
Intracellular autofluorescence: a biomarker for epithelial cancer stem cells
Upload
independentCategory
view
2download
0
Neurobiology of Aging xx (2012) xxx
0h
Tau pathology modulates Pin1 post-translational modifications and maybe relevant as biomarker
Kunie Andoa,b, Pierre Dourlena,b, Anne-Véronique Samboa,b, Alexis Brettevillea,b,Karim Bélarbia,b, Valérie Vingtdeuxa,b, Sabiha Eddarkaouia,b, Hervé Drobecqb,c,
Antoine Ghestema,b, Séverine Bégarda,b, Emmanuelle Demey-Thomasd, Patricia Melnykb,c,Caroline Smete, Guy Lippense, Claude-Alain Mauragea,b,f, Marie-Laure Caillet-Boudina,b,Yann Verdierd, Joelle Vinhd, Isabelle Landrieue, Marie-Christine Galasa,b, David Bluma,b,
Malika Hamdanea,b, Nicolas Sergeanta,b,f,*, Luc Buéea,b,f,*a Inserm, UMR 837, Alzheimer & Tauopathies, Jean-Pierre Aubert Research Centre, Faculty of Medicine, Institute of Predictive Medicine and
Therapeutic Research, Lille, Franceb Univ. Lille Nord de France, UDSL, Lille, France
c CNRS, UMR 8161, Institut Pasteur de Lille, Lille, Franced ESPCI Biological Mass Spectrometry and Proteomics USR 3149 CNRS/ESPCI ParisTech, Paris, France
e CNRS, UMR 8576, Univ Lille1, Villeneuve d’Ascq, Francef CHR-Lille, Department of anatomo-pathology, Lille, France
Received 17 September 2011; received in revised form 18 July 2012; accepted 2 August 2012
Abstract
A prerequisite to dephosphorylation at Ser–Pro or Thr–Pro motifs is the isomerization of the imidic peptide bond preceding the proline.The peptidyl-prolyl cis/trans isomerase named Pin1 catalyzes this mechanism. Through isomerization, Pin1 regulates the function of agrowing number of targets including the microtubule-associated tau protein and is supposed to be deregulated Alzheimer’s disease (AD).Using proteomics, we showed that Pin1 is posttranslationally modified on more than 5 residues, comprising phosphorylation, N-acetylation,and oxidation. Although Pin1 expression remained constant, Pin1 posttranslational two-dimensional pattern was modified by tau overex-pression in a tau-inducible neuroblastoma cell line, in our THY-Tau22 mouse model of tauopathy as well as in AD. Interestingly, in all ofthese systems, Pin1 modifications were very similar. In AD brain tissue when compared with control, Pin1 is hyperphosphorylated at serine16 and found in the most insoluble hyperphosphorylated tau fraction of AD brain tissue. Furthermore, in all tau pathology conditions,acetylation of Pin1 may also contribute to the differences observed. In conclusion, Pin1 displays several posttranslational modifications,which are specific in tauopathies and may be useful as biomarker.© 2012 Elsevier Inc. All rights reserved.
Keywords: Alzheimer’s disease; Tauopathy; Pin1; Microtubule-associated protein tau; Proteomics; Phosphorylation; Acetylation; Posttranslational modifi-cation; Biomarker
* Corresponding author at: Inserm, UMR 837, Alzheimer & Tauopathies, Univ. Lille Nord de France, Centre Jean-Pierre Aubert, Institut de MédecinePrédictive et Recherche Thérapeutique, Faculté de Médecine, 59045 Lille cedex, France. Tel.: �00 33 (0) 320 622074; fax: �00 33 (0) 320 622079.
E-mail address: [email protected] (L. Buée).* Alternate corresponding author at: Inserm, UMR 837, Alzheimer & Tauopathies, Univ. Lille Nord de France, Centre Jean-Pierre Aubert, Institut de
Médecine Prédictive et Recherche Thérapeutique, Faculté de Médecine, 59045 Lille cedex, France. Tel.: �00 33 (0) 320 622074; fax: �00 33 (0) 320 622079.
www.elsevier.com/locate/neuaging
E-mail address: [email protected] (N. Sergeant).
197-4580/$ – see front matter © 2012 Elsevier Inc. All rights reserved.ttp://dx.doi.org/10.1016/j.neurobiolaging.2012.08.004

cpptycpgsr
fct
osciw
macfiesofimpmgma
2
2
wt1MfFa1(b
2 K. Ando et al. / Neurobiology of Aging xx (2012) xxx
1. Introduction
The peptidyl-prolyl cis/trans isomerase Pin1 (EC5.2.1.8) is a cytosolic protein that isomerizes the peptidebond of a phosphorylated-serine or a phosphorylated-threonine followed by a proline (pSer-/pThr-Pro). Pin1catalyzes the cis-/trans-isomerization of its substrates,onsequently potentiating the accessibility of the phos-hate residue for further dephosphorylation by proteinhosphatases such as the protein phosphatase PP2A. Al-ernatively, the binding of Pin1 to other highly phosphor-lated substrates can repress their dephosphorylation byalcineurin. Therefore, through isomerization of pSer-/Thr-Pro, Pin1 regulates the function or degradation of arowing number of proteins including cytoskeletal, tran-cription factors, mitotic, or proapoptotic proteins (foreview see Rudrabhatla and Pant, 2010). Pin1 consists of
2 functional domains. The binding domain corresponds tothe amino terminal region consisting of a group IV WWdomain (Trp-Trp domain) that specifically binds to pSer-/pThr-Pro motifs. The carboxyl terminal region is thecatalytic domain (for review, Wulf et al., 2005).
Pin1 substrate-binding and isomerase activity are regu-lated by phosphorylation. Indeed, 3 phosphorylation sites ofPin1 are yet characterized. Serine 16 is located in the WWdomain and is phosphorylated by protein kinase A. Phos-phorylation of Pin1 at serine 16 represses the substraterecognition (Lu et al., 2002). Pin1 is also phosphorylated atserine 65 by Polo-like kinase 1 (Eckerdt et al., 2005) and atserine 71 by death-associated protein kinase 1 (Lee et al.,2011). Phosphorylation at serine 65 stabilizes Pin1 andpreserves the isomerase activity (Eckerdt et al., 2005)whereas phosphorylation at serine 71 precludes the isomer-ase activity (Lee et al., 2011). Pin1 has several additionalputative phosphorylation sites (e.g., human Pin1 has 29residues of serine or threonine, and 3 tyrosines). For in-stance, phosphorylation at serine 18 may also regulate Pin1substrate recognition and binding (Landrieu et al., 2006).Similarly, a catalytic mutant of Pin1, where serine 115 isreplaced by a glutamate, represses the isomerase activity(Galas et al., 2006). Point mutations in the WW domain liketyrosine 23 (Y23A, or W34A) also reduce Pin1 bindingactivity (Lu et al., 1999b; Nishi et al., 2011). Apart fromphosphorylation, a whole proteome study of lysine acetyla-tion has identified the lysine 46 of Pin1 WW domain aspotentially acetylated (Choudhary et al., 2009). The bindingof 14-3-3 protein to phosphorylated motifs is repressed byacetylation. By similarity, acetylation of Pin1 could alsomodify the binding of Pin1 to its substrates, but this remainsto be demonstrated.
Alzheimer disease (AD) is characterized by neurofi-brillary tangles (NFT) and extracellular amyloid deposits.NFT are composed of intraneuronal aggregates of hyper-phosphorylated tau proteins and amyloid deposits are
made of amyloid-beta peptide (A�) peptides that result arom the sequential cleavage of its transmembrane pre-ursor named amyloid precursor protein. Pin1 facilitatesau dephosphorylation (Zhou et al., 2000) and regulated
amyloid precursor protein metabolism, thus suggesting apotential neuroprotective function of Pin1 against AD(Galas et al., 2006; Hamdane et al., 2006; Liou et al.,2003; Lu et al., 1999a; Pastorino et al., 2006). However,Pin1 activity may be repressed by oxidation in AD (But-terfield et al., 2006; Sultana et al., 2006). Furthermore,the soluble pool of Pin1 is depleted in AD (Lu et al.,1999a), and Pin1 is localized to granular vesicles in ADand frontotemporal dementias but not to tau aggregates(Dakson et al., 2011; Holzer et al., 2002; Lu et al., 1999a;Ramakrishnan et al., 2003; Thorpe et al., 2001, 2004).This mislocalization of Pin1 could be related to an ex-cessive amount of hyperphosphorylated tau (Thorpe etal., 2001). In contrast, a recent study suggests no rela-tionship between Pin1 mislocalization and tau or amyloidpathology (Dakson et al., 2011). An abnormal regulation
f Pin1 in AD might be instrumental even if the relation-hip with the severity of brain lesions remains elusive. Inontrast, we can also hypothesize that tau pathology maympair Pin1 activity because Pin1 is observed associatedith granular vesicles in AD and other tauopathies.Pin1 appears as a marker associated with the develop-
ent of several neurological disorders but less is knownbout Pin1 itself in these disease-associated conditions. Be-ause Pin1 activity is regulated by posttranslational modi-cations, we analyzed Pin1 using two-dimensional (2D) gellectrophoresis coupled to Western blotting as well as masspectrometry (MS). In the present study, we show thatverexpression of tau modifies the 2D electrophoretic pro-le of Pin1. Similarly, in our THY-Tau22 transgenic mouseodel of tau pathology, Pin1 was found to be hypophos-
horylated. Moreover, we also showed that posttranslationalodifications of Pin1 are altered in AD brain tissue. To-
ether, we show for the first time that Pin1 posttranslationalodifications of Pin1 are altered in AD and in cellular and
nimal models of tau pathology.
. Methods
.1. Antibodies
Anti-Pin1 polyclonal antibody H123 (dilution 1:1000)as from Santa Cruz Biotechnology (Clinisciences, Nan-
erre, France). Anti-Pin1 polyclonal antibody (dilution:2000) was from Upstate Biotechnology (Euromedex,undolshem, France). Anti-phospho-serine 16 of Pin1 was
rom Cell Signaling (Ozyme, Saint Quentin Yvelines,rance) and used at final dilution of 1:4000. A well-char-cterized anti-human tau against amino acid sequence 1 to9 M19G polyclonal antibody (1:20,000) was also usedBussière et al., 1999). Anti-tau pThr231 monoclonal anti-ody AT180 was purchased from Innogenetics. Anti-�-
ctin antibody and all secondary antibodies coupled with
H(ntat
2b
hmA
HtuEt
m
3K. Ando et al. / Neurobiology of Aging xx (2012) xxx
horseradish peroxidase were from Sigma Immunochemical(Saint-Quentin Fallavier, France).
2.2. Human brain subjects
All the human brain autopsy materials used in this studywere from the Lille Neurobank (Regional Hospital Centreand University of Lille). Tissue was removed according toLocal Ethics Committee guidelines, and informed consentfor brain donation was obtained from next of kin. Fordemographic details of cases, see Table 1. Frontal corticalbrain areas showing neuropathological characteristics ofAD were used.
2.3. Brain tissue fractionation
Brain tissues were processed as previously described(Sergeant et al., 2003b). Briefly, brain tissue samples werehomogenized in 10 mM Tris-HCl pH 6.8 solution (1/10wt/vol) using a Teflon potter homogenizer. The homogenatewas centrifuged at 100,000 � g for 1 hour at 4 °C; thesupernatant was collected and the pellet was homogenizedin the starting buffer added with detergents of a growingsolubilization strength. A series of solubilization and cen-trifugation was performed with the starting buffer addedwith (1) 0.5%, (2) 2% Triton X-100, (3) 0.5% SDS (sodiumdodecyl sulfate), and (4) the final pellet was homogenized inLaemmli sample buffer. The supernatants recovered afterthe 4 centrifugations were added with 1 volume of a 2�Laemmli SDS-polyacrilamyde gel electrophoresis (PAGE)
Table 1Demographic characteristics of the AD patients and control subjects
Age (y) Gender PMI (h) Tau A�42-40
Control 1 72 M 12 S3 S0, S0Control 2 78 M 24 S1 S0, S0Control 3 89 F 16 S2 S0, S0Control 4 95 M 60 S4 S0, S0Control 5 46 F 24 S0 S0, S0Control 6 43 M — S0 S1, S0Control 7 42 M — �S4 S0, S0Control 8 26 F 15 S0 S0, S0Control 9 36 M 23 S0 S0, S0Control 10 65 F — S3 S1, S0AD 1 71 F 6 S10 S9, S3AD 2 74 M 4 S10 S9, S8AD 3 78 M 15 S10 S7, S8AD 4 80 F 12 S10 S8, S5AD 5 73 M — S10 S8, S6AD 6 73 M 9 S10 S7, S0AD 7 82 F — S10 S6, S0AD 8 54 F — S10 S10, S7AD 9 65 F 6 S9 S9, S4AD 10 81 M — S10 S8, S4AD 11 91 F — S8 S7, S7
Nondemented subjects used as controls; — indicates unknown PMI. S0–S10 indicates stages of amyloid and tau pathology, as described in Dela-courte et al. (2002).Key: A�, amyloid-beta peptide; AD, Alzheimer’s disease; F, female; M,
ale; PMI, postmortem interval.
loading buffer. The protein concentration in each fraction P
was determined using the BCA protein assay kit (FischerScientific SAS, Illkrich, France), in accordance with themanufacturer’s instructions.
2.4. Mouse brain subjects
Whole brains of transgenic mice overexpressing humantau G272V P301S (THY-Tau22) were dissected by separat-ing the hippocampus from the cortex and thalamus as de-scribed (Schindowski et al., 2006). Hippocampal tissueswere used for this study as this area showed high levels ofhuman tau expression. Brain tissue homogenates were pre-pared following the same protocol as for human brain tissue.All protocols followed European animal welfare regulationsand were approved by the local ethical committee (Ap-proval n° AF 06/2010, 31/3/2010).
2.5. Cell culture and pharmacological treatment
Tau and Pin1 inducible SY5Y cell lines were previouslydescribed (Bretteville et al., 2009; Hamdane et al., 2006).To inhibit phosphatase activities, after 48 hours of tetracy-cline-induction SY5Y-Pin1 cells were treated with 125 nMokadaic acid for 2 hours as described (Lefebvre et al., 2003).
istone deacetylase were inhibited with trichostatin ATSA, Sigma-Aldrich, Sigma-Aldrich, Lyon, France), aonselective inhibitor. SY5Y Pin1-inducible cells werereated for 24 or 48 hours with 0.1, 0.5, or 1 �M of TSAfter induction of Pin1 expression during 24 hours withetracycline.
.6. SDS-PAGE, 2D gel electrophoresis, and Westernlotting
For SDS-PAGE analysis, 200 mg of brain tissue wasomogenized in 2 mL Laemmli sample buffer containing 20M of Dithiothreitol (DTT) (Fluka Biochemica, Sigma-ldrich) and incubated at 100 °C for 10 minutes. Fifty �g
of protein was loaded on Tris-Tricine 16.5% gel followedby Western blotting as described (Vingtdeux et al., 2005).For 2D gel electrophoresis, 200 mg of brain tissue washomogenized in 2 mL 2D buffer containing 7 M urea, 2 Mthiourea, 4% Triton X and 10 mM DTT. Brain lysate (500�g) was loaded on immobilized pH gradient (IPG) Strip3–10 (GE Healthcare, Buc, France); isoelectric focusingwas performed with the isoelectric focusing IPGPhore (GEHealthcare) by applying a total of 75 kV per hour, accordingto the manufacturer’s instructions. The IPG strip was equil-ibrated 3 times for 15 minutes in Laemmli sample bufferand was layered over a 16.5% Tris–Tricine SDS-PAGE.Western blot films were digitized using a Umax. scanner (GE
ealthcare). The scanner was calibrated for optical densi-ies. Image-Master 1D Elite Software (GE Healthcare) wassed to quantify the signal, and data were collected usingxcel:Mac2011 Software (Microsoft, Les Ulis, France). T-
est analysis was performed using Prism 5 software (Graph
ad Software, Inc., San Diego, CA, USA). Results from
tMp
p
BDibb
2
Smcbpdsr0
iv(pftsprP
4 K. Ando et al. / Neurobiology of Aging xx (2012) xxx
either representative experiments or the mean (� standarderror of the mean) of all experiments are shown.
2.7. Protein expression and purification and PKA in vitrophosphorylation
Pin1 protein with an N-terminal histidine-tag fusion wereproduced in Escherichia coli BL21(DE3) strain, carryinghe recombinant Pin1-pET15b plasmid (Novagen, Merck-
illipore, Darmstadt, Germany). The recombinant Pin1roteins were produced and purified as described (Smet et
al., 2004). In vitro protein kinase A (PKA; EC 2.7.11.11)hosphorylation was performed as described (Leroy et al.,
2010).
2.8. In vitro dephosphorylation with �-phosphatase
Phosphorylated Pin1 was incubated at 37 °C, for 80minutes with �-protein phosphatase (EC 3.1.3.16) in reac-tion buffer (2 mM MgCl2, reaction buffer 1�, New England
iolab, Evry, France) as described (Bretteville et al., 2009).ephosphorylation was stopped on ice and samples were
mmediately mixed to LDS (lithium dodecyl sulfate) sampleuffer for SDS-PAGE or dissolved in a 2D electrophoresisuffer for 2D analysis.
.9. Mass spectrometry
Two-dimensional electrophoresis was performed forY5Y Pin1 cell lysates, followed by coloration with Coo-assie brilliant blue. The stained polypeptide spots that
orresponded to the positive spots recognized by Westernlotting using anti-Pin1 antibodies were cut in 1-mm2 gelieces and analyzed by MALDI-TOF (matrix-assisted laseresorption-ionisation and time-of-flight) as previously de-cribed (Sergeant et al., 2003a). Briefly, the trypsin (Rocheef. 1418025, La Rochelle, France) was resuspended in TFA.01% (trifluoroacetate) at a final concentration of 0.1 �g/
�L. The spots were treated with DTT and the reducedcysteines were alkylated with iodoacetamide at 37 °C for 2hours and 20 minutes on an orbital shaker. After dehydra-tion of the gel-spots 6 to 12 ng of trypsin was added anddigestion was stopped by the addition of 40 �L of 10%formic acid. The peptide mixture was analyzed by MALDI-TOF (Applera Applied Biosystems, Inc., Framingham, MA,USA), FTICR (Fourier transform ion cyclotron resonancemass spectrometry) (LTQ-Ft, Thermo, Fisher Scientific)and MALDI-Orbitrap (LTQ-Orbitrap, Thermo Electron).Peak-lists were generated using GPS 3.6 Explorer Softwarefor MALDI-TOF data, and using Exalibur 2.0.7 forMALDI-Orbitrap and FTICR acquisitions. Protein identifi-cation was performed using Mascot 2/2/04 (Matrix Science)software, using the following parameters: enzyme used:database: Swissprot (2010)-05, taxon, homo sapiens(20,280 entries), trypsin; 2 miss cleavages permitted, nofixed modification, variable modifications: carbamidom-ethylation (C), oxidation (M), acetylation N-terminal; mass
tolerance for precursor ions: 25 ppm for MALDI-TOF spec-tra, 5 ppm for MALDI-Orbitrap and FTICR spectra; masstolerance for fragment ions: 0.3 Da for MALDI-TOF, 0.9Da for FTICR. The following peptides from matrix, keratinand trypsin were excluded: 806.08, 917.29, 1707.79,2163.06, and 2273.18. Proteins with a threshold p � 0.05were validated.
3. Results
3.1. Posttranslational modifications of Pin1
Although recent studies have shown that Pin1 Ser16 andSer65/71 are phosphorylated, the overall phosphorylationstate or post-translational modifications of Pin1 remainedunknown. To determine the potential number of phospho-sites on Pin1 in vivo, we analyzed Pin1 isovariants using 2Dgel electrophoresis followed by Western blotting in a well-characterized Pin1 inducible SY5Y neuroblastoma cell line(Hamdane et al., 2002, 2006). According to the primaryamino acid sequence, theoretical isoelectric point of Pin1 isof 8.95. For 2D analysis, Pin1 isovariants were resolvedusing a pH range between 3 and 11. Endogenous SY5YPin1 was resolved as 4–5 isovariants on 2D Western blot-ting (Fig. 1A and Supplementary Fig. 1, SY5Y native). InSY5Y cells stably expressing Pin1, 6 isovariants were ob-served likely due to a greater sensitivity of the detection(Fig. 1A and Supplementary Fig. 1, SY5Y Pin1). The firstisovariant on the right corresponded to native Pin1 becauseits pI was close to 9 or Pin1 was even stacked at the basicextremity of the strip (Fig. 1A). Starting from the nativeform of Pin1, 5 acidic isovariants were detected suggestingthat 5 posttranslational modifications of Pin1 were associ-ated with the increase acidity of Pin1. Several posttransla-tional modifications can confer an anionic charge to a poly-peptide including phosphorylation, acetylation of lysine,oxidation, or even nitrosylation. To determine the contribu-tion of phosphorylation, SY5Y cells stably expressing Pin1were treated with okadaic acid, an inhibitor of serine/thre-onine-protein phosphatases (Fig. 1A). In addition to Pin1sovariants visualized in the control condition, additionalery acidic isovariants with a shift toward higher MWmolecular weight) were observed further supporting phos-horylation as a major posttranslational modification. Tourther demonstrate that phosphorylation was involved inhe 2D pattern of Pin1, a phospho-dependent Pin1 antibodypecific of Pin phosphorylation at serine 16 was used (Sup-lementary Fig. 2). First, to assess the antibody specificity,ecombinant-Pin protein was phosphorylated in vitro withKA followed or not by dephosphorylation with the �-phos-
phatase. Pin1 proteins were then analyzed by 2D-gel elec-trophoresis. Without treatment, the recombinant protein wasresolved as a single spot stacked at the basic part of the pHgradient. This spot was not detected with phosphorylated-serine 16 Pin1 antibody. In sharp contrast, an increasedacidity of Pin1 was observed following phosphorylation of
Pin1 with PKA and the acidic isovariants were detected
dfiFepBps
wotmoaf
5K. Ando et al. / Neurobiology of Aging xx (2012) xxx
with the phosphorylated-serine 16 antibody. Moreover, thissignal was abrogated following the treatment with �-phos-phatase and the number of acidic isovariants of Pin1 wasalso reduced, therefore demonstrating the specificity of thephosphorylated-serine 16 Pin1-antibody. We analyzed Pin1phosphorylation in SY5Y Pin1 homogenate treated or notwith the �-phosphatase. In the control condition, a band wasetected at 18 kDa and this band was no more detectedollowing �-phosphatase treatment demonstrating that Pin1s efficiently dephosphorylated at serine 16 (Supplementaryig. 3A). The same homogenates were used for 2D-gellectrophoresis followed by Western blotting with Pin1hosphoserine 16 or Pin1 antibodies (Supplementary Fig. 3). Four among the 6 Pin1 isovariants were stained with thehosphoserine 16 Pin1 and the staining was abrogated inample treated with the �-phosphatase. Together, our data
Fig. 1. Two-dimensional (2D) characterization of multiple Pin1 isovariantPin1 overexpressing cells and SY5Y Pin1 overexpressing cells treated withisovariant (0) to the most acidic (1–5). (B) Mass spectrometry. Tryptic pepti(matrix-assisted laser desorption-ionisation and time-of-flight). MS/MS adiphosphate kinase (�), and Trypsin (T). As shown in the enlargement boxm/z from distinct peptides. (C) Sequence coverage of Pin1 obtained by malines underlying Pin1 sequence corresponds to the spots (indicated by colomethionine residues, the phosphorylated sites are indicated by ℗ and N-acycolored in light blue and purple respectively.
demonstrate that the 3 most acidic isovariants of Pin1 are c
phosphorylated at serine 16 as well as the first isovariantadjacent to the native protein and therefore, posttransla-tional modifications other than phosphorylation may con-tribute to the diversity of Pin1 isovariants.
We next isolated 4 isovariants and analyzed those bymass spectrometry following in-gel-trypsin digestion.Note that the 2 most acidic spots were not visualized onthe Coomassie-stained gel and they were therefore notanalyzed (Fig. 1A). A similar mass spectrometry pattern
as obtained for the 4 spots (not shown). Peptides massesverlapped more than 90% of Pin1 sequence. For post-ranslational modifications, oxidation was identified atethionine 130 and 146 (Fig. 1B and C). N-acetylation
f the primary amine of second alanine residue and N-cetylation the lysine 46 of Pin1 have been identified. Tourther demonstrate that Pin1 is N-acetylated, SY5Y Pin1
epresentative 2D Western blots for Pin1 show Pin1 isovariants in SY5Yokadaic acid for 2 hours. Pin1 isovariants are numbered from the native
fingerprint from 2D spot number 3 (in Da, m/z) obtained by MALDI-TOFdemonstrates the presence of Pin1 (2), cyclophilin A (�), nucleotideI Orbitrap very high resolution allows the accurate measurement of close
trometry (MALDI Orbitrap and MALDI-TOF analyses). The color of thebers) where the peptides were characterized. Asterisks indicated oxidizedresidues are indicated by (NAc). The WW domain and PPiase domain are
s. (A) R125 nM
de massnalysis, MALDss spec
red numtelated
ells were treated with the nonselective histone deacety-
2HsAwtatPrssdposTtdiv
gcp
6 K. Ando et al. / Neurobiology of Aging xx (2012) xxx
lase inhibitor TSA. We first determined the experimentalconditions to efficiently repress histone H3 deacetylation(Supplementary Fig. 4). SY5Y Pin1 cells were treated for
4 or 48 hours with incremental doses of TSA. Histone3 acetylation was determined using an antibody that
pecifically recognizes N-acetylated form of histone H3.s shown on Supplementary Fig. 2, the optimal conditionas achieved at 0.1 �M of TSA during 48 hours showing
he strongest N-acetylated H3 staining. The globalmount of histone H3 or Pin1 was not modified by thereatment (Supplementary Fig. 4). Next, we analyzedin1 by 2D-gel electrophoresis and used an antibodyaised against N-acetylated motif to determine if Pin1pots are detected with this antibody. Isovariants with theame pI and MW as Pin1 were detected in control con-itions and TSA treatment enhanced the labeling (Sup-lementary Fig. 4). An increased acidity of Pin1 was alsobserved in SY5Y Pin1 cells treated with TSA thereforehowing that Pin1 is acetylated. Moreover, additionalau isovariants stained by the phospho-Thr231 were de-
ected following TSA treatment suggesting a decreasedephosphorylation of tau at that site but not a globalncreased tau phosphorylation because more acidic iso-
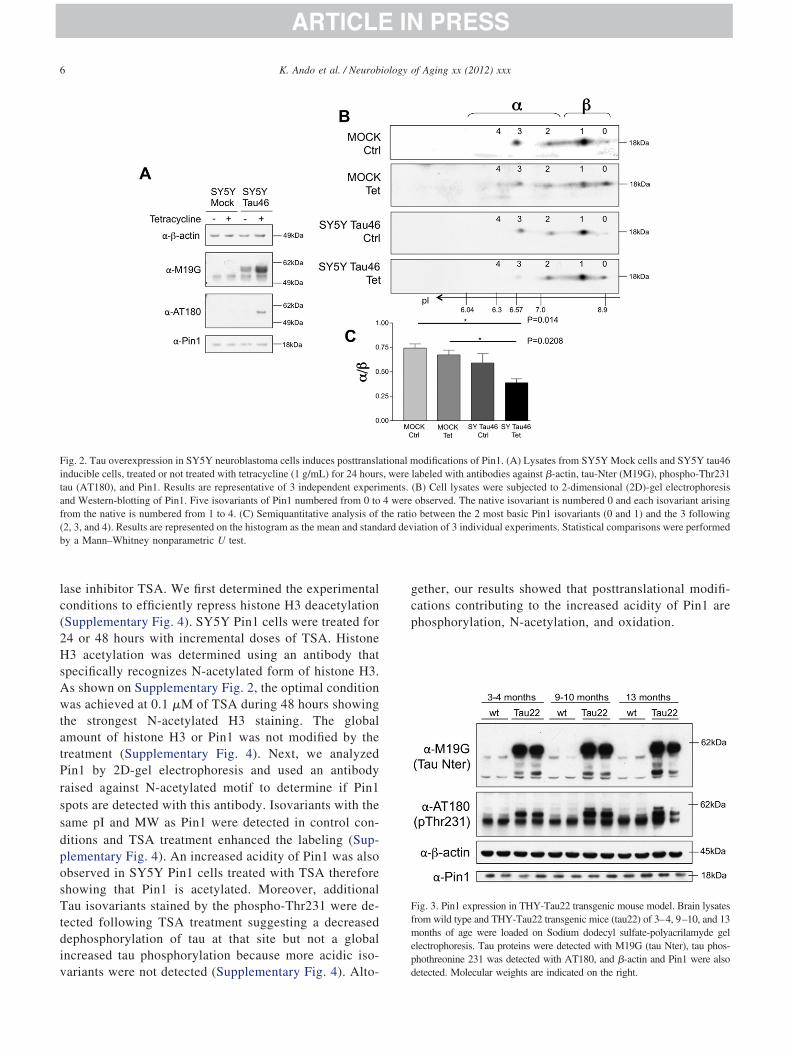
Fig. 2. Tau overexpression in SY5Y neuroblastoma cells induces posttranslainducible cells, treated or not treated with tetracycline (1 g/mL) for 24 hourtau (AT180), and Pin1. Results are representative of 3 independent experiand Western-blotting of Pin1. Five isovariants of Pin1 numbered from 0 tofrom the native is numbered from 1 to 4. (C) Semiquantitative analysis of(2, 3, and 4). Results are represented on the histogram as the mean and standby a Mann–Whitney nonparametric U test.
ariants were not detected (Supplementary Fig. 4). Alto- d
ether, our results showed that posttranslational modifi-ations contributing to the increased acidity of Pin1 arehosphorylation, N-acetylation, and oxidation.
Fig. 3. Pin1 expression in THY-Tau22 transgenic mouse model. Brain lysatesfrom wild type and THY-Tau22 transgenic mice (tau22) of 3–4, 9–10, and 13months of age were loaded on Sodium dodecyl sulfate-polyacrilamyde gelelectrophoresis. Tau proteins were detected with M19G (tau Nter), tau phos-phothreonine 231 was detected with AT180, and �-actin and Pin1 were also
odifications of Pin1. (A) Lysates from SY5Y Mock cells and SY5Y tau46labeled with antibodies against �-actin, tau-Nter (M19G), phospho-Thr231(B) Cell lysates were subjected to 2-dimensional (2D)-gel electrophoresisobserved. The native isovariant is numbered 0 and each isovariant arising
o between the 2 most basic Pin1 isovariants (0 and 1) and the 3 followingiation of 3 individual experiments. Statistical comparisons were performed
tional ms, werements.4 werethe ratiard dev
etected. Molecular weights are indicated on the right.

tptlitP1hT2mc
ppeeSvtpwiisr
RapWtoP
ric Man
7K. Ando et al. / Neurobiology of Aging xx (2012) xxx
3.2. Tau accumulation modifies Pin1 posttranslationalmodifications
To further investigate relationships between Pin1 post-translational modifications and its substrates, we usedSY5Y Tau46 inducible cell model and analyzed endog-enous Pin1 using 2D-gel electrophoresis and Westernblotting. The human tau inducible cells, SY5Y tau46cells, were generated from SH-SY5Y neuroblastomacells (Bretteville et al., 2009). After incubation withetracycline for 24 hours, SY5Y tau46 cells overex-ressed tau, whereas tetracycline treatment did not alterau expression level in Mock cells (Fig. 2A). Neverthe-ess, tau was expressed at low levels in SY5Y tau46 cellsn the absence of tetracycline indicating a small leak ofhe system (Fig. 2A). The phosphothreonine 231 is ain1-recognition motif (Landrieu et al., 2006; Lu et al.,999a; Smet et al., 2004; Wintjens et al., 2001). After 24ours of induction, an increased level of phospho-hr231-immunoreactive tau protein was observed (Fig.A; AT180). The enhanced expression of tau did notodify the amount of Pin1 in SY5Y tau46 cells when
Fig. 4. Two-dimensional (2D) gel analysis of Pin1 in wild type (WT) and(tau22) were resolved by 2D-gel electrophoresis followed by Western bisovariants 0 and 1) and the 3 following (�-group: isovariants 2, 3, and 4(tau22; n � 3) transgenic mice of 3 groups of ages (3–4, 9–10, and 13 mstandard deviation. Statistical analyses are performed using a nonparamet
ompared with noninduced conditions as well as in com- n
arison with SY5Y mock cells (Fig. 2A). The overex-ression of tau did not therefore alter the endogenousxpression of Pin1 and we next analyzed Pin1 by 2D-gellectrophoresis. Pin1 was studied in SY5Y Mock andY5Y tau inducible cells lines (Fig. 2B). Five Pin1 iso-ariants (numbered from 0 to 4) were observed. In SY5Yau46 inducible cell line, without tetracycline, the 2Drofile was different. The most acidic isovariants 3 and 4ere slightly reduced. Following 24 hours of tetracycline
nduction, this reduction was even more pronounced,sovariant 4 was no more detected, and isovariant 3 wastrongly diminished. We calculated the staining intensityatio between isovariants 4, 3, and 2 (Fig. 2B, �-group of
spots) and isovariants 1 and 0 (Fig. 2B, �-group of spots).esults are illustrated by the histogram on Fig. 2Bnd statistical analyses of 3 individual experiments wereerformed using a nonparametric Mann–Whitney U test.e observed a significant reduction of the ratio when
au expression was induced (Fig. 2B), suggesting that tauverexpression was associated to a modified pattern ofin1 isovariants and the disappearance of the most acidic
au22 brains. Hippocampal brain lysates from WT and THY-Tau22 miceof Pin1. The ratio between the 2 most basic Pin1 isovariants (�-group:calculated. This analysis was performed in WT (n � 3) and THY-Tau22of age). Results are reported on histogram and expressed as the mean �n–Whitney U test.
THY-Tlotting) wereonths
ot attributed to an overall loss of Pin1 expression

eamahPpo3epowc
smmaistftcompm
3
bwTbbbhf2sP
(icdib
Sswsptw
8 K. Ando et al. / Neurobiology of Aging xx (2012) xxx
but rather due to a change of posttranslational modifica-tions.
3.3. Change of Pin1 posttranslational modifications inTHY-Tau22 transgenic mouse model
Overexpression of tau in SY5Y cells induced a mod-ified pattern of posttranslational modifications of Pin1. InNFT, tau accumulates to form fibrillar aggregates madeof hyper- and abnormally phosphorylated tau proteins.We developed a transgenic mouse model of hippocampalneurofibrillary degeneration, named THY-Tau22 (Belarbit al., 2009; Schindowski et al., 2006; Van der Jeugd etl., 2011). We studied Pin1 in this transgenic mouseodel that overexpresses the Human 4 repeat tau mutated
t G272V and P301S (numbering according to the longestuman isoform of tau). We first analyzed endogenousin1 expression levels in the tau transgenic model com-ared with those of control littermates. No difference wasbserved between wild type and THY-Tau22 mice (Fig.) showing that overexpression of tau did not alter thendogenous Pin1 expression. Noteworthy, tau was highlyhosphorylated at Thr231 in THY-Tau22 mice thus dem-nstrating that the amount of tau phosphothreonine 231as increased in transgenic mice when compared with
ontrol littermates (Fig. 3).We next analyzed endogenous Pin1 2D-gel pattern in
THY-Tau22 of growing ages (Fig. 4). Pin1 2D profile con-isted of the 5 isovariants showing that the posttranslationalodification of Pin1 is homogeneous among the presentodels. When comparing wild type and THY-Tau22 mice,reduced number of isovariants was significantly observed
n the latter. Noticeably, the most acidic isovariants (Fig. 4,pot 4 and 3) were reduced in THY-Tau22 mice. With age,his difference was more noticeable and extended to theollowing less acidic isovariant 3, which was almost unde-ectable at 13 months. This was not due to an aging-asso-iated mechanism because this loss of isovariants was notbserved in wild type mice with aging. Posttranslationalodifications of Pin1 are modified in correlation with the
resence of tau pathology in our THY-Tau22 transgenicouse model.
.4. Pin1 in AD brains
As Pin1 posttranslational modifications are reduced inoth SY5YTau46 and the THY-Tau22 transgenic model,e analyzed the 2D-profile of Pin1 in human AD brains.o examine the quantity of Pin1 in normal and Alzheimerrain samples, we first performed 1-dimensional Westernlot analysis on cortical tissues from control and ADrains. As shown previously, Pin1 levels are slightlyeterogeneous among samples but not significantly dif-erent between controls and AD brains (Hamdane et al.,002; Lu et al., 1999a; Wang et al., 2007) (Fig. 5). Weelected samples with a well-preserved expression of
in1 to further analyze Pin1 by 2D-gel electrophoresis iFig. 6). Interestingly, human brain Pin1 consisted of 5sovariants, showing that Pin1 posttranslational modifi-ations are not dramatically modified by the postmortemelay. In AD brain, we observed that the most acidicsovariants 3 and 4 were strongly reduced. The ratioetween �- and �-group of spots was significantly dimin-
ished in AD when compared with controls (Fig. 6A and6B). As previously shown, tau is hyperphosphorylated inAD brain samples and strongly stained with antibodiesagainst phosphothreonine 231 (Spillantini et al., 1996;Supplementary Fig. 4). Together, our results thereforesuggested that Pin1 posttranslational modifications arereduced in Alzheimer disease.
3.5. Enhanced phosphorylation of Pin1 at serine 16 inAD brain
The characterization of Pin1 posttranslational modifica-tions suggest that the 3 most acidic isovariants correspondwith phosphorylated variants. We therefore studied thephosphorylation status of Pin1 at the serine 16 (Fig. 7).
urprisingly, the ratio between phosphoserine 16 and Pin1taining was significantly increased in AD when comparedith controls (Fig. 7A). To further analyze this phospho-
ite, 2D isovariants of Pin1 were stained with Pin1 phos-hoserine 16 Pin1-antibody. In the control brain tissue, onlyhe 3 most acidic Pin1 isovariants 3, 4, and 5 were stainedith the phosphorylated-serine 16 Pin1 antibody. The same
Fig. 5. Pin1 expression in Alzheimer’s disease. Representative Westernblotting of brain lysates from control (Ctrl) and Alzheimer (AD) corticaltissue probed with anti-�-actin monoclonal antibody and anti-Pin1 poly-clonal antibody. The graphs show the ratios of the densitometry valuesmeasured for Pin1 and �-actin for 10 Ctrl and 11 AD patients. The valueof 100% was given to the average ratio of the control subjects.
sovariants were detected in AD brain tissue but the signal

sAPpp
TPmatueosrtbftt
tRemdpfsmsato
4
ccPmitpierp
cautnpsvOipttTdtte
tsptaP
dtwrd
9K. Ando et al. / Neurobiology of Aging xx (2012) xxx
was stronger whereas these isovariants 3, 4, and 5 were inlower amount in AD brain (Fig. 7B). Our data thereforeuggest that disappearances of acidic isovariants of Pin1 inD are likely not related to a reduced phosphorylation ofin1, which at opposite, is rather enhanced at serine-16hospho-site. Other posttranslational modifications may ex-lain this disappearance.
Pin1 has been shown to colocalize with granular vesicle.his localization may modify the biochemical properties ofin1, which should be found in insoluble fraction as otherarkers such as tau and A� (Sergeant et al., 2003b). To
ddress this hypothesis, human brain was processed in ordero obtain different fractions reflecting the progressive insol-bility of proteins (Supplementary Fig. 5). Thus, by differ-ntial solubility centrifugations, the most insoluble materialf AD brain tissue is concentrated in the last pellet. Ashown on Supplementary Fig. 1, insoluble hyperphospho-ylated tau and A� peptide is visualized in this pellet ma-erial (Supplementary Fig. 5, pellet lane). In contrast, solu-le tau is mainly found in the Triton X-100 and the SDSractions, both in control and Alzheimer brain tissue ex-racts. In the control brain, tau protein is not detected with
Fig. 6. Two-dimensional (2D) gel analysis of Pin1 in Alzheimer’s disease(AD). (A) Cortical brain tissue lysates from controls (n � 8) and ADpatients (n � 8) were resolved by 2D-gel electrophoresis and Pin1 wasetected by Western blotting. The 5 isovariants of Pin1 are numbered fromhe native form to the most acidic from 0 to 4. The apparent moleculareight and pI are indicated on the y- and x-axis of the 2D Western blots,
espectively. (B) The volume and intensity of each spot was calculated byensitometry and the ratio between the �- and �-group was determined.
The means � standard deviations are represented on the histogram and anonparametric Mann–Whitney U test was performed between the 2 groups.
he phospho-dependent antibodies against phosphorylated a
hreonine 231 and phosphorylated serines 396 and 404.egarding the distribution of Pin1, we observed that Pin1 isssentially found in the SDS and pellet fractions (Supple-entary Fig. 5, SDS and pellet lanes). There was no obvious
ifference in between control and AD. In sharp contrast,hosphorylated-Pin1 at serine 16 was found in the pelletraction in AD brain tissue extract but not in the control,uggesting that Pin1 phosphorylated at serine 16 cosedi-ented with the most insoluble fraction. Together our data
uggest that the most phosphorylated fraction of Pin1 isssociated with the most insoluble fraction of AD brainissue extracts relevant to the granulovacuolar localizationf Pin1 in AD.
. Discussion
Herein, we report that Pin1 posttranslational modifi-ations are altered in response to tau overexpression in aell system, in THY-Tau22 mouse model, and in AD.in1 is known to be a modulator of tau metabolism anday contribute to the pathophysiology of tauopathies
ncluding AD. Our data suggest that Pin1 posttransla-ional modifications are deregulated in response to tauathology. This specific pattern of posttranslational mod-fications gives to Pin1 a signature that may be consid-red as a biomarker of tau pathology. Finally, it may alsoeflect a dysfunction of Pin1 in Alzheimer’s disease andossibly in other tauopathies.
Posttranslational modifications are postgenomic modifi-ations that contribute to the diversity of the proteome butre also strong modulators of protein functions. Herein, wesed 2D-gel electrophoresis and mass spectrometry to de-ermine Pin1 posttranslational modifications that occur in aeuroblastoma inducible cell system. First of all, the 2Drofile of Pin1 when overexpressed in SY5Y inducibleystem is very similar to the endogenous protein but alsoery similar to that observed in the mouse and human brain.ur data strongly suggest that Pin1 posttranslational mod-
fications are similar among the systems used. Moreover,ostmortem delay does not profoundly modify these post-ranslational modifications because Pin1 profile is globallyhe same between mouse and postmortem human brains.he analysis of Pin1 is observed without any postmortemelay in mice. Together, we suggest that the characteriza-ion of Pin1 by 2D pattern and posttranslational modifica-ions characterized in SY5Y Pin1-inducible cells can bextrapolated to the experimental conditions herein analyzed.
Because Pin1 has at least 4 major isovariants in additiono the native polypeptide, this means that Pin1 has 4 (pos-ibly more) posttranslational modifications including phos-horylation at 3 sites (Ser16 and Ser65/Ser71), N-acetyla-ion (amino-terminus and Lys46) and oxidation (Met130nd 146). Regarding phosphorylation, our data suggest thatin1 can be phosphorylated on 3 sites. By inhibiting the
ctivity of phosphatases with okadaic acid, we expect to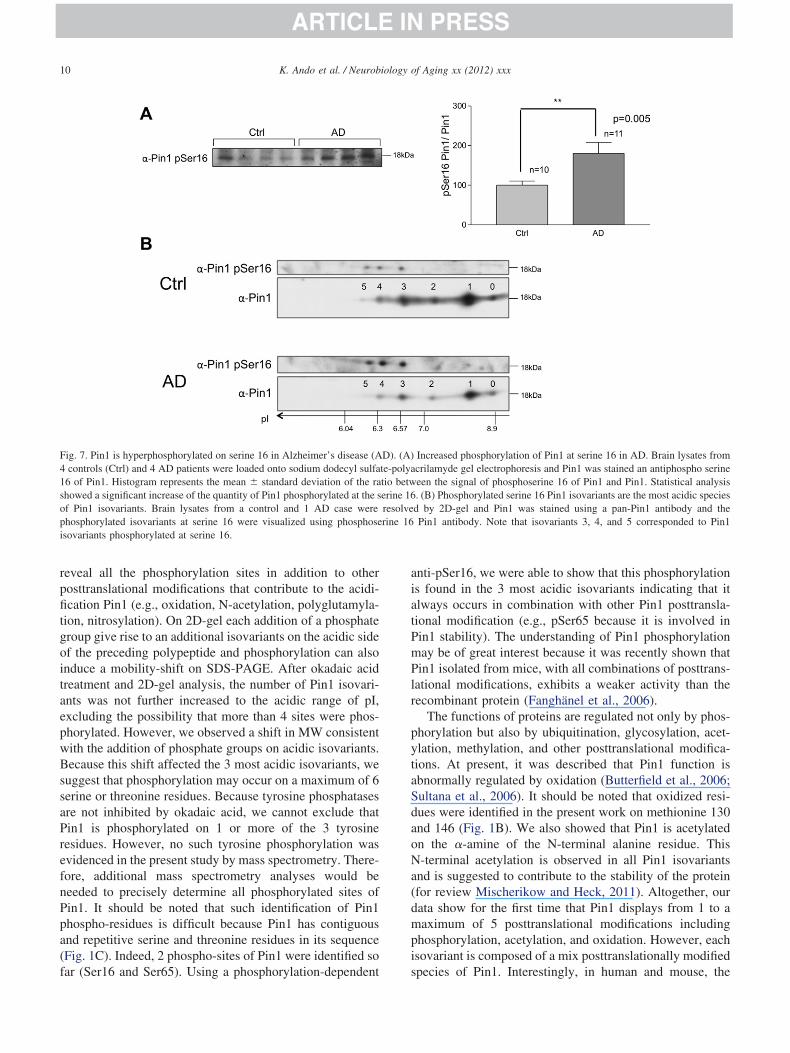
10 K. Ando et al. / Neurobiology of Aging xx (2012) xxx
reveal all the phosphorylation sites in addition to otherposttranslational modifications that contribute to the acidi-fication Pin1 (e.g., oxidation, N-acetylation, polyglutamyla-tion, nitrosylation). On 2D-gel each addition of a phosphategroup give rise to an additional isovariants on the acidic sideof the preceding polypeptide and phosphorylation can alsoinduce a mobility-shift on SDS-PAGE. After okadaic acidtreatment and 2D-gel analysis, the number of Pin1 isovari-ants was not further increased to the acidic range of pI,excluding the possibility that more than 4 sites were phos-phorylated. However, we observed a shift in MW consistentwith the addition of phosphate groups on acidic isovariants.Because this shift affected the 3 most acidic isovariants, wesuggest that phosphorylation may occur on a maximum of 6serine or threonine residues. Because tyrosine phosphatasesare not inhibited by okadaic acid, we cannot exclude thatPin1 is phosphorylated on 1 or more of the 3 tyrosineresidues. However, no such tyrosine phosphorylation wasevidenced in the present study by mass spectrometry. There-fore, additional mass spectrometry analyses would beneeded to precisely determine all phosphorylated sites ofPin1. It should be noted that such identification of Pin1phospho-residues is difficult because Pin1 has contiguousand repetitive serine and threonine residues in its sequence(Fig. 1C). Indeed, 2 phospho-sites of Pin1 were identified so
Fig. 7. Pin1 is hyperphosphorylated on serine 16 in Alzheimer’s disease (A4 controls (Ctrl) and 4 AD patients were loaded onto sodium dodecyl sulfa16 of Pin1. Histogram represents the mean � standard deviation of the rashowed a significant increase of the quantity of Pin1 phosphorylated at the sof Pin1 isovariants. Brain lysates from a control and 1 AD case werephosphorylated isovariants at serine 16 were visualized using phosphoseisovariants phosphorylated at serine 16.
far (Ser16 and Ser65). Using a phosphorylation-dependent
anti-pSer16, we were able to show that this phosphorylationis found in the 3 most acidic isovariants indicating that italways occurs in combination with other Pin1 posttransla-tional modification (e.g., pSer65 because it is involved inPin1 stability). The understanding of Pin1 phosphorylationmay be of great interest because it was recently shown thatPin1 isolated from mice, with all combinations of posttrans-lational modifications, exhibits a weaker activity than therecombinant protein (Fanghänel et al., 2006).
The functions of proteins are regulated not only by phos-phorylation but also by ubiquitination, glycosylation, acet-ylation, methylation, and other posttranslational modifica-tions. At present, it was described that Pin1 function isabnormally regulated by oxidation (Butterfield et al., 2006;Sultana et al., 2006). It should be noted that oxidized resi-dues were identified in the present work on methionine 130and 146 (Fig. 1B). We also showed that Pin1 is acetylatedon the �-amine of the N-terminal alanine residue. ThisN-terminal acetylation is observed in all Pin1 isovariantsand is suggested to contribute to the stability of the protein(for review Mischerikow and Heck, 2011). Altogether, ourdata show for the first time that Pin1 displays from 1 to amaximum of 5 posttranslational modifications includingphosphorylation, acetylation, and oxidation. However, eachisovariant is composed of a mix posttranslationally modified
Increased phosphorylation of Pin1 at serine 16 in AD. Brain lysates fromcrilamyde gel electrophoresis and Pin1 was stained an antiphospho serineeen the signal of phosphoserine 16 of Pin1 and Pin1. Statistical analysis. (B) Phosphorylated serine 16 Pin1 isovariants are the most acidic speciesd by 2D-gel and Pin1 was stained using a pan-Pin1 antibody and thePin1 antibody. Note that isovariants 3, 4, and 5 corresponded to Pin1
D). (A)te-polyatio betwerine 16resolverine 16
species of Pin1. Interestingly, in human and mouse, the

tNtn1
11K. Ando et al. / Neurobiology of Aging xx (2012) xxx
mono- and trimodified Pin1 species were the most abundantisovariants among the 5.
In all experimental conditions, including tau-overex-pressing cells, tau transgenic mice and AD brains, globallevels of Pin1 posttranslational modifications were de-creased compared with control conditions. Our data showhere for the first time that Pin1 posttranslational modifica-tions are altered by the overexpression of tau or the presenceof tau pathology. Thus, a reciprocal regulation may existbetween tau and Pin1. Pin1 binds to phosphorylated Thr231of tau and facilitates the dephosphorylation of phospho-Thr231 through isomerization (Galas et al., 2006; Hamdaneet al., 2006; Lu et al., 1999a). Phosphorylation at Thr231 onau is associated with the early events of tau aggregation andFT (Augustinack et al., 2002). Pin1 binds and isomerizes
he proline imidic peptide bond following the phosphothreo-ine 231 (Galas et al., 2006; Hamdane et al., 2006; Lu et al.,999a; Smet et al., 2004; Wintjens et al., 2001). In AD
brains, tau proteins are hyperphosphorylated and aggregatedinto an insoluble fibrous structure, referred to as pairedhelical filament. It was discussed in several reports thatabundant phosphorylated tau and paired helical filament inthe cytoplasm may pull out Pin1 from the nucleus to thecytoplasm in human brains (Lu et al., 1999a; Ramakrishnanet al., 2003; Thorpe et al., 2001, 2004), implying that tauamount in the cytoplasm can actually alter the localizationof Pin1 in a certain manner. Nevertheless, we recentlydemonstrated in human neuroblastoma cells that overex-pression of phospho-tau does not modify Pin1 cellular dis-tribution in the nucleus or the cytosol (Hamdane et al.,2006). In fact, phosphorylation of Pin1 at Ser16 inhibits itsnuclear localization possibly through inhibition of Pin1 sub-strate-binding property (Lu et al., 2002) while phosphory-lation at Ser65 does not change activity of localization butstabilizes Pin1 by preventing its ubiquitination (Eckerdt etal., 2005). Therefore, phosphorylation of Pin1 must be a keyfactor for regulating its localization, function, and metabo-lism. In this report, we have found, for the first time, that tauis involved in the regulation of balance between phospho-rylation-dephosphorylation of Pin1. Interestingly, in ADpatients, we observed a hyperphosphorylation of Pin1 onserine 16 suggesting that dysfunction of Pin1 metabolismmay occur through phosphorylation, in part. However, wedo not yet know if serine-16 phosphorylation is associatedwith the granulovesicular localization of Pin1 in AD. Nev-ertheless, the hyperphosphorylation of Pin1 at serine 16may certainly reduce its affinity for its substrate includingtau. Our results suggest that a fraction of Pin1 is inefficientto isomerase its substrates and therefore to promote taudephosphorylation. However, all isovariants are not phos-phorylated at serine 16 and a little proportion may also beassociated with hyperphosphorylated tau or associated withthe most insoluble fraction of AD brain tissue as shownherein.
In conclusion, the dysregulation of Pin1 is crucial in AD
etiopathogenesis and strongly linked to the tau pathology.Such changes in Pin1 posttranslational modifications mayalso represent interesting biomarkers to follow the severityof AD and tauopathies (Fig. 7).
Disclosure statement
The authors disclose no conflicts of interest.All protocols followed European animal welfare regula-
tions and were approved by the local ethical committee(Approval n° AF 06/2010, 31/3/2010). All the human brainautopsy materials used in this study were from the LilleNeurobank (Regional Hospital Centre and University ofLille). Tissue was removed according to Local Ethics Com-mittee guidelines, and informed consent for brain donationwas obtained from next of kin.
Acknowledgements
This work was supported by Inserm, CNRS, IMPRT,University of Lille, Nord de France, Lille County Hos-pital (CHRU-Lille), Région Nord/Pas-de-Calais, FEDER,MEDIALZ, Labex (excellence laboratory) DISTALZ (De-velopment of Innovative Strategies for a Transdisciplinaryapproach to ALZheimer’s disease), ANR-05-BLANC-0320-01, France Alzheimer, GIS-Longévité, LECMA, Fé-dération pour la Recherche sur le Cerveau, France Alzhei-mer, and grants from the European Community: cNEUPRO(FP6 contract LSHM-CT-2007-037950) and MEMOSAD(FP7 contract). P.D. and K.A. are recipients of a scholarshipfrom the French Research Ministry and the French ForeignAffairs Ministry respectively, and A.B. and K.B. recipientsfrom a scholarship cosponsored by both Région Nord/Pas-de-Calais and CHR-Lille.
Appendix A. Supplementary data
Supplementary data associated with this article can befound, in the online version, at http://dx.doi.org/10.1016/j.neurobiolaging.2012.08.004.
References
Augustinack, J.C., Schneider, A., Mandelkow, E.M., Hyman, B.T., 2002.Specific tau phosphorylation sites correlate with severity of neuronalcytopathology in Alzheimer’s disease. Acta Neuropathol. 103, 26–35.
Belarbi, K., Schindowski, K., Burnouf, S., Caillierez, R., Grosjean, M.E.,Demeyer, D., Hamdane, M., Sergeant, N., Blum, D., Buée, L., 2009.Early Tau pathology involving the septo-hippocampal pathway in aTau transgenic model: relevance to Alzheimer’s disease. Curr. Alzhei-mer Res. 6, 152–157.
Bretteville, A., Ando, K., Ghestem, A., Loyens, A., Bégard, S., Beauvil-lain, J.C., Sergeant, N., Hamdane, M., Buée, L., 2009. Two-dimen-sional electrophoresis of Tau mutants reveals specific phosphorylationpattern likely linked to early Tau conformational changes. PLoS One 4,e4843.
Bussière, T., Hof, P.R., Mailliot, C., Brown, C.D., Caillet-Boudin, M.L.,
Perl, D.P., Buée, L., Delacourte, A., 1999. Phosphorylated serine422 on
D
E
F
G
H
H
H
L
L
L
L
L
L
L
L
M
N
P
R
R
S
S
S
S
S
S
T
T
12 K. Ando et al. / Neurobiology of Aging xx (2012) xxx
Tau proteins is a pathological epitope found in several diseases withneurofibrillary degeneration. Acta Neuropathol. 97, 221–230.
Butterfield, D.A., Poon, H.F., St Clair, D., Keller, J.N., Pierce, W.M.,Klein, J.B., Markesbery, W.R., 2006. Redox proteomics identificationof oxidatively modified hippocampal proteins in mild cognitive impair-ment: insights into the development of Alzheimer’s disease. Neurobiol.Dis. 22, 223–232.
Choudhary, C., Kumar, C., Gnad, F., Nielsen, M.L., Rehman, M., Walther,T.C., Olsen, J.V., Mann, M., 2009. Lysine acetylation targets proteincomplexes and co-regulates major cellular functions. Science 325,834–840.
Dakson, A., Yokota, O., Esiri, M., Bigio, E.H., Horan, M., Pendleton, N.,Richardson, A., Neary, D., Snowden, J.S., Robinson, A., Davidson,Y.S., Mann, D.M., 2011. Granular expression of prolyl-peptidylisomerase PIN1 is a constant and specific feature of Alzheimer’sdisease pathology and is independent of tau, A� and TDP-43 pathol-ogy. Acta Neuropathol. 121, 635–649.
elacourte, A., Sergeant, N., Champain, D., Wattez, A., Maurage, C.A.,Lebert, F., Pasquier, F., David, J.P., 2002. Nonoverlapping but syner-getic tau and APP pathologies in sporadic Alzheimer’s disease. Neu-rology. 59, 398–407.
ckerdt, F., Yuan, J., Saxena, K., Martin, B., Kappel, S., Lindenau, C.,Kramer, A., Naumann, S., Daum, S., Fischer, G., Dikic, I., Kaufmann,M., Strebhardt, K., 2005. Polo-like kinase 1-mediated phosphorylationstabilizes Pin1 by inhibiting its ubiquitination in human cells. J. Biol.Chem. 280, 36575–36583.
anghänel, J., Akiyama, H., Uchida, C., Uchida, T., 2006. Comparativeanalysis of enzyme activities and mRNA levels of peptidyl prolylcis/trans isomerases in various organs of wild type and Pin1-/- mice.FEBS Lett. 580, 3237–3245.
alas, M.C., Dourlen, P., Bégard, S., Ando, K., Blum, D., Hamdane, M.,Buée, L., 2006. The peptidylprolyl cis/trans-isomerase Pin1 modulatesstress-induced dephosphorylation of Tau in neurons. Implication in apathological mechanism related to Alzheimer disease. J. Biol. Chem.281, 19296–19304.
amdane, M., Dourlen, P., Bretteville, A., Sambo, A.V., Ferreira, S.,Ando, K., Kerdraon, O., Bégard, S., Geay, L., Lippens, G., Sergeant,N., Delacourte, A., Maurage, C.A., Galas, M.C., Buée, L., 2006. Pin1allows for differential Tau dephosphorylation in neuronal cells. Mol.Cell. Neurosci. 32, 155–160.
amdane, M., Smet, C., Sambo, A.V., Leroy, A., Wieruszeski, J.M.,Delobel, P., Maurage, C.A., Ghestem, A., Wintjens, R., Bégard, S.,Sergeant, N., Delacourte, A., Horvath, D., Landrieu, I., Lippens, G.,Buée, L., 2002. Pin1: a therapeutic target in Alzheimer neurodegen-eration. J. Mol. Neurosci. 19, 275–287.
olzer, M., Gärtner, U., Stöbe, A., Härtig, W., Gruschka, H., Brückner,M.K., Arendt, T., 2002. Inverse association of Pin1 and Tau accumu-lation in Alzheimer’s disease hippocampus. Acta Neuropathol. 104,471–481.
andrieu, I., Smet, C., Wieruszeski, J.M., Sambo, A.V., Wintjens, R.,Buée, L., Lippens, G., 2006. Exploring the molecular function of PIN1by nuclear magnetic resonance. Curr. Protein Pept. Sci. 7, 179–194.
ee, T.H., Chen, C.H., Suizu, F., Huang, P., Schiene-Fischer, C., Daum, S.,Zhang, Y.J., Goate, A., Chen, R.H., Zhou, X.Z., Lu, K.P., 2011.Death-associated protein kinase 1 phosphorylates pin1 and inhibits itsprolyl isomerase activity and cellular function. Mol. Cell 42, 147–159.
efebvre, T., Ferreira, S., Dupont-Wallois, L., Bussière, T., Dupire, M.J.,Delacourte, A., Michalski, J.C., Caillet-Boudin, M.L., 2003. Evidenceof a balance between phosphorylation and O-GlcNAc glycosylation ofTau proteins—a role in nuclear localization. Biochim. Biophys. Acta1619, 167–176.
eroy, A., Landrieu, I., Huvent, I., Legrand, D., Codeville, B., Wierusz-eski, J.M., Lippens, G., 2010. Spectroscopic studies of GSK3{beta}phosphorylation of the neuronal tau protein and its interaction withthe N-terminal domain of apolipoprotein E. J. Biol. Chem. 285,
33435–33444.iou, Y.C., Sun, A., Ryo, A., Zhou, X.Z., Yu, Z.X., Huang, H.K., Uchida,T., Bronson, R., Bing, G., Li, X., Hunter, T., Lu, K.P., 2003. Role ofthe prolyl isomerase Pin1 in protecting against age-dependent neuro-degeneration. Nature 424, 556–561.
u, P.J., Wulf, G., Zhou, X.Z., Davies, P., Lu, K.P., 1999a. The prolylisomerase Pin1 restores the function of Alzheimer-associated phos-phorylated Tau protein. Nature 399, 784–788.
u, P.J., Zhou, X.Z., Liou, Y.C., Noel, J.P., Lu, K.P., 2002. Critical role ofWW domain phosphorylation in regulating phosphoserine binding ac-tivity and Pin1 function. J. Biol. Chem. 277, 2381–2384.
u, P.J., Zhou, X.Z., Shen, M., Lu, K.P., 1999b. Function of WW domainsas phosphoserine- or phosphothreonine-binding modules. Science 283,1325–1328.
ischerikow, N., Heck, A.J., 2011. Targeted large-scale analysis of proteinacetylation. Proteomics 11, 571–589.
ishi, M., Akutsu, H., Masui, S., Kondo, A., Nagashima, Y., Kimura, H.,Perrem, K., Shigeri, Y., Toyoda, M., Okayama, A., Hirano, H.,Umezawa, A., Yamamoto, N., Lee, S.W., Ryo, A., 2011. A distinct rolefor pin1 in the induction and maintenance of pluripotency. J. Biol.Chem. 286, 11593–11603.
astorino, L., Sun, A., Lu, P.J., Zhou, X.Z., Balastik, M., Finn, G., Wulf,G., Lim, J., Li, S.H., Li, X., Xia, W., Nicholson, L.K., Lu, K.P., 2006.The prolyl isomerase Pin1 regulates amyloid precursor protein process-ing and amyloid-beta production. Nature 440, 528–534.
amakrishnan, P., Dickson, D.W., Davies, P., 2003. Pin1 colocalizationwith phosphorylated Tau in Alzheimer’s disease and other Tauopathies.Neurobiol. Dis. 14, 251–264.
udrabhatla, P., Pant, H.C., 2010. Phosphorylation-specific peptidyl-prolylisomerization of neuronal cytoskeletal proteins by Pin1: implicationsfor therapeutics in neurodegeneration. J. Alzheimers Dis. 19, 389–403.
chindowski, K., Bretteville, A., Leroy, K., Bégard, S., Brion, J.P.,Hamdane, M., Buée, L., 2006. Alzheimer’s disease-like Tau neuro-pathology leads to memory deficits and loss of functional synapsesin a novel mutated Tau transgenic mouse without any motor deficits.Am. J. Pathol. 169, 599 – 616.
ergeant, N., Bombois, S., Ghestem, A., Drobecq, H., Kostanjevecki, V.,Missiaen, C., Wattez, A., David, J.P., Vanmechelen, E., Sergheraert,C., Delacourte, A., 2003a. Truncated beta-amyloid peptide species inpre-clinical Alzheimer’s disease as new targets for the vaccinationapproach. J. Neurochem. 85, 1581–1591.
ergeant, N., Wattez, A., Galván-valencia, M., Ghestem, A., David, J.P.,Lemoine, J., Sautiére, P.E., Dachary, J., Mazat, J.P., Michalski, J.C.,Velours, J., Mena-López, R., Delacourte, A., 2003b. Association ofATP synthase alpha-chain with neurofibrillary degeneration in Alzhei-mer’s disease. Neuroscience 117, 293–303.
met, C., Sambo, A.V., Wieruszeski, J.M., Leroy, A., Landrieu, I., Buée,L., Lippens, G., 2004. The peptidyl prolyl cis/trans-isomerase Pin1recognizes the phospho-Thr212-Pro213 site on Tau. Biochemistry 43,2032–2040.
pillantini, M.G., Crowther, R.A., Goedert, M., 1996. Comparison of theneurofibrillary pathology in Alzheimer’s disease and familial preseniledementia with tangles. Acta Neuropathol. 92, 42–48.
ultana, R., Boyd-Kimball, D., Poon, H.F., Cai, J., Pierce, W.M., Klein, J.B.,Markesbery, W.R., Zhou, X.Z., Lu, K.P., Butterfield, D.A., 2006. Oxida-tive modification and down-regulation of Pin1 in Alzheimer’s diseasehippocampus: a redox proteomics analysis. Neurobiol. Aging 27, 918–925.
horpe, J.R., Morley, S.J., Rulten, S.L., 2001. Utilizing the peptidyl-prolyl cis-trans isomerase Pin1 as a probe of its phosphorylatedtarget proteins. Examples of binding to nuclear proteins in a humankidney cell line and to Tau in Alzheimer’s diseased brain. J. His-tochem. Cytochem. 49, 97–108.
horpe, J.R., Mosaheb, S., Hashemzadeh-Bonehi, L., Cairns, N.J., Kay,
J.E., Morley, S.J., Rulten, S.L., 2004. Shortfalls in the peptidyl-prolyl
13K. Ando et al. / Neurobiology of Aging xx (2012) xxx
cis-trans isomerase protein Pin1 in neurons are associated withfrontotemporal dementias. Neurobiol. Dis. 17, 237–249.
Van der Jeugd, A., Ahmed, T., Burnouf, S., Belarbi, K., Hamdame, M.,Grosjean, M.E., Humez, S., Balschun, D., Blum, D., Buée, L.,D’Hooge, R., 2011. Hippocampal tauopathy in tau transgenic micecoincides with impaired hippocampus-dependent learning and mem-ory, and attenuated late-phase long-term depression of synaptic trans-mission. Neurobiol. Learn. Mem. 95, 296–304.
Vingtdeux, V., Hamdane, M., Gompel, M., Bégard, S., Drobecq, H.,Ghestem, A., Grosjean, M.E., Kostanjevecki, V., Grognet, P., Van-mechelen, E., Buée, L., Delacourte, A., Sergeant, N., 2005. Phosphor-ylation of amyloid precursor carboxy-terminal fragments enhancestheir processing by a gamma-secretase-dependent mechanism. Neuro-
biol. Dis. 20, 625–637.Wang, S., Simon, B.P., Bennett, D.A., Schneider, J.A., Malter, J.S., Wang,D.S., 2007. The significance of Pin1 in the development of Alzheimer’sdisease. J. Alzheimers Dis. 11, 13–23.
Wintjens, R., Wieruszeski, J.M., Drobecq, H., Rousselot-Pailley, P., Buée,L., Lippens, G., Landrieu, I., 2001. 1H NMR study on the binding ofPin1 Trp-Trp domain with phosphothreonine peptides. J. Biol. Chem.276, 25150–25156.
Wulf, G., Finn, G., Suizu, F., Lu, K.P., 2005. Phosphorylation-specificprolyl isomerization: is there an underlying theme? Nat. Cell Biol. 7,435–441.
Zhou, X.Z., Kops, O., Werner, A., Lu, P.J., Shen, M., Stoller, G., Küllertz,G., Stark, M., Fischer, G., Lu, K.P., 2000. Pin1-dependent prolylisomerization regulates dephosphorylation of Cdc25C and tau proteins.
Mol. Cell 6, 873–883.Copyright © 2022 FDOKUMEN




















