
Taking organelles apart, putting them back together and creating new ones: Lessons from the...
48
Progress in Histochemistry and Cytochemistry 46 (2011) 1–48 PROGRESS IN HISTOCHEMISTRY AND CYTOCHEMISTRY www.elsevier.de/proghi Available online at www.sciencedirect.com Review Taking organelles apart, putting them back together and creating new ones: Lessons from the endoplasmic reticulum Christine Lavoie a , Line Roy b , Joël Lanoix c , Mariam Taheri d , Robin Young d , Geneviève Thibault d , Carol Abi Farah e , Nicole Leclerc d , Jacques Paiement d,∗ a Département de pharmacologie, Faculté de médecine, Université de Sherbrooke, Sherbrooke, QC, Canada b Proteomics Platform, Genome Quebec, Montreal, Quebec, Canada c Caprion Proteomics Inc., Montreal, Canada d Département de pathologie et biologie cellulaire, Université de Montréal, Montréal, Canada e Department of Neurology and Neurosurgery, Montreal Neurological Institute, McGill University, Montreal, Quebec, Canada Accepted 2 April 2011 Abstract The endoplasmic reticulum (ER) is a highly dynamic organelle. It is composed of four subcom- partments including nuclear envelope (NE), rough ER (rER), smooth ER (sER) and transitional ER Abbreviations: AAA, ATPase associated with different cellular activities; ATPase, adenosine triphosphatase; AP, acute pancreatitis; BiP, 78-kDa glucose-regulated protein; CoA, coenzyme A; COPI, coatomer protein I; COPII, coatomer protein II; CMP, cytosine monophosphate; CTP, cytosine triphosphate; ER, endoplasmic reticu- lum; ERGIC, ER-Golgi intermediate compartment; GTPS, guanosine 5 -3-O-(thio)triphosphate; LDM, smooth microsomes and low density rough microsomes; NE, nuclear envelope; p97, valosin-containing protein; PDI, pro- tein disulfide isomerase; PUFAs, polyunsaturated free fatty acids; rER, rough endoplasmic reticulum; sER, smooth endoplasmic reticulum; SNARE, SNAP receptor; SRM, stripped rough microsomes; SRPr, signal recognition particle receptor beta; tER, transitional endoplasmic reticulum; VCP, valosin-containing protein; VTCs, vesicular tubular clusters. ∗ Corresponding author. Département de pathologie et biologie cellulaire, Université de Montréal, C.P. 6128, Succ. Centre-ville, Montréal, QC, Canada, H3C 3J7 Tel.: +514-343-7259; fax: +514-343-5755. E-mail address: [email protected] (J. Paiement). 0079-6336/$ – see front matter © 2011 Elsevier GmbH. All rights reserved. doi:10.1016/j.proghi.2011.04.001
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Taking organelles apart, putting them back together and creating new ones: Lessons from the...

Progress in Histochemistry and Cytochemistry 46 (2011) 1–48
PROGRESS IN HISTOCHEMISTRY
AND CYTOCHEMISTRY
www.elsevier.de/proghi
Available online at www.sciencedirect.com
Review
Taking organelles apart, putting them back together andcreating new ones: Lessons from the endoplasmic reticulum
Christine Lavoie a, Line Roy b, Joël Lanoix c, Mariam Taheri d,Robin Young d, Geneviève Thibault d, Carol Abi Farah e,Nicole Leclerc d, Jacques Paiement d,∗a Département de pharmacologie, Faculté de médecine, Université de Sherbrooke,Sherbrooke, QC, Canadab Proteomics Platform, Genome Quebec, Montreal, Quebec, Canadac Caprion Proteomics Inc., Montreal, Canadad Département de pathologie et biologie cellulaire, Université de Montréal, Montréal, Canadae Department of Neurology and Neurosurgery, Montreal Neurological Institute,McGill University, Montreal, Quebec, Canada
Accepted 2 April 2011
Abstract
The endoplasmic reticulum (ER) is a highly dynamic organelle. It is composed of four subcom-partments including nuclear envelope (NE), rough ER (rER), smooth ER (sER) and transitional ER
Abbreviations: AAA, ATPase associated with different cellular activities; ATPase, adenosine triphosphatase;AP, acute pancreatitis; BiP, 78-kDa glucose-regulated protein; CoA, coenzyme A; COPI, coatomer protein I;COPII, coatomer protein II; CMP, cytosine monophosphate; CTP, cytosine triphosphate; ER, endoplasmic reticu-lum; ERGIC, ER-Golgi intermediate compartment; GTP�S, guanosine 5′-3-O-(thio)triphosphate; LDM, smoothmicrosomes and low density rough microsomes; NE, nuclear envelope; p97, valosin-containing protein; PDI, pro-tein disulfide isomerase; PUFAs, polyunsaturated free fatty acids; rER, rough endoplasmic reticulum; sER, smoothendoplasmic reticulum; SNARE, SNAP receptor; SRM, stripped rough microsomes; SRPr�, signal recognitionparticle receptor beta; tER, transitional endoplasmic reticulum; VCP, valosin-containing protein; VTCs, vesiculartubular clusters.
∗ Corresponding author. Département de pathologie et biologie cellulaire, Université de Montréal, C.P. 6128,Succ. Centre-ville, Montréal, QC, Canada, H3C 3J7 Tel.: +514-343-7259; fax: +514-343-5755.
E-mail address: [email protected] (J. Paiement).
0079-6336/$ – see front matter © 2011 Elsevier GmbH. All rights reserved.doi:10.1016/j.proghi.2011.04.001

2 C. Lavoie et al. / Progress in Histochemistry and Cytochemistry 46 (2011) 1–48
(tER). The subcompartments are interconnected, can fragment and dissociate and are able to reassem-ble again. They coordinate with cell function by way of protein regulators in the surrounding cytosol.The activity of the many associated molecular machines of the ER as well as the fluid nature of thelimiting membrane of the ER contribute extensively to the dynamics of the ER. This review examinesthe properties of the ER that permit its isolation and purification and the physiological conditions thatpermit reconstitution both in vitro and in vivo in normal and in disease conditions.© 2011 Elsevier GmbH. All rights reserved.
Keywords: Endoplasmic reticulum; Subcellular fractionation; Cell-free assembly; In vitro and in vivoreconstitution; Electron microscopy; Cytochemistry; Immunocytochemistry; Organelle transplanta-tion; Organelle proteomics; Transformed membranes; Cancer biomarkers
Contents
1. Introduction . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 32. Taking apart the ER: subcellular fractionation of the ER . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 33. Taking apart the ER to see if it is well or not: the proteome of the ER in health and
in disease . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 44. Putting the ER back together after fragmentation and isolation from the cellular milieu . . 65. Reconstitution of the ER: Nuclear envelope (NE) assembly . . . . . . . . . . . . . . . . . . . . . . . . . . . 7
5.1. In vitro assays which helped define the steps in NE assembly . . . . . . . . . . . . . . . . . . . 75.2. Fusion of outer NE membranes with ER membranes . . . . . . . . . . . . . . . . . . . . . . . . . . . 95.3. Fusion of outer NE membranes . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 95.4. Interaction between outer NE membranes and microtubules . . . . . . . . . . . . . . . . . . . . 9
6. Reconstitution of the ER: Fusion of rough ER (rER) membranes . . . . . . . . . . . . . . . . . . . . . 126.1. Role of ribosomes in rER assembly . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 146.2. Lipids involved in rER assembly and lipid droplet formation: Creating a new
organelle . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 156.3. Role of G-proteins in rER assembly . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 18
6.3.1. A ras-like GTPase involved in rER membrane fusion . . . . . . . . . . . . . . . . . 196.3.2. Signal recognition particle receptor beta (SRPr�) and ER assembly . . . . 19
7. Reconstitution of the ER: Fusion of smooth ER (sER) membranes . . . . . . . . . . . . . . . . . . . . 277.1. Role of the ATPase p97 and the SNARE protein syntaxin 5 in the assembly of
sER membranes . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 317.2. Summary of molecular factors involved in the assembly of rough ER cisternae
and smooth ER tubules . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 338. sER assembled in vitro is similar to transitional ER (tER). . . . . . . . . . . . . . . . . . . . . . . . . . . . 34
8.1. Formation of ER exit sites . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 348.2. Formation of ER exit sites from tER in vitro: Role of COPI . . . . . . . . . . . . . . . . . . . . 358.3. Formation of ER exit sites from tER in vitro: Studies with COPII . . . . . . . . . . . . . . 37
9. ER reconstitution in live cells . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 389.1. In vivo reconstitution of the rER . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 389.2. In vivo reconstitution of the sER . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 409.3. In vivo reconstitution of the ER in dividing cells . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 42
10. Concluding remarks . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 43Acknowledgements . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 44References . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 44

C. Lavoie et al. / Progress in Histochemistry and Cytochemistry 46 (2011) 1–48 3
1. Introduction
Electron microscopic radioautography, immunocytochemistry and cell fractionation pro-vided key experiments that first defined the secretory pathway. From initial experimentscarried out in a variety of laboratories around the world we now know that secretory pro-teins are synthesized in the ER in association with ribosomes, are transported to the Golgicomplex, and then are packaged into secretion granules which ultimately fuse with theplasma membrane and release their content to the extracellular milieu (Revel and Hay,1963; Warshawsky et al., 1963; Caro and Palade, 1964; Nadler et al., 1964; Whur et al.,1969; Young and Droz, 1968; Ashley and Peters, 1969; Glaumann and Ericsson, 1970;Meldolesi et al., 1971; Roth et al., 1981; Bendayan et al., 1986). To explain protein trans-port between the various compartments of the secretory pathway Palade (1975) suggestedthat transport of secretory proteins between these distinct organelle compartments occursvia vesicular carriers which implied the existence of membrane traffic between the vari-ous membrane compartments involved. In other words, transfer from the ER to the Golgirequires the formation of a transport vesicle by a budding event at the “donor” organelle(ER) and the subsequent fusion of the carrier at the “acceptor” organelle (the Golgi).
Experiments in which the vectorial transport of proteins into the lumen of the ER werereconstituted represented the first true in vitro reconstitutions of complex biological activityrelated to membrane traffic (Sabatini and Blobel, 1970; Blobel and Dobberstein, 1975a andb). Such “in vitro assays” facilitated identification of important protein components involvedin these processes, components whose eventual physiological relevance was confirmed bygenetic analysis of the same processes in yeast (Deshaies and Schekman, 1987).
Today, in vitro reconstitution is among the most important and widely used methodsin studying membrane traffic. Cell-free reconstitution is now often used in combinationwith morphological, cytochemical, genetic, and molecular biological approaches. In vitroreconstitution is providing not only information on protein sorting, for example acrossthe ER membrane but also information on processes such as membrane fusion, vesicleformation and organelle biogenesis (Mellman and Warren, 2000).
Thus taking organelles apart allows the means to isolate and purify their derivatives fromtheir complex intracellular environments. Once isolated the organelle derivatives can befurther analyzed for content and in addition can serve as source of material for reconstitutionof organelle structure and function in a cell-free microenvironment using in vitro incubation.
2. Taking apart the ER: subcellular fractionation of the ER
The biochemical study of ER derivatives started in the 1930 s. Albert Claude by dif-ferential centrifugation isolated a subcellular fraction made up of very small particles(Claude, 1938) later called microsomes (Claude, 1943). Rough vesicles, smooth vesiclesand RNA particles (ribosomes) were described in this fraction by Palade and Siekevitz(Palade and Siekevitz, 1956a, 1956b). Initially microsomes were subfractionated by a vari-ety of techniques including by differential centrifugation in homogeneous medium, througha stabilizing density gradient or through sucrose layers and by isopycnic equilibration in

4 C. Lavoie et al. / Progress in Histochemistry and Cytochemistry 46 (2011) 1–48
density gradients (reviewed in Beaufay et al., 1974a). Differential centrifugation of tissuhomogenates leads to the production of two main particulate fractions: a large granules ormitochondria fraction and a small granules or microsomal fraction. Density gradient cen-trifugation of the microsomal fraction yields rough microsomes and smooth microsomeswith the rough microsomes characterized by high content of the enzyme markers for the ERand the smooth microsomes characterized by the additional content of the enzyme markersfor mitochondria, Golgi and plasma membrane (Beaufay et al., 1974a). Rough microsomesbecause of their associated content of RNA have equilibrium densities greater than those ofsmooth microsomes and this property has been used to advantage to isolate and characterizea spectrum of rough and smooth microsomes by physical, chemical and biochemical assays(Beaufay et al., 1974b). A two step procedure involving differential centrifugation (to sepa-rate large granules from microsomes) and ultracentrifugation of the microsomes through adiscontinuous sucrose gradient was adapted from several earlier methods (Paiement et al.,1980; Paiement and Bergeron, 1983; Lavoie et al., 1996) to separate rough microsomesfrom smooth microsomes (Fig. 1). Further characterization of the smooth microsomal frac-tion by in vitro assay indicated that these constitute a subcompartment of the transitionalendoplasmic reticulum of hepatocytes (Lavoie et al., 1996; Lavoie et al., 1999; Roy et al.,2000).
3. Taking apart the ER to see if it is well or not: the proteome of theER in health and in disease
The protein content of each of the subcompartments of the ER has been studied wellusing subcellular fractions from normal cells and organelle proteomics (reviewed in Lavoieand Paiement, 2008 and in Chen et al., 2010a). In contrast the protein content of similar ERsubcompartments from cells in disease states has not been studied as well. Indeed it is nowappropriate to study ER composition in disease tissues because the ER is being implicated asa key determinant in an increasingly greater number of human pathologies. For example theER has been implicated in carcinogenesis and is now being targeted for cancer treatment(Healy et al., 2009; Wlodkowic et al., 2009; Hoeller and Dikic, 2009). Other diseasesin which the ER has been implicated as a key contributor to malfunction include diabetes(Fonseca et al., 2010), neurodegeneration (Matus et al., 2011), arthrosclerosis (Tabas, 2010),pancreatitis (Chen et al., 2010b) and heart disease (Prins and Michalak, 2009). With regardsto the NE subcompartment of the ER many inherited diseases and syndromes have beenlinked to lamin A and/or C and certain associated NE transmembrane proteins (Schirmerand Gerace, 2005). Of course in all these disease states ER can be studied by a multitude ofapproaches including organelle proteomics. The advantage of this approach is that it allowssimultaneous examination of numerous members of protein families and complete proteincontent of molecular machines which are involved in ER function.
Endoplasmic reticulum protein families and molecular machines are now being exam-ined using organelle proteomics in both ER from tissues after drug treatment and in ER fromdisease tissues. Galeva and Altermann (2002) have used proteomics to study the response ofcytochrome p450 family members after drug treatment and observed differential expressionof cytochrome p450 2B1 and 2B2. In addition proteomics permitted these authors to show

C. Lavoie et al. / Progress in Histochemistry and Cytochemistry 46 (2011) 1–48 5
Fig. 1. Isolation of rough and smooth microsomes. (A) Subcellular fractionation protocol to purifyrough and smooth microsomal fractions from rat liver. LSS: low speed supernatant; N: cellulardebris and nuclei; ML: mitochondria and lysosome; ISS: intermediate-speed supernatant; TM: totalmicrosomes; S: supernatant. (B-C) Electron micrographs of purified rough and smooth microsomalfractions. (B) Rough microsomes contain more than 80% rough vesicles as determined by quantifi-cation of the number of vesicles with associated ribosomes. (C) Smooth microsomes contain 60%smooth microsomal vesicles and 40% rough vesicles which have few (one to five) ribosomes asso-ciated with their membranes. Both microsomal fractions are enriched in enzyme markers for the ERand reduced in enzyme marker of the Golgi and plasma membrane derivatives. (Figures modifiedfrom Paiement et al. (2006).
that protein disulfide isomerase A3 and A6, and 78-kDa glucose-regulated protein wereoverexpressed following phenobarbitol treatment. Already in these earlier studies Galevaand Altermann (2002) concluded that proteomics has the advantage of detecting the responseof multiple protein families to drug treatment. Morand et al. (2005) used proteomics analysisof ER microsomes from the liver of a fructose-fed, insulin-resistant hamster model and wereable to show overexpression of chaperones (GRP94 and PDI) in this animal model. Theauthors concluded that there is a close association between acquisition of insulin-resistance

6 C. Lavoie et al. / Progress in Histochemistry and Cytochemistry 46 (2011) 1–48
and cellular stress as defined by the unfolded protein response. Chen et al. (2010b) quan-titatively compared the protein compositions of pancreatic rER between normal and acutepancriatitis (AP) animals and observed a great decrease in amount of digestive enzymes,little change in chaperone content and an increase in proteins involved in translation andtranslocation in rER from arginine-induced AP animals. The authors concluded that earlystages of AP involve changes in multiple rER proteins that may affect the synthesis andprocessing of digestive enzymes. Interestingly the proteomic studies of Roy et al. (2010)of tER in hepatocellular carcinoma also revealed changes in proteins involved in proteintranslation. Whether the protein translation machinery of the ER is modified in many diseasestates is worthy of further analysis.
The studies of tER in hepatocellular carcinoma by Roy et al. (2010) revealed changesin secretory proteins, protein translation as well as in protein secretion. Proteins of theCOPII protein complex as well as proteins of the dynein/dynactin protein complexes wereobserved overexpressed in tER from tumor nodules. Transport vesicles are formed at ERexit sites from COPII protein complex composed of Sar1p, Sec23p-Sec24p and Sec13p-Sec31p (Gürkan et al., 2006) and are subsequently translocated to the Golgi Apparatusalong microtubules by the dynein/dynactin protein complexes (Presley et al., 1997). Thusorganelle proteomics of tER fractions as applied by Roy et al. (2010) was able to detectoverexpression of proteins known to be involved in the assembly of transport vesicles as wellas proteins known to be involved in the translocation of these same structures. Althoughdata exists implicating COPII coupled systems in human health and disease (Routledgeet al., 2010) to the best of our knowledge no reports have appeared as of yet implicatingCOPII coupled systems in cancer.
4. Putting the ER back together after fragmentation and isolationfrom the cellular milieu
Following the description of the secretion pathway within cells (Palade, 1975) a varietyof in vitro assays were developed to determine the identity of the molecular components thatallowed membrane traffic between closed compartments. As mentioned above experimentsin which protein insertion into the ER was reproduced represented among the first in vitroreconstitutions of complex biological activity related to membrane traffic (Sabatini andBlobel, 1970; Blobel and Dobberstein, 1975a, b). Since then assays for the reconstitutionof membrane fusion in vitro have been described for ER-ER membranes (Paiement et al.,1980; Dabora and Sheetz, 1988; Kachar and Reese, 1988; Vale and Hotani, 1988; Watkinset al., 1993), for ER-NE membranes (Paiement, 1984; Latterich and Schekman, 1994), andfor NE-NE membranes (Paiement, 1981a, b; Vigers and Lohka, 1991; Boman et al., 1992;Newport and Dunphy, 1992).
Assays for the reconstitution of membrane fusion in vitro have been described for therough and smooth subcompartments of the ER. This phenomenon occurs at 37 ◦C withoutthe addition of cytoplasmic proteins. However, the nucleotides GTP and ATP have beenshown to be essential. GTP was shown to be required for the fusion of rough ER mem-branes (Paiement et al., 1980) and ATP was shown to be required for the fusion smooth ERmembranes (Lavoie et al., 1996). A GTPase involved ER fusion has recently been identified

C. Lavoie et al. / Progress in Histochemistry and Cytochemistry 46 (2011) 1–48 7
as atlastin (Orso et al., 2009) and an ATPase involved in smooth ER fusion was previouslyidentified as p97 (Roy et al., 2000).
Both in vitro and in vivo reconstitution approaches can be used to study organelleassembly and membrane traffic. Such reconstitutions are often used in combination withmorphological, cytochemical, genetic, and molecular biological approaches. Reconstitutionis providing not only information on protein sorting, for example across the ER membranebut also information on processes such as membrane fusion, vesicle formation, organellebiogenesis and membrane traffic. While cell-free reconstitution provides a powerful tool toidentify important protein components involved in each of these various processes, in vivoreconstitution such as that provided by cell transplantation via microinjection help confirmin vitro phenomenon in the living cell. Similarly genetic manipulation of specific proteincomponents help confirm physiological relevance in the intact cell.
5. Reconstitution of the ER: Nuclear envelope (NE) assembly
The NE is composed of an inner nuclear membrane, which contains a specific subsetof transmembrane proteins, and an outer nuclear membrane which is continuous with theER (Hetzer, 2010). NE growth and surface expansion in interphase cells are thought tooccur via an input of lipids and resident proteins from the ER (Anderson and Hetzer, 2007).Input of these components can occur via lateral diffusion in the membrane from the ER tothe outer nuclear membrane. New inner nuclear membrane and outer nuclear membraneproteins are translated in the rough ER and diffuse throughout the ER network until theyreach the outer nuclear membrane. Proteins destined for the inner nuclear membrane dif-fuse passively through channels of nuclear pores (Soullam and Worman, 1995) or activelythrough nuclear pores via a process involving the Ran cycle and import factors (King et al.,2006). It is clear that continuity between ER and nuclear membranes plays an importantrole in NE biogenesis, what is less clear is how continuity between these two compartmentsis established and what are the regulating factors.
5.1. In vitro assays which helped define the steps in NE assembly
ER contributes to NE assembly in post-mitotic cells (Hetzer, 2010). Based purely on mor-phological considerations NE assembly was previously described as a sequence of eventsinvolving ER membrane interactions with nuclear chromatin (Paiement, 1984). Fig. 2 sum-marizes the steps which include: 1- ER and NE membrane fragments align themselves alongchromosomes forming flattened membranous cisternae, 2- the chromosomes aggregate andcoalesce to form a continuous nuclear substratum or nuclear shell, 3- lateral segments ofaligned membranous cisterns make contact and fuse with one another in a GTP-dependentmanner to form continous nuclear membrane shell with newly differentiated inner and outermembranes.
Membrane binding to nuclear chromatin as defined in Step 1 was first shown byusing demembranated Xenopus sperm and Xenopus oocyte vesicle fractions (Viger andLohka,1991). In this first study vesicle binding to chromatin was observed to be ATP-independent and membrane fusion at the surface of the nucleus required a second population

8 C. Lavoie et al. / Progress in Histochemistry and Cytochemistry 46 (2011) 1–48
Fig. 2. Diagrammatic representation of nuclear envelope formation in post-mitotic cells. ER mem-brane fragments (with few associated ribosomes and without pores shown on the top left) and nuclearenvelope membrane fragments (with residual pores (Hetzer, 2010), shown on the top right) at telophaseaggregate and align themselves as flattened cisterns along chromosomal masses (ch). The chromoso-mal masses then coalesce forming a nuclear shell (ns) delimited by closely apposed flattened cisternsof both ER and nuclear envelope origin. The aligned cisterns along the chromosomal masses createa polarity whereby that portion of the respective cisterns in contact with the nuclear shell becomesthe prospective inner membrane (im) of the nuclear envelope and that portion away from the chro-mosomes becomes the prospective outer membrane (om). Subsequent to establishment of membranecontact between the lateral segments (ls) of the juxtaposed cisterns GTP stimulates membrane fusionleading to the formation of a new and continuous nuclear envelope. Reprinted from Fig. 14 in Paiement(1984).
of vesicles and was stimulated by ATP. Newport and Dunphy (1992) used this same in vitromodel and observed specific binding of 70 nm-diameter vesicles to chromatin and sub-sequent fusion of these vesicles in the presence of ATP and GTP. Membrane binding tonuclear chromatin has since been shown to involve DNA-binding-NE-specific membraneproteins which are associated with post-mitotic ER membrane fragments (Ulbert et al.,2006; Anderson and Hetzer, 2007).
Chromatin changes alluded to in Step 2 include chromatin decondensation and thisprocess has recently been shown to be an ATPase Cdc48/p97-dependent process (Ramadanet al., 2007). This could be an important ATP-dependent step in nuclear NE assembly whichoccurs prior or coincident with membrane fusion?
The contribution of NE-NE membrane fusion to NE assembly (Step 3) is known to bea GTP-dependent event and the GTPase Ran has been implicated (Hetzer et al., 2000).Whether Ran promotes ER-NE fusion in interphase cells remains to be shown?

C. Lavoie et al. / Progress in Histochemistry and Cytochemistry 46 (2011) 1–48 9
5.2. Fusion of outer NE membranes with ER membranes
Nuclear envelope growth and surface expansion in interphase cells was suggested to occurvia an input of lipids and resident proteins from the ER. New inner nuclear membrane andouter nuclear membrane proteins are translated in the rough ER and diffuse throughout theER network until they reach the outer nuclear membrane (Anderson and Hetzer, 2007).Thus ER-NE membrane fusion is expected to occur often within interphase cells and by sodoing contribute to the renewal of newly synthesized NE membrane proteins. Assays forthe reconstitution of membrane fusion in vitro have been described for ER-NE membranes(Paiement, 1984; Latterich and Schekman, 1994). Fig. 3A shows the effect of incubatingnuclei with ER membrane derivatives. Incubation of isolated nuclei in the presence of roughmicrosomes stripped of associated ribosomes leads to the fusion of outer membranes ofnuclei with microsomes to form large irregular outer nuclear membrane extensions. Nucleiincubated in the presence of GTP but in the absence of ER membranes can undergo NE-NEmembrane fusion but they do not show formation of large nuclear membrane extentions(Fig. 3B). ER derivatives incubated in the presence of GTP but in the absence of nucleifuse to form vesicles with a heterogeneous population of sizes (Fig. 3C). Thus ER-NEmembrane fusion is a GTP-dependent event. The GTPase involved in this event has yet tobe identified?
5.3. Fusion of outer NE membranes
The incubation of isolated nuclei at 37 ◦C in the presence of physiological concentrationsof GTP leads to the formation of nuclear aggregates which include paired nuclei withcontinuous outer membranes (Paiement, 1981a, b; Paiement, 1984). Confluent perinuclearspaces and inverted vesicles attached to the inner nuclear membrane via nuclear poresare observed in the zone of contact between the fused nuclei (properties summarized inFig. 4). The formation of continuous outer nuclear membranes between two nuclei as wellas confluent perinuclear cisternae (spaces between vesicles in regions of nuclear contact) hasbeen reported during karyogamy of gametes (Jensen, 1964; Longo and Anderson, 1968;Longo, 1973; Iwamatsu and Kobayashi, 2002) and represents an early stage of gametefusion. The initial step of nuclear fusion between gametes has been well documented byelectron microscopy however little is known about the molecular mechanism involved inthis process and it remains to be shown whether a GTPase is involved?
5.4. Interaction between outer NE membranes and microtubules
The intracellular positioning and shape of the nucleus are determined by nuclear sur-face interaction with the cytoskeleton. The outer nuclear membrane proteins nesprin-1 andnesprin-2 interact with actin, nesprin-3 can connect the nucleus to the intermediate filamentnetwork via its interaction with plectin and the motor proteins kinesin-1 and dynein controlnuclear positioning and movement along microtubules in opposite directions (reviewed inFriedl et al., 2010). Cell polarity genes and microtubule-associated proteins have also beenimplicated in nuclear dynamics (Tsai and Gleeson, 2005).

10 C. Lavoie et al. / Progress in Histochemistry and Cytochemistry 46 (2011) 1–48
Fig. 3. In vitro reconstitution of NE assembly. (A) Fusion of outer NE membranes with ER mem-branes. Electron micrograph showing the periphery of a nucleus after incubation in the presence ofSRM and GTP. The lumina of large irregular membrane extentions is shown in regions confluent withthe perinuclear space (arrows). Arrowheads indicate points of deflection of the outer nuclear mem-brane to form the large irregular membrane extensions. The latter are observed in different sizes andin certain cases are associated with small regular unfused microsomal vesicles. dg, dense chromatingranules. Bar, 1 �m. (B) Fusion of outer NE membranes. Electron micrograph of rat liver nucleiincubated in the presence of GTP. A zone of close apposition of two nuclei is shown. The outermembranes of the two nuclei have fused to form continuous outer membranes (arrows) confluentperinuclear spaces (stars) and vesicles adjoining adjacent nuclei at the level of their respective innermembranes (arrowheads). Bar, 1 �m. (C) Fusion of ER membranes. Stripped rough microsomes afterincubation in the presence of GTP. The vesicle profiles are devoid of ribosomes, many are aggregatedand highly variable in size. Fusion of the normal-sized vesicles has led to the formation of very largemembrane-bounded profiles (asterisks). Bar, 1 �m. From Figs. 4, 5 and 8 in Paiement (1984).
Outer nuclear membrane interaction with microtubules has been shown in vitro by elec-tron microscopy using incubation of isolated liver nuclei in the presence of purified tubulin(Paiement, 1981a, b). Tubulin was prepared from rat brain homogenates by the microtubuledisassembly-reassembly method of Shelanski et al. (1973). Microtubules were incubatedwith purified nuclei prepared by the Blobel and Potter (1966) procedure. Purified rat livernuclei were incubated in the presence of pre-polymerized microtubules and then centrifugedat 1000 x g and the mixture processed for electron microscopic analysis. Following incuba-

C. Lavoie et al. / Progress in Histochemistry and Cytochemistry 46 (2011) 1–48 11
Fig. 4. Diagrammatic representation of outer nuclear membrane fusion. The zone of contact betweentwo nuclei is shown with its typical membrane arrangement. Arrows point to continuous outer nuclearmembranes at the limits of the zone of contact. The outer membranes within the zone of contact are nolonger continuous and exist as vesicular profiles (asterisks) attached to the inner nuclear membranesby means of nuclear pores (arrowheads). Ribosomes can be seen on the inner aspect of the vesicleprofiles as well as on the outside of the outer membrane away from the zone of contact.
tion microtubules were observed as cross sections or oblique sections and in close apposition(< 25 nm) to the outer nuclear membrane of different nuclei (Fig. 5).
Farah et al. (2005) used a similar assay to show outer nuclear membrane interaction withmicrotubules. CLIMP-63 an outer nuclear membrane protein and microtubule-associatedprotein-2 (MAP-2) were implicated in promoting nuclear-microtubule interaction (Farah
Fig. 5. In vitro reconstitution of the interaction between outer NE membranes and microtubules.Electron micrograph showing the periphery of a rat liver nucleus after incubation in the presenceof twice assembled microtubule proteins (prepared by the Shelanski method). Arrowheads point toribosomes on the outer nuclear membrane. Arrows point to cross-sections and oblique sections ofmicrotubules located at the surface of outer nuclear membrane (ONM). Bar, 0.5 �m.

12 C. Lavoie et al. / Progress in Histochemistry and Cytochemistry 46 (2011) 1–48
Fig. 6. In vitro reconstitution of the interaction between outer NE membranes and MAP2-pre-polymerized microtubules. Electron micrographs showing the periphery of nuclei after incubationin the presence of MAP2-pre-polymerized microtubules. Arrows point to cross-sections and obliquesections of microtubules located at the surface of associated outer nuclear membranes. All micrographshave the same magnification. Bar, 0.5 �m.
et al., 2005). MAP2-containing microtubules were observed to bind specifically to outernuclear membranes (see examples in Fig. 6) in contrast to microtubules containing tau, theaxonal MAP (not shown). Pretreatment of nuclei with antibodies to CLIMP-63 diminishedthe capacity of the nuclei to bind MAP-2-containing microtubules thus implicating thistransmembrane protein in microtubule binding at the nuclear surface. CLIMP-63 has beenshown to be associated with the rough ER and the outer nuclear membrane (Schweizeret al., 1995) and was shown to be involved in the interaction of ER with microtubules usingp63 overexpression (Klopfenstein et al., 1998) thus it is likely that CLIMP-63 in the outernuclear membrane plays a role in binding interactions between nuclei and microtubules invivo.
6. Reconstitution of the ER: Fusion of rough ER (rER) membranes
ER assembly occurs by a variety of mechanisms including de novo synthesis of allthe membrane components during interphase and by membrane fusion of existing ERmembrane fragments as is known to occur in post-mitotic cells and during interphase(Palade, 1975; Baumann and Walz, 2001; Estrada de Martin et al., 2005; Borgese et al.,2006).
As a consequence of work from numerous laboratories it is clear that membrane fusioncontributes to ER assembly however it is less clear to what extent membrane fusion con-tributes to the assembly of the different subcompartments of the ER. Few studies of fusionof ER membranes distinguish whether fusion occurs between membrane derivatives of

C. Lavoie et al. / Progress in Histochemistry and Cytochemistry 46 (2011) 1–48 13
Fig. 7. In vitro reconstitution of rER. Electron micrographs showing SRM incubated in the absenceof GTP (A and C) or in the presence of 0.5 mM GTP (B and D). Arrows in B and D indicate very largefusion products produced in the presence of GTP. Freeze-fracture electron micrographs in C and Dreveal the concave cytoplasmic fracture face (PF) and convex lumenal fracture face of small unfusedvesicles. The PF fracture face of a large membrane fusion product dominates the micrograph in D.Figures A and B reprinted from Biochem. Biophys. Res. Commun: Fig. 2A and B (Paiement et al.,1991). Figure D reprinted from the J Cell Biol: Fig 6 (Paiement et al., 1980). Bars, 0.5 �m.
the rER or between membrane derivatives of the sER or between membrane derivativesof both subcompartments. Because rER is defined by the presence of ribosomes and sERby the absence of ribosomes (Palade and Siekevitz, 1956a and b) electron microscopy isa key tool that can be used to distinguish between the rough and smooth ER membranecompartments.
Paiement et al. (1980) incubated purified rat liver rough microsomes in the presence ofGTP and Mg2+. This led to membrane fusion. Fusion was defined as the formation of largemembrane-bounded elements by fusion of membranes of small rER vesicles. Formationof large fusion products from rough microsomes required removal of part or all of themembrane-associated ribosomal particles. Fusion has been shown by thin-section electronmicroscopy of fixed and embedded microsomes (Fig. 7A and B), by freeze-fracture electronmicroscopy of rapidly frozen and cleaved membranes (Fig. 7C and D) as well as by lightmicroscopy of unfixed membranes using fluorescent lipid probes (not shown). Fusion hasalso been defined quantitatively as an increase in membrane surface area as evaluatedby morphometric measurement (Fig. 8) and compared with GTP-stimulated glycosylation(Paiement and Bergeron, 1983) and with GTP-stimulated accumulation of polyunsaturatedfree fatty acids (Lavoie et al., 1991).

14 C. Lavoie et al. / Progress in Histochemistry and Cytochemistry 46 (2011) 1–48
Fig. 8. Morphometric analysis of fusion of rER membranes. Stripped rough microsomes were incu-bated in the presence of GTP and then treated for analysis by electron microscopy. Large fusionproducts are highlighted in white. For morphometry, vesicles in electron micrographs were definedas membrane-limited structures that were often circular in profile, with a lumen that was mainlydevoid of content. Adjacent, non-overlapping micrographs were taken of the embedded and sectionedmembrane pellicle at a magnification of 11 500x. Negatives were scanned at a resolution of 400 dpiand converted to a positive image. Vesicles were highlighted in white using Photoshop 7.0 imagingsoftware (Adobe Systems Inc., Mountain View, CA) and an Intuos2 digital writing tablet (WacomTechnology Corp., Vancouver, WA). Highlighting in white of large vesicles in this figure was doneusing this method. The brightness and contrast of the regions excluding the vesicles was then loweredso that there were no other white objects on the image. The image was imported into the digitalmeasurement system, Sigma Scan Pro (SPSS Science). The number of highlighted vesicles was auto-matically stored in computer memory and Sigma Scan Pro calculated the diameter of each vesicle.A total of 1000 vesicles were measured for each sample in each experiment. The data generated bySigma Scan Pro was imported into statistics software package SPSS for statistical analysis. Bar, 1 �m.
6.1. Role of ribosomes in rER assembly
Although it is well known that ribosomal particles participate in the synthesis of ER pro-teins it is less well known how such particles influence membrane fusion and ER assembly.As indicated above rough microsomal vesicles will fuse in the presence of GTP if all orpart (> 60%) of the amount of associated RNA is removed from the membrane (Paiementet al., 1980). Puromycin-treatment of rough microsomes releases greater than two thirds ofthe RNA from rough microsomes but the remaining third of the RNA can be observed byelectron microscopy as ribosomal remnants associated with the membrane of the vesicles(Paiement et al., 1980, Fig. 9). Two possibilities may explain the impairment of fusion byribosomes, either ribosomal particles prevent fusion because their presence along the sur-face of the vesicles create steric hindrance and prevent membranes from coming into contactto allow fusion or the membrane fusogen has a restricted location beneath membrane-boundpolyribosomes and thus cannot take part in the fusion process. The role of ribosomes inrER membrane fusion was studied using purified rough microsomes and ribosome cap-

C. Lavoie et al. / Progress in Histochemistry and Cytochemistry 46 (2011) 1–48 15
Fig. 9. The role of ribosomes in rER membrane fusion. Electron micrograph of puromycin-treatedrough microsomes incubated at 37 ◦C for 120 min in the presence of 0.5 mM GTP. The vesicle profilesare aggregated and reveal a heterogeneity in size because many vesicles have undergone vesicle fusion.Ribosomal remnants, representing about one third of the original RNA associated with the microsomesare observed along the periphery of a fusion product (arrows). Arrowheads outline an en face view ofnumerous ribosomes on the surface of a fused vesicle. Bar, 0.5 �m.
ping experiments (Paiement et al., 1987). Ojakian et al. (1977) demonstrated that treatmentof rough microsomes with low concentrations of ribonuclease led to the agregation ofpolysomes creating clusters of ribosomes on the surface of the microsome. We have usedribonuclease treatment to create ribosome-free regions on the surface of rough microsomes.Electron microscope morphometry of rough microsomes treated with ribonuclease led tothe production of vesicles having as much as 50% of their surface area devoid of associatedribosomal particles (Paiement et al., 1987). When ribonuclease-treated microsomes wereincubated in the presence of GTP it was found that they were unable to fuse despite theincreased membrane surface area available for membrane interaction. Control experimentsconfirmed that RNAse treatment did not inhibit fusion of stripped rough microsomes. Itwas concluded that a cytosolically oriented protein involved in rER membrane fusion wasprobably hidden beneath the ribosome (Paiement et al., 1987). Thus ribosomes may act asnegative regulators for membrane interaction and ER assembly.
6.2. Lipids involved in rER assembly and lipid droplet formation: Creating a neworganelle
The contribution of lipids to the GTP-dependent fusion of rER membranes has beenexamined. The accumulation of polyunsaturated free fatty acids (PUFAs) was observedto occur coincident with GTP-dependent fusion of liver rough microsomes (Lavoie et al.,1991). Modulation of the content of unsaturated fatty acids induced coincident changes inGTP-dependent membrane fusion (Paiement et al., 1994). In the presence of GTP cytidinenucleotides were used to stimulate unsaturated fatty acid (linoleic acid and arachidonic acid)

16 C. Lavoie et al. / Progress in Histochemistry and Cytochemistry 46 (2011) 1–48
Fig. 10. The role of lipids in rER membrane fusion. In vitro modulation of the content of polyunsatu-rated free fatty acids induces coincident changes in GTP-dependent rER membrane fusion. Incubationof SRM in the presence of GTP (to promote GTP-dependent membrane fusion) and CTP (to promoteaccumulation of PUFAs) stimulates membrane fusion and leads to the production of large membrane-bound fusion products (top). Incubation in the presence of GTP and CTP plus ATP and coenzyme-A(to promote synthesis of triglycerides) led to a marked inhibition of membrane fusion (bottom) aswell as a reduction in free fatty acids. Bars, 0.5 �m. Figures reprinted from Biochim. Biophys. Acta:(Paiement et al., 1994).
accumulation in stripped rough microsomes (SRM) and ATP and coenzyme A were usedto reduce production of unsaturated fatty acids. Electron microscopy of SRM incubatedunder identical conditions revealed coincident stimulated or inhibited membrane fusion(Fig. 10, Paiement et al., 1994). It was concluded that unsaturated fatty acids have dramaticeffects on GTP-dependent membrane fusion and on rER membrane structure. The exactmechanism whereby PUFAs affect rER membrane fusion is not known but recent findingshave implicated omega-3 and omega-6 PUFAs in syntaxin-dependent membrane fusion(reviewed in Darios et al., 2007). Whether a similar mechanism is operational in the rERremains to be determined.
Biochemical analysis of SRM incubated in the presence of GTP, CMP, ATP plus increas-ing concentrations of CoA showed the accumulation of increasing amounts of triacylglyceroland decreasing amounts of unsaturated fatty acids. Electron microscope analysis of these

C. Lavoie et al. / Progress in Histochemistry and Cytochemistry 46 (2011) 1–48 17
Fig. 11. In vitro formation of triacylglycerol lipid droplets. (A-D) Effect of increasing concentrationsof CoA on the structure of SRM. Stripped rough microsomes were incubated in the presence of 0.5 mMGTP, 5 mM MgCl2, 0.5 mM CMP, 2 mM ATP and increasing concentrations (0 - 5 �M) of CoA for120 min at 37 ◦C. Bars, 0.5 �m. Figures reprinted from Biochim. Biophys. Acta: Fig. 8 (Paiementet al., 1994).
SRM confirmed the decreasing amounts of GTP-dependent membrane fusion and alsorevealed the formation of electron-dense structures on the cytoplasmic side of the vesiclemembranes (Fig. 11A-D). The electron-dense structures formed in the presence of CoAstained heavily when treated with osmium tetroxide (Fig. 12) and resemble intracellularosmiophilic lipid droplets described in hepatocytes in situ (Ashworth et al., 1966). Thus weinterpret these structures to represent triacylglycerol lipid droplets (Paiement et al., 1994).Stripped rough microsomes and cell-free reconstitution can thus be used to examine boththe role of fusion in ER assembly and lipid droplet formation. This model system allowsthe use of natural ER membranes to investigate the enzymes and substrates involved in thebirth of an organelle that of the lipid droplet.
But how is the lipid droplet made? Electron micrographs from a variety of cell typeshave shown that lipid droplets are often tightly associated with the ER and the enzymes thatcatalyze the synthesis of neutral lipids in the droplet core are localized predominantly to the

18 C. Lavoie et al. / Progress in Histochemistry and Cytochemistry 46 (2011) 1–48
Fig. 12. Osmiophilia of electron-dense amorphous structures formed in the presence of ATP andCoA. Stripped rough microsomes were incubated in the presence of 0.5 mM GTP, 5 mM MgCl2,0.5 mM CMP, 2 mM ATP and 1 mM CoA, 120 min at 37 ◦C. The microsomes were then filtered ontoMillipore membranes (Millipore, Bedford, MA, USA) and treated with a 1% OsO4 solution on icefor 60 min. They were then processed for electron microscopy and analysis of thin sections was donewithout further staining. Arrows point to large osmiophilic structures located at the periphery of largemembrane-bound fusion products. Bar, 0.5 �m. Reprinted from Biochim. Biophys. Acta: (Paiementet al., 1994).
ER (reviewed in Walther and Farese, 2009). The observation of accumulation of electron-dense osmiophilic structures at the surface of the membrane of purified rER derivatives(Fig. 12) coincident with stimulated triglyceride synthesis in vitro gives strong support tothe notion that the ER is the origin of lipid droplets (Paiement et al., 1994). The mechanismby which newly synthesized neutral lipids are transfered from the lipid bilayer of the ERmembrane to the growing lipid droplet on the cytoplasmic surface of the ER could be studiedto advantage by using purified ER membranes and cell-free incubation in combination withbiophysical approaches permitting the analysis of growing triglyceride aggregates withinthe interior of lipid bilayers as was done for triolein in synthetic membranes (Khandeliaet al., 2010).
6.3. Role of G-proteins in rER assembly
Using electron microscopy work from several laboratories have indicated that homotypicrER fusion is a GTP-dependent event (Paiement et al., 1980; Sokoloff et al., 1995; Dreierand Rapoport, 2000; Orso et al., 2009). The identity of the GTPase(s) involved in thisphenomenon has been a quest for many years. Recent studies from Orso et al (2009) haveimplicated the dynamin-like GTPase atlastin in GTP-dependent fusion of ER membranes.These investigators showed that downregulation of atlastin in Drosophila cells led to ERfragmentation and overexpression of this same protein led to expanded ER and expandednuclear envelope cisternae. Although the studies of Orso et al. (2009) did not specify whichER membrane domain was GTPase-sensitive the electron micrographs presented in theirpaper clearly show ER membranes with associated ribosomes (see Figs. 2a and 3b in Orso

C. Lavoie et al. / Progress in Histochemistry and Cytochemistry 46 (2011) 1–48 19
et al., 2009). Other authors have suggested that atlastin promotes membrane interaction(fusion or fission) of the reticular ER based on light microscope data (reviewed in Farhanand Hauri, 2009). In these studies, reticular ER was not defined in terms of rough andsmooth domains of the ER thus the possibility cannot be excluded that atlastin may infact be required for the fusion and biogenesis of a reticular (smooth or transitional) ERfrom flattened cisternal sheets of rER. Indeed, since the rER is composed of two differentdomains, one defined by the presence of ribosomes (the outer nuclear envelope membraneand flattened rER cisternae) and one defined by the membrane junction between rER andsER (transitional ER), different GTPases could be involved in membrane fusion at eachdomain. Whereas atlastin may regulate fusion and biogenesis of the reticular ER (Hu et al.,2009) a separate GTPase may regulate homotypic fusion of rER membranes.
6.3.1. A ras-like GTPase involved in rER membrane fusionWe have previously identified many ras-related proteins (GTP-binding proteins) in rough
microsomal membranes (Lanoix et al., 1989). Since numerous ras-related proteins havebeen implicated in membrane fusion during vesicular trafficking (Hutagalung and Novick,2011) and since ras-related proteins exhibit varying degrees of homology with ras p21(Valencia et al., 1991) we have examined the effects of several anti-ras p21 antibodies onthe GTP-dependent fusion of rER membranes in vitro. When stripped rough microsomeswere incubated at 37 ◦C in neutral buffer plus MgCl2 but in the absence of GTP they aggre-gated into small clumps (Fig. 13A). Incubation of stripped rough microsomes in the samemedium plus GTP led to aggregation as well as to the fusion of small microsomal vesiclesto form very large membrane-bounded vesicles (Fig. 13B). Pretreatment of stripped roughmicrosomes with monoclonal antibody 6B7 led to a strong inhibition of GTP-dependentfusion as indicated by the absence of large fusion products (Fig. 13C). This inhibition waspartially reversed when monoclonal antibody 6B7 was pre-incubated in the presence ofras protein prior to incubation with membranes (Fig. 13 D). These results were confirmedusing a quantitative assay employing morphometric analysis (Table 1). In contrast, othermonoclonal antibodies including 142-24E05 (Fig. 13E), M-90 (Fig. 13 F), Y13-259 andM-157 (data not shown) had no effect as indicated by the formation of large fusion productssimilar to those observed when stripped rough microsomes were incubated with GTP alone(compare for example the range of vesicle sizes in Fig. 13E and F with that of vesicles inFig. 13B). Control pretreatments of microsomes with 5 �g of non-specific mouse IgG, 5 �gof non-specific rat IgG or with the preservatives added to the monoclonal antibody 6B7solutions (0.1% sodium azide and serum albumin) had negligible effect on GTP-stimulatedrER membrane fusion (data not shown). These results suggest that a ras-related proteinmaybe involved in GTP-dependent fusion of rER membranes in vitro. The 6B7 antibodywas most effective in inhibiting fusion. Since this antibody recognizes the effector domainof the ras protein (Wong et al., 1986) this suggests that the GTPase involved in rER mem-brane fusion contains a peptide sequence that is similar to the putative effector region of rasp21.
6.3.2. Signal recognition particle receptor beta (SRPrβ) and ER assemblyWe next examined the possibility that SRPr� might be involved in GTP-dependent fusion
of rER membranes. Signal recognition particle receptor � is a transmembrane protein, a

20 C. Lavoie et al. / Progress in Histochemistry and Cytochemistry 46 (2011) 1–48
Fig. 13. Effect of various anti-ras monoclonal antibodies on GTP-dependent fusion of rER. Strippedrough microsomes (150 �g) were pre-incubated in the presence of 20 �l of sucrose imidazole buffer(A-B) or 20 �l of various anti-ras monoclonal antibodies (C-F) for 30 min at 10 ◦C. Subsequentlysucrose-imidazole (A) or GTP (0.5 mM final concentration, B-F) and MgCl2 (7.5 mM final concen-tration) were added to the incubation medium and incubation continued for an additional 120 min at37 ◦C. After incubation, membranes were fixed, dehydrated and processed for electron microscopy. Inpanel D anti-ras monoclonal antibody 6B7 (20 �l) was neutralized in the presence of 5 �g of purifiedras protein (v-Ha-ras, a gift from J.B. Gibbs, Merck Sharp and Dohme Research Laboratories, WestPoint, PA, U.S.A.) 30 min at 10 ◦C prior to addition to the microsomes and incubation for 120 minat 37 ◦C in the presence of GTP. Asterisks indicate large vesicles formed during incubation as con-sequence of membrane fusion between small unfused vesicles. 6B7 monoclonal antibody preparedagainst a synthetic peptide corresponding to amino acids residues 29 through 44 of normal ras proteinwas from Cetus Corporation, Emeryville, CA, U.S.A. Rat monoclonal antibody Y13-259 generatedagainst Harvey K-ras p21 which recognizes all ras p21 species was from Oncogene Science Inc.,Manhasset, NY, U.S.A. Monoclonal antibody M-90 which recognizes amino acids 107 to 130 ofnormal ras was a gift from J.C. Lacal, Madrid Spain. Bars, 1 �m.

C. Lavoie et al. / Progress in Histochemistry and Cytochemistry 46 (2011) 1–48 21
Table 1. Morphometric analysis of the effects of anti-ras monoclonal antibody 6B7 on GTP-dependent fusion of stripped rough microsomal membranes. Results are shown from three separateexperiments using three different batches of rough microsomes.
Incubationcondition
Total membrane(�m)
Mean membranelength (�m)
Standarddeviation
Fusionindex (%)
−GTP 263.6 .263 0.194 0285.2 .285 0.171 0234.5 .235 0.165 0
+GTP 425.8 .426 1.092 38.1548.1 .548 1.396 47.9461.0 .461 1.097 49.1
+GTP 188.1 .188 0.171 0+6B7 264.0 .264 0.180 0
259.1 .259 0.196 9.5+GTP 372.5 .372 0.659 29.3+6B7 374.4 .374 0.453 23.8+ras 318.7 .319 0.524 26.4
GTPase, found in the rER and is involved in the targeting of nascent polypeptide chains tothe protein translocation machinery in the endoplasmic reticulum membrane (Egea et al.,2005). Three main pieces of information inspired us to examine the possible involvementof SRPr� in the fusion of rER membranes. One, SRPr� has a peptide sequence that exhibits40% amino acid identity with the amino acids in the ras effector region as determinedusing the amino acid alignment tool developed by Pearson et al. (1997), two, a reportindicating that SRPr� controls ER structure in yeast (Prinz et al., 2000) and three, ourprevious demonstration that the GTPase involved in the fusion of rough microsomes islocated beneath the ribosome (Paiement et al., 1987).
Rabbit polyclonal antibodies were generated against the peptide sequence CSAKG-GRGDTGSADIQDLEK which corresponds to the C-terminal sequence region (amino acids244-264) of rat SRPr� (NCBI gi|number 61676217). Western blot was performed to testreactivity and purity. Antibody was observed to bind to a single protein band of approximatemolecular weight of 30 kDa in stripped rough microsomes, rough microsomes and smoothmicrosomes from rat liver (Fig. 14A). Digestion of stripped rough microsomes in vitro withincreasing concentrations of Proteinase K led to the gradual disappearance of anti- SRPr�antibody binding on immunoblots but did not affect BiP antibody staining in the same blotconfirming protein latency for BiP and indicating that the epitope recognized by the anti-SRPr� was on the cytosolic side of the ER microsomes (Fig. 14B). Immunofluorescencelabeling of cultured Hela cells using anti- SRPr� revealed a subcellular staining patternresembling classical ER distribution (Fig. 15).
We next determined the effect of anti-SRPr� antibody on ER membrane fusion in vitroby antibody neutralization study. Stripped rough microsomes (high density microsomesstripped of associated ribosomes by high salt treatment) were incubated with GTP in thepresence or absence of anti-SRPr� antibody. When SRM were incubated with GTP in the

22 C. Lavoie et al. / Progress in Histochemistry and Cytochemistry 46 (2011) 1–48
Fig. 14. Characterization of anti-SRPr� antibodies. (A) Anti-SRPr� antibodies reacted with proteinbands in the 30 kDa molecular weight range in stripped rough microsomes (SRM), in rough micro-somes (RM) and smooth microsomes (SM) from rat liver. (B) Site of location of the SRPr� antigenin the ER membrane. Protease digestion of stripped rough microsomes (70 �g) with increasing con-centrations of proteinase K (0.01-0.1 �g/ml). The anti-SRPr� antibody signal disappears followingtreatment with increasing concentrations of Proteinase K whereas the anti-BiP antibody signal doesnot change proportionally. This indicates that anti-SRPr� antibodies react with an epitope on thecytosolic side of the stripped rough microsomes. Since BiP is within the lumen of the same micro-somes they are protected from Proteinase K digestion. Anti-SRPr� antibodies were prepared withthe help of Dr. Hugh Bennett (Sheldon Biotechnology Centre, McGill University, Montreal, Canada)when Line Roy and Jacques Paiement were members of the Montreal proteomics network directed byDr. John Bergeron (Department of Anatomy and Cell Biology, McGill University, Montreal, Canada).
Fig. 15. Localization of SRPr� in Hela cells. Cells were fixed with 4% formaldehyde washed, thentreated with anti- SRPr� as primary antibody followed by fluorescein-congugated anti- rabbit IgGand examined in the fluorescent microscope. The montage reveals a fluorescent staining patternreminiscent of that for ER protein marker distribution.
absence of anti-SRPr� antibody, electron micrographs revealed that GTP stimulated fusionof small SRM and caused formation of large vesicle fusion products (Fig. 16A). Incubationof stripped rough microsomes with GTP and pre-immune antibody led to the formationof large vesicles of similar phenotype (not shown). However incubation of stripped roughmicrosomes with GTP and anti-SRPr� antibody led to significant decrease of membranefusion and caused formation of much smaller fusion products (Fig. 16B).
These results were confirmed by quantification using morphometric analysis. Usingelectron micrographs of incubated microsomes we measured the membrane lengths or

C. Lavoie et al. / Progress in Histochemistry and Cytochemistry 46 (2011) 1–48 23
Fig. 16. Effect of anti-SRPr� antibody on in vitro rER membrane fusion. Electron micrographs ofSRM after incubation for 120 min in the presence of 0.5 mM GTP without antibody (A) or in thepresence of anti- SRPr� (B). Incubation led to the fusion of small vesicles and the formation of largefusion products (arrows). Fusion products produced in the presence of anti- SRPr� (arrows, B) aremuch smaller than those produced during control incubations (compare fusion products in A withthose in B). Bars, 0.5 �m.
perimeters of incubated vesicles, for microsomes incubated under normal conditions andtreated with anti-SRPr� antibody. The perimeter measurements were then converted todiameter values. The mean diameter values and the standard deviations from the meansare shown in Table 2. Since the sizes of the vesicles depend on the amount of membranefusion, the mean vesicle diameter was then used to compare relative amounts of fusion.When compared statistically control-incubated stripped rough microsomes exhibited greatercapacity to fuse in the presence of GTP than stripped rough microsomes incubated with GTPand in the presence of anti-SRPr� antibody (Table 2). These in vitro studies suggest thatGTP-dependent fusion of rough ER membranes can be controlled by the GTPase SRPr�.
We also determined the effect of anti-SRPr� antibody on the fusion of nuclear envelopemembranes in vitro. Fusion between outer nuclear membranes is monitored by countingclosely apposed nuclear pairs that exhibit fused outer membranes (Paiement, 1981a, b).When isolated nuclei are incubated in the absence GTP they aggregate and form nuclearpairs with closely applied outer nuclear membranes (not shown). When isolated nuclei are

24 C. Lavoie et al. / Progress in Histochemistry and Cytochemistry 46 (2011) 1–48
Table 2. Morphometric analysis of the effect of anti-SRPr� antibody on in vitro rER membranefusion. Stripped rough microsomes were incubated in the presence of GTP in the absence or presence ofanti-SRPr� for 120 min. Following incubation the membranes were processed for analysis by electronmicroscopy. Electron micrographs taken at 45,000 times magnification were used for measurement ofvesicles sizes. Hundreds of vesicles were measured as described in the legend to Fig. 8. The presenceof anti-SRPr� led to inhibition of GTP-dependent membrane fusion and the formation of smallervesicles as shown by a reduction in the average diameters. Results are shown from three separateexperiments using three different batches of rough microsomes. SRM, stripped rough microsomes.
IncubationSRM
No. vesiclesmeasured
Mean diameter(�m)
Standard errorof the mean
Maximum vesiclesize (�m)
− antibodyExp1
1013 0.331 0.353 3.14
+ anti-SRPr�Exp1
993 0.281 0.238 1.92
− antibodyExp2
1302 0.288 0.317 2.76
+ anti-SRPr�Exp2
1302 0.215 0.214 1.98
− antibodyExp3
969 0.307 0.116 3.49
+ anti-SRPr�Exp3
969 0.246 0.008 1.89
incubated with GTP they aggregate and form nuclear pairs with continuous outer mem-branes, confluent perinuclear spaces and inverted vesicles with ribosomes in their lumenwhich interconnect the fused nuclei at the level of nuclear pores (Figs. 3B and 4). Amountof GTP-dependent fusion of outer nuclear membranes was calculated following incubationin the absence or presence of anti-SRPr� antibodies (Table 3). Incubation in the presence ofanti-SRPr� antibodies led to greater than 50% inhibition of outer nuclear membrane fusionwhen compared to control (Table 3). Thus SRPr� antibodies were able to inhibit GTP-dependent fusion of outer nuclear membranes as observed in the case of GTP-dependentfusion of rER membranes. This data is consistent with SRPr� being associated with theouter nuclear membrane and with the possibility that SRPr� may control GTP-dependentouter nuclear membrane fusion as well. GTP-dependent fusion may play an important rolein NE growth and nuclear surface expansion in interphase cells.
To investigate the possibility that SRPr� can influence ER structure in vivo we specifi-cally suppressed SRPr� expression by treatment with three short interfering RNA (siRNA)targeted against SRPr� sequence. Immunoblot analysis of HeLa cell lysates from timecourse experiments revealed depletion of SRPr� in Hela cells as early as 6 h after trans-fection (not shown). Fig. 17 shows immunoblot results obtained 24 h after transfection.SRPr� expression was knocked down by the three SRPr�-specific siRNA probes used inour studies whereas the control nonspecific siRNA probe had no effect. To indicate theeffect of SRPr� knockdown on ER structure immunocytochemistry was performed usinganti-calnexin antibody (Fig. 18, left panels & 18, right panels). Hela cells were transfected

C. Lavoie et al. / Progress in Histochemistry and Cytochemistry 46 (2011) 1–48 25
Table 3. Morphometric analysis of the effect of anti-SRPr� antibody on in vitro GTP-dependentouter nuclear membrane fusion. Rat liver nuclei were incubated in the presence of GTP in the absenceor presence of anti- SRPr� antibodies for 120 min, they were then fixed with glutaraldehyde andprocessed for analysis by electron microscopy. GTP-dependent fusion of outer nuclear membraneswas quantified as previously described (Paiement, 1981). Results are shown from three separateexperiments using three different batches of nuclei. ONM, outer nuclear membrane.
IncubationONM
No. of pairednuclei
No. of pairs withfused ONM
% of pairs withfused ONM
− antibodyExp1
84 60 71
+ anti-SRPr�Exp1
102 33 32
− antibodyExp2
81 46 56
+ anti-SRPr�Exp2
83 28 34
− antibodyExp3
93 67 72
+ anti-SRPr�Exp3
98 37 38
Fig. 17. Efficiency of short interfering RNA (siRNA) targeted against SRPr� sequence. Immunoblotof Hela whole-cell lysates 24 h after triplicate transfections with SRPr�-targeting or control siRNAs.Transfections were performed 4 times with a 12 h interval. Blots were probed with anti-SRPr�. Cellswere maintained in medium supplemented with 10% foetal calf serum. Three siRNAs targeting SRPr�mRNA (purchased from Invitrogen Canada Inc.) comprised of 25 base RNA nucleotide sequences(Oligo 1, uuaacaaccugacaaagagcaacgu, Oligo 2, uuaaaccgcucuaagaacugaagcc, Oligo 3, cagaaacucagc-cacaucuuucacc). All three siRNAs gave similar results. The control siRNA gave no target specificityin the human genome (oligo 1, acguugcucuuugucagguuguuaa, oligo 2, ggcuucaguucuuagagcgguu-uaa, oligo 3, ggugaaagauguggcugaguuucug, SRPr� reversed). siRNA duplex was transfected intocells using Lipofectamine RNAIMAX (Invitrogen Canada Inc.). Proteins were detected with HRP-conjugated secondary antibodies and chemiluminescent substrate. rm = hepatic rough microsomesused as positive control for SRPr� detection.
with negative control siRNA and three SRPr�-specific siRNA. 6, 12, 18, 24, and 48 hafter transfection, cells were fixed and incubated with anti-calnexin antibody and subse-quently with rhodamine anti-rabbit IgG and examined with the fluorescence microscope. Weobserved significant changes in the structure of the ER in Hela cells transfected with SRPr�siRNA whereas negative control siRNA had no effect on ER structure. Cells transfected with

26 C. Lavoie et al. / Progress in Histochemistry and Cytochemistry 46 (2011) 1–48
Fig. 18. SRPr� knockdown induces fragmentation of the rER in vivo as suggested by fluorescencemicroscopy. HeLa, cells were transfected with negative control siRNA (left panels) or with SRPr�siRNA (right panels). 6, 12, 18 and 24 h after transfection, cells were fixed and incubated withanti-calnexin antibody and subsequently with rhodamine anti-rabbit IgG and examined with the flu-orescence microscope. In control cells (left panels), arrowheads point to fluorescent calnexin-stainedtubules radiating out into Hela cell cytoplasmic extensions and observed at different times after trans-fection. Cells treated with SRPr� siRNA (right panels) reveal two new phenotypes 1) cells are morerounded and 2) they reveal a fragmented calnexin-labeled ER distribution when compared to controlsiRNA treated cells.
negative control siRNA revealed normal tubular ER structure (Fig. 18, left panels) whereascells transfected with SRPr� siRNA revealed a fragmented and vesicular ER distribution(Fig. 18, right panels). This effect was detected as early as 6 h after transfection.
To further examine the effect SRPr� knockdown on ER structure, electron microscopywas carried out. Hela cells were transfected with negative control siRNA or three SRPr�-specific siRNA. 6, 9 and 12 h after transfection, cells were fixed and processed for electronmicroscopy. Cells transfected with negative control siRNA revealed the presence of longuninterrupted ER cisternae (Fig. 19A, B and C) whereas cells transfected with SRPr�siRNA revealed the presence of short interrupted ER cisternae (Fig. 19D, E and F). Theshort interrupted cisternae observed in cross-section of Hela cell extensions in electronmicrographs (Fig. 19D, E and F) are thought to correspond to the fragmented ER structuresobserved by en face view in fluorescence micrographs (Fig. 18 right panels). The electronmicroscopy data is consistent with the immunofluorescence data and suggests that SRPr�knockdown leads to modification of ER structure in Hela cells.
These results suggest that SRPr� may control ER structure in vivo and are consistentwith the previous report suggesting that SRPr� controls ER structure in yeast (Prinz et al.,2000). Put into perspective with the results obtained by the in vitro fusion assays (see above),SRPr� may in fact control ER structure by being able to promote ER membrane fusion.Thus in addition to its well known role in the control of the site of ribosome binding inthe rER, SRPr� may control fusion of the rER with itself (i.e. homotypic fusion). Such

C. Lavoie et al. / Progress in Histochemistry and Cytochemistry 46 (2011) 1–48 27
Fig. 19. SRPr� knockdown induces fragmentation of the rER in vivo as suggested by electronmicroscopy. (A, B and C) control siRNA treated cells. Hela cells were transfected with negativecontrol siRNA. 6, 12, and 24 h after transfection, cells were fixed and processed for analysis byelectron microscopy. Figures A, B and C show examples of Hela cell cytoplasmic extensions. Theendoplasmic reticulum (ER) cisternae within these extensions appear long and uninterrupted. (C, Dand E) SRPr� siRNA treated cells. Hela cells were transfected with SRPr� siRNA. 6, 12, and 24 hafter transfection, cells were fixed and processed for analysis by electron microscopy. Figures D, Eand F show examples of Hela cell cytoplasmic extensions. The endoplasmic reticulum (ER) cisternaewithin these extensions appear short and disrupted. m, mitochondrion, n, nucleus. Bars, 1 �m.
rER fusion would ensure maintenance of the concentration of ribosome binding sites ina localized region of the ER such as flattened cisternae where all the cofactors of proteinsynthesis (including the ribosome, the signal recognition particle, the mRNA etc.) recycleon and off the rER membrane.
7. Reconstitution of the ER: Fusion of smooth ER (sER) membranes
As discussed above numerous laboratories have provided evidence for the fusion of ERmembranes however few of these studies have determined to what extent membrane fusionoccurs in different subcompartments of the ER. Indeed most studies have not concernedthemselves whether fusion occurs between membrane derivatives of the rER or betweenmembrane derivatives of the sER or between membrane derivatives of both subcompart-

28 C. Lavoie et al. / Progress in Histochemistry and Cytochemistry 46 (2011) 1–48
ments. This lack of distinction between the fusing compartments of the ER may in fact be dueto the varying definitions used for the subcompartments of the ER. Electron microscopy usesthe ribosome as the morphological marker for the rER and interconnecting tubules devoidof associated ribosomes and with tripartite junctions as definition for the sER (Palade andSiekevitz, 1956a and b; Fawcett, 1965; Baumann and Walz, 2001). More recent studiesusing cell culture, molecular biology techniques and confocal microscopy have redefinedthe subregions of the ER as sheets and tubules corresponding to the rough and smoothsubcompartments of the ER (Shibata et al., 2010; Farhan and Hauri, 2009). Endoplasmicreticulum tubules are thought to be shaped by the curvature-stabilizing proteins reticulonsand DP1/Yop1p and Climp63 is suggested to be an important contributor to the formationof rER sheets (Shibata et al., 2010). Whether these shape-regulating membrane proteinsdefine the subcompartments of the ER in all cell types is not known.
A number of cell types (such as hepatocytes and steroid-synthesizing cells) have a muchmore extensive sER consisting of an elaborate network of interconnecting tubules limitedby membranes devoid of associated ribosomes. It is not known if the ER subcompartmentsof these specialized cells exhibit similar molecular composition nor whether they fuse undersimilar conditions. Subcellular fractionation and proteomic analysis can provide informationon the former and cell-free reconstitution and electron microscopy can inform us on thelatter.
An in vitro assay was developed to study simultaneously the fusion properties of bothrough and smooth ER membrane subcompartments (Lavoie et al., 1996). In order to beable to study reconstitution of both rER and sER membranes simultaneously it was neces-sary to isolate a microsomal fraction that contained both rough and smooth ER membranederivatives.
A subfraction of rat liver microsomes that contained low-density rough microsomesas well as smooth microsomes was recovered from the 1.38 M sucrose step above theresidual pellet (high-density rough microsomes) generated after ultracentrifugation of totalmicrosomes in a sucrose step gradient (Fig. 1). Electron microscope morphometry (Lavoieet al., 1996) revealed this fraction to contain 57.8% smooth-surfaced vesicles with an aver-age diameter of 0.08 �m and 39.4% rough-surfaced vesicles with an average diameter of0.12 �m. The rough-surfaced vesicles revealed 1-5 ribosomal particles per vesicle. Incu-bation of this mixture of microsomes in the presence of 5 mM MgCl2, 0.5 mM GTP and2 mM ATP for 120 min at 37 ◦C led to the formation of membrane networks characterizedby the presence of large rough membrane cisternae at their periphery and an interconnectingnetwork of smooth tubules in their center (Fig. 20). GTP was observed to be required forreconstitution of rough membrane cisternae and both GTP and ATP were observed to berequired for smooth tubule formation. Morphometric studies indicated sequential forma-tion of rough membrane cisternae (0-60 min) followed by appearance of interconnectingsmooth tubules (>60 min). In addition the morphometric studies revealed that the proportionof rough membrane cisternae within the membrane networks diminished after 60 minutes,concomitant with an increase in smooth tubules while the sum of the two types of membraneremained constant (Lavoie et al., 1996). Thus assembly of smooth tubules may occur as anoutgrowth (i.e. via tubulation) from rough membranes. GTP is thought to promote tubula-tion from rough ER cisternae and ATP stimulates fusion of smooth tubules to generate aninterconnecting network and this is regulated by p97 (discussed below). This observation

C. Lavoie et al. / Progress in Histochemistry and Cytochemistry 46 (2011) 1–48 29
Fig. 20. In vitro reconstitution of sER. Low-density microsomes were incubated in the presenceof 0.5 mM GTP, 2 mM ATP and 5 mM MgCl2 for 120 min at 37 ◦C. This led to reconstitution ofER microsomes into membrane networks. The membrane networks shown in the three panels arecomposed of rough cisternae (rm), interconnecting smooth tubules (st) and fenestration (f). Asterisksindicate tritubular junctions. Bars, 0.5 �m.
is consistent with the RER-SER transformation model previously defined by the group ofPalade studying postnatal differentiation of hepatocyte ER in rat (Dallner et al., 1966a,b;Omura et al., 1967).
In vitro reconstituted ER networks have been characterized using a variety of elec-tron microscope cytochemical, immunocytochemical and radioautographic techniques.Glucose-6-phosphatase cytochemistry revealed reaction product on the luminal surfaceof the interconnecting tubules thus confirming the presence of glucose-6-phosphatase thebiochemical marker for the ER in reconstituted tubules (Fig. 21). A variety of secretoryproteins and membrane proteins were localized to in vitro reconstituted ER networks usingimmunogold labeling and ultrathin cryosections. Albumin and transferrin localization areshown in Fig. 22 A and B respectively, �2p24 and p58 localization are shown in Fig. 22Cand D respectively. Quantification by gold particle counting revealed these proteins to be inhigher concentration over the interconecting smooth tubules while the counts for ribophorinand ribosomal particles were in higher concentration in association with rough ER cister-nae (Lavoie et al., 1999). Thus opposing protein gradients were established and maintainedduring reconstitution of the ER membrane networks. A preembedding immunolabeling pro-cedure was also used to study protein composition in the in vitro reconstituted ER membranenetworks. This procedure was developed to examine protein antigenicity on the cytosolicsurface of purified and reconstituted membranes (Dominguez et al., 1991). Preembeddingimmunocytochemical technique shows calnexin antigenicity on both the surfaces of parallelrough membranes and interconnecting smooth tubules (Fig. 23).

30 C. Lavoie et al. / Progress in Histochemistry and Cytochemistry 46 (2011) 1–48
Fig. 21. Glucose-6-phosphatase cytochemistry of in vitro reconstituted sER. An interconnecting net-work of membrane tubules was generated by incubation of smooth microsomes in the presence ofGTP, ATP and MgCl2 and was subsequently treated for glucose-6-phosphatase detection. Reactionproduct indicative of the presence of enzyme activity is evident as electron opaque deposits. Arrowspoint to reaction product that fills or partially fills the lumen of interconnecting tubules and roughcisternae (rm). Modified from J. Cell Sci: Fig. 9 in Lavoie et al., 1996. Bar, 0.5 �m.
Fig. 22. Localization of albumin, transferrin, �2p24 and p58 (ERGIC-53) to in vitro reconstitutedsER networks. Antibodies against albumin (A), transferrin (B), �2p24 (C) and p58 (D) were appliedto ultrathin cryosections. Gold particle labeling is observed over interconnecting ER tubules (ser) andrough ER cisternae (rer). Inset in C shows �2p24 labeling associated with a vesicle at the periphery ofa reconstituted network. rer, parallel rough cisternae; ser, interconnecting smooth ER tubules. Bars,0.2 �m. Reprinted from J. Cell Biol: Fig. 2 (Lavoie et al., 1999).

C. Lavoie et al. / Progress in Histochemistry and Cytochemistry 46 (2011) 1–48 31
Fig. 23. Localization of calnexin antigens associated with in vitro reconstituted sER networks. Lowdensity microsomes were incubated in the presence of GTP, ATP and MgCl2 to allow reconstitution ofrough ER membranes and interconnecting smooth tubules. Anticalnexin antibodies were added to theincubation medium and the membranes further incubated for 30 min at room temperature. Membraneswere then fixed with 0.05% glutaraldehyde for 30 min then filtered onto Millipore membranes, theywere washed in saline solution and finally incubated with 3% bovine serum albumin. Following thisprotein-A gold was applied to the membrane pellicles and these were then washed and processed forelectron microscopy. Bar, 0.2 �m. Reprinted from J. Cell Biol: Fig. 4o (Lavoie et al.1999).
In summary, we have defined a cell-free system that reconstitutes an ER membranenetwork consisting of parallel rER cisternae enriched in ribophorin. The rER cisternae arein continuity with interconnecting smooth ER tubules enriched in soluble cargo albuminand transferrin as well as the transmembrane proteins �2p24 and p58. This reconstitutedER network is remarkably similar in structural organization to that of the ER in intacthepatocytes (Krstic, 1979; Fawcett, 1981).
7.1. Role of the ATPase p97 and the SNARE protein syntaxin 5 in the assembly ofsER membranes
Formation of interconnecting sER tubules in vitro is an ATP-dependent event sinceno interconnecting tubules are formed when ATP is omitted from the incubation mediumor when incubation is carried out in the presence of ATP and the ATP hexokinase D-Hexose-6-phosphotransferase (Roy et al., 2000). Because smooth tubule formation wasN-ethylmaleimide sensitive and because Latterich et al. (1995) had previously shown theinvolvement of the ATPase Cdc48p in both ER-ER and ER- nuclear envelope fusion in yeastthe involvement of p97 (the mammalian homologue of Cdc48p) in smooth tubule assemblywas investigated.
Antibodies to p97 were used to assess sER tubule formation in vitro. When low densitymicrosomes were incubated in the presence of GTP, ATP and MgCl2 with antibodies againstp97 rough microsomes fused to reconstitute ER membrane networks composed only of

32 C. Lavoie et al. / Progress in Histochemistry and Cytochemistry 46 (2011) 1–48
Fig. 24. Role of p97 and p47 on in vitro reconstitution of sER networks. KCl-treated low-densitymicrosomes were incubated in the presence of GTP, ATP and MgCl2 alone (A) or in the presenceof exogenous p97/p47 (B). Dilated ER cisternae were observed in the absence of p97/p47 (A) whileinterconnecting smooth tubules (st) and fenestrations (f) were observed in the presence of p97/p47.Bar, 0.5 �m. Reprinted from Molec. Biol. cell: Fig. 5B and C (Roy et al., 2000.).
parallel rough ER cisternae. The p97 antibodies inhibited the assembly of interconnectingsmooth tubules (Roy et al., 2000). Since the t-SNARE syntaxin 5 had been shown to berequired for the formation of pre-Golgi intermediates (Rowe et al., 1998) and since syntaxin5 was shown to be required for p97-mediated membrane fusion events (Rabouille et al.,1998) we next examined the involvement of syntaxin 5 in the production of ER networkswith interconnecting tubules. Antibodies to the cytoplasmic domain of syntaxin 5 were ableto inhibit smooth tubule formation. Tests of specificity were employed for both anti-p97 andfor anti-syntaxin 5 antibodies and confirmed identical inhibitory effects on smooth tubuleformation (Roy et al., 2000). It was concluded that p97 and syntaxin 5 are required for thefusion of smooth microsomes and thus for the assembly of smooth tubular ER networks invitro.
p97 ATPase protein was shown to bind reversibly to smooth ER membranes (Royet al., 2000). Low density microsomes that had been washed with high salt (2 M KCl)lost their content of p97 protein and were unable to reconstitute interconnecting smoothtubules (Fig. 24A). In contrast salt -washed microsomes incubated in the presence ofexogenously added purified p97 with associated p47 were able to form membrane net-works with smooth tubules (Fig. 24B). It was suggested that p97 dissociation fromER membranes following membrane fusion may play a role in the modulation of theamount of membrane fusion and consequently of the size of reconstituted ER net-works (Roy et al., 2000). A subsequent study revealed that tyrosine phosphorylationof p97 regulated p97 binding to ER membranes and thus assembly of ER membranesof the early secretory pathway (Lavoie et al., 2000). Phosphorylation/dephosphorylationof p97 was suggested to be able to control the number of rounds of fusion and

C. Lavoie et al. / Progress in Histochemistry and Cytochemistry 46 (2011) 1–48 33
Fig. 25. Model for SRPr�, atlastin, p97 and Syntaxin 5 involvement in ER assembly. The GTPaseSRPr� is suggested to play a role in the fusion of ER membranes permitting formation of parallelrough cisternae. The GTPase atlastin promotes formation of smooth tubules from rough ER cisternaeand the ATPase p97 and the SNARE protein syntaxin 5 stimulate fusion between the membrane ofthe tubules leading to the formation of a network of interconnecting smooth tubules continuous withrough ER cisternae.
this in turn would regulate the amount or extent of ER membranes in a cytoplasmicmilieu.
p97 forms a complex with p47 and syntaxin 5, VCP135 binds to the p97/p47/syntaxin5complex and dissociates it via p97 catalyzed ATP hydrolysis (Uchiyama et al., 2002).Uchiyama et al. (2002) microinjected antibodies to either VCP135 or to p47 into live CHOcells and observed a reduction in ER network formation defined quantitatively as amountof three-way junctions viewed in cells expressing GFP-tagged HSP47, an ER marker. Thisdata confirmed the in vitro studies of Roy et al. (2000) and Lavoie et al. (2000) and providedthe first in vivo evidence for an implication of p97 in sER assembly.
7.2. Summary of molecular factors involved in the assembly of rough ERcisternae and smooth ER tubules
Fig. 25 summarizes the controlling factors potentially involved in ER assembly. SinceGTP is required for assembly of rER membranes and since GTP and rER cisternae are bothrequired for smooth tubule formation in vitro (Lavoie et al., 1996; Roy et al., 2000) twoGTPases are postulated to be involved in ER assembly. SRPr� is a proposed new candidateregulator of rER fusion based on in vitro antibody neutralization studies using anti-SRPr�antibodies (Figs. 14-16, Tables 2 and 3) and based on siRNA SRPr�-knock-out experimentswith cell cultures (Figs. 17-19). SRPr� has previously been implicated in the control of ERstructure in yeast (Prinz et al., 2000). Here we suggest that it has a similar role in mammaliancells and that it does this by controlling fusion of NE membranes in postmitotic cells aswell as fusion between rER membranes and fusion between rER and NE membranes ininterphase cells. Because both GTP and rER membranes are required for the formation

34 C. Lavoie et al. / Progress in Histochemistry and Cytochemistry 46 (2011) 1–48
of smooth tubules and because morphometric studies have previously shown sequentialformation of rough membrane cisternae followed by appearance of interconnecting smoothtubules (Lavoie et al., 1996), a second ER GTPase is postulated to control smooth tubuleformation from the rER. Atlastin is the GTPase likely involved in smooth tubule formation(Hu et al., 2009; Orso et al., 2009). Atlastins are thought to directly affect branching of theER tubules either by fusion or by fission (Hu et al., 2009). A role for atlastin in the fissionof ER membranes and generation of smooth tubules from rough ER is proposed in ourmodel and the AAA ATPase p97 and syntaxin 5 are suggested to stimulate fusion betweensmooth tubules and thus complete the last step of ER assembly the formation of a networkof interconnecting tubules.
8. sER assembled in vitro is similar to transitional ER (tER)
Membrane networks consisting of parallel rough ER cisternae continuous with inter-connecting smooth tubules assembled in vitro (Lavoie et al., 1996; Lavoie et al., 1999;Roy et al., 2000) exhibit properties similar to tER. In vitro reconstituted networks werepreviously defined as tER based on (1) morphological criteria, i.e., a segregated smoothtubular compartment in direct continuity with rough ER cisternae, (2) the higher contentof secretory cargo, albumin and transferrin, in the smooth tubular compartment, (3) thehigher content of the recycling membrane proteins ERGIC53/p58 and the p24 family mem-ber �2p24 and (4) the smooth tubular domain of the reconstituted membrane networkscan transform into pleimorphic vesicular tubular clusters (VTCs) after incubation of pre-assembled ER networks in the presence of cytosol (Lavoie et al., 1999). Thus, reconstitutedtER defined by our cell-free incubation system consists of continuous rough and smoothmembrane domains; it contains molecular markers of the ER and the Golgi intermediatecompartments; and one of the subdomains of the tER, the smooth tubular domain, has thecapacity to transform into VTCs.
8.1. Formation of ER exit sites
The ER is the first compartment in the secretory pathway and is responsible for thetransfer of newly synthesized protein and lipid to the Golgi (Palade, 1975). Transportof membranes and proteins to the Golgi Apparatus occurs via membrane-bound vesiclesformed by COPII coatomer coat complex. COPII coatomer coats were first identified as aconsequence of yeast genetics and ER-Golgi transport assays and represent markers for ERexit sites (Fromme et al., 2008; Stagg et al., 2008). A second type of vesicle formed by COPIcoatomer coat complex is found between the ER and the Golgi Apparatus. COPI coatomercoat and vesicle formation was originally described using Golgi cell-free transport assays(Rothman, 1996; Beck et al., 2009). COPI-generated vesicles are primarily involved in ret-rograde traffic from the Golgi to the ER, are responsible for retrieving escaped ER proteinsfrom the Golgi (Cosson et al., 1998), promote membrane protein sorting (Dominguez et al.,1998) and are involved in recycling of the machinery that mediates vesicle formation andfusion (Lee et al., 2004).

C. Lavoie et al. / Progress in Histochemistry and Cytochemistry 46 (2011) 1–48 35
Fig. 26. Cytosol-dependent formation of VTCs. (A) tER formed in the presence of 0.5 mM GTP,0.5 mM ATP and 5 mM MgCl2, showing smooth ER tubules (st) continuous with peripherally locatedrough membrane cisternae (rm). (B,C) Subsequent incubation of tER in the presence of rat livercytosol (30 min) led to formation of vesicular tubular clusters (VTCs) and concomitant reduction ofsER tubules. rm, rough ER membranes. Bars, 0.2 �m.
8.2. Formation of ER exit sites from tER in vitro: Role of COPI
A cell-free reconstitution assay was developed to study ER cargo exit sites (Lavoie et al.,1999). This assay involved sequential assembly of tER from starting ER-derived low densitymicrosomes and subsequent selective generation of VTCs from interconnecting smooth ERtubules located within the tER by adding cytosol. In the first step, incubation of low densitymicrosomes in the presence of GTP, ATP and MgCl2 leads to microsomal fusion and thereconstitution of ER membrane networks consisting of parallel rER cisternae continuouswith interconnecting smooth ER tubules (Fig. 26A). In the second step, reconstituted tERis incubated in the presence of cytosol and mixed nucleotides which leads to selective andspecific transformation of interconnecting smooth tubules and the appearance of VTCs(Fig. 26B and C).
Antibodies to the COOH-terminal tail of �2p24, but not antibodies to the COOH-terminaltail of calnexin were able to inhibit the first step of ER reconstitution and COPI proteinwas implicated in the second step of reconstitution, the cytosol-dependent formation ofVTCs, as revealed by the fact that COPI-depleted cytosol inhibited VTC formation (Lavoieet al., 1999). Anti-�2p24 antibodies and the preembedding immunogold labeling proce-dure revealed �2p24 antigenic sites in association with the VTCs generated in the presenceof cytosol (Fig. 27). Anti-�-COP antibodies and the preembedding immunogold labelingprocedure revealed �-COP antigenic sites in association with the VTCs generated in thepresence of cytosol (Fig. 28). Thus the p24 family member, �2p24, and its cytosolic coatligand COPI coatomer, play a role in the de novo formation of VTCs and the regenerationof ER cargo exit sites. Consistent with this in vitro data overexpressed p23, a p24 familymember, in cultured cells was shown to stimulate proliferation of smooth ER tubules (Rojoet al., 2000) and more recently Mitrovic et al. (2008) used siRNA to silence p25/�2p24protein in cultured cells and they obtained a reduced number of ERGIC clusters and frag-

36 C. Lavoie et al. / Progress in Histochemistry and Cytochemistry 46 (2011) 1–48
Fig. 27. �2p24 localization in VTCs formed in the presence of cytosol. tER were incubated in thepresence of rat liver cytosol to form VTCs. These membrane networks were next incubated withanti-�2p24 antibody and then fixed in 0.05% glutaraldehyde before incubation with gold-secondaryantibody. �2p24 antigenicity is found within the VTCs of the two ER networks shown. Bars, 0.2 �m.Fig. 27A is reprinted from J. Cell Biol:Fig. 5 (Lavoie et al., 1999).
Fig. 28. COPI localization in VTCs formed in the presence of cytosol. (A) Immunoblot indicatingthe recruitment of �-COP, a COPI component, on reconstituted tER networks after incubation withor without cytosol, GTP and GTP�S. GTP�S is a non-hydrolyzable analog of GTP that stabilizes themembrane association of coatomers. Reconstituted tER networks were incubated in the absence (B) orpresence (C) of rat liver cytosol to form vesicular tubular clusters. These membrane networks were nextincubated with anti-�-COP antibody and then fixed in 0.05% glutaraldehyde before incubation withgold-secondary antibody. �-COP antigenicity is found mainly within the VTCs of the tER networksincubated with cytosol (C). Arrowheads indicate gold particles over clusters of vesicles (VTCs), arrowindicates gold particle over smooth membranes. (Panels A, B and C are reprinted from J.Cell Biol:Fig. 6 and 7 (Lavoie et al., 1999).
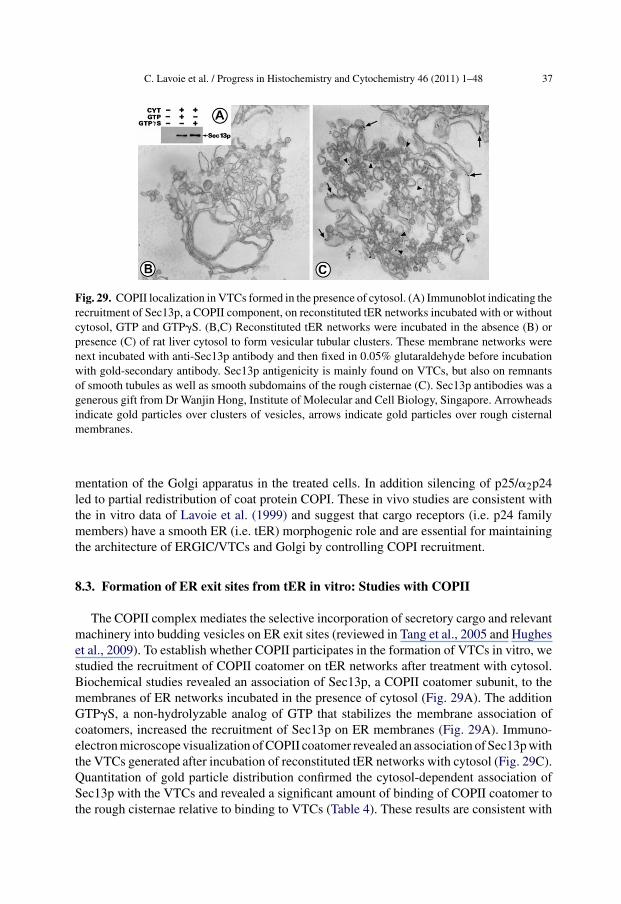
C. Lavoie et al. / Progress in Histochemistry and Cytochemistry 46 (2011) 1–48 37
Fig. 29. COPII localization in VTCs formed in the presence of cytosol. (A) Immunoblot indicating therecruitment of Sec13p, a COPII component, on reconstituted tER networks incubated with or withoutcytosol, GTP and GTP�S. (B,C) Reconstituted tER networks were incubated in the absence (B) orpresence (C) of rat liver cytosol to form vesicular tubular clusters. These membrane networks werenext incubated with anti-Sec13p antibody and then fixed in 0.05% glutaraldehyde before incubationwith gold-secondary antibody. Sec13p antigenicity is mainly found on VTCs, but also on remnantsof smooth tubules as well as smooth subdomains of the rough cisternae (C). Sec13p antibodies was agenerous gift from Dr Wanjin Hong, Institute of Molecular and Cell Biology, Singapore. Arrowheadsindicate gold particles over clusters of vesicles, arrows indicate gold particles over rough cisternalmembranes.
mentation of the Golgi apparatus in the treated cells. In addition silencing of p25/�2p24led to partial redistribution of coat protein COPI. These in vivo studies are consistent withthe in vitro data of Lavoie et al. (1999) and suggest that cargo receptors (i.e. p24 familymembers) have a smooth ER (i.e. tER) morphogenic role and are essential for maintainingthe architecture of ERGIC/VTCs and Golgi by controlling COPI recruitment.
8.3. Formation of ER exit sites from tER in vitro: Studies with COPII
The COPII complex mediates the selective incorporation of secretory cargo and relevantmachinery into budding vesicles on ER exit sites (reviewed in Tang et al., 2005 and Hugheset al., 2009). To establish whether COPII participates in the formation of VTCs in vitro, westudied the recruitment of COPII coatomer on tER networks after treatment with cytosol.Biochemical studies revealed an association of Sec13p, a COPII coatomer subunit, to themembranes of ER networks incubated in the presence of cytosol (Fig. 29A). The additionGTP�S, a non-hydrolyzable analog of GTP that stabilizes the membrane association ofcoatomers, increased the recruitment of Sec13p on ER membranes (Fig. 29A). Immuno-electron microscope visualization of COPII coatomer revealed an association of Sec13p withthe VTCs generated after incubation of reconstituted tER networks with cytosol (Fig. 29C).Quantitation of gold particle distribution confirmed the cytosol-dependent association ofSec13p with the VTCs and revealed a significant amount of binding of COPII coatomer tothe rough cisternae relative to binding to VTCs (Table 4). These results are consistent with

38 C. Lavoie et al. / Progress in Histochemistry and Cytochemistry 46 (2011) 1–48
Table 4. Quantitation of COPII labeling over tER networks. Reconstituted tER networks were incu-bated in the absence of cytosol (−Cyt), the presence of cytosol and GTP (+Cyt+GTP), or in thepresence of cytosol and GTP�S (+Cyt+GTP�S). These membrane networks were next incubated withanti-Sec13p antibody and then fixed in 0.05% glutaraldehyde before incubation with gold-secondaryantibody. Counts are expressed as mean number of gold particles per membrane network over roughER cisternae (RER) and over vesicular tubular complexes (SER/VTC).
Incubation tER RER SER/VTC # Networks
− Cyt 6.1 2.1 10+ Cyt + GTP 14.8 8.9 13+ Cyt + GTP�S 15.3 10 12
COPII being involved in formation of vesicles from the rough domain of the ER and furthersupport our premiss that in vitro reconstituted ER networks correspond to tER. Consistentwith the in vitro data obtained with reconstituted tER networks, both COPII and COPI havebeen found on VTCs in mammalian cells (Aridor et al., 1995; Scales et al., 1997). However,the dynamic relationship between these coats in ER-to-Golgi transport remains unclear.
9. ER reconstitution in live cells
Results obtained from in vitro studies are always subject to the question whether theyrepresent actual phenomenon that occurs in live cells. One may ask what is the physiologicalrelevance of the cell-free reconstitution of fragmented ER membranes into rER, sER andtER? Is it possible that fragmented ER membranes have actually been modified during cellfractionation and thus undergo aggregation and fusion to reconstitute modified structures inthe test tube? Or on the contrary, is it possible that fragmentation has not altered the basicproperties of the membranes and that incubation using physiological conditions allowsstudy of growth or reduction in size of large ER saccules and tubules that in vivo are knownto be constantly fluctuating between states of fusion and fission? In agreement with thelatter explanation is the fact that artificially induced fragments of ER have been shown toreconstitute into rER, sER and tER after microinjection into live cells in a heterologousmembrane transplant system (Paiement et al., 1988; Paiement et al., 1990)
9.1. In vivo reconstitution of the rER
Fragments of rER purified from rat liver homogenates were microinjected into Xenopusoocytes and the sites of microinjection analyzed by electron microscopy at different timespostinjection. The site of microinjection was determined by using morphological, cytochem-ical and immunocytochemical markers characteristic of injected microsomes (Paiementet al., 1988). Rough microsomes were identified by their association with peroxisomalcores in ultrathin sections of microinjected oocytes. Peroxisomal cores are contaminatingelements of rat liver rough microsomal fractions and because they are not found in frog

C. Lavoie et al. / Progress in Histochemistry and Cytochemistry 46 (2011) 1–48 39
Fig. 30. In vivo reconstitution of the rER. (A-D) Cytoplasmic regions of injection sites in Xenopusoocytes fixed at different times after injection of SRM from rat liver. (A) Region fixed immediately afterinsertion of SRM. A peroxisomal core (Pc) is observed in close proximity to many small membrane-bounded vesicles resembling freshly isolated microsomes on the basis of size and the presence ofinvaginations (arrowheads). Ribosomes are seen at the periphery of the small vesicles and in thesurrounding cytoplasm. Portions of a lipid droplet (Lp) and a mitochondrion (m) can also be seen. (B)Region fixed 30 min after microinjection. Peroxisomal cores (Pc) can be seen surrounded by smallmembrane-bounded vesicles, some showing invaginations (arrowheads) and others displaying anelongated tubular form. Double-headed arrows indicate the linear array of tubular elements in the field.(m) mitochondria. (C) Region fixed 60 min after microinjection. Two peroxisomal cores (Pc) are seenin a field containing endoplasmic reticulum (Er) cisternae. m, mitochondria. Structures resemblingelongated rough microsomes are indicated (arrows). (D) Region fixed 120 min after microinjection.Peroxisomal cores (Pc) are found near two cisternae of endoplasmic reticulum (Er) and mitochondria(m). Bars = 0.5 �m. Reprinted from J. Histochem. Cytochem: Figs. 1–4 (Paiement et al., 1988).
oocytes they provide a convenient morphological marker for the identity of microinjec-tion sites. At times 0 to 30 min postinjection, rough microsomal vesicles were recognizedwithin the injection site. These resembled the stripped rough microsomal vesicles used formicroinjection based on the fact that they contained invaginations as well as electron denselipoprotein particles, the morphological marker for hepatocyte ER (Fig. 30A and B). At latertimes (60 and 120 min) post injection, microinjection sites had reduced amounts of smallvesicles and increased amounts of parallel rER cisternae (Fig. 30C and D). Morphometricanalysis of rough microsomes in the injection sites revealed sequential disappearance ofrough microsomes and appearance of rER cisternae suggesting the transformation by fusionof the microsomes and formation of large parallel cisternae.

40 C. Lavoie et al. / Progress in Histochemistry and Cytochemistry 46 (2011) 1–48
Fig. 31. Immunogold labeling of the reconstituted rat liver rER in Xenopus oocytes. (A-F) Serialsections of oocyte cytoplasm near the injection site 3 hr after injection of rat liver rough microsomes.The sections were reacted with rabbit anti-rat liver rER antibodies (A-C) or pre-immune serum (D-F)and subsequently with protein-A gold complexes, and were counterstained with uranyl acetate andlead citrate and fixed with 3% formaldehyde and 1% glutaraldehyde. GMA embedding, 11-nm goldparticles. (A) Oocyte cytoplasm showing many rat liver peroxisomal cores. Many gold particles areobserved over the cores (arrows). LD, lipid droplet. (B) Cytoplasmic region containing immunolabeledperoxisomal cores (PC) as well as immunolabeled endoplasmic reticulum (ER). LD, lipid droplet.(C) Cytoplasmic region showing heavily labeled endoplasmic reticulum (ER). m, mitochondrion.(D-F) Oocyte cytoplasm showing background immunolabeling over peroxisomal cores (Pc), adjacentcytoplasm and ER cisternae (arrows). No label is observed over endogenous oocyte endoplasmicreticulum (ER). m, mitochondrion. Bars = 0.5 �m. Reprinted from J. Histochem. Cytochem: Figs.9–11 (Paiement et al., 1988).
Immunogold immunocytochemistry was employed to confirm the identity of the recon-stituted rER. When specific anti-rat liver rER antibodies and protein A-gold complexes wereapplied to thin sections from oocytes fixed 3 hr after microinjection of rough microsomesfrom rat liver, gold partides were observed mainly over peroxisomal cores, as well as overflattened and parallel ER cisternae (Fig. 31A, B and C). Treatment of serial sections identi-cally but with pre-immune sera instead of with specific antibodies led to background labelingover these same structures (Fig. 31D, E and F). These studies indicated that fragmentedrER from rat liver were able to reconstitute by membrane fusion after in vivo incubationinto cytoplasmic structures resembling those of rat hepatocyte rER from which they werederived.
9.2. In vivo reconstitution of the sER
The injection of rat liver smooth microsomes (tER subfraction) into Xenopus oocytesled to the formation of yolk platelet-free regions in oocyte cytoplasm which could be

C. Lavoie et al. / Progress in Histochemistry and Cytochemistry 46 (2011) 1–48 41
Fig. 32. In vivo reconstitution of the sER. (A-B) Cytoplasmic region of an injection site in a Xenopusoocyte fixed 120 min after injection of smooth microsomes from rat liver. (A) Low power micrographshowing three regions concentrated with reconstituted smooth ER (SER). Arrowheads indicate theposition of rER cisternae. m, mitochondrion. (B) Higher magnification of one of the networks ofinterconnecting smooth tubules shown in A. Asterisks indicate tritubular junctions and arrows point tolipoprotein particles, the morphological marker for rat hepatocyte ER, within the tubules. Arrowheadsindicate rER cisternae at the periphery of the sER networks. Bars = 0.5 �m.
recognized even in semi-thin sections by light microscopy. Analysis of these sites by electronmicroscopy revealed them to contain large areas filled with both parallel rER cisternaeand interconnecting smooth tubules (Fig. 32). Electron microscope immunocytochemistryusing antibodies specific to rat liver rER confirmed the presence of rat hepatocyte ERantigenicity associated with these reconstituted membranes (Paiement et al., 1988). Thefate of the smooth microsomes after in vivo incubation was determined by morphometry ofthe microsomes at different times postinjection (Paiement et al., 1990). Fig. 33 shows that

42 C. Lavoie et al. / Progress in Histochemistry and Cytochemistry 46 (2011) 1–48
Fig. 33. Kinetics of the sER reconstitution following insertion of rat-liver smooth microsomes intoXenopus oocyte cytoplasm. Relative proportions of vesicles (squares), branching tubules (circles),and cisternae (triangles) in yolk-platelet-free regions after various times of in vivo incubation ofmicroinjected rat liver smooth microsomes. Reprinted from Am. J. Anat: Fig. 2 (Paiement et al.,1990).
at time 0 min postinjection, only vesicles were observed. From 0 to 40 min, a decrease inthe number of vesicles occured coincident with an increase in the number of ER tubules andcisternae. Thereafter the number of vesicles and cisternae did not change but the numberof tubular junctions continued to increase. These studies indicated that fragmented sERfrom rat liver were able to reconstitute by membrane fusion after in vivo incubation intorER cisternae and interconnecting smooth tubules characterized by the presence of tripartitejunctions.
9.3. In vivo reconstitution of the ER in dividing cells
Fragments of rER and sER from normal adult rat liver as well as from dissected livertumors and from liver after hepatectomy were microinjected into one cell of cleaving two-cell Xenopus laevis embryos and the effects on development were examined during earlycleavage by morphological analysis (Paiement et al., 1992). The effect of microinjectionof membranes was examined by light microscopy, by electron microscopy as well as byscanning electron microscopy. By scanning electron microscopy after control microinjec-tion with sucrose-imidazole buffer, the surface of the animal hemisphere of 6-h embryosappeared normal and resembled that of uninjected control embryos (Fig. 34A). Embryosthat were microinjected with ER derivatives from normal adult rat liver had distinctly differ-ent surface morphologies and were characterized by the presence of large cells (Fig. 34B).Microinjection of microsomes from adult rat liver led to delayed cleavage in the microin-jected blastomeres and eventual arrest of embryonic development (Paiement et al., 1992).In contrast microinjection of microsomes from dissected liver tumors or from liver afterhepatectomy did not prevent embryonic development. It was concluded that membranes

C. Lavoie et al. / Progress in Histochemistry and Cytochemistry 46 (2011) 1–48 43
Fig. 34. Effect of microinjection of adult rat liver rER into one cell of a two cell Xenopus embryo.(A) Scanning electron micrograph of the animal pole of a Xenopus embryo after microinjection with4 mM imidazole buffer (pH 7.4) containing 0.25 M sucrose and then allowed to develop for 6 h.Normal cleavage has led to the formation of numerous small cells which appear uniform in size.(B) Scanning electron micrograph of the animal pole of a Xenopus embryo after microinjection withrough microsomes from normal adult rat liver and then allowed to develop for 6 h. Three hundrednanograms of membrane protein was injected. Delay in cleavage has led to the appearance of largeirregular cells on one side of the embryo. The arrows point towards the borders observed betweenlarge cells and small nomal-sized cells. Bars = 100 �m. Modified from Biochem. Cell Biol: Fig. 6Aand B (Paiement et al., 1992).
from proliferating tissues have no effect on embryonic development and thus have proper-ties that are more compatible with the cytoplasmic milieu of rapidly dividing blastomeres.These microinjection studies can be put into perspective with what is now known about thedifference in protein content between adult rat liver ER and ER from liver tumors (Roy et al.,2010). The observation that ER from liver tumors contains many antiapoptotic proteins thathave been upregulated could in part explain the neutral effect of these membranes aftermicroinjection into rapidly dividing blastomeres.
10. Concluding remarks
The endoplasmic reticulum is an amazing organelle. Our capacity to take it apart largelydepends on the physical properties of its different subcompartments. This organelle isremarkable in its capacity to function out of context of the intracellular milieu. The keyto being able to put the ER back together in the test tube is to ensure incubation of puri-fied components under physiological conditions and to add appropriate substrates. Underspecific conditions of incubation not only are we able to reconstitute the structure of thisorganelle but we can drive metabolic pathways that permit the creation of new organellessuch as lipid droplets from rER and VTCs from tER. Membrane transplantation method-ology has confirmed the capacity of the ER to undergo reconstitution in vivo. Organelleproteomics has revealed specific molecular profiles for each subcompartment and is startingto show us that the profiles may be significantly altered in disease. Further studies usingpurified ER subfractions are expected to increase our knowledge of the multiple functionsof this organelle. Indeed the ER has no doubt reserved many more fascinating lessons forus in the future.

44 C. Lavoie et al. / Progress in Histochemistry and Cytochemistry 46 (2011) 1–48
Acknowledgements
The current work was supported along the years by research grants from the CanadianInstitutes of Health Research and the Fonds de la Recherche en Santé du Québec.
References
Anderson DJ, Hetzer MW. Nuclear envelope formation by chromatin-mediated reorganization of the endoplasmicreticulum. Nat Cell Biol 2007;9:1160–6.
Aridor M, Bannykh SI, Rowe T, Balch WE. Sequential coupling between COPII and COPI vesicle coats inendoplasmic reticulum to Golgi transport. J Cell Biol 1995;131:875–93.
Ashley CA, Peters Jr T. Electron microscopic radioautographic detection of sites of protein synthesis and migrationin liver. J Cell Biol 1969;43:237–49.
Ashworth CT, Leonard JS, Eigenbrodt EH, Wrightsman FJ. Hepatic intracellular osmiophilic droplets: Effect ofLipid Solvents during Tissue Preparation. J Cell Biol 1966;31:301–18.
Baumann O, Walz B. Endoplasmic reticulum of animal cells and its organization into structural and functionaldomains. Int Rev Cytol 2001;205:149–214.
Beaufay H, Amar-Costesec A, Feytmans E, Thinès-Sempoux D, Wibo M, Robbi M, Berthet J. Analytical studyof microsomes and isolated subcellular membranes from rat liver. I. Biochemical methods. J Cell Biol1974a;61:188–200.
Beaufay H, Amar-Costesec A, Thinès-Sempoux D, Wibo M, Robbi M, Berthet J. Analytical study of microsomesand isolated subcellular membranes from rat liver. 3. Subfractionation of the microsomal fraction by isopycnicand differential centrifugation in density gradients. J Cell Biol 1974b;61:213–31.
Beck R, Rawet M, Wieland FT, Cassel D. The COPI system: molecular mechanisms and function. FEBS Lett2009;583:2701–9.
Bendayan M, Nanci A, Herbener GH, Grégoire S, Duhr MA. A review of the study of protein secretion applyingthe protein A-gold immunocytochemical approach. Am J Anat 1986;175:379–400.
Blobel G, Potter VR. Nuclei from rat liver: isolation method that combines purity with high yield. Science1966;154:1662–5.
Blobel G, Dobberstein B. Transfer of proteins across membranes. I. Presence of proteolytically processed andunprocessed nascent immunoglobulin light chains on membrane-bound ribosomes of murine myeloma. J CellBiol 1975a;67:835–51.
Blobel G, Dobberstein B. Transfer of proteins across membranes. II. Reconstitution of functional rough microsomesfrom heterologous components. J Cell Biol 1975b;67:852–62.
Boman AL, Taylor TC, Melancon P, Wilson KL. A role for ADP-ribosylation factor in nuclear vesicle dynamics.Nature 1992;358:512–4.
Borgese N, Francolini M, Snapp E. Endoplasmic reticulum architecture: structures in flux. Curr Opin Cell Biol2006;18:358–64.
Caro LG, Palade GE. Protein synthesis, storage, and discharge in the pancreatic exocrine cell. An autoradiographicstudy. J Cell Biol 1964;20:473–95.
Chen X, Karnovsky A, Sans MD, Andrews PC, Williams JA. Molecular characterization of the endoplasmicreticulum: insights from proteomic studies. Proteomics 2010a;10:4040–52.
Chen X, Sans MD, Strahler JR, Karnovsky A, Ernst SA, Michailidis G, Andrews PC, Williams JA. Quantitativeorganellar proteomics analysis of rough endoplasmic reticulum from normal and acute pancreatitis rat pancreas.J Proteome Res 2010b;9:885–96.
Claude A. A fraction from normal chick embryo similar to the tumor producing fraction of chicken tumor. ProcSoc Exp Biol Med 1938;39:398.
Claude A. The constitution of protoplasm. Science 1943;97:451–6.Cosson P, Lefkir Y, Démollière C, Letourneur F. New COP1-binding motifs involved in ER retrieval. EMBO J
1998;17:6863–70.Dabora SL, Sheetz MP. The microtubule-dependent formation of a tubulovesicular network with characteristics
of the ER from cultured cell extracts. Cell 1988;54:27–35.Dallner G, Siekevitz P, Palade GE. Biogenesis of endoplasmic reticulum membranes. I. Structural and chemical
differentiation in developing rat hepatocyte. J Cell Biol 1966a;30:73–96.Dallner G, Siekevitz P, Palade GE. Biogenesis of endoplasmic reticulum membranes. II. Synthesis of constitutive
microsomal enzymes in developing rat hepatocyte. J Cell Biol 1966b;30:97–117.

C. Lavoie et al. / Progress in Histochemistry and Cytochemistry 46 (2011) 1–48 45
Darios F, Connell E, Davletov B. Phospholipases and fatty acid signalling in exocytosis. J Physiol2007;585:699–704.
Deshaies RJ, Schekman R. A yeast mutant defective at an early stage in import of secretory protein precursorsinto the endoplasmic reticulum. J Cell Biol 1987;105:633–45.
Dominguez JM, Lanoix J, Paiement J. Localization of ras antigenicity in rat hepatocyte plasma membrane andrough endoplasmic reticulum fractions. Exp Cell Res 1991;192:137–47.
Dominguez M, Dejgaard K, Füllekrug J, Dahan S, Fazel A, Paccaud JP, Thomas DY, Bergeron JJ, Nilsson T.gp25L/emp24/p24 protein family members of the cis-Golgi network bind both COP I and II coatomer. J CellBiol 1998;140:751–65.
Dreier L, Rapoport TA. In vitro formation of the endoplasmic reticulum occurs independently of microtubules bya controlled fusion reaction. J Cell Biol 2000;148:883–98.
Egea PF, Stroud RM, Walter P. Targeting proteins to membranes: structure of the signal recognition particle. CurrOpin Struct Biol 2005;15:213–20.
Estrada de Martin P, Novick P, Ferro-Novick S. The organization, structure, and inheritance of the ER in higherand lower eukaryotes. Biochem Cell Biol 2005;83:752–61.
Farah CA, Liazoghli D, Perreault S, Desjardins M, Guimont A, Anton A, Lauzon M, Kreibich G, Paiement J,Leclerc N. Interaction of microtubule-associated protein-2 and p63: a new link between microtubules andrough endoplasmic reticulum membranes in neurons. J Biol Chem 2005;280:9439–49.
Farhan H, Hauri HP. Membrane biogenesis: networking at the ER with atlastin. Curr Biol 2009;19:R906–8.Fawcett DW.Seno S, Cowdry EV, editors. Intracellular membranous structure. Okayama: Japan society for cell
biology; 1965. p. 15–40.Fawcett DW. The cell. Philadelphia: WB Saunders Company Ltd; 1981.Fonseca SG, Urano F, Burcin M, Gromada J. Stress hypERactivation in the �-cell. Islets 2010;2:1–9.Friedl P, Wolf K, Lammerding J. Nuclear mechanics during cell migration. Curr Opin Cell Biol 2010;23:1–10.Fromme JC, Orci L, Schekman R. Coordination of COPII vesicle trafficking by Sec23. Trends Cell Biol
2008;18:330–6.Galeva N, Altermann M. Comparison of one-dimensional and two-dimensional gel electrophoresis as a separa-
tion tool for proteomic analysis of rat liver microsomes: cytochromes P450 and other membrane proteins.Proteomics 2002;2:713–22.
Glaumann H, Ericsson JL. Evidence for the participation of the Golgi apparatus in the intracellular transport ofnascent albumin in the liver cell. J Cell Biol 1970;47:555–67.
Gürkan C, Stagg SM, Lapointe P, Balch WE. The COPII cage: unifying principles of vesicle coat assembly. NatRev Mol Cell Biol 2006;7:727–38.
Healy SJ, Gorman AM, Mousavi-Shafaei P, Gupta S, Samali A. Targeting the endoplasmic reticulum-stressresponse as an anticancer strategy. Eur J Pharmacol 2009;625:234–46.
Hetzer M, Bilbao-Cortes D, Walther TC, Gruss OJ, Mattaj IW. GTP hydrolysis by Ran is required for nuclearenvelope assembly. Mol Cell 2000;5:1013–24.
Hetzer MW. The nuclear envelope. Cold Spring Harb Perspect Biol 2010;2:a000539.Hoeller D, Dikic I. Targeting the ubiquitin system in cancer therapy. Nature 2009;458:438–44.Hu J, Shibata Y, Zhu PP, Voss C, Rismanchi N, Prinz WA, Rapoport TA, Blackstone C. A class of dynamin-like
GTPases involved in the generation of the tubular ER network. Cell 2009;138:549–61.Hughes H, Budnik A, Schmidt K, Palmer KJ, Mantell J, Noakes C, Johnson A, Carter DA, Verkade P, Watson P,
Stephens DJ. Organisation of human ER-exit sites: requirements for the localisation of Sec16 to transitionalER. J Cell Sci 2009;122:2924–34.
Hutagalung AH, Novick PJ. Role of rab GTPases in membrane traffic and cell physiology. Physiol Rev2011;91:119–49.
Iwamatsu T, Kobayashi H. Electron microscopic observations of karyogamy in the fish egg. Dev Growth Differ2002;44:357–63.
Jensen WA. Observations on the fusion of nuclei in plants. J Cell Biol 1964;23:669–72.Kachar B, Reese TS. The mechanism of cytoplasmic streaming in characean algal cells: sliding of endoplasmic
reticulum along actin filaments. J Cell Biol 1988;106:1545–52.Khandelia H, Duelund L, Pakkanen KI, Ipsen JH. Triglyceride blisters in lipid bilayers: implications
for lipid droplet biogenesis and the mobile lipid signal in cancer cell membranes. PLoS One 2010.,doi:10.1371/journal.pone.0012811.
King MC, Lusk CP, Blobel G. Karyopherin-mediated import of integral inner nuclear membrane proteins. Nature2006;442:1003–7.
Klopfenstein DR, Kappeler F, Hauri HP. A novel direct interaction of endoplasmic reticulum with microtubules.EMBO J 1998;17:6168–77.
Krstic RV. Ultrastructure of the Mammalian Cell. An Atlas. Berlin: Springer-Verlag; 1979.

46 C. Lavoie et al. / Progress in Histochemistry and Cytochemistry 46 (2011) 1–48
Lanoix J, Roy L, Paiement J. Detection of GTP-binding proteins in purified derivatives of rough endoplasmicreticulum. Biochem J 1989;262:497–503.
Latterich M, Schekman R. The karyogamy gene KAR2 and novel proteins are required for ER-membrane fusion.Cell 1994;78:87–98.
Latterich M, Fröhlich KU, Schekman R. Membrane fusion and the cell cycle: Cdc48p participates in the fusionof ER membranes. Cell 1995;82:885–93.
Lavoie C, Jolicoeur M, Paiement J. Accumulation of polyunsaturated free fatty acids coincident with the fusionof rough endoplasmic reticulum membranes. Biochim Biophys Acta 1991;1070:274–8.
Lavoie C, Lanoix J, Kan FW, Paiement J. Cell-free assembly of rough and smooth endoplasmic reticulum. J CellSci 1996;109:1415–25.
Lavoie C, Paiement J, Dominguez M, Roy L, Dahan S, Gushue JN, Bergeron JJ. Roles for alpha(2)p24 and COPIin endoplasmic reticulum cargo exit site formation. J Cell Biol 1999;146:285–99.
Lavoie C, Chevet E, Roy L, Tonks NK, Fazel A, Posner BI, Paiement J, Bergeron JJ. Tyrosine phosphory-lation of p97 regulates transitional endoplasmic reticulum assembly in vitro. Proc Natl Acad Sci U S A2000;97:13637–42.
Lavoie C, Paiement J. Topology of molecular machines of the endoplasmic reticulum: a compilation of proteomicsand cytological data. Histochem Cell Biol 2008;129:117–28.
Lee MC, Miller EA, Goldberg J, Orci L, Schekman R. Bi-directional protein transport between the ER and Golgi.Annu Rev Cell Dev Biol 2004;20:87–123.
Longo FJ, Anderson E. The fine structure of pronuclear development and fusion in the sea urchin, Arbaciapunctulata. J Cell Biol 1968;39:339–68.
Longo FJ. Fertilization: a comparative ultrastructural review. Biol Reprod 1973;9:149–215.Matus S, Glimcher LH, Hetz C. Protein folding stress in neurodegenerative diseases: a glimpse into the ER. Curr
Opin Cell Biol 2011 Jan 31.Meldolesi J, Jamieson JD, Palade GE. Composition of cellular membranes in the pancreas of the guinea pig. 3.
Enzymatic activities. J Cell Biol 1971;49:150–8.Mellman I, Warren G. The road taken: past and future foundations of membrane traffic. Cell 2000;100:99–112.Mitrovic S, Ben-Tekaya H, Koegler E, Gruenberg J, Hauri HP. The cargo receptors Surf4, endoplasmic reticulum-
Golgi intermediate compartment (ERGIC)-53, and p25 are required to maintain the architecture of ERGICand Golgi. Mol Biol Cell 2008;19:1976–90.
Morand JP, Macri J, Adeli K. Proteomic profiling of hepatic endoplasmic reticulum-associated proteins in ananimal model of insulin resistance and metabolic dyslipidemia. J Biol Chem 2005;280:17626–33.
Nadler NJ, Young BA, Leblond CP, Mitmaker B. Elaboration of thyroglobulin in the thyroid follicle. Endocrinology1964;74:333–54.
Newport J, Dunphy W. Characterization of the membrane binding and fusion events during nuclear envelopeassembly using purified components. J Cell Biol 1992;116:295–306.
Ojakian GK, Kreibich G, Sabatini DD. Mobility of ribosomes bound to microsomal membranes. A freeze-etchand thin-section electron microscope study of the structure and fluidity of the rough endoplasmic reticulum. JCell Biol 1977;72:530–51.
Omura T, Siekevitz P, Palade GE. Turnover of constituents of the endoplasmic reticulum membranes of rathepatocytes. J Biol Chem 1967;242:2389–96.
Orso G, Pendin D, Liu S, Tosetto J, Moss TJ, Faust JE, Micaroni M, Egorova A, Martinuzzi A, McNew JA, DagaA. Homotypic fusion of ER membranes requires the dynamin-like GTPase atlastin. Nature 2009;460:978–83.
Paiement J, Beaufay H, Godelaine D. Coalescence of microsomal vesicles from rat liver: a phenomenon occurringin parallel with enhancement of the glycosylation activity during incubation of stripped rough microsomeswith GTP. J Cell Biol 1980;86:29–37.
Paiement J. Interactions of microtubule proteins with outer nuclear membranes. J Cell Biol 1981a;91:329a.Paiement J. GTP-dependent fusion of outer nuclear membranes in vitro. Exp Cell Res 1981b;134:93–102.Paiement J, Bergeron JJ. Localization of GTP-stimulated core glycosylation to fused microsomes. J Cell Biol
1983;96:1791–6.Paiement J. Physiological concentrations of GTP stimulate fusion of the endoplasmic reticulum and the nuclear
envelope. Exp Cell Res 1984;151:354–66.Paiement J, Rindress D, Smith CE, Poliquin L, Bergeron JJ. Properties of a GTP sensitive microdomain in rough
microsomes. Biochim Biophys Acta 1987;898:6–22.Paiement J, Kan FW, Lanoix J, Blain M. Cytochemical analysis of the reconstitution of endoplasmic reticu-
lum after microinjection of rat liver microsomes into Xenopus oocytes. J Histochem Cytochem 1988;36:1263–73.
Paiement J, Dominguez JM, McLeese J, Bernier J, Roy L, Bergeron M. Morphogenesis of endoplasmic reticulumin Xenopus oocytes after microinjection of rat liver smooth microsomes. Am J Anat 1990;187:183–92.

C. Lavoie et al. / Progress in Histochemistry and Cytochemistry 46 (2011) 1–48 47
Paiement J, Dominguez JM, Guénette A, Roy L. GTP-dependent membrane fusion during hepatocarcinogenesisand liver regeneration. Biochem Biophys Res Commun 1991;176:1494–500.
Paiement J, Dominguez JM, Messier PE. Cytoplasmic membrane transplants in early Xenopus embryos indicatecell-cycle-specific effects. Biochem Cell Biol 1992;70:1290–300.
Paiement J, Lavoie C, Gavino GR, Gavino VC. Modulation of GTP-dependent fusion by linoleic and arachi-donic acid in derivatives of rough endoplasmic reticulum from rat liver. Biochim Biophys Acta 1994;1190:199–212.
Paiement J, Young R, Roy L, Bergeron JJM. Isolation of rough and smooth membrane domains of the endoplasmicreticulum from rat liver. In: Celis JE, editor. Cell Biology: A Laboratory Handbook., Vol. 2, 3rd ed. ElsevierScience; 2006. p. 41–4.
Palade G, Siekevitz P. Liver microsomes; an integrated morphological and biochemical study. J Biophys BiochemCytol 1956a;2:171–200.
Palade G, Siekevitz P. Pancreatic microsomes; an integrated morphological and biochemical study. J BiophysBiochem Cytol 1956b;2:671–90.
Palade G. Intracellular aspects of the process of protein synthesis. Science 1975;189:347–58.Pearson WR, Wood T, Zhang Z, Miller W. Comparison of DNA sequences with protein sequences. Genomics
1997;46:24–36.Presley JF, Cole NB, Schroer TA, Hirschberg K, Zaal KJ, Lippincott-Schwartz J. ER-to-Golgi transport visualized
in living cells. Nature 1997;389:81–5.Prinz WA, Grzyb L, Veenhuis M, Kahana JA, Silver PA, Rapoport TA. Mutants affecting the structure of the
cortical endoplasmic reticulum in Saccharomyces cerevisiae. J Cell Biol 2000;150:461–74.Prins D, Michalak M. Endoplasmic reticulum proteins in cardiac development and dysfunction. Can J Physiol
Pharmacol 2009;87:419–25.Rabouille C, Kondo H, Newman R, Hui N, Freemont P, Warren G. Syntaxin 5 is a common component of the
NSF- and p97-mediated reassembly pathways of Golgi cisternae from mitotic Golgi fragments in vitro. Cell1998;92:603–10.
Ramadan K, Bruderer R, Spiga FM, Popp O, Baur T, Gotta M, Meyer HH. Cdc48/p97 promotes reformation ofthe nucleus by extracting the kinase Aurora B from chromatin. Nature 2007;450:1258–62.
Revel JP, Hay ED. An autoradiographic and electron microscopic study of collagen synthesis in differentiatingcartilage. Z Zellforsch Mikrosk Anat 1963;61:110–44.
Rojo M, Emery G, Marjomäki V, McDowall AW, Parton RG, Gruenberg J. The transmembrane protein p23contributes to the organization of the Golgi apparatus. J Cell Sci 2000;113:1043–57.
Roth J, Ravazzola M, Bendayan M, Orci L. Application of the protein A-gold technique for electron microscopicdemonstration of polypeptide hormones. Endocrinology 1981;108:247–53.
Rothman JE. The protein machinery of vesicle budding and fusion. Protein Sci 1996;5:185–94.Routledge KE, Gupta V, Balch WE. Emergent properties of proteostasis-COPII coupled systems in human health
and disease. Mol Membr Biol 2010;27:385–97.Rowe T, Dascher C, Bannykh S, Plutner H, Balch WE. Role of vesicle-associated syntaxin 5 in the assembly of
pre-Golgi intermediates. Science 1998;279:696–700.Roy L, Bergeron JJ, Lavoie C, Hendriks R, Gushue J, Fazel A, Pelletier A, Morré DJ, Subramaniam VN, Hong
W, Paiement J. Role of p97 and syntaxin 5 in the assembly of transitional endoplasmic reticulum. Mol BiolCell 2000;11:2529–42.
Roy L, Laboissière S, Abdou E, Thibault G, Hamel N, Taheri M, Boismenu D, Lanoix J, Kearney RE, PaiementJ. Proteomic analysis of the transitional endoplasmic reticulum in hepatocellular carcinoma: an organelleperspective on cancer. Biochim Biophys Acta 2010;1804:1869–81.
Sabatini DD, Blobel G. Controlled proteolysis of nascent polypeptides in rat liver cell fractions. II. Location ofthe polypeptides in rough microsomes. J Cell Biol 1970;45:146–57.
Scales SJ, Pepperkok R, Kreis TE. Visualization of ER-to-Golgi transport in living cells reveals a sequential modeof action for COPII and COPI. Cell 1997;90:1137–48.
Schirmer EC, Gerace L. The nuclear membrane proteome: extending the envelope. Trends Biochem Sci2005;30:551–8.
Schweizer A, Rohrer J, Slot JW, Geuze HJ, Kornfeld S. Reassessment of the subcellular localization of p63. J CellSci 1995;108:2477–85.
Shelanski ML, Gaskin F, Cantor CR. Microtubule assembly in the absence of added nucleotides. Proc Natl AcadSci U S A 1973;70:765–88.
Shibata Y, Shemesh T, Prinz WA, Palazzo AF, Kozlov MM, Rapoport TA. Mechanisms determining the morphologyof the peripheral ER. Cell 2010;143:774–88.
Sokoloff AV, Whalley T, Zimmerberg J. Characterization of N-ethylmaleimide-sensitive thiol groups required forthe GTP-dependent fusion of endoplasmic reticulum membranes. Biochem J 1995;312:23–30.

48 C. Lavoie et al. / Progress in Histochemistry and Cytochemistry 46 (2011) 1–48
Soullam B, Worman HJ. Signals and structural features involved in integral membrane protein targeting to theinner nuclear membrane. J Cell Biol 1995;130:15–27.
Stagg SM, LaPointe P, Razvi A, Gürkan C, Potter CS, Carragher B, Balch WE. Structural basis for cargo regulationof COPII coat assembly. Cell 2008;134:474–84.
Tabas I. The role of endoplasmic reticulum stress in the progression of atherosclerosis. Circ Res 2010;107:839–50.Tang BL, Wang Y, Ong YS, Hong W. COPII and exit from the endoplasmic reticulum. Biochim Biophys
2005;1744:293–303.Tsai LH, Gleeson JG. Nucleokinesis in neuronal migration. Neuron 2005;46:383–8.Uchiyama K, Jokitalo E, Kano F, Murata M, Zhang X, Canas B, Newman R, Rabouille C, Pappin D, Freemont
P, Kondo H. VCIP135, a novel essential factor for p97/p47-mediated membrane fusion, is required for Golgiand ER assembly in vivo. J Cell Biol 2002;159:855–66.
Ulbert S, Platani M, Boue S, Mattaj IW. Direct membrane protein-DNA interactions required early in nuclearenvelope assembly. J Cell Biol 2006;173:469–76.
Vale RD, Hotani H. Formation of membrane networks in vitro by kinesin-driven microtubule movement. J CellBiol 1988;107:2233–41.
Valencia A, Chardin P, Wittinghofer A, Sander C. The ras protein family: evolutionary tree and role of conservedamino acids. Biochemistry 1991;30:4637–48.
Vigers GP, Lohka MJ. A distinct vesicle population targets membranes and pore complexes to the nuclear envelopein Xenopus eggs. J Cell Biol 1991;112:545–56.
Walther TC, Farese Jr RV. The life of lipid droplets. Biochim Biophys Acta 2009;1791:459–66.Warshawsky H, Leblond CP, Droz B. Synthesis and migration of proteins in the cells of the exocrine pancreas as
revealed by specific activity determination from radioautographs. J Cell Biol 1963;16:1–24.Watkins JD, Hermanowski AL, Balch WE. Oligomerization of immunoglobulin G heavy and light chains in vitro.
A cell-free assay to study the assembly of the endoplasmic reticulum. J Biol Chem 1993;268:5182–92.Whur P, Herscovics A, Leblond CP. Radioautographic visualization of the incorporation of galactose-3H and
mannose-3H by rat thyroids in vitro in relation to the stages of thyroglobulin synthesis. J Cell Biol1969;43:289–311.
Wlodkowic D, Skommer J, McGuinness D, Hillier C, Darzynkiewicz Z. ER-Golgi network-a future target foranti-cancer therapy. Leuk Res 2009;33:1440–7.
Wong G, Arnheim N, Clark R, McCabe P, Innis M, Aldwin L, Nitecki D, McCormick F. Detection of activatedMr 21,000 protein, the product of ras oncogenes, using antibodies with specificity for amino acid 12. CancerRes 1986;46:6029–33.
Young RW, Droz B. The renewal of protein in retinal rods and cones. J Cell Biol 1968;39:169–84.




















