
Characterization of a chaperone ClpB homologue of Paracoccidioides brasiliensis
Upload
independentCategory
view
1download
0
Target recognition RNA methylation activity andtranscriptional regulation of the Dictyosteliumdiscoideum Dnmt2-homologue (DnmA)Sara Muller1 Indra M Windhof1 Vladimir Maximov1 Tomasz Jurkowski2
Albert Jeltsch2 Konrad U Forstner3 Cynthia M Sharma3 Ralph Graf4 and
Wolfgang Nellen1
1Department of Genetics University of Kassel Heinrich-Plett-Str 40 34132 Kassel Germany 2Institute ofBiochemistry University Stuttgart Pfaffenwaldring 55 70569 Stuttgart Germany 3Research Center forInfectious Diseases (ZINF) University of Wurzburg Josef-Schneider-Str 2Bau D15 97080 Wurzburg and4Universitat Potsdam Institut fur Biochemie und Biologie Abt Zellbiologie Karl-Liebknecht-Strasse 24-2514476 Potsdam ndash Golm
Received December 12 2012 Revised June 24 2013 Accepted June 26 2013
ABSTRACT
Although the DNA methyltransferase 2 family ishighly conserved during evolution and recentreports suggested a dual specificity with strongeractivity on transfer RNA (tRNA) than DNA sub-strates the biological function is still obscure Weshow that the Dictyostelium discoideum Dnmt2-homologue DnmA is an active tRNA methyltrans-ferase that modifies C38 in tRNAAsp(GUC) in vitroand in vivo By an ultraviolet-crosslinking andimmunoprecipitation approach we identifiedfurther DnmA targets This revealed specific tRNAfragments bound by the enzyme and identifiedtRNAGlu(CUCUUC) and tRNAGly(GCC) as new butweaker substrates for both human Dnmt2 andDnmA in vitro but apparently not in vivo Dnmt2enzymes form transient covalent complexes withtheir substrates The dynamics of complex forma-tion and complex resolution reflect methylation effi-ciency in vitro Quantitative PCR analyses revealedalterations in dnmA expression during developmentcell cycle and in response to temperature stressHowever dnmA expression only partially correlatedwith tRNA methylation in vivo Strikingly dnmAexpression in the laboratory strain AX2 was sig-nificantly lower than in the NC4 parent strain
As expression levels and binding of DnmA to atarget in vivo are apparently not necessarilyaccompanied by methylation we propose an add-itional biological function of DnmA apart frommethylation
INTRODUCTION
Dnmt2 is a member of the eukaryotic DNAmethyltransferase family A few model organisms espe-cially Drosophila melanogaster Schizosaccharomycespombe Entamoeba histolytica and Dictyosteliumdiscoideum contain only one Dnmt2 homologue but lackthe more active homologues Dnmt1 and Dnmt3 Thoughhighly conserved during evolution loss of Dnmt2 homo-logues has no obvious phenotypic effects in Mus musculus(1) D melanogaster (2) S pombe (3) and D discoideum(45) In Danio rerio disruption of dnmt2 has beenreported to cause pleiotropic effects (6) and inE histolytica a gene disruption could not be obtainedsuggesting that Dnmt2 was required for viability (7) Oncloser inspection more subtle long-term effects of Dnmt2loss were observed in D melanogaster for exampleH4K20me3 was strongly reduced and telomeric sequenceswere lost (2) Durdevic et al (8) recently showed that inD melanogaster Dnmt2 was also involved in viruscontrol At least in D melanogaster E histolytica andD discoideum low amounts of DNA methylation
To whom correspondence should be addressed Tel +49 561 8044805 Fax +49 561 8044800 Email nellenuni-kasseldePresent addressesSara Muller Institute for Biochemistry and Molecular Cell Biology Georg-August-University Humboldtallee 23 37073 Gottingen Germany
The authors wish it to be known that in their opinion the first two authors should be regarded as joint First Authors
Vladimir Maximov Department of Plant Biology and Forest Genetics Uppsala BioCenter Swedish University of Agricultural Sciences PO-Box7080 SE-75007 Uppsala Sweden
Nucleic Acids Research 2013 1ndash13doi101093nargkt634
The Author(s) 2013 Published by Oxford University PressThis is an Open Access article distributed under the terms of the Creative Commons Attribution Non-Commercial License (httpcreativecommonsorglicensesby-nc30) which permits non-commercial re-use distribution and reproduction in any medium provided the original work is properly cited For commercialre-use please contact journalspermissionsoupcom
Nucleic Acids Research Advance Access published July 22 2013 at U
niversitaumlt Wuumlrzburg on A
ugust 5 2013httpnaroxfordjournalsorg
Dow
nloaded from
especially on retroelements were detected and attributedto Dnmt2 although these data are controversially dis-cussed (9ndash11)In 2006 Bestorrsquos group reported that Dnmt2 rather
methylates transfer RNA (tRNA) than DNA inM musculus D melanogaster and Arabidopsis thalianaSpecifically they found C38 in tRNAAsp as the major oronly substrate for the enzyme (1) A dual role of Dnmt2had been discussed (12) and Jurkowski et al (13) couldshow that tRNAAsp was methylated by an enzymaticmechanism characteristic for DNA methyltransferasesrather than by the reaction pathways of enzymes thatmethylate RNAstRNAAsp methylation activity was also reported for
the Dnmt2-homologues from E histolytica (14) andD melanogaster (15) Schaefer et al (15) showed thatD melanogaster tRNAVal(CAC) and tRNAGly(GCC) werealso methylated at position C38 in vivo and by humanDnmt2 (hDnmt2) in vitro They further found thatdnmt2 knockout flies were more sensitive to oxidativestress A more detailed analysis of tRNAAsp(GUC) andtRNAGly(GCC) suggested that methylation protectedtRNAs from stress-induced cleavage (15) Double-knockout mutant mice of Dnmt2 and Nsun2 the secondknown m5C-tRNA-methyltransferase in higher eukary-otes showed a phenotype with impaired cellular differen-tiation an overall reduction in protein synthesis and earlylethality (16) Recently we identified tRNAGlu(UUC) as anadditional novel substrate of Pmt1 the Dnmt2-homo-logue in S pombe (17) Pmt1-dependent tRNA methyla-tion seemed to be regulated by nutrient conditionsNutritional control was also reported for Ehmeth theDnmt2 homologue from E histolytica that is inhibitedby the glycolytic enzyme enolase (1418)Here we demonstrate that recombinant D discoideum
DnmA and hDnmt2 can methylate tRNAAsp(GUC)tRNAGlu(UUC) and tRNAGly(GCC) from D discoideumin vitro with different efficiencies Both enzymes formedcovalent complexes [22] with specific tRNAs with similarkinetics but they were significantly slower for the minorsubstrate tRNAGlu than for the major substrate tRNAAspUltraviolet (UV)-crosslinking and immunoprecipitation(CLIP) experiments showed that specific fragments ofthe three target tRNAs associated with DnmA in vivoHowever in vivo only methylation of tRNAAsp(GUC) byDnmA was confirmed whereas methylation of the targetnucleotide C38 in other substrates was not detectable bybisulfite sequencingAs revealed by quantitative reverse transcription (RT)-
PCR dnmA is differentially expressed in development cellcycle and in the recovery phase after temperature stressThe increase in dnmA expression levels correlated withelevated tRNAAsp methylation in development but notafter temperature stress The other targets identified byin vitro methylation and by CLIP were apparently alsonot methylated in vivo in developmentOur data document that additional RNA molecules can
serve as substrates for Dnmt2 binding and that the fullrange of targets is probably not yet recognized The resultsalso suggest that binding of the methyltransferase to an
RNA molecule not necessarily results in methylation butmay have different biological functions
MATERIALS AND METHODS
Dictyostelium discoideum cultures and nomenclature
Dictyostelium discoideum AX2-214 was grown in HL5+medium (ForMedium) containing 50 mgml Ampicillin025 mgml Amphotericin-B 100 mgml PenicillinStreptomycin (PAA) at 22C constant light under select-ive conditions as required NC4 cells were grown in a sus-pension of Klebsiella aerogenes in phosphate buffer Whenindicated AX2 cells were also grown in bacterial suspen-sion to allow for comparison with NC4
For cold treatment cells (1 106ml) were shaken at 4Cfor 2ndash14 h Cells were allowed to recover at 22C for 2frac12 hbefore RNA isolation For synchronization cells at adensity of 1ndash3 105ml were incubated at 4C overnight Before cold treatment cells were briefly cooleddown in a water bath with ice For synchronizationcells were then warmed up in a 25C water bath beforecultivating at 22C Synchronization was measured bycounting cells every 30 min during a period of at least7 h Samples for RNA isolation were taken at the timesindicated
Gene names and accession numbers refer to DictyBase(httpdictybaseorg Jan 3rd 2013) Complete sequencesfor tRNAs relevant for the experiments are given inSupplementary Table S2 In silico folded clover leafstructures for tRNAAsp(GUC) tRNAGlu(UUC) andtRNAGly(GCC) are given in Supplementary Figure S1
Nucleotides in tRNAs have been numbered accordingto Motorin et al (19)
Expression and purification of recombinant DnmA andhuman hDnmt2
DnmA and hDnmt2 were both cloned with N-terminalHis6-tags in pET28a(+) (Novagen) into the NdeI andXhoI sites Recombinant proteins were expressed inEscherichia coli (DE3) Rosetta pLysS cells Inductionand protein purification were done as described previously(13) Briefly the cells were induced at an OD600 of 06 with1mM IPTG and incubated for 3 h at 37C for hDnmt2and at 22C for DnmA After harvesting cells were resus-pended in sonication buffer [30 mM KPi (pH 70)300mM KCl 10 (vv) glycerol 10mM imidazole01mM Dithiotreitol (DTT)] containing protease inhibi-tors (complete mini Roche) and lysed by sonication with aDr Hielscher up200s sonifier (5ndash10 times cycle 05ampli-tude 50) The cell lysate was cleared by centrifugation(10000 RCF) and the supernatant was incubated onNiNTA-Sepharose for 15min at 4C After washing withsonication buffer His6-tagged proteins were eluted with abuffer containing 30mM KPi (pH 70) 300mM KCl10 glycerol 200mM imidazole and 01mM DTT Theeluate was dialysed in two steps [30mM KPi (pH 70)200mM KCl 20 glycerol 1mM DTT 01mM EDTAand 30mM KPi (pH 70) 100mM KCl 50 glycerol1mM DTT 01mM EDTA]
2 Nucleic Acids Research 2013
at Universitaumlt W
uumlrzburg on August 5 2013
httpnaroxfordjournalsorgD
ownloaded from
UV-crosslinking and RNA immunoprecipitation
For UV CLIP 2 108 D discoideum cells expressingDnmA-GFP in the dnmAKO-background and cells ex-pressing GFP only were harvested (390 RCF 5min)washed once and resuspended in 20ml of phosphatebuffer Ten microlitre aliquots of the suspension werespread in 10 cm petri dishes and UV irradiated (254 nm)with 250mJcm2 Cells were spun down and collected in4ml of modified RIPA-Buffer [50mM TrisHCl (pH 75)150mM NaCl 05 NP40 05 sodium deoxycholate1mM EDTA] (20) Lysis was supported by three timessonication (cycle 05 and amplitude 50) with a DrHielscher up200s sonifier Samples were centrifuged(20 000 RCF 15min 4C) and the supernatant wastransferred to a fresh tube it was again centrifuged andthe supernatant collected for a third centrifugation stepHereafter the supernatant was pre-incubated with 250 mlSephadexTM G-50 Medium (GE Healthcare) and thentransferred to 50 ml GFP-trap beads (ChromoTekMartinsried) (21) Incubation was carried out for 15minat 4C on a rotating wheel followed by two washing stepswith 15ml stringency RIPA buffer A [50mM TrisHCl(pH 75) 1M NaCl 1 NP40 1 sodium deoxycholate1mM EDTA 01 SDS 2M Urea] and two washingsteps with stringency RIPA buffer B [50mM TrisHCl(pH 75) 1M NaCl 1 NP40 1 sodium deoxycholate1mM EDTA 01 SDS 1M Urea]
For further analysis aliquots were taken from input(supernatant after preincubation with Sephadex G50)and the last wash Beads were equilibrated [50mM TrisHCl (pH 75) 300mM NaCl 05 NP40 05 sodiumdeoxycholate 1mM EDTA 01 SDS] and resuspendedin RIPA elution buffer [50mM TrisHCl (pH 75)300mM NaCl 05 NP40 05 sodium deoxycholate5mM EDTA 05 SDS] To each sample proteinase Kwas added at a final concentration of 1mgml andincubated for 1 h at 42C and 5 h at 56C Samples wereextracted with phenolchloroformisoamyl alcoholsolution (Roth) and RNA was precipitated with isopro-panol and 20 mg of glycogen
Deep sequencing of co-immunoprecipitated RNAs
cDNA synthesis and Illumnia sequencing was carried outaccording to previously published protocols (22)Specifically the two RNA samples from the CLIP experi-ment were poly(A)-tailed using poly(A) polymerase Afterthat the 50PPP was converted into 50P using tobacco acidpyrophosphatase Then an RNA adapter was ligated tothe 50-phosphate of the RNA First-strand cDNA synthe-sis was performed using an oligo(dT)-adapter primer andthe M-MLV reverse transcriptase The resulting cDNAswere PCR-amplified to 20ndash30 ngml using a high-fidelityDNA polymerase Adaptors that were used for ligationare listed in Supplementary Table S1
The cDNA libraries were sequenced using an IlluminaGAIIx machine with 100 cycles resulting in 5 674 433(DnmA-GFP pull down) and 5 540 257 (GFP control)reads After removal of the barcodes and adaptorsequences the reads were aligned to the genome ofD discoideum (sequence accession numbers NC_000895
NC_001889 NC_007087 NC_007088 NC_007089NC_007090 NC_007091 NC_007092) using the shortread mapper segemehl (23) For unmapped readspolyA-tails were computationally searched in the 30-endand clipped if found The resulting sequences weremapped again to the reference sequences In all1 249 579 reads of the DnmA-GFP pulldown and514 809 reads of the control sample could be successfullyaligned Enrichment of RNAs was calculated by compari-son of normalized read numbers from the DnmA-GFPpull down and the control In case of multiple genes re-sulting in the same transcript the reads are mapped to thefirst copy in the genome starting from chromosome 1
Cloning and in vitro transcription of RNAs
The genes for tRNAAsp(GUC-1) (DDB_G0294707)tRNAPhe(GAA-2) (DDB_G0294663) tRNALeu(UAG-1)
(DDB_0234827) tRNALeu(AAG-8) (DDB_0235066)tRNAVal(CAC-1) (DDB_0234789) and tRNAGly(GCC-1)
(DDB_0234894) were cloned into pJET1blunt vector(Fermentas) according to the suppliers suggestions Inthe case of multiple identical genes for one tRNA onlyone gene is listed here The annotation of tRNAs se-quences potential isoacceptor tRNAs and in silicofolding are shown in Supplementary Table S2 andSupplementary Figure S1 Genes were produced by recur-sive PCR using overlapping oligos (Supplementary Table
S1) For tRNAAsp(GUC) tRNALeu(UAG) tRNALeu(AAG)tRNAVal(CAC) and tRNAGly(GCC) the respective oligos 2and 3 served as template and oligo 1 and 4 as primers Arestriction site was added at the 30-end and a T7-promotorat the 50-end For tRNAPhe(GAA) oligo 1 2 and 3 are usedas template Amplification was done with oligo 4 and 5
The tRNAGlu(UUC) (DDB_G0294709) and suppressor
tRNAGlu(CUA) (kindly provided by T Winckler Jena)were recloned into pJET1blunt vector from pGEMTvector The mutated versions tRNAAsp(GUC)C38A andtRNAGlu(UUC)C38A were constructed by using the respect-ive oligonucleotides (Supplementary Table S1)For in vitro transcription 500 ng of linearized plasmid
was incubated at 37C for 2 h in 50 ml with 1 ml T7-RNApolymerase 1mM NTPs 1 transcription buffer(Fermentas) and RiboLockTM RNase Inhibitor(Fermentas) After transcription samples were treatedwith DNaseI for 15min extracted with an equal volumeof phenolchloroformisoamyl alcohol (Roth) and purifiedover a Sephadex G-50 spin columnFor radio-labelled in vitro transcription 2 mg of
linearized plasmid were incubated at 37C for 2 h in100 ml volume with 2 ml T7-RNA polymerase 2mMATP GTP and CTP 1mM UTP and 2 ml [a32P] UTP(110 TBqmmol Hartmann) 1 transcription bufferand RiboLockTM RNase Inhibitor (Fermentas) Aftertranscription samples were treated as aforementioned
Preparation of small RNA fraction
RNA isolation was done using the Trizol-method (24)For enrichment of the small RNA fraction 05M NaCl5 PEG 8000 was added to the RNA solution (in DEPCtreated H2O) and incubated at 20C for 30min After
Nucleic Acids Research 2013 3
at Universitaumlt W
uumlrzburg on August 5 2013
httpnaroxfordjournalsorgD
ownloaded from
centrifugation (10000 RCF 4C 30min) the supernatantwas collected in a fresh tube and precipitated with threevolumes of 100 EtOH
SDS-EMSA
Detection of denaturant-resistant DnmA-tRNA wascarried out by SDS-electrophoretic mobility shift assayas described elsewhere (25) with some modificationsBriefly DnmA (1ndash9 mg) was incubated in 20 ml of 10mMKPi (pH 70) 225 glycerol 25mM KCl 10mM DTT2mM MgCl2 and 100ndash500 mM S-adenosyl-methionine(SAM) with the respective 32P labelled tRNA (100ndash665 ng) at 22C Reactions were terminated at theindicated times by addition of 10 ml of SDS-stop mixture[150mM TrisndashHCl (pH 80) 6 SDS (wv) 30mMEDTA 0025 Bromophenol blue] and heated for10min at 65C Samples were cooled down to room tem-perature and subjected to electrophoresis on 6 or 12SDSndashpolyacrylamide gels sealed in Saran Wrap andexposed to an imager plate for 12ndash40 h at RT Plateswere read in a phosphoimager (Fujifilm FLA-7000) Insome cases samples were transferred from the gel to anitrocellulose membrane for immunodetection of DnmAusing an anti-His antibody and a secondary mouseantibody coupled to alkaline phosphatase
In vitro methylation assay
Methylation of RNA substrates was carried out asdescribed previously (13) 3 mM of recombinant hDnmt2and 3ndash12 mM of DnmA were incubated with 500 ng oftRNA-transcript (or 10 mg of small enriched RNA) 1methylation buffer [50mM TrisHCl (pH 70) 50mMNaCl 5mM MgCl2 1mM DTT] and 17mM [3H]-SAM(Hartmann) 1 uml RiboLockTM RNase Inhibitor(Fermentas) at 22C for 1min to 15 h as indicated inthe text The reaction was stopped by adding 1mM non-radioactive SAM and 2mgml proteinase K for 30minLoading buffer [formamide 5mM EDTA (pH 74)bromophenol blue] was added and samples were run ona 12 denaturing (7M Urea) polyacrylamide gel (1mm)for 2ndash5 h at 400V The gel was stained withethidiumbromide fixed (10 methanol 10 aceticacid) and immersed for 1 h in amplify solution(Amersham) dried and exposed to X-ray film (ContatypCX-BL+ Medical X-ray film) for 16 h up to 2 weeks at80C
In vitro blocked methylation assay
The in vitro blocked methylation assay is based on theassumption that RNA methylation is inhibited if thetarget site is covered by hybridization with a complemen-tary DNA oligo RNA (500 ng of transcript or 10 mg ofsmall enriched RNA) was mixed with 2 mM of the respect-ive oligonucleotide in 1 STE-buffer [10mM TrisHCl(pH 8) 50mM NaCl 1mM EDTA] heated up to 80Cin a thermocycler and then cooled down stepwise to roomtemperature The blocked RNA was subsequently used ina 3H-methylation assay as described earlier in the textBlocking oligos are shown in Supplementary Table S1(oligo 2 for tRNAAsp and tRNAGlu respectively)
dnmA expression analysis by real-time quantitative PCR
For analysing dnmA expression levels RNA was extractedby the Trizol-method (24) qPCR was performed in anEppendorf realplex light cycler with the SensiFAST SYBRNo-ROX One-Step Kit (Bioline) according to the sup-pliers instructions Relative quantification was done withthe Ct-method using gapdh (DDB_G0275153) as anormalization control (housekeeping gene)
Microscopy
For fluorescence analysis cells were fixed in 4 (wv)paraformaldehyde dissolved in 20mM phosphate buffer(pH 67) for 10min at 22C and stained with DAPI
Images were acquired on a Leica DMIRB invertedmicroscope equipped with a DC350 camera and IM50Acquisition software (Leica Microsystems WetzlarGermany)
For the movie DnmA-GFP cells (in DnmAKO back-ground) were analysed with a Zeiss Cell Observer SDwith Rapp UGA-40-2 L Scanner Images of living cellswere taken in 15 s intervals at 7 layers with a 08mMdistance between each layer during a period of 120minFurther image analysis was done with ImageJ 146 Themovie is displayed with eight frames per second
RNA bisulfite sequencing
RNA bisulfite sequencing was carried out as describedelsewhere (26) Primer sequences are listed in theSupplementary Material (Supplementary Table S1)
RESULTS
DnmA is an active tRNAAsp methyltransferase in vitroand in vivo
Previous experiments have shown that hDnmt2 canmethylate a D discoideum RNA in the size range oftRNAs in vitro (13) Here we investigated whether theD discoideum Dnmt2-homologue DnmA is also anactive tRNAAsp methyltransferase In vitro transcribedD discoideum tRNAAsp(GUC) was used as a substrate formethylation by DnmA and hDnmt2 in vitro The assaysshowed that the tRNA was efficiently methylated by bothenzymes (Figure 1A) The reaction was time dependentthe activity was sensitive to Mg2+ concentrations andalmost lost at 10mM Mg2+ In contrast to the humanenzyme DnmA was inactive at 37C (data not shown)Therefore all reactions were carried out at 22C wherehDnmt2 still showed good activity
To investigate methylation in vivo RNA bisulfiteanalysis of tRNAAsp(GUC) from D discoideum wasperformed Figure 1B shows that in wild-type cellstRNAAsp(GUC) was methylated at position C38 to30 whereas methylation was at background levels inDnmA knockout strains Ectopic overexpression ofhDnmt2 in wild-type cells increased tRNAAsp(GUC) C38methylation to 50 indicating cross-species activity ofDnmt2-homologues C49 was methylated at 90 inde-pendently of DnmA and served as an internal control forthe bisulfite reaction
4 Nucleic Acids Research 2013
at Universitaumlt W
uumlrzburg on August 5 2013
httpnaroxfordjournalsorgD
ownloaded from
As tRNAAsp appeared to be not the only target forDnmt2 in other organisms (eg D melanogaster) weexamined if more cellular-derived RNAs could bemethylated in vitro (ex vivo methylation experiments)Total RNA from dnmAKO AX2 cells was subjected toin vitro methylation by recombinant DnmA for varioustimes Methylated RNAs were separated by gelelectro-phoresis and visualized by autoradiography Figure 1Cshows that a substantial RNA fraction in the tRNA sizerange could be methylated At longer incubation times asecond minor methylated RNA species appeared
indicating that at least one further substrate could bemethylated in vitro
Identification of further substratesmdashin vitro binding andmethylation of tRNAGlu
The group of Helm (personal communication) hadproposed a sequence pattern (C32 C34 A37 C40) thatmay serve as a target recognition motif for C38 methyla-tion by hDnmt2 We therefore examined all D discoideumtRNAs in silico to identify candidates for the additionalmethylated band in the ex vivo experiment In addition to
Figure 1 In vitro methylation of tRNAAsp(GUC) (A) In vitro methylation of tRNAAsp(GUC) by DnmA at 2mM 5mM and 10mM MgCl2 and byhDnmt2 at 5mM Mg2+ The upper panel shows ethidium bromide staining of the in vitro transcripts separated in a denaturing polyacrylamide gelthe lower panel shows incorporated 3H-Me in the tRNAs Reactions were run for the times indicated (B) In vivo methylation of cytosines intRNAAsp(GUC) from different D discoideum strains Results of the RNA bisulfite sequencing (454 pyrosequencing) are given in percentage of readsNumbers of sequence reads are shown in brackets C49 is methylated by a different methyltransferase and thus serves as an internal standard All 22tRNAAsp(GUC) genes result in the same transcript and no isoacceptors are encoded in the D discoideum genome (C) In vitro methylation of smallenriched RNA of a dnmAKO strain (ex vivo methylation) The methylation reaction was done for the times indicated
Nucleic Acids Research 2013 5
at Universitaumlt W
uumlrzburg on August 5 2013
httpnaroxfordjournalsorgD
ownloaded from
tRNAAsp(GUC) we found tRNAGlu(UUC) andtRNAGlu(CUC) to match the pattern (Figure 2 andSupplementary Table S2)To test for tRNAGlu as a potential substrate we
modified the methylation assay by addition of a 50 ntDNA antisense oligonucleotide to tRNAAsp(GUC)
covering the methylation target site Figure 3A shows asa control that methylation of in vitro transcribedtRNAAsp(GUC) was completely blocked We then per-formed ex vivo methylation assays with enriched smallRNAs isolated from D discoideum after blockingtRNAAsp(GUC) with the antisense oligo Figure 3B showsthat the majority of 3H-incorporation disappearedHowever with both DnmA and hDnmt2 a significantamount of methylation was still detectable in the sizerange of tRNAs Assuming that this signal containedtRNAGlu we used a second antisense oligo directedagainst tRNAGlu(UUC) for blocking methylation Theoligo also covered the isoacceptor tRNAGlu(CUC) with
minor mismatches (Supplementary Figure S2) A furtherreduction of ex vivo methylation was observed Notablythe lower band in the tRNA size range disappeared (arrowin Figure 3B) suggesting that this represented a tRNAGlu
population that could be methylated at least in vitro Theresidual methylation after blocking with both oligos mayrepresent additional tRNAs or other unknown targets Aswe only concentrated on small RNAs larger methylatedmolecules and less abundant targets may have escapeddetection
To further analyse methylation on tRNAAsp andtRNAGlu (exemplified by tRNAGlu(UUC)) we carried outassays with wild-type sequences and C38A mutatedin vitro transcripts (Figure 4) We also included a suppres-sor tRNAGlu(CUA) that differed in only 2 nt fromtRNAGlu(UUC) (Figure 2) Assays were done in parallelwith the recombinant hDnmt2 and D discoideumDnmA The experiment showed that tRNAAsp(GUC) wasthe best substrate for both DnmA and hDnmt2 (Figure 4)Substantial methylation was also observed fortRNAGlu(UUC) and suppressor tRNAGlu(CUA) Moreoverthe complete loss of methylation in the C38A mutations oftRNAAsp(GUC) and tRNAGlu(UUC) confirmed that methy-lation occurred at position C38 and largely ruled out thatmethyl group incorporation was due to another
Figure 3 Ex vivo methylation and blocking assay (A) Methylation of in vitro transcribed tRNAAsp(GUC) was completely blocked when a comple-mentary antisense oligo was hybridized (B) Using enriched small RNA from dnmAKO cells ex vivo methylation in the tRNA size class wasdifferentially lost when antisense oligos to tRNAAsp(GUC) and tRNAGlu(UUC) were hybridized before the methylation reaction The oligo againsttRNAGlu(UUC) also covers tRNAGlu(CUC) with minor mismatches (see Supplementary Figure S2) Even with both oligos a significant amount of 3Hincorporation still remained The upper panel shows the ethidiumbromide stained gel to demonstrate equal loading the lower panel shows thefluorogram The arrow marks a band that was specifically lost when the anti tRNAGlu(UUC) oligo was used
Figure 2 Target site sequences of tRNAs used for in vitro methylationThe position of the putative signature nucleotides are indicated andprinted in bold if present The anticodon is underlined tRNAGlu(CUC)
(not shown) also harbours the nucleotide pattern Except for the anti-codon both tRNAGlu are identical in the anticodon stem-loop region(Supplementary Figure S2) Only a few differences appear in theacceptor stem
Figure 4 C38 is the target for in vitro methylation of tRNAAsp(GUC)
and tRNAGlu(UUC) In vitro methylation of tRNAAsp(GUC)tRNAGlu(UUC) and a suppressor tRNAGlu(CUA) C38A mutations con-firmed C38 as the target nucleotide tRNAAsp(GUC) methylation wasmost efficient
6 Nucleic Acids Research 2013
at Universitaumlt W
uumlrzburg on August 5 2013
httpnaroxfordjournalsorgD
ownloaded from
contaminating methyltransferase in the recombinantprotein preparation (17)
The methylation experiments using in vitro transcribedtRNAs showed that no other modifications were necessaryfor C38 methylation by DnmA and hDnmt2 (13) and thatthe processed ends of a mature tRNA were not requiredfor target recognition as the tRNA transcripts contain no30-CCA Even tRNAGlu embedded in a long dsRNAmolecule (1000 nt) with small stem-loop structures atthe ends (27) was a specific substrate of DnmA andhDnmt2 (Supplementary Figure S3)
By EMSA no difference in in vitro binding could beseen for the specific target tRNAGlu(UUC) and non-targetRNAs indicating that DnmA associated non-specificallywith any nucleic acid (data not shown) However specificcovalent enzyme-substrate intermediates can be detectedby denaturing SDSndashPAGE (25) Figure 5A andSupplementary Figure S4 show that radioactivelylabelled tRNA transcripts that can be methylatedin vitro by DnmA formed denaturation resistantcomplexes while non-target RNAs did not To confirmthat the denaturation resistant complexes were really dueto binding of the enzyme to the substrate SDS gels wereblotted and His-tagged DnmA was detected by a specificanti-His antibody A small amount of the tagged proteinwas shifted to the same position as the labelled tRNA(Supplementary Figure S5)
We then performed time courses for the reaction andfound that covalent complex formation was only transientunder the conditions used For tRNAAsp(GUC) adductsreached a maximum after 1min and were undetectableafter 15min whereas the tRNAGlu(UUC) complex onlybegan to accumulate after 1min but persisted for almost60min (Figure 5A and B) This coincided with the highmethylation activity on tRNAAsp(GUC-1) and a lower oneon tRNAGlu(UUC-2) The human enzyme appeared to beslightly faster in the reaction but otherwise showed asimilar behaviour compared with D discoideum DnmA
tRNAAsp(GUC) tRNAGlu(UUCCUC) and tRNAGly(GCC)
bind specifically to DnmA in vivo
The blocking assay (Figure 3) suggested that further sub-strates could be targeted by DnmA To identify suchtargets and to further investigate the interaction betweenDnmA and RNAs in vivo we established a UV-CLIP pro-cedure for D discoideumDnmA-GFP expressing cells andcontrol cells expressing GFP only were UV cross-linkedand complexes were precipitated under denaturing condi-tions by a GFP nanobody (21) Then co-precipitatedRNAs were isolated converted into strand-specificcDNA libraries and analyzed by Illumina sequencing(Figure 6 and Supplmentary Figure S6)
Various tRNA fragments were enriched in the DnmACLIP sample compared with the control By their lengthand position fragments appeared not to be random butspecific We found mid-fragments of 20ndash24 nt (position32ndash36 to 53ndash56) that covered the C38 methylation site inthe respective tRNAs 50-fragments of 17 nt (nucleotideposition 1ndash17) 30-fragments of 2021 nt (nucleotideposition 54 to the end) and 30-half tRNAs of 41ndash43 nt
(position 30ndash32 to the end) Mid-fragments oftRNAAsp(GUC) tRNAGlu(UUCCUC) and tRNAGly(GCC)
were significantly enriched in the DnmA-GFP fraction incomparison with the control CLIP assay tRNAAsp(GUC)tRNAGlu(UUC) and tRNAGlu(CUC) harbour the sequencepattern (C32 C34 A37 C40) which has been suggestedas a signature for C38 methylation (Figure 2 Helmpersonal communication) tRNAGly(GCC) contains thepattern except for C40 For tRNAAla(AGC) which doesnot contain C38 and the signature pattern a differentfragment from the 30-end was enriched in the DnmA-CLIP sample For other tRNAs no significant accumula-tion was observed in the experiment (Figure 6 andSupplementary Figure S6) For several tRNAs varyingamounts of full-length molecules were found but thesewere never enriched in the DnmA sample in comparisonwith the control
tRNAGly(GCC) is an additional methylation substratein vitro
The CLIP data suggested that tRNAGly(GCC) was anothertarget for DnmA We therefore cloned this and several othertRNA genes that contained the pattern only partially(Figure 2) We generated in vitro transcripts and examinedmethylation by DnmA and hDnmt2 in vitro IndeedtRNAGly(GCC) was methylated by both enzymes (Figure 7)while none of the other tRNAs (tRNALeu(UAG-1)tRNALeu(AAG-8) tRNAVal(CAC-1) and tRNAPhe(GAA-2))resulted in detectable methylation signals in the assay(Supplementary Figure S7) Interestingly the in vitro methy-lation efficiency of the three tRNA substrates
Figure 5 Formation and turnover of tRNA-methyltransferasecomplexes Turnover of covalent complexes of tRNA substrates withDnmA and hDnmt2 (A) Examples of time courses on covalentcomplex formation with tRNAAsp(GUC) and tRNAGlu(UUC) (B)Complex bands in A were quantified and presented on a time scale
Nucleic Acids Research 2013 7
at Universitaumlt W
uumlrzburg on August 5 2013
httpnaroxfordjournalsorgD
ownloaded from
(tRNAAsp(GUC) tRNAGlu(UUC) tRNAGly(GCC)) correlatedroughly with the number of tRNA fragments found in theRNA-CLIP experimentThe data supported the conclusion from the ex vivo
blocking assay (Figure 3B) that in addition toD discoideum tRNAAsp and tRNAGlu other RNAs weresubstrates for DnmA and hDnmt2 in vitro
Expression of dnmA is regulated during developmentcell cycle and on temperature stress
DnmA expression in growing vegetative D discoideumcells is low To examine tRNA methylation in vivo it
was therefore advantageous to determine conditionsunder which dnmA was most highly expressed Promptedby previous studies (141517) we assumed that the biolo-gical function of DnmA was connected to developmentalandor environmental conditions We therefore examinedexpression levels of dnmA under different regimes dnmAmRNA was quantified by qPCR over the 24 h develop-mental cycle of D discoideum (data not shown) and con-firmed the data deposited in Dictyexpress (httpdictyexpressbiolabsi)
Interestingly an 5-fold increase in expression wasobserved at 16 h in development This time coincides
Figure 6 tRNA and tRNA fragments associated with DnmA-GFP After UV crosslinking RNA bound to DnmA was co-immunoprecipitatedunder denaturing conditions (CLIP) The number of normalised reads for tRNAs detected by Illumina sequencing is shown for DnmA CLIP and forthe control CLIP with GFP In addition to full-length tRNAs four classes of tRNA fragments were found and these are indicated by differentshading of the bars Size and localization of fragments is shown schematically on the simplified clover leaf structure tRNAs that are not significantlyenriched with DnmA are shown in Supplementary Figure S6 In case of multiple gene copys resulting in the same RNA transcript the sequencingreads were mapped to the first copy in the genome starting from chromsome 1 Only this copy is listed Sequences and detailed information on genecopies and potential isoacceptors are listed in Supplementary Table S2
8 Nucleic Acids Research 2013
at Universitaumlt W
uumlrzburg on August 5 2013
httpnaroxfordjournalsorgD
ownloaded from
with a presumed synchronous mitotic division (28) wherea multitude of cell-cycle regulated genes are expressed[(29) and McWilliams personal Communication]
Therefore we synchronized vegetative D discoideumcells to examine dnmA expression during the cell cycleFor these experiments the D discoideum wild-typestrain NC4 the parent of the axenic laboratory strainAX2 was used (30) This was prompted by the observationthat D melanogaster and E histolytica laboratory strainsmay lose dnmt2 expression (Reuter Ankri personalcommunication)
The data confirmed our assumption of a cell-cycle-de-pendent transcriptional regulation of dnmA (Figure 8A)The peak of expression coincided with late mitosis or earlyS-phase as D discoideum does not display a G1 phaseUsing AX2 cells cotransformed with PCNA-RFP andDnmA-GFP we observed GFP accumulation in thenucleus during S phase (indicated by PCNA expression)and a loss of DnmA from the nucleus during mitosis(Figure 8B) This is further exemplified by the movie inthe Supplementary Material (Supplementary Movie S1)
In other organisms dnmt2 was observed to beupregulated during stress response (315) We thereforeexamined expression after anoxia heat shock and coldshock but did not find any significant changes (data notshown) Only during recovery from cold shock wedetected a strong transient increase in dnmA mRNA by47-fold (Figure 8C)
Thus dnmA expression was differentially regulatedduring the cell cycle development and on temperaturestress Furthermore DnmA localization changed specific-ally during the cell cycle
Though the basic expression in vegetative cells was in-distinguishable dnmA expression in NC4 was 30-foldhigher at 16 h of development whereas developmentalupregulation was only 5-fold in AX2 The data aresummarized in Figure 8D
NC4 the parent strain of AX2 displays increasedtRNA
Asp methylation during development
Based on these observations we asked the questionwhether expression levels correlated with tRNA
methylation activity in vivo Bisulfite sequencing wasdone with primers for tRNAAsp tRNAGlu and tRNAGly
on RNA isolated from vegetative AX2 cells AX2 cellsdeveloped for 16 h AX2 cells subjected to cold shockAX2 recovered for 25 h after cold shock vegetativeNC4 cells and NC4 cells developed for 16 h The resultson C38 methylation of tRNAAsp(GUC) are summarized inFigure 8E The methylation status of all C residues fromall tested tRNAs in this study is shown in SupplementaryFigure S8 Indeed tRNAAsp(GUC) methylation at C38increased to 76 in development in NC4 cells Noincrease was observed in AX2 cells during developmentpossibly due to insufficient sensitivity of the assayInterestingly no increase in tRNAAsp(GUC) methylationwas observed at the highest dnmA expression levelsduring stress recovery In Figure 8E the 30ndash35 methy-lation level in all tRNAAsp(GUC) samples represents signifi-cant methylation as compared with other C residues (seeSupplementary Figure S8)We did however not detect any significant methylation
of tRNAGlu(UUCCUC) and tRNAGly(GCC) under any con-ditions (Supplementary Figure S8) In contrast C49C50methylation which does not depend on Dnmt2 enzymeswas found in all samples and thus served as an internalcontrol In all 1175 bisulfite-sequence reads wereexamined for tRNAGlu(UUCCUC) and 4040 fortRNAGly(GCC) No methylation above background wasfound Though it cannot be excluded that the failure todetect C38 methylation of tRNAGlu and tRNAGly was dueto inefficient reverse transcription or low levels of methy-lation (see lsquoDiscussionrsquo section) we conclude thattRNAAsp is the major or possibly the only in vivo targetfor methylation by DnmA in D discoideum
DISCUSSION
The biological function of Dnmt2 methyltransferases islargely unknown and its role in DNA methylation is con-troversially discussed (910) In contrast tRNA methyla-tion by Dnmt2 is robust in vitro and in vivo andevolutionary conserved In addition to the originallydescribed tRNAAsp(GUC) (1) two further tRNAs targets(tRNAVal(CAC) and tRNAGly(GCC)) have been defined inD melanogaster (15) and tRNAGlu(UUC) in S pombe (17)To get insight into potential functions of the
D discoideum Dnmt2 homologue DnmA we examinedexpression levels in development in the cell cycle andunder different environmental conditions We confirmedby qPCR an expression peak at 16 h of developmentWeijer and co-workers (28) had previously proposed asynchronous cell division at this developmental stageWe therefore investigated dnmA expression in the cellcycle and found a strong transient increase of mRNA inS-phase or shortly thereafter The expression peak wasaccompanied by reaccumulation of DnmA in thenucleus We assume that DnmA is degraded duringmitosis and re-synthesised after cell divisionIn S pombe as well as in D melanogaster tRNA methy-
lation changes in response to environmental challengeslike temperature stress anoxia or nutrients We tested
Figure 7 In vitro methylation of tRNAGly(GCC) by recombinant DnmAand hDnmt2 In vitro transcribed tRNAs were methylated in vitro asdescribed before The top panel shows the ethidium-bromide stainedgel the bottom panel the fluorogram of 3H-labelled methylated tRNAsAs a control tRNAAsp(GUC) methylation is shown
Nucleic Acids Research 2013 9
at Universitaumlt W
uumlrzburg on August 5 2013
httpnaroxfordjournalsorgD
ownloaded from
Figure 8 Expression localization and tRNAAsp(GUC) methylation activity of DnmA under various conditions (A) dnmA expression is regulatedduring the cell cycle Cells were arrested in the cell cycle by cold treatment and then released by transfer to 22C Cells were counted every 30min(grey dots and grey line) and samples were taken for qPCR (black bars) After cell division dnmA expression increased 5-fold and then rapidlydeclined to basal levels Normalization was done on vegetative growing cells dnmA expression is only shown from 3 to 45 h during recovery(B) DnmA is lost from the nucleus during mitosis (arrow) The three cells on the right are in S-phase as indicated by the RFP-PCNA marker andaccumulate DnmA in the nucleus (for further details see movie in Supplementary Material) (C) Relative quantification of dnmA expression levels inAX2 cells after 25 h recovery from cold shock at 4C Expression of dnmA increased gt40-fold and returned within 30min to basal levels (n=3)(D) At 16 h of development dnmA expression increased 46-fold in the NC4 strain In AX2 cells expression increased only 5-fold (E) At 16 h ofdevelopment in vivo methylation of C38 in tRNAAsp(GUC) increased up to 75 in the D discoideum strain NC4 whereas no significant increase wasobserved in AX2 cells
10 Nucleic Acids Research 2013
at Universitaumlt W
uumlrzburg on August 5 2013
httpnaroxfordjournalsorgD
ownloaded from
heat shock cold shock anoxia and various nutrient mediabut did not observe altered levels Only during therecovery phase after cold treatment a transient almost50-fold increase in mRNA abundance was seen
Reuter and Ankri (personal communication) hadobserved a loss of Dnmt2 expression in laboratorystrains of D melanogaster and E histolytica We thereforeexamined D discoideum NC4 the parent of the axeniclaboratory strain AX2 The developmental regulation inAX2 and NC4 was qualitatively similar but expressionlevels were much higher in NC4 The observationsupports the assumption that Dnmt2 expression wasreduced under optimal growth conditions in the labora-tory strain and may actually be selected against
We show that the D discoideum Dnmt2 homologueDnmA is an active tRNAAsp(GUC) methyltransferasein vitro and in vivo To identify further potential targetsfor DnmA we established a UV-CLIP method to analyseDnmAndashRNA interaction in vivo In addition to the knowntarget tRNAAsp(GUC) fragments of tRNAGlu(UUC) andtRNAGly(GCC) were most abundant in comparison withthe control Unexpectedly complete tRNAs were notenriched in the DnmA CLIP fraction but rather specificlsquomid-fragmentsrsquo that covered the putative methylation siteand the recognition pattern proposed by Helm (personal
communication) A minor exception was tRNAGly(GCC)which lacks C40 We do not know whether these frag-ments were directly or indirectly generated by DnmAin vivo or whether they were protected by DnmA fromdegradation during the experimental procedure As noco-crystal of Dnmt2 and a tRNA substrate is availablethe exact interface for the interaction is unknownJurkowski et al (31) mapped the binding site on Dnmt2by mutational analysis A manual placement of a tRNAinto the crystal structure suggests that Dnmt2 mainlycontacts the anticodon stem loop Assuming that mid-frag-ments are footprints of DnmA our data support this notionbut suggest that the interaction surface extends into theD-loop which is partially covered by the mid-fragments
We examined the putative DnmA targets in vitro (bymethylation of in vitro transcribed tRNAs) ex vivo(by methylation of RNA isolated from D discoideum)and in vivo (by bisulfite sequencing) Furthermore weinvestigated by SDSndashPAGE the kinetics of covalentcomplex formation between tRNAAsp and tRNAGlu withDnmA and hDnmt2
All three tRNAs (tRNAAsp tRNAGlu tRNAGly) werereadily methylated by recombinant DnmA and hDnmt2in vitro although the activity was much higher (estimatedat least 10-fold) on tRNAAsp compared with tRNAGlu andtRNAGly For tRNAAsp and tRNAGlu this correlated wellwith the formation and decay of covalent RNAmdashproteincomplexes where kinetics for tRNAAsp were much fasterthan for tRNAGlu Instead of C methylation the covalentintermediate may also proceed to a hydrogen exchangereaction or to a deamination of the cytosine In bothcases an intermediate will be detected but it will notresult in a methylated product (32) It may be thatslower off rates represent these alternative pathways thatwould result in unmethylated tRNAs
Using antisense oligos we showed that tRNAAsptRNAGlu and at least one additional RNA could bemethylated ex vivo A quantification of the results wasnot possible the apparently low methylation oftRNAGlu ex vivo could be due to the low activity of theenzyme on this substrate (as suggested by the in vitroexperiments see Figure 5B)The data for the examination of in vivo methylation by
bisulfite sequencing were rather unexpected AlthoughtRNAAsp(GUC) was consistently found to be methylatedat C38 we never detected any methylation at C38 oftRNAGlu(UUCCUC) or tRNAGly(GCC) There could betwo reasons to explain the failure to detect C38 methyla-tion for these in vitro substrates (i) due to the low activityof DnmA on tRNAGlu(UUCCUC) and tRNAGly(GCC) asshown in vitro there was low or no significant methylationin vivo and (ii) methylation escaped detection becauseother unknown modifications blocked reverse transcrip-tion of the methylated molecules However if this wasthe case one has to assume that C49C50 methylationoccurs independently of these unknown modificationswhereas C38 methylation strictly depends on them AsC38 methylation does not require any other modificationsin vitro we believe that this scenario is less likely If in vitromethylation activity was comparable with in vivo methy-lation 10 methylation of tRNAGlu(UUCCUC) andtRNAGly(GCC) could be expected relative totRNAAsp(GUC) As tRNAAsp was methylated to 30 a3 methylation of tRNAGlu and tRNAGly could be belowthe detection level of the bisulfite assayWe find that high expression of dnmA did not necessar-
ily correlate with increased methylation of tRNAs Duringrecovery from cold shock D discoideum cells displayedthe highest level of dnmA expression but this had noeffect on tRNAAsp methylation Only in developmentand with the high expressing NC4 strain a significantincrease of tRNAAsp methylation to 70 was observedIt has to be considered however that bisulfite sequencingdoes not necessarily allow for quantitative evaluation ofdata detected C-methylation depends on reverse tran-scription efficiency which may be hampered by othermodifications that could be different under differentconditions Furthermore the completeness of bisulfite-mediated conversion could also depend on RNA struc-ture which may be altered by other modificationsHowever incomplete conversion will lead to an overesti-mation of C-methylation as maybe the case for forexample C30 C31 C35 in tRNAGly in some samplesTaken together the data from CLIP complex forma-
tion and methylation show that all three substrates can bebound and methylated in vitro The CLIP data show thatthere is a specific interaction of DnmA with tRNA mid-fragments of tRNAAsp(GUC) tRNAGlu(UUCCUC) andtRNAGly(GCC) but not with other tRNAs In contrastbisulfite sequencing could only detect methylation ontRNAAsp(GUC) even at the highest expression levels ofdnmA Even though low amounts of methylation mayhave escaped detection in bisulfite sequencing there isstill a discrepancy between dnmA expression levels (egduring cold shock recovery) DnmA binding to substrates(as determined by CLIP) and methylation in vivo
Nucleic Acids Research 2013 11
at Universitaumlt W
uumlrzburg on August 5 2013
httpnaroxfordjournalsorgD
ownloaded from
We therefore suggest that DnmA binding to specifictargets could elicit functions in addition to RNA methy-lation eg by promoting processing or degradation andor recruitment of other proteins At least for some rRNAmethyltransferases such as Bud23 or Nep1 fromSaccharomyces cerevisiae it is known that they executefunctions independent from the methylation activity(33ndash35) As shown by Schaefer and co-workers (15)methylation protects tRNAs from degradation or process-ing after stress By a different pathway binding of DnmAmay initiate processing that leads to specific tRNA frag-ments that could serve for other biological functions (36)Interestingly tRNAGlu(UUC) is also a substrate for theDnmt2 homologue Pmt1 in S pombe (17) HowevertRNAGlu(UUC) is found to be methylated to a low butsignificant extend in vivo in fission yeast when Pmt1 isoverexpressedThe detection of tRNAAla(AGC) fragments in the CLIP
assay brings up a new question tRNAAla(AGC) has a Tinstead of a C in position 38 and does not contain thesignature nucleotides in the anticodon stem-loopInstead of a mid-fragment a distinct 30 fragment wasassociated with DnmA suggesting that a recognitionmotif was located in this part of the moleculeAs methylation also occurred in a large artificial
molecule with embedded tRNAGlu (SupplementaryFigure S2) it could be that other longer RNAs containingan appropriate recognition motif may also be bound bythe enzyme We thus propose that the biological functionsof Dnmt2 proteins could extend far beyond our presentknowledge Investigation of further targets and of the sig-nificance of RNA fragments associated with Dnmt2should shed light on the importance of this enzyme
SUPPLEMENTARY DATA
Supplementary Data are available at NAR Online
ACKNOWLEDGEMENTS
The authors thank Thomas Winckler (Friedrich-Schiller-University Jena) for providing the tRNAGlu(UUC) and sup-pressor tRNAGlu(CUA) plasmids Susanne Junk for cloningtRNAAspC38A Matthias Schafer (DKFZ) and RobinLorenz (University of Kassel) are acknowledged forRNA bisulfite sequencing and Richard Reinhardt (MaxPlanck Genome Centre Cologne) for deep sequencingMark Helm (Johannes-Gutenberg-University Mainz) gen-erously provided data before publication CarstenSeehafer (University of Kassel) helped with the analysisof deep-sequencing data
FUNDING
Deutsche Forschungsgemeinschaft [FOR 1082 to WNand AJ] and the Studienstiftung des Deutschen Volkes(to SM) Funding for open access charge FOR1082
Conflict of interest statement None declared
REFERENCES
1 GollMG KirpekarF MaggertKA YoderJA HsiehCLZhangX GolicKG JacobsenSE and BestorTH (2006)Methylation of tRNAAsp by the DNA methyltransferasehomolog Dnmt2 Science 311 395ndash398
2 PhalkeS NickelO WalluscheckD HortigF OnoratiMCand ReuterG (2009) Retrotransposon silencing and telomereintegrity in somatic cells of Drosophila depends on the cytosine-5methyltransferase DNMT2 Nat Genet 41 696ndash702
3 WilkinsonCR BartlettR NurseP and BirdAP (1995) Thefission yeast gene pmt1+ encodes a DNA methyltransferasehomologue Nucleic Acids Res 23 203ndash210
4 KatohM CurkT XuQ ZupanB KuspaA and ShaulskyG(2006) Developmentally regulated DNA methylation inDictyostelium discoideum Eukaryot Cell 5 18ndash25
5 KuhlmannM BorisovaBE KallerM LarssonP StachDNaJ EichingerL LykoF AmbrosV SoderbomF et al(2005) Silencing of retrotransposons in Dictyostelium by DNAmethylation and RNAi Nucleic Acids Res 33 6405ndash6417
6 RaiK ChidesterS ZavalaCV ManosEJ JamesSRKarpfAR JonesDA and CairnsBR (2007) Dnmt2 functionsin the cytoplasm to promote liver brain and retina developmentin zebrafish Genes Dev 21 261ndash266
7 FisherO Siman-TovR and AnkriS (2006) Pleiotropicphenotype in Entamoeba histolytica overexpressing DNAmethyltransferase (Ehmeth) Mol Biochem Parasitol 147 48ndash54
8 DurdevicZ HannaK GoldB PollexT CherryS LykoFand SchaeferM (2013) Efficient RNA virus control in Drosophilarequires the RNA methyltransferase Dnmt2 EMBO Rep 14269ndash275
9 SchaeferM and LykoF (2010) Lack of evidence for DNAmethylation of Invader4 retroelements in Drosophila andimplications for Dnmt2-mediated epigenetic regulation NatGenet 42 920ndash921 author reply 921
10 KraussV and ReuterG (2011) DNA methylation in Drosophila -a critical evaluation Prog Mol Biol Transl Sci 101 177ndash191
11 RaddatzG GuzzardoPM OlovaN FantappieMRRamppM SchaeferM ReikW HannonGJ and LykoF(2013) Dnmt2-dependent methylomes lack defined DNAmethylation patterns Proc Natl Acad Sci USA 110 8627ndash8631
12 JeltschA NellenW and LykoF (2006) Two substrates arebetter than one dual specificities for Dnmt2 methyltransferasesTrends Biochem Sci 31 306ndash308
13 JurkowskiTP MeusburgerM PhalkeS HelmM NellenWReuterG and JeltschA (2008) Human DNMT2 methylatestRNA(Asp) molecules using a DNA methyltransferase-likecatalytic mechanism RNA 14 1663ndash1670
14 TovyA Siman TovR GaentzschR HelmM and AnkriS(2010) A new nuclear function of the Entamoeba histolyticaglycolytic enzyme enolase the metabolic regulation of cytosine-5methyltransferase 2 (Dnmt2) activity PLoS Pathog 6 e1000775
15 SchaeferM PollexT HannaK TuortoF MeusburgerMHelmM and LykoF (2010) RNA methylation by Dnmt2protects transfer RNAs against stress-induced cleavage GenesDev 24 1590ndash1595
16 TuortoF LiebersR MuschT SchaeferM HofmannSKellnerS FryeM HelmM StoecklinG and LykoF (2012)RNA cytosine methylation by Dnmt2 and NSun2 promotestRNA stability and protein synthesis Nat Struct Mol Biol 19900ndash905
17 BeckerM MullerS NellenW JurkowskiTP JeltschA andEhrenhofer-MurrayAE (2012) Pmt1 a Dnmt2 homolog inSchizosaccharomyces pombe mediates tRNA methylation inresponse to nutrient signaling Nucleic Acids Res 4011648ndash11658
18 TovyA HertzR Siman-TovR SyanS FaustD GuillenNand AnkriS (2011) Glucose starvation boosts Entamoebahistolytica virulence PLoS Negl Trop Dis 5 e1247
19 MotorinY and HelmM (2011) RNA nucleotide methylationWiley Interdiscip Rev RNA 2 611ndash631
20 NiranjanakumariS LasdaE BrazasR and Garcia-BlancoMA(2002) Reversible cross-linking combined with
12 Nucleic Acids Research 2013
at Universitaumlt W
uumlrzburg on August 5 2013
httpnaroxfordjournalsorgD
ownloaded from
immunoprecipitation to study RNA-protein interactions in vivoMethods 26 182ndash190
21 RothbauerU ZolghadrK MuyldermansS SchepersACardosoMC and LeonhardtH (2008) A versatile nanotrapfor biochemical and functional studies with fluorescent fusionproteins Mol Cell Proteomics 7 282ndash289
22 SittkaA LucchiniS PapenfortK SharmaCM RolleKBinnewiesTT HintonJC and VogelJ (2008) Deep sequencinganalysis of small noncoding RNA and mRNA targets of theglobal post-transcriptional regulator Hfq PLoS Genet 4e1000163
23 HoffmannS OttoC KurtzS SharmaCM KhaitovichPVogelJ StadlerPF and HackermullerJ (2009) Fast mapping ofshort sequences with mismatches insertions and deletions usingindex structures PLoS Comput Biol 5 e1000502
24 ConnollyMA ClausenPA and LazarJG (2006) Purificationof RNA from animal cells using trizol Cold Spring HarborProtoc 2006 pii pdbprot4104
25 DongA YoderJA ZhangX ZhouL BestorTH andChengX (2001) Structure of human DNMT2 an enigmaticDNA methyltransferase homolog that displays denaturant-resistant binding to DNA Nucleic Acids Res 29 439ndash448
26 SchaeferM PollexT HannaK and LykoF (2009) RNAcytosine methylation analysis by bisulfite sequencing NucleicAcids Res 37 e12
27 BoninM OberstrassJ VogtU WasseneggerM and NellenW(2001) Binding of IRE-BP to its cognate RNA sequence SFMstudies on a universal RNA backbone for the analysis of RNA-protein interaction Biol Chem 382 1157ndash1162
28 ZimmermanW and WeijerCJ (1993) Analysis of cell cycleprogression during the development of Dictyostelium and itsrelationship to differentiation Dev Biol 160 178ndash185
29 StrasserK BloomfieldG MacWilliamsA CeccarelliAMacWilliamsH and TsangA (2012) A retinoblastomaorthologue is a major regulator of S-phase mitotic anddevelopmental gene expression in Dictyostelium PLoS One 7e39914
30 WattsDJ and AshworthJM (1970) Growth of myxameobae ofthe cellular slime mould Dictyostelium discoideum in axenicculture Biochem J 119 171ndash174
31 JurkowskiTP ShanmugamR HelmM and JeltschA (2012)Mapping the tRNA binding site on the surface of humanDNMT2 methyltransferase Biochemistry 51 4438ndash4444
32 SvedruzicZM and ReichNO (2005) DNA cytosine C5methyltransferase Dnmt1 catalysis-dependent release of allostericinhibition Biochemistry 44 9472ndash9485
33 ThomasSR KellerCA SzykA CannonJR and Laronde-LeblancNA (2011) Structural insight into the functionalmechanism of Nep1Emg1 N1-specific pseudouridinemethyltransferase in ribosome biogenesis Nucleic Acids Res 392445ndash2457
34 LeulliotN BohnsackMT GrailleM TollerveyD andVan TilbeurghH (2008) The yeast ribosome synthesisfactor Emg1 is a novel member of the superfamily of alphabeta knot fold methyltransferases Nucleic Acids Res 36629ndash639
35 WhiteJ LiZ SardanaR BujnickiJM MarcotteEM andJohnsonAW (2008) Bud23 methylates G1575 of 18S rRNA andis required for efficient nuclear export of pre-40S subunits MolCell Biol 28 3151ndash3161
36 SobalaA and HutvagnerG (2013) Small RNAs derived fromthe 50-end of tRNA can inhibit protein translation in humancells RNA Biol 10 525ndash535
Nucleic Acids Research 2013 13
at Universitaumlt W
uumlrzburg on August 5 2013
httpnaroxfordjournalsorgD
ownloaded from

especially on retroelements were detected and attributedto Dnmt2 although these data are controversially dis-cussed (9ndash11)In 2006 Bestorrsquos group reported that Dnmt2 rather
methylates transfer RNA (tRNA) than DNA inM musculus D melanogaster and Arabidopsis thalianaSpecifically they found C38 in tRNAAsp as the major oronly substrate for the enzyme (1) A dual role of Dnmt2had been discussed (12) and Jurkowski et al (13) couldshow that tRNAAsp was methylated by an enzymaticmechanism characteristic for DNA methyltransferasesrather than by the reaction pathways of enzymes thatmethylate RNAstRNAAsp methylation activity was also reported for
the Dnmt2-homologues from E histolytica (14) andD melanogaster (15) Schaefer et al (15) showed thatD melanogaster tRNAVal(CAC) and tRNAGly(GCC) werealso methylated at position C38 in vivo and by humanDnmt2 (hDnmt2) in vitro They further found thatdnmt2 knockout flies were more sensitive to oxidativestress A more detailed analysis of tRNAAsp(GUC) andtRNAGly(GCC) suggested that methylation protectedtRNAs from stress-induced cleavage (15) Double-knockout mutant mice of Dnmt2 and Nsun2 the secondknown m5C-tRNA-methyltransferase in higher eukary-otes showed a phenotype with impaired cellular differen-tiation an overall reduction in protein synthesis and earlylethality (16) Recently we identified tRNAGlu(UUC) as anadditional novel substrate of Pmt1 the Dnmt2-homo-logue in S pombe (17) Pmt1-dependent tRNA methyla-tion seemed to be regulated by nutrient conditionsNutritional control was also reported for Ehmeth theDnmt2 homologue from E histolytica that is inhibitedby the glycolytic enzyme enolase (1418)Here we demonstrate that recombinant D discoideum
DnmA and hDnmt2 can methylate tRNAAsp(GUC)tRNAGlu(UUC) and tRNAGly(GCC) from D discoideumin vitro with different efficiencies Both enzymes formedcovalent complexes [22] with specific tRNAs with similarkinetics but they were significantly slower for the minorsubstrate tRNAGlu than for the major substrate tRNAAspUltraviolet (UV)-crosslinking and immunoprecipitation(CLIP) experiments showed that specific fragments ofthe three target tRNAs associated with DnmA in vivoHowever in vivo only methylation of tRNAAsp(GUC) byDnmA was confirmed whereas methylation of the targetnucleotide C38 in other substrates was not detectable bybisulfite sequencingAs revealed by quantitative reverse transcription (RT)-
PCR dnmA is differentially expressed in development cellcycle and in the recovery phase after temperature stressThe increase in dnmA expression levels correlated withelevated tRNAAsp methylation in development but notafter temperature stress The other targets identified byin vitro methylation and by CLIP were apparently alsonot methylated in vivo in developmentOur data document that additional RNA molecules can
serve as substrates for Dnmt2 binding and that the fullrange of targets is probably not yet recognized The resultsalso suggest that binding of the methyltransferase to an
RNA molecule not necessarily results in methylation butmay have different biological functions
MATERIALS AND METHODS
Dictyostelium discoideum cultures and nomenclature
Dictyostelium discoideum AX2-214 was grown in HL5+medium (ForMedium) containing 50 mgml Ampicillin025 mgml Amphotericin-B 100 mgml PenicillinStreptomycin (PAA) at 22C constant light under select-ive conditions as required NC4 cells were grown in a sus-pension of Klebsiella aerogenes in phosphate buffer Whenindicated AX2 cells were also grown in bacterial suspen-sion to allow for comparison with NC4
For cold treatment cells (1 106ml) were shaken at 4Cfor 2ndash14 h Cells were allowed to recover at 22C for 2frac12 hbefore RNA isolation For synchronization cells at adensity of 1ndash3 105ml were incubated at 4C overnight Before cold treatment cells were briefly cooleddown in a water bath with ice For synchronizationcells were then warmed up in a 25C water bath beforecultivating at 22C Synchronization was measured bycounting cells every 30 min during a period of at least7 h Samples for RNA isolation were taken at the timesindicated
Gene names and accession numbers refer to DictyBase(httpdictybaseorg Jan 3rd 2013) Complete sequencesfor tRNAs relevant for the experiments are given inSupplementary Table S2 In silico folded clover leafstructures for tRNAAsp(GUC) tRNAGlu(UUC) andtRNAGly(GCC) are given in Supplementary Figure S1
Nucleotides in tRNAs have been numbered accordingto Motorin et al (19)
Expression and purification of recombinant DnmA andhuman hDnmt2
DnmA and hDnmt2 were both cloned with N-terminalHis6-tags in pET28a(+) (Novagen) into the NdeI andXhoI sites Recombinant proteins were expressed inEscherichia coli (DE3) Rosetta pLysS cells Inductionand protein purification were done as described previously(13) Briefly the cells were induced at an OD600 of 06 with1mM IPTG and incubated for 3 h at 37C for hDnmt2and at 22C for DnmA After harvesting cells were resus-pended in sonication buffer [30 mM KPi (pH 70)300mM KCl 10 (vv) glycerol 10mM imidazole01mM Dithiotreitol (DTT)] containing protease inhibi-tors (complete mini Roche) and lysed by sonication with aDr Hielscher up200s sonifier (5ndash10 times cycle 05ampli-tude 50) The cell lysate was cleared by centrifugation(10000 RCF) and the supernatant was incubated onNiNTA-Sepharose for 15min at 4C After washing withsonication buffer His6-tagged proteins were eluted with abuffer containing 30mM KPi (pH 70) 300mM KCl10 glycerol 200mM imidazole and 01mM DTT Theeluate was dialysed in two steps [30mM KPi (pH 70)200mM KCl 20 glycerol 1mM DTT 01mM EDTAand 30mM KPi (pH 70) 100mM KCl 50 glycerol1mM DTT 01mM EDTA]
2 Nucleic Acids Research 2013
at Universitaumlt W
uumlrzburg on August 5 2013
httpnaroxfordjournalsorgD
ownloaded from
UV-crosslinking and RNA immunoprecipitation
For UV CLIP 2 108 D discoideum cells expressingDnmA-GFP in the dnmAKO-background and cells ex-pressing GFP only were harvested (390 RCF 5min)washed once and resuspended in 20ml of phosphatebuffer Ten microlitre aliquots of the suspension werespread in 10 cm petri dishes and UV irradiated (254 nm)with 250mJcm2 Cells were spun down and collected in4ml of modified RIPA-Buffer [50mM TrisHCl (pH 75)150mM NaCl 05 NP40 05 sodium deoxycholate1mM EDTA] (20) Lysis was supported by three timessonication (cycle 05 and amplitude 50) with a DrHielscher up200s sonifier Samples were centrifuged(20 000 RCF 15min 4C) and the supernatant wastransferred to a fresh tube it was again centrifuged andthe supernatant collected for a third centrifugation stepHereafter the supernatant was pre-incubated with 250 mlSephadexTM G-50 Medium (GE Healthcare) and thentransferred to 50 ml GFP-trap beads (ChromoTekMartinsried) (21) Incubation was carried out for 15minat 4C on a rotating wheel followed by two washing stepswith 15ml stringency RIPA buffer A [50mM TrisHCl(pH 75) 1M NaCl 1 NP40 1 sodium deoxycholate1mM EDTA 01 SDS 2M Urea] and two washingsteps with stringency RIPA buffer B [50mM TrisHCl(pH 75) 1M NaCl 1 NP40 1 sodium deoxycholate1mM EDTA 01 SDS 1M Urea]
For further analysis aliquots were taken from input(supernatant after preincubation with Sephadex G50)and the last wash Beads were equilibrated [50mM TrisHCl (pH 75) 300mM NaCl 05 NP40 05 sodiumdeoxycholate 1mM EDTA 01 SDS] and resuspendedin RIPA elution buffer [50mM TrisHCl (pH 75)300mM NaCl 05 NP40 05 sodium deoxycholate5mM EDTA 05 SDS] To each sample proteinase Kwas added at a final concentration of 1mgml andincubated for 1 h at 42C and 5 h at 56C Samples wereextracted with phenolchloroformisoamyl alcoholsolution (Roth) and RNA was precipitated with isopro-panol and 20 mg of glycogen
Deep sequencing of co-immunoprecipitated RNAs
cDNA synthesis and Illumnia sequencing was carried outaccording to previously published protocols (22)Specifically the two RNA samples from the CLIP experi-ment were poly(A)-tailed using poly(A) polymerase Afterthat the 50PPP was converted into 50P using tobacco acidpyrophosphatase Then an RNA adapter was ligated tothe 50-phosphate of the RNA First-strand cDNA synthe-sis was performed using an oligo(dT)-adapter primer andthe M-MLV reverse transcriptase The resulting cDNAswere PCR-amplified to 20ndash30 ngml using a high-fidelityDNA polymerase Adaptors that were used for ligationare listed in Supplementary Table S1
The cDNA libraries were sequenced using an IlluminaGAIIx machine with 100 cycles resulting in 5 674 433(DnmA-GFP pull down) and 5 540 257 (GFP control)reads After removal of the barcodes and adaptorsequences the reads were aligned to the genome ofD discoideum (sequence accession numbers NC_000895
NC_001889 NC_007087 NC_007088 NC_007089NC_007090 NC_007091 NC_007092) using the shortread mapper segemehl (23) For unmapped readspolyA-tails were computationally searched in the 30-endand clipped if found The resulting sequences weremapped again to the reference sequences In all1 249 579 reads of the DnmA-GFP pulldown and514 809 reads of the control sample could be successfullyaligned Enrichment of RNAs was calculated by compari-son of normalized read numbers from the DnmA-GFPpull down and the control In case of multiple genes re-sulting in the same transcript the reads are mapped to thefirst copy in the genome starting from chromosome 1
Cloning and in vitro transcription of RNAs
The genes for tRNAAsp(GUC-1) (DDB_G0294707)tRNAPhe(GAA-2) (DDB_G0294663) tRNALeu(UAG-1)
(DDB_0234827) tRNALeu(AAG-8) (DDB_0235066)tRNAVal(CAC-1) (DDB_0234789) and tRNAGly(GCC-1)
(DDB_0234894) were cloned into pJET1blunt vector(Fermentas) according to the suppliers suggestions Inthe case of multiple identical genes for one tRNA onlyone gene is listed here The annotation of tRNAs se-quences potential isoacceptor tRNAs and in silicofolding are shown in Supplementary Table S2 andSupplementary Figure S1 Genes were produced by recur-sive PCR using overlapping oligos (Supplementary Table
S1) For tRNAAsp(GUC) tRNALeu(UAG) tRNALeu(AAG)tRNAVal(CAC) and tRNAGly(GCC) the respective oligos 2and 3 served as template and oligo 1 and 4 as primers Arestriction site was added at the 30-end and a T7-promotorat the 50-end For tRNAPhe(GAA) oligo 1 2 and 3 are usedas template Amplification was done with oligo 4 and 5
The tRNAGlu(UUC) (DDB_G0294709) and suppressor
tRNAGlu(CUA) (kindly provided by T Winckler Jena)were recloned into pJET1blunt vector from pGEMTvector The mutated versions tRNAAsp(GUC)C38A andtRNAGlu(UUC)C38A were constructed by using the respect-ive oligonucleotides (Supplementary Table S1)For in vitro transcription 500 ng of linearized plasmid
was incubated at 37C for 2 h in 50 ml with 1 ml T7-RNApolymerase 1mM NTPs 1 transcription buffer(Fermentas) and RiboLockTM RNase Inhibitor(Fermentas) After transcription samples were treatedwith DNaseI for 15min extracted with an equal volumeof phenolchloroformisoamyl alcohol (Roth) and purifiedover a Sephadex G-50 spin columnFor radio-labelled in vitro transcription 2 mg of
linearized plasmid were incubated at 37C for 2 h in100 ml volume with 2 ml T7-RNA polymerase 2mMATP GTP and CTP 1mM UTP and 2 ml [a32P] UTP(110 TBqmmol Hartmann) 1 transcription bufferand RiboLockTM RNase Inhibitor (Fermentas) Aftertranscription samples were treated as aforementioned
Preparation of small RNA fraction
RNA isolation was done using the Trizol-method (24)For enrichment of the small RNA fraction 05M NaCl5 PEG 8000 was added to the RNA solution (in DEPCtreated H2O) and incubated at 20C for 30min After
Nucleic Acids Research 2013 3
at Universitaumlt W
uumlrzburg on August 5 2013
httpnaroxfordjournalsorgD
ownloaded from
centrifugation (10000 RCF 4C 30min) the supernatantwas collected in a fresh tube and precipitated with threevolumes of 100 EtOH
SDS-EMSA
Detection of denaturant-resistant DnmA-tRNA wascarried out by SDS-electrophoretic mobility shift assayas described elsewhere (25) with some modificationsBriefly DnmA (1ndash9 mg) was incubated in 20 ml of 10mMKPi (pH 70) 225 glycerol 25mM KCl 10mM DTT2mM MgCl2 and 100ndash500 mM S-adenosyl-methionine(SAM) with the respective 32P labelled tRNA (100ndash665 ng) at 22C Reactions were terminated at theindicated times by addition of 10 ml of SDS-stop mixture[150mM TrisndashHCl (pH 80) 6 SDS (wv) 30mMEDTA 0025 Bromophenol blue] and heated for10min at 65C Samples were cooled down to room tem-perature and subjected to electrophoresis on 6 or 12SDSndashpolyacrylamide gels sealed in Saran Wrap andexposed to an imager plate for 12ndash40 h at RT Plateswere read in a phosphoimager (Fujifilm FLA-7000) Insome cases samples were transferred from the gel to anitrocellulose membrane for immunodetection of DnmAusing an anti-His antibody and a secondary mouseantibody coupled to alkaline phosphatase
In vitro methylation assay
Methylation of RNA substrates was carried out asdescribed previously (13) 3 mM of recombinant hDnmt2and 3ndash12 mM of DnmA were incubated with 500 ng oftRNA-transcript (or 10 mg of small enriched RNA) 1methylation buffer [50mM TrisHCl (pH 70) 50mMNaCl 5mM MgCl2 1mM DTT] and 17mM [3H]-SAM(Hartmann) 1 uml RiboLockTM RNase Inhibitor(Fermentas) at 22C for 1min to 15 h as indicated inthe text The reaction was stopped by adding 1mM non-radioactive SAM and 2mgml proteinase K for 30minLoading buffer [formamide 5mM EDTA (pH 74)bromophenol blue] was added and samples were run ona 12 denaturing (7M Urea) polyacrylamide gel (1mm)for 2ndash5 h at 400V The gel was stained withethidiumbromide fixed (10 methanol 10 aceticacid) and immersed for 1 h in amplify solution(Amersham) dried and exposed to X-ray film (ContatypCX-BL+ Medical X-ray film) for 16 h up to 2 weeks at80C
In vitro blocked methylation assay
The in vitro blocked methylation assay is based on theassumption that RNA methylation is inhibited if thetarget site is covered by hybridization with a complemen-tary DNA oligo RNA (500 ng of transcript or 10 mg ofsmall enriched RNA) was mixed with 2 mM of the respect-ive oligonucleotide in 1 STE-buffer [10mM TrisHCl(pH 8) 50mM NaCl 1mM EDTA] heated up to 80Cin a thermocycler and then cooled down stepwise to roomtemperature The blocked RNA was subsequently used ina 3H-methylation assay as described earlier in the textBlocking oligos are shown in Supplementary Table S1(oligo 2 for tRNAAsp and tRNAGlu respectively)
dnmA expression analysis by real-time quantitative PCR
For analysing dnmA expression levels RNA was extractedby the Trizol-method (24) qPCR was performed in anEppendorf realplex light cycler with the SensiFAST SYBRNo-ROX One-Step Kit (Bioline) according to the sup-pliers instructions Relative quantification was done withthe Ct-method using gapdh (DDB_G0275153) as anormalization control (housekeeping gene)
Microscopy
For fluorescence analysis cells were fixed in 4 (wv)paraformaldehyde dissolved in 20mM phosphate buffer(pH 67) for 10min at 22C and stained with DAPI
Images were acquired on a Leica DMIRB invertedmicroscope equipped with a DC350 camera and IM50Acquisition software (Leica Microsystems WetzlarGermany)
For the movie DnmA-GFP cells (in DnmAKO back-ground) were analysed with a Zeiss Cell Observer SDwith Rapp UGA-40-2 L Scanner Images of living cellswere taken in 15 s intervals at 7 layers with a 08mMdistance between each layer during a period of 120minFurther image analysis was done with ImageJ 146 Themovie is displayed with eight frames per second
RNA bisulfite sequencing
RNA bisulfite sequencing was carried out as describedelsewhere (26) Primer sequences are listed in theSupplementary Material (Supplementary Table S1)
RESULTS
DnmA is an active tRNAAsp methyltransferase in vitroand in vivo
Previous experiments have shown that hDnmt2 canmethylate a D discoideum RNA in the size range oftRNAs in vitro (13) Here we investigated whether theD discoideum Dnmt2-homologue DnmA is also anactive tRNAAsp methyltransferase In vitro transcribedD discoideum tRNAAsp(GUC) was used as a substrate formethylation by DnmA and hDnmt2 in vitro The assaysshowed that the tRNA was efficiently methylated by bothenzymes (Figure 1A) The reaction was time dependentthe activity was sensitive to Mg2+ concentrations andalmost lost at 10mM Mg2+ In contrast to the humanenzyme DnmA was inactive at 37C (data not shown)Therefore all reactions were carried out at 22C wherehDnmt2 still showed good activity
To investigate methylation in vivo RNA bisulfiteanalysis of tRNAAsp(GUC) from D discoideum wasperformed Figure 1B shows that in wild-type cellstRNAAsp(GUC) was methylated at position C38 to30 whereas methylation was at background levels inDnmA knockout strains Ectopic overexpression ofhDnmt2 in wild-type cells increased tRNAAsp(GUC) C38methylation to 50 indicating cross-species activity ofDnmt2-homologues C49 was methylated at 90 inde-pendently of DnmA and served as an internal control forthe bisulfite reaction
4 Nucleic Acids Research 2013
at Universitaumlt W
uumlrzburg on August 5 2013
httpnaroxfordjournalsorgD
ownloaded from
As tRNAAsp appeared to be not the only target forDnmt2 in other organisms (eg D melanogaster) weexamined if more cellular-derived RNAs could bemethylated in vitro (ex vivo methylation experiments)Total RNA from dnmAKO AX2 cells was subjected toin vitro methylation by recombinant DnmA for varioustimes Methylated RNAs were separated by gelelectro-phoresis and visualized by autoradiography Figure 1Cshows that a substantial RNA fraction in the tRNA sizerange could be methylated At longer incubation times asecond minor methylated RNA species appeared
indicating that at least one further substrate could bemethylated in vitro
Identification of further substratesmdashin vitro binding andmethylation of tRNAGlu
The group of Helm (personal communication) hadproposed a sequence pattern (C32 C34 A37 C40) thatmay serve as a target recognition motif for C38 methyla-tion by hDnmt2 We therefore examined all D discoideumtRNAs in silico to identify candidates for the additionalmethylated band in the ex vivo experiment In addition to
Figure 1 In vitro methylation of tRNAAsp(GUC) (A) In vitro methylation of tRNAAsp(GUC) by DnmA at 2mM 5mM and 10mM MgCl2 and byhDnmt2 at 5mM Mg2+ The upper panel shows ethidium bromide staining of the in vitro transcripts separated in a denaturing polyacrylamide gelthe lower panel shows incorporated 3H-Me in the tRNAs Reactions were run for the times indicated (B) In vivo methylation of cytosines intRNAAsp(GUC) from different D discoideum strains Results of the RNA bisulfite sequencing (454 pyrosequencing) are given in percentage of readsNumbers of sequence reads are shown in brackets C49 is methylated by a different methyltransferase and thus serves as an internal standard All 22tRNAAsp(GUC) genes result in the same transcript and no isoacceptors are encoded in the D discoideum genome (C) In vitro methylation of smallenriched RNA of a dnmAKO strain (ex vivo methylation) The methylation reaction was done for the times indicated
Nucleic Acids Research 2013 5
at Universitaumlt W
uumlrzburg on August 5 2013
httpnaroxfordjournalsorgD
ownloaded from
tRNAAsp(GUC) we found tRNAGlu(UUC) andtRNAGlu(CUC) to match the pattern (Figure 2 andSupplementary Table S2)To test for tRNAGlu as a potential substrate we
modified the methylation assay by addition of a 50 ntDNA antisense oligonucleotide to tRNAAsp(GUC)
covering the methylation target site Figure 3A shows asa control that methylation of in vitro transcribedtRNAAsp(GUC) was completely blocked We then per-formed ex vivo methylation assays with enriched smallRNAs isolated from D discoideum after blockingtRNAAsp(GUC) with the antisense oligo Figure 3B showsthat the majority of 3H-incorporation disappearedHowever with both DnmA and hDnmt2 a significantamount of methylation was still detectable in the sizerange of tRNAs Assuming that this signal containedtRNAGlu we used a second antisense oligo directedagainst tRNAGlu(UUC) for blocking methylation Theoligo also covered the isoacceptor tRNAGlu(CUC) with
minor mismatches (Supplementary Figure S2) A furtherreduction of ex vivo methylation was observed Notablythe lower band in the tRNA size range disappeared (arrowin Figure 3B) suggesting that this represented a tRNAGlu
population that could be methylated at least in vitro Theresidual methylation after blocking with both oligos mayrepresent additional tRNAs or other unknown targets Aswe only concentrated on small RNAs larger methylatedmolecules and less abundant targets may have escapeddetection
To further analyse methylation on tRNAAsp andtRNAGlu (exemplified by tRNAGlu(UUC)) we carried outassays with wild-type sequences and C38A mutatedin vitro transcripts (Figure 4) We also included a suppres-sor tRNAGlu(CUA) that differed in only 2 nt fromtRNAGlu(UUC) (Figure 2) Assays were done in parallelwith the recombinant hDnmt2 and D discoideumDnmA The experiment showed that tRNAAsp(GUC) wasthe best substrate for both DnmA and hDnmt2 (Figure 4)Substantial methylation was also observed fortRNAGlu(UUC) and suppressor tRNAGlu(CUA) Moreoverthe complete loss of methylation in the C38A mutations oftRNAAsp(GUC) and tRNAGlu(UUC) confirmed that methy-lation occurred at position C38 and largely ruled out thatmethyl group incorporation was due to another
Figure 3 Ex vivo methylation and blocking assay (A) Methylation of in vitro transcribed tRNAAsp(GUC) was completely blocked when a comple-mentary antisense oligo was hybridized (B) Using enriched small RNA from dnmAKO cells ex vivo methylation in the tRNA size class wasdifferentially lost when antisense oligos to tRNAAsp(GUC) and tRNAGlu(UUC) were hybridized before the methylation reaction The oligo againsttRNAGlu(UUC) also covers tRNAGlu(CUC) with minor mismatches (see Supplementary Figure S2) Even with both oligos a significant amount of 3Hincorporation still remained The upper panel shows the ethidiumbromide stained gel to demonstrate equal loading the lower panel shows thefluorogram The arrow marks a band that was specifically lost when the anti tRNAGlu(UUC) oligo was used
Figure 2 Target site sequences of tRNAs used for in vitro methylationThe position of the putative signature nucleotides are indicated andprinted in bold if present The anticodon is underlined tRNAGlu(CUC)
(not shown) also harbours the nucleotide pattern Except for the anti-codon both tRNAGlu are identical in the anticodon stem-loop region(Supplementary Figure S2) Only a few differences appear in theacceptor stem
Figure 4 C38 is the target for in vitro methylation of tRNAAsp(GUC)
and tRNAGlu(UUC) In vitro methylation of tRNAAsp(GUC)tRNAGlu(UUC) and a suppressor tRNAGlu(CUA) C38A mutations con-firmed C38 as the target nucleotide tRNAAsp(GUC) methylation wasmost efficient
6 Nucleic Acids Research 2013
at Universitaumlt W
uumlrzburg on August 5 2013
httpnaroxfordjournalsorgD
ownloaded from
contaminating methyltransferase in the recombinantprotein preparation (17)
The methylation experiments using in vitro transcribedtRNAs showed that no other modifications were necessaryfor C38 methylation by DnmA and hDnmt2 (13) and thatthe processed ends of a mature tRNA were not requiredfor target recognition as the tRNA transcripts contain no30-CCA Even tRNAGlu embedded in a long dsRNAmolecule (1000 nt) with small stem-loop structures atthe ends (27) was a specific substrate of DnmA andhDnmt2 (Supplementary Figure S3)
By EMSA no difference in in vitro binding could beseen for the specific target tRNAGlu(UUC) and non-targetRNAs indicating that DnmA associated non-specificallywith any nucleic acid (data not shown) However specificcovalent enzyme-substrate intermediates can be detectedby denaturing SDSndashPAGE (25) Figure 5A andSupplementary Figure S4 show that radioactivelylabelled tRNA transcripts that can be methylatedin vitro by DnmA formed denaturation resistantcomplexes while non-target RNAs did not To confirmthat the denaturation resistant complexes were really dueto binding of the enzyme to the substrate SDS gels wereblotted and His-tagged DnmA was detected by a specificanti-His antibody A small amount of the tagged proteinwas shifted to the same position as the labelled tRNA(Supplementary Figure S5)
We then performed time courses for the reaction andfound that covalent complex formation was only transientunder the conditions used For tRNAAsp(GUC) adductsreached a maximum after 1min and were undetectableafter 15min whereas the tRNAGlu(UUC) complex onlybegan to accumulate after 1min but persisted for almost60min (Figure 5A and B) This coincided with the highmethylation activity on tRNAAsp(GUC-1) and a lower oneon tRNAGlu(UUC-2) The human enzyme appeared to beslightly faster in the reaction but otherwise showed asimilar behaviour compared with D discoideum DnmA
tRNAAsp(GUC) tRNAGlu(UUCCUC) and tRNAGly(GCC)
bind specifically to DnmA in vivo
The blocking assay (Figure 3) suggested that further sub-strates could be targeted by DnmA To identify suchtargets and to further investigate the interaction betweenDnmA and RNAs in vivo we established a UV-CLIP pro-cedure for D discoideumDnmA-GFP expressing cells andcontrol cells expressing GFP only were UV cross-linkedand complexes were precipitated under denaturing condi-tions by a GFP nanobody (21) Then co-precipitatedRNAs were isolated converted into strand-specificcDNA libraries and analyzed by Illumina sequencing(Figure 6 and Supplmentary Figure S6)
Various tRNA fragments were enriched in the DnmACLIP sample compared with the control By their lengthand position fragments appeared not to be random butspecific We found mid-fragments of 20ndash24 nt (position32ndash36 to 53ndash56) that covered the C38 methylation site inthe respective tRNAs 50-fragments of 17 nt (nucleotideposition 1ndash17) 30-fragments of 2021 nt (nucleotideposition 54 to the end) and 30-half tRNAs of 41ndash43 nt
(position 30ndash32 to the end) Mid-fragments oftRNAAsp(GUC) tRNAGlu(UUCCUC) and tRNAGly(GCC)
were significantly enriched in the DnmA-GFP fraction incomparison with the control CLIP assay tRNAAsp(GUC)tRNAGlu(UUC) and tRNAGlu(CUC) harbour the sequencepattern (C32 C34 A37 C40) which has been suggestedas a signature for C38 methylation (Figure 2 Helmpersonal communication) tRNAGly(GCC) contains thepattern except for C40 For tRNAAla(AGC) which doesnot contain C38 and the signature pattern a differentfragment from the 30-end was enriched in the DnmA-CLIP sample For other tRNAs no significant accumula-tion was observed in the experiment (Figure 6 andSupplementary Figure S6) For several tRNAs varyingamounts of full-length molecules were found but thesewere never enriched in the DnmA sample in comparisonwith the control
tRNAGly(GCC) is an additional methylation substratein vitro
The CLIP data suggested that tRNAGly(GCC) was anothertarget for DnmA We therefore cloned this and several othertRNA genes that contained the pattern only partially(Figure 2) We generated in vitro transcripts and examinedmethylation by DnmA and hDnmt2 in vitro IndeedtRNAGly(GCC) was methylated by both enzymes (Figure 7)while none of the other tRNAs (tRNALeu(UAG-1)tRNALeu(AAG-8) tRNAVal(CAC-1) and tRNAPhe(GAA-2))resulted in detectable methylation signals in the assay(Supplementary Figure S7) Interestingly the in vitro methy-lation efficiency of the three tRNA substrates
Figure 5 Formation and turnover of tRNA-methyltransferasecomplexes Turnover of covalent complexes of tRNA substrates withDnmA and hDnmt2 (A) Examples of time courses on covalentcomplex formation with tRNAAsp(GUC) and tRNAGlu(UUC) (B)Complex bands in A were quantified and presented on a time scale
Nucleic Acids Research 2013 7
at Universitaumlt W
uumlrzburg on August 5 2013
httpnaroxfordjournalsorgD
ownloaded from
(tRNAAsp(GUC) tRNAGlu(UUC) tRNAGly(GCC)) correlatedroughly with the number of tRNA fragments found in theRNA-CLIP experimentThe data supported the conclusion from the ex vivo
blocking assay (Figure 3B) that in addition toD discoideum tRNAAsp and tRNAGlu other RNAs weresubstrates for DnmA and hDnmt2 in vitro
Expression of dnmA is regulated during developmentcell cycle and on temperature stress
DnmA expression in growing vegetative D discoideumcells is low To examine tRNA methylation in vivo it
was therefore advantageous to determine conditionsunder which dnmA was most highly expressed Promptedby previous studies (141517) we assumed that the biolo-gical function of DnmA was connected to developmentalandor environmental conditions We therefore examinedexpression levels of dnmA under different regimes dnmAmRNA was quantified by qPCR over the 24 h develop-mental cycle of D discoideum (data not shown) and con-firmed the data deposited in Dictyexpress (httpdictyexpressbiolabsi)
Interestingly an 5-fold increase in expression wasobserved at 16 h in development This time coincides
Figure 6 tRNA and tRNA fragments associated with DnmA-GFP After UV crosslinking RNA bound to DnmA was co-immunoprecipitatedunder denaturing conditions (CLIP) The number of normalised reads for tRNAs detected by Illumina sequencing is shown for DnmA CLIP and forthe control CLIP with GFP In addition to full-length tRNAs four classes of tRNA fragments were found and these are indicated by differentshading of the bars Size and localization of fragments is shown schematically on the simplified clover leaf structure tRNAs that are not significantlyenriched with DnmA are shown in Supplementary Figure S6 In case of multiple gene copys resulting in the same RNA transcript the sequencingreads were mapped to the first copy in the genome starting from chromsome 1 Only this copy is listed Sequences and detailed information on genecopies and potential isoacceptors are listed in Supplementary Table S2
8 Nucleic Acids Research 2013
at Universitaumlt W
uumlrzburg on August 5 2013
httpnaroxfordjournalsorgD
ownloaded from
with a presumed synchronous mitotic division (28) wherea multitude of cell-cycle regulated genes are expressed[(29) and McWilliams personal Communication]
Therefore we synchronized vegetative D discoideumcells to examine dnmA expression during the cell cycleFor these experiments the D discoideum wild-typestrain NC4 the parent of the axenic laboratory strainAX2 was used (30) This was prompted by the observationthat D melanogaster and E histolytica laboratory strainsmay lose dnmt2 expression (Reuter Ankri personalcommunication)
The data confirmed our assumption of a cell-cycle-de-pendent transcriptional regulation of dnmA (Figure 8A)The peak of expression coincided with late mitosis or earlyS-phase as D discoideum does not display a G1 phaseUsing AX2 cells cotransformed with PCNA-RFP andDnmA-GFP we observed GFP accumulation in thenucleus during S phase (indicated by PCNA expression)and a loss of DnmA from the nucleus during mitosis(Figure 8B) This is further exemplified by the movie inthe Supplementary Material (Supplementary Movie S1)
In other organisms dnmt2 was observed to beupregulated during stress response (315) We thereforeexamined expression after anoxia heat shock and coldshock but did not find any significant changes (data notshown) Only during recovery from cold shock wedetected a strong transient increase in dnmA mRNA by47-fold (Figure 8C)
Thus dnmA expression was differentially regulatedduring the cell cycle development and on temperaturestress Furthermore DnmA localization changed specific-ally during the cell cycle
Though the basic expression in vegetative cells was in-distinguishable dnmA expression in NC4 was 30-foldhigher at 16 h of development whereas developmentalupregulation was only 5-fold in AX2 The data aresummarized in Figure 8D
NC4 the parent strain of AX2 displays increasedtRNA
Asp methylation during development
Based on these observations we asked the questionwhether expression levels correlated with tRNA
methylation activity in vivo Bisulfite sequencing wasdone with primers for tRNAAsp tRNAGlu and tRNAGly
on RNA isolated from vegetative AX2 cells AX2 cellsdeveloped for 16 h AX2 cells subjected to cold shockAX2 recovered for 25 h after cold shock vegetativeNC4 cells and NC4 cells developed for 16 h The resultson C38 methylation of tRNAAsp(GUC) are summarized inFigure 8E The methylation status of all C residues fromall tested tRNAs in this study is shown in SupplementaryFigure S8 Indeed tRNAAsp(GUC) methylation at C38increased to 76 in development in NC4 cells Noincrease was observed in AX2 cells during developmentpossibly due to insufficient sensitivity of the assayInterestingly no increase in tRNAAsp(GUC) methylationwas observed at the highest dnmA expression levelsduring stress recovery In Figure 8E the 30ndash35 methy-lation level in all tRNAAsp(GUC) samples represents signifi-cant methylation as compared with other C residues (seeSupplementary Figure S8)We did however not detect any significant methylation
of tRNAGlu(UUCCUC) and tRNAGly(GCC) under any con-ditions (Supplementary Figure S8) In contrast C49C50methylation which does not depend on Dnmt2 enzymeswas found in all samples and thus served as an internalcontrol In all 1175 bisulfite-sequence reads wereexamined for tRNAGlu(UUCCUC) and 4040 fortRNAGly(GCC) No methylation above background wasfound Though it cannot be excluded that the failure todetect C38 methylation of tRNAGlu and tRNAGly was dueto inefficient reverse transcription or low levels of methy-lation (see lsquoDiscussionrsquo section) we conclude thattRNAAsp is the major or possibly the only in vivo targetfor methylation by DnmA in D discoideum
DISCUSSION
The biological function of Dnmt2 methyltransferases islargely unknown and its role in DNA methylation is con-troversially discussed (910) In contrast tRNA methyla-tion by Dnmt2 is robust in vitro and in vivo andevolutionary conserved In addition to the originallydescribed tRNAAsp(GUC) (1) two further tRNAs targets(tRNAVal(CAC) and tRNAGly(GCC)) have been defined inD melanogaster (15) and tRNAGlu(UUC) in S pombe (17)To get insight into potential functions of the
D discoideum Dnmt2 homologue DnmA we examinedexpression levels in development in the cell cycle andunder different environmental conditions We confirmedby qPCR an expression peak at 16 h of developmentWeijer and co-workers (28) had previously proposed asynchronous cell division at this developmental stageWe therefore investigated dnmA expression in the cellcycle and found a strong transient increase of mRNA inS-phase or shortly thereafter The expression peak wasaccompanied by reaccumulation of DnmA in thenucleus We assume that DnmA is degraded duringmitosis and re-synthesised after cell divisionIn S pombe as well as in D melanogaster tRNA methy-
lation changes in response to environmental challengeslike temperature stress anoxia or nutrients We tested
Figure 7 In vitro methylation of tRNAGly(GCC) by recombinant DnmAand hDnmt2 In vitro transcribed tRNAs were methylated in vitro asdescribed before The top panel shows the ethidium-bromide stainedgel the bottom panel the fluorogram of 3H-labelled methylated tRNAsAs a control tRNAAsp(GUC) methylation is shown
Nucleic Acids Research 2013 9
at Universitaumlt W
uumlrzburg on August 5 2013
httpnaroxfordjournalsorgD
ownloaded from
Figure 8 Expression localization and tRNAAsp(GUC) methylation activity of DnmA under various conditions (A) dnmA expression is regulatedduring the cell cycle Cells were arrested in the cell cycle by cold treatment and then released by transfer to 22C Cells were counted every 30min(grey dots and grey line) and samples were taken for qPCR (black bars) After cell division dnmA expression increased 5-fold and then rapidlydeclined to basal levels Normalization was done on vegetative growing cells dnmA expression is only shown from 3 to 45 h during recovery(B) DnmA is lost from the nucleus during mitosis (arrow) The three cells on the right are in S-phase as indicated by the RFP-PCNA marker andaccumulate DnmA in the nucleus (for further details see movie in Supplementary Material) (C) Relative quantification of dnmA expression levels inAX2 cells after 25 h recovery from cold shock at 4C Expression of dnmA increased gt40-fold and returned within 30min to basal levels (n=3)(D) At 16 h of development dnmA expression increased 46-fold in the NC4 strain In AX2 cells expression increased only 5-fold (E) At 16 h ofdevelopment in vivo methylation of C38 in tRNAAsp(GUC) increased up to 75 in the D discoideum strain NC4 whereas no significant increase wasobserved in AX2 cells
10 Nucleic Acids Research 2013
at Universitaumlt W
uumlrzburg on August 5 2013
httpnaroxfordjournalsorgD
ownloaded from
heat shock cold shock anoxia and various nutrient mediabut did not observe altered levels Only during therecovery phase after cold treatment a transient almost50-fold increase in mRNA abundance was seen
Reuter and Ankri (personal communication) hadobserved a loss of Dnmt2 expression in laboratorystrains of D melanogaster and E histolytica We thereforeexamined D discoideum NC4 the parent of the axeniclaboratory strain AX2 The developmental regulation inAX2 and NC4 was qualitatively similar but expressionlevels were much higher in NC4 The observationsupports the assumption that Dnmt2 expression wasreduced under optimal growth conditions in the labora-tory strain and may actually be selected against
We show that the D discoideum Dnmt2 homologueDnmA is an active tRNAAsp(GUC) methyltransferasein vitro and in vivo To identify further potential targetsfor DnmA we established a UV-CLIP method to analyseDnmAndashRNA interaction in vivo In addition to the knowntarget tRNAAsp(GUC) fragments of tRNAGlu(UUC) andtRNAGly(GCC) were most abundant in comparison withthe control Unexpectedly complete tRNAs were notenriched in the DnmA CLIP fraction but rather specificlsquomid-fragmentsrsquo that covered the putative methylation siteand the recognition pattern proposed by Helm (personal
communication) A minor exception was tRNAGly(GCC)which lacks C40 We do not know whether these frag-ments were directly or indirectly generated by DnmAin vivo or whether they were protected by DnmA fromdegradation during the experimental procedure As noco-crystal of Dnmt2 and a tRNA substrate is availablethe exact interface for the interaction is unknownJurkowski et al (31) mapped the binding site on Dnmt2by mutational analysis A manual placement of a tRNAinto the crystal structure suggests that Dnmt2 mainlycontacts the anticodon stem loop Assuming that mid-frag-ments are footprints of DnmA our data support this notionbut suggest that the interaction surface extends into theD-loop which is partially covered by the mid-fragments
We examined the putative DnmA targets in vitro (bymethylation of in vitro transcribed tRNAs) ex vivo(by methylation of RNA isolated from D discoideum)and in vivo (by bisulfite sequencing) Furthermore weinvestigated by SDSndashPAGE the kinetics of covalentcomplex formation between tRNAAsp and tRNAGlu withDnmA and hDnmt2
All three tRNAs (tRNAAsp tRNAGlu tRNAGly) werereadily methylated by recombinant DnmA and hDnmt2in vitro although the activity was much higher (estimatedat least 10-fold) on tRNAAsp compared with tRNAGlu andtRNAGly For tRNAAsp and tRNAGlu this correlated wellwith the formation and decay of covalent RNAmdashproteincomplexes where kinetics for tRNAAsp were much fasterthan for tRNAGlu Instead of C methylation the covalentintermediate may also proceed to a hydrogen exchangereaction or to a deamination of the cytosine In bothcases an intermediate will be detected but it will notresult in a methylated product (32) It may be thatslower off rates represent these alternative pathways thatwould result in unmethylated tRNAs
Using antisense oligos we showed that tRNAAsptRNAGlu and at least one additional RNA could bemethylated ex vivo A quantification of the results wasnot possible the apparently low methylation oftRNAGlu ex vivo could be due to the low activity of theenzyme on this substrate (as suggested by the in vitroexperiments see Figure 5B)The data for the examination of in vivo methylation by
bisulfite sequencing were rather unexpected AlthoughtRNAAsp(GUC) was consistently found to be methylatedat C38 we never detected any methylation at C38 oftRNAGlu(UUCCUC) or tRNAGly(GCC) There could betwo reasons to explain the failure to detect C38 methyla-tion for these in vitro substrates (i) due to the low activityof DnmA on tRNAGlu(UUCCUC) and tRNAGly(GCC) asshown in vitro there was low or no significant methylationin vivo and (ii) methylation escaped detection becauseother unknown modifications blocked reverse transcrip-tion of the methylated molecules However if this wasthe case one has to assume that C49C50 methylationoccurs independently of these unknown modificationswhereas C38 methylation strictly depends on them AsC38 methylation does not require any other modificationsin vitro we believe that this scenario is less likely If in vitromethylation activity was comparable with in vivo methy-lation 10 methylation of tRNAGlu(UUCCUC) andtRNAGly(GCC) could be expected relative totRNAAsp(GUC) As tRNAAsp was methylated to 30 a3 methylation of tRNAGlu and tRNAGly could be belowthe detection level of the bisulfite assayWe find that high expression of dnmA did not necessar-
ily correlate with increased methylation of tRNAs Duringrecovery from cold shock D discoideum cells displayedthe highest level of dnmA expression but this had noeffect on tRNAAsp methylation Only in developmentand with the high expressing NC4 strain a significantincrease of tRNAAsp methylation to 70 was observedIt has to be considered however that bisulfite sequencingdoes not necessarily allow for quantitative evaluation ofdata detected C-methylation depends on reverse tran-scription efficiency which may be hampered by othermodifications that could be different under differentconditions Furthermore the completeness of bisulfite-mediated conversion could also depend on RNA struc-ture which may be altered by other modificationsHowever incomplete conversion will lead to an overesti-mation of C-methylation as maybe the case for forexample C30 C31 C35 in tRNAGly in some samplesTaken together the data from CLIP complex forma-
tion and methylation show that all three substrates can bebound and methylated in vitro The CLIP data show thatthere is a specific interaction of DnmA with tRNA mid-fragments of tRNAAsp(GUC) tRNAGlu(UUCCUC) andtRNAGly(GCC) but not with other tRNAs In contrastbisulfite sequencing could only detect methylation ontRNAAsp(GUC) even at the highest expression levels ofdnmA Even though low amounts of methylation mayhave escaped detection in bisulfite sequencing there isstill a discrepancy between dnmA expression levels (egduring cold shock recovery) DnmA binding to substrates(as determined by CLIP) and methylation in vivo
Nucleic Acids Research 2013 11
at Universitaumlt W
uumlrzburg on August 5 2013
httpnaroxfordjournalsorgD
ownloaded from
We therefore suggest that DnmA binding to specifictargets could elicit functions in addition to RNA methy-lation eg by promoting processing or degradation andor recruitment of other proteins At least for some rRNAmethyltransferases such as Bud23 or Nep1 fromSaccharomyces cerevisiae it is known that they executefunctions independent from the methylation activity(33ndash35) As shown by Schaefer and co-workers (15)methylation protects tRNAs from degradation or process-ing after stress By a different pathway binding of DnmAmay initiate processing that leads to specific tRNA frag-ments that could serve for other biological functions (36)Interestingly tRNAGlu(UUC) is also a substrate for theDnmt2 homologue Pmt1 in S pombe (17) HowevertRNAGlu(UUC) is found to be methylated to a low butsignificant extend in vivo in fission yeast when Pmt1 isoverexpressedThe detection of tRNAAla(AGC) fragments in the CLIP
assay brings up a new question tRNAAla(AGC) has a Tinstead of a C in position 38 and does not contain thesignature nucleotides in the anticodon stem-loopInstead of a mid-fragment a distinct 30 fragment wasassociated with DnmA suggesting that a recognitionmotif was located in this part of the moleculeAs methylation also occurred in a large artificial
molecule with embedded tRNAGlu (SupplementaryFigure S2) it could be that other longer RNAs containingan appropriate recognition motif may also be bound bythe enzyme We thus propose that the biological functionsof Dnmt2 proteins could extend far beyond our presentknowledge Investigation of further targets and of the sig-nificance of RNA fragments associated with Dnmt2should shed light on the importance of this enzyme
SUPPLEMENTARY DATA
Supplementary Data are available at NAR Online
ACKNOWLEDGEMENTS
The authors thank Thomas Winckler (Friedrich-Schiller-University Jena) for providing the tRNAGlu(UUC) and sup-pressor tRNAGlu(CUA) plasmids Susanne Junk for cloningtRNAAspC38A Matthias Schafer (DKFZ) and RobinLorenz (University of Kassel) are acknowledged forRNA bisulfite sequencing and Richard Reinhardt (MaxPlanck Genome Centre Cologne) for deep sequencingMark Helm (Johannes-Gutenberg-University Mainz) gen-erously provided data before publication CarstenSeehafer (University of Kassel) helped with the analysisof deep-sequencing data
FUNDING
Deutsche Forschungsgemeinschaft [FOR 1082 to WNand AJ] and the Studienstiftung des Deutschen Volkes(to SM) Funding for open access charge FOR1082
Conflict of interest statement None declared
REFERENCES
1 GollMG KirpekarF MaggertKA YoderJA HsiehCLZhangX GolicKG JacobsenSE and BestorTH (2006)Methylation of tRNAAsp by the DNA methyltransferasehomolog Dnmt2 Science 311 395ndash398
2 PhalkeS NickelO WalluscheckD HortigF OnoratiMCand ReuterG (2009) Retrotransposon silencing and telomereintegrity in somatic cells of Drosophila depends on the cytosine-5methyltransferase DNMT2 Nat Genet 41 696ndash702
3 WilkinsonCR BartlettR NurseP and BirdAP (1995) Thefission yeast gene pmt1+ encodes a DNA methyltransferasehomologue Nucleic Acids Res 23 203ndash210
4 KatohM CurkT XuQ ZupanB KuspaA and ShaulskyG(2006) Developmentally regulated DNA methylation inDictyostelium discoideum Eukaryot Cell 5 18ndash25
5 KuhlmannM BorisovaBE KallerM LarssonP StachDNaJ EichingerL LykoF AmbrosV SoderbomF et al(2005) Silencing of retrotransposons in Dictyostelium by DNAmethylation and RNAi Nucleic Acids Res 33 6405ndash6417
6 RaiK ChidesterS ZavalaCV ManosEJ JamesSRKarpfAR JonesDA and CairnsBR (2007) Dnmt2 functionsin the cytoplasm to promote liver brain and retina developmentin zebrafish Genes Dev 21 261ndash266
7 FisherO Siman-TovR and AnkriS (2006) Pleiotropicphenotype in Entamoeba histolytica overexpressing DNAmethyltransferase (Ehmeth) Mol Biochem Parasitol 147 48ndash54
8 DurdevicZ HannaK GoldB PollexT CherryS LykoFand SchaeferM (2013) Efficient RNA virus control in Drosophilarequires the RNA methyltransferase Dnmt2 EMBO Rep 14269ndash275
9 SchaeferM and LykoF (2010) Lack of evidence for DNAmethylation of Invader4 retroelements in Drosophila andimplications for Dnmt2-mediated epigenetic regulation NatGenet 42 920ndash921 author reply 921
10 KraussV and ReuterG (2011) DNA methylation in Drosophila -a critical evaluation Prog Mol Biol Transl Sci 101 177ndash191
11 RaddatzG GuzzardoPM OlovaN FantappieMRRamppM SchaeferM ReikW HannonGJ and LykoF(2013) Dnmt2-dependent methylomes lack defined DNAmethylation patterns Proc Natl Acad Sci USA 110 8627ndash8631
12 JeltschA NellenW and LykoF (2006) Two substrates arebetter than one dual specificities for Dnmt2 methyltransferasesTrends Biochem Sci 31 306ndash308
13 JurkowskiTP MeusburgerM PhalkeS HelmM NellenWReuterG and JeltschA (2008) Human DNMT2 methylatestRNA(Asp) molecules using a DNA methyltransferase-likecatalytic mechanism RNA 14 1663ndash1670
14 TovyA Siman TovR GaentzschR HelmM and AnkriS(2010) A new nuclear function of the Entamoeba histolyticaglycolytic enzyme enolase the metabolic regulation of cytosine-5methyltransferase 2 (Dnmt2) activity PLoS Pathog 6 e1000775
15 SchaeferM PollexT HannaK TuortoF MeusburgerMHelmM and LykoF (2010) RNA methylation by Dnmt2protects transfer RNAs against stress-induced cleavage GenesDev 24 1590ndash1595
16 TuortoF LiebersR MuschT SchaeferM HofmannSKellnerS FryeM HelmM StoecklinG and LykoF (2012)RNA cytosine methylation by Dnmt2 and NSun2 promotestRNA stability and protein synthesis Nat Struct Mol Biol 19900ndash905
17 BeckerM MullerS NellenW JurkowskiTP JeltschA andEhrenhofer-MurrayAE (2012) Pmt1 a Dnmt2 homolog inSchizosaccharomyces pombe mediates tRNA methylation inresponse to nutrient signaling Nucleic Acids Res 4011648ndash11658
18 TovyA HertzR Siman-TovR SyanS FaustD GuillenNand AnkriS (2011) Glucose starvation boosts Entamoebahistolytica virulence PLoS Negl Trop Dis 5 e1247
19 MotorinY and HelmM (2011) RNA nucleotide methylationWiley Interdiscip Rev RNA 2 611ndash631
20 NiranjanakumariS LasdaE BrazasR and Garcia-BlancoMA(2002) Reversible cross-linking combined with
12 Nucleic Acids Research 2013
at Universitaumlt W
uumlrzburg on August 5 2013
httpnaroxfordjournalsorgD
ownloaded from
immunoprecipitation to study RNA-protein interactions in vivoMethods 26 182ndash190
21 RothbauerU ZolghadrK MuyldermansS SchepersACardosoMC and LeonhardtH (2008) A versatile nanotrapfor biochemical and functional studies with fluorescent fusionproteins Mol Cell Proteomics 7 282ndash289
22 SittkaA LucchiniS PapenfortK SharmaCM RolleKBinnewiesTT HintonJC and VogelJ (2008) Deep sequencinganalysis of small noncoding RNA and mRNA targets of theglobal post-transcriptional regulator Hfq PLoS Genet 4e1000163
23 HoffmannS OttoC KurtzS SharmaCM KhaitovichPVogelJ StadlerPF and HackermullerJ (2009) Fast mapping ofshort sequences with mismatches insertions and deletions usingindex structures PLoS Comput Biol 5 e1000502
24 ConnollyMA ClausenPA and LazarJG (2006) Purificationof RNA from animal cells using trizol Cold Spring HarborProtoc 2006 pii pdbprot4104
25 DongA YoderJA ZhangX ZhouL BestorTH andChengX (2001) Structure of human DNMT2 an enigmaticDNA methyltransferase homolog that displays denaturant-resistant binding to DNA Nucleic Acids Res 29 439ndash448
26 SchaeferM PollexT HannaK and LykoF (2009) RNAcytosine methylation analysis by bisulfite sequencing NucleicAcids Res 37 e12
27 BoninM OberstrassJ VogtU WasseneggerM and NellenW(2001) Binding of IRE-BP to its cognate RNA sequence SFMstudies on a universal RNA backbone for the analysis of RNA-protein interaction Biol Chem 382 1157ndash1162
28 ZimmermanW and WeijerCJ (1993) Analysis of cell cycleprogression during the development of Dictyostelium and itsrelationship to differentiation Dev Biol 160 178ndash185
29 StrasserK BloomfieldG MacWilliamsA CeccarelliAMacWilliamsH and TsangA (2012) A retinoblastomaorthologue is a major regulator of S-phase mitotic anddevelopmental gene expression in Dictyostelium PLoS One 7e39914
30 WattsDJ and AshworthJM (1970) Growth of myxameobae ofthe cellular slime mould Dictyostelium discoideum in axenicculture Biochem J 119 171ndash174
31 JurkowskiTP ShanmugamR HelmM and JeltschA (2012)Mapping the tRNA binding site on the surface of humanDNMT2 methyltransferase Biochemistry 51 4438ndash4444
32 SvedruzicZM and ReichNO (2005) DNA cytosine C5methyltransferase Dnmt1 catalysis-dependent release of allostericinhibition Biochemistry 44 9472ndash9485
33 ThomasSR KellerCA SzykA CannonJR and Laronde-LeblancNA (2011) Structural insight into the functionalmechanism of Nep1Emg1 N1-specific pseudouridinemethyltransferase in ribosome biogenesis Nucleic Acids Res 392445ndash2457
34 LeulliotN BohnsackMT GrailleM TollerveyD andVan TilbeurghH (2008) The yeast ribosome synthesisfactor Emg1 is a novel member of the superfamily of alphabeta knot fold methyltransferases Nucleic Acids Res 36629ndash639
35 WhiteJ LiZ SardanaR BujnickiJM MarcotteEM andJohnsonAW (2008) Bud23 methylates G1575 of 18S rRNA andis required for efficient nuclear export of pre-40S subunits MolCell Biol 28 3151ndash3161
36 SobalaA and HutvagnerG (2013) Small RNAs derived fromthe 50-end of tRNA can inhibit protein translation in humancells RNA Biol 10 525ndash535
Nucleic Acids Research 2013 13
at Universitaumlt W
uumlrzburg on August 5 2013
httpnaroxfordjournalsorgD
ownloaded from

UV-crosslinking and RNA immunoprecipitation
For UV CLIP 2 108 D discoideum cells expressingDnmA-GFP in the dnmAKO-background and cells ex-pressing GFP only were harvested (390 RCF 5min)washed once and resuspended in 20ml of phosphatebuffer Ten microlitre aliquots of the suspension werespread in 10 cm petri dishes and UV irradiated (254 nm)with 250mJcm2 Cells were spun down and collected in4ml of modified RIPA-Buffer [50mM TrisHCl (pH 75)150mM NaCl 05 NP40 05 sodium deoxycholate1mM EDTA] (20) Lysis was supported by three timessonication (cycle 05 and amplitude 50) with a DrHielscher up200s sonifier Samples were centrifuged(20 000 RCF 15min 4C) and the supernatant wastransferred to a fresh tube it was again centrifuged andthe supernatant collected for a third centrifugation stepHereafter the supernatant was pre-incubated with 250 mlSephadexTM G-50 Medium (GE Healthcare) and thentransferred to 50 ml GFP-trap beads (ChromoTekMartinsried) (21) Incubation was carried out for 15minat 4C on a rotating wheel followed by two washing stepswith 15ml stringency RIPA buffer A [50mM TrisHCl(pH 75) 1M NaCl 1 NP40 1 sodium deoxycholate1mM EDTA 01 SDS 2M Urea] and two washingsteps with stringency RIPA buffer B [50mM TrisHCl(pH 75) 1M NaCl 1 NP40 1 sodium deoxycholate1mM EDTA 01 SDS 1M Urea]
For further analysis aliquots were taken from input(supernatant after preincubation with Sephadex G50)and the last wash Beads were equilibrated [50mM TrisHCl (pH 75) 300mM NaCl 05 NP40 05 sodiumdeoxycholate 1mM EDTA 01 SDS] and resuspendedin RIPA elution buffer [50mM TrisHCl (pH 75)300mM NaCl 05 NP40 05 sodium deoxycholate5mM EDTA 05 SDS] To each sample proteinase Kwas added at a final concentration of 1mgml andincubated for 1 h at 42C and 5 h at 56C Samples wereextracted with phenolchloroformisoamyl alcoholsolution (Roth) and RNA was precipitated with isopro-panol and 20 mg of glycogen
Deep sequencing of co-immunoprecipitated RNAs
cDNA synthesis and Illumnia sequencing was carried outaccording to previously published protocols (22)Specifically the two RNA samples from the CLIP experi-ment were poly(A)-tailed using poly(A) polymerase Afterthat the 50PPP was converted into 50P using tobacco acidpyrophosphatase Then an RNA adapter was ligated tothe 50-phosphate of the RNA First-strand cDNA synthe-sis was performed using an oligo(dT)-adapter primer andthe M-MLV reverse transcriptase The resulting cDNAswere PCR-amplified to 20ndash30 ngml using a high-fidelityDNA polymerase Adaptors that were used for ligationare listed in Supplementary Table S1
The cDNA libraries were sequenced using an IlluminaGAIIx machine with 100 cycles resulting in 5 674 433(DnmA-GFP pull down) and 5 540 257 (GFP control)reads After removal of the barcodes and adaptorsequences the reads were aligned to the genome ofD discoideum (sequence accession numbers NC_000895
NC_001889 NC_007087 NC_007088 NC_007089NC_007090 NC_007091 NC_007092) using the shortread mapper segemehl (23) For unmapped readspolyA-tails were computationally searched in the 30-endand clipped if found The resulting sequences weremapped again to the reference sequences In all1 249 579 reads of the DnmA-GFP pulldown and514 809 reads of the control sample could be successfullyaligned Enrichment of RNAs was calculated by compari-son of normalized read numbers from the DnmA-GFPpull down and the control In case of multiple genes re-sulting in the same transcript the reads are mapped to thefirst copy in the genome starting from chromosome 1
Cloning and in vitro transcription of RNAs
The genes for tRNAAsp(GUC-1) (DDB_G0294707)tRNAPhe(GAA-2) (DDB_G0294663) tRNALeu(UAG-1)
(DDB_0234827) tRNALeu(AAG-8) (DDB_0235066)tRNAVal(CAC-1) (DDB_0234789) and tRNAGly(GCC-1)
(DDB_0234894) were cloned into pJET1blunt vector(Fermentas) according to the suppliers suggestions Inthe case of multiple identical genes for one tRNA onlyone gene is listed here The annotation of tRNAs se-quences potential isoacceptor tRNAs and in silicofolding are shown in Supplementary Table S2 andSupplementary Figure S1 Genes were produced by recur-sive PCR using overlapping oligos (Supplementary Table
S1) For tRNAAsp(GUC) tRNALeu(UAG) tRNALeu(AAG)tRNAVal(CAC) and tRNAGly(GCC) the respective oligos 2and 3 served as template and oligo 1 and 4 as primers Arestriction site was added at the 30-end and a T7-promotorat the 50-end For tRNAPhe(GAA) oligo 1 2 and 3 are usedas template Amplification was done with oligo 4 and 5
The tRNAGlu(UUC) (DDB_G0294709) and suppressor
tRNAGlu(CUA) (kindly provided by T Winckler Jena)were recloned into pJET1blunt vector from pGEMTvector The mutated versions tRNAAsp(GUC)C38A andtRNAGlu(UUC)C38A were constructed by using the respect-ive oligonucleotides (Supplementary Table S1)For in vitro transcription 500 ng of linearized plasmid
was incubated at 37C for 2 h in 50 ml with 1 ml T7-RNApolymerase 1mM NTPs 1 transcription buffer(Fermentas) and RiboLockTM RNase Inhibitor(Fermentas) After transcription samples were treatedwith DNaseI for 15min extracted with an equal volumeof phenolchloroformisoamyl alcohol (Roth) and purifiedover a Sephadex G-50 spin columnFor radio-labelled in vitro transcription 2 mg of
linearized plasmid were incubated at 37C for 2 h in100 ml volume with 2 ml T7-RNA polymerase 2mMATP GTP and CTP 1mM UTP and 2 ml [a32P] UTP(110 TBqmmol Hartmann) 1 transcription bufferand RiboLockTM RNase Inhibitor (Fermentas) Aftertranscription samples were treated as aforementioned
Preparation of small RNA fraction
RNA isolation was done using the Trizol-method (24)For enrichment of the small RNA fraction 05M NaCl5 PEG 8000 was added to the RNA solution (in DEPCtreated H2O) and incubated at 20C for 30min After
Nucleic Acids Research 2013 3
at Universitaumlt W
uumlrzburg on August 5 2013
httpnaroxfordjournalsorgD
ownloaded from
centrifugation (10000 RCF 4C 30min) the supernatantwas collected in a fresh tube and precipitated with threevolumes of 100 EtOH
SDS-EMSA
Detection of denaturant-resistant DnmA-tRNA wascarried out by SDS-electrophoretic mobility shift assayas described elsewhere (25) with some modificationsBriefly DnmA (1ndash9 mg) was incubated in 20 ml of 10mMKPi (pH 70) 225 glycerol 25mM KCl 10mM DTT2mM MgCl2 and 100ndash500 mM S-adenosyl-methionine(SAM) with the respective 32P labelled tRNA (100ndash665 ng) at 22C Reactions were terminated at theindicated times by addition of 10 ml of SDS-stop mixture[150mM TrisndashHCl (pH 80) 6 SDS (wv) 30mMEDTA 0025 Bromophenol blue] and heated for10min at 65C Samples were cooled down to room tem-perature and subjected to electrophoresis on 6 or 12SDSndashpolyacrylamide gels sealed in Saran Wrap andexposed to an imager plate for 12ndash40 h at RT Plateswere read in a phosphoimager (Fujifilm FLA-7000) Insome cases samples were transferred from the gel to anitrocellulose membrane for immunodetection of DnmAusing an anti-His antibody and a secondary mouseantibody coupled to alkaline phosphatase
In vitro methylation assay
Methylation of RNA substrates was carried out asdescribed previously (13) 3 mM of recombinant hDnmt2and 3ndash12 mM of DnmA were incubated with 500 ng oftRNA-transcript (or 10 mg of small enriched RNA) 1methylation buffer [50mM TrisHCl (pH 70) 50mMNaCl 5mM MgCl2 1mM DTT] and 17mM [3H]-SAM(Hartmann) 1 uml RiboLockTM RNase Inhibitor(Fermentas) at 22C for 1min to 15 h as indicated inthe text The reaction was stopped by adding 1mM non-radioactive SAM and 2mgml proteinase K for 30minLoading buffer [formamide 5mM EDTA (pH 74)bromophenol blue] was added and samples were run ona 12 denaturing (7M Urea) polyacrylamide gel (1mm)for 2ndash5 h at 400V The gel was stained withethidiumbromide fixed (10 methanol 10 aceticacid) and immersed for 1 h in amplify solution(Amersham) dried and exposed to X-ray film (ContatypCX-BL+ Medical X-ray film) for 16 h up to 2 weeks at80C
In vitro blocked methylation assay
The in vitro blocked methylation assay is based on theassumption that RNA methylation is inhibited if thetarget site is covered by hybridization with a complemen-tary DNA oligo RNA (500 ng of transcript or 10 mg ofsmall enriched RNA) was mixed with 2 mM of the respect-ive oligonucleotide in 1 STE-buffer [10mM TrisHCl(pH 8) 50mM NaCl 1mM EDTA] heated up to 80Cin a thermocycler and then cooled down stepwise to roomtemperature The blocked RNA was subsequently used ina 3H-methylation assay as described earlier in the textBlocking oligos are shown in Supplementary Table S1(oligo 2 for tRNAAsp and tRNAGlu respectively)
dnmA expression analysis by real-time quantitative PCR
For analysing dnmA expression levels RNA was extractedby the Trizol-method (24) qPCR was performed in anEppendorf realplex light cycler with the SensiFAST SYBRNo-ROX One-Step Kit (Bioline) according to the sup-pliers instructions Relative quantification was done withthe Ct-method using gapdh (DDB_G0275153) as anormalization control (housekeeping gene)
Microscopy
For fluorescence analysis cells were fixed in 4 (wv)paraformaldehyde dissolved in 20mM phosphate buffer(pH 67) for 10min at 22C and stained with DAPI
Images were acquired on a Leica DMIRB invertedmicroscope equipped with a DC350 camera and IM50Acquisition software (Leica Microsystems WetzlarGermany)
For the movie DnmA-GFP cells (in DnmAKO back-ground) were analysed with a Zeiss Cell Observer SDwith Rapp UGA-40-2 L Scanner Images of living cellswere taken in 15 s intervals at 7 layers with a 08mMdistance between each layer during a period of 120minFurther image analysis was done with ImageJ 146 Themovie is displayed with eight frames per second
RNA bisulfite sequencing
RNA bisulfite sequencing was carried out as describedelsewhere (26) Primer sequences are listed in theSupplementary Material (Supplementary Table S1)
RESULTS
DnmA is an active tRNAAsp methyltransferase in vitroand in vivo
Previous experiments have shown that hDnmt2 canmethylate a D discoideum RNA in the size range oftRNAs in vitro (13) Here we investigated whether theD discoideum Dnmt2-homologue DnmA is also anactive tRNAAsp methyltransferase In vitro transcribedD discoideum tRNAAsp(GUC) was used as a substrate formethylation by DnmA and hDnmt2 in vitro The assaysshowed that the tRNA was efficiently methylated by bothenzymes (Figure 1A) The reaction was time dependentthe activity was sensitive to Mg2+ concentrations andalmost lost at 10mM Mg2+ In contrast to the humanenzyme DnmA was inactive at 37C (data not shown)Therefore all reactions were carried out at 22C wherehDnmt2 still showed good activity
To investigate methylation in vivo RNA bisulfiteanalysis of tRNAAsp(GUC) from D discoideum wasperformed Figure 1B shows that in wild-type cellstRNAAsp(GUC) was methylated at position C38 to30 whereas methylation was at background levels inDnmA knockout strains Ectopic overexpression ofhDnmt2 in wild-type cells increased tRNAAsp(GUC) C38methylation to 50 indicating cross-species activity ofDnmt2-homologues C49 was methylated at 90 inde-pendently of DnmA and served as an internal control forthe bisulfite reaction
4 Nucleic Acids Research 2013
at Universitaumlt W
uumlrzburg on August 5 2013
httpnaroxfordjournalsorgD
ownloaded from
As tRNAAsp appeared to be not the only target forDnmt2 in other organisms (eg D melanogaster) weexamined if more cellular-derived RNAs could bemethylated in vitro (ex vivo methylation experiments)Total RNA from dnmAKO AX2 cells was subjected toin vitro methylation by recombinant DnmA for varioustimes Methylated RNAs were separated by gelelectro-phoresis and visualized by autoradiography Figure 1Cshows that a substantial RNA fraction in the tRNA sizerange could be methylated At longer incubation times asecond minor methylated RNA species appeared
indicating that at least one further substrate could bemethylated in vitro
Identification of further substratesmdashin vitro binding andmethylation of tRNAGlu
The group of Helm (personal communication) hadproposed a sequence pattern (C32 C34 A37 C40) thatmay serve as a target recognition motif for C38 methyla-tion by hDnmt2 We therefore examined all D discoideumtRNAs in silico to identify candidates for the additionalmethylated band in the ex vivo experiment In addition to
Figure 1 In vitro methylation of tRNAAsp(GUC) (A) In vitro methylation of tRNAAsp(GUC) by DnmA at 2mM 5mM and 10mM MgCl2 and byhDnmt2 at 5mM Mg2+ The upper panel shows ethidium bromide staining of the in vitro transcripts separated in a denaturing polyacrylamide gelthe lower panel shows incorporated 3H-Me in the tRNAs Reactions were run for the times indicated (B) In vivo methylation of cytosines intRNAAsp(GUC) from different D discoideum strains Results of the RNA bisulfite sequencing (454 pyrosequencing) are given in percentage of readsNumbers of sequence reads are shown in brackets C49 is methylated by a different methyltransferase and thus serves as an internal standard All 22tRNAAsp(GUC) genes result in the same transcript and no isoacceptors are encoded in the D discoideum genome (C) In vitro methylation of smallenriched RNA of a dnmAKO strain (ex vivo methylation) The methylation reaction was done for the times indicated
Nucleic Acids Research 2013 5
at Universitaumlt W
uumlrzburg on August 5 2013
httpnaroxfordjournalsorgD
ownloaded from
tRNAAsp(GUC) we found tRNAGlu(UUC) andtRNAGlu(CUC) to match the pattern (Figure 2 andSupplementary Table S2)To test for tRNAGlu as a potential substrate we
modified the methylation assay by addition of a 50 ntDNA antisense oligonucleotide to tRNAAsp(GUC)
covering the methylation target site Figure 3A shows asa control that methylation of in vitro transcribedtRNAAsp(GUC) was completely blocked We then per-formed ex vivo methylation assays with enriched smallRNAs isolated from D discoideum after blockingtRNAAsp(GUC) with the antisense oligo Figure 3B showsthat the majority of 3H-incorporation disappearedHowever with both DnmA and hDnmt2 a significantamount of methylation was still detectable in the sizerange of tRNAs Assuming that this signal containedtRNAGlu we used a second antisense oligo directedagainst tRNAGlu(UUC) for blocking methylation Theoligo also covered the isoacceptor tRNAGlu(CUC) with
minor mismatches (Supplementary Figure S2) A furtherreduction of ex vivo methylation was observed Notablythe lower band in the tRNA size range disappeared (arrowin Figure 3B) suggesting that this represented a tRNAGlu
population that could be methylated at least in vitro Theresidual methylation after blocking with both oligos mayrepresent additional tRNAs or other unknown targets Aswe only concentrated on small RNAs larger methylatedmolecules and less abundant targets may have escapeddetection
To further analyse methylation on tRNAAsp andtRNAGlu (exemplified by tRNAGlu(UUC)) we carried outassays with wild-type sequences and C38A mutatedin vitro transcripts (Figure 4) We also included a suppres-sor tRNAGlu(CUA) that differed in only 2 nt fromtRNAGlu(UUC) (Figure 2) Assays were done in parallelwith the recombinant hDnmt2 and D discoideumDnmA The experiment showed that tRNAAsp(GUC) wasthe best substrate for both DnmA and hDnmt2 (Figure 4)Substantial methylation was also observed fortRNAGlu(UUC) and suppressor tRNAGlu(CUA) Moreoverthe complete loss of methylation in the C38A mutations oftRNAAsp(GUC) and tRNAGlu(UUC) confirmed that methy-lation occurred at position C38 and largely ruled out thatmethyl group incorporation was due to another
Figure 3 Ex vivo methylation and blocking assay (A) Methylation of in vitro transcribed tRNAAsp(GUC) was completely blocked when a comple-mentary antisense oligo was hybridized (B) Using enriched small RNA from dnmAKO cells ex vivo methylation in the tRNA size class wasdifferentially lost when antisense oligos to tRNAAsp(GUC) and tRNAGlu(UUC) were hybridized before the methylation reaction The oligo againsttRNAGlu(UUC) also covers tRNAGlu(CUC) with minor mismatches (see Supplementary Figure S2) Even with both oligos a significant amount of 3Hincorporation still remained The upper panel shows the ethidiumbromide stained gel to demonstrate equal loading the lower panel shows thefluorogram The arrow marks a band that was specifically lost when the anti tRNAGlu(UUC) oligo was used
Figure 2 Target site sequences of tRNAs used for in vitro methylationThe position of the putative signature nucleotides are indicated andprinted in bold if present The anticodon is underlined tRNAGlu(CUC)
(not shown) also harbours the nucleotide pattern Except for the anti-codon both tRNAGlu are identical in the anticodon stem-loop region(Supplementary Figure S2) Only a few differences appear in theacceptor stem
Figure 4 C38 is the target for in vitro methylation of tRNAAsp(GUC)
and tRNAGlu(UUC) In vitro methylation of tRNAAsp(GUC)tRNAGlu(UUC) and a suppressor tRNAGlu(CUA) C38A mutations con-firmed C38 as the target nucleotide tRNAAsp(GUC) methylation wasmost efficient
6 Nucleic Acids Research 2013
at Universitaumlt W
uumlrzburg on August 5 2013
httpnaroxfordjournalsorgD
ownloaded from
contaminating methyltransferase in the recombinantprotein preparation (17)
The methylation experiments using in vitro transcribedtRNAs showed that no other modifications were necessaryfor C38 methylation by DnmA and hDnmt2 (13) and thatthe processed ends of a mature tRNA were not requiredfor target recognition as the tRNA transcripts contain no30-CCA Even tRNAGlu embedded in a long dsRNAmolecule (1000 nt) with small stem-loop structures atthe ends (27) was a specific substrate of DnmA andhDnmt2 (Supplementary Figure S3)
By EMSA no difference in in vitro binding could beseen for the specific target tRNAGlu(UUC) and non-targetRNAs indicating that DnmA associated non-specificallywith any nucleic acid (data not shown) However specificcovalent enzyme-substrate intermediates can be detectedby denaturing SDSndashPAGE (25) Figure 5A andSupplementary Figure S4 show that radioactivelylabelled tRNA transcripts that can be methylatedin vitro by DnmA formed denaturation resistantcomplexes while non-target RNAs did not To confirmthat the denaturation resistant complexes were really dueto binding of the enzyme to the substrate SDS gels wereblotted and His-tagged DnmA was detected by a specificanti-His antibody A small amount of the tagged proteinwas shifted to the same position as the labelled tRNA(Supplementary Figure S5)
We then performed time courses for the reaction andfound that covalent complex formation was only transientunder the conditions used For tRNAAsp(GUC) adductsreached a maximum after 1min and were undetectableafter 15min whereas the tRNAGlu(UUC) complex onlybegan to accumulate after 1min but persisted for almost60min (Figure 5A and B) This coincided with the highmethylation activity on tRNAAsp(GUC-1) and a lower oneon tRNAGlu(UUC-2) The human enzyme appeared to beslightly faster in the reaction but otherwise showed asimilar behaviour compared with D discoideum DnmA
tRNAAsp(GUC) tRNAGlu(UUCCUC) and tRNAGly(GCC)
bind specifically to DnmA in vivo
The blocking assay (Figure 3) suggested that further sub-strates could be targeted by DnmA To identify suchtargets and to further investigate the interaction betweenDnmA and RNAs in vivo we established a UV-CLIP pro-cedure for D discoideumDnmA-GFP expressing cells andcontrol cells expressing GFP only were UV cross-linkedand complexes were precipitated under denaturing condi-tions by a GFP nanobody (21) Then co-precipitatedRNAs were isolated converted into strand-specificcDNA libraries and analyzed by Illumina sequencing(Figure 6 and Supplmentary Figure S6)
Various tRNA fragments were enriched in the DnmACLIP sample compared with the control By their lengthand position fragments appeared not to be random butspecific We found mid-fragments of 20ndash24 nt (position32ndash36 to 53ndash56) that covered the C38 methylation site inthe respective tRNAs 50-fragments of 17 nt (nucleotideposition 1ndash17) 30-fragments of 2021 nt (nucleotideposition 54 to the end) and 30-half tRNAs of 41ndash43 nt
(position 30ndash32 to the end) Mid-fragments oftRNAAsp(GUC) tRNAGlu(UUCCUC) and tRNAGly(GCC)
were significantly enriched in the DnmA-GFP fraction incomparison with the control CLIP assay tRNAAsp(GUC)tRNAGlu(UUC) and tRNAGlu(CUC) harbour the sequencepattern (C32 C34 A37 C40) which has been suggestedas a signature for C38 methylation (Figure 2 Helmpersonal communication) tRNAGly(GCC) contains thepattern except for C40 For tRNAAla(AGC) which doesnot contain C38 and the signature pattern a differentfragment from the 30-end was enriched in the DnmA-CLIP sample For other tRNAs no significant accumula-tion was observed in the experiment (Figure 6 andSupplementary Figure S6) For several tRNAs varyingamounts of full-length molecules were found but thesewere never enriched in the DnmA sample in comparisonwith the control
tRNAGly(GCC) is an additional methylation substratein vitro
The CLIP data suggested that tRNAGly(GCC) was anothertarget for DnmA We therefore cloned this and several othertRNA genes that contained the pattern only partially(Figure 2) We generated in vitro transcripts and examinedmethylation by DnmA and hDnmt2 in vitro IndeedtRNAGly(GCC) was methylated by both enzymes (Figure 7)while none of the other tRNAs (tRNALeu(UAG-1)tRNALeu(AAG-8) tRNAVal(CAC-1) and tRNAPhe(GAA-2))resulted in detectable methylation signals in the assay(Supplementary Figure S7) Interestingly the in vitro methy-lation efficiency of the three tRNA substrates
Figure 5 Formation and turnover of tRNA-methyltransferasecomplexes Turnover of covalent complexes of tRNA substrates withDnmA and hDnmt2 (A) Examples of time courses on covalentcomplex formation with tRNAAsp(GUC) and tRNAGlu(UUC) (B)Complex bands in A were quantified and presented on a time scale
Nucleic Acids Research 2013 7
at Universitaumlt W
uumlrzburg on August 5 2013
httpnaroxfordjournalsorgD
ownloaded from
(tRNAAsp(GUC) tRNAGlu(UUC) tRNAGly(GCC)) correlatedroughly with the number of tRNA fragments found in theRNA-CLIP experimentThe data supported the conclusion from the ex vivo
blocking assay (Figure 3B) that in addition toD discoideum tRNAAsp and tRNAGlu other RNAs weresubstrates for DnmA and hDnmt2 in vitro
Expression of dnmA is regulated during developmentcell cycle and on temperature stress
DnmA expression in growing vegetative D discoideumcells is low To examine tRNA methylation in vivo it
was therefore advantageous to determine conditionsunder which dnmA was most highly expressed Promptedby previous studies (141517) we assumed that the biolo-gical function of DnmA was connected to developmentalandor environmental conditions We therefore examinedexpression levels of dnmA under different regimes dnmAmRNA was quantified by qPCR over the 24 h develop-mental cycle of D discoideum (data not shown) and con-firmed the data deposited in Dictyexpress (httpdictyexpressbiolabsi)
Interestingly an 5-fold increase in expression wasobserved at 16 h in development This time coincides
Figure 6 tRNA and tRNA fragments associated with DnmA-GFP After UV crosslinking RNA bound to DnmA was co-immunoprecipitatedunder denaturing conditions (CLIP) The number of normalised reads for tRNAs detected by Illumina sequencing is shown for DnmA CLIP and forthe control CLIP with GFP In addition to full-length tRNAs four classes of tRNA fragments were found and these are indicated by differentshading of the bars Size and localization of fragments is shown schematically on the simplified clover leaf structure tRNAs that are not significantlyenriched with DnmA are shown in Supplementary Figure S6 In case of multiple gene copys resulting in the same RNA transcript the sequencingreads were mapped to the first copy in the genome starting from chromsome 1 Only this copy is listed Sequences and detailed information on genecopies and potential isoacceptors are listed in Supplementary Table S2
8 Nucleic Acids Research 2013
at Universitaumlt W
uumlrzburg on August 5 2013
httpnaroxfordjournalsorgD
ownloaded from
with a presumed synchronous mitotic division (28) wherea multitude of cell-cycle regulated genes are expressed[(29) and McWilliams personal Communication]
Therefore we synchronized vegetative D discoideumcells to examine dnmA expression during the cell cycleFor these experiments the D discoideum wild-typestrain NC4 the parent of the axenic laboratory strainAX2 was used (30) This was prompted by the observationthat D melanogaster and E histolytica laboratory strainsmay lose dnmt2 expression (Reuter Ankri personalcommunication)
The data confirmed our assumption of a cell-cycle-de-pendent transcriptional regulation of dnmA (Figure 8A)The peak of expression coincided with late mitosis or earlyS-phase as D discoideum does not display a G1 phaseUsing AX2 cells cotransformed with PCNA-RFP andDnmA-GFP we observed GFP accumulation in thenucleus during S phase (indicated by PCNA expression)and a loss of DnmA from the nucleus during mitosis(Figure 8B) This is further exemplified by the movie inthe Supplementary Material (Supplementary Movie S1)
In other organisms dnmt2 was observed to beupregulated during stress response (315) We thereforeexamined expression after anoxia heat shock and coldshock but did not find any significant changes (data notshown) Only during recovery from cold shock wedetected a strong transient increase in dnmA mRNA by47-fold (Figure 8C)
Thus dnmA expression was differentially regulatedduring the cell cycle development and on temperaturestress Furthermore DnmA localization changed specific-ally during the cell cycle
Though the basic expression in vegetative cells was in-distinguishable dnmA expression in NC4 was 30-foldhigher at 16 h of development whereas developmentalupregulation was only 5-fold in AX2 The data aresummarized in Figure 8D
NC4 the parent strain of AX2 displays increasedtRNA
Asp methylation during development
Based on these observations we asked the questionwhether expression levels correlated with tRNA
methylation activity in vivo Bisulfite sequencing wasdone with primers for tRNAAsp tRNAGlu and tRNAGly
on RNA isolated from vegetative AX2 cells AX2 cellsdeveloped for 16 h AX2 cells subjected to cold shockAX2 recovered for 25 h after cold shock vegetativeNC4 cells and NC4 cells developed for 16 h The resultson C38 methylation of tRNAAsp(GUC) are summarized inFigure 8E The methylation status of all C residues fromall tested tRNAs in this study is shown in SupplementaryFigure S8 Indeed tRNAAsp(GUC) methylation at C38increased to 76 in development in NC4 cells Noincrease was observed in AX2 cells during developmentpossibly due to insufficient sensitivity of the assayInterestingly no increase in tRNAAsp(GUC) methylationwas observed at the highest dnmA expression levelsduring stress recovery In Figure 8E the 30ndash35 methy-lation level in all tRNAAsp(GUC) samples represents signifi-cant methylation as compared with other C residues (seeSupplementary Figure S8)We did however not detect any significant methylation
of tRNAGlu(UUCCUC) and tRNAGly(GCC) under any con-ditions (Supplementary Figure S8) In contrast C49C50methylation which does not depend on Dnmt2 enzymeswas found in all samples and thus served as an internalcontrol In all 1175 bisulfite-sequence reads wereexamined for tRNAGlu(UUCCUC) and 4040 fortRNAGly(GCC) No methylation above background wasfound Though it cannot be excluded that the failure todetect C38 methylation of tRNAGlu and tRNAGly was dueto inefficient reverse transcription or low levels of methy-lation (see lsquoDiscussionrsquo section) we conclude thattRNAAsp is the major or possibly the only in vivo targetfor methylation by DnmA in D discoideum
DISCUSSION
The biological function of Dnmt2 methyltransferases islargely unknown and its role in DNA methylation is con-troversially discussed (910) In contrast tRNA methyla-tion by Dnmt2 is robust in vitro and in vivo andevolutionary conserved In addition to the originallydescribed tRNAAsp(GUC) (1) two further tRNAs targets(tRNAVal(CAC) and tRNAGly(GCC)) have been defined inD melanogaster (15) and tRNAGlu(UUC) in S pombe (17)To get insight into potential functions of the
D discoideum Dnmt2 homologue DnmA we examinedexpression levels in development in the cell cycle andunder different environmental conditions We confirmedby qPCR an expression peak at 16 h of developmentWeijer and co-workers (28) had previously proposed asynchronous cell division at this developmental stageWe therefore investigated dnmA expression in the cellcycle and found a strong transient increase of mRNA inS-phase or shortly thereafter The expression peak wasaccompanied by reaccumulation of DnmA in thenucleus We assume that DnmA is degraded duringmitosis and re-synthesised after cell divisionIn S pombe as well as in D melanogaster tRNA methy-
lation changes in response to environmental challengeslike temperature stress anoxia or nutrients We tested
Figure 7 In vitro methylation of tRNAGly(GCC) by recombinant DnmAand hDnmt2 In vitro transcribed tRNAs were methylated in vitro asdescribed before The top panel shows the ethidium-bromide stainedgel the bottom panel the fluorogram of 3H-labelled methylated tRNAsAs a control tRNAAsp(GUC) methylation is shown
Nucleic Acids Research 2013 9
at Universitaumlt W
uumlrzburg on August 5 2013
httpnaroxfordjournalsorgD
ownloaded from
Figure 8 Expression localization and tRNAAsp(GUC) methylation activity of DnmA under various conditions (A) dnmA expression is regulatedduring the cell cycle Cells were arrested in the cell cycle by cold treatment and then released by transfer to 22C Cells were counted every 30min(grey dots and grey line) and samples were taken for qPCR (black bars) After cell division dnmA expression increased 5-fold and then rapidlydeclined to basal levels Normalization was done on vegetative growing cells dnmA expression is only shown from 3 to 45 h during recovery(B) DnmA is lost from the nucleus during mitosis (arrow) The three cells on the right are in S-phase as indicated by the RFP-PCNA marker andaccumulate DnmA in the nucleus (for further details see movie in Supplementary Material) (C) Relative quantification of dnmA expression levels inAX2 cells after 25 h recovery from cold shock at 4C Expression of dnmA increased gt40-fold and returned within 30min to basal levels (n=3)(D) At 16 h of development dnmA expression increased 46-fold in the NC4 strain In AX2 cells expression increased only 5-fold (E) At 16 h ofdevelopment in vivo methylation of C38 in tRNAAsp(GUC) increased up to 75 in the D discoideum strain NC4 whereas no significant increase wasobserved in AX2 cells
10 Nucleic Acids Research 2013
at Universitaumlt W
uumlrzburg on August 5 2013
httpnaroxfordjournalsorgD
ownloaded from
heat shock cold shock anoxia and various nutrient mediabut did not observe altered levels Only during therecovery phase after cold treatment a transient almost50-fold increase in mRNA abundance was seen
Reuter and Ankri (personal communication) hadobserved a loss of Dnmt2 expression in laboratorystrains of D melanogaster and E histolytica We thereforeexamined D discoideum NC4 the parent of the axeniclaboratory strain AX2 The developmental regulation inAX2 and NC4 was qualitatively similar but expressionlevels were much higher in NC4 The observationsupports the assumption that Dnmt2 expression wasreduced under optimal growth conditions in the labora-tory strain and may actually be selected against
We show that the D discoideum Dnmt2 homologueDnmA is an active tRNAAsp(GUC) methyltransferasein vitro and in vivo To identify further potential targetsfor DnmA we established a UV-CLIP method to analyseDnmAndashRNA interaction in vivo In addition to the knowntarget tRNAAsp(GUC) fragments of tRNAGlu(UUC) andtRNAGly(GCC) were most abundant in comparison withthe control Unexpectedly complete tRNAs were notenriched in the DnmA CLIP fraction but rather specificlsquomid-fragmentsrsquo that covered the putative methylation siteand the recognition pattern proposed by Helm (personal
communication) A minor exception was tRNAGly(GCC)which lacks C40 We do not know whether these frag-ments were directly or indirectly generated by DnmAin vivo or whether they were protected by DnmA fromdegradation during the experimental procedure As noco-crystal of Dnmt2 and a tRNA substrate is availablethe exact interface for the interaction is unknownJurkowski et al (31) mapped the binding site on Dnmt2by mutational analysis A manual placement of a tRNAinto the crystal structure suggests that Dnmt2 mainlycontacts the anticodon stem loop Assuming that mid-frag-ments are footprints of DnmA our data support this notionbut suggest that the interaction surface extends into theD-loop which is partially covered by the mid-fragments
We examined the putative DnmA targets in vitro (bymethylation of in vitro transcribed tRNAs) ex vivo(by methylation of RNA isolated from D discoideum)and in vivo (by bisulfite sequencing) Furthermore weinvestigated by SDSndashPAGE the kinetics of covalentcomplex formation between tRNAAsp and tRNAGlu withDnmA and hDnmt2
All three tRNAs (tRNAAsp tRNAGlu tRNAGly) werereadily methylated by recombinant DnmA and hDnmt2in vitro although the activity was much higher (estimatedat least 10-fold) on tRNAAsp compared with tRNAGlu andtRNAGly For tRNAAsp and tRNAGlu this correlated wellwith the formation and decay of covalent RNAmdashproteincomplexes where kinetics for tRNAAsp were much fasterthan for tRNAGlu Instead of C methylation the covalentintermediate may also proceed to a hydrogen exchangereaction or to a deamination of the cytosine In bothcases an intermediate will be detected but it will notresult in a methylated product (32) It may be thatslower off rates represent these alternative pathways thatwould result in unmethylated tRNAs
Using antisense oligos we showed that tRNAAsptRNAGlu and at least one additional RNA could bemethylated ex vivo A quantification of the results wasnot possible the apparently low methylation oftRNAGlu ex vivo could be due to the low activity of theenzyme on this substrate (as suggested by the in vitroexperiments see Figure 5B)The data for the examination of in vivo methylation by
bisulfite sequencing were rather unexpected AlthoughtRNAAsp(GUC) was consistently found to be methylatedat C38 we never detected any methylation at C38 oftRNAGlu(UUCCUC) or tRNAGly(GCC) There could betwo reasons to explain the failure to detect C38 methyla-tion for these in vitro substrates (i) due to the low activityof DnmA on tRNAGlu(UUCCUC) and tRNAGly(GCC) asshown in vitro there was low or no significant methylationin vivo and (ii) methylation escaped detection becauseother unknown modifications blocked reverse transcrip-tion of the methylated molecules However if this wasthe case one has to assume that C49C50 methylationoccurs independently of these unknown modificationswhereas C38 methylation strictly depends on them AsC38 methylation does not require any other modificationsin vitro we believe that this scenario is less likely If in vitromethylation activity was comparable with in vivo methy-lation 10 methylation of tRNAGlu(UUCCUC) andtRNAGly(GCC) could be expected relative totRNAAsp(GUC) As tRNAAsp was methylated to 30 a3 methylation of tRNAGlu and tRNAGly could be belowthe detection level of the bisulfite assayWe find that high expression of dnmA did not necessar-
ily correlate with increased methylation of tRNAs Duringrecovery from cold shock D discoideum cells displayedthe highest level of dnmA expression but this had noeffect on tRNAAsp methylation Only in developmentand with the high expressing NC4 strain a significantincrease of tRNAAsp methylation to 70 was observedIt has to be considered however that bisulfite sequencingdoes not necessarily allow for quantitative evaluation ofdata detected C-methylation depends on reverse tran-scription efficiency which may be hampered by othermodifications that could be different under differentconditions Furthermore the completeness of bisulfite-mediated conversion could also depend on RNA struc-ture which may be altered by other modificationsHowever incomplete conversion will lead to an overesti-mation of C-methylation as maybe the case for forexample C30 C31 C35 in tRNAGly in some samplesTaken together the data from CLIP complex forma-
tion and methylation show that all three substrates can bebound and methylated in vitro The CLIP data show thatthere is a specific interaction of DnmA with tRNA mid-fragments of tRNAAsp(GUC) tRNAGlu(UUCCUC) andtRNAGly(GCC) but not with other tRNAs In contrastbisulfite sequencing could only detect methylation ontRNAAsp(GUC) even at the highest expression levels ofdnmA Even though low amounts of methylation mayhave escaped detection in bisulfite sequencing there isstill a discrepancy between dnmA expression levels (egduring cold shock recovery) DnmA binding to substrates(as determined by CLIP) and methylation in vivo
Nucleic Acids Research 2013 11
at Universitaumlt W
uumlrzburg on August 5 2013
httpnaroxfordjournalsorgD
ownloaded from
We therefore suggest that DnmA binding to specifictargets could elicit functions in addition to RNA methy-lation eg by promoting processing or degradation andor recruitment of other proteins At least for some rRNAmethyltransferases such as Bud23 or Nep1 fromSaccharomyces cerevisiae it is known that they executefunctions independent from the methylation activity(33ndash35) As shown by Schaefer and co-workers (15)methylation protects tRNAs from degradation or process-ing after stress By a different pathway binding of DnmAmay initiate processing that leads to specific tRNA frag-ments that could serve for other biological functions (36)Interestingly tRNAGlu(UUC) is also a substrate for theDnmt2 homologue Pmt1 in S pombe (17) HowevertRNAGlu(UUC) is found to be methylated to a low butsignificant extend in vivo in fission yeast when Pmt1 isoverexpressedThe detection of tRNAAla(AGC) fragments in the CLIP
assay brings up a new question tRNAAla(AGC) has a Tinstead of a C in position 38 and does not contain thesignature nucleotides in the anticodon stem-loopInstead of a mid-fragment a distinct 30 fragment wasassociated with DnmA suggesting that a recognitionmotif was located in this part of the moleculeAs methylation also occurred in a large artificial
molecule with embedded tRNAGlu (SupplementaryFigure S2) it could be that other longer RNAs containingan appropriate recognition motif may also be bound bythe enzyme We thus propose that the biological functionsof Dnmt2 proteins could extend far beyond our presentknowledge Investigation of further targets and of the sig-nificance of RNA fragments associated with Dnmt2should shed light on the importance of this enzyme
SUPPLEMENTARY DATA
Supplementary Data are available at NAR Online
ACKNOWLEDGEMENTS
The authors thank Thomas Winckler (Friedrich-Schiller-University Jena) for providing the tRNAGlu(UUC) and sup-pressor tRNAGlu(CUA) plasmids Susanne Junk for cloningtRNAAspC38A Matthias Schafer (DKFZ) and RobinLorenz (University of Kassel) are acknowledged forRNA bisulfite sequencing and Richard Reinhardt (MaxPlanck Genome Centre Cologne) for deep sequencingMark Helm (Johannes-Gutenberg-University Mainz) gen-erously provided data before publication CarstenSeehafer (University of Kassel) helped with the analysisof deep-sequencing data
FUNDING
Deutsche Forschungsgemeinschaft [FOR 1082 to WNand AJ] and the Studienstiftung des Deutschen Volkes(to SM) Funding for open access charge FOR1082
Conflict of interest statement None declared
REFERENCES
1 GollMG KirpekarF MaggertKA YoderJA HsiehCLZhangX GolicKG JacobsenSE and BestorTH (2006)Methylation of tRNAAsp by the DNA methyltransferasehomolog Dnmt2 Science 311 395ndash398
2 PhalkeS NickelO WalluscheckD HortigF OnoratiMCand ReuterG (2009) Retrotransposon silencing and telomereintegrity in somatic cells of Drosophila depends on the cytosine-5methyltransferase DNMT2 Nat Genet 41 696ndash702
3 WilkinsonCR BartlettR NurseP and BirdAP (1995) Thefission yeast gene pmt1+ encodes a DNA methyltransferasehomologue Nucleic Acids Res 23 203ndash210
4 KatohM CurkT XuQ ZupanB KuspaA and ShaulskyG(2006) Developmentally regulated DNA methylation inDictyostelium discoideum Eukaryot Cell 5 18ndash25
5 KuhlmannM BorisovaBE KallerM LarssonP StachDNaJ EichingerL LykoF AmbrosV SoderbomF et al(2005) Silencing of retrotransposons in Dictyostelium by DNAmethylation and RNAi Nucleic Acids Res 33 6405ndash6417
6 RaiK ChidesterS ZavalaCV ManosEJ JamesSRKarpfAR JonesDA and CairnsBR (2007) Dnmt2 functionsin the cytoplasm to promote liver brain and retina developmentin zebrafish Genes Dev 21 261ndash266
7 FisherO Siman-TovR and AnkriS (2006) Pleiotropicphenotype in Entamoeba histolytica overexpressing DNAmethyltransferase (Ehmeth) Mol Biochem Parasitol 147 48ndash54
8 DurdevicZ HannaK GoldB PollexT CherryS LykoFand SchaeferM (2013) Efficient RNA virus control in Drosophilarequires the RNA methyltransferase Dnmt2 EMBO Rep 14269ndash275
9 SchaeferM and LykoF (2010) Lack of evidence for DNAmethylation of Invader4 retroelements in Drosophila andimplications for Dnmt2-mediated epigenetic regulation NatGenet 42 920ndash921 author reply 921
10 KraussV and ReuterG (2011) DNA methylation in Drosophila -a critical evaluation Prog Mol Biol Transl Sci 101 177ndash191
11 RaddatzG GuzzardoPM OlovaN FantappieMRRamppM SchaeferM ReikW HannonGJ and LykoF(2013) Dnmt2-dependent methylomes lack defined DNAmethylation patterns Proc Natl Acad Sci USA 110 8627ndash8631
12 JeltschA NellenW and LykoF (2006) Two substrates arebetter than one dual specificities for Dnmt2 methyltransferasesTrends Biochem Sci 31 306ndash308
13 JurkowskiTP MeusburgerM PhalkeS HelmM NellenWReuterG and JeltschA (2008) Human DNMT2 methylatestRNA(Asp) molecules using a DNA methyltransferase-likecatalytic mechanism RNA 14 1663ndash1670
14 TovyA Siman TovR GaentzschR HelmM and AnkriS(2010) A new nuclear function of the Entamoeba histolyticaglycolytic enzyme enolase the metabolic regulation of cytosine-5methyltransferase 2 (Dnmt2) activity PLoS Pathog 6 e1000775
15 SchaeferM PollexT HannaK TuortoF MeusburgerMHelmM and LykoF (2010) RNA methylation by Dnmt2protects transfer RNAs against stress-induced cleavage GenesDev 24 1590ndash1595
16 TuortoF LiebersR MuschT SchaeferM HofmannSKellnerS FryeM HelmM StoecklinG and LykoF (2012)RNA cytosine methylation by Dnmt2 and NSun2 promotestRNA stability and protein synthesis Nat Struct Mol Biol 19900ndash905
17 BeckerM MullerS NellenW JurkowskiTP JeltschA andEhrenhofer-MurrayAE (2012) Pmt1 a Dnmt2 homolog inSchizosaccharomyces pombe mediates tRNA methylation inresponse to nutrient signaling Nucleic Acids Res 4011648ndash11658
18 TovyA HertzR Siman-TovR SyanS FaustD GuillenNand AnkriS (2011) Glucose starvation boosts Entamoebahistolytica virulence PLoS Negl Trop Dis 5 e1247
19 MotorinY and HelmM (2011) RNA nucleotide methylationWiley Interdiscip Rev RNA 2 611ndash631
20 NiranjanakumariS LasdaE BrazasR and Garcia-BlancoMA(2002) Reversible cross-linking combined with
12 Nucleic Acids Research 2013
at Universitaumlt W
uumlrzburg on August 5 2013
httpnaroxfordjournalsorgD
ownloaded from
immunoprecipitation to study RNA-protein interactions in vivoMethods 26 182ndash190
21 RothbauerU ZolghadrK MuyldermansS SchepersACardosoMC and LeonhardtH (2008) A versatile nanotrapfor biochemical and functional studies with fluorescent fusionproteins Mol Cell Proteomics 7 282ndash289
22 SittkaA LucchiniS PapenfortK SharmaCM RolleKBinnewiesTT HintonJC and VogelJ (2008) Deep sequencinganalysis of small noncoding RNA and mRNA targets of theglobal post-transcriptional regulator Hfq PLoS Genet 4e1000163
23 HoffmannS OttoC KurtzS SharmaCM KhaitovichPVogelJ StadlerPF and HackermullerJ (2009) Fast mapping ofshort sequences with mismatches insertions and deletions usingindex structures PLoS Comput Biol 5 e1000502
24 ConnollyMA ClausenPA and LazarJG (2006) Purificationof RNA from animal cells using trizol Cold Spring HarborProtoc 2006 pii pdbprot4104
25 DongA YoderJA ZhangX ZhouL BestorTH andChengX (2001) Structure of human DNMT2 an enigmaticDNA methyltransferase homolog that displays denaturant-resistant binding to DNA Nucleic Acids Res 29 439ndash448
26 SchaeferM PollexT HannaK and LykoF (2009) RNAcytosine methylation analysis by bisulfite sequencing NucleicAcids Res 37 e12
27 BoninM OberstrassJ VogtU WasseneggerM and NellenW(2001) Binding of IRE-BP to its cognate RNA sequence SFMstudies on a universal RNA backbone for the analysis of RNA-protein interaction Biol Chem 382 1157ndash1162
28 ZimmermanW and WeijerCJ (1993) Analysis of cell cycleprogression during the development of Dictyostelium and itsrelationship to differentiation Dev Biol 160 178ndash185
29 StrasserK BloomfieldG MacWilliamsA CeccarelliAMacWilliamsH and TsangA (2012) A retinoblastomaorthologue is a major regulator of S-phase mitotic anddevelopmental gene expression in Dictyostelium PLoS One 7e39914
30 WattsDJ and AshworthJM (1970) Growth of myxameobae ofthe cellular slime mould Dictyostelium discoideum in axenicculture Biochem J 119 171ndash174
31 JurkowskiTP ShanmugamR HelmM and JeltschA (2012)Mapping the tRNA binding site on the surface of humanDNMT2 methyltransferase Biochemistry 51 4438ndash4444
32 SvedruzicZM and ReichNO (2005) DNA cytosine C5methyltransferase Dnmt1 catalysis-dependent release of allostericinhibition Biochemistry 44 9472ndash9485
33 ThomasSR KellerCA SzykA CannonJR and Laronde-LeblancNA (2011) Structural insight into the functionalmechanism of Nep1Emg1 N1-specific pseudouridinemethyltransferase in ribosome biogenesis Nucleic Acids Res 392445ndash2457
34 LeulliotN BohnsackMT GrailleM TollerveyD andVan TilbeurghH (2008) The yeast ribosome synthesisfactor Emg1 is a novel member of the superfamily of alphabeta knot fold methyltransferases Nucleic Acids Res 36629ndash639
35 WhiteJ LiZ SardanaR BujnickiJM MarcotteEM andJohnsonAW (2008) Bud23 methylates G1575 of 18S rRNA andis required for efficient nuclear export of pre-40S subunits MolCell Biol 28 3151ndash3161
36 SobalaA and HutvagnerG (2013) Small RNAs derived fromthe 50-end of tRNA can inhibit protein translation in humancells RNA Biol 10 525ndash535
Nucleic Acids Research 2013 13
at Universitaumlt W
uumlrzburg on August 5 2013
httpnaroxfordjournalsorgD
ownloaded from

centrifugation (10000 RCF 4C 30min) the supernatantwas collected in a fresh tube and precipitated with threevolumes of 100 EtOH
SDS-EMSA
Detection of denaturant-resistant DnmA-tRNA wascarried out by SDS-electrophoretic mobility shift assayas described elsewhere (25) with some modificationsBriefly DnmA (1ndash9 mg) was incubated in 20 ml of 10mMKPi (pH 70) 225 glycerol 25mM KCl 10mM DTT2mM MgCl2 and 100ndash500 mM S-adenosyl-methionine(SAM) with the respective 32P labelled tRNA (100ndash665 ng) at 22C Reactions were terminated at theindicated times by addition of 10 ml of SDS-stop mixture[150mM TrisndashHCl (pH 80) 6 SDS (wv) 30mMEDTA 0025 Bromophenol blue] and heated for10min at 65C Samples were cooled down to room tem-perature and subjected to electrophoresis on 6 or 12SDSndashpolyacrylamide gels sealed in Saran Wrap andexposed to an imager plate for 12ndash40 h at RT Plateswere read in a phosphoimager (Fujifilm FLA-7000) Insome cases samples were transferred from the gel to anitrocellulose membrane for immunodetection of DnmAusing an anti-His antibody and a secondary mouseantibody coupled to alkaline phosphatase
In vitro methylation assay
Methylation of RNA substrates was carried out asdescribed previously (13) 3 mM of recombinant hDnmt2and 3ndash12 mM of DnmA were incubated with 500 ng oftRNA-transcript (or 10 mg of small enriched RNA) 1methylation buffer [50mM TrisHCl (pH 70) 50mMNaCl 5mM MgCl2 1mM DTT] and 17mM [3H]-SAM(Hartmann) 1 uml RiboLockTM RNase Inhibitor(Fermentas) at 22C for 1min to 15 h as indicated inthe text The reaction was stopped by adding 1mM non-radioactive SAM and 2mgml proteinase K for 30minLoading buffer [formamide 5mM EDTA (pH 74)bromophenol blue] was added and samples were run ona 12 denaturing (7M Urea) polyacrylamide gel (1mm)for 2ndash5 h at 400V The gel was stained withethidiumbromide fixed (10 methanol 10 aceticacid) and immersed for 1 h in amplify solution(Amersham) dried and exposed to X-ray film (ContatypCX-BL+ Medical X-ray film) for 16 h up to 2 weeks at80C
In vitro blocked methylation assay
The in vitro blocked methylation assay is based on theassumption that RNA methylation is inhibited if thetarget site is covered by hybridization with a complemen-tary DNA oligo RNA (500 ng of transcript or 10 mg ofsmall enriched RNA) was mixed with 2 mM of the respect-ive oligonucleotide in 1 STE-buffer [10mM TrisHCl(pH 8) 50mM NaCl 1mM EDTA] heated up to 80Cin a thermocycler and then cooled down stepwise to roomtemperature The blocked RNA was subsequently used ina 3H-methylation assay as described earlier in the textBlocking oligos are shown in Supplementary Table S1(oligo 2 for tRNAAsp and tRNAGlu respectively)
dnmA expression analysis by real-time quantitative PCR
For analysing dnmA expression levels RNA was extractedby the Trizol-method (24) qPCR was performed in anEppendorf realplex light cycler with the SensiFAST SYBRNo-ROX One-Step Kit (Bioline) according to the sup-pliers instructions Relative quantification was done withthe Ct-method using gapdh (DDB_G0275153) as anormalization control (housekeeping gene)
Microscopy
For fluorescence analysis cells were fixed in 4 (wv)paraformaldehyde dissolved in 20mM phosphate buffer(pH 67) for 10min at 22C and stained with DAPI
Images were acquired on a Leica DMIRB invertedmicroscope equipped with a DC350 camera and IM50Acquisition software (Leica Microsystems WetzlarGermany)
For the movie DnmA-GFP cells (in DnmAKO back-ground) were analysed with a Zeiss Cell Observer SDwith Rapp UGA-40-2 L Scanner Images of living cellswere taken in 15 s intervals at 7 layers with a 08mMdistance between each layer during a period of 120minFurther image analysis was done with ImageJ 146 Themovie is displayed with eight frames per second
RNA bisulfite sequencing
RNA bisulfite sequencing was carried out as describedelsewhere (26) Primer sequences are listed in theSupplementary Material (Supplementary Table S1)
RESULTS
DnmA is an active tRNAAsp methyltransferase in vitroand in vivo
Previous experiments have shown that hDnmt2 canmethylate a D discoideum RNA in the size range oftRNAs in vitro (13) Here we investigated whether theD discoideum Dnmt2-homologue DnmA is also anactive tRNAAsp methyltransferase In vitro transcribedD discoideum tRNAAsp(GUC) was used as a substrate formethylation by DnmA and hDnmt2 in vitro The assaysshowed that the tRNA was efficiently methylated by bothenzymes (Figure 1A) The reaction was time dependentthe activity was sensitive to Mg2+ concentrations andalmost lost at 10mM Mg2+ In contrast to the humanenzyme DnmA was inactive at 37C (data not shown)Therefore all reactions were carried out at 22C wherehDnmt2 still showed good activity
To investigate methylation in vivo RNA bisulfiteanalysis of tRNAAsp(GUC) from D discoideum wasperformed Figure 1B shows that in wild-type cellstRNAAsp(GUC) was methylated at position C38 to30 whereas methylation was at background levels inDnmA knockout strains Ectopic overexpression ofhDnmt2 in wild-type cells increased tRNAAsp(GUC) C38methylation to 50 indicating cross-species activity ofDnmt2-homologues C49 was methylated at 90 inde-pendently of DnmA and served as an internal control forthe bisulfite reaction
4 Nucleic Acids Research 2013
at Universitaumlt W
uumlrzburg on August 5 2013
httpnaroxfordjournalsorgD
ownloaded from
As tRNAAsp appeared to be not the only target forDnmt2 in other organisms (eg D melanogaster) weexamined if more cellular-derived RNAs could bemethylated in vitro (ex vivo methylation experiments)Total RNA from dnmAKO AX2 cells was subjected toin vitro methylation by recombinant DnmA for varioustimes Methylated RNAs were separated by gelelectro-phoresis and visualized by autoradiography Figure 1Cshows that a substantial RNA fraction in the tRNA sizerange could be methylated At longer incubation times asecond minor methylated RNA species appeared
indicating that at least one further substrate could bemethylated in vitro
Identification of further substratesmdashin vitro binding andmethylation of tRNAGlu
The group of Helm (personal communication) hadproposed a sequence pattern (C32 C34 A37 C40) thatmay serve as a target recognition motif for C38 methyla-tion by hDnmt2 We therefore examined all D discoideumtRNAs in silico to identify candidates for the additionalmethylated band in the ex vivo experiment In addition to
Figure 1 In vitro methylation of tRNAAsp(GUC) (A) In vitro methylation of tRNAAsp(GUC) by DnmA at 2mM 5mM and 10mM MgCl2 and byhDnmt2 at 5mM Mg2+ The upper panel shows ethidium bromide staining of the in vitro transcripts separated in a denaturing polyacrylamide gelthe lower panel shows incorporated 3H-Me in the tRNAs Reactions were run for the times indicated (B) In vivo methylation of cytosines intRNAAsp(GUC) from different D discoideum strains Results of the RNA bisulfite sequencing (454 pyrosequencing) are given in percentage of readsNumbers of sequence reads are shown in brackets C49 is methylated by a different methyltransferase and thus serves as an internal standard All 22tRNAAsp(GUC) genes result in the same transcript and no isoacceptors are encoded in the D discoideum genome (C) In vitro methylation of smallenriched RNA of a dnmAKO strain (ex vivo methylation) The methylation reaction was done for the times indicated
Nucleic Acids Research 2013 5
at Universitaumlt W
uumlrzburg on August 5 2013
httpnaroxfordjournalsorgD
ownloaded from
tRNAAsp(GUC) we found tRNAGlu(UUC) andtRNAGlu(CUC) to match the pattern (Figure 2 andSupplementary Table S2)To test for tRNAGlu as a potential substrate we
modified the methylation assay by addition of a 50 ntDNA antisense oligonucleotide to tRNAAsp(GUC)
covering the methylation target site Figure 3A shows asa control that methylation of in vitro transcribedtRNAAsp(GUC) was completely blocked We then per-formed ex vivo methylation assays with enriched smallRNAs isolated from D discoideum after blockingtRNAAsp(GUC) with the antisense oligo Figure 3B showsthat the majority of 3H-incorporation disappearedHowever with both DnmA and hDnmt2 a significantamount of methylation was still detectable in the sizerange of tRNAs Assuming that this signal containedtRNAGlu we used a second antisense oligo directedagainst tRNAGlu(UUC) for blocking methylation Theoligo also covered the isoacceptor tRNAGlu(CUC) with
minor mismatches (Supplementary Figure S2) A furtherreduction of ex vivo methylation was observed Notablythe lower band in the tRNA size range disappeared (arrowin Figure 3B) suggesting that this represented a tRNAGlu
population that could be methylated at least in vitro Theresidual methylation after blocking with both oligos mayrepresent additional tRNAs or other unknown targets Aswe only concentrated on small RNAs larger methylatedmolecules and less abundant targets may have escapeddetection
To further analyse methylation on tRNAAsp andtRNAGlu (exemplified by tRNAGlu(UUC)) we carried outassays with wild-type sequences and C38A mutatedin vitro transcripts (Figure 4) We also included a suppres-sor tRNAGlu(CUA) that differed in only 2 nt fromtRNAGlu(UUC) (Figure 2) Assays were done in parallelwith the recombinant hDnmt2 and D discoideumDnmA The experiment showed that tRNAAsp(GUC) wasthe best substrate for both DnmA and hDnmt2 (Figure 4)Substantial methylation was also observed fortRNAGlu(UUC) and suppressor tRNAGlu(CUA) Moreoverthe complete loss of methylation in the C38A mutations oftRNAAsp(GUC) and tRNAGlu(UUC) confirmed that methy-lation occurred at position C38 and largely ruled out thatmethyl group incorporation was due to another
Figure 3 Ex vivo methylation and blocking assay (A) Methylation of in vitro transcribed tRNAAsp(GUC) was completely blocked when a comple-mentary antisense oligo was hybridized (B) Using enriched small RNA from dnmAKO cells ex vivo methylation in the tRNA size class wasdifferentially lost when antisense oligos to tRNAAsp(GUC) and tRNAGlu(UUC) were hybridized before the methylation reaction The oligo againsttRNAGlu(UUC) also covers tRNAGlu(CUC) with minor mismatches (see Supplementary Figure S2) Even with both oligos a significant amount of 3Hincorporation still remained The upper panel shows the ethidiumbromide stained gel to demonstrate equal loading the lower panel shows thefluorogram The arrow marks a band that was specifically lost when the anti tRNAGlu(UUC) oligo was used
Figure 2 Target site sequences of tRNAs used for in vitro methylationThe position of the putative signature nucleotides are indicated andprinted in bold if present The anticodon is underlined tRNAGlu(CUC)
(not shown) also harbours the nucleotide pattern Except for the anti-codon both tRNAGlu are identical in the anticodon stem-loop region(Supplementary Figure S2) Only a few differences appear in theacceptor stem
Figure 4 C38 is the target for in vitro methylation of tRNAAsp(GUC)
and tRNAGlu(UUC) In vitro methylation of tRNAAsp(GUC)tRNAGlu(UUC) and a suppressor tRNAGlu(CUA) C38A mutations con-firmed C38 as the target nucleotide tRNAAsp(GUC) methylation wasmost efficient
6 Nucleic Acids Research 2013
at Universitaumlt W
uumlrzburg on August 5 2013
httpnaroxfordjournalsorgD
ownloaded from
contaminating methyltransferase in the recombinantprotein preparation (17)
The methylation experiments using in vitro transcribedtRNAs showed that no other modifications were necessaryfor C38 methylation by DnmA and hDnmt2 (13) and thatthe processed ends of a mature tRNA were not requiredfor target recognition as the tRNA transcripts contain no30-CCA Even tRNAGlu embedded in a long dsRNAmolecule (1000 nt) with small stem-loop structures atthe ends (27) was a specific substrate of DnmA andhDnmt2 (Supplementary Figure S3)
By EMSA no difference in in vitro binding could beseen for the specific target tRNAGlu(UUC) and non-targetRNAs indicating that DnmA associated non-specificallywith any nucleic acid (data not shown) However specificcovalent enzyme-substrate intermediates can be detectedby denaturing SDSndashPAGE (25) Figure 5A andSupplementary Figure S4 show that radioactivelylabelled tRNA transcripts that can be methylatedin vitro by DnmA formed denaturation resistantcomplexes while non-target RNAs did not To confirmthat the denaturation resistant complexes were really dueto binding of the enzyme to the substrate SDS gels wereblotted and His-tagged DnmA was detected by a specificanti-His antibody A small amount of the tagged proteinwas shifted to the same position as the labelled tRNA(Supplementary Figure S5)
We then performed time courses for the reaction andfound that covalent complex formation was only transientunder the conditions used For tRNAAsp(GUC) adductsreached a maximum after 1min and were undetectableafter 15min whereas the tRNAGlu(UUC) complex onlybegan to accumulate after 1min but persisted for almost60min (Figure 5A and B) This coincided with the highmethylation activity on tRNAAsp(GUC-1) and a lower oneon tRNAGlu(UUC-2) The human enzyme appeared to beslightly faster in the reaction but otherwise showed asimilar behaviour compared with D discoideum DnmA
tRNAAsp(GUC) tRNAGlu(UUCCUC) and tRNAGly(GCC)
bind specifically to DnmA in vivo
The blocking assay (Figure 3) suggested that further sub-strates could be targeted by DnmA To identify suchtargets and to further investigate the interaction betweenDnmA and RNAs in vivo we established a UV-CLIP pro-cedure for D discoideumDnmA-GFP expressing cells andcontrol cells expressing GFP only were UV cross-linkedand complexes were precipitated under denaturing condi-tions by a GFP nanobody (21) Then co-precipitatedRNAs were isolated converted into strand-specificcDNA libraries and analyzed by Illumina sequencing(Figure 6 and Supplmentary Figure S6)
Various tRNA fragments were enriched in the DnmACLIP sample compared with the control By their lengthand position fragments appeared not to be random butspecific We found mid-fragments of 20ndash24 nt (position32ndash36 to 53ndash56) that covered the C38 methylation site inthe respective tRNAs 50-fragments of 17 nt (nucleotideposition 1ndash17) 30-fragments of 2021 nt (nucleotideposition 54 to the end) and 30-half tRNAs of 41ndash43 nt
(position 30ndash32 to the end) Mid-fragments oftRNAAsp(GUC) tRNAGlu(UUCCUC) and tRNAGly(GCC)
were significantly enriched in the DnmA-GFP fraction incomparison with the control CLIP assay tRNAAsp(GUC)tRNAGlu(UUC) and tRNAGlu(CUC) harbour the sequencepattern (C32 C34 A37 C40) which has been suggestedas a signature for C38 methylation (Figure 2 Helmpersonal communication) tRNAGly(GCC) contains thepattern except for C40 For tRNAAla(AGC) which doesnot contain C38 and the signature pattern a differentfragment from the 30-end was enriched in the DnmA-CLIP sample For other tRNAs no significant accumula-tion was observed in the experiment (Figure 6 andSupplementary Figure S6) For several tRNAs varyingamounts of full-length molecules were found but thesewere never enriched in the DnmA sample in comparisonwith the control
tRNAGly(GCC) is an additional methylation substratein vitro
The CLIP data suggested that tRNAGly(GCC) was anothertarget for DnmA We therefore cloned this and several othertRNA genes that contained the pattern only partially(Figure 2) We generated in vitro transcripts and examinedmethylation by DnmA and hDnmt2 in vitro IndeedtRNAGly(GCC) was methylated by both enzymes (Figure 7)while none of the other tRNAs (tRNALeu(UAG-1)tRNALeu(AAG-8) tRNAVal(CAC-1) and tRNAPhe(GAA-2))resulted in detectable methylation signals in the assay(Supplementary Figure S7) Interestingly the in vitro methy-lation efficiency of the three tRNA substrates
Figure 5 Formation and turnover of tRNA-methyltransferasecomplexes Turnover of covalent complexes of tRNA substrates withDnmA and hDnmt2 (A) Examples of time courses on covalentcomplex formation with tRNAAsp(GUC) and tRNAGlu(UUC) (B)Complex bands in A were quantified and presented on a time scale
Nucleic Acids Research 2013 7
at Universitaumlt W
uumlrzburg on August 5 2013
httpnaroxfordjournalsorgD
ownloaded from
(tRNAAsp(GUC) tRNAGlu(UUC) tRNAGly(GCC)) correlatedroughly with the number of tRNA fragments found in theRNA-CLIP experimentThe data supported the conclusion from the ex vivo
blocking assay (Figure 3B) that in addition toD discoideum tRNAAsp and tRNAGlu other RNAs weresubstrates for DnmA and hDnmt2 in vitro
Expression of dnmA is regulated during developmentcell cycle and on temperature stress
DnmA expression in growing vegetative D discoideumcells is low To examine tRNA methylation in vivo it
was therefore advantageous to determine conditionsunder which dnmA was most highly expressed Promptedby previous studies (141517) we assumed that the biolo-gical function of DnmA was connected to developmentalandor environmental conditions We therefore examinedexpression levels of dnmA under different regimes dnmAmRNA was quantified by qPCR over the 24 h develop-mental cycle of D discoideum (data not shown) and con-firmed the data deposited in Dictyexpress (httpdictyexpressbiolabsi)
Interestingly an 5-fold increase in expression wasobserved at 16 h in development This time coincides
Figure 6 tRNA and tRNA fragments associated with DnmA-GFP After UV crosslinking RNA bound to DnmA was co-immunoprecipitatedunder denaturing conditions (CLIP) The number of normalised reads for tRNAs detected by Illumina sequencing is shown for DnmA CLIP and forthe control CLIP with GFP In addition to full-length tRNAs four classes of tRNA fragments were found and these are indicated by differentshading of the bars Size and localization of fragments is shown schematically on the simplified clover leaf structure tRNAs that are not significantlyenriched with DnmA are shown in Supplementary Figure S6 In case of multiple gene copys resulting in the same RNA transcript the sequencingreads were mapped to the first copy in the genome starting from chromsome 1 Only this copy is listed Sequences and detailed information on genecopies and potential isoacceptors are listed in Supplementary Table S2
8 Nucleic Acids Research 2013
at Universitaumlt W
uumlrzburg on August 5 2013
httpnaroxfordjournalsorgD
ownloaded from
with a presumed synchronous mitotic division (28) wherea multitude of cell-cycle regulated genes are expressed[(29) and McWilliams personal Communication]
Therefore we synchronized vegetative D discoideumcells to examine dnmA expression during the cell cycleFor these experiments the D discoideum wild-typestrain NC4 the parent of the axenic laboratory strainAX2 was used (30) This was prompted by the observationthat D melanogaster and E histolytica laboratory strainsmay lose dnmt2 expression (Reuter Ankri personalcommunication)
The data confirmed our assumption of a cell-cycle-de-pendent transcriptional regulation of dnmA (Figure 8A)The peak of expression coincided with late mitosis or earlyS-phase as D discoideum does not display a G1 phaseUsing AX2 cells cotransformed with PCNA-RFP andDnmA-GFP we observed GFP accumulation in thenucleus during S phase (indicated by PCNA expression)and a loss of DnmA from the nucleus during mitosis(Figure 8B) This is further exemplified by the movie inthe Supplementary Material (Supplementary Movie S1)
In other organisms dnmt2 was observed to beupregulated during stress response (315) We thereforeexamined expression after anoxia heat shock and coldshock but did not find any significant changes (data notshown) Only during recovery from cold shock wedetected a strong transient increase in dnmA mRNA by47-fold (Figure 8C)
Thus dnmA expression was differentially regulatedduring the cell cycle development and on temperaturestress Furthermore DnmA localization changed specific-ally during the cell cycle
Though the basic expression in vegetative cells was in-distinguishable dnmA expression in NC4 was 30-foldhigher at 16 h of development whereas developmentalupregulation was only 5-fold in AX2 The data aresummarized in Figure 8D
NC4 the parent strain of AX2 displays increasedtRNA
Asp methylation during development
Based on these observations we asked the questionwhether expression levels correlated with tRNA
methylation activity in vivo Bisulfite sequencing wasdone with primers for tRNAAsp tRNAGlu and tRNAGly
on RNA isolated from vegetative AX2 cells AX2 cellsdeveloped for 16 h AX2 cells subjected to cold shockAX2 recovered for 25 h after cold shock vegetativeNC4 cells and NC4 cells developed for 16 h The resultson C38 methylation of tRNAAsp(GUC) are summarized inFigure 8E The methylation status of all C residues fromall tested tRNAs in this study is shown in SupplementaryFigure S8 Indeed tRNAAsp(GUC) methylation at C38increased to 76 in development in NC4 cells Noincrease was observed in AX2 cells during developmentpossibly due to insufficient sensitivity of the assayInterestingly no increase in tRNAAsp(GUC) methylationwas observed at the highest dnmA expression levelsduring stress recovery In Figure 8E the 30ndash35 methy-lation level in all tRNAAsp(GUC) samples represents signifi-cant methylation as compared with other C residues (seeSupplementary Figure S8)We did however not detect any significant methylation
of tRNAGlu(UUCCUC) and tRNAGly(GCC) under any con-ditions (Supplementary Figure S8) In contrast C49C50methylation which does not depend on Dnmt2 enzymeswas found in all samples and thus served as an internalcontrol In all 1175 bisulfite-sequence reads wereexamined for tRNAGlu(UUCCUC) and 4040 fortRNAGly(GCC) No methylation above background wasfound Though it cannot be excluded that the failure todetect C38 methylation of tRNAGlu and tRNAGly was dueto inefficient reverse transcription or low levels of methy-lation (see lsquoDiscussionrsquo section) we conclude thattRNAAsp is the major or possibly the only in vivo targetfor methylation by DnmA in D discoideum
DISCUSSION
The biological function of Dnmt2 methyltransferases islargely unknown and its role in DNA methylation is con-troversially discussed (910) In contrast tRNA methyla-tion by Dnmt2 is robust in vitro and in vivo andevolutionary conserved In addition to the originallydescribed tRNAAsp(GUC) (1) two further tRNAs targets(tRNAVal(CAC) and tRNAGly(GCC)) have been defined inD melanogaster (15) and tRNAGlu(UUC) in S pombe (17)To get insight into potential functions of the
D discoideum Dnmt2 homologue DnmA we examinedexpression levels in development in the cell cycle andunder different environmental conditions We confirmedby qPCR an expression peak at 16 h of developmentWeijer and co-workers (28) had previously proposed asynchronous cell division at this developmental stageWe therefore investigated dnmA expression in the cellcycle and found a strong transient increase of mRNA inS-phase or shortly thereafter The expression peak wasaccompanied by reaccumulation of DnmA in thenucleus We assume that DnmA is degraded duringmitosis and re-synthesised after cell divisionIn S pombe as well as in D melanogaster tRNA methy-
lation changes in response to environmental challengeslike temperature stress anoxia or nutrients We tested
Figure 7 In vitro methylation of tRNAGly(GCC) by recombinant DnmAand hDnmt2 In vitro transcribed tRNAs were methylated in vitro asdescribed before The top panel shows the ethidium-bromide stainedgel the bottom panel the fluorogram of 3H-labelled methylated tRNAsAs a control tRNAAsp(GUC) methylation is shown
Nucleic Acids Research 2013 9
at Universitaumlt W
uumlrzburg on August 5 2013
httpnaroxfordjournalsorgD
ownloaded from
Figure 8 Expression localization and tRNAAsp(GUC) methylation activity of DnmA under various conditions (A) dnmA expression is regulatedduring the cell cycle Cells were arrested in the cell cycle by cold treatment and then released by transfer to 22C Cells were counted every 30min(grey dots and grey line) and samples were taken for qPCR (black bars) After cell division dnmA expression increased 5-fold and then rapidlydeclined to basal levels Normalization was done on vegetative growing cells dnmA expression is only shown from 3 to 45 h during recovery(B) DnmA is lost from the nucleus during mitosis (arrow) The three cells on the right are in S-phase as indicated by the RFP-PCNA marker andaccumulate DnmA in the nucleus (for further details see movie in Supplementary Material) (C) Relative quantification of dnmA expression levels inAX2 cells after 25 h recovery from cold shock at 4C Expression of dnmA increased gt40-fold and returned within 30min to basal levels (n=3)(D) At 16 h of development dnmA expression increased 46-fold in the NC4 strain In AX2 cells expression increased only 5-fold (E) At 16 h ofdevelopment in vivo methylation of C38 in tRNAAsp(GUC) increased up to 75 in the D discoideum strain NC4 whereas no significant increase wasobserved in AX2 cells
10 Nucleic Acids Research 2013
at Universitaumlt W
uumlrzburg on August 5 2013
httpnaroxfordjournalsorgD
ownloaded from
heat shock cold shock anoxia and various nutrient mediabut did not observe altered levels Only during therecovery phase after cold treatment a transient almost50-fold increase in mRNA abundance was seen
Reuter and Ankri (personal communication) hadobserved a loss of Dnmt2 expression in laboratorystrains of D melanogaster and E histolytica We thereforeexamined D discoideum NC4 the parent of the axeniclaboratory strain AX2 The developmental regulation inAX2 and NC4 was qualitatively similar but expressionlevels were much higher in NC4 The observationsupports the assumption that Dnmt2 expression wasreduced under optimal growth conditions in the labora-tory strain and may actually be selected against
We show that the D discoideum Dnmt2 homologueDnmA is an active tRNAAsp(GUC) methyltransferasein vitro and in vivo To identify further potential targetsfor DnmA we established a UV-CLIP method to analyseDnmAndashRNA interaction in vivo In addition to the knowntarget tRNAAsp(GUC) fragments of tRNAGlu(UUC) andtRNAGly(GCC) were most abundant in comparison withthe control Unexpectedly complete tRNAs were notenriched in the DnmA CLIP fraction but rather specificlsquomid-fragmentsrsquo that covered the putative methylation siteand the recognition pattern proposed by Helm (personal
communication) A minor exception was tRNAGly(GCC)which lacks C40 We do not know whether these frag-ments were directly or indirectly generated by DnmAin vivo or whether they were protected by DnmA fromdegradation during the experimental procedure As noco-crystal of Dnmt2 and a tRNA substrate is availablethe exact interface for the interaction is unknownJurkowski et al (31) mapped the binding site on Dnmt2by mutational analysis A manual placement of a tRNAinto the crystal structure suggests that Dnmt2 mainlycontacts the anticodon stem loop Assuming that mid-frag-ments are footprints of DnmA our data support this notionbut suggest that the interaction surface extends into theD-loop which is partially covered by the mid-fragments
We examined the putative DnmA targets in vitro (bymethylation of in vitro transcribed tRNAs) ex vivo(by methylation of RNA isolated from D discoideum)and in vivo (by bisulfite sequencing) Furthermore weinvestigated by SDSndashPAGE the kinetics of covalentcomplex formation between tRNAAsp and tRNAGlu withDnmA and hDnmt2
All three tRNAs (tRNAAsp tRNAGlu tRNAGly) werereadily methylated by recombinant DnmA and hDnmt2in vitro although the activity was much higher (estimatedat least 10-fold) on tRNAAsp compared with tRNAGlu andtRNAGly For tRNAAsp and tRNAGlu this correlated wellwith the formation and decay of covalent RNAmdashproteincomplexes where kinetics for tRNAAsp were much fasterthan for tRNAGlu Instead of C methylation the covalentintermediate may also proceed to a hydrogen exchangereaction or to a deamination of the cytosine In bothcases an intermediate will be detected but it will notresult in a methylated product (32) It may be thatslower off rates represent these alternative pathways thatwould result in unmethylated tRNAs
Using antisense oligos we showed that tRNAAsptRNAGlu and at least one additional RNA could bemethylated ex vivo A quantification of the results wasnot possible the apparently low methylation oftRNAGlu ex vivo could be due to the low activity of theenzyme on this substrate (as suggested by the in vitroexperiments see Figure 5B)The data for the examination of in vivo methylation by
bisulfite sequencing were rather unexpected AlthoughtRNAAsp(GUC) was consistently found to be methylatedat C38 we never detected any methylation at C38 oftRNAGlu(UUCCUC) or tRNAGly(GCC) There could betwo reasons to explain the failure to detect C38 methyla-tion for these in vitro substrates (i) due to the low activityof DnmA on tRNAGlu(UUCCUC) and tRNAGly(GCC) asshown in vitro there was low or no significant methylationin vivo and (ii) methylation escaped detection becauseother unknown modifications blocked reverse transcrip-tion of the methylated molecules However if this wasthe case one has to assume that C49C50 methylationoccurs independently of these unknown modificationswhereas C38 methylation strictly depends on them AsC38 methylation does not require any other modificationsin vitro we believe that this scenario is less likely If in vitromethylation activity was comparable with in vivo methy-lation 10 methylation of tRNAGlu(UUCCUC) andtRNAGly(GCC) could be expected relative totRNAAsp(GUC) As tRNAAsp was methylated to 30 a3 methylation of tRNAGlu and tRNAGly could be belowthe detection level of the bisulfite assayWe find that high expression of dnmA did not necessar-
ily correlate with increased methylation of tRNAs Duringrecovery from cold shock D discoideum cells displayedthe highest level of dnmA expression but this had noeffect on tRNAAsp methylation Only in developmentand with the high expressing NC4 strain a significantincrease of tRNAAsp methylation to 70 was observedIt has to be considered however that bisulfite sequencingdoes not necessarily allow for quantitative evaluation ofdata detected C-methylation depends on reverse tran-scription efficiency which may be hampered by othermodifications that could be different under differentconditions Furthermore the completeness of bisulfite-mediated conversion could also depend on RNA struc-ture which may be altered by other modificationsHowever incomplete conversion will lead to an overesti-mation of C-methylation as maybe the case for forexample C30 C31 C35 in tRNAGly in some samplesTaken together the data from CLIP complex forma-
tion and methylation show that all three substrates can bebound and methylated in vitro The CLIP data show thatthere is a specific interaction of DnmA with tRNA mid-fragments of tRNAAsp(GUC) tRNAGlu(UUCCUC) andtRNAGly(GCC) but not with other tRNAs In contrastbisulfite sequencing could only detect methylation ontRNAAsp(GUC) even at the highest expression levels ofdnmA Even though low amounts of methylation mayhave escaped detection in bisulfite sequencing there isstill a discrepancy between dnmA expression levels (egduring cold shock recovery) DnmA binding to substrates(as determined by CLIP) and methylation in vivo
Nucleic Acids Research 2013 11
at Universitaumlt W
uumlrzburg on August 5 2013
httpnaroxfordjournalsorgD
ownloaded from
We therefore suggest that DnmA binding to specifictargets could elicit functions in addition to RNA methy-lation eg by promoting processing or degradation andor recruitment of other proteins At least for some rRNAmethyltransferases such as Bud23 or Nep1 fromSaccharomyces cerevisiae it is known that they executefunctions independent from the methylation activity(33ndash35) As shown by Schaefer and co-workers (15)methylation protects tRNAs from degradation or process-ing after stress By a different pathway binding of DnmAmay initiate processing that leads to specific tRNA frag-ments that could serve for other biological functions (36)Interestingly tRNAGlu(UUC) is also a substrate for theDnmt2 homologue Pmt1 in S pombe (17) HowevertRNAGlu(UUC) is found to be methylated to a low butsignificant extend in vivo in fission yeast when Pmt1 isoverexpressedThe detection of tRNAAla(AGC) fragments in the CLIP
assay brings up a new question tRNAAla(AGC) has a Tinstead of a C in position 38 and does not contain thesignature nucleotides in the anticodon stem-loopInstead of a mid-fragment a distinct 30 fragment wasassociated with DnmA suggesting that a recognitionmotif was located in this part of the moleculeAs methylation also occurred in a large artificial
molecule with embedded tRNAGlu (SupplementaryFigure S2) it could be that other longer RNAs containingan appropriate recognition motif may also be bound bythe enzyme We thus propose that the biological functionsof Dnmt2 proteins could extend far beyond our presentknowledge Investigation of further targets and of the sig-nificance of RNA fragments associated with Dnmt2should shed light on the importance of this enzyme
SUPPLEMENTARY DATA
Supplementary Data are available at NAR Online
ACKNOWLEDGEMENTS
The authors thank Thomas Winckler (Friedrich-Schiller-University Jena) for providing the tRNAGlu(UUC) and sup-pressor tRNAGlu(CUA) plasmids Susanne Junk for cloningtRNAAspC38A Matthias Schafer (DKFZ) and RobinLorenz (University of Kassel) are acknowledged forRNA bisulfite sequencing and Richard Reinhardt (MaxPlanck Genome Centre Cologne) for deep sequencingMark Helm (Johannes-Gutenberg-University Mainz) gen-erously provided data before publication CarstenSeehafer (University of Kassel) helped with the analysisof deep-sequencing data
FUNDING
Deutsche Forschungsgemeinschaft [FOR 1082 to WNand AJ] and the Studienstiftung des Deutschen Volkes(to SM) Funding for open access charge FOR1082
Conflict of interest statement None declared
REFERENCES
1 GollMG KirpekarF MaggertKA YoderJA HsiehCLZhangX GolicKG JacobsenSE and BestorTH (2006)Methylation of tRNAAsp by the DNA methyltransferasehomolog Dnmt2 Science 311 395ndash398
2 PhalkeS NickelO WalluscheckD HortigF OnoratiMCand ReuterG (2009) Retrotransposon silencing and telomereintegrity in somatic cells of Drosophila depends on the cytosine-5methyltransferase DNMT2 Nat Genet 41 696ndash702
3 WilkinsonCR BartlettR NurseP and BirdAP (1995) Thefission yeast gene pmt1+ encodes a DNA methyltransferasehomologue Nucleic Acids Res 23 203ndash210
4 KatohM CurkT XuQ ZupanB KuspaA and ShaulskyG(2006) Developmentally regulated DNA methylation inDictyostelium discoideum Eukaryot Cell 5 18ndash25
5 KuhlmannM BorisovaBE KallerM LarssonP StachDNaJ EichingerL LykoF AmbrosV SoderbomF et al(2005) Silencing of retrotransposons in Dictyostelium by DNAmethylation and RNAi Nucleic Acids Res 33 6405ndash6417
6 RaiK ChidesterS ZavalaCV ManosEJ JamesSRKarpfAR JonesDA and CairnsBR (2007) Dnmt2 functionsin the cytoplasm to promote liver brain and retina developmentin zebrafish Genes Dev 21 261ndash266
7 FisherO Siman-TovR and AnkriS (2006) Pleiotropicphenotype in Entamoeba histolytica overexpressing DNAmethyltransferase (Ehmeth) Mol Biochem Parasitol 147 48ndash54
8 DurdevicZ HannaK GoldB PollexT CherryS LykoFand SchaeferM (2013) Efficient RNA virus control in Drosophilarequires the RNA methyltransferase Dnmt2 EMBO Rep 14269ndash275
9 SchaeferM and LykoF (2010) Lack of evidence for DNAmethylation of Invader4 retroelements in Drosophila andimplications for Dnmt2-mediated epigenetic regulation NatGenet 42 920ndash921 author reply 921
10 KraussV and ReuterG (2011) DNA methylation in Drosophila -a critical evaluation Prog Mol Biol Transl Sci 101 177ndash191
11 RaddatzG GuzzardoPM OlovaN FantappieMRRamppM SchaeferM ReikW HannonGJ and LykoF(2013) Dnmt2-dependent methylomes lack defined DNAmethylation patterns Proc Natl Acad Sci USA 110 8627ndash8631
12 JeltschA NellenW and LykoF (2006) Two substrates arebetter than one dual specificities for Dnmt2 methyltransferasesTrends Biochem Sci 31 306ndash308
13 JurkowskiTP MeusburgerM PhalkeS HelmM NellenWReuterG and JeltschA (2008) Human DNMT2 methylatestRNA(Asp) molecules using a DNA methyltransferase-likecatalytic mechanism RNA 14 1663ndash1670
14 TovyA Siman TovR GaentzschR HelmM and AnkriS(2010) A new nuclear function of the Entamoeba histolyticaglycolytic enzyme enolase the metabolic regulation of cytosine-5methyltransferase 2 (Dnmt2) activity PLoS Pathog 6 e1000775
15 SchaeferM PollexT HannaK TuortoF MeusburgerMHelmM and LykoF (2010) RNA methylation by Dnmt2protects transfer RNAs against stress-induced cleavage GenesDev 24 1590ndash1595
16 TuortoF LiebersR MuschT SchaeferM HofmannSKellnerS FryeM HelmM StoecklinG and LykoF (2012)RNA cytosine methylation by Dnmt2 and NSun2 promotestRNA stability and protein synthesis Nat Struct Mol Biol 19900ndash905
17 BeckerM MullerS NellenW JurkowskiTP JeltschA andEhrenhofer-MurrayAE (2012) Pmt1 a Dnmt2 homolog inSchizosaccharomyces pombe mediates tRNA methylation inresponse to nutrient signaling Nucleic Acids Res 4011648ndash11658
18 TovyA HertzR Siman-TovR SyanS FaustD GuillenNand AnkriS (2011) Glucose starvation boosts Entamoebahistolytica virulence PLoS Negl Trop Dis 5 e1247
19 MotorinY and HelmM (2011) RNA nucleotide methylationWiley Interdiscip Rev RNA 2 611ndash631
20 NiranjanakumariS LasdaE BrazasR and Garcia-BlancoMA(2002) Reversible cross-linking combined with
12 Nucleic Acids Research 2013
at Universitaumlt W
uumlrzburg on August 5 2013
httpnaroxfordjournalsorgD
ownloaded from
immunoprecipitation to study RNA-protein interactions in vivoMethods 26 182ndash190
21 RothbauerU ZolghadrK MuyldermansS SchepersACardosoMC and LeonhardtH (2008) A versatile nanotrapfor biochemical and functional studies with fluorescent fusionproteins Mol Cell Proteomics 7 282ndash289
22 SittkaA LucchiniS PapenfortK SharmaCM RolleKBinnewiesTT HintonJC and VogelJ (2008) Deep sequencinganalysis of small noncoding RNA and mRNA targets of theglobal post-transcriptional regulator Hfq PLoS Genet 4e1000163
23 HoffmannS OttoC KurtzS SharmaCM KhaitovichPVogelJ StadlerPF and HackermullerJ (2009) Fast mapping ofshort sequences with mismatches insertions and deletions usingindex structures PLoS Comput Biol 5 e1000502
24 ConnollyMA ClausenPA and LazarJG (2006) Purificationof RNA from animal cells using trizol Cold Spring HarborProtoc 2006 pii pdbprot4104
25 DongA YoderJA ZhangX ZhouL BestorTH andChengX (2001) Structure of human DNMT2 an enigmaticDNA methyltransferase homolog that displays denaturant-resistant binding to DNA Nucleic Acids Res 29 439ndash448
26 SchaeferM PollexT HannaK and LykoF (2009) RNAcytosine methylation analysis by bisulfite sequencing NucleicAcids Res 37 e12
27 BoninM OberstrassJ VogtU WasseneggerM and NellenW(2001) Binding of IRE-BP to its cognate RNA sequence SFMstudies on a universal RNA backbone for the analysis of RNA-protein interaction Biol Chem 382 1157ndash1162
28 ZimmermanW and WeijerCJ (1993) Analysis of cell cycleprogression during the development of Dictyostelium and itsrelationship to differentiation Dev Biol 160 178ndash185
29 StrasserK BloomfieldG MacWilliamsA CeccarelliAMacWilliamsH and TsangA (2012) A retinoblastomaorthologue is a major regulator of S-phase mitotic anddevelopmental gene expression in Dictyostelium PLoS One 7e39914
30 WattsDJ and AshworthJM (1970) Growth of myxameobae ofthe cellular slime mould Dictyostelium discoideum in axenicculture Biochem J 119 171ndash174
31 JurkowskiTP ShanmugamR HelmM and JeltschA (2012)Mapping the tRNA binding site on the surface of humanDNMT2 methyltransferase Biochemistry 51 4438ndash4444
32 SvedruzicZM and ReichNO (2005) DNA cytosine C5methyltransferase Dnmt1 catalysis-dependent release of allostericinhibition Biochemistry 44 9472ndash9485
33 ThomasSR KellerCA SzykA CannonJR and Laronde-LeblancNA (2011) Structural insight into the functionalmechanism of Nep1Emg1 N1-specific pseudouridinemethyltransferase in ribosome biogenesis Nucleic Acids Res 392445ndash2457
34 LeulliotN BohnsackMT GrailleM TollerveyD andVan TilbeurghH (2008) The yeast ribosome synthesisfactor Emg1 is a novel member of the superfamily of alphabeta knot fold methyltransferases Nucleic Acids Res 36629ndash639
35 WhiteJ LiZ SardanaR BujnickiJM MarcotteEM andJohnsonAW (2008) Bud23 methylates G1575 of 18S rRNA andis required for efficient nuclear export of pre-40S subunits MolCell Biol 28 3151ndash3161
36 SobalaA and HutvagnerG (2013) Small RNAs derived fromthe 50-end of tRNA can inhibit protein translation in humancells RNA Biol 10 525ndash535
Nucleic Acids Research 2013 13
at Universitaumlt W
uumlrzburg on August 5 2013
httpnaroxfordjournalsorgD
ownloaded from

As tRNAAsp appeared to be not the only target forDnmt2 in other organisms (eg D melanogaster) weexamined if more cellular-derived RNAs could bemethylated in vitro (ex vivo methylation experiments)Total RNA from dnmAKO AX2 cells was subjected toin vitro methylation by recombinant DnmA for varioustimes Methylated RNAs were separated by gelelectro-phoresis and visualized by autoradiography Figure 1Cshows that a substantial RNA fraction in the tRNA sizerange could be methylated At longer incubation times asecond minor methylated RNA species appeared
indicating that at least one further substrate could bemethylated in vitro
Identification of further substratesmdashin vitro binding andmethylation of tRNAGlu
The group of Helm (personal communication) hadproposed a sequence pattern (C32 C34 A37 C40) thatmay serve as a target recognition motif for C38 methyla-tion by hDnmt2 We therefore examined all D discoideumtRNAs in silico to identify candidates for the additionalmethylated band in the ex vivo experiment In addition to
Figure 1 In vitro methylation of tRNAAsp(GUC) (A) In vitro methylation of tRNAAsp(GUC) by DnmA at 2mM 5mM and 10mM MgCl2 and byhDnmt2 at 5mM Mg2+ The upper panel shows ethidium bromide staining of the in vitro transcripts separated in a denaturing polyacrylamide gelthe lower panel shows incorporated 3H-Me in the tRNAs Reactions were run for the times indicated (B) In vivo methylation of cytosines intRNAAsp(GUC) from different D discoideum strains Results of the RNA bisulfite sequencing (454 pyrosequencing) are given in percentage of readsNumbers of sequence reads are shown in brackets C49 is methylated by a different methyltransferase and thus serves as an internal standard All 22tRNAAsp(GUC) genes result in the same transcript and no isoacceptors are encoded in the D discoideum genome (C) In vitro methylation of smallenriched RNA of a dnmAKO strain (ex vivo methylation) The methylation reaction was done for the times indicated
Nucleic Acids Research 2013 5
at Universitaumlt W
uumlrzburg on August 5 2013
httpnaroxfordjournalsorgD
ownloaded from
tRNAAsp(GUC) we found tRNAGlu(UUC) andtRNAGlu(CUC) to match the pattern (Figure 2 andSupplementary Table S2)To test for tRNAGlu as a potential substrate we
modified the methylation assay by addition of a 50 ntDNA antisense oligonucleotide to tRNAAsp(GUC)
covering the methylation target site Figure 3A shows asa control that methylation of in vitro transcribedtRNAAsp(GUC) was completely blocked We then per-formed ex vivo methylation assays with enriched smallRNAs isolated from D discoideum after blockingtRNAAsp(GUC) with the antisense oligo Figure 3B showsthat the majority of 3H-incorporation disappearedHowever with both DnmA and hDnmt2 a significantamount of methylation was still detectable in the sizerange of tRNAs Assuming that this signal containedtRNAGlu we used a second antisense oligo directedagainst tRNAGlu(UUC) for blocking methylation Theoligo also covered the isoacceptor tRNAGlu(CUC) with
minor mismatches (Supplementary Figure S2) A furtherreduction of ex vivo methylation was observed Notablythe lower band in the tRNA size range disappeared (arrowin Figure 3B) suggesting that this represented a tRNAGlu
population that could be methylated at least in vitro Theresidual methylation after blocking with both oligos mayrepresent additional tRNAs or other unknown targets Aswe only concentrated on small RNAs larger methylatedmolecules and less abundant targets may have escapeddetection
To further analyse methylation on tRNAAsp andtRNAGlu (exemplified by tRNAGlu(UUC)) we carried outassays with wild-type sequences and C38A mutatedin vitro transcripts (Figure 4) We also included a suppres-sor tRNAGlu(CUA) that differed in only 2 nt fromtRNAGlu(UUC) (Figure 2) Assays were done in parallelwith the recombinant hDnmt2 and D discoideumDnmA The experiment showed that tRNAAsp(GUC) wasthe best substrate for both DnmA and hDnmt2 (Figure 4)Substantial methylation was also observed fortRNAGlu(UUC) and suppressor tRNAGlu(CUA) Moreoverthe complete loss of methylation in the C38A mutations oftRNAAsp(GUC) and tRNAGlu(UUC) confirmed that methy-lation occurred at position C38 and largely ruled out thatmethyl group incorporation was due to another
Figure 3 Ex vivo methylation and blocking assay (A) Methylation of in vitro transcribed tRNAAsp(GUC) was completely blocked when a comple-mentary antisense oligo was hybridized (B) Using enriched small RNA from dnmAKO cells ex vivo methylation in the tRNA size class wasdifferentially lost when antisense oligos to tRNAAsp(GUC) and tRNAGlu(UUC) were hybridized before the methylation reaction The oligo againsttRNAGlu(UUC) also covers tRNAGlu(CUC) with minor mismatches (see Supplementary Figure S2) Even with both oligos a significant amount of 3Hincorporation still remained The upper panel shows the ethidiumbromide stained gel to demonstrate equal loading the lower panel shows thefluorogram The arrow marks a band that was specifically lost when the anti tRNAGlu(UUC) oligo was used
Figure 2 Target site sequences of tRNAs used for in vitro methylationThe position of the putative signature nucleotides are indicated andprinted in bold if present The anticodon is underlined tRNAGlu(CUC)
(not shown) also harbours the nucleotide pattern Except for the anti-codon both tRNAGlu are identical in the anticodon stem-loop region(Supplementary Figure S2) Only a few differences appear in theacceptor stem
Figure 4 C38 is the target for in vitro methylation of tRNAAsp(GUC)
and tRNAGlu(UUC) In vitro methylation of tRNAAsp(GUC)tRNAGlu(UUC) and a suppressor tRNAGlu(CUA) C38A mutations con-firmed C38 as the target nucleotide tRNAAsp(GUC) methylation wasmost efficient
6 Nucleic Acids Research 2013
at Universitaumlt W
uumlrzburg on August 5 2013
httpnaroxfordjournalsorgD
ownloaded from
contaminating methyltransferase in the recombinantprotein preparation (17)
The methylation experiments using in vitro transcribedtRNAs showed that no other modifications were necessaryfor C38 methylation by DnmA and hDnmt2 (13) and thatthe processed ends of a mature tRNA were not requiredfor target recognition as the tRNA transcripts contain no30-CCA Even tRNAGlu embedded in a long dsRNAmolecule (1000 nt) with small stem-loop structures atthe ends (27) was a specific substrate of DnmA andhDnmt2 (Supplementary Figure S3)
By EMSA no difference in in vitro binding could beseen for the specific target tRNAGlu(UUC) and non-targetRNAs indicating that DnmA associated non-specificallywith any nucleic acid (data not shown) However specificcovalent enzyme-substrate intermediates can be detectedby denaturing SDSndashPAGE (25) Figure 5A andSupplementary Figure S4 show that radioactivelylabelled tRNA transcripts that can be methylatedin vitro by DnmA formed denaturation resistantcomplexes while non-target RNAs did not To confirmthat the denaturation resistant complexes were really dueto binding of the enzyme to the substrate SDS gels wereblotted and His-tagged DnmA was detected by a specificanti-His antibody A small amount of the tagged proteinwas shifted to the same position as the labelled tRNA(Supplementary Figure S5)
We then performed time courses for the reaction andfound that covalent complex formation was only transientunder the conditions used For tRNAAsp(GUC) adductsreached a maximum after 1min and were undetectableafter 15min whereas the tRNAGlu(UUC) complex onlybegan to accumulate after 1min but persisted for almost60min (Figure 5A and B) This coincided with the highmethylation activity on tRNAAsp(GUC-1) and a lower oneon tRNAGlu(UUC-2) The human enzyme appeared to beslightly faster in the reaction but otherwise showed asimilar behaviour compared with D discoideum DnmA
tRNAAsp(GUC) tRNAGlu(UUCCUC) and tRNAGly(GCC)
bind specifically to DnmA in vivo
The blocking assay (Figure 3) suggested that further sub-strates could be targeted by DnmA To identify suchtargets and to further investigate the interaction betweenDnmA and RNAs in vivo we established a UV-CLIP pro-cedure for D discoideumDnmA-GFP expressing cells andcontrol cells expressing GFP only were UV cross-linkedand complexes were precipitated under denaturing condi-tions by a GFP nanobody (21) Then co-precipitatedRNAs were isolated converted into strand-specificcDNA libraries and analyzed by Illumina sequencing(Figure 6 and Supplmentary Figure S6)
Various tRNA fragments were enriched in the DnmACLIP sample compared with the control By their lengthand position fragments appeared not to be random butspecific We found mid-fragments of 20ndash24 nt (position32ndash36 to 53ndash56) that covered the C38 methylation site inthe respective tRNAs 50-fragments of 17 nt (nucleotideposition 1ndash17) 30-fragments of 2021 nt (nucleotideposition 54 to the end) and 30-half tRNAs of 41ndash43 nt
(position 30ndash32 to the end) Mid-fragments oftRNAAsp(GUC) tRNAGlu(UUCCUC) and tRNAGly(GCC)
were significantly enriched in the DnmA-GFP fraction incomparison with the control CLIP assay tRNAAsp(GUC)tRNAGlu(UUC) and tRNAGlu(CUC) harbour the sequencepattern (C32 C34 A37 C40) which has been suggestedas a signature for C38 methylation (Figure 2 Helmpersonal communication) tRNAGly(GCC) contains thepattern except for C40 For tRNAAla(AGC) which doesnot contain C38 and the signature pattern a differentfragment from the 30-end was enriched in the DnmA-CLIP sample For other tRNAs no significant accumula-tion was observed in the experiment (Figure 6 andSupplementary Figure S6) For several tRNAs varyingamounts of full-length molecules were found but thesewere never enriched in the DnmA sample in comparisonwith the control
tRNAGly(GCC) is an additional methylation substratein vitro
The CLIP data suggested that tRNAGly(GCC) was anothertarget for DnmA We therefore cloned this and several othertRNA genes that contained the pattern only partially(Figure 2) We generated in vitro transcripts and examinedmethylation by DnmA and hDnmt2 in vitro IndeedtRNAGly(GCC) was methylated by both enzymes (Figure 7)while none of the other tRNAs (tRNALeu(UAG-1)tRNALeu(AAG-8) tRNAVal(CAC-1) and tRNAPhe(GAA-2))resulted in detectable methylation signals in the assay(Supplementary Figure S7) Interestingly the in vitro methy-lation efficiency of the three tRNA substrates
Figure 5 Formation and turnover of tRNA-methyltransferasecomplexes Turnover of covalent complexes of tRNA substrates withDnmA and hDnmt2 (A) Examples of time courses on covalentcomplex formation with tRNAAsp(GUC) and tRNAGlu(UUC) (B)Complex bands in A were quantified and presented on a time scale
Nucleic Acids Research 2013 7
at Universitaumlt W
uumlrzburg on August 5 2013
httpnaroxfordjournalsorgD
ownloaded from
(tRNAAsp(GUC) tRNAGlu(UUC) tRNAGly(GCC)) correlatedroughly with the number of tRNA fragments found in theRNA-CLIP experimentThe data supported the conclusion from the ex vivo
blocking assay (Figure 3B) that in addition toD discoideum tRNAAsp and tRNAGlu other RNAs weresubstrates for DnmA and hDnmt2 in vitro
Expression of dnmA is regulated during developmentcell cycle and on temperature stress
DnmA expression in growing vegetative D discoideumcells is low To examine tRNA methylation in vivo it
was therefore advantageous to determine conditionsunder which dnmA was most highly expressed Promptedby previous studies (141517) we assumed that the biolo-gical function of DnmA was connected to developmentalandor environmental conditions We therefore examinedexpression levels of dnmA under different regimes dnmAmRNA was quantified by qPCR over the 24 h develop-mental cycle of D discoideum (data not shown) and con-firmed the data deposited in Dictyexpress (httpdictyexpressbiolabsi)
Interestingly an 5-fold increase in expression wasobserved at 16 h in development This time coincides
Figure 6 tRNA and tRNA fragments associated with DnmA-GFP After UV crosslinking RNA bound to DnmA was co-immunoprecipitatedunder denaturing conditions (CLIP) The number of normalised reads for tRNAs detected by Illumina sequencing is shown for DnmA CLIP and forthe control CLIP with GFP In addition to full-length tRNAs four classes of tRNA fragments were found and these are indicated by differentshading of the bars Size and localization of fragments is shown schematically on the simplified clover leaf structure tRNAs that are not significantlyenriched with DnmA are shown in Supplementary Figure S6 In case of multiple gene copys resulting in the same RNA transcript the sequencingreads were mapped to the first copy in the genome starting from chromsome 1 Only this copy is listed Sequences and detailed information on genecopies and potential isoacceptors are listed in Supplementary Table S2
8 Nucleic Acids Research 2013
at Universitaumlt W
uumlrzburg on August 5 2013
httpnaroxfordjournalsorgD
ownloaded from
with a presumed synchronous mitotic division (28) wherea multitude of cell-cycle regulated genes are expressed[(29) and McWilliams personal Communication]
Therefore we synchronized vegetative D discoideumcells to examine dnmA expression during the cell cycleFor these experiments the D discoideum wild-typestrain NC4 the parent of the axenic laboratory strainAX2 was used (30) This was prompted by the observationthat D melanogaster and E histolytica laboratory strainsmay lose dnmt2 expression (Reuter Ankri personalcommunication)
The data confirmed our assumption of a cell-cycle-de-pendent transcriptional regulation of dnmA (Figure 8A)The peak of expression coincided with late mitosis or earlyS-phase as D discoideum does not display a G1 phaseUsing AX2 cells cotransformed with PCNA-RFP andDnmA-GFP we observed GFP accumulation in thenucleus during S phase (indicated by PCNA expression)and a loss of DnmA from the nucleus during mitosis(Figure 8B) This is further exemplified by the movie inthe Supplementary Material (Supplementary Movie S1)
In other organisms dnmt2 was observed to beupregulated during stress response (315) We thereforeexamined expression after anoxia heat shock and coldshock but did not find any significant changes (data notshown) Only during recovery from cold shock wedetected a strong transient increase in dnmA mRNA by47-fold (Figure 8C)
Thus dnmA expression was differentially regulatedduring the cell cycle development and on temperaturestress Furthermore DnmA localization changed specific-ally during the cell cycle
Though the basic expression in vegetative cells was in-distinguishable dnmA expression in NC4 was 30-foldhigher at 16 h of development whereas developmentalupregulation was only 5-fold in AX2 The data aresummarized in Figure 8D
NC4 the parent strain of AX2 displays increasedtRNA
Asp methylation during development
Based on these observations we asked the questionwhether expression levels correlated with tRNA
methylation activity in vivo Bisulfite sequencing wasdone with primers for tRNAAsp tRNAGlu and tRNAGly
on RNA isolated from vegetative AX2 cells AX2 cellsdeveloped for 16 h AX2 cells subjected to cold shockAX2 recovered for 25 h after cold shock vegetativeNC4 cells and NC4 cells developed for 16 h The resultson C38 methylation of tRNAAsp(GUC) are summarized inFigure 8E The methylation status of all C residues fromall tested tRNAs in this study is shown in SupplementaryFigure S8 Indeed tRNAAsp(GUC) methylation at C38increased to 76 in development in NC4 cells Noincrease was observed in AX2 cells during developmentpossibly due to insufficient sensitivity of the assayInterestingly no increase in tRNAAsp(GUC) methylationwas observed at the highest dnmA expression levelsduring stress recovery In Figure 8E the 30ndash35 methy-lation level in all tRNAAsp(GUC) samples represents signifi-cant methylation as compared with other C residues (seeSupplementary Figure S8)We did however not detect any significant methylation
of tRNAGlu(UUCCUC) and tRNAGly(GCC) under any con-ditions (Supplementary Figure S8) In contrast C49C50methylation which does not depend on Dnmt2 enzymeswas found in all samples and thus served as an internalcontrol In all 1175 bisulfite-sequence reads wereexamined for tRNAGlu(UUCCUC) and 4040 fortRNAGly(GCC) No methylation above background wasfound Though it cannot be excluded that the failure todetect C38 methylation of tRNAGlu and tRNAGly was dueto inefficient reverse transcription or low levels of methy-lation (see lsquoDiscussionrsquo section) we conclude thattRNAAsp is the major or possibly the only in vivo targetfor methylation by DnmA in D discoideum
DISCUSSION
The biological function of Dnmt2 methyltransferases islargely unknown and its role in DNA methylation is con-troversially discussed (910) In contrast tRNA methyla-tion by Dnmt2 is robust in vitro and in vivo andevolutionary conserved In addition to the originallydescribed tRNAAsp(GUC) (1) two further tRNAs targets(tRNAVal(CAC) and tRNAGly(GCC)) have been defined inD melanogaster (15) and tRNAGlu(UUC) in S pombe (17)To get insight into potential functions of the
D discoideum Dnmt2 homologue DnmA we examinedexpression levels in development in the cell cycle andunder different environmental conditions We confirmedby qPCR an expression peak at 16 h of developmentWeijer and co-workers (28) had previously proposed asynchronous cell division at this developmental stageWe therefore investigated dnmA expression in the cellcycle and found a strong transient increase of mRNA inS-phase or shortly thereafter The expression peak wasaccompanied by reaccumulation of DnmA in thenucleus We assume that DnmA is degraded duringmitosis and re-synthesised after cell divisionIn S pombe as well as in D melanogaster tRNA methy-
lation changes in response to environmental challengeslike temperature stress anoxia or nutrients We tested
Figure 7 In vitro methylation of tRNAGly(GCC) by recombinant DnmAand hDnmt2 In vitro transcribed tRNAs were methylated in vitro asdescribed before The top panel shows the ethidium-bromide stainedgel the bottom panel the fluorogram of 3H-labelled methylated tRNAsAs a control tRNAAsp(GUC) methylation is shown
Nucleic Acids Research 2013 9
at Universitaumlt W
uumlrzburg on August 5 2013
httpnaroxfordjournalsorgD
ownloaded from
Figure 8 Expression localization and tRNAAsp(GUC) methylation activity of DnmA under various conditions (A) dnmA expression is regulatedduring the cell cycle Cells were arrested in the cell cycle by cold treatment and then released by transfer to 22C Cells were counted every 30min(grey dots and grey line) and samples were taken for qPCR (black bars) After cell division dnmA expression increased 5-fold and then rapidlydeclined to basal levels Normalization was done on vegetative growing cells dnmA expression is only shown from 3 to 45 h during recovery(B) DnmA is lost from the nucleus during mitosis (arrow) The three cells on the right are in S-phase as indicated by the RFP-PCNA marker andaccumulate DnmA in the nucleus (for further details see movie in Supplementary Material) (C) Relative quantification of dnmA expression levels inAX2 cells after 25 h recovery from cold shock at 4C Expression of dnmA increased gt40-fold and returned within 30min to basal levels (n=3)(D) At 16 h of development dnmA expression increased 46-fold in the NC4 strain In AX2 cells expression increased only 5-fold (E) At 16 h ofdevelopment in vivo methylation of C38 in tRNAAsp(GUC) increased up to 75 in the D discoideum strain NC4 whereas no significant increase wasobserved in AX2 cells
10 Nucleic Acids Research 2013
at Universitaumlt W
uumlrzburg on August 5 2013
httpnaroxfordjournalsorgD
ownloaded from
heat shock cold shock anoxia and various nutrient mediabut did not observe altered levels Only during therecovery phase after cold treatment a transient almost50-fold increase in mRNA abundance was seen
Reuter and Ankri (personal communication) hadobserved a loss of Dnmt2 expression in laboratorystrains of D melanogaster and E histolytica We thereforeexamined D discoideum NC4 the parent of the axeniclaboratory strain AX2 The developmental regulation inAX2 and NC4 was qualitatively similar but expressionlevels were much higher in NC4 The observationsupports the assumption that Dnmt2 expression wasreduced under optimal growth conditions in the labora-tory strain and may actually be selected against
We show that the D discoideum Dnmt2 homologueDnmA is an active tRNAAsp(GUC) methyltransferasein vitro and in vivo To identify further potential targetsfor DnmA we established a UV-CLIP method to analyseDnmAndashRNA interaction in vivo In addition to the knowntarget tRNAAsp(GUC) fragments of tRNAGlu(UUC) andtRNAGly(GCC) were most abundant in comparison withthe control Unexpectedly complete tRNAs were notenriched in the DnmA CLIP fraction but rather specificlsquomid-fragmentsrsquo that covered the putative methylation siteand the recognition pattern proposed by Helm (personal
communication) A minor exception was tRNAGly(GCC)which lacks C40 We do not know whether these frag-ments were directly or indirectly generated by DnmAin vivo or whether they were protected by DnmA fromdegradation during the experimental procedure As noco-crystal of Dnmt2 and a tRNA substrate is availablethe exact interface for the interaction is unknownJurkowski et al (31) mapped the binding site on Dnmt2by mutational analysis A manual placement of a tRNAinto the crystal structure suggests that Dnmt2 mainlycontacts the anticodon stem loop Assuming that mid-frag-ments are footprints of DnmA our data support this notionbut suggest that the interaction surface extends into theD-loop which is partially covered by the mid-fragments
We examined the putative DnmA targets in vitro (bymethylation of in vitro transcribed tRNAs) ex vivo(by methylation of RNA isolated from D discoideum)and in vivo (by bisulfite sequencing) Furthermore weinvestigated by SDSndashPAGE the kinetics of covalentcomplex formation between tRNAAsp and tRNAGlu withDnmA and hDnmt2
All three tRNAs (tRNAAsp tRNAGlu tRNAGly) werereadily methylated by recombinant DnmA and hDnmt2in vitro although the activity was much higher (estimatedat least 10-fold) on tRNAAsp compared with tRNAGlu andtRNAGly For tRNAAsp and tRNAGlu this correlated wellwith the formation and decay of covalent RNAmdashproteincomplexes where kinetics for tRNAAsp were much fasterthan for tRNAGlu Instead of C methylation the covalentintermediate may also proceed to a hydrogen exchangereaction or to a deamination of the cytosine In bothcases an intermediate will be detected but it will notresult in a methylated product (32) It may be thatslower off rates represent these alternative pathways thatwould result in unmethylated tRNAs
Using antisense oligos we showed that tRNAAsptRNAGlu and at least one additional RNA could bemethylated ex vivo A quantification of the results wasnot possible the apparently low methylation oftRNAGlu ex vivo could be due to the low activity of theenzyme on this substrate (as suggested by the in vitroexperiments see Figure 5B)The data for the examination of in vivo methylation by
bisulfite sequencing were rather unexpected AlthoughtRNAAsp(GUC) was consistently found to be methylatedat C38 we never detected any methylation at C38 oftRNAGlu(UUCCUC) or tRNAGly(GCC) There could betwo reasons to explain the failure to detect C38 methyla-tion for these in vitro substrates (i) due to the low activityof DnmA on tRNAGlu(UUCCUC) and tRNAGly(GCC) asshown in vitro there was low or no significant methylationin vivo and (ii) methylation escaped detection becauseother unknown modifications blocked reverse transcrip-tion of the methylated molecules However if this wasthe case one has to assume that C49C50 methylationoccurs independently of these unknown modificationswhereas C38 methylation strictly depends on them AsC38 methylation does not require any other modificationsin vitro we believe that this scenario is less likely If in vitromethylation activity was comparable with in vivo methy-lation 10 methylation of tRNAGlu(UUCCUC) andtRNAGly(GCC) could be expected relative totRNAAsp(GUC) As tRNAAsp was methylated to 30 a3 methylation of tRNAGlu and tRNAGly could be belowthe detection level of the bisulfite assayWe find that high expression of dnmA did not necessar-
ily correlate with increased methylation of tRNAs Duringrecovery from cold shock D discoideum cells displayedthe highest level of dnmA expression but this had noeffect on tRNAAsp methylation Only in developmentand with the high expressing NC4 strain a significantincrease of tRNAAsp methylation to 70 was observedIt has to be considered however that bisulfite sequencingdoes not necessarily allow for quantitative evaluation ofdata detected C-methylation depends on reverse tran-scription efficiency which may be hampered by othermodifications that could be different under differentconditions Furthermore the completeness of bisulfite-mediated conversion could also depend on RNA struc-ture which may be altered by other modificationsHowever incomplete conversion will lead to an overesti-mation of C-methylation as maybe the case for forexample C30 C31 C35 in tRNAGly in some samplesTaken together the data from CLIP complex forma-
tion and methylation show that all three substrates can bebound and methylated in vitro The CLIP data show thatthere is a specific interaction of DnmA with tRNA mid-fragments of tRNAAsp(GUC) tRNAGlu(UUCCUC) andtRNAGly(GCC) but not with other tRNAs In contrastbisulfite sequencing could only detect methylation ontRNAAsp(GUC) even at the highest expression levels ofdnmA Even though low amounts of methylation mayhave escaped detection in bisulfite sequencing there isstill a discrepancy between dnmA expression levels (egduring cold shock recovery) DnmA binding to substrates(as determined by CLIP) and methylation in vivo
Nucleic Acids Research 2013 11
at Universitaumlt W
uumlrzburg on August 5 2013
httpnaroxfordjournalsorgD
ownloaded from
We therefore suggest that DnmA binding to specifictargets could elicit functions in addition to RNA methy-lation eg by promoting processing or degradation andor recruitment of other proteins At least for some rRNAmethyltransferases such as Bud23 or Nep1 fromSaccharomyces cerevisiae it is known that they executefunctions independent from the methylation activity(33ndash35) As shown by Schaefer and co-workers (15)methylation protects tRNAs from degradation or process-ing after stress By a different pathway binding of DnmAmay initiate processing that leads to specific tRNA frag-ments that could serve for other biological functions (36)Interestingly tRNAGlu(UUC) is also a substrate for theDnmt2 homologue Pmt1 in S pombe (17) HowevertRNAGlu(UUC) is found to be methylated to a low butsignificant extend in vivo in fission yeast when Pmt1 isoverexpressedThe detection of tRNAAla(AGC) fragments in the CLIP
assay brings up a new question tRNAAla(AGC) has a Tinstead of a C in position 38 and does not contain thesignature nucleotides in the anticodon stem-loopInstead of a mid-fragment a distinct 30 fragment wasassociated with DnmA suggesting that a recognitionmotif was located in this part of the moleculeAs methylation also occurred in a large artificial
molecule with embedded tRNAGlu (SupplementaryFigure S2) it could be that other longer RNAs containingan appropriate recognition motif may also be bound bythe enzyme We thus propose that the biological functionsof Dnmt2 proteins could extend far beyond our presentknowledge Investigation of further targets and of the sig-nificance of RNA fragments associated with Dnmt2should shed light on the importance of this enzyme
SUPPLEMENTARY DATA
Supplementary Data are available at NAR Online
ACKNOWLEDGEMENTS
The authors thank Thomas Winckler (Friedrich-Schiller-University Jena) for providing the tRNAGlu(UUC) and sup-pressor tRNAGlu(CUA) plasmids Susanne Junk for cloningtRNAAspC38A Matthias Schafer (DKFZ) and RobinLorenz (University of Kassel) are acknowledged forRNA bisulfite sequencing and Richard Reinhardt (MaxPlanck Genome Centre Cologne) for deep sequencingMark Helm (Johannes-Gutenberg-University Mainz) gen-erously provided data before publication CarstenSeehafer (University of Kassel) helped with the analysisof deep-sequencing data
FUNDING
Deutsche Forschungsgemeinschaft [FOR 1082 to WNand AJ] and the Studienstiftung des Deutschen Volkes(to SM) Funding for open access charge FOR1082
Conflict of interest statement None declared
REFERENCES
1 GollMG KirpekarF MaggertKA YoderJA HsiehCLZhangX GolicKG JacobsenSE and BestorTH (2006)Methylation of tRNAAsp by the DNA methyltransferasehomolog Dnmt2 Science 311 395ndash398
2 PhalkeS NickelO WalluscheckD HortigF OnoratiMCand ReuterG (2009) Retrotransposon silencing and telomereintegrity in somatic cells of Drosophila depends on the cytosine-5methyltransferase DNMT2 Nat Genet 41 696ndash702
3 WilkinsonCR BartlettR NurseP and BirdAP (1995) Thefission yeast gene pmt1+ encodes a DNA methyltransferasehomologue Nucleic Acids Res 23 203ndash210
4 KatohM CurkT XuQ ZupanB KuspaA and ShaulskyG(2006) Developmentally regulated DNA methylation inDictyostelium discoideum Eukaryot Cell 5 18ndash25
5 KuhlmannM BorisovaBE KallerM LarssonP StachDNaJ EichingerL LykoF AmbrosV SoderbomF et al(2005) Silencing of retrotransposons in Dictyostelium by DNAmethylation and RNAi Nucleic Acids Res 33 6405ndash6417
6 RaiK ChidesterS ZavalaCV ManosEJ JamesSRKarpfAR JonesDA and CairnsBR (2007) Dnmt2 functionsin the cytoplasm to promote liver brain and retina developmentin zebrafish Genes Dev 21 261ndash266
7 FisherO Siman-TovR and AnkriS (2006) Pleiotropicphenotype in Entamoeba histolytica overexpressing DNAmethyltransferase (Ehmeth) Mol Biochem Parasitol 147 48ndash54
8 DurdevicZ HannaK GoldB PollexT CherryS LykoFand SchaeferM (2013) Efficient RNA virus control in Drosophilarequires the RNA methyltransferase Dnmt2 EMBO Rep 14269ndash275
9 SchaeferM and LykoF (2010) Lack of evidence for DNAmethylation of Invader4 retroelements in Drosophila andimplications for Dnmt2-mediated epigenetic regulation NatGenet 42 920ndash921 author reply 921
10 KraussV and ReuterG (2011) DNA methylation in Drosophila -a critical evaluation Prog Mol Biol Transl Sci 101 177ndash191
11 RaddatzG GuzzardoPM OlovaN FantappieMRRamppM SchaeferM ReikW HannonGJ and LykoF(2013) Dnmt2-dependent methylomes lack defined DNAmethylation patterns Proc Natl Acad Sci USA 110 8627ndash8631
12 JeltschA NellenW and LykoF (2006) Two substrates arebetter than one dual specificities for Dnmt2 methyltransferasesTrends Biochem Sci 31 306ndash308
13 JurkowskiTP MeusburgerM PhalkeS HelmM NellenWReuterG and JeltschA (2008) Human DNMT2 methylatestRNA(Asp) molecules using a DNA methyltransferase-likecatalytic mechanism RNA 14 1663ndash1670
14 TovyA Siman TovR GaentzschR HelmM and AnkriS(2010) A new nuclear function of the Entamoeba histolyticaglycolytic enzyme enolase the metabolic regulation of cytosine-5methyltransferase 2 (Dnmt2) activity PLoS Pathog 6 e1000775
15 SchaeferM PollexT HannaK TuortoF MeusburgerMHelmM and LykoF (2010) RNA methylation by Dnmt2protects transfer RNAs against stress-induced cleavage GenesDev 24 1590ndash1595
16 TuortoF LiebersR MuschT SchaeferM HofmannSKellnerS FryeM HelmM StoecklinG and LykoF (2012)RNA cytosine methylation by Dnmt2 and NSun2 promotestRNA stability and protein synthesis Nat Struct Mol Biol 19900ndash905
17 BeckerM MullerS NellenW JurkowskiTP JeltschA andEhrenhofer-MurrayAE (2012) Pmt1 a Dnmt2 homolog inSchizosaccharomyces pombe mediates tRNA methylation inresponse to nutrient signaling Nucleic Acids Res 4011648ndash11658
18 TovyA HertzR Siman-TovR SyanS FaustD GuillenNand AnkriS (2011) Glucose starvation boosts Entamoebahistolytica virulence PLoS Negl Trop Dis 5 e1247
19 MotorinY and HelmM (2011) RNA nucleotide methylationWiley Interdiscip Rev RNA 2 611ndash631
20 NiranjanakumariS LasdaE BrazasR and Garcia-BlancoMA(2002) Reversible cross-linking combined with
12 Nucleic Acids Research 2013
at Universitaumlt W
uumlrzburg on August 5 2013
httpnaroxfordjournalsorgD
ownloaded from
immunoprecipitation to study RNA-protein interactions in vivoMethods 26 182ndash190
21 RothbauerU ZolghadrK MuyldermansS SchepersACardosoMC and LeonhardtH (2008) A versatile nanotrapfor biochemical and functional studies with fluorescent fusionproteins Mol Cell Proteomics 7 282ndash289
22 SittkaA LucchiniS PapenfortK SharmaCM RolleKBinnewiesTT HintonJC and VogelJ (2008) Deep sequencinganalysis of small noncoding RNA and mRNA targets of theglobal post-transcriptional regulator Hfq PLoS Genet 4e1000163
23 HoffmannS OttoC KurtzS SharmaCM KhaitovichPVogelJ StadlerPF and HackermullerJ (2009) Fast mapping ofshort sequences with mismatches insertions and deletions usingindex structures PLoS Comput Biol 5 e1000502
24 ConnollyMA ClausenPA and LazarJG (2006) Purificationof RNA from animal cells using trizol Cold Spring HarborProtoc 2006 pii pdbprot4104
25 DongA YoderJA ZhangX ZhouL BestorTH andChengX (2001) Structure of human DNMT2 an enigmaticDNA methyltransferase homolog that displays denaturant-resistant binding to DNA Nucleic Acids Res 29 439ndash448
26 SchaeferM PollexT HannaK and LykoF (2009) RNAcytosine methylation analysis by bisulfite sequencing NucleicAcids Res 37 e12
27 BoninM OberstrassJ VogtU WasseneggerM and NellenW(2001) Binding of IRE-BP to its cognate RNA sequence SFMstudies on a universal RNA backbone for the analysis of RNA-protein interaction Biol Chem 382 1157ndash1162
28 ZimmermanW and WeijerCJ (1993) Analysis of cell cycleprogression during the development of Dictyostelium and itsrelationship to differentiation Dev Biol 160 178ndash185
29 StrasserK BloomfieldG MacWilliamsA CeccarelliAMacWilliamsH and TsangA (2012) A retinoblastomaorthologue is a major regulator of S-phase mitotic anddevelopmental gene expression in Dictyostelium PLoS One 7e39914
30 WattsDJ and AshworthJM (1970) Growth of myxameobae ofthe cellular slime mould Dictyostelium discoideum in axenicculture Biochem J 119 171ndash174
31 JurkowskiTP ShanmugamR HelmM and JeltschA (2012)Mapping the tRNA binding site on the surface of humanDNMT2 methyltransferase Biochemistry 51 4438ndash4444
32 SvedruzicZM and ReichNO (2005) DNA cytosine C5methyltransferase Dnmt1 catalysis-dependent release of allostericinhibition Biochemistry 44 9472ndash9485
33 ThomasSR KellerCA SzykA CannonJR and Laronde-LeblancNA (2011) Structural insight into the functionalmechanism of Nep1Emg1 N1-specific pseudouridinemethyltransferase in ribosome biogenesis Nucleic Acids Res 392445ndash2457
34 LeulliotN BohnsackMT GrailleM TollerveyD andVan TilbeurghH (2008) The yeast ribosome synthesisfactor Emg1 is a novel member of the superfamily of alphabeta knot fold methyltransferases Nucleic Acids Res 36629ndash639
35 WhiteJ LiZ SardanaR BujnickiJM MarcotteEM andJohnsonAW (2008) Bud23 methylates G1575 of 18S rRNA andis required for efficient nuclear export of pre-40S subunits MolCell Biol 28 3151ndash3161
36 SobalaA and HutvagnerG (2013) Small RNAs derived fromthe 50-end of tRNA can inhibit protein translation in humancells RNA Biol 10 525ndash535
Nucleic Acids Research 2013 13
at Universitaumlt W
uumlrzburg on August 5 2013
httpnaroxfordjournalsorgD
ownloaded from

tRNAAsp(GUC) we found tRNAGlu(UUC) andtRNAGlu(CUC) to match the pattern (Figure 2 andSupplementary Table S2)To test for tRNAGlu as a potential substrate we
modified the methylation assay by addition of a 50 ntDNA antisense oligonucleotide to tRNAAsp(GUC)
covering the methylation target site Figure 3A shows asa control that methylation of in vitro transcribedtRNAAsp(GUC) was completely blocked We then per-formed ex vivo methylation assays with enriched smallRNAs isolated from D discoideum after blockingtRNAAsp(GUC) with the antisense oligo Figure 3B showsthat the majority of 3H-incorporation disappearedHowever with both DnmA and hDnmt2 a significantamount of methylation was still detectable in the sizerange of tRNAs Assuming that this signal containedtRNAGlu we used a second antisense oligo directedagainst tRNAGlu(UUC) for blocking methylation Theoligo also covered the isoacceptor tRNAGlu(CUC) with
minor mismatches (Supplementary Figure S2) A furtherreduction of ex vivo methylation was observed Notablythe lower band in the tRNA size range disappeared (arrowin Figure 3B) suggesting that this represented a tRNAGlu
population that could be methylated at least in vitro Theresidual methylation after blocking with both oligos mayrepresent additional tRNAs or other unknown targets Aswe only concentrated on small RNAs larger methylatedmolecules and less abundant targets may have escapeddetection
To further analyse methylation on tRNAAsp andtRNAGlu (exemplified by tRNAGlu(UUC)) we carried outassays with wild-type sequences and C38A mutatedin vitro transcripts (Figure 4) We also included a suppres-sor tRNAGlu(CUA) that differed in only 2 nt fromtRNAGlu(UUC) (Figure 2) Assays were done in parallelwith the recombinant hDnmt2 and D discoideumDnmA The experiment showed that tRNAAsp(GUC) wasthe best substrate for both DnmA and hDnmt2 (Figure 4)Substantial methylation was also observed fortRNAGlu(UUC) and suppressor tRNAGlu(CUA) Moreoverthe complete loss of methylation in the C38A mutations oftRNAAsp(GUC) and tRNAGlu(UUC) confirmed that methy-lation occurred at position C38 and largely ruled out thatmethyl group incorporation was due to another
Figure 3 Ex vivo methylation and blocking assay (A) Methylation of in vitro transcribed tRNAAsp(GUC) was completely blocked when a comple-mentary antisense oligo was hybridized (B) Using enriched small RNA from dnmAKO cells ex vivo methylation in the tRNA size class wasdifferentially lost when antisense oligos to tRNAAsp(GUC) and tRNAGlu(UUC) were hybridized before the methylation reaction The oligo againsttRNAGlu(UUC) also covers tRNAGlu(CUC) with minor mismatches (see Supplementary Figure S2) Even with both oligos a significant amount of 3Hincorporation still remained The upper panel shows the ethidiumbromide stained gel to demonstrate equal loading the lower panel shows thefluorogram The arrow marks a band that was specifically lost when the anti tRNAGlu(UUC) oligo was used
Figure 2 Target site sequences of tRNAs used for in vitro methylationThe position of the putative signature nucleotides are indicated andprinted in bold if present The anticodon is underlined tRNAGlu(CUC)
(not shown) also harbours the nucleotide pattern Except for the anti-codon both tRNAGlu are identical in the anticodon stem-loop region(Supplementary Figure S2) Only a few differences appear in theacceptor stem
Figure 4 C38 is the target for in vitro methylation of tRNAAsp(GUC)
and tRNAGlu(UUC) In vitro methylation of tRNAAsp(GUC)tRNAGlu(UUC) and a suppressor tRNAGlu(CUA) C38A mutations con-firmed C38 as the target nucleotide tRNAAsp(GUC) methylation wasmost efficient
6 Nucleic Acids Research 2013
at Universitaumlt W
uumlrzburg on August 5 2013
httpnaroxfordjournalsorgD
ownloaded from
contaminating methyltransferase in the recombinantprotein preparation (17)
The methylation experiments using in vitro transcribedtRNAs showed that no other modifications were necessaryfor C38 methylation by DnmA and hDnmt2 (13) and thatthe processed ends of a mature tRNA were not requiredfor target recognition as the tRNA transcripts contain no30-CCA Even tRNAGlu embedded in a long dsRNAmolecule (1000 nt) with small stem-loop structures atthe ends (27) was a specific substrate of DnmA andhDnmt2 (Supplementary Figure S3)
By EMSA no difference in in vitro binding could beseen for the specific target tRNAGlu(UUC) and non-targetRNAs indicating that DnmA associated non-specificallywith any nucleic acid (data not shown) However specificcovalent enzyme-substrate intermediates can be detectedby denaturing SDSndashPAGE (25) Figure 5A andSupplementary Figure S4 show that radioactivelylabelled tRNA transcripts that can be methylatedin vitro by DnmA formed denaturation resistantcomplexes while non-target RNAs did not To confirmthat the denaturation resistant complexes were really dueto binding of the enzyme to the substrate SDS gels wereblotted and His-tagged DnmA was detected by a specificanti-His antibody A small amount of the tagged proteinwas shifted to the same position as the labelled tRNA(Supplementary Figure S5)
We then performed time courses for the reaction andfound that covalent complex formation was only transientunder the conditions used For tRNAAsp(GUC) adductsreached a maximum after 1min and were undetectableafter 15min whereas the tRNAGlu(UUC) complex onlybegan to accumulate after 1min but persisted for almost60min (Figure 5A and B) This coincided with the highmethylation activity on tRNAAsp(GUC-1) and a lower oneon tRNAGlu(UUC-2) The human enzyme appeared to beslightly faster in the reaction but otherwise showed asimilar behaviour compared with D discoideum DnmA
tRNAAsp(GUC) tRNAGlu(UUCCUC) and tRNAGly(GCC)
bind specifically to DnmA in vivo
The blocking assay (Figure 3) suggested that further sub-strates could be targeted by DnmA To identify suchtargets and to further investigate the interaction betweenDnmA and RNAs in vivo we established a UV-CLIP pro-cedure for D discoideumDnmA-GFP expressing cells andcontrol cells expressing GFP only were UV cross-linkedand complexes were precipitated under denaturing condi-tions by a GFP nanobody (21) Then co-precipitatedRNAs were isolated converted into strand-specificcDNA libraries and analyzed by Illumina sequencing(Figure 6 and Supplmentary Figure S6)
Various tRNA fragments were enriched in the DnmACLIP sample compared with the control By their lengthand position fragments appeared not to be random butspecific We found mid-fragments of 20ndash24 nt (position32ndash36 to 53ndash56) that covered the C38 methylation site inthe respective tRNAs 50-fragments of 17 nt (nucleotideposition 1ndash17) 30-fragments of 2021 nt (nucleotideposition 54 to the end) and 30-half tRNAs of 41ndash43 nt
(position 30ndash32 to the end) Mid-fragments oftRNAAsp(GUC) tRNAGlu(UUCCUC) and tRNAGly(GCC)
were significantly enriched in the DnmA-GFP fraction incomparison with the control CLIP assay tRNAAsp(GUC)tRNAGlu(UUC) and tRNAGlu(CUC) harbour the sequencepattern (C32 C34 A37 C40) which has been suggestedas a signature for C38 methylation (Figure 2 Helmpersonal communication) tRNAGly(GCC) contains thepattern except for C40 For tRNAAla(AGC) which doesnot contain C38 and the signature pattern a differentfragment from the 30-end was enriched in the DnmA-CLIP sample For other tRNAs no significant accumula-tion was observed in the experiment (Figure 6 andSupplementary Figure S6) For several tRNAs varyingamounts of full-length molecules were found but thesewere never enriched in the DnmA sample in comparisonwith the control
tRNAGly(GCC) is an additional methylation substratein vitro
The CLIP data suggested that tRNAGly(GCC) was anothertarget for DnmA We therefore cloned this and several othertRNA genes that contained the pattern only partially(Figure 2) We generated in vitro transcripts and examinedmethylation by DnmA and hDnmt2 in vitro IndeedtRNAGly(GCC) was methylated by both enzymes (Figure 7)while none of the other tRNAs (tRNALeu(UAG-1)tRNALeu(AAG-8) tRNAVal(CAC-1) and tRNAPhe(GAA-2))resulted in detectable methylation signals in the assay(Supplementary Figure S7) Interestingly the in vitro methy-lation efficiency of the three tRNA substrates
Figure 5 Formation and turnover of tRNA-methyltransferasecomplexes Turnover of covalent complexes of tRNA substrates withDnmA and hDnmt2 (A) Examples of time courses on covalentcomplex formation with tRNAAsp(GUC) and tRNAGlu(UUC) (B)Complex bands in A were quantified and presented on a time scale
Nucleic Acids Research 2013 7
at Universitaumlt W
uumlrzburg on August 5 2013
httpnaroxfordjournalsorgD
ownloaded from
(tRNAAsp(GUC) tRNAGlu(UUC) tRNAGly(GCC)) correlatedroughly with the number of tRNA fragments found in theRNA-CLIP experimentThe data supported the conclusion from the ex vivo
blocking assay (Figure 3B) that in addition toD discoideum tRNAAsp and tRNAGlu other RNAs weresubstrates for DnmA and hDnmt2 in vitro
Expression of dnmA is regulated during developmentcell cycle and on temperature stress
DnmA expression in growing vegetative D discoideumcells is low To examine tRNA methylation in vivo it
was therefore advantageous to determine conditionsunder which dnmA was most highly expressed Promptedby previous studies (141517) we assumed that the biolo-gical function of DnmA was connected to developmentalandor environmental conditions We therefore examinedexpression levels of dnmA under different regimes dnmAmRNA was quantified by qPCR over the 24 h develop-mental cycle of D discoideum (data not shown) and con-firmed the data deposited in Dictyexpress (httpdictyexpressbiolabsi)
Interestingly an 5-fold increase in expression wasobserved at 16 h in development This time coincides
Figure 6 tRNA and tRNA fragments associated with DnmA-GFP After UV crosslinking RNA bound to DnmA was co-immunoprecipitatedunder denaturing conditions (CLIP) The number of normalised reads for tRNAs detected by Illumina sequencing is shown for DnmA CLIP and forthe control CLIP with GFP In addition to full-length tRNAs four classes of tRNA fragments were found and these are indicated by differentshading of the bars Size and localization of fragments is shown schematically on the simplified clover leaf structure tRNAs that are not significantlyenriched with DnmA are shown in Supplementary Figure S6 In case of multiple gene copys resulting in the same RNA transcript the sequencingreads were mapped to the first copy in the genome starting from chromsome 1 Only this copy is listed Sequences and detailed information on genecopies and potential isoacceptors are listed in Supplementary Table S2
8 Nucleic Acids Research 2013
at Universitaumlt W
uumlrzburg on August 5 2013
httpnaroxfordjournalsorgD
ownloaded from
with a presumed synchronous mitotic division (28) wherea multitude of cell-cycle regulated genes are expressed[(29) and McWilliams personal Communication]
Therefore we synchronized vegetative D discoideumcells to examine dnmA expression during the cell cycleFor these experiments the D discoideum wild-typestrain NC4 the parent of the axenic laboratory strainAX2 was used (30) This was prompted by the observationthat D melanogaster and E histolytica laboratory strainsmay lose dnmt2 expression (Reuter Ankri personalcommunication)
The data confirmed our assumption of a cell-cycle-de-pendent transcriptional regulation of dnmA (Figure 8A)The peak of expression coincided with late mitosis or earlyS-phase as D discoideum does not display a G1 phaseUsing AX2 cells cotransformed with PCNA-RFP andDnmA-GFP we observed GFP accumulation in thenucleus during S phase (indicated by PCNA expression)and a loss of DnmA from the nucleus during mitosis(Figure 8B) This is further exemplified by the movie inthe Supplementary Material (Supplementary Movie S1)
In other organisms dnmt2 was observed to beupregulated during stress response (315) We thereforeexamined expression after anoxia heat shock and coldshock but did not find any significant changes (data notshown) Only during recovery from cold shock wedetected a strong transient increase in dnmA mRNA by47-fold (Figure 8C)
Thus dnmA expression was differentially regulatedduring the cell cycle development and on temperaturestress Furthermore DnmA localization changed specific-ally during the cell cycle
Though the basic expression in vegetative cells was in-distinguishable dnmA expression in NC4 was 30-foldhigher at 16 h of development whereas developmentalupregulation was only 5-fold in AX2 The data aresummarized in Figure 8D
NC4 the parent strain of AX2 displays increasedtRNA
Asp methylation during development
Based on these observations we asked the questionwhether expression levels correlated with tRNA
methylation activity in vivo Bisulfite sequencing wasdone with primers for tRNAAsp tRNAGlu and tRNAGly
on RNA isolated from vegetative AX2 cells AX2 cellsdeveloped for 16 h AX2 cells subjected to cold shockAX2 recovered for 25 h after cold shock vegetativeNC4 cells and NC4 cells developed for 16 h The resultson C38 methylation of tRNAAsp(GUC) are summarized inFigure 8E The methylation status of all C residues fromall tested tRNAs in this study is shown in SupplementaryFigure S8 Indeed tRNAAsp(GUC) methylation at C38increased to 76 in development in NC4 cells Noincrease was observed in AX2 cells during developmentpossibly due to insufficient sensitivity of the assayInterestingly no increase in tRNAAsp(GUC) methylationwas observed at the highest dnmA expression levelsduring stress recovery In Figure 8E the 30ndash35 methy-lation level in all tRNAAsp(GUC) samples represents signifi-cant methylation as compared with other C residues (seeSupplementary Figure S8)We did however not detect any significant methylation
of tRNAGlu(UUCCUC) and tRNAGly(GCC) under any con-ditions (Supplementary Figure S8) In contrast C49C50methylation which does not depend on Dnmt2 enzymeswas found in all samples and thus served as an internalcontrol In all 1175 bisulfite-sequence reads wereexamined for tRNAGlu(UUCCUC) and 4040 fortRNAGly(GCC) No methylation above background wasfound Though it cannot be excluded that the failure todetect C38 methylation of tRNAGlu and tRNAGly was dueto inefficient reverse transcription or low levels of methy-lation (see lsquoDiscussionrsquo section) we conclude thattRNAAsp is the major or possibly the only in vivo targetfor methylation by DnmA in D discoideum
DISCUSSION
The biological function of Dnmt2 methyltransferases islargely unknown and its role in DNA methylation is con-troversially discussed (910) In contrast tRNA methyla-tion by Dnmt2 is robust in vitro and in vivo andevolutionary conserved In addition to the originallydescribed tRNAAsp(GUC) (1) two further tRNAs targets(tRNAVal(CAC) and tRNAGly(GCC)) have been defined inD melanogaster (15) and tRNAGlu(UUC) in S pombe (17)To get insight into potential functions of the
D discoideum Dnmt2 homologue DnmA we examinedexpression levels in development in the cell cycle andunder different environmental conditions We confirmedby qPCR an expression peak at 16 h of developmentWeijer and co-workers (28) had previously proposed asynchronous cell division at this developmental stageWe therefore investigated dnmA expression in the cellcycle and found a strong transient increase of mRNA inS-phase or shortly thereafter The expression peak wasaccompanied by reaccumulation of DnmA in thenucleus We assume that DnmA is degraded duringmitosis and re-synthesised after cell divisionIn S pombe as well as in D melanogaster tRNA methy-
lation changes in response to environmental challengeslike temperature stress anoxia or nutrients We tested
Figure 7 In vitro methylation of tRNAGly(GCC) by recombinant DnmAand hDnmt2 In vitro transcribed tRNAs were methylated in vitro asdescribed before The top panel shows the ethidium-bromide stainedgel the bottom panel the fluorogram of 3H-labelled methylated tRNAsAs a control tRNAAsp(GUC) methylation is shown
Nucleic Acids Research 2013 9
at Universitaumlt W
uumlrzburg on August 5 2013
httpnaroxfordjournalsorgD
ownloaded from
Figure 8 Expression localization and tRNAAsp(GUC) methylation activity of DnmA under various conditions (A) dnmA expression is regulatedduring the cell cycle Cells were arrested in the cell cycle by cold treatment and then released by transfer to 22C Cells were counted every 30min(grey dots and grey line) and samples were taken for qPCR (black bars) After cell division dnmA expression increased 5-fold and then rapidlydeclined to basal levels Normalization was done on vegetative growing cells dnmA expression is only shown from 3 to 45 h during recovery(B) DnmA is lost from the nucleus during mitosis (arrow) The three cells on the right are in S-phase as indicated by the RFP-PCNA marker andaccumulate DnmA in the nucleus (for further details see movie in Supplementary Material) (C) Relative quantification of dnmA expression levels inAX2 cells after 25 h recovery from cold shock at 4C Expression of dnmA increased gt40-fold and returned within 30min to basal levels (n=3)(D) At 16 h of development dnmA expression increased 46-fold in the NC4 strain In AX2 cells expression increased only 5-fold (E) At 16 h ofdevelopment in vivo methylation of C38 in tRNAAsp(GUC) increased up to 75 in the D discoideum strain NC4 whereas no significant increase wasobserved in AX2 cells
10 Nucleic Acids Research 2013
at Universitaumlt W
uumlrzburg on August 5 2013
httpnaroxfordjournalsorgD
ownloaded from
heat shock cold shock anoxia and various nutrient mediabut did not observe altered levels Only during therecovery phase after cold treatment a transient almost50-fold increase in mRNA abundance was seen
Reuter and Ankri (personal communication) hadobserved a loss of Dnmt2 expression in laboratorystrains of D melanogaster and E histolytica We thereforeexamined D discoideum NC4 the parent of the axeniclaboratory strain AX2 The developmental regulation inAX2 and NC4 was qualitatively similar but expressionlevels were much higher in NC4 The observationsupports the assumption that Dnmt2 expression wasreduced under optimal growth conditions in the labora-tory strain and may actually be selected against
We show that the D discoideum Dnmt2 homologueDnmA is an active tRNAAsp(GUC) methyltransferasein vitro and in vivo To identify further potential targetsfor DnmA we established a UV-CLIP method to analyseDnmAndashRNA interaction in vivo In addition to the knowntarget tRNAAsp(GUC) fragments of tRNAGlu(UUC) andtRNAGly(GCC) were most abundant in comparison withthe control Unexpectedly complete tRNAs were notenriched in the DnmA CLIP fraction but rather specificlsquomid-fragmentsrsquo that covered the putative methylation siteand the recognition pattern proposed by Helm (personal
communication) A minor exception was tRNAGly(GCC)which lacks C40 We do not know whether these frag-ments were directly or indirectly generated by DnmAin vivo or whether they were protected by DnmA fromdegradation during the experimental procedure As noco-crystal of Dnmt2 and a tRNA substrate is availablethe exact interface for the interaction is unknownJurkowski et al (31) mapped the binding site on Dnmt2by mutational analysis A manual placement of a tRNAinto the crystal structure suggests that Dnmt2 mainlycontacts the anticodon stem loop Assuming that mid-frag-ments are footprints of DnmA our data support this notionbut suggest that the interaction surface extends into theD-loop which is partially covered by the mid-fragments
We examined the putative DnmA targets in vitro (bymethylation of in vitro transcribed tRNAs) ex vivo(by methylation of RNA isolated from D discoideum)and in vivo (by bisulfite sequencing) Furthermore weinvestigated by SDSndashPAGE the kinetics of covalentcomplex formation between tRNAAsp and tRNAGlu withDnmA and hDnmt2
All three tRNAs (tRNAAsp tRNAGlu tRNAGly) werereadily methylated by recombinant DnmA and hDnmt2in vitro although the activity was much higher (estimatedat least 10-fold) on tRNAAsp compared with tRNAGlu andtRNAGly For tRNAAsp and tRNAGlu this correlated wellwith the formation and decay of covalent RNAmdashproteincomplexes where kinetics for tRNAAsp were much fasterthan for tRNAGlu Instead of C methylation the covalentintermediate may also proceed to a hydrogen exchangereaction or to a deamination of the cytosine In bothcases an intermediate will be detected but it will notresult in a methylated product (32) It may be thatslower off rates represent these alternative pathways thatwould result in unmethylated tRNAs
Using antisense oligos we showed that tRNAAsptRNAGlu and at least one additional RNA could bemethylated ex vivo A quantification of the results wasnot possible the apparently low methylation oftRNAGlu ex vivo could be due to the low activity of theenzyme on this substrate (as suggested by the in vitroexperiments see Figure 5B)The data for the examination of in vivo methylation by
bisulfite sequencing were rather unexpected AlthoughtRNAAsp(GUC) was consistently found to be methylatedat C38 we never detected any methylation at C38 oftRNAGlu(UUCCUC) or tRNAGly(GCC) There could betwo reasons to explain the failure to detect C38 methyla-tion for these in vitro substrates (i) due to the low activityof DnmA on tRNAGlu(UUCCUC) and tRNAGly(GCC) asshown in vitro there was low or no significant methylationin vivo and (ii) methylation escaped detection becauseother unknown modifications blocked reverse transcrip-tion of the methylated molecules However if this wasthe case one has to assume that C49C50 methylationoccurs independently of these unknown modificationswhereas C38 methylation strictly depends on them AsC38 methylation does not require any other modificationsin vitro we believe that this scenario is less likely If in vitromethylation activity was comparable with in vivo methy-lation 10 methylation of tRNAGlu(UUCCUC) andtRNAGly(GCC) could be expected relative totRNAAsp(GUC) As tRNAAsp was methylated to 30 a3 methylation of tRNAGlu and tRNAGly could be belowthe detection level of the bisulfite assayWe find that high expression of dnmA did not necessar-
ily correlate with increased methylation of tRNAs Duringrecovery from cold shock D discoideum cells displayedthe highest level of dnmA expression but this had noeffect on tRNAAsp methylation Only in developmentand with the high expressing NC4 strain a significantincrease of tRNAAsp methylation to 70 was observedIt has to be considered however that bisulfite sequencingdoes not necessarily allow for quantitative evaluation ofdata detected C-methylation depends on reverse tran-scription efficiency which may be hampered by othermodifications that could be different under differentconditions Furthermore the completeness of bisulfite-mediated conversion could also depend on RNA struc-ture which may be altered by other modificationsHowever incomplete conversion will lead to an overesti-mation of C-methylation as maybe the case for forexample C30 C31 C35 in tRNAGly in some samplesTaken together the data from CLIP complex forma-
tion and methylation show that all three substrates can bebound and methylated in vitro The CLIP data show thatthere is a specific interaction of DnmA with tRNA mid-fragments of tRNAAsp(GUC) tRNAGlu(UUCCUC) andtRNAGly(GCC) but not with other tRNAs In contrastbisulfite sequencing could only detect methylation ontRNAAsp(GUC) even at the highest expression levels ofdnmA Even though low amounts of methylation mayhave escaped detection in bisulfite sequencing there isstill a discrepancy between dnmA expression levels (egduring cold shock recovery) DnmA binding to substrates(as determined by CLIP) and methylation in vivo
Nucleic Acids Research 2013 11
at Universitaumlt W
uumlrzburg on August 5 2013
httpnaroxfordjournalsorgD
ownloaded from
We therefore suggest that DnmA binding to specifictargets could elicit functions in addition to RNA methy-lation eg by promoting processing or degradation andor recruitment of other proteins At least for some rRNAmethyltransferases such as Bud23 or Nep1 fromSaccharomyces cerevisiae it is known that they executefunctions independent from the methylation activity(33ndash35) As shown by Schaefer and co-workers (15)methylation protects tRNAs from degradation or process-ing after stress By a different pathway binding of DnmAmay initiate processing that leads to specific tRNA frag-ments that could serve for other biological functions (36)Interestingly tRNAGlu(UUC) is also a substrate for theDnmt2 homologue Pmt1 in S pombe (17) HowevertRNAGlu(UUC) is found to be methylated to a low butsignificant extend in vivo in fission yeast when Pmt1 isoverexpressedThe detection of tRNAAla(AGC) fragments in the CLIP
assay brings up a new question tRNAAla(AGC) has a Tinstead of a C in position 38 and does not contain thesignature nucleotides in the anticodon stem-loopInstead of a mid-fragment a distinct 30 fragment wasassociated with DnmA suggesting that a recognitionmotif was located in this part of the moleculeAs methylation also occurred in a large artificial
molecule with embedded tRNAGlu (SupplementaryFigure S2) it could be that other longer RNAs containingan appropriate recognition motif may also be bound bythe enzyme We thus propose that the biological functionsof Dnmt2 proteins could extend far beyond our presentknowledge Investigation of further targets and of the sig-nificance of RNA fragments associated with Dnmt2should shed light on the importance of this enzyme
SUPPLEMENTARY DATA
Supplementary Data are available at NAR Online
ACKNOWLEDGEMENTS
The authors thank Thomas Winckler (Friedrich-Schiller-University Jena) for providing the tRNAGlu(UUC) and sup-pressor tRNAGlu(CUA) plasmids Susanne Junk for cloningtRNAAspC38A Matthias Schafer (DKFZ) and RobinLorenz (University of Kassel) are acknowledged forRNA bisulfite sequencing and Richard Reinhardt (MaxPlanck Genome Centre Cologne) for deep sequencingMark Helm (Johannes-Gutenberg-University Mainz) gen-erously provided data before publication CarstenSeehafer (University of Kassel) helped with the analysisof deep-sequencing data
FUNDING
Deutsche Forschungsgemeinschaft [FOR 1082 to WNand AJ] and the Studienstiftung des Deutschen Volkes(to SM) Funding for open access charge FOR1082
Conflict of interest statement None declared
REFERENCES
1 GollMG KirpekarF MaggertKA YoderJA HsiehCLZhangX GolicKG JacobsenSE and BestorTH (2006)Methylation of tRNAAsp by the DNA methyltransferasehomolog Dnmt2 Science 311 395ndash398
2 PhalkeS NickelO WalluscheckD HortigF OnoratiMCand ReuterG (2009) Retrotransposon silencing and telomereintegrity in somatic cells of Drosophila depends on the cytosine-5methyltransferase DNMT2 Nat Genet 41 696ndash702
3 WilkinsonCR BartlettR NurseP and BirdAP (1995) Thefission yeast gene pmt1+ encodes a DNA methyltransferasehomologue Nucleic Acids Res 23 203ndash210
4 KatohM CurkT XuQ ZupanB KuspaA and ShaulskyG(2006) Developmentally regulated DNA methylation inDictyostelium discoideum Eukaryot Cell 5 18ndash25
5 KuhlmannM BorisovaBE KallerM LarssonP StachDNaJ EichingerL LykoF AmbrosV SoderbomF et al(2005) Silencing of retrotransposons in Dictyostelium by DNAmethylation and RNAi Nucleic Acids Res 33 6405ndash6417
6 RaiK ChidesterS ZavalaCV ManosEJ JamesSRKarpfAR JonesDA and CairnsBR (2007) Dnmt2 functionsin the cytoplasm to promote liver brain and retina developmentin zebrafish Genes Dev 21 261ndash266
7 FisherO Siman-TovR and AnkriS (2006) Pleiotropicphenotype in Entamoeba histolytica overexpressing DNAmethyltransferase (Ehmeth) Mol Biochem Parasitol 147 48ndash54
8 DurdevicZ HannaK GoldB PollexT CherryS LykoFand SchaeferM (2013) Efficient RNA virus control in Drosophilarequires the RNA methyltransferase Dnmt2 EMBO Rep 14269ndash275
9 SchaeferM and LykoF (2010) Lack of evidence for DNAmethylation of Invader4 retroelements in Drosophila andimplications for Dnmt2-mediated epigenetic regulation NatGenet 42 920ndash921 author reply 921
10 KraussV and ReuterG (2011) DNA methylation in Drosophila -a critical evaluation Prog Mol Biol Transl Sci 101 177ndash191
11 RaddatzG GuzzardoPM OlovaN FantappieMRRamppM SchaeferM ReikW HannonGJ and LykoF(2013) Dnmt2-dependent methylomes lack defined DNAmethylation patterns Proc Natl Acad Sci USA 110 8627ndash8631
12 JeltschA NellenW and LykoF (2006) Two substrates arebetter than one dual specificities for Dnmt2 methyltransferasesTrends Biochem Sci 31 306ndash308
13 JurkowskiTP MeusburgerM PhalkeS HelmM NellenWReuterG and JeltschA (2008) Human DNMT2 methylatestRNA(Asp) molecules using a DNA methyltransferase-likecatalytic mechanism RNA 14 1663ndash1670
14 TovyA Siman TovR GaentzschR HelmM and AnkriS(2010) A new nuclear function of the Entamoeba histolyticaglycolytic enzyme enolase the metabolic regulation of cytosine-5methyltransferase 2 (Dnmt2) activity PLoS Pathog 6 e1000775
15 SchaeferM PollexT HannaK TuortoF MeusburgerMHelmM and LykoF (2010) RNA methylation by Dnmt2protects transfer RNAs against stress-induced cleavage GenesDev 24 1590ndash1595
16 TuortoF LiebersR MuschT SchaeferM HofmannSKellnerS FryeM HelmM StoecklinG and LykoF (2012)RNA cytosine methylation by Dnmt2 and NSun2 promotestRNA stability and protein synthesis Nat Struct Mol Biol 19900ndash905
17 BeckerM MullerS NellenW JurkowskiTP JeltschA andEhrenhofer-MurrayAE (2012) Pmt1 a Dnmt2 homolog inSchizosaccharomyces pombe mediates tRNA methylation inresponse to nutrient signaling Nucleic Acids Res 4011648ndash11658
18 TovyA HertzR Siman-TovR SyanS FaustD GuillenNand AnkriS (2011) Glucose starvation boosts Entamoebahistolytica virulence PLoS Negl Trop Dis 5 e1247
19 MotorinY and HelmM (2011) RNA nucleotide methylationWiley Interdiscip Rev RNA 2 611ndash631
20 NiranjanakumariS LasdaE BrazasR and Garcia-BlancoMA(2002) Reversible cross-linking combined with
12 Nucleic Acids Research 2013
at Universitaumlt W
uumlrzburg on August 5 2013
httpnaroxfordjournalsorgD
ownloaded from
immunoprecipitation to study RNA-protein interactions in vivoMethods 26 182ndash190
21 RothbauerU ZolghadrK MuyldermansS SchepersACardosoMC and LeonhardtH (2008) A versatile nanotrapfor biochemical and functional studies with fluorescent fusionproteins Mol Cell Proteomics 7 282ndash289
22 SittkaA LucchiniS PapenfortK SharmaCM RolleKBinnewiesTT HintonJC and VogelJ (2008) Deep sequencinganalysis of small noncoding RNA and mRNA targets of theglobal post-transcriptional regulator Hfq PLoS Genet 4e1000163
23 HoffmannS OttoC KurtzS SharmaCM KhaitovichPVogelJ StadlerPF and HackermullerJ (2009) Fast mapping ofshort sequences with mismatches insertions and deletions usingindex structures PLoS Comput Biol 5 e1000502
24 ConnollyMA ClausenPA and LazarJG (2006) Purificationof RNA from animal cells using trizol Cold Spring HarborProtoc 2006 pii pdbprot4104
25 DongA YoderJA ZhangX ZhouL BestorTH andChengX (2001) Structure of human DNMT2 an enigmaticDNA methyltransferase homolog that displays denaturant-resistant binding to DNA Nucleic Acids Res 29 439ndash448
26 SchaeferM PollexT HannaK and LykoF (2009) RNAcytosine methylation analysis by bisulfite sequencing NucleicAcids Res 37 e12
27 BoninM OberstrassJ VogtU WasseneggerM and NellenW(2001) Binding of IRE-BP to its cognate RNA sequence SFMstudies on a universal RNA backbone for the analysis of RNA-protein interaction Biol Chem 382 1157ndash1162
28 ZimmermanW and WeijerCJ (1993) Analysis of cell cycleprogression during the development of Dictyostelium and itsrelationship to differentiation Dev Biol 160 178ndash185
29 StrasserK BloomfieldG MacWilliamsA CeccarelliAMacWilliamsH and TsangA (2012) A retinoblastomaorthologue is a major regulator of S-phase mitotic anddevelopmental gene expression in Dictyostelium PLoS One 7e39914
30 WattsDJ and AshworthJM (1970) Growth of myxameobae ofthe cellular slime mould Dictyostelium discoideum in axenicculture Biochem J 119 171ndash174
31 JurkowskiTP ShanmugamR HelmM and JeltschA (2012)Mapping the tRNA binding site on the surface of humanDNMT2 methyltransferase Biochemistry 51 4438ndash4444
32 SvedruzicZM and ReichNO (2005) DNA cytosine C5methyltransferase Dnmt1 catalysis-dependent release of allostericinhibition Biochemistry 44 9472ndash9485
33 ThomasSR KellerCA SzykA CannonJR and Laronde-LeblancNA (2011) Structural insight into the functionalmechanism of Nep1Emg1 N1-specific pseudouridinemethyltransferase in ribosome biogenesis Nucleic Acids Res 392445ndash2457
34 LeulliotN BohnsackMT GrailleM TollerveyD andVan TilbeurghH (2008) The yeast ribosome synthesisfactor Emg1 is a novel member of the superfamily of alphabeta knot fold methyltransferases Nucleic Acids Res 36629ndash639
35 WhiteJ LiZ SardanaR BujnickiJM MarcotteEM andJohnsonAW (2008) Bud23 methylates G1575 of 18S rRNA andis required for efficient nuclear export of pre-40S subunits MolCell Biol 28 3151ndash3161
36 SobalaA and HutvagnerG (2013) Small RNAs derived fromthe 50-end of tRNA can inhibit protein translation in humancells RNA Biol 10 525ndash535
Nucleic Acids Research 2013 13
at Universitaumlt W
uumlrzburg on August 5 2013
httpnaroxfordjournalsorgD
ownloaded from

contaminating methyltransferase in the recombinantprotein preparation (17)
The methylation experiments using in vitro transcribedtRNAs showed that no other modifications were necessaryfor C38 methylation by DnmA and hDnmt2 (13) and thatthe processed ends of a mature tRNA were not requiredfor target recognition as the tRNA transcripts contain no30-CCA Even tRNAGlu embedded in a long dsRNAmolecule (1000 nt) with small stem-loop structures atthe ends (27) was a specific substrate of DnmA andhDnmt2 (Supplementary Figure S3)
By EMSA no difference in in vitro binding could beseen for the specific target tRNAGlu(UUC) and non-targetRNAs indicating that DnmA associated non-specificallywith any nucleic acid (data not shown) However specificcovalent enzyme-substrate intermediates can be detectedby denaturing SDSndashPAGE (25) Figure 5A andSupplementary Figure S4 show that radioactivelylabelled tRNA transcripts that can be methylatedin vitro by DnmA formed denaturation resistantcomplexes while non-target RNAs did not To confirmthat the denaturation resistant complexes were really dueto binding of the enzyme to the substrate SDS gels wereblotted and His-tagged DnmA was detected by a specificanti-His antibody A small amount of the tagged proteinwas shifted to the same position as the labelled tRNA(Supplementary Figure S5)
We then performed time courses for the reaction andfound that covalent complex formation was only transientunder the conditions used For tRNAAsp(GUC) adductsreached a maximum after 1min and were undetectableafter 15min whereas the tRNAGlu(UUC) complex onlybegan to accumulate after 1min but persisted for almost60min (Figure 5A and B) This coincided with the highmethylation activity on tRNAAsp(GUC-1) and a lower oneon tRNAGlu(UUC-2) The human enzyme appeared to beslightly faster in the reaction but otherwise showed asimilar behaviour compared with D discoideum DnmA
tRNAAsp(GUC) tRNAGlu(UUCCUC) and tRNAGly(GCC)
bind specifically to DnmA in vivo
The blocking assay (Figure 3) suggested that further sub-strates could be targeted by DnmA To identify suchtargets and to further investigate the interaction betweenDnmA and RNAs in vivo we established a UV-CLIP pro-cedure for D discoideumDnmA-GFP expressing cells andcontrol cells expressing GFP only were UV cross-linkedand complexes were precipitated under denaturing condi-tions by a GFP nanobody (21) Then co-precipitatedRNAs were isolated converted into strand-specificcDNA libraries and analyzed by Illumina sequencing(Figure 6 and Supplmentary Figure S6)
Various tRNA fragments were enriched in the DnmACLIP sample compared with the control By their lengthand position fragments appeared not to be random butspecific We found mid-fragments of 20ndash24 nt (position32ndash36 to 53ndash56) that covered the C38 methylation site inthe respective tRNAs 50-fragments of 17 nt (nucleotideposition 1ndash17) 30-fragments of 2021 nt (nucleotideposition 54 to the end) and 30-half tRNAs of 41ndash43 nt
(position 30ndash32 to the end) Mid-fragments oftRNAAsp(GUC) tRNAGlu(UUCCUC) and tRNAGly(GCC)
were significantly enriched in the DnmA-GFP fraction incomparison with the control CLIP assay tRNAAsp(GUC)tRNAGlu(UUC) and tRNAGlu(CUC) harbour the sequencepattern (C32 C34 A37 C40) which has been suggestedas a signature for C38 methylation (Figure 2 Helmpersonal communication) tRNAGly(GCC) contains thepattern except for C40 For tRNAAla(AGC) which doesnot contain C38 and the signature pattern a differentfragment from the 30-end was enriched in the DnmA-CLIP sample For other tRNAs no significant accumula-tion was observed in the experiment (Figure 6 andSupplementary Figure S6) For several tRNAs varyingamounts of full-length molecules were found but thesewere never enriched in the DnmA sample in comparisonwith the control
tRNAGly(GCC) is an additional methylation substratein vitro
The CLIP data suggested that tRNAGly(GCC) was anothertarget for DnmA We therefore cloned this and several othertRNA genes that contained the pattern only partially(Figure 2) We generated in vitro transcripts and examinedmethylation by DnmA and hDnmt2 in vitro IndeedtRNAGly(GCC) was methylated by both enzymes (Figure 7)while none of the other tRNAs (tRNALeu(UAG-1)tRNALeu(AAG-8) tRNAVal(CAC-1) and tRNAPhe(GAA-2))resulted in detectable methylation signals in the assay(Supplementary Figure S7) Interestingly the in vitro methy-lation efficiency of the three tRNA substrates
Figure 5 Formation and turnover of tRNA-methyltransferasecomplexes Turnover of covalent complexes of tRNA substrates withDnmA and hDnmt2 (A) Examples of time courses on covalentcomplex formation with tRNAAsp(GUC) and tRNAGlu(UUC) (B)Complex bands in A were quantified and presented on a time scale
Nucleic Acids Research 2013 7
at Universitaumlt W
uumlrzburg on August 5 2013
httpnaroxfordjournalsorgD
ownloaded from
(tRNAAsp(GUC) tRNAGlu(UUC) tRNAGly(GCC)) correlatedroughly with the number of tRNA fragments found in theRNA-CLIP experimentThe data supported the conclusion from the ex vivo
blocking assay (Figure 3B) that in addition toD discoideum tRNAAsp and tRNAGlu other RNAs weresubstrates for DnmA and hDnmt2 in vitro
Expression of dnmA is regulated during developmentcell cycle and on temperature stress
DnmA expression in growing vegetative D discoideumcells is low To examine tRNA methylation in vivo it
was therefore advantageous to determine conditionsunder which dnmA was most highly expressed Promptedby previous studies (141517) we assumed that the biolo-gical function of DnmA was connected to developmentalandor environmental conditions We therefore examinedexpression levels of dnmA under different regimes dnmAmRNA was quantified by qPCR over the 24 h develop-mental cycle of D discoideum (data not shown) and con-firmed the data deposited in Dictyexpress (httpdictyexpressbiolabsi)
Interestingly an 5-fold increase in expression wasobserved at 16 h in development This time coincides
Figure 6 tRNA and tRNA fragments associated with DnmA-GFP After UV crosslinking RNA bound to DnmA was co-immunoprecipitatedunder denaturing conditions (CLIP) The number of normalised reads for tRNAs detected by Illumina sequencing is shown for DnmA CLIP and forthe control CLIP with GFP In addition to full-length tRNAs four classes of tRNA fragments were found and these are indicated by differentshading of the bars Size and localization of fragments is shown schematically on the simplified clover leaf structure tRNAs that are not significantlyenriched with DnmA are shown in Supplementary Figure S6 In case of multiple gene copys resulting in the same RNA transcript the sequencingreads were mapped to the first copy in the genome starting from chromsome 1 Only this copy is listed Sequences and detailed information on genecopies and potential isoacceptors are listed in Supplementary Table S2
8 Nucleic Acids Research 2013
at Universitaumlt W
uumlrzburg on August 5 2013
httpnaroxfordjournalsorgD
ownloaded from
with a presumed synchronous mitotic division (28) wherea multitude of cell-cycle regulated genes are expressed[(29) and McWilliams personal Communication]
Therefore we synchronized vegetative D discoideumcells to examine dnmA expression during the cell cycleFor these experiments the D discoideum wild-typestrain NC4 the parent of the axenic laboratory strainAX2 was used (30) This was prompted by the observationthat D melanogaster and E histolytica laboratory strainsmay lose dnmt2 expression (Reuter Ankri personalcommunication)
The data confirmed our assumption of a cell-cycle-de-pendent transcriptional regulation of dnmA (Figure 8A)The peak of expression coincided with late mitosis or earlyS-phase as D discoideum does not display a G1 phaseUsing AX2 cells cotransformed with PCNA-RFP andDnmA-GFP we observed GFP accumulation in thenucleus during S phase (indicated by PCNA expression)and a loss of DnmA from the nucleus during mitosis(Figure 8B) This is further exemplified by the movie inthe Supplementary Material (Supplementary Movie S1)
In other organisms dnmt2 was observed to beupregulated during stress response (315) We thereforeexamined expression after anoxia heat shock and coldshock but did not find any significant changes (data notshown) Only during recovery from cold shock wedetected a strong transient increase in dnmA mRNA by47-fold (Figure 8C)
Thus dnmA expression was differentially regulatedduring the cell cycle development and on temperaturestress Furthermore DnmA localization changed specific-ally during the cell cycle
Though the basic expression in vegetative cells was in-distinguishable dnmA expression in NC4 was 30-foldhigher at 16 h of development whereas developmentalupregulation was only 5-fold in AX2 The data aresummarized in Figure 8D
NC4 the parent strain of AX2 displays increasedtRNA
Asp methylation during development
Based on these observations we asked the questionwhether expression levels correlated with tRNA
methylation activity in vivo Bisulfite sequencing wasdone with primers for tRNAAsp tRNAGlu and tRNAGly
on RNA isolated from vegetative AX2 cells AX2 cellsdeveloped for 16 h AX2 cells subjected to cold shockAX2 recovered for 25 h after cold shock vegetativeNC4 cells and NC4 cells developed for 16 h The resultson C38 methylation of tRNAAsp(GUC) are summarized inFigure 8E The methylation status of all C residues fromall tested tRNAs in this study is shown in SupplementaryFigure S8 Indeed tRNAAsp(GUC) methylation at C38increased to 76 in development in NC4 cells Noincrease was observed in AX2 cells during developmentpossibly due to insufficient sensitivity of the assayInterestingly no increase in tRNAAsp(GUC) methylationwas observed at the highest dnmA expression levelsduring stress recovery In Figure 8E the 30ndash35 methy-lation level in all tRNAAsp(GUC) samples represents signifi-cant methylation as compared with other C residues (seeSupplementary Figure S8)We did however not detect any significant methylation
of tRNAGlu(UUCCUC) and tRNAGly(GCC) under any con-ditions (Supplementary Figure S8) In contrast C49C50methylation which does not depend on Dnmt2 enzymeswas found in all samples and thus served as an internalcontrol In all 1175 bisulfite-sequence reads wereexamined for tRNAGlu(UUCCUC) and 4040 fortRNAGly(GCC) No methylation above background wasfound Though it cannot be excluded that the failure todetect C38 methylation of tRNAGlu and tRNAGly was dueto inefficient reverse transcription or low levels of methy-lation (see lsquoDiscussionrsquo section) we conclude thattRNAAsp is the major or possibly the only in vivo targetfor methylation by DnmA in D discoideum
DISCUSSION
The biological function of Dnmt2 methyltransferases islargely unknown and its role in DNA methylation is con-troversially discussed (910) In contrast tRNA methyla-tion by Dnmt2 is robust in vitro and in vivo andevolutionary conserved In addition to the originallydescribed tRNAAsp(GUC) (1) two further tRNAs targets(tRNAVal(CAC) and tRNAGly(GCC)) have been defined inD melanogaster (15) and tRNAGlu(UUC) in S pombe (17)To get insight into potential functions of the
D discoideum Dnmt2 homologue DnmA we examinedexpression levels in development in the cell cycle andunder different environmental conditions We confirmedby qPCR an expression peak at 16 h of developmentWeijer and co-workers (28) had previously proposed asynchronous cell division at this developmental stageWe therefore investigated dnmA expression in the cellcycle and found a strong transient increase of mRNA inS-phase or shortly thereafter The expression peak wasaccompanied by reaccumulation of DnmA in thenucleus We assume that DnmA is degraded duringmitosis and re-synthesised after cell divisionIn S pombe as well as in D melanogaster tRNA methy-
lation changes in response to environmental challengeslike temperature stress anoxia or nutrients We tested
Figure 7 In vitro methylation of tRNAGly(GCC) by recombinant DnmAand hDnmt2 In vitro transcribed tRNAs were methylated in vitro asdescribed before The top panel shows the ethidium-bromide stainedgel the bottom panel the fluorogram of 3H-labelled methylated tRNAsAs a control tRNAAsp(GUC) methylation is shown
Nucleic Acids Research 2013 9
at Universitaumlt W
uumlrzburg on August 5 2013
httpnaroxfordjournalsorgD
ownloaded from
Figure 8 Expression localization and tRNAAsp(GUC) methylation activity of DnmA under various conditions (A) dnmA expression is regulatedduring the cell cycle Cells were arrested in the cell cycle by cold treatment and then released by transfer to 22C Cells were counted every 30min(grey dots and grey line) and samples were taken for qPCR (black bars) After cell division dnmA expression increased 5-fold and then rapidlydeclined to basal levels Normalization was done on vegetative growing cells dnmA expression is only shown from 3 to 45 h during recovery(B) DnmA is lost from the nucleus during mitosis (arrow) The three cells on the right are in S-phase as indicated by the RFP-PCNA marker andaccumulate DnmA in the nucleus (for further details see movie in Supplementary Material) (C) Relative quantification of dnmA expression levels inAX2 cells after 25 h recovery from cold shock at 4C Expression of dnmA increased gt40-fold and returned within 30min to basal levels (n=3)(D) At 16 h of development dnmA expression increased 46-fold in the NC4 strain In AX2 cells expression increased only 5-fold (E) At 16 h ofdevelopment in vivo methylation of C38 in tRNAAsp(GUC) increased up to 75 in the D discoideum strain NC4 whereas no significant increase wasobserved in AX2 cells
10 Nucleic Acids Research 2013
at Universitaumlt W
uumlrzburg on August 5 2013
httpnaroxfordjournalsorgD
ownloaded from
heat shock cold shock anoxia and various nutrient mediabut did not observe altered levels Only during therecovery phase after cold treatment a transient almost50-fold increase in mRNA abundance was seen
Reuter and Ankri (personal communication) hadobserved a loss of Dnmt2 expression in laboratorystrains of D melanogaster and E histolytica We thereforeexamined D discoideum NC4 the parent of the axeniclaboratory strain AX2 The developmental regulation inAX2 and NC4 was qualitatively similar but expressionlevels were much higher in NC4 The observationsupports the assumption that Dnmt2 expression wasreduced under optimal growth conditions in the labora-tory strain and may actually be selected against
We show that the D discoideum Dnmt2 homologueDnmA is an active tRNAAsp(GUC) methyltransferasein vitro and in vivo To identify further potential targetsfor DnmA we established a UV-CLIP method to analyseDnmAndashRNA interaction in vivo In addition to the knowntarget tRNAAsp(GUC) fragments of tRNAGlu(UUC) andtRNAGly(GCC) were most abundant in comparison withthe control Unexpectedly complete tRNAs were notenriched in the DnmA CLIP fraction but rather specificlsquomid-fragmentsrsquo that covered the putative methylation siteand the recognition pattern proposed by Helm (personal
communication) A minor exception was tRNAGly(GCC)which lacks C40 We do not know whether these frag-ments were directly or indirectly generated by DnmAin vivo or whether they were protected by DnmA fromdegradation during the experimental procedure As noco-crystal of Dnmt2 and a tRNA substrate is availablethe exact interface for the interaction is unknownJurkowski et al (31) mapped the binding site on Dnmt2by mutational analysis A manual placement of a tRNAinto the crystal structure suggests that Dnmt2 mainlycontacts the anticodon stem loop Assuming that mid-frag-ments are footprints of DnmA our data support this notionbut suggest that the interaction surface extends into theD-loop which is partially covered by the mid-fragments
We examined the putative DnmA targets in vitro (bymethylation of in vitro transcribed tRNAs) ex vivo(by methylation of RNA isolated from D discoideum)and in vivo (by bisulfite sequencing) Furthermore weinvestigated by SDSndashPAGE the kinetics of covalentcomplex formation between tRNAAsp and tRNAGlu withDnmA and hDnmt2
All three tRNAs (tRNAAsp tRNAGlu tRNAGly) werereadily methylated by recombinant DnmA and hDnmt2in vitro although the activity was much higher (estimatedat least 10-fold) on tRNAAsp compared with tRNAGlu andtRNAGly For tRNAAsp and tRNAGlu this correlated wellwith the formation and decay of covalent RNAmdashproteincomplexes where kinetics for tRNAAsp were much fasterthan for tRNAGlu Instead of C methylation the covalentintermediate may also proceed to a hydrogen exchangereaction or to a deamination of the cytosine In bothcases an intermediate will be detected but it will notresult in a methylated product (32) It may be thatslower off rates represent these alternative pathways thatwould result in unmethylated tRNAs
Using antisense oligos we showed that tRNAAsptRNAGlu and at least one additional RNA could bemethylated ex vivo A quantification of the results wasnot possible the apparently low methylation oftRNAGlu ex vivo could be due to the low activity of theenzyme on this substrate (as suggested by the in vitroexperiments see Figure 5B)The data for the examination of in vivo methylation by
bisulfite sequencing were rather unexpected AlthoughtRNAAsp(GUC) was consistently found to be methylatedat C38 we never detected any methylation at C38 oftRNAGlu(UUCCUC) or tRNAGly(GCC) There could betwo reasons to explain the failure to detect C38 methyla-tion for these in vitro substrates (i) due to the low activityof DnmA on tRNAGlu(UUCCUC) and tRNAGly(GCC) asshown in vitro there was low or no significant methylationin vivo and (ii) methylation escaped detection becauseother unknown modifications blocked reverse transcrip-tion of the methylated molecules However if this wasthe case one has to assume that C49C50 methylationoccurs independently of these unknown modificationswhereas C38 methylation strictly depends on them AsC38 methylation does not require any other modificationsin vitro we believe that this scenario is less likely If in vitromethylation activity was comparable with in vivo methy-lation 10 methylation of tRNAGlu(UUCCUC) andtRNAGly(GCC) could be expected relative totRNAAsp(GUC) As tRNAAsp was methylated to 30 a3 methylation of tRNAGlu and tRNAGly could be belowthe detection level of the bisulfite assayWe find that high expression of dnmA did not necessar-
ily correlate with increased methylation of tRNAs Duringrecovery from cold shock D discoideum cells displayedthe highest level of dnmA expression but this had noeffect on tRNAAsp methylation Only in developmentand with the high expressing NC4 strain a significantincrease of tRNAAsp methylation to 70 was observedIt has to be considered however that bisulfite sequencingdoes not necessarily allow for quantitative evaluation ofdata detected C-methylation depends on reverse tran-scription efficiency which may be hampered by othermodifications that could be different under differentconditions Furthermore the completeness of bisulfite-mediated conversion could also depend on RNA struc-ture which may be altered by other modificationsHowever incomplete conversion will lead to an overesti-mation of C-methylation as maybe the case for forexample C30 C31 C35 in tRNAGly in some samplesTaken together the data from CLIP complex forma-
tion and methylation show that all three substrates can bebound and methylated in vitro The CLIP data show thatthere is a specific interaction of DnmA with tRNA mid-fragments of tRNAAsp(GUC) tRNAGlu(UUCCUC) andtRNAGly(GCC) but not with other tRNAs In contrastbisulfite sequencing could only detect methylation ontRNAAsp(GUC) even at the highest expression levels ofdnmA Even though low amounts of methylation mayhave escaped detection in bisulfite sequencing there isstill a discrepancy between dnmA expression levels (egduring cold shock recovery) DnmA binding to substrates(as determined by CLIP) and methylation in vivo
Nucleic Acids Research 2013 11
at Universitaumlt W
uumlrzburg on August 5 2013
httpnaroxfordjournalsorgD
ownloaded from
We therefore suggest that DnmA binding to specifictargets could elicit functions in addition to RNA methy-lation eg by promoting processing or degradation andor recruitment of other proteins At least for some rRNAmethyltransferases such as Bud23 or Nep1 fromSaccharomyces cerevisiae it is known that they executefunctions independent from the methylation activity(33ndash35) As shown by Schaefer and co-workers (15)methylation protects tRNAs from degradation or process-ing after stress By a different pathway binding of DnmAmay initiate processing that leads to specific tRNA frag-ments that could serve for other biological functions (36)Interestingly tRNAGlu(UUC) is also a substrate for theDnmt2 homologue Pmt1 in S pombe (17) HowevertRNAGlu(UUC) is found to be methylated to a low butsignificant extend in vivo in fission yeast when Pmt1 isoverexpressedThe detection of tRNAAla(AGC) fragments in the CLIP
assay brings up a new question tRNAAla(AGC) has a Tinstead of a C in position 38 and does not contain thesignature nucleotides in the anticodon stem-loopInstead of a mid-fragment a distinct 30 fragment wasassociated with DnmA suggesting that a recognitionmotif was located in this part of the moleculeAs methylation also occurred in a large artificial
molecule with embedded tRNAGlu (SupplementaryFigure S2) it could be that other longer RNAs containingan appropriate recognition motif may also be bound bythe enzyme We thus propose that the biological functionsof Dnmt2 proteins could extend far beyond our presentknowledge Investigation of further targets and of the sig-nificance of RNA fragments associated with Dnmt2should shed light on the importance of this enzyme
SUPPLEMENTARY DATA
Supplementary Data are available at NAR Online
ACKNOWLEDGEMENTS
The authors thank Thomas Winckler (Friedrich-Schiller-University Jena) for providing the tRNAGlu(UUC) and sup-pressor tRNAGlu(CUA) plasmids Susanne Junk for cloningtRNAAspC38A Matthias Schafer (DKFZ) and RobinLorenz (University of Kassel) are acknowledged forRNA bisulfite sequencing and Richard Reinhardt (MaxPlanck Genome Centre Cologne) for deep sequencingMark Helm (Johannes-Gutenberg-University Mainz) gen-erously provided data before publication CarstenSeehafer (University of Kassel) helped with the analysisof deep-sequencing data
FUNDING
Deutsche Forschungsgemeinschaft [FOR 1082 to WNand AJ] and the Studienstiftung des Deutschen Volkes(to SM) Funding for open access charge FOR1082
Conflict of interest statement None declared
REFERENCES
1 GollMG KirpekarF MaggertKA YoderJA HsiehCLZhangX GolicKG JacobsenSE and BestorTH (2006)Methylation of tRNAAsp by the DNA methyltransferasehomolog Dnmt2 Science 311 395ndash398
2 PhalkeS NickelO WalluscheckD HortigF OnoratiMCand ReuterG (2009) Retrotransposon silencing and telomereintegrity in somatic cells of Drosophila depends on the cytosine-5methyltransferase DNMT2 Nat Genet 41 696ndash702
3 WilkinsonCR BartlettR NurseP and BirdAP (1995) Thefission yeast gene pmt1+ encodes a DNA methyltransferasehomologue Nucleic Acids Res 23 203ndash210
4 KatohM CurkT XuQ ZupanB KuspaA and ShaulskyG(2006) Developmentally regulated DNA methylation inDictyostelium discoideum Eukaryot Cell 5 18ndash25
5 KuhlmannM BorisovaBE KallerM LarssonP StachDNaJ EichingerL LykoF AmbrosV SoderbomF et al(2005) Silencing of retrotransposons in Dictyostelium by DNAmethylation and RNAi Nucleic Acids Res 33 6405ndash6417
6 RaiK ChidesterS ZavalaCV ManosEJ JamesSRKarpfAR JonesDA and CairnsBR (2007) Dnmt2 functionsin the cytoplasm to promote liver brain and retina developmentin zebrafish Genes Dev 21 261ndash266
7 FisherO Siman-TovR and AnkriS (2006) Pleiotropicphenotype in Entamoeba histolytica overexpressing DNAmethyltransferase (Ehmeth) Mol Biochem Parasitol 147 48ndash54
8 DurdevicZ HannaK GoldB PollexT CherryS LykoFand SchaeferM (2013) Efficient RNA virus control in Drosophilarequires the RNA methyltransferase Dnmt2 EMBO Rep 14269ndash275
9 SchaeferM and LykoF (2010) Lack of evidence for DNAmethylation of Invader4 retroelements in Drosophila andimplications for Dnmt2-mediated epigenetic regulation NatGenet 42 920ndash921 author reply 921
10 KraussV and ReuterG (2011) DNA methylation in Drosophila -a critical evaluation Prog Mol Biol Transl Sci 101 177ndash191
11 RaddatzG GuzzardoPM OlovaN FantappieMRRamppM SchaeferM ReikW HannonGJ and LykoF(2013) Dnmt2-dependent methylomes lack defined DNAmethylation patterns Proc Natl Acad Sci USA 110 8627ndash8631
12 JeltschA NellenW and LykoF (2006) Two substrates arebetter than one dual specificities for Dnmt2 methyltransferasesTrends Biochem Sci 31 306ndash308
13 JurkowskiTP MeusburgerM PhalkeS HelmM NellenWReuterG and JeltschA (2008) Human DNMT2 methylatestRNA(Asp) molecules using a DNA methyltransferase-likecatalytic mechanism RNA 14 1663ndash1670
14 TovyA Siman TovR GaentzschR HelmM and AnkriS(2010) A new nuclear function of the Entamoeba histolyticaglycolytic enzyme enolase the metabolic regulation of cytosine-5methyltransferase 2 (Dnmt2) activity PLoS Pathog 6 e1000775
15 SchaeferM PollexT HannaK TuortoF MeusburgerMHelmM and LykoF (2010) RNA methylation by Dnmt2protects transfer RNAs against stress-induced cleavage GenesDev 24 1590ndash1595
16 TuortoF LiebersR MuschT SchaeferM HofmannSKellnerS FryeM HelmM StoecklinG and LykoF (2012)RNA cytosine methylation by Dnmt2 and NSun2 promotestRNA stability and protein synthesis Nat Struct Mol Biol 19900ndash905
17 BeckerM MullerS NellenW JurkowskiTP JeltschA andEhrenhofer-MurrayAE (2012) Pmt1 a Dnmt2 homolog inSchizosaccharomyces pombe mediates tRNA methylation inresponse to nutrient signaling Nucleic Acids Res 4011648ndash11658
18 TovyA HertzR Siman-TovR SyanS FaustD GuillenNand AnkriS (2011) Glucose starvation boosts Entamoebahistolytica virulence PLoS Negl Trop Dis 5 e1247
19 MotorinY and HelmM (2011) RNA nucleotide methylationWiley Interdiscip Rev RNA 2 611ndash631
20 NiranjanakumariS LasdaE BrazasR and Garcia-BlancoMA(2002) Reversible cross-linking combined with
12 Nucleic Acids Research 2013
at Universitaumlt W
uumlrzburg on August 5 2013
httpnaroxfordjournalsorgD
ownloaded from
immunoprecipitation to study RNA-protein interactions in vivoMethods 26 182ndash190
21 RothbauerU ZolghadrK MuyldermansS SchepersACardosoMC and LeonhardtH (2008) A versatile nanotrapfor biochemical and functional studies with fluorescent fusionproteins Mol Cell Proteomics 7 282ndash289
22 SittkaA LucchiniS PapenfortK SharmaCM RolleKBinnewiesTT HintonJC and VogelJ (2008) Deep sequencinganalysis of small noncoding RNA and mRNA targets of theglobal post-transcriptional regulator Hfq PLoS Genet 4e1000163
23 HoffmannS OttoC KurtzS SharmaCM KhaitovichPVogelJ StadlerPF and HackermullerJ (2009) Fast mapping ofshort sequences with mismatches insertions and deletions usingindex structures PLoS Comput Biol 5 e1000502
24 ConnollyMA ClausenPA and LazarJG (2006) Purificationof RNA from animal cells using trizol Cold Spring HarborProtoc 2006 pii pdbprot4104
25 DongA YoderJA ZhangX ZhouL BestorTH andChengX (2001) Structure of human DNMT2 an enigmaticDNA methyltransferase homolog that displays denaturant-resistant binding to DNA Nucleic Acids Res 29 439ndash448
26 SchaeferM PollexT HannaK and LykoF (2009) RNAcytosine methylation analysis by bisulfite sequencing NucleicAcids Res 37 e12
27 BoninM OberstrassJ VogtU WasseneggerM and NellenW(2001) Binding of IRE-BP to its cognate RNA sequence SFMstudies on a universal RNA backbone for the analysis of RNA-protein interaction Biol Chem 382 1157ndash1162
28 ZimmermanW and WeijerCJ (1993) Analysis of cell cycleprogression during the development of Dictyostelium and itsrelationship to differentiation Dev Biol 160 178ndash185
29 StrasserK BloomfieldG MacWilliamsA CeccarelliAMacWilliamsH and TsangA (2012) A retinoblastomaorthologue is a major regulator of S-phase mitotic anddevelopmental gene expression in Dictyostelium PLoS One 7e39914
30 WattsDJ and AshworthJM (1970) Growth of myxameobae ofthe cellular slime mould Dictyostelium discoideum in axenicculture Biochem J 119 171ndash174
31 JurkowskiTP ShanmugamR HelmM and JeltschA (2012)Mapping the tRNA binding site on the surface of humanDNMT2 methyltransferase Biochemistry 51 4438ndash4444
32 SvedruzicZM and ReichNO (2005) DNA cytosine C5methyltransferase Dnmt1 catalysis-dependent release of allostericinhibition Biochemistry 44 9472ndash9485
33 ThomasSR KellerCA SzykA CannonJR and Laronde-LeblancNA (2011) Structural insight into the functionalmechanism of Nep1Emg1 N1-specific pseudouridinemethyltransferase in ribosome biogenesis Nucleic Acids Res 392445ndash2457
34 LeulliotN BohnsackMT GrailleM TollerveyD andVan TilbeurghH (2008) The yeast ribosome synthesisfactor Emg1 is a novel member of the superfamily of alphabeta knot fold methyltransferases Nucleic Acids Res 36629ndash639
35 WhiteJ LiZ SardanaR BujnickiJM MarcotteEM andJohnsonAW (2008) Bud23 methylates G1575 of 18S rRNA andis required for efficient nuclear export of pre-40S subunits MolCell Biol 28 3151ndash3161
36 SobalaA and HutvagnerG (2013) Small RNAs derived fromthe 50-end of tRNA can inhibit protein translation in humancells RNA Biol 10 525ndash535
Nucleic Acids Research 2013 13
at Universitaumlt W
uumlrzburg on August 5 2013
httpnaroxfordjournalsorgD
ownloaded from

(tRNAAsp(GUC) tRNAGlu(UUC) tRNAGly(GCC)) correlatedroughly with the number of tRNA fragments found in theRNA-CLIP experimentThe data supported the conclusion from the ex vivo
blocking assay (Figure 3B) that in addition toD discoideum tRNAAsp and tRNAGlu other RNAs weresubstrates for DnmA and hDnmt2 in vitro
Expression of dnmA is regulated during developmentcell cycle and on temperature stress
DnmA expression in growing vegetative D discoideumcells is low To examine tRNA methylation in vivo it
was therefore advantageous to determine conditionsunder which dnmA was most highly expressed Promptedby previous studies (141517) we assumed that the biolo-gical function of DnmA was connected to developmentalandor environmental conditions We therefore examinedexpression levels of dnmA under different regimes dnmAmRNA was quantified by qPCR over the 24 h develop-mental cycle of D discoideum (data not shown) and con-firmed the data deposited in Dictyexpress (httpdictyexpressbiolabsi)
Interestingly an 5-fold increase in expression wasobserved at 16 h in development This time coincides
Figure 6 tRNA and tRNA fragments associated with DnmA-GFP After UV crosslinking RNA bound to DnmA was co-immunoprecipitatedunder denaturing conditions (CLIP) The number of normalised reads for tRNAs detected by Illumina sequencing is shown for DnmA CLIP and forthe control CLIP with GFP In addition to full-length tRNAs four classes of tRNA fragments were found and these are indicated by differentshading of the bars Size and localization of fragments is shown schematically on the simplified clover leaf structure tRNAs that are not significantlyenriched with DnmA are shown in Supplementary Figure S6 In case of multiple gene copys resulting in the same RNA transcript the sequencingreads were mapped to the first copy in the genome starting from chromsome 1 Only this copy is listed Sequences and detailed information on genecopies and potential isoacceptors are listed in Supplementary Table S2
8 Nucleic Acids Research 2013
at Universitaumlt W
uumlrzburg on August 5 2013
httpnaroxfordjournalsorgD
ownloaded from
with a presumed synchronous mitotic division (28) wherea multitude of cell-cycle regulated genes are expressed[(29) and McWilliams personal Communication]
Therefore we synchronized vegetative D discoideumcells to examine dnmA expression during the cell cycleFor these experiments the D discoideum wild-typestrain NC4 the parent of the axenic laboratory strainAX2 was used (30) This was prompted by the observationthat D melanogaster and E histolytica laboratory strainsmay lose dnmt2 expression (Reuter Ankri personalcommunication)
The data confirmed our assumption of a cell-cycle-de-pendent transcriptional regulation of dnmA (Figure 8A)The peak of expression coincided with late mitosis or earlyS-phase as D discoideum does not display a G1 phaseUsing AX2 cells cotransformed with PCNA-RFP andDnmA-GFP we observed GFP accumulation in thenucleus during S phase (indicated by PCNA expression)and a loss of DnmA from the nucleus during mitosis(Figure 8B) This is further exemplified by the movie inthe Supplementary Material (Supplementary Movie S1)
In other organisms dnmt2 was observed to beupregulated during stress response (315) We thereforeexamined expression after anoxia heat shock and coldshock but did not find any significant changes (data notshown) Only during recovery from cold shock wedetected a strong transient increase in dnmA mRNA by47-fold (Figure 8C)
Thus dnmA expression was differentially regulatedduring the cell cycle development and on temperaturestress Furthermore DnmA localization changed specific-ally during the cell cycle
Though the basic expression in vegetative cells was in-distinguishable dnmA expression in NC4 was 30-foldhigher at 16 h of development whereas developmentalupregulation was only 5-fold in AX2 The data aresummarized in Figure 8D
NC4 the parent strain of AX2 displays increasedtRNA
Asp methylation during development
Based on these observations we asked the questionwhether expression levels correlated with tRNA
methylation activity in vivo Bisulfite sequencing wasdone with primers for tRNAAsp tRNAGlu and tRNAGly
on RNA isolated from vegetative AX2 cells AX2 cellsdeveloped for 16 h AX2 cells subjected to cold shockAX2 recovered for 25 h after cold shock vegetativeNC4 cells and NC4 cells developed for 16 h The resultson C38 methylation of tRNAAsp(GUC) are summarized inFigure 8E The methylation status of all C residues fromall tested tRNAs in this study is shown in SupplementaryFigure S8 Indeed tRNAAsp(GUC) methylation at C38increased to 76 in development in NC4 cells Noincrease was observed in AX2 cells during developmentpossibly due to insufficient sensitivity of the assayInterestingly no increase in tRNAAsp(GUC) methylationwas observed at the highest dnmA expression levelsduring stress recovery In Figure 8E the 30ndash35 methy-lation level in all tRNAAsp(GUC) samples represents signifi-cant methylation as compared with other C residues (seeSupplementary Figure S8)We did however not detect any significant methylation
of tRNAGlu(UUCCUC) and tRNAGly(GCC) under any con-ditions (Supplementary Figure S8) In contrast C49C50methylation which does not depend on Dnmt2 enzymeswas found in all samples and thus served as an internalcontrol In all 1175 bisulfite-sequence reads wereexamined for tRNAGlu(UUCCUC) and 4040 fortRNAGly(GCC) No methylation above background wasfound Though it cannot be excluded that the failure todetect C38 methylation of tRNAGlu and tRNAGly was dueto inefficient reverse transcription or low levels of methy-lation (see lsquoDiscussionrsquo section) we conclude thattRNAAsp is the major or possibly the only in vivo targetfor methylation by DnmA in D discoideum
DISCUSSION
The biological function of Dnmt2 methyltransferases islargely unknown and its role in DNA methylation is con-troversially discussed (910) In contrast tRNA methyla-tion by Dnmt2 is robust in vitro and in vivo andevolutionary conserved In addition to the originallydescribed tRNAAsp(GUC) (1) two further tRNAs targets(tRNAVal(CAC) and tRNAGly(GCC)) have been defined inD melanogaster (15) and tRNAGlu(UUC) in S pombe (17)To get insight into potential functions of the
D discoideum Dnmt2 homologue DnmA we examinedexpression levels in development in the cell cycle andunder different environmental conditions We confirmedby qPCR an expression peak at 16 h of developmentWeijer and co-workers (28) had previously proposed asynchronous cell division at this developmental stageWe therefore investigated dnmA expression in the cellcycle and found a strong transient increase of mRNA inS-phase or shortly thereafter The expression peak wasaccompanied by reaccumulation of DnmA in thenucleus We assume that DnmA is degraded duringmitosis and re-synthesised after cell divisionIn S pombe as well as in D melanogaster tRNA methy-
lation changes in response to environmental challengeslike temperature stress anoxia or nutrients We tested
Figure 7 In vitro methylation of tRNAGly(GCC) by recombinant DnmAand hDnmt2 In vitro transcribed tRNAs were methylated in vitro asdescribed before The top panel shows the ethidium-bromide stainedgel the bottom panel the fluorogram of 3H-labelled methylated tRNAsAs a control tRNAAsp(GUC) methylation is shown
Nucleic Acids Research 2013 9
at Universitaumlt W
uumlrzburg on August 5 2013
httpnaroxfordjournalsorgD
ownloaded from
Figure 8 Expression localization and tRNAAsp(GUC) methylation activity of DnmA under various conditions (A) dnmA expression is regulatedduring the cell cycle Cells were arrested in the cell cycle by cold treatment and then released by transfer to 22C Cells were counted every 30min(grey dots and grey line) and samples were taken for qPCR (black bars) After cell division dnmA expression increased 5-fold and then rapidlydeclined to basal levels Normalization was done on vegetative growing cells dnmA expression is only shown from 3 to 45 h during recovery(B) DnmA is lost from the nucleus during mitosis (arrow) The three cells on the right are in S-phase as indicated by the RFP-PCNA marker andaccumulate DnmA in the nucleus (for further details see movie in Supplementary Material) (C) Relative quantification of dnmA expression levels inAX2 cells after 25 h recovery from cold shock at 4C Expression of dnmA increased gt40-fold and returned within 30min to basal levels (n=3)(D) At 16 h of development dnmA expression increased 46-fold in the NC4 strain In AX2 cells expression increased only 5-fold (E) At 16 h ofdevelopment in vivo methylation of C38 in tRNAAsp(GUC) increased up to 75 in the D discoideum strain NC4 whereas no significant increase wasobserved in AX2 cells
10 Nucleic Acids Research 2013
at Universitaumlt W
uumlrzburg on August 5 2013
httpnaroxfordjournalsorgD
ownloaded from
heat shock cold shock anoxia and various nutrient mediabut did not observe altered levels Only during therecovery phase after cold treatment a transient almost50-fold increase in mRNA abundance was seen
Reuter and Ankri (personal communication) hadobserved a loss of Dnmt2 expression in laboratorystrains of D melanogaster and E histolytica We thereforeexamined D discoideum NC4 the parent of the axeniclaboratory strain AX2 The developmental regulation inAX2 and NC4 was qualitatively similar but expressionlevels were much higher in NC4 The observationsupports the assumption that Dnmt2 expression wasreduced under optimal growth conditions in the labora-tory strain and may actually be selected against
We show that the D discoideum Dnmt2 homologueDnmA is an active tRNAAsp(GUC) methyltransferasein vitro and in vivo To identify further potential targetsfor DnmA we established a UV-CLIP method to analyseDnmAndashRNA interaction in vivo In addition to the knowntarget tRNAAsp(GUC) fragments of tRNAGlu(UUC) andtRNAGly(GCC) were most abundant in comparison withthe control Unexpectedly complete tRNAs were notenriched in the DnmA CLIP fraction but rather specificlsquomid-fragmentsrsquo that covered the putative methylation siteand the recognition pattern proposed by Helm (personal
communication) A minor exception was tRNAGly(GCC)which lacks C40 We do not know whether these frag-ments were directly or indirectly generated by DnmAin vivo or whether they were protected by DnmA fromdegradation during the experimental procedure As noco-crystal of Dnmt2 and a tRNA substrate is availablethe exact interface for the interaction is unknownJurkowski et al (31) mapped the binding site on Dnmt2by mutational analysis A manual placement of a tRNAinto the crystal structure suggests that Dnmt2 mainlycontacts the anticodon stem loop Assuming that mid-frag-ments are footprints of DnmA our data support this notionbut suggest that the interaction surface extends into theD-loop which is partially covered by the mid-fragments
We examined the putative DnmA targets in vitro (bymethylation of in vitro transcribed tRNAs) ex vivo(by methylation of RNA isolated from D discoideum)and in vivo (by bisulfite sequencing) Furthermore weinvestigated by SDSndashPAGE the kinetics of covalentcomplex formation between tRNAAsp and tRNAGlu withDnmA and hDnmt2
All three tRNAs (tRNAAsp tRNAGlu tRNAGly) werereadily methylated by recombinant DnmA and hDnmt2in vitro although the activity was much higher (estimatedat least 10-fold) on tRNAAsp compared with tRNAGlu andtRNAGly For tRNAAsp and tRNAGlu this correlated wellwith the formation and decay of covalent RNAmdashproteincomplexes where kinetics for tRNAAsp were much fasterthan for tRNAGlu Instead of C methylation the covalentintermediate may also proceed to a hydrogen exchangereaction or to a deamination of the cytosine In bothcases an intermediate will be detected but it will notresult in a methylated product (32) It may be thatslower off rates represent these alternative pathways thatwould result in unmethylated tRNAs
Using antisense oligos we showed that tRNAAsptRNAGlu and at least one additional RNA could bemethylated ex vivo A quantification of the results wasnot possible the apparently low methylation oftRNAGlu ex vivo could be due to the low activity of theenzyme on this substrate (as suggested by the in vitroexperiments see Figure 5B)The data for the examination of in vivo methylation by
bisulfite sequencing were rather unexpected AlthoughtRNAAsp(GUC) was consistently found to be methylatedat C38 we never detected any methylation at C38 oftRNAGlu(UUCCUC) or tRNAGly(GCC) There could betwo reasons to explain the failure to detect C38 methyla-tion for these in vitro substrates (i) due to the low activityof DnmA on tRNAGlu(UUCCUC) and tRNAGly(GCC) asshown in vitro there was low or no significant methylationin vivo and (ii) methylation escaped detection becauseother unknown modifications blocked reverse transcrip-tion of the methylated molecules However if this wasthe case one has to assume that C49C50 methylationoccurs independently of these unknown modificationswhereas C38 methylation strictly depends on them AsC38 methylation does not require any other modificationsin vitro we believe that this scenario is less likely If in vitromethylation activity was comparable with in vivo methy-lation 10 methylation of tRNAGlu(UUCCUC) andtRNAGly(GCC) could be expected relative totRNAAsp(GUC) As tRNAAsp was methylated to 30 a3 methylation of tRNAGlu and tRNAGly could be belowthe detection level of the bisulfite assayWe find that high expression of dnmA did not necessar-
ily correlate with increased methylation of tRNAs Duringrecovery from cold shock D discoideum cells displayedthe highest level of dnmA expression but this had noeffect on tRNAAsp methylation Only in developmentand with the high expressing NC4 strain a significantincrease of tRNAAsp methylation to 70 was observedIt has to be considered however that bisulfite sequencingdoes not necessarily allow for quantitative evaluation ofdata detected C-methylation depends on reverse tran-scription efficiency which may be hampered by othermodifications that could be different under differentconditions Furthermore the completeness of bisulfite-mediated conversion could also depend on RNA struc-ture which may be altered by other modificationsHowever incomplete conversion will lead to an overesti-mation of C-methylation as maybe the case for forexample C30 C31 C35 in tRNAGly in some samplesTaken together the data from CLIP complex forma-
tion and methylation show that all three substrates can bebound and methylated in vitro The CLIP data show thatthere is a specific interaction of DnmA with tRNA mid-fragments of tRNAAsp(GUC) tRNAGlu(UUCCUC) andtRNAGly(GCC) but not with other tRNAs In contrastbisulfite sequencing could only detect methylation ontRNAAsp(GUC) even at the highest expression levels ofdnmA Even though low amounts of methylation mayhave escaped detection in bisulfite sequencing there isstill a discrepancy between dnmA expression levels (egduring cold shock recovery) DnmA binding to substrates(as determined by CLIP) and methylation in vivo
Nucleic Acids Research 2013 11
at Universitaumlt W
uumlrzburg on August 5 2013
httpnaroxfordjournalsorgD
ownloaded from
We therefore suggest that DnmA binding to specifictargets could elicit functions in addition to RNA methy-lation eg by promoting processing or degradation andor recruitment of other proteins At least for some rRNAmethyltransferases such as Bud23 or Nep1 fromSaccharomyces cerevisiae it is known that they executefunctions independent from the methylation activity(33ndash35) As shown by Schaefer and co-workers (15)methylation protects tRNAs from degradation or process-ing after stress By a different pathway binding of DnmAmay initiate processing that leads to specific tRNA frag-ments that could serve for other biological functions (36)Interestingly tRNAGlu(UUC) is also a substrate for theDnmt2 homologue Pmt1 in S pombe (17) HowevertRNAGlu(UUC) is found to be methylated to a low butsignificant extend in vivo in fission yeast when Pmt1 isoverexpressedThe detection of tRNAAla(AGC) fragments in the CLIP
assay brings up a new question tRNAAla(AGC) has a Tinstead of a C in position 38 and does not contain thesignature nucleotides in the anticodon stem-loopInstead of a mid-fragment a distinct 30 fragment wasassociated with DnmA suggesting that a recognitionmotif was located in this part of the moleculeAs methylation also occurred in a large artificial
molecule with embedded tRNAGlu (SupplementaryFigure S2) it could be that other longer RNAs containingan appropriate recognition motif may also be bound bythe enzyme We thus propose that the biological functionsof Dnmt2 proteins could extend far beyond our presentknowledge Investigation of further targets and of the sig-nificance of RNA fragments associated with Dnmt2should shed light on the importance of this enzyme
SUPPLEMENTARY DATA
Supplementary Data are available at NAR Online
ACKNOWLEDGEMENTS
The authors thank Thomas Winckler (Friedrich-Schiller-University Jena) for providing the tRNAGlu(UUC) and sup-pressor tRNAGlu(CUA) plasmids Susanne Junk for cloningtRNAAspC38A Matthias Schafer (DKFZ) and RobinLorenz (University of Kassel) are acknowledged forRNA bisulfite sequencing and Richard Reinhardt (MaxPlanck Genome Centre Cologne) for deep sequencingMark Helm (Johannes-Gutenberg-University Mainz) gen-erously provided data before publication CarstenSeehafer (University of Kassel) helped with the analysisof deep-sequencing data
FUNDING
Deutsche Forschungsgemeinschaft [FOR 1082 to WNand AJ] and the Studienstiftung des Deutschen Volkes(to SM) Funding for open access charge FOR1082
Conflict of interest statement None declared
REFERENCES
1 GollMG KirpekarF MaggertKA YoderJA HsiehCLZhangX GolicKG JacobsenSE and BestorTH (2006)Methylation of tRNAAsp by the DNA methyltransferasehomolog Dnmt2 Science 311 395ndash398
2 PhalkeS NickelO WalluscheckD HortigF OnoratiMCand ReuterG (2009) Retrotransposon silencing and telomereintegrity in somatic cells of Drosophila depends on the cytosine-5methyltransferase DNMT2 Nat Genet 41 696ndash702
3 WilkinsonCR BartlettR NurseP and BirdAP (1995) Thefission yeast gene pmt1+ encodes a DNA methyltransferasehomologue Nucleic Acids Res 23 203ndash210
4 KatohM CurkT XuQ ZupanB KuspaA and ShaulskyG(2006) Developmentally regulated DNA methylation inDictyostelium discoideum Eukaryot Cell 5 18ndash25
5 KuhlmannM BorisovaBE KallerM LarssonP StachDNaJ EichingerL LykoF AmbrosV SoderbomF et al(2005) Silencing of retrotransposons in Dictyostelium by DNAmethylation and RNAi Nucleic Acids Res 33 6405ndash6417
6 RaiK ChidesterS ZavalaCV ManosEJ JamesSRKarpfAR JonesDA and CairnsBR (2007) Dnmt2 functionsin the cytoplasm to promote liver brain and retina developmentin zebrafish Genes Dev 21 261ndash266
7 FisherO Siman-TovR and AnkriS (2006) Pleiotropicphenotype in Entamoeba histolytica overexpressing DNAmethyltransferase (Ehmeth) Mol Biochem Parasitol 147 48ndash54
8 DurdevicZ HannaK GoldB PollexT CherryS LykoFand SchaeferM (2013) Efficient RNA virus control in Drosophilarequires the RNA methyltransferase Dnmt2 EMBO Rep 14269ndash275
9 SchaeferM and LykoF (2010) Lack of evidence for DNAmethylation of Invader4 retroelements in Drosophila andimplications for Dnmt2-mediated epigenetic regulation NatGenet 42 920ndash921 author reply 921
10 KraussV and ReuterG (2011) DNA methylation in Drosophila -a critical evaluation Prog Mol Biol Transl Sci 101 177ndash191
11 RaddatzG GuzzardoPM OlovaN FantappieMRRamppM SchaeferM ReikW HannonGJ and LykoF(2013) Dnmt2-dependent methylomes lack defined DNAmethylation patterns Proc Natl Acad Sci USA 110 8627ndash8631
12 JeltschA NellenW and LykoF (2006) Two substrates arebetter than one dual specificities for Dnmt2 methyltransferasesTrends Biochem Sci 31 306ndash308
13 JurkowskiTP MeusburgerM PhalkeS HelmM NellenWReuterG and JeltschA (2008) Human DNMT2 methylatestRNA(Asp) molecules using a DNA methyltransferase-likecatalytic mechanism RNA 14 1663ndash1670
14 TovyA Siman TovR GaentzschR HelmM and AnkriS(2010) A new nuclear function of the Entamoeba histolyticaglycolytic enzyme enolase the metabolic regulation of cytosine-5methyltransferase 2 (Dnmt2) activity PLoS Pathog 6 e1000775
15 SchaeferM PollexT HannaK TuortoF MeusburgerMHelmM and LykoF (2010) RNA methylation by Dnmt2protects transfer RNAs against stress-induced cleavage GenesDev 24 1590ndash1595
16 TuortoF LiebersR MuschT SchaeferM HofmannSKellnerS FryeM HelmM StoecklinG and LykoF (2012)RNA cytosine methylation by Dnmt2 and NSun2 promotestRNA stability and protein synthesis Nat Struct Mol Biol 19900ndash905
17 BeckerM MullerS NellenW JurkowskiTP JeltschA andEhrenhofer-MurrayAE (2012) Pmt1 a Dnmt2 homolog inSchizosaccharomyces pombe mediates tRNA methylation inresponse to nutrient signaling Nucleic Acids Res 4011648ndash11658
18 TovyA HertzR Siman-TovR SyanS FaustD GuillenNand AnkriS (2011) Glucose starvation boosts Entamoebahistolytica virulence PLoS Negl Trop Dis 5 e1247
19 MotorinY and HelmM (2011) RNA nucleotide methylationWiley Interdiscip Rev RNA 2 611ndash631
20 NiranjanakumariS LasdaE BrazasR and Garcia-BlancoMA(2002) Reversible cross-linking combined with
12 Nucleic Acids Research 2013
at Universitaumlt W
uumlrzburg on August 5 2013
httpnaroxfordjournalsorgD
ownloaded from
immunoprecipitation to study RNA-protein interactions in vivoMethods 26 182ndash190
21 RothbauerU ZolghadrK MuyldermansS SchepersACardosoMC and LeonhardtH (2008) A versatile nanotrapfor biochemical and functional studies with fluorescent fusionproteins Mol Cell Proteomics 7 282ndash289
22 SittkaA LucchiniS PapenfortK SharmaCM RolleKBinnewiesTT HintonJC and VogelJ (2008) Deep sequencinganalysis of small noncoding RNA and mRNA targets of theglobal post-transcriptional regulator Hfq PLoS Genet 4e1000163
23 HoffmannS OttoC KurtzS SharmaCM KhaitovichPVogelJ StadlerPF and HackermullerJ (2009) Fast mapping ofshort sequences with mismatches insertions and deletions usingindex structures PLoS Comput Biol 5 e1000502
24 ConnollyMA ClausenPA and LazarJG (2006) Purificationof RNA from animal cells using trizol Cold Spring HarborProtoc 2006 pii pdbprot4104
25 DongA YoderJA ZhangX ZhouL BestorTH andChengX (2001) Structure of human DNMT2 an enigmaticDNA methyltransferase homolog that displays denaturant-resistant binding to DNA Nucleic Acids Res 29 439ndash448
26 SchaeferM PollexT HannaK and LykoF (2009) RNAcytosine methylation analysis by bisulfite sequencing NucleicAcids Res 37 e12
27 BoninM OberstrassJ VogtU WasseneggerM and NellenW(2001) Binding of IRE-BP to its cognate RNA sequence SFMstudies on a universal RNA backbone for the analysis of RNA-protein interaction Biol Chem 382 1157ndash1162
28 ZimmermanW and WeijerCJ (1993) Analysis of cell cycleprogression during the development of Dictyostelium and itsrelationship to differentiation Dev Biol 160 178ndash185
29 StrasserK BloomfieldG MacWilliamsA CeccarelliAMacWilliamsH and TsangA (2012) A retinoblastomaorthologue is a major regulator of S-phase mitotic anddevelopmental gene expression in Dictyostelium PLoS One 7e39914
30 WattsDJ and AshworthJM (1970) Growth of myxameobae ofthe cellular slime mould Dictyostelium discoideum in axenicculture Biochem J 119 171ndash174
31 JurkowskiTP ShanmugamR HelmM and JeltschA (2012)Mapping the tRNA binding site on the surface of humanDNMT2 methyltransferase Biochemistry 51 4438ndash4444
32 SvedruzicZM and ReichNO (2005) DNA cytosine C5methyltransferase Dnmt1 catalysis-dependent release of allostericinhibition Biochemistry 44 9472ndash9485
33 ThomasSR KellerCA SzykA CannonJR and Laronde-LeblancNA (2011) Structural insight into the functionalmechanism of Nep1Emg1 N1-specific pseudouridinemethyltransferase in ribosome biogenesis Nucleic Acids Res 392445ndash2457
34 LeulliotN BohnsackMT GrailleM TollerveyD andVan TilbeurghH (2008) The yeast ribosome synthesisfactor Emg1 is a novel member of the superfamily of alphabeta knot fold methyltransferases Nucleic Acids Res 36629ndash639
35 WhiteJ LiZ SardanaR BujnickiJM MarcotteEM andJohnsonAW (2008) Bud23 methylates G1575 of 18S rRNA andis required for efficient nuclear export of pre-40S subunits MolCell Biol 28 3151ndash3161
36 SobalaA and HutvagnerG (2013) Small RNAs derived fromthe 50-end of tRNA can inhibit protein translation in humancells RNA Biol 10 525ndash535
Nucleic Acids Research 2013 13
at Universitaumlt W
uumlrzburg on August 5 2013
httpnaroxfordjournalsorgD
ownloaded from

with a presumed synchronous mitotic division (28) wherea multitude of cell-cycle regulated genes are expressed[(29) and McWilliams personal Communication]
Therefore we synchronized vegetative D discoideumcells to examine dnmA expression during the cell cycleFor these experiments the D discoideum wild-typestrain NC4 the parent of the axenic laboratory strainAX2 was used (30) This was prompted by the observationthat D melanogaster and E histolytica laboratory strainsmay lose dnmt2 expression (Reuter Ankri personalcommunication)
The data confirmed our assumption of a cell-cycle-de-pendent transcriptional regulation of dnmA (Figure 8A)The peak of expression coincided with late mitosis or earlyS-phase as D discoideum does not display a G1 phaseUsing AX2 cells cotransformed with PCNA-RFP andDnmA-GFP we observed GFP accumulation in thenucleus during S phase (indicated by PCNA expression)and a loss of DnmA from the nucleus during mitosis(Figure 8B) This is further exemplified by the movie inthe Supplementary Material (Supplementary Movie S1)
In other organisms dnmt2 was observed to beupregulated during stress response (315) We thereforeexamined expression after anoxia heat shock and coldshock but did not find any significant changes (data notshown) Only during recovery from cold shock wedetected a strong transient increase in dnmA mRNA by47-fold (Figure 8C)
Thus dnmA expression was differentially regulatedduring the cell cycle development and on temperaturestress Furthermore DnmA localization changed specific-ally during the cell cycle
Though the basic expression in vegetative cells was in-distinguishable dnmA expression in NC4 was 30-foldhigher at 16 h of development whereas developmentalupregulation was only 5-fold in AX2 The data aresummarized in Figure 8D
NC4 the parent strain of AX2 displays increasedtRNA
Asp methylation during development
Based on these observations we asked the questionwhether expression levels correlated with tRNA
methylation activity in vivo Bisulfite sequencing wasdone with primers for tRNAAsp tRNAGlu and tRNAGly
on RNA isolated from vegetative AX2 cells AX2 cellsdeveloped for 16 h AX2 cells subjected to cold shockAX2 recovered for 25 h after cold shock vegetativeNC4 cells and NC4 cells developed for 16 h The resultson C38 methylation of tRNAAsp(GUC) are summarized inFigure 8E The methylation status of all C residues fromall tested tRNAs in this study is shown in SupplementaryFigure S8 Indeed tRNAAsp(GUC) methylation at C38increased to 76 in development in NC4 cells Noincrease was observed in AX2 cells during developmentpossibly due to insufficient sensitivity of the assayInterestingly no increase in tRNAAsp(GUC) methylationwas observed at the highest dnmA expression levelsduring stress recovery In Figure 8E the 30ndash35 methy-lation level in all tRNAAsp(GUC) samples represents signifi-cant methylation as compared with other C residues (seeSupplementary Figure S8)We did however not detect any significant methylation
of tRNAGlu(UUCCUC) and tRNAGly(GCC) under any con-ditions (Supplementary Figure S8) In contrast C49C50methylation which does not depend on Dnmt2 enzymeswas found in all samples and thus served as an internalcontrol In all 1175 bisulfite-sequence reads wereexamined for tRNAGlu(UUCCUC) and 4040 fortRNAGly(GCC) No methylation above background wasfound Though it cannot be excluded that the failure todetect C38 methylation of tRNAGlu and tRNAGly was dueto inefficient reverse transcription or low levels of methy-lation (see lsquoDiscussionrsquo section) we conclude thattRNAAsp is the major or possibly the only in vivo targetfor methylation by DnmA in D discoideum
DISCUSSION
The biological function of Dnmt2 methyltransferases islargely unknown and its role in DNA methylation is con-troversially discussed (910) In contrast tRNA methyla-tion by Dnmt2 is robust in vitro and in vivo andevolutionary conserved In addition to the originallydescribed tRNAAsp(GUC) (1) two further tRNAs targets(tRNAVal(CAC) and tRNAGly(GCC)) have been defined inD melanogaster (15) and tRNAGlu(UUC) in S pombe (17)To get insight into potential functions of the
D discoideum Dnmt2 homologue DnmA we examinedexpression levels in development in the cell cycle andunder different environmental conditions We confirmedby qPCR an expression peak at 16 h of developmentWeijer and co-workers (28) had previously proposed asynchronous cell division at this developmental stageWe therefore investigated dnmA expression in the cellcycle and found a strong transient increase of mRNA inS-phase or shortly thereafter The expression peak wasaccompanied by reaccumulation of DnmA in thenucleus We assume that DnmA is degraded duringmitosis and re-synthesised after cell divisionIn S pombe as well as in D melanogaster tRNA methy-
lation changes in response to environmental challengeslike temperature stress anoxia or nutrients We tested
Figure 7 In vitro methylation of tRNAGly(GCC) by recombinant DnmAand hDnmt2 In vitro transcribed tRNAs were methylated in vitro asdescribed before The top panel shows the ethidium-bromide stainedgel the bottom panel the fluorogram of 3H-labelled methylated tRNAsAs a control tRNAAsp(GUC) methylation is shown
Nucleic Acids Research 2013 9
at Universitaumlt W
uumlrzburg on August 5 2013
httpnaroxfordjournalsorgD
ownloaded from
Figure 8 Expression localization and tRNAAsp(GUC) methylation activity of DnmA under various conditions (A) dnmA expression is regulatedduring the cell cycle Cells were arrested in the cell cycle by cold treatment and then released by transfer to 22C Cells were counted every 30min(grey dots and grey line) and samples were taken for qPCR (black bars) After cell division dnmA expression increased 5-fold and then rapidlydeclined to basal levels Normalization was done on vegetative growing cells dnmA expression is only shown from 3 to 45 h during recovery(B) DnmA is lost from the nucleus during mitosis (arrow) The three cells on the right are in S-phase as indicated by the RFP-PCNA marker andaccumulate DnmA in the nucleus (for further details see movie in Supplementary Material) (C) Relative quantification of dnmA expression levels inAX2 cells after 25 h recovery from cold shock at 4C Expression of dnmA increased gt40-fold and returned within 30min to basal levels (n=3)(D) At 16 h of development dnmA expression increased 46-fold in the NC4 strain In AX2 cells expression increased only 5-fold (E) At 16 h ofdevelopment in vivo methylation of C38 in tRNAAsp(GUC) increased up to 75 in the D discoideum strain NC4 whereas no significant increase wasobserved in AX2 cells
10 Nucleic Acids Research 2013
at Universitaumlt W
uumlrzburg on August 5 2013
httpnaroxfordjournalsorgD
ownloaded from
heat shock cold shock anoxia and various nutrient mediabut did not observe altered levels Only during therecovery phase after cold treatment a transient almost50-fold increase in mRNA abundance was seen
Reuter and Ankri (personal communication) hadobserved a loss of Dnmt2 expression in laboratorystrains of D melanogaster and E histolytica We thereforeexamined D discoideum NC4 the parent of the axeniclaboratory strain AX2 The developmental regulation inAX2 and NC4 was qualitatively similar but expressionlevels were much higher in NC4 The observationsupports the assumption that Dnmt2 expression wasreduced under optimal growth conditions in the labora-tory strain and may actually be selected against
We show that the D discoideum Dnmt2 homologueDnmA is an active tRNAAsp(GUC) methyltransferasein vitro and in vivo To identify further potential targetsfor DnmA we established a UV-CLIP method to analyseDnmAndashRNA interaction in vivo In addition to the knowntarget tRNAAsp(GUC) fragments of tRNAGlu(UUC) andtRNAGly(GCC) were most abundant in comparison withthe control Unexpectedly complete tRNAs were notenriched in the DnmA CLIP fraction but rather specificlsquomid-fragmentsrsquo that covered the putative methylation siteand the recognition pattern proposed by Helm (personal
communication) A minor exception was tRNAGly(GCC)which lacks C40 We do not know whether these frag-ments were directly or indirectly generated by DnmAin vivo or whether they were protected by DnmA fromdegradation during the experimental procedure As noco-crystal of Dnmt2 and a tRNA substrate is availablethe exact interface for the interaction is unknownJurkowski et al (31) mapped the binding site on Dnmt2by mutational analysis A manual placement of a tRNAinto the crystal structure suggests that Dnmt2 mainlycontacts the anticodon stem loop Assuming that mid-frag-ments are footprints of DnmA our data support this notionbut suggest that the interaction surface extends into theD-loop which is partially covered by the mid-fragments
We examined the putative DnmA targets in vitro (bymethylation of in vitro transcribed tRNAs) ex vivo(by methylation of RNA isolated from D discoideum)and in vivo (by bisulfite sequencing) Furthermore weinvestigated by SDSndashPAGE the kinetics of covalentcomplex formation between tRNAAsp and tRNAGlu withDnmA and hDnmt2
All three tRNAs (tRNAAsp tRNAGlu tRNAGly) werereadily methylated by recombinant DnmA and hDnmt2in vitro although the activity was much higher (estimatedat least 10-fold) on tRNAAsp compared with tRNAGlu andtRNAGly For tRNAAsp and tRNAGlu this correlated wellwith the formation and decay of covalent RNAmdashproteincomplexes where kinetics for tRNAAsp were much fasterthan for tRNAGlu Instead of C methylation the covalentintermediate may also proceed to a hydrogen exchangereaction or to a deamination of the cytosine In bothcases an intermediate will be detected but it will notresult in a methylated product (32) It may be thatslower off rates represent these alternative pathways thatwould result in unmethylated tRNAs
Using antisense oligos we showed that tRNAAsptRNAGlu and at least one additional RNA could bemethylated ex vivo A quantification of the results wasnot possible the apparently low methylation oftRNAGlu ex vivo could be due to the low activity of theenzyme on this substrate (as suggested by the in vitroexperiments see Figure 5B)The data for the examination of in vivo methylation by
bisulfite sequencing were rather unexpected AlthoughtRNAAsp(GUC) was consistently found to be methylatedat C38 we never detected any methylation at C38 oftRNAGlu(UUCCUC) or tRNAGly(GCC) There could betwo reasons to explain the failure to detect C38 methyla-tion for these in vitro substrates (i) due to the low activityof DnmA on tRNAGlu(UUCCUC) and tRNAGly(GCC) asshown in vitro there was low or no significant methylationin vivo and (ii) methylation escaped detection becauseother unknown modifications blocked reverse transcrip-tion of the methylated molecules However if this wasthe case one has to assume that C49C50 methylationoccurs independently of these unknown modificationswhereas C38 methylation strictly depends on them AsC38 methylation does not require any other modificationsin vitro we believe that this scenario is less likely If in vitromethylation activity was comparable with in vivo methy-lation 10 methylation of tRNAGlu(UUCCUC) andtRNAGly(GCC) could be expected relative totRNAAsp(GUC) As tRNAAsp was methylated to 30 a3 methylation of tRNAGlu and tRNAGly could be belowthe detection level of the bisulfite assayWe find that high expression of dnmA did not necessar-
ily correlate with increased methylation of tRNAs Duringrecovery from cold shock D discoideum cells displayedthe highest level of dnmA expression but this had noeffect on tRNAAsp methylation Only in developmentand with the high expressing NC4 strain a significantincrease of tRNAAsp methylation to 70 was observedIt has to be considered however that bisulfite sequencingdoes not necessarily allow for quantitative evaluation ofdata detected C-methylation depends on reverse tran-scription efficiency which may be hampered by othermodifications that could be different under differentconditions Furthermore the completeness of bisulfite-mediated conversion could also depend on RNA struc-ture which may be altered by other modificationsHowever incomplete conversion will lead to an overesti-mation of C-methylation as maybe the case for forexample C30 C31 C35 in tRNAGly in some samplesTaken together the data from CLIP complex forma-
tion and methylation show that all three substrates can bebound and methylated in vitro The CLIP data show thatthere is a specific interaction of DnmA with tRNA mid-fragments of tRNAAsp(GUC) tRNAGlu(UUCCUC) andtRNAGly(GCC) but not with other tRNAs In contrastbisulfite sequencing could only detect methylation ontRNAAsp(GUC) even at the highest expression levels ofdnmA Even though low amounts of methylation mayhave escaped detection in bisulfite sequencing there isstill a discrepancy between dnmA expression levels (egduring cold shock recovery) DnmA binding to substrates(as determined by CLIP) and methylation in vivo
Nucleic Acids Research 2013 11
at Universitaumlt W
uumlrzburg on August 5 2013
httpnaroxfordjournalsorgD
ownloaded from
We therefore suggest that DnmA binding to specifictargets could elicit functions in addition to RNA methy-lation eg by promoting processing or degradation andor recruitment of other proteins At least for some rRNAmethyltransferases such as Bud23 or Nep1 fromSaccharomyces cerevisiae it is known that they executefunctions independent from the methylation activity(33ndash35) As shown by Schaefer and co-workers (15)methylation protects tRNAs from degradation or process-ing after stress By a different pathway binding of DnmAmay initiate processing that leads to specific tRNA frag-ments that could serve for other biological functions (36)Interestingly tRNAGlu(UUC) is also a substrate for theDnmt2 homologue Pmt1 in S pombe (17) HowevertRNAGlu(UUC) is found to be methylated to a low butsignificant extend in vivo in fission yeast when Pmt1 isoverexpressedThe detection of tRNAAla(AGC) fragments in the CLIP
assay brings up a new question tRNAAla(AGC) has a Tinstead of a C in position 38 and does not contain thesignature nucleotides in the anticodon stem-loopInstead of a mid-fragment a distinct 30 fragment wasassociated with DnmA suggesting that a recognitionmotif was located in this part of the moleculeAs methylation also occurred in a large artificial
molecule with embedded tRNAGlu (SupplementaryFigure S2) it could be that other longer RNAs containingan appropriate recognition motif may also be bound bythe enzyme We thus propose that the biological functionsof Dnmt2 proteins could extend far beyond our presentknowledge Investigation of further targets and of the sig-nificance of RNA fragments associated with Dnmt2should shed light on the importance of this enzyme
SUPPLEMENTARY DATA
Supplementary Data are available at NAR Online
ACKNOWLEDGEMENTS
The authors thank Thomas Winckler (Friedrich-Schiller-University Jena) for providing the tRNAGlu(UUC) and sup-pressor tRNAGlu(CUA) plasmids Susanne Junk for cloningtRNAAspC38A Matthias Schafer (DKFZ) and RobinLorenz (University of Kassel) are acknowledged forRNA bisulfite sequencing and Richard Reinhardt (MaxPlanck Genome Centre Cologne) for deep sequencingMark Helm (Johannes-Gutenberg-University Mainz) gen-erously provided data before publication CarstenSeehafer (University of Kassel) helped with the analysisof deep-sequencing data
FUNDING
Deutsche Forschungsgemeinschaft [FOR 1082 to WNand AJ] and the Studienstiftung des Deutschen Volkes(to SM) Funding for open access charge FOR1082
Conflict of interest statement None declared
REFERENCES
1 GollMG KirpekarF MaggertKA YoderJA HsiehCLZhangX GolicKG JacobsenSE and BestorTH (2006)Methylation of tRNAAsp by the DNA methyltransferasehomolog Dnmt2 Science 311 395ndash398
2 PhalkeS NickelO WalluscheckD HortigF OnoratiMCand ReuterG (2009) Retrotransposon silencing and telomereintegrity in somatic cells of Drosophila depends on the cytosine-5methyltransferase DNMT2 Nat Genet 41 696ndash702
3 WilkinsonCR BartlettR NurseP and BirdAP (1995) Thefission yeast gene pmt1+ encodes a DNA methyltransferasehomologue Nucleic Acids Res 23 203ndash210
4 KatohM CurkT XuQ ZupanB KuspaA and ShaulskyG(2006) Developmentally regulated DNA methylation inDictyostelium discoideum Eukaryot Cell 5 18ndash25
5 KuhlmannM BorisovaBE KallerM LarssonP StachDNaJ EichingerL LykoF AmbrosV SoderbomF et al(2005) Silencing of retrotransposons in Dictyostelium by DNAmethylation and RNAi Nucleic Acids Res 33 6405ndash6417
6 RaiK ChidesterS ZavalaCV ManosEJ JamesSRKarpfAR JonesDA and CairnsBR (2007) Dnmt2 functionsin the cytoplasm to promote liver brain and retina developmentin zebrafish Genes Dev 21 261ndash266
7 FisherO Siman-TovR and AnkriS (2006) Pleiotropicphenotype in Entamoeba histolytica overexpressing DNAmethyltransferase (Ehmeth) Mol Biochem Parasitol 147 48ndash54
8 DurdevicZ HannaK GoldB PollexT CherryS LykoFand SchaeferM (2013) Efficient RNA virus control in Drosophilarequires the RNA methyltransferase Dnmt2 EMBO Rep 14269ndash275
9 SchaeferM and LykoF (2010) Lack of evidence for DNAmethylation of Invader4 retroelements in Drosophila andimplications for Dnmt2-mediated epigenetic regulation NatGenet 42 920ndash921 author reply 921
10 KraussV and ReuterG (2011) DNA methylation in Drosophila -a critical evaluation Prog Mol Biol Transl Sci 101 177ndash191
11 RaddatzG GuzzardoPM OlovaN FantappieMRRamppM SchaeferM ReikW HannonGJ and LykoF(2013) Dnmt2-dependent methylomes lack defined DNAmethylation patterns Proc Natl Acad Sci USA 110 8627ndash8631
12 JeltschA NellenW and LykoF (2006) Two substrates arebetter than one dual specificities for Dnmt2 methyltransferasesTrends Biochem Sci 31 306ndash308
13 JurkowskiTP MeusburgerM PhalkeS HelmM NellenWReuterG and JeltschA (2008) Human DNMT2 methylatestRNA(Asp) molecules using a DNA methyltransferase-likecatalytic mechanism RNA 14 1663ndash1670
14 TovyA Siman TovR GaentzschR HelmM and AnkriS(2010) A new nuclear function of the Entamoeba histolyticaglycolytic enzyme enolase the metabolic regulation of cytosine-5methyltransferase 2 (Dnmt2) activity PLoS Pathog 6 e1000775
15 SchaeferM PollexT HannaK TuortoF MeusburgerMHelmM and LykoF (2010) RNA methylation by Dnmt2protects transfer RNAs against stress-induced cleavage GenesDev 24 1590ndash1595
16 TuortoF LiebersR MuschT SchaeferM HofmannSKellnerS FryeM HelmM StoecklinG and LykoF (2012)RNA cytosine methylation by Dnmt2 and NSun2 promotestRNA stability and protein synthesis Nat Struct Mol Biol 19900ndash905
17 BeckerM MullerS NellenW JurkowskiTP JeltschA andEhrenhofer-MurrayAE (2012) Pmt1 a Dnmt2 homolog inSchizosaccharomyces pombe mediates tRNA methylation inresponse to nutrient signaling Nucleic Acids Res 4011648ndash11658
18 TovyA HertzR Siman-TovR SyanS FaustD GuillenNand AnkriS (2011) Glucose starvation boosts Entamoebahistolytica virulence PLoS Negl Trop Dis 5 e1247
19 MotorinY and HelmM (2011) RNA nucleotide methylationWiley Interdiscip Rev RNA 2 611ndash631
20 NiranjanakumariS LasdaE BrazasR and Garcia-BlancoMA(2002) Reversible cross-linking combined with
12 Nucleic Acids Research 2013
at Universitaumlt W
uumlrzburg on August 5 2013
httpnaroxfordjournalsorgD
ownloaded from
immunoprecipitation to study RNA-protein interactions in vivoMethods 26 182ndash190
21 RothbauerU ZolghadrK MuyldermansS SchepersACardosoMC and LeonhardtH (2008) A versatile nanotrapfor biochemical and functional studies with fluorescent fusionproteins Mol Cell Proteomics 7 282ndash289
22 SittkaA LucchiniS PapenfortK SharmaCM RolleKBinnewiesTT HintonJC and VogelJ (2008) Deep sequencinganalysis of small noncoding RNA and mRNA targets of theglobal post-transcriptional regulator Hfq PLoS Genet 4e1000163
23 HoffmannS OttoC KurtzS SharmaCM KhaitovichPVogelJ StadlerPF and HackermullerJ (2009) Fast mapping ofshort sequences with mismatches insertions and deletions usingindex structures PLoS Comput Biol 5 e1000502
24 ConnollyMA ClausenPA and LazarJG (2006) Purificationof RNA from animal cells using trizol Cold Spring HarborProtoc 2006 pii pdbprot4104
25 DongA YoderJA ZhangX ZhouL BestorTH andChengX (2001) Structure of human DNMT2 an enigmaticDNA methyltransferase homolog that displays denaturant-resistant binding to DNA Nucleic Acids Res 29 439ndash448
26 SchaeferM PollexT HannaK and LykoF (2009) RNAcytosine methylation analysis by bisulfite sequencing NucleicAcids Res 37 e12
27 BoninM OberstrassJ VogtU WasseneggerM and NellenW(2001) Binding of IRE-BP to its cognate RNA sequence SFMstudies on a universal RNA backbone for the analysis of RNA-protein interaction Biol Chem 382 1157ndash1162
28 ZimmermanW and WeijerCJ (1993) Analysis of cell cycleprogression during the development of Dictyostelium and itsrelationship to differentiation Dev Biol 160 178ndash185
29 StrasserK BloomfieldG MacWilliamsA CeccarelliAMacWilliamsH and TsangA (2012) A retinoblastomaorthologue is a major regulator of S-phase mitotic anddevelopmental gene expression in Dictyostelium PLoS One 7e39914
30 WattsDJ and AshworthJM (1970) Growth of myxameobae ofthe cellular slime mould Dictyostelium discoideum in axenicculture Biochem J 119 171ndash174
31 JurkowskiTP ShanmugamR HelmM and JeltschA (2012)Mapping the tRNA binding site on the surface of humanDNMT2 methyltransferase Biochemistry 51 4438ndash4444
32 SvedruzicZM and ReichNO (2005) DNA cytosine C5methyltransferase Dnmt1 catalysis-dependent release of allostericinhibition Biochemistry 44 9472ndash9485
33 ThomasSR KellerCA SzykA CannonJR and Laronde-LeblancNA (2011) Structural insight into the functionalmechanism of Nep1Emg1 N1-specific pseudouridinemethyltransferase in ribosome biogenesis Nucleic Acids Res 392445ndash2457
34 LeulliotN BohnsackMT GrailleM TollerveyD andVan TilbeurghH (2008) The yeast ribosome synthesisfactor Emg1 is a novel member of the superfamily of alphabeta knot fold methyltransferases Nucleic Acids Res 36629ndash639
35 WhiteJ LiZ SardanaR BujnickiJM MarcotteEM andJohnsonAW (2008) Bud23 methylates G1575 of 18S rRNA andis required for efficient nuclear export of pre-40S subunits MolCell Biol 28 3151ndash3161
36 SobalaA and HutvagnerG (2013) Small RNAs derived fromthe 50-end of tRNA can inhibit protein translation in humancells RNA Biol 10 525ndash535
Nucleic Acids Research 2013 13
at Universitaumlt W
uumlrzburg on August 5 2013
httpnaroxfordjournalsorgD
ownloaded from
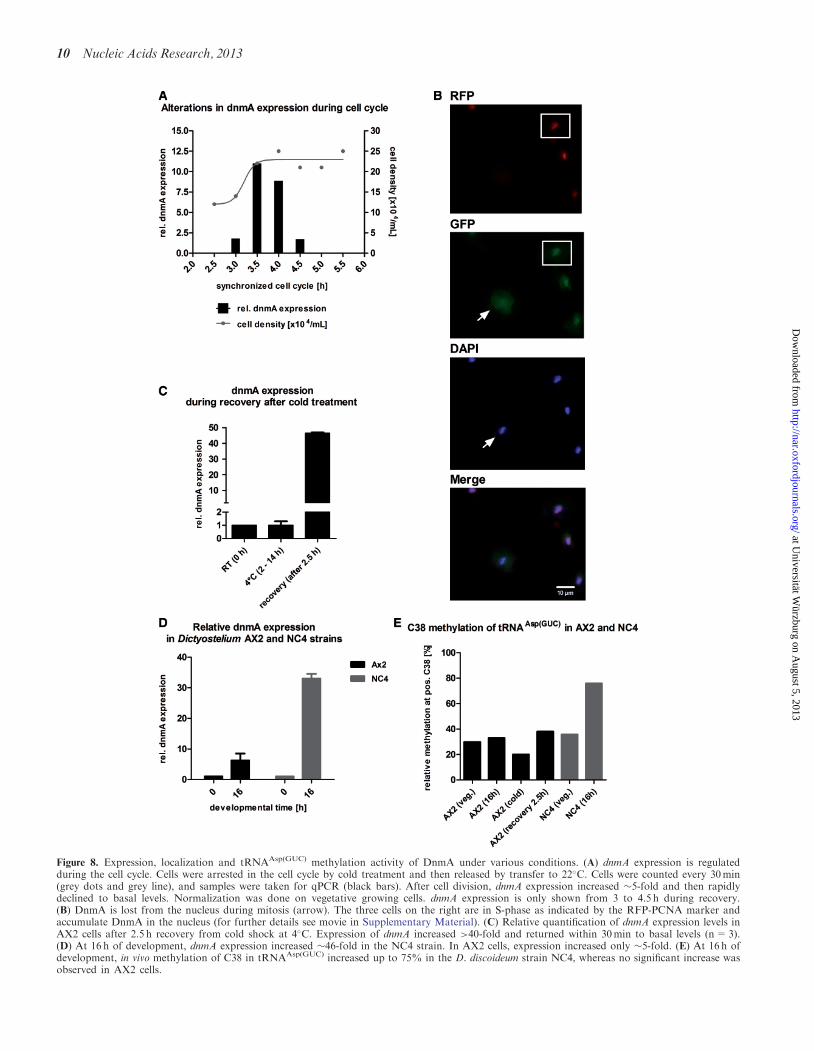
Figure 8 Expression localization and tRNAAsp(GUC) methylation activity of DnmA under various conditions (A) dnmA expression is regulatedduring the cell cycle Cells were arrested in the cell cycle by cold treatment and then released by transfer to 22C Cells were counted every 30min(grey dots and grey line) and samples were taken for qPCR (black bars) After cell division dnmA expression increased 5-fold and then rapidlydeclined to basal levels Normalization was done on vegetative growing cells dnmA expression is only shown from 3 to 45 h during recovery(B) DnmA is lost from the nucleus during mitosis (arrow) The three cells on the right are in S-phase as indicated by the RFP-PCNA marker andaccumulate DnmA in the nucleus (for further details see movie in Supplementary Material) (C) Relative quantification of dnmA expression levels inAX2 cells after 25 h recovery from cold shock at 4C Expression of dnmA increased gt40-fold and returned within 30min to basal levels (n=3)(D) At 16 h of development dnmA expression increased 46-fold in the NC4 strain In AX2 cells expression increased only 5-fold (E) At 16 h ofdevelopment in vivo methylation of C38 in tRNAAsp(GUC) increased up to 75 in the D discoideum strain NC4 whereas no significant increase wasobserved in AX2 cells
10 Nucleic Acids Research 2013
at Universitaumlt W
uumlrzburg on August 5 2013
httpnaroxfordjournalsorgD
ownloaded from
heat shock cold shock anoxia and various nutrient mediabut did not observe altered levels Only during therecovery phase after cold treatment a transient almost50-fold increase in mRNA abundance was seen
Reuter and Ankri (personal communication) hadobserved a loss of Dnmt2 expression in laboratorystrains of D melanogaster and E histolytica We thereforeexamined D discoideum NC4 the parent of the axeniclaboratory strain AX2 The developmental regulation inAX2 and NC4 was qualitatively similar but expressionlevels were much higher in NC4 The observationsupports the assumption that Dnmt2 expression wasreduced under optimal growth conditions in the labora-tory strain and may actually be selected against
We show that the D discoideum Dnmt2 homologueDnmA is an active tRNAAsp(GUC) methyltransferasein vitro and in vivo To identify further potential targetsfor DnmA we established a UV-CLIP method to analyseDnmAndashRNA interaction in vivo In addition to the knowntarget tRNAAsp(GUC) fragments of tRNAGlu(UUC) andtRNAGly(GCC) were most abundant in comparison withthe control Unexpectedly complete tRNAs were notenriched in the DnmA CLIP fraction but rather specificlsquomid-fragmentsrsquo that covered the putative methylation siteand the recognition pattern proposed by Helm (personal
communication) A minor exception was tRNAGly(GCC)which lacks C40 We do not know whether these frag-ments were directly or indirectly generated by DnmAin vivo or whether they were protected by DnmA fromdegradation during the experimental procedure As noco-crystal of Dnmt2 and a tRNA substrate is availablethe exact interface for the interaction is unknownJurkowski et al (31) mapped the binding site on Dnmt2by mutational analysis A manual placement of a tRNAinto the crystal structure suggests that Dnmt2 mainlycontacts the anticodon stem loop Assuming that mid-frag-ments are footprints of DnmA our data support this notionbut suggest that the interaction surface extends into theD-loop which is partially covered by the mid-fragments
We examined the putative DnmA targets in vitro (bymethylation of in vitro transcribed tRNAs) ex vivo(by methylation of RNA isolated from D discoideum)and in vivo (by bisulfite sequencing) Furthermore weinvestigated by SDSndashPAGE the kinetics of covalentcomplex formation between tRNAAsp and tRNAGlu withDnmA and hDnmt2
All three tRNAs (tRNAAsp tRNAGlu tRNAGly) werereadily methylated by recombinant DnmA and hDnmt2in vitro although the activity was much higher (estimatedat least 10-fold) on tRNAAsp compared with tRNAGlu andtRNAGly For tRNAAsp and tRNAGlu this correlated wellwith the formation and decay of covalent RNAmdashproteincomplexes where kinetics for tRNAAsp were much fasterthan for tRNAGlu Instead of C methylation the covalentintermediate may also proceed to a hydrogen exchangereaction or to a deamination of the cytosine In bothcases an intermediate will be detected but it will notresult in a methylated product (32) It may be thatslower off rates represent these alternative pathways thatwould result in unmethylated tRNAs
Using antisense oligos we showed that tRNAAsptRNAGlu and at least one additional RNA could bemethylated ex vivo A quantification of the results wasnot possible the apparently low methylation oftRNAGlu ex vivo could be due to the low activity of theenzyme on this substrate (as suggested by the in vitroexperiments see Figure 5B)The data for the examination of in vivo methylation by
bisulfite sequencing were rather unexpected AlthoughtRNAAsp(GUC) was consistently found to be methylatedat C38 we never detected any methylation at C38 oftRNAGlu(UUCCUC) or tRNAGly(GCC) There could betwo reasons to explain the failure to detect C38 methyla-tion for these in vitro substrates (i) due to the low activityof DnmA on tRNAGlu(UUCCUC) and tRNAGly(GCC) asshown in vitro there was low or no significant methylationin vivo and (ii) methylation escaped detection becauseother unknown modifications blocked reverse transcrip-tion of the methylated molecules However if this wasthe case one has to assume that C49C50 methylationoccurs independently of these unknown modificationswhereas C38 methylation strictly depends on them AsC38 methylation does not require any other modificationsin vitro we believe that this scenario is less likely If in vitromethylation activity was comparable with in vivo methy-lation 10 methylation of tRNAGlu(UUCCUC) andtRNAGly(GCC) could be expected relative totRNAAsp(GUC) As tRNAAsp was methylated to 30 a3 methylation of tRNAGlu and tRNAGly could be belowthe detection level of the bisulfite assayWe find that high expression of dnmA did not necessar-
ily correlate with increased methylation of tRNAs Duringrecovery from cold shock D discoideum cells displayedthe highest level of dnmA expression but this had noeffect on tRNAAsp methylation Only in developmentand with the high expressing NC4 strain a significantincrease of tRNAAsp methylation to 70 was observedIt has to be considered however that bisulfite sequencingdoes not necessarily allow for quantitative evaluation ofdata detected C-methylation depends on reverse tran-scription efficiency which may be hampered by othermodifications that could be different under differentconditions Furthermore the completeness of bisulfite-mediated conversion could also depend on RNA struc-ture which may be altered by other modificationsHowever incomplete conversion will lead to an overesti-mation of C-methylation as maybe the case for forexample C30 C31 C35 in tRNAGly in some samplesTaken together the data from CLIP complex forma-
tion and methylation show that all three substrates can bebound and methylated in vitro The CLIP data show thatthere is a specific interaction of DnmA with tRNA mid-fragments of tRNAAsp(GUC) tRNAGlu(UUCCUC) andtRNAGly(GCC) but not with other tRNAs In contrastbisulfite sequencing could only detect methylation ontRNAAsp(GUC) even at the highest expression levels ofdnmA Even though low amounts of methylation mayhave escaped detection in bisulfite sequencing there isstill a discrepancy between dnmA expression levels (egduring cold shock recovery) DnmA binding to substrates(as determined by CLIP) and methylation in vivo
Nucleic Acids Research 2013 11
at Universitaumlt W
uumlrzburg on August 5 2013
httpnaroxfordjournalsorgD
ownloaded from
We therefore suggest that DnmA binding to specifictargets could elicit functions in addition to RNA methy-lation eg by promoting processing or degradation andor recruitment of other proteins At least for some rRNAmethyltransferases such as Bud23 or Nep1 fromSaccharomyces cerevisiae it is known that they executefunctions independent from the methylation activity(33ndash35) As shown by Schaefer and co-workers (15)methylation protects tRNAs from degradation or process-ing after stress By a different pathway binding of DnmAmay initiate processing that leads to specific tRNA frag-ments that could serve for other biological functions (36)Interestingly tRNAGlu(UUC) is also a substrate for theDnmt2 homologue Pmt1 in S pombe (17) HowevertRNAGlu(UUC) is found to be methylated to a low butsignificant extend in vivo in fission yeast when Pmt1 isoverexpressedThe detection of tRNAAla(AGC) fragments in the CLIP
assay brings up a new question tRNAAla(AGC) has a Tinstead of a C in position 38 and does not contain thesignature nucleotides in the anticodon stem-loopInstead of a mid-fragment a distinct 30 fragment wasassociated with DnmA suggesting that a recognitionmotif was located in this part of the moleculeAs methylation also occurred in a large artificial
molecule with embedded tRNAGlu (SupplementaryFigure S2) it could be that other longer RNAs containingan appropriate recognition motif may also be bound bythe enzyme We thus propose that the biological functionsof Dnmt2 proteins could extend far beyond our presentknowledge Investigation of further targets and of the sig-nificance of RNA fragments associated with Dnmt2should shed light on the importance of this enzyme
SUPPLEMENTARY DATA
Supplementary Data are available at NAR Online
ACKNOWLEDGEMENTS
The authors thank Thomas Winckler (Friedrich-Schiller-University Jena) for providing the tRNAGlu(UUC) and sup-pressor tRNAGlu(CUA) plasmids Susanne Junk for cloningtRNAAspC38A Matthias Schafer (DKFZ) and RobinLorenz (University of Kassel) are acknowledged forRNA bisulfite sequencing and Richard Reinhardt (MaxPlanck Genome Centre Cologne) for deep sequencingMark Helm (Johannes-Gutenberg-University Mainz) gen-erously provided data before publication CarstenSeehafer (University of Kassel) helped with the analysisof deep-sequencing data
FUNDING
Deutsche Forschungsgemeinschaft [FOR 1082 to WNand AJ] and the Studienstiftung des Deutschen Volkes(to SM) Funding for open access charge FOR1082
Conflict of interest statement None declared
REFERENCES
1 GollMG KirpekarF MaggertKA YoderJA HsiehCLZhangX GolicKG JacobsenSE and BestorTH (2006)Methylation of tRNAAsp by the DNA methyltransferasehomolog Dnmt2 Science 311 395ndash398
2 PhalkeS NickelO WalluscheckD HortigF OnoratiMCand ReuterG (2009) Retrotransposon silencing and telomereintegrity in somatic cells of Drosophila depends on the cytosine-5methyltransferase DNMT2 Nat Genet 41 696ndash702
3 WilkinsonCR BartlettR NurseP and BirdAP (1995) Thefission yeast gene pmt1+ encodes a DNA methyltransferasehomologue Nucleic Acids Res 23 203ndash210
4 KatohM CurkT XuQ ZupanB KuspaA and ShaulskyG(2006) Developmentally regulated DNA methylation inDictyostelium discoideum Eukaryot Cell 5 18ndash25
5 KuhlmannM BorisovaBE KallerM LarssonP StachDNaJ EichingerL LykoF AmbrosV SoderbomF et al(2005) Silencing of retrotransposons in Dictyostelium by DNAmethylation and RNAi Nucleic Acids Res 33 6405ndash6417
6 RaiK ChidesterS ZavalaCV ManosEJ JamesSRKarpfAR JonesDA and CairnsBR (2007) Dnmt2 functionsin the cytoplasm to promote liver brain and retina developmentin zebrafish Genes Dev 21 261ndash266
7 FisherO Siman-TovR and AnkriS (2006) Pleiotropicphenotype in Entamoeba histolytica overexpressing DNAmethyltransferase (Ehmeth) Mol Biochem Parasitol 147 48ndash54
8 DurdevicZ HannaK GoldB PollexT CherryS LykoFand SchaeferM (2013) Efficient RNA virus control in Drosophilarequires the RNA methyltransferase Dnmt2 EMBO Rep 14269ndash275
9 SchaeferM and LykoF (2010) Lack of evidence for DNAmethylation of Invader4 retroelements in Drosophila andimplications for Dnmt2-mediated epigenetic regulation NatGenet 42 920ndash921 author reply 921
10 KraussV and ReuterG (2011) DNA methylation in Drosophila -a critical evaluation Prog Mol Biol Transl Sci 101 177ndash191
11 RaddatzG GuzzardoPM OlovaN FantappieMRRamppM SchaeferM ReikW HannonGJ and LykoF(2013) Dnmt2-dependent methylomes lack defined DNAmethylation patterns Proc Natl Acad Sci USA 110 8627ndash8631
12 JeltschA NellenW and LykoF (2006) Two substrates arebetter than one dual specificities for Dnmt2 methyltransferasesTrends Biochem Sci 31 306ndash308
13 JurkowskiTP MeusburgerM PhalkeS HelmM NellenWReuterG and JeltschA (2008) Human DNMT2 methylatestRNA(Asp) molecules using a DNA methyltransferase-likecatalytic mechanism RNA 14 1663ndash1670
14 TovyA Siman TovR GaentzschR HelmM and AnkriS(2010) A new nuclear function of the Entamoeba histolyticaglycolytic enzyme enolase the metabolic regulation of cytosine-5methyltransferase 2 (Dnmt2) activity PLoS Pathog 6 e1000775
15 SchaeferM PollexT HannaK TuortoF MeusburgerMHelmM and LykoF (2010) RNA methylation by Dnmt2protects transfer RNAs against stress-induced cleavage GenesDev 24 1590ndash1595
16 TuortoF LiebersR MuschT SchaeferM HofmannSKellnerS FryeM HelmM StoecklinG and LykoF (2012)RNA cytosine methylation by Dnmt2 and NSun2 promotestRNA stability and protein synthesis Nat Struct Mol Biol 19900ndash905
17 BeckerM MullerS NellenW JurkowskiTP JeltschA andEhrenhofer-MurrayAE (2012) Pmt1 a Dnmt2 homolog inSchizosaccharomyces pombe mediates tRNA methylation inresponse to nutrient signaling Nucleic Acids Res 4011648ndash11658
18 TovyA HertzR Siman-TovR SyanS FaustD GuillenNand AnkriS (2011) Glucose starvation boosts Entamoebahistolytica virulence PLoS Negl Trop Dis 5 e1247
19 MotorinY and HelmM (2011) RNA nucleotide methylationWiley Interdiscip Rev RNA 2 611ndash631
20 NiranjanakumariS LasdaE BrazasR and Garcia-BlancoMA(2002) Reversible cross-linking combined with
12 Nucleic Acids Research 2013
at Universitaumlt W
uumlrzburg on August 5 2013
httpnaroxfordjournalsorgD
ownloaded from
immunoprecipitation to study RNA-protein interactions in vivoMethods 26 182ndash190
21 RothbauerU ZolghadrK MuyldermansS SchepersACardosoMC and LeonhardtH (2008) A versatile nanotrapfor biochemical and functional studies with fluorescent fusionproteins Mol Cell Proteomics 7 282ndash289
22 SittkaA LucchiniS PapenfortK SharmaCM RolleKBinnewiesTT HintonJC and VogelJ (2008) Deep sequencinganalysis of small noncoding RNA and mRNA targets of theglobal post-transcriptional regulator Hfq PLoS Genet 4e1000163
23 HoffmannS OttoC KurtzS SharmaCM KhaitovichPVogelJ StadlerPF and HackermullerJ (2009) Fast mapping ofshort sequences with mismatches insertions and deletions usingindex structures PLoS Comput Biol 5 e1000502
24 ConnollyMA ClausenPA and LazarJG (2006) Purificationof RNA from animal cells using trizol Cold Spring HarborProtoc 2006 pii pdbprot4104
25 DongA YoderJA ZhangX ZhouL BestorTH andChengX (2001) Structure of human DNMT2 an enigmaticDNA methyltransferase homolog that displays denaturant-resistant binding to DNA Nucleic Acids Res 29 439ndash448
26 SchaeferM PollexT HannaK and LykoF (2009) RNAcytosine methylation analysis by bisulfite sequencing NucleicAcids Res 37 e12
27 BoninM OberstrassJ VogtU WasseneggerM and NellenW(2001) Binding of IRE-BP to its cognate RNA sequence SFMstudies on a universal RNA backbone for the analysis of RNA-protein interaction Biol Chem 382 1157ndash1162
28 ZimmermanW and WeijerCJ (1993) Analysis of cell cycleprogression during the development of Dictyostelium and itsrelationship to differentiation Dev Biol 160 178ndash185
29 StrasserK BloomfieldG MacWilliamsA CeccarelliAMacWilliamsH and TsangA (2012) A retinoblastomaorthologue is a major regulator of S-phase mitotic anddevelopmental gene expression in Dictyostelium PLoS One 7e39914
30 WattsDJ and AshworthJM (1970) Growth of myxameobae ofthe cellular slime mould Dictyostelium discoideum in axenicculture Biochem J 119 171ndash174
31 JurkowskiTP ShanmugamR HelmM and JeltschA (2012)Mapping the tRNA binding site on the surface of humanDNMT2 methyltransferase Biochemistry 51 4438ndash4444
32 SvedruzicZM and ReichNO (2005) DNA cytosine C5methyltransferase Dnmt1 catalysis-dependent release of allostericinhibition Biochemistry 44 9472ndash9485
33 ThomasSR KellerCA SzykA CannonJR and Laronde-LeblancNA (2011) Structural insight into the functionalmechanism of Nep1Emg1 N1-specific pseudouridinemethyltransferase in ribosome biogenesis Nucleic Acids Res 392445ndash2457
34 LeulliotN BohnsackMT GrailleM TollerveyD andVan TilbeurghH (2008) The yeast ribosome synthesisfactor Emg1 is a novel member of the superfamily of alphabeta knot fold methyltransferases Nucleic Acids Res 36629ndash639
35 WhiteJ LiZ SardanaR BujnickiJM MarcotteEM andJohnsonAW (2008) Bud23 methylates G1575 of 18S rRNA andis required for efficient nuclear export of pre-40S subunits MolCell Biol 28 3151ndash3161
36 SobalaA and HutvagnerG (2013) Small RNAs derived fromthe 50-end of tRNA can inhibit protein translation in humancells RNA Biol 10 525ndash535
Nucleic Acids Research 2013 13
at Universitaumlt W
uumlrzburg on August 5 2013
httpnaroxfordjournalsorgD
ownloaded from

heat shock cold shock anoxia and various nutrient mediabut did not observe altered levels Only during therecovery phase after cold treatment a transient almost50-fold increase in mRNA abundance was seen
Reuter and Ankri (personal communication) hadobserved a loss of Dnmt2 expression in laboratorystrains of D melanogaster and E histolytica We thereforeexamined D discoideum NC4 the parent of the axeniclaboratory strain AX2 The developmental regulation inAX2 and NC4 was qualitatively similar but expressionlevels were much higher in NC4 The observationsupports the assumption that Dnmt2 expression wasreduced under optimal growth conditions in the labora-tory strain and may actually be selected against
We show that the D discoideum Dnmt2 homologueDnmA is an active tRNAAsp(GUC) methyltransferasein vitro and in vivo To identify further potential targetsfor DnmA we established a UV-CLIP method to analyseDnmAndashRNA interaction in vivo In addition to the knowntarget tRNAAsp(GUC) fragments of tRNAGlu(UUC) andtRNAGly(GCC) were most abundant in comparison withthe control Unexpectedly complete tRNAs were notenriched in the DnmA CLIP fraction but rather specificlsquomid-fragmentsrsquo that covered the putative methylation siteand the recognition pattern proposed by Helm (personal
communication) A minor exception was tRNAGly(GCC)which lacks C40 We do not know whether these frag-ments were directly or indirectly generated by DnmAin vivo or whether they were protected by DnmA fromdegradation during the experimental procedure As noco-crystal of Dnmt2 and a tRNA substrate is availablethe exact interface for the interaction is unknownJurkowski et al (31) mapped the binding site on Dnmt2by mutational analysis A manual placement of a tRNAinto the crystal structure suggests that Dnmt2 mainlycontacts the anticodon stem loop Assuming that mid-frag-ments are footprints of DnmA our data support this notionbut suggest that the interaction surface extends into theD-loop which is partially covered by the mid-fragments
We examined the putative DnmA targets in vitro (bymethylation of in vitro transcribed tRNAs) ex vivo(by methylation of RNA isolated from D discoideum)and in vivo (by bisulfite sequencing) Furthermore weinvestigated by SDSndashPAGE the kinetics of covalentcomplex formation between tRNAAsp and tRNAGlu withDnmA and hDnmt2
All three tRNAs (tRNAAsp tRNAGlu tRNAGly) werereadily methylated by recombinant DnmA and hDnmt2in vitro although the activity was much higher (estimatedat least 10-fold) on tRNAAsp compared with tRNAGlu andtRNAGly For tRNAAsp and tRNAGlu this correlated wellwith the formation and decay of covalent RNAmdashproteincomplexes where kinetics for tRNAAsp were much fasterthan for tRNAGlu Instead of C methylation the covalentintermediate may also proceed to a hydrogen exchangereaction or to a deamination of the cytosine In bothcases an intermediate will be detected but it will notresult in a methylated product (32) It may be thatslower off rates represent these alternative pathways thatwould result in unmethylated tRNAs
Using antisense oligos we showed that tRNAAsptRNAGlu and at least one additional RNA could bemethylated ex vivo A quantification of the results wasnot possible the apparently low methylation oftRNAGlu ex vivo could be due to the low activity of theenzyme on this substrate (as suggested by the in vitroexperiments see Figure 5B)The data for the examination of in vivo methylation by
bisulfite sequencing were rather unexpected AlthoughtRNAAsp(GUC) was consistently found to be methylatedat C38 we never detected any methylation at C38 oftRNAGlu(UUCCUC) or tRNAGly(GCC) There could betwo reasons to explain the failure to detect C38 methyla-tion for these in vitro substrates (i) due to the low activityof DnmA on tRNAGlu(UUCCUC) and tRNAGly(GCC) asshown in vitro there was low or no significant methylationin vivo and (ii) methylation escaped detection becauseother unknown modifications blocked reverse transcrip-tion of the methylated molecules However if this wasthe case one has to assume that C49C50 methylationoccurs independently of these unknown modificationswhereas C38 methylation strictly depends on them AsC38 methylation does not require any other modificationsin vitro we believe that this scenario is less likely If in vitromethylation activity was comparable with in vivo methy-lation 10 methylation of tRNAGlu(UUCCUC) andtRNAGly(GCC) could be expected relative totRNAAsp(GUC) As tRNAAsp was methylated to 30 a3 methylation of tRNAGlu and tRNAGly could be belowthe detection level of the bisulfite assayWe find that high expression of dnmA did not necessar-
ily correlate with increased methylation of tRNAs Duringrecovery from cold shock D discoideum cells displayedthe highest level of dnmA expression but this had noeffect on tRNAAsp methylation Only in developmentand with the high expressing NC4 strain a significantincrease of tRNAAsp methylation to 70 was observedIt has to be considered however that bisulfite sequencingdoes not necessarily allow for quantitative evaluation ofdata detected C-methylation depends on reverse tran-scription efficiency which may be hampered by othermodifications that could be different under differentconditions Furthermore the completeness of bisulfite-mediated conversion could also depend on RNA struc-ture which may be altered by other modificationsHowever incomplete conversion will lead to an overesti-mation of C-methylation as maybe the case for forexample C30 C31 C35 in tRNAGly in some samplesTaken together the data from CLIP complex forma-
tion and methylation show that all three substrates can bebound and methylated in vitro The CLIP data show thatthere is a specific interaction of DnmA with tRNA mid-fragments of tRNAAsp(GUC) tRNAGlu(UUCCUC) andtRNAGly(GCC) but not with other tRNAs In contrastbisulfite sequencing could only detect methylation ontRNAAsp(GUC) even at the highest expression levels ofdnmA Even though low amounts of methylation mayhave escaped detection in bisulfite sequencing there isstill a discrepancy between dnmA expression levels (egduring cold shock recovery) DnmA binding to substrates(as determined by CLIP) and methylation in vivo
Nucleic Acids Research 2013 11
at Universitaumlt W
uumlrzburg on August 5 2013
httpnaroxfordjournalsorgD
ownloaded from
We therefore suggest that DnmA binding to specifictargets could elicit functions in addition to RNA methy-lation eg by promoting processing or degradation andor recruitment of other proteins At least for some rRNAmethyltransferases such as Bud23 or Nep1 fromSaccharomyces cerevisiae it is known that they executefunctions independent from the methylation activity(33ndash35) As shown by Schaefer and co-workers (15)methylation protects tRNAs from degradation or process-ing after stress By a different pathway binding of DnmAmay initiate processing that leads to specific tRNA frag-ments that could serve for other biological functions (36)Interestingly tRNAGlu(UUC) is also a substrate for theDnmt2 homologue Pmt1 in S pombe (17) HowevertRNAGlu(UUC) is found to be methylated to a low butsignificant extend in vivo in fission yeast when Pmt1 isoverexpressedThe detection of tRNAAla(AGC) fragments in the CLIP
assay brings up a new question tRNAAla(AGC) has a Tinstead of a C in position 38 and does not contain thesignature nucleotides in the anticodon stem-loopInstead of a mid-fragment a distinct 30 fragment wasassociated with DnmA suggesting that a recognitionmotif was located in this part of the moleculeAs methylation also occurred in a large artificial
molecule with embedded tRNAGlu (SupplementaryFigure S2) it could be that other longer RNAs containingan appropriate recognition motif may also be bound bythe enzyme We thus propose that the biological functionsof Dnmt2 proteins could extend far beyond our presentknowledge Investigation of further targets and of the sig-nificance of RNA fragments associated with Dnmt2should shed light on the importance of this enzyme
SUPPLEMENTARY DATA
Supplementary Data are available at NAR Online
ACKNOWLEDGEMENTS
The authors thank Thomas Winckler (Friedrich-Schiller-University Jena) for providing the tRNAGlu(UUC) and sup-pressor tRNAGlu(CUA) plasmids Susanne Junk for cloningtRNAAspC38A Matthias Schafer (DKFZ) and RobinLorenz (University of Kassel) are acknowledged forRNA bisulfite sequencing and Richard Reinhardt (MaxPlanck Genome Centre Cologne) for deep sequencingMark Helm (Johannes-Gutenberg-University Mainz) gen-erously provided data before publication CarstenSeehafer (University of Kassel) helped with the analysisof deep-sequencing data
FUNDING
Deutsche Forschungsgemeinschaft [FOR 1082 to WNand AJ] and the Studienstiftung des Deutschen Volkes(to SM) Funding for open access charge FOR1082
Conflict of interest statement None declared
REFERENCES
1 GollMG KirpekarF MaggertKA YoderJA HsiehCLZhangX GolicKG JacobsenSE and BestorTH (2006)Methylation of tRNAAsp by the DNA methyltransferasehomolog Dnmt2 Science 311 395ndash398
2 PhalkeS NickelO WalluscheckD HortigF OnoratiMCand ReuterG (2009) Retrotransposon silencing and telomereintegrity in somatic cells of Drosophila depends on the cytosine-5methyltransferase DNMT2 Nat Genet 41 696ndash702
3 WilkinsonCR BartlettR NurseP and BirdAP (1995) Thefission yeast gene pmt1+ encodes a DNA methyltransferasehomologue Nucleic Acids Res 23 203ndash210
4 KatohM CurkT XuQ ZupanB KuspaA and ShaulskyG(2006) Developmentally regulated DNA methylation inDictyostelium discoideum Eukaryot Cell 5 18ndash25
5 KuhlmannM BorisovaBE KallerM LarssonP StachDNaJ EichingerL LykoF AmbrosV SoderbomF et al(2005) Silencing of retrotransposons in Dictyostelium by DNAmethylation and RNAi Nucleic Acids Res 33 6405ndash6417
6 RaiK ChidesterS ZavalaCV ManosEJ JamesSRKarpfAR JonesDA and CairnsBR (2007) Dnmt2 functionsin the cytoplasm to promote liver brain and retina developmentin zebrafish Genes Dev 21 261ndash266
7 FisherO Siman-TovR and AnkriS (2006) Pleiotropicphenotype in Entamoeba histolytica overexpressing DNAmethyltransferase (Ehmeth) Mol Biochem Parasitol 147 48ndash54
8 DurdevicZ HannaK GoldB PollexT CherryS LykoFand SchaeferM (2013) Efficient RNA virus control in Drosophilarequires the RNA methyltransferase Dnmt2 EMBO Rep 14269ndash275
9 SchaeferM and LykoF (2010) Lack of evidence for DNAmethylation of Invader4 retroelements in Drosophila andimplications for Dnmt2-mediated epigenetic regulation NatGenet 42 920ndash921 author reply 921
10 KraussV and ReuterG (2011) DNA methylation in Drosophila -a critical evaluation Prog Mol Biol Transl Sci 101 177ndash191
11 RaddatzG GuzzardoPM OlovaN FantappieMRRamppM SchaeferM ReikW HannonGJ and LykoF(2013) Dnmt2-dependent methylomes lack defined DNAmethylation patterns Proc Natl Acad Sci USA 110 8627ndash8631
12 JeltschA NellenW and LykoF (2006) Two substrates arebetter than one dual specificities for Dnmt2 methyltransferasesTrends Biochem Sci 31 306ndash308
13 JurkowskiTP MeusburgerM PhalkeS HelmM NellenWReuterG and JeltschA (2008) Human DNMT2 methylatestRNA(Asp) molecules using a DNA methyltransferase-likecatalytic mechanism RNA 14 1663ndash1670
14 TovyA Siman TovR GaentzschR HelmM and AnkriS(2010) A new nuclear function of the Entamoeba histolyticaglycolytic enzyme enolase the metabolic regulation of cytosine-5methyltransferase 2 (Dnmt2) activity PLoS Pathog 6 e1000775
15 SchaeferM PollexT HannaK TuortoF MeusburgerMHelmM and LykoF (2010) RNA methylation by Dnmt2protects transfer RNAs against stress-induced cleavage GenesDev 24 1590ndash1595
16 TuortoF LiebersR MuschT SchaeferM HofmannSKellnerS FryeM HelmM StoecklinG and LykoF (2012)RNA cytosine methylation by Dnmt2 and NSun2 promotestRNA stability and protein synthesis Nat Struct Mol Biol 19900ndash905
17 BeckerM MullerS NellenW JurkowskiTP JeltschA andEhrenhofer-MurrayAE (2012) Pmt1 a Dnmt2 homolog inSchizosaccharomyces pombe mediates tRNA methylation inresponse to nutrient signaling Nucleic Acids Res 4011648ndash11658
18 TovyA HertzR Siman-TovR SyanS FaustD GuillenNand AnkriS (2011) Glucose starvation boosts Entamoebahistolytica virulence PLoS Negl Trop Dis 5 e1247
19 MotorinY and HelmM (2011) RNA nucleotide methylationWiley Interdiscip Rev RNA 2 611ndash631
20 NiranjanakumariS LasdaE BrazasR and Garcia-BlancoMA(2002) Reversible cross-linking combined with
12 Nucleic Acids Research 2013
at Universitaumlt W
uumlrzburg on August 5 2013
httpnaroxfordjournalsorgD
ownloaded from
immunoprecipitation to study RNA-protein interactions in vivoMethods 26 182ndash190
21 RothbauerU ZolghadrK MuyldermansS SchepersACardosoMC and LeonhardtH (2008) A versatile nanotrapfor biochemical and functional studies with fluorescent fusionproteins Mol Cell Proteomics 7 282ndash289
22 SittkaA LucchiniS PapenfortK SharmaCM RolleKBinnewiesTT HintonJC and VogelJ (2008) Deep sequencinganalysis of small noncoding RNA and mRNA targets of theglobal post-transcriptional regulator Hfq PLoS Genet 4e1000163
23 HoffmannS OttoC KurtzS SharmaCM KhaitovichPVogelJ StadlerPF and HackermullerJ (2009) Fast mapping ofshort sequences with mismatches insertions and deletions usingindex structures PLoS Comput Biol 5 e1000502
24 ConnollyMA ClausenPA and LazarJG (2006) Purificationof RNA from animal cells using trizol Cold Spring HarborProtoc 2006 pii pdbprot4104
25 DongA YoderJA ZhangX ZhouL BestorTH andChengX (2001) Structure of human DNMT2 an enigmaticDNA methyltransferase homolog that displays denaturant-resistant binding to DNA Nucleic Acids Res 29 439ndash448
26 SchaeferM PollexT HannaK and LykoF (2009) RNAcytosine methylation analysis by bisulfite sequencing NucleicAcids Res 37 e12
27 BoninM OberstrassJ VogtU WasseneggerM and NellenW(2001) Binding of IRE-BP to its cognate RNA sequence SFMstudies on a universal RNA backbone for the analysis of RNA-protein interaction Biol Chem 382 1157ndash1162
28 ZimmermanW and WeijerCJ (1993) Analysis of cell cycleprogression during the development of Dictyostelium and itsrelationship to differentiation Dev Biol 160 178ndash185
29 StrasserK BloomfieldG MacWilliamsA CeccarelliAMacWilliamsH and TsangA (2012) A retinoblastomaorthologue is a major regulator of S-phase mitotic anddevelopmental gene expression in Dictyostelium PLoS One 7e39914
30 WattsDJ and AshworthJM (1970) Growth of myxameobae ofthe cellular slime mould Dictyostelium discoideum in axenicculture Biochem J 119 171ndash174
31 JurkowskiTP ShanmugamR HelmM and JeltschA (2012)Mapping the tRNA binding site on the surface of humanDNMT2 methyltransferase Biochemistry 51 4438ndash4444
32 SvedruzicZM and ReichNO (2005) DNA cytosine C5methyltransferase Dnmt1 catalysis-dependent release of allostericinhibition Biochemistry 44 9472ndash9485
33 ThomasSR KellerCA SzykA CannonJR and Laronde-LeblancNA (2011) Structural insight into the functionalmechanism of Nep1Emg1 N1-specific pseudouridinemethyltransferase in ribosome biogenesis Nucleic Acids Res 392445ndash2457
34 LeulliotN BohnsackMT GrailleM TollerveyD andVan TilbeurghH (2008) The yeast ribosome synthesisfactor Emg1 is a novel member of the superfamily of alphabeta knot fold methyltransferases Nucleic Acids Res 36629ndash639
35 WhiteJ LiZ SardanaR BujnickiJM MarcotteEM andJohnsonAW (2008) Bud23 methylates G1575 of 18S rRNA andis required for efficient nuclear export of pre-40S subunits MolCell Biol 28 3151ndash3161
36 SobalaA and HutvagnerG (2013) Small RNAs derived fromthe 50-end of tRNA can inhibit protein translation in humancells RNA Biol 10 525ndash535
Nucleic Acids Research 2013 13
at Universitaumlt W
uumlrzburg on August 5 2013
httpnaroxfordjournalsorgD
ownloaded from

We therefore suggest that DnmA binding to specifictargets could elicit functions in addition to RNA methy-lation eg by promoting processing or degradation andor recruitment of other proteins At least for some rRNAmethyltransferases such as Bud23 or Nep1 fromSaccharomyces cerevisiae it is known that they executefunctions independent from the methylation activity(33ndash35) As shown by Schaefer and co-workers (15)methylation protects tRNAs from degradation or process-ing after stress By a different pathway binding of DnmAmay initiate processing that leads to specific tRNA frag-ments that could serve for other biological functions (36)Interestingly tRNAGlu(UUC) is also a substrate for theDnmt2 homologue Pmt1 in S pombe (17) HowevertRNAGlu(UUC) is found to be methylated to a low butsignificant extend in vivo in fission yeast when Pmt1 isoverexpressedThe detection of tRNAAla(AGC) fragments in the CLIP
assay brings up a new question tRNAAla(AGC) has a Tinstead of a C in position 38 and does not contain thesignature nucleotides in the anticodon stem-loopInstead of a mid-fragment a distinct 30 fragment wasassociated with DnmA suggesting that a recognitionmotif was located in this part of the moleculeAs methylation also occurred in a large artificial
molecule with embedded tRNAGlu (SupplementaryFigure S2) it could be that other longer RNAs containingan appropriate recognition motif may also be bound bythe enzyme We thus propose that the biological functionsof Dnmt2 proteins could extend far beyond our presentknowledge Investigation of further targets and of the sig-nificance of RNA fragments associated with Dnmt2should shed light on the importance of this enzyme
SUPPLEMENTARY DATA
Supplementary Data are available at NAR Online
ACKNOWLEDGEMENTS
The authors thank Thomas Winckler (Friedrich-Schiller-University Jena) for providing the tRNAGlu(UUC) and sup-pressor tRNAGlu(CUA) plasmids Susanne Junk for cloningtRNAAspC38A Matthias Schafer (DKFZ) and RobinLorenz (University of Kassel) are acknowledged forRNA bisulfite sequencing and Richard Reinhardt (MaxPlanck Genome Centre Cologne) for deep sequencingMark Helm (Johannes-Gutenberg-University Mainz) gen-erously provided data before publication CarstenSeehafer (University of Kassel) helped with the analysisof deep-sequencing data
FUNDING
Deutsche Forschungsgemeinschaft [FOR 1082 to WNand AJ] and the Studienstiftung des Deutschen Volkes(to SM) Funding for open access charge FOR1082
Conflict of interest statement None declared
REFERENCES
1 GollMG KirpekarF MaggertKA YoderJA HsiehCLZhangX GolicKG JacobsenSE and BestorTH (2006)Methylation of tRNAAsp by the DNA methyltransferasehomolog Dnmt2 Science 311 395ndash398
2 PhalkeS NickelO WalluscheckD HortigF OnoratiMCand ReuterG (2009) Retrotransposon silencing and telomereintegrity in somatic cells of Drosophila depends on the cytosine-5methyltransferase DNMT2 Nat Genet 41 696ndash702
3 WilkinsonCR BartlettR NurseP and BirdAP (1995) Thefission yeast gene pmt1+ encodes a DNA methyltransferasehomologue Nucleic Acids Res 23 203ndash210
4 KatohM CurkT XuQ ZupanB KuspaA and ShaulskyG(2006) Developmentally regulated DNA methylation inDictyostelium discoideum Eukaryot Cell 5 18ndash25
5 KuhlmannM BorisovaBE KallerM LarssonP StachDNaJ EichingerL LykoF AmbrosV SoderbomF et al(2005) Silencing of retrotransposons in Dictyostelium by DNAmethylation and RNAi Nucleic Acids Res 33 6405ndash6417
6 RaiK ChidesterS ZavalaCV ManosEJ JamesSRKarpfAR JonesDA and CairnsBR (2007) Dnmt2 functionsin the cytoplasm to promote liver brain and retina developmentin zebrafish Genes Dev 21 261ndash266
7 FisherO Siman-TovR and AnkriS (2006) Pleiotropicphenotype in Entamoeba histolytica overexpressing DNAmethyltransferase (Ehmeth) Mol Biochem Parasitol 147 48ndash54
8 DurdevicZ HannaK GoldB PollexT CherryS LykoFand SchaeferM (2013) Efficient RNA virus control in Drosophilarequires the RNA methyltransferase Dnmt2 EMBO Rep 14269ndash275
9 SchaeferM and LykoF (2010) Lack of evidence for DNAmethylation of Invader4 retroelements in Drosophila andimplications for Dnmt2-mediated epigenetic regulation NatGenet 42 920ndash921 author reply 921
10 KraussV and ReuterG (2011) DNA methylation in Drosophila -a critical evaluation Prog Mol Biol Transl Sci 101 177ndash191
11 RaddatzG GuzzardoPM OlovaN FantappieMRRamppM SchaeferM ReikW HannonGJ and LykoF(2013) Dnmt2-dependent methylomes lack defined DNAmethylation patterns Proc Natl Acad Sci USA 110 8627ndash8631
12 JeltschA NellenW and LykoF (2006) Two substrates arebetter than one dual specificities for Dnmt2 methyltransferasesTrends Biochem Sci 31 306ndash308
13 JurkowskiTP MeusburgerM PhalkeS HelmM NellenWReuterG and JeltschA (2008) Human DNMT2 methylatestRNA(Asp) molecules using a DNA methyltransferase-likecatalytic mechanism RNA 14 1663ndash1670
14 TovyA Siman TovR GaentzschR HelmM and AnkriS(2010) A new nuclear function of the Entamoeba histolyticaglycolytic enzyme enolase the metabolic regulation of cytosine-5methyltransferase 2 (Dnmt2) activity PLoS Pathog 6 e1000775
15 SchaeferM PollexT HannaK TuortoF MeusburgerMHelmM and LykoF (2010) RNA methylation by Dnmt2protects transfer RNAs against stress-induced cleavage GenesDev 24 1590ndash1595
16 TuortoF LiebersR MuschT SchaeferM HofmannSKellnerS FryeM HelmM StoecklinG and LykoF (2012)RNA cytosine methylation by Dnmt2 and NSun2 promotestRNA stability and protein synthesis Nat Struct Mol Biol 19900ndash905
17 BeckerM MullerS NellenW JurkowskiTP JeltschA andEhrenhofer-MurrayAE (2012) Pmt1 a Dnmt2 homolog inSchizosaccharomyces pombe mediates tRNA methylation inresponse to nutrient signaling Nucleic Acids Res 4011648ndash11658
18 TovyA HertzR Siman-TovR SyanS FaustD GuillenNand AnkriS (2011) Glucose starvation boosts Entamoebahistolytica virulence PLoS Negl Trop Dis 5 e1247
19 MotorinY and HelmM (2011) RNA nucleotide methylationWiley Interdiscip Rev RNA 2 611ndash631
20 NiranjanakumariS LasdaE BrazasR and Garcia-BlancoMA(2002) Reversible cross-linking combined with
12 Nucleic Acids Research 2013
at Universitaumlt W
uumlrzburg on August 5 2013
httpnaroxfordjournalsorgD
ownloaded from
immunoprecipitation to study RNA-protein interactions in vivoMethods 26 182ndash190
21 RothbauerU ZolghadrK MuyldermansS SchepersACardosoMC and LeonhardtH (2008) A versatile nanotrapfor biochemical and functional studies with fluorescent fusionproteins Mol Cell Proteomics 7 282ndash289
22 SittkaA LucchiniS PapenfortK SharmaCM RolleKBinnewiesTT HintonJC and VogelJ (2008) Deep sequencinganalysis of small noncoding RNA and mRNA targets of theglobal post-transcriptional regulator Hfq PLoS Genet 4e1000163
23 HoffmannS OttoC KurtzS SharmaCM KhaitovichPVogelJ StadlerPF and HackermullerJ (2009) Fast mapping ofshort sequences with mismatches insertions and deletions usingindex structures PLoS Comput Biol 5 e1000502
24 ConnollyMA ClausenPA and LazarJG (2006) Purificationof RNA from animal cells using trizol Cold Spring HarborProtoc 2006 pii pdbprot4104
25 DongA YoderJA ZhangX ZhouL BestorTH andChengX (2001) Structure of human DNMT2 an enigmaticDNA methyltransferase homolog that displays denaturant-resistant binding to DNA Nucleic Acids Res 29 439ndash448
26 SchaeferM PollexT HannaK and LykoF (2009) RNAcytosine methylation analysis by bisulfite sequencing NucleicAcids Res 37 e12
27 BoninM OberstrassJ VogtU WasseneggerM and NellenW(2001) Binding of IRE-BP to its cognate RNA sequence SFMstudies on a universal RNA backbone for the analysis of RNA-protein interaction Biol Chem 382 1157ndash1162
28 ZimmermanW and WeijerCJ (1993) Analysis of cell cycleprogression during the development of Dictyostelium and itsrelationship to differentiation Dev Biol 160 178ndash185
29 StrasserK BloomfieldG MacWilliamsA CeccarelliAMacWilliamsH and TsangA (2012) A retinoblastomaorthologue is a major regulator of S-phase mitotic anddevelopmental gene expression in Dictyostelium PLoS One 7e39914
30 WattsDJ and AshworthJM (1970) Growth of myxameobae ofthe cellular slime mould Dictyostelium discoideum in axenicculture Biochem J 119 171ndash174
31 JurkowskiTP ShanmugamR HelmM and JeltschA (2012)Mapping the tRNA binding site on the surface of humanDNMT2 methyltransferase Biochemistry 51 4438ndash4444
32 SvedruzicZM and ReichNO (2005) DNA cytosine C5methyltransferase Dnmt1 catalysis-dependent release of allostericinhibition Biochemistry 44 9472ndash9485
33 ThomasSR KellerCA SzykA CannonJR and Laronde-LeblancNA (2011) Structural insight into the functionalmechanism of Nep1Emg1 N1-specific pseudouridinemethyltransferase in ribosome biogenesis Nucleic Acids Res 392445ndash2457
34 LeulliotN BohnsackMT GrailleM TollerveyD andVan TilbeurghH (2008) The yeast ribosome synthesisfactor Emg1 is a novel member of the superfamily of alphabeta knot fold methyltransferases Nucleic Acids Res 36629ndash639
35 WhiteJ LiZ SardanaR BujnickiJM MarcotteEM andJohnsonAW (2008) Bud23 methylates G1575 of 18S rRNA andis required for efficient nuclear export of pre-40S subunits MolCell Biol 28 3151ndash3161
36 SobalaA and HutvagnerG (2013) Small RNAs derived fromthe 50-end of tRNA can inhibit protein translation in humancells RNA Biol 10 525ndash535
Nucleic Acids Research 2013 13
at Universitaumlt W
uumlrzburg on August 5 2013
httpnaroxfordjournalsorgD
ownloaded from

immunoprecipitation to study RNA-protein interactions in vivoMethods 26 182ndash190
21 RothbauerU ZolghadrK MuyldermansS SchepersACardosoMC and LeonhardtH (2008) A versatile nanotrapfor biochemical and functional studies with fluorescent fusionproteins Mol Cell Proteomics 7 282ndash289
22 SittkaA LucchiniS PapenfortK SharmaCM RolleKBinnewiesTT HintonJC and VogelJ (2008) Deep sequencinganalysis of small noncoding RNA and mRNA targets of theglobal post-transcriptional regulator Hfq PLoS Genet 4e1000163
23 HoffmannS OttoC KurtzS SharmaCM KhaitovichPVogelJ StadlerPF and HackermullerJ (2009) Fast mapping ofshort sequences with mismatches insertions and deletions usingindex structures PLoS Comput Biol 5 e1000502
24 ConnollyMA ClausenPA and LazarJG (2006) Purificationof RNA from animal cells using trizol Cold Spring HarborProtoc 2006 pii pdbprot4104
25 DongA YoderJA ZhangX ZhouL BestorTH andChengX (2001) Structure of human DNMT2 an enigmaticDNA methyltransferase homolog that displays denaturant-resistant binding to DNA Nucleic Acids Res 29 439ndash448
26 SchaeferM PollexT HannaK and LykoF (2009) RNAcytosine methylation analysis by bisulfite sequencing NucleicAcids Res 37 e12
27 BoninM OberstrassJ VogtU WasseneggerM and NellenW(2001) Binding of IRE-BP to its cognate RNA sequence SFMstudies on a universal RNA backbone for the analysis of RNA-protein interaction Biol Chem 382 1157ndash1162
28 ZimmermanW and WeijerCJ (1993) Analysis of cell cycleprogression during the development of Dictyostelium and itsrelationship to differentiation Dev Biol 160 178ndash185
29 StrasserK BloomfieldG MacWilliamsA CeccarelliAMacWilliamsH and TsangA (2012) A retinoblastomaorthologue is a major regulator of S-phase mitotic anddevelopmental gene expression in Dictyostelium PLoS One 7e39914
30 WattsDJ and AshworthJM (1970) Growth of myxameobae ofthe cellular slime mould Dictyostelium discoideum in axenicculture Biochem J 119 171ndash174
31 JurkowskiTP ShanmugamR HelmM and JeltschA (2012)Mapping the tRNA binding site on the surface of humanDNMT2 methyltransferase Biochemistry 51 4438ndash4444
32 SvedruzicZM and ReichNO (2005) DNA cytosine C5methyltransferase Dnmt1 catalysis-dependent release of allostericinhibition Biochemistry 44 9472ndash9485
33 ThomasSR KellerCA SzykA CannonJR and Laronde-LeblancNA (2011) Structural insight into the functionalmechanism of Nep1Emg1 N1-specific pseudouridinemethyltransferase in ribosome biogenesis Nucleic Acids Res 392445ndash2457
34 LeulliotN BohnsackMT GrailleM TollerveyD andVan TilbeurghH (2008) The yeast ribosome synthesisfactor Emg1 is a novel member of the superfamily of alphabeta knot fold methyltransferases Nucleic Acids Res 36629ndash639
35 WhiteJ LiZ SardanaR BujnickiJM MarcotteEM andJohnsonAW (2008) Bud23 methylates G1575 of 18S rRNA andis required for efficient nuclear export of pre-40S subunits MolCell Biol 28 3151ndash3161
36 SobalaA and HutvagnerG (2013) Small RNAs derived fromthe 50-end of tRNA can inhibit protein translation in humancells RNA Biol 10 525ndash535
Nucleic Acids Research 2013 13
at Universitaumlt W
uumlrzburg on August 5 2013
httpnaroxfordjournalsorgD
ownloaded from
Copyright © 2022 FDOKUMEN




















