
cGMP Signalling Mediates Water Sensation (Hydrosensation ...
Upload
independentCategory
view
2download
0
Systemic signalling of environmental cues inArabidopsis leaves
S. A. Coupe1, B. G. Palmer1, J. A. Lake1, S. A. Overy1,*, K. Oxborough2, F. I. Woodward1, J. E. Gray3 and
W. P. Quick1,†
1 Department of Animal and Plant Sciences, The University of Sheffield, Western Bank, Sheffield S10 2TN, UK2 Department of Biological Sciences, University of Essex, Colchester CO4 3SQ, UK3 Department of Molecular Biology and Biotechnology, University of Sheffield, Sheffield S10 2TN, UK
Received 15 July 2005; Accepted 20 October 2005
Abstract
Light intensity and atmospheric CO2 partial pressure
are two environmental signals known to regulate
stomatal numbers. It has previously been shown that
if a mature Arabidopsis leaf is supplied with either
elevated CO2 (750 ppm instead of ambient at 370 ppm)
or reduced light levels (50 lmol m22 s21 instead of
250 lmol m22 s21), the young, developing leaves that
are not receiving the treatment grow with a stomatal
density as if they were exposed to the treatment. But
the signal(s) that it is believed is generated in the
mature leaves and transmitted to developing leaves are
largely unknown. Photosynthetic rates of treated,
mature Arabidopsis leaves increased in elevated CO2
and decreased when shaded, as would be expected.
Similarly, the levels of sugars (glucose, fructose, and
sucrose) in the treated mature leaves increased in
elevated CO2 and decreased with shade treatment. The
levels of sugar in developing leaves were also meas-
ured and it was found that they mirrored this result
even though they were not receiving the shade or
elevated CO2 treatment. To investigate the effect of
these treatments on global gene expression patterns,
transcriptomics analysis was carried out using Affy-
metrix, 22K, and ATH1 arrays. Total RNA was extracted
from the developing leaves after the mature leaves had
received either the ambient control treatment, the
elevated CO2 treatment, or the shade treatment, or
both elevated CO2 and shade treatments for 2, 4, 12, 24,
48, or 96 h. The experiment was replicated four times.
Two other experiments were also conducted, one to
compare and contrast gene expression in response to
plants grown at elevated CO2 and the other to look at
the effect of these treatments on the mature leaf. The
data were analysed and 915 genes from the untreated,
signalled leaves were identified as having expression
levels affected by the shade treatment. These genes
were then compared with those whose transcript
abundance was affected by the shade treatment in
the mature treated leaves (1181 genes) and with 220
putative ‘stomatal signalling’ genes previously identi-
fied from studies of the yoda mutant. The results of
these experiments and how they relate to environmen-
tal signalling are discussed, as well as possible
mechanisms for systemic signalling.
Key words: Elevated CO2, light, microarrays, mutants, shade,
stomata, systemic signal, transcriptomics.
Introduction
Stomata develop from meristemoids, which are locatedin the developing epidermal cell layer of plant leaves. Themeristemoid produces, directly or through a number ofasymmetric cell divisions, a guard mother cell, the imme-diate stomatal precursor cell. An equal division of the guardmother cell then leads to the formation of two guard cells,which, in conjunction with additional neighbouring epider-mal cells, constitute the stomatal complex (Serna andFenoll, 1997) that regulates the aperture of a stomatal pore.
* Present address: Department of Biology, University of York, PO Box 373, York YO10 5YW, UK.y To whom correspondence should be addressed. E-mail: [email protected]: CO2, carbon dioxide; IRGA, infrared gas analyser; PCA, principal components analysis; ppm, parts per million; NASC, Nottingham ArabidopsisStock Centre.
ª The Author [2005]. Published by Oxford University Press [on behalf of the Society for Experimental Biology]. All rights reserved.For Permissions, please e-mail: [email protected]
Journal of Experimental Botany, Vol. 57, No. 2, pp. 329–341, 2006
Phenotypic Plasticity and the Changing Environment Special Issue
doi:10.1093/jxb/erj033 Advance Access publication 5 December, 2005
by guest on May 7, 2015
http://jxb.oxfordjournals.org/D
ownloaded from

The stomatal complex represents a group of highlyspecialized cell types in the epidermal cell layer of severalhigher plant organs including leaves, stems, and reproduct-ive structures (Willmer and Fricker, 1996). This complexcontrols gas exchange between the plant and the atmos-phere. Proper control of pore size is critical for optimalplant performance. The guard cells of the stomatal complexsense and integrate many different exogenous signals(including CO2, light and water deficit) as well as en-dogenous signals (like phytohormone concentrations) inorder to balance CO2 uptake versus water loss in continu-ally changing environmental conditions (Schroeder et al.,2001) and these signalling pathways are likely to beintegrated into more complex scale-free networks ratherthan independent stand-alone pathways (Hetherington andWoodward, 2003).
The observation that increases in atmospheric CO2
concentration also affect stomatal numbers was the firstpiece of in situ evidence for a developmental response tothe increased CO2 emissions during the industrial era(Woodward, 1987). Changes in stomatal development,recorded as changes in stomatal density (stomatal numbersper unit area), have also provided unique information aboutthe CO2 environment during leaf development which isrecorded in fossils over geological time scales (Beerlingand Woodward, 1997). A critical feature of the develop-mental stomatal response to the environment is that theenvironmental signal is detected by mature leaves and thensignalled systemically to developing leaves. This has beenshown to occur for both CO2 concentration and lightintensity in Arabidopsis (Lake et al., 2001) and for lightintensity in tobacco (Thomas et al., 2004). This detection ofenvironmental signals by mature parts of the plant ensuresthat the most accurate measurement of the environmentaround the plant is determined by a wholly unfurled andmature leaf, rather than by (or in addition to) the developingleaf growing in a less representative environment (Larcher,1995). As well as CO2 and light, the same mature leavesalso detect a range of independent environmental chal-lenges such as pathogens, wounding, ethylene, and lowtemperatures that are also signalled systemically and havebeen reviewed elsewhere (Shinozaki et al., 2003; Alonsoand Stepanova, 2004; Braam, 2005; Glazebrook, 2005).More general and rapid systemic signals between leaveshave also been shown in Arabidopsis that involve theproduction of hydrogen peroxide within the vasculature inresponse to excess light (Karpinski et al., 1999), but theseresponses have not been demonstrated to target developingtissue and subsequent leaf development.
A number of insights have been made with respect tothe pathways regulating stomatal development. For ex-ample, a mitogen-activated protein kinase kinase kinase(MAPKKK) called YODA has been identified as a keyintermediary in stomatal formation, with mutation ofYODA in yoda mutant plants leading to ectopic stomatal
formation, and expression of a constitutively active form ofYODA (DN-YODA) resulting in the generation of leaveslacking stomata (Bergmann et al., 2004). With respect topatterning of stomata across the epidermal surface, TMM(the mutation of which leads to the formation of stomatalclusters) has been shown to encode a leucine-rich repeat-like receptor (LRR), suggesting that the protein acts asa component of a receptor for some unknown signal(Nadeau and Sack, 2002). It has been suggested that thissignal may be peptide-based as another gene involved instomatal patterning, SDD1, encodes a putative subtilisin-like serine protease (Berger and Altmann, 2000). Sucha protease may well be involved in signal processing. Fattyacid-derived signals have also been implicated in theenvironmental modulation of stomatal frequency as theHIC (high carbon dioxide) gene, which encodes a putative3-ketoacyl CoA synthase (KCS) believed to be involved inthe synthesis of cuticle waxes, affects the response ofstomatal development to changing CO2 concentration inArabidopsis (Gray et al., 2000). When the expression ofHIC is reduced, plants respond to elevated CO2 witha large increase in stomatal density (stomatal numbers perunit area) and index (stomatal numbers as the fraction ofall epidermal cells). This contrasts with decreases instomatal density and index for wild-type plants grown atelevated CO2. A model linking these components ofstomatal development has recently been proposed (Grayand Hetherington, 2004) but the signals that are perceivedand passed to these downstream components still remainunknown.
More recently, a family of three ERECTA (ER) genesthat encode leucine-rich repeat-receptor-like kinases havebeen shown to regulate two important steps in stomataldevelopment (Shpak et al., 2005): Firstly, the specificationof stomatal cell fate by influencing the initial decision ofa protodermal cell to undertake an asymmetric division toform a meristemoid cell. Secondly, the decision of themeristemoid terminally to differentiate into a guard mothercell. Interestingly, the three genes were shown to give riseto independent phenotypes and to act synergistically tocontrol stomatal development. TMM was also shown tointeract with ER and the authors speculated that the twoputative classes of membrane-bound receptor-like proteinsmay interact to form heterodimers that are inactive andprevent signalling. One interesting aspect of this work isthat the single er mutant or the double mutants er-er1 ander-er2 all led to increased numbers of stomatal cells withoutthe formation of stomatal clusters, whereas the triple mutanter-er1-er2 formed very large clusters of stomata (Shpaket al., 2005). Systemic signalling also regulates stomatalnumbers without the formation of clusters and this couldindicate an interaction with the ER gene family andsignalling pathway.
The roles of phytohormones (abscisic acid, auxin, bras-sinosteroids, cytokinins, ethylene, gibberellins, jasmonates)
330 Coupe et al.
by guest on May 7, 2015
http://jxb.oxfordjournals.org/D
ownloaded from

in regulating responses to environmental stimuli have beenknown for some time (Taiz and Zeiger, 2002). Recently,studies of mutants have begun to elucidate the mechanismsby which phytohormones integrate with biochemical signaltransduction pathways to regulate plant processes (Gray,2004), but there are still many gaps in our knowledge withrespect to receptors, signalling and integration of signallingpathways. Other signalling molecules, such as nutrients,Ca2+ and sugars, have also been implicated (Lake et al.,2002). It is likely that the CO2 response controllingstomatal development in Arabidopsis is affected by inputfrom systemic and local responses to other environmentalchallenges such as light, wounding, fungal infection,ethylene, and defoliation. The stomatal development re-sponse to CO2 has also been shown to be restored in mu-tants that are insensitive to these challenges (Lake et al.,2002).
There are several other possible sources of systemicsignals; leaves exposed to high atmospheric CO2 con-centrations typically increase their rates of carbohydrateproduction. Under these conditions, carbohydrate accumu-lation is known to regulate the expression of photosyntheticgenes such as ribulose-1,5-bisphosphate carboxylase/oxygenase and this has been proposed as a sugar-sensingmechanism (Sheen et al., 1999). In addition, sugar signal-ling is frequently found to affect/interact with otherhormonal signalling pathways mentioned earlier (Rollandand Sheen, 2005). High atmospheric CO2 treatments andlow light intensity treatments generally induce stomatalclosure and hence reduce transpiration rates (Taiz andZeiger, 2002). This might also affect the delivery ofhormones and sugars that are transported to leaves via thetranspiration stream (Pons et al., 2001).
The hypothesis was that the mature leaves wouldrespond to these treatments at the physiological, biochem-ical, and molecular level and that it would be possible todetect biochemical and molecular changes in the untreateddeveloping leaves and gain some insights into the environ-mental signalling network that modulates stomatal de-velopment. A cuvette system was used (Lake et al.,2001) where the mature leaves of an Arabidopsis plantwere enclosed and the growing environment could bemanipulated independently of the developing centralrosette leaves that were exposed to the environment ofthe growth chamber. Ambient air and air containingelevated levels of CO2 were supplied and a neutral densityfilter was used as a shade treatment to reduce the irradiancereceived by the mature leaves while the developing leavesreceived ambient air and were fully illuminated. Photosyn-thesis, carbohydrate content, and chlorophyll fluorescencewere measured and a large-scale transcriptomics experi-ment was conducted to assess the impact of these treatmentson global Arabidopsis gene expression, not only in themature, treated leaves but also in the untreated, developingleaves over a 4-d period.
Materials and methods
Plant growth
Seeds of Arabidopsis thaliana (ecotype Columbia, Col-0) werepurchased from Lehle Seeds (Texas, USA) and were germinated inHumax multi-purpose compost (Humax Horticulture Ltd., Cumbria,UK). The seedlings and subsequent plants were grown in two, side-by-side, constant temperature rooms; both kept at 20 8C, with lights at250 lmol m�2 s�1 supplied from cool fluorescent tubes (TLD 58W/840 Reflex, Philips, Netherlands), 10 h photoperiod starting at 09.00 hand relative humidity at 50%. Plants were watered every second day.The ambient room had a CO2 level of 370 ppm and the elevated roomnext door had a CO2 level of 750 ppm supplied from a CO2 cylinder(BOC Edwards, Surry, UK) attached to a Tylan mass flow controller(Munich, Germany) and filtered through a column of potassiumpermanganate to oxidize volatile organic compounds such asethylene. After 7 d the germinated seedlings were pricked out into8313 cell plug trays (LBS horticulture, Lancs, UK) and grown fora further 2 weeks before potting up into 10 cm pots. The plants thenhad the first half of the cuvette system added (Lake et al., 2001) whichwas the lid of a standard, 9 cm Petri dish with a 2.5 cm hole cut in themiddle and a foam ring added around it. This can be seen in Fig. 1.The foam (West Tapes, Sheffield, UK) rings were cut from a 2.5 m rollthat was 25 cm wide and 3 mm thick using two punches so that theinternal diameter was 2.5 cm and the external diameter was 3.5 cm.The plant continued growing through the hole and the mature leavesspread out over the foam disc. After 4 weeks, when the plants were atgrowth stage 3.50 (Boyes et al., 2001), leaves 5 to 13 were enclosedusing the lower part of an upturned Petri dish that had a 2.5 cm holeand two foam rings attached. The two halves of the Petri dish werethen reunited so that the sets of foam discs met in the middle to isolatethe mature leaves whilst the young developing leaves could grow outof the top. The upper and lower parts of the cuvette system were thenattached to each other and sealed with black insulation tape. The upperpart of the cuvette system had four 0.5 cm holes drilled into them andfour sets of plastic tubing were glued in place so that there was one gasinlet and three outlets (Fig. 1). Continual air was supplied from an oil-free compressor (Machine Mart, Sheffield, UK) with a 35 l reserve,using 4 mm silicone tubing (Portex, SLS, Nottingham, UK) that wasthen passed through water to humidify it and then was split into twolines of tubing to two manifolds (Air Accessories, Sheffield, UK) thatsupplied six flow-meters (K1100, Solartron Mobrey, Berks., UK)each, that allowed the air to be supplied at a controlled rate of 500ml min�1. A duplicate air flow system was set up in an adjacent roommaintained at 750 ppm CO2 and delivered to the ambient room viatubing. This meant that 12 plants in cuvettes could be supplied withCO2 at 370 ppm and 12 plants could be supplied with CO2 at 750 ppmthat was then externally exhausted from the growth room. The CO2
levels in each room were continually monitored using an IRGA(LCA3, ADC, Herts., UK) and were found to be constant even whenthe elevated CO2 was being pumped into the treatment cuvettes.The target developing leaves (insertions 16–19, see Fig. 1) were
marked with a dot of white acrylic paint before the mature leaves weresealed into the cuvette at 2 h after the start of the photoperiod (10.00 h).The plants were then left in the cuvette for 24 h to acclimatize andduring this time all 24 plants were continually supplied with ambientair containing 370 ppm CO2. 24 h later the target developing leavesfrom four plants were harvested and immediately frozen in liquid N2
(zero h control samples). Ten of the plants were switched over to aircontaining 750 ppm CO2 and the remaining ten plants received aircontaining ambient CO2. Each of these groups was then furthersubdivided into two groups of five plants with half receiving a shadetreatment which consisted of a 12 cm square piece of neutral densityfilter (Cat. No. 210, 0.6ND, Lee Filters, Hampshire, UK), that had hada 3.5 cm hole punched out of the middle, and was overlaid on top of
Systemic signalling in Arabidopsis 331
by guest on May 7, 2015
http://jxb.oxfordjournals.org/D
ownloaded from
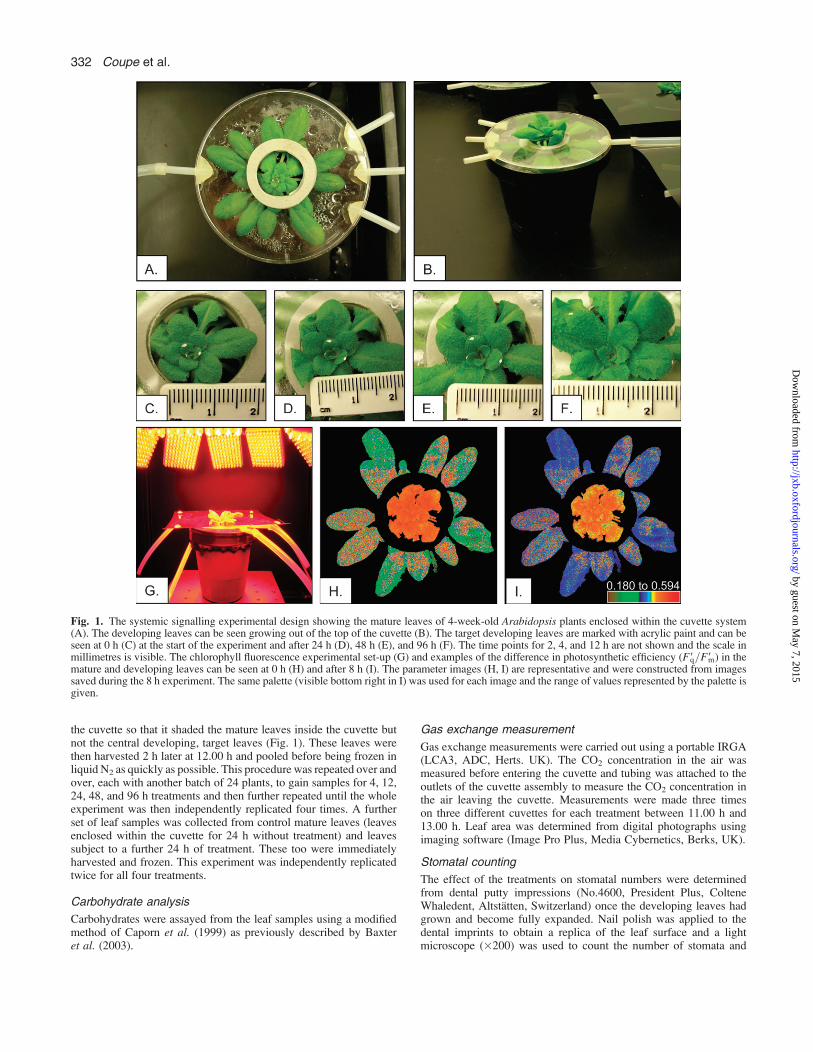
the cuvette so that it shaded the mature leaves inside the cuvette butnot the central developing, target leaves (Fig. 1). These leaves werethen harvested 2 h later at 12.00 h and pooled before being frozen inliquid N2 as quickly as possible. This procedure was repeated over andover, each with another batch of 24 plants, to gain samples for 4, 12,24, 48, and 96 h treatments and then further repeated until the wholeexperiment was then independently replicated four times. A furtherset of leaf samples was collected from control mature leaves (leavesenclosed within the cuvette for 24 h without treatment) and leavessubject to a further 24 h of treatment. These too were immediatelyharvested and frozen. This experiment was independently replicatedtwice for all four treatments.
Carbohydrate analysis
Carbohydrates were assayed from the leaf samples using a modifiedmethod of Caporn et al. (1999) as previously described by Baxteret al. (2003).
Gas exchange measurement
Gas exchange measurements were carried out using a portable IRGA(LCA3, ADC, Herts. UK). The CO2 concentration in the air wasmeasured before entering the cuvette and tubing was attached to theoutlets of the cuvette assembly to measure the CO2 concentration inthe air leaving the cuvette. Measurements were made three timeson three different cuvettes for each treatment between 11.00 h and13.00 h. Leaf area was determined from digital photographs usingimaging software (Image Pro Plus, Media Cybernetics, Berks, UK).
Stomatal counting
The effect of the treatments on stomatal numbers were determinedfrom dental putty impressions (No.4600, President Plus, ColteneWhaledent, Altstatten, Switzerland) once the developing leaves hadgrown and become fully expanded. Nail polish was applied to thedental imprints to obtain a replica of the leaf surface and a lightmicroscope (3200) was used to count the number of stomata and
Fig. 1. The systemic signalling experimental design showing the mature leaves of 4-week-old Arabidopsis plants enclosed within the cuvette system(A). The developing leaves can be seen growing out of the top of the cuvette (B). The target developing leaves are marked with acrylic paint and can beseen at 0 h (C) at the start of the experiment and after 24 h (D), 48 h (E), and 96 h (F). The time points for 2, 4, and 12 h are not shown and the scale inmillimetres is visible. The chlorophyll fluorescence experimental set-up (G) and examples of the difference in photosynthetic efficiency (F9q=F9m) in themature and developing leaves can be seen at 0 h (H) and after 8 h (I). The parameter images (H, I) are representative and were constructed from imagessaved during the 8 h experiment. The same palette (visible bottom right in I) was used for each image and the range of values represented by the palette isgiven.
332 Coupe et al.
by guest on May 7, 2015
http://jxb.oxfordjournals.org/D
ownloaded from

epidermal cells per mm2. The number of stomatal and epidermal cellswere counted in 10 fields of view from the three marked leaves offive individual plants for each treatment.
Chlorophyll fluorescence
The experiments detailed earlier were re-created inside a chlorophyllfluorescence imaging system (FluorImager, Technologica Ltd.,Colchester, UK). The aim was to quantify the treatment effects ofelevated CO2 concentration or reduced light intensity on the enclosedmature and untreated, developing leaves. Images of chlorophyllfluorescence parameters were obtained using this system that hasbeen described extensively elsewhere (Barbagallo et al., 2003;Oxborough, 2004; von Caemmerer et al., 2004). Two chlorophyllfluorescence parameters were calculated from the images to assesswhether changes in photosynthesis of the mature leaves resulted ina change in photosynthesis of the developing leaves. F9q=F9m is de-fined as the PSII photochemical efficiency (an indicator of photo-synthetic efficiency),F9q=F9v as the PSII photochemical factor (relatingthe maximum PSII photochemical efficiency, given by F9v=F9mand the actual PSII photochemical efficiency, given by F9q=F9mand is affected by the proportion of PSII reaction centres in theoxidized state.
Transcriptomics
Total RNA was extracted from the frozen leaf samples using theQiagen RNeasy kit (Qiagen, Crawley, UK) and each extract wasanalysed on an Agilent 2100 Bioanalyser (Agilent Technologies UK.,Wokingham, UK) to verify lack of rRNA degradation and used ina RT-PCR reaction with Actin2 primers, before being submitted toNASC (http://affymetrix.arabidopsis.info/) for hybridization to theATH1 Affymetrix gene chip arrays. The procedures used for theAffymetrix data production, collection and analysis (Craigon et al.,2004) were performed as described in the documentation, providedby NASC (http://affymetrix.arabidopsis.info). The data from 112Affymetrix hybridization experiments have been made publiclyavailable and downloadable from the NASC web-site at the follow-ing URLs:
http://affymetrix.arabidopsis.info/narrays/experimentpage.pl?experimentid=156
http://affymetrix.arabidopsis.info/narrays/experimentpage.pl?experimentid=157
http://affymetrix.arabidopsis.info/narrays/experimentpage.pl?experimentid=158
http://affymetrix.arabidopsis.info/narrays/experimentpage.pl?experimentid=159
http://affymetrix.arabidopsis.info/narrays/experimentpage.pl?experimentid=160
The data were deposited in October 2004 and were later distributedon Affywatch CDs 24–26 (NASC Affymetrix data subscriptionservice).
Gene chip data analysis
Hybridization data from the transcriptomics experiments were im-ported directly into GeneSpring software (ver 7.0, Agilent Technolo-gies, Wokingham, UK). The 112 data sets were normalized to the50th percentile (per chip) and to the median (per gene) and the datawere first analysed using the PCA tool and conditions set with allgenes and all 112 data sets included. The data were also normalizedper chip to the 50th percentile and per gene to the control samples(24 h ambient for the mature leaf data and the respective ambientcontrol at each time point for the developing leaf data). The datawere then filtered using four different filters: (i) by flags removinggenes that were absent in any replicate (no signal detected); (ii) by
expression level to remove those genes that were deemed to beunchanging between log values 0.8 and 1.2 (>1.5-fold difference);(iii) by a word filter to remove the Affymetrix control genes (AFFX);and (iv) by confidence using a one-sample t-test and a P-value cut-offof 0.05 so that any gene with a P-value of 0.05 or less when comparedto the normalized control was regarded as statistically significant, i.e.up- or down-regulated compared to the expression baseline of 1.(All genes were centred on 1 after normalization). To determine thosegenes that were statistically, differentially expressed between groupsof samples, GeneSpring’s ANOVA statistical analysis function(a Welch t-test) was applied to the filtered data set.
Results
Leaf development
Mature leaf insertions, 5 to 13, were enclosed in a plasticcuvette at 4 weeks (Fig. 1A, B). These leaves were exposedto elevated CO2 concentration or were shaded to reducelight intensity, or were subject to both treatments. The con-trol leaves were exposed to the same ambient conditions asthe developing leaves. Developing leaf insertions, 16–19,which were not inside the cuvette, were marked with a dotof acrylic paint (Fig. 1C) at 0 h, when they were 1–3 mm inlength and their growth was monitored. The growth ofleaves after 2, 4, and 12 h was minimal and hence are notshown. After 24 h the leaves had grown to 2–5 mm in length(Fig. 1D), after 48 h the leaves had grown to 3–8 mm inlength (Fig. 1E) and after 96 h they had grown to 6–12 mm(Fig. 1F). The shade treatment of mature leaves was foundto slow the growth of developing leaves (leaf length after4 dwas reduced by 26%)whereas treatment ofmature leaveswith elevated CO2 caused the developing leaves to growslightly faster (leaf length after 4 d was increased by 18%).
Stomatal counting
Plants were set up in the cuvette system and grown con-tinuously in the treatments until the marked developingleaves reached full expansion (2 weeks); these were thenharvested and the stomata counted. All of the results inFig. 2 are for developing or mature-treated leaves where themature leaves were maintained in ambient conditions (A),given a shade treatment (AS), or exposed to increased CO2
concentration (EA).Figure 2A shows data for stomatal density; as reported
previously increasing the CO2 concentration of matureleaves decreased the stomatal density of the developingleaves on both the adaxial and abaxial leaf surfaces (datanot shown). Similarly the shade treatment also causeda marked reduction in the stomatal density of the unshadeddeveloping leaves (Fig. 2A).
Gas exchange
When mature leaves were enclosed in the cuvette system(Fig. 1A, B) and treated with ambient CO2 and ambientlight (treatment A), the photosynthetic rate was found to be9.32 lmol m�2 s�1 (Fig. 2B). When a shade treatment (AS,
Systemic signalling in Arabidopsis 333
by guest on May 7, 2015
http://jxb.oxfordjournals.org/D
ownloaded from

which reduced the light intensity from 250 lmol m�2 s�1 to50 lmol m�2 s�1) was applied, this immediately reducedphotosynthesis to 1.98 lmol m�2 s�1. When air containingelevated CO2 was supplied (treatment EA) to the cuvettesystem, the photosynthetic rate was increased to 13.25lmol m�2 s�1 (Fig. 2B).
Carbohydrate content of mature and developing leaves
Mature leaves that were enclosed in the cuvettes and sup-plied with air containing elevated CO2 (AE) containedmore soluble carbohydrates, especially after 2–4 d, than
those receiving ambient CO2 (A) (see Fig. 2C). By contrast,leaves that were shaded (AS) contained significantly lesssoluble carbohydrates. When the sugar content of the de-veloping, untreated leaves was measured, the same trend ofhigher sugar concentrations in the plants where the matureleaves were receiving elevated CO2 was seen within 2 h oftreatment when compared with the ambient control (Fig.2D). The plants where the mature leaves were shaded hadsimilar amounts of sugars as the ambient control in thedeveloping leaves except at 4 h after treatment when sig-nificantly reduced levels of sugars were recorded (Fig. 2D).
0
5
10
15
AA AS EATreatment
Ph
oto
syn
th
etic R
ate
(µ
mo
l m
-2 s
-1)
0
100
200
300
400
A abx AS abx A adx AS adxTreatment
Sto
matal D
en
sity
0
20
40
60
80
100
Time (h)
To
tal S
ug
ar C
on
ten
t
(n
mo
ls m
g-1 F
W)
To
tal S
ug
ar C
on
ten
t
(n
mo
ls m
g-1 F
W)
A
AE
AS
A
AE
AS
05
101520253035
0 2 4 12 24 48 960 2 4 12 24 48 96Time (h)
0
0.2
0.4
0.6
0.8
1
Time (h)
Fq
'/F
m'
Fq
'/F
m'A
AEAS
AAEAS
AAEAS
AAEAS
0
0.2
0.4
0.6
0.8
1
1 2 3 4 5 6 7 8
1 2 3 4 5 6 7 81 2 3 4 5 6 7 8
1 2 3 4 5 6 7 8Time (h)
0
0.2
0.4
0.6
0.8
1
Time (h)
Fq
'/F
v' (q
P)
Fq
'/F
v' (q
P)
0
0.2
0.4
0.6
0.8
1
Time (h)
A. B.
C. D.
E. F.
G. H.
●
●●
●
Fig. 2. Arabidopsis stomatal density (A) and photosynthetic rate (B) in response to ambient (A or AA), shade (AS), or elevated CO2 (EA) treatments onabaxial (abx) or adaxial (adx) leaf surfaces. Total sugar content in mature (C) or developing/signalled (D) leaves in response to the signalling treatments.Chlorophyll fluorescence parameters of mature (E, H) and developing leaves (F, I) to assess photosynthetic performance and efficiency in response to thetreatments applied to mature leaves. Arrows denote at what point the treatments were started. Data are the means of three replicates 6SE.
334 Coupe et al.
by guest on May 7, 2015
http://jxb.oxfordjournals.org/D
ownloaded from

Chlorophyll fluorescence
Images of chlorophyll fluorescence were recorded todetermine fluorescence quenching parameters (Barbagalloet al., 2003). Using a FluorImager (Fig. 1G) images ofF9q=F9m for a plant in the cuvette system receiving ambientair can be seen in Fig. 1H, I. When a shade treatment wasapplied to the mature leaves, F9q=F9m increased in theseleaves (Fig. 1I). This effect can be seen more clearly in Fig.2E–I, where there is a clear increase in F9q=F9m and also inF9q=F9v in the shaded mature leaves, relative to the ambientcontrol and a decrease in F9q=F9v in response to elevatedCO2 concentration (Fig. 2E, H). By contrast, F9q=F9v andF9q=F9m were unchanged in the untreated, developingleaves for both elevated CO2 and shade treatment of matureleaves (Fig. 2F, I).
Microarray data analysis
Over 2000 plants and 200 RNA extractions were performedto generate samples with replicates that were hybridized to112 Affymetrix gene chip arrays. The data were importedinto the GeneSpring data analysis package and, after theinitial standard normalization, the data were subjected toa principal components analysis (PCA) (data not shown). Itwas clear from the PCA (Fig. 3) that the data from themature leaf samples group vary differently from thedeveloping leaf samples. It was also clear that the time ofsampling, independent of treatment, was an importantparameter perhaps reflecting a circadian rhythm. For thepurposes of this study the focus was only on those genes
whose expression changed in response to the shadetreatment; there is insufficient space within the article topresent the data concerning the effects of CO2 and thecombination of shade and CO2; these will be the subject ofa future article. The focus was also on those genes whoseexpressions were altered in mature leaves that received theshade treatment directly (treated mature) and genes whoseexpressions were altered in developing leaves that did notreceive the shade treatment (signalled leaves).
Genes that showed high variation in expression betweenreplicates or were not present in all of the 112 sampleswere removed and this reduced the number of genes to16 397 in signalled leaves and 14 972 genes in treatedmature leaves. The data were then filtered to remove thosegenes whose expression level changed up or down lessthan 1.5-fold of the control treatment at each respectivetime point; this further reduced the number of genes to 12837 in signalled leaves and 10 719 in treated matureleaves. Lastly, the data were analysed using a t-test toidentify those genes whose expression changed and hada low P-value score (up to 0.05 in at least 1 condition andno multiple testing corrections) which left 11 397 genes insignalled leaves and 4163 genes in treated mature leaves.A second t-test was then performed using the Benjaminiand Hochberg false discovery rate and using ‘variances notassumed equal’ along with a Tukey post hoc test and thisreturned 915 genes that change expression in signalledleaves and 1181 in treated mature leaves that werestatistically significantly different between the ambientand shade treatments.
Fig. 3. Principal components analysis of data obtained from hybridization experiments with 112 Affymetrix microarray chips and mature anddeveloping leaf extracts over the period of 96 h after shade and elevated CO2 treatments were applied to mature leaves. The PCA plot was generatedusing GeneSpring v. 7.0.
Systemic signalling in Arabidopsis 335
by guest on May 7, 2015
http://jxb.oxfordjournals.org/D
ownloaded from

Functional classification and candidatesignalling genes
The two groups of genes that showed at least a 1.5-foldchange in expression level in at least one of the time pointsfollowing shading of the mature leaves were classifiedaccording to their putative functions. This was based ontheir classification in the Munich Information Centre forProtein Sequence (MIPS) database to determine whetheruntreated developing leaves differed in their gene expres-sion patterns compared with the treated mature leaves.Figure 4A and B show a summary of the functionalcharacterization of those transcripts identified as havingaltered expression patterns in treated mature and untreateddeveloping leaves. The data show that, where gene functioncan be assigned, the majority of transcripts where expres-sion was altered encode proteins that were associatedwith protein turnover and metabolism in the shaded matureleaves and metabolism and cell wall/development in thedeveloping, untreated leaves. Seven-fold more genes in-volved in photosynthesis were altered in the mature leavescompared with the untreated developing leaves. The re-maining categorieswere quite similarwith the exception that5% of the genes in the mature leaves were predicted to beinvolved in translation compared with only 1% in thedeveloping leaves.
When the two sets of gene lists (i.e. genes with alteredexpression by the shading of mature leaves) were comparedand contrasted using a Venn diagram in Fig. 4C, there were998 genes specific to the shaded mature leaves and 732genes that were specific to the signalled developing leaves,with 183 genes common to both. Examples from these threegroups of genes can be seen in Table 1. There were severalsuitable candidate genes in the ‘treated-mature’ list thatmight be involved in signalling to developing leaves, forexample, eight auxin-related genes, one brassinosteroid-related gene, one gibberellin-related gene, a putative mitogenactivated protein kinase kinase (MAPKK), and a putativesugar transporter. Examination of the untreated developingleaf gene list (Table 1) shows there were several interest-ing candidate genes that might interact with a signal fromthe mature leaves and these include CER2, EIN3, andSDD1.
The inclusion of SDD1 and the closely related, SDD1-like (At1g01900), in the list of ‘signalled’ genes thatrespond in developing leaves to the shade treatment in themature leaves is interesting and fits in with previouslyproposed models of stomatal development in developingArabidopsis leaves (Bergmann, 2004). There was a need tofind out if any other genes that have been implicated insignalling stomatal development were present in the twolists. Recently, a microarray experiment was carried outusing a mutant as controlling stomatal development andpatterning and encoding a putative MAPKKK calledYODA (Bergmann et al., 2004). 220 genes were identified
that changed expression pattern in opposing directions inthe mutant yoda (excess stomata) and plants expressing theDN-YODA protein (no stomata). The list of these 220YODA-related genes imported into GeneSpring are com-pared with the two gene lists described earlier (Table 1),again using a Venn diagram (Fig. 4D). Twelve of theYODA-related genes were specific to the mature leaf list,nine were specific to the untreated developing leaf list, andsix were common to both (Table 2).
Discussion
The search for a systemic signal
Stomatal density and index generally decrease with de-creasing light intensity. It has previously been shown thatshading of only mature Arabidopsis leaves also leads tountreated developing leaves growing as if they havereceived the shade treatment with a lower stomataldensity/index when compared with control plants (Lakeet al., 2001). It has been shown that this also occurs onboth surfaces of the developing leaf in the shade signal-ling system used here. These results suggest that signalstriggered by a change in irradiance are transmitted frommature to young leaves where they act to regulate stomataldevelopment (Lake et al., 2001; Thomas et al., 2004). Theresults also indicate that the effect of changing CO2
concentration (on the mature leaves) has an effect onstomatal development in young leaves and that the effectinteracts directly with the light signal (data not shown),suggesting an interdependence of the respective signallingmechanisms.
These CO2 and light treatments caused large changes tothe gas exchange parameters and photosynthetic rates of themature leaves enclosed within the treatment cuvette whencompared with ambient controls. The reduction in photo-synthesis correlated well with the reduction in light in-tensity caused by the shade treatment (Fig. 2B) whilst theincrease in response to elevated CO2 was also as expected.The rates of gas exchange observed for these plants arewithin the range of values previously published (Lake,2004).
When the total soluble sugar content of the enclosed,treated, mature leaves was measured there was a markeddecrease in the level of sugar in response to the shadetreatment even after 2 h and the trend continued throughoutthe 96 h time-course (Fig. 2C). Increased sugar concen-trations (sucrose, glucose, and fructose) were measured inmature leaves in response to the elevated CO2 treatment asexpected with increased photosynthesis. Altered sucroseconcentrations are likely to directly impact the rate of sugarexport. This may at least partly explain why the developingleaves grew at different rates in response to the twotreatments. This was further supported by the measurementof the sugar content of the untreated developing leaves
336 Coupe et al.
by guest on May 7, 2015
http://jxb.oxfordjournals.org/D
ownloaded from

(Fig. 2D) which were much higher (even after 2 h) in theplants where the mature leaves had been supplied withelevated CO2 up to 24 h. The trend was less obvious wherethe mature leaves had been shaded, but a clear decreasecould be seen at 4 h. There was therefore a direct effect ofthis study’s treatments to mature leaves on the sugar statusand growth rate of the developing leaves concomitant withaltered rates of phloem transport between the mature anddeveloping leaves.
Chlorophyll fluorescence parameters were also measuredin the mature and developing leaves to determine if any ofthe treatments were causing changes in photosynthesis of
the developing leaves that could not be enclosed in themeasuring cuvette of this laboratory’s gas-exchange equip-ment (Fig. 2E–I). Chlorophyll fluorescence imaging gavethe ability to assess both the enclosed mature leaves andthe exposed developing leaves simultaneously during thetreatment period. Chlorophyll fluorescence, measures ofphotosynthetic efficiency (F9q=F9m and F9q=F9v), increased inthe mature leaves in response to shade, suggesting de-creased reduction of PSII reaction centres and reducedlevels of non-photosynthetic energy dissipation, as expected(Fig. 2E, H). The developing leaves, however, showedno change, suggesting that there was no photosynthetic
Fig. 4. A pie-chart comparison of predicted functional distribution of shade-regulated genes in treated mature (A) and untreated developing (B) leaves.Distribution of mature and developing leaf profiles among 12 major classes was performed using the MIPs database (http://mips.gsf.de/proj/thal/db/tables/tables_func_frame.html). (C) Venn diagram to compare and contrast the 1181 shade-responsive genes present in the mature leaf and the 915shade-responsive genes present in the signalled leaves. 20 897 remaining genes were filtered out as they were not consistently expressed or notresponsive to the treatment and were not included in the analysis. (D) Venn diagram to compare and contrast the shade-responsive genes from the matureand developing leaves with the 220 YODA-related genes thought to be important in controlling stomatal fate (Bergmann et al., 2004). The Venndiagrams were generated using GeneSpring v. 7.0.
Systemic signalling in Arabidopsis 337
by guest on May 7, 2015
http://jxb.oxfordjournals.org/D
ownloaded from

response to the treatments of the mature leaves (Fig. 2F, I).So, clearly, the treatments are causing changes in photosyn-thesis, sugar content, and chlorophyll fluorescence parame-ters of the mature leaf and the end result of that is a change instomatal numbers and sugar content of the untreated de-veloping leaves. It is likely therefore that the differences insugar content may be one source of the signal being passedfrom mature to developing leaves. However, it should benoted that increased CO2 and shade have opposite effects onthe sugar concentration of the mature leaves yet both treat-ments lead to a decline in stomatal numbers of the devel-oping leaves.
Can transcriptomics help identify signallingcandidates?
It is clear from the explosion of transcriptomics data inrecent years that characterizing an organism’s transcrip-tomic response to a treatment is a powerful way of ident-
ifying many new genes that respond to a specific stimuli.Treatments such as circadian rhythms (Harmer et al., 2000),nitrate (Wang et al., 2003), shade (Devlin et al., 2003),sucrose starvation (Contento et al., 2004), pathogens(Eulgem et al., 2004), senescence (Buchanan-Wollastonet al., 2005), and hormones such as abscisic acid (Leonhardtet al., 2004), auxin and brassinosteroids (Goda et al., 2004)have been used to identify candidate genes that might beinvolved in these processes. Transcriptomic approacheshave also been used to characterize those differencesthat a mutation in a specific gene can cause in the widertranscriptome, as recently shown for the yoda mutant(Bergmann et al., 2004). These experiments, however, allfocus on changes in gene expression occurring directly inthe tissues that were receiving the treatment or carried themutation. The responses of a plant’s transcriptome havebeen measured not only in the mature leaves receiving theshade and elevated CO2 treatments but also in the untreated
Table 1. Examples of gene annotations from the 1181 treated mature leaf and 915 untreated developing leaf shade-responsive genelists generated by analysis of the microarray experiments
Those genes in bold show statistically significant increases in expression and those not in bold show statistically significant decreases in expression.
Mature leaf only Both Signalled leaves only
Description AGI Description AGI Description AGI
Auxin transport At2g01420 Carbonic anhydrase At3g08510 Phospholipase C At1g58180Auxin responsive 7 At3g23050 Light regulated protein At3g26740 CER2 At4g24510Auxin response factor At1g19220 Auxin-regulated At2g33830 EIN3 At3g20770Auxin PIN7 protein At1g23080 Senescence protein At3g22550 SDD1 At1g04110Auxin regulatory protein At4g39400 SEN1 protein At4g35770 Expansin At1g66350Auxin PIN3 protein At1g70940 Zinc finger protein At5g22920 GA3 signalling At3g29030Auxin ARF1 protein At2g46530 Calnexin 1 At5g61790 SDD1-like At1g01900Auxin protein IAA1 At4g14560 Glucose transporter At1g11260 GA3 SPINDLY At3g11540MAPKK At4g08500 Myb T.F. protein At1g22640 MAPK17 At2g01450Brassinosteroid 1 At4g39400 Protein phosphatase At2g25620 Scarecrow T.F. At1g21450Sugar transporter At2g48020 Amino acid transport At2g40420 Sugar transporter At4g04750Gibberellin signalling At1g14920 Neoxanthin protein At4g19170 Auxin protein At3g07760Aquaporin PIP1;5 At4g23400 Drought protein At1g56280 Dof protein At5g60850Aquaporin TIP2;1 At3g16240 EIN3 related 1 At2g27050 Aquaporin TIP2;2 At4g17340
Table 2. Results of the microarray data analysis to show the distribution of YODA-related genes within the 1181 mature and 915signalled leaf genes that respond to shade
Yoda mature leaf only Yoda both Yoda signalled leaves only
Description AGI Description AGI Description AGI
Kinase At1g21920 AP2 domain protein At1g79700 SDD1 At1g04110Auxin protein At1g29510 Glycine- rich protein At2g05540 Gibberellin 3-b-hydroxylase At1g15550Unknown At1g47400 Serine peptidase At2g22980 Histone protein At2g18050Lysine decarboxylase At2g28300 Zinc finger protein At2g25900 Tropinine reductase At2g29290Unknown At2g38210 Unknown At4g19160 Protein kinase At2g41820PEPC At2g42600 E2 ubiquitin protein At5g41700 Acyl-CoA synthase At2g47240bZIP T.F. At3g58120 Glycosyl hydrolase 36 At3g57520Glycosyl hydrolase 19 At3g61060 DHFR At5g42800Peroxidase At4g01700 Unknown At5g54970Unknown At4g08390Unknown At5g02020Arabinogalactan At5g44130
338 Coupe et al.
by guest on May 7, 2015
http://jxb.oxfordjournals.org/D
ownloaded from

developing leaves over the course of 4 d with the aim ofidentifying genes that may be involved in the systemicsignalling of environmental stimuli.
Initial observations of the microarray data showed that itwould easily be possible to discriminate between themature leaf samples and the signalled leaves using PCAanalysis (Fig. 3). PCA has previously been shown to bea useful tool for revealing subtle differences in largemetabolic datasets (Overy et al., 2005). It was also foundthat time was a significant principal component that couldbe used to distinguish the treatments in the time-course.Given that circadian rhythms have already been shown toinfluence gene expression significantly (Harmer et al.,2000), it was necessary to include an untreated control ateach time point and that the data were normalized to thesecontrol time points rather than a zero h time point.
After the data were filtered and analysed statistically,1181 genes were identified from the mature leaf experi-ments that were differentially expressed between theambient and shade treatments. 915 genes were differentiallyexpressed between the ambient and shade treatments in theuntreated developing leaves. Those genes whose expres-sion was altered by shading mature leaves were groupedinto functional classes (Fig. 4A, B). The classifications ofboth sets of affected genes are quite similar but there areseveral differences, for example, the mature leaf set hasseven photosynthesis genes that form part of the metabol-ism category, but the signalled leaves show only one. Thisis perhaps not surprising as it has already been shown (bychlorophyll fluorescence, Fig. 2) that the signalled leavesdo not show a photosynthetic response when the matureleaves were treated. There were also more differences ingenes involved in protein turnover and translation in themature leaves compared with the developing leaves and thismay be related to the changes in gene expression inresponse to the treatments, as opposed to the untreateddeveloping leaves. The developing leaves have a largerproportion of affected genes involved in cell wall synthesisand cell development which reflect the different stage ofgrowth and development.
The Venn diagram (Fig. 4C) identifies 998 genes withexpression patterns specifically affected in the treatedmature leaves, 732 in the untreated developing leaves and183 genes in both. A high proportion of auxin signallinggenes; for example, PIN3, PIN7, and IAA1 (Table 1) wereobserved for mature leaves. There was also a putativeMAPKK in the list of genes affected in mature leaves. Thismay be of interest as a MAPK signalling pathway is im-plicated in stomatal development (Bergmann et al., 2004).There were also components of the brassinosteroid (BRI1)and gibberellin (GA, GAI) signalling pathways that wereaffected. Many of the genes in this list have previously beenshown to be regulated by shade (Devlin et al., 2003) andthis convergence of light and hormone signalling might beparticularly interesting when looking for candidates that
might be involved in environmental signalling from matureto developing leaves.
From a list of known genes specifically affected in un-treated developing leaves (Table I) it is interesting to notethe presence of EIN3 and SDD1 as well as components ofthe GA signalling pathway. These two sets of genes werecompared with the 220 genes that contained 109 up-regulated genes in yoda and down-regulated in DN-YODAand 111 up-regulated genes in DN-YODA and down-regulated in yoda that were hypothesized to be closely in-volved in guard cell specification and function (Bergmannet al., 2004). The Venn diagram (Fig. 4) showed that 12of these genes were present in the treated mature leafgene list and nine were present in the untreated developingleaf gene list and six were present in both gene lists (Table2). Again auxin and GA signalling related genes are presentas well as putative transcription factors, kinases andglycosyl hydrolases but one of the most interesting findingswas the presence of SDD1. SDD1 encodes a putativesubtilisin-like serine protease and mutations in SDD1 leadto elevated stomatal density and frequent violations of the1-cell spacing rule that normally prevents individualstomata developing next to each other (Nadeau and Sack,2003). The full role that SDD1 plays is unknown, but it isthought to act in a common signalling pathway (along withTMM, YODA, and ERECTA proteins) to cleave a ligandthat is perceived by TMM and a co-receptor kinase (Nadeauand Sack, 2003). In these results SDD1 and the closelyrelated SDD1-like (At1g01900) are both down-regulated inthe signalled leaves in response to the shade treatmentof mature leaves (data not shown). These two genes share43% identity at the amino acid level and out of the 56Arabidopsis subtilisin-like genes are the most closelyrelated (see http://web.uni-frankfurt.de/fb15/botanik/mcb/AFGN/altmann.htm) and have been renamed AtSBT1.1(SDD1-like) and AtSBT1.2 (SDD1). The subtilisins havediverse roles in plant development and have been impli-cated in epidermal development in Arabidopsis embryos(Tanaka et al., 2001) and in auxin-induced root hairformation (Xie et al., 2000). The photosynthetic perform-ance of sdd1 has been evaluated (Schluter et al., 2003)and they found that the mutant accumulated 30% morestarch and sugar than wild-type under increasing light butconclude that SDD1 is not responsible for environmentallymediated regulation of stomatal density. These preliminaryresults suggest that SDD1 expression is affected in de-veloping leaves by shading mature leaves.
In summary, this study of a systemic signalling systemhas suggested that both sugars and hormones may playa part in systemic signalling but that further work is requiredto discover the precise mechanism. It has recently beenshown that leaf morphogenesis can be controlled bymicroRNAs (Palatnik et al., 2003) and these have beenimplicated in auxin signalling (Kidner and Martienssen,2005). Ten per cent of the 915 genes identified as having
Systemic signalling in Arabidopsis 339
by guest on May 7, 2015
http://jxb.oxfordjournals.org/D
ownloaded from

altered expression in treated developing leaves and 70%of the forty putative transcription factors from this genelist contain high homology microRNA binding domainsas identified on the Arabidopsis small RNA web site(http://asrp.cgrb.oregonstate.edu/). It has also been demon-strated that an mRNA member of the GRAS gene family,a negative regulator of gibberellic acid responses, isdelivered by the phloem from developing leaves to leafmeristems and alters subsequent leaf development. Thissuggests possible mRNA candidates as systemic signal-lers. Furthermore, this mRNA did not affect all newlydeveloped leaves but rather was position-dependent ingrafting experiments; increasing distance from the sourceof mRNA reduced both the quantity of mRNA arriving atthe meristem and efficacy in altering leaf development(Haywood et al., 2005). This type of system seemsappropriate in ensuring that only signals from ‘nearby’mature leaves are effective in this systemic signallingpathway and provides a mechanism whereby developingleaves are optimized for performance under the prevailingenvironmental conditions.
Acknowledgements
The authors would like to thank NASC for carrying out theAffymetrix microarray hybridization experiments, Dr Steven Rolfefor help with the chlorophyll fluorescence image analysis, andundergraduate students Michael Hahn, Jonathon Freer, and DanielHungerford, for help with the growing of Arabidopsis plants, andDr Bela Tiwari and the bioinformatics team at CEH, Oxford for helpwith data anlysis and access to GeneSpring through the NERCGenomics Thematic Programme. This work was funded by NERCand BBSRC grants to JE Gray, WP Quick and FI Woodward.
References
Alonso JM, Stepanova AN. 2004. The ethylene signaling pathway.Science 306, 1513–1515.
Barbagallo RP, Oxborough K, Pallett KE, Baker NR. 2003.Rapid, noninvasive screening for perturbations of metabolismand plant growth using chlorophyll fluorescence imaging. PlantPhysiology 132, 485–493.
Baxter CJ, Foyer CH, Turner J, Rolfe SA, Quick WP. 2003.Elevated sucrose-phosphate synthase activity in transgenic tobaccosustains photosynthesis in older leaves and alters development.Journal of Experimental Botany 54, 1813–1820.
Beerling DJ, Woodward FI. 1997. Changes in land plant functionover the Phanerozoic: reconstructions based on the fossil record.Botanical Journal of the Linnean Society 124, 137–153.
Berger D, Altmann T. 2000. A subtilisin-like serine proteaseinvolved in the regulation of stomatal density and distribution inArabidopsis thaliana. Genes and Development 14, 1119–1131.
Bergmann DC. 2004. Integrating signals in stomatal development.Current Opinion in Plant Biology 7, 26.
Bergmann DC, Lukowitz W, Somerville CR. 2004. Stomataldevelopment and pattern controlled by a MAPKK kinase. Science304, 1494–1497.
Boyes DC, Zayed AM, Ascenzi R, McCaskill AJ, Hoffman NE,Davis KR, Gorlach J. 2001. Growth stage-based phenotypic
analysis of Arabidopsis: a model for high throughput functionalgenomics in plants. The Plant Cell 13, 1499–1510.
Braam J. 2005. In touch: plant responses to mechanical stimuli.New Phytologist 165, 373–389.
Buchanan-Wollaston V, Page T, Harrison E, et al. 2005. Com-parative transcriptome analysis reveals significant differences ingene expression and signalling pathways between developmentaland dark/starvation-induced senescence in Arabidopsis. The PlantJournal 42, 567–585.
Caporn SJM, Brooks AL, Press MC, Lee JA. 1999. Effects oflong-term exposure to elevated CO2 and increased nutrient supplyon bracken (Pteridium equilinum). Functional Ecology 13,107–115.
Contento AL, Kim S-J, Bassham DC. 2004. Transcriptome pro-filing of the response of Arabidopsis suspension culture cells to sucstarvation. Plant Physiology 135, 2330–2347.
Craigon DJ, James N, Okyere J, Higgins J, Jotham J, May S.2004. NASCArrays: a repository for microarray data generatedby NASC’s transcriptomics service. Nucleic Acids Research 32,D575–577.
Devlin PF, Yanovsky MJ, Kay SA. 2003. A genomic analysis ofthe shade avoidance response in Arabidopsis. Plant Physiology133, 1617–1629.
Eulgem T, Weigman VJ, Chang H-S, McDowell JM, Holub EB,Glazebrook J, Zhu T, Dangl JL. 2004. Gene expression sig-natures from three genetically separable resistance gene signalingpathways for downy mildew resistance. Plant Physiology 135,1129–1144.
Glazebrook J. 2005. Contrasting mechanisms of defense againstbiotrophic and necrotrophic pathogens. Annual Review of Phyto-pathology 43, 9.1–9.23.
Goda H, Sawa S, Asami T, Fujioka S, Shimada Y, Yoshida S.2004. Comprehensive comparison of auxin-regulated andbrassinosteroid-regulated genes in Arabidopsis. Plant Physiology134, 1555–1573.
Gray JE, Hetherington AM. 2004. Plant development: YODA thestomatal switch. Current Biology 14, R488.
Gray JE, Holroyd GH, van der Lee FM, Bahrami AR, SijmonsPC, Woodward FI, Schuch W, Hetherington AM. 2000.The HIC signalling pathway links CO2 perception to stomataldevelopment. Nature 408, 713.
Gray WM. 2004. Hormonal regulation of plant growth and de-velopment. Public Library of Science, Biology 2, e311.
Harmer SL, Hogenesch JB, Straume M, Chang H-S, Han B,Zhu T, Wang X, Kreps JA, Kay SA. 2000. Orchestratedtranscription of key pathways in Arabidopsis by the circadianclock. Science 290, 2110–2113.
Haywood V, Yu T-S, Huang N-C, Lucas WJ. 2005. Phloem long-distance trafficking of GIBBERELLIC ACID-INSENSITIVERNA regulates leaf development. The Plant Journal 42, 49–68.
Hetherington AM, Woodward FI. 2003. The role of stomata insensing and driving environmental change. Nature 424, 901–908.
Karpinski S, Reynolds H, Karpinska B, Creissen G, Wingsle G,Mullineaux P. 1999. Systemic signalling and acclimation in re-sponse to excess excitation energy in Arabidopsis. Science 284,654–657.
Kidner CA, Martienssen RA. 2005. The developmental role ofmicroRNA in plants. Current Opinion in Plant Biology 8, 38.
Lake JA. 2004. Gas exchange: new challenges with Arabidopsis.New Phytologist 162, 1–3.
Lake JA, Quick WP, Beerling DJ, Woodward FI. 2001.Plant development: signals from mature to new leaves. Nature411, 154.
Lake JA, Woodward FI, Quick WP. 2002. Long-distance CO2
signalling in plants. Journal of Experimental Botany 53, 183–193.
340 Coupe et al.
by guest on May 7, 2015
http://jxb.oxfordjournals.org/D
ownloaded from

Larcher W. 1995. Physiological plant ecology, 3rd edn. Germany:Springer-Verlag.
Leonhardt N, Kwak JM, Robert N, Waner D, Leonhardt G,Schroeder JI. 2004. Microarray expression analyses of Arabi-dopsis guard cells and isolation of a recessive abscisic acidhypersensitive protein phosphatase 2C mutant. The Plant Cell16, 596–615.
Nadeau JA, Sack FD. 2002. Control of stomatal distribution on theArabidopsis leaf surface. Science 296, 1697–1700.
Nadeau JA, Sack FD. 2003. Stomatal development: cross talk putsmouths in place. Trends in Plant Science 8, 294.
Overy SA, Walker HJ, Malone S, Howard TP, Baxter CJ,Sweetlove LJ, Hill SA, Quick WP. 2005. Application ofmetabolite profiling to the identification of traits in a populationof tomato introgression lines. Journal of Experimental Botany 56,287–296.
Oxborough K. 2004. Imaging of chlorophyll a fluorescence:theoretical and practical aspects of an emerging technique for themonitoring of photosynthetic performance. Journal of Experi-mental Botany 55, 1195–1205.
Palatnik JF, Allen E,WuX, Schommer C, Schwab R, CarringtonJC, Weigel D. 2003. Control of leaf morphogenesis by micro-RNAs. Nature 425, 257–263.
Pons TL, Jordi W, Kuiper D. 2001. Acclimation of plants to lightgradients in leaf canopies: evidence for a possible role forcytokinins transported in the transpiration stream. Journal ofExperimental Botany 52, 1563–1574.
Rolland F, Sheen J. 2005. Sugar sensing and signalling networks inplants. Biochemical Society Transactions 33, 269–271.
Schluter U, Muschak M, Berger D, Altmann T. 2003. Photosyn-thetic performance of an Arabidopsis mutant with elevatedstomatal density (sdd1-1) under different light regimes. Journalof Experimental Botany 54, 867–874.
Schroeder JI, Allen GI, Hugouvieux V, Kwak JM, Warner D.2001. Guard cell signal transduction. Annual Review of PlantPhysiology and Plant Molecular Biology 52, 627–658.
Serna L, Fenoll C. 1997. Tracing the ontogeny of stomatal clustersin Arabidopsis with molecular markers. The Plant Journal 12,747–755.
Sheen J, Zhou L, Jang JC. 1999. Sugars as signalling molecules.Current Opinion in Plant Biology 2, 410–418.
Shinozaki K, Yamaguchi-Shinozaki K, Seki M. 2003. Regulatorynetwork of gene expression in the drought and cold stressresponses. Current Opinion in Plant Biology 6, 410.
Shpak ED, Messmer McAbee J, Pillitteri LJ, Torii KU. 2005.Stomatal patterning and differentiation by synergistic interactionsof receptor kinases. Science 309, 290–293.
Taiz L, Zeiger E. 2002. Plant physiology, 2nd edn. Sunderland,Massachusetts: Sinauer Associates Inc., Publishing.
Tanaka H, Onouchi H, Kondo M, Hara-Nishimura I, NishimuraM, Machida C, Machida Y. 2001. A subtilisin-like serineprotease is required for epidermal surface formation in Arabidopsisembryos and juvenile plants. Development 128, 4681–4689.
Thomas PW, Woodward FI, Quick WP. 2004. Systemic irradiancesignalling in tobacco. New Phytologist 161, 193–198.
von Caemmerer S, Lawson T, Oxborough K, Baker NR,Andrews TJ, Raines CA. 2004. Stomatal conductance does notcorrelate with photosynthetic capacity in transgenic tobacco withreduced amounts of Rubisco. Journal of Experimental Botany55, 1157–1166.
Wang R, Okamoto M, Xing X, Crawford NM. 2003. Microarrayanalysis of the nitrate response in Arabidopsis roots and shootsreveals over 1000 rapidly responding genes and new linkages toglucose, trehalose-6-phosphate, iron, and sulfate metabolism.Plant Physiology 132, 556–567.
Willmer C, Fricker M. 1996. Stomata, 2nd edn. London: Chapmanand Hall.
Woodward FI. 1987. Stomatal numbers are sensitive to increses inCO2 from pre-industrial levels. Nature 327, 617–618.
Xie Q, Frugis G, Colgan D, Chua N-H. 2000. Arabidopsis NAC1transduces auxin signal downstream of TIR1 to promote lateralroot development. Genes and Development 14, 3024–3036.
Systemic signalling in Arabidopsis 341
by guest on May 7, 2015
http://jxb.oxfordjournals.org/D
ownloaded from
Copyright © 2022 FDOKUMEN




















