
Enhanced effects from tiny flexible in-wall blips and shear flow
Upload
independentCategory
view
4download
0
This article appeared in a journal published by Elsevier. The attachedcopy is furnished to the author for internal non-commercial researchand education use, including for instruction at the authors institution
and sharing with colleagues.
Other uses, including reproduction and distribution, or selling orlicensing copies, or posting to personal, institutional or third party
websites are prohibited.
In most cases authors are permitted to post their version of thearticle (e.g. in Word or Tex form) to their personal website orinstitutional repository. Authors requiring further information
regarding Elsevier’s archiving and manuscript policies areencouraged to visit:
http://www.elsevier.com/copyright

Author's personal copy
Systematic relationships of Oriental tiny frogs of the family Microhylidae(Amphibia, Anura) as revealed by mtDNA genealogy
Masafumi Matsui a,⇑, Amir Hamidy a,b, Daicus M. Belabut c,d, Norhayati Ahmad d,e, Somsak Panha f,Ahmad Sudin g, Wichase Khonsue f, Hong-Shik Oh h, Hoi-Sen Yong c, Jian-Ping Jiang i, Kanto Nishikawa a
a Graduate School of Human and Environmental Studies, Kyoto University, Kyoto 606-8501, Japanb Museum Zoologicum Bogoriense, Research Center for Biology, Indonesian Institute of Sciences, Gd. Widyasatwaloka, Jl. Raya Jakarta Bogor km 46, Cibinong West Java, Indonesiac Institute of Biological Sciences, Faculty of Science, University of Malaya, 50603 Kuala Lumpur, Malaysiad Institute for Environment and Development (LESTARI), Universiti Kebangsaan Malaysia, 43600 UKM Bangi, Selangor, Malaysiae Faculty of Science and Technology, Universiti Kebangsaan Malaysia, 43600 UKM Bangi, Selangor, Malaysiaf Department of Biology, Faculty of Science, Chulalongkorn University, Bangkok 10330, Thailandg Institute for Tropical Biology and Conservation, University Malaysia Sabah, Kota Kinabalu 88999, Sabah, Malaysiah Department of Science Education, Cheju National University, 690 756 Cheju, South Koreai Chengdu Institute of Biology, Chinese Academy of Sciences, Chengdu 610041, China
a r t i c l e i n f o
Article history:Received 17 February 2011Revised 24 May 2011Accepted 25 May 2011Available online 31 May 2011
Keywords:AsiaGastrophrynoidesmtDNAPapua-Australian elementPhrynellaSubgeneric classification
a b s t r a c t
We estimated the genealogical relationships and assessed systematic relationships among 45 out of 89named species and four unnamed taxa from 11 of 14 genera of the Oriental microhylids from 1767 bpsequences of the mitochondrial DNA genes 12S rRNA and 16S rRNA using maximum parsimony, maxi-mum likelihood, and Bayesian inference methods. Monophyly was rejected for the subfamily Microhyli-nae, and our data reveal four well-supported clades whose relationships to each other are unresolved: (A)Microhyla, Calluella, and Glyphoglossus, (B) Chaperina, (C) Kaloula, Phrynella, and Metaphrynella, and (D)Micryletta. They were genetically as divergent from each other as from another Oriental subfamily Kal-ophryninae, and could be recognized as distinct subfamilies. Within Clade A, our data reveal threewell-supported subclades whose relationships to each other are unresolved: (AI) Microhyla-I, (AII) Callu-ella and Glyphoglossus, and (AIII) Microhyla-II. Of the two enigmatic Malaysian genera, whose subfamilialplacement has been undetermined, Phrynella was found to be the sister species of Metaphrynella in CladeC, whereas Gastrophrynoides was grouped in the Papua-Australian subfamily Asterophryinae. Currentlyrecognized subgenera and species groups within Microhyla based on morphology were not supportedphylogenetically, and require thorough reassessments.
� 2011 Elsevier Inc. All rights reserved.
1. Introduction
The family Microhylidae exhibits circumtropic distribution andis diagnosed by many unique musculature and larval characteris-tics, including the presence of a posteromedially positioned spira-cle and absence of keratodonts and keratinized jaw sheath inlarvae (Frost et al., 2006). The family is now considered to contain11 subfamilies: Asterophryinae from Philippines, Indonesia, NewGuinea and Australia; Cophylinae from Madagascar; Dyscophinaefrom Madagascar; Gastrophryninae from North and South Amer-ica; Hoplophryninae from Africa; Kalophryninae from Southeast-ern and Southern Asia; Melanobatrachinae from Southern Asia;Microhylinae from Eastern, Southeastern and Southern Asia;Otophryninae from South America, Phrynomerinae from Africa;and Scaphiophryninae from Madagascar (Frost, 2010). Besides
species grouped in these subfamilies, there remain several specieswhose subfamilial placement is undetermined.
Microhylinae is distributed widely in the Oriental region, fromIndia and Korea to the Greater Sunda Islands, and it includes about70 species in nine genera (Calluella, Chaperina, Glyphoglossus, Kalo-ula, Metaphrynella, Microhyla, Micryletta, Ramanella, and Uperodon:Frost, 2010; Matsui, 2011). Another subfamily Kalophryninae con-tains about 16 species in one genus Kalophrynus that occur fromSouthern China to Java and Philippines, and northern India (Frost,2010; Dehling, 2011). Besides, one species of Phrynella from MalayPeninsula and two species of Gastrophrynoides from PeninsularMalaysia and Borneo, both in unknown subfamilies (Frost et al.,2006), about eight species of Oreophryne in Asterophryinae fromPhilippines, Celebes, and the Lesser Sunda Islands, and one speciesof Melanobatrachus in Melanobatrachinae from Southern India arefound within the Oriental region (Frost, 2010).
Separation of microhylids into distinct subfamilies on the basisof morphological characteristics has a long and complicatedhistory (e.g., Gadow, 1901; Parker, 1934; Laurent, 1986), but more
1055-7903/$ - see front matter � 2011 Elsevier Inc. All rights reserved.doi:10.1016/j.ympev.2011.05.015
⇑ Corresponding author. Fax: +81 75 753 6846.E-mail address: [email protected] (M. Matsui).
Molecular Phylogenetics and Evolution 61 (2011) 167–176
Contents lists available at ScienceDirect
Molecular Phylogenetics and Evolution
journal homepage: www.elsevier .com/ locate /ympev
Author's personal copy
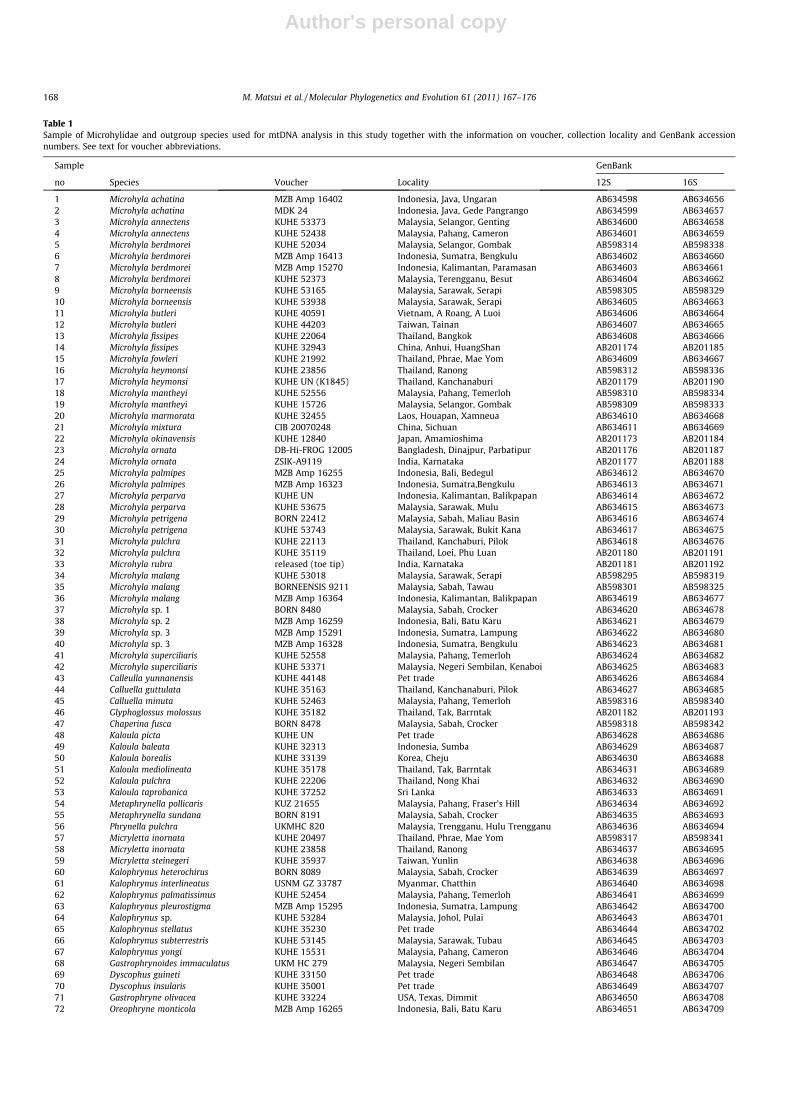
Table 1Sample of Microhylidae and outgroup species used for mtDNA analysis in this study together with the information on voucher, collection locality and GenBank accessionnumbers. See text for voucher abbreviations.
Sample GenBank
no Species Voucher Locality 12S 16S
1 Microhyla achatina MZB Amp 16402 Indonesia, Java, Ungaran AB634598 AB6346562 Microhyla achatina MDK 24 Indonesia, Java, Gede Pangrango AB634599 AB6346573 Microhyla annectens KUHE 53373 Malaysia, Selangor, Genting AB634600 AB6346584 Microhyla annectens KUHE 52438 Malaysia, Pahang, Cameron AB634601 AB6346595 Microhyla berdmorei KUHE 52034 Malaysia, Selangor, Gombak AB598314 AB5983386 Microhyla berdmorei MZB Amp 16413 Indonesia, Sumatra, Bengkulu AB634602 AB6346607 Microhyla berdmorei MZB Amp 15270 Indonesia, Kalimantan, Paramasan AB634603 AB6346618 Microhyla berdmorei KUHE 52373 Malaysia, Terengganu, Besut AB634604 AB6346629 Microhyla borneensis KUHE 53165 Malaysia, Sarawak, Serapi AB598305 AB59832910 Microhyla borneensis KUHE 53938 Malaysia, Sarawak, Serapi AB634605 AB63466311 Microhyla butleri KUHE 40591 Vietnam, A Roang, A Luoi AB634606 AB63466412 Microhyla butleri KUHE 44203 Taiwan, Tainan AB634607 AB63466513 Microhyla fissipes KUHE 22064 Thailand, Bangkok AB634608 AB63466614 Microhyla fissipes KUHE 32943 China, Anhui, HuangShan AB201174 AB20118515 Microhyla fowleri KUHE 21992 Thailand, Phrae, Mae Yom AB634609 AB63466716 Microhyla heymonsi KUHE 23856 Thailand, Ranong AB598312 AB59833617 Microhyla heymonsi KUHE UN (K1845) Thailand, Kanchanaburi AB201179 AB20119018 Microhyla mantheyi KUHE 52556 Malaysia, Pahang, Temerloh AB598310 AB59833419 Microhyla mantheyi KUHE 15726 Malaysia, Selangor, Gombak AB598309 AB59833320 Microhyla marmorata KUHE 32455 Laos, Houapan, Xamneua AB634610 AB63466821 Microhyla mixtura CIB 20070248 China, Sichuan AB634611 AB63466922 Microhyla okinavensis KUHE 12840 Japan, Amamioshima AB201173 AB20118423 Microhyla ornata DB-Hi-FROG 12005 Bangladesh, Dinajpur, Parbatipur AB201176 AB20118724 Microhyla ornata ZSIK-A9119 India, Karnataka AB201177 AB20118825 Microhyla palmipes MZB Amp 16255 Indonesia, Bali, Bedegul AB634612 AB63467026 Microhyla palmipes MZB Amp 16323 Indonesia, Sumatra,Bengkulu AB634613 AB63467127 Microhyla perparva KUHE UN Indonesia, Kalimantan, Balikpapan AB634614 AB63467228 Microhyla perparva KUHE 53675 Malaysia, Sarawak, Mulu AB634615 AB63467329 Microhyla petrigena BORN 22412 Malaysia, Sabah, Maliau Basin AB634616 AB63467430 Microhyla petrigena KUHE 53743 Malaysia, Sarawak, Bukit Kana AB634617 AB63467531 Microhyla pulchra KUHE 22113 Thailand, Kanchaburi, Pilok AB634618 AB63467632 Microhyla pulchra KUHE 35119 Thailand, Loei, Phu Luan AB201180 AB20119133 Microhyla rubra released (toe tip) India, Karnataka AB201181 AB20119234 Microhyla malang KUHE 53018 Malaysia, Sarawak, Serapi AB598295 AB59831935 Microhyla malang BORNEENSIS 9211 Malaysia, Sabah, Tawau AB598301 AB59832536 Microhyla malang MZB Amp 16364 Indonesia, Kalimantan, Balikpapan AB634619 AB63467737 Microhyla sp. 1 BORN 8480 Malaysia, Sabah, Crocker AB634620 AB63467838 Microhyla sp. 2 MZB Amp 16259 Indonesia, Bali, Batu Karu AB634621 AB63467939 Microhyla sp. 3 MZB Amp 15291 Indonesia, Sumatra, Lampung AB634622 AB63468040 Microhyla sp. 3 MZB Amp 16328 Indonesia, Sumatra, Bengkulu AB634623 AB63468141 Microhyla superciliaris KUHE 52558 Malaysia, Pahang, Temerloh AB634624 AB63468242 Microhyla superciliaris KUHE 53371 Malaysia, Negeri Sembilan, Kenaboi AB634625 AB63468343 Calleulla yunnanensis KUHE 44148 Pet trade AB634626 AB63468444 Calluella guttulata KUHE 35163 Thailand, Kanchanaburi, Pilok AB634627 AB63468545 Calluella minuta KUHE 52463 Malaysia, Pahang, Temerloh AB598316 AB59834046 Glyphoglossus molossus KUHE 35182 Thailand, Tak, Barrntak AB201182 AB20119347 Chaperina fusca BORN 8478 Malaysia, Sabah, Crocker AB598318 AB59834248 Kaloula picta KUHE UN Pet trade AB634628 AB63468649 Kaloula baleata KUHE 32313 Indonesia, Sumba AB634629 AB63468750 Kaloula borealis KUHE 33139 Korea, Cheju AB634630 AB63468851 Kaloula mediolineata KUHE 35178 Thailand, Tak, Barrntak AB634631 AB63468952 Kaloula pulchra KUHE 22206 Thailand, Nong Khai AB634632 AB63469053 Kaloula taprobanica KUHE 37252 Sri Lanka AB634633 AB63469154 Metaphrynella pollicaris KUZ 21655 Malaysia, Pahang, Fraser’s Hill AB634634 AB63469255 Metaphrynella sundana BORN 8191 Malaysia, Sabah, Crocker AB634635 AB63469356 Phrynella pulchra UKMHC 820 Malaysia, Trengganu, Hulu Trengganu AB634636 AB63469457 Micryletta inornata KUHE 20497 Thailand, Phrae, Mae Yom AB598317 AB59834158 Micryletta inornata KUHE 23858 Thailand, Ranong AB634637 AB63469559 Micryletta steinegeri KUHE 35937 Taiwan, Yunlin AB634638 AB63469660 Kalophrynus heterochirus BORN 8089 Malaysia, Sabah, Crocker AB634639 AB63469761 Kalophrynus interlineatus USNM GZ 33787 Myanmar, Chatthin AB634640 AB63469862 Kalophrynus palmatissimus KUHE 52454 Malaysia, Pahang, Temerloh AB634641 AB63469963 Kalophrynus pleurostigma MZB Amp 15295 Indonesia, Sumatra, Lampung AB634642 AB63470064 Kalophrynus sp. KUHE 53284 Malaysia, Johol, Pulai AB634643 AB63470165 Kalophrynus stellatus KUHE 35230 Pet trade AB634644 AB63470266 Kalophrynus subterrestris KUHE 53145 Malaysia, Sarawak, Tubau AB634645 AB63470367 Kalophrynus yongi KUHE 15531 Malaysia, Pahang, Cameron AB634646 AB63470468 Gastrophrynoides immaculatus UKM HC 279 Malaysia, Negeri Sembilan AB634647 AB63470569 Dyscophus guineti KUHE 33150 Pet trade AB634648 AB63470670 Dyscophus insularis KUHE 35001 Pet trade AB634649 AB63470771 Gastrophryne olivacea KUHE 33224 USA, Texas, Dimmit AB634650 AB63470872 Oreophryne monticola MZB Amp 16265 Indonesia, Bali, Batu Karu AB634651 AB634709
168 M. Matsui et al. / Molecular Phylogenetics and Evolution 61 (2011) 167–176

Author's personal copy
recent molecular phylogenetic studies have drastically changed thetraditional systematic arrangements within the family (VanBocxlaer et al., 2006; Frost et al., 2006; Van der Meijden et al.,2007). Results of these studies are not always in agreement, andthe basal resolution is usually insufficient. This latter point, how-ever, indicates very early diversification of clades recognized, andled some authors to consider elevating subfamilies to the familialrank (Bossuyt and Roelants, 2009).
Previous molecular phylogenetic studies of microhylid taxawere made from higher levels among anurans (see above) to thelower, infrageneric level (e.g., Hoskin, 2004). Among them, someauthors closely studied samples from a smaller part of the distribu-tion of the family (e.g., Africa: Loader et al., 2004; Papua-New Gui-nea: Köhler and Günther, 2008; Madagascar: Wollenberg et al.,2008). In contrast, the Oriental region suffers from inadequacy ofphylogenetic studies based on taxon-rich sampling of the familyuntil now.
In this paper, we present a molecular phylogeny of the OrientalMicrohylidae, sampling about half of the named species and sev-eral unnamed taxa from all but three of 14 known genera. Ourmain purpose is not to elucidate basal relationships of subfamilieswithin Microhylidae, which would be not easy from analyses ofonly short sequences of mtDNA. Instead, in order to evaluate theposition of the Oriental taxa, we compared our data with availableGenBank data of known microhylid subfamilies. Based on our find-ings, we also examined the current taxonomic grouping within thegenus Microhyla, the most species-rich genus in the OrientalMicrohylidae.
2. Materials and methods
2.1. Sampling design
We examined a total of 76 partial DNA sequences of the mito-chondrial DNA genes 12S and 16S rRNA from 50 named (45 fromAsia and five from other regions) and four unnamed taxa of Micro-hylidae and three outgroup species (Table 1). Undescribed orunidentified taxa included: Microhyla sp. 1, a Bornean species mor-phologically similar to M. borneensis; Microhyla sp. 2 from Bali,Indonesia, also similar to M. borneensis but lacks toe webs; Micro-hyla sp. 3 from Sumatra, morphologically similar to M. achatina;and Kalophrynus sp. from Peninsular Malaysia that has been con-fused with K. pleurostigma (Matsui, unpublished). We used Kassinasenegalensis, Hyperolius viridiflavus, and Rhacophorus schlegelii(GenBank AB 202078: Sano et al., 2005) as outgroup taxa. Kassinaand Hyperolius are hyperoliids in the Afrobatrachia, which is thesister clade of Microhylidae, and together forms Allodapanura,whereas Rhacophorus is a rhacophorid, which belongs to Natatan-ura, the sister group of Allodapanura (Frost et al., 2006).
Voucher specimens/tissues are stored in BORN (BORNEENSIScollection, Institute for Tropical Biology and Conservation, Univer-sity Malaysia Sabah), CIB (Chengdu Institute of Biology, ChineseAcademy of Sciences), DB-HI-FROG (Laboratory for AmphibianBiology, Hiroshima University), KUHE (Kyoto University, GraduateSchool of Human and Environmental Studies), KUZ (Department of
Zoology, Graduate School of Science, Kyoto University), MZB(Museum Zoologicum Bogoriense), MDK (Department of Conserva-tion and Ecotourism, Faculty of Forestry, Bogor Agricultural Insti-tute), UKMHC (Herpetological Collection, University KebangsaanMalaysia), USNM GZ (George Zug field number, United StatesNational Museum), and ZSIK (Zoological survey of India, Kolkata).
2.2. Preparation of DNA, PCR and DNA sequencing
Tissues were obtained from frozen or ethanol (95–99%) pre-served specimens and total genomic DNA extracted using standardphenol–chloroform extraction procedure (Hillis et al., 1996). Wehomogenized tissues in 0.6 ml STE buffer containing 10 mM Tris/HCl, pH 8.0, 100 mM NaCl and 1 mM EDTA, pH 8.0. We addedProteinase K (0.1 mg/ml) to the homogenate solutions and digestedproteins for 4–12 h at 55 �C. The solution was treated with phenoland chloroform/isoamyl alcohol, and DNA was precipitated withethanol. DNA precipitates were dried and then resuspended in0.6 ml TE (10 mM Tris/HCl, 1 mM EDTA, pH 8.0) and 1 ll was sub-jected to polymerase chain reaction (PCR). The PCR cycle includedan initial denaturation step of 5 min at 94 �C and 33cycles of dena-turation for 30 s at 94 �C, primer annealing for 30 s at 48–50 �C,and extension for 1 min 30 s at 72 �C. Primers used in PCR areshown in Table 2. The PCR products purified using polyethyleneglycol (PEG, 13%) precipitation procedures were used directly astemplates for Cycle Sequencing Reactions with fluorescent-dye-la-beled terminator (ABI Big Dye Terminators v.3.1 cycle sequencingkit). The sequencing reaction products were purified by ethanolprecipitation following the manufacturer’s protocol and then runon an ABI PRISM 3130 genetic analyzer. All samples were se-quenced in both directions using the same primers as for PCR.
2.3. Phylogenetic analysis
Aligned, concatenated sequences of 12S and 16S rRNA yielded atotal 1767 bp positions. We used Chromas Pro software (Technely-sium Pty Ltd., Tewntin, Australia) to edit the sequences, and wealigned them using the ClustalW option of Bioedit (Hall, 1998).The initial alignments were then checked by eye and adjustedslightly. Phylogenetic trees were constructed using maximumparsimony (MP), maximum likelihood (ML), and Bayesian infer-ence (BI). MP trees, obtained using PAUP�4.0b10 (Swofford,2002), involved a heuristic search with the tree bisection reconnec-tion (TBR) branch-swapping algorithm, and 100 random additionsreplicates. Transitions and transversions were equally weighted,and gaps were treated as missing data. ML analyses wereperformed by Treefinder version June 2007 (Jobb et al., 2004), withthe general time-reversible (GTR) model of DNA evolution with agamma shape parameter (G), identified as the best-fitting modelunder the Akaike information criterion implemented in Kakusan3 (Tanabe, 2007). BI and Bayesian posterior probabilities (BPP)were estimated using MrBayes 3.0b4 (Huelsenbeck and Ronquist,2001), under the GTR model with G and proportion of invariablesites (I), selected by MrModeltest2.2 (Nylander, 2004). BI used foursimultaneous Metropolis coupled Monte Carlo Markov chains for
Table 1 (continued)
Sample GenBank
no Species Voucher Locality 12S 16S
73 Phrynomantis bifasciatus KUHE 33277 Pet trade AB634652 AB63471074 Scaphiophryne gottlebei KUHE 34977 Pet trade AB634653 AB63471175 Hyperolius viridiflavus KUHE 43562 Zambia AB634654 AB63471276 Kassina senegalensis KUHE 12756 Pet trade AB634655 AB63471377 Rhacophorus schlegelii Japan, Hiroshima AB202078 AB202078
M. Matsui et al. / Molecular Phylogenetics and Evolution 61 (2011) 167–176 169

Author's personal copy
6,000,000 generations. We sampled a tree every 100 generationsand calculated a consensus topology for 30,001 trees after discard-ing the first 30,000 trees (burn-in = 3,000,000).
We used strength of nodal support in the MP analyses and usednon-parametric bootstrapping (MPBS; Felsenstein, 1985) with1000 pseudoreplicates in PAUP�, and 500 replicates for the ML tree(MLBS). We a priori regarded tree nodes with bootstrap value 70%or greater as sufficiently resolved (Huelsenbeck and Hillis, 1993),and those between 50% and 70% as tendencies. In the BI analysis,we considered nodes with a BPP of 95% or greater significant(Leaché and Reeder, 2002).
3. Results
3.1. Sequence and statistics
Sequence statistics for the two gene fragments and for the com-bined alignment including all nucleotide positions are provided inTable 3. The aligned 12S and 16S rRNA data set consisted of 1767characters, in which 948 sites were variable, and 758 potentiallyphylogenetically informative. MP analysis yielded two most parsi-monious trees of 6691 steps, a consistency index of 0.247, andretention index of 0.581. The ML analysis produced a topology withlnL �29636.124 (gamma shape parameter = 0.242; nucleotide fre-quencies: A = 0.317, C = 0.263, G = 0.191, and T = 0.229). BI calcu-lated average parameter estimates for nucleotide frequencies ofA = 0.409, C = 0.228, G = 0.125 and T = 0.237, a gamma shapeparameter 0.652, and proportion of invariable sites of 0.372.
3.2. Phylogenetic relationships and genetic divergence
All analyses produced essentially the same topologies. They dif-fered only in associations at poorly supported nodes. The ML tree(Fig. 1) infers the following sets of relationships:
(i) A clade representing all Microhylidae with respect to Kassi-na, Hyperolius, and Rhacophorus received very weak boot-strap support in all trees.
(ii) Monophyly of the Oriental microhylids was not supportedand they were divided into five monophyletic groups whoserelationships to each other were unresolved: Clade A (Micro-hyla, Calluella, and Glyphoglossus: MPBS = 94%, MLBS = 99%,BPP = 100%), Clade B (Chaperina), Clade C (Kaloula, Phrynella,and Metaphrynella; MPBS = 85%, MLBS = 98%, BPP = 100%),and Clade D (Micryletta; MPBS, MLBS, BPP all 100%) of micro-hylines, and Clade E (Kalophryninae; all supportvalues = 100%).
(iii) Besides these five monophyletic groups, Gastrophrynoidesand Oreophryne formed a clade, although not universallysupported (MPBS = 59%, MLBS = 93%, BPP = 100%). The twogenera differed from each other by a large uncorrected p-distance in 16S rRNA of 18.0%.
(iv) In Clade A, three subclades (I–III) were recovered. SubcladesAI and AII tended to form a monophyletic group, but the sup-port was insufficient (MPBS = 70%, MLBS = 75%, BPP = 85%),and relationships among the three subclades wereunresolved.
(v) Subclade AI, AII, and AIII, respectively, contained Microhyla I(Microhyla except for four species in AIII: MPBS = 96%,MLBS = 100%, BPP = 100%), Calluella and Glyphoglossus (all100%), and Microhyla II (M. petrigena, M. perparva, M. annec-tens, and M. marmorata: all 100%).
(vi) In Subclade AI, four monophyletic groups, (a) M. palmipes,(b) M. ornata and M. rubra (MPBS = 99%, MLBS = 99%,BPP = 100%), (c) M. butleri and M. superciliaris (MPBS = 92%,MLBS = 92%, BPP = 100%), and (d) the remaining Microhyla(MPBS = 79%, MLBS = 92%, BPP = 100%) showed unresolvedinterrelationships. In Group AIb, M. ornata from India andBangladesh showed a large divergence with a p-distance of6.7%, while in Group AIc, p-distances were as large as11.6–12.0% between M. butleri and M. superciliaris.
(vii) In Group AId, two monophyletic subgroups, (d1) M. berdmorei,M. fowleri, and M. pulchra (all 100%), and (d2) the remainingMicrohyla (MPBS = 79%, MLBS = 98%, BPP = 100%) showedsister-group relationships. In Subgroup AId1, uncorrected p-distances were not large (3.9–4.4%) between M. berdmoreiand M. fowleri, but were very large (9.7–10.8%) between M.pulchra and the other two species.
(viii) In Subgroup AId2, three monophyletic groups, (a) M. fissipes,M. okinavensis, and M. mixtura (MPBS = 99%, MLBS = 99%,BPP = 100%), (b) M. heymonsi, and (c) M. achatina, Microhylasp. 3, M. mantheyi, Microhyla sp. 2, M. borneensis, Microhylasp. 1, and M. malang (MPBS = 93%, MLBS = 99%, BPP =100%), showed unresolved relationships. In AId2a uncor-rected p-distance between M. okinavensis and M. mixturawas not small (6.4%). In AId2c, uncorrected p-distancesbetween M. achatina and Microhyla sp. 3 were 5.5–6.6%,and were 7.0–7.2% between M. mantheyi and Microhyla sp.
Table 3Alignment statistics for fragments of the 12S rRNA and 16S rRNA (all nucleotidepositions included); number of base bairs (bp), number of variable sites (vs), numberof parsimony informative sites (pi), the transition–transversion ratio given foringroups only (ti/tv).
bp vs pi ti/tv
12S rRNA 888 459 357 1.15316S rRNA 879 489 401 1.153Combined 1767 948 758 1.138
Table 2Primers used in this study.
Target Primer Sequence (50-30) Reference
12S rRNA 12SL 1 RYAGTGYTGAAAACGTAAGA Matsui et al. (2010a)12SA-L AAACTGGGATTAGATACCCCACTAT Palumbi et al. (1991)L1507 TACACACCGCCCGTCACCCTCTT Matsui et al. (2010b)12SF CATTGCTCGTAATTCCCTGGCG Goebel et al. (1999)H1548 TACCATGTTACGACTTTCCTCTTCT Matsui et al. (2005)
16S rRNA L1879 CGTACCTTTTGCATCATGGTC Matsui et al. (2010b)L2188 AAAGTGGGCCTAAAAGCAGCCA Matsui et al. (2006)16L-1 CTGACCGTGCAAAGGTAGCGTAATCACT Hedges (1994)H1923 AAGTAGCTCGCTTAGTTTCGG Matsui et al. (2010b)H2317 TTCTTGTTACTAGTTCTAGCAT Shimada et al. (2011)will6 CCCTCGTGATGCCGTTGATAC Wilkinson et al. (2002)16H1 CTCCGGTCTGAACTCAGATCACGTAGG Hedges (1994)
170 M. Matsui et al. / Molecular Phylogenetics and Evolution 61 (2011) 167–176

Author's personal copy
2, 4.3% between and M. borneensis and Microhyla sp. 1, and5.3–6.3% between M. borneensis plus Microhyla sp. 1 and M.malang.
(ix) In Subclade AII, Calluella yunnanensis, Glyphoglossus, and twoother Calluella species exhibited unresolved relationships toeach other. Uncorrected p-distances between C. yunnanensisand two other Calluella species (7.6–10.2%) were larger than
the distance observed between the latter two species (6.3%).Glyphoglossus was closer to C. yunnanensis (7.6%) than to thetwo other Calluella species (8.4–10.1%).
(x) In Subclade AIII, M. petrigena and M. perparva (MPBS = 98%,MLBS = 99%, BPP = 100%), and M. annectens and M. marmora-ta (MPBS = 87%, MLBS = 92%, BPP = 100%), respectively, weremonophyletic. Uncorrected p-distances were 6.8–8.2%
Fig. 1. Maximum likelihood phylogram of 1767 bp of 12S rRNA and 16S rRNA mitochondrial genes for samples of Oriental microhylids and related species. Voucher samplenumbers are included in Table 1. Numbers near branches represent bootstrap supports for MP/ML/Bayesian inferences. Asterisks indicate nodes with significant bootstrapsupports for ML and MP (>70%) and Bayesian posterior probabilities (BPP>95%). A–D and E indicate major clades recognized in Microhylinae and Kalophryninae, respectively(see text for detail).
M. Matsui et al. / Molecular Phylogenetics and Evolution 61 (2011) 167–176 171

Author's personal copy
between M. petrigena and M. perparva and 8.4–8.6% betweenM. annectens and M. marmorata. Intraspecific divergence wasalso large, with the uncorrected p-distance of 6.5% and 5.5%between paired samples of M. petrigena and M. perparva,respectively.
(xi) In Clade C, Phrynella proved to be the sister species to Meta-phrynella (MPBS = 92%, MLBS = 99%, BPP = 100%). Kaloulawas paraphyletic with respect to the group of Phrynellaand Metaphrynella. Relationships among K. taprobanica,other taxa of Kaloula, and the Phrynella and Metaphrynellaclade were unresolved. Kaloula taprobanica was distant fromthe congeners with very large uncorrected p-distances of11.0–12.9%. Uncorrected p-distance between the two spe-cies of Metaphrynella was 7.5%.
(xii) In Clade D, Micryletta inornata was paraphyletic with thesample from southern Thailand (M. i. lineata) being the sistertaxon to the group of the sample from northern Thailand (M.i. inornata) and M. steinegeri. Uncorrected p-distances of thesouthern Thai sample from the other two samples (4.9–5.1%)were much larger than the distance between the latter twosamples (2.9%).
(xiii) In Clade E (Kalophryninae), relationships among eight taxaof Kalophrynus were unresolved. Uncorrected p-distancesamong taxa examined varied from 4.9% to 10.5%.
(xiv) Two species of Dyscophus from Madagascar formed a mono-phyletic group (all 100%), but their sister taxon could not beidentified.
4. Discussion
4.1. Known phylogenetic relationships of Oriental microhylids
Van Bocxlaer et al. (2006) studied nuclear and mitochondrialgene phylogeny of Microhylidae and obtained subfamilial relation-ships. As to Oriental members, they found that: Kalophrynus (ourClade E) forms a clade with Plethodontohyla and Melanobatrachus,and is placed outside of the microhyline clade (our Clades A andC); Together with Uperodon and Ramanella, Kaloula and Metaphry-nella (our Clade C) form a clade, which is the sister taxon to theclade containing Calluella, Glyphoglossus, and Microhyla (our CladeA); Calluella guttulata and Glyphoglossus molossus (part of our Subc-lade AII) are sister species, and are the sister taxon to Microhyla(our Clade AI); and Malagasy Dyscophus is the sister species tothe Microhylinae (including Calluella). In contrast to Van Bocxlaeret al. (2006), Van der Meijden et al. (2007) could not resolve basalbranches in their study of nuclear gene phylogeny of world Micro-hylidae. The phylogenetic relationships among the subfamilies re-mained unknown, but as to Oriental microhylids, they similarlyfound sister relationships of the Calluella plus Glyphoglossus cladeand the Microhyla clade, and distant relationships of Kalophrynusfrom the microhyline clade. Van der Meijden et al. (2007) alsodemonstrated sister-group relationships of the Microhylinae andDyscophus as reported by Van Bocxlaer et al. (2006). Among Micro-hyla, they found that M. pulchra and M. heymonsi (our SubcladeAId) formed a monophyletic group, and were the sister taxon toM. butleri (our Subclade AIc). Micryletta (our Clade D) was placedas the sister taxon of the Microhyla, Calluella, and Glyphoglossusclade (our Clade A). Before this study, Frost et al. (2006) proposeda new anuran taxonomic system by combining morphological andnuclear and mitochondrial gene data, and placed Kaloula, Chaperi-na, Calluella, and Microhyla in their Microhylinae, but the subfam-ilial placement of Kalophrynus was undetermined. Van derMeijden et al. (2007) reanalyzed molecular data from Frost et al.(2006) and obtained a different result from the original one.According to the result of reanalysis, monophyly was supportedonly between Microhyla and Calluella, and Frost et al.’s (2006)
Microhylinae was rejected, although Chaperina and Kaloula tendedto be grouped with Microhyla and Calluella.
4.2. Proposed taxonomic relationships of Oriental microhylids
From these previous major studies, monophyly in each of (1)Kaloula and Metaphrynella, (2) Calluella and Glyphoglossus, (3) theCalluella and Glyphoglossus clade and the Microhyla clade, (4) theKaloula and Metaphrynella clade and the Calluella, Glyphoglossus,and Microhyla clade, and (5) Dyscophus and the Microhylinae,seems to be almost certain. In addition, (6) non-monophyly ofCalluella and Dyscophus and (7) distant relationships of Microhyli-nae and Kalophrynus are also universally accepted.
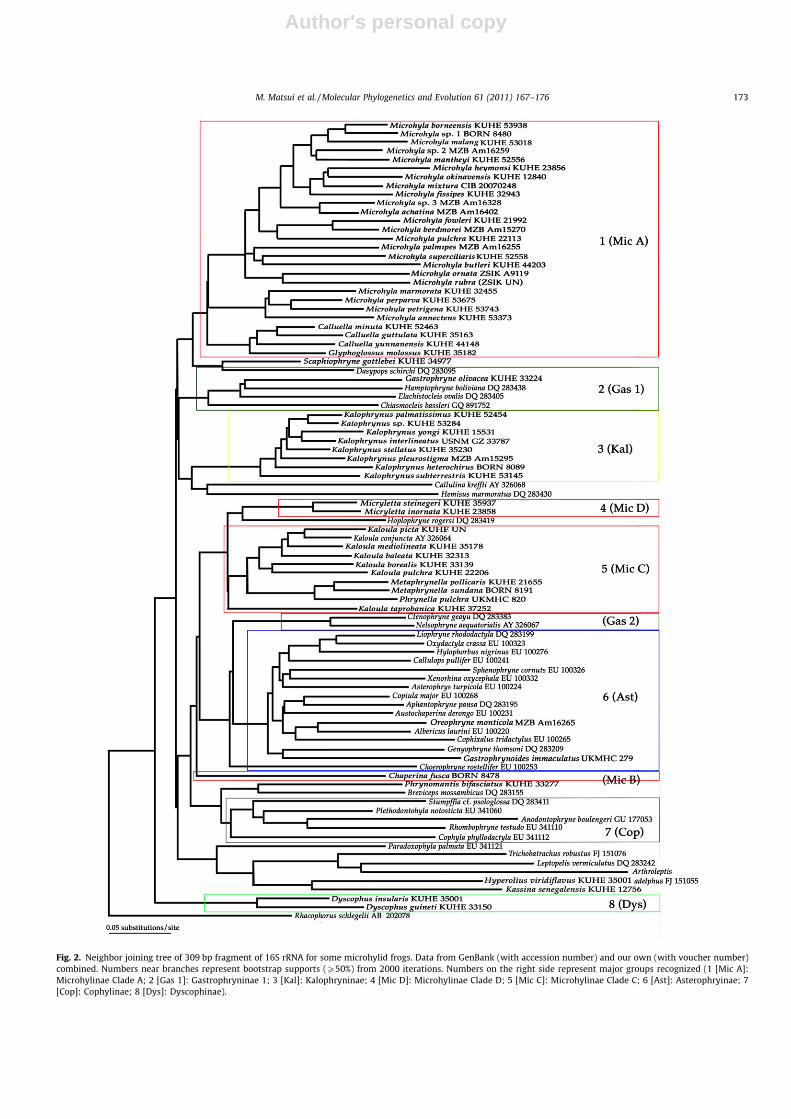
However, relationships among many other taxa in the family re-mained unknown from these previous studies. We therefore triedto evaluate relationships of Oriental microhylids by using GenBankdata. Unfortunately, published sequences for microhylid frogsgreatly differ from each other in lengths and aligned sites, as faras we have checked. Thus, the available common region is verylimited as exemplified in Fig. 2, which could be constructed usingonly 309 bp of 16S rRNA partial sequences that were comparableamong GenBank data (Darst and Cannatella, 2004; Frost et al.,2006; Blackburn, 2008; Wollenberg et al., 2008; Köhler andGünther, 2008; Funk and Cannatella, 2009; Vences et al., 2010).As shown in the figure, basal resolution was chaotic and supportsfor terminal branches were also very weak.
From the figure, the following main groups could be recognized,although monophyly in each of them is rarely strongly supported:(1) Microhyla Clade (our Clade A); (2) Gastrophryninae (Gastroph-ryne, Hamptophryne, Elachistocleis, and Chiasmocleis); (3) Kalophry-nus Clade (our Clade E); (4) Micryletta (our Clade D); (5) KaloulaClade (our Clade C); (6) Asterophryinae, including Gastrophryno-ides; (7) Cophylinae; and (8) Dyscophinae. The remaining microhy-lid taxa (Chaperina [our Clade B], part of Gastrophryninae[Ctenophryne, Nelsonophryne, and Dasypops], Phrynomerinae[Phrynomantis], Scaphiophryninae [Scaphiophryne and Paradoxo-phyla], and Hoplophryninae), as well as outgroup taxa of Brevici-pitidae, Hemisotiadae, Arthroleptidae, and Hyperoliidae wereplaced among the above main groups.
This grouping has many problems especially with regard to theposition of non-microhylid taxa, but at the same time indicatesinteresting relationships of the Oriental microhylids that strength-en our own results. Each of our four microhyline clades and Kal-ophryninae formed a compact group, and positioned distantlyfrom each other. Sister species relationships of Phrynella and Meta-phrynella in Group 5, as well as distant relationships of Calluella inGroup 1 and Dyscophinae (Group 8), were again ascertained. Mostimportantly, Gastrophrynoides was deeply nested within the groupof 14 asterophryine genera other than Choerophryne in Group 6.This strengthens our own result using a single asterophryine Ore-ophryne, and Gastrophrynoides should be considered a member ofAsterophryinae. This is interesting because no Papua-Australiananuran elements are by now known to have invaded into Borneoand Malay Peninsula (Inger, 1999).
Rejection of sister-group relationships of Calluella (in our Subc-lade AII) and Dyscophus conforms to previous authors, and justifiedFei et al.’s (2005) removal of Calluella from the subfamily Dyscophi-nae. However, their proposal to place Calluella in a distinct subfam-ily Calluellinae is not justified. If we admit such a classification,two other subclades in our Clade A (Subclades AI and AII of Micro-hyla) should be also separated in different subfamilies, which issurely over-splitting. In contrast, four microhyline clades recog-nized in our results are as distant from each other as are currentlyrecognized subfamilies of Microhylidae.
If we admit the major clades recognized in our phylogenetictrees (Fig. 1) as representing subfamilies, the following taxonomic
172 M. Matsui et al. / Molecular Phylogenetics and Evolution 61 (2011) 167–176
Author's personal copy
Fig. 2. Neighbor joining tree of 309 bp fragment of 16S rRNA for some microhylid frogs. Data from GenBank (with accession number) and our own (with voucher number)combined. Numbers near branches represent bootstrap supports (P50%) from 2000 iterations. Numbers on the right side represent major groups recognized (1 [Mic A]:Microhylinae Clade A; 2 [Gas 1]: Gastrophryninae 1; 3 [Kal]: Kalophryninae; 4 [Mic D]: Microhylinae Clade D; 5 [Mic C]: Microhylinae Clade C; 6 [Ast]: Asterophryinae; 7[Cop]: Cophylinae; 8 [Dys]: Dyscophinae).
M. Matsui et al. / Molecular Phylogenetics and Evolution 61 (2011) 167–176 173

Author's personal copy
changes could be proposed. The subfamily Microhylinae is re-stricted to Clade A (Microhyla, Calluella, and Glyphoglossus). Calluel-la and Glyphoglossus form a monophyletic group, which is close toMicrohyla, and their placement in the subfamily Microhylinae isjustified. Because Calluella is paraphyletic with respect to Glyphog-lossus, the most distinct taxon C. yunnanensis may be placed in adistinct genus separate from the other species, or Glyphoglossusshould be synonymized with Calluella. Each of Chaperina (CladeB) and Micryletta (Clade D) is much divergent from Microhyla andshould be removed from the subfamily Microhylinae to form dis-tinct monotypic subfamilies. Clade C (Metaphrynella and Kaloula)forms a group distinct from Microhylinae and should be placedin a distinct subfamily. Phrynella should be grouped in this clade,because its sister-species relationship to Metaphrynella is unambig-uous. Members of Kalophrynus are monophyletic and much diver-gent from other clades, justifying current recognition of a distinctsubfamily Kalophryninae. Gastrophrynoides should be placed inthe subfamily Asterophryinae as noted above.
4.3. Taxonomic considerations of several taxa
Our phylogenetic analysis remedied several taxonomic prob-lems at the species/subspecies level in some Oriental microhylids.In our results Micryletta inornata was paraphyletic with respect toM. steinegeri. The sample of M. inornata from northern Thailand(M. i. inornata) and M. steinegeri were genetically close (uncor-rected p-distance in 16S of 2.9%), while the sample of M. inornatafrom southern Thailand (M. i. lineata of Taylor, 1962) was moredivergent from them with uncorrected p-distances of 4.9–5.1%,which values exceed those reported between different frog species(Fouquet et al., 2007). These results support the separation of twosubspecies of M. inornata into two distinct species, but such a revi-sion includes problems. Distribution range of M. inornata is verywide from India, through Myanmar, Thailand, and southern Chinato Malay Peninsula and Sumatra. However, the type localities ofthe two subspecies, Sumatra in M. inornata and Nakhon Si Thamm-arat, southern Thailand in M. i. lineata, are close to each other. Thus,topotypic samples of the nominal subspecies from Sumatra are ex-pected to be genetically closer to our sample of M. i. lineata than tonorthern Thailand populations of M. i. inornata. If this were thecase, the name M. i. lineata would be synonymized with the nom-inal subspecies, but at the same time northern populations requirenew name(s). Future studies on many populations, especially fromSumatra, are badly needed to solve taxonomic problems in thisspecies. In relation to this, distinct specific status of M. steinegeriendemic to Taiwan requires reassessment, because the species isgenetically not very distinct from the northern population of M.inornata.
Kalophrynus stellatus from Basilan (Stejneger, 1908) and K. acu-tirostris from Samar (Inger, 1954), both in the Philippines, havebeen synonymized with K. pleurostigma originally described fromSumatra (Parker, 1934; Inger, 1954, 1966; Alcala, 1986; Ohlerand Grosjean, 2005). Identification of our single sample from anunknown locality of the Philippines is tentative, but the samplewas genetically divergent from a topotypic sample of K. pleurostig-ma (p-distance of 4.9%). Probably, wide-ranging K. pleurostigmamay include several cryptic species, and at least Philippine popula-tions are thought to be not conspecific with K. pleurostigma. An un-named species of Kalophrynus from southern Malay Peninsula hasbeen confused with K. pleurostigma (e.g., Berry, 1975), but is mor-phologically and acoustically quite distinct from it (Matsui, unpub-lished). Large genetic divergence between them (p-distance of9.4%) supports their heterospecific status.
Species of Kaloula (Clade C) were paraphyletic with respect tothe group of Phrynella and Metaphrynella, and K. taprobanica fromSouthern Asia was much divergent (p-distances of 11.0–12.9%)
from other Kaloula taxa from East and Southeast Asia. Recentmolecular studies in various lineages of frogs have revealed dis-tinct status of South Asian members from former congeners, andwarranted drastic taxonomic changes (e.g., Ansonia and Ghatoph-ryne in Bufonidae: Biju et al., 2009; Limnonectes in Dicroglossidaeand Lankanectes in Nyctibatrachidae: Dubois and Ohler, 2001; Phil-autus and Pseudophilautus in Rhacophoridae: Bossuyt and Dubois,2001). Large genetic divergence of K. taprobanica from the non-In-dian congeners suggests its distinct generic status.
Microhyla fowleri was originally described from Chiang Mai,Siam (=Thailand: Taylor, 1934), but was later synonymized withM. berdmorei by the author himself (Taylor, 1962). However, someauthors still treat it as a valid species, without close comparison ofthe two taxa (Dubois, 1987; Yang and Rao, 2008; Fei et al., 2009).Our sample of M. fowleri was obtained from Phrae, not distant fromthe type locality, but was not much divergent from M. berdmorei,with p-distance of 3.9–4.4%. This result favors synonymizing M.fowleri with M. berdmorei as already done by Taylor (1962).
Within Microhyla genetic divergences as measured by uncor-rected p-distances were large between sister species, ranging from10.8% between M. pulchra and M. fowleri to 6.4% between M. oki-navensis and M. mixtura. However, in some species, intraspecificdivergences were also large as seen between samples of M. ornatafrom India and Bangladesh (6.7%) and isolated samples of M. petri-gena (6.5%) and M. perparva (5.5%). These results indicate the pres-ence of cryptic species in these species. Divergences of two of threeunidentified or unnamed samples (Microhyla spp. 2 and 3) fromtheir sister taxa were also large, ranging from 7.0% to 7.2% betweenM. mantheyi and Microhyla sp. 2 to 5.5–6.6% between M. achatinaand Microhyla sp. 3, and indicated their distinct species status.The distance between M. borneensis and Microhyla sp. 1 (4.3%)was smaller than these values, and was equivalent to 3.9–4.4% be-tween M. fowleri and M. berdmorei (see above). Thus the taxonomicposition of Microhyla sp. 1 requires confirmation through genetic,as well as non-genetic analyses of more samples.
4.4. Taxonomic arrangement within Microhyla
Parker (1934) provided a key to species of Microhyla, then rec-ognized on the basis of morphological characteristics such as pala-tine bone, digital tips, and toe webs, and he recognized two majorgroups (I and II) that differ in snout length. Group I consisted ofthree groups (A–C), and most Asian members were componentsof A and B, while M. inornata and American species were groupedin C. Later, American species (Group II and IC, except for M. inornat-a) were split from the genus (Carvalho, 1954). Thus the genusMicrohyla was confined to Asia, and consisted of Parker’s (1934)Group IA–C. Within Group IA, Parker (1934) also recognized twogroups (1 and 2), but he never formally named groups shown inhis key. By adding species and character states not listed in Parker(1934), Dubois (1987) classified these groups and proposed a newtaxonomy. He removed Microhyla inornata (Group IC of Parker,1934) from the genus to establish a distinct genus Micryletta. Healso split the genus Microhyla into two subgenera, Microhyla sensustricto (Group IA of Parker, 1934) and Diplopelma (Group IB ofParker, 1934), and recognized two species groups in the subgenusMicrohyla (Group IA1 and IA2 of Parker, 1934). The M. berdmoreigroup (Parker’s IA1) contains M. annamensis, M. annectens, M. berd-morei, M. borneensis, M. butleri, M. fowleri, M. mixtura, M. palmipes,M. perparva, M. petrigena, and M. superciliaris. Also, M. mantheyi andM. malang (both split from M. borneensis: Das et al., 2007 andMatsui, 2011, respectively) can be placed in the M. berdmorei groupby implication. The M. achatina group (Parker’s IA2) comprisesM. achatina, M. chakrapanii, M. fusca, M. heymonsi, and M. zeylanica.Another subgenus Diplopelma contains M. okinavensis, M. ornata,M. picta, M. pulchra, and M. rubra. Also, M. fissipes (resurrected from
174 M. Matsui et al. / Molecular Phylogenetics and Evolution 61 (2011) 167–176

Author's personal copy
M. ornata: Matsui et al., 2005) should be grouped in Diplopelma byimplication. Placement of the following taxa, described afterDubois (1987) is undetermined: M. erythropoda, M. karunaratnei,M. maculifera, M. marmorata, M. nanapollexa, M. pulverata, and M.sholigari. By contrast, Fei et al. (2005) proposed a classificationfor the Chinese members different from that of Dubois (1987).They did not recognize subgenera and assigned all species toMicrohyla, which was split into the Microhyla butleri group andthe Microhyla ornata group on the basis of digital morphology.Microhyla fowleri, M. mixtura, M. butleri, and M. heymonsi were as-signed to their Microhyla butleri group, and M. pulchra and M. ornat-a (=M. fissipes) to their Microhyla ornata group.
However, results of our phylogenetic analysis do not supportDubois’ (1987) classification based solely on the morphologicalkey of Parker (1934). The sister–taxon relationship of M. rubraand M. ornata was always supported, but the two species did notgroup with M. okinavensis and M. pulchra to form Dubois’ (1987)distinct subgenus Diplopelma. Instead, M. pulchra was alwaysplaced as the sister species of M. berdmorei. Furthermore, M. berd-morei and M. achatina, representing two species groups of Dubois(1987), were grouped within the Microhyla main Subclade (AI) asa group AId. Likewise, our results are not consistent with Feiet al.’s (2005) classification.
From our results, each of the two subclades, AI and AIII, shouldbe treated as a subgenus or possibly a distinct genus, and the fourgroupings, AIa–AId in the Subclade AI, should be considered spe-cies groups. We suggest recognizing the subgenus Microhyla (Subc-lade AI, including M. achatina, the type species of the genus) and anunnamed subgenus (Subclade AIII, including M. annectens, M. per-parva, M. petrigena, and M. marmorata) in the genus Microhyla. Inthe newly defined subgenus Microhyla, four species groups, theM. palmipes group (M. palmipes: AIa), the M. ornata group (M. ornat-a and M. rubra: AIb), the M. butleri group (M. butleri and M. super-ciliaris: AIc), and the M. achatina group (M. berdmorei, M. pulchra, M.fissipes, M. heymonsi, M. okinavensis, M. mixtura, M. achatina, Micro-hyla sp. 3, M. mantheyi, Microhyla sp. 2, M. borneensis, Microhyla sp.1, and M. malang: AId) can be recognized from the phylogeneticviewpoint.
Our results suggest that taxonomy in the genus Microhyla hasbeen misled by a high frequency of morphological homoplasiesboth in adults and larvae. Validity of our taxonomic arrangementshould be tested by both phylogenetically examining additionaltaxa unavailable in this study, and finding non-genetic (e.g., mor-phological and acoustic) evidence to diagnose each grouprecognized.
Acknowledgments
MM is grateful to the following for their encouragements and/orpermission to conduct research and field companionship: L. Apin,K. Araya, Md. S. Azman, the late A.-A. Hamid, T. Hikida, H. Ota, T.Kusano, D. Labang, M.B. Lakim, M. Maryati, the late J. Nabitabhata,T. Shimada, T. Sugahara, T. Tachi, M. Toda, and A. Tominaga. MM isalso indebted to M. Hori, H. Ito, Y. Kadota, M. Kageyama, M. Kato,S.K. Saidapur, A. de Silva, J.-Y. Song, M. Sumida, T. Sota, and G.R.Zug for tissue samples, and N. Kuraishi for laboratory assistance.The Economic-Planning Unit (former Socio-Economic ResearchUnit) of Malaysia, the State Government of Sarawak, Sabah Parks,The National Research Council of Thailand, and the Royal ForestDepartment of Thailand kindly permitted MM to conduct the pro-ject, and University Malaya, Universiti Malaysia Sabah, UniversitiKebangsaan Malaysia (UKM), JICA, the Forest Department, Sara-wak, and Chulalongkorn University kindly provided all the facili-ties for conducting research. Field trips by MM were madepossible by grants from The Monbusho International Scientific Re-search Program (Field Research, 01041051, 02041051, 04041068,
06041066, 08041144), The Monbukagakusho through the JapaneseSociety for the Promotion of Sciences (JSPS: Field Research,15370038, 20405013), UKM (OUP-PLW-14-59/2008), and TJTTP-OECF. AH thanks M.D. Kusrini, S. Kirono, and D. Susanto for provid-ing tissue samples and to the Monbukagakusho for scholarshipfunding. Research export permits were obtained through the ForestDepartment, Sarawak and the Chinese Academy of Sciences. Wethank two anonymous reviewers and A. Larson for improving themanuscript.
References
Alcala, A.C., 1986. Guide to Philippine Flora and Fauna. Amphibians and Reptiles,vol. X. Natural Resource Management Center, Ministry of Natural Resources andthe University of the Philippines, Manila.
Berry, P.Y., 1975. The Amphibian Fauna of Peninsular Malaysia. Tropical Press, KualaLumpur.
Biju, S.D., Van Bocxlaer, I., Giri, V.B., Loader, S.P., Bossuyt, F., 2009. Two new endemicgenera and a new species of toad (Anura: Bufonidae) from the Western Ghats ofIndia. BMC Res. Notes 2 (241), 1–10.
Blackburn, D.C., 2008. Biogeography and evolution of body size and life history ofAfrican frogs: phylogeny of squeakers (Arthroleptis) and long-fingered frogs(Cardioglossa) estimated from mitochondrial data. Mol. Phylogenet. Evol. 49,806–826.
Bossuyt, F., Dubois, A., 2001. A review of the frog genus Philautus Gistel, 1848(Amphibia, Anura, Ranidae, Rhacophorinae). Zeylanica 6, 1–112.
Bossuyt, F., Roelants, K., 2009. Anura. In: Hedges, S.B., Kumar, S. (Eds.), Timetree ofLife. Oxford University Press, New York, pp. 357–364.
Carvalho, A.L., 1954. A preliminary synopsis of the genera of American microhylidfrogs. Occas. Pap. Mus. Zool. Univ. Michigan 555, 1–19.
Darst, C.R., Cannatella, D.C., 2004. Novel relationships among hyloid frogs inferredfrom 12S and 16S mitochondrial DNA sequences. Mol. Phylogenet. Evol. 31,462–475.
Das, I., Yaakob, N., Sukumaran, J., 2007. A new species of Microhyla (Anura:Microhylidae) from the Malay Peninsula. Hamadryad 32, 304–314.
Dehling, J.M., 2011. A new karst-dwelling species of Kalophrynus (Anura:Microhylidae) from Gunung Mulu National Park, Borneo, Malaysia. Zootaxa2737, 49–60.
Dubois, A., 1987. Miscellanea taxinomica batrachologica (II). Alytes 6, 1–9.Dubois, A., Ohler, A., 2001. A new genus for an aquatic ranid (Amphibia, Anura) from
Sri Lanka. Alytes 19, 81–106.Fei, L., Hu, S.-Q., Ye, C.-Y., Huang, Y.-Z. (Eds.), 2009. Fauna Sinica. Amphibia, vol 2.
Anura. Science Press, Beijing.Fei, L., Ye, C.-Y., Jiang, J., 2005a. On Kalophrynus pleurostigma interlineatus and
taxonomic discussion on the genus Calluella with description of a newsubfamily. In: Fei, L., Ye, C.-Y., Jiang, J., Xie, F., Huang, Y. (Eds.), An IllustratedKey to Chinese Amphibians. Sichuan Publishing House of Science andTechnology, Chengdu, pp. 271–278.
Fei, L., Ye, C.-Y., Jiang, J., Xie, F., Huang, Y. (Eds.), 2005b. An Illustrated Key to ChineseAmphibians. Sichuan Publishing House of Science and Technology, Chengdu.
Felsenstein, J., 1985. Confidence limits on phylogenies: an approach using thebootstrap. Evolution 39, 783–791.
Fouquet, A., Gilles, A., Vences, M., Marty, C., Blanc, M., Gemmell, N.J., 2007.Underestimation of species richness in Neotropical frogs revealed by mtDNAanalyses. PLoS ONE 2, e1109.
Frost, D.R., 2010. Amphibian Species of the World: an Online Reference, version 5.4(8 April, 2010). American Museum of Natural History, New York, USA. <http://research.amnh.org/vz/herpetology/amphibia/> .
Frost, D.R., Grant, T., Faivovich, J.N., Bain, R.H., Haas, A., Haddad, C.F.B., de Sá, R.O.,Channing, A., Wilkinson, M., Donnellan, S.C., Raxworthy, C.J., Campbell, J.A.,Blotto, B.L., Moler, P., Drewes, R.C., Nussbaum, R.A., Lynch, J.D., Green, D.M.,Wheeler, W.C., 2006. The amphibian tree of life. Bull. Am. Mus. Nat. Hist. 297, 1–370.
Funk, W.C., Cannatella, D.C., 2009. A new, large species of Chiasmocleis Mehely 1904(Anura: Microhylidae) from the Iquitos region, Amazonian Peru. Zootaxa 2247,37–50.
Gadow, H., 1901. Amphibia and Reptiles. Macmillan, London.Goebel, A.M., Donnelly, J.M., Atz, M.E., 1999. PCR primers and amplification
methods for 12S ribosomal DNA, the control region, cytochrome oxidase I,and cytochrome b in bufonids and other frogs, and an overview of PCR primerswhich have amplified DNA in amphibians successfully. Mol. Phylogenet. Evol.11, 163–199.
Hall, T.A., 1998. BioEdit: a user-friendly biological sequence alignment editor andanalysis program for Windows 95/98/NT. Nucl. Acids Symp. Ser. 41, 95–98.
Hedges, S.B., 1994. Molecular evidence for the origin of birds. Proc. Nat. Acad. Sci.USA 91, 2621–2624.
Hillis, D.M., Mable, B.K., Larson, A., Davis, S.K., Zimmer, E.A., 1996. Nucleic acids IV:sequencing and cloning. In: Hillis, D.M., Mable, B.K., Moritz, C. (Eds.), MolecularSystematics, second ed. Sinauer, Sunderland, pp. 321–406.
Hoskin, C.J., 2004. Australian microhylid frogs (Cophixalus and Austrochaperina):Phylogeny, taxonomy, calls, distributions and breeding biology. Aust. J. Zool. 52,237–269.
M. Matsui et al. / Molecular Phylogenetics and Evolution 61 (2011) 167–176 175

Author's personal copy
Huelsenbeck, J.P., Hillis, D.M., 1993. Success of phylogenetic methods in the four-taxon case. Syst. Biol. 42, 247–264.
Huelsenbeck, J.P., Ronquist, F.R., 2001. MrBayes: Bayesian inference of phylogenetictrees. Bioinformatics 17, 754–755.
Inger, R.F., 1954. Systematics and zoogeography of Philippine Amphibia. Fieldiana:Zool. 33, 183–531.
Inger, R.F., 1966. The systematics and zoogeography of the Amphibia of Borneo.Fieldiana: Zool. 52, 1–402.
Inger, R.F., 1999. Distribution of amphibians in Southern Asia and adjacent islands.In: Duellman, W.E. (Ed.), Patterns of Distribution of Amphibians: A GlobalPerspective. Johns Hopkins Univ. Press, Baltimore and London, pp. 445–482.
Jobb, G., von Haeseler, A., Strimmer, K., 2004. Treefinder: a powerful graphicalanalysis environment for molecular phylogenetics. BMC Evol. Biol. 4, 18.
Köhler, F., Günther, R., 2008. The radiation of microhylid frogs (Amphibia: Anura) onNew Guinea: a mitochondrial phylogeny reveals parallel evolution ofmorphological and life history traits and disproves the current morphology-based classification. Mol. Phylogenet. Evol. 47, 363–365.
Laurent, R.F., 1986. Sous-classe des Lissamphibiens (Lissamphibia). Systématique.In: Grassé, P.P., Delsol, M. (Eds.), Traité de zoologie, Tome XIV, Batraciens, Fasc.I-B. Masson, Paris, pp. 594–797.
Leaché, A.D., Reeder, T.W., 2002. Molecular systematics of the eastern fence lizard(Sceloporus undulatus): a comparison of parsimony, likelihood, and Bayesianapproaches. Syst. Biol. 51, 44–68.
Loader, S.P., Gower, D.J., Howell, K.M.N.D., Rödel, M.-O., Clarke, B.T., De Sá, R., Cohen,B.L., Wilkinson, M., 2004. Phylogenetic relationships of African microhylid frogsinferred from DNA sequences of mitochondrial 12S and 16S rRNA genes. Org.Divers. Evol. 4, 227–235.
Matsui, M., 2011. Taxonomic revision of one of the Old World’s smallest frogs, withdescription of a new Bornean Microhyla (Amphibia, Microhylidae). Zootaxa2814, 33–49.
Matsui, M., Ito, H., Shimada, T., Ota, H., Saidapur, S.K., Khonsue, W., Tanaka-Ueno, T.,Wu, G.-F., 2005. Taxonomic relationships within the Pan-Oriental narrow-mouth toad Microhyla ornata as revealed by mtDNA analysis (Amphibia, Anura,Microhylidae). Zool. Sci. 22, 489–495.
Matsui, M., Shimada, T., Liu, W.Z., Maryati, M., Khonsue, W., Orlov, N., 2006.Phylogenetic relationships of Oriental torrent frogs in the genus Amolops and itsallies (Amphibia, Anura, Ranidae). Mol. Phylogenet. Evol. 38, 659–666.
Matsui, M., Kuraishi, N., Jiang, J.-P., Ota, H., Hamidy, A., Orlov, N.L., Nishikawa, K.,2010a. Systematic reassessments of fanged frogs from China and adjacentregions (Anura: Dicroglossidae). Zootaxa 2345, 33–42.
Matsui, M., Hamidy, A., Murphy, R.W., Khonsue, W., Yambun, P., Shimada, T.,Norhayati, A., Daicus, B.M., Jiang, J.-P., 2010b. Phylogenetic relationships ofmegophryid frogs of the genus Leptobrachium (Amphibia, Anura) as revealed bymtDNA gene sequences. Mol. Phylogenet. Evol. 56, 259–272.
Nylander, J.A.A., 2004. MrModeltest v2. Program Distributed by the Author.Evolutionary Biology Centre, Uppsala University.
Ohler, A., Grosjean, S., 2005. Color pattern and call variation in Kalophrynus fromSouth-east Asia (Anura: Microhylidae). Herpetozoa 18, 99–106.
Palumbi, S.R., Martin, A., Romano, S., McMillan, W.O., Stice, L., Grabowski, G., 1991.The simple fool’s guide to PCR, version 2.0. Privately published, compiled byPalumbi S., Department of Zoology, University of Hawaii, Honolulu.
Parker, B.A., 1934. A Monograph of the Frogs of the Family Microhylidae. The BritishMuseum, London.
Sano, N., Kurabayashi, A., Fujii, T., Yonekawa, H., Sumida, M., 2005. Completenucleotide sequence of the mitochondrial genome of Schlegel’s tree frogRhacophorus schlegelii (family Rhacophoridae): duplicated control regions andgene rearrangements. Genes Genet. Syst. 80, 213–224.
Shimada, T., Matsui, M., Yambun, P., Sudin, A., 2011. A taxonomic study ofWhitehead’s torrent frog, Meristogenys whiteheadi, with descriptions of two newspecies (Amphibia: Ranidae). Zool. J. Linn. Soc. 161, 157–183.
Stejneger, L., 1908. Two new species of toads from the Philippines. Proc. US Nat.Mus. 33, 573–576.
Swofford, D.L., 2002. PAUP�. Phylogenetic Analysis Using Parsimony (�and OtherMethods), Version 4. Sinauer Associates, Sunderland, MA.
Tanabe, A.S., 2007. Kakusan: a computer program to automate the selection of anucleotide substitution model and the configuration of a mixed model onmultilocus data. Mol. Ecol. Notes 7, 962–964.
Taylor, E.H., 1934. Zoological results of the third De Schauensee Siamese Expedition,Part III. Amphibians and reptiles. Proc. Acad. Nat. Sci. Philadelphia 86, 281–310.
Taylor, E.H., 1962. The amphibian fauna of Thailand. Univ. Kansas Sci. Bull. 63, 265–599.
Van Bocxlaer, I., Roelants, K., Biju, S.D., Nagaraju, J., Bossuyt, F., 2006. LateCretaceous vicariance in Gondwanan amphibians. PLoS ONE 1, e74.
Van der Meijden, A., Vences, M., Hoegg, S., Boistel, R., Channing, A., Meyer, A., 2007.Nuclear gene phylogeny of narrow-mouthed toads (Family: Microhylidae) and adiscussion of competing hypotheses concerning their biogeographical origins.Mol. Phylogenet. Evol. 44, 1017–1030.
Vences, M., Glaw, F., Koehler, J., Wollenberg, K.C., 2010. Molecular phylogeny,morphology and bioacoustics reveal five additional species of arborealmicrohylid frogs of the genus Anodonthyla from Madagascar. Contrib. Zool.79, 1–32.
Wilkinson, J.A., Drewes, R.C., Tatum, O.L., 2002. A molecular phylogenetic analysis ofthe family Rhacophoridae with an emphasis on the Asian and African genera.Mol. Phylogenet. Evol. 24, 265–273.
Wollenberg, K.C., Vieites, D.R., Van der Meijden, A., Glaw, F., Cannatella, D.C., Vences,M., 2008. Patterns of endemism and species richness in Malagasy cophylinefrogs support a key role of mountainous areas for speciation. Evolution 62,1890–1907.
Yang, D.-T., Rao, D. (Eds.), 2008. Amphibia and Reptilia of Yunnan. Yunnan Scienceand Technology Press, Kunming.
176 M. Matsui et al. / Molecular Phylogenetics and Evolution 61 (2011) 167–176
Copyright © 2022 FDOKUMEN




















