion de un estımulo mecanico macroscopico.

9.4 Desarrollos futuros 411
En relacion al modelo del implante
a) Simular el proceso de degradacion de la osteointegracion. Considerar por un lado la
posibilidad de que la union se rompa, si el cortante o la traccion en la interfase hueso -
implante son elevados o simplemente estan por encima de la resistencia de la union. Hay
que tener en cuenta tambien que dicha resistencia puede disminuir como consecuencia de la
degradacion del hueso. Por otro lado, hay que considerar que la osteointegracion perdida en
un determinado momento puede recuperarse si se dan las condiciones para ello.
b) En este mismo sentido, debe analizarse mas en detalle el comportamiento del tejido
fibroso que rodea al implante, a traves del cual se produce la osteointegracion en realidad.
Conviene considerar igualmente el callo oseo que se forma en la zona intervenida quirurgi-
camente, por encima del nivel crestal (ver figura 3.12). Hay que analizar su crecimiento,
consolidacion y posterior reabsorcion.
c) Analizar como influye el proceso quirurgico y el posterior perıodo de curacion en
el hueso intervenido. Una vez que se conozca como han podido afectar ambos, se podran
establecer unas condiciones iniciales para la simulacion mas realistas que las usadas en esta
tesis.
d) Simular la reabsorcion osea crestal mediante remodelacion osea externa. El fenomeno
aquı observado es la disminucion de la densidad osea, pero, en realidad, esa disminucion
va acompanada de un retroceso de la superficie osea alveolar, que solo puede simularse
considerando la remodelacion externa.
e) Comparar el efecto de la geometrıa del implante. Serıa conveniente estudiar distintos
tipos de implante comerciales y analizar la influencia de diversos parametros como la longi-
tud, el diametro, la posicion dentro del arcada dentaria, la superestructura que los soporta,
etc. Especial enfasis debe hacerse en implantes roscados que no se han analizado aquı, pero
que son los de uso mas extendido hoy dıa y tambien en los microrroscados, que han aparecido
en los ultimos anos y que se caracterizan por tener roscada toda su longitud, pero con un
paso de rosca mucho menor en la zona crestal.
f) Validar clınicamente los resultados numericos. Para ello conviene hacer una serie de
seguimientos radiograficos que permitan evaluar la evolucion temporal de la reabsorcion
osea alrededor del implante y compararlas con los resultados aquı obtenidos. Para evaluar la
osteointegracion del implante es conveniente hacer estudios histomorfomericos mas o menos
exhaustivos, pero ello obliga a acometer los estudios en animales, con la evidente limitacion

412 Conclusiones y desarrollos futuros
que esto supone para la comparacion entre resultados numericos y experimentales.

Apendice A
Modelo de Stanford basado en
la energıa: Revision.
En el capıtulo 5 se desarrollan las ideas principales del modelo de remodelacion interna
anisotropa de Stanford basado en la energıa. La formulacion que desarrolla Jacobs en su
tesis tiene algunos fallos que se van a revisar en este apendice.
La idea basica del modelo es que la remodelacion osea se produce de tal manera que
se maximiza la eficiencia, definida en el capıtulo 5, teniendo que cumplir las condiciones
impuestas por los criterios de remodelacion. Las ecuaciones que permiten calcular C y ρ
son:
C : εεε = −n∑
j=1
γj∂fj∂εεε
y Θ′′ρ = −n∑
j=1
γj∂fj∂ρ
, (A.1)
que junto a las condiciones de Kuhn-Tucker cierran el problema de optimizacion:
γj ≥ 0 , fj ≤ 0 , γjfj = 0 (A.2)
El criterio de remodelacion se escribe de la siguiente forma, separando la dependencia
de la deformacion y de la densidad:
fj(εεε,C, ρ) = φj(εεε,C)−Rj(ρ) (A.3)
y si la funcion φj es homogenea de grado uno en la deformacion, como ocurre en el criterio
de remodelacion usado en el modelo isotropo de Stanford:
φj(εεε,C) =∂φj(εεε,C)
∂εεε: εεε (A.4)
413

414 Modelo de Stanford basado en la energıa: Revision.
de la cual se obtiene directamente:
∂fj∂εεε
=∂φj∂εεε
( 1
φj
∂φj∂εεε
: εεε)
(A.5)
Si se sustituye esta derivada en la ecuacion (A.1) se obtiene:
C : εεε = −n∑
j=1
( γjφj
∂φj∂εεε⊗ ∂φj
∂εεε
): εεε (A.6)
que se debe cumplir para cualquier εεε, quedando ası C de la siguiente forma:
C = −n∑
j=1
( γjφj
∂φj∂εεε⊗ ∂φj
∂εεε
), (A.7)
expresion que se diferencia en el signo de la dada por Jacobs [166].
A continuacion se particulariza esta expresion para los criterios de remodelacion intro-
ducidos en el modelo isotropo de Stanford:
fr(εεε,C, ρ) = (−e− w)ρ2−β/2 = φr(εεε,C)−Rr(ρ)fa(εεε,C, ρ) = (e− w)ρ2−β/2 = φa(εεε,C)−Ra(ρ)
(A.8)
El error de remodelacion se definio en el modelo isotropo de Stanford. Usando las ecua-
ciones (5.28), (5.29) y (5.30):
e = Ψt −Ψ∗
t = Ψ
(ρ
ρ
)2
−Ψ∗
t = n1/mc
√2EU
(ρ
ρ
)2
−Ψ∗
t (A.9)
Recordando la correlacion entre el modulo de Young y la densidad aparente, que tiene
la forma general E = Bρβ :
e = K
√εεε : C : εεε
ρ2−β/2−Ψ∗
t donde K = ρ2n1/m√B (A.10)
Sustituyendo en la ecuacion (A.8) se pueden identificar las funciones φa, φr, Ra y Rr:
φr(εεε,C) = −K√εεε : C : εεε
φa(εεε,C) = K√εεε : C : εεε
Rr(ρ) = (w −Ψ∗
t )ρ2−β/2
Ra(ρ) = (w + Ψ∗
t )ρ2−β/2
(A.11)
Diferenciando las funciones φj y sustituyendo en (A.7) se obtiene C:
C =
γrK√σσσ : εεε
σσσ ⊗ σσσσσσ : εεε
si fr ≥ 0 y fa < 0
0 si fa < 0 y fr < 0
−γaK√σσσ : εεε
σσσ ⊗ σσσσσσ : εεε
si fa ≥ 0 y fr < 0
(A.12)

415
que se diferencia de la expresion dada por Jacobs en los signos, al igual que la ecuacion
(A.7), de la que proviene, y en el escalar que acompana a γj . Este escalar no tiene mayor
importancia puesto que los γj se van a calcular mediante regularizacion viscosa [306], como
se vera a continuacion. Sı es mas importante el signo, porque segun las condiciones de Kuhn-
Tucker (A.2) γj ≥ 0 y eso implica que en reabsorcion aumenta la rigidez y en formacion
disminuye, lo que no tiene sentido alguno.
Garcıa sugiere en su tesis [115] que los materiales vivos cumplen el principio de mıni-
ma disipacion. Si se minimiza la disipacion mecanica, esto es la eficiencia Ξ, en lugar de
maximizarla como hace Jacobs, se llega a la siguiente expresion de C:
C =
n∑
j=1
( γjφj
∂φj∂εεε⊗ ∂φj
∂εεε
), (A.13)
que particularizada para el criterio de remodelacion (A.8) queda:
C =
−γrK√σσσ : εεε
σσσ ⊗ σσσσσσ : εεε
si fr ≥ 0 y fa < 0
0 si fa < 0 y fr < 0
γaK√σσσ : εεε
σσσ ⊗ σσσσσσ : εεε
si fa ≥ 0 y fr < 0
(A.14)
que ahora sı tiene sentido fısico.
Regularizacion viscosa
A continuacion se muestra el proceso de regularizacion viscosa, mediante el cual se cal-
culan los parametros γj . Estos deben hacer que el modelo anisotropo reproduzca el modelo
isotropo de Stanford cuando se aplique a un material isotropo sometido a un estado de
tension hidrostatico σσσ = p I.
Por un lado, en el modelo isotropo de Stanford la ley de comportamiento se escribe:
E = Bρβ (A.15a)
C =E
1 + ν
(ν
1− 2ν1⊗ 1 + I
)(A.15b)
donde se supondra que el coeficiente de Poisson no depende de la densidad aparente, salvo
en la transicion entre hueso cortical y trabecular. De aquı, teniendo en cuenta que
E =βρ
ρE, (A.16a)

416 Modelo de Stanford basado en la energıa: Revision.
resulta inmediato obtener el tensor C:
C =βρ
ρ
E
1 + ν
(ν
1− 2ν1⊗ 1 + I
)(A.16b)
Por otro lado se aplica el modelo anisotropo a un estado tensional hidrostatico, σσσ = p1.
La energıa de deformacion es en este caso:
U =1
2σσσ : εεε =
p2
κ(A.17)
donde κ =E
3(1− 2ν)es el modulo de rigidez volumetrica. Aplicando (5.54) para reabsorcion,
por ejemplo, se obtiene:
C = −γrK√σσσ : εεε
κ 1⊗ 1 (A.18)
A continuacion se despejarıa γr igualando (A.18) y (A.16b). Se observa facilmente, que
no es posible igualar ambas ecuaciones porque el termino que acompana a I no existe en
la ecuacion (A.18). Se concluye de esto que no es posible reproducir el modelo isotropo de
Stanford con esta formulacion en ningun caso.
Si a pesar de ello se insiste en igualar los terminos que sı se pueden igualar, los que
acompanan a 1⊗ 1, resulta la siguiente expresion de C:
C =3ν
1 + ν
βρ
ρ
σσσ ⊗ σσσσσσ : εεε
, (A.19)
que es valida tanto en reabsorcion como en formacion. Se observa que esta expresion es
parecida, pero no igual a la ecuacion dada por Jacobs, salvo en el caso de un material
incompresible, para el que el primer factor de la ecuacion (A.19) es igual a la unidad.
Como se ha visto, son muchas las incongruencias que presenta la formulacion dada por
Jacobs en su tesis [166], por lo que no es de extranar que al implementarla se obtengan
resultados poco satisfactorios.

Bibliografıa
[1] http://courses.washington.edu/bonephys/.
[2] http://www.medmayor.cl/odontologia/segundo/fisiologia/masticaion.doc.
[3] American academy of implant dentistry glossary of terms, 1986.
[4] O. A. Abu-Hammad, A. Harrison, and D. Williams. The effect of a hydroxyapatite-
reinforced polyethylene stress distributor in a dental implant on compressive stress
levels in surrounding bone. Int. J. Oral. Max. Impl., 15 (4):559–564, 2000.
[5] R. Adell, U. Lehholm, B. Rockler, and P-I. Branemark. A 15-year study of osseointe-
grated implants in the treatment of the edentulous jaw. Int. J. Oral Surg., 10:387–416,
1981.
[6] K. Akca, M. C. Cehreli, and H. Iplikcioglu. A comparison of three-dimensional finite
element stress analysis with in vitro strain gauge measurements on dental implants.
Int. J. Prosthodont., 15 (2):115–121, 2002.
[7] E. Alarcon. Diario El Paıs, 22 octubre:40, 2003.
[8] T. Albrektsson, P-I. Branemark, H. A. Hansson, B. Kasemo, K. Larsson, I. Lundstrom,
et al. The interface zone of inorganic implant in vivo: titanium implants in bone. Ann.
Biomed. Eng., 11:1–27, 1983.
[9] T. Albrektsson, E. Dahl, S. Engevall, B. Engquist, A. R. Eriksson, et al. Osseoin-
tegrated oral implants: A swedish multicenter study of 8139 consecutively inserted
nobelpharma implants. J. Periodontol., 59:287–296, 1988.
417

418 BIBLIOGRAFIA
[10] F. J. Arendts and C. Sigolotto. Standardabmessungen, Elastizitatskennwerte und Fes-
tigkeitsverhalten des Human-Uterkiefers, ein Betrag zur Darstellung der Biomechanik
der Unterkiefer - Teil i. Biomed. Tech., 34:248–255, 1989.
[11] F. J. Arendts and C. Sigolotto. Mechanische Kennwerte des Human-Unterkiefers und
Untersuchung zum “in-vivo”-Verhalten des kompakten Knochengewebes, ein Betrag
zur Darstellung der Biomechanik der Unterkiefer - Teil ii. Biomed. Tech., 35:123–130,
1990.
[12] M. Arnold. Bruxism and the occlusion. Dent. Clin. North Am., 25:395–407, 1981.
[13] A. Ascenzi and E. Bonucci. The ultimate tensile strength of single osteons. Acta
Anatomica, 58:160–183, 1964.
[14] A. Ascenzi and E. Bonucci. The tensile properties of single osteons. Anatomical Record,
158:375–386, 1967.
[15] A. Ascenzi and E. Bonucci. The compressive properties of single osteons. Anatomical
Record, 161:377–391, 1968.
[16] A. Ascenzi and E. Bonucci. The shearing properties of single osteons. Anatomical
Record, 172:499–510, 1972.
[17] J. E. Aubin and E. Bonnelye. Osteoprotegerin and its ligand: a new paradigm for
regulation of osteoclastogenesis and bone resorption. Medscape Womens Heal., 5:5,
2000.
[18] B. J. Ausk, T. S. Gross, and S. Srinivasan. An agent based model for real-time signaling
induced in osteocytic networks by mechanical stimuli (in press). J. Biomech, 2006.
[19] G. E. Bacon, P. J. Bacon, and R. K. Griffiths. A neutron diffraction study of the
bones of the foot. J. Anatomy, 139:265–273, 1984.
[20] M. Bagge. Remodelling of bone structures. PhD thesis, Technical University of Den-
mark, 1999.
[21] M. Bagge. A model of bone adaptation as an optimization process. J. Biomech.,
33:1349–1357, 2000.

BIBLIOGRAFIA 419
[22] R. Baron, R. Tross, and A. Vignery. Evidence of sequential remodeling in rat tra-
becular bone morphology, dynamic histomorphometry, and changes during skeletal
maturation. Anat. Rec., 208:137–145, 1984.
[23] O. Barou, M. H. Lafage-Proust, S. Palle, L. Vico, and C. Alexandre. Effects of im-
mobilization on preosteoblast proliferation assessed histomorphometrically using brdu
uptake on epon-embedded rat bone (abstract). Bone, 19:131 S, 1996.
[24] R. H. K. Batenburg, H. J. A. Meijer, G. M. Raghoebar, R. P. van Oort, and G. Boering.
Mandibular overdentures supported by two Branemark, IMZ or ITI implants. Clin.
Oral Implan. Res., 9:374–383, 1998.
[25] R. H. K. Batenburg, R. P. van Oort, H. Reintsema, T. J. Brouwer, G. M. Raghoebar,
and G. Boering. Overdentures supported by two implants in the lower jaw. Clin. Oral
Implan. Res., 5:207–212, 1994.
[26] G. S. Beaupre, T. E. Orr, and D. R. Carter. An approach for time-dependent bone
modelling and remodelling - theoretical development. J. Orthopaed. Res., 8 (5):651–
661, 1990.
[27] G. S. Beaupre, T. E. Orr, and D. R. Carter. An approach for time-dependent bone
modelling and remodelling - application: A preliminary remodeling simulation. J.
Orthopaed. Res., 8 (5):662–670, 1990.
[28] C. Bell. Animal Mechanics, or Proofs of Design in the Animal Frame. Morrill Wyman,
Cambridge, MA, 1827.
[29] U. C. Belser and A. G. Hannam. The contribution of the deep fibers of the masseter
muscle to selected tooth clenching and chewing tasks. J. Prosthet. Dent., 56:629–635,
1986.
[30] M. P. Bendsøe. Optimization of Structural Topology Shape and Material. Springer,
Berlin, 1995.
[31] V. Bentolila, T. M. Boyce, D. P. Fyhrie, R. Drumb, T. M. Skerry, and M. B. Schaffler.
Intracortical remodeling in adult rat long bones after fatigue loading. J. Orthopaed.
Res., 8(5):651–661, 1990.

420 BIBLIOGRAFIA
[32] D. H. Birkenhager-Frenkel and J. C. Birkenhager. Bone appositional rate and per-
centage of doubly and singly labeled surfaces: comparison of data from 5 and 20 µm.
Bone, 8:7–12, 1987.
[33] D. B. Bogy and K. C. Wang. Stress singularities at interface corners in bonded dis-
similar isotropic elastic materials. Int. J. Solids Struct., 7:993–1005, 1971.
[34] G. A. Borelli. On the Movement of Animals (Traducido por P. Maquet). Springer-
Verlag, Berlin, 1989.
[35] J. M. Bourgery. Traite Complet de l’Anatomie de l’Homme. I. Osteologie, Paris, 1832.
[36] J. Braga and Y. Heuze. Quantifying variation in human dental developmental se-
quencies. an evo-devo perspective. In S. E. Bailey and J. J. Hublin, editors, Dental
Perspectives on Human Evolution: State of the Art Research in Dental Paleoanthro-
pology. Springer, Netherlands, 2006.
[37] P-I. Branemark. Introducction to osseointegration. In P-I. Branemark, G. Zarb,
and T. Albrektsson, editors, Tissue-integrated Prostheses. Osseointegration in Clinical
Dentistry, pages 1–76. Quintessence, Chicago, 1985.
[38] P-I. Branemark, G. Zarb, and T. Albrektsson. Tissue integrates prostheses: osseoin-
tegration in clinical dentistry. Quintessence, Chicago, USA, 1985.
[39] R. Branemark, L. O. Ohrnell, R. Skalak, L. Carlsson, and P-I. Branemark. Biome-
chanical characterization of osseointegration: an experimental in-vivo investigation in
the beagle dog. J. Orthopaed. Res., 16:61–69, 1998.
[40] H. Brockstedt, J. Bollerslev, F. Melsen, and L. Mosekilde. Cotical bone remodeling in
autosomal dominant osteopetrosis: a study of two different phenotypes. Bone, 18:67–
72, 1996.
[41] H. Brockstedt, M. Kassen, E. F. Eriksen, L. Mosekilde, and F. Melsen. Age- and
sex-related changes in iliac cortical bone mass and remodeling. Bone, 14:681–691,
1993.
[42] C. Ten Bruggenkate, W. A. M. Van der Kwast, and H. S. Oosterbeek. Success criteria
in oral implantology - a review of the literature. Int. J. Oral Implant., 7:45–53, 1990.

BIBLIOGRAFIA 421
[43] J. B. Brunski, A. F. Moccia, S. R. Pollack, E. Korostoff, and D. I. Trachtenberg. The
influence of functional use of endosseous implants on the tissue-implant interface. I.
Histological aspects. J. Dent. Res., 58:1953–1969, 19.
[44] J. B. Brunski, A. F. Moccia, S. R. Pollack, E. Korostoff, and D. I. Trachtenberg. The
influence of functional use of endosseous implants on the tissue-implant interface. II.
Clinical aspects. J. Dent. Res., 58:1970–1980, 19.
[45] J. A. Buckwalter and R. R. Cooper. Bone structure and function. In Instrumental
Courses Lectures, volume 36, pages 29–45. The American Academy of Orthopaedic
Surgeons, 1987.
[46] J. A. Buckwalter, M. J. Glincher, R. R. Cooper, and R. Recker. Bone biology. part i:
Structure, blood supply, cells, matrix and mineralization. J. Bone Joint Surg. Am.,
77-A(8):1256–1275, 1995.
[47] E. H. Burger, J. Klein-Nulend, and S. C. Cowin. Mechanotransduction in bone. Adv.
Organ Biol., 5A:123–136, 1998.
[48] D. B. Burr and R. B. Martin. Calculating the probability that microcracks initiate
resorption spaces. J. Biomech., 26:613–616, 1993.
[49] D. B. Burr, C. H. Turner, P. Naick, M. R. Forwood, W. Ambrosius, et al. Does
microdamage accumulation affect the mechanical properties of bone? J. Biomech.,
31(4):337–345, 1998.
[50] A. H. Burstein, J. D. Currey, V. H. Frankel, and T. D. Reilly. The ultimate properties
of bone tissue: the effects of yielding. J. Biomech., 5:35–44, 1972.
[51] D. Buser, R. K. Schenk, S. Steinenmann, Fiorellini, et al. Influence of surface char-
acteristics on bone integration of titanium implants. a histomorphometric study in
miniature pigs. J. Biomed. Mater. Res., 25:889, 1991.
[52] H. U. Cameron, R. M. Pilliar, and I. MacNab. The effect of movement on the bonding
of porous metal to bone. J. Biomed. Mater. Res., 7:301–311, 1973.
[53] S. Carlsoo. Nervous coordination and mechanical function of the mandibular elevators:
an emg study of the activity and anatomic analysis of the mechanics of the muscles.
Acta Odont. Scand., 10 (11):1–132, 1952.

422 BIBLIOGRAFIA
[54] A. B. Carr, D. A. Gerard, and P. E. Larsen. Quantitative histomorphometric descrip-
tion of implant anchorage for three types of dental implants following 3 months of
healing in baboons. Int. J. Oral. Max. Impl., 12 (6):777–784, 1997.
[55] D. R. Carter. Mechanical loading histories and cortical bone remodeling. Calcified
Tissue Int., 36 (Supplement 1):19–24, 1984.
[56] D. R. Carter and G. S. Beaupre. Skeletal function and form. Cambridge University
Press, Cambridge, 2001.
[57] D. R. Carter, D. P. Fyhrie, and R. T. Whalen. Trabecular bone density and loading
history: regulation of tissue biology by mechanical energy. J. Biomech., 20:785–795,
1987.
[58] D. R. Carter and W. C. Hayes. The behavior of bone as a two-phase porous structure.
J. Bone Joint Surg., 59 (A):954–962, 1977.
[59] D. R. Carter, T. E. Orr, and D. P. Fyhrie. Relationship between loadingt history and
femoral cancellous bone architecture. J. Biomech., 22(3):231–244, 1989.
[60] M. C. Cehreli, H. Iplikcioglu, and O. G. Bilir. The influence of the location of load
transfer on strains around implants supporting four unit cement-retained fixed prosthe-
ses: in vitro evaluation of axial versus off-set loading. J. Oral Rehabil., 29 (4):394–400,
2002.
[61] T. J. Chambers and K. Fuller. Bone cells predispose bone surfaces to resorption by
exposure of mineral to osteoclastic contact. J. Cell. Sci., 6:155–165, 1985.
[62] G. Chaushu, S. Chaushu, A. Tzohar, and D. Dayan. Inmediate loading of single tooth
implants. inmediate versus non inmediate implantation. A. clinical report. Int. J. Oral.
Maxillofac. Implants, 16:267–272, 2001.
[63] L. P. Y. Chin, F. D. Aker, and K. Zarrinnia. The viscoelastic properties of the human
temporomandibular joint disc. J. Oral Maxil. Surg., 54:315–318, 1996.
[64] C. K. Ching, D. H. Kohn, and S. J. Hollister. Micromechanics of implant-tissue inter-
faces. J. Oral Implant., 18(3):220–230, 1992.

BIBLIOGRAFIA 423
[65] K. Choi, J. L. Khun, M. J. Ciarelli, and S. A. Goldstein. The elastic moduli of human
subchondral, trabecular and cortical tissue and the size-dependency of cortical bone
modulus. J. Biomech., 23:1103–1113, 1990.
[66] H. J. Chun, S. Y. Cheong, J. H. Han, S. J. Heo, et al. Evaluation of design parameters
of osseointegrated dental implants using finite element analysis. J. Oral Rehabil.,
29(6):565–574, 2002.
[67] Y. Ciftci and S. Canay. The effect of veneering materials on stress distribution in
implant-supported fixed prosthetic restorations. Int. J. Oral. Max. Impl., 15 (4):571–
582, 2000.
[68] D. L. Cochran. A comparison of endosseous dental implant surfaces. J. Periodontol.,
70 (12):1523–1539, 1999.
[69] J. P. Cordebois and F. Sideroff. Damage induced elastic anisotropy. In Mechanical
Behavior of Anisotropic Solids, Proc. EUROMECH Colloque, volume 115, pages 761–
774, 1982.
[70] S. C. Cowin. The relationship between the elasticity tensor and the fabric tensor.
Mech. Mater., 4:137–147, 1985.
[71] S. C. Cowin. Wolff’s law of trabecular architecture at remodeling equilibrium. J.
Biomech. Eng., 108:83–88, 1986.
[72] S. C. Cowin. The false premise of wolff’s law. Forma, 12:247–262, 1997.
[73] S. C. Cowin. Mechanosensation and fluid transport in living bone. J. Muskuloske.
Neuron Interact., 2(3):256–260, 2002.
[74] S. C. Cowin and D. H. Hegedus. Bone remodeling i: A theory of adaptive elasticity.
J. Elasticity, 6:313–326, 1976.
[75] S. C. Cowin, A. M. Sadegh, and G. M. Luo. An evolutionary wolff’s law for trabecular
architecture. J. Biomech. Eng., 114:129–136, 1992.
[76] J. F. Cox and G. A. Zarb. The longitudinal clinical efficacy of osseointegrated dental
implants: a 3-year report. Int. J. Oral Maxillofacial Implants, 2:91–100, 1987.
[77] R. G. Craig. Restorative dental materials. CV Mosby, St Louis, 6 edition, 1980.

424 BIBLIOGRAFIA
[78] R. G. Craig and F. A. Peyton. Elastic and mechanical properties of human dentin. J.
Dent. Res., 37:710–718, 1958.
[79] L. M. Cruz-Orive, L. M. Karlsson, S. E. Larsen, and F. Wainschtein. Characterizing
anisotropy: a new concept. Micron Microsc. Acta, 23:75–76, 1992.
[80] K. Culmann. Die Graphische Statik. Verlag von Meyer & Zeller, Zurich, 1866.
[81] J. D. Currey. The mechanical consequences of variation in the mineral content of bone.
J. Biomech., 2:1–11, 1969.
[82] J. D. Currey. Mechanical properties of bone with greatly differing functions. J.
Biomech., 12:313–319, 1979.
[83] J. D. Currey. The mechanical adaptations of bones. Princeton University Press, New
Jersey, 1984.
[84] J. D. Currey. The effect of porosity and mineral content on the young’s modulus of
elasticity of compact bone. J. Biomech., 21:131–139, 1988.
[85] D.T. Davy and K.J. Jepsen. Bone Damage Mechanics, chapter 18, pages 18.1–18.25.
Bone Mechanics Handbook, 2nd Ed. CRC Press, 2001.
[86] Instituto Nacional de Estadıstica. http://www.ine.es.
[87] A. Perez del Palomar and M. Doblare. Finite element analysis of the temporomandibu-
lar joint during internal excursion of the mandible (in press). J.Biomech., 2005.
[88] A. Perez del Palomar and M. Doblare. On the numerical simulation of the mechanical
behaviour of articular cartilage (in press). Int. J. Numer. Meth. Eng., 2005.
[89] V. del Valle, G. Faulkner, and J. Wolfaardt. Craniofacial osseointegrated implant-
induced strain distribution: A numerical study. Int. J. Oral. Max. Impl., 12:200–210,
1997.
[90] J. M. H. Dibbets. One century of wolff’s law. In D. S. Carlson and S. A. Goldstein,
editors, Bone Biodynamics in Orthodontic and Orthopaedic Treatment, volume 27 of
Craniofacial Growth Series, pages 1–13. University of Michigan, Ann Arbor, 1992.

BIBLIOGRAFIA 425
[91] M. Doblare and J. M. Garcıa. Anisotropic bone remodelling model based on a contin-
uum damage-repair theory. J. Biomech., 35 (1):1–17, 2002.
[92] M. Doblare, S. Ramtani, and J. M. Garcıa. Computer simulation of an adaptive
damage-bone remodeling law applied to three unit-bone bars structure. Comput. Biol.
Med., 34(3):259–273, 2004.
[93] T. Dostalova, L. Himmlova, M. Jelinek, and C. Grivas. Osseointegration of loaded
dental implant with krf laser hydroxylapatite films on ti6al4v alloy by minipigs. J.
Biomed. Opt., 6 (2):239–243, 2001.
[94] E. L. DuBrul. Origin and evolution of the oral apparatus. Front Oral Physiol., 1:1–30,
1974.
[95] P. Ducheyne, P. DeMeester, E. Aernoudt, M. Martens, and C. Mulier. Influence of
a functional dynamic loading on bone ingrowth into surface pores of orthopaedic im-
plants. J. Biomed. Mater. Res., 11(6):811–838, 1977.
[96] J. Duyck, H. J. Rønold, H. V. Oosterwyck, I. Naert, et al. The influence of static
and dynamic loading on the marginal bone behaviour around implants: an animal
experimental study. Clin. Oral Implant Res., 12:207–218, 2001.
[97] E. F. Eriksen. Normal and pathological remodeling of human trabecular bone: three
dimensional reconstruction of the remodeling sequence in normals and in metabolic
bone disease. Endocr. Rev., 7:379–408, 1986.
[98] E. F. Eriksen, S. F. Hodgson, R. Eastell, S. L. Cedel, et al. Cancellous bone remodeling
in type i (postmenopausal) osteoporosis: quantitative assesment of rates of formation,
resorption and bone loss at tissue and cellular levels. J. Bone Miner. Res., 5:311–319,
1990.
[99] K. Firoozbaksh and S. C. Cowin. Devolution of inhomogeneities in bone structure:
Predictions of adaptive elasticity theory. J. Biomech. Eng., 102:287–293, 1980.
[100] K. Firoozbaksh and S. C. Cowin. An analytical model of pauwels functional adaptation
mechanism for bone. J. Biomech. Eng., 103:246–252, 1981.

426 BIBLIOGRAFIA
[101] J. Folgado, P. R. Fernandes, J. M. Guedes, and H. C. Rodrigues. Evaluation of osteo-
porotic bone quality by a computational model for bone remodeling. Computers and
Structures, 82:1381–1388, 2004.
[102] The American Society for Bone and Mineral Research (ASBMR). Bone curriculum.
http://depts.washington.edu/bonebio/ASBMRed/ASBMRed.html.
[103] P. Fridez. Modelisation de l’adaptation osseuse externe. In Physics Department. EPFL,
1996.
[104] H. M. Frost. Observations on osteoid seams: the existence of a resting state. Henry
Ford Hos. Med. B.e, 8:220–224, 1960.
[105] H. M. Frost. The Laws of Bone Structure. Charles C. Thomas, Springfield IL., 1964.
[106] H. M. Frost. Mathematical Elements of Lamellar Bone Remodelling. Charles C.
Thomas, Springfield IL., 1964.
[107] H. M. Frost. Tetracycline-based histological analysis of bone remodeling. Calc. Tiss.
Res., 3:211–237, 1969.
[108] H. M. Frost. Bone Remodelling and its Relationship to Metabolic Diseases. Charles
C. Thomas, Springfield IL., 1973.
[109] H. M. Frost. The mechanostat: a proposed pathogenic mechanism of osteoporosis and
the bone mass effect of mechanical and non-mechanical agents. Bone Mineral, 2:73–85,
1987.
[110] H. M. Frost and C. H. Turner. Toward a mathematical description of bone biology:
The principle of cellular accommodation. Calc. Tissue Int., 67(2):184–187, 2000.
[111] Y. C. Fung. Biomechanics. Mechanical properties of living tissues. Springer Verlag,
1993.
[112] D. P. Fyhrie and D. R. Carter. A unifying principle relating stress to trabecular bone
morphology. J. Orthopaed. Res., 4:304–317, 1986.
[113] D. P. Fyhrie and M. B. Schaffler. The adaptation of bone apparent density to applied
load. J. Biomech., 28(2):135–146, 1995.

BIBLIOGRAFIA 427
[114] G. Galileo. Discorsi e dimonstrazioni matematiche, intorno a due nuove scienze atten-
tanti alla meccanica ed a muovementi locali. University of Wisconsin Press, Madison,
1638.
[115] J. M. Garcıa. Modelos de remodelacion osea: analisis numerico y aplicacion al diseno
de fijaciones de fracturas del femur proximal. PhD thesis, Universidad de Zaragoza,
1999.
[116] J. M. Garcıa-Aznar, J. H. Kuiper, M. J. Gomez, M. Doblare, and J. B. Richardson.
Computational simulation of fracture healing. J. Biomech., 121:9–14, 2005.
[117] J. M. Garcıa-Aznar, T. Rueberg, and M. Doblare. A bone remodelling model coupling
microdamage growth and repair by 3d bmu activity. Biomech. Model. Mechan., 4(2-
3):147–167, 2005.
[118] T. Gedrange, C. Bourauel, C. Kobel, and W. Harzer. Three-dimensional analysis of
endosseous palatal implants and bones after vertical, horizontal, and diagonal force
application. Eur. J. Orthodont., 25 (2):109–115, 2003.
[119] F. Genna, C. Paganelli, S. Salgarello, and P. Sapelli. 3-d numerical analysis of the
stress state caused by short-term loading of a fixed dental implant containing a pdl-
like nonlinear elastic internal layer. Cmes-Comput. Model. Eng. Sci., 4 (3-4):405–420,
2003.
[120] F. Genna, C. Paganelli, S. Salgarello, and P. Sapelli. Mechanical response of bone
under short-term loading of a dental implant with an internal layer simulating the
nonlinear behaviour of the periodontal ligament. Comp. Meth. Biomech. Biomed.
Eng., 6:305–318, 2003.
[121] C. H. Gibbs. Electromyographic activity during the motionless period of chewing. J.
Prosthet. Dent., 34:35–40, 1975.
[122] C. H. Gibbs, P. E. Mahan, T. M. Wilkinson, and A. Mauderli. Emg activity of the
superior belly of the lateral pterygoid muscle in relation to other jaw muscles. J.
Prosthet. Dent., 51:691–702, 1984.
[123] L. J. Gibson. The mechanical behavior of cancellous bone. J. Biomech., 18:317–328,
1985.

428 BIBLIOGRAFIA
[124] M. M. Giraud-Guille. Twisted plywood architecture of collagen fibrils on human
compact bone osteons. Calcified Tissue Int., 42:167–180, 1988.
[125] J. Goldhahn, J. Seebeck, R. Frei, M. Frenz, I. Antoniadis, and E. Schneider. New
implants designs for fracture fixation in osteoporotic bone. Osteoporosis Int., 16:S112–
S119, 2005.
[126] M. J. Gomez, J. M. Garcıa-Aznar, J. H. Kuiper, and M. Doblare. Influence of fracture
gap size on the pattern of long bone healing: A computational study. J. Theor. Biol.,
235:105–119, 2005.
[127] C. J. Goodacre, G. Bernal, K. Rungcharassaeng, and J. Y. K. Kan. Clinical com-
plications with implants and implant prostheses. J. Prosthet. Dent., 90(2):121–132,
2003.
[128] H. Graf. Occlusal forces during function. In N. H. Rowe, editor, Occlusion: Research in
form and function. University of Michigan School of Dentistry and the Dental Research
Institute, 1975.
[129] C. J. Griffin and C. J. Sharpe. The structure of the adult temporomandibular meniscus.
Aust. Dent. J., 5:190–195, 1960.
[130] J. M. Guedes. User’s manual for the program PREMAT3D. IDMEC-Instituto Superior
Tecnico, Lisboa, 1995.
[131] J. M. Guedes and N. Kikuchi. Preprocessing and postprocessing for materials based
on the homogenization method with adaptive finite element methods. Comput. Meth.
Appl. M., 83:143–198, 1990.
[132] T. M. Hale, B. B. Boretsky, M. J. Scheidt, M. J. McQuade, S. L. Strong, and T. E. van
Dyke. Evaluation of titanium dental implant osseointegration in posterior edentulous
areas of micro swine. J. Oral Implant., 17:118–124, 1991.
[133] C. H. Han, C. B. Johansson, A. Wennerberg, and T. Albrektsson. Quantitative and
qualitative investigations of surface enlarged titanium and titanium alloy implants.
Clin. Oral Implants Res., 9:1–10, 1998.
[134] S. Hansson. The implant neck: smooth or provided with retention elements - a biome-
chanical approach. Clinical Oral Impl. Res., 10(5):394–405, 1999.

BIBLIOGRAFIA 429
[135] S. Hansson. A conical implant - abutment interface at the level of the marginal bone
improves the distribution of stresses in the supporting bone - an axisymmetric finite
element analysis. Clinical Oral Impl. Res., 14(3):286–293, 2003.
[136] T. P. Harrigan and R. W. Mann. Characterization of microstructural anisotropy in
orthotropic materials using a second rank tensor. J. Mater. Sci, 19:761–767, 1984.
[137] R. T. Hart, V. V. Hennebel, N. Thongpreda, W. C. van Buskirk, and R. C. Anderson.
Modeling the biomechanics of the mandible: a three-dimensional finite element study.
J. Biomech., 25 (3):261–286, 1992.
[138] H. A. Haxton. Absolute muscle force in the ankle flexors of man. J. Physiol., 103:267–
273, 1944.
[139] W.C. Hayes and B. D. Snyder. Toward a quantitative formulation of wolff’s law in
trabecular bone. In S. C. Cowin, editor, Mechanical Properties of Bone. Vol. AMD
45, pages 43–68. American Society of Mechanical Engineers, New York, 1981.
[140] S. J. Hazelwood, R. B. Martin, M. M. Rashid, and J. J. Rodrigo. A mechanistic
model for internal bone remodeling exhibits different dynamic responses in disuse and
overload. J. Biomech., 34:299–308, 2001.
[141] H. S. Hedia. Stress and strain distribution behavior in the bone due to the effect of
cancellous bone, dental implant material and the bone height. Bio-Med. Mater. Eng.,
12 (2):111–119, 2002.
[142] D. H. Hegedus and S. C. Cowin. Bone remodeling ii: Small strain adaptive elasticity.
J. Elasticity, 6:337–352, 1976.
[143] E. Helkimo. Bite force and functional state of the masticatory system in young men.
Swed. Dent. J., 2:167–175, 1978.
[144] C. J. Hernandez. Simulation of bone remodeling during the development and treatment
of osteoporosis. PhD thesis, Stanford University, Stanford, California, 2001.
[145] C. J. Hernandez, G. S. Beaupre, T. S. Keller, and D. R. Carter. The influence of bone
volume fraction and ash fraction on bone strength and modulus. Bone, 29(1):74–78,
2001.

430 BIBLIOGRAFIA
[146] K. Heydenrijk, G. M. Raghoebar, H. J. A. Meijer, W. A. van der Reijden, A. J. van
Winkelhoff, and B. Stegenga. Two-stage IMZ implants and ITI implants inserted in a
single-stage procedure. Clin. Oral Impl. Res., 13:371–380, 2002.
[147] K. M. Hiiemae. Mammalian mastication: A review of the activity of the jaw muscles
and the movement they produce in chewing. In K. A. Joysey and P. M. Butler, editors,
Development, Function and Evolution of Teeth. Academic Press, London, 1978.
[148] L. Himmlova, T. Dostalova, A. Kacovsky, and S. Konvickova. Influence of implant
length and diameter on stress distribution: A finite element analysis. J. Prosthet.
Dent., 91 (1):20–25, 2004.
[149] M. Hisamatsu. Studies on mastication and biting forces of differences from occlusal
form of artificial teeth of complete dentures. J. Jpn. Prosthodont., 22:15–35, 1978.
[150] L. C. Hofbauer, S. Khosla, C. R. Dunstan, D. L. Lacey, et al. The roles of osteopro-
tegerin and osteoprotegerin ligand in the paracrine regulation of bone resorption. J.
Bone Miner. Res., 15:2–12, 2000.
[151] G. L. Honee. The anatomy of the lateral pterygoid muscle. Acta Morphol. Neer. Sc.,
10:331–340, 1972.
[152] S. J. Hoshaw, J. B. Brunski, and G. V. B. Cochran. Mechanical loading of Branemark
implants affects interfacial bone modelling and remodelling. Int. J. Oral. Max. Impl.,
9:345–360, 1994.
[153] H. M. Huang, S. Y. Lee, C. Y. Yeh, and C. T. Lin. Resonance frequency assessment
of dental implant stability with various bone qualities: a numerical approach. Clin.
Oral Implant. Res., 13 (1):65–74, 2002.
[154] R. Huiskes. The various stress patterns of press-fit, ingrown and cemented femoral
stems. Clin. Orthop., 261:27–38, 1990.
[155] R. Huiskes and R. Boeklagen. The appliaction of numerical shape optimization to
artificial joint design. In R. L. Spilker and B. R. Simon, editors, Computational Methods
in Bioengineering, page 185. The Am. Soc. Mech. Engrs., New York, 1988.
[156] R. Huiskes and R. Boeklagen. Mathematical shape optimization of hip-prosthesis
design. J. Biomech., 22:793–804, 1989.

BIBLIOGRAFIA 431
[157] R. Huiskes, R. Ruimerman, G. H. van Lethe, and J. D. Janssen. Effects of mechanical
forces on maintenance and adaptation of form in trabecular bone. Nature, 405:704–706,
2000.
[158] R. Huiskes, H. Weinans, H. J. Grootenboer, M. Dalstra, et al. Adaptive bone-
remodeling theory applied to prosthetic-design analysis. J. Biomech., 20 (11/12):1135–
1150, 1987.
[159] S. S. Huja, H. H. Qian, W. E. Roberts, and T. R. Katona. Effects of callus and bonding
on strains in bone surrounding an implant under bending. Int. J. Oral. Max. Impl.,
13 (5):630–638, 1998.
[160] W. L. Hylander. Functional anatomy. In B. G. Sarnat and D. Laskin, editors, The
Temporomandibular Joint: A Biologic Basis for Clinical Practice, pages 60–92. W.B.
Saunders Co., Philadelphia, 1992.
[161] W. L. Hylander and K. R. Johnson. Temporalis and masseter function in humans and
macaques during incision. Int. J. Primatol., 6:289–322, 1985.
[162] W. L. Hylander, K. R. Johnson, and A. W. Crompton. Loading patterns and jaw
movements during mastication in macaca fascicularis: A bone-strain electromyographic
and cineradiographic analysis. Am. J. Phys. Anthropol., 72:287–314, 1987.
[163] F. Iamoni, G. Rasperini, P. Trisi, and M. Simion. Histomorphometric analysis of a
half hydroxiapatite-coated implant in humans: A pilot study. Int. J. Oral Maxillofac.
Implants, 14:729–735, 1999.
[164] H. Iplikcioglu and K. Akca. Comparative evaluation of the effect of diameter, length
and number of implants supporting three-unit fixed partial prostheses on stress distri-
bution in the bone. J. Dent., 30 (1):41–46, 2002.
[165] F. Isidor. Histological evaluation of peri-implant bone at implants subjected to oc-
clussal overload or plaque accumulation. Clin. Oral Implant Res., 8:1–9, 1997.
[166] C. R. Jacobs. Numerical simulation of bone adaptation to mechanical loading. PhD
thesis, Stanford University, 1994.

432 BIBLIOGRAFIA
[167] C. R. Jacobs, J. C. Simo, G. S. Beaupre, and D. R. Carter. Adaptive bone remod-
eling incorporating simultaneous density and anisotropy considerations. J. Biomech.,
30(6):603–613, 1997.
[168] B. Jankelson, G. M. Hoffman, and J. A. Hendron. Physiology of the stomatognathic
system. J. Am. Dent. Assoc., 46:375–386, 1953.
[169] B. Jaramaz and A. M. DiGoia III. Pre-operative surgical simulation of an acetabular
press fit: assembly strains in bone. In Proceedings of the International Conference of
the IEEE Engineering in Medicine and Biology Society, pages 1101–1102, 1993.
[170] Z. F. Jaworski and E. Lok. The rate of osteoclastic bone erosion in haversian remod-
eling sites of adult dog’s rib. Calc. Tiss. Res., 10:103–112, 1972.
[171] Z. F. G. Jaworski, B. Duck, and G. Sekaly. Kinetics of osteoclasts and their nuclei in
evolving secondary haversian systems. J. Anatomy, 133:397–405, 1981.
[172] Z. F. G. Jaworski and C. Hooper. Study of cell kinetics within evolving secondary
haversian systems. J. Anatomy, 131:91–102, 1980.
[173] Z. F. G. Jaworski and H. K. Uhthoff. Reversibility of nontraumatic disuse osteoporosis
during its active phase. Bone, 7:431–439, 1986.
[174] W. S. S. Jee. The skeletal tissues. In L. Weiss, editor, Histology: Cell and Tissue
Biology. Elsevier, 5th edition edition, 1983.
[175] K.J. Jepsen, D.T. Davy, and O. Akkus. Observations of damage in bone, chapter 17,
pages 17.1–17.18. Bone Mechanics Handbook, 2nd Ed. CRC Press, 2001.
[176] L. C. Johnson. Morphologic analysis in pathology: the kinetics of disease and general
biology in bone. In H. M. Frost, editor, Bone Biodynamics. Little, Brown, 1964.
[177] H. Plenk Jr. and H. Zitter. Material considerations. In G. Watzek, editor, Endosseous
Implants: Scientific and Clinical Aspects, pages 63–99. Quintessence Publishing Co.,
Inc., 1996.
[178] L. M. Kachanov. Time of the rupture process under creep conditions. IVZ Akad. Nauk
S.S.R. Otd Tech. Nauk, 8:26–31, 1958.

BIBLIOGRAFIA 433
[179] W. Kahle, H. Leonhardt, and W. Platzer. Atlas de Anatomıa (para estudiantes y
medicos). Tomo I. Ediciones Omega, Barcelona, 1993.
[180] J. L. Katz. Hard tissue as a composite material. i. bounds on the elastic behavior. J.
Biomech., 4:455–473, 1971.
[181] R. F. Kay and K. M. Hiiemae. Jaw movement and tooth use in recent and fossil
primates. Am. J. Phys. Anthropol., 40:227–256, 1974.
[182] A. Kirsch and K. L. Ackermann. The IMZ osteointegrated implant system. Dent.
Clin. North. Am., 33:733–791, 1989.
[183] C. P. Klein, P. Patka, H. B. van der Lubbe, J. G. Wolke, and K. de Groot. Plasma-
sprayed coatings of tetracalciumphosphate, hydroxyl-apatite and alpha-tcp on titani-
um alloy: an interface study. J. Biomed. Mater. Res., 25:53–65, 1991.
[184] J. Klein-Nulend, A. Van der Plas, C. M. Semeins, N. E. Ajubi, J. A. Frangos, et al.
Sensitivity of osteocytes to biomechanical stress in vitro. FASEB J., 9:441–445, 1995.
[185] Y. M. Kong, D. H. Kim, H. E. Kim, S. J. Heo, and J. Y. Koak. Hydroxyapatite-based
composite for dental implants: an in vivo removal torque experiment. J. Biomed.
Mater. Res., 63 (6):714–721, 714-721.
[186] T. W. P. Korioth, D. P. Romilly, and A. G. Hannam. Three-dimensional finite element
stress analysis of the dentate human mandible. Am. J. Phys. Anthropol., 88:69–96,
1992.
[187] M. Kregzde. A method of selecting the best implant prosthesis design option using
three-dimensional finite element analysis. Int. J. Oral. Max. Impl., 8:662–673, 1993.
[188] J. L. Kuhn, S. A. Goldstein, M. J. Ciarelli, and L. S. Mathhews. The limitations of
canine trabecular bone as a model for human: A biomechanical study. J. Biomech.,
22:95–107, 1989.
[189] B. K. F. Kummer. Photoelastic studies on the functional structure of bone. Fol.
Biotheoret., 6:31–40, 1966.
[190] B. K. F. Kummer. Computer simulation of the adaptation of bone to mechanical stress.
In Proceedings of the San Diego Biomedical Symposium, pages 5–12, San Diego, CA,
1971.

434 BIBLIOGRAFIA
[191] B. K. F. Kummer. Biomechanics of bone: mechanical properties, functional struc-
ture, functional adaptation. In Y. C. Fung, editor, Biomechanics: Its Foundation and
Objectives, pages 237–271. Prentice-Hall, Englewood Cliffs, NJ, 1972.
[192] B. K. F. Kummer. The musculo-skeletal system in a man: Development structure and
function in dependence of gravity, and potential limitations for long term space flights.
Adv. Space Res., 6(12):323–330, 1986.
[193] H. K. Kupeyan and K. B. May. Implant and provisional crown placement: a one stage
protocol. Imp. Dent., 7:213–217, 1998.
[194] L. E. Lanyon. Using functional loading to influence bone mass and architecture:
objectives mechanisms and relationship with estrogen of the mechanically adaptive
process in bone. Bone, 1996:37–43, 18.
[195] L. E. Lanyon, A. E. Goodship, C. J. Pye, and J. H. MacFie. Mechanically adaptive
bone remodelling. J. Biomech., 15:141–154, 1982.
[196] L. E. Lanyon and C. T. Rubin. Static versus dynamic loads as an influence on bone
remodeling. J. Biomech., 17:897–906, 1984.
[197] A. Latif. An electromyographic study of the temporalis muscle in normal persons
during selected positions and movements of the mandible. Amer. J. Orthodontics,
43:577–591, 1957.
[198] T.C. Lee, F.J. O’Brien, and D. Taylor. The nature of fatigue damage in bone. Int. J.
Fatigue, 22:847–853, 2000.
[199] J. Lemaitre. A continuous damage mechanic model for ductile fracture. J. Eng. Mater.
Technol., 107:83–89, 1985.
[200] J. Lemaitre and J.L. Chaboche. Mechanics of solid materials. Cambridge University
Press, Cambridge, 1990.
[201] M. E. Levenston. Temporal stability of node-based internal bone adaptation simula-
tions. J. Biomech., 30 (4):403–407, 1997.
[202] I. Lewinstein, L. Banks-Sills, and R. Eliasi. Finite element analysis of a new system
(il) for supporting an implant-retained cantilever prosthesis. Int. J. Oral. Max. Impl.,
10:355–366, 1995.

BIBLIOGRAFIA 435
[203] C. L. Lin and J. C. Wang. Nonlinear finite element analysis of a splinted implant with
various connectors and occlusal forces. Int. J. Oral. Max. Impl., 18 (3):331–340, 2003.
[204] L. Lindquist, B. Rockler, and G. E. Carlsson. Bone resorption around fixtures in
edentulous patients treated with mandibular fixed tissue-integrated prosthesis. J.
Prosthet. Dent., 59:59–63, 1988.
[205] H. M. Liversidge. Variation in modern human dental development. In J. L. Thompson,
G. E. Krovitz, and A. J. Nelson, editors, Patterns of Growth and Development in the
Genus Homo, pages 73–113. Cambridge University Press, Cambridge, 2003.
[206] J. L. Lozada, M. F. Abbate, F. A. Pizzarello, and R. A. James. Comparative three-
dimensional analysis of two finite-element endosseous implant designs. J. Oral Im-
plant., 20(4):315–321, 1994.
[207] J. W. C. MacDonald. Relationship between specific occlusal contacts and jaw clos-
ing muscle activity during parafunctional clenching tasks in man. Master’s thesis,
University of British Columbia, 1982.
[208] J. W. C. MacDonald and A. G. Hannam. Natural tooth contact and activity in the
jaw musculature during clenching. J. Prosthet. Dent., 52:862–867, 1984.
[209] J. W. C. MacDonald and A. G. Hannam. Relationship between occlusal contacts and
jaw closing muscle activity durng tooth clenching. J. Prosthet. Dent., 52:718–729,
1984.
[210] P. E. Mahan, T. M. Wilkinson, C. H. Gibbs, A. Mauderli, and L. S. Brannon. Superior
and inferior lateral pterygoid emg activity at basic jaw positions. J. Prosthet. Dent.,
50:710–717, 1983.
[211] A. Manns and G. Dıaz. Sistema estomatognatico. Sociedad Grafica Almagro Ltda.,
Santiago de Chile, 1988.
[212] J. D. Manson and N. E. Waters. Observations on the rate of maturation of the cat
osteon. J. Anatomy, 99:539–549, 1965.
[213] R. B. Martin. Porosity and specific surface of bone. In CRC Critical Reviews in
Biomedical Engineering, pages 179–222. CRC Press, Boca Raton, FL, 1984.

436 BIBLIOGRAFIA
[214] R. B. Martin. Mathematical model for repair of fatigue damage and stress fracture in
osteonal bone. J. Orthopaed. Res., 13:309–316, 1995.
[215] R. B. Martin. Toward a unifying theory of bone remodeling. Bone, 26(1):1–6, 2000.
[216] R. B. Martin. Fatigue damage, remodeling, and the minimization of skeletal weight.
J. Theor. Biol., 220(2):271–276, 2003.
[217] R. B. Martin. Fatigue microdamage as an essential element of bone mechanics and
biology. Calcified Tissue Int., 6, 2003.
[218] R. B. Martin and D. L. Boardman. The effects of collagen fiber orientation, porosity,
density, and mineralization on bovine cortical bone bending properties. J. Biomech.,
26:1047–1054, 1993.
[219] R. B. Martin and D. B. Burr. Structure, Function and Adaptation of compact bone.
Raven Press, New York, 1989.
[220] R. B. Martin, D. B. Burr, and N. A. Sharkey. Skeletal Tissue Mechanics. Springer
Verlag New York Inc., New York, USA, 1998.
[221] R. B. Martin, G. A. Danucci, and S. J. Hood. Bone apposition rate differences in os-
teonal and trabecular bone. Transactions of the Orthopaedic Research Society, 12:178,
1987.
[222] R. B. Martin and J. Ishida. The relative effects of collagen fiber orientation, porosity,
density, and mineralization on bone strength. J. Biomech., 22:419–426, 1989.
[223] J. Martınez, J. Domınguez, J. M. Garcıa, and M. Doblare. The obtaining of the bone
distribution in a human mandible by means of the application of a remodelling model
based on damage mechanics. In International Congress on Computational Bioengi-
neering, volume 2, pages 689–698, 2003.
[224] G. O. Matheson, D. B. Clement, D. C. McKenzie, J. E. Taunton, et al. Stress fractures
in athletes. a study of 320 cases. Am. J. Sports Med., 15(1):46–58, 1987.
[225] F. McClintock. A criterion for ductile fracture by the growth of holes. J. Appl. Mech.,
35:363–371, 1968.

BIBLIOGRAFIA 437
[226] J. H. McElhaney. Dynamic response of bone and muscle tissue. J. Appl. Physiol.,
21:1231–1236, 1966.
[227] H. J. A. Meijer, J. H. Kuiper, F. J. M. Starmans, and F. Bosman. Stress distribution
around dental implants: Influence of superestructure, length of implants and height of
mandible. J. Prosthet. Dent., 68:96–102, 1992.
[228] H. J. A. Meijer, F. J. M. Starmans, F. Bosman, and W. H. A. Steen. A comparison
of three finite models of an edentulous mandible provided with implants. J. Oral
Rehabil., 20:147–157, 1993.
[229] H. J. A. Meijer, F. J. M. Starmans, C. De Putter, and C. A. van Blitterswijk. The
influence of a flexible coating on the bone stress around dental implants. J. Oral
Rehabil., 22 (2):105–111, 1995.
[230] H. J. A. Meijer, F. J. M. Starmans, W. H. A. Steen, and F. Bosman. A three-
dimensional finite element study on two versus four implants in an edentulous
mandible. Int. J. Prosthodont., 7:271–279, 1994.
[231] H. J. A. Meijer, F. J. M. Starmans, W. H. A. Steen, and F. Bosman. Loading conditions
of endosseous implants in an edentulous human mandible: a three-dimensional, finite-
element study. J. Oral Rehabil., 23:757–763, 1996.
[232] A. Mellal, H. W. A. Wiskott, J. Botsis, S. S. Scherrer, and U. C. Belser. Stimulating
effect of implant loading on surrounding bone - comparison of three numerical models
and validation by in vivo data. Clin. Oral Implant. Res., 15 (2):239–248, 2004.
[233] M. Meyenberg, S. Kubik, and S. Palla. Relationships of the the muscles of mastication
to the articular disc of the temporomandibular joint. Helv. Odontol. Acta, 30:1–20,
1986.
[234] B. Mikic and D. R. Carter. Bone strain gage data and theoretical models of functional
adaptation. J. Biomech., 28(4):465–469, 1995.
[235] C. Milgrom, M. Giladi, M. Stein, H. Kashtan, J. Y. Margulies, et al. Stress fractures
in military recruits. a prospective study showing an unuasully high incidence. J. Bone
Joint Surg. Br., 67(5):732–735, 1985.

438 BIBLIOGRAFIA
[236] S. C. Miller and W. S. S. Jee. Bone lining cells. In B. K. Hall, editor, Bone, pages
1–19. CRC Press, 1992.
[237] J. R. E. Mills. The relationship between tooth patterns and jaw movements in the
hominoidea. In K. A. Joysey and P. M. Butler, editors, Development, Function and
Evolution of Teeth. Academic Press, London, 1978.
[238] C. E. Misch. Contemporary Implant Dentistry, 1st edition. Mosby-Year Book, Inc.,
Saint Louis, USA, 1999.
[239] E. Møller. The chewing apparatus: An electomyographic study of the action of the
muscles of mastication and its correlation to facial morphology. Acta Physiol. Scand.,
69 (Supl. 280):1–229, 1966.
[240] E. Møller. Action of the muscles of mastication. In Y. Kawamura, editor, Frontiers of
Oral Physiology. Karger, Basel, 1974.
[241] S. Mori and D. B. Burr. Increased intracortical remodeling following fatigue damage.
Bone, 14(2):103–109, 1993.
[242] S. Mori, R. Harruff, W. Ambrosius, and D. B. Burr. Trabecular bone volume and
microdamage accumulation in the femoral heads of women with and without femoral
neck fractures. Bone, 21:521–526, 1997.
[243] T. Mori and K. Tanaka. Average stress in matrix and average elastic energy of mate-
rials with misfitting inclusions. Acta Metall. Mater., 21:571–574, 1973.
[244] T. R. Morneburg and P. A. Proschel. Measurement of masticatory forces and implant
loads: a methodologic clinical study. Int. J. Prosthodont., 15 (1):20–27, 2002.
[245] M. Moss. Functional anatomy of the temporomandibular joint. In L. Schwartz, edi-
tor, Disorders of the Temporomandibular Joint: Diagnosis, Management, Relation to
Occlusion of Teeth. W. B. Saunders, Philadelphia, 1959.
[246] T. Nagasao, M. Kobayashi, Y. Tsuchiya, T. Kaneko, and T. Nakajima. Finite element
analysis of the stresses around fixtures in various reconstructed mandibular models -
part ii (effect of horizontal load). J. Cranio Maxillofac. Surg., 31 (3):168–175, 2003.

BIBLIOGRAFIA 439
[247] A. N. Natali, E. A. Meroi, K. R. Williams, and L. Calabrese. Investigation of the
integration process of dental implants by means of a numerical analysis of dynamic
response. Dent. Mater., 13 (5-6):325–332, 1997.
[248] J. L. Neil, T. C. Demos, J. L. Stone, and W. C. Hayes. Tensile and compressive
properties of vertebral trabecular bone. Trans. Orthopaed. Res. Soc., 8:344, 1983.
[249] G. J. Nelson. Three dimensional computer modeling of human mandibular biomechan-
ics. PhD thesis, University of British Columbia, 1986.
[250] K. Nevakari. An analysis of the mandibular movement from rest to occlusal position: A
roentgenographic-cephalometric investgation. Acta Odontol. Scand., 14 (Suppl 19):1–
129, 1956.
[251] M. Nordin and V. H. Frankel. Biomechanics of whole bones and bone tissue. In Basic
Biomechanics of the Skeletal System, pages 15–61. Lea and Febiger, 1980.
[252] J. A. O’Connor, L. E. Lanyon, and H. MacFie. The influence of strain rate on adaptive
bone remodeling. 1982, 15:767–781, J. Biomech.
[253] A. Odgaard, E. B. Jensen, and H. J. G. Gundersen. Estimation of structural anisotropy
based on volume orientation. a new concept. J. Microsc., 157:149–182, 1990.
[254] A. Odgaard, J. Kabel, B. van Rietbergen, M. Dalstra, and R. Huiskes. Fabric and
elastic principal directions of cancellous bone are closely related. J. Biomech., 30
(5):487–495, 1997.
[255] K. Okada, T. Murata, D. Nawata, et al. The estimation of the biting force during
chewing and swallowing. J. Jpn. Prosthodont., 42:193, 1999.
[256] H. V. Oosterwyck, J. V. Sloten, J. Duyck, and I. Naert. Bone loading and adaptation
around oral implants. In E.B. de Las Casas and D. Pamplona, editors, Computational
Models in Biomechanics, pages 1–40. CIMNE, Barcelona, 2003.
[257] S. Orava. Stress fractures. Br. J. Sports Med. Mar., 14(1):40–44, 1980.
[258] M. Owen. The origin of bone cells in the post-natal organism. Arthritis Rheum.,
23:1073–1079, 1980.

440 BIBLIOGRAFIA
[259] G. Papavasiliou, P. Kamposiora, S. C. Bayne, and D. A. Felton. Three-dimensional
finite element analysis of stress distribution around single tooth implants as a function
of bony support, prosthesis type and loading during function. J. Prosthet. Dent.,
76:633–640, 1996.
[260] A. M. Parfitt. The physiologic and clinical significance of bone hisotomorphometric
data, pages 143–223. Bone Histomorphometry Techniques and Interpretation. CRC
Press, Boca Raton, 1983.
[261] A. M. Parfitt. Bone and plasma calcium homeostasis. Bone, 8:S1–S8, 1987.
[262] A. M. Parfitt. Osteonal and hemi-osteonal remodeling: the spatial and temporal frame-
work for signal traffic in adult human bone. J. Cell. Biochem., 55(3):273–86, 1994.
[263] A. M. Parfitt, G. R. Mundy, G. D. Roodman, D.E. Hughes, and B. F. Boyce. A new
model for the regulation of bone resorption, with particular reference to the effects of
biphosphonates. J. Bone Miner. Res., 11(2):150–159, 1996.
[264] G. R. Parr, D. E. Steflik, A. L. Sisk, and A. Aguero. Clinical and histological obser-
vations of failed two-stage titanium alloy basket implants. Int. J. Oral Maxillofacial
Implants, 3:49–55, 1988.
[265] C. A. Pattin, W. E. Caler, and D. R. Carter. Cyclic mechanical property degradation
during fatigue loading of cortical bone. J. Biomech., 29(1):69–79, 1996.
[266] F. Pauwels. Biomechanics of the Locomotor Apparatus: Contributions on the Func-
tional Anatomy of the Locomotor Apparatus. Springer-Verlag, Berlin, 1980.
[267] M. A. Perez, J. M. Garcıa, and M. Doblare. Analysis of the debonding of the stem-
cemented interface in intramedullary fixation using a non-linear fracture mechanics
approach. Eng. Fract. Mech., 72:1125–1147, 2004.
[268] B. Peter. Orthopaedic implants used as drug delivery systems: numerical, in vitro and
in vivo studies. PhD thesis, Ecole Polytechnique Federale de Lausanne, 2004.
[269] H. E. Pettermann, T. J. Reiter, and F. G. Rammerstorfer. Computational simulation
of internal bone remodeling. Arch. Comp. Meth. Eng., 4 (4):295–323, 1997.

BIBLIOGRAFIA 441
[270] P.Moreo, M. A. Perez, J. M. Garcıa, and M. Doblare. Modelling the mechanical
behaviour of living bony interfaces (in press). Comp. Meth. Applied Mech. Eng., 2006.
[271] M. Portigliatti-Barbos, P. Bianco, and A. Ascenzi. Distribution of osteonic and inter-
stitial components in the human femoral shaft with reference to structure, calcification
and mechanical properties. Acta Anat., 115:178–186, 1983.
[272] M. Portigliatti-Barbos, P. Bianco, A. Ascenzi, and A. Boyde. Collagen orientation in
compact bone: Ii. distribution of lamellae in the whole of the human femoral shaft with
reference to its mechanical properties,. Metab. Bone Dis. Relat., 5:309–315, 1984.
[273] P. J. Prendergast. Finite element models in tissue mechanics and orthopaedic implante
design. Clin. Biomech., 12:343–366, 1997.
[274] P. J. Prendergast and D. Taylor. Prediction of bone adaptation using damage accu-
mulation. J. Biomech., 27:1067–1076, 1994.
[275] G. J. Pruim, H. J. De Jongh, and J. J. Ten Bosch. Forces acting on the mandible
during bilateral static bite at different bite force levels. J. Biomech., 13:755–763, 1980.
[276] M. Quirynen, I. Naert, and D. van Steenberghe. Fixture design and overload influence
marginal bone loss and fixture success in the Branemark system. Clin. Oral Implant
Res., 3:104–111, 1992.
[277] W. J. Ralph. Tensile behaviour of the periodontal ligament. J. Periodontal Res.,
17:423–426, 1982.
[278] H. J. Ralston, M. J. Polissar, V. T. Inman, J. R. Close, and B. Feinstein. Dynamic
features of human isolated voluntary muscles. J. Appl. Physiol., 1:526–533, 1949.
[279] S. Ramtani and M. Zidi. A theoretical model of the effect of continuum damage on a
bone adaptation model. J. Biomech., 34(4):471–479, 2001.
[280] T. D. Reilly and A. H. Burstein. The elastic and ultimate properties of compact bone
tissue. J. Biomech., 8:393–405, 1975.
[281] T. D. Reilly, A. H. Burstein, and V. H. Frankel. The elastic modulus for bone. J.
Biomech., 7:271–275, 1974.

442 BIBLIOGRAFIA
[282] J. Y. Rho and R. B. Ashman abd C. H. Turner. Young’s modulus of trabecular
and cortical bone material: ultrasonic and microtensile measurements. J. Biomech.,
26:111–119, 1993.
[283] J. C. Rice, S. C. Cowin, and J. A. Bowman. On the dependency of the elasticity and
strength of cancellous bone on apparent density. J. Biomech., 21:155–168, 1988.
[284] J. R. Rice and D. M. Tracey. On ductile enlargement of voids in triaxial stress fields.
J., 17:201–217, 1969.
[285] M. R. Rieger, W. K. Adams, and G. L. Kinzel. A finite element survey of eleven
endosseous implants. J. Prosthet. Dent., 63:457–465, 1990.
[286] M. R. Rieger, M. Mayberry, and M. O. Brose. Finite element analysis of six endosseous
implants. J. Prosthet. Dent., 63:671–676, 1990.
[287] W. E. Roberts. Bone tissue interface. J. Dent. Education, 52 (12):804–809, 1988.
[288] W. E. Roberts, E. Morey-Holton, and M. R. Gonsalves. Sensitivity of bone cell pop-
ulations to weightlessness and simulated weightlessness. In The Gravity Relevance in
Bone Mineralisation Processes. European Space Agency, 1984.
[289] W. E. Roberts, P. G. Mozsary, and E. Klingler. Nuclear size as a cell-kinetic marker
for osteoblast differentiation. Am. J. Anat., 165:373–384, 1982.
[290] H. Rodrigues and P. R. Fernendes. Computational Bioengineering (Current Trends
and Applications), pages 135–161. Imperial College Press, 2004.
[291] H. Roesler. The history of some fundamental concepts in bone biomechanics. J.
Biomech., 20:1025–1034, 1987.
[292] L. Rohl, E. Larsen, F. Linde, A. Odgaard, and J. Jorgensen. Tensile and compressive
properties of cancellous bone. J. Biomech., 24:1143–1149, 1991.
[293] W. Roux. Gesammelte Abhandlungen ”uber die Entwicklungsmechanik der Organis-
men. Wilhelm Engelmann, Leipzig, 1885.
[294] C. Rubin, A. S. Turner, S. Bain, C. Mallinckrodt, and K. McLeod. Low mechanical
signals strengthen long bones. Nature, 412:603–604, 2001.

BIBLIOGRAFIA 443
[295] C. T. Rubin and L. E. Lanyon. Regulation of bone formation by applied dynamic
loads. J. Bone Jt. Surg., 66A:397–402, 1984.
[296] C. T. Rubin and L. E. Lanyon. Osteoregulatory nature of mechanical stimuli: function
as a determinant for adaptive bone remodeling. J. Orthopaed. Res., 5:300–310, 1987.
[297] S. Sahin, Y. Akagawa, M. Wadamoto, and Y. Sato. The three-dimensional bone
interface of an osseointegrated implant. ii: A morphometric evaluation after three
months of loading. J. Prosthet. Dent., 76:176–180, 1996.
[298] E. Sanchez-Palencia. Non-homegeneous media and vibration theory. In Lectures Notes
in Physics 127. Springer, Berlin, 1980.
[299] M. B. Schaffler and D. B. Burr. Stiffness of compact bone: effects of porosity and
density. J. Biomech., 21:13–16, 1988.
[300] M. B. Schaffler, K. Choi, and C. Milgrom. Aging and matrix microdamage accumula-
tion in human compact bone. Bone, 17:521–525, 1995.
[301] C. L. Schwartz-Dabney, P. C. Dechow, and R. B. Ashman. Elastic properties of the
human mandibular symphisis. J. Dent. Res., 70:2019, 1991.
[302] A. Sertgoz and S. Guvener. Finite element analysis of the effect of cantilever and
implant length on stress distribution in an implant-supported fixed prosthesis. J.
Prosthet. Dent., 76:165–169, 1996.
[303] I. M. Sheppard. Incisive and related movements of the mandible. J. Prosthet. Dent.,
14:898–906, 1964.
[304] D. Siegele and U. Soltesz. Numerical investigations of the influence of implant shape
on stress distribution in the jaw bone. Int. J. Oral. Max. Impl., 4:333–340, 1989.
[305] O. Sigmund. Design of material structures using topology optimization. PhD thesis,
Technical University of Denmark, 1994.
[306] J. C. Simo. Topics on the numerical analysis and simulations of classical plasticity. In
P. Ciarlet and J. Lions, editors, Handbook for Numerical Analysis. Vol. III. Elsevier
Science Publisher (North-Holland), Amsterdam, 1993.

444 BIBLIOGRAFIA
[307] J. C. Simo and J. W. Ju. Strain- and stress-based continuum damage models. i.
formulation. Int. J. Solids Str., 23:821–840, 1987.
[308] J. C. Simo and J. W. Ju. Strain- and stress-based continuum damage models. ii.
computational aspects. Int. J. Solids Str., 23:841–870, 1987.
[309] B. H. Smith. Growth and development and its significance for early hominid behaviour.
Ossa, 14:63–96, 1989.
[310] S. Srinivasan, D. A. Weimer, S. C. Agans, S. D. Bain, and T. S. Gross. Low-magnitude
mechanical loading becomes osteogenic when rest is inserted between each load cycle.
J. Bone Miner. Res., 17:1613–1620, 2002.
[311] D. E. Steflik, D. C. Koth, and R. V. McKinney. Human clinical trials with the single
crystal sapphire endosteal implant. J. Oral Implant., 13:39–53, 1986.
[312] D. E. Steflik, R. V. McKinney, and D. C. Koth. Ultraestructural (tem) observations
of the gingival response to the single crystal endosteal implant. J. Dent. Res., 61:231,
1982.
[313] S. G. Steinemann, J. Eulenberger, P. A. Maeusli, and A. Schroeder. Adhesion of bone
to titanium. In P. Christel, A. Meunier, and A. C. J. Lee, editors, Biological and
Biomechanical Performance of Biomaterials, pages 409–414. Elsevier Science Publish-
ers, Amsterdam, 1986.
[314] T. Steiniche, P. Christiansen, A. Versterby, C. Hasling, et al. Marked changes in iliac
crest bone structure in postmenopausal osteoporotic patients without any signs of
disturbed bone remodeling or balance. Bone, 15:73–79, 1994.
[315] M. A. Stulpner, B. D. Reddy, G. R. Starke, and A. Spirakis. A three-dimensional finite
analysis of adaptive remodelling in the proximal femur. J. Biomech., 30 (10):1063–
1066, 1997.
[316] S. D. Stout and R. Lueck. Bone remodeling rates and skeletal maturation in three
archaeological skeletal populations. Am. J. Phys. Anthropol., 98:161–171, 1995.
[317] S. Tada, R. Stegaroiu, E. Kitamura, O. Miyakawa, and H. Kusakari. Influence of
implant design and bone quality on stress/strain distribution in bone around implants:

BIBLIOGRAFIA 445
A 3-dimensional finite element analysis. Int. J. Oral Maxillofac. Implants, 18 (3):357–
368, 2003.
[318] S. Takata and N. Yasui. Disuse osteoporosis. J. Med. Invest., 48(3-4):147–156, 2001.
[319] D. Taylor. Fatigue of bone and bones: an analysis based on stressed volume. J.
Orthopaed. Res., 16(2):163–169, 1998.
[320] D. Taylor, E. Casolari, and C. Bignardi. Predicting stress fractures using a probabilistic
model of damage, repair and adaptation. J. Orthopaed. Res., 22:487–494, 2004.
[321] D. Taylor, J. G. Hazenberg, and T. C. Lee. The cellular transducer in damage-
stimulated bone remodelling: a theoretical investigation using fracture mechanics. J.
Theor. Biol., 225:65–75, 2003.
[322] D. Taylor and A. Tilmans. Stress intensity variations in bone microcracks during the
repair process. J. Theor. Biol., 229:169–177, 2004.
[323] G. Tepper, R. Haas, W. Zechner, W. Krach, and G. Watzek. Three-dimensional finite
element analysis of implant stability in the atrophic posterior maxilla - a mathematical
study of the sinus floor augmentation. Clin. Oral Implant Res., 13 (6):657–665, 2002.
[324] K. A. Thomas and S. D. Cook. An evaluation of variables influencing implant fixation
by direct bone apposition. J. Biomed. Mater. Res., 19:875–901, 1985.
[325] H. Triepel. Die Architekturen der menschlichen Knochenspongiosa. Verlag der J. F.
Bergmann, Munich, 1922.
[326] K. Tsubota, T. Adachi, and Y. Tomita. Simulation study on model parameters
of trabecular surface remodelling model. In J. Middleton, M. L. Jones, N. G.
Shrive, and G. N. Pande, editors, Computer Methods in Biomechanics and Biomedical
Engineering-3, pages 129–134. Gordon and Breach Science Publishers, London, 2001.
[327] K. Tsubota, T. Adachi, and Y. Tomita. Functional adaptation of cancellous bone in
human proximal femur predicted by trabecular surface remodeling simulation toward
uniform stress state. J. Biomech., 35(12):1541–1551, 2002.
[328] C. H. Turner. Functional determinants of bone structures: Beyond wolff’s law of bone
transformation. Bone, 13:403–409, 1992.

446 BIBLIOGRAFIA
[329] C. H. Turner. Toward a mathematical description of bone biology: The principle of
cellular accomodation. Calcified Tissue Int., 65:466–471, 1999.
[330] C. H. Turner, I. Owan, and Y. Takano. Mechanotransduction in bone: role of strain
rate. Am. J. Physiol., 269:E438–E442, 1995.
[331] H. Vaillancourt, R. M. Pilliar, and D. McCammond. Finite element analysis of crestal
bone loss around porous-coated dental implants. J Applied Biomaterials, 6:267–282,
1995.
[332] O. Verbogt, G. J. Gibson, and M. B. Schaffler. Loss of osteocyte integrity in association
with microdamage and bone remodelling after fatigue in vivo. J. Bone Miner. Res.,
15:60–67, 2000.
[333] A. R. Villanueva and H. M. Frost. Evaluation of factors affecting the tissue-level
haversian bone formation rate in man. J. Dent. Res, 49:836–846, 1970.
[334] M. Vitti and J. V. Basmajian. Integrated actions of masticatory muscles: Simultaneous
emg from eight intramuscular electrodes. Anat. Rec., 187:173–189, 1977.
[335] G. H. von Meyer. Die architektur der spongiosa. Arch. Anat. Physiol. wiss. Med.,
34:615–628, 1867.
[336] G. P. Vose and A. L. Kubala. Bone strength - its relationship to x-ray-determined ash
content. Hum. Biol., 31:261–270, 1959.
[337] M. Wadamoto, Y. Akagawa, Y. Sato, and T. Kubo. The three-dimensional bone
interface of an osseointegrated implant. i: A morphometric evaluation in initial healing.
J. Prosthet. Dent., 76:170–175, 1996.
[338] F. O. Ward. Outlines of Human Osteology. London, 1838.
[339] W. A. Weijs. Biomechanical models and the analysis of form: A study of the mam-
malian masticatory apparatus. Am. Zool., 20:707–719, 1980.
[340] W. A. Weijs and B. Hillen. Relationship between the physiological cross-section of the
human jaw muscles and their cross-sectional in computer tomograms. Acta Anat., 118
(3):129–138, 1984.

BIBLIOGRAFIA 447
[341] H. Weinans, R. Huiskes, and H. J. Grootenboer. The behaviour of adaptive bone-
remodeling simulation models. J. Biomech., 25 (12):1425–1441, 1992.
[342] H. Weinans and P. J. Prendergast. Tissue adaptation as a dynamical process far from
equilibrium. Bone, 19:143–149, 1996.
[343] R. T. Whalen and D. R. Carter. Influence of physical activity on the regulation of
bone density. J. Biomech., 21(10):825–837, 1988.
[344] W. J. Whitehouse and E. D. Dyson. Scanning electron microscope studies of trabecular
bone in the proximal end of the human femur. J. Anat., 118:417–423, 1974.
[345] G. E. O. Widera, J. A. Tesk, and E. Privitzer. Interaction effects among cortical bone,
cancellous bone and periodontal membrane of natural teeth and implants. J. Biomed.
Mater. Res. Symp., 7:613–623, 1976.
[346] S. E. Wildmalm, J. H. Lillie, and M. M. Ash Jr. Anatomical and electromyographical
studies of the lateral pterygoid muscle. J. Oral Rehabil., 14:429–446, 1987.
[347] H. J. Wilke, L. Claes, and S. Steinenmann. The influence of various titanium surfaces
on the interface shear strength between implants and bone. Clin. Implant Mater.,
9:309, 1990.
[348] T. M. Wilkinson. The relationship between the disk and the lateral pterygoid muscle
in the human temporomandibular joint. J. Prosthet. Dent., 60:715–724, 1988.
[349] J. Willer, N. Noack, and J. Hoffmann. Survival rate of IMZ implants: a prospective
10-year analysis. J. Oral Maxillofac. Surg., 61:691–695, 2003.
[350] K. R. Williams and W. M. Murphy. A finite element stress analysis of the transman-
dibular implant system. Int. J. Prosthodont., 14 (2):115–119, 2001.
[351] K. R. Williams and A. D. C. Williams. Impulse response of a dental implant in bone
by numerical analysis. Biomaterials, 18 (10):715–719, 1997.
[352] M. L. Williams. Stress singularities from various boundary conditions in angular
corners of plates in extension. J. Appl. Mech., 19:526–528, 1952.

448 BIBLIOGRAFIA
[353] H. W. A. Wiskott and U. C. Belser. Lack of integration of smooth titanium surfaces:
a working hypothesis based on strains generated in the surrounding bone. Clin. Oral
Implant Res., 10 (6):429–444, 1999.
[354] J. Wolff. The Law of Bone Remodelling (Das Gesetz der Transformation der Knochen).
Springer Verlag, Berlin, 1986. Traducido por Maquet y Furlong.
[355] W. W. Wood, K. Takada, and A. G. Hannam. The electromyographic activity of the
inferior part of the human lateral pterygoid muscle during clenching and chewing. Arc.
Oral Biol., 31:245–253, 1985.
[356] H. Yamada. Strength of Biological Materials. Krieger, Huntington, 1973.
[357] H. S. Yang, H. J. Chung, and Y. J. Park. Stress analysis of a cantilevered fixed partial
denture with normal and reduced bone support. J. Prosthet. Dent., 76:424–430, 1996.
[358] F. A. Young, K. R. Williams, and R. Draughn R. Strohaver. Design of prosthetic
cantilever bridgework supported by osseointegrated implants using the finite element
method. Dental Materials, 14 (1):37–43, 1998.
[359] O. C. Zienkiewicz and J. Z. Zhu. The superconvergent patch recovery and a posteriori
error estimates. part 1: The recovery technique. Int. J. Numer. Meth. Eng., 33:1331–
1364, 1992.
[360] P. Zioupos and J. D. Currey. Changes in the stiffness, strength, and toughness of
human cortical bone with age. Bone, 22(1):57–66, 1998.
[361] P. Zioupos and J.D. Currey. Cumulative damage and the response of human bone in
two-step loading fatigue. J Biomech, 31:825–833, 1998.
[362] P. K. Zysset and A. Curnier. An alternative model for anisotropic elasticity based on
fabric tensors. Mech. Mater., 21:243–250, 1995.
[363] P. K. Zysset, R. W. Goulet, and S. J. Hollister. A global relationship between trabecu-
lar bone morphology and homogenized elastic properties. J. Biomech. Eng., 120:640–
646, 1998.




















