
Synthesis and conformational studies of peptides from new C-linked carbo-β-amino acids (β-Caas)...
11
PLASMID 12, 41-51 (1984) Interaction between Mitochondrial Genes in Yeast: Evidence for Novel box Effect(s) L ,AMBERT A. M. HENSGENS, GERDA VAN DER HORST, AND LESLIE A. GRIVELL Section for Molecular Biology, Laboratory of Biochemistry, University of Amsterdam, Kruislaan 318, 1098 SA4Amsterdam, The Netherlands Received January 25, 1984; revised April 10, 1984 The effectsof mutations have been studied within the apocytochrome b gene on the processing of transcripts from the gene for subunit 1of cytochrome c oxidase (~0x1).Most mutations which affect the expression of the reading frame encoded by the fourth intron of the apocytochrome b gene (b14)result in a failure to remove intron a14from precursor transcripts of cox1. Mutations in other apocytochrome b introns result in additional and complex defects in the processing of subunit I transcripts. Mutants Ml233 and Ml282 are mutated within the second intron (b12) of the apocytochrome b gene and have OX13 transcripts of 4900, 6100, and 6500 nucleotides. Thesetranscripts are absent from the wild-type strain and do not hybridize with all exon sequences of this gene. In mutant M 1392 (mutated within the third intron of the apocytochrome b gene), two OX13transcripts of 2200 and 2800 nucleotidesare presentwhich hybridize only with sequences downstream of the fifth exon of this gene (A%). We propose that all these transcripts result from distinctive cut-no-splice events, occurring at different intron-exon borders of OX13 pre- RNAs depending on the mutational site within the apocytochrome b gene. The box9 mutant M4458 and the box7 mutant M 143 1 lack detectable 18s mRNA for subunit 1 of cytochrome c oxidase. The box9 mutants M4751 and M4701 contain reduced amounts of this mRNA. The fact that these loci complement each other (B. Weiss-Brummer, G. Riidel, R. J. Schweyen, and F. Kaudewitz (1982) Cell 29, 527-536), therefore, suggests that mutations within the different functional domains of b14 lead to different defects in the processing of OX13 transcripts. This, together with the defects observed in bI2 and b13 mutants, implies that the box effect (i.e., the interaction between these two split genes) is not mediated by the box7 element alone. The possibility is discussedthat mutated apocytochrome b intronic reading frame products lead to these aberrant events in the processing of transcripts of the gene for subunit I of cytochrome c oxidase. The presenceof unassigned reading frames (URFs)’ within the introns which split some structural genes is one of the intriguing aspects of fungal mitochondrial genomes (see for re- views Dujon, 1981, and Grivell, 1983). Ex- tensive genetic studies backed up by DNA sequence analysis have provided evidence that the URFs presentwithin the secondand fourth introns (b12 and b14) of the apocytochrome b gene in yeast encode proteins required for the splicing of these introns from precursors of the cytochrome b messenger RNA (mRNA) (Lazowska et al., 1980;De la Salle et al., 1982; ’ Abbreviations used: bp, Base. pair(s); DBM-cellulose, diazobenzyloxymethyl-cell~ose; mRNA, messenger RNA; URF, unassigned reading frame. Weiss-Brummer et al., 1982). Other intronic URFs probably encode similar proteins (Hensgens et al., 1983b).The b14intron prod- uct is also involved in the splicing of transcripts from the gene for subunit I of cytochrome c ox&se. This link in the expression of the two genes has been termed the ‘box’ effect. Evi- dence for its existence is based on the obser- vation that mutations which disturb the expressionof the b14URF a&ct the expression of both the apocytochrome b gene and the gene for subunit I of cytochrome c oxidase (Pajot et al., 1976; Mahler et al., 1978, 1982; Dhawale et al., 1981; Haid et al., 1979; La- bouesse and Slonimski, 1983; Dujardin et al., 1982).The most likely role of the b14maturase is an involvement in the splicing of intron a14. 41 0147-619X/84 $3.00 Copyright Q 1984 by Academic Pm& Inc. All rights of reproduction in any form reserved.
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Synthesis and conformational studies of peptides from new C-linked carbo-β-amino acids (β-Caas)...

PLASMID 12, 41-51 (1984)
Interaction between Mitochondrial Genes in Yeast: Evidence for Novel box Effect(s)
L ,AMBERT A. M. HENSGENS, GERDA VAN DER HORST, AND LESLIE A. GRIVELL
Section for Molecular Biology, Laboratory of Biochemistry, University of Amsterdam, Kruislaan 318, 1098 SA4 Amsterdam, The Netherlands
Received January 25, 1984; revised April 10, 1984
The effects of mutations have been studied within the apocytochrome b gene on the processing of transcripts from the gene for subunit 1 of cytochrome c oxidase (~0x1). Most mutations which affect the expression of the reading frame encoded by the fourth intron of the apocytochrome b gene (b14) result in a failure to remove intron a14 from precursor transcripts of cox1. Mutations in other apocytochrome b introns result in additional and complex defects in the processing of subunit I transcripts. Mutants Ml233 and Ml282 are mutated within the second intron (b12) of the apocytochrome b gene and have OX13 transcripts of 4900, 6100, and 6500 nucleotides. These transcripts are absent from the wild-type strain and do not hybridize with all exon sequences of this gene. In mutant M 1392 (mutated within the third intron of the apocytochrome b gene), two OX13 transcripts of 2200 and 2800 nucleotides are present which hybridize only with sequences downstream of the fifth exon of this gene (A%). We propose that all these transcripts result from distinctive cut-no-splice events, occurring at different intron-exon borders of OX13 pre- RNAs depending on the mutational site within the apocytochrome b gene. The box9 mutant M4458 and the box7 mutant M 143 1 lack detectable 18s mRNA for subunit 1 of cytochrome c oxidase. The box9 mutants M4751 and M4701 contain reduced amounts of this mRNA. The fact that these loci complement each other (B. Weiss-Brummer, G. Riidel, R. J. Schweyen, and F. Kaudewitz (1982) Cell 29, 527-536), therefore, suggests that mutations within the different functional domains of b14 lead to different defects in the processing of OX13 transcripts. This, together with the defects observed in bI2 and b13 mutants, implies that the box effect (i.e., the interaction between these two split genes) is not mediated by the box7 element alone. The possibility is discussed that mutated apocytochrome b intronic reading frame products lead to these aberrant events in the processing of transcripts of the gene for subunit I of cytochrome c oxidase.
The presence of unassigned reading frames (URFs)’ within the introns which split some structural genes is one of the intriguing aspects of fungal mitochondrial genomes (see for re- views Dujon, 1981, and Grivell, 1983). Ex- tensive genetic studies backed up by DNA sequence analysis have provided evidence that the URFs present within the second and fourth introns (b12 and b14) of the apocytochrome b gene in yeast encode proteins required for the splicing of these introns from precursors of the cytochrome b messenger RNA (mRNA) (Lazowska et al., 1980; De la Salle et al., 1982;
’ Abbreviations used: bp, Base. pair(s); DBM-cellulose, diazobenzyloxymethyl-cell~ose; mRNA, messenger RNA; URF, unassigned reading frame.
Weiss-Brummer et al., 1982). Other intronic URFs probably encode similar proteins (Hensgens et al., 1983b). The b14 intron prod- uct is also involved in the splicing of transcripts from the gene for subunit I of cytochrome c ox&se. This link in the expression of the two genes has been termed the ‘box’ effect. Evi- dence for its existence is based on the obser- vation that mutations which disturb the expression of the b14 URF a&ct the expression of both the apocytochrome b gene and the gene for subunit I of cytochrome c oxidase (Pajot et al., 1976; Mahler et al., 1978, 1982; Dhawale et al., 1981; Haid et al., 1979; La- bouesse and Slonimski, 1983; Dujardin et al., 1982). The most likely role of the b14 maturase is an involvement in the splicing of intron a14.
41 0147-619X/84 $3.00 Copyright Q 1984 by Academic Pm& Inc. All rights of reproduction in any form reserved.

42 HENSGENS, VAN DER HORST, AND GRIVELL
This intron displays high sequence homology with b14 (Bonitz et al., 1980) including the sequences which appear to act as processing signals (De la Salle et al., 1982; Netter et al., 1982; Weiss-Brummer et al., 1983; Davies et al., 1982; Michel and Dujon, 1983). In support of this it has been claimed that in a yeast strain in which some of the introns (but not a14 and b14) are missing, mutations in the b14 intron lead to arrested processing of the a14 intron (Bonitz et al., 1980). Most of these observa- tions are indirect, however, and a study of transcripts in these mutants is not available. We have, therefore, systematically analysed the effects of mutations in different introns of the apocytochrome b gene on the processing of OX13 transcripts. We show that the pro- cessing of the a14 intron is indeed disturbed in most cytochrome b intron mutants studied. However, some mutations have additional ef- fects on the processing of precursor transcripts of the subunit I gene. These defects vary de- pending on the apocytochrome b gene intron in which the mutation is located. We suggest that they are the consequence of the structural similarities that exist between the various in- trons and between the proteins required for their processing.
MATERIALS AND METHODS
Yeast strains. The yeast strains used are listed in Table 1. Diploids of the parental wild- type strain Saccharomyces cerevisiae 777-3A and the mutants (gifts from Dr. R. J. Schweyen) were constructed by crossing with a mutant of S. cerevisiae KL14-4A iacking mtDNA.
Isolation of nucleic acids. Diploid cells of S. cerevisiae 777-3A and the mutants were grown in 2% (w/v) galactose, 1% (w/v) yeast extract, and 1% (w/v) Bacto peptone. Yeast cells were harvested in mid-to-late exponential phase. mtRNA was isolated and treated with pronase and deoxyribonuclease as described by Van Ommen et al. (1979).
Electrophoresis and hybridization. Electro- phoresis, staining, photography, size calibra- tion of transcripts, transfer to diazobenzyl- oxymethyl (DBM)-cellulose paper, and hy-
bridization were performed according to Van Ommen et al. ( 1979) and Hensgens et al. (1983a). Transcript lengths are given in nu- cleotides. Additionally, major transcripts identified previously are also identified by an approximate SE value.
In vitro labeling of nucleic acids. DNA was labeled by nick translation as described by Jefheys and Flavell (1977).
Probes used in hybridizations. Plasmid preparations were used (Hensgens et al., 1983a) and in some cases DNA fragments from these isolated as described by Van Ommen et al. (1980). D* is an 465bp Sau- 3A fragment isolated from pKLD. 11 l.E is an isolated 350-bp Sau3A fragment from pKLll1. 109* is an isolated 600-bp Sau3A fragment from pKL109. For intron aI5j3 an M 13 clone 24 11, containing intron sequences only, was used after labeling by primed, sec- ond-strand synthesis in vitro (Hensgens et al., 1983b).
RESULTS
Information on the localization and nature of the studied apocytochrome b mutants is given in Fig. 1 and Table 1. To study the transcripts of the apocytochrome b gene (en- coded by the COB region) and the gene for the subunit I of cytochrome c oxidase (encoded by the OX13 region), mtRNA from the pa- rental wild-type S. cerevisiae 777-3A and from the mutant strains was isolated, fractionated by electrophoresis, blotted onto DBM paper and hybridized with different 32P-labeled probes (see Materials and Methods and Hens- gens et al. ( 1983a) for further details). As probe for COB transcripts, mtDNA of the petite mutant Al 5-1 was used. This petite retains all exon and intron sequences of the gene (Van Ommen et al., 1980). For the detection of OX13 transcripts the mtDNA of the petite strain LH30-A5 was used. This probe also de- tects the 22s and 24s transcripts of the genes for the subunits 6 and 8 of the ATPase com- plex (Van Ommen et al., 1980; Hensgens et al., 1983a; Macreadie et al., 1983). To study the composition of the different OX13 tran-

MULTIPLE box EFFECTS 43
bll bl2 bl3 blL bl5
q,lgmu; E
Ml233 WLLSB ML751
lML701J
HL921
rho-A%-1
FIG. 1. The organization of the apocytochrome b gene in 777-3A. Black bars represent sequences coding for apo- cytochrome b (exons Bl-6) and open bars represent the long URFs present within the introns. The region of wild- type mtDNA retained in the repeating unit of the petite mutant Al 5-1 (Van Ommen et al., 1980) is also shown, together with the positions of the different mutations. M475 1 contains a G-A transition 341 bp downstream of exon B4 (Weiss-Brummer et al., 1983), M4458 a CG- TT transition 340 and 341 bp downstream of B4 (Weiss- Brummer et al., 1982), and Ml431 a G-A transition 779 bp downstream of B4 (Jacq ef al.. 1982). For M4751 a suppressor has been isolated which contains a C-T tran- sition 47 bp downstream of B4 (Weiss-Brummer et al., 1983). Only the G-A transition in Ml431 gives rise to a nonsense codon in the b14 URF. The b13 mutant Ml392 is mutated within a &-acting sequence (Haid et al., 1980) which might fulfil a function in the splicing of b13 similar to the box2 and box9 sequences in splicing of b14. The position of this and other unsequenced mutants is based on genetic mapping described by Bechmann et al. ( 198 1).
scripts, the RNA blots were hybridized with probes specific for certain regions of the sub- unit I gene (see Fig. 2 for the location of se- quences retained).
The results obtained are discussed below.
Transcripts of the Apocytochrome b Gene
To establish the effects of mutations in the cytochrome b gene at the transcript level, mu- tant transcripts were analyzed with probe A 15 1 (Fig. 3). The patterns found confirm ex- pectations based on genetic analysis (see also Van Ommen et al., 1980, Church et al., 1979, and Haid et al., 1980. Mutations in the b12 intron (M 1233 and M 1282; Fig. 3, lanes j and k) prevent excision of the b12 intron and of the downstream introns b13 and b14. In these mutants a precursor of 6200 nucleotides ac- cumulates still containing these intron se- quences (Van Ommen et al., 1980). The b13 mutant Ml392 (Fig. 3, lane b) is blocked in the processing of the b13 and b14 introns, re-
sulting primarily in the accumulation of a 4800nucleotide transcript (exons + b13 + b14 sequences). The mutant M4924 (presumed to be mutated in B4) and the box9 mutant M4458 do not excise b14, giving rise to a tran- script of 3400 nucleotides (exons + b14 se- quences; Fig. 3, lanes c-e).
The other b14 mutants (M470 1, M475 1, and M 143 1; Fig. 3, lanes f-i) show a more complex pattern. In M470 1 and M475 1 pro- cessing of the b14 intron is strongly inhibited and both mutants contain a high concentra- tion of transcripts with the unexcised intron. Mature 18s mRNA is still present, however (Van Ommen et al., 1980). The temperature- sensitive box7 mutant M 143 1 (Fig. 3, lane h) contains a lowered concentration of 18s apo- cytochrome b mRNA with respect to wild type. Excision of the b14 intron is reduced in this mutant also, since a high concentration of a 3400-nucleotide transcript (exons + b14 sequences) is detected.
As might be expected, the bI1 and b15 in- trons are processed normally in all mutants. Neither intron contains a reading frame and processing does not require mitochondrial protein synthesis (Halbreich et al., 1980; Church and Gilbert, 1980).
Transcripts of the Gene for Subunit I of Cytochrome c Oxidase The mRNA for subunit I of cytochrome c
oxidase is a 18s transcript (2 100 nucleotides) (Hensgens et al., 1983a). The presence or ab- sence of this and other ~0x1 transcripts in the mutants was verified by the experiments shown in Figs. 4-6. A mutant is considered to contain 18s mRNA if it contains a tran- script of that size that hybridizes with the 5’ (Al) and 3’ (A6-8) exon probes (pIU41 and pKL114; see Fig. 2).
The b12 mutants Ml282 and Ml233 (Fig. 4, lanes a, b, and j), the B4 mutant M4924, the b14 mutants M4458, and Ml431 (Fig. 5, lanes c-e and h) clearly lack an 18s mRNA for subunit I. The box9 mutants M4701 and M475 1 contain 18s RNA in a very low con- centration (Fig. 5, lanes f and g; Fig. 4, lanes a, b, and j). From Figs. 4-6 it is evident that
44 HENSGENS, VAN DER HORST, AND GRIVELL
TABLE I
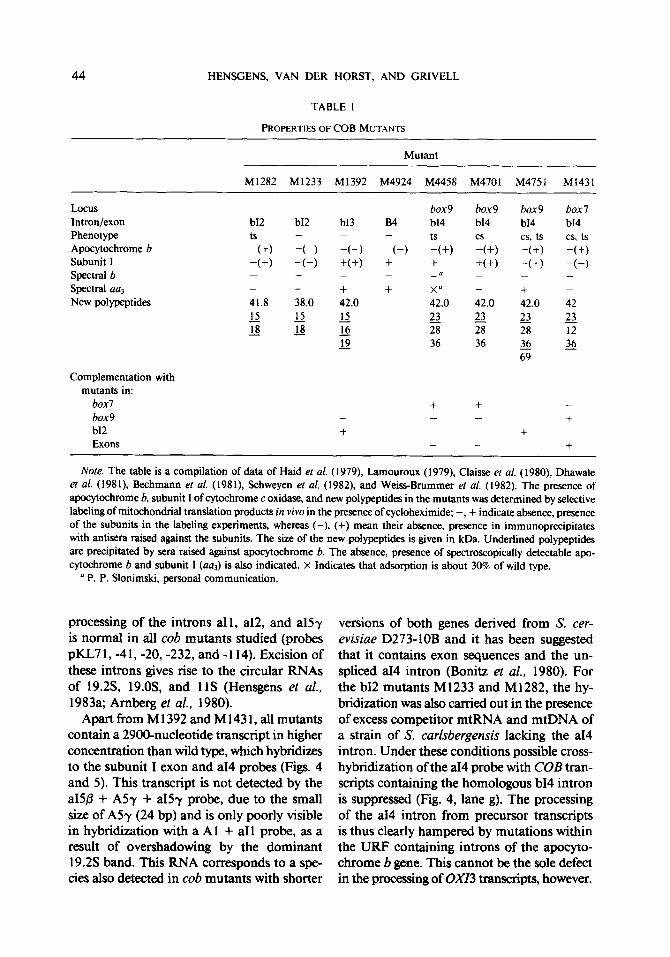
PROPERTIES OF COB MUTANTS
Mutant
MI282 Ml233 Ml392 M4924 M4458 M4701 M4751 Ml431
Locus Intron/exon Phenotype Apocytochrome b Subunit I Spectral b Spectral aa3 New polypeptides
box9 box9 box9 box7 b12 b12 b13 B4 b14 b14 b14 b14 ts - - - ts cs cs, ts cs, ts -(+I -(-) 4-l -c-j --(+I -(+) -(+) -(+) -(+I -(-I +(+I + + +(+I -(+) -(-) - - - - -u - - - - - + + X0 - + -
41.8 38.0 42.0 42.0 42.0 42.0 42
II II rs 23 22 22 22 18 18 16 28 28 28 12
22 36 36 36 36 69
Complementation with mutants in:
box7 box9 b12 Exons
+ + - - - - + + + - - - +
No@. The table is a compilation of data of Haid et al. (1979), Lamouroux (1979), Claisse ef al. ( 1980), Dhawale et al. (1981), Bechmann et al. (1981), Schweyen ef al. (1982), and Weiss-Bntmmer et al. (1982). The presence of apocytochrome b, subunit I of cytochrome c oxidase, and new polypeptides in the mutants was determined by selective labeling of mitochondtial translation products in viva in the presence of cycloheximide; -, + indicate absence, presence of the subunits in the labeling experiments, whereas (-), (+) mean their absence, presence in immunoprecipitates with antisera raised against the subunits. The size of the new polypeptides is given in kDa. Underlined polypeptides are precipitated by sera raised against apocytochrome 6. The absence, presence of spectroscopically detectable apo- cytochrome b and subunit I (a~) is also indicated. X Indicates that adsorption is about 30% of wild type.
a P. P. Slonimski, personal communication.
processing of the introns a1 1, a12, and aI5y is normal in all cob mutants studied (probes pKL7 1, -4 1, -20, -232, and - 114). Excision of these introns gives rise to the circular RNAs of 19.2S, 19.OS, and 11s (Hensgens et al., 1983a; Arnberg et al., 1980).
Apart from M 1392 and M 143 1, all mutants contain a 29OOnucleotide transcript in higher concentration than wild type, which hybridizes to the subunit I exon and a14 probes (Figs. 4 and 5). This transcript is not detected by the aI58 + A5y + aI5y probe, due to the small size of A5y (24 bp) and is only poorly visible in hybridization with a A 1 + a1 1 probe, as a result of overshadowing by the dominant 19.2s band. This RNA corresponds to a spe- cies also detected in cob mutants with shorter
versions of both genes derived from S. cer- evisiae D273- 10B and it has been suggested that it contains exon sequences and the un- spliced a14 intron (Bonitz et al., 1980). For the b12 mutants M 1233 and M 1282, the hy- bridization was also carried out in the presence of excess competitor mtRNA and mtDNA of a strain of S. carisbergensis lacking the a14 intron. Under these conditions possible cross- hybridization of the a14 probe with COB tran- scripts containing the homologous b14 intron is suppressed (Fig. 4, lane g). The processing of the a14 intron from precursor transcripts is thus clearly hampered by mutations within the URF containing introns of the apocyto- chrome b gene. This cannot be the sole defect in the processing of OX13 transcripts, however.

MULTIPLE box EFFECTS 45
on al4 am OISY OLl2
A2 A3 A4 A5y A6-6
Pm.25 -l9S
-11s
2600 ... 2100 -- > _ MI392
6100 ._.. -- ---_ ---.----- -.ssoo.!!!!~ _...-..__.__.. - ,,,233,J,,2@
, 41 I I?( I3 &I ‘8 I!% 121(11
, c , D 1 , 106 , I14 J
d&l
rho- LH30-A5
FIG. 2. Organization of the gene for subunit I of cytochrome c ox&se. Black bars represent sequences coding for subunit I (exons Al-8) and open bars represent the URFs present within the introns (ail-a15j3). The positions of the DNA sequences contained within the mtDNA of petite mutant LH30-A5 the different clones, and the isolated fragments used as probes in the hybridization experiments are given (see also Hensgens et al., 1983a,b). The position of the sequences coding for circular RNAs are indicated. Likely positions of the transcripts arising by aberrant processing in M 1233, M 1282, and M I392 are indicated by (-), with areas of uncertainty given by ( . - - ). *Indicates that the probe used consisted of a restriction fragment isolated from the designated clone (see Materials and Methods).
abcdafah ai jk
1
a5
kb
FIG. 3. COB transcripts in wild-type and cob mutants. mtRNA was prepared from S. cerevisiae 777-3A (a), and the cob mutants M 1392 (b), M4924 (c), M4458 grown at the permissive 28°C (d) and nonpermissive 35°C (e) temperatures, M4701 grown at permissive 28°C (f) and nonpermissive 35°C (g) temperatures, Ml431 (h), M4751 (i), Ml233 (i) and Ml282 (k). 1 pg mtRNA ofeach strain was electrophoresed through 1.25% (w/v) agarose gels and transferred to DBM paper. The RNA was hybridized with 32P-Iabelled mtDNA from the COB petite A 15- I. See Fig. 1 for location of the mutations. Electrophoretic mobilities (SE values) were calculated relative to 2 IS and 15s rRNAs.

46 HENSGENS, VAN DER HORST, AND GRlVELL
WT Ml233 Ml282 I I
kb
21 - 1s c I8 - 1s -
11 -
S,
WT
a
ML751
abd f j
6
L 3
2
kb
FIG. 4. Analysis of OX13 transcripts in different strains. mtRNA of the wild-type strain Saccharomyces cerevisiae 777-3A and of the cob mutants M 1233, M 1282, and M475 1 was electrophoresed, blotted, and hybridized with different probes from the OX13 region. The probe used in lane (a) (LH30-A5) also detects the 22s and 24s transcripts encoded by the genes for ATPase subunits 6 and 8. Sequences retained in the OX13 clones used as probes are shown in Fig. 2. A, Probe 1ll.E in the presence of an approximate 50- fold excess of mtRNA from S. carlsbergensis. ti indicates the positions of transcripts that possibly arise by cut-no-splice events (see text).
0x13-OL12 I Al41 1 a14
3 177-3A c ML92L e MLL50 U5T) g ML701 135’ Cl 5 Ml392 d MLL50 f ML701 h Ml131 135’Cl
bcdeloh obcdefrtl
0.5
kb
aisp o1-S+l.A6y*alSy.A6-0
FIG. 5. Analysis of OX13 transcripts in different apocytochrome b mutants. mtRNA was prepared from wild-type S. cerevisiae 777-3A (a), mutants Ml392 (b), M4924 (c), M4458 grown at permissive (d) and nonpermissive temperature (e), M4701 grown at permissive (f) and nonpermissive temperature (g), and Ml431 (h). mtRNA (1 pg) of each strain was electrophoresed, blotted, and hybridized with 32P-labeled probes from the gene for subunit 1. Probes used were as follows: 0X13-OLI2, LH30-AS; A 1 + a1 1, pKL4 1; a14, 350-bp SadA fragment from pKLll1; aISa, 600-bp Sau3A fragment from pKLlO9; aIS/ + A5y + aISy, pKL232; a15j3, Ml3 clone 2411; a15,9 + A5y + aI5y + A6-8, pKLl14.

MULTIPLE box EFFECTS 47
FIG. 6. Analysis of OX13 transcripts in different apocytochrome b mutants. Probes used were: a1 1, pKL7 1; a12, pKL20, a13, 465-bp Sau3A fragment from pKLD. See legend to Fig. 5 for further details.
The mutant transcripts display a size and con- centration distribution which differs from each other and from the pattern present in wild type (Fig. 4, lane a; Fig. 5, probe 0X13-OL12). This indicates that the level and/or mode of inhibition of transcript processing differs from mutant to mutant (see below).
box3 and box10 Mutants Display Novel box Eficts
Three specific OX13 transcripts are present in the b12 mutants M 1233 and M 1282 (ar- rowed in Fig. 4a). These do not correspond either in size or hybridization behavior to wild- type OX13 transcripts or OX13 transcripts in other cob mutants studied. Two transcripts of 4900 and 6500 nucleotides, respectively, hy- bridize with aISo, aIS@, and aI5y probes but not with the leader exon Al and intron a11 probe pKL41 and with the a13, A4, and a14 probe pKLD (Fig. 4). A transcript of 6100 nucleotides hybridizes only with pKLD, pKL20, and pKL41 probes (Fig. 4; these probes are all upstream of a14). These data suggest that the 4900- and 65OOnucleotide transcripts contain only 3’ portions of the gene and that the 6100-nucleotide transcript con- tains only 5’ portions of the gene, respectively. As such, they could arise by cut-no-splice events in the region between A4 and A5a.
In the b13 mutant Ml392 a very strong hybridization is detected with two OX13 tran- scripts of approximately 2200 and 2800 nu- cleotides. These hybridize with a probe con- taining sequences of aIS@ only, with probe pKL232 (aI5& aISy, and A57), and with probe pKLl14 (aI5/3, A5y, aISy, and A6-8; see Fig. 2 for localization and Fig. 5). These transcripts are not detected by the same probes in the wild-type strain, or the other mutants. Both RNAs hybridize equally strongly to probes containing sequences of aI5-y (pKL 114, 232), but in hybridizations with an aIS@ probe, the signal with the 2200nucleotide RNA is much stronger. These observations, combined with the size of the transcripts suggests that the only difference between these two tran- scripts is the absence of intron aI5y sequences from the shorter transcript. This intron has presumably been removed by nuclear-encoded splicing enzymes (Hensgens et al., 1983a). These data strongly suggest that in the b13 mutant, the 2200- and 2800nucleotide tran- scripts arise by a cut-no-splice event at the 5’ end of the aI50 intron. Some normal pro- cessing must occur, however, as this mutant also shows hybridization at the 18s mRNA position with an Al + a11 probe. Our failure to detect the 5’ products of the cut-no-splice event might be due to their instability or their

48 HENSGENS, VAN DER HORST, AND GRIVELL
being masked by transcripts with similar mo- bilities and hybridization behavior.
Although the procedure used in the exper- iments of Figs. 3-6 has provided useful in- formation, the method has its limitations. We have not been able, for example, to precisely define the nature of the defect in the processing of OX13 transcripts in the box7 mutant M 143 1. Many high-molecular-weight tran- scripts have been detected in this mutant and they are larger than the expected transcript of 2900 nucleotides (Fig. 5, probe 0X13-OLZ2, lane h). These RNAs fail to hybridize with an a14 probe, however (panel a14, see also Dis- cussion). Another puzzling result is observed in the mutants M1392, M4924, M4458, and M4701. When probes retaining aISa, aI5& and aI5y intron sequences are used, these mu- tants are found to contain many transcripts that are absent from the corresponding wild- type strain 777-3A (Fig. 5, lanes b-g). The exact composition of these transcripts cannot be resolved by the data available at present. The same probes detect transcripts with similar mobilities present in very low concentration in petite mutants lacking mitochondrial pro- tein synthesis. It is possible that they arise by the action of nuclear-encoded enzymes in cells in which mitochondrially encoded maturase(s) are inactive (or absent). Further experimen- tation is needed to settle these issues.
DISCUSSION
Is the cob-box Effect Solely Mediated by the “box7” Maturase?
It has been proposed that a maturase en- coded by the b14 URF of the apocytochrome b gene is required for expression of the gene for subunit I of cytochrome c oxidase (Bonitz et al., 1980; De la Salle et al., 1982; Dujardin et al., 1982; Dhawale et al., 1982; Jacq et al., 1982; Labouesse and Slonimski, 1983; Iamb et al., 1983; Mahler et al., 1982; Weiss-Brum- mer et al., 1982). In this report we have studied the effect of mutations in b14 and other do- mains of the apocytochrome b gene on the processing of transcripts of the gene for subunit I. We show that in most cob mutants in which
the expression of the b14 URF is disturbed (as judged by transcript analysis of the COB gene), processing of the a14 intron is also ham- pered. The concentration of precursors con- taining this intron is unexpectedly low, how- ever, possibly as a result of a high turnover rate of these transcripts and/or competition from the 2 1 S ribosomal RNA band for binding to the DBM paper, since this RNA migrates at approximately the same position as the 2900nucleotide (exons + a14) precursor.
Somewhat surprisingly, no transcripts con- taining aI sequences are observed in the only box7 mutant studied (M1431), even though this mutant-which is both cold- and heat- sensitive-accumulates other high-molecular- weight transcripts. Leakiness most probably accounts for this absence, although compe- tition for binding to DBM paper as described above may also contribute. The pattern of high-molecular-weight transcripts differs from that of wild-type and the other mutants (Fig. 5), suggesting that at least one other processing step is disturbed, however.
Our results show that besides b14 other parts of the apocytochrome b gene are involved in correct expression of the subunit I gene, since mutations outside the b14 intron also result in alterations in the pattern of OX13 process- ing, superimposed on a block in a14 excision. These other mutations do not necessarily lead to the complete absence of subunit I 18s mRNA. Clear examples of the consequences of such mutations are the accumulation of 2200- and 2800nucleotide transcripts in the b13 mutant Ml392 (Fig. 5, lane b) and the transcripts of 4900,6 100 and 6500 nucleotides present in the b12 mutants M 1233 and M 1282 (Fig. 4, lanes b, d-f, and g-i). These transcripts most likely result from cut-no-splice events at the ASP-aIS/ and at the A4-a14 or a13 bor- ders, respectively. The fact that b12 and b13 mutants contain different abnormal OX13 transcripts indicates that at least two different apocytochrome b gene products can interfere with OX13 processing. A shortened b12 ma- turase is present in the b12 mutants (wild type, a 42-kDa; M1282, a 41.8-kDa; M1233, a 38- kDa protein: Bechmann et al., 198 1; Claisse

MULTIPLE box EFFECTS 49
et al., 1980) and a putatively mutated b13 ma- turase in the b13 mutant M 1392 (Haid et al., 1980; Kreike, 1982). The mutations prevent the maturases from processing the introns which encode them, but their involvement in the processing of OX13 transcripts is unclear. They may normally play a role in splicing OX13 transcripts, in which case a mutation could interfere with activity. Alternatively, they may not be required for OX13 processing in wild type and only interfere in mutated form with the splicing machineries of 0X13, either directly or by producing a high amount of unusual COB precursor transcripts. In this respect it is noteworthy that in previous studies of COB transcripts by Van Ommen et al. (1980) the excision of upstream introns is hampered in some cases by a downstream in- tron mutation. This observation may serve as an indication that mutated intron products or the presence of large amounts of splicing intermediates can interfere with correct pro- cessing of other introns. Comparison of our results with those of Mahler et al. (1982) and Labouesse and Slonimski ( 1983) provides fur- ther evidence in this direction. In the latter studies no effect on the expression of the sub- unit I gene was found when COB introns other than b14 were deleted. Such deletions would clearly prevent accumulation of faulty ma- turases and/or unspliced intron sequences. Since the processing of b13 and b14 is ham- pered in both b12 and b13 mutants and the observed defects in OX13 processing are dif- ferent, it is possible that the mutated bI3-en- coded “maturase” rather than the presence of unspliced b13 and b14 intron sequences is re- sponsible for the defects in the b13 mutant M1392.
box9 Mutants For the box9 mutants (especially M4458
and M4701) it has been shown that they can complement box7 mutants (Haid et al., 1980; Weiss-Brummer et al., 1982). These results suggest that in box9 mutants an active box7- encoded trans-acting element is made. Nev- ertheless, processing of OX13 transcripts is disturbed in these box9 mutants (Fig. 5). No
~0x1 18s mRNA is detected in the mutant M4458, while its concentration is low in the conditional mutants M470 1 and M475 1. It has been suggested that the mutations they contain (missense mutations for M4458 and M475 1; Weiss-Brummer et al., 1982, 1983) prevent the formation of a secondary structure of the b14 intron, necessary for proper excision (see also Davies et al., 1982, and Michel and Dujon, 1983). For M4751 a suppressor mu- tation has been found in the b14 intron 47 bp downstream from the B4 exon. This mutation (C-T transition) presumably restores the re- quired secondary structure (Weiss-Brummer et al., 1983). Genetic studies and sequence comparisons (Jacq et al., 1982; Weiss-Brum- mer et al., 1982, 1983; Hensgens et al., 1983b) suggest that this part of the b14 URF is not important for the activity of the trans-acting box7 maturase. Although we cannot precisely define which steps in the processing of OX13 transcripts are disturbed in these box9 mu- tants, it is unlikely that only the processing of a14 is inhibited. The transcript patterns de- tected with the aISa, aI5& and A6-8 probes differ from the patterns detected in the box7 mutant Ml431 and the wild-type strain. A high concentration of unspliced b14 sequences or of a mutant box7 maturase (containing the missense mutation) might interfere with the processing of OX13 introns as discussed above for the b12 and b13 mutants.
We have observed that processing of the aI1, a12, and aI5y introns, which belong to group II mitochondrial introns, is not dis- turbed in any of the mutants studied. The disturbance of OX13 processing only occurs within the sequences of the a13, a14, aISa, and aI5fl introns. In the mutants studied, the le- sions are present in the b12, b13, or b14 introns of the apocytochrome b gene. All these seven introns belong to one class of mitochondrial introns, which share sequence and potential secondary structure homology (Davies et al., 1982; Michel and Dujon, 1983) and which contain URFs coding for proteins likely to be involved in intron removal. These putative maturases have areas of considerable sequence homology at the amino acid level (Hensgens

50 HENSGENS, VAN DER HORST, AND GRIVELL
et al., 1983b), suggesting both that they are derived from a common ancestor and that they possess functional homology. Such ho- mology could provide an explanation for the pleiotropic effects on splicing that we have observed.
ACKNOWLEDGMENTS
We are indebted to Dr. R. J. Schweyen (Genetisches Institut, Maria-Ward-Strasse la, D-8000 Miinchen 19, Germany) for generous gifts of mutant and wild-type strains. We thank Mr. M. De Haan for his help in some of these experiments and Dr. R. Benne for critical reading of the manuscript. This work was supported in part by a grant to P. Borst-L. A. Grivell from The Netherlands Foundation for Chemical Research (SON) with financial aid from The Netherlands Organization for the Advance- ment of Pure Research (ZWO).
REFERENCES
ARNBERG, A. C., VAN OMMEN, G. J. B., GRIVELL, L. A., VAN BRUGGEN, E. F. J., AND BORST, P. (1980). Some yeast mitochondrial RNAs are circular. Cell 19, 313-319.
BECHMANN, H., HAID, A., SCHWEYEN, R. J., MATHEWS, S., AND KAUDEWITZ, F. (1981). Expression of the split gene COB in yeast mtDNA. Translation of intervening sequences in mutant strains. J. Biol. Chem. 256,3525- 3531.
BONITZ, S. G., CORUZZI, G., THALENFELD, B. E., TZA- GOLOFF, A., AND MACINO, G. (1980). Assembly of the mitochondrial membrane system, structure and nu- cleotide sequence of the gene coding for subunit I of yeast cytochrome oxidase. J. Biol. Chem. 255, Il927- 11941.
CHURCH, G. M., AND GILBERT, W. (1980). Yeast mi- tochondrial intron products required in trans for RNA splicing. In “Mobilization and Reassembly of Genetic Information” (D. R. Joseph, J. Schultz, W. A. Scott, and R. Werner, eds.), pp. 379-394. Academic Press, New York.
CHURCH, G. M., SLONIMSKI, P. P., AND GILBERT, W. (1979). Pleiotropic mutations within two yeast mito- chondrial cytochrome genes block mRNA processing. Cell 18, 1209-1215.
CLAISSE, M. L., SLONIMSKI, P. P., JOHNSON, J., AND MAHLER, H. R. (1980). Mutations within an intron and its flanking sites: Patterns of novel polypeptides generated by mutants in one segment of the cob-box region of yeast mitochondrial DNA. Mol. Gen. Genet. 177, 375-387.
DAVIES, R. W., WARING, R. B., RAY, J. A., BROWN, T. A., AND SCAZZOCCHIO, C. (1982). Making ends meet: A model for RNA splicing in fungal mitochondria. Na- lure (London) 300, 7 19-724.
DE LA SALLE, H., JACQ, C., AND SLONIMSKI, P. P. (1982). Critical sequences within mitochondrial introns: Pleio- tropic mRNA maturase and cisdominant signals of the box intron controlling reductase and oxidase. Cell 213, 721-732.
DHAWALE, S., HANSON, D. K., ALEXANDER, N. J., PERL- MAN, P. S., AND MAHLER, H. R. (1981). Regulatory interactions between mitochondrial genes: Interactions between two mosaic genes. Proc. NaB Acad. Sci. USA. 78, 1778-1782.
DUJARDIN, G., JACQ, C., AND SLONIMSKI, P. P. (1982). Single base substitution in an intron of oxidase gene compensates splicing defects of the cytochrome b gene. Nature (London) 298, 628-632.
DUJON, B. (I 98 I). Mitochondrial genetics and functions. In “Molecular Biology of the Yeast Saccharomyces: Life Cycle and Inheritance” (J. N. Strathem, E. W. Jones, and J. R. Broach, eds.), pp. 505-635. Cold Spring Harbor Laboratory, Cold Spring Harbor, N. Y.
GRIVELL, L. A. (1983). Mitochondrial DNA. Sci. Amer. 248, 78-89.
HAID, A., GROSCH, G., SCHMELZER, C., SCHWEYEN, R. J., AND KAUDEWITZ, F. (1980). Expression of the split gene COB in yeast mtDNA. Mutational arrest in the pathway of transcript splicing. Curr. Genet. 1, l55- 161.
HAID, A., SCHWEYEN, R. J., BECHMANN, H., KWDEWITZ, F., SOLIOZ, M., AND SCHATZ, G. (1979). The mito- chondrial COB region in yeast codes for apocytochrome b and is mosaic. Eur. J. Biochem. 94, 451-464.
HALBREICH, A., PAJOT, P., FOUCHER, M., GRANDCHAMP, C., AND SLONIMSKI, P. P. (1980). A pathway of cy-
tochrome b mRNA processing in yeast mitochondria: specific splicing steps and an intron-derived circular RNA. Cell 19, 32 l-329.
HENSCENS, L. A. M., ARNBERG, A. C., ROOSENDAAL, E., VAN DER HORST, G., VAN DER VEEN, R., VAN OMMEN, G. J. B., AND GRIVELL, L. A. (1983a). Vari- ation, transcription and circular RNAs of the mito- chondrial gene for subunit I of cytochrome c oxidase. J. Mol. Biol. 164, 35-58.
HENSGENS, L. A. M., B~NEN, L., DE HAAN, M., VAN DER HORST, G., AND GRIVELL, L. A. (1983b). Two intron sequences in yeast mitochondrial COXI gene: Homology among URF-containing introns and strain- dependent variation in flanking exons. Cell 32, 379- 389.
JACQ, C., PAJOT, P., LAZOWSKA, J., DUJARDIN, G., CLAISSE, M., GROUDINSKY, O., DE LA SALLE, H., GRANDCHAMP, C., LABOUESSE, M., GARGOURI, A., GUIARD, B., SPYRIDAKIS, A., DREYFW., M., AND SLONIMSKI, P. P. (1982). Role of introns in the yeast cytochrome b gene: Cis- and transacting signals, intron manipulations, expression and intergenic communi- cations. In “Mitochondrial Genes” (P. P. Slonimski, P. Borst, and G. Attardi, eds.), pp. 155-183. Cold Spring Harbor Laboratory, Cold Spring Harbor, N. Y.
JEFFREYS, A. J., AND FLAVELL, R. A. (1977). The rabbit

MULTIPLE box EFFECTS 51
&lobin gene contains a large insert in the coding se- quence. Cell 12, 1097-I 108.
KREIKE, J. (1982). “The Biosynthesis of Complex III in Yeast.” Ph.D. Thesis, Rodopi, Amsterdam.
LABOUESSE, M., AND SLONIMSKI, P. P. (1983). Construe- tion of novel cytochrome b genes in yeast mitochondtia by subtraction or addition of introns. EMBO .I. 2,269- 276.
LAMB, M. R., ANZIANO, P. Q., GLAUS, K. R., HANSON, D. K., KLAPPER, H. J., PERLMAN, P. S., AND MAHLER, H. R. (1983). Functional domains in introns. RNA processing intermediates in cis- and trans-acting mutants in the penultimate intron of the mitochondrial gene for cytochrome b. J. BioI. Chem. 258, 199 I - 1999.
LAMOUROUX, A. (1979). “Compl6mentation in vivo: Moyen d&de de l’expression du gene morcele mi- tochondrial codant pour l’apocytochrome b chez Suc- charomyces cerevisiae.” Ph.D. Thesis, Universite Paris Xl, Paris, France.
LAZOWSKA, J., JACQ, C., AND SLONIMSKI, P. P. (1980). Sequence of introns and flanking exons in wild-type and box-3 mutants of cytochrome b reveals an interlaced splicing protein coded by an intron. Cell 22, 333-348.
MACREADIE, I. G., NOVITSKI, C. E., MAXWELL, R. J., ULRIK, J., 001, B.-G., MCMULLEN, G. L., LUKINS, H. B., LINNANE, A. W., AND NAGLEY, P. (1983). Bio- genesis of mitochondria: the mitochondrial gene (aapl) coding for mitochondrial ATPase subunit 8 in Sac- charomyces cerevisiae. Nucleic Acids Res. It, 4435 4451.
MAHLER, H. R., HANSON, D., MILLER, D., LIN, C. C., ALEXANDER, N. J., VINCENT, R. D., AND KRLMAN, P. S. (1978). Regulatory aspects of mitochondrial bio- genesis. In “Biochemistry and Genetics of Yeast: Pure and Applied Aspects” (M. Bacila, B. L. Horecker, and A. 0. M. Stoppani, eds.), pp. 5 13-547. Academic Press, New York.
MAHLER, H. R., HANSON, D. K., LAMB, M. R., PERLMAN, P. S., ANZIANO, P. Q., GLAUS, K. R., AND HALDI, M. L. (1982). Regulatory interactions between mito- chondrial genes: Expressed introns-Their function and regulation. In “Mitochondrial Genes” (P. P. Slonimski, P. Borst, and G. Attardi, eds.), pp. 185-199. Cold Spring Harbor Laboratory, Cold Spring Harbor, N. Y.
MICHEL, F., AND DUJON, B. (1983). Conservation of RNA secondary structures in two intron families including mitochondrial-, chloroplast- and nuclear-encoded members. EMBO J. 2, 33-38.
NETTER, P., JACQ, C., CARIGNANI, G., AND SLONIMSKI, P. P. (1982). Critical sequences within mitochondrial introns: C&dominant mutations of the “cytochrome- b-like” intron of the oxidase gene. Cell 28, 733-738.
PAJOT, P., WAMBIER-KLUPPEL, M. L., KOTYLAK, Z., AND SLONIMSKI, P. P. (1976). Regulation of cytochrome oxidase formation by mutations in a mitochondrial gene for cytochrome b. In “Genetics and Biogenesis of Chlo- roplasts and Mitochondria” (T. Biicher, W. Neupert, W. Sebald, and S. Werner, eds.), pp. 443-451. North- Holland, Amsterdam.
SCHWEYEN, R. J., FRANCISCI, S., HAID, A., OSTERMAYR, R., R~)DEL,G.,SCHMELZER, CSCHROEDER, R., WEISS- BRUMMER, B., AND KAUDEWITZ, F. (1982). Transcripts of yeast mitochondrial DNA: Processing of a split-gene transcript and expression of RNA species during ad- aptation and differentiation processes. In “Mitochon- drial Genes” (P. Slonimski, P. Borst, and G. Attardi, eds.), pp. 20 1-2 12. Cold Spring Harbor Laboratory, Cold Spring Harbor, N. Y.
VAN OMMEN, G. J. B., GROOT, G. S. P., AND GRIVELL, L. A. (1979). Transcription maps of mtDNAs of two strains of Saccharomyces: Transcription of strain-specific insertions; complex RNA maturation and splicing. Cell 18, 511-523.
VAN OMMEN, G. J. B., BOER, P. H., GROOT, G. S. P., DE HAAN, M., ROOSENDAAL, E., GRIVELL, L. A., HAID, A., AND SCHWEYEN, R. J. (1980). Mutations affecting RNA splicing and the interaction of gene expression of the yeast mitochondrial loci COB and 0X13. Cell 20, 173-183.
WEISS-BRUMMER, B., R~DEL, G., SCHWEYEN, R. J., AND KAUDEWITZ, F. (1982). Expression of the split gene cob in yeast: Evidence for a precursor of a L‘maturase” protein translated from intron 4 and preceding exons. Cell 29, 527-563.
WEISS-BRUMMER, B., HOLL, J., SCHWEYEN, R. J., R~DEL, G., AND KAUDEWITZ, F. (1983). Processing of yeast mitochondrial RNA: Involvement of intramolecular hybrids in splicing of cob intron 4 RNA by mutation and reversion. Cell 33, 195-202.




















