
TAVŞANLARDA BALON HASARI SONRASI DAMAR DÜZ KAS HÜCRELERİNDE TELOMER DEVAMLILIĞI VE OKSİDAN...
179
i T.C. EGE ÜNİVERSİTESİ SAĞLIK BİLİMLERİ ENSTİTÜSÜ TAVŞANLARDA BALON HASARI SONRASI DAMAR DÜZ KAS HÜCRELERİNDE TELOMER DEVAMLILIĞI VE OKSİDAN STRESİN ROLÜ Eczacılık Fakültesi Farmakoloji Anabilim Dalı Doktora Tezi Uzm. Ecz. Gönen ÖZŞARLAK SÖZER Danışman: Prof. Dr. Zeliha KERRY İZMİR 2008
Transcript of TAVŞANLARDA BALON HASARI SONRASI DAMAR DÜZ KAS HÜCRELERİNDE TELOMER DEVAMLILIĞI VE OKSİDAN...

i
T.C. EGE ÜNİVERSİTESİ
SAĞLIK BİLİMLERİ ENSTİTÜSÜ
TAVŞANLARDA BALON HASARI SONRASI
DAMAR DÜZ KAS HÜCRELERİNDE TELOMER DEVAMLILIĞI VE OKSİDAN STRESİN ROLÜ
Eczacılık Fakültesi Farmakoloji Anabilim Dalı Doktora Tezi
Uzm. Ecz. Gönen ÖZŞARLAK SÖZER
Danışman:
Prof. Dr. Zeliha KERRY
İZMİR 2008

ii

iii
DEĞERLENDİRME KURULU ÜYELERİ
Başkan : Prof.Dr. Zeliha KERRY
.......................................................
Üye : Prof.Dr.Aslı ÖZER
.......................................................
Üye : Prof.Dr.Zeki TOPÇU .......................................................
Üye : Doç Dr. Metiner TOSUN ........................................................
Üye : Prof.Dr.Gülgün OKTAY ........................................................
Doktora Tezinin kabul edildiği tarih: .............................................

iv
ÖNSÖZ
Tüm akademik yaşamım boyunca her zaman desteğini gördüğüm anabilim dalı başkanımız değerli hocam Prof. Dr. Aslı Özer’e,
Bilimsel gelişimime her zaman destek olup yön veren, istediğim konuda çalışmam için beni teşvik eden ve gerçekleşmesi için sabırla ve özveri ile bütün imkanları sunan, her zaman manevi
desteğini hissettiğim değerli danışman hocam Prof. Dr. Zeliha Kerry’ye
Beni telomer dünyası ile tanıştıran, ve bilmediğim moleküler biyoloji dünyasında bana ışık tutan ve yön veren, laboratuvarının tüm olanaklarını kullanmamı sağlayan ve her zaman bilgi ve
deneyimlerini paylaşan 2. danışmanım, değerli hocam Prof. Dr. Zeki Topçu’ya,
Balon anjiyoplasti modelini bana öğreten, bilgi ve deneyimlerini paylaşan, modelin oturtulmasında ve sonrasında her zaman sonsuz yardımlarını gördüğüm değerli hocam Prof. Dr.
İsmail Oran’a,
Benimle her zaman bilgi ve deneyimlerini paylaşıp, tezimin her aşamısında katkıları ile beni motive eden değerli hocam Doç. Dr. Metiner Tosun’a,
Balon anjiyoplasti operasyonları sırasında bana yardım eden sevgili arkadaşım Göksel Gökçe’ye ve gerektiğinde yardımıma koşan diğer tüm hocalarım ve arkadaşlarıma,
Manevi destekleri ile her zaman yanımda olan sevgili aileme, ve her zaman sonsuz sabır ve anlayışıyla yanımda olan, bu zor sürecin kolaylaşmasını sağlayan sevgili eşim Naci Sözer’e
Teşekkürlerimi sunarım.
İzmir, 2008 Gönen Özşarlak Sözer

v
İÇİNDEKİLER
1. GİRİŞ VE GENEL BİLGİLER 1
1.1. GİRİŞ 1
1.2. GENEL BİLGİLER 3
1.2.1. Ateroskleroz 3
1.2.1.1. Tanım 3
1.2.1.2. Ateroskleroz Patojenezi 4
1.2.2. Vasküler Proliferasyon ve Ateroskleroz 7
1.2.3. Deneysel İntimal Kalınlaşma Modelleri 9
1.2.3.1. Deneysel Modellere Genel Bakış 9
1.2.3.2. Perkütanöz Tranlüminal Koroner Anjiyoplasti (PTCA) 13
1.2.4. Telomerler 17
1.2.4.1. Telomer Devamlığının Korunması 19
1.2.4.2. Telomer Devamlılığı ve Apoptosis 21
1.2.5. Telomeraz 22
1.2.5.1. Telomerazın RNA Komponenti 23
1.2.5.2. Telomerazın Katalitik Komponenti 24
1.2.5.3. Telomerazın Regülasyonu 25
1.2.5.4. Telomeraz İnhibitörleri 26
1.2.5.5. Telomeraz ile Etkileşen Bileşikler 26
1.2.6. Alternatif Telomer Uzaması (ALT) 29
1.2.7. Oksidatif Stres ve Reaktif Oksijen Türleri 29

vi
1.2.8. Reaktif Oksijen Türleri ve Hücresel Kaynakları 30
1.2.9. Reaktif Oksijen Türlerinin Düzenlenmesinde Rol Alan Oksidan Ve Antioksidan
Enzim Sistemleri 32
1.2.9.1. Oksidan Enzim Sistemleri 32
1.2.9.1.1. NAD(P)H Oksidaz 32
1.2.9.1.2. Ksantin Oksidaz 33
1.2.9.1.3. Endoteliyel Nitrik Oksit Sentaz 33
1.2.9.2. Antioksidan Enzim Sistemleri 34
1.2.9.2.1. Süperoksit Dismutaz (SOD) 34
1.2.9.2.2. Glutatyon Peroksidaz 34
1.2.9.2.3. Katalaz 35
1.2.9.2.4. Tiyoredoksin Reküktaz 36
1.2.10. Oksidan Stres ve Kardiyovasküler Hastalıklar 36
1.2.10.1. Restenoz ve Serbest Oksijen Radikalleri 39
1.2.11. Oksidan Stres ve Telomerler 40
1.2.12.Glutatyon ve BSO 43
1.2.12.1. Hücresel GSH Düzeylerinin Düzenlenmesi 44
1.2.13. Taurin 46
2. GEREÇ ve YÖNTEM 48
2.1. Tavşanların Gruplandırılması 48
2.2. Balon Hasarı Oluşturulması 50
2.3. Doku Örneklerinin İzolasyonu 53
2.4. Telomerik Tekrar Restriksiyonel Fragmantasyon Deneyleri:Terminal

vii
Restriction Fragments (TRF) 54
2.4.1. Dokuların Homojenizasyonu 55
2.4.2. DNA İzolasyonu 56
2.4.3. DNA Miktarının Belirlenmesi 57
2.4.4. DNA’nın Agaroz Jelde Yürütülmesi ve Görüntülenmesi 58
2.4.4.1. DNA’nın Jel Elektroforezinin Temeli 58
2.4.4.2. Agaroz Jel Elektroforezi 59
2.4.5. DNA’nın Restriksiyon Enzimleri ile Kesilmesi 60
2.4.6. Restriksiyon Enzimleri ile Kesilmiş DNA Fragmanlarının
Agaroz Jelde Yürütülmesi 61
2.4.6.1. Agaroz Jelin (% 0.8) Hazırlanması 61
2.4.6.2. Jelin Yürütülmesi 62
2.4.7. Southern Blot Yöntemi ile DNA’nın Jelden Membrana Transfer Edilmesi 63
2.4.7.1. Jelin Transfere Hazırlanması 64
2.4.7.2. Transfer 65
2.4.7.3. Transfer Öncesi ve Transferde Kullanılan Tamponlar 66
2.4.8. Membran Üzerine DNA’nın Sabitlenmesi 66
2.4.9. Prehibridizasyon 67
2.4.10. Hibridizasyon 67
2.4.11. Hibridize Olmuş Telomerik DNA’nın Belirenmesi 68
2.4.12. Hibridizasyonda Kullanılan Çözeltiler 70
2.4.13. DNA’nın Jelden Geri Alınması 70
2.5. Telomerik Tekrar Amplifikasyon Protokolü (Telomeric Repeat

viii
Amplification Protocol, TRAP) 71
2.5.1. Ekstraktların Hazırlanması 72
2.5.2. Total Protein Miktar Tayini 73
2.5.3. Kontroller 76
2.5.4. TRAP Analizinin PCR Basamağı 78
2.5.5. Oluşan PCR Ürünlerinin Poliakrilamit Jelde Yürütülmesi 80
2.6. Histolojik Analizler 81
2.6.1. Morfometrik İncelemeler 83
2.6.2. İmmünohistokimyasal İncelemeler 84
2.7. Kan Örneklerinde Yapılan Ölçümler 86
2.7.1. Plazma Total Glutatyon (GSH) Miktarının Belirlenmesi 86
2.7.2. Plazma Glutatyon /Okside Glutatyon (GSH/GSSG) Oranının Tayini 87
2.7.3. Plazma Glutatyon Peroksidaz (GPx) Enzim Aktivitesinin Tayini 88
2.8. Aygıtlar 88
2.7. Verilerin Değerlendirilmesi 90
3. BULGULAR 91
3.1. Bölüm I 91
3.1.1. Morfometrik İncelemeler 91
3.1.1.1. İntima alanı 92
3.1.1.2. Medya Alanı 93
3.1.1.3. Lümen Alanı 93
3.1.2. İmmünohistokimyasal İncelemeler 94
3.1.2.1. Ki-67 Ekspresyonu 94

ix
3.1.2.2. PCNA (Proliferating Cell Nuclear Antigen) Ekspresyonu 95
3.1.2.3. α-Aktin Ekspresyonu 95
3.1.3. Telomer Fragmanlarının Uzunluğu 96
3.1.3.1. DNA İzolasyonu ve Miktarının Saptanması 96
3.1.3.2. DNA’nın Restriksiyon Enzimleri İle Kesilmesi 97
3.1.3.3. DNA’nın Jelden Membrana Transfer Edilmesi 97
3.1.3.3.1. Radyoaktif Olmayan İşaretleme ile Yapılan Deneyler 97
3.1.3.3.2. Radyoaktif İşaretleme ile Yapılan Deneyler 102
3.1.3.4. Telomerik Fragmanların Kantitatif Değerlendirilmesi 104
3.1.4. Telomeraz Biyoaktivitesi 106
3.2. Bölüm II 109
3.2.1. Morfometrik İncelemeler 109
3.2.2. İmmünohistokimyasal İncelemeler 112
3.2.2.1. Ki-67 Ekspresyonu 112
3.2.2.2. PCNA (Proliferating Cell Nuclear Antigen) Ekspresyonu 113
3.2.2.3. α-Aktin Ekspresyonu 114
3.2.3. Kanda Yapılan Ölçümler 115
3.2.3.1. Plazma Glutatyon (GSH) Miktarı 115
3.2.3.2. Plazma Glutatyon/ Okside Glutatyon (GSH/GSSG) Oranları 116
3.2.3.3. Plazma Glutatyon Peroksidaz (GPx) Enzim Aktivitesi 117
3.2.4. Telomer Fragmanlarının uzunluğu 118
3.2.4.1. DNA İzolasyonu ve Miktarının Saptanması 118
3.2.4.2. DNA’nın Restriksiyon Enzimleri ile Kesilmesi 120

x
3.2.4.3. DNA’nın Jelden Membrana Transfer Edilmesi 122
3.2.4.4. Telomerik Fragmanların Kantitatif Değerlendirilmesi 127
4. TARTIŞMA 128
5. SONUÇ VE ÖNERİLER 142
ÖZET 143
ABSTRACT 145
KAYNAKLAR 147

xi
TABLO LİSTESİ
Tablo 1.1. İntimal kalınlaşma ve restenoz modelleri 13
Tablo 2.1. Damar örneklerinden elde edilen DNA konsantarsyonları 57
Tablo 2.2. Değişik boyutlarda DNA fargmanlarının ayırımı için
uygun agaroz konsantrasyonları 59
Tablo 2.3. Kullanılan tamponlar 66
Tablo 2.4. Hibridizasyonda kullanılan çözeltiler 70
Tablo 2.5. Total protein miktar tayininde kullanılan dilüsyonlar 74
Tablo 2.6. Standartların belirli konsantrasyonlarına karşı ölçülen absorbans değerleri 75
Tablo 2.7. %12.5’luk Poliakrilamit Jelin Hazırlanması 80
Tablo 2.8. Birinci Seri Deneylerde Kullanılan Aygıtlar 89
Tablo 3.1. DNA izolasyonu sonrası elde edilen absorbans değerleri 98
Tablo 3.2. Şekil 3.8’deki blot görüntüsüne ait ortalama telomerik
fragman (OTF) değerleri 105
Tablo 3.3. Şekil 4.9’daki blot görüntüsüne ait OTF değerleri 105
Tablo 3.4. Şekil 4.10’da radyoaktif işaretleme sonucu elde edilen blot görüntüsü
verilen örneklerin ortalama rölatif telomerik fragman (OTF) değerleri 106
Tablo 3.5. Plazmadaki Totatl Glutatyon Konsantrasyonu 115
Tablo 3.6. Plazma GSH/GSSG Oranları 116
Tablo 3.7. Plazma GPx Enzim Aktivitesi 117
Tablo 3.8. DNA izolasyonu sonrası elde edilen örneklerin absorbans değerleri 119
Tablo 3.9. Ortalama Telomer Boylarının Yüzdesi 127

xii
ŞEKİL LİSTESİ
Şekil 1.1. Farklı ateroskleroz evrelerinde damar düz kas
hücrelerinin fonksiyonları 11
Şekil 1.2. Uç replikasyon sorunu 20
Şekil 1.3. Telomeraz enziminin üç boyutlu yapısı 23
Şekil 1.4. Telomerik kompleklsler, telomeraz ve telomer sinyallenmesinde
görev alan proteinlerin kompozisyonun şematik gösterimi 27
Şekil 1.5. Reaktif oksijen türleri ve doku hasarı 31
Şekil 1.6. Reaktif oksijen türlerinin etkileri 38
Şekil 1.7. Glutatyonun (GSH) kimyasal yapısı 43
Şekil 1.8. Glutatyonun Metabolizması 44
Şekil 2.1. Deney şeması 50
Şekil 2.2. Total protein miktarı standart grafiği 76
Şekil 3.1. Balon anjiyoplasti sonrası 0. günde hasar uygulanan ve
uygulanmayan tavşan ilyak arterlerine örnek kesitler 92
Şekil 3.2. Balon anjiyoplasti sırasında hasar uygulanan ve uygulanmayan
tavşan ilyak arterlerinin anjiyoplastiyi izleyen 14. günde alınan
örnek kesitleri 92
Şekil 3.3. Balon anjiyoplasti uygulaması sonrası 14. günde izole
Edilerek parafin ile sabitlendirilmiş ve Ki-67 ile etkileştirilmiş
ilyak arter kesitinde intimal kalınlaşma alanı 94

xiii
Şekil 3.4. Balon anjiyoplasti uygulaması sonrası 14. günde izole
Edilerek parafin ile sabitlendirilmiş ve PCNA ile etkileştirilmiş
ilyak arter kesitinde intimal kalınlaşma alanı 95
Şekil 3.5. Balon anjiyoplasti sırasında hasar verilen ve uygulama Sonrası
14. günde izole edilen damarlarda gelişen intimal
kalınlaşmada α-aktin ekspresyonu 96
Şekil 3.6. DNA izolasyonu sonrası genomik DNA’nın %0.8’lik
agaroz jele uygulanması sonucu elde edilen 1. jele ilişkin
görüntü 98
Şekil 3.7. DNA izolasyonu ve restriksiyonu yapılanörneklerin (1. jel)
Southern blot ve hibridizasyon sonrası elde edilen görüntüsü 99
Şekil 3.8.. Telomerik DNA fragmanlarının büyük ölçüde uzun olup
olmadığının kontrol edilmesi amacıyla %0.8’lik agaroz jelde
yürütülen ve ETBr ile görünür hale getirilmiş genomik DNA’nın
restriksiyon öncesi (A) ve sonrası elde edilen fragmanların (B)
jeldeki görüntüleri 100
Şekil 3.9. İkinci denemeye ilişkin (2. jel) DNA izolasyonu ve restriksiyonu
yapılan örneklerin Southern blot ve hibridizasyon sonrası
elde edilen görüntü 101
Şekil 3.10. DNA izolasyonu ve restriksiyon sonrası Southern blot ve
hibridizasyonu yapılan bir diğer deney serisine ilişkin
jelin (3. jel) görüntüsü 102
Şekil 3.11. 32P işaretleme ile elde edilen radyografi 103

xiv
Şekil 3.12. TRAP analizi sonrası elde edilen poliakrilamid jel fotoğrafı 107
Şekil 3.13. TRAPeze (Chemicon, ABD) kiti ile elde edilen telomeraz
aktivitesine ilişkin jel görüntüsü 108
Şekil 3.14. Hematoksilin-eosin ile boyanmış tavşan iliyak arter kesitleri 110
Şekil 3.15. İntimal Alan eDğerleri 111
Şekil 3.16. Lümen Alan Değrleri 111
Şekil 3.17. Balon anjiyoplasti uygulaması sonrası 14. günde izole
Edilerek parafin ile sabitlendirilmiş ve Ki-67 ile etkileştirilmiş
ilyak arter kesitinde intimal kalınlaşma alanı 112
Şekil 3.18. Balon anjiyoplasti uygulaması sonrası 14. günde izole
Edilerek parafin ile sabitlendirilmiş ve PCNA ile etkileştirilmiş
ilyak arter kesitinde intimal kalınlaşma alanı 113
Şekil 3.19. α-actin boyması 114
Şekil3.20. Telomer analizi öncesi jel fotoğrafı 120
Şekil 3.21. Restriksiyon Sonrası Jel Fotoğraf 121
Şekil 3.22.Transfer ve hibridizasyon sonrası film görüntüsü 123
Şekil 3.23. Hibridizasyonun yoğun olduğu üst bölgeler çıkarıldıktan sonra elde edilen
membran froğrafı 124
Şekil 3.24. 2. denemeye ait restriksiyon sonrası jel foroğrafı 125
Şekil 3.25. 2. membrana ait film görüntüsü 126

xv
RESİM LİSTESİ
Resim 2.1. Tavşan ilyak arter (A) ve femorel arter (B) anjiyogramı 51
Resim 2.2. Bir branül yardımı ile femoral artere girilmesi 52
Resim 2.3. Balon kateterin damar içerisinde kılavuz tel yardımı ile
İlerletilmesi 53

1. GİRİŞ VE GENEL BİLGİLER
1.1. Giriş
Ateroskleroz ve neden olduğu kardiyovasküler komplikasyonlar ülkemizde ve
tüm dünyada başta gelen ölüm nedenlerinden birini oluşturmaktadır (47). Hastalığın
insan sağlığını tehdit eden özelliğinin ortaya konması, hasta ve hasta yakınlarına
psikolojik ve sosyal sorunlar yüklemesinin yanı sıra neden olduğu sağlık
problemlerinin maliyetinin çok büyük boyutlara ulaşması, önlenmesi ve denetim
altına alınması yönündeki çabaları artırmıştır. Günümüzde artık ateroskleroz ile ilgili
risk faktörlerinin büyük bir bölümü bilinmekte ve erken aterosklerotik lezyonların
gelişimi ile ilgili mekanizmaların aydınlatılmasına çalışılmaktadır (35, 61).
Damar düz kas hücre proliferasyonu ve migrasyonuna bağlı olarak meydana
gelen intimal hiperplazinin aterosklerozun ve balon anjiplasti sonrası oluşan
restenozun patojenezinden sorumlu olduğu bilinmektedir (52). Vasküler düz hücre
migrasyon ve proliferasyonu düzenleyen moleküler yolakları aydınlatmaya ve
vasküler düz kas hücre hiperplazisini önlemeye yönelik stratejileri geliştirmek üzere
çok sayıda çalışma yapılmıştır. Bununla birlikte neden olan mekanizmalar büyük
oranda bilinmemekte ve etkin bir tedavi de oluşturulamamış durumdadır (105, 107).
Telomeraz enzimi kromozomların uçlarının devamından sorumlu bir
ribonükleoproteindir. Yetişkin somatik hücrelerde her hücre bölünmesi telomerik

2
DNA’dan 30-150 bç’lik kayba neden olur. RNA’ya bağımlı DNA polimeraz olan
telomeraz varlığında, telomer boyu uzatılır veya devam ettirilir ve böylece yinelenen
eskime (repetitive senesence) engellenir. Kanser hücrelerinin büyük çoğunluğunda
ayrıca hızlı çoğalan hematopoietik hücreler, endometriyal hücreler ve trofoblastlarda
değişen oranlarda telomeraz enzim aktivitesi saptanmıştır (104).
Yakın zamanda yapılan çalışmalarda telomeraz aktivasyonu ve telomer
devamlılığının genetik hipertansiyonda vasküler yeniden modellenmenin
oluşumunun temelinde yatan artan damar düz kas hücre proliferasyonunu
yönlendirdiği ileri sürülmektedir (33). Benzer şekilde, vasküler düz kas hücrelerinde,
hücre proliferasyonu sürecinde telomerazın aktive olduğu, buna karşılık telomeraz
aktivitesinin inhibisyonu ile hücre büyümesinin azaldığı da gösterilmiştir (126). Bu
kapsamda, gerek aterosklerotik lezyonlar gerekse balon anjiyoplasti sonrası oluşan
restenozda telomer devamlılığının ve telomeraz aktivitesinin rolü henüz
bilinmemektedir (108).
Bu bilgilerden hareketle bu araştırma projesinde endotel hasarı sonrası gelişen
düz kas hücre proliferasyonu ve/veya migrasyonunda telomeraz enziminin olası rolü
ile telomer devamlılığı ve restenoz arasındaki olası ilişki araştırılmıştır. Bu amaçla
tavşanlarda ilyak arterine balon anjiyoplasti uygulanmış (178) ve daha sonra izole
edilen hasar gören ve hasar görmeyen damar segmentlerinde karşılaştırmalı olarak
telomeraz enzim aktivitesi ve telomer devamlılığı araştırılmıştır. Bu kapsamda damar
segmentlerinden DNA izole edilmiş, restriksiyon enzimleri ile kesilmiş ve Southern
blot ile telomerik DNA fragmanlarının uzunluğu kantitatif olarak analiz edilmiştir.
Ayrıca telomerik fragmanların boy uzunluğu belirlenen örneklerde telomerik tekrar
amplifikasyon protokolü (TRAP) analizi ile telomeraz biyoaktivitesi araştırılmıştır.

3
Bunun yanı sıra ilyak arter segmentlerinde intimal hiperplazi oluşumunun
saptanmasına yönelik morfometrik ve immünohistokimyasal analizler yapılmıştır.
1.2. Genel Bilgiler
1.2.1. Ateroskleroz
1.2.1.1. Tanım
Ateroskleroz, endotel hasarı, lipit birikimi, monosit infiltrasyonu, düz kas
hücre proliferasyonu ve migrasyonu ve ekstraselüler matriks birikimi ile karekterize
kompleks bir arter hastalığıdır. Günümüzde ülkemizde ve tüm dünyada meydana
gelen ölüm vakalarının % 50’sinden ateroskleroz ve ona bağlı olarak gelişen
kardiyovasküler komplikasyonlar sorumlu tutulmaktadır (137, 155).
Ateroskleroz ilk kez 1950’lerin sonlarında Virchow tarafından hücrelerin
katılımı ile gerçekleşen proliferatif bir hastalık olarak tanımlanmıştır (189). O
zamandan günümüze dek aterosklerotik plak evrimini, plak komplikasyonunun
determinantlarını, akut iskemik hastalıkların sebebini anlamaya yönelik olarak
lezyonların morfolojik tipleri üzerinde çalışmalar yapılmıştır (20, 47, 161). Bu
patolojik çalışmalar aterosklerotik lezyonun hücresel elementleri arasında geniş
spektrumlu moleküler etkileşimler bulunduğunu ortaya koymuştur. Son yıllarda artan
moleküler düzeydeki biyolojik çalışmalar özellikle aterosklerozun genetik
predispozisyonu konusuna yoğunlaşmıştır. Genetik yatkınlık hastalığın gelişiminde

4
önemli bir faktör olmasına karşın çevresel, beslenmeye dayalı ve hemodinamik
faktörler de bu patojenezde önemli rol oynamaktadır. Bu nedenle aterosklerozun
daha iyi anlaşılması hastalığın evrimini yavaşlatmak, etkin terapötik yaklaşımlar ve
koruyucu stratejiler geliştirmek açısından son derece önemlidir (91).
Aterosklerotik lezyonlar daha yaşamın erken dönemlerinde, çocukluk
çağlarında çoğunlukla büyük ya da orta büyüklükteki elastik ve müsküler arterlerin
intimasında düşük dansiteli lipoprotein (LDL) ve monositlerden türeyen
makrofajların birikimi sonucu oluşmaya başlar (155). Yağ izleri olarak adlandırılan
bu en erken lezyon tipi uzun süre belirti vermeksizin ilerleyerek yaşamın ileri
dönemlerinde fibröz plak olarak adlandırılan kompleks ilerlemiş lezyonlar haline
dönüşür (153). Oluşan bu lezyonlar kalp, beyin ve ekstremitelerde iskemiye yol
açabilir ve infarktüs ile sonuçlanabilir (155). Dislipidemi, hipertansiyon, diyet,
diyabet, stres, sigara içme alışkanlığı, cinsiyet, genetik faktörler ve yaş hastalığın
patojenezi ve klinik seyri ile yakından ilişkili risk faktörlerini oluşturmaktadır (90).
1.2.1.2. Ateroskleroz Patojenezi
Ateroskleroz, endotel disfonksiyonu, inflamasyon, vasküler proliferasyon ve
matriks değişimini içeren çok yönlü kompleks bir süreçtir. Hastalığın patojenezinin
açıklanmasına yönelik çok sayıda hipotez ileri sürülmüştür. Bunlardan başlıcaları
hasara cevap, monoklonal hücre büyümesi, organize trombus ve infeksiyon
inflamasyon hipotezidir (20, 91,156, 161). Ancak bunlardan en çok kabul gören Ross
tarafından 1976’da ileri sürülen ve 1986’da yine Ross tarafından modifiye edilen

5
hasara cevap hipotezidir (154, 156). Günümüzde ateroskleroz Ross’un “modifiye
hasara cevap hipotezi” ile açıklanmaktadır. Bu hipoteze göre hasar ya da diğer bir
uyaran arterdeki hücrelerin (endotel hücreleri, monosit ve makrofajlar, trombositler)
otokrin ya da parakrin stimülasyonuna yol açarak büyüme faktörleri salınımını
stimüle etmektedir (154). Bu hipotezde birbirine alternatif iki yolağın düz kas hücre
proliferasyonu sonrası lezyon oluşumuna yol açabileği vurgulanmıştır.
Bunlardan ilkinde kronik hiperkolesterolemi başlatıcı faktör olabilir. Endotel
hücreleri kronik olarak yüksek LDL ve VLDL düzeyleri ile karşı karşıya kalırsa
endotel hücrelerinin plazma membranları ile hızlı kolesterol değişimi meydana
gelebilir. Bu olay kolesterol, fosfolipid ve protein oranlarında karmaşık değişikliklere
ve bu hücrelerin plazma membranlarının viskozitesinde değişmelere, endotel
hücrelerinden çeşitli büyüme faktörlerinin salınımına, monositlerin endotel hücre
yüzeyine yapışmasına yol açar. Monositler endotel yüzeyine yapışıp büyüme
faktörleri salıverirler daha sonra endotelyal hücreler arasından subendotelyal bölgeye
geçerek orada lokalize olurlar. Subendotelyal bölgede makrofajlara dönüşürler.
Endotelyal modifiye LDL subendotelyal makrofajlar için bir stimulus olabilir.
Bu makrofajlar reseptör aracılı endositoz ile modifiye LDL’yi bünyelerine alıp
“köpük hücreleri” ni oluştururlar. Damar duvarında görülen “yağ izleri” bu şekilde
oluşur. Makrofajlar da monositler gibi çeşitli büyüme faktörleri, kemoatraktan ve
mitojenleri salıverme yeteneğindedirler. Bu arada trombositler de değişen endotelyal
hücre yüzeyi ile etkileşir ve kemoatraktan maddeler (trombosit kaynaklı büyüme
faktörü; PDGF) salıverirler. Bu maddeler düz kas hücrelerinin arteriyel intimaya
migrasyonu ve orada prolifere olması için yeterli uyaranı oluşturur. Prolifere olan
intimal düz kas hücreleri lipidlerle yüklenerek “miyojenik köpük hücreleri” ne

6
dönüşürler. Sonuçta sürekli devam eden çeşitli büyüme faktörlerinin salınımı,
monosit yapışması ve düz kas hücre birikimi lezyonun “fibröz plak” haline
dönüşmesine yol açar. Bu plak bir bağ dokusu matriksinin çevrelediği değişik
miktarda ekstraselüler lipid içeren intimal düz kas hücrelerinden oluşur.
Makrofajlardan salınan maddeler endotel hücreleri üzerindeki hasarı daha da
artırabilir ve hatta endotel hücre kayıplarına neden olabilir. Bu nedenle trombosit
adezyon ve agregasyonu görülebilir. Büyüme faktörlerinin diğer bir kaynağını
oluşturan trombositler ve trombusun fibröz maddeleri plak içine katıldığı zaman
“kompleks aterosklerotik lezyonlar” oluşabilir (154, 156).
Hasara-cevap hipotezinin ikinci yolağına göre endotelyum kimyasal,
immünolojik, viral ya da diğer zararlı etmenlerle hasar görmesine karşın bütünlüğünü
korur. Endotel hücrelerinde oluşan zedelenme bu hücrelerin turnover’ını artırarak
endotel kaynaklı büyüme faktörleri üretmelerine neden olur. Buna PDGF’nin düz kas
hücreleri tarafından endojen olarak üretimi eşlik eder. Bu büyüme faktörleri düz kas
hücrelerinin medyadan intimaya migrasyonlarını uyarır. Bu yolakta başlangıçta bir
monosit adezyonu olmadan ve trombositler subendotelyal yapılara yapışmadan
doğrudan fibröz plak gelişebilir (154). Bu yolak diyabet, hipertansiyon, sigara içme
alışkanlığı veya ateroskleroz insidansını artıran diğer olgular için önem taşır (153).
Sonuç olarak, endotel hasarı ve endotelyal işlevdeki bozulma hasara-cevap teorisine
göre aterosklerozun başlangıç ve ilerlemesinde esansiyel rol oynar (48).

7
1.2.2. Vasküler Proliferasyon ve Ateroskleroz
Ateroskleroz endotel disfonksiyonu, enflamasyon, vasküler proliferasyon ve
matriks değişikliklerini içeren çok basamaklı süreçlerden oluşurmaktadır (3).
Vasküler porliferasyon ateroskleroz patobiyolojisinde rol oynar ve enflamasyon,
apoptosis ve matriks değişiklikleri gibi diğer hücresel süreçlerle içiçedir. Özellikle
vasküler proliferasyonun in-stent restenoz, transplante damar hastalığı ve ven bypass
graft yetmezliğinin patofizyolojisine olan katkısı oldukça önemlidir. Bu tür
rahatsızlıkların tedavisinde yeni yaklaşım hücre döngüsünün hedeflenmesi ile
vasküler proliferasyonun önlenmesidir (23, 48).
Yapılan son çalışmalar aterosklerozun tüm evrelerinde enflamasyonun yer
aldığını göstermektedir. Enflamasyonun yanı sıra damar düz kas hücre
proliferasyonu da çok önemli bir süreci oluşturmaktadır. Şekil 1.1.’de (47) farklı
ateroskleroz evrelerinde damar düz kas hücrelerinin fonksiyonları görülmektedir.
Erken aterosklerozda damar düz kas hücreleri monosit kemoattraktan protein 1
(MCP–1) ve vasküler hücre adhezyon molekülü (VCAM) gibi enflamatuar
medyatörleri üreterek ve lipoprotein tutulması için gerekli matriks moleküllerini
sentezlenmesi yoluyla aterom oluşumuna katkıda bulunduğu düşünülmektedir.
Bunların yanında, damar düz kas hücreleri fibröz bir kapak oluşturarak plağın
stabilitesinin korunmasında da önemli rol oynadıkları düşünülmektedir. Fibröz
kapağın zayıf ve ince olduğu bölgelerde damar düz kas hücrelerinin apoptosise
uğraması bunun bir kanıtı olarak gösterilmektedir (164). İleri lezyonlarda
fibroblastlar ve damar düz kas hücreleri ekstraselüler kalsifikasyon ile fibrokalsifik
bir plak oluşturmaktadırlar. Büyüme faktörleri ve damar düz kas hücrelerinin damar

8
hasarına yanıtlarının anlaşılmasına yönelik çalışmalar genel olarak hayvan arteriyel
hasar modellerini içermektedir. Hastalardan direkt veri elde etmek güçtür. Sıçan
modelinde ölmekte olan damar hücrelerinden salınan temel fibroblast büyüme
faktörü damar düz kas hücrelerinin medyal proliferasyonunu başlatırken, trombosit
kaynaklı büyüme faktörü (PDGF) damar düz kas hücrelerinin intimaya göçüne neden
olmaktadırlar. Trombosit kaynaklı büyüme faktörü, TGFβ, anjiyotensin II, epidermal
büyüme faktörü ve insülin-benzeri büyüme faktörü-1 intimal proliferasyon ve
matriks birikimini etkilemektedirler. Ayrıca, endotel hücrelerden nitrik oksit (NO)
salınımının azalması, reaktif oksijen türleri tarafından NO’in inakive edilmesi veya
heparan sülfat proteoglikan sentezindeki değişiklik nedeniyle ortaya çıkan büyüme
inhibitör faktörlerinin kaybı da damar düz kas hücrelerini göç etmesine
proliferasyonuna ve enflamatuar yanıtlarının artmasına katkıda bulunabilmektedirler.
Bütün bu olaylarda damar düz kas proliferasyonun öneminin algılanması ve hücre
proliferasyonunun moleküler ve hücresel mekanizmalarının anlaşılmasına yönelik
gelişmeler, antiproliferatif tedavi yaklaşımını araştırma ve geliştirmenin hedefi haline
getirmiştir (89, 91, 107, 138).
Vasküler proliferatif hastalıkların önlenmesinde denenen ilaçlar arasında
heparin, anjiyotensin dönüştürücü enzim inhibitörleri, terbinafin ve trapidil gibi
büyüme faktörü antagonistleri, bir somatostatin analoğu olan anjiyopeptin, etoposit
ve doksorubisin gibi sitostatik ajanlar ve kalsiyum-kalmodulin antagonistleri
sayılabilir. Bu ilaçlarda hayvan deneylerinden elde edilen sonuçlar klinik
denemelerde beklenen başarıyı göstermemiştir. Bunun üzerine sitotoksik veya
sitostatik tedavi yaklaşımına gidilmiştir. Bunların arasında en ümit vaadeden ilaç
makrolit grubu bir antibiyotik olan rapamisindir. Son çalışmalar rapamisinin hücre

9
döngüsünün düzenlenmesi ve neointima gelişimini önlenmesindeki etkilerini ve
damar düz kas hücrelerine olan moleküler etkilerini ortaya çıkarmıştır (24, 59). Ümit
veren bir başka ilaç da paklitaksel (taksol)’dir. Paklitaksel tübülinin proliferasyonuna
neden olur ve böylece mikrotübüller oluşamaz ve hücre bölünmesini engelleyerek
hücrenin ölümüne neden olmaktadır. Paklitaksel-kaplı stentlerle yapılan klinik
denemeler lezyonun azaldığını göstermektedir.
Antiproliferatif tedavi yaklaşımları şimdiye dek restenoz ve graft yetmezliği
gibi hızlı gelişen ve lokalize proliferatif damar hastalıklarına odaklanmıştır. Oysa
ateroskleroz yavaş gelişen difüz bir hastalıktır. Bu nedenle hücre siklusunu inhibe
eden ilaçların lokal olarak kısa süreli verilmesi yaklaşımının bu durumda olasılıkla
etkin olması beklenmemelidir. Uzun süreli antiproliferatif tedavi aterosklerotik
lezyonlardaki fibröz kapağı zayıflatarak yırtılmalara açık hale getirebilir. Bunun
yanında antiproliferatif tedavi in-stent restenozun ve ven bypass graft tıkanmasının
önlenmesinde mantıklı bir tedavi yaklaşımı olabilir (29, 46).
1.2.3. Deneysel İntimal Kalınlaşma Modelleri
1.2.3.1. Deneysel Modellere Genel Bakış
Atreoskleroz gelişiminden önceki erken ve temel aşama olarak kabul edilen
intimal kalınlaşmanın patojenezinde vasküler hasar önemli rol oynar (12, 105). Bu
nedenle endotel bütünlüğünün önemini kavramak, endotelyal hasar sonrası oluşan
hücresel çoğalma cevaplarını, bu cevapların farklı vasküler hastalıkların
patojenezindeki rollerini araştırmak için çeşitli deneysel vasküler hasar modelleri

10
geliştirilmiştir (35, 43, 46, 52,58, 134, 195). Fizyolojik koşullarda küçük hayvan
türlerinin çoğunun arterlerinde diffüz intimal kalınlaşma veya intimal yastıkçıklar
oluşmaz ya da hayvanın yaşlanması ile birlikte çok yavaş olarak gelişir. Buna karşın
değişik nitelikteki hasarlara yanıt olarak tüm arterlerde intimal kalınlaşma oluşumu
görülür. Hayvanlarda restenoz ve neointima oluşumu modelleri iki ana gruba
ayrılarak incelenebilir (Tablo 1.1.).
Birinci gruptaki modellerde normal sağlıklı arterlerde endotel tabakasında
değişik tekniklerle hasar oluşturulması (denüdasyon oluşturulması) esastır.
Endotelyumun zedelenmesi balon embolektomi kateterinin hassas bir şekilde damar
içinden geçirilmesi (balon denüdasyon), balon anjiyoblasti kateterinin şişirilerek
arterin gerilmesi (balon anjiyoplasti), döner tel geçirilmesi, elektriksel stimülasyon,
damar üzerine baskı uygulanması, arterin içinden hava akımı geçirilmesi gibi değişik
yöntemlerle sağlanabilir (35, 43). Bu teknikler lipidden zengin bir diyetle kombine
edilebilir (211). Kendi içinde bu gruptaki teknikleri de intravasküler teknikler ve
perivasküler teknikler olarak ayırmak mümkündür. Sert polietilen yaka uygulaması,
elektriksel stimülasyon gibi intravasküler tekniklerin uygulanması ile hasara yanıt
olarak gelişen yeni intima, “neointima” olarak adlandırılır (7). Bunun nedeni orijinal
intimanın hasara uğratılması ya da tümüyle yok edilmesidir. Ancak yumuşak silikon
yaka modelinde olduğu gibi orijinal intimaya doğrudan hasar vermeyen tekniklerle
oluşan yanıta “intimal kalınlaşma” adı verilir.

11
Şekil 1.1.: Farklı ateroskeroz evrelerinde damar düz kas hücrelerinin fonksiyonları (VSMC: Damar düz kas hücreleri) (47).

12
İkinci gruptaki modellerde damarlara ikinci bir hasar oluşturulur. Bunun için
önce ilk grupta belirtilen tekniklerden herhangi biri uygulanarak primer lezyon
oluşturulur. Daha sonra balon anjiyoplasti uygulanarak ikinci lezyon gerçekleştirilir.
Restenoz terimi önceden daralmanın söz konusu olduğu bu durumdaki damarlar için
kullanılır (87).
Hiperkolesterolemi modeli tek başına ya da bu modellere ek olarak
uygulanabilir. Hiperkolesterolemi aslında en yaygın olarak uygulanan intimal
kalınlaşma modelidir (211). Çoğunlukla tavşanlara uygulanmakla birlikte
hiperkolesterolemik primat ve domuz modelleri insan aterosklerotik lezyonlarını en
iyi temsil eden modeller olarak kabul edilmektedir. Bu model endotel hücre
denüdasyonu ve endotelyumda morfolojik değişiklikler oluşturmaksızın intimal
lezyon oluşumuna neden olmaktadır (58). Yukarıda yapılan sınıflandırma en yaygın
olarak kullanılan intimal kalınlaşma modellerini içermektedir. Bununla birlikte bu
sınıflandırmada değinilmeyen ve endotel denüdasyonu oluşturmayan diğer intimal
kalınlaşma modelleri arasında viral hasar (herpes virüs, sitomegalo virüs ile
infeksiyon), kronik karbon monoksit inhalasyonu, endotelin ve anjiyotensin gibi
vazoaktif maddelerin kronik olarak uygulanması sayılabilir. Ayrıca yabancı protein
enjeksiyonu sonucu oluşan ve endotel denüdasyonu oluşturan immun hasar modeli
de bu deneysel modellere eklenebilir (43).

13
Tablo 1.1.: İntimal kalınlaşma ve restenoz modelleri (43)
I. DAMARA TEK HASAR UYGULANAN MODELLER
(hiperlipidemi diyetli / hiperlipidemi diyetsiz)
İntravasküler metodlar Perivasküler metodlar
EH denüdasyonu +
kısmi mediyal hasar
EH denüdasyonu +
şiddetli mediyal hasar
İntimaya direkt hasar İntimaya indirekt
hasar
balon denüdasyon
döner tel
balon anjiyoplasti
havayla kurutma
baskı uygulama
stent yerleştirme
elektriksel stimülasyon
sert polietilen yaka
yumuşak silikon yaka
II. DAMARA İKİ HASAR UYGULANAN MODELLER
Yukarıdaki metodlardan biri ile oluşturulan primer bir lezyon üzerinde anjiyoplasti oluşturulması
1.2.3.1 Perkütanöz Translüminal Koroner Anjiyoplasti (PTCA)
İlk kez deneysel olarak 1963 yılında Baumgartner (11) tarafından tanıtılmakla
birlikte bir balon katater kullanılarak uygulanan koroner anjiyoplastinin ilk klinik
uygulaması 1978 yılında Grutenzig (74) tarafından gerçekleştirilmiştir. Tanıtımından
sonra da balon anjiyoplastinin emin ve etkin olduğu gösterilmiştir. Koroner arter
bypass graft (CABG) operasyonu ile karşılaştırıldığında, PTCA kabul edilebilir bir
alternatif olarak ele alınmaktadır (52). PTCA uygulanan ve CABG operasyonu
geçiren hastaları karşılaştıran çalışmalarda ölüm ve öldürücü-olmayan miyokard
infarktüsü gibi klinik sonlanmalar açısından benzer veriler elde edilmiştir (13).
Ancak PTCA uygulanan hastalarda belirgin şekilde fazla oranda yöntemin tekrarı
gerekmiş ve hastaların büyük çoğunluğunda angina belirtileri gözlenmiştir. PTCA

14
günümüzde de her yıl 500 000’den fazla hastaya uygulanmakta ve etkin olarak
kullanılmaktadır. Ancak akut kapanma ve ilk andaki başarısızlık oranı çok düşük
olmakla birlikte, uzun süreli komplikasyon açısından restenoz hala bir sorun olmayı
sürdürmektedir (29).
Restenoz balon dilatasyonu yapılan damar bölgesinde damarın daralmasıdır.
Uygulamadan hemen sonra gelişir ve belirtilerin tekrarı ile birlikte artar. Restenozun
tam olarak mekanizması henüz aydınlatılamamıştır ancak hastalar arasında
değişkenlik göstermekle birlikte arter duvarının yeniden modellenmesi (remodeling)
ile intimal hiperplazinin bir karışımı sonucu oluştuğu bilinmektedir. Restenozu
incelemek üzere çeşitli hayvan modelleri geliştirilmiştir (8, 58, 107). Bu modeller
aracılığı ile restenozun hücresel gelişim mekanizmaları konusunda yoğun bilgiler
elde edilmiştir (95).
İnsanda ve deneysel hayvan modellerinde yapılan çalışmalar restenozun
karakteristik histolojik özelliğinin neointimada düz kas hücre birikiminin olduğunu
göstermektedir. Balon hasarı başlangıçta medyal düz kas hücre ölümü ile birlikte
gelişir ve bunu aşağıda özetlenen olaylar izler;
i. Trombositler ve lizize uğrayan hücrelerden salıverilen büyüme faktöleri
tarafından medyal düz kas hücrelerinde proliferasyon başlar,
ii. Trombosit-kaynaklı büyüme faktörü (PDGF) başta olmak üzere kemo-
atraktanlara yanıt olarak medyal düz kas hücreleri intimaya göç eder,
iii. İntimal düz kas hücrelerinde proliferasyon gelişir,
iv. Sentetik bir fenotipe dönüşen düz kas hücrelerince kondroitin sülfat ve
hyalüronan gibi ekstraselüler matriks moleküllerinin yapımı artar,

15
v. Matriks metaloproteinazlar tarafından ekstraselüler matriksin aşırı yıkımı
ile vasküler yeniden modellenme gelişir.
Bu olayların bir birleşimi olarak meydana gelen ve sonuçta luminal daralmaya
neden olan olay ve bunun yanı sıra bu süreçte her aşamada rol oynayan faktörlerin
anlaşılabilmesi restenozun terapötik yaklaşımında yeni hedeflerin belirlenebilmesine
olanak sağlayacaktır. Bu açıdan bakıldığında balon hasarı sonrası salıverilen
mitojenler ve kemoatraktanlara verdikleri yanıt olarak medyal düz kas hücrelerinin
farklılık gösterdiği açıktır. Düz kas hücrelerinin proliferasyonu ve fenotipik
modülasyonu restenozun deneysel modellerinde arteriyal yanıtın en önemli
özelliklerini oluşturmaktadırlar (115, 196).
Hayvan modelleri
Restenozun mekanizmasını aydınlatmaya yönelik olarak gerçekleştirilen
deneysel çalışmaların büyük çoğunluğu sıçan ve tavşan karotid arteri kullanılarak
uygulanmıştır. Kullanılan bu arterlerin çapının insan koroner arterinden daha az
olması nedeniyle bir anjiyoplasti kateteri kullanılması mümkün olmamış, bu
çalışmaların büyük çoğunluğunda bir Fogarty embolektomi kateteri kullanılmıştır.
Tam olarak bir de-endotelizasyon (endotelsizleştirme) oluşturabilmek için, kısmen
şişirilmiş bir kateterin arter yatağında tekrarlanarak çekilmesi gerekmiştir. Bu
anlamda arter duvarının maruz kaldığı kuvvet büyüklük ve doğası açısından klinik
uygulamada gerçekleştirilenden farklılık göstermektedir. Bir diğer sorun ise, koroner
anjiyopasti daha önceden var olan bir hastalığa karşı arterleri dilate etmek için

16
uygulanırken, deneysel hayvan modelleri kullanılan çalışmaların bulgularının normal
arterleri yansıtması ile ilişkilidir.
Tavşan Modeli
Balon hasarına ilişkin tavşan modeli ilk kullanılan deneysel yöntemlerden
birisidir (211). Tavşanlar aterojenik diyete çok çabuk ve kolay yanıt verirler, diyete
başlandıktan kısa bir süre sonra hiperlipidemi ve erken lezyonlar gelişir. Bu anlamda
ilk çalışmalar Tashiro ve arkadaşları (184) tarafında yapılmıştır ve bu araştırıcılar
doymuş yağ asidlerinin az miktarları ile beslenen tavşanlarda serum lipid düzeyleri
değişmeksizin intimal kalınlaşma geliştiğini saptamışlardır.
İzleyen yıllarda tavşanlarda balon hasarı sonrası oluşan hücre kinetikleri ile
ilgili ayrıntılı veriler sunulmuştur. Bu verilere göre 14. gün sonunda rendotelizayon
tamamlanır, ayrıca vasküler permeabilite faktörlerinin katkısı ile artabilir. Hücre
proliferasyonu medyal hücreler açısından 7., intimal hücreler açısından 14. günde
maksimuma ulaşır; bir ay içinde de bu değerler normal döner (39, 41).

17
1.2.4. Telomerler
Telomerler ökaryotik kromozomların fiziksel uçlarıdır. İlk olarak 1938’de
Muller tarafından kromozom uçlarının özelleşmiş yapılar olduğu Drosophila’da
yaptığı çalışmalar sonucunda önerilmiş ve telomer terimi kullanılmıştır (16).
Telomerler kromozomun yapısal bütünlüğünün korunmasında önemli rol oynarlar
(17, 145). DNA uçlarının tam olarak replike olmasını ve stabilitesini sağlarlar (67).
Kromozomun replikasyon sürecine aracılık ederler. Hücre yaşlanma sürecini
düzenlerler böylece hücrenin ömrünü belirlerler (110). İnsan kromozomlarının
telomerleri 15 kilobaza kadar uzayabilen guanozinden zengin basit telomerik
hekzanükleotid tekrar dizilerinden oluşur. Telomerik DNA genel yapısı (T veya
A)m(G)n şeklindedir. Omurgalılarda ise (TTAGGG)n tekrar dizilerinden oluşur (110).
Telomerik baz dizileri hücresel DNA’yı kararlı halde tutar ve onları yeni
kombinasyonlardan (end to end fusion) korur (149). Tek bir hücrede türüne bağlı
olarak 40.000–1.000.000 telomerik tekrar bulunabilir (67, 148).
Telomerler normal hücrelerde in vivo koşullarda yaş ile birlikte veya normal
hücre kültürlerinde kromozomal DNA’nın bir fonksiyonu olarak progresif bir
kısalma ile metabolize olurlar. Konvansiyonel DNA polimerazların sadece 3’-5’
doğrultusunda DNA’yı replike edebilmeleri nedeniyle telomerlerin en uçta kalan
kısmını yenilemedeki yetersizlikleri sonucu her bir kromozom replikasyon
döngüsünde 5’ ucundan ~150 baz çifti kaybederler. Yeni sentez edilen DNA
dizisinin sentezi küçük RNA primerlernin kullanılması ile gerçekleşir. Bütün DNA
dizisi sentezlenip 5’ ucunun sonuna gelindiğinde primer en son kısma bağlanır fakat
karşı tarafta sentezlenecek kısım kalmadığı için ayrılır ve degrade olur (122).

18
Böylece DNA replikasyonun sonunda tamamlanamayan replikasyon sonucu tek
taraflı dizi kalır. Bu da sentezlenen DNA’nın 5’ ucunun kalıp DNA’nın 3’ ucundan
daha kısa olacağı anlamına gelir. Bu durum “uç replikasyon sorunu” (end replication
problem) olarak bilinir (40) (Şekil 1.2.). Böylece hücre bölünme sayısı artıkça,
telomer boyu kısalır. Bir popülasyonda hücrelerin bölündüğü ölçüde telomerler
kısalır. DNA replikasyonuna bağlı telomer boyundaki bu kısalma hücre yaşlanma
sinyalinin ortaya çıkmasında mitotik bir saat görevi görür. Telomerler hücre
bölünmesiyle birlikte kritik bir noktaya kadar kısalmaya devam ederler. O kritik
noktada hücre yaşlanma sürecine girer ve daha fazla bölünmeyip hücre bölünme
siklusundan çıkar (159). İn vitro koşullarda kültüre edilen normal memeli
hücrelerinin de sınırlı sayıda prolifere olduğu bilinmektedir.
Buna “Hayflick Limiti” denir (81). Daha sonra yaşlanma (state of senescence)
sürecine girerler. Metabolik olarak aktiftirler fakat replike olamazlar. İrreversibl
olarak hücre siklusunun G1 fazında bloke edilirler ve mitojenik uyarıya yanıt
vermezler, fakat canlıdırlar ve metabolik olarak aktiftirler (78, 98). Hücrelerin bu
yaşlanmış fenotipleri normal hücrelere göre morfoloji, nükleer yapı, gen
ekspresyonu, protein işleyişi ve metabolizması açısından farklılık göstermektedirler.
Bu hücrelerin önemli bir kısmı yaşlanma ile ilişkili β-galaktozidaz ile boyanırlar ve
tümör baskılayıcı genler olan p53, p21 ve p16 upregüle olur (10,127). Telomerlerin
yaşlanma süreci ve yaşlanma ile ilişkili hastalıklarda biyobelirteç olabileceği öne
sürülmüştür (27, 57, 128, 194).

19
Organizmalar belirli koşullarda telomer kısalmasına karşı telomer kaybını kompanse
etmek için kompleks mekanizmalar geliştirmişlerdir. Şu ana dek 3 tip telomer
devamlılığı mekanizması saptanmıştır (110, 145):
1. İnsan hücrelerinde telomeraz ile telomerlerin yeniden sentezi öncül baskın
mekanizmadır.
2. Drosophila melanogaster’de telomerik DNA kromozom uçlarına spesifik
retrotranspozonların transpozisyonu ile telomerler uzatılabilmektedir.
3. Mayada ise non-resiprokol telomerik DNA’nın telomerler arasına
rekombinasyonu ile telomerler uzatılabilmektedir.
1.2.4.1. Telomer Devamlığının Korunması
Belli bir sürede ökaryotik bir organizmada telomeri kısaltan ve uzatan
mekanizmalar denge halindedir. Telomerler türe spesifik korunmuş DNA
dizilerinden ve bu dizilere bağlanan telomer bağlayan proteinler (Telomere-binding
proteins, TBP) olarak bilinen proteinlerden oluşmuşlardır (159). Telomerik DNA
TBP’ler ile birlikte özel bir yapı oluşturur ve bu yapı kromozomların tek başına
varlığını korur. Telomerik DNA’nın yapısı bozulduğunda zamanla fonksiyonlarını
kaybederler ve kromozomları tek bir bütün yapıda tutamazlar (203). Telomer
fonksiyon kaybının ilk belirtisi uç uca kromozom füzyonunun bir sonucu olarak
disentrik kromozomların ortaya çıkmasıdır. Bu nedenle telomer fonksiyon kaybının
en önemli belirteci telomer kısalmasının normal sınırların altına düşmesidir. İnsan ve
hayvan deneyleri bu bulguyu desteklemektedir (111).

20
Şekil 1.2: Uç replikasyon sorunu (End replication problem) (40).
Telomer dengesinin düzenlenmesinde başlıca düzenleyiciler TBP’lerdir.
Memeli hücrelerinde bunlardan en önemlileri telomer tekrar faktörü 1 (telomere
repeat factor1-TRF1) ve TRF2’dir (187). Bu proteinlerin aşırı ekspresyonu
telomerlerin kısalmasına neden olur. Bu da telomeri uzatan faktörlerin negatif
düzenleyicileri oldukları anlamına gelmektedir. Telomerlerde TRF1 ve TRF2
moleküllerinin sayıca kritik bir düzeyin altına düşmesi telomerazı uyarır. Buradan da
anlaşılacağı gibi, memeli hücreleri protein sayımını içeren bir mekanizma ile telomer
boyunu düzenlerler. Bu mekanizmada görev alan diğer telomerik proteinler tankiraz,
TIN2, hRAP1, TANK2 ve pot1’dir. Bu moleküllerden bazıları telomer boyunun
düzenlenmesinde TRF1 ve TRF2 ile birlikte çalışırlar (42, 75).

21
Telomerin fonksiyonu telomerin yapısı ile yönlendirilir. Gerek maya gerekse
insan telomerlerinde 3’ uzantılar bulunmaktadır. Bu uzantılar aslında DNA çift
sarmak kırıklarından farksızdır ve DNA hasarını belirleyen hücresel
mekanizmalardan korunmalıdır. Memeli telomerleri elektron mikroskobu
incelendiğinde telomerlerde bazı kıvrımların varlığı gösterilmiştir. 3’ uzantılar geriye
doğru katlanır. DNA çift sarmalı üzerinde dolanarak T-kıvrımını oluşturur (73).
Daha sonra serbest olan 3’ ucu iki sarmalın arasına girer ve D-kıvrımını oluşturur. T-
kıvrımının oluşmasına TRF2 aracılık etmektedir. Diğer TBP’ler de kıvrım
içerisindeki yerlerini alırlar. Kıvrım yapısı serbest kalan 3’-ucunu koruyarak DNA
hasarını ve onarım sinyallerinin aktive olmasını önler. Böylelikle kromozoma
stabilite sağlar (171).
1.2.4.2. Telomer Devamlılığı ve Apoptosis (159, 171)
Telomer devamlılığı ve apoptosun kontrolü arasında bir bağlantı olduğu
düşünülmektedir. Telomeraz eksikliği olan farelerde telomer kısalması ve telomerik
füzyonlar ile birlikte apoptosis seviyesinin da artığı gösterilmiştir. Apoptosis üreme
ve homeopoetik sistem gibi yüksek oranda proliferatif dokularda gözlenmektedir
(159). İnsan hücrelerinin TBF’lerinin genetik olarak manipüle edildiği bir çalışma
telomer devamlılığı ve apoptozun kontrolü arasındaki bağlantıyı ortaya çıkarmıştır.
Bu çalışmada TRF2’yi kodlayan gen mutasyona uğratıldığında apoptos düzeyinin
artığı gösterilmiştir. TRF2’nin telomer boyunu düzenlediği, telomerik füzyonları
önlediği ve T-kıvrımlarının oluşumuna katkı sağladığı bilinmektedir. TRF2
yokluğunda telomerler fonksiyon kaybına uğrarlar. Fonksiyon kaybı olan

22
telomerlerin G-uzantıları olmayabilir ve bu nedenle de T-kıvrımlarını oluşturamazlar.
Bu da disentrik kromozomların ortaya çıkmasına neden olur. Yapıdaki bu
değişiklikler hücre bölünmesinin normal akışını etkileyerek hücre yaşlanması veya
apoptosa yol açar. Bazı durumlarda Werner Sendromu gibi prematüre yaşlanma
sendromuna neden olabilir (159).
1.2.5. Telomeraz
Telomeraz telomerik DNA sentezlemede kendi RNA kalıbını kullanan bir
revers transkriptaz enzimidir. RNA altünitesi ve çeşitli protein komponentlerini
içeren büyük bir ribonükleoprotein kompleksidir (37). İlk olarak 1985’te Greider ve
Blackburn adlı araştırmacılar tarafından Tetrahymena thermophila’da varlığı
gösterilmiştir (65). Çoğu normal somatik hücrede telomeraz aktivitesi bulunmazken
kanser hücrelerinin ve in vitro ölümsüzleştirilmiş hücrelerin %90’ında telomeraz
aktivitesi bulunmaktadır (40, 97, 168). Epidermis bazal tabakası (208), menstrüel
siklusun proliferatif fazında endometriyal doku (104) ve oral mukoza hücreleri (93)
gibi yüksek proliferatif özellik gösteren bazı normal hücrelerde de telomeraz
aktivitesi saptanmıştır (130).

23
Şekil 1.3.: Telomeraz enziminin 3 boyutlu yapısı. Enzim en az bir RNA altbirimi ve protein altbirimi (katalitik altbirim) içerir. RNA altbirimi telomer sentezi için gerekli RNA kalıbını içerir. Telomer üzerinde dkG-quadruplex yapısı bulunmaktadır.
Telomerazın iki ana komponenti bulunur (144):
1. Fonksiyonel RNA komponenti
2. Katalitik komponenti
1.2.5.1. Telomerazın RNA komponenti
İnsanda kısaca hTR veya hTERC olarak gösterilir. RNA komponenti telomerik
DNA’nın sentezinde kalıp görevi görmektedir. İnsan telomeraz RNA’sı (hTR) 451
nükleotid uzunluğundadır. hTR’yi kodlayan gen 3. kromozomda bulunmaktadır. 11
nükleotidlik kalıp bir dizi (5’-CUAACCCUAAC–3’) telomerik tekrarları
(TTAGGG)n kodlar (114). Bazı spesifik nükleotidler RNA’yı enzimin aktif
bölgesinde tutabilmek için DNA primer substratındaki yapısal komponentler ve
protein altbirimleri ile etkileşirler (66).

24
1.2.5.2. Telomerazın Katalitik Komponenti
İnsanda hTERT olarak gösterilir. Revers transkriptaz aktivitesi gösteren
katalitik kısımdır. İnsan telomeraz revers transkriptazı veya hTERT 127 kDa’luk bir
polipeptiddir ve 5. kromozomda bulunan bir gen tarafından kodlanmıştır (66).
hTR ve hTERT telomerazın minimum çekirdek yapısını oluştururlar. hTR
telomeraz aktivitesinden bağımsız olarak hemen her dokuda eksprese edilir. Kanser
hücrelerinde ise genel olarak hTR ekspresyonu normal hücrelere göre artmıştır.
Katalitik komponent olan hTERT’in ekspresyonu ise hücredeki telomeraz
aktivitesine bağımlıdır. hTERT normal hücrelerde baskılanırken tümör dokusunda ve
ölümsüz hücrelerde upregüle olur. Telomeraz-negatif hücrelerde ektopik olarak
hTERT ekspresyonu telomeraz aktivitesini başlatmak, telomerleri uzatmak ve hücre
yaşam süresini artırmak için yeterlidir. Bu da hTERT’in, enzim aktivitesinin başlıca
belirleyicisi olduğunu göstermektedir (40).
Telomerazın protein komponentinde yer alan başka bir protein ise telomeraza
bağımlı protein-telomerase associated protein (TEP1)’dir. TEP’in telomeraz
enziminin yapısını kontrol etmede ve çeşitli regülatör faktörün enzime bağlanmasını
düzenlemede rolünün olduğu düşünülmektedir (37).

25
1.2.5.3. Telomerazın Regülasyonu
Telomeraz enzim aktivitesi birkaç farklı düzeyde kontrol edilir.
1. Transkripsiyonel regülasyon:
hTR ve hTEP1’in normal dokularda eksprese olmaslarına karşın, hTERT çoğu
normal somatik dokuda represe edilmektedir. Tümör dokularında ise aktif hale
geçmektedir. Bu da hTEP1 ve hTR’nin inaktif bir kompleks halinde bulıunduğu ve
enzim aktivasyonu için hTERT’in varlığına ihtiyaç duyulduğunu düşündürmektedir.
hTERT farklı durumlarda çeşitli transkripsyon faktörleri düzenlenir (2). C-Myc’nin
telomerazı aktive ettiği gösterilmiştir. NO’in telomerazı aktive ederek endotel hücre
yaşlanmasını geciktirdiği bildirilmiştir (188). Östrojen de telomerazı aktive eden
diğer bir faktördür. Bunun yanında hTERT’in promoter kısmının metilasyonu veya
ilişkili histonun deasetilasyonu genin transkripsiyonunu represe etmektedir (68, 79).
2. Diğer proteinlerle etkileşim
Telomerazın regülasyonunda posttranslasyonel mekanizmalar da etkilidir.
Telomerazın optimal konformasyonu ve aktivitesi diğer proteinlerle etkileşmesini
gerektirmektedir. hTR ve hTERT’in transkripsiyonal düzeyde düzenlenmesi
belirleyici ana faktördür. Bunun yanında post-transkripsiyonel mekanizmalar da rol
oynar. hTEP1’in NH2 bölgesinden bir peptid olan TEIPP1’in spesifik olarak
telomeraz aktivitesini inhibe ettiği gösterilmiştir. Bu da hTEP’in düzenleyici
faktörler ile etkileşip enzimin konformasyonunu etkileyerek regülatör bir rol
oynadığı fikrini desteklemektedir (84, 79).

26
3. Fosforilasyonun rolü
hTERT’in fosforilasyon durumu da telomerazın katalitik aktivitesinin
düzenlenmesinde etkilidir. Protein fosfotaz 2A (PP2A) ve c-Abl tirozin kinaz
telomeraz fonksiyonunu negatif yönde regüle ederken PKC ve Akt protein kinaz
telomeraz aktivitesini upregüle eder (96). Bu bulgular fosforilasyon ile telomerazın
aktive olduğunu, defosforilasyon ile de telomerazın inaktive olduğunu
göstermektedir (75).
1.2.5.4. Telomeraz İnhibitörleri
Telomeraz inhibitörleri etki yerlerine göre 2 gruba ayrılabilirler: Telomeraz ile
etkileşen bileşikler ve telomer ile etkileşen bileşikler.
1.2.5.5. Telomeraz ile Etkileşen Bileşikler
1. Antisense Oligonükletidler: Bunlar telomerik tekrarları taklit eden
deoksiriboz oligonükleotidlerdir ve hTR’nin kalıp bölgesine karşı
yönlendirilmişlerdir. Bu grupta en çok çalışılan ilaç fosforotiyoattır. RNA’ya bağlı
iken RNA’nın bağlanmasını önler ve translasyonu durdurur (37, 79).
2. Peptid Nükleik Asitler (PNA’lar): PNA’lar non-iyonik bir omurga içeren
modifiye oligonükleotidlerdir. Nükleotidler ve PNA’lar arasındaki bağın yüksüz
oluşu hedeflenen nükleik asitler ile hibridizasyon hızını ve afinitelerini artırır. Bu da
proteazlar ve nükleazlar ile degrade olmaya karşı daha etkin bir şekilde karşı

27
koymalarını sağlar. PNA’lar antisense etkilerini RNA’nın işleyişini bloke ederek
gösterirler. PNA’lar telomerazın RNA komponentini (hTR) hedeflerler ve spesifik
olarak hTR’nin aktivitesini inhibe ederler (37, 79).
3. 2’-O-methyl-RNA molekülleri: Bu moleküller telomerazın çok güçlü
inhibitörleridir. Kimyasal yapılarının DNA’ya benzemesi nedeniyle telomeraz
kompleksi tarafından substrat olarak tanınmada avantaj sağlarlar (37, 79).
Şekil 1.4: Telomerik kompleksler, telomeraz ve telomer sinyallenmesinde görev alan proteinlerin kompozisyonunun şematik gösterimi (96).

28
4. Antibiyotikler: Son gelişmeler hTR’ye karşı bazı antibiyotiklerin de
inhibitör etkiye sahip olabileceğini göstermektedir. Neomisin gibi aminoglikozitler
ve kinolon antibiyotikleri yüksek konsantrasyonlarda insan tranisyonel karsinoma
hücre kültüründe büyüme inhibisyonuna ve telomeraz aktivitesinde azalmaya neden
olmuşlardır (37).
5. Proteinler: HIV gp120, TGF-β1 ve pRb gibi bazı iyi karakterize edilmiş
proteinlerin telomerazı inhibe ettikleri gösterilmiştir. Prostaglandin A1’in de
telomerazı inhibe ettiği gösterilmiştir. Proteinler hücre içine hedeflenmesi zor
bileşiklerdir. Bu nedenle de yakın zamanda klinik uygulamada yer almaları mümkün
görünmemekle birlikte telomerazın düzenlenmesinde ve ileride modifiye bileşiklerin
geliştirilmesinde rol oynayabilirler (37).
6. Gen Modülatörleri: Retroviral vektörler kullanılarak hTERT’in inaktif
mutantını kodlayan bir genin hücre içine sokulması tümör hücrelerinde var olan
telomeraz aktivitesinin kesintiye uğramasına neden olmuştur. Bu durum hem in vitro
hem de in vivo koşullarda gösterilmiştir (37).
7. “Hammerhead” Ribozimler: Bu bileşikler 40-50 bazlık katalitik RNA
dizileridir; GUC gibi trinükleotid dizilerine karşı aktivite gösteren katalitik bölgesi
bulunur (37, 96)
8. Nükleozid ve Non-nükleozid Revers Trankriptaz İnhibitörleri: Bu bileşikler
telomerazın polimeraz aktif bölgesi ile etkileşirler. HIV’e karşı etkili olduğu bilinen
bileşiklerdir. AZT ve ddG’nin uzun süreli hücre kültürlerinde telomerazı inhibe
ettikleri gösterilmiştir (37).

29
1.2.6. Alternatif Telomer Uzaması (ALT)
Tümör hücrelerinin %90’ında telomer devamlılığı telomeraz aktivitesi ile
sağlanır (121). İnsan kanser olgularının ~%10’unda belirlenebilen bir telomeraz
aktivitesi yoktur (14, 19). Bu durumda telomer devamlılığı alternatif telomer uzaması
(ALT) olarak adlandırılan telomerazdan-bağımsız mekanizmalar ile gerçekleştirilir
(146). Bunlardan en önemlisi rekombinasyondur. Rekombinasyonal Telomer
Uzunlaması (RTE) birçok farklı durumda telomer devamlılığının sağlanmasından
sorumludur. ALT tümörleri ve hücre kültürlerinde boyları yüksek oranda heterojenite
gösteren telomerlere sahiplerdir. Boyları 1kb’dan 20kb’a dek farklılık gösterebilir.
Diğer bir özellikleri de ALT ile ilişkili promiyelositik lösemi (PML) cisimciklerinin
bulunmasıdır (26, 82).
1.2.7. Oksidatif Stres Ve Reaktif Oksijen Türleri
Reaktif oksijen / nitrojen türleri (ROS / RNS) ve serbest radikaller normalde
vücutta sürekli olarak oluşmakta ve anti oksidan savunma sistemi ile ortadan
kaldırılmaktadır. Belli bir miktara kadar ROS /RNS üretimi sağlık için gereklidir.
Örneğin ROS /RNS immün sistemin mikroorganizmaları atmasında yardımcıdır (4).
Sağlıklı bireylerde serbest radikallerin oluşum hızı ile bunların ortadan kaldırılma
hızı bir denge içerisindedir ve bu durum “oksidatif denge” olarak adlandırılır.
Oksidatif denge sağlandığı sürece organizma, serbest radikallerden
etkilenmemektedir. Bu radikallerin oluşum hızında bir artma ya da ortadan
kaldırılma hızında bir düşme bu dengenin bozulmasına neden olur. “Oksidatif stres”

30
olarak adlandırılan bu durum özetle: serbest radikal oluşumu ile antioksidan savunma
mekanizması arasındaki ciddi dengesizliği göstermekte olup, sonuçta doku hasarına
yol açmaktadır (166) (Şekil 1.5). Bazı hastalıklarda artan oksidatif /nitrosatif stres
hastalık patolojisine önemli katkıda bulunabileceği gibi hastalık patolojisinden de
kaynaklanabilir. Oksidatif dengenin ROS /RNS lehine artması sonucu oluşan fazla
miktarda serbest radikal hücre yapısını ve canlılığını ciddi şekilde etkileyen lipidler,
proteinler ve nükleik asit bazları gibi hemen hemen her tür biyolojik moleküller ile
reaksiyona girer. Özellikle tiyoller, proteinler ve lipidler gibi hücre sinyallemesinde
önemli role sahip hücresel hedefler serbest radikallerden etkilenirler (4).
1.2.8. Reaktif Oksijen Türleri Ve Hücresel Kaynakları
Birçok ROS /RNS çiftlenmemiş elektrona sahiptir ve bu nedenle de serbest
radikaldir. Bunlar süperoksit anyonu (.O2-), hidroksil radikali, (.OH), nitrik oksit
(.NO), nitrojen dioksit (.NO2) ve lipid radikalleridir. Diğer ROS /RNS ise hidrojen
peroksit (H2O2), peroksinitrit (ONOO-), peroksinitröz asittir (HOCl). Bunlar serbest
radikal değillerdir fakat oksidatif strese katkıda bulunan oksitleyici etkileri
bulunmaktadır. (209)

31
Şekil 1.5: Reaktif oksijen türleri ve doku hasarı
Hücresel olarak herhangi bir reaktif oksijen türünün üretilmesi radikal zincir
reaksiyonları vasıtasıyla diğerlerinin de üretilmesine yol açabilir. Örneğin, hücre
membranında çoklu doymamış yağ asitleri ile radikallerin reaksiyonu sonucu yağ
asidi peroksil radiakalleri (R-COO·) oluşabilir. Bunlar da komşu yağ asidi yan
zincirlerine saldırıp diğer lipid radikallerinin üretimini başlatabilirler (31). Zincir
reaksiyon sonucu üretilen lipid radikalleri hücre mebranında birikirler ve
plazmolemma sızıntısı ve veya membrana bağlı reseptörlerin disfonksiyonu gibi bir
çok hücresel fonksiyonu etkilerler. Lipid peroksidasyonunun son ürünleri olan
doymamış aldehitler ve diğer metabolitler sitotoksik ve mutajenik özelliklere
sahiptirler (31).

32
1.2.9. Reaktif Oksijen Türlerinin Düzenlenmesinde Rol Alan Oksidan ve
Antioksidan Enzim Sistemleri
1.2.9.1. Oksidan Enzin Sistemleri
Memeli hücrelerinde, ROS üretimine neden olan potansiyel enzim kaynakları,
araşidonik asit yolağı enzimleri lipoksijenaz ve siklooksijenaz, sitokrom P450,
ksantin oksidaz, NADH/NADPH oksidaz, nitrik oksit sentaz, peroksidazlar ve
hemoproteinlerdir (197). Bunlardan üçü kardiyovasküler sistemde yaygın olarak
çalışılmıştır:
1.2.9.1.1. NAD(P)H Oksidaz
NADH/NADPH oksidaz damarlarda süperoksit üretiminden sorumlu başlıca
enzimdir. NADH/NADPH oksidaz vasküler patojenezde rol oynayan sitokinler,
hormonlar ve mekanik güçlerle düzenlenir. Vasküler düz kas hücrelerinin
anjiyotensin II, trombin, trombosit kaynaklı büyüme faktörü, tümör büyüme faktörü-
α ve laktozilseramit vasküler ROS üretimini ve NADH/NADPH oksidaz üretimini
artırırlar (31). Bu enzimin alt üniteleri kesin olarak tanımlanamamıştır, bu nedenle de
nasıl etkileşim gösterdikleri tam olarak anlaşılamamıştır. Bununla beraber aktive
olduğunda süperoksit üretimini artırarak NO biyoyararlanımını azaltarak endotel
disfonksiyona neden olduğu ve çeşitli patofizyolojik koşullarda vasküler
hastalıkların oluşumunda önemli bir role sahip olduğunu düşünülmektedir (31).

33
1.2.9.1.2. Ksantin Oksidaz
Ksantin oksidoredüktaz pürin metabolizmasında ksantin ve hipoksantinin
oksidasyonunu katalize eden bir enzimdir. Birbirine dönüşebilen iki formda
bulunabilir: Ksantin dehidrogenaz veya ksantin oksidaz. (31). Ksantin oksidaz
moleküler oksijeni redükleyerek süperoksit ve hidrojen peroksit oluşumuna neden
olur. Patolojik koşullarda ksantin oksidaz aktivitesi artar ve yüksek miktarlarda
reaktif oksijen türlerinin üretimine neden olur. Enzim sadece vasküler düz kas
hücrelerinde eksprese olmaz. Aynı zamanda plazmada dolaşarak endotel hücrelerin
ekstraselüler matriksine bağlanır (196).
1.2.9.1.3. Endoteliyel Nitrik Oksit Sentaz
Fizyolojik koşullarda endoteiyel nitrik oksit sentaz L-arjininin L-sitrüline
oksidasyonunu katalizler. Bu reaksiyon sonucunda yan ürün olarak nitrik oksit (NO)
açığa çıkar (196) Bazı koşullarda NOS nitrik okside ilave olarak süperoksit
oluşumuna neden olabilir (183). eNOS elektron transportu için oksijenaz
bölgesindeki hem grubuna yakın bağlanma bölgesi bulunan tetrahidrobiyopterin
kofaktörüne ihtiyaç duymaktadır. L-arjinin veya tetrahidrobiyopterin yetersizliğinde
veya tetrahidrobiyopterin’in yükseltgendiği durumlarda ortaklanmamış elektronlar
moleküler oksijeni NO yerine süperoksite dönüştürürler. (183, 196). Özellikle
hipertansiyonda NAD(P)H oksidazların aktive olması tetrahidrobiyopterinin
oksidasyonuna neden olur ve yüksek miktarlarda süperoksit açığa çıkar (183)

34
1.2.9.2. Antioksidan Enzim Sistemleri
1.2.9.2.1. Superoksit Dismutaz (SOD)
Süperoksit dismutazlar (SOD) bütün vasküler hücrelerde süperoksite karşı
başlıca savunma sistemini oluştururlar. Bu enzimler katalitik merkezlerinde redox
metaller içerirler ve süperoksit radikallerinin hidrojen peroksite dismutasyonunu
katalizlerler. Reaksiyonu aşağıdaki gibidir:
2H+ + 2O2ֿ H2O2 + O2
Günümüze dek üç farklı SOD izoformu tanımlanmıştır: Sitozolik bakır/çinko-
içeren SOD (Cu/ZnSOD veya SOD-1), mitokondriyal mangan-içeren SOD (MnSOD,
SOD-2) ve ekstraselüler SOD (ecSOD, SOD-3). EcSOD da aslında bakır/çinko-
içeren bir enzim olmasına rağmen genel olarak damar düz kas hücrelerinden
salgılanıp, endoteliyel hücre yüzeyinde bulunan vasküler ekstraselüler matriks
içerisindeki glikozaminoglikanlara bağlanır. EcSOD
Vasküler interstisyumun oksidan durumunun düzenlenmesinde önemli rol
oynamaktadır (196).
1.2.9.2.2. Glutatyon Peroksidaz
Glutatyon peroksidaz (GPx) selenyum içeren antioksidan bir enzimdir.
Hidrojen peroksidin ve lipit peroksitlerin sırasıyla suya ve lipit alkollere
indirgenmesini katalize etmektedir (5). Bu reaksiyon sırasında aynı zamanda

35
indirgenmiş glutatyonu glutatyon disülfite yükseltgemektedir. Ortamda yeterince
glutatyon veya glutatyon peroksidazın bulunmadığı durumlarda hidrojen peroksit ve
lipit peroksitleri detoksifiye edilemez ve Fe+2 gibi geçiş elementleri aracılığı ile
hidroksil radikallerine ve lipit peroksil radikallerine dönüşebilirler (196).
1.2.9.2.3. Katalaz
Katalaz çoğunlukla hücresel peroksizomlarda, az miktarlarda da sitozolde
lokalize olan hücreiçi bir antioksidan enzimdir. Hidrojen peroksidin suya ve
moleküler oksijene dönüşümünü katalize etmektedir. Reaksiyonu aşağıdaki gibidir;
2H2O2 2H2O + O2
Hidrojen peroksidi yok ederek SOD tarafından hidrojen perokside
dönüştürülen süperoksit radikallerini de indirekt olarak detoksifiye etmektedir.
Katalaz enzimi aynı zamanda peroksidaz enzim aktivitesine de sahiptir. Organik
peroksitlerle ve hidrojen donörleri ile reaksiyona girerek onları suya ve organik
alkollere dönüştürür. Yüksek düzeydeki oksidatif streste çok etkindir ve hücreyi
hücre içinde oluşan hidrojen peroksitten korur (196). Katalaz enzimi ortamda
glutatyon içeriğinin veya az GPx aktivitesinin yetersiz olduğu durumlarda özel
öneme sahiptir ve hücrelerin adaptif yanıtında oksidan strese tolerans gelişiminde
büyük rol oynamaktadır (60).

36
1.2.9.2.4. Tiyoredoksin Redüktaz
Tiyoredüksin redüktaz tiyole bağlı hücresel indirgenme süreçlerinde görev
yapan antioksidan bir enzimdir. Bu enzim yükseltgenme reaksiyonlarında redüktör
olarak görev alan yükseltgenmiş tiyoredüksinin tiyoredoksine geri indirgenmesini
katalize etmektedir. Ayrıca lipit hidroperoksitleri indirgeme özelliği de
bulunmaktadır. Glutatyona bağlı disülfit redüksiyonunu katalize eden tiyoredoksin ve
glutaredoksin plazma glutatyon peroksidazın redüktanlarıdır. Bu da ortamda
glutatyon seviyesinin düşük olduğu durumlarda glutatyon peroksidin hidroperoksidi
indirgemesini sağlar. Ayrıca tiyoredeoksinin MnSOD ekspresyonunu artırdığı
gösterilmiştir. Tiyoredoksin sistemi oksidatif stres ile inaktive olan proteinleri etkin
şekilde rejenere edebilmektedir (196)
1.2.10. Oksidan Stres ve Kardiyovasküler Hastalıklar
Artmış serbest oksijen radikallerinin ve lipid peroksidasyonunun, birçok
hastalığın patogenezinde rol aldığı bilinmektedir. Miyokard infarktüsü gibi
kardiyolojik hastalıklar, nörolojik hastalıklar, astım, diabetes mellitus, romatoid artrit
gibi romatolojik hastalıklar, kanser, yaşlanma dahil birçok hastalığın oksidatif stres
ile ilişkisi gösterilmiştir (50, 51,80, 206, 207).
Son yıllarda yapılan birçok çalışma oksidatif stresin hiperlipidemi, diabetes mellitus,
hipertansiyon, iskemik kalp hastalığı ve kronik kalp yetmezliği gibi birçok hastalıkla
bağlantılı bulunan kardiyovasküler disfonksiyonun başlamasında ve gelişiminde

37
önemli rol oynadığını göstermektedir. Oksidatif stres fazla miktardaki reaktif oksijen
türlerinin (ROS) endojen antioksidan sistemlere baskın geldiği durum olarak
tanımlanır. ROS damar yapısındaki herbir hücre türünde farklı fonksiyonel etkilere
sahiptir ve hem fizyolojik hem de patofizyolojik rol oynar (184). Damar yapısındaki
en önemli reaktif oksijen türlerinden biri süperoksittir ( .O2- ). Oksijenin tek
değerlikli oksijene indirgenmesi ile oluşur. Bu reaksiyona NAD(P)H oksidaz,
ksantin oksidaz gibi çeşitli enzim sistemleri aracılık eder. Süperoksit vasküler
fonksiyonda kendi bizzat etkiler gösterebilmekle birlikte diğer reaktif türlerin
oluşumunda da ana etkendir. Süperoksidin NO ile etkileşiminden peroksinitrit oluşur.
Süperoksitin süperoksit dismutaz tarafından dismutasyona uğratılması sonucunda da
daha stabil bir ROS olan hidrojen peroksit oluşur. Hidrojen peroksit daha sonra
katalaz ve glutatyon peroksidaz enzimleri vasıtasıyla suya çevrilir. Hidrojen peroksit
aynı zamanda redükte geçiş metalleri ile reaksiyona girerek yüksek oranda rekatif
olan hidroksil radikaline dönüştürülebilir veya miyeloperoksidaz (MPO) ile
metabolize edilerek hipoklorik asit oluşturabilir. Hemen bütün vasküler hücre tipleri
süperoksit ve hidrojen peroksit üretebilirler. NAD(P)H oksidazlar, ksantin oksidazlar
(XO), nitrik oksit sentazlar (NOS) ve miyeloperoksidazlar (MPO) gibi damar
yapısında bulunan çeşitli enzimatik sistemler süperoksit ve türevlerini üretebilirler.
Bu herbir enzimin biribirine göreli olarak önemi damar yapısının fizyolojik
durumuna bağlıdır (184).
Oksidatif stres NO inaktivasyonu, DNA ve proteinlerin oksidatif
modifikasyonu, lipid oksidasyonu, vasküler hücrelerin mitojenliğinin artması,
vasküler hücrelerin apoptozu gibi birçok hücresel olaya yol açabilir. Ayrıca
kardiyovasküler hastalıkların farklı süreçlerinde görev alan okside LDL , adhezyon

38
molekülleri, kemotaktik faktörler, proenflamatuvar sitokinler hücre siklusu sürecinde
görev alan düzenleyiciler ve matriks metaloproteinazlar gibi redoksa duyarlı genlerin
ekspresyonunda ve aktivasyonunda artışa neden olabilirler (Şekil 1.6). Reaktif
oksijen türlerinin yukarıda bahsedilen olaylara etkisinin ve NO biyokaktivitesinin
azalmasının vasküler hücrelerde ateroskerozun tüm evrelerinde hastalığın gelişmesi
ve ilerlemesinde katkısının olduğu düşünülmektedir (Şekil 1.6).
Şekil 1.6: Reaktif oksijen türlerinin etkileri (www.tkd.org.tr)
Ateroskerotik plaklarda hastalığa sahip arter duvarının tüm katmanlarında
reaktif oksijen türlerinin düzeyinin arttığı çeşitli çalışmalarda gösterilmiştir (45, 152,
183). Gerek ateroskerotik hayvanlarda, gerek insanlarda, vasküler hasar sonrası

39
vasküler NAD(P)H oksidazın bazı altünitelerinin upregüle olduğu ve enzim
aktivitesinin arttığı gösterilmiştir (173, 177, 183). NAD(P)H okidazdaki bozulmanın
yanı sıra, koroner arter hastalığında gerek koroner arterlerde gerekse plazmada
ksantin oksidaz aktivitesinin arttığı ve ecSOD aktivitesinin de azaldığı gösterilmiştir
(106, 177). Bütün bu bulgularla birlikte diğer birçok çalışmanın bulguları bir araya
getirildiğinde, oksidan strese neden olan oksidan ve antioksidan enzimlerin
modülasyonuun aterosklerotik hastalığın patojenezinde önemli rol oynadığı
görülmektedir. Bunların yanında GPx aktivitesindeki azalma ve miyeloperoksit
düzeyindeki yükselmenin de koroner arter hastalığına sahip hastalardaki
kardiyovasküler olaylarda bağımsız risk faktörleri olduğunu göstermektedir (18).
1.2.10.1.Restenoz ve Serbest Oksijen Radikalleri
Restenoz anjiyoplasti sonrası karşılaşılan önemli bir komplikasyondur. Bu
patolojik süreçte vasküler düz kas hücreleri apoptoza, proliferasyona ve migrasyona
uğrarlar. Adventisyal fibroblastlar da miyofibroblastlara dönüşüp neointimaya göç
ederek sürece katkıda bulunurlar (201). Serbest oksijen radikalleri de bu fenotipik
değişikliklere neden olarak restenozun patojenezine katkıda bulunabilirler.
Balon hasarı sonrası özellikle medyal ve neointimal düz kas hücrelerinde ve
adventisyel fibroblastlarda süperoksit üretimi artmaktadır. Neointima gelişiminin
antioksidanlar ile inhibe olması, serbest oksijen radikallerinin önemini
göstermektedir (9). Balon hasarı sonrası reaktif oksijen türlerinin gelişiminden

40
sorumlu oksidazlar tam olarak aydınlatılamamak ile birlikte NAD(P)H oksidazın rolü
ortaya konmuştur (169).
1.2.11. Oksidan Stres ve Telomerler
Hücre doğal hücre bölünmesi sonucu yaşlanma sürecine girebildikleri gibi
(replicative senescence), oksidatif stres, radyasyon, DNA hasarına neden olan çeşitli
kimyasallar gibi stres oluşturan çeşitli fakörlere yanıt olarak da “stres kaynaklı
prematüre yaşlanma” (SIPS) sürecine girebilirler (36, 62, 181).
Süperoksit anyonları, hidrojen peroksit ve hidroksil radikallerinin hücre
içerisinde üretilmesi DNA kırıklarına ve çeşitli DNA bazlarının modifikasyonuna
neden olabilir (45, 83, 147). Bunlardan en önemlisi guanin rezidülerinin yüksek
oranda mutajenik olduğu 7,8 dihidro 8-oxo-guanin (8-oxoG)’dir (62).
Telomerler yüksek guanin içerikleri nedeni ile oksidatif stresin neden olduğu
hasara yüksek oranda duyarlılık gösterirler. Bir hücrenin ömrü süresince bazlara
yönelik oksidatif hasarın biriktiği ve hücre yaşlanmasına önemli etkisinin olduğu
bildirilmiştir (143, 150, 158, 191, 193). Reaktif oksijen türleri, özellikle de hidroksil
radikalleri, tekli sarmal kırıklarına neden olurlar (85). Telomerik DNA; genomik
DNA’dan farklı olarak tek sarmal kırıklarını tamir edememektedir. Bunun sonucu
olarak da özellikle reaktif oksijen türlerinin neden olduğu 8-oxodG kırıkları birikime
uğramaktadır (143, 193). Tamir edilemeyen bazların varlığı replikasyonu
etkilemektedir. Bu durum oksidaitif stresin telomer kısalmasını arttırmasının bir

41
nedeni olarak önerilmiştir (192). Telomerik DNA’nın tamir edilememesine bir
açıklama olarak TRF2’nin telomerlere bağlanmasının DNA tamir enzimlerinin
telomerik sarmal kırıklarına ulaşamaması gösterilmiştir (151).
Autosomal Resesiv Ataxia Telangiectasia (A-T) kronik oksidatif stres ve
telomer kısalması arasındaki ilişkiye kanıt oluşturabilecek bir hastalıktır. A-T
immünyetmezlik ve prematüre yaşlanma ile karakterize bir rahatsızlıktır. Bu
hastalıktan sorumlu olan gen “Ataxia telangiectaisa mutasyona uğramış gen” (ATM)
‘dir. ATM geni normalde DNA kırıklarına yanıt olarak aktive olan ve kromozomal
stabilite ve temomer bütünlüğünün korunması için gerekli bir gen olduğu
düşünülmektedir. A-T hastalarında hücrler kronik bir oksidatif stres durumu
içerisindedirler ve bu nedenle de telomerik DNA kırıklarını onaramazlar ve telomer
kısalmasının artmasına neden olurlar (185).
Passos ve ark. tarafından yapılan bir çalışmada genç ve yaşlı insan
fibroblastları karşılaştırılmıştır. Karşılaştırmanın sonucunda yaşlanmış hücrelerde
reaktif oksijen türlerinin daha yüksek seviyelerde olduğu, mitokondriyal
disfonksiyon olduğu, daha fazla DNA çift-sarmal kırıklarına ve daha kısa telomerlere
sahip oldukları görülmüştür (142)
Glutatyon redoks siklusunun bozulması ile oluşurulan hafif oksidatif stresin
insan umbilikal ven endoteliyel hücrelerinde telomeraz aktivitesinin
downregülasyonunu hızlandırdığı, telomer aşınmasını artırdığı ve replikatif yaşlanma
sürecini prematüre olarak başlattığı gösterilmiştir (103).

42
Hücre kültüründe yapılan farklı çalımalarda çeşitli antioksidanların ortama
ilave edilmesinin hücre ömrünü artırdığı, telomer kısalmasını ve yaşlanma sürecini
yavaşlattığı gösterilmiştir (56, 94).
Homosisteinin aterojenik etkiler göstermektedir ve bunun da hidrojen peroksit
oluşumunu artırarak yaptığı önerilmiştir. Homosisteine maruz bırakılan insan
umblikal ven endotel hücrelerinde telomer kısalma hızının arttığı ve bu etkinin
katalaz ilavesi ile zayıfladığı gösterilmiştir. Kronik olarak homosistein alan bu
hücrelerde ayrıca intraselüler adhezyon molekülü-1 (ICAM-1) ve plazminojen
aktivatör inhibitörü 1 (PAI-1) ekspresyonunun arttığı gözlemlenmiştir. Bu markırlar
yaşlanmış hücrelerde çok fazla eksprese edilmektedir. Bunun da ateroskeleroza
katkısının olabileceği düşünülmektedir (205).
1.2.12 Glutatyon ve BSO
Glutatyon (GSH) glutamat, sistein ve glisinden oluşan bir tripeptiddir.
Glutamat ve sistein arasında bir γ-peptid bağı içerir (Şekil 1.7 ). Bu bağ GSH’ı çoğu
peptidazın hidrolizinden korur. GSH çoğu memeli hücresinde bulunur ve hücre
içerisinde en yaygın olarak bulunan tiyoldür (0.2-10 mM). GSH hücre içerisinde
NADPH’a bağımlı bir enzim olan glutatyon disülfit (GSSG) redüktaz tarafından tiyol
formunda tutulur. GSH birçok öemli hücresel fonksiyonlara sahiptir (Şekil 1.8). GSH
bir koenzim olarak görev alır ve amino asid transportuna katılır. Ayrıca askorbik
asidin redükte formda tutulmasında ve ribonükleik asitlerin oluşumunda görev alır.
Toksik bileşikler ile enzimatik veya non-enzimatik olarak etkileşerek GSH

43
konjugatları oluşturur. Bunun yanında metabolik olaylarda doğal olarak oluşan
reaktif oksijen türlerinin neden olduğu oksidatif hasara karşı korur. GSH reaktif
oksijen türleri ile non-enzimatik olarak reaksiyona girebilir. Hidrojen peroksit ve
diğer peroksitler de gluatyon peroksidaz ile detoksifiye olurlar (5).
Şekil 1.7: Glutatyonun (GSH) kimyasal yapısı (L-γ-glutamil-sisteinil glisin)
Şekil 1.8: Glutatyon metabolizması

44
1.2.12.1. Hücresel GSH Düzeylerinin Düzenlenmesi
Glutatyon biyosentezinden sorumlu olan γ-glutamilsistein sentetaz ve GSH
sentetaz enzimerinin doğuştan eksikliği GSH eksikliğine yol açan nedenlerden
biridir. Hemolitik anemi ve nörolojik semptomlar ile karakterizedir. GSH
eksikliğinin etkilerinin tam olarak anlaşılabilmesi amacıyla deneysel olarak in vivo
GSH eksikliğinin oluşturulduğu çalışmalara gerek duyulmuştur. Metiyonin
sülfoksimin (MSO) yapısal olarak γ-glutamil bir bileşik olan glutamine
benzemektedir ve glutamin sentetazı inhibe ettiği gösterilmiştir (157).
Glutaminsentetaz ve γ-glutamilsistein sentetaz benzer reaksiyonları katalizlerler. Bu
nedenle de MSO aynı zamanda γ-glutamilsistein sentetazı da inhibe etmektedir. (69).
MSO’ya benzer diğer bir molekül ise bütiyonin sülfoksimindir ve γ-glutamilsistein
sentetazı selektif olarak inhibe eder (70). GSH çoğu hücrede dışarı taşınır, bu
nedenle BSO verilerek GSH sentezi inhibe edildiğinde GSH düzeyleri azalır.
Rodentlerin ve/veya hücrelerin BSO’ya maruz bırakılması onları radyasyon,
isklofosfamit, civa iyonları, kadmiyum iyonları ve sisplatine karşı duyarlı hale
getirir. (180). İnsan lenfositleri ve T-hücreleri BSO ile muamele edildiğinde
aktivitelerini kaybederler (32). Yeni doğmuş rodentlere BSO verildiğinde katarakt
gelişmektedir (32). Farelere uzun süreli BSO uygulandığında ciddi GSH eksikiği
geliştiği görülmüştür. Mitokondri GSH sentezlemez ve GSH ihtiyacını hücre içi
GSH’tan karşılamaktadır (72). Elektron transport sistemi oldukça etkin olmasına
rağmen bir kısım ROS sızıntıya uğrayabilir. Mitokondrilerde katalaz enzimi
bulunmaması sebebiyle, ROS toksisitesinden korunmak için GSH peroksidaza ve
GSH’ın non-enzimatik reaksiyonlarına bağımlıdır. Uzun süreli BSO uygulaması ile
GSH düzeyleri ciddi anlamda tüketildiğinde mitokondri şişer, vakuoller oluşur ve

45
endojen oksidatif stres modelini oluşturur (117, 118). Oluşan bu oksidatif hasar GSH
esterleri ve askorbat ile önlenebilir. (118).
Düşük GSH düzeyleri HIV, hepatit C, Tip II diyabet, ülseratif kolit, erişkin
solunum güçlüğü sendromu (ARDS) ve katarakt gibi birçok hastalığın patolojisi ile
ilişkilendirilmiştir (5). GSH reaktif oksijen türlerine karşı korunmada önemli bir
proteindir. Hücredeki GSH düzeylerini arttırılması amacıyla çeşitli bileşikler
önerilmiştir. Bunlardan N-asetil sistein, 2-oksotiyazolidin-4 karboksilat gibi sistein
prekürsörü maddeler GSH sentezinde görevli enzimlere ihtiyaç duymaktadırlar ve
sentezlenen GSH hücre dışında degrade olmakta, bir orana kadar hücre içerisine
alınmaktadır. Bu nedenle GSH analogları olan GSH esterler önerilmiştir (5). Bu
bileşikler hücreiçine kolayca taşınıp hücrede GSH’a çevrilirler ve birçok dokuya iyi
dağılım gösterirler.
1.2.13. Taurin
2-aminoetansulfonik asit olarak da bilinen taurin çoğu memeli dokusunda
milimolar konsantrasyonlarda bulunan serbest bir amino asittir (21). Hayvan
dokusunda en bol bulunan serbest amino asitlerden olmasına karşın bazı alg türleri
dışında bitkilerde bulunmamaktadır (88). İnsanlarda ise karaciğerde metiyonin ve
sisteinden sentezlenebilmesine rağmen daha çok taırin transporter Tau (TauT) yolula
gıdalardan alınması gerekmektedir. (86). Taurin hiçbir memeli proteininin
komponenti olmamakla birlikte insanlarda gelişme sürecinde önemli rol
oynamaktadır (179). Osmoregülasyon, immünomodülasyon, ve safra tuzu oluşumu

46
gibi birçok fizyolojik olayda yer almaktadır. Son zamanlarda yapılan birçok çalışma
diyetle taurin alımının gerekliliğine işaret etmektedir (21)
Diyabetes mellitus’ta taurinin etkileri yaygın olarak çalışılmıştır. Bir tip II
diyabet modeli olan yüksek fruktoz ile beslenmiş sıçanlarda taurinin insülin
rezistansını, hiperinsülinemiyi, lipid peroksidasyonunu, glukoz serum
konsantrasyonunu azalttığı gösterilmiştir (131, 132, 133). Taurinin aynı zamanda
insanlardan saflaştırılan insülin reseptörüne insülinin bağlanma etkilerini
potansiyalize ettiği gösterilmiştir (120). Bu da daha iyi bir metabolizma kontrolünü
sağlamaktadır. Diyabetik sıçanlar ve obez sıçanlarda taurin uygulaması serum lipid
konsantrasyonundaki azalma ile ilişkilendirilmiştir (210). Taurinin aynı zamanda
Langerhans adacıklarında bulunduğu ve insülin sekresyonunu stimule ettiği fetal β
hücrelerde in vitro olarak gösterilmiştir (38). In vivo yapılan birçok çalışma ise taurin
uygulamasının diabetes mellitusun neden olduğu retina, lens, ve periferal sinirlerdeki
değişiklikleri azalttığını göstermektedir (55)
Taurinin antioksidan etkilerini gösteren birçok çalışma yapılmıştır (21). Taurin
reaktif oksijen türlerine karşı direk süpürücü etki gösterememekte fakat diğer
hücresel antioksidan fonksiyonları artırmaktadır. Nötrofil ve lökositlerde daha stabil
bir formu olan taurin-kloramine (Tau-Cl) dönüşmektedir. Tau-Cl lökositlerde ve
makrofajlarda süperoksit anyonunun ve IL-6 ve Il-8 gibi pro-enflamatuvar
sitokinlerin oluşumunun önlenmesi amacıyla TNF-α oluşumunu ve
lipopolisakkaritlerin neden olduğu NO üretimini baskılamaktadır (165, 202). Fare
demir-yükleme modelinde taurinin oksidatif stresi azalttığı ve kardiyovasküler
fonksiyonları düzelttiği gösterilmiştir (139).

47
Taurinin önemli biyolojik aktivitelerinden biri de safra tuzu oluşumudur. Safra
tuzları barsak sindirimi ve lipidlerin absorpsiyonu için esastır. Taurin ve glisin
kolesterol türevleri ile konjugat oluşturmaktadır. İnsanlarda tauroşölat plazmadan
kolesterolün temizlenmesine neden olan ana safra tuzudur. Sonuç olarak, taurinin
vücutta daha az bulunması, kolesterolün kandan daha az temizlenmesi dolayısıyla
daha çok birikmesi anlamına gelmektedir. Bu da ateroskleroz riskini artırmaktadır
(21).

48
2. GEREÇ VE YÖNTEM
2.1. Tavşanların Gruplandırılması
Deneylerde her iki cinsten (n=40) ortalama 2,5-3 kg ağırlığındaki Yeni Zelanda
tipi beyaz tavşanlar kullanılmıştır. Deney hayvanları protokol gereği tüm deney
süresince ayrı kafeslerde tutulmuş, standart yem ile beslenmişler ve serbestçe su
içmelerine olanak sağlanmıştır. Çalışmanın amacı ve kapsamı doğrultusunda
deneyler balon hasarı sonrası vasküler düz kas hücrelerinde telomer devamlılığının
saptanmasına (Bölüm I) ve oksidan stresin etkisinin araştırılmasına yönelik (Bölüm
II) olmak üzere iki bölümde gerçekleştirilmiştir.
Bölüm I
Balon hasarı sonrası vasküler düz kas hücrelerinde telomer devamlılığı ve
telomeraz aktivitesinin incelenebilmesi amacıyla farklı lezyon süreleri sonrasında (0.
gün, 2. gün, 7. gün ve 14. gün) hasarlı damar ve karşı damar izole edilmiştir. Bu
bölümdeki deneylerde çalışma protokolü uyarınca telomer uzunluğu ve telomeraz
ekspresyonu için doku homojenatı hazırlanması ile morfometrik ve
immünohistokimyasal analizler için doku kesitlerinin alınabilmesi amacıyla iki farklı
seri deney grubu oluşturulmuştur:

49
Seri I: Birinci seri deneylerden elde edilen dokular telomer uzunluğu ve telomeraz
ekspresyonu için doku homojenatı elde edilmesinde kullanılmıştır. Bu seride
kullanılan tavşanlar ayrıca oluşturulan lezyonun süresine göre 0. gün (n=3), 2. gün
(n=3), 7. gün (n=3) ve 14. gün (n=3) olmak üzere 4 gruba ayrılmıştır.
Seri II: İkinci seri deneylerden elde edilen dokular morfometrik ve
immünohistokimyasal analizler için kullanılmışlardır. Bu seride yine ilk seride
olduğu gibi tavşanlar ayrıca oluşturulan lezyonun süresine göre 0. gün (n=4), 2. gün
(n=4), 7. gün (n=4) ve 14. gün (n=4) olmak üzere 4 gruba ayrılmıştır.
Bölüm II
Balon hasarı sonrası vasküler düz kas hücrelerinde telomer uzunluğuna
oksidatif stresin etkisinin araştırılması amacıyla 4 farklı deney grubu
oluşturulmuştur:
1. Bütiyonin sülfoksimin, BSO grubu (n=3): Bu gruptaki tavşanlara oksidan stres
oluşturmak amacıyla daha önceden ön denemeler ile 14 günlük uygulama sonunda
yaklaşık %40-50 oranında GSH inhibisyonu sağladığı belirlenen 75 mg/kg/gün
dozda, s.c. yoldan BSO uygulanmıştır.
2. Taurin grubu (n=3): Bu gruba antioksidan bir madde olduğu bilinen taurin içme
suyunda %10 (ağırlık/hacim) oranında uygulanmıştır.
3. BSO + Taurin grubu (n=3): Bu gruptaki tavşanlara aynı yoldan ve aynı dozda
BSO ve taurin verilmiştir.
4. Kontrol grubu (n=3): Bu gruptaki tavşanlara deney süresince herhangi bir tedavi
uygulanmamıştır.

50
Şekil 2.1’de oluşturulan bütün deney grupları ve oluşturulan deney gruplarına
göre yapılan işlemlerin şematik olarak özeti verilmiştir.
Tavşanların Gruplandırılması (n=40)
Bölüm 1•Gruplar: 0., 2., 7., 14. günSeri I: Telomer boyu icin TRF
Telomeraz aktivitesi için TRAP analiziSeri II: Morfometri ve İmmünohistokimya
Balon Hasarı Oluşturulması
Bölüm 2•Gruplar: BSO, Taurin, BSO+Taurin, Kontrol (14 gün)
-Telomer boyu icin TRF analizi-Morfometri ve İmmunohistokimya- 0., 8. ve 14. günlerde oksidan stres markırlarının
belirlenmesi amacıyla kan örneklerinin alınması
Tavşanların Gruplandırılması (n=40)
Bölüm 1•Gruplar: 0., 2., 7., 14. günSeri I: Telomer boyu icin TRF
Telomeraz aktivitesi için TRAP analiziSeri II: Morfometri ve İmmünohistokimya
Bölüm 1•Gruplar: 0., 2., 7., 14. günSeri I: Telomer boyu icin TRF
Telomeraz aktivitesi için TRAP analiziSeri II: Morfometri ve İmmünohistokimya
Balon Hasarı OluşturulmasıBalon Hasarı Oluşturulması
Bölüm 2•Gruplar: BSO, Taurin, BSO+Taurin, Kontrol (14 gün)
-Telomer boyu icin TRF analizi-Morfometri ve İmmunohistokimya- 0., 8. ve 14. günlerde oksidan stres markırlarının
belirlenmesi amacıyla kan örneklerinin alınması
Şekil 2.1.: Deney şeması
2.2. Balon Hasarı Oluşturulması
Balon hasarının etkin ve doğru bir şekilde uygulanabilmesi için Ege
Üniversitesi Tıp Fakültesi Radyodiagnostik Anabilim Dalında ön denemeler
yapılmıştır. Bu amaçla deneylerde kullanılacak tavşanlara Prof. Dr. İsmail Oran
tarafından anjiyografi uygulanmıştır. Bir cetvel yardımıyla tavşanların femoral ve
ilyak arterlerinin yerleşimi belirlenmiştir (Resim 2.1.A ve B). Bu şekilde balon

51
anjiyoplasti uygulanması sırasında kılavuz tel ile damar içerinde kaç cm ilerlenmesi
gerektiği belirlenmiştir.
A) B)
Resim 2.1.: Tavşan ilyak arter (A) ve femoral arter (B) anjiyogramı.
Balon anjiyoplasti uygulama sürecinde, deney hayvanları, deney başlangıcında
sodyum pentobarbital (25 mg/kg, iv) ile anestezi edilmişlerdir. Femoral arterleri
cerrahi girişim ile açığa çıkarılmış ve 21G branül yardımı ile damar içerisine
girilmiştir (Resim 2.2.). Branül içerisinden damar içerisine kılavuz tel ile girildikten
sonra tel damar içerisinde ilerletilmiştir. Daha sonra branül çıkarılarak 2 cm
uzunluğunda 3 mm çapında Fogarty balon dilatasyon kateteri kılavuz tel aracılığı ile
damar içerisine girilmiştir (Resim 2.3.). Kateter ilyak arterde ön denemelerde
saptanan miktar kadar ilerletilmiş ve balon hasarı uygulanmıştır. Bunun için balon
sırasıyla 4 kez 6, 8, 4 ve 10 atmosfer basınçla 1 dakika boyunca şişirilmiştir. Daha
sonra balon çıkarılmış ve insizyon cerrahi olarak dikilerek kapatılmıştır. Diğer

52
femoral arterde ise aynı işlemlerle insizyon yapılmış, ancak balon kateterle
girilmeksizin kapatılmıştır (178). Cerrahi girişim sonrası femoral arter ligate edilip
kesilen dokular cerrahi iplikle dikilerek kapatılmıştır. Balon anjiyoplasti uygulanması
sonucu tavşanlara 14 gün süre ile yukarıda belirtildiği biçimde BSO, taurin ve
BSO+taurin uygulaması sürdürülmüştür.
Resim 2.2.: Bir branül yardımı ile femoral artere girilmesi

53
Resim 2.3. :Balon kateterinin damar içerisinde kılavuz tel yardımı ile
ilerletilmesi
2.3. Doku Örneklerinin İzolasyonu
1. Bölüm’deki deneyler kapsamında daha önce de belirtildiği gibi telomer
uzunluğu ve telomeraz aktivitesi analizleri için (Seri I) ve morfometri ve
immünohistokimya analizleri için (Seri II) olmak üzere 2 farklı deney serisi
oluşturulmuştur. Belirlenen süreler sonunda (0. 2. 7. ve 14. günler) tavşanlar aşırı
doz sodyum pentobarbital ile öldürülmüş sağ ve sol ilyak arterleri izole edilmiştir.
Damarların izolasyonu sonrası yapılan işlemler gerçekleştirilecek analize göre 2
seride farklılık göstermektedir:
Seri I: İzole edilen damar segmentleri çevrelerindeki bağ dokusundan
temizlenmişlerdir. Daha sonra damar dokuları telomer ve telomeraz enzimi analizi

54
yapılmak üzere 5-6 mm uzunluğunda iki eşit halkaya ayrılmıştır. Elde edilen damar
örnekleri ayrı ayrı düşük ısı ve yüksek basınca dayanıklı önceden darası alınmış
cryotüplere konarak tartılmış ve derhal sıvı azotta dondurulmuştur. Örnekler 3-4 saat
sonra sıvı azot tankından çıkarılarak derin dondurucuya (-80ºC) alınmış ve analiz
gününe dek derin dondurucuda bekletilmiştir.
Seri II: Bu seride damarlar izolasyon sonrası çevrelerindeki bağ dokulardan
temizlenmiş ve morfometrik ölçümler ve immünohistokimyasal incelemeler
yapılmak üzere %4’lük formalin ile fikse edilmişlerdir.
2. Bölümde ise telomer uzunluğu ile morfometri ve immünohistokimya
analizleri için farklı seriler tasarlanmamış, izole edilen damar segmentlerinin 2/3’ü
telomer analizi için kullanılmak üzere 1. Bölüm’dekine benzer şekilde derin
dondurucuda (-80ºC) bekletilmiştir. 1/3’lük kısmı ise morfometrik ölçümler ve
immünohistokimyasal incelemeler yapılmak üzere %4’lük formalin ile fikse
edilmiştir.
2.4. Telomerik Tekrar Restriksiyonel Fragmantasyon Deneyleri: Terminal
Restriction Fragments (TRF)
Bu deneyler Bölüm 1’de Seri I sonucu elde edilen ve -80ºC’de saklanan damar
segmentlerinin her bir damara ait ringlerden birer tanesi kullanılarak Bölüm 2’de ise
morfometri ve immünohistokimya için kullanılacak damar segmenti ayrıldıktan
sonra geri kalan ve -80ºC’de saklanan 2/3’lük damar segmentinde

55
gerçekleştirilmiştir. TRF deneylerinde amaç telomerik fragmanların uzunluğunun
saptanmasıdır. Deney basamakları kısaca aşağıdaki gibidir (100).
TRF Deney Basamakları:
1. Dokuların homojenizasyonu
2. Homojenize edilen dokulardan DNA izolasyonu
3. DNA miktarının belirlenmesi
4. Kromozomal DNA’nın agaroz jelde yürütülmesi ve görüntülenmesi
5. Kromozomal DNA’nın restriksiyon enzimleri ile kesilmesi
6. Kesilen fragmanların agaroz jelde yürütülmesi
7. Southern Blot yöntemi ile DNA’nın jelden membrana transfer edilmesi
8. Membrana transfer olan DNA’nın telomerik oligonükleotid ile hibridizasyonu
9. Radyografi filmine aktarılarak görüntülenmesi
2.4.1. Dokuların Homojenizasyonu
Derin dondurucudan çıkarılan dokular çözündürülüp steril bir petri kabı
içerisinde steril bir bistüri yardımıyla küçük parçacıklara ayrılmıştır. Daha sonra
doku parçacıkları içerisinde DNA izolasyonu için gerekli olan liziz tamponunu
içeren tüplere (eppendorf) aktarılmıştır. Parçalanma ve homojenizasyonu sağlamak
amacıyla tüp içerisine steril tungsten boncuklar ilave edilmiş ve homojenizatörde
(Qiagen TissuLyser®- Retech, Haan, Almanya) homojenize edilmiştir.

56
2.4.2. DNA İzolasyonu
DNA izolasyonun gerçekleştirilmesi amacıyla Qiagen DNeasy® Tissue Kit’i
(Haan, Almanya) kullanılmıştır. Bu yöntem ile DNA izolasyonu DNA’nın silica-gel
membrana bağlanarak diğer fragmanlardan ayrılması esasına dayanır. Klasik DNA
izolasyon yönteminde olduğu gibi DNA’nın ekstraksiyonu ve etanolle çöktürülme
işlemini içermez.
Temel çalışma prensibi: Örnekler liziz tamponu içerisinde homojenize
edildikten sonra proteinaz K çözeltisi ilave edilerek hücrelerin liziz olması sağlanır.
Daha sonra lizat DNeasy mini spin kolonlara yüklenir ve santrifüj edilir. Santrifüj
işlemi sırasında DNA selektif olarak silika membrana bağlanır. Diğer kontaminantlar
ve enzim inhibitörleri ise membrandan geçer. Kalan kontaminantlar ve enzim
inhibitörlerinin ise iki basamaklı yıkama işleminden geçirilerek uzaklaşmaları
sağlanır. Daha sonra membrana bağlanmış halde olan DNA, TE (Tris-EDTA)
tamponu (10mM Tris.Cl, pH 7.5 + 1mM EDTA, pH 8.0) içerisinde ayrıştırılır.
2.4.3. DNA Miktarının Belirlenmesi
İzolasyonu sonucu elde edilen elüattaki DNA’nın nanogram/mikrogram
düzeyinde belirlenmesi amacıyla spektrofotometrik yöntem kullanıldı. Bu amaçla
elde edilen DNA örnekleri önceden belirlenen oranlarda sulandırılarak UV geçirgen
kuvartz mikroküvetlerde 260nm dalga boyundaki absorbans değerleri okundu.

57
Absorbans değerlerinin 260 nm dalga boyunda okunmasının sebebi nükleotidlerin
heterosiklik halkalarının 260 nm dalga boyunda maksimum absorbans göstermesidir.
Çift zincirli DNA molekülleri için, 1 optik dansitenin (OD) 50 µg/ml’ye
karşılık geldiği bilinmektedir. Buna göre çift zincirli DNA için miktar
belirlenmesinde aşağıdaki formül kullanılmıştır:
DNA (µg/ml) = A260 x seyreltme oranı x 50
Tablo 2.1.: Damar örneklerinden elde edilen DNA konsantrasyonları
Örnek No Absorbans (A260) Konsantrasyon (µg/ml)
1 0.063 63
2 0.044 44
3 0.080 80
4 0.044 44
5 0.046 46
6 0.045 45
7 0.042 42
8 0.048 48

58
2.4.4. DNA’nın Agaroz Jelde Yürütülmesi ve Görüntülenmesi
2.4.4.1. DNA’nın Jel Elektroforezinin Temeli
Kromozomal DNA’nın jel üzerinde görüntülenmesi ve restriksiyon öncesi
spektrofotometre ile belirlenen miktar tayininin doğrulanması amacıyla DNA
örneklerinin bir kısmına agaroz jel elektroforezi uygulanmıştır.
Bu yöntemle UV ışığı altında fluoresan etki gösteren etidyum bromür (EtdBr)
boyası kullanılarak DNA’nın jel üzerindeki yerini belirlemek mümkündür. DNA’nın
elektroforetik analizinin temeli, elektriksel bir alanda, jel üzerindeki göçüne dayanır.
Bu göç hızı molekülün büyüklüğüne, yapısına, jelde kullanılan maddenin
konsantrasyonuna, iyonik kuvvete ve uygulanan akıma bağlı olarak değişmektedir.
Jel elektroforezinde kullanılan destekleyici madde (matriks) mekaniksel etkiyi
engellemek ve molekülleri büyüklüklerine göre ayırmak için kullanılır. Poliakrilamid
jeller genellikle küçük DNA fragmanları (5-500 baz çifti) ve RNA moleküllerinin
ayrımında kullanılır. Bu tip jeller, ayırım güçleri çok fazla olduğundan, boyut
bakımından birbirine çok yakın DNA fragmanlarını analizleri için daha uygundur.
Nükleik asitler için en çok kullanılan yöntem agaroz jel elektroforezidir.

59
2.4.4.2. Agaroz Jel Elektroforezi
Agaroz, bir kırmızı alg türü olan Agar agar’dan izole edilen doğrusal bir
polisakkariddir. Agaroz sıcak suda çözünür ve soğutulduğu zaman polimerde
karşılıklı hidrojen bağlarının oluşumu sonucu jel yapısı oluşur. Bu oluşum geri
dönüşümlüdür. Agaroz konsantrasyonu değiştirilerek jelin por çapı ayarlanabilir.
Böylece, küçük DNA fragmanları için yüksek, büyük DNA fragmanları için düşük
agaroz konsantrasyonu kullanılarak DNA’nın jelde en uygun şekilde yürümesi
sağlanabilir. DNA’nın jelde görünür hale gelebilmesi EtdBr’ün interkalasyon özelliği
ile DNA bağları arasına bağlanarak 300 veya 360 nm’de ışığı absorbe etmesi sonucu
flüoresan etki göstermesi ile olur. Bu etki DNA konsantrasyonuna bağlı olarak
kuvvetli veya zayıf olabilir.
Tablo 2.2.: Değişik boyutlarda DNA fragmanlarının ayırımı için uygun agaroz
konsantrasyonları
Agaroz konsantrasyonu (% ağırlık/hacim) Doğrusal DNA molekülü (kb)
0.3 5.0 – 60.0
0.6 1.0 – 20.0
0.7 0.8 – 10.0
0.9 0.5 – 7.0
1.2 0.4 – 6.0
1.5 0.2 – 3.0
2.0 0.1 – 2.0

60
2.4.5. DNA’nın Restriksiyon Enzimleri ile Kesilmesi
DNA’nın enzimatik kesimi “restriksiyon endonükleazları” kullanılarak yapılır.
Restriksiyon endonükleazları, çift iplikli sarmal DNA’da özel nükleotid dizilerini
tanıyan ve DNA’nın her iki ipliğini kesen enzimlerdir. Bakterilerde bulunan bu
enzimler DNA’da özgün kısa dizileri tanır ve kesme işlemini enzime göre değişmek
üzere, tanıma yerinde veya bu yerin dışında başka özel bir dizide gerçekleştirirler.
Kesim sonucunda küt veya yapışkan uçlu DNA fragmanları oluşur. Günümüzde
1000- 1200 restriksiyon enzimi ticari olarak üretilmektedir. Bir ünite restriksiyon
enzim aktivitesi, uygun koşullar altında 1 µg DNA’yı kesebilen enzim olarak
tanımlanmaktadır. Restriksiyon enzimlerinin büyük çoğunluğu %50 gliserol içeren
tamponlarda ve -20˚C’de saklanır. Enzimlerin aktif olarak çalışacağı koşulları
sağlayan özel tamponlar genellikle 10X konsantrasyondadır ve kullanım sırasında 1x
şeklinde sulandırılır. Restriksiyon enzimlerinin çoğu 37˚C’de aktifken, bazıları
60˚C’de optimum aktivite gösterir. Kural olarak 20 µl reaksiyon karışımı içerisinde
0.2-1 µg DNA bulunmalıdır. DNA konsantrasyonu fazla ise reaksiyon hacmi
artırılmalıdır.
Star Aktivitesi: Restriksiyon enzimlerinin uygun olmayan koşullarda özel kesim
dizilerine benzer dizilerde kesim yapmaları sonucunda istenmeyen fragmanların
oluşması olayıdır.

61
DNA izolasyonu sonucu elde edilen DNA örneklerinden telomerik restriksiyon
fragmanlarının saptanması amacıyla bu ve bundan sonraki basamaklarda
TeloTAGGG “Telomere Length Assay” Kit (Roche, Almanya) kullanılmıştır.
Dokulardan izole edilmiş olan genomik DNA örnekleri öncelikle HinfI ve RsaI
restriksiyon enzimleri ile muamele edildiler. Bu iki enzimin özelliği telomerik
DNA’yı ve subtelomerik DNA’yı spesifik tekrar dizi (TTAGGG) özelliğinden dolayı
kesmeyip, DNA’nın geri kalan kısımlarını (non-telomerik kısımlar) düşük moleküler
ağırlık fragmanlarına kadar kesmeleridir. Telomer boyunun saptanmasına yönelik
deneylerde sıklıkla bu 2 enzim birlikte kullanılır. Bu amaçla 1-2 µg DNA final hacmi
17 µl olacak şekilde nükleaz içermeyen su ile seyreltildi. HinfI ve RsaI enzimlerinin
eşit hacimde karışımları hazırlandı. Her bir tüpe 2 µl kesim tamponu ve 1 µl enzim
karışımı ilave edildi ve karıştırıldı. Reaksiyon karışımı 37˚C’de 2 saat inkübe edildi.
İnkübasyon süresi sonunda reaksiyonu durdurmak amacıyla tüp içerisine 4 µl jel
elektroforezi yükleme tamponu (5X) kondu ve karıştırıcıda karıştırıldı.
2.4.6. Restriksiyon Enzimleri ile Kesilmiş DNA Fragmanlarının Agaroz Jelde
Yürütülmesi
2.4.6.1. Agaroz Jelin (%0.8) Hazırlanması
0.8 mg agaroz tartıldı. 100 ml 0.5xTBE (Tris-borat EDTA tamponu) içerisinde
mikrodalga fırında çözüldü. Elle tutulacak sıcaklığa (50-55˚C) ulaştığında kenarları
kapatılmış yatay elektroforez kabına sızıntı ve hava kabarcığı olmamasına dikkat

62
edilerek döküldü. Cepleri oluşturacak tarak yerleştirilerek donması beklendi. Tarak
ve kenarlıklar dikkatlice uzaklaştırılarak jel elektroforez tankına yerleştirildi.
2.4.6.2. Jelin Yürütülmesi
Her bir kuyucuğa 1-2 µg DNA içerecek şekilde restriksiyon enzimleri ile
kesilen DNA örnekleri yüklendi. Telomer uzunluğunun sağlıklı bir şekilde
ölçülebilmesi için her bir örneğin eşit miktarda DNA içermesine özen gösterildi.
Yaklaşık 500 ng moleküler ağırlık belirteci, 12 µl nükleaz içermeyen su ve 4 µl 5X
yükleme tamponu karıştırıldı. Telomer uzunluğunun belirlenmesinde referans
oluşturması için örneklerin her iki tarafına ilk ve son kuyucuğa 10’ar µl yüklendi.
Daha sonra jel, 0.5xTBE tamponu içerisinde yükleme tamponu içerisindeki
bromfenol mavisinin neden olduğu mavi boya yükleme yerinden 10 cm uzaklaşana
dek yürütüldü (5V/cm). Daha sonra jel EtdBr (10 µg/ml) ile muamele edilerek
UV’de görüntülendi.
0.5xTBE çözeltisinin hazırlanması
0.5xTBE 5xTBE stok çözeltisinden 1:10 oranında seyreltilerek hazırlandı.
1 litre 5 x TBE için: (0,45 M Tris-borate ve 0,01 M EDTA)
54 g Tris-base
27,5 borik asit
20 ml 0,5 M EDTA

63
2.4.7. Southern Blot Yöntemi ile DNA’nın Jelden Membrana Transfer Edilmesi
Aranılan DNA dizisinin saptanması Southern blot tekniği ile mümkün
olabilmektedir. Bu teknik ilk olarak adını aldığı Southern adlı araştırmacı tarafından
1975 yılında tanımlanmıştır (174). Bu tekniğin temeli, istenilen DNA parçasının ona
tamamlayıcı olan radyoaktif veya radyoaktif olmayan bir takım belirleyicilerle
işaretlenmiş olan problar (tanımlanmak istenen nükleik asit bölgesine tamamlayıcı
olan, belirleyici olarak kullanılan tek iplikli nükleik asit parçaları) kullanılarak
hibridizasyonuna ve bunun ardından belirleyicinin özelliğine göre radyoaktif,
immünolojik, kimyasal ya da flüoresan yöntemlerle görünür hale getirilmesine
dayanır.
Probların radyoaktif işaretlemesinde en çok 32P kullanılır ve radyoaktivite
otoradyografi ile belirlenir. Radyoaktif maddelerle çalışmanın getirdiği zorluklar ve
tehlikeler nedeniyle radyoaktif olmayan işaretleme ve belirleme sistemleri
geliştirilmiştir. Deneylerimizde kullandığımız digoksigenin-anti-digoksigenin sistemi
bu tür sistemlerden biridir. Digoksigenin-anti-digoksigenin sistemi, otoradyografi ile
aynı temel prensibe dayanır. Radyoaktif madde gerektirmemesi, kolay uygulanabilir
olması, hızlı sonuç vermesi, özgül ve duyarlı olması gibi avantajlara sahiptir.
Digoksigenin-anti-digoksigenin sisteminde DNA, RNA ve oligonükleotidlerin
işaretlenmesinde Digitalis bitkisinden elde edilen bir kardenolid steroid hapten olan
digoksigenin kullanılır. Prob hazırlamada kullanılan digoksigenin nükleotid trifosfat
analoğu olan digoksigenin – dUTP (DIG-11-dUTP) yapısında, urasile bağlı olarak
yer alır. Belirlenmesi istenen DNA dizisinin tamamlayıcısı olan prob in vitro sentez

64
edilirken ortamda 4 çeşit dNTP’ye ek olarak DIG-11-dUTP de bulunduğundan yeni
sentezlenen prob, kalıp DNA zincirindeki her adenin nükleotidinin karşılığı olarak ya
dTTP ya da DIG-11-dUTP taşır.
2.4.7.1. Jelin Transfere Hazırlanması
UV’de görüntülenip kontrol edilen agaroz jel temiz bir kaba alındı. Transfer
öncesi sırasıyla aşağıdaki işlem basamakları uygulandı ve bütün basamaklar oda
sıcaklığında ve jel hafif çalkalanarak gerçekleştirildi:
1. Jel 10 dakika bromfenol mavisi sarıya dönene kadar 0.2 N HCl çözeltisi
içerisinde depürinasyona tabi tutuldu.
2. İki kez su ile yıkandı.
3. 2x15 dakika denatürasyon çözeltisinde bekletildi.
4. Tekrar iki kez su ile yıkandı.
5. 2x15 dakika nötralizasyon çözeltisi içerisinde bekletildi.
6. Jeli transfer çözeltisine hazırlamak amacıyla adaptasyon çözeltisi içerisinde
(2xSSC-sodyum sitrat/sodyum klorür) 5 dakika bekletildi.
7. Transfer işlemine geçildi.

65
2.4.7.2. Transfer
Jel üzerindeki DNA kapiler aktarım yoluyla naylon membrana transfer edildi.
Jel düzenek üzerindeki iki ucu tampona değen blot kağıdı (Hybond®-Amersham,
Pharmacia Biotech, Buckingamshire, İngiltere) üzerine ters ve arada hava kabarcığı
kalmayacak şekilde yerleştirildi. Jel ile aynı boyutta kesilen naylon membran
(Hybond-N+ positively charged nylon membrane, Amersham Biosciences, İngiltere)
suda ıslatıldıktan sonra transfer çözeltisi olan 10xSSC’den (sodyum sitrat-sodyum
klorür) geçirildi. Jel üzerine arada hava kabarcığı kalmayacak şekilde kapatıldı.
Üzeri yine aynı boyuttaki blot kağıtları ile örtüldü ve çok katlı kağıt havlular
dizildikten sonra en üste ağırlık konarak gece boyunca (yaklaşık 16 saat) transfere
bırakıldı. Süre sonunda ıslanmış kâğıt havlular ve blot kâğıtları kaldırıldı. Membran
bir köşesinden işaretlendikten sonra pens ile ters çevrilip bir filtre kâğıdı üzerine
alındı. Aktarımın başarılı olup olmadığının anlaşılması amacıyla jel UV’de
görüntülendi. Transferin tam olup olmadığını anlamak için işlem sonrası jelde
belirgin bir bantın kalmadığı kontrol edildi.

66
2.4.7.3. Transfer Öncesi ve Transferde Kullanılan Tamponlar
Tablo 2.3.: Kullanılan tamponlar
Tampon İçeriği Formülasyon
Apürinasyon Tamponu 0.2 N HCl %37’lik HCl’den 1:50
Denatürasyon Tamponu 0.5N NaOH/1.5 NaCl 300 ml 5M NaCl
100 ml 5N NaOH
600 ml distile su (1litre için)
Nötralizasyon Tamponu 1M Tris (pH 7.5) /1.5M NaCl 225 ml 5M NaCl
500 ml 1.5M Tris (pH 7.5)
distile su ile 750 ml’ye
tamamlanır.
Adaptasyon Tamponu 2x SSC 20xSSC’den 1:10 hazırlanır.
Transfer Tamponu 10xSSC 20xSSC’den 1:2 hazırlanır.
20xSSC stok çözeltisi NaCl-Sodyum sitrat tamponu
(pH:7.0)
175,3 g NaCl
88,2 g Sodyum sitrat
800 ml’ye dH2O ile tamamlanır.
pH :7,0’ye ayarlanır.
Final hacmi:1lt.
2.4.8. Membran Üzerine DNA’nın Sabitlenmesi
Transfer olan DNA’nın membrana sabitlenmesi amacıyla “UV-crosslinker”
cihazında 120 mJ’e tabi tutuldu.

67
2.4.9. Prehibridizasyon
Hibridizasyon işlemi sırasında probların membrana ve DNA örneği üzerinde
özgün olmayan bölgelere bağlanmasını en aza indirgemek ya da önlemek amacıyla
hibridizasyon öncesi işlemler yapılır (prehibridizasyon). Bu amaçla 200 cm2’lik bir
membran için 18 ml olacak şekilde önceden 42˚C’ye ısıtılmış DIG Easy Hyb (kit
içerisinde hazır) çözeltisi içerisinde 30-60 dakika 42˚C’de düşük devirde hafif
çalkalanarak “Thermoshaker”da inkübe edildi.
2.4.10. Hibridizasyon
Her 5 ml DIG Easy Hyb çözeltisine 1 µl DIG işaretli telomer probu olacak
şekilde hibridizasyon çözeltisi hazırlandı. Prehibridizasyon işleminden sonra
prehibridizasyon çözeltisi ortamdan uzaklaştırıldıktan sonra membran hazırlanan
hibridizasyon çözeltisi içerisinde DIG işaretli telomerik tekrar dizinlere spesifik prob
ile 3 saat 42˚C’de düşük devirde hafif çalkalanarak “Thermoshaker”da
hibridizasyona bırakıldılar. 200 cm2 membran için yaklaşık 6.5 ml hibridizasyon
çözeltisi gerekmektedir. Probla nitroselüloz membran üzerindeki DNA tek zincir
fragmanlarından tamamlayıcısı olan arasında, hidrojen bağları oluşumu ile çift
zincirli DNA parçası meydana geldi.
Hibridizasyon süresi sonunda hibridizasyon çözeltisi uzaklaştırıldı ve önce 2
defa 5’er dakika hibridizasyon sonrası 1. yıkama çözeltisi içerisinde oda sıcaklığında
yıkandı. Daha sonra hibridizasyon sonrası 2. yıkama çözeltisi içerisinde 2 defa 15’er

68
dakika önceden 50˚C’ye ayarlanmış “Thermoshaker”da düşük devirde hafif
çalkalanarak yıkandı. Bu yıkama işlemlerinin sonunda hibridize olmayan fakat
membranın yüzeyine tutunan problar ortamdan uzaklaştırıldılar.
2.4.11. Hibridize Olmuş Telomerik DNA’nın Belirlenmesi
Membran yeni temiz bir kaba alındı. Oda sıcaklığında 100 ml yıkama
tamponunda 5 dakika yıkandı. Yıkama tamponu uzaklaştırıldı ve membran 30 dakika
oda sıcaklığında hafif çalkalanarak taze hazırlanmış bloklama çözeltisi içerisinde
inkübe edildi. Bloklama çözeltisi uzaklaştırıldı. Daha sonra 75mU/ml olacak şekilde
(1/10,000 oranında) bloklama çözeltisi içerisinde sulandırılmış Anti-DIG-Alkalen
fosfataz (AP) çözeltisi hazırlandı. Membran bu çözelti ile probun yapısındaki
digoksigenin ile anti-DIG arasında kompleks oluşumu için oda sıcaklığında 30
dakika hafif çalkalanarak inkübe edildi. İnkübasyon süresi sonunda Anti-DIG-AP
antikor çözeltisi uzaklaştırılarak membran bağlanmayan fakat membran yüzeyine
tutunmuş olan antikorları uzaklaştırmak için 2 defa 15’er dakika 200 ml yıkama
tamponu (kitte hazır) içerisinde yıkandı. Yıkama tamponu uzaklaştırıldıktan sonra
membran 2-5 dakika oda sıcaklığında 100 ml belirleme tamponu içerisinde inkübe
edildi. Belirleme tamponu (kitte hazır) uzaklaştırıldıktan sonra fazla sıvının
uzaklaştırılması amacıyla membran absorban bir kâğıt üzerine kondu. Daha sonra
membran DNA bağlı yüzü üste gelecek şekilde bir naylon film üzerine yerleştirildi.
Bu işlem sırasında membran yüzeyi kurumamalıdır. Membranın üzerine
alkalenfosfataz enziminin substratı olan CDP-star maddesini içeren çözeltiden 40
damla damlatıldı ve membranın bütün yüzeyine homojen dağılması sağlandı.

69
Membranın üzeri arada hava kalmayacak ve düzgün bir yüzey oluşturacak şekilde
tekrar naylon film ile kapatıldı. Daha sonra bir kaset içerisinde karanlık bir odada
ışık geçirmeyecek şekilde yaklaşık 30 dakika radyografi filmi ile etkileştirildi. Daha
sonra film banyo edilerek telomerik fragmanlara ait bant görüntüleri elde edildi.
Bu yöntem kemilüminesans adını alır ve esası alkalen fosfatazın
kemilüminesan özellik gösteren substratı ile reaksiyona girmesi sonucu substratını
ışık sinyalleri (kemilüminesans) oluşturacak ürünlere dönüştürmesi esasına dayanır.
Oluşan bu ışık sinyallerinin radyografi filmini bozundurması sonucu membrandaki
bant görüntüsü filme yansır. Daha sonra filmdeki telomerik fragmanlere ait bölgeler
moleküler ağırlık standartı ile karşılaştırılarak dansitometrik olarak tayin edildi (77).
Bu amaçla aşağıdaki formül kullanıldı:
Ortalama Telomer uzunluğu=∑(ODi)/ ∑(ODi/Li)
ODi: Kemilüminesan sinyal (optik dansite)
Li: i pozisyonunda telomer fragmanının boyu (TeloTAGG Telomer Lenght Assay
kiti, Roche, Almanya)

70
2.4.12. Hibridizayonda Kullanılan Çözeltiler
Tablo 2.4.: Hibridizasyonda kullanılan çözeltiler
Tampon İçeriği
1. yıkama çözeltisi 2xSSC
%0.1 SDS
2. yıkama çözeltisi 0.2xSSC
%0.1 SDS
Bloklama tamponu Kit içerisinde hazır verilen 10x bloklama
tamponundan maleik asit ile 1:10 oranında
seyreltilerek hazırlanır.
Anti-DIG-AP tamponu Kit içerisinde verilen anti-DIG-AP çözeltisi daha
önceden hazırlanmış olan bloklama tamponu ile
1:10 oranında seyreltilir.
2.4.13. DNA’nın Jelden Geri Alınması
TRF analizlerimiz sırasında alınan ilk membran görüntüsünde görüntüde kısa
telomerik fargmanların beklendiği membran bölgesinde daha az yoğunlukta “smear”
(sürüntü) olmuştur. Bunun nedeninin restriksiyon kesiminin yetersiz kalarak
telomerik tekrarların yüksek moleküler ağırlıklı DNA içerisinde taşınması
olabileceği ihtimalini test etmek için, yüksek moleküler ağırlıklı DNA bandı
kesilerek jelden izole edilmiş ve restriksiyon enzimleri ile yeniden kesilerek
incelenmiştir.

71
DNA’nın jelden geri kazanılmasına yönelik oldukça farklı yöntemler
geliştirilmiştir. Bu yöntemlerle istenilen boyuttaki DNA fragmanlarını saf olarak elde
etmek veya DNA’yı herhangi bir aşamada saflaştırmak olasıdır. Jelden kestiğimiz
yüksek ağırlıklı DNA bandı bu yöntemlerden biri olan ticari olarak üretilen kolonlar
(BioBasic® SK131 Uniq-10 Spin Column) kullanılarak jelden izole edilmiştir. Bu
amaçla önce yüksek moleküler ağırlıklı DNA fragmanının olduğu jel bölgesi bistüri
ile kesilip çıkartılmıştır. 1.5 ml’lik bir tüp içerisinde kitle birlikte verilen tampon
çözeltisi içerisinde 50-60˚C’de 10 dakika inkübe edilmiştir. Agaroz tamamen
çözündükten sonra elde edilen çözelti kolonlara aktarılmış ve 8000 g’de 1 dakika
santrifüj edilmiştir. Böylece DNA’nın kolona bağlanması sağlanmıştır. Daha sonra
ayrıştırma tamponu kullanılarak DNA kolondan geri alınmıştır.
2.5. Telomerik Tekrar Amplifikasyon Protokolü (Telomeric Repeat
Amplification Protocol, TRAP)
Bu yöntem 1994’te Kim ve arkadaşları tarafından tanımlanmıştır (97). Hücre
ve dokulardan elde edilen homojenat bir PCR tüpünde, özel hazırlanmış primerler ile
muamele edilir. Eğer homojenat telomeraz içeriyorsa ortama konan primerleri
uzatacaktır. Daha sonra bu uzama ürünleri (extension product) “reverse” primer
kullanılarak PCR sisteminde amplifiye edilir. Poliakrilamid jelde yürütülür.
Telomeraz enzimi varlığında elektroforez sonucu tipik olarak 6 bç (baz çifti) içeren
tekrar bantları oluşur. Her zaman, çalışılan örneklerin ısı ile veya RNaz ile denatüre
edilmiş olanları da paralel olarak çalışılır. Eğer görünen bantlar telomerazın
telomerik DNA sentezlemek için kendi RNA kalıbını kullanması sonucu (telomeraz

72
aktivitesi) ortaya çıkıyorsa, RNA’ları denatüre edilmiş örneklerin bu tekrar bantları
oluşturmaması beklenir.
TRAP yöntemi ile telomeraz aktivitesinin ölçülmesine yönelik bu deneylerde
telomeraz enziminin içerdiği RNA komponentinin DNA primerlerin uzatılmasında
kalıp RNA olarak kullanılması ve RNA’nın çok çabuk denatüre olabilmesi nedeniyle
sterilizasyona son derece dikkat edilmelidir.
2.5.1. Ekstraktların Hazırlanması
Telomeraz aktivitesine yönelik bu seri deneyler için ayrılan, sıvı azotta
dondurulup daha sonra -80˚C’de bekletilmiş olan dokular çıkarılarak buz üzerinde
çözünmeleri sağlanmıştır. Daha sonra dokular steril bir petri kabı içerisinde steril bir
bistüri yardımıyla küçük parçacıklara ayrılmış daha sonra bu parçacıklar içerisinde
40-100 mg dokuya 200 µl 1xCHAPS liziz tamponu olacak şekilde bu tamponu içeren
tüplere (eppendorf) aktarılmıştır. Daha sonra parçalanma ve homojenizasyonu
sağlamak amacıyla içerisine steril tungsten boncuklar ilave edilmiş ve homojenize
edilmiştir. Bu aşamada ısı oluşunu önlemek son derece önemlidir. Bu amaçla
homojenizatörün hazneleri önceden -20˚C’de tutulmuş, homojenizasyon işleminin
hemen öncesinde çıkarılmıştır. Homojenizasyon işlemi ısı oluşumunu önlemek
amacıyla belirli aralıklarla gerçekleştirilmiştir.

73
Homojenizasyon işlemi sonunda suspansiyon haline gelmiş olan dokular buz
üzerinde 30 dakika inkübe edildiler. Daha sonra 12000 g’de 4˚C’de 20 dakika
santrifüj edildiler. Santrifüj işlemi sonucunda oluşan süpernatantın 160 µl’si yeni
steril bir tüpe alındı ve Biçinkoninik asid (BCA) yöntemi ile protein miktar tayini
yapılmak üzere ayrıldı. Kalan ekstrakt ise tekrar -80˚C’ye kaldırıldı.
2.5.2. Total Protein Miktar Tayini
Elde edilen ekstrakttaki toplam protein miktarını belirleyebilmek için total
protein miktar tayini yapılmıştır. Bu amaçla Biçinkoninik Asit (BCA) yöntemi
uygulanmıştır (200). Biçinkoninik asit (BCA) analizinin prensibi Lowry protein
tayinine benzemektedir (113). Her iki yöntemin temeli alkali ortamda Cu+2-protein
kompleksinin oluşumuna ve Cu+2’nin Cu+1’e indirgenmesine dayanmaktadır. Örnekte
var olan protein miktarı indirgenmiş Cu+1 miktarı ile doğru orantılıdır. Protein
yapısında yer alan sistein, sistin, triptofan, tirozin ve peptid bağlarının Cu+2’yi Cu+1’e
indirgediği gösterilmiştir. BCA, alkali ortamda Cu+1 ile mor-mavi kompleks
oluşturur, böylelikle alkali Cu+2’nin proteinler tarafından indirgenmesinin
izlenmesini mümkün kılmaktadır (172).
BCA protein analizi, Lowry (113) protein yönteminden daha duyarlıdır. Ayrıca,
Bradford (22) protein yöntemine göre daha stabil değerler elde edilir. BCA protein
yöntemi diğer protein yöntemlerine göre pek çok avantaja sahiptir;

74
• Kullanımı kolaydır,
• Renkli kompleks stabildir,
• Deterjanlardan daha az etkilenir,
• Geniş aralıkta protein konsantrasyonlarına uygulanabilir.
Çalışma reaktifinin hazırlanışı:
Reaktif A (Biçinkoninik asit çözeltisi) ve Reaktif B (%4’lük Cu(II)sülfat pentahidrat
çözeltisi) 50:1 oranında karıştırılır.
Standartların hazırlanışı:
Protein standartı olarak Bovine Serum Albumin (BSA) ve lizis tamponu kullanılarak
aşağıdaki şekilde dilüsyonlar hazırlanmıştır:
Tablo 2.5.:Total protein miktar tayininde kullanılan dilüsyonlar
Küvet No BSA final miktarı
(µg)
BSA Std. Hacmi
(0.1 mg/ml stoktan)
BSA Std Hacmi (1.0
mg/ml stoktan)
Lizis Tamponu
Hacmi
1 0 0 0 50 µl
2 1 10µl 0 40 µl
3 2 20 µl 0 30 µl
4 5 50 µl 0 0
5 5 0 5 µl 45 µl
6 10 0 10 µl 40 µl
7 15 0 15 µl 35 µl
8 20 0 20 µl 30 µl

75
Örneklerin hazırlanışı:
10 µl örneğe 40 µl lizis tamponu eklenerek dilüsyonlar hazırlanmıştır. Daha
sonra hazırlanan 50 µl final hacmine sahip kör, standart ve örnek dilüzyonlarının
üzerine 1’er ml çalışma reaktifi eklenir. 37˚C’de 30 dakika bekletildikten sonra
spektrofotometrede 562 nm’de örnek ve standartların absorbans değerleri köre karşı
okundu.
Tablo 2.6.’da standartların belirli konsantrasyonlarına karşı ölçülen absorbans
değerleri görülmektedir. Daha sonra bu değerlere göre standart grafiği çizilmiştir
(Şekil 2.2.). Absorbansı ölçülen örneklerin protein miktarları, örnek için ölçülen
absorbans değerine karşılık gelen konsantrasyon değeri standart grafik denkleminden
hesaplanarak belirlenmiştir.
Tablo 2.6.:Standartların belirli konsantrasyonlarına karşı ölçülen absorbans değerleri
Standart Konsantrasyon (µg/ml) Absorbans (nm)
1 0 0
2 1 0.032
3 2 0.079
4 ve 5 5 0.126
6 10 0.177
7 15 0.200
8 20 0.262

76
protein miktar tayini y = 0,0119x + 0,0353R2 = 0,9312
00,050,1
0,150,2
0,250,3
0 5 10 15 20 25 konsantrasyon
abso
rban
s (5
62 n
m'd
e)
Şekil 2.2.: Total protein miktarı standart grafiği örneği
2.5.3. Kontroller
2.5.3.1. Negatif kontroller
Telomeraz ısıya duyarlı bir enzimdir. Bu nedenle negatif kontrol olarak her bir
örnek için ısıya maruz bırakılarak telomeraz enzimi inaktive edilmiş bir seri
hazırlandı. Bunun için her bir örnekten 10 µl alınarak 10 dakika 85˚C’de inkübe
edildi.
2.5.3.2. Telomeraz-pozitif hücre ekstraktı ile kontrol
Kitin içerisindeki hücre pelletleri (106 hücre) ile 200 µl 1xCHAPS lizis
tamponu kullanılarak telomeraz pozitif hücre ekstraktları hazırlandı. Bu stok çözelti
80˚C’de saklanıp her deney öncesi çıkarılıp 1:20 1xCHAPS lizis tamponu ile
seyreltilerek her bir reaksiyon için 2 µl kullanıldı (2 µl =500 hücre).

77
2.5.3.3. PCR Amlifikasyon kontrolü
Kullandığımız kit her bir reaksiyon tüpündeki PCR inhibisyonun gözlenmesi
amacıyla internal standart içermektedir. Bazı örnekler Taq polimeraz inhibitörleri
içerebilirler ve bu nedenle yanlış-negatif sonuç verebilirler. Bunu diğer
problemlerden ayırt etmek için kitteki TRAPeze primer karışımı TS primeri ile
birlikte internal kontrol oligonükleotidleri olan K1 ve TSK1’i içerir. Bunların normal
PCR koşullarında (Taq polimeraz inhibitörü yokluğunda) her bir sırada 36 bç’lik
bant (S-IC) oluşturması beklenir. S-IC bandı aynı zamanda amlifikasyonun
etkinliğini de göstermektedir.
2.5.3.4. Primer-dimer /PCR kontaminasyon kontrolü
Kalıp reaksiyon ortamında bulunan primerler kalıp DNA olmaksızın da
kalıptan bağımsız PCR ürünleri oluşturabilirler. Bu tür PCR ürünlerine primer-dimer
PCR artifaktları denir. Bu tür artifaktların neden olduğu kontaminasyonların test
edilmesi amacıyla her bir deney için hücre/doku ekstraktı yerine 2 µl 1xCHAPS lizis
tamponu kullanılarak reaksiyon kuruldu. Deneyin en uygun koşullarda çalıştığı
durumlarda primer-dimer/PCR kontaminasyon hatlarında 36 bç’lik internal kontrol
bandından başka bir ürün bulunmamasına dikkat edildi.

78
2.5.3.5. Telomeraz kantitasyon kontrol kalıbı-TSR8
TSR8 TS primerle aynı diziye sahip 8 telomerik tekrarla AG(GGTTAG)7
uzatılmış bir oligonükleotidtir. Bu kontrol telomerik tekrarların telomeraz ile
uzatıldığı verilen bir ekstraktta TS primerlerin miktarını tahmin etmek için standart
bir kontroldür. Örnek ekstraktı yerine 1-2 µl TSR8 kontrol kalıbı kullanılarak
reaksiyon kuruldu.
2.5.4. TRAP Analizinin PCR Basamağı
Her bir reaksiyon tüpü için aşağıdaki formülasyona göre reaksiyon karışımı
hazırlandı:
10xTRAP Reaksiyon Tamponu 5.0 µl
50x dNTP Karışımı 1.0 µl
TS primer 1.0 µl
TRAP primer karışımı 1.0 µl
Taq polimeraz (5 ünite/µl) 0.4 µl
Su 39.6 µl
Toplam hacim 48.0 µl

79
Bu karışımın 48 µl’si steril bir PCR tüpüne kondu ve üzerine 2 µl örnek ekstraktı,
ısı ile inaktive edilmiş örnek ekstraktı veya kontroller ilave edildi:
• Örnek ekstraktları: Her bir örnek tüpüne 2 µl ekstrakt kondu.
• Isı ile inaktive edilmiş örnek ekstraktı: Her bir örnekten 4-5 µl ekstrakt
85˚C’de 10 dakika inkübe edildi. Bu örnekler için ayrılmış her bir tüpe 2’şer
µl ısı ile inaktive edilmiş ekstrakt kondu.
• Telomeraz-pozitif kontroller: Pozitif kontrol hücre ekstraktlarından 250
hücre/µl konsantrasyonda olacak şekilde 2 µl kondu.
• Primer-dimer/PCR kontaminasyon kontrolü: Bu amaçla ayrılmış tüplere
1xCHAPS Lizis tamponu kondu.
• Kantitasyon Kontrolü: Kontrol tüplerinden birine 1 µl TSR8 (kontrol kalıbı)
ve 1 µl su diğerine 2 µl TSR8 kondu.
PCR Amplifikasyonu
Tüpler termal bloğuna yerleştirilip ve örneklerdeki olası telomeraz enzimi
etkisi ile primerlerin uzatılması amacıyla 30˚C’de 30 dakika inkübe edildi.
Daha sonra denatürasyon basamağı 94˚C’de 30 saniye, primerlerin bağlanması
basamağı 59˚C’de 30 saniye ve uzatma basamağı 72˚C’de 1 dakika olacak şekilde 32
döngü PCR işlemi uygulandı. Telomerik tekrarlara (GGTTAG) komplementer
oligonükleotid primerler kullanılan kitin üretici firması (TRAPeze, Chemicon, ABD)
tarafından sağlanmıştır.

80
2.5.5. Oluşan PCR Ürünlerinin Poliakrilamit Jelde Yürütülmesi
Poliakrilamid jel elektroforezi (PAGE)
Akrilamitin polimerizasyonu ile hazırlanan poliakrilamit jeller küçük ya da orta
boydaki nükleik asit ve proteinler için yüksek ayrıştırma gücüne sahiptir.
Poliakrilamit jeller akrilamit ve çapraz bağlayıcı N,N’-metilen-bis-akrilamidin
serbest radikal polimerizasyonu ile hazırlanır. Kimyasal polimerizasyon bir başlatıcı
(amonyum persülfat) ve katalizör (Tetrametilen etilen diamin, TEMED) tarafından
kontrol edilir.
Tablo 2.7: % 12,5’luk Poliakrilamit jelin hazırlanması (50 ml jel için)
Akrilamit/bisakrilamit (%40’lık) 15 ml
5xTBE 5 ml
%10 amonyum persülfat 250 µl
TEMED 50 µl
Distile su 30 ml
Herbir reaksiyon tüpüne 5 µl bromfenol mavisi ve ksilen siyanol içeren
yükleme boyası ilave edildi ve dikey elektroforez sistemine önceden dökülüp tarakla
birlikte dondurulmuş olan poliakrilamit jeldeki kuyucuklara yüklendi. 0.5xTBE’de
400V’da ksilen siyanol boyası 10-12 cm’lik jelin %70-75’ini katedene kadar
yaklaşık 1,5 saat yürütüldü. Elektroforez sonunda jel etidyum bromür ile yaklaşık 30
dakika boyandı ve daha sonra bağlanmamış boyanın uzaklaştırılması amacıyla

81
deiyonize su ile yaklaşık 20-30 dakika yıkanarak (destaining) UV’de görüntülendi.
S-IC internal kontrol bandı 36 bç ve en küçük PCR ürünü 50 bç olmalıdır.
Geçerli bir TRAP deneyi deney sonunda jel görüntülendiğinde şu koşulları
karşılamalıdır:
• Primer-dimer/PCR kontaminasyon kontrolü: 36 bç’lik internal kontrol bantı
dışında herhangi başka bir ürün görünmemelidir.
• Telomeraz-pozitif kontroller: 36 bç’lik internal kontrol bandı ve 50 bç’den
başlamak üzere 6 bazlık artışlar gösteren ardışık bantlar görünmelidir.
• Isı uygulanarak inaktive edilmiş örnekler: 36 bç’lik internal kontrol bantı
dışında herhangi başka bir ürün görünmemelidir.
Eğer yukarıdaki koşullar karşılanıyorsa, örnek eğer telomeraz-pozitif ise
telomeraz-pozitif kontroller ile aynı bant görüntülerine sahip olmalıdır (50 bç’den
başlamak üzere 6 bazlık artışlar gösteren ardışık bantlar). Eğer örnek telomeraz-
negatif ise sadece 36 bç’lik internal kontrol bandı görünmelidir.
2.6. Histolojik Analizler
Morfometrik inceleme için ayrılan damar segmentleri %4’lük formalin ile fikse
edilmiştir. Daha sonra 2 mm kalınlıkta olacak şekilde enine en az 4 örnek elde
edilerek doku takip işlemine alınmıştır. Doku takip cihazında (Intelsint, RV1, İtalya)
sırasıyla; artan derecedeki alkoller, ksilol ve parafin istasyonlarından geçirilerek

82
dokular parafine gömülmüştür. Histokimyasal işlemlerin detayı aşağıda
açıklanmıştır:
1. %80 alkol 2 saat
2. %95 alkol I 2 saat
3. %95 alkol II 1 saat
4. Absolu alkol I 1 saat
5. Absolu alkol II 2 saat
6. Ksilol I 20 dk
7. Ksilol II 20 dk
8. Ksilol III 20 dk
9. Parafin I 45 dk
10. Parafin II 1 saat
11. Parafin III 1 saat
Parafin bloklardan elde edilen 4 mikron kalınlığındaki kesitler bir tanesi klasik
diğer 4 tanesi ise polilisinli olmak üzere 5 lama alındı. Klasik preparatlar
Hematoksilen Eozin (HE), lizinli preparatlar ise düz kas aktini (DKA) (138),
prolifere olan hücre nükleer antijeni (proliferating cell nuclear antigen = PCNA)
(170) ve Ki67 (7) antikorları ile immunohistokimyasal boyama için ayrıldı.

83
2.6.1. Morfometrik İncelemeler
Klasik lama alınan preparatlar morfometrik incelemeler yapmak amacıyla HE
ile boyandı (101).
HE ile boyama işlemlerinin detayı aşağıda açıklanmıştır:
Prepratlar 60 ºC ayarlı etüvde 3 saat bekletildi.
Deparafinizasyon Ksilol I 10 dk
Deparafinizasyon Ksilol II 15 dk
Absolü alkol I 5 dk
Absolü alkol II 5 dk
%95 alkol 5 dk
%80 alkol 5 dk
Distile su 5 dk
Hematoksilen 4 dk
Çeşme suyu 2 dk
Asit-alkol 2 kez batırıp-çıkarma
Çesme suyu 2 kez batırıp-çıkarma
Amoyaklı su 2 kez batırıp-çıkarma
Çeşme suyu 2 kez batırıp-çıkarma
Distile su 3 dk
Eozin 1 dk
%95 alkol 2 dk
Absolü alkol 2 dk
Absolü alkol II 2 dk
Kurutma
Şeffaflandırma ksilol I 5 dk
Şeffaflandırma ksilol II 5 dk
Lam üzerine entellan damlatma
Lamel ile kaplama

84
Her bir örnek için rastlantısal olarak seçilen 3 farklı damar kesitine ait x4
büyütmedeki görüntüler yüksek çözünürlüklü video kamera (VKC220E, Hitachi,
Tokyo, Japonya) donanımı olan bir ışık mikroskobundan (Labophot-2, Nikon,
Tokyo, Japonya) IBM uyumlu bilgisayara aktarılmış ve bir görüntü analiz programı
(BS 200 Docu Version 2.0, BAB Imaging Systems, Ankara) kullanılarak lümen,
intima ve medya alanları monitör üzerinden çizilerek ölçülmüştür.
2.6.2. İmmünohistokimyasal İncelemeler
İmmunohistokimyasal uygulamada primer antikor olarak DKA (141) için 1A4
klonu (M0851, Dako; Glostrup, Danimarka) 1/200sulandırılarak, PCNA (25) için
PC10 klonu (M0879, Dako; Glostrup, Danimarka) 1/400 sulandırılarak ve Ki67 (44)
için SP6 klonu (RM-9106, Labvision; Fremont, Kanada) 1/200 sulandırılarak
kullanıldı. Ayrıca Ki67 için “clone” farkını ortaya koyabilmek amacıyla MIB-1
(N1633, Dako; Glostrup, Danimarka) için de ayrı bir boyama uygulandı. Universal
kit olarak işaretli Avidin-biyotin-peroksidaz metodu (LSAB) (Dako; Glostrup,
Danimarka), renklendirici olarak ise DAB (Zymed; South San Francisco, ABD)
kullanıldı. İmmunohistokimyasal boyamanın detayı aşağıda açıklanmıştır:
1. Parafin bloklardan distile suya kadar getirilen kesitler PBS’de (Phosphate Buffered
Saline Solution) (1 saat)
2. %3 H²O² (5 dk)
3. Çeşme suyunda yıkama
4. PBS (5 dk)
5. Blokan antikor (10 dk)
6. PBS (5 dk)
7. 10mMol sitrat solution (pH 8.5) içerisinde buhar tenceresinde 20 dk kaynatma ve
20 dk soğutma

85
8. PBS (5 dk)
9. Primer antikor (2 saat)
10. PBS (5 dk)
11. Biotinli sekonder antikor (45 dk)
12. PBS (5 dk)
13. Streptavidin/peoksidaz kompleksi (45 dk)
14. PBS (5 dk)
15. Diaminobenzidin tetrahidroklorür (DAB) ile 5 dk renklendirme
16. Çeşme suyu (5 dk)
17. Meyer Hematoksilen 2 dk zıt boyama
18. Çeşme suyu (2 dk)
19. %80 alkol (2 dk)
20. %95 alkol (2 dk)
21. Absolü alkol (2 dk)
22. Kurutma
23. Entellan ile lamelleme
İmmunohistokimyasal boyama sonucunda DKA için sitoplazmik, PCNA ve
Ki67 için nükleer kahverenkli boyanmalar pozitif kabul edilerek değerlendirmeye
gidilmiştir.
İmmüno boyama için kullanılan yöntem strept(avidin)- biotin peroksidaz
yöntemine dayanır. Bu yöntemde türe özgü primer antikor örnekle inkübe edildikten
sonra primer antikora bağlanması için biyotinlenmiş sekonder antikor kullanılır.
Daha sonra ortama eklenen “Horse Radish Peroxidase” (HRP) ile işaretlenmiş
streptavidin, biyotinli sekonder anikora bağlanır. Oluşan antijen/antikor/enzim
kompleksine son aşamada substrat/kromojen maddesi diaminobenzidin (DAB) ilave
edilerek peroksidaz enzimi aracılığı ile kahverengi renk reaksiyonu meydana
getirilir. Avidin yumurta akından elde edilen ve biotine karşı aşırı ilgisi olan bir

86
glikoproteindir. Avidinin yerine kullanılabilen bir molekül olan streptavidin ise
Streptococcus avidini’den elde edilir. Avidinden farklı olarak yüksüz bir moleküldür
ve özgün olmayan elektrostatik bağlanmalar daha az oluşur. Biotin ise antikor gibi
çeşitli biyolojik moleküllere konjüge olabilen düşük molekül ağırlıklı bir vitamindir.
Bu yöntem immünohistokimyasal yöntemler içerisinde en yaygın olarak kullanılan
yöntem olarak bilinmektedir (77).
2.7. Kan Örneklerinde Yapılan Ölçümler
II. Bölümde deney süresince bütiyonin sülfoksimin uygulanması sonucu kanda
indirgenmiş glutatyon/ yükseltgenmiş glutatyon oranının (GSH/GSSG), total
glutatyonun ve glutatyon peroksidazın enzim düzeylerindeki değişikliği
belirleyebilmek amacıyla tavşanların kulak arterinden 0. gün, 8. gün ve 14. günde
olmak üzere EDTA’lı tüplere alınan kan örneklerine, ölçümlerde kullanılan kitlerin
tanımladığı şekilde muamele edilmiş ve kan örnekleri ölçümlerin yapılacağı güne
dek -80˚C’de saklanmıştır.
2.7.1. Plazma Total Glutatyon (GSH) Miktarının Belirlenmesi
Alınan kan örneklerindeki total GSH miktarının belirlenmesi amacıyla
Bioxytech GSH-420 (Oxis, ABD) kiti kullanılmıştır. Bu yönteme göre örnek ilk önce
indirgen bir ajan olan tris(2-karboksietil)fosfin (TCEP) ilave edilerek örnekte
bulunan okside glutatyonun (GSSG) redükte forma indirgenmesi sağlanır. Daha
sonra kromojen, 4-kloro-1-metil-7-triflorometilkinolinyum metilsulfat ilave edilerek

87
tiyoeterler oluşturulur. Ortam bazik hale getirilerek kromoforik tiyon oluşturulur ve
420 nm’de absorbans okunur (30).
2.7.2. Plazma Glutatyon /Okside Glutatyon (GSH/GSSG) Oranının Tayini
Alınan kan örneklerindeki GSH/GSSG oranının tayini Bioxytech GSH/GSSG-
412 (Oxis, ABD) kiti kullanılarak yapılmıştır.Memeli hücreleri fazla oksidatif strese
maruz kaldıklarında GSH/GSSG oranı GSSG’nın birikimine bağlı olarak azalacaktır.
GSSG düzeyinin veya GSH/GSSG oranının ölçülmesi oksidatif stres için yararlı bir
indikatördür. Antioksidanlar ile girişim yapıldığında antioksidanların etkinliğinin
gözlenmesine olanak verir (71). Bu yöntemde tiyol süpürücü bir reaktif olan 1-metil-
2-vinilpiridinyum triflorometansülfonat (M2PV) kullanılarak GSSG ölçümü
yapılacak örnek hemen M2PV ile etkileştirilerek ortamdaki GSH’un uzaklaştırılması
sağlanır. Daha sonra ortama glutatyon redüktaz, NADPH ve kromojen olarak 5, 5’-
ditiyobis-(2-nitrobenzoik asit) (DTNB) ilave edilerek GSSG’un GSH’a
dönüştürülmesi sağlanır ve bu dönüşüm sırasında oluşan renkteki değişim 412 nm’de
kaydedilir. Yöntemde yer alan tepkime döngüsü aşağıdaki gibidir.

88
2.7.3. Plazma Glutatyon Peroksidaz (GPx) Enzim Aktivitesinin Tayini
Alınan kan örneklerindeki glutatyon peroksidaz enzim aktivitesinin tayini
Bioxytech GPx-340 (Oxis, ABD) kiti kullanılarak yapılmıştır. Glutatyon peroksidaz
enzimi vücutta hidrojen peroksidin suya ve reaktif radikaller oluşturabilen organik
peroksitlerin de stabil organik alkollere indirgenmesini katalizler. Bu raksiyon için
indirgeyici ajan olarak glutatyonu (GSH) kullanır. Yöntemin esası GPx tarafından
organik bir peroksidin indirgenmesi sırasında ortaya çıkan okside glutatyonun
(GSSG) glutatyon redüktaz enzimi ile tekrar redükte formu olan GSH’a
indirgenmesine dayanan dolaylı bir ölçüme dayanır (140). Bu indirgenme reaksiyonu
sonucunda NADPH da NADP+’ye dönüşür. NADPH’ın NADP+’ya dönüşümüne
340nm’teki absorbansta azalma eşlik eder. Örnek; glutatyon, glutatyon redüktaz ve
NADPH bulunan ortama ilave edilir. Substrat (tert-bütil hidroperoksit) eklendiğinde
reaksiyon başlar ve 340 nm’deki absorbans 3 dk. boyunca kaydedilir. Yönteme
ilişkin reaksiyon basamakları aşağıdaki gibidir.
2.8. Aygıtlar
Birinci seri deneylerimizde kullandığımız aygıtlar Tablo 2.8 ‘de verilmiştir.

89
Tablo 2.8.: Birinci seri deneylerde kullanılan aygıtlar
Aygıt Marka
Otoklav Hirayama Hiclave HV-50L, Japonya
Jel karıştırıcı IKA K5501D, ABD
Isıtıcılı karıştırıcı kabin Thermoshake, Gerhardt, Almanya
Homojenizatör Qiagen TissuLyser®- Retech, Haan, Almanya
Vorteks IKA, MS1 Minishaker, Amerika
Manyetik karıştırıcı Elektro-mag, M22, Türkiye
Tartım cihazı Sartorius BP121S, Almanya
Santrifüj cihazı Hettich EBA12, Almanya
Soğutuculu santrifüj cihazı Hettich Mikro 200R, Almanya
Thermal cycler Techne TC-512, Cambridge, İngiltere
pHmetre Inolab pHlevel1, Wheilheim, Almanya
Derin dondurucu Nuaire -86˚C Ultralow freezer, Revco Technologies,
Asheville,NC, ABD
UVcrosslinker cihazı Vilber Lourmat, Cedex, Fransa
UV görüntüleyici Vilber Lourmat, Cedex, Fransa
Mikrodalga fırın Ariston MW211W, İtalya
Elektroforez güç kaynağı 1 EC-250-90, Thermo, New York, ABD
Elektroforez güç kaynağı 2 EC-1000-90 Thermo, New York, ABD
Yatay elektroforez sistemi 1 Thermo EC Maxicell Primo EC340,New York,ABD
Yatay elektroforez sistemi 2 Thermo EC Midicell Primo EC330,New York,ABD
Dikey elektroforez sistemi Atto AE-6200 SlabEP Chamber ve AE-6210 Gel cast,
Tokyo, Japonya
UV spektrofotometre Shimadzu, UV-1208, Japonya
UV spektrofotometre modülü Perkin Elmer Victor3, ABD

90
2.9. Verilerin Değerlendirilmesi
Veriler ortalama±ortalamanın standart hatası şeklinde verilmiştir; n damarların
alındığı tavşan sayısını göstermektedir. Çoklu karşılaştırmalar varyans analiz modülü
(one way analysis of variance, ANOVA) ile gerçekleştirilmiştir. Gruplar arasındaki
karşılaştırmalar Student t testi ile analiz edilmiştir. P değerleri 0.05’ten küçük
bulunduğunda fark istatistiksel olarak anlamlı kabul edilmiştir.

91
3. BULGULAR
3.1. Bölüm 1
Bu bölümde tavşan iliyak arterine anjiyoplasti uygulaması sonrası farklı lezyon
sürelerinde (0. gün, 2. gün, 7. gün ve 14. gün) vasküler düz kas hücrelerinde telomer
devamlılığı ve telomeraz aktivitesinin incelenmesine ilişkin deneyler yapılmıştır.
Elde edilen bulgular aşağıdaki gibidir;
3.1.1. Morfometrik İncelemeler
3.1.1.1. İntima alanı
Tavşan ilyak arterine balon anjiyoplasti uygulamasına bağlı olarak intimal
kalınlaşma gelişimini saptamak üzere, uygulamanın 0., 2., 7. ve 14. günü sonunda
intimal alan ölçümleri üzerinde istatistiksel analizler yapılmıştır. Analiz sonuçları
balon anjiyoplasti uygulamasının, uygulama sonrası, 0., 2., ve 7. günlerde, intimal
kalınlaşma üzerinde istatistiksel olarak anlamlı bir etkisinin olmadığını ortaya
koymaktadır (Şekil 3.1., sayısal veriler sunulmamıştır). Bunun yanı sıra, analizler
balon uygulamasının 14. gün sonunda intimal kalınlaşmayı anlamlı bir şekilde
artırdığını göstermektedir (mm2: 0.75±0.08, 0.25±0.06, sırasıyla balon anjiyoplasti
uygulanmış ve uygulanmamış ilyak arterler, Student t testi, P<0.01, n=3) (Şekil 3.2.).

92
A B
Şekil 3.1.: Balon anjiyoplasti sonrası 0. günde hasar uygulanan (A) ve uygulanmayan (B) tavşan ilyak arterlerine örnek kesitler (x 10 büyütmede alınmıştır).
A) B)
Şekil 3.2.: Balon anjiyoplasti sırasında hasar uygulanan (A) ve uygulanmayan (B) tavşan ilyak arterlerinin anjiyoplastiyi izleyen 14. günde alınan örnek kesitleri (x10 büyütmede alınmıştır).

93
3.1.1.2. Medya Alanı
Tavşan ilyak arterine balon anjiyoplasti uygulamasına bağlı olarak,
uygulamanın 0., 2., 7. ve 14. günü sonunda medya alanı üzerindeki etkilerini
saptamak üzere istatistiksel analizler yapılmıştır. Analizi sonuçları balon anjiyoplasti
uygulamasının 0., 2., 7. ve 14. günlerinde medya alanı üzerinde istatistiksel olarak
anlamlı bir etkisinin olmadığını ortaya koymaktadır (Sayısal veriler sunulmamıştır).
3.1.1.3. Lümen Alanı
Tavşan ilyak arterine balon anjiyoplasti uygulamasına bağlı olarak,
uygulamanın 0., 2., 7. ve 14. günü sonunda lümen üzerindeki etkilerini saptamak
üzere istatistiksel analizler yapılmıştır. Analiz sonuçları balon anjiplasti
uygulamasının 0., 2., 7. günlerinde balon anjiyoplasti uygulamasının lümen alanı
üzerine herhangi bir etkisinin olmadığını ortaya koymuştur (sayısal veriler
sunulmamıştır). Buna karşılık balon anjiyoplasti uygulaması sonrası 14. gününde
lümen alanının anlamlı bir şekilde azaldığı belirlenmiştir (mm2: 0.52±0.07,
0.195±0.03; sırasıyla balon anjiyoplasti uygulanmış ve uygulanmamış ilyak arterler,
Student t testi, P<0.001, n=3).

94
3.1.2. İmmunohistokimyasal İncelemeler
3.1.2.1. Ki–67 Ekspresyonu
Tavşan ilyak arterine balon uygulamasına bağlı olarak 0., 2., 7. ve 14. günler
sonunda proliferasyon belirteci olan Ki-67’nin MIB-1 klonu (Dako, Glostrup,
Danimarka) ile herhangi bir boya görülmezken (Şekil 3.3 A), Ki-67’nin SP6 klonu
(Labvision, Fremont, Kanada) ile nükleer alanda pozitif boyanma saptanmıştır
(Şekil 3.3 B).
A) B)
Şekil 3.3.: Balon anjiyoplasti uygulaması sonrası 14. günde izole edilerek parafin ile sabitlendirilmiş ve A) Ki-67 MIB-1 klonu ile etkileştirilmiş ilyak arter damar kesiti -boyama (-) (X20 büyütme) B) Ki-67 SP6 klonu ile etkileştirilmiş ilyak arter damar kesiti –Boyama (+) (X40 büyütme)

95
3.1.2.2. Prolifere Olan Hücre Nükleer Antijeni (Proliferating Cell Nuclear
Antigen = PCNA) Ekspresyonu
Tavşan ilyak arterine balon uygulamasına bağlı olarak 0., 2., 7. ve 14. günler
sonunda proliferasyon belirteci olan PCNA ile nükleer alanda pozitif bir boyanma
saptanmıştır (Şekil 3.4.).
A) B)
C) D)
Şekil 3.4.: Balon anjiyoplasti uygulaması sonrası A)14. günde B) 7. günde C) 2. günde D) 0. günde izole edilerek parafin ile sabitlendirilmiş ve PCNA ile etkileştirilmiş ilyak arter damar kesitinde intimal kalınlaşma alanında PCNA ile pozitif boyanma (X10 büyütme)
3.1.2.3. α-Aktin Ekspresyonu
Balon anjiyoplasti uygulaması sonrası intimal kalınlaşma gelişen tavşan ilyak
arterlerinde düz kasa spesifik α-aktin ekspresyonu saptanmıştır (Şekil 3.5.).

96
Şekil 3.5.: Balon anjiyoplasti sırasında hasar verilen ve uygulama sonrası 14. günde izole edilen damarlarda gelişen intimal kalınlaşmada α-aktin ekspresyonu saptanmıştır (kahve renkli boyanma).
3.1.3. Telomer Fragmanlarının Uzunluğu
3.1.3.1. DNA İzolasyonu ve Miktarının Saptanması
Tavşan ilyak arterine balon anjiyoplasti uygulanmasının telomer boyu üzerine
olan etkilerini araştırmak üzere telomerik tekrar restriksiyonel fragmantasyon
(Terminal Restriction Fragments, TRF) deneyleri gerçekleştirilmiştir (Bakınız Gereç
ve Yöntem). Bu amaçla homojenize edilen dokulardan Gereç ve Yöntem bölümünde
ayrıntılı olarak belirtildiği gibi DNA izolasyonu yapılmıştır. DNA izolasyonu sonucu
elde edilen örneklerde DNA düzeyi spektrofotometrik yöntem ile belirlenmiştir.
Tablo 3.1.’de örneklerin absorbans değerleri ve DNA konsantrasyonları verilmiştir.
Bu veriler telomerik fragmanların incelenmesine yeterli olacak düzeyde genomik
DNA elde edildiğini göstermektedir. İzole edilen DNA örneklerinin agaroz jelde
yürütülmesi ve görüntülenmesi sonucu elde edilen analiz de Şekil 3.6’da
verilmektedir.

97
3.1.3.2. DNA’nın Restriksiyon Enzimleri ile Kesilmesi
Uygulanan protokol gereği izole edilen genomik DNA Gereç ve Yöntem
bölümünde ayrıntılı olarak belirtildiği gibi restriksiyon endonükleazları ile
kesilmiştir. Restriksiyon sonrası elde edilen görüntü Şekil 3.6.’te verilmektedir.
Restriksiyon öncesi ve sonrasına ait olan görüntüler (Şekil 3.6. A ve B)
karşılaştırıldığında genomik DNA moleküllerinin uygulanan restriksiyon kesimine
yanıt verdiği, telomerik fragmanları kapsayan moleküllerin daha kısa fragmanlara
dönüştüğü görülmektedir.
3.1.3.3. DNA’nın Jelden Membrana Transfer Edilmesi
3.1.3.3.1. Radyoaktif Olmayan İşaretleme ile Yapılan Deneyler
Restriksiyon sonrası görüntülenen moleküllerin telomerik fragmanlara ait olup
olmadığını saptamak üzere, agaroz jelleri Southern blot tekniği ile nitroselüloz
membranlara transfer edilmişlerdir. Bu amaçla membranlar radyoaktif olmayan bir
işaretleyici olan digoksigenin ile işaretli telomerik oligo ile hibridize edilmiş ve
tabedilmiştir (Şekil 3.7). Bu görüntü incelendiğinde kısa telomerik fragmanların
beklendiği membran bölgesinde daha az yoğunlukta sürüntüler görülmektedir.

98
Tablo 3.1.: DNA izolasyonu sonrası elde edilen örneklerin absorbans değerleri.
Örnek No* Absorbans Konsantrasyon (µg/µl)
1 0.073 0.073
2 0.044 0.044
3 0.050 0.050
4 0.044 0.044
5 0.046 0.046
6 0.045 0.045
7 0.042 0.042
8 0.048 0.048
* Örnekler jele uygulama sırasına göre belirtilmiştir.
A) B) 1 2 3 4 5 6 7 8 1 2 3 4 5 6 7 8 9 10 11
Şekil 3.6.: A) DNA izolasyonu sonrası genomik DNA’nın %0.8’lik agaroz jele uygulanması sonucu elde edilen 1. jele ilişkin görüntü ( 40 V/saat; toplam 160 V); B) Restriksiyon sonrası elde edilen görüntü. 1. sıra moleküler büyüklük standardı, 2. ve 3. sıra yüksek ve düşük standartlar.

99
1 2 3 4 5 6 7 8 9 10 11
Şekil 3. 7.: DNA izolasyonu ve restriksiyonu yapılan örneklerin (1. jel) Southern blot ve hibridizasyon sonrası elde edilen görüntüsü.
Örneklerdeki telomerik DNA fragmanlarının uzun olup olmadığını test etmek
amacıyla deney aynı koşullarda yinelenmiştir. Bu denemeye ilişkin restriksiyon
öncesi ve sonrası elde edilen jel görüntüleri Şekil 3.8.A ve B’de verilmektedir (2.
jel). Şekil 3.8.B’de, daha önce yapılan deney sonuçlarına (Şekil 3.6.B) benzer şekilde
genomik DNA moleküllerinin uygulanan restriksiyon kesimine yanıt verdiği ve
telomerik fragmanları kapsayan moleküllerin daha kısa fragmanlara dönüştüğü
görülmektedir.
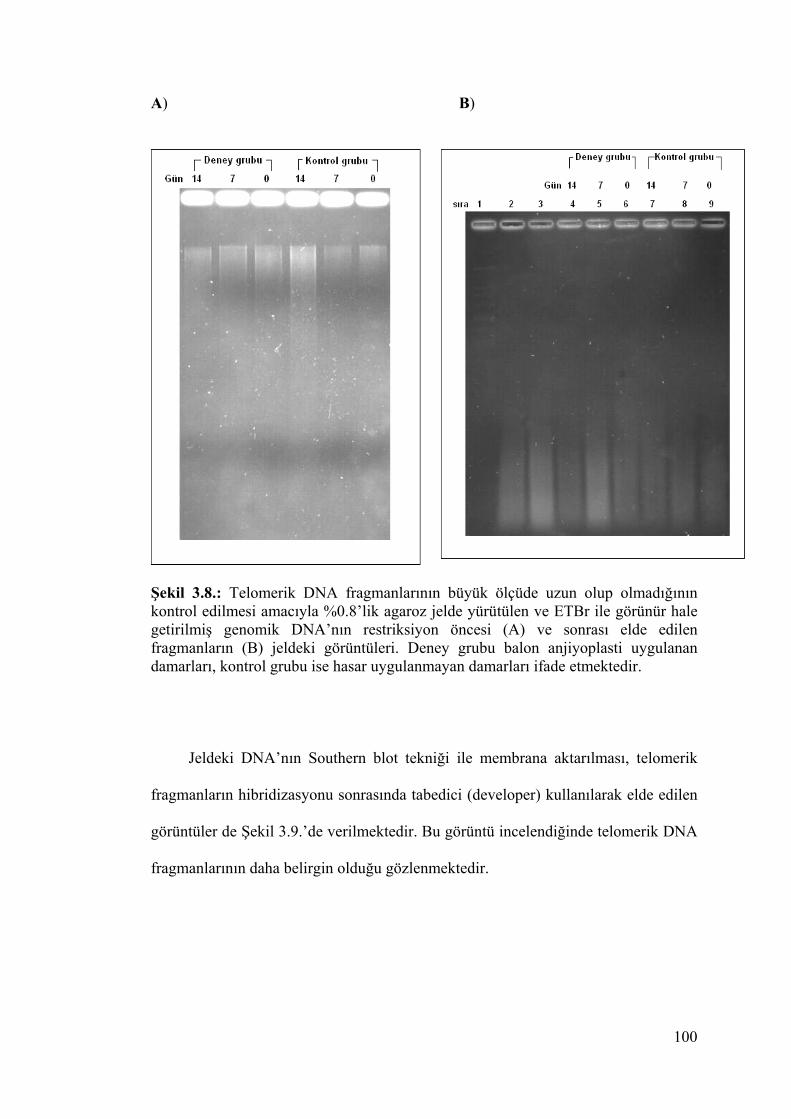
100
A) B)
Şekil 3.8.: Telomerik DNA fragmanlarının büyük ölçüde uzun olup olmadığının kontrol edilmesi amacıyla %0.8’lik agaroz jelde yürütülen ve ETBr ile görünür hale getirilmiş genomik DNA’nın restriksiyon öncesi (A) ve sonrası elde edilen fragmanların (B) jeldeki görüntüleri. Deney grubu balon anjiyoplasti uygulanan damarları, kontrol grubu ise hasar uygulanmayan damarları ifade etmektedir.
Jeldeki DNA’nın Southern blot tekniği ile membrana aktarılması, telomerik
fragmanların hibridizasyonu sonrasında tabedici (developer) kullanılarak elde edilen
görüntüler de Şekil 3.9.’de verilmektedir. Bu görüntü incelendiğinde telomerik DNA
fragmanlarının daha belirgin olduğu gözlenmektedir.

101
Şekil 3.9.: İkinci denemeye ilişkin (2. jel) DNA izolasyonu ve restriksiyonu yapılan örneklerin Southern blot ve hibridizasyon sonrası elde edilen görüntü (Deney grubu balon anjiyoplasti uygulanan damarları, kontrol grubu hasar uygulanmayan damarları ifade etmektedir). 1. sıra moleküler büyüklük standardı, 2. ve 3. sıra yüksek ve düşük standartlar.
Gereç ve Yöntem’de belirtilen protokol izlenerek bir diğer deney serisinden (3. jel)
elde edilen blot görüntüsü de Şekil 3.10.’da verilmektedir.

102
Şekil 3.10.: DNA izolasyonu ve restriksiyon sonrası Southern blot ve hibridizasyonu yapılan bir diğer deney serisine ilişkin jelin (3. jel) görüntüsü (Deney grubu balon anjiyoplasti uygulanan damarları, kontrol grubu hasar uygulanmayan damarları ifade etmektedir). 1. sıra moleküler büyüklük standardı.
3.1.3.3.2. Radyoaktif İşaretleme ile Yapılan Deneyler
Bu amaçla 32P işaretli DNA dizisi kullanılmıştır. Digoksigenin ile işaretli
telomerik oligo ile gerçekleştirilen deneylerin sonucunda elde edilen görüntülerin
duyarlığını test etmek amacıyla, aynı deney protokolu uygulanarak hazırlanan
dördüncü bir jel membrana aktarılarak ve Georgia Üniversitesi Genetik Bölümü’ne
(Dr. McEachern, ABD) gönderilmiştir. Bu deneye ilişkin sonuçlar Şekil 3.11.’da
verilmiştir.

103
Şekil 3.11.: 32P işaretleme ile elde edilen radyografi (Deney grubu balon anjiyoplasti uygulanan damarları, kontrol grubu hasar uygulanmayan damarları ifade etmektedir).

104
3.1.3.4. Telomerik Fragmanların Kantitatif Değerlendirilmesi
İkinci ve üçüncü jelin blotlanması sonucu elde edilen Şekil 3.9. ve Şekil
3.10.’da verilen telomerik fragman analizlerinin kantitatif değerlendirilmesi (Bakınız
Gereç ve Yöntem) yapılmıştır (100). Elde edilen sonuçlar sırasıyla Tablo 3.2. ve
3.3.’te verilmektedir. Yapılan incelemeler ilyak artere balon anjiyoplasti uygulaması
sonucu telomer boyunun ilk ikinci denemede 7. ve 14. günlerde sırasıyla % 71 ve %
77; üçüncü denemede 2., 7. ve 14. günlerde hasar görmeyen ilyak artere göre
sırasıyla % 64, % 85 ve % 63 oranlarında azalma gösterdiği belirlenmiştir.
Benzer şekilde radyoaktif işaretleme yapılarak elde edilen blot görüntüsündeki
(Şekil 3.11.) rölatif ortalama telomer boylarına (% OTF) ilişkin sonuçlar Tablo
3.4.’de verilmiştir. Yapılan analizler ortalama telomer uzunluğunun balon
anjiyoplasti uygulanan ilyak arterlerde, uygulamanın 2. ve 14. günlerinde %53 ve
%74 oranında anlamlı bir şekilde azaldığını göstermektedir (OTF, kbç:3.03±0.83,
1.61±0.09, 2.66±0.29, 2.27±0.19 sırasıyla 0., 2., 7. ve 14. günler, P<0.01, 0. ve 2.
gün arasında; P<0.05 0. ve 14. gün arasında, Student t testi, n=3).

105
Tablo 3.2.: Şekil 3.9’deki blot görüntüsüne ait ortalama telomerik fragman (OTF) değerleri (OTF değerleri deneysel grupta kontrol gruba göre yüzdelenmiştir).
(OTF) (%)
Gün Deneysel Grup Kontrol
Grubu
0 100 100
7 71 100
14 77 100
Tablo 3.3.: Şekil 3.10’daki blot görüntüsüne ait OTF değerleri Kontrol grubuna göre yüzdelendiğinde elde edilen % değerler.
OTF (%)
Gün Deneysel Grup Kontrol
Grubu
0 100 100
2 64 100
7 85 100
14 63 100

106
Tablo 3.4.: Şekil 3.11’de radyoaktif işaretleme sonucu elde edilen blot görüntüsü verilen örneklerin ortalama rölatif telomerik fragman (OTF) değerleri
OTF (%)
Günler Deneysel Grup Kontrol Grubu
0 100 100
2 53 73
7 87 82
14 74 88
3.1.4. Telomeraz Biyoaktivitesi
Tavşan ilyak arterine balon anjiyoplasti uygulaması sonucu telomer boy
uzunluğunu belirlemek için kullanılan örneklerde TRAP analizi yapılarak telomeraz
biyoaktivitesi araştırılmıştır. Sonuçlar Şekil 3.12.’de verilmektedir. Bulgular,
TRAPeze kitini üreten firmanın telomeraz ekspresyonu varlığında beklenen jel
görüntüsünden farklıdır (Şekil 3.13.) Buradan hareketle, TRAP analizi deneyleri
ilyak artere balon anjiyoplasti uygulamasının telomeraz ekspresyonuna neden
olmadığını ortaya koymuştur.

107
Şekil 3.12: TRAP analizi sonucu elde edilen poliakrilamid jel fotoğrafı. Deney grubu balon anjiyoplasti uygulanan damarları, kontrol grubu hasar uygulanmayan damarları ifade etmektedir.

108
Şekil 3.13. TRAPeze (Chemicon, ABD) kiti ile elde edilen telomeraz aktivitesine ilişkin ideal jel görüntüsü.

109
3.2. Bölüm II
Bu bölümde tavşan iliyak arterine balon anjiyoplasti uygulanması sonrası
vasküler düz kas hücrelerinde telomer uzunluğuna oksidatif stresin etkisinin
araştırılmasına ilişkin deneyler yapılmıştır. Elde edilen bulgular aşağıdaki gibidir;
3.2.1. Morfometrik İncelemeler
Alınan kesitlerden, intima, medya, lümen alanı ölçümleri yapılmıştır. Aşağıda
her gruba ait deney hayvanlarından birer örnek kesit verilmektedir (Şekil 3.14).
Yapılan istatistiksel değerlendirmeler;
a. Balon hasarı oluşturulmuş damarlarda, BSO ile işlem görmenin
intimal hiperplaziyi istatistiksel olarak anlamlı bir şekilde artırdığını (p<0.01,
ANOVA sonrası Bonferoni testi),
b. BSO uygulamasının tek başına intimal hiperplazi oluşumuna
katkıda bulunmadığını,
c. Taurin ile tedavinin BSO ile artan intimal hiperplaziyi istatistiksel
olarak anlamlı şekilde azalttığını (p<0.01, ANOVA sonrası Bonferoni testi)ortaya
koymaktadır.
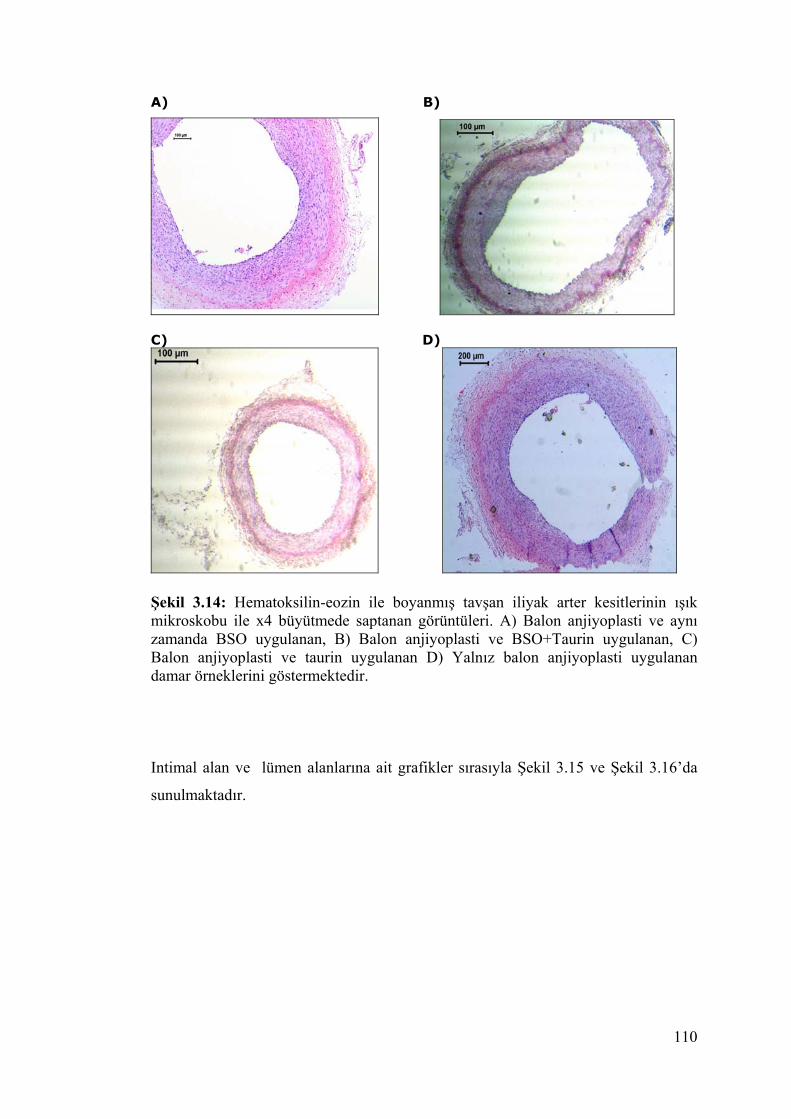
110
A) B)
C) D)
Şekil 3.14: Hematoksilin-eozin ile boyanmış tavşan iliyak arter kesitlerinin ışık mikroskobu ile x4 büyütmede saptanan görüntüleri. A) Balon anjiyoplasti ve aynı zamanda BSO uygulanan, B) Balon anjiyoplasti ve BSO+Taurin uygulanan, C) Balon anjiyoplasti ve taurin uygulanan D) Yalnız balon anjiyoplasti uygulanan damar örneklerini göstermektedir.
Intimal alan ve lümen alanlarına ait grafikler sırasıyla Şekil 3.15 ve Şekil 3.16’da
sunulmaktadır.

111
Şekil 3.15 İntimal alan (mm2) değerleri (Kontrol: Hasar görmemiş ilyak arter, B: Balon uygulaması, T: Taurin BT: BSO ve Taurinin birlikte uygulanması) (ANOVA sonrası Bonferoni test) (Herbir grup için n=3)
Şekil 3.16: Lümen alanı (mm2) değerleri (Kontrol:Herhangi bir işlem görmemiş damar, B: Balon uygulaması, T: Taurin BT: BSO ve Taurinin birlikte uygulanması) (ANOVA sonrası Bonferoni test) (Herbir grup için n=3)

112
3.2.2. İmmünohistokimyasal İncelemeler
3.2.2.1. Ki–67 Ekspresyonu
Tavşan ilyak arterine balon uygulamasına bağlı olarak her dört grubun balon
anjiyoplasti uygulanmış damarlarında proliferasyon belirteci olan Ki-67 (SP6 klonu)
ile nükleer alanda pozitif bir boyanma saptanmıştır (Şekil 3.17.).
Şekil 3.17.: Balon anjiyoplasti uygulaması sonrası 14. günde izole edilerek parafin ile sabitlendirilmiş ve Ki-67 ile etkileştirilmiş ilyak arter damar kesitinde intimal kalınlaşma alanında. Ki-67 ile pozitif boyama (X40 büyütme)

113
3.1.2.2. Prolifere Olan Hücre Nükleer Antijeni (Proliferating Cell Nuclear
Antigen = PCNA) Ekspresyonu
Tavşan ilyak arterine balon uygulamasına bağlı olarak her dört grubun balon
anjiyoplasti uygulanmış damarlarında proliferasyon belirteci olan PCNA ile nükleer
alanda pozitif bir boyanma saptanmıştır (Şekil 3.18.).
A) B)
C) D)
Şekil 3.18.: Balon anjiyoplasti uygulaması sonrası 14. günde izole edilerek parafin ile sabitlendirilmiş ve PCNA ile etkileştirilmiş ilyak arter damar kesitinde intimal kalınlaşma alanında. PCNA ile pozitif boyama (X10 büyütme) A)Balon+BSO B)Balon+BSO+T C)Balon D)Balon+Taurin

114
3.1.2.3. α-Aktin Ekspresyonu
Tavşan ilyak arterine balon uygulamasına bağlı olarak her dört grubun balon
anjiyoplasti uygulanmış damarlarında da düz kasa spesifik α-aktin’a spesigik
boyanma saptanmıştır (Şekil 3.19.).
A) B)
C) D)
Şekil 3.19.: Balon anjiyoplasti uygulaması sonrası 14. günde izole edilerek parafin ile sabitlendirilmiş ve α-aktin ile etkileştirilmiş ilyak arter damar kesitinde intimal kalınlaşma alanında α-aktin ile pozitif boyanma (X10 büyütme) A)Balon+BSO B)Balon+BSO+T C)Balon D)Balon+T

115
3.2.3. Kanda Yapılan Ölçümler
3.2.3.1. Plazma Total Glutatyon (GSH) Miktarı
Ölçümler sonucunda yapılan istatistiksel analizler BSO ile glutatyonun
tüketilmesinin balon hasarı sonrası 7. gün ve 14. günlerde alınan kan örneklerinde
total GSH konsantrasyonunu istatiksel olarak anlamlı bir şekilde azalttığını
göstermektedir. BSO ve taurinin birlikte uygulanması bu azalmayı anlamlı bir
şekilde geri döndürmektedir (Tablo 3.5).
Tablo 3.5.: Plazmadaki total glutatyon konstrasyonu (µM)
0.gün 7.gün 14.gün
Kontrol (n=3) 1111.3±67.96 981±52.75* 1052±109.81**
BSO (n=3) 1098.1±66.18 675.73±28.06+ 431.2±34.23++,#
BSO+Taurin (n=3) 1072.1±102.96 968.81±25.61## 1049.9±53.57***
Taurin (n=3) 1259.8±60.32 1119.7±87.43§ 1036.7±56.63§§
*Kontrol 7. gün vs BSO 7. gün (p<0.05) **Kontrol 14. gün vs BSO 14. gün (p<0.001) +BSO 0. gün vs BSO 7. gün (p<0.01) ++BSO 0. gün vs BSO 14. gün (p<0.001) #BSO 7. gün vs BSO 14. gün (p<0.05) ## BSO 7. gün vs BSO+Taurin 7. gün (p<0.05) ***BSO 14. gün vs BSO+Taurin 14. gün (p<0.001) §BSO 7. gün vs Taurin 7. gün (p<0.01) §§BSO 14. gün vs Taurin 14. gün (p<0.01) (Değerler ortalama ± ortalamanın standart hatası olarak verilmiştir. ANOVA sonrası Bonferoni test kullanılmıştır)

116
3.2.3.2. Plazma Glutatyon /Okside Glutatyon (GSH/GSSG) Oranları
Ölçümler sonucunda yapılan istatistiksel analizler BSO ile glutatyonun
tüketilmesinin balon hasarı sonrası 7. gün ve 14. günlerde alınan kan örneklerinde
GSH/GSSG oranını istatistiksel olarak anlamlı bir şekilde azalttığını göstermektedir.
BSO ve taurinin birlikte uygulanması bu oranı istatistiksel olarak anlamlı bir şekilde
normalize etmektedir (Tablo3.6)
Tablo 3.6.: Plazma GSH/GSSG Oranları
0.gün 7.gün 14.gün
Kontrol (n=3) 176.99±3.49 171.72±3.28* 166.49±1.97**
BSO (n=3) 166.96±2.4 24.68±2.25+ 78.85±7.79++,#
BSO+Taurin (n=3) 172.55±5.05 164.78±4.27## 156.21±5.50***
Taurin (n=3) 175.56±3.74 173.80±2.73§ 175.19±2.92§§
*Kontrol 7. gün vs BSO 7. gün (p<0.01) **Kontrol 14. gün vs BSO 14. gün (p<0.001) +BSO 0. gün vs BSO 7. gün (p<0.001) ++BSO 0. gün vs BSO 14. gün (p<0.001) #BSO 7. gün vs BSO 14. gün (p<0.001) ## BSO 7. gün vs BSO+Taurin 7. gün (p<0.01) *** BSO 14. gün vs BSO+Taurin 14. gün (p<0.01) §BSO 7. gün vs Taurin 7. gün (p<0.01) §§BSO 14. gün vs Taurin 14. gün (p<0.01)
(Değerler ortalama ± ortalamanın standart hatası olarak verilmiştir. ANOVA sonrası Bonferoni test kullaılmıştır)

117
3.2.3.3. Plazma Glutatyon Peroksidaz (GPx) Enzim Aktivitesi
Ölçümler sonucunda yapılan istatistiksel analizler BSO ile glutatyonun
tüketilmesinin balon hasarı sonrası 7. gün ve 14. günlerde alınan kan örneklerinde
glutatyon peroksidaz enziminin aktivitesini istatistiksel olarak anlamlı bir şekilde
azalttığını göstermektedir. BSO ve taurin’in birlikte uygulanması bu azalmayı
istatistiksel olarak anlamlı bir şekilde normalize etmektedir (Tablo 3.7)
Tablo 3.7: Plazma GPx (µmol/NADPH/dk/ml) Enzim Aktivitesi
0.gün 7.gün 14.gün
Kontrol (n=3) 2.80±0.03 2.75±0.03* 2.76±0.02**
BSO (n=3) 2.72±0.04 2.12±0.08+ 1.64±0.12++,#
BSO+Taurin (n=3) 2.76±0.07 2.69±0.02## 2.65±0.09***
Taurin (n=3) 2.69±0.10 2.70±0.12§ 2.80±0.04§§
*Kontrol 7. gün vs BSO 7. gün (p<0.01) **Kontrol 14. gün vs BSO 14. gün (p<0.001) +BSO 0. gün vs BSO 7. gün (p<0.01) ++BSO 0. gün vs BSO 14. gün (p<0.01) #BSO 7. gün vs BSO 14. gün (p<0.001) ## BSO 7. gün vs BSO+Taurin 7. gün (p<0.01) ***BSO 14. gün vs BSO+Taurin 14. gün (p<0.01) §BSO 7. gün vs Taurin 7. gün (p<0.01)
§§BSO 14. gün vs Taurin 14. gün (p<0.01) (Değerler ortalama ± ortalamanın standart hatası olarak verilmiştir. ANOVA sonrası Bonferoni test kullaılmıştır)

118
3.2.4. Telomer Fragmanlarının Uzunluğu
3.2.4.1. DNA İzolasyonu ve Miktarının Saptanması
Tavşan iliyak arterine uygulanan balon anjiyoplasti sonrası vasküler düz kas
hücrelerinde telomer uzunluğuna oksidatif stresin etkisinin araştırılması amacıyla bir
önceki bölümde olduğu gibi telomerik tekrar restriksiyonel fragmantasyon (Terminal
Restriction Fragments, TRF) deneyleri gerçekleştirilmiştir (Bakınız Gereç ve
Yöntem). Bu amaçla homojenize edilen dokulardan Gereç ve Yöntem bölümünde
ayrıntılı olarak belirtildiği gibi DNA izolasyonu yapılmıştır. DNA izolasyonu sonucu
elde edilen örneklerde DNA düzeyi spektrofotometrik yöntem ile belirlenmiştir.
Tablo 3.8’de örneklerin absorbans değerleri ve DNA konsantrasyonları verilmiştir.
Bu veriler telomerik fragmanların incelenmesine yeterli olacak düzeyde genomik
DNA elde edildiğini göstermektedir.
Daha sonra izole edilen ve miktarları belirlenen DNA örnekleri eşit miktarlarda
olacak şekilde % 0.8’lik agaroz jelde yürütülmüştür. Aynı örneklerin bir kısmı ise
EcoRI enzimi ile 1 saat 37˚C’de inkübe edildikten sonra % 0.8’lik jelde
yürütülmüştür. Aynı örneklere her iki uygulamanın karşılaştırılmalı olarak aynı jelde
görülebileceği örnek jel fotoğrafı Şekil 3.20’de verilmektedir. Bu jelden elde edilen
görüntü spektrofotometrik olarak yapılan miktar tayini ile paralellik göstermektedir
ve Southern blot yönteminde iyi bir jel görüntüsü elde edebilmek için her bir
kuyucuğa ~1 µg DNA uygulamasının yeterli olacağı sonucuna varılmıştır.

119
Tablo 3.8: DNA izolasyonu sonrası elde edilen örneklerin absorbans değerleri.
Örnek No. Absorbans Konsantrasyon
(µg/ml)
1 0.062 62 2 0.069 69 3 0.077 77 4 0.062 62 5 0.056 56 6 0.051 51 7 0.058 58 8 0.057 57 9 0.066 66
10 0.063 63 11 0.123 123 12 0.085 85 13 0.088 88 14 0.097 97 15 0.145 145 16 0.07 70 17 0.094 94 18 0.144 144 19 0.116 116 20 0.062 62 21 0.065 65 22 0.068 68 23 0.091 91 24 0.054 54

120
Şekil 3.20: Telomer analizi öncesi DNA miktarının gözlenebilmesi amacıyla elde edilen jel fotoğrafı. Üst sıra: EcoRI ile kesilmiş DNA, alt sıra: Aynı örneklere ait herhangi bir işlem görmemiş genomik DNA(uygulama sırası aynıdır).
3.2.4.2. DNA’nın Restriksiyon Enzimleri ile Kesilmesi
Uygulanan protokol gereği izole edilen genomik DNA Gereç ve Yöntem
bölümünde ayrıntılı olarak belirtildiği gibi restriksiyon endonükleazları ile
kesilmiştir. Restriksiyon sonrası elde edilen görüntü Şekil 3.21’da verilmektedir.

121
Telomerik fragmanları kapsayan moleküllerin daha kısa fragmanlara dönüştüğü
görülmektedir. BSO almış tavşanlara ait DNA örneklerinin bir kısmında
restriksiyonun tam olmadığı, hala kesilmemiş DNA’nın kaldığı görülmektedir.
Şekil 3.21: Restriksiyon sonrası jel fotoğrafı. (mw: moleküler ağırlık belirteci, B: Balon angiyoplasti yapılan damar, N: Hasar uygulanmamış damar)

122
3.2.4.3. DNA’nın Jelden Membrana Transfer Edilmesi
Restriksiyon sonrası görüntülenen moleküllerin telomerik fragmanlara ait olup
olmadığını saptamak üzere, agaroz jelleri Southern blot tekniği ile nitroselüloz
membranlara transfer edilmişlerdir. Bu amaçla membranlar radyoaktif olmayan bir
işaretleyici olan digoksigenin ile işaretli telomerik oligo ile hibridize edilmiş ve
tabedilmiştir (Şekil 3.22).
Daha önce Bölüm I’de yapılan deney sonuçlarına benzer şekilde genomik
DNA moleküllerinin uygulanan restriksiyon kesimine yanıt verdiği ve telomerik
fragmanları kapsayan moleküllerin genel daha kısa fragmanlara dönüştüğü
görülmektedir. Daha önce transfer sonrası elde edilen görüntülerden farklı olarak üst
bölgelerde de yoğun telomerik işaretlenme görülmektedir. Yine Bölüm I’de yapılan
telomer analizlerinden farklı olarak bu defa telomerik fragmanların görünür hale
gelmesi için filmin daha uzun süre (8 saat) membrana maruz bırakılması gerekmiştir.
Üst bölgelerde edilen yoğun bağlanmanın nonspesifik bağlanma olup
olmadığının belirlenmesi ve bunun doğru olduğu durumda daha kısa telomerik
bölgelere probun daha yüksek yoğunlukta bağlanması ve de böylelikle daha kısa
fragmanları da görünür hale getirebilmek amacıyla membranın kuyucuklara yakın üst
bölgeleri kesilerek hibridizasyon deneyi tekrarlanmıştır. Bu amaçla üst kısımları
kesilen membrandan bağlı substrat, enzim ve prob kompleksinin ayrılması amacıyla
membran önce denatüre edilmiş, daha sonra probla yeniden hibridize edilerek tekrar
filme aktarılmıştır. Elde edilen yeni görüntünde, beklediği gibi daha önceki
görüntüye göre fragmanların daha belirgin hale gelmediği görülmüştür. (Şekil 3.23)

123
Şekil 3.22: Transfer ve hibridizasyon sonrası film görüntüsü (mw: moleküker ağırlık belirteci, B: Balon angiyoplasti yapılan damar, N: Hasar uygulanmamış damar)(Film mebrana 8 saat boyunca maruz bırakılmıştır)

124
Şekil 3.23: 1. membranda hibridizasyonun yoğun olduğu üst bölgeler çıkarıldıktan sonra elde edilen membran fotoğrafı (film membrana 16 saat boyunca maruz bırakılmıştır)

125
Şekil 3.24: 2. denemeye ait restriksiyon sonrası jel görüntüsü (mw: moleküker ağırlık belirteci, B: Balon angiyoplasti yapılan damar, N: Hasar uygulanmamış damar)
2. denemeye ait restriksiyon sonrası jel fotoğrafında da (Şekil 3.24) telomerik
fragmanları kapsayan moleküllerin daha kısa fragmanlara dönüştüğü görülmektedir.
BSO almış tavşanlara ait DNA örneklerinin bir kısmında birinci denemeye benzer

126
şekilde restriksiyonun tam olmadığı, hala kesilmemiş bir kısım DNA’nın kaldığı
görülmektedir. Jel membrana transfer edilip hibridize edildikten sonraki görüntü
1.denemedeki membran görüntüsü ile paralellik göstermektedir (Şekil 3.25).
Şekil 3.25: 2. jelde transfer ve hibridizasyon sonrası membrana ait film görüntüsü (mw: moleküker ağırlık belirteci, B: Balon angiyoplasti yapılan damar, N: Karşı damar) (film membrana 16 saat boyunca maruz bırakılmıştır)

127
3.2.4.4. Telomerik Fragmanların Kantitatif Değerlendirilmesi
Birinci jelin blotlanması sonucu elde edilen Şekil 3.23 ve Şekil 3.24’de verilen
telomerik fragman analizlerinin kantitatif değerlendirilmesi (Bakınız Gereç ve
Yöntem) yapılmıştır. Elde edilen sonuçlar Tablo 3.9 da verilmektedir. Yapılan
incelemeler bireysel farklılığı ortadan kaldırmak amacıyla hasarlı damara ait OTF
değeri karşı damara ait OTF değeri ile yüzdelediğinde BSO’lu damarların
telomerlerinin diğer gruplara göre anlamlı olarak bir şekilde daha kısa olduğunu
ortaya çıkmaktadır (Tablo 3.9)
Tablo 3.9: Ortalama Telomer Boylarının Yüzdesi
% OTF
Deneysel Grubun Karşı Damara Göre Yüzdesi
Kontrol (n=3) 72.33±1.453
BSO (n=3) 56.67±2.028*
BSO+Taurin (n=3) 66.67±2.603**
Taurin (n=3) 77.00±2.082+, ++
*
BSO vs Kontrol p<0.01, **BSO vs BT p<0.05, +BSO vs T p<0.001, ++BT vs T p<0.05

128
4. TARTIŞMA
Damar düz kas hücre proliferasyonu ve migrasyonuna bağlı olarak meydana
gelen intimal hiperplazinin aterosklerozun ve balon anjiyoplasti sonrası oluşan
restenozun patojenezinden sorumlu olduğu bilinmektedir (28, 29, 48, 92). Vasküler
düz hücre migrasyon ve proliferasyonunu düzenleyen moleküler yolakları
aydınlatmaya ve vasküler düz kas hücre hiperplazisini önlemeye yönelik stratejileri
geliştirmek üzere çok sayıda çalışma yapılmıştır. Bununla birlikte neden olan
mekanizmaların büyük oranda bilinmemesi nedeniyle etkin bir tedavi
geliştirilememiştir (52, 105).
Yakın zamanda yapılan çalışmalarda telomeraz aktivasyonu ve telomer
devamlılığının genetik hipertansiyonda vasküler yeniden modellenmenin
oluşumunun temelinde yatan artan damar düz kas hücre proliferasyonunu
yönlendirdiği ileri sürülmektedir (124, 190). Benzer şekilde, vasküler damar düz kas
hücrelerinde, hücre proliferasyonu sürecinde telomerazın aktive olduğu, buna
karşılık telomeraz aktivitesinin inhibisyonu ile hücre büyümesinin azaldığı da
gösterilmiştir (20). Diğer taraftan yapılan birçok çalışmada vasküler hastalıklarda
hücre yaşlanma sürecinin hastalık patojenezine olan katkısı üzerinde de
durulmaktadır (36, 116, 119).

129
Telomerlerin replikatif olarak kısalmasının hücre yaşlanma sürecini başlattığı
bilinmektedir. Son yıllarda yapılan çalışmalar oksidatif stresin bu süreci kısaltarak
“Stres kaynaklı prematüre yaşlanma” (Stress-induced premature senescence= SIPS)
ya neden olduğunu ortaya koymaktadır (15, 36, 62, 181). Ateroskerozda damar
duvarının bütün katmanlarında reaktif oksijen türlerinin (reactive oxygen species=
ROS) varlığı gösterilmiştir. Bütün bu bilgiler ışığında çalışmamızın sonuçları iki
farklı başlık altında tartışılabilir:
Bölüm I: Tavşan Balon Anjiyoplasti Modelinde Telomeraz Aktivitesi ve
Telomer Devamlılığının Lezyon Süresi ile İlişkisi
Morfometri ve İmmünohistokimyasal İncelemeler
Çalışmamızda insan koroner arterine büyük oranda benzerlik gösteren tavşan
ilyak arteri kullanılmıştır. Balon anjiyoplasti sonrası, farklı zaman dilimlerinde (2., 7.
ve 14. gün) hasar gören ve görmeyen (kontrol) ilyak arterlerden alınan seri kesitlerde
gerek Ki-67 gerek PCNA antikorları ile yapılan immünohistokimyasal boyamalarda
hasar gören ilyak arterdeki intimal alanda tüm zaman dilimlerinde proliferasyon
saptanmıştır. Arter duvarı hasarı veya aterosklerozun erken döneminde vasküler düz
kas hücreleri çeşitli mitojenlere yanıt olarak kontraktil bir fenotipten sentetik bir
fenotipe geçerek prolifere olabilirler (153, 163). İnsan Ki-67 proteini
ekspresyonunun hücre proliferasyonu ile doğrudan ilişkili olduğu gösterilmiştir
(162). İnterfaz süresinde nükleus içinde bol miktarda saptanabilen antijen mitoz
sırasında kromozomların yüzeyinde yerleşim gösterir. Ki-67 proteininin G1, S, G2 ve
mitoz gibi hücre siklüsünün tüm aktif fazlarında eksprese edildiği ancak istirahat

130
fazındaki (G0) hücrelerde eksprese edilmediği bilinmektedir (49). Bu özelliği
nedeniyle de protein hücre proliferasyonunun bir belirteci olarak değerlendirilmekte
ayrıca bazı kanser türlerinde de yüksek orandaki ekspresyonu ile kanser prognozu
arasında bir ilişki olduğu bildirilmektedir (186). Çalışmamızda Ki-67 ile ilgili iki
farklı bulgu söz konusudur. Çalışmamızda kullanılan iki anti-Ki-67 klonundan MIB-
1 klonu ile boyanma olmamış, buna karşılık SP6 klonu ile ise boyanma görülmüştür.
Bu farklılık tavşan iliyak arter balon anjiyoplasti modelinin anti-Ki-67 antikorunun
MIB-1 klonuna karşı dirençli olabileceğini düşündürmektedir.
Diğer bir proliferasyon belirteci olan PCNA’in ise geç G1 fazından erken S
fazına kadar eksprese olabilen bir nükleer antijen olduğu bilinmektedir (25, 44).
Aoyagi ve arkardaşları (6) tavşan karotid arteri balon hasarı modelinde tümör
baskılayıcı bir protein olan p53 proteini ile proliferasyona yanıt olarak PCNA’nın
aynı hücrelerde birlikte boyanma gösterdiğini ortaya koymuşlardır . PCNA’nın
Ki-67’ ye göre daha yoğun boyanma göstermesi bu hücrelerin hücre siklusunun G1
fazında kalıp S fazına geçmeyen sessiz (quiescent) hücreler olabileceğini
düşündürmektedir. Bu bulgu özellikle aterosklerotik hastalıklarda ateroskleroz
gelişimine hücre yaşlanma sürecinin katkısı düşünüldüğünde (27, 57, 127, 128) hücre
yaşlanması açısından önemli olabilir.
Çalışmamızda balon anjiyoplasti uygulaması sonrası 14. günde gelişen intimal
kalınlaşmada düz kasa özgül bir protein olan alfa-aktin ekspresyonunun saptanması
uyguladığımız modelde gelişen intimal kalınlaşmada medyadan intimaya göç eden
düz kas hücrelerinin de katkısı olduğunu düşündürmektedir. Yukarıda da belirtildiği
gibi değişik deneysel modellerde gelişen intimal hiperplaziye migrasyon ve

131
proliferasyonun katkısı tam olarak belirlenememiştir (7, 196, 199). Bu durum, ayrıca
hücre proliferasyonunu inhibe ettiği bilinen ve gösterilen bazı ilaçların, değişik
modellerdeki intimal hiperplaziyi engelleyememeleri bulgusu ile de uyum
göstermektedir (28, 196).
Telomeraz enzim biyoaktivitesi
Projemizde tavşanlara ilyak arter balon anjiyoplasti uygulaması sonucu gelişen
vasküler hasarda telomeraz aktivitesinin araştırılmasına yönelik olarak
gerçekleştirdiğimiz deneyler sonucunda hasar gören ve görmeyen ilyak arterlerde
telomeraz biyoaktivitesi saptanmamıştır. Son bulgular normal ve hipoksik koşullarda
insan vasküler düz kas hücrelerinin telomeraz aktivasyonu ve insan telomeraz ters
transkriptaz (human telomerase reverse transcriptase, hTERT) aracılığı ile prolifere
olduğunu göstermiştir (124,125,126). Bu bulgular telomerazın vasküler yeniden
modellenmede bir rolünün olabileceğini düşündürmektedir. Liu ve arkadaşları insan
aterosklerotik epikardiyal koroner arterlerinde yaptıkları çalışmada telomeraz
biyoaktivesi ve telomerazın katalitik birimi olan hTERT protein ekspresyonunun
upregüle olduğunu göstermişlerdir(110).
Çalışmamızın morfometrik ve immunohistokimyasal sonuçları balon
anjiyoplasti uygulanan ilyak arterlerde gelişen intimal kalınlaşmanın düz kas hücre
migrasyonu ve proliferasyonuna bağlı olarak ortaya çıktığını düşündürmektedir.
Bununla birlikte Liu ve arkadaşları yakın bir zamanda yaptıkları araştırmalarında
koroner aterosklerozlu hastaların bazılarında intima ve medyada Ki–67 ile
saptanabilen proliferasyon olmamakla birlikte telomeraz enzim aktivitesi

132
saptamışlardır (111). Ancak bu araştırmacılar koroner hastalığı olan hastaların büyük
çoğunluğunda proliferasyon ve telomeraz biyoaktivitesinin birlikte var olduğunu ve
hTERT immunoreaktivitesinin de geliştiğini göstermişlerdir.
Çalışmamızda uygulanan yöntemin duyarlık sınırları içerisinde tavşanlarda
ilyak artere balon anjiyoplasti uygulamasının telomeraz ekspresyonuna neden
olmadığı belirlenmiştir.
Telomerik Fragmanların Boyu
Hipertansiyon, ateroskleroz ve kalp yetmezliğinin patojenezinde telomer
disfonksiyonun önemli bir faktör olduğu son yıllarda giderek artan bir şekilde çok
çeşitli sayıdaki bilimsel araştırmaların konusunu oluşturmaktadır (127, 129, 160,
167). Benetos ve arkadaşları (15) son yılarda yaptıkları çalışmalarında insanda,
kronik hipertansiyon varlığında beyaz kan hücrelerinde saptanan daha kısa telomer
boylarının artan karotid arter aterosklerozu ile ilişkili olduğunu göstermişlerdir.
Diğer yandan yaşlanmanın önemli bir kardiyovasküler risk olduğu bilinmektedir.
Okuda ve arkadaşları (135) yaş ve ateroskleroz arasındaki ilişkiyi insan abdominal
aortasındaki telomer kısalmasını saptayarak araştırmışlardır. Bu çalışmanın sonuçları
telomer uzunluğunun aterosklerotik derece ile negatif korelasyon gösterdiğini ancak
bu ilişkinin yaş ile anlamlı olmadığını ortaya koymuştur.
Çalışmamızda balon hasarı uygulanan ve uygulanmayan ilyak arterlerde
yapılan denemelerde telomerik fragmanlar kantitatif olarak değerlendirilmiştir. Bu
değerlendirmelerde ortalama telomer uzunluğunun kontrol grubuna göre 2., 7., ve 14.

133
günlerde sırasıyla %64, %85 ve %63 oranında azalma gösterdiği görülmüştür
(Bakınız Bulgular; Tablo 3.3.). Benzer şekilde çalışmamızda farklı örnekler
kullanılarak elde edilen bir diğer analizde de deney grubunun ortalama telomer
uzunluğunun 7. ve 14. günler için sırasıyla %71 ve %77 oranında azalma
görülmüştür. Ancak bu azalmanın uygulamanın günleri açısından farklı olmadığı
anlaşılabilir; bu durum da kullanılan örneklerin farklı bireylere ait olması ile
ilişkilendirilebilir. Genel anlamda radyoaktif işaretli olmayan TRF sonuçlarının
telomer boy uzunluğunu, uygulanan işleme göre azalttığını her iki grup deneyde de
ifade etmek mümkün görünmektedir. Bununla birlikte bu deneylerden elde ettiğimiz
sonuçlar telomer boyundaki azalmayı balon uygulamasından sonraki (örneğin 2., 7.
ve 14. gün boyları gibi) süreler ile ilişkilendirmek açısından yeterli değildir.
Diğer yandan radyoaktif prob kullanılarak yapılan işaretlemenin radyoaktif
olmayan işaretlemeye göre çok daha duyarlı olduğu bilinmektedir. Gereç ve
Yöntem’de açıklandığı gibi radyoaktif olmayan işaretlemede telomerik fragmanı
hibridize edecek proba digoksigenin gibi non-radyoaktif bir işaretleyici bağlanır ve
bu işaretleyicinin substratı ile reaksiyona girmesi sonucu yaydığı ışımanın
(elektrokemiluminesans) röntgen filmini bozundurmasından yararlanılır. Bu yöntem
oldukça dolaylı bir yöntem olduğu için substrat konsantrasyonu, filmin reaksiyona
maruziyet süresi ve ortamdaki ışık gibi pek çok faktöre bağımlıdır. Radyoaktif
işaretlemede ise 32P işaretli prob kullanılır. Bu işaretleme ve izleyen aşamada
görüntünün fosfor görüntüleyici (phosphoimager) ile alınması çok daha etkin bir
yöntemdir.

134
Çalışmamızda radyoaktif işaretli olan ve olmayan prob kullanılarak elde edilen
sonuçları karşılaştırdığımızda duyarlık açısından çok önemli farklılık
gözlenmemektedir. Ortalama telomer uzunluğunun balon anjiyoplasti uygulamasının
2., 7., ve 14. günlerinde %53, %87 ve %74 olarak azaldığı saptanan radyoaktif
işaretli prob kullanılarak gerçekleştirilen deney sonuçları radyoaktif olmayan prob
kullanılarak elde edilen sonuçlarla uyum içindedir (175). Ancak radyoaktif işaretli
prob kullanılarak elde edilen sonuçlarda, telomer boyunun uygulamanın 2. ve 14.
günlerinde farklılık gösterdiği belirlenmektedir. Bununla birlikte genel anlamda
radyoaktif olmayan işaretleme ile gerçekleştirilen denemelerde kullanılan telomer
boyunu saptamaya yönelik kitin optimum koşulları sağladığı düşünülebilir.
Bu kapsamda metodolojik olarak aşağıda özetlenen teknik kavramlar ışığında
balon anjiyoplasti uygulanan ilyak arter örneklerinde telomeraz aktivitesinin
saptanamamasının uygulanan yöntem (ve/veya kullanılan kit) ile ilişkili
olamayabileceği düşünülmektedir. Bu düşüncemizin nedeni Şekil 3.11 de düşük ve
yüksek kopya sayılı internal standartlar kullanıldığında (Şekil 3.11, 2. ve 3.
uygulamalar) PCR işleminin beklenildiği gibi elde edilmesidir. Bütünüyle aynı
işlemlerden geçirilmiş örneklerimizle yapılan bu iki uygulama metodolojiye bağlı
kuşkuları giderecek niteliktedir. Ayrıca TRF deneyleri açısından aşağıda kısaca
özetlenen teknik ayrıntılar ışığında bu deneylerin uygulanan metodoloji ile ilgili bir
sınırlama yaratmayacağını da düşünmekteyiz:

135
i. Telomer uzunluğunun saptanması için DNA izolasyonu yapılan örnekler
Bulgular bölümünde Şekil 3.5.A’da verilmektedir. Bu şekil incelendiğinde belirli bir
miktar DNA’nın kuyucuklarda kaldığı görülmektedir. Bu genomik DNA
preparasyonlarında sıklıkla karşılaşılan bir durumdur. DNA moleküllerinin agaroz
jeli içerisindeki hareketliliği birçok başka faktörlerin yanı sıra moleküler ağırlık ve
jeldeki agaroz yüzdesi tarafından belirlendiği için yüksek molekül ağırlıklı genomik
DNA’nın bir bölümü kullandığımız agaroz jeli içinde ayrıştırılamamıştır. Ancak, bu
çalışmamızda ilgilendiğimiz DNA’nın genomik DNA olması nedeniyle bu görüntü
sonuçların yorumlanabilmesinde herhangi bir sorun oluşturmamıştır.
ii. Genomik DNA izolasyonlarının bilinen bir özelliği olarak bazı örnekler
“smear” olarak adlandırılan sürüntü görünümü göstermişlerdir (Bakınız Bulgular;
Şekil 3.5.A,). Özellikle 3 no.lu uygulamada gözlenen bu görüntünün uyguladığımız
voltajın yanı sıra, bu örneğin diğer örneklerden miktar olarak fazla olmasından
kaynaklandığını düşünmekteyiz. Gerçekten de DNA izolasyonları yapılan örneklerin
jele uygulama öncesi 260 nm’deki absorbansları incelediğinde bu görüntünün
beklenebileceği düşünülebilir. Bununla birlikte Tablo 1’deki örneklerin hepsinin
DNA konsantrasyonları ile jel üzerindeki görüntülerinin örtüşmediği de
gözlenmektedir. Bu durumun 260 nm UV aralığında spektrofotometrik ölçümlerin
DNA dışındaki aromatik yapıları da kapsamasından kaynaklanabileceği
düşünülebilir. Tüm bu teknik parametreler göz önüne alındığında telomerik
restriksiyonel frgmanların uzunluğunu belirlemeye yönelik deneylerimizin başarılı
olduğu görülmektedir.

136
Balon hasarı sonrası gelişen vasküler düz kas hücre proliferasyonunda telomer
devamlılığını aydınlatmayı amaçlayan bu bölümdeki bulgularımız telomer boyundaki
azalmanın balon anjiyoplasti uygulaması ile birlikte geliştiğini göstermektedir. 2. 7.
ve 14. günlerde gözlenen telomer boyundaki farklılık bireysel farklılıklardan
kaynaklanabileceği gibi daha önce genel bilgiler bölümünde ayrıntılı olarak söz
edilen telomerlerde oluşabilecek uç uca füzyon veya “Alternatif Telomer Uzaması”
(Alternative Lengthening of Telomeres=ALT) mekanizmalarından da
kaynaklanabileceği düşünülebilir (82, 136, 146).
Bölüm II: Tavşan Balon Anjiyoplasti Modelinde Telomer Boyu ve Oksidatif
Stres
Reaktif oksijen türlerinin hiperlipidemi, diabetes mellitus, hipertansiyon,
iskemik kalp hastalığı, kronik kalp yetmezliği ile ilişkilendirilen kardiyovasküler
disfonksiyonun ortaya çıkması ve progresyonundaki süreçte yer alan birçok olayda
rol oynadığı son yıllarda birçok çalışma ile ortaya konmuştur (1, 9, 18, 34, 99,182).
çalışmamızın amaçları kapsamında oksidatif stresin balon anjiyoplasti sonrası
vasküler düz kas hücrelerinde telomer devamlılığı üzerine etkisinin araştırılmasına
yönelik olarak balon anjiyoplasti hasarında BSO ile oluşturulan oksidatif stres
modeli uygulanmıştır. BSO ile endojen oksidatif stres oluşturma modeli hayvanlara
uzun süreli BSO verilerek glutatyon depolarının sürekli olarak tüketilmesine
dayanmaktadır (53, 198).

137
Oksidatif Stres Belirteçleri
Gluatatyonun sürekli olarak tüketilmesinin glutatyon tarafından
indirgenemeyen reaktif oksijen türlerinin açığa çıkmasına neden olduğu
bilinmektedir (198).
Çalışmamızda da kullanılan bu modelde 14 gün süre ile uygulanan BSO’nun
(75 mg/kg/gün) 14 gün süresince glutatyon metabolizmasına etkisi araştırılmıştır. Bu
amaçla deneyin başlangıcında (0. gün), ortasında (7. gün) ve son gününde (14. gün)
alınan kanda yapılan ölçümler BSO uygulamasının hem total glutatyonun, hem de
redükte glutatyon/okside glutatyon disülfit (GSH/GSSG) oranını zamana bağlı olarak
azalttığını göstermektedir. Bunun yanı sıra glutatyon peroksidaz enziminin düzeyinin
de azaldığı saptanmıştır. Bu sonuçlar aynı model kullanılarak yapılan önceki
deneylerimiz ile uyumludur (63). Ayrıca çalışmamızın sonuçları ile uyumlu bir
şekilde, söz konusu model kullanılarak yapılan çalışmalardan elde edilen sonuçlar da
BSO’nun karaciğer, böbrek, kalp ve dalak gibi dokularda da GSH düzeyini
azalttığını göstermektedir (53, 109, 198). Diğer yandan önceki çalışmamızda da
bildirdiğimiz gibi (64) BSO uygulaması ile azalan glutatyon düzeylerinin taurin ile
normalize edildiği bulgusu, taurinin antioksidan özelliği ile paralellik göstermektedir
(55, 86).
Morfometrik ve İmmünohistokimyasal İncelemeler
Daha önceki çalışmamız ile benzer şekilde (63) damar örneklerinde yapılan
morfometrik incelemeler BSO uygulanan grupta kontrol gruba göre intimal

138
kalınlaşmanın daha fazla olduğunu göstermektedir. Gerçekten de reaktif oksijen
türlerinin üretiminin hipertansiyon, ateroskleroz ve balon anjiyoplasti sonrası
restenozda vasküler düz kas hücrelerinde hipertrofi ve proliferasyonu ile ilişkili
olduğunu gösteren çalışmalar bulunmaktadır (57, 83, 152). Benzer şekilde konstriktif
vazoaktif bir madde olan anjiyotensin II’nin oluşturduğu hipertansiyona reaktif
oksijen türlerinin aracılık ettiği de gösterilmiştir (1). Bunun yanı sıra çeşitli büyüme
faktörlerinin oluşturduğu proliferasyonda da reaktif oksijen türlerinin rol oynadığı
gösterilmiştir (57,102). Bu kapsamda yukarıda bahsi geçen çalışmalarda olduğu gibi
bizim çalışmamızda da morfometri ve immünohistokimyasal incelemelere yönelik
olarak yapılan araştırmalardan elde edilen bulgular bu alandaki çok sayıdaki literatür
bilgileri ile uyum içindedir .
Telomer Fragmanlarının Boyu
Oksidatif stresin genel anlamda yaşlanma, endotel disfonksiyonu ve
aterosklerozda rolü olduğu son yıllarda yapılan çok sayıdaki çalışmada gösterilmiştir
(119, 143, 194). Bu açıdan, uzun yıllardan beri vasküler sistem ile ilişkili
hastalıkların başlamasının ve ilerlemesinin yaş ile ilişkili olabileceği düşünülmüştür.
Benzer bir yaklaşımla, yaşlanmanın intraselüler reaktif oksijen türleri artışı ile
telomeraz ters transkriptaz aktivitesinin kaybolması ile birlikte geliştiği de
bilinmektedir (76). Bu kapsamda çalışmamızda oksidatif stres, telomer boyu ve
antioksidan tedavi arasındaki olası ilişki tavşan ilyak arteri balon anjiyoplasti
modelinde araştırılmıştır.

139
Alınan damar örneklerinde her gruptaki telomer boylarının kontrol damara
göre yüzdelenmesi (34) ile elde ettiğimiz sonuçlar, BSO uygulamasının telomer
boyunu kısalttığını ortaya koymaktadır (176). Kurz ve arkadaşları tarafından insan
umblikal ven endotel hücre kültüründe (HUVEC) yapılan çalışmada normal durumda
telomer aşınmasının sabit (55 bç/PD) (PD-population doubling[hücre sayısının iki
katına çıkması]) olduğu ancak bu hücreler BSO ile inkübe edildiğinde telomerik
fragmanların aşınma hızının 110 bç/PD’ye yükseldiği gösterilmiştir (103). Diğer
yandan, Von Zglicki ve arkadaşları (192) tarafından insan fibroblastlarında yapılan
bir çalışmada ise oksidatif streste hafif bir artışın (%40) yaşlanma sürecini
hızlandırdığı ve hücre bölünme hızını 3 katına çıkardığı bildirilmiştir. Araştırmacılar
fibroblast hücrelerde ortaya çıkan erken yaşlanmanın telomerlere olan oksidatif
hasardan kaynaklandığını ileri sürmüşlerdir. Aynı çalışma grubundan Richter ve Von
Zglinicki (156) araştırmalarını insan ve koyundan elde edilen fibroblastlarda
sürdürmüşler ve tür farkı gözetmeden oksidatif stres ile telomer kısalma hızı arasında
pozitif bir korelasyon olduğunu bildirmişlerdir.
Diğer taraftan çalışmamızdan elde ettiğimiz diğer bir bulgu BSO alan grupta
daha uzun telomer fragmanlarına da rastlanmasıdır. Yapılan çalışmalarda hem BSO
ile glutatyon depolarının tüketilmesinin hem de oluşan serbest oksijen radikallerinin
DNA hasarına neden olabildiği gösterilmiştir (45, 83, 147). Özellikle guaninden
zengin telomer tekrarlarının reaktif oksijen türlerinin oluşturduğu DNA hasarına
hassas olduğu bilinmektedir. Çalışmamızdan elde ettiğimiz bu sonuç telomere
yönelik olarak kullanılan probun nonspesifik bağlanması olabileceği gibi BSO’nun
DNA hasarı sonucu oluşturabileceği DNA kırıkları da olabilir. Kurz ve arkadaşları,
tarafından HUVEC’te yapılan çalışmada da benzer bulgular rapor edilmektedir

140
(103). Telomerler TRF ile analiz edildiğinde ortalama telomer boyu oksidatif stres
etkisi ile prematüre kısalmaya uğrarken telomer fragmanlarının bifazikleştiği
gözlemlenmiştir. Kurz ve arkadaşları söz konusu çalışmada telomeraz biyoaktivitesi
de bulunamadığı için bu durumun alternatif telomer uzamasından (ALT)
kaynaklanabileceğini ileri sürmüşler ve immünofloresan mikroskobi ile PML
cisimciklerinin varlığını gösterilmişlerdir. Bir diğer çalışma Hensen ve arkadaşları
tarafından bildirilmiştir (82). Bu araştırma grubu ALT yolağında DNA hasar onarım
mekanizmasının devreye girdiğini saptamış ve telomerler de hasara yatkın bölgeler
olduğu için rekombinasyona uğrayarak alternatif telomer uzamasının gerçekleştiğini
ileri sürmüşlerdir. Genel olarak ALT mekanizmasının gerçekleşmesi için PML
cisimciklerinin TRF1 ve TRF2 ile asosiye halde bulunması gerektiği
düşünülmektedir (143). Diğer taraftan telomer tekrar kalıbının tüm omurgalılarda
korunmuş olduğu bilinmekle birlikte tür farklılığı da göz önünde bulundurulmalıdır
(42, 54, 123, 204). Çalışmamızda uzun telomer fragmanlarının varlığı ALT ve/veya
DNA hasar onarım mekanizmalarının rolünün olabileceğini düşündürmekle birlikte
çalışmanın amacı kapsamında bu yönde deneyler tasarlanmamıştır. Buradan
hareketle de uzun telomer fragmanları ve ALT ilişkisinin kullandığımız modelde
araştırılması için ayrıntılı deneylerin yapılmasının gerektiği düşünülmektedir.
Çalışmamızda bir antioksidan olan taurinin BSO uygulaması ile oluşan telomer
kısalmasını normalize ettiği gösterilmiştir. Taurinin çeşitli deney koşullarında, farklı
türlerde bir antioksidan olarak işlev gördüğüne ilişkin çok sayıda araştırma
bulunmaktadır. Bu açıdan bakıldığında taurinin, BSO ile oluşan telomer kısalmasını
normalize ettiği bulgusu ilk kez bu çalışmada gösterilmiştir ve antioksidan

141
mekanizmasının telomer kısalmasını yavaşlatıcı etkisini bir kez daha gündeme
getirmektedir (112).
Sonuç olarak çalışmamızda elde ettiğimiz bulguları kısaca yeniden değerlendirirsek;
i. Histopatoljik bulgularımız tavşan ilyak arterine uygulanan balon
anjiyoplastinin intimal kalınlaşmaya neden olduğunu ve söz konusu kalınlaşmanın
intimaya göç eden ve prolifere olan düz kas hücrelerinden kaynaklandığını
göstermektedir. BSO verilmesi intimal kalınlaşmayı artırmaktadır. Taurin ise bu
intimal kalınlaşmayı geri döndürmektedir.
ii. Ekspresyon çalışmalarımız ilyak artere balon anjiyoplasti uygulaması
sonucu oluşan damar düz kas hücre proliferasyonu ile ilişkili olarak telomeraz
biyoaktivitesi saptanmadığını göstermektedir.
iii. Bulgularımız telomer boyundaki azalmanın balon anjiyoplasti uygulaması
ile birlikte geliştiğini göstermektedir. Ancak azalmanın fizyolojik telomer kısalması
ile ilişkili olabileceği de göz ardı edilmemelidir.
iv. BSO uygulaması ortalama telomer boyunun azalmasına neden olmuştur.
Bir antioksidan olan taurin BSO’nun ortalama telomer boyunda oluşturduğu
azalmayı kısmen geri döndürmüştür.

142
5. SONUÇ VE ÖNERİLER
Bu sonuçlar kapsamında, ileriye dönük hedefler açısından, balon anjiyoplasti
sonrası endotel ve/veya düz kas hücre kültürü ile elde edilebilecek sonuçların
özellikle yaşlanma süreci de gözönüne alındığında restenoz ve aterosklerotik
hastalıkların oluşum mekanizmalarının aydınlatılmasına büyük katkı sağlayacağını
düşünmekteyiz. Deneysel olarak uygulanan tavşan ilyak arter balon anjiyoplasti
modelinin insanda koroner ater perkütanöz transluminal anjiyoplasi girişimi ile yakın
benzerlik gösterdiği genel olarak kabul edilmektedir. Bu açıdan hareketle,
çalışmamızdan elde edilen sonuçların klinik uygulamaya dönük yaklaşımlara destek
sağlayabileceğini düşünmekteyiz. Diğer yandan sonuçlarımız aterosklerotik
hastalıklarda antioksidan kullanımınn önemini bir kez daha gündeme getirebilir
niteliktedir.

143
ÖZET
Düz kas hücre proliferasyon ve/veya migrasyonuna bağlı olarak gelişen intimal
hiperplazinin ateroskleroz patojenezi ile balon anjiyoplasti sonrası oluşan restenozdan
sorumlu olduğu bilinmektedir. Ökaryotik hücrelerde, kromozomların uçlarında bulunan
telomerler kromozomal bütünlüğünü korumak üzere etkinlik gösterirler. Söz konusu
telomerler kromozomal replikasyon sürecinin sürdürülmesinde ve yine hücresel
yaşlanma sürecinin zamanlanmasında anahtar bir rol oynarlar. Telomer devamlılığı
telomeraz enziminin aktivitesi ile sürdürülür. Yüksek oranda proliferasyon gösteren
hücrelerde telomeraz aktivitesinin arttığı gösterilmiştir. Bunun yanı sıra, damar düz kas
hücrelerinde telomeraz aktivitesinin arttığı ve telomeraz inhibisyonunun ise damar düz
kas hücre büyümesini engellediği de gösterilmiştir.
Bu açıdan bakıldığında, ateroskleroz ve balon anjiyoplasti sonrası gelişen
restenozun patojenezindeki telomeraz aktivitesi ve telomer devamlılığının rolü tam
olarak saptanamamıştır. Diğer taraftan hücrelerdeki telomer boyu yaşlanma süreci
ilişkilendirilmiştir. Oksidatif stresin de yaşlanma sürecine katkısı birçok çalışmada
ortaya konmuştur. Bu çalışmada balon aanjiyoplasti yöntemi ile balon hasarı oluşturulan
tavşan iliyak arterlerde telomer devamlılığının araştırılması ve oksidatif stresin bu
süreçteki etkisinin araştırılması amaçlanmıştır. Ayrıca olası bir etkinin antioksidanlar ile
modüle olup olmadığı da araştıma kapsamındadır.

144
Bu amaçla çalışmanın ilk bölümünde tavşan ilyak arterine balon anjiyoplasti
uygulanmış ve daha sonra hasar gören ve görmeyen ilyak arterler izole edilerek
telomeraz aktivitesi ve telomer devamlılığı araştırılmıştır. Çalışmanın ikinci bölümüne
ise balon anjiyoplasti uygulanan tavşanlarda BSO ile oksidaitif stres oluşturulmuş ve
telomer devamlılığına etkisi araştırılmıştır. Bunun yanı sıra her iki bölümde de ilyak
arter segmentlerinde intimal hiperplaziyi saptamak üzere morfometrik ve
immünohistokimyasal analizler gerçekleştirilmiştir.
Çalışmamızın sonuçları ilyak artere balon anjiyoplasti uygulamasının, hasar
görmüş arterlerde damar düz kas hücrelerinde proliferasyona neden olarak intimal
hiperplaziye yol açtığını göstermiştir. Ayrıca, hasar gören ilyak arter segmentlerinde
telomeraz aktivitesi saptanmamıştır. Çalışmamızın sonuçları, bunların yanı sıra, hasar
görmüş ilyak arter damar segmentlerinde telomerik restriksiyonel fragmanların
kısaldığına işaret etmektedir. BSO uygulaması ortalama telomer boyunun azalmasına
neden olmuştur. Bir antioksidan olan taurin BSO’nun ortalama telomer boyunda
oluşturduğu azalmayı kısmen geri döndürmüştür.

145
ABSTRACT
It has been reported that intimal hyperplasia, due to smooth muscle cell
proliferation and migration, is responsible for the pathogenesis of atherosclerosis and
restenosis occuring after balloon angioplasty. In eucaryotic cells, the telomer is found at
the end of linear chromosomes and acts to preserve chromosomal integrity. Telomeres
play a key role in mediating the processes of chromosomal replication and in regulating
the timing of cellular aging processes. Telomeres are maintained by the activity of the
enzyme telomerase. Telomerase activity is found to be increased in highly proliferative
cells. Besides, it has been shown that telomerase activity is increased in vascular smooth
muscle cells whereas inhibition of telomerase diminished growth of vascular smooth
muscle cells.
In this context, the role of telomere maintenance and telomerase activity in the
pathogenesis of atherosclerosis and restenosis occuring after balloon angioplasty has not
been fully determined. On the other hand, telomere length is associated with cellular
aging. The contribution of oxidative stress is also stated in many studies. This study
investigates the telomere length regulation of vascular smooth muscle cells after balloon
injury in the balloon angioplasty model and to investigate the effects of oxidative stress
in this phenomenon. The study was extended to investigate the modulation of the
possible effects of oxidative stress by antioxidants.
For this purpose, in the first part of the study, balloon angioplasty was performed in the
iliac artery of the rabbits and thereafter telomerase activity and telomer maintenance
were investigated in the isolated injured and non-injured arteries. In the second part of
the study, oxidative stress was induced by BSO and telomere maintenance was

146
investigated. In addition, morphometric and immunohistochemical analyses were
conducted in order to determine intimal hyperplasia in the arterial segments for both
parts of the study.
Our results demonstrated that balloon angioplasty in iliac arteries resulted in
intimal hyperplasia due to proliferation of the smooth muscle cells. In addition,
telomerase activity has not been detected in injured arteries. Small size telomeric
restrictional fragments were evident in injured arteries. BSO induced even shorter
telomeric fragments. In addition, an antioxidant agent, taurin,partially reversed this
effect.

147
KAYNAKLAR
1. Agarwal R., Campbell R.C., Warnock D.G. (2004). Oxidative Stress in Hypertension and Chronic Kidney Disease: Role of Angiotensin II, Semin. Nephrol., 24(2):101-14
2. Aisner D.L., Wright W.E., Shay J.W. (2002). Telomerase Regulation: Not Just Flipping the Switch, Curr. Opin. Genet. Dev., 12: 80-85
3. Alexander R.W., Dzau V.J. (2000). Vascular Biology: The Past 50 Years, Circulation,102:IV-112-IV-116
4. Altan N., Dinçel A.S., Koca C. (2006). Diabetes Mellitus and Oxidative Stress, Turk. J. Biochem., 31(2): 51-56
5. Anderson M.E. (1998). Gluthathione: An Overview of Bisynthesis and Modulation, Chem-Biol. Interact., 111-112: 1-14
6. Aoyagi M., Yamamoto M., Azuma H., Nagashima G., Niimi Y., Hirakawa K., Yamamoto K. (1997). Expression of p53 Protein and p53 Gene Transcripts in Rabbit Carotid Arteries After Balloon Denudation, Histochem. Cell. Biol., 107: 365-70
7. Arthur Y.F., Yin Z.L., Young H.M., Dusting G.J. (1997). Induction of Nitric Oxide Synthase in the Neointima Induced by a Periarterial Collar in Rabbits, Arterioscler Thromb Vasc Biol,; 17:737-40
8. Assadnia S., Rapp J.P., Nestor A.L., Pringle T., Cerilli G.J., Gunning W.T., Webb T.H., Kligman M., Allison D.C. (1999). Strain Differences in Neointimal Hyperplasia in the Rat, Circ. Res, 84: 1252–7
9. Azevedo L.C., Pedro M.A., Souza L.C., de Souza H.P., Janiszewski M., da Luz P.L., Laurindo F.R. (2000). Oxidative Stress As a Signaling Mechanism of The Vascular Response To Injury: The Redox Hypothesis of Restenosis, Cardiovasc. Res., 47: 436-45
10. Balasubramanyam M., Adaikalakoteswari A., Finny-Monickaraj S., Mohan V. (2007). Telomere Shortening and Metabolic/Vascular Diseases, Indian J. Med. Res., 125: 441-50
11. Baumgartner H.R. (1963). Eine Neue Methode Zur Erzeungung Vor Thromben Durch Gesilete Uberdehnung der Gefasswand, Z. Ges. Exp. Med.,137: 227
12. Bayes-Genis A., Campbell J.H., Carlson P.J., Holes D.R., Schwartz R.S. (2002). Macrophages, Myofibroblasts and Neointimal Hyperplasia After Coronary Artery Injury and Repair, Atherosclerosis, 163: 89-98

148
13. Bayes-Genis A., Kantor, B., Keelan, P.C., Altman, J.D., Lubbe, D.F., Kang, J.H., Schwartz, R.S. (2000). Restenosis and Hyperplasia: Animal Models, Cur.r Interv. Cardiol. Rep., 2(4): 303-8
14. Belair C.D., Yeager, T.R., Lopez, P.M., Reznikofff, C.A. (1997). Telomerase Activity:A Biomarker of Cell Proliferation, not Malignant Transformation, Proc. Natl. Acad. Sci, 94: 13677-82
15. Benetos A., Gardner J.P., Zureik M., Labat C., Xiaobin L., Adamopoulos C., Temmar M., Bean K.E., Thomas F., Aviv A. (2004). Short Telomerase are Associated with Increased Carotid Atherosclerosis in Hypertensive Subjects, Hypertension, 43: 182-5
16. Blackburn E.H. (1990). Telomeres: Structure and Synthesis, J. Biol. Chem., 265(11): 5919-21
17. Blackburn E.H. (1991). Structure and Function of Telomeres, Nature, 350(6319): 569-73
18. Blankenberg S., Rupprecht H.J., Bickel C., Torzewski M., Hafner G., Tiret L., Smieja M.,Cambien F., Meyer J., Lackner K.J. (2003). Gluthathione Peroxidase 1 Activity and Cardiovascular Events in Patients with Coronary Artery Disease, N. Engl. J. Med., 349:1605-13
19. Blasco M.A., Lee H.W., Hande M.P., Samper E., Lansdorp P.M., DePinho R.A., Greider C.W. (1997). Telomere Shortening and Tumor Formation by Mouse Cells Lacking Telomerase RNA, Cell, 91(1): 25-34
20. Bonin L.R., Madden K., Shera K., Ihle J., Matthews C., Aziz S. (1999). Perez-Reyes N., McDougall J.K., Conroy S.C. Generation and Characterization of Human Smooth Muscle Cell Lines Derived From Atherosclerotic Plaque, Arterioscler. Thromb. Vasc. Biol., 19: 575-87
21. Bouckenooghe T., Remacle C., Reusens B. (2006). Is Taurine a Functional Nutrient?. Curr. Opin. Clin. Nutr. Metab. Care, 9: 728-33
22. Bradford M.M. (1976). A Rapid and Sensitive Method for the Quantitation of Microgram quantities of Protein Utilizing the Principle of Protein-dye Binding, Anal. Biochem., 72: 248-54
23. Braun-Dullaeus R.C, Mann M.J., Sedding D.G., Sherwood S.W., von der Leyen H.E., Dzau V.J. (2004). Cell Cycle-Dependent Regulation of Smooth Muscle Cell Activation, Arterioscler. Tromb. Vasc. Biol., 24: 845-50
24. Braun-Dullaeus R.C. (2001). Cell Cycle Protein Expression in Vascular Smooth Muscle Cells in Vitro and in Vivo is Regulated Through Phosphatidylinositol 3-kinase and Mammalian Target of Rapamycin, Arterioscler. Thromb. Vasc. Biol., 21: 1152-58

149
25. Bravo R., Macdonald-Bravo H. (1987). Existence of two populations of cyclin/proliferating cell nuclear antigen during the cell cycle: Association with DNA Replication Sites, J. Cell Biol., 105: 1549-54
26. Breitschopf K., Zeiher A.M., Dimmeler S. (2001). Pro-atherogenic Factors Induce Telomerase Inactivation in Endothelial Cells Through an Akt-dependent Mechanism, FEBS Lett, 493: 21-25
27. Broilette S., Moore J.S., McMahon A., Thompson J.R., Ford I., Shepherd J., Packard C. J., Samani N.J. (2007). Telomere Length, Risk of Coronary Heart Disease and Statin Treatment in the West of Scotland Primary Prevention Study: A Nested Case-control Study, Lancet, 369: 107-14
28. Bruijins R.H.J., Bult H. (2001). Effects of Local Cytochalasin D Delivery on Smooth Muscle Cell Migration and on Collar-Induced Intimal Hyperplasia in the Rabbit Carotid Artery, Br. J. Pharmacol., 134: 473-83
29. Bult H. (2000). Restenosis: A Challenge for Pharmacology, TIPS, 21: 274-8
30. Burns J.A. Butler J.C.,Moran J., Whitesides G.M. (1991). Selective Reduction of Disulfides by Tris(2-carboxyethyl)phospine, J. Org. Chem., 56: 2648-50
31. Cai H., Harrison D.G. (2000). Endothelial Dysfunction in Cardiovascular Diseases: The Role of Oxidant Stress, Circ. Res., 87: 840-44
32. Calvin H.L., Medvedovsky C., Worgul B.V. (1986). Near-total Glutathione Depletion and Age-specific Cataracts Induced by Buthionine Sulfoximine in Mice, Science, 233: 553-55
33. Cao Y., Li H., Mu F-T., Ebisui O., Funder J.W., Liu J-P. (2002). Telomerase Activation Causes Smooth Muscle Cell Prolifaration in Genetic Hypertension, FASEB J., 16: 96-8
34. Cattan V., Mercier N., Gardner J.P., Regnault V., Labat C., Maki-Jouppila J., Nzietchueng R., Benetos A., Kimura M., Aviv A., Lacolley P. (2008). Chronic Oxidative Stress Induces a Tissue-specific Reduction in Telomere Length in CAST/Ei Mice, Free Radical Biol. Med., doi:10.1016/j.freeradbiomed.2008.01.007
35. Chan A.W., Chew D.P., Lincoff A.M. (2001). Update on Pharmacology For Restenosis, Curr. Int. Cardiovasc. Rep, 3: 149-55
36. Chang E., Harley C.B. (1995). Telomere Length and Replicative Aging in Human Vascular Tissues, Proc. Natl. Acad. Sci. U.S.A., 92: 11190-94
37. Chatziantoniou V.D. (2001). Telomerase: Biological Function and Potential Role in Cancer Management, Pathol. Oncol. Res., 7(3): 161-70

150
38. Cherif H., Reusens B., Dahri S. (1996). Stimulatory Effects of Taurine on Insulin Secretion by Fetal Rat Islets Cultured in vitro, J. Endocrinol., 151: 501-506
39. Christen T., Verin V., Bochaton-Piallat M-L., Popowski Y., Ramaekers F., Debruyne P., Comenzind E., van Eys G., Gabbiani G. (2001). Mechanisms of Neointima Formation and Remodeling in the Porcine Coronary Artery, Circulation, 103: 882-8
40. Cong Y.S., Wright W.E., Shay J.W. (2002). Human Telomerase and Its Regulation, Microbiol. Mol. Biol. R., 66(3):407-25
41. Corseaux D., Le Tourneau T., Six I., Ezekowitz MD., Mc Fedden E.P., Meurice T., Asseman P., Bauters C., Jude B. (1998). Enhanced Monocyte Tissue Factor Response After Experimental Balloon Angioplasty in Hypercholesterolemic Rabbit: Inhibition with Dietary L-Arginine, Circulation, 98: 1776-82
42. De Lange T. (2002). Protection of Mammalian Telomeres, Oncogene, 21(4): 532-40
43. De Meyer G.R., Bult H. (1997). Mechanisms of Neointima Formation--Lessons From Experimental Models, Vasc. Med, 2:179-89
44. Diendorck J.H. van, Wijsman J.H., Keijser R., Velde C.J.H. van de, Cornellise C.J. (1991). Cell cycle-related staining patterns of anti-proliferating cell nuclear antigen monoclonal antibodies. Comparison with BrdU labeling and Ki-67 staining, Am.J. Pathol., 138: 1165-72
45. Duan J., Duan J., Zhang Z., Tong T. (2005). Irreversible Cellular Senescence Induced by Prolonged Exposure to H2O2 involves DNA-damage-and-repair Genes and Telomere Shortening, Int. J. Biochem. Cell. Biol., 37: 1407-1420
46. Durand E., Mallat Z., Addad F., Vilde F., Desnos M., Guérot C., Tedgui A., Lafont A. (2002). Time Course of Apoptosis and Cell Proliferation and Their Relationship to Arterial Remodeling and Restenosis After Angioplasty in an Atherosclerotic Rabbit Model. J Am. Coll. Cardiol., 39:1680-5
47. Dzau V.J., Braun-Dullaeus R.C., Sedding D.G. (2002). Vascular Proliferation and Atherosclerosis: New Perspectives and Therapeutic Strategies, Nat. Med., 8(11): 1249-56
48. Dzau V.J., Gnecchi M.,Pachori A.S., Morello F., Melo L.G. (2005). Therapeutic Potential of Andothelial Progenitor Cells in Cardiovascular Diseases, Hypertension, 46: 7-18
49. Endl E., Gerdes J. (2000). The Ki-67 Protein: Fascinating Froms and Unknown Function, Exper. Cell. Res., 257: 231-7

151
50. Engin A., Altan N. (2000). Effects of Obstructive Jaundice on the Antioxidative Capacity of Human Red Blood Cells, Hematologia, 30(2): 91-96
51. Engin A., Altan N., Isik E. (2005). Erythrocyte Gluthathione Levels in Lithium-induced Hypothyroidism, Drug Dev. Res., 6(1):35-40
52. Ferns G.A.A., Avades T.Y. (2000). The Mechanisms of Coronary Restenosis: Insights from Experimental Models, Cardiovasc. Pathol., 81: 63-88
53. Ford R.J., Graham D.A., Denniss S.G., Quadrilatero J., Rush J.W.E. (2006). Glutathione Depletion in vivo Enhances Contraction and Attenuates Endothelium-dependent Relaxation of Isolated Rat Aorta, Free Radic. Biol. Med., 40:670-78
54. Fradiani P.A., Ascenzioni F., Lavitrano M., Donini P. (2004). Telomeres and Telomerase Activity in Pig Tissues, Biochimie, 86: 7-12
55. Franconi F., Di Leo M.A., Bennardini F., Ghirlanda G. (2004). Is Taurine Beneficial in Reducing Risk Factors for Diabetes Mellitus?, Neurochem. Res., 29(1):143-50
56. Furumoto K., Inoue E., Nagao N., Hiyama E., Miwa N. (1998). Age-dependent Telomere Shortening is Slowed Down by Enrichment of Intracellular Vitamin C via Suppression of Oxidative Stress, Life Sci., 3: 935-48
57. Fuster J.J., Andres V. (2006). Telomere Biology and Cardiovascular Disease, Circ. Res., 99: 1167-80
58. Gabeler E.E., van Hillegersberg R., van Eps R.G.S., Sluiter W. Gussenhoven E.J., Mulder P., van Urk H. (2002). A Comparison of Balloon Injury Models of Endovascular Lesions in Rat, BMC Cardiovasc. Disorders., 2:16-27
59. Gallo R. (1999). Inhibition of Intimal Thickening After Balloon Angioplasty in Porcine Coronary Arteries by Targeting Regulators of the Cell Cycle, Circulation, 99: 2164-70
60. Ganea E., Harding J.J. (2006). Glutathione-Related Enzymes and The Eye, Curr. Eye. Res., 31: 1-11
61. Goldschmidt-Clermont P.J., Creager M.A., Losordo D.W., Lam G.K.W., Wassef M., Dzau V.J. (2005). Atherosclerosis 2005: Recent Discoveries and Novel Hypothesis, Circulation, 112: 3348–53
62. Gorenne I., Kavurma M., Scott M., Scott S., Bennett M. (2006). Vascular Smooth Muscle Cell Senescence in Atherosclerosis, Cardiovasc. Res., 72: 9-17

152
63. Gökçe, G., Özşarlak-Sözer, G., Oran, İ., Oktay, G., Özkal, S., Kerry, Z. Oksidatif stres varlığında balon hasarı oluşturulan tavşanlarda taurinin intimal hiperplazi üzerine etkileri.Türk Farmakoloji Derneği, 19. Ulusal Farmakooji Kongresi, 24-27 Ekim 2007, Trabzon (Poster sunumu).
64. Gökçe, G., Özşarlak-Sözer, G., Oran, İ., Oktay, G., Özkal, S., Kerry, Z. Oksidatif stres varlığında izole tavşan damarlarında endotel kaynaklı gevşeme yanıtlarının taurin ile düzenlenmesi. .Türk Farmakoloji Derneği, 19. Ulusal Farmakooji Kongresi, 24-27 Ekim 2007, Trabzon (Poster sunumu).
65. Greider C.W., Blackburn E.H. (1985). Identification of a Specific Telomere Terminal Tansferase Activity in Tetrahymena Extracts, Cell, 43(2 Pt 1): 405-13
66. Greider C.W., Blackburn E.H. (1989)., A Telomeric Sequence in the RNA of Tetrahymena Telomerase Required for Telomere Repeat Synthesis, Nature, 337(6205):331-37
67. Greider C.W. (1996). Telomere Length Regulation, Annu. Rev. Biochem., 65: 337-65
68. Greider C.W. (1999). Telomerase Activation: One Step on the Road to Cancer?, Trends Genet., 15(3): 109-12
69. Griffith O.W., Meister A. (1978). Differential Inhibition of Glutamine and γ-glutamylcysteine Synthetases by α-alkyl Analogs of Methionine Sulfoximine That Induce Convulsions, J. Biol. Chem., 253: 2333-38
70. Griffith O.W., meister A. (1979). Potent and Specific Inhibition of glutathione Synthesis by Buthionine Sulfoximine (S-n-butyl homocysteine sulfoximine), J. Biol. Chem. 254: 7558-60
71. Griffith O.W. (1980). Determination of Glutathione and Glutathione DisulfideUsing Glutathione Reductase and 2-Vinylpiridine, Analytical Chemistry, 106(1):207-12
72. Griffith O.W., Meister A. (1985). Origin and turnover of Mitochondrial Glutathione, Proc. Natl. Acad. Sci. U.S.A., 82: 4668-72
73. Griffith J.D., Cameau L., Rosenfield L., Stansel R.M., Bianchi A., Moss H., de Lange T. (1999). Mammalian Telomeres End in a Large Duplex Loop, Cell, 97(4): 503-14
74. Gruentzig A.R. (1978). Transluminal Dilation of Coronary Artery Stenosis, Lancet, I: 263
75. Haendeler J., Hoffmann J., Rahman S., Zeiher A.M., Dimmeler S. (2003). Regulation of Telomerase Activity and Anti-apoptotic Function by Protein-protein Interaction and Phosphorylation, FEBS Lett., 536: 180-86

153
76. Haendeler J., Hoffmann J., Diehl J.F., Vasa M., Spyridopoulos, Zeiher A.M., Dimmeler S. (2004). Antioxidants Inhibit Nuclear Export of Telomerase Reverse Transcriptase and Delay Replicative Senescence of Endothelial Cells, Circ. Res., 94: 768-75
77. Harley C.B., Futcher A.B., Greider C.W. (1990). Telomeres Shorten During Aging of Human Fibroblasts, Nature, 345: 458-60
78. Harley C.B, Vaziri H., Counter C.M., Allsopp R.C. (1992). The Telomere Hypothesis of Cellular Aging, Exp. Gerontol., 27: 375-82
79. Harrington L. (2003). Biomedical Aspects of Telomerase Function, Cancer Letters, 194: 139–54
80. Hasanoglu E., Altan N., Sindel P., Ongun C.O., Bali M., Altintas E. (1994). The Relationship Between Erythrocyte Superoxide Dismutase Activity and Plasma Levels of Some Trace Elements (Al, Cu, Zn) of Dialysis Patients, General Pharmacology, 25(1): 107-10
81. Hayflick L. (1965). The Limited in vitro Life Time of Human Diploid Cell Strains, Exp. Cell Res., 37: 614-36
82. Henson J.D., Neumann A.A., Yeager T.R., Reddel R.R. (2002). Alternative Lengthening of Telomeres in Mammalian Cells, Oncogene, 21: 598-610
83. Herbert K.E., Mistry Y., Hastings R., Poolman T., Niklason L., Williams B. (2008). Angiotensin II-Mediated Oxidative DNA Damage Accelerates Cellular Senescence in Cultured Human Vascular Smooth Muscle Cells via Telomere-Depent and Independent Pathways, Circ. Res. 102: 201-208
84. Holt S.E., Wright W.E., Shay J.W. (1997). Multiple Pathways for the Regulation of Telomerase Activity, Eur. J. Cancer, 33(5): 761-66
85. Houben J.M.J., Moonen H.J.J., van Schooten F.J., Hageman G.J. (2008) Telomere Length Assesment: Biomarker of Chronic Oxidative Stress?, Free Radical Bio Med, 44:235-46
86. Huxtable R.J. (1992). Physiological Actions of Taurine, Physiol. Rev., 72: 101-163
87. Inoue S., Koyama H., Miyata T., Shigematsu H. (2002). Cell Replication Induces in-Stent Lesion Growth in Rabbit Carotid Artery with Preexisting Intimal Hyperplasia., Atherosclerosis, 162: 345–53
88. Jacobsen J.G., Smith L.H. (1968). Biochemistry and Physiology of Taurine and Taurine Derivatives, Physiol. Rev., 48: 424-511
89. Janero D.R, Ewing J.F. (2000). Nitric Oxide and Postangioplasty Restenosis: Pathological Correlates and Therapeutic Potential, Free Radical Biol & Med, 29(12): 1199–221

154
90. John H.I.P., Fuster V., Badimon L., Badimon J., Taubman M.B., Chesebro J.H. (1990). Syndromes of Accelerated Atherosclerosis: Role of Vascular Injury and Smooth Muscle Cell Proliferation, 15: 1667-87
91. Kadar A., Glasz T. (2001). Development of Atherosclerosis and Plaque Biology, J. Cardiovasc. Surg, 9(2): 109-21
92. Kamenz J., Seibold W., Wohlfrom M., Hanke S., Heise N., Lenz C., Hanke H. (2000). Incidence of Intimal Proliferation and Apoptosis Following Balloon Angioplasty in an Atherosclerotic Rabbit Model, Cardiovasc. Res., 45: 766-76
93. Kannan S., Tahara H., Yokozaki H., Matthew B., Nalinakumari K.R. Nair M.K., Tahara E. (1997). Telomerase Activity in Premalignant and Malignant Lesions of Human Oral Mucosa, Cancer Epidemiol Biomarkers Prev, 6(6): 413-20
94. Kashino G., Kodama S., Nakayama Y., Suzuki K., Fukase K., Goto M., Watanabe M. (2003). Relief of Oxidative Stress by Acorbic Acid Delays Cellular Senescence of Normal and Werner Syndrome Fibroblast Cells, Free Radical Biol. Med., 35: 438-43
95. Kawamoto R., Yamashita A., Nishihira K., Furukoji, E., Hatakeyama K., Ishikawa T., Imamura T., Itabe, H., Eto T., Asada Y. (2006). Different Inflammatory Response and Oxidative Stres in Neointimal Hyperplasia After Balloon Angioplasty and Stent Implantation in Cholesterol-fed Rabbits, Pathol. Res. Pract. 202: 447–56
96. Keith, W.N., Bilsland A., Evans T.R.J., and Glasspool R.M. (2002). Telomerase-directed molecular therapeutics, Expert Reviews in Molecular Medicine,; http://www.expertreviews.org/02004507h.htm
97. Kim NW, Piatyszek MA, Prowse KR, Harley CB, West MD, Ho PLC, Coviello GM, Wright WE, Weinrich SL, Shay JW. (1994). Specific Association of Human Telomerase Activity with Immortal Cells and Cancer, Science, 266: 2011-5
98. Kiyokawa H. (2005). Senescence and Cell Cycle Control, Results Probl. Cell Differ., 42: 257-70
99. Knopfholz, Précoma D.B., Brofman P.R.S., Bier G.E., Kuenzer C. Da Silva R.F., Silva A.P.K., Lemos A.C., Silva, L Lemos A.C.,de Olivera Silva P., Dompsin de Mores J. (2006). The Effect of L-Arginine on Neointimal Proliferation and Artery Remodeling on an Iliac Artery Lesion Caused by a Balloon Catheter in Hypercholesterolemic Rabbits, Arq. Bras. Cardiol., 87: 472-6
100. Kojima H., Miyazaki H., Shiwa M., Tanaka Y., Moriyama H. (2001). Molecular Biological Diagnosis of Congenital and Acquired Cholesteatoma

155
on the Basis of Differences in Telomere Length, Laryngoscope, 111(5): 867-73
101. Kumral A., Ozer E., Yilmaz O., Akhisarlioglu M., Gokmen N., Duman N., Ulukus C., Genc S., Ozkan H. (2003). Neuroprotective effect of erythropoietin on hypoxic-ischemic injury in neonatal rats, Biol. Neonate, 83(3): 224-8
102. Kurz D.J., Hong Y., Trivier E., Huang H., Decary S., Zang G.H., Lüscher T.F., Erusalimsky J.D. (2003). Fibroblast Growth Factor-2, But Not Vascular Endothelial Growth Factor, Upregulates Telomerase Activity in Human Endothelial Cells, Arterioscler. Thromb. Vasc. Biol., 23: 748-54
103. Kurz D.J., Decary S., Hong Y., Trivier E., Akhmedov A., Erusalimsky J.D. (2004). Chronic Oxidaitive Stress Compromises Telomere Integrity and Accelerates the Onset of Senescence in Human Endothelial Cells, J. Cell Sci., 117: 2417-26
104. Kyo S., Takakura M., Kohama T., Inoue M. (1997). Telomerase activity in human endometrium, Cancer Res., 57(4): 610-4
105. Lafont A., Faxon D. (1998). Why do Animal Models of Post-Angioplasty Restenosis Sometimes Poorly Predict the Outcome of Clinical Trials? Cardiovasc. Res., 39,50-9
106. Landmesser U., Merten R., Spiekermann S., Buttner K., Drexler H., Hornig B. (2000). Vascular Extracellular Superoxide Dismutase Activity in Patients with Coronary Artery Disease: Relation To Endothelium-dependent Vasodilation, Circulation, 101: 2264-70
107. Lehoux S., Tedgui A. (2003). Cellular Mechanics and Gene Expression in Blood Vessels, J. Biomechanics, 36: 631–43
108. Li J-J, Gao R-L. (2005).Should Atherosclerosis be Considered a Cancer of the Vascular Wall? Med. Hypothesis, 64: 694-8
109. Limon-Pacheco J.H., Hernandez N.A., Fanjul-Moles J.L., Gonsebatt M.E. (2007). Glutathione depletion activates mitogen-activated protein kinase (MAPK) pathway that display organ-specific responses and brain protection in mice, Free Radic. Biol. Med., 43(9): 1335-47
110. Liu J.P. (1999). Studies of the Molecular Mechanisms in the Regulation of Telomerase Activity, FASEB J., 13: 2091-104
111. Liu S-C., Wang S-S., Wu M-Z., Wu D-C., Yu F-J., Chen W-J., Chiang F-T., Yu M-F. (2005). Activation of Telomerase and Expression of Human Telomerase Reverse Transcriptase in Coronary Atherosclerosis, Cardiovasc. Pathol., 14: 232-40

156
112. Lorenz M., Saretzki G., Sitte N., Metzkow S., von Zglinicki T. (2001). BJ Fibroblasts Display High Antioxidant Capacity and Slow Telomere Shortening Independent of hTERT Transfection, Free Radic Biol. Med., 31(6): 824-831
113. Lowry O.H., Rosebrough N.J., Farr A.L., Randall R.J. (1951). Protein Measurement with the Folin Phenol Reagent, J Biol Chem, 193(1): 265-75
114. Lustig A.J. (1999). Crisis Intervention: The Role of Telomerase, Proc. Natl. Acad. Sci. U.S.A., 96: 3339-41
115. Mach F. (2000). Toward New Thrapeutic Strategies Against Neointimal Formation in Restenosis, Arteroscler Thromb Vasc. Biol, 20: 1699-1700
116. Malik N., Francis S.F., Holt C.M., Gunn J., Thomas G.L., Shepherd L., Chamberlain J., Newman C.M.H., Cumberland D.C., Crossman D.C.(1998). Apoptosis and Cell Proliferation After Porcine Coronary Angioplasty, Circulation, 98:1657-65
117. Martensson J., Meister A. (1989). Mitochondrial Damage in Muscle Occurs After Marked Depletion of Gluathione and is Prevented by Giving Glutathione Monoester, Proc. Natl. Acad. Sci. U.S.A., 86: 471-75
118. Martensson J., Jain A., Stole E., Frayer W., Auld P.A.M., Meister A. (1991). Inhibiton of Glutathione Synthesis in the Newborn Rat: A Model For Endogenously Produced Oxidative Stress, Proc. Natl. Acad. Sci. U.S.A., 88: 9360-64
119. Matthews C., Gorenne I., Scott S., Figg N., Kirkpatrick P., Ritchie A., Goddard M., Bennett M. (2006). Vascular Smooth Muscle Cells Undergo Telomere-Based Senescence in Human Atherosclerosis: Effects of Telomerase and Oxidative Stress, Circ. Res., 99:156-64
120. Maturo J., Kulakowski E.C.(1988). Taurine Binding to the Purified Insulin Receptor, Biochem. Pharmacol., 37: 3755-60
121. Mergny J-L., Riou J-F., Mailliet P., Teulade-Fichou M-P.,Gilson E. (2002). Natural and Pharmacological Regulation of Telomerase, Nucl. Acid. Res., 30(4): 839-65
122. Meyerson M. (2000). Role of Telomerase in Normal and Cancer Cells, J. Clin. Oncol., 18 (13): 2626-34
123. Meyne J., Ratliff R.L., Moyzis R.K. (1989). Conservation of the Human Telomere Sequence (TTAGGG)n among Vertebrates, Proc. Natl. Acad.Sci. U.S.A., 86(18):7049-53
124. Minamino, T., Kourembanas S. (2001). Mechanisms of Telomerase Induction During Vascular Smooth Muscle Cell Proliferation, Circ. Res., 89: 237–43

157
125. Minamino T., Komuro I. (2002). Role of Telomere in Endothelial Dysfuction in Atherosclerosis, Curr. Opin. Lipidol., 13: 537-43
126. Minamino T., Komura I. (2003). The Role of Telomerase Activation in the Regulation of Vascular Smooth Muscle Cell Proliferation, Drug News Perspec., 16(5): 1-6
127. Minamino T., Komuro I. (2008). Role of Telomeres in Vascular Senescence, Front. Biosci., 13: 2971-79
128. Mognoford J.E., Liu W.R., Reid R., Chiu C., Said H., Chen S., Harley C.B., Mustoe T.A. (2006). Adenoviral Human Telomerase Reverse Transcriptase Dramatically Improves Ischemic Wound Healing Without Detrimental Immune Response in an Aged Rabbit Model, Hum. Gene Ther., 17: 651-60
129. Muller F.L., Lustgarten M.S:, Jang Y., Richardson A., van Remmen H. (2007). Trends in Oxidative Aging Theories, Free Redical Bio Med, 43:477-503
130. Nakamura T.M., Cech T.R. (1998). Reversing Time: Origin of Telomerase, Cell, 92: 587-90
131. Nandhini A.T., Thirunavukkarasu V., Anuradha C.V. (2004). Stimulation of Glucose Utilization and Inhibition of Protein Glycation and AGE Products by Taurine, Acta Physiol. Scand., 181: 297-303
132. Nandhini A.T., Thirunavukkarasu V., Ravichandran M.K. (2005). Effect of Taurine on Biomarkers of Oxidative Stress in Tissues of Fructose-fed Insulin-resitant Rats, Singapore Med. J., 46: 82-87
133. Nandhini A.T., Thirunavukkarasu V., Anuradha C.V. (2005). Taurine Prevents Collagen Abnormalities in High Fructose-fed Rats, Indian J. Med. Res., 122: 171-77
134. Narayanaswamy M., Wright K., Kandarpa K. (2000). Animal Models for Atherosclerosis, Restenosis, and Endovascular Graft Research, J. Vasc. Int. Radiology, 11: 5-17
135. Okuda K., Khan Y., Skurnick J., Kimura M., Aviv H., Aviv A. (2000). Telomere attrition of the Human Abdominal Aorta: relationships with age and atherosclerosis, Atherosclerosis, 152: 391-8
136. Omaska L., McEachern M.J., Nosek J. (2004). Alternatives to Telomerase: Keeping Linear Chromosomes via Telomeric Circles, FEBS Lett, 567: 142-6
137. Onat A. (2001). Risk Factors and Cardiovascular Disease in Turkey, Atherosclerosis, 156(1): 1-10

158
138. Orford J.L., Selwyn A.P., Ganz P., Popma J.J., Rogers C. (2000). The Comparative Pathobiology of Atherosclerosis and Restenosis, Am. J. Cardiol., 86: 6H-11H
139. Oudit G.Y., Trivieri M.G., Khaper N., Husain T., Wilson G.J., Liu P., Sole M.J., Backx P.H. (2004). Taurine Supplementation Reduces Oxidative Stress and Improves Cardiovascular Function in an Iron-overload Murine Model, Circulation, 109: 1877-85.
140. Paglia D.E., Valentine W.N. (1967). Studies on the quantitative and qualitative characterization of erythrocyte glutathione peroxide, J. Lab. Clin. Med., 70(1): 158-69.
141. Pakala R., Dilcher C., Baffour R., Hellinga D., Seabron R., Joner M., Kolodgie F., Virmani R., Waksman R. (2006). Peroxisome Proliferator-Activated Receptor γ Ligand Pioglitazone Alters Neointimal Composition in a Balloon-Denuded and Radiated Hypercholesterolemic Rabbit, J. Cardivasc. Pharmacol., 48: 299-305
142. Passos J. F., von Zgilinicki T. (2005). Mitochondria, Telomeres and Cell Senescence, Exp. Gerontol., 40: 466-72
143. Passos J.F., Von Zglinicki T. (2006). Oxygen Free Radicals in Cell Senescence: Are They Signal Transducers?, Free Radical Res., 40 (12): 1277-83
144. Poole J.C, Andrews L.G, Tollefsbol T.O. (2001). Activity, Function and Gene Regulation of the Catalytic Subunit of Telomerase (hTERT), Gene, 269: 1-12
145. Price C.M. (1999) Telomere and telomerase: Broad Effects on Cell Growth, Curr. Opin. Genet. Dev., 9: 218-24
146. Reddel R.R., Bryan T.M., Colgin L.M., Perrem K.T., Yeager T.R. (2001). Alternative Lengthening of Telomerase in Human Cells. Radiation Res., 155: 194-200
147. Reliene R., Schiestl R.H. (2006). Gluthatione Depletion by Buthionine Sulfoximine Induces DNA Deletion in Mice, Carcinogenesis, 27(2):240-4
148. Rhodes D., Fairall L., Simonsson T., Court R., Chapman L. (2002). Telomere Architecture, EMBO Rep., 3(12): 1139-45
149. Rhyu M.S. (1995). Telomeres, Telomerase and Immortality, J Natl Cancer Inst, 87(12): 884-94
150. Richter T., von Zglinicki T. (2007). A Continous Correlation Between Oxidative Stress and Telomere Shortening in Fibroblasts, Exp. Gerontol., 42: 1039-42

159
151. Richter T.S.G., Nelson G., Melcher M., Olijslagers S., von Zglinicki T. (2007). TRF2 Overexpression Diminishes Repair of Telomeric Single Strand Breaks and Accelerates Telomere Shortening in Human Fibroblasts, Mech. Aging Dev., 128: 340-45
152. Richter T., Proctor C. (2007). The Role of Intracellular Peroxide Levels On The Development and Maintenance of Telomere-dependent Senescence, Exp. Gerontol., 42(11): 1043-52
153. Ross R. (1993). The Pathogenesis of Atherosclerosis: a Perspective for the 1990s, Nature, 362: 801-9
154. Ross R. (1986). The Pathogenesis of Atherosclerosis--an update, N.Engl. J. Med, 314: 488-500
155. Ross R. (1999).Atherosclerosis-An inflamatory disease, N Engl J Med, 340: 115-26,
156. Ross R., Glomset, J.A. (1976). The Pathogenesis of Atherosclerosis, N Engl J Med, 295(7): 369-77
157. Rowe W.B., Meister A. (1973). Studies on the Inhibition of Glutamine Sythethase by Methionine Sulfone, Biochemistry, 12: 1578-82
158. Rudolph K.L., Chang S., Lee L.W., Blasco M., Gottlieb G.J., Greider C., DePinho R.A. (1999) Longevity, Stress Response, and Cancer in Aging Telomerase-dependent Mice, Cell, 96(5):701-12
159. Saldanha S.N., Andrews L.G., Tollefsboll T.O. (2003). Assessment of Telomer Length and Factors That Contribute to its Stability., Eur. J. Biochem., 270: 389-403
160. Samani N.J., Boultby R., Butler R., Thompson J.R., Goodall A.H. (2001). Telomere Shortening in Atherosclerosis, Lancet, 358: 472-73
161. Sata M. (2003). Circulating Vascular Progenitor Cells Contribute to Vascular Repair, Remodeling, and Lesion Formation, Trends Cardiovasc. Med., 13: 249-53
162. Scholzen T., Gerdes J. (2000). The Ki-67 Protein: From the Known and the Unknown, J. Cell. Physiol, 182: 311-22
163. Schwarz S.M., Campbell G.R., Campbell J.H. (1986). Replication of Smooth Mmuscle Cells in Vacular Disease, Circ. Res., 58: 427-44
164. Schwartz, S.M., deBlois D., O’Brien E.R.M. (1995). The intima. Soil for Atherosclerosis and Restenosis, Circ. Res., 77: 445-65
165. Schuller-Levis G.B., Park E. (2004). Taurine and its Chloramine: Modulators of Immunity, Neurochem. Res., 29: 117-26

160
166. Serafini M., Del Rio D. (2004). Understanding the Association Between Dietary Antioxidants, Redox Status and Disease: Is the Total Antioxidant Capacity the Right Tool?, RedoxRep., 9(3):145-52
167. Serano A.L., Andres V. (2004). Telomerase and Cardiovascular Disease, Circ. Res., 94: 575-84
168. Shay J.W., Gazdar A.F. (1997). Telomerase in the Early Detection of Cancer, J. Clin. Pathol., 50: 106-09
169. Shi Y., Niculescu R., Wang D., Patel S., Davenpeck K.L., Zalewski A. (2001). Increased NAD(P)H Oxidase and Reactive Oxygen Species in Coronary Arteries After Balloon Injury, Arterioscler., Thromb. Vasc. Biol., 22: 21-27
170. Shinohara M., Kawashiwa S., Yamashita T., Takaya T., Toh R., Ishida T., Ueyama T., Inoue N., Hirata K., Yokoyama M. (2005). Xenogenic Smooth Muscle Cell Immunization Reduces Neointimal Formation in Balloon-injured Rabbit Carotid Arteries, Cardiovasc. Res., 68: 249-58
171. Slipcevic P. (2003). Telomeres and the Control of Cell Division, Cell Proliferation and Apoptosis, Eds: David Hughes and Huseyin Mehmet, Scientific Publishers Limited, Oxford, p:123.
172. Smith P.K., Krohn R.I., Hermanson G.T., Mallia A.K., Gartner F.H., Provenzano M.D., Fujimoto E.K., Goeke N.M., Olson B.J., Klenk D.C. (1985). Measurement of Protein Using Bicinchoninic Acid, Anal Biochem, 150(1): 76-85
173. Sorescu D. Weiss D., Lassegue B., Clempus R.E., Szöcs K., Sorescu G.P., Valppu L., Quinn M.T., Lambeth J.D., Vega J.D., Taylor W.R., Griendling K.K. (2002). Superoxide Production and Expression of Nox Family Proteins in Human Atherosclerosis, Circulation, 105: 1429-35
174. Southern, E.M. (1975). Detection of Specific Sequences Among DNA Fragments Seperated by Gel Electrophoresis, J. Mol. Biology, 98(3): 503-17
175. Sozer G.O., Kerry Z., Oran I., Gokce G., McEachern J., Topcu Z.(2005) Telomere Length Regulation in the Proliferation of the Vascular Smooth Muscle Cells, FEBS Journal, 272:p129-129 (poster sunumu)
176. Sozer G.O. Gokce G., Oran I., Topcu Z., Kerry Z. Telomere Length Regulation of Vascular Smooth Muscle Cells in a Balloon Injury Model: Effects of Oxidative Stress, 77th EAS Congress, 26-29 Nisan 2008, Istanbul (poster sunumu)
177. Spiekermann S., Landmesser U., Dikalov S., Bredt M., Gamez G., Tatge H., Reepschlager N., Hornig B., Drexler H., Harrison D.G. (2003). Electron Spin Resonance Characterization of Vascular Xanthine and NAD(P)H Oxidase

161
Activity in Patients with Coronary Artery Disease: Relation to Endothelium-dependent Vasodilation, Circulation, 105: 1429-35
178. Strauss B.H., Chisholm R.J., Keeley F.W., Gotlieb A.I., Logan R.A., Armstrong P.W. (1994). Extracellular Matrix Remodeling After Balloon Angioplasty Injury in a Rabbit Model of Restenosis, Circ. Res., 75: 650-8
179. Sturman J.A., (1993)., Taurine in Development, Physiol. Rev., 73: 119-47
180. Suthanthiran M. Anderson M.E., Sharma V.K., Meister A. (1990). Glutathione Regulates Activation-dependent DNA Synthesis in Highly Purified T-lymphocytes Stimulated via CD2 ad CD3 Antigens, Proc. Natl. Acad. Sci. U.S.A., 87: 3343-47
181. Suzuki M., Boothman D.A. (2008). Stress-Induced Premature Senescence (SIPS), J. Radiat. Res., doi:10.1269/jrr.07081
182. Szöcs K., Lassegue B., Sorescu D., Hilenski L.L., Valppu L.L., Couse T.L., Wilcox J.N., Quinn M.T., Lambeth J.D., Griendling K.K. (2002). Upregulation of Nox-based NAD(P)H oxidases in Restenosis After Carotid Injury, Arterioscler. Thromb. Vasc. Biol., 22:21-27
183. Taniyama Y., Griendling K.K. (2003). Reactive Oxygen Species in The Vasculature: Molecular and Cellular Mechanisms, Hypertension, 42: 1075-81
184. Tashiro J., Takahashi K., Yokote K. (1998). Administration of a Small Amount of Lard Enhances Intimal Thickening in the Balloon Catheter Injury Model Without Affecting Serum Lipids, Scan. J. Clin. Lab. Invest., 58: 149-54
185. Tchirkov A., Lansdorp P.M. (2003). Role of Oxidative Stress in Telomere Shortening in Cultured Fibroblasts From Normal Individuals and Patients with Ataxia-Telangiectasia, Hum. Mol. Genet., 12: 227-32
186. Urruticoechea A., Smith I.E., Dowsett M. (2005). Proliferation Marker Ki-67 in Early Breast Cancer, J. Clin. Oncology, 23(28): 7212-720
187. Van Steensel B., Smogorzewska A., de Lange T. (1998). TRF2 Protects Human Telomeres From End-to-end Fusions, Cell, 92(3): 401-13
188. Vasa M., Breitschof K., Zeiher A.M., Dimmeler S. (2000). Nitric Oxide Activates Telomerase and Delays Endothelial Cell Senescence, Circ. Res., 87: 540-42
189. Virchow R. (1989). Cellular Pathology.As Based upon Physiological and Pathological Histology. Lecture XVI-Atheromatous affection of arteries, Nutr. Rev., 47(1): 23-5
190. Vodovotz, Y., Mitchell J. B., Scott Lucia M., McKinney L., Kollum M., Cottin Y., Chan R. C., Barsellos-Hoff M. H., Waksman R. (1999).

162
Modulation of Protein Expression and Activity by Radiation: Relevance to Intracoronary Radiation for the Prevention of Restenosis, Cardiovas. Rad. Med.,1(4): 336-43
191. Von Zglinicki T., Saretzki G., Docke W., Lotze C. (1995). Mild Hyperoxia Shortens Telomeres and Inhibits Proliferation of Fibroblasts: A Model for Senescence?, Exp. Cell Res., 220: 186-93
192. Von Zglinicki T., Pilger R., Sitte N. (2000). Accumulation of Single-Strand Breaks is the Major Cause of Telomere Shortening in Human Fibroblasts, Free Radical Biol. Med., 28: 64-74
193. Von Zglinicki T. (2005). Oxidative Stress Shortens Telomeres, Trends Biochem. Sci., 27(7): 339-44
194. Von Zglinicki T., Martin-Ruiz C.M. (2005). Telomeres as Biomarkers for Ageing and Age-related Diseases, Curr. Mol. Med., 5(2): 197-203
195. Walsh K. (2000). Building a Better Mouse Model, J Mol. Cell Cardiol., 32: 1921–2
196. Walter J.R., Murphy G.J., Bicknell G.R., Sandford R., Margolin S.B., Nicholson ML. (2002). Pirfenidone Inhibits Early Myointimal Proliferation but has No Effect on Late Lesion Size in Rats, Eur J. Endovasc. Surg.,23: 234-40
197. Wassmann S., Wassmann K., Nickening G. (2004). Modulation of Oxidant and Antioxidant Enzyme Expression and Function in Vacular Cells, Hypertension, 44: 381-86
198. Watanabe T., Sagisaka H., Arakawa S., Shibaya Y., Watanabe M., Igarashi I., Tanaka K., Totsuka S., Takasaki W., Manabe S. (2003). A Novel Model of Continuous Depletion of Glutathione in Mice Treated with L-Buthionine (S,R)-Sulfoximine, J. Toxico. Sci., 28(5): 455-69
199. Wentzel J.J., Gijsen F.J.H., Stergiopulos N., Serruys P.W., Slager C.J., Krams R. (2003). Shear Stress, Vascular Remodeling and Neointimal Formation, J. Biomechanics, 35,681–8
200. Wiechelman K.J., Braun R.D., Fitzpatrick J.D. (1988). Investigation of the Bicinchoninic Acid Protein Assay: Identification of the Groups Responsible for Color Formation, Anal Biochem, 15,175(1): 231-7
201. Wilcox J.N., Cipolla G.D., Martin F.H., Simonet L., Dunn B., Ross C.E., Scott N.A. (1997). Contribution of Adventitial Myofibroblasts To Vascular Remodeling and Lesion Formation After Experimental Angioplasty in Pig Coronary Arteries, Ann. N. Y. Acad. Sci., 811: 437-47

163
202. Wojtecka-Lukasik E., Czuprynska K., Maslinska D. (2006). Taurine-chloramine Is a Potent Antiinflammatory Substance, Inflamm. Res., 55: S17-S18
203. Wood J.G., Sinclair D.A. (2002). TPE or not TPE? It’s No Longer a Question, Trends Pharmacol. Sci., 23(1): 1-4
204. Xiang H., Wang J., Mao Y., Li D.W: (2000). hTERT Can Function With Rabbit Telomerase RNA: Regulation of Gene Expression and Attenuation of Apoptosis, Biochem. Bioph. Res.Co., 278: 503-10
205. Xu D., Neville R., Finkel T. (2000). Homocysteine Accelerates Endothelial Cell Senescence, FEBS Lett., 470: 20-24
206. Yardim-Akaydin S., Sepici A., Ozkan Y., Torun M., Simsek B., Sepici V. (2004). Oxidation of Uric Acid in Rheumatoid Arthritis: Is Allantoin a Marker of Oxidative Stress?, Free Radical Res., 38(6): 623-28
207. Yardim-Akaydin S., Sepici A., Ozkan Y., Simsek B., Sepici V. (2006). Evaluation of Allontoin Levels As a New Marker of Oxidative Stress in Behcet’s Disease, Scand. J. Rheumatol., 35(1):61-64
208. Yasumoto, S., Kunimura C., Kikuchi K., Tahara H., Ohji H., Yamamoto H., Ide T., Utakoji T., (1996). Telomerase Activity in Normal Human Epithelial Cells, Oncogene, 13(2): 433-9
209. Yucel D., Şeneş M., Topkaya B.C., Zengi O.(2006). Oxidative Stress/Nitrosative Stress in Chronic Heart Failure: A Critical Review, Tur. J. Biochem., 31(2): 86-95
210. Zhang M., Bi L.F., Fang J.H. (2004). Beneficial Effects of Taurine on Serum Lipids in Overweight or Obese Nondiabetic Subjects, Amino Acids, 26: 267-71
211. Zou J., Huang Y., Cao K., Yang G., Yin H., Len J., Hsieh T-chen, Wu J.M. (2000). Effect of Resveratrol on Intimal Hyperplasia After Endothelial Denudation in an Experimental Rabbit Model, Life Sci.,68: 153-63

164
ÖZGEÇMİŞ
1992 yılında Ege Üniversitesi Eczacılık Fakültesi’ne girdim ve 1997 yılında aynı
fakülteden mezun oldum. 1997’de Ege Üniversitesi Sağlık Bilimleri Enstitüsü
Farmakoloji Yüksek Lisans Programı’na kabul edildim. Aynı yıl Ege Üniversitesi
Eczacılık Fakültesi Farmakoloji Anabilim Dalı’na araştırma görevlisi olarak atandım.
2000 yılında “Tavşan İzole Damar Preparatlarında Çeşitli Kastırıcı Ajanların
Yanıtlarının Endotel ile Etkileşiminin İncelenmesi” isimli tezim ile yüksek lisans
tezimi tamamladım.Halen aynı anabilim dalında görevimi sürdürmekteyim.




















