
Temporal changes of soft-bottom zoobenthic communities in and around Alsancak Harbor (Izmir Bay,...
18
ORIGINAL ARTICLE Temporal changes of soft-bottom zoobenthic communities in and around Alsancak Harbor (Izmir Bay, Aegean Sea), with special attention to the autecology of exotic species Melih Ertan C ¸ inar, Tuncer Katagan, Bilal O ¨ ztu ¨ rk, O ¨ zdemir Egemen, Zeki Ergen, Ahmet Kocatas, Mesut O ¨ nen, Fevzi Kirkim, Kerem Bakir, Gu ¨ ley Kurt, Ertan Dagli, Asli Kaymakc ¸i, Sermin Ac ¸ik, Alper Dogan & Tahir O ¨ zcan Department of Hydrobiology, Faculty of Fisheries, Ege University, Bornova, Izmir, Turkey Problem Anthropogenically introduced alien species are one of the major factors affecting biodiversity. The number and diver- sity of non-native species (exotics) vary both at regional and global scales according to the complex combination of differences in history, timing, and diversity of transport vectors, traits of the introduced species, abiotic and biotic features of the invaded environment, and sampling strat- egies (Wonham & Carlton 2005). A study performed in San Francisco Bay indicated that the rate of invasions increased from an average of one new species established every 55 weeks from 1851 to 1960, to an average of one species every 14 weeks from 1961 to 1995 (Cohen & Carl- ton 1998). Species-poor communities, such as those inha- biting polluted or physically degraded environments, are more vulnerable to invasion than are other communities (Elton 1958). Shea & Chesson (2002) demonstrated that a weakening of a species fundamentally important in a native ecosystem structure or those in inherently unstable and variable environments often result in multiple introduc- tions. Native species might not be adapted to the altered Keywords Community structure; exotics; Izmir Bay; Mediterranean Sea; pollution; temporal variation; zoobenthos. Correspondence Melih Ertan C ¸ inar, Department of Hydrobiology, Faculty of Fisheries, Ege University, 35100 Bornova, Izmir, Turkey E-mail: [email protected] Accepted: 15 June 2006 doi:10.1111/j.1439-0485.2006.00102.x Abstract Temporal and spatial variation in soft-bottom benthic communities following recovery from a pollution episode were studied between January and Septem- ber 2004 in and around Alsancak Harbor, located in the polluted part of Izmir Bay (Aegean Sea, eastern Mediterranean). Samples were collected at seven sta- tions by van Veen grab. Three additional stations were sampled by means of a beam trawl to take into account large mobile animals and for a better estimate of the local biodiversity. A total of 231 species belonging to 10 zoobenthic groups were found. Polychaetes contributed 90% of the total faunal popula- tions and mollusks 87% of the total biomass in the area. Community parame- ters varied significantly among stations and sampling periods; number of species ranged from 2 to 79 per 0.1 m 2 grab sample; density from 20 to 81,720 indÆm )2 ; biomass from 0.1 to 4190 gÆm )2 ; Shannon–Wiener diversity index (log 2 base) from 0.4 to 4.4; and Pielou’s evenness index from 0.11 to 1.0. Collections indicate that a number of species, including those sensitive to pol- lution, have colonized the area where azoic conditions had been previously reported. A total of six exotic species, Streblospio gynobranchiata, Polydora cor- nuta, Hydroides dianthus, Hydroides elegans, Anadara demiri and Fulvia fragilis, probably transferred to the area via ballast water or hull fouling, dominated soft or hard substrata in and near Alsancak Harbor. The first two species accounted for more than 70% of the total population in the area, while A. demiri contributed the most to the biomass (93%, at station 7). Marine Ecology. ISSN 0173-9565 Marine Ecology 27 (2006) 229–246 ª 2006 The Authors. Journal compilation ª 2006 Blackwell Publishing Ltd 229
Transcript of Temporal changes of soft-bottom zoobenthic communities in and around Alsancak Harbor (Izmir Bay,...

ORIGINAL ARTICLE
Temporal changes of soft-bottom zoobenthic communitiesin and around Alsancak Harbor (Izmir Bay, Aegean Sea),with special attention to the autecology of exotic speciesMelih Ertan Cinar, Tuncer Katagan, Bilal Ozturk, Ozdemir Egemen, Zeki Ergen, Ahmet Kocatas,Mesut Onen, Fevzi Kirkim, Kerem Bakir, Guley Kurt, Ertan Dagli, Asli Kaymakci, Sermin Acik, AlperDogan & Tahir Ozcan
Department of Hydrobiology, Faculty of Fisheries, Ege University, Bornova, Izmir, Turkey
Problem
Anthropogenically introduced alien species are one of the
major factors affecting biodiversity. The number and diver-
sity of non-native species (exotics) vary both at regional
and global scales according to the complex combination of
differences in history, timing, and diversity of transport
vectors, traits of the introduced species, abiotic and biotic
features of the invaded environment, and sampling strat-
egies (Wonham & Carlton 2005). A study performed in
San Francisco Bay indicated that the rate of invasions
increased from an average of one new species established
every 55 weeks from 1851 to 1960, to an average of one
species every 14 weeks from 1961 to 1995 (Cohen & Carl-
ton 1998). Species-poor communities, such as those inha-
biting polluted or physically degraded environments, are
more vulnerable to invasion than are other communities
(Elton 1958). Shea & Chesson (2002) demonstrated that a
weakening of a species fundamentally important in a native
ecosystem structure or those in inherently unstable and
variable environments often result in multiple introduc-
tions. Native species might not be adapted to the altered
Keywords
Community structure; exotics; Izmir Bay;
Mediterranean Sea; pollution; temporal
variation; zoobenthos.
Correspondence
Melih Ertan Cinar, Department of
Hydrobiology, Faculty of Fisheries, Ege
University, 35100 Bornova, Izmir, Turkey
E-mail: [email protected]
Accepted: 15 June 2006
doi:10.1111/j.1439-0485.2006.00102.x
Abstract
Temporal and spatial variation in soft-bottom benthic communities following
recovery from a pollution episode were studied between January and Septem-
ber 2004 in and around Alsancak Harbor, located in the polluted part of Izmir
Bay (Aegean Sea, eastern Mediterranean). Samples were collected at seven sta-
tions by van Veen grab. Three additional stations were sampled by means of a
beam trawl to take into account large mobile animals and for a better estimate
of the local biodiversity. A total of 231 species belonging to 10 zoobenthic
groups were found. Polychaetes contributed 90% of the total faunal popula-
tions and mollusks 87% of the total biomass in the area. Community parame-
ters varied significantly among stations and sampling periods; number of
species ranged from 2 to 79 per 0.1 m2 grab sample; density from 20 to
81,720 indÆm)2; biomass from 0.1 to 4190 gÆm)2; Shannon–Wiener diversity
index (log2 base) from 0.4 to 4.4; and Pielou’s evenness index from 0.11 to 1.0.
Collections indicate that a number of species, including those sensitive to pol-
lution, have colonized the area where azoic conditions had been previously
reported. A total of six exotic species, Streblospio gynobranchiata, Polydora cor-
nuta, Hydroides dianthus, Hydroides elegans, Anadara demiri and Fulvia fragilis,
probably transferred to the area via ballast water or hull fouling, dominated
soft or hard substrata in and near Alsancak Harbor. The first two species
accounted for more than 70% of the total population in the area, while
A. demiri contributed the most to the biomass (93%, at station 7).
Marine Ecology. ISSN 0173-9565
Marine Ecology 27 (2006) 229–246 ª 2006 The Authors. Journal compilation ª 2006 Blackwell Publishing Ltd 229

environmental conditions, lessening their ability to reduce
resource availability uniformly in time and space, and thus
providing resource opportunities for invaders. Invaders
that alter ecosystem processes such as primary production,
decomposition, hydrology, geomorphology, nutrient
cycling and/or disturbance regimes do not simply compete
with or consume native species – they change the rules of
existence for all species (Vitousek et al. 1997).
The inner part of Izmir Bay, where Alsancak Harbor is
located, has been subjected to various pollution discharges
since the 1960s and is considered to be one of the most pol-
luted environments of the Mediterranean Sea over the last
two decades. The main pollutants affecting the bay water
quality are organic matter, suspended matter, hydrocar-
bons, metals, and pathogenic organisms, all derived mainly
from domestic and industrial wastes (50%), rainfall (15%),
rivers and streams (10%), agricultural sources (10%) and
others (15%) (UNEP 1994). Intense marine transportation
around Alsancak Harbor also contributes to pollution in
the area. According to the statistical data maintained by the
Alsancak Harbor Authority in 2003, a total of 3640 vessels
approached the harbor for loading or unloading. The
majority of the vessels came from Mediterranean countries
but almost 2% came from outside of the Mediterranean.
Because of intense pollution and degradation, azoic condi-
tions together with intense odor have previously been
reported from the area (Muezzinoglu et al. 2000; Dogan
et al. 2005).
Environmental instability and maritime traffic make the
area more susceptible to invasion by exotics. Previous stud-
ies have reported a total of seven exotic species in the inner
part of Izmir Bay: Pseudonereis anomala, Streblospio gyno-
branchiata, Polydora cornuta, Hydroides dianthus, Hydroides
elegans, Anadara demiri, and Fulvia fragilis. Hydroides dian-
thus has been known from the area for more than a century
(Quatrefages 1865) and Ergen (1976) first reported H. ele-
gans from the polluted part of Izmir Bay. These two species
are considered as the primary fouling organisms in the
Mediterranean Sea (Kocak et al. 1999), derived from the
northern Atlantic (H. dianthus) and the tropical/subtrop-
ical region (H. elegans) by shipping (Zibrowius 1971). The
other well-known exotic species from Izmir Bay is A. dem-
iri, which was first reported in Izmir Bay (Demir 1977) and
subsequently from the Greek and Adriatic coasts (Zenetos
1994; Morello & Solustri 2001). This species was possibly
introduced into the area from the Indian Ocean by ship-
ping (Zenetos et al. 2003). More recently, Cinar et al.
(2005) found dense populations of two spionid polycha-
etes, S. gynobranchiata and P. cornuta, around Alsancak
Harbor, and emphasized that they were exotic species ori-
ginated from the South Atlantic coast of the US (type local-
ity of S. gynobranchiata) and the northern Atlantic (P.
cornuta). Cinar & Ergen (2005) reported the Lessepsian
species (i.e. Red Sea species migrated to the Mediterranean
through the Suez Canal) P. anomala on mussels attached to
the piles of Alsancak Harbor and rocks. Another Lessepsian
species, F. fragilis, was found to build up a dense popula-
tion on the polluted soft bottom of Izmir Bay (Ozturk &
Poutiers 2005).
A sudden increase in the number of exotics reported
from the inner part of Izmir Bay coincided with the
improvement in the water quality of the area after the
establishment of ‘The Grand Canal Project’ in February
2000. The project treated wastewater being discharged
into the bay. A recovery in the sediment and water qual-
ity was observed and no azoic conditions have been
detected in the area since 2002 (Ergen et al. 2006). The
disappearance of azoic conditions may create empty
niches for both opportunistic and exotic species. How-
ever, no detailed study regarding this situation has been
carried out within the area.
The objectives of the present study were (i): to investi-
gate the spatial and temporal patterns of soft-bottom ben-
thic communities after an episode of severe pollution and
the establishment of a wastewater treatment plant in the
area, (ii) to examine the possible recovery in water and
sediment quality in the area, (iii) to assess the environ-
mental factors affecting the structure of benthic commu-
nities in the area, and (iv) to determine the importance
of previously known exotic species within the communi-
ties and their spatio-temporal patterns in the area.
Material and Methods
Soft sediments were collected in various periods between
January and September 2004 by a van Veen grab (samp-
ling an area of 0.1 m)2) at seven stations located in and
around Alsancak Harbor (Fig. 1). At each station, three
Fig. 1. Map of the investigated area with the location of sampling
sites. The numbers 1–7 indicate stations sampled with a van Veen
grab, while B1, B2, and B2 are beam-trawl hauls.
Zoobentos and exotics in Alsancak Harbor Cinar, Katagan, Ozturk, Egemen, Ergen, Kocatas, Onen, Kirkim, Bakir, Kurt, Dagli, Kaymakci, Acik,
Dogan & Ozcan
230 Marine Ecology 27 (2006) 229–246 ª 2006 The Authors. Journal compilation ª 2006 Blackwell Publishing Ltd
grab replicates were taken for benthic community analysis
and an additional sample was collected for chemical and
granulometric analyses of sediments. Stations 1–5 were
selected near Alsancak Harbor, station 6 in Pasaport Har-
bor and station 7 off Inciralti, distant from the Alsancak
Harbor. To better estimate the local biodiversity in the
area, three additional stations were selected near Alsancak
Harbor and sampled by means of a beam trawl to capture
large, mobile animals such as crabs and shrimps that can-
not be sampled by using the grab. A presence–absence
matrix was constructed for species found in the beam-
trawl samples, but was not used for further community
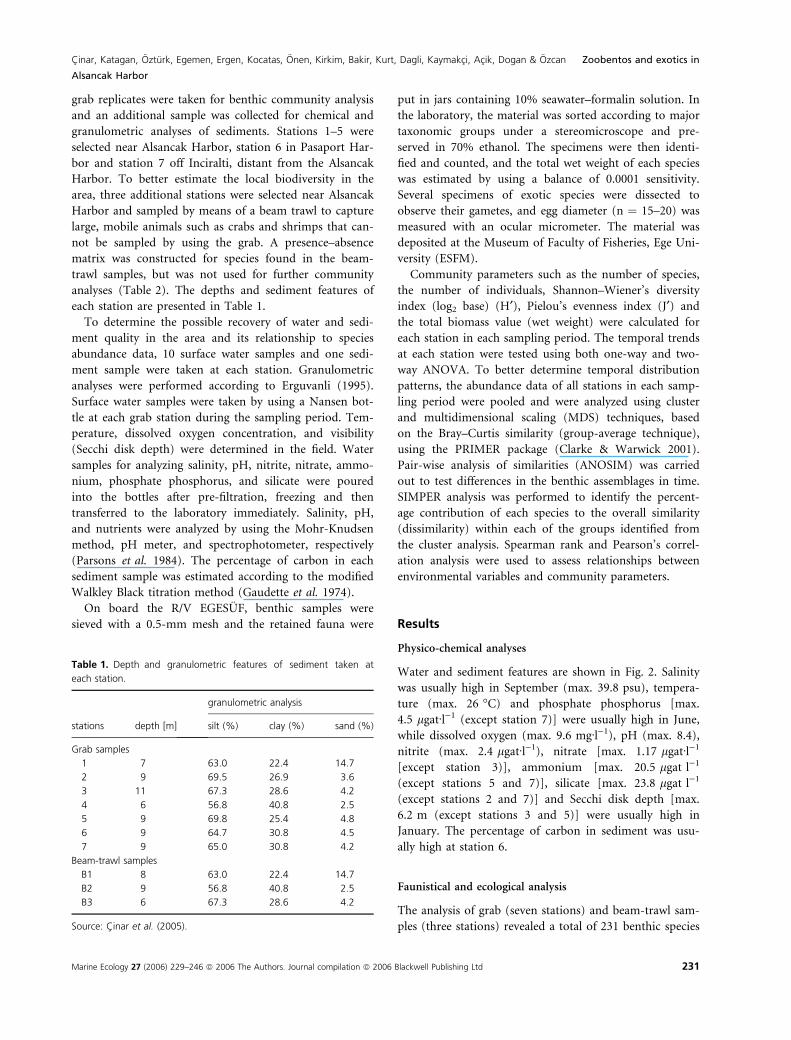
analyses (Table 2). The depths and sediment features of
each station are presented in Table 1.
To determine the possible recovery of water and sedi-
ment quality in the area and its relationship to species
abundance data, 10 surface water samples and one sedi-
ment sample were taken at each station. Granulometric
analyses were performed according to Erguvanli (1995).
Surface water samples were taken by using a Nansen bot-
tle at each grab station during the sampling period. Tem-
perature, dissolved oxygen concentration, and visibility
(Secchi disk depth) were determined in the field. Water
samples for analyzing salinity, pH, nitrite, nitrate, ammo-
nium, phosphate phosphorus, and silicate were poured
into the bottles after pre-filtration, freezing and then
transferred to the laboratory immediately. Salinity, pH,
and nutrients were analyzed by using the Mohr-Knudsen
method, pH meter, and spectrophotometer, respectively
(Parsons et al. 1984). The percentage of carbon in each
sediment sample was estimated according to the modified
Walkley Black titration method (Gaudette et al. 1974).
On board the R/V EGESUF, benthic samples were
sieved with a 0.5-mm mesh and the retained fauna were
put in jars containing 10% seawater–formalin solution. In
the laboratory, the material was sorted according to major
taxonomic groups under a stereomicroscope and pre-
served in 70% ethanol. The specimens were then identi-
fied and counted, and the total wet weight of each species
was estimated by using a balance of 0.0001 sensitivity.
Several specimens of exotic species were dissected to
observe their gametes, and egg diameter (n ¼ 15–20) was
measured with an ocular micrometer. The material was
deposited at the Museum of Faculty of Fisheries, Ege Uni-
versity (ESFM).
Community parameters such as the number of species,
the number of individuals, Shannon–Wiener’s diversity
index (log2 base) (H¢), Pielou’s evenness index (J¢) and
the total biomass value (wet weight) were calculated for
each station in each sampling period. The temporal trends
at each station were tested using both one-way and two-
way ANOVA. To better determine temporal distribution
patterns, the abundance data of all stations in each samp-
ling period were pooled and were analyzed using cluster
and multidimensional scaling (MDS) techniques, based
on the Bray–Curtis similarity (group-average technique),
using the PRIMER package (Clarke & Warwick 2001).
Pair-wise analysis of similarities (ANOSIM) was carried
out to test differences in the benthic assemblages in time.
SIMPER analysis was performed to identify the percent-
age contribution of each species to the overall similarity
(dissimilarity) within each of the groups identified from
the cluster analysis. Spearman rank and Pearson’s correl-
ation analysis were used to assess relationships between
environmental variables and community parameters.
Results
Physico-chemical analyses
Water and sediment features are shown in Fig. 2. Salinity
was usually high in September (max. 39.8 psu), tempera-
ture (max. 26 �C) and phosphate phosphorus [max.
4.5 lgatÆl)1 (except station 7)] were usually high in June,
while dissolved oxygen (max. 9.6 mgÆl)1), pH (max. 8.4),
nitrite (max. 2.4 lgatÆl)1), nitrate [max. 1.17 lgatÆl)1
[except station 3)], ammonium [max. 20.5 lgat l)1
(except stations 5 and 7)], silicate [max. 23.8 lgat l)1
(except stations 2 and 7)] and Secchi disk depth [max.
6.2 m (except stations 3 and 5)] were usually high in
January. The percentage of carbon in sediment was usu-
ally high at station 6.
Faunistical and ecological analysis
The analysis of grab (seven stations) and beam-trawl sam-
ples (three stations) revealed a total of 231 benthic species
Table 1. Depth and granulometric features of sediment taken at
each station.
stations depth [m]
granulometric analysis
silt (%) clay (%) sand (%)
Grab samples
1 7 63.0 22.4 14.7
2 9 69.5 26.9 3.6
3 11 67.3 28.6 4.2
4 6 56.8 40.8 2.5
5 9 69.8 25.4 4.8
6 9 64.7 30.8 4.5
7 9 65.0 30.8 4.2
Beam-trawl samples
B1 8 63.0 22.4 14.7
B2 9 56.8 40.8 2.5
B3 6 67.3 28.6 4.2
Source: Cinar et al. (2005).
Cinar, Katagan, Ozturk, Egemen, Ergen, Kocatas, Onen, Kirkim, Bakir, Kurt, Dagli, Kaymakci, Acik, Dogan & Ozcan Zoobentos and exotics in
Alsancak Harbor
Marine Ecology 27 (2006) 229–246 ª 2006 The Authors. Journal compilation ª 2006 Blackwell Publishing Ltd 231
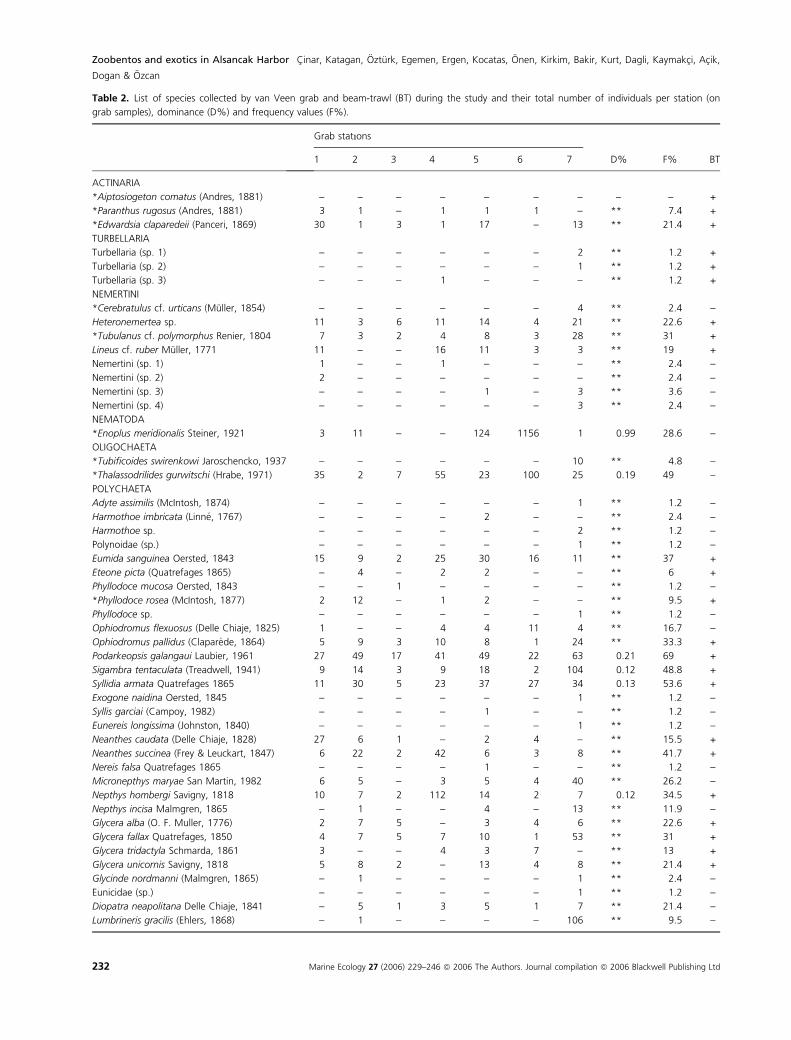
Table 2. List of species collected by van Veen grab and beam-trawl (BT) during the study and their total number of individuals per station (on
grab samples), dominance (D%) and frequency values (F%).
Grab statıons
D% F% BT1 2 3 4 5 6 7
ACTINARIA
*Aiptosiogeton comatus (Andres, 1881) ) ) ) ) ) ) ) ) ) +
*Paranthus rugosus (Andres, 1881) 3 1 ) 1 1 1 ) ** 7.4 +
*Edwardsia claparedeii (Panceri, 1869) 30 1 3 1 17 ) 13 ** 21.4 +
TURBELLARIA
Turbellaria (sp. 1) ) ) ) ) ) ) 2 ** 1.2 +
Turbellaria (sp. 2) ) ) ) ) ) ) 1 ** 1.2 +
Turbellaria (sp. 3) ) ) ) 1 ) ) ) ** 1.2 +
NEMERTINI
*Cerebratulus cf. urticans (Muller, 1854) ) ) ) ) ) ) 4 ** 2.4 )Heteronemertea sp. 11 3 6 11 14 4 21 ** 22.6 +
*Tubulanus cf. polymorphus Renier, 1804 7 3 2 4 8 3 28 ** 31 +
Lineus cf. ruber Muller, 1771 11 ) ) 16 11 3 3 ** 19 +
Nemertini (sp. 1) 1 ) ) 1 ) ) ) ** 2.4 )Nemertini (sp. 2) 2 ) ) ) ) ) ) ** 2.4 )Nemertini (sp. 3) ) ) ) ) 1 ) 3 ** 3.6 )Nemertini (sp. 4) ) ) ) ) ) ) 3 ** 2.4 )NEMATODA
*Enoplus meridionalis Steiner, 1921 3 11 ) ) 124 1156 1 0.99 28.6 )OLIGOCHAETA
*Tubificoides swirenkowi Jaroschencko, 1937 ) ) ) ) ) ) 10 ** 4.8 )*Thalassodrilides gurwitschi (Hrabe, 1971) 35 2 7 55 23 100 25 0.19 49 )POLYCHAETA
Adyte assimilis (McIntosh, 1874) ) ) ) ) ) ) 1 ** 1.2 )Harmothoe imbricata (Linne, 1767) ) ) ) ) 2 ) ) ** 2.4 )Harmothoe sp. ) ) ) ) ) ) 2 ** 1.2 )Polynoidae (sp.) ) ) ) ) ) ) 1 ** 1.2 )Eumida sanguinea Oersted, 1843 15 9 2 25 30 16 11 ** 37 +
Eteone picta (Quatrefages 1865) ) 4 ) 2 2 ) ) ** 6 +
Phyllodoce mucosa Oersted, 1843 ) ) 1 ) ) ) ) ** 1.2 )*Phyllodoce rosea (McIntosh, 1877) 2 12 ) 1 2 ) ) ** 9.5 +
Phyllodoce sp. ) ) ) ) ) ) 1 ** 1.2 )Ophiodromus flexuosus (Delle Chiaje, 1825) 1 ) ) 4 4 11 4 ** 16.7 )Ophiodromus pallidus (Claparede, 1864) 5 9 3 10 8 1 24 ** 33.3 +
Podarkeopsis galangaui Laubier, 1961 27 49 17 41 49 22 63 0.21 69 +
Sigambra tentaculata (Treadwell, 1941) 9 14 3 9 18 2 104 0.12 48.8 +
Syllidia armata Quatrefages 1865 11 30 5 23 37 27 34 0.13 53.6 +
Exogone naidina Oersted, 1845 ) ) ) ) ) ) 1 ** 1.2 )Syllis garciai (Campoy, 1982) ) ) ) ) 1 ) ) ** 1.2 )Eunereis longissima (Johnston, 1840) ) ) ) ) ) ) 1 ** 1.2 )Neanthes caudata (Delle Chiaje, 1828) 27 6 1 ) 2 4 ) ** 15.5 +
Neanthes succinea (Frey & Leuckart, 1847) 6 22 2 42 6 3 8 ** 41.7 +
Nereis falsa Quatrefages 1865 ) ) ) ) 1 ) ) ** 1.2 )Micronepthys maryae San Martin, 1982 6 5 ) 3 5 4 40 ** 26.2 )Nepthys hombergi Savigny, 1818 10 7 2 112 14 2 7 0.12 34.5 +
Nepthys incisa Malmgren, 1865 ) 1 ) ) 4 ) 13 ** 11.9 )Glycera alba (O. F. Muller, 1776) 2 7 5 ) 3 4 6 ** 22.6 +
Glycera fallax Quatrefages, 1850 4 7 5 7 10 1 53 ** 31 +
Glycera tridactyla Schmarda, 1861 3 ) ) 4 3 7 ) ** 13 +
Glycera unicornis Savigny, 1818 5 8 2 ) 13 4 8 ** 21.4 +
Glycinde nordmanni (Malmgren, 1865) ) 1 ) ) ) ) 1 ** 2.4 )Eunicidae (sp.) ) ) ) ) ) ) 1 ** 1.2 )Diopatra neapolitana Delle Chiaje, 1841 ) 5 1 3 5 1 7 ** 21.4 )Lumbrineris gracilis (Ehlers, 1868) ) 1 ) ) ) ) 106 ** 9.5 )
Zoobentos and exotics in Alsancak Harbor Cinar, Katagan, Ozturk, Egemen, Ergen, Kocatas, Onen, Kirkim, Bakir, Kurt, Dagli, Kaymakci, Acik,
Dogan & Ozcan
232 Marine Ecology 27 (2006) 229–246 ª 2006 The Authors. Journal compilation ª 2006 Blackwell Publishing Ltd

Table 2. Continued.
Grab statıons
D% F% BT1 2 3 4 5 6 7
Lumbrineris latreilli Audouin & M. Edwards, 1834 ) ) ) ) ) ) 4 ** 3.6 )Schistomeringos rudolphii (Delle Chiaje, 1828) 67 9 30 7 6 3 12 0.11 34.5 +
Drilonereis filum (Claparede, 1868) ) 1 ) ) ) ) ) ** 1.2 )Aonides oxycephala (Sars, 1862) ) ) ) ) ) ) 4 ** 3.6 )Laonice cirrata (Sars, 1851) ) ) ) ) ) ) 1 ** 1.2 )Malacoceros fuliginosus (Claparede, 1868) 4 ) 13 ) ) 5 ) ** 15.5 )Polydora coeca (Oersted, 1843) ) ) ) ) ) ) 10 ** 2.4 )Polydora cornuta Bosc, 1802 203 256 106 247 304 141 8 0.97 69 +
Polydora sp. 1 ) ) ) ) 2 ) 4 ** 5.6 +
Polydora sp. 2 ) ) ) ) ) ) 1 ** 1.2 +
Prionospio fallax Soderstrom, 1920 62 135 26 127 83 198 265 0.68 67.9 +
Prionospio multibranchiata Berkeley, 1927 308 1452 398 893 620 951 1688 4.84 91.7 +
Prionospio sp. ) ) ) 7 ) ) ) ** 2.4 )Pseudopolydora antennata (Claparede, 1870) 653 1102 77 2343 1925 1441 15 5.8 66.7 +
*Pseudopolydora pulchra (Carazzi, 1895) 5 23 2 2 9 2 ) ** 21.4 +
Spio decoratus Bobretzky, 1870 105 92 37 378 187 123 7 0.71 48.8 +
Streblospio gynobranchiata Rice & Levin 1998 17956 15678 14151 13572 16765 6337 1069 66 95.2 +
Aricidea fragilis mediterranea Laubier & Ramos, 1974 ) 6 ) ) 14 ) 37 ** 20.2 +
Aricidea suecica meridionalis Laubier & Ramos, 1974 ) 1 1 ) 2 ) 43 ** 15.5 )Cirrophorus furcatus (Hartman, 1957) 1 ) ) ) 15 ) 26 ** 11.9 )Paradoneis lyra (Southern, 1914) 3 ) ) ) 4 ) 17 ** 11.9 )Poecilochaetus sp. ) ) ) ) 2 ) 1 ** 3.6 )Spiochaetopterus costarum (Claparede, 1870) ) ) ) ) ) ) 17 ** 9.5 )Magelona alleni Wilson, 1958 ) ) ) ) ) ) 2 ** 2.4 )Magelona minuta Eliason, 1962 ) ) ) ) ) ) 11 ** 7.1 )Capitella capitata (Fabricius 1780) 1963 217 359 310 827 1114 61 3.72 71.4 +
Heteromastus filiformis (Claparede, 1864) 1269 729 79 1086 1284 314 782 4.26 78.6 +
Mediomastus fragilis Rasmussen, 1973 ) 1 ) 12 274 ) 871 0.89 16.7 +
Mediomastus sp. 8 ) ) 1 ) ) 2 ** 7.1 )Notomastus latericeus Sars, 1851 ) ) ) ) ) ) 2 ** 2.4 )Ctenodrilus serratus (Scmidt, 1857) ) ) ) ) ) 148 ) 0.11 3.6 )Aphelochaeta spp. ) ) ) ) ) ) 64 ** 7.1 )Caulleriella bioculata (Keferstein, 1862) ) ) ) ) ) ) 3 ** 2.4 )Chaetozone gibber Woodham & Chambers, 1994 1 ) ) ) 1 ) 18 ** 11.9 +
Chaetozone sp. 1 ) ) ) ) ) ) 15 ** 7.1 )Chaetozone sp. 2 ) ) ) ) ) ) 2 ** 2.4 )Cirriformia cf. tentaculata (Montagu, 1808) 1 ) ) ) ) ) 29 ** 7.1 )*Dodecaceria cf. sextentaculata (Delle Chiaje, 1822) ) ) ) ) ) ) 10 ** 1.2 )Dodecaceria sp. ) ) ) ) ) ) 1 ** 1.2 )Monticellina dorsobranchialis (Kirkegaard, 1959) ) ) ) ) ) ) 5 ** 2.4 )Monticellina heterochaeta Laubier, 1961 ) ) ) ) ) ) 24 ** 4.8 )Monticellina sp. ) ) ) ) ) ) 1 ** 1.2 )Protocirrineris sp. ) ) ) ) ) ) 1 ** 1.2 )Timarete sp. ) ) ) ) ) ) 53 ** 3.6 )*Ophelina modesta Stop)Bowitz, 1958 16 26 18 6 29 4 14 ** 25 +
Polyophthalmus pictus (Dujardin, 1839) ) ) ) ) ) ) 1 ** 1.2 )Cossura soyeri Laubier, 1963 ) ) ) ) ) ) 29 ** 8.3 )Owenia fusiformis Delle Chiaje, 1842 1 ) ) ) ) 2 5 ** 5.6 )Lagis koreni (Malmgren, 1866) 1 13 8 ) 77 2 8 ** 26.2 +
Sabellaria alveolata (Linne, 1767) ) ) ) ) ) ) ) ) ) +
*Sabellaria alcocki Gravier, 1906 ) ) ) 1 ) ) 21 ** 3.6 +
Sabellaria spinulosa Leuckart, 1849 ) ) ) ) ) ) 1 ** 1.2 )Melinna palmata Grube, 1870 ) ) ) ) ) ) 19 ** 10.7 )Lanice conchylega (Pallas, 1766) ) 2 1 13 25 ) ) ** 19 +
Terebella lapidaria Linne 1767 1 ) ) ) ) ) 52 ** 6 +
Cinar, Katagan, Ozturk, Egemen, Ergen, Kocatas, Onen, Kirkim, Bakir, Kurt, Dagli, Kaymakci, Acik, Dogan & Ozcan Zoobentos and exotics in
Alsancak Harbor
Marine Ecology 27 (2006) 229–246 ª 2006 The Authors. Journal compilation ª 2006 Blackwell Publishing Ltd 233

Table 2. Continued.
Grab statıons
D% F% BT1 2 3 4 5 6 7
Polycirrus sp. ) ) ) ) ) ) 61 ** 7.1 )Chone filicaudata Southern, 1914 ) ) ) ) ) ) 1 ** 1.2 )Demonax brachychone (Claparede, 1870) ) ) ) 1 ) ) ) ** 1.2 )Demonax tenuicollaris (Grube, 1870) ) ) ) ) ) ) 2 ** 2.4 +
*Demonax cf. langerhansi Knight)Jones, 1983 ) ) ) ) ) ) 1 ** 1.2 +
Demonax sp. ) ) ) 1 ) ) ) ** 1.2 )Euchone rosea Langerhans, 1884 ) ) ) ) ) ) 20 ** 3.6 )*Myxicola infundibulum (Renier, 1804) ) ) ) ) ) ) 2 ** 1.2 )Hydroides dianthus (Verrill, 1873) 11 2 ) 25 1 ) 83 ** 15.5 +
Hydroides elegans (Haswell, 1883) ) 2 ) 12 1 3 10 ** 16.7 +
Spirobranchus polytrema (Philippi, 1844) ) ) ) ) ) ) 4 ** 1.2 )Vermiliopsis infundibulum (Gmelin, 1788) ) ) ) ) ) ) ) ) ) +
PHORONIDA
Phoronis cf. muelleri Selys-Longchamps, 1903 ) 10 ) 1 11 ) 28 ** 22.6 +
Phoronis sp. 1 ) ) ) ) ) 140 0.11 6 +
CRUSTACEA
Balanus amphitrite (Darwin, 1854) ) ) ) ) ) ) ) ) ) +
Balanus eburneus Gould, 1841 ) ) ) ) ) ) ) ) ) +
Ampelisca pseudosarsi B Santini&K Malka, 1977 ) ) ) ) ) ) 45 ** 3.6 )Ampelisca sarsi Chevreux, 1888 ) ) ) ) ) ) 2 ** 2.4 )Corophium acherusicum Costa, 1851 ) 2 ) ) ) ) ) ** 2.4 +
Corophium acutum Chevreux, 1908 ) 6 ) 14 30 1 ) ** 15.5 +
Elasmopus affinis (Della Valle, 1893) ) 1 ) ) ) 1 ) ** 2.4 )Elasmopus rapax A.Costa, 1853 ) ) ) 5 ) ) 2 ** 2.4 +
Erichthonius brasiliensis (Dana, 1855) ) ) ) ) ) ) 141 0.11 3.6 +
Jassa marmorata Holmes, 1903 ) ) ) ) ) ) ) ) ) +
Leucothoe serraticarpa Della Valle, 1893 ) ) ) ) ) ) 34 ** 3.6 )Maera grossimana Montagu, 1808 ) ) ) ) ) ) 11 ** 2.4 )Maera inaequipes (Costa, 1857) ) ) ) ) ) ) 19 ** 2.4 )Maera hamigera Haswell, 1880 ) ) ) ) ) ) ) ) ) +
Maera shiecki Karaman & Ruffo, 1971 ) ) ) ) ) ) 12 ** 2.4 )Microdeutopus gryllotalpa (Costa, 1853) ) ) ) ) ) ) 7 ** 2.4 )Microdeutopus sp. 1 ) ) ) ) ) ) ** 1.2 +
Pereinotus testudo (Montagu, 1808) ) ) ) ) ) ) 1 ** 1.2 )Phtisica marina Slabber, 1749 16 61 ) 10 149 9 6 0.19 23.8 +
Synchelidium haplocheles (Grube, 1864) ) ) ) ) ) ) 9 ** 7.1 )Synchelidium longidigitatum Ruffo, 1947 ) ) ) ) ) ) 5 ** 2.4 )Cymodoce tuberculata Costa in Hope, 1851 ) ) ) ) ) ) 4 ** 1.2 +
Gnathia vorax (Lucas 1849) ) ) ) ) ) ) 26 ** 3.6 )Leptochelia savignyi (Kroyer, 1842) ) ) ) ) ) ) 30 ** 2.4 )Iphinoe serrata Norman, 1867 2 4 7 5 1 ) 13 ** 15.5 )Iphinoe tenella (G. O. Sars, 1878) ) ) 1 ) 1 ) 5 ** 6 )Mesopodopsis slabberi (P. J. Van Beneden, 1861) ) 2 ) ) ) ) ) ** 2.4 )Paramysis helleri (G. O. Sars, 1877) ) ) ) 4 ) ) 4 ** 3.6 )Mysis sp. ) ) ) ) ) ) 1 ** 1.2 +
Squilla mantis Linne, 1758 ) ) ) ) ) ) ) ) ) +
Crangon crangon (Linne, 1758) ) ) ) ) ) ) ) ) ) +
Palaemon adspersus (Rathke, 1837) ) ) ) ) ) ) ) ) ) +
Melicertus kerathurus (Forskal, 1775) ) ) ) ) ) ) ) ) ) +
Sicyonia carinata (Brunnich, 1768) ) ) ) ) ) ) ) ) ) +
Galathea cenarroi (Zariquiey Alvarez, 1968) ) ) ) ) ) ) ) ) ) +
Upogebia pusilla (Petagna, 1792) ) ) ) ) ) ) 1 ** 1.2 )Brachynotus sexdentatus (Risso, 1827) 5 16 ) 12 3 ) 9 ** 32.1 +
Zoobentos and exotics in Alsancak Harbor Cinar, Katagan, Ozturk, Egemen, Ergen, Kocatas, Onen, Kirkim, Bakir, Kurt, Dagli, Kaymakci, Acik,
Dogan & Ozcan
234 Marine Ecology 27 (2006) 229–246 ª 2006 The Authors. Journal compilation ª 2006 Blackwell Publishing Ltd

Table 2. Continued.
Grab statıons
D% F% BT1 2 3 4 5 6 7
Carcinus aestuarii Nardo, 1847 ) ) ) ) ) ) ) ) ) +
Polybius vernalis (Risso, 1827) ) ) ) ) ) ) ) ) ) +
Pisidia bluteli (Risso, 1816) ) ) ) ) ) ) 1 ** 1.2 +
MOLLUSCA
Cerithium vulgatum Brugiere, 1792 ) ) ) ) 3 ) ) ** 1.2 )Bittium latreillii (Payraudeau, 1826) ) 2 ) ) 1 ) ) ** 3.6 )Bittium reticulatum (Da Costa, 1778) ) 48 1 ) 26 ) 13 ** 16.7 +
Bittium scabrum (Olivi, 1792) ) 1 ) ) ) ) ) ** 1.2 +
Skeneopsis sp. 3 1 3 ) 1 ) ) ** 4.8 )Alvania geryonia (Mardo, 1847 ex Chiereghini) ) 21 1 1 7 ) ) ** 10.7 )Crisilla semistriata (Montagu, 1808) ) ) ) ) 1 ) ) ** 1.2 )Pusillina lineolata (Michaud 1832) 5 5 3 ) 2 20 3 ** 21.4 +
Ventrosia ventrosa (Montagu, 1803) ) 1 ) ) ) ) ) ** 1.2 )Hyala vitrea (Montagu, 1803) ) ) ) ) ) ) 1 ** 1.2 )Aporrhais pespelecani (Linne, 1767) ) ) ) ) ) ) 2 ** 1.2 )Calyptraea chinensis (Linne, 1758) ) ) ) ) ) ) 1 ** 1.2 )Euspira guillemini (Payraudeau, 1826) ) 1 ) ) ) ) ) ** 1.2 )Euspira pulchella (Risso, 1826) ) ) ) ) ) ) 1 ** 1.2 )Monophorus perversus (Linne, 1758) ) 2 ) ) ) ) ) ** 2.4 )Cerithiopsis tubercularis (Montagu, 1803) ) 3 ) ) ) ) ) ** 1.2 )Epitonium commune (Lamarck, 1822) ) 1 ) ) 1 ) ) ** 2.4 )Eulima glabra (Da Costa, 1778) ) 7 ) ) ) ) ) ** 6 )Hexaplex trunculus (Linne, 1758) ) 3 ) ) ) ) ) ** 2.4 +
Cyclope neritea (Linne, 1758) ) ) ) ) ) ) ) ) ) +
Nassarius incrassatus (Stroem, 1768) ) 3 ) ) ) ) ) ** 1.2 )Nassarius pygmaeus (Lamarck, 1822) ) 58 ) 1 9 3 7 ** 19 +
Nassarius reticulatus (Linne, 1758) 2 2 ) ) ) 1 1 ** 7.1 +
Bela brachystoma (Philippi, 1844) 1 5 ) ) ) ) 1 ** 6 )Bela nebula (Montagu, 1803) 2 3 ) ) ) ) 1 ** 6 )Mangelia attenuata (Montagu, 1803) ) 4 ) ) ) ) 1 ** 2.4 )Mangelia nuperrima (Tiberi, 1855) ) 1 ) ) ) ) ) ** 1.2 )Mangelia unifasciata (Deshayes, 1835) ) 14 ) 1 3 ) 1 ** 11.9 )Acteon tornatilis (Linne, 1758) ) ) ) ) 1 ) ) ** 1.2 )Ringicula conformis (Monterosato, 1877) ) 1 ) ) ) ) ) ** 1.2 )Weinkauffia turgidula (Forbes, 1844) ) ) ) ) 1 ) ) ** 1.2 )Philine aperta (Linne 1767) 6 ) ) 1 ) ) ) ** 3.6 +
Nucula nitidosa Winckworth, 1931 1 2 ) ) 3 ) ) ** 6 )Lembulus pellus (Linne, 1758) ) 22 1 ) 5 ) ) ** 9.5 )Anadara demiri (Piani, 1981) 5 85 3 22 21 1 110 0.19 46.4 +
Mytilus galloprovincialis (Lamarck, 1819) 3 1 ) 2 2 3 2 ** 9.5 +
Modiolus barbatus (Linne, 1758) 3 ) 6 ) 2 ) ) ** 7.1 +
Modiolus adriaticus (Lamarck, 1819) ) ) ) ) ) 1 ) ** 1.2 )Modiolarca subpicta (Cantraine, 1835) ) ) ) ) ) ) ) ) ) +
Anomia ephippium Linne, 1758 1 14 ) 158 37 2 21 0.18 25 +
Mimachlamys varia (Linne, 1758 ) ) ) ) ) ) 1 ** 1.2 )Ctena decussata (Costa, 1829) ) 1 ) ) ) ) ) ** 1.2 )Lucinella divaricata (Linne, 1758) ) ) ) ) ) 1 ) ** 1.2 )Anadontia fragilis (Philippi, 1836) ) ) ) ) ) ) 2 ** 2.4 )Mysella bidentata (Montagu, 1803) 9 ) 5 78 4 17 99 0.16 20.2 +
Acanthocardia paucicostata (Sowerby, 1841) 6 ) 2 9 7 9 11 ** 25 +
Parvicardium exiguum (Gmelin, 1791) 1 3 ) 8 ) ) 1 ** 4.8 +
Cerastoderma glaucum (Poiret, 1789) 4 1 2 40 3 16 1 ** 27.4 +
Fulvia fragilis (Forsskal, 1775) ) ) ) 1 4 ) 2 ** 6 +
Spisula subtruncata (Da Costa, 1778) 5 1 2 27 13 11 9 ** 25 +
Tellina nitida Poli, 1791 ) 2 ) 3 1 ) ) ** 4.8 +
Cinar, Katagan, Ozturk, Egemen, Ergen, Kocatas, Onen, Kirkim, Bakir, Kurt, Dagli, Kaymakci, Acik, Dogan & Ozcan Zoobentos and exotics in
Alsancak Harbor
Marine Ecology 27 (2006) 229–246 ª 2006 The Authors. Journal compilation ª 2006 Blackwell Publishing Ltd 235

belonging to Actinaria (three species, 72 individuals),
Turbellaria (three species, four individuals), Nemertini
(eight species, 184 individuals), Nematoda (one species,
1295 individuals), Oligochaeta (two species, 257 individu-
als), Polychaeta (98 species, 116,991 individuals), Phoron-
ida (two species, 191 individuals), Crustacea (40 species,
757 individuals), Mollusca (62 species, 10,432 individu-
als), Echinodermata (five species, 48 individuals), and Tu-
nicata (seven species, 12 individuals). Grab samples
included more species (211 species) than beam-trawl sam-
ples (107 species) (Table 2). A total of 80 species were
found both in grab and beam-trawl samples. This study
added 17 new records to the inventory of the marine
fauna inhabiting the coasts of Turkey (see Table 2).
Polychaeta accounted for 90% of the total number of
specimens, followed by Mollusca (8%). The dominant
species in the area were Streblospio gynobranchiata (66%
of total individuals), Corbula gibba (6.6%), Pseudopolydo-
ra antennata (5.8%), Prionospio multibranchiata (4.8%),
Heteromastus filiformis (4.3%) and Capitella capitata
(3.7%), all comprising 92% of total specimens (Table 2).
The most frequent species (all belonging to Polychaeta)
in all grab samples were S. gynobranchiata (present in
95% of the samples), P. multibranchiata (92%), H. filifor-
mis (79%), Polydora cornuta (69%), Podarkeopsis galan-
gaui (69%), Prionospio fallax (68%) and P. antennata
(67%) (Table 2).
Mollusca accounted for 87% of the total biomass
(1891 g) in grab samples, followed by Polychaeta (4%)
and Tunicata (4%). The species with the highest individ-
ual biomass were A. demiri (38% of total biomass), Ceras-
toderma glaucum (16%), Anomia ephippium (15%) and C.
gibba (12%).
Temporal variations in number of individuals, num-
ber of species, biomass, and diversity and evenness val-
ues at all stations are presented in Figs 3 and 4. The
mean number of individuals encountered at all stations
was always high in April. Stations 1 and 2 showed the
highest number of individuals. Except at station 7,
community parameters at all stations varied significantly
between sampling periods (P < 0.01, see Figs 3 and 4,
and Table 3). However, temporal changes in diversity
values at stations 1, 3, and 7 were not statistically sig-
nificant (P > 0.05). No difference was found in the
value of biomass between time and the combined effect
of time and station (Table 3).
Table 2. Continued.
Grab statıons
D% F% BT1 2 3 4 5 6 7
Tellina pulchella Lamarck, 1818 ) 2 ) ) 1 1 6 ** 7.1 +
Abra alba (Wood, 1802) 74 9 36 28 34 47 32 0.2 41.7 +
Abra prismatica (Montagu, 1808) 8 2 7 9 5 1 14 ** 26.2 +
Gouldia minima (Montagu, 1803) ) ) ) ) ) ) 1 ** 1.2 +
Dosinia exoleta (Linne, 1758) ) 1 ) ) 1 ) 2 ** 3.6 )Dosinia lupinus (Linneaus, 1758) 1 10 ) 3 7 1 4 ** 19 )Pitar rudis (Poli, 1795) ) 1 ) ) ) 1 ) ** 2.4 )Paphia rhomboides (Pennant, 1777) 2 13 ) 18 20 19 18 ** 38 +
Sphenia binghami Turton, 1822 ) ) ) ) ) 1 3 ** 2.4 )Corbula gibba (Olivi, 1792) 1749 266 396 719 1567 3624 311 6.63 86.9 +
Dentalium inaequicostatum Dautzenberg 1891 ) 48 1 ) 10 1 5 ** 21.4 +
ECHINODERMATA
Astropecten spinulosus (Philippi, 1837) ) ) ) ) ) ) ) ) ) +
Amphipholis squamata (Delle Chiaje, 1829) ) ) ) ) ) ) 3 ** 1.2 )Brissopsis lyrifera (Forbes, 1841) ) ) ) ) ) ) 42 ** 9.5 )Psammechinus microtuberculatus (Blainville, 1825) ) ) ) ) ) ) 2 ** 1.2 )Holothuroidea (sp.) ) ) ) ) ) ) 1 ** 1.2 )TUNICATA
Ascidia mentula O. F. Muller, 1776 ) ) ) ) ) ) 4 ** 1.2 )Ciona intestinalis (Linne, 1767) ) ) ) ) ) ) ) ) ) +
Botryllus schlosseri (Pallas, 1766) ) ) ) ) 1 1 ) ** 2.4 )Phallusia mammilata (Cuvier, 1815) ) ) ) ) ) ) 1 ** 1.2 )*Pyura dura Heller, 1877 ) ) ) ) ) ) ) ) ) +
*Styela canopus Stimson, 1852 ) ) ) ) ) ) 1 ** 1.2 )*Styela plicata (Lesueur, 1823) ) ) ) 2 ) ) 2 ** 2.4 +
*Species that represent new records for the Turkish fauna; **species with a dominance value lower than 0.10%.
Zoobentos and exotics in Alsancak Harbor Cinar, Katagan, Ozturk, Egemen, Ergen, Kocatas, Onen, Kirkim, Bakir, Kurt, Dagli, Kaymakci, Acik,
Dogan & Ozcan
236 Marine Ecology 27 (2006) 229–246 ª 2006 The Authors. Journal compilation ª 2006 Blackwell Publishing Ltd

The highest biomass values were generally found at sta-
tions 4 and 7, because of the presence of C. glaucum
(max. 1138 gÆm)2 at station 4 in April) and Anadara
demiri (max. 3330 gÆm)2 at station 7 in April). The lowest
biomass values were encountered at stations 1 and 3,
which were dominated by small-sized polychaete species.
The highest number of species, and the highest diver-
sity and evenness were found at station 7, the lowest at
stations 1 and 3 (Fig. 4). The diversity value was always
higher than 3 at station 7, showing a relatively undis-
turbed or recovering condition. There were large differ-
ences in these variables among sampling periods, with
September having generally lower and April higher num-
ber of species.
Based on Bray–Curtis similarity values higher than
50%, five groups of stations (A–E) can be recognized
(Fig. 5). Six species were the most responsible for the
similarity between groups, which correspond quite well to
different sampling periods (Table 4). The density of S. gy-
nobranchiata was the main factor affecting the similarity
or dissimilarity of the samples taken in the whole area
(Table 4). ANOSIM detected a significant difference
Fig. 2. Sediment and water characteristics for each grab station during the various sampling periods.
Cinar, Katagan, Ozturk, Egemen, Ergen, Kocatas, Onen, Kirkim, Bakir, Kurt, Dagli, Kaymakci, Acik, Dogan & Ozcan Zoobentos and exotics in
Alsancak Harbor
Marine Ecology 27 (2006) 229–246 ª 2006 The Authors. Journal compilation ª 2006 Blackwell Publishing Ltd 237

(P < 0.05) between September and April, and June and
April (Table 5).
Spearman’s rank correlation analysis between the mean
scores of community parameters and the environmental
variables indicated that the number of species was posi-
tively (P < 0.05) correlated with the Secchi disk depth
(qr ¼ 0.40) and negatively with the percentage of carbon
in sediment (qr ¼ )0.40); the diversity index negatively
with the ammonium concentration (qr ¼ )0.38).
A total of six exotic species, all previously known from
the area, were collected. These were S. gynobranchiata, P.
cornuta, Hydroides dianthus, H. elegans, A. demiri, and
Fulvia fragilis. The importance of these exotic species in
terms of the number of individuals and biomass at all sta-
tions and in all sampling periods is shown in Fig. 3. The
majority of individuals collected at stations near Alsancak
Harbor belonged to these species (e.g. station 3 with 92%
of the individuals). There was a weak positive correlation
between the total density of exotic species and that of
native ones (Pearson’s correlation r ¼ 0.33, P < 0.05). A.
demiri occurred patchily in the area, reaching its highest
density at station 7 (580 indÆm)2) in April and station 2
(520 indÆm)2) in January. However, weights of specimens
of A. demiri found at station 7 were higher than those
found at station 2, accounting for 93% of the mean total
biomass at station 7 in January. The contribution of exo-
tic species to total biomass values at stations 1 and 6 was
negligible (Fig. 3), except for samples taken in September
at station 6 where exotics comprised 97% of the total bio-
mass. The correlation between the total biomass value of
exotics versus natives was positive and significant,
although low (qr ¼ 0.29, P < 0.05).
The density and biomass values of the exotics at all sta-
tions and in all periods are shown in Fig. 6. The mean
Fig. 3. Temporal fluctuations in the mean
faunal densities (number of individuals per
0.1 m)2) and biomass (g wet weight per
0.1 m2) with +1 standard error. Dark bars
show the mean density and biomass of exotic
species, and white bars those of native ones.
The percentage of exotics in the total faunal
density and biomass estimated at each station
is indicated on the bars. One-way ANOVA
was used to find out if the mean scores for
density and biomass are significant or not
with regard to sampling time at each station
(*P < 0.01, **P < 0.001, ns ¼ not
significant). Ja ¼ January; A ¼ April; J ¼June; S ¼ September.
Zoobentos and exotics in Alsancak Harbor Cinar, Katagan, Ozturk, Egemen, Ergen, Kocatas, Onen, Kirkim, Bakir, Kurt, Dagli, Kaymakci, Acik,
Dogan & Ozcan
238 Marine Ecology 27 (2006) 229–246 ª 2006 The Authors. Journal compilation ª 2006 Blackwell Publishing Ltd

density and biomass values of S. gynobranchiata and P.
cornuta attained their maximum levels in April at stations
1 and 2, and gradually diminished with increasing dis-
tance from the harbor (from station 1 to 7). As station 6
is located in Pasaport Harbor, the species also had high
abundance and biomass. The other exotic polychaetes, H.
dianthus and H. elegans, occurring at all stations, except
station 3, were found on living or dead bivalve shells
embedded in muddy substrates. They were abundant at
stations 7 and 4 where shells of large bivalves A. demiri
and A. ephippium occurred. There were two density and
biomass peaks for A. demiri at stations 2 (520 indÆm)2;
680 gÆm)2 in January) and 7 (580 indÆm)2; 3330 gÆm)2 in
April). Fulvia fragilis was found only at stations 4, 5, and
Fig. 4. Temporal fluctuations in the mean
number of species, diversity index and
evenness index at each station, with +1
standard error. One-way ANOVA was used to
find out if the mean values of these
parameters are significant according to
sampling time at each station (*P < 0.01,
**P < 0.001, ns ¼ not significant).
Cinar, Katagan, Ozturk, Egemen, Ergen, Kocatas, Onen, Kirkim, Bakir, Kurt, Dagli, Kaymakci, Acik, Dogan & Ozcan Zoobentos and exotics in
Alsancak Harbor
Marine Ecology 27 (2006) 229–246 ª 2006 The Authors. Journal compilation ª 2006 Blackwell Publishing Ltd 239

7, with the highest density and biomass at stations 5
(20 indÆm)2, 2.5 gÆm)2 in April) and 7 (20 indÆm)2,
0.1 gÆm)2 in June).
Spearman’s rank correlation between abundances of the
six exotic species and abiotic variables indicated that the
number of individuals of P. cornuta was significantly
(P < 0.05) and negatively correlated with pH (qr ¼)0.42) and positively correlated with the silicate concen-
tration (qr ¼ 0.43); S. gynobranchiata was negatively cor-
related with the salinity (qr ¼ )0.39), and positively with
ammonium (qr ¼ 0.50) and silicate (qr ¼ 0.63) concen-
trations; A. demiri was negatively correlated with the per-
centage of carbon in sediment (qr ¼ )0.39).
No correlation was estimated between densities of each
exotic species and those of natives while abundance of H.
dianthus was well correlated with that of A. demiri (qr ¼0.63, P < 0.05), and abundance of H. elegans with that of
A. ephippium (qr ¼ 0.65, P < 0.05).
The reproductive periods of S. gynobranchiata and P.
cornuta occurred throughout the year, with maximum
oocyte diameter being calculated as 120 lm (mean ± SE ¼91 lm ± 2.89, n ¼ 15) in January for S. gynobranchiata
and as 60 lm (53 lm ± 1.05, n ¼ 20) for P. cornuta.
Oocytes within coelomic cavities of H. dianthus (max.
diameter ¼ 65 lm, mean ± SE ¼ 56 lm ± 1.35, n ¼ 15)
Table 3. Result of two-way ANOVA of the community parameters.
sources of variation df
number of
species
(C ¼ 0.346**)
number of
individuals
(C ¼ 0.361***)
evenness
index (J¢)(C ¼ 0.323***)
diversity
index (H¢)(C ¼ 0.152*)
biomass
(C ¼ 0.811***)
MS P MS P MS P MS P MS P
stations 6 1408.3 *** 3,119,553 *** 0.099 *** 6.83 *** 7092.4 *
post hoc test All 7 7 3, 7 7
sampling periods 3 1844.1 *** 40,452,695 *** 0.093 *** 1.03 *** 5196.8 ns
post hoc test All A A, S J, S ns
stations · periods 18 67.3 ns 2,153,805 *** 0.122 *** 1.43 *** 2122.6 ns
residual 56 38 618,929 0.010 0.14 2445.4
*P < 0.05; **P < 0.01; ***P < 0.001.
MS ¼ mean square; df ¼ degrees of freedom; C ¼ Cochrane’s test; ns ¼ not significant.
Post hoc test (Tukey’s HSD) shows the stations and sampling periods which determine statistical differences.
Fig. 5. Multidimensional scaling plot showing the similarity among
temporal samples. Associations were determined according to the
results of Bray–Curtis similarity analysis (each association has a similarity
higher than 50%). Ja ¼ January; A ¼ April; J ¼ June; S ¼ September.
Table 4. Species contributing to similarity among sampling periods,
and their average similarity.
average similarity
September June April January
29 MD 32 MD 50 MD 40 MD
Prionospio
multibranchiata
10 1220 4 1160 5 4300 12 2320
Pseudopolydora
antennata
– – 12 4340 5 5820 – –
Streblospio
gynobranchiata
70 9270 45 10770 72 74390 78 27730
Heteromastus
filiformis
11 1420 19 4690 – – – –
Capitella capitata – – – – 4 4940 – –
Corbula gibba – – 12 2030 6 8730 – –
Bold numbers indicate the highest score of species contribution in
each assemblage.
MD ¼ mean density (individuals Æm)2).
Table 5. The result of ANOSIM testing for differences among samp-
ling periods.
months R-value P-value
September versus June 0.07 ns
September versus April 0.43 *
September versus January 0.09 ns
June versus April 0.36 *
June versus January 0.16 ns
April versus January 0.23 ns
*P < 0.05; ns ¼ not significant.
Zoobentos and exotics in Alsancak Harbor Cinar, Katagan, Ozturk, Egemen, Ergen, Kocatas, Onen, Kirkim, Bakir, Kurt, Dagli, Kaymakci, Acik,
Dogan & Ozcan
240 Marine Ecology 27 (2006) 229–246 ª 2006 The Authors. Journal compilation ª 2006 Blackwell Publishing Ltd

and H. elegans (max. diameter ¼ 50 lm, mean ± SE ¼40.4 lm ± 0.96, n ¼ 15) were only found in June. No
reproductive individuals of A. demiri and F. fragilis were
observed.
Discussion
Analysis of benthic samples showed that the investigated
area has a diverse benthic fauna with a total number of
231 species. The innermost part of Izmir Bay was previ-
ously known to be one of the most polluted regions in
Turkey (Geldiay & Ergen 1972; Ergen 1979; Geldiay et al.
1979; Kocatas et al. 1987; Kocak et al. 1999). Azoic condi-
tions (Dogan et al. 2005; based on the material collected
in 1995 and 1996) and odorous biogenic sulfurous gases
(Muezzinoglu et al. 2000) were reported in the area in
the 1980s and 1990s. The muddy bottom of the area was
previously dominated only by opportunistic species such
as Capitella capitata, Malacoceros fuliginosus, Prionospio
fallax, and Ophiodromus pallidus (Ergen 1979; Dogan
et al. 2005). However, an improvement in the environ-
mental quality of the bay was anticipated once a new
wastewater treatment plant came into operation in early
2000. Ergen et al. (2006) examined the benthic materials
collected periodically in the area between 1997 and 2002
and showed such an improvement in 2002. In this year,
no azoic condition occurred in the area, even in summer,
and polychaetes other than pollution indicators were also
encountered. The present study also observed several spe-
cies such as Syllis garciai, Owenia fusiformis, Squilla man-
tis, Melicertus kerathurus, and Amphipholis squamata in
and around Alsancak Harbor, which are sensitive to
Fig. 6. Temporal fluctuations of the mean values of density and biomass of exotic species found at stations with +1 standard error (Ja ¼ January;
A ¼ April; J ¼ June; S ¼ September).
Cinar, Katagan, Ozturk, Egemen, Ergen, Kocatas, Onen, Kirkim, Bakir, Kurt, Dagli, Kaymakci, Acik, Dogan & Ozcan Zoobentos and exotics in
Alsancak Harbor
Marine Ecology 27 (2006) 229–246 ª 2006 The Authors. Journal compilation ª 2006 Blackwell Publishing Ltd 241

intense pollution. The number of species near Alsancak
Harbor (stations 1–5) ranged between seven (station 3, in
September) and 70 (station 2, April). The station farthest
from Alsancak Harbor (station 7) had a rich faunal com-
ponent, comprised of 107 species in April. Kocatas et al.
(1987) reported that 20 zoobenthic species inhabited the
area surrounding Alsancak Harbor in 1974; however, only
nine species were recorded in 1983. Ergen et al. (2006)
found azoic conditions near Alsancak Harbor (station 5
in the present study) every summer (except for 2002).
The number of species found at this station in our study
ranged from 17 (January) to 41 (April). In this survey, we
reported 231 soft-bottom benthic organisms in the inner
part of Izmir Bay, whereas only 21 species were reported
by Kocatas (1980), 23 species by Palaz (1989), 14 poly-
chaete species by Ergen (1992) and nine species by Dogan
et al. (2005). This result clearly shows that there has been
an environmental improvement, possibly because of the
beneficial effect of the treatment plant in reducing pol-
lution and making the area more favorable, even in sum-
mer, for the settlement of benthic organisms.
The chemical analyses of water and sediment samples
during this study also showed that there was a decline in
the concentrations of nitrite, nitrate, and ammonium over
time, especially after the establishment of the wastewater
treatment plant in 2000. The concentration of ammo-
nium in surface water, which was the dominant nitrogen
source in the area, was reported to reach maximum levels
of 42.4 lgatÆl)1 in 1987 (Uysal & Yaramaz 1987),
65.7 lgatÆl)1 in 2000 (Kaymakci et al. 2000), 50 lgatÆl)1
in 2001 (Kontas et al. 2004) and 20.5 lgatÆl)1 (mean ± -
SE ¼ 3.3 ± 0.8 lgatÆl)1) in 2004 (this study). However,
the concentration of phosphate phosphorus, which is an
important source of eutrophication in the bay (Kucuksez-
gin et al. 2001), remained relatively unchanged over the
years, possibly because of the effects of the polluted
creeks’ discharge into the area; max. 5.58 lgatÆl)1 in 1987
(Uysal & Yaramaz 1987), 10 lgatÆl)1 in 2001 (Kontas
et al. 2004) and 5.9 lgatÆl)1 in 2004 (this study). Simi-
larly, Kontas et al. (2004) pointed out that the capacity of
the wastewater plant was sufficient for removal of nitro-
gen from wastes, whereas it was inadequate for the
removal of phosphate. Similar results were also seen in
terms of the percentage of carbon in sediments in that
the highest levels were found in this present study (max.
27%) when compared with the previous study undertaken
by DEU (1997) (7%). The main reason for this difference
might be that in this study, the harbor environment
where the water circulation is weaker and the sedimenta-
tion is higher was also sampled.
In the present study, a clear temporal trend in the
abundance of benthic organisms, with peaks during April,
followed by a striking decline throughout June and
September was determined. At all stations (except for sta-
tion 7), the mean values of community parameters varied
significantly, showing the effects of sampling time, especi-
ally in the more polluted areas. In April, the benthic
assemblages were characterized by having a high number
of juveniles and a high percentage of reproducing speci-
mens belonging to the dominant species such as Streblos-
pio gynobranchiata, Capitella capitata, Pseudopolydora
antennata, Prionospio multibranchiata, and Corbula gibba.
Sarda et al. (1999) found a similar result in the abun-
dance and biomass of macroinfaunal assemblages at sub-
littoral stations of the Bay of Blanes, with peaks during
spring, and concluded that the reproductive cycle of most
species was the main factor affecting density fluctuations
in time. The temporal dynamics of the benthic fauna was
reported to be influenced by the availability of food and
the presence of predators (Sarda et al. 1995). However,
Oug (2000) examined the soft-bottom macrofauna from
fjords of northern Norway during 1979–1982 and repor-
ted that species number and densities varied stochastically
with peak values occurring at different times of the year.
He concluded that the lack of a temporal pattern was
mainly because of the stable physical conditions and the
irregular nutrient input in the fjord basin. This assump-
tion could also be applied to this study because of the
fact that community parameters show significant tem-
poral differences at stations in or around Alsancak Har-
bor (stations 1–6), where marked fluctuations in
environmental variables were observed in time. In con-
trast, the community parameters at station 7, which is
relatively far from pollution sources, did not show any
significant differences. In addition, the mean score for the
diversity index was always <3 at stations 1–6 (except for
station 2 in January), whereas it was always >3 at station
7. Low values of the diversity index indicate that the
majority of the study area could still be considered as an
impacted environment.
The community parameters were also affected by the
environmental factors. For example, the number of spe-
cies was negatively correlated with percentage of carbon
in sediment and positively correlated with transparency
(Secchi disk depth). A negative correlation was found
between diversity values and the ammonium concentra-
tion, which had higher values in January at stations near
Alsancak Harbor. Bourget et al. (2003) clearly showed
that water temperature and transparency, plus in some
cases primary production, were the variables having a sig-
nificant influence on the total biomass of epibenthic
assemblages in the Gulf of St. Lawrence. Je et al. (2003)
showed the importance of DDT, low aromatic hydrocar-
bons, lindane and grain size in the distribution of benthic
organisms in Vancouver Harbor. Similarly, pollution gra-
dient and granulometric parameters were reported to be
Zoobentos and exotics in Alsancak Harbor Cinar, Katagan, Ozturk, Egemen, Ergen, Kocatas, Onen, Kirkim, Bakir, Kurt, Dagli, Kaymakci, Acik,
Dogan & Ozcan
242 Marine Ecology 27 (2006) 229–246 ª 2006 The Authors. Journal compilation ª 2006 Blackwell Publishing Ltd

the main factors affecting polychaete distribution in a
harbor at Ceuta located in the Strait of Gibraltar (Guerra-
Garcıa & Garcıa-Gomez 2004). In the present study, the
significant difference in community parameters between
sampling periods and stations suggests that benthic com-
munities occurring in and near Alsancak Harbor were
dynamic and greatly affected by both environmental fac-
tors, and biological and ecological features of organisms.
Soft sediments in and around Alsancak Harbor have
been exposed to an intense invasion by two spionid poly-
chaetes, S. gynobranchiata and Polydora cornuta, which
were previously known from the area (Cinar et al. 2005).
The morphological, biometric, and reproductive features
of populations of these two species in this area were given
by Cinar et al. (2005). The importance of these two spe-
cies was high, they constituted more than 90% of faunal
populations in the majority of samples collected at sta-
tions 1–5. However, their biomass was low because of
their small body sizes (largest 8.3 mm long). Streblospio
gynobranchiata was previously reported not only from the
western Atlantic Ocean in muddy sand at a depth of 1 m,
but also in estuarine areas (Rice & Levin 1998). The first
appearance of this species in the inner part of Izmir Bay
was in 2002 when the azoic conditions disappeared. The
population density of this species was reported to be very
low in its type locality; while its density in this survey
reached up to 60,480 indÆm)2 at station 1 in April.
Organically polluted sediments of the inner part of Izmir
Bay seem to provide a suitable habitat for settlement and
growth. The reproductive period for the species in the
area continued throughout the year. This species together
with P. cornuta, Anadara demiri and Fulvia fragilis might
have been introduced to the area via ballast water. Evi-
dence for this includes: (i) a huge gap in their distribu-
tional patterns, (ii) their absence from previous material
taken from the area, which has been continuously monit-
ored since the 1970s, and (iii) the extensive maritime traf-
fic present from different parts of the world.
Polydora cornuta, which is apparently a cosmopolitan
species (Radashevsky 2005), was first reported from the
Valencia Harbor (Spain) in the Mediterrranean Sea (Tena
et al. 1991), and subsequently from Izmir Bay (Cinar
et al. 2005) and the Black Sea (Radashevsky 2005). Previ-
ous studies performed in the Mediterranean Sea cited P.
ciliata as one of the dominant species from polluted envi-
ronments (Ergen 1979; Magni et al. 2004). As taxonomic
confusion exists for species of Polydora, some previous
records of P. ciliata from the polluted soft sediments of
the Mediterranean Sea might indeed be P. cornuta. This
species was also one of the dominant species in the area
near Alsancak Harbor and reproduced continuously
throughout the year, suggesting that it has acclimatized
well in the region.
Hydroides dianthus and H. elegans were found to be
encrusted on shells of the bivalves A. ephippium and A.
demiri, and reached their maximum densities at station 7
where these bivalve species occurred. They dominate arti-
ficial hard substrates in polluted environments of the
Mediterranean Sea (Kocak et al. 1999).
The exotic bivalve A. demiri has been known from
Izmir Bay since the 1970s (Demir 1977). It was previously
found at densities of 300 indÆm)2 on gray mud,
30 indÆm)2 on black mud (Demir 1977), and 180 indÆm)2
in the polluted bottom of Thessaloniki Bay at 15–22 m
(Zenetos 1994). Our findings showed that this species
preferred semi-polluted bottoms and was negatively cor-
related with the percentage of carbon in sediment (qr ¼)0.39).
Fulvia fragilis, which is a Lessepsian bivalve, was
recently reported from Izmir Bay (Ozturk & Poutiers
2005) and is known both in the eastern and western
Mediterranean Sea (Ozturk & Poutiers 2005). These
authors emphasized that its introduction into Izmir Bay
was most probably via ballast waters, and found a dense
population at station 4 collected using a beam trawl in
July 2003.
The impact of exotics on benthic community structure
in the inner part of Izmir Bay was clear, contributing to
the majority of specimens and biomass in all sampling
periods. Streblospio gynobranchiata is the key species
responsible for the similarity/dissimilarity levels of benthic
samples in all periods. The occurrence and abundance of
this species do not seem to interfere with the pattern of
distribution and abundance of other species. Ranasinghe
et al. (2005) outlined a similar finding in a southern Cali-
fornia embayment in that abundance of exotics was posi-
tively correlated with the abundance and richness of other
species and that exotics did not have a negative impact
on the overall community, possibly because of the bio-
genic structures built by them. The wealth of detritus
material in the inner part of Izmir Bay might hinder any
possible competition between species and the still pol-
luted conditions restrict the settlement of pollution-sensi-
tive species, which have greater competitive abilities. This
was more evident at station 7, where exotic spionids had
very low abundances as they were probably out-competed
from the area by the natives.
Extreme anthropogenic disturbance may create a mis-
match between the traits of native species and their
environment, so that some invading species are as well
adapted to the altered environment as are the compet-
ing native species (Byers 2002). These disturbances
increase invasion success not only by creating new
microhabitats, introducing propagules, and decreasing
the populations of native species that can resist inva-
sion, but also by weakening the ‘per capita’ ability of
Cinar, Katagan, Ozturk, Egemen, Ergen, Kocatas, Onen, Kirkim, Bakir, Kurt, Dagli, Kaymakci, Acik, Dogan & Ozcan Zoobentos and exotics in
Alsancak Harbor
Marine Ecology 27 (2006) 229–246 ª 2006 The Authors. Journal compilation ª 2006 Blackwell Publishing Ltd 243

native biota to resist invaders (Kocak et al. 1999; Byers
2002). A recent study of macrofouling organisms dis-
covered many more species in a polluted rather than a
non-polluted marina, and that the exotic serpulid H.
elegans, which made up 65% of the population in the
polluted marina, was occasionally found in the non-
polluted marina (Kocak et al. 1999). A healthy pristine
community represents a natural barrier to bio-invasion
and thus environmental conservation plays a fundamen-
tal role in preventing further spread of non-indigenous
species (Occhipinti-Ambrogi & Savini 2003). Holeck
et al. (2004) also demonstrated that previous introduc-
tions of exotics can facilitate the introduction and suc-
cess of subsequent arrivals. Therefore, halting the
introduction of any one exotic (e.g. through control of
ballast water discharge and other vectors) can inhibit a
potentially larger number of other invaders whose suc-
cess and impact are magnified by that species.
The present study shows that the pollution intensity in
the area greatly diminished after the establishment of a
wastewater plant, facilitating the settlement of many
native and exotic species in the area. The exotics invaded
the soft sediments near Alsancak Harbor and played an
important role in the benthic community structure.
Conclusions
The soft-bottom samples collected from several stations
(seven with van Veen grab and three with beam trawl)
in and around Alsancak Harbor, in various periods
between January and September 2004, included a total
of 231 zoobenthic species, 17 of which are new records
for the Turkish coast. The collection included six exotic
species: Streblospio gynobranchiata, Polydora cornuta,
Hydroides dianthus, H. elegans, Anadara demiri and Ful-
via fragilis which were previously known from the area.
The area was also highly colonized by opportunistic
species such as Capitella capitata and Corbula gibba. In
general, polychaetes were the dominant benthic compo-
nents in the area, accounting for 90% of the total
faunal populations, followed by mollusks (8%). The
most dominant species in the area was the exotic spe-
cies S. gynobranchiata (66% of the total individuals),
which contributed extensively to the similarity or dissi-
milarity of stations in time. Community parameters
varied significantly between periods and stations. April
was characterized by relatively high species richness and
dense populations of some species. The importance of
exotics in benthic fauna diminished when the distance
from the harbor increased. Community and chemical
analyses showed that, where azoic conditions had been
previously reported, recovery had occurred after the
establishment of a wastewater treatment plant in the
area in 2000. However, relatively low values of diversity
(H¢ < 3) at stations near Alsancak Harbor indicated
that the area still suffers from the pollution impact.
Acknowledgements
We are much indebted to Dr. Christer Erseus (Sweden)
for the identification of oligochaetes, to Dr. Dimitris Vaf-
idis (Greece) for the identification of actinarians, to Drs.
Juan Junoy (Spain) and Jon L. Norenberg (USA) for the
identification of nemerteans, to Dr. Duane Hope (USA)
for the identification of the nematode Enoplus meridional-
is, to Dr. Alfonso Ramos (Spain) for the identification of
ascidian species, to two anonymous referees for their con-
structive comments on the manuscript, and to personnel
of R/V EGESUF for their help in collecting the benthic
material. This work has been financially supported by Sci-
entific Research Projects of Ege University (Project num-
ber: 03 SUF 005).
References
Bourget E., Ardisson P.-L., Lapointe L., Daigle G. (2003) Envi-
ronmental factors as predictors of epibenthic assemblage
biomass in the St. Lawrence system. Estuarine, Coastal and
Shelf Science, 57, 641–652.
Byers J.E. (2002) Impact of non-indigenous species on natives
enhanced by anthropogenic alteration of selection regimes.
Oikos, 97, 449–458.
Cinar M.E., Ergen Z. (2005) Lessepsian migrants expanding
their distributional ranges; Pseudonereis anomala (Polychaet-
a: Nereididae) in Izmir Bay (Aegean Sea). Journal of the
Marine Biological Associations of the United Kingdom, 85,
313–321.
Cinar M.E., Ergen Z., Dagli E., Petersen M.E. (2005) Alien spe-
cies of spionid polychaetes (Streblospio gynobranchiata and
Polydora cornuta) in Izmir Bay, eastern Mediterranean. Jour-
nal of Marine Biological Associations of the United Kingdom,
85, 821–827.
Clarke K.R., Warwick R.M. (2001) Change in marine communi-
ties: an approach to statistical analysis and interpretation, 2nd
edition. PRIMER–E, Plymouth.
Cohen A.N., Carlton J.T. (1998) Accelerating invasion rate in
a highly invaded estuary. Science, 279, 555–557.
Demir M. (1977) On the presence of Arca (Scapharca) amygd-
alum Philippi, 1847 in the harbour of Izmir, Turkey. Istan-
bul Universitesi Fen Fakultesi Mecmuasi Serisi, 42, 197–202.
DEU (1997) Izmir Korfezi 1994–1995 Deniz Arastirmalari Yillik
Raporu. Deniz Bilimleri ve Teknolojisi Enstitusu Projesi, In-
ciralti: 98 pp.
Dogan A., Cinar M.E., Onen M., Ergen Z., Katagan T. (2005)
Seasonal analysis of soft bottom zoobenthic communities in
polluted and unpolluted areas of Izmir Bay (Aegean Sea).
Senckenbergiana Maritima, 35, 133–145.
Zoobentos and exotics in Alsancak Harbor Cinar, Katagan, Ozturk, Egemen, Ergen, Kocatas, Onen, Kirkim, Bakir, Kurt, Dagli, Kaymakci, Acik,
Dogan & Ozcan
244 Marine Ecology 27 (2006) 229–246 ª 2006 The Authors. Journal compilation ª 2006 Blackwell Publishing Ltd

Elton C.S. (1958) The ecology of invasions by animals and
plants. The University of Chicago Press, Chicago: 181 pp.
Ergen Z. (1976) Izmir Korfezi ve civari poliketleri uzerine tak-
sonomik ve ekolojik arastirmalar. Ege Universitesi Fen Fakul-
tesi Ilmi Raporlar Serisi, 209, 1–73.
Ergen Z. (1979) _Izmir Korfezi’nin Pollusyonunun Poliket
(Polychaeta-Annelida) Populasyonunun Dagilimi Uzerine
Etkileri. TUJJB Yayinlari, 11, 77–82.
Ergen Z. (1992) The latest status of Polychaeta in the soft sub-
strate of Izmir Bay. Rapport de la Commission International
Exploration de la Mer Mediterranee, 33,36.
Ergen Z., Cinar M.E., Dagli E., Kurt G. (2006) Seasonal
dynamics of soft-bottom polychaetes in Izmir Bay (Aegean
Sea, eastern Mediterranean). Scientia Marina, in press.
Erguvanli K. (1995) Muhendislik Jeolojisi. Sec Yayin Dagitim, 4.
Baski, _Istanbul: 590 pp.
Gaudette H.E., Flight W.R., Toner L., Folger W. (1974) An in-
expensive titration method for the determination of organic
carbon in recent sediments. Journal of Sedimentary Petrology,
44, 249–253.
Geldiay R., Ergen Z. (1972) Deniz biyolojisi arastirma labo-
ratuvari onundeki bentik poliket faunasi uzerinde on musa-
hedeler. Ege Universitesi Fen Fakultesi _Ilmi Raporlar Serisi,
134, 1–10.
Geldiay R., Ergen Z., Kocatas A. (1979) Some effects of
pollution on the benthic communities of the soft subtrate in
the Gulf of Izmir (Turkey). Rapport de la Commission
International Exploration de la Mer Mediterranee, 25/26,
193–194.
Guerra-Garcıa J.M., Garcıa-Gomez J.C. (2004) Polychaete
assemblages and sediment pollution in a harbour with two
opposing entrances. Helgolander Marine Research, 58, 183–
191.
Holeck K.T., Mills E.L., McIsaac H.J., Dochoda M.R., Colautti
R.I., Ricciardi A. (2004) Bridging troubled waters: biological
invasions, transoceanic shipping, and the Laurentian Great
Lakes. BioScience, 54, 919–929.
Je J.G., Belan T., Levings C., Koo B.J. (2003) Changes in ben-
thic communities along a presumed pollution gradient in
Vancouver Harbour. Marine Environmental Research, 57,
121–135.
Kaymakci A., Sunlu U., Egemen O. (2000) Assessment of
nutrient pollution caused by land based activities in Izmir
Bay, Turkey. Meeting on Interdependency between Agriculture,
Urbanization: Conflicts on Sustainable Use of Soil, Water,
Tunisia: 41–49.
Kocak F., Ergen Z., Cinar M.E. (1999) Fouling organisms and
their developments in a polluted and an unpolluted marina
in the Aegean Sea (Turkey). Ophelia, 50, 1–20.
Kocatas A. (1980) Evolution cyclique du Benthos dans les zones
de pollution du golfe d’Izmir (Turquie). Ves Journees Etude
Pollutions. Rapport de la Commission International Explora-
tion de la Mer Mediterranee, Cagliari: 643–648.
Kocatas A., Ergen Z., Katagan T. (1987) Changes in the
benthic communities due to various pollutants in Izmir Bay
(Turkey). FAO Fisheries Report Number, 352(Suppl.), 112–
122.
Kontas A., Kucuksezgin F., Altay O., Uluturhan E. (2004)
Monitoring of eutrophication and nutrient limitation in
Izmir Bay (Turkey) before and after wastewater treatment
plant. Environmental International, 29, 1057–1062.
Kucuksezgin F., Kontas A., Altay O., Uluturhan E. (2001) Eutro-
phication in Izmir Bay (eastern Aegean): nutrient limitation
and monitoring of long-term effects. Rapport de la Commission
International Exploration de la Mer Mediterranee, 36, 397.
Magni P., Micheletti S., Casu D., Floris A., De Falco G., Cas-
telli A. (2004) Macrofaunal community structure and distri-
bution in a muddy coastal lagoon. Chemistry and Ecology,
20, 397–409.
Morello E., Solustri C. (2001) First record of Anadara demiri
(Piani, 1981) (Bivalvia: Arcidae) in Italian waters. Bollettino
Malacologico, 37, 231–234.
Muezzinoglu A., Sponza D., Koken I., Alparslan N., Akyarli A.,
Ozture N. (2000) Hydrogen sulfide and odour control in
Izmir Bay. Water, Air and Soil Pollution, 123, 245–257.
Occhipinti-Ambrogi A., Savini D. (2003) Biological invasions
as a component of global change in stressed marine ecosys-
tems. Marine Pollution Bulletin, 46, 542–551.
Oug E. (2000) Soft-bottom macrofauna in the high-latitude
ecosystem of Balsfjord, northern Norway: species composi-
tion, community structure and seasonal variability. Sarsia,
85, 1–13.
Ozturk B., Poutiers J.-M. (2005) Fulvia fragilis (Bivalvia:
Cardiidae): a Lessepsian mollusk species from Izmir Bay
(Aegean Sea). Journal of Marine Biological Associations of the
United Kingdom, 85, 351–356.
Palaz M. (1989) _Izmir Korfezi’nin ic bolgesindeki zoobentozun
dinamikleri. Yuksek Lisans Tezi, 9 Eylul Universitesi: 37 pp.
Parsons T.R., Matia Y., Lalli C.M. (1984) A manual of chemical
and biological methods for seawater analysis. Pergamon Press,
New York: 173 pp.
Quatrefages A. (1865) Histoire naturelle des Anelees d’eau
douce et marin. Paris, 1, 1–588.
Radashevsky V.I. (2005) On adult and larval morphology of
Polydora cornuta Bosc, 1802 (Annelida: Spionidae). Zootaxa,
1064, 1–24.
Ranasinghe J.A., Mikel T.K., Velarde R.G., Weisberg S.B.,
Montagne D.E., Cadien D.B., Dalkey A. (2005) The
prevalence of non-indigenous species in southern Califor-
nia embayments and their effects on benthic macroinver-
tebrate communities. Biological Invasions, 7, 679–686.
Rice S.A., Levin L.A. (1998) Streblospio gynobranchiata, a
new spionid polychaete species (Annelida: Polychaeta)
from Florida and the Gulf of Mexico with an analysis of
phylogenetic relationships within the genus Streblospio.
Proceedings of the Biological Society of Washington, 111,
694–707.
Sarda R., Martin D., Pinedo S., Dueso A., Cardell M.J. (1995)
Seasonal dynamics of shallow soft-bottom communities in
western Mediterranean. In: Eleftheriou A., Ansell A.D.,
Cinar, Katagan, Ozturk, Egemen, Ergen, Kocatas, Onen, Kirkim, Bakir, Kurt, Dagli, Kaymakci, Acik, Dogan & Ozcan Zoobentos and exotics in
Alsancak Harbor
Marine Ecology 27 (2006) 229–246 ª 2006 The Authors. Journal compilation ª 2006 Blackwell Publishing Ltd 245

Smith C.J. (Eds), The biology and ecology of shallow coastal
waters. Olsen and Olsen, Fredensborg: 191–198.
Sarda R., Pinedo S., Martin D. (1999) Seasonal dynamics of
macroinfaunal key species inhabiting shallow soft-bottoms
in the Bay of Blanes (NW Mediterranean). Acta Oecologica,
20, 315–326.
Shea K., Chesson P. (2002) Community ecology theory as a
framework for biological invasions. Trends in Ecology and
Evolution, 17, 170–176.
Tena J., Capaccioni-Azzati R., Porras R., Torres-Gavila F.J.
(1991) Cuatro especies de poliquetos nuevas para las costas
Mediterraneas en los sedimentos del antepuerto de Valencia.
Miscellania Zoologica, 15, 29–41.
UNEP (1994) Integrated management study for the area of
Izmir. MAP Technical Reports Series No. 84, United
Nations Environmental Programme, Split: 1–130. pp.
Uysal H., Yaramaz O. (1987) Izmir Korfezi’nin farkli
bolgelerinde bazi polluantlarin mukayeseli olarak
arastirilmasi. CEVRE 87 Symposium, EBSO, Izmir.
Vitousek P.M., D’Antonio C.M., Loope L.L., Rejmanek M.,
Westbrooks R. (1997) Introduced species: a significant com-
ponent of human-caused global change. New Zealand Jour-
nal of Ecology, 21, 1–16.
Wonham M.J., Carlton J.T. (2005) Trends in marine biolo-
gical invasions at local and regional scales: the Northeast
Pacific Ocean as a model system. Biological Invasions, 7,
369–392.
Zenetos A. (1994) Scapharca demiri (Piani, 1981): first finding
in the North Aegean Sea. La Conchiglia, 271, 37–38.
Zenetos A., Gofas S., Russo G.F., Templado J. (2003) CIESM
atlas of exotic species in the Mediterranean. Volume 3. Mol-
luscs. CIESM Publishers, Monaco: 376 pp.
Zibrowius H. (1971) Les especes Mediterraneennes du genre
Hydroides (Polychaete Serpulidae) remarques sur le preten-
du polymorphisme de Hydroides uncinata. Tethys, 2, 691–
745.
Zoobentos and exotics in Alsancak Harbor Cinar, Katagan, Ozturk, Egemen, Ergen, Kocatas, Onen, Kirkim, Bakir, Kurt, Dagli, Kaymakci, Acik,
Dogan & Ozcan
246 Marine Ecology 27 (2006) 229–246 ª 2006 The Authors. Journal compilation ª 2006 Blackwell Publishing Ltd




















