
Lymphokines Enhance the Capacity of Human Monocytes to Secrete Reactive Oxygen Intermediates
Upload
independentCategory
view
0download
0
Synergistic stimulation of human monocytes anddendritic cells by Toll-like receptor 4 and NOD1- andNOD2-activating agonistsJ�rg H. Fritz1, Stephen E. Girardin2, Catherine Fitting3, Catherine Werts1, Dominique Mengin-Lecreulx4, Martine Caroff5, Jean-Marc Cavaillon3, Dana J. Philpott1 and Minou Adib-Conquy3
1 Groupe d'Immunit� Inn�e et Signalisation, Institut Pasteur, Paris, France2 Unit� de Pathog�nie Microbienne Mol�culaire, Institut Pasteur, Paris, France3 Unit� Postulante Cytokines and Inflammation, Institut Pasteur, Paris, France4 Enveloppes Bact�riennes et Antibiotiques, IBBMC, UMR 8619 CNRS, Universit� Paris-Sud, Orsay, France5 Equipe Endotoxines, IBBMC, UMR 8619 CNRS, Universit� Paris-Sud, Orsay, France
Muropeptides are degradation products of bacterial peptidoglycan (PG) sensed bynucleotide-binding oligomerization domain 1 (NOD1) and NOD2, members of arecently discovered family of pattern recognition molecules (PRM). One of thesemuropeptides, muramyl dipeptide (MDP) mediates cell signaling by NOD2, exertsadjuvant activity and synergizes with lipopolysaccharide (LPS) to induce pro-inflammatory responses in vitro and in vivo. In contrast, few and contradictory resultsexist about the stimulatory capacity of NOD1 agonists. Thus, the ability of NOD1(MurNAc-L-Ala-c-D-Glu-meso-diaminopimelic acid, MtriDAP) and NOD2 (MurNAc-L-Ala-D-isoGln, MDP; MurNAc-L-Ala-c-D-Glu-L-Lys, MtriLYS) agonists to activate primaryhuman myeloid cells was examined. We show that both CD14+ monocytes and CD1a+
immature dendritic cells (DC) express NOD1 and NOD2 mRNA. Stimulation of primaryhuman monocytes and DC with highly purified muropeptides (MtriDAP, MDP andMtriLYS) induces release of pro-inflammatory cytokines. We reveal here that NOD1 aswell as NOD2 agonists act cooperatively with LPS to stimulate the release of both pro-and anti-inflammatory cytokines in these myeloid cell subsets. Finally, we report thatNOD1 as well as NOD2 agonists synergize with sub-active doses of LPS to induce DCmaturation, demonstrating that NOD agonists act cooperatively with molecules sensedby Toll-like receptor 4 to instruct the onset of adaptive immune responses.
Introduction
Innate immune detection of pathogens relies on specificsensing of conserved pathogen-associated molecularpatterns (PAMP). Several classes of proteins, termedpattern recognition molecules (PRM), have beenassigned to participate in these recognition events.Among the group of PRM, Toll-like receptors (TLR) andNACHT-LRR (domain present in NAIP, CIITA, HET-E andTP1with leucine-rich repeat) proteins (NLR) possess thecapability to convert the information gleaned fromdetection of microbes to elicit appropriate downstreamsignaling events [1, 2]. Defining the minimal structuresof microbial molecules required for full-fledged innate
Correspondence:Dana J. Philpott, Groupe d'Immunit� Inn�e etSignalisation, Institut Pasteur, 25–28 rue du Dr Roux, 75724Paris, France, Fax: +33-1-4061-3963, e-mail: [email protected]
Received 8/4/05Revised 23/5/05
Accepted 8/6/05
[DOI 10.1002/eji.200526286]
Key words:NOD proteins
� Purified LPS � Innateimmune recognition
� Monocytes� Dendritic cells
Abbreviations: CIITA: MHC class II transactivator � DAP: meso-diaminopimelic acid � FK156: D-lactyl-L-Ala-c-D-Glu-meso-DAP-Gly � GMtriDAP: GlcNAc-MurNAc-L-Ala-c-D-Glu-meso-diaminopimelic acid � HET-E: plant het gene product involvedin vegetative incompatibility � iE-DAP: c-D-Glu-meso-DAP �LAL: limulus amebocyte assay � LRR: leucine-rich repeat �LTA: lipoteichoic acid � MDP: muramyl dipeptide (MurNAc-L-Ala-D-isoGln) � MtriDAP: MurNAc-L-Ala-c-D-Glu-meso-diami-nopimelic acid � MtriLYS: MurNAc-L-Ala-c-D-Glu-L-Lys �NACHT: domain present in NAIP, CIITA, HET-E and TP1 �NAIP: neuronal apoptosis inhibitor protein � NLR: NACHT-LRRproteins � NOD: nucleotide-binding oligomerization domain �PAMP: pathogen-associated molecular patterns �PG: peptidoglycan � PRM: pattern recognition molecules �TP1: telomerase-associated protein 1
Eur. J. Immunol. 2005. 35: 2459–2470 Innate immunity 2459
f 2005 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.eji.de

immune activation is of extreme importance forgathering knowledge of downstream pathways inducedby individual PAMP-PRM engagement. During thecourse of the last years, the microbial motifs for manyTLR have been deciphered, while quite recently the firstNLR, NOD1 and NOD2, have been described to beinvolved in innate immune sensing by recognizingspecific fragments of bacterial peptidoglycan (PG), alsoknown as muropeptides [3]. Earlier reports suggestedthat TLR2 detects PG [4]. However, it has been shownrecently that TLR2-mediated PG sensing is due toimpurities in the preparations, indicating that PG devoidof contaminating lipopeptides or lipoteichoic acid (LTA)is not sensed by TLR2 [5]. Thus, together with PGrecognition proteins, NLRs such as NOD1 and NOD2 arekey sensors towards bacterial PG [3].
It has been known well that, whereas a lysine (Lys)residue is found in third position of the PG stem peptidein most Gram-positive bacteria, an unusual amino acid –meso-diaminopimelic acid (DAP) – replaces Lys in Gram-negative bacteria [6]. Interestingly, these differences aresensed by NOD1 and NOD2, indicating that these PRMmight have non-overlapping specificities towards PG.NOD1 senses a naturally occurring PG breakdownproduct GlcNAc-MurNAc-L-Ala-c-D-Glu-meso-diamino-pimelic acid (GMtriDAP) foundmostly in Gram-negativebacteria [7]. Using synthetic peptides, is has been shownthat the minimal structure detected by NOD1 is thedipeptide c-D-Glu-meso-DAP (iE-DAP), neither aminoacid being present in eukaryotes. In contrast, biochem-ical and functional analyses reveal that muramyldipeptide MurNAc-L-Ala-D-isoGln (MDP) is the minimalmotif sensed by NOD2 [7]. Further analysis of the PGstructural requirements allowing sensing by NOD2 haveshown that, in addition to MDP, NOD2 can detectMtriLYS (MurNAc-L-Ala-c-D-Glu-L-Lys), a PG structurepresent in Gram-positive bacteria [7]. However, MDP isthe minimal component common to the PG of bothGram-negative and Gram-positive bacteria, identifyingNOD2 as a general sensor of bacterial infection throughthe recognition of MDP [8].
Several reports have demonstrated that MDP canstimulate murine and human macrophages and mono-cytes for production of cytokines and chemokines(reviewed in [9]). In addition, it has been shown thatMDP can act synergistically with lipopolysaccharide(LPS) to induce inflammatory cytokine production invitro [10, 11]. Furthermore, it was reported that MDP-primed mice display enhanced sensitivity to LPS-induced septic shock and anaphylactic reactions[12–14]. In naturally occurring pathological conditionssuch as septic shock or bacterial infection, MDP and LPSare likely to be found simultaneously in the same tissues,which further reinforces the notion that such synergymight be of great physiological importance [15].
In contrast to studies done with the NOD2 agonistMDP, few and contradictory reports about the reactivityof NOD1 agonists towards myeloid cells exist. In mousemacrophages, iE-DAP stimulation was reported toinduce TNF-a and IL-6 secretion, and when combinedwith LPS generated a larger IL-6 response than LPSalone [16]. Furthermore, studies with the humanmyelomonocytic cell line THP-1 revealed that a DAP-containing lactoylpeptide, FK156 (D-lactyl-L-Ala-c-D-Glu-meso-DAP-Gly), induces IL-8 production by itselfand in synergy with TLR2, TLR4 and TLR9 ligands [17].However, a recent report showed that NOD1 agonistswere not able to synergize with LPS for TNF-a release bywhole human blood cells [11].
In this study, the capacity of the NOD1 agonistMtriDAP, and the NOD2 agonists MDP and MtriLYS toinduce cytokine and chemokine release by primaryhuman monocytes and DC was examined. The extent towhich NOD1, as well as NOD2 agonists, are able tostimulate cytokine and chemokine release in synergywith LPS in thesemyeloid cell populations was analyzed.For the first time, the capacity of various muropeptidesto induce DC maturation alone or in synergy with LPSwas investigated.
Results
Assessment of the purity of LPS and muropeptidepreparations and of the sensitivity of NOD1 andNOD2 agonist detection
A major obstacle in studying interactions betweendifferent bacterial cell wall components like LPS, PG,or lipoproteins is the difficulty to obtain highly purifiedmolecules free of contamination. Such purity is difficultto achieve with conventional isolation procedures, but isessential for studying synergistic interactions betweenbacterial components. Thus, the purity of LPS prepara-tions used in our assays was assessed using HEK293Tcells transiently transfected with TLR2, NOD1 or NOD2(Fig. 1A). The highly purified LPS used in all experi-ments was devoid of lipopeptide or PG contaminations,as no NF-jB activation was observed in TLR2-, NOD1- orNOD2-transfected cells. In contrast, the three commer-cially available LPS preparations that we tested werecontaminated with lipopeptides (LPS crude 2), or bothPG and lipopeptides (LPS crude 1 and crude 3, Fig. 1A).Pam3CSK4 was used as a positive control for NF-jBactivation through TLR2. To fully ensure the ability todetect contaminants, we further assessed the sensitivityof the applied assay. As shown in Fig. 1B, muropeptides,which included both NOD1- as well as NOD2-activatingagonists, are very potent activators of the NF-jBtranscription factor. Maximal activation was observed
J�rg H. Fritz et al. Eur. J. Immunol. 2005. 35: 2459–24702460
f 2005 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.eji.de
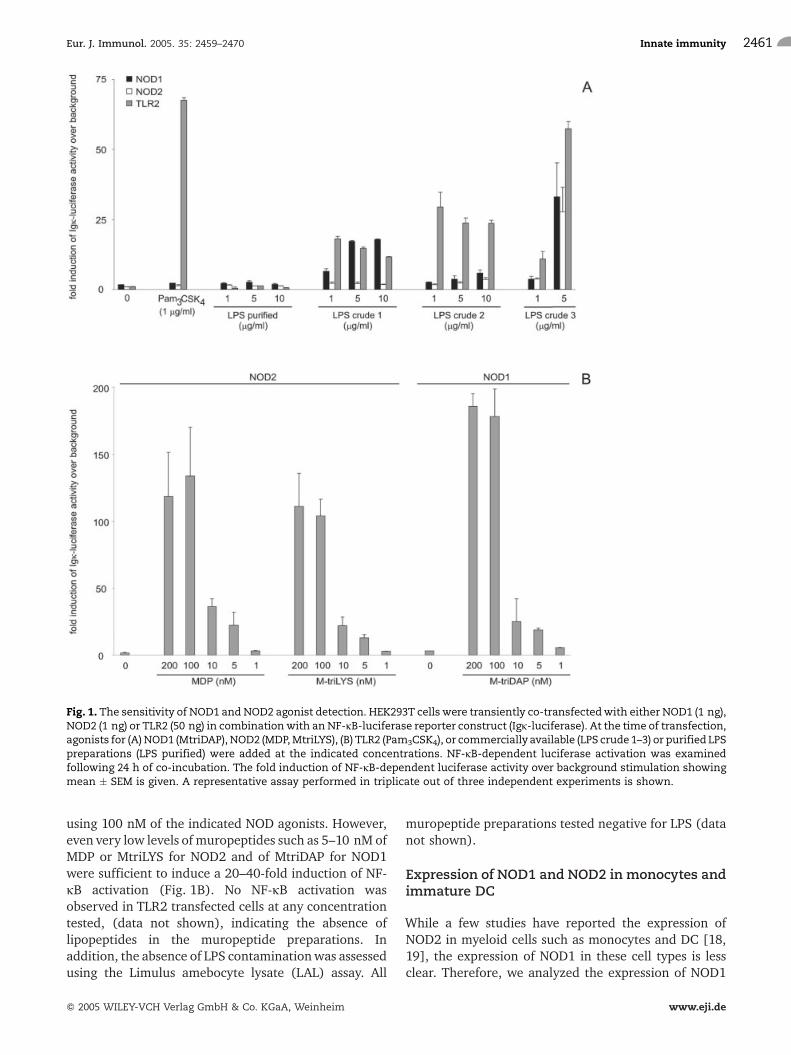
using 100 nM of the indicated NOD agonists. However,even very low levels of muropeptides such as 5–10 nM ofMDP or MtriLYS for NOD2 and of MtriDAP for NOD1were sufficient to induce a 20–40-fold induction of NF-jB activation (Fig. 1B). No NF-jB activation wasobserved in TLR2 transfected cells at any concentrationtested, (data not shown), indicating the absence oflipopeptides in the muropeptide preparations. Inaddition, the absence of LPS contaminationwas assessedusing the Limulus amebocyte lysate (LAL) assay. All
muropeptide preparations tested negative for LPS (datanot shown).
Expression of NOD1 and NOD2 in monocytes andimmature DC
While a few studies have reported the expression ofNOD2 in myeloid cells such as monocytes and DC [18,19], the expression of NOD1 in these cell types is lessclear. Therefore, we analyzed the expression of NOD1
Fig. 1. The sensitivity of NOD1 and NOD2 agonist detection. HEK293T cells were transiently co-transfectedwith either NOD1 (1 ng),NOD2 (1 ng) or TLR2 (50 ng) in combinationwith an NF-jB-luciferase reporter construct (Igj-luciferase). At the time of transfection,agonists for (A)NOD1 (MtriDAP), NOD2 (MDP,MtriLYS), (B) TLR2 (Pam3CSK4), or commercially available (LPS crude 1–3) or purified LPSpreparations (LPS purified) were added at the indicated concentrations. NF-jB-dependent luciferase activation was examinedfollowing 24 h of co-incubation. The fold induction of NF-jB-dependent luciferase activity over background stimulation showingmean � SEM is given. A representative assay performed in triplicate out of three independent experiments is shown.
Eur. J. Immunol. 2005. 35: 2459–2470 Innate immunity 2461
f 2005 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.eji.de

and NOD2 in monocytes and immature DC by RT-PCR.Fig. 2 shows the result of NOD1 and NOD2 mRNAanalysis of monocytes and DC from the same donor(lanes 1 and 2), for DC from a separate donor (lane 4)and for the monocytic cell line THP-1 (lane 3). All cellsexpress high amounts of mRNA coding for NOD2, asseen in Fig. 2. NOD1 mRNA was also found to beexpressed in all cell types. However, the expression ofNOD1 in primary monocytes appears to be lower than inTHP-1 and corresponding DC (Fig. 2).
Cytokine production by primary humanmonocytes in response toNOD1 orNOD2 agonistsalone or in synergy with LPS
First, we tested the capacity of NOD1 (MtriDAP) andNOD2 agonists (MDP, MtriLYS) to stimulate primaryhuman monocytes for production of pro- as well as anti-inflammatory cytokines. As shown in Fig. 3, productionof pro-inflammatory cytokines such as IL-1b, IL-6 andTNF-a, as well as the chemokine IL-8, was observed afterstimulation with all tested muropeptides in a dose-dependent manner. However, no significant amounts ofthe anti-inflammatory cytokine IL-10 were found after
Fig. 2.Analysis ofNOD1andNOD2mRNA inhumanmonocytesand DC. Total RNA from CD14+ monocytes isolated by MACSfrom PBMC and CD1a+CD14– DC from the same donor (lane 1and 2, respectively), the myelomonocytic cell line THP-1 (lane3) and CD1a+CD14– DC from a separate donor (lane 4) wasobtained and analyzed for NOD1 and NOD2 mRNA levels bysemi-quantitative RT-PCR. GAPDH mRNA was used as anamplification control. Negative control usingwater as templateis shown in lane 5. One representative experiment out of threeperformed on different donors is shown.
Fig. 3. Cytokine production of human CD14+ monocytes after stimulation with NOD1 and NOD2 agonists. Various concentrationsofmuropeptides activating NOD1 (MtriDAP) or NOD2 (MDP, MtriLYS) were used to stimulate human CD14+ monocytes sorted fromPBMC. Release of TNF-a, IL-1b, IL-6 and IL-8 after 24 h of stimulation was measured by ELISA. A representative experiment out ofthree different donors is shown.
J�rg H. Fritz et al. Eur. J. Immunol. 2005. 35: 2459–24702462
f 2005 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.eji.de
stimulation of primary monocytes with various con-centrations of NOD1 and NOD2 agonists (data notshown). Interestingly, lower amounts of NOD2 agonists,MDP and MtriLYS, were needed to obtain cytokine andchemokine production, as compared to the NOD1agonist MtriDAP. At the maximal concentrations used(1000 nM of MDP, MtriLYS and MtriDAP corresponds to
492, 622 and 666 ng/mL, respectively), muropeptidesinduced up to 100–200 pg/mL TNF-a, 300 pg/mL IL-6,50 pg/mL of IL-1b, and 35 ng/mL of IL-8 (Fig. 3). Incomparison, 40 or 100 pg/mL purified LPS induceseveral nanograms per milliliter of all of these cytokines(Fig. 4).
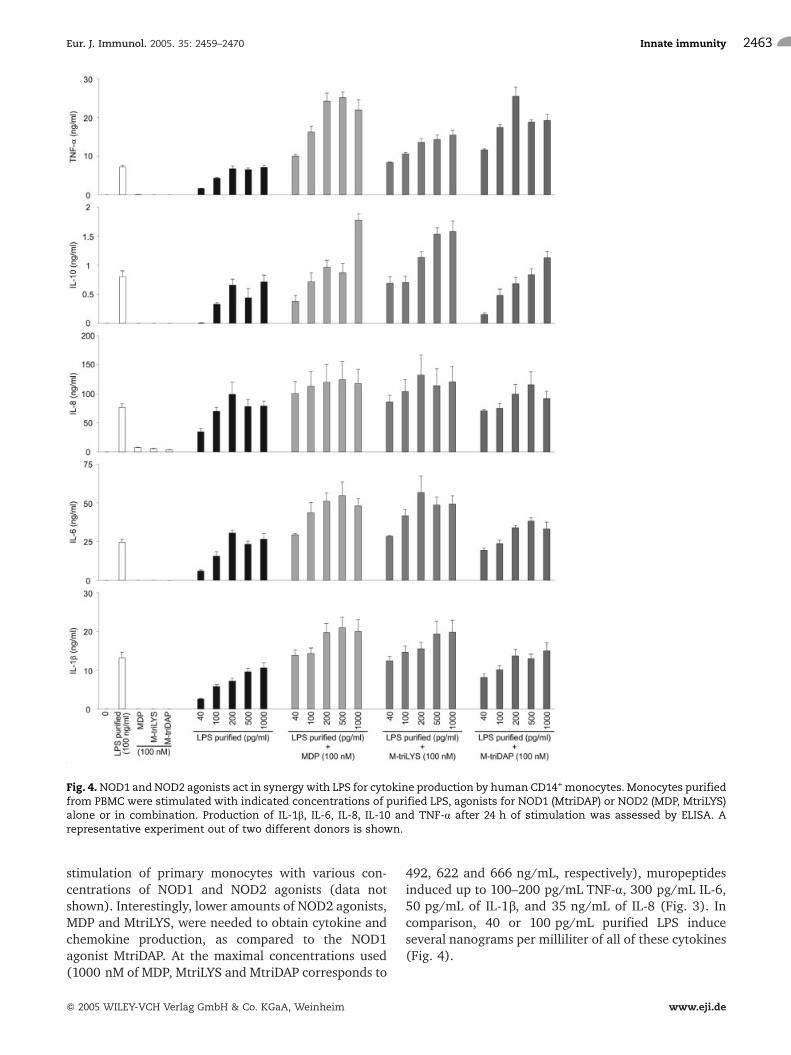
Fig. 4.NOD1 and NOD2 agonists act in synergy with LPS for cytokine production by human CD14+ monocytes. Monocytes purifiedfrom PBMC were stimulated with indicated concentrations of purified LPS, agonists for NOD1 (MtriDAP) or NOD2 (MDP, MtriLYS)alone or in combination. Production of IL-1b, IL-6, IL-8, IL-10 and TNF-a after 24 h of stimulation was assessed by ELISA. Arepresentative experiment out of two different donors is shown.
Eur. J. Immunol. 2005. 35: 2459–2470 Innate immunity 2463
f 2005 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.eji.de

Upon interaction of microbes with host cells such asmonocytes and DC, a multitude of diverse PAMP-PRMrecognition events takes place. Thus, we studied in detailthe effects of simultaneous stimulation with PG frag-ments such as NOD1 or NOD2 agonists and LPS for theproduction of pro- as well as anti-inflammatorycytokines. The duration of stimulation (24 h formonocytes and 40 h for dendritic cells) was chosenbased on preliminary studies that showed peak levels ofsecretion of all dosed mediators at these time points(data not shown). As can be seen in Fig. 4, NOD1(MtriDAP) as well as NOD2 agonists (MDP, MtriLYS) actin synergy with LPS and considerably amplify IL-1b, IL-6, IL-8, IL-10 and TNF-a production by primary humanmonocytes. This synergistic effect was seen withconcentrations of purified LPS ranging from 40 pg/mL up to 1 ng/mL. Interestingly, the simultaneouspresence of low amounts of both LPS and PG fragmentspermitted the induction of higher amounts of all dosedmediators (Fig. 4) than those obtained with a high doseof LPS alone (100 ng/mL). In general, the synergisticeffects examined with NOD2 agonists MDP and MtriLYSwere more potent than that with the NOD1 agonistMtriDAP.
The synergistic effects observed after simultaneousstimulation of primary human monocytes with PGfragments and LPSwas highest with the use of 40 pg/mLpurified LPS in combination with 100 nM of variousmuropeptides (Fig. 4). Co-application of higheramounts of muropeptides with purified LPS did notyield heightened cytokine or chemokine production(data not shown). Thus, these agonist concentrationswere chosen to study the donor dependency of thedescribed effects. By testing the reactivity of monocytesfrom eight different donors, a significant increase of thecytokines IL-1b, IL-6, IL-10 and TNF-a (Fig. 5) as well asthe chemokine IL-8 (data not shown) upon simulta-neous stimulation with MDP, MtriLYS or MtriDAP andLPS was observed. Moreover, no differences in thepattern of induction and the overall magnitude ofcytokine and chemokine production was observed bycomparing monocytes isolated via plastic adherence andmonocyte preparations enriched by CD14+ magneticcell sorting (data not shown).
Stimulation of primary human DC with NOD1 orNOD2 agonists alone or in synergy with LPS
Next, the capacity of NOD1 (MtriDAP) and NOD2agonists (MDP, MtriLYS) alone or in combination withLPS to stimulate primary human immature DC gener-ated from CD14+ monocytes was examined. Theproduction of pro- as well as anti-inflammatorycytokines was determined by ELISA, and the levels ofcostimulatory molecule expression and maturation of
DC was examined by FACS analysis. Similar tomonocytes, the production of cytokines of inflammationsuch as IL-6 and TNF-a as well as the chemokine IL-8was observed after stimulation with all tested muropep-tides in a dose-dependent manner, while no significantamounts of IL-1b and IL-10 were found (data not
Fig. 5. Statistical significance of the synergy of NOD1 andNOD2agonistswith LPS for cytokine production by CD14+monocytes.CD14+ monocytes purified from PBMC were stimulated with40 pg/mL purified LPS, alone or in combination with 100 nM ofagonists for NOD1 (MtriDAP) or NOD2 (MDP, MtriLYS). Produc-tion of IL-1b, IL-6, IL-10 and TNF-a after 24 h of stimulationwasassessed by ELISA. Values are mean � SEM of eight differentdonors. *p<0.05 using the Wilcoxon signed-rank test.
J�rg H. Fritz et al. Eur. J. Immunol. 2005. 35: 2459–24702464
f 2005 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.eji.de

shown). The pattern of induction and the overallmagnitude of cytokine and chemokine production ofDC stimulated with NOD1 and NOD2 agonists werecomparable to the observed effects with monocytes(data not shown). Moreover, DC stimulated with MDP,MtriLYS or MtriDAP produced IL-12p40, yielding levelssimilar to that observed with IL-6. Furthermore, none ofthe tested muropeptides induced IL-12p70 production(data not shown).
In addition, we studied the effects of simultaneousstimulation with PG fragments and LPS. NOD1 (Mtri-DAP) as well as NOD2 agonists (MDP, MtriLYS) acted in
synergy with LPS for production of IL-6, IL-8, IL-12p40and TNF-a by DC (Fig. 6). By comparing the results ofeight different donors, synergistic IL-6 and IL-8production was observed with concentrations of pur-ified LPS ranging from 40 pg/mL to 1 ng/mL incombination with 100 nM of the indicated muropep-tides. In contrast, a more restricted range of enhancedIL-12p40 and TNF-a production was observed aftersimultaneous stimulation with 100 nM NOD1 as well asNOD2 agonists (Fig. 6). No enhancement of IL-12p70release above levels with LPS alone was observed. Likefor monocytes, the overall synergistic effects examined
Fig. 6. NOD1 and NOD2 agonists act in synergy with LPS for cytokine production by human CD1a+CD14– DC. DC were stimulatedwith various concentrations of purified LPS, muropeptides activating NOD1 (MtriDAP) or NOD2 (MDP, MtriLYS) and indicatedcombinations. Release of IL-6, IL-8, IL-12p40 and TNF-a after 40 h of stimulationwas determined by ELISA. Values aremean� SEMof eight different donors. *p<0.05 using the Wilcoxon signed-rank test.
Eur. J. Immunol. 2005. 35: 2459–2470 Innate immunity 2465
f 2005 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.eji.de

for the NOD2 agonists, MDP and MtriLYS, were morepotent than that with the NOD1 agonist, MtriDAP.
In addition to cytokine production, we analyzedchanges in the maturation profile of stimulated DC byexamining cell-surface expression levels of HLA-DR,CD80, CD86, CD40, CD54 and CD83. As shown in Fig. 7,immature monocyte derived CD1a+CD14– DC expressedalready considerable levels of MHC class II (HLA-DR),CD40 and CD54, while cells were only weakly labeled forCD80 and CD86. Moreover, the immature phenotype ofthe DC populationwas demonstrated, since the obtainedcells did not stain positively for CD83, a knowmarker forthe maturation of DC [20]. After stimulation withMtriDAP, cell-surface expression levels for CD40 andCD54 were slightly enhanced, while no changes wereobserved for the cell-surface expression of HLA-DR,CD80, CD86 and CD83 (Fig. 7). Similar results wereobserved with MDP and MtriLYS, and by titrating theamounts of muropeptides ranging from 10 nM to 20 lM(data not shown). In contrast, stimulation of DC withLPS induced the up-regulation of all molecules in a dose-dependent manner. Simultaneous stimulation with the
NOD1 agonist MtriDAP and low levels of purified LPS(40 pg/mL) considerably enhanced the expression ofHLA-DR, CD86, CD54 and CD83 (Fig. 7). In addition,this combination of agonists induced slight cell-surfaceexpression of CD80 and CD40 compared to stimulationwith individual agonists alone. Similar results wereobtained by costimulation with a low dose of purifiedLPS with NOD2-activating agonists, MDP or MtriLYS(data not shown), indicating that NOD agonists incooperation with LPS amplify maturation of DC.
Discussion
Recent findings have demonstrated that PG devoid oflipopeptides or LTA contamination is not sensed byTLR2, placing NOD1 and NOD2 in the limelight forinnate immune sensing of PG in mammals [3]. Whileseveral reports have shown that the NOD2-activatingagonist, MDP, can stimulate myeloid cells for cytokineand chemokine production [10], the role of NOD1agonists remained largely unknown. Thus, we aimed at
Fig. 7. NOD1 and NOD2 agonists act in synergy with LPS for DC maturation. CD1a+CD14– DC were stimulated with variousconcentrations of purified LPS, muropeptides activating NOD1 (MtriDAP) or NOD2 (MDP, MtriLYS) and indicated combinations.After 40 h of stimulation cell-surface expression level of HLA-DR, CD80, CD86, CD40, CD54 and CD83 was determined by FACSanalysis using FITC-labeled antibodies specific for the indicatedmolecules. Stainingwith isotype control antibodies is representedin thin dashed lines, stained unstimulated cells are given as thin lines, while labeling of stimulated cells are specified by bold lines.One representative experiment out of five is shown.
J�rg H. Fritz et al. Eur. J. Immunol. 2005. 35: 2459–24702466
f 2005 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.eji.de

investigating the capacity of the NOD1 agonist MtriDAPto stimulate primary human myeloid cell subsets aloneor in combination with the TLR4 agonist LPS.
By testing the purity of several LPS preparations, weobserved that most of the commercially available LPScontain impurities stimulating signaling by TLR2, NOD1or NOD2. This implies that results of previous studies, inwhich the purity of the used agonists was not tested,might be at least partially interpreted as a cooperativeeffect between TLR and NOD signaling. It is shown herethat the possibilities of testing PRM agonists forcontaminations stimulating NOD1 or NOD2 are avail-able, ensuring a quick and sensitive measurement of thesubstance to be tested. Access to highly purified LPS andmuropeptides enabled us to study the stimulatorycapacity of muropeptides to stimulate primary humanmonocytes and DC, and to investigate the cooperativeeffect of these PG fragments with LPS.
We demonstrate here that NOD1 (MtriDAP) andNOD2 (MDP, MtriLYS) agonists stimulate primaryhuman monocytes for production of pro-inflammatorycytokines such as IL-1b, IL-6 and TNF-a as well as thechemokine IL-8, in a dose-dependent manner. More-over, the production of IL-6, IL-12p40, TNF-a and IL-8was observed after stimulation of DC with NOD1- andNOD2-activating agonists. However, no significantamounts of the anti-inflammatory cytokine IL-10 werefoundwithmonocytes and DC after stimulationwith anyof the tested muropeptides, indicating predominantly apro-inflammatory role of these agonists. While we founda significant induction of IL-1b release by monocytes, nointra- and extracellular IL-1b production by DC wasobserved after stimulation with NOD agonists. Ourresults are in agreement with a recent report showingthat MDP stimulates IL-1b release by human monocytes[21]. In contrast, wewere not able to confirm the findingthat highly purified LPS is unable to stimulate mono-cytes for IL-1b release [21], even though we ensured thelack of impurities stimulating TLR2, NOD1 or NOD2signaling. These findings therefore challenge the ideathat the inflammasome [22] is activated exclusively byMDP [21]. Alternatively, theremay be a distinct pathwaytriggered by LPS leading to IL-1b secretion. It remains tobe determined, however, if these conflicting results aredue to experimental differences, since diverse LPSpreparations and different cell isolation procedureswere used. Nevertheless, we show here for the first timethat the NOD1 agonist MtriDAP as well as the NOD2agonist MtriLYS induce IL-1b release by primary humanmonocytes, suggesting that bothmuropeptides likeMDP[21] can activate the inflammasome.
Some studies conducted with cell lines or wholehuman blood have reported that the NOD2 agonist MDPcan cooperate with LPS for TNF-a production [10, 11].We extend these studies by comparing the potential of
NOD1 aswell as NOD2 agonists to synergize with LPS forpro- and anti-inflammatory cytokine production byenriched primary myeloid cell populations. It is shownhere that NOD1 (MtriDAP) and NOD2 agonists (MDP,MtriLYS) act cooperatively with LPS, and considerablyamplify production of pro-inflammatory cytokines suchas IL-1b, IL-6, IL-8 and TNF-a, and also of the anti-inflammatory cytokine IL-10 by primary human mono-cytes.
Although a recent report showed that NOD1 agonistswere not able to synergize with LPS for TNF-a release bywhole human blood cells [11], we observed thatMtriDAP by itself and cooperatively with LPS stimulatescytokine and chemokine production in primary humanmyeloid cell subsets. These differences might be due tolower sensitivity in detection of cytokines and chemo-kines when using whole blood, or by the interference ofother cell types or of plasma components. Indeed, it hasbeen reported that mouse macrophages stimulated withiE-DAP secrete TNF-a and IL-6, and that this moleculewhen combined with LPS generated a larger IL-6response than LPS alone [16]. Furthermore, studieswith the human myelomonocytic cell line THP-1revealed that a DAP-containing lactoylpeptide, FK156(D-lactyl-L-Ala-c-D-Glu-meso-DAP-Gly), induced IL-8production by itself and in synergy with TLR2, TLR4or TLR9 ligands [17]. These two studies both considerthat the used compounds are recognized equally well bymouse and human NOD1. However, it must be notedthat species dependency of MDP sensing has beenreported [10, 23], opening up the possibility that speciesdifferences may exist for agonist specificity of bothNOD1 and NOD2.
Interestingly, all tested PG fragments by themselvesor in synergy with LPS were able to highly induce therelease of the chemokine IL-8 by monocytes and DC.IL-8 has been shown to recruit and activate neutrophilgranulocytes [24], and stimulate IL-2-activated naturalkiller cells [25] and lymphokine-activated killer cells[26]. Recently, a subset of CD8+ T lymphocytes has beendescribed, which is poised for immediate cytotoxicityand early recruitment by IL-8 into sites of innateimmune activation [27, 28]. Induction of IL-8 bymuropeptides, therefore, may define a bacterial-specificresponse to enhance clearance from the infected host.
To initiate immune responses, DC need to activatena�ve T lymphocytes, which only occurs if DC undergomaturation [29]. DC maturation increases the levels ofsurface expression of costimulatory and adhesionmolecules, such as CD40, CD80, CD86 and CD54.Furthermore, DC maturation influences the generationand transport of MHC-peptide complexes to the cellsurface and is paralleled by increased cell-surfaceexpression of CD83. We show here for the first timethat NOD1 as well as NOD2 agonists can stimulate DC
Eur. J. Immunol. 2005. 35: 2459–2470 Innate immunity 2467
f 2005 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.eji.de

for production of cytokines of inflammation such asTNF-a, IL-1b and IL-6, mediators known to positivelyinfluence the maturation process of DC [29]. However,muropeptides were not able to trigger full maturation ofDC. Only slight enhancement of CD40 and CD54 cell-surface expression was observed, indicating that thelevels of pro-inflammatory cytokines induced by mur-opeptides are not sufficient to yield full maturation ofDC.
This report is also the first demonstrating that therelease of IL-6, IL-8, IL-12p40 and TNF-a was sig-nificantly amplified after cooperative stimulation withLPS and muropeptides (MDP, MtriLYS, MtriDAP) by DC.In addition to cytokine production, we analyzed DCmaturation and observed that NOD1 and NOD2 agonistsamplify the maturation of DC, synergistically with lowdose LPS, indicating that cooperative engagement ofTLR4 and NOD1 or NOD2 can prime DC for the onset ofadaptive immunity.
Interestingly, the simultaneous presence of lowamounts of both LPS and PG fragments of both Gram-positive and Gram-negative bacteria amplifies theamount of cytokines secreted by monocytes and DCcompared to those obtained with a high dose of LPSalone (100 ng/mL). Moreover, beside the synergisticstimulation of monocytes and dendritic cells of MDPwith LPS, we show that MtriLYS, a PG breakdownproduct from Gram-positive bacteria, yields similarresults. During a bacterial infection, NOD1 and NOD2agonists, as well as LPS, are likely to be foundsimultaneously in the same tissue, which furtherreinforces the notion that the observed synergy mightbe of great physiological importance [15]. Sepsis isdefined as an infection with evidence of systemicinflammation [30]. The occurrence of Gram-negativesepsis has diminished over the years, whereas that ofGram-positive and polymicrobial infections has in-creased and accounted in the year 2000 for 30–50%and 25% of cases, respectively [31, 32]. Although sepsisinvolves several factors that interact in a long chain ofevents, pathogen recognition by multiple PAMP-PRMengagement, like that shown here, might constitute akey event for the onset of the resulting exacerbatedinflammatory response. Thus, the findings we show herehave important implications for the priming of adaptiveimmunity and also for naturally occurring pathologicalconditions such as septic shock and bacterial infection.
Material and methods
Reagents
Muramyl dipeptide (MDP) was purchased from EMD Bios-ciences (San Diego, CA). MtriLYS and MtriDAP were preparedas described [7]. All muropeptide preparations tested negative
for LPS contamination (detection limit: 0.1 EU/mL = 10 pg/mL) by the LAL (QCL-1000, BioWhittaker, Verviers, Belgium).Pam3CSK4 was purchased from EMC Microcollections (T�bin-gen, Germany). Various LPS preparations (Escherichia coliO111:B4) were obtained from Sigma-Aldrich (St. Louis, MO),Invivogen (San Diego, CA) and ALEXIS Corp. (Lausen,Switzerland). Highly purified LPS (from E. coli O119) wasprepared according to [33].
Reporter assay for NF-jB activation
Human embryonic kidney HEK293T cells (ATCC, Manassas,VA) were cultured in Dulbecco's modified Eagle's mediumsupplemented with 2 mM L-glutamine, 1 mM sodium pyr-uvate, 1� nonessential amino acids, (all purchased fromInvitrogen, Carlsbad, CA) and 5% fetal calf serum (FCS;Hyclone, Logan, UT). HEK293T cells were seeded into 24-wellplates at a density of 105 cells/mL and transfectionwith variousexpression plasmids was carried out as described previously[7]. Briefly, HEK293T cells were transfected overnight using1% of FuGENE 6 transfection reagent (Roche Diagnostics,Germany) with 75 ng reporter plasmid Igj-luciferase alone orplus the following vectors: 1 ng NOD1 or NOD2, 50 ng TLR2.The pcDNA3.1 vector was used to balance the transfected DNAconcentration. At the time of transfection, muropeptides orTLR2 agonist Pam3CSK4 were added to the cell culturemedium, and the synergistic NF-jB-dependent luciferaseactivation was examined following 24 h of co-incubation.NF-jB-dependent luciferase assays were performed in tripli-cates and data represent at least three independent experi-ments. Data show mean � SEM.
RT-PCR analysis
Total RNA was prepared using the RNeasy Mini Kit (Qiagen,Valencia, CA). Purified RNA was reverse-transcribed withSuperscript II RNase H (Invitrogen) according to themanufacturer's protocol. The generated cDNA was amplifiedby using primers for human NOD1 (forward-primer: 50-GTAAAGGTGCTAAGCGAAGA-30, reverse-primer: 50-TCTGAT-TCTGGATAAGCCAT-30), human NOD2 (forward-primer: 50-CTCAGTCTCGCTTCCTCAGTAC-30, reverse-primer: 50-GGTTA-ACAGGACACGGTCAGG-30) and glyceraldehyde-3-phosphate-dehydrogenase (GAPDH, forward-primer: 50-TGAAGGTCG-GAGTCAACGGATTTGGT-30, reverse-primer: 50-CATGTGGGC-CATGAGGTCCACCAC-30). The cDNA samples were heatdenatured (95�C for 5 min) and then amplified with 35cycles, each comprising successive incubations at 95�C for40 s, 55�C for 40 s and 72�C for 60 s. A further extension stepwas done at 72�C for 5 min. The amplicons (GAPDH: 983 bp,NOD1: 490 bp, NOD2: 450 bp) were size fractionated onto a2% agarose gel and stained with ethidium bromide.
PBMC and monocyte isolation, dendritic cell culture
Peripheral blood mononuclear cells (PBMC) were preparedfrom fresh blood samples of healthy donors drawn on citrate/phosphate/dextrose (Etablissement Fran�ais du Sang, Paris,France). Blood was diluted 1:2 in RPMI 1640 Glutamaxmedium (BioWhittaker) and centrifuged over Ficoll (MSL;
J�rg H. Fritz et al. Eur. J. Immunol. 2005. 35: 2459–24702468
f 2005 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.eji.de

Eurobio, Les Ulis, France) for 20 min at 15�C and 600�g.Human monocytes were selected from PBMC by adherence.PBMCwere plated at 6�106 cells/mL and allowed to adhere for1 h at 37�C in a 5% CO2 air incubator in a humidifiedatmosphere. Nonadherent cells were removed, adherent cellswere washed with RPMI and cultured in RPMI supplementedwith antibiotics (100 IU/mL penicillin; 100 g/mL streptomy-cin) and 10% FCS (Hyclone). In some experiments, monocyteswere isolated using anti-CD14 MACS beads (Miltenyi Biotec,Bergisch Gladbach, Germany) according to the manufacturer'sprotocol. Purity of monocytes was determined by FACSanalysis, routinely yielding >98% purity of CD14+ cells.
For generation of immature DC, CD14+ monocytes wereseeded in 6-well plates at 2�106 cells/well in RPMI 1640Glutamax medium supplemented with 1 mM sodium pyru-vate, 1� nonessential amino acids, 0.5 mM 2-mercaptoethanol(all purchased from Invitrogen), 10% FCS (Hyclone), 800 U/mL recombinant human GM-CSF (rhu-GM-CSF, Leukine,Aesca, Traiskirchen, Austria) and 1000 U/mL IL-4 (rhu-IL-4,Novartis, Basel, Switzerland; kind gift of Dr ChristianDemeure, Institut Pasteur). At day 2 of culture, cells weresupplemented with fresh medium (�1 mL) supplementedwith 1600 U/mL rhu-GM-CSF. On day 7, cells were harvestedand washed with culture medium without cytokines, andseeded in 24-well plates at 4�105 cells/well. Purity of DC wasdetermined by FACS analysis, routinely yielding >95% purityof CD14– CD1a+cells.
Monocytes and DCwere stimulatedwith various amounts ofrepurified LPS and/or muropeptides (MDP,MtriLYS, MtriDAP)for 24 or 40 h, respectively, before collection of supernatants.
ELISA
The concentration in the culture supernatants of IL-1b, IL-6, IL-8, IL-10, IL-12p40, IL-12p70 and TNF-a was determined byELISA (DuoSet; R&D Systems, Minneapolis, MN) as specifiedby the manufacturer.
Flow cytometry
After stimulation, cells were harvested and resuspended in ice-cold PBS supplemented with 0.1% bovine serum albumin(BSA) and 0.2% NaN3 (both purchased from Sigma-Aldrich).To block unspecific staining, human IgG was added (Beriglo-bin, Behring, Marburg, Germany) and samples were incubatedfor 15 min on ice. Subsequently, FITC-labeled antibodiesspecific for CD14, CD40, CD54, CD80, CD83, CD86 and HLA-DR, PE-labeled anti-CD1a antibody, or isotype-matchingcontrol antibodies were added and incubated on ice for30 min (all antibodies were purchased from Becton Dickinson,Franklin Lakes, NJ). Cells were washed extensively andresuspended in PBS/0.1% BSA and analyzed by FACS(FACScan, Becton Dickinson). Dead cells were excluded fromanalysis by propidium iodide staining. Datawas analyzed usingWinMDI 2.8 software.
Statistical analysis
The results are given as mean � SEM. Statistical analysis wasperformed using the nonparametric Wilcoxon signed-rank test
and the Statview software (SAS, Cary, NC); p<0.05 wasconsidered significant.
Acknowledgements: We thank Christian Demeure forproviding rhu-IL-4. We are grateful to Lionel LeBourhis,Ronan Kapetanovic and Thomas Kufer for helpfulcomments and critically reading the manuscript. Wethank Muguette J�hanno for help with HEK cellexperiments and Martine Parmier for excellent techni-cal help and plasmid preparations. J�rg H. Fritz wassupported by the Association Fran�ois Aupetit and bythe 'Programme Transversal de Recherche 153' from theInstitut Pasteur.
References
1 Akira, S. and Takeda, K., Toll-like receptor signalling. Nat. Rev. Immunol.2004. 4: 499–511.
2 Ting, J. P. and Davis, B. K., CATERPILLER: A novel gene family important inimmunity, cell death, and diseases. Annu. Rev. Immunol. 2005. 23: 387–414.
3 Girardin, S. E. and Philpott, D. J., Mini-review: The role of peptidoglycanrecognition in innate immunity. Eur. J. Immunol. 2004. 34: 1777–1782.
4 Takeuchi, O., Hoshino, K., Kawai, T., Sanjo, H., Takada, H., Ogawa, T.,Takeda, K. and Akira, S., Differential roles of TLR2 and TLR4 in recognitionof gram-negative and gram-positive bacterial cell wall components.Immunity 1999. 11: 443–451.
5 Travassos, L. H., Girardin, S. E., Philpott, D. J., Blanot, D., Nahori, M. A.,Werts, C. and Boneca, I. G., Toll-like receptor 2-dependent bacterial sensingdoes not occur via peptidoglycan recognition. EMBO Rep. 2004. 5:1000–1006.
6 Schleifer, K. H. and Kandler, O., Peptidoglycan types of bacterial cell wallsand their taxonomic implications. Bacteriol. Rev. 1972. 36: 407–477.
7 Girardin, S. E., Travassos, L. H., Herve, M., Blanot, D., Boneca, I. G.,Philpott, D. J., Sansonetti, P. J. and Mengin-Lecreulx, D., Peptidoglycanmolecular requirements allowing detection by Nod1 and Nod2. J. Biol.Chem. 2003. 278: 41702–41708.
8 Girardin, S. E., Boneca, I. G., Viala, J., Chamaillard, M., Labigne, A.,Thomas, G., Philpott, D. J. and Sansonetti, P. J., Nod2 is a general sensorof peptidoglycan through muramyl dipeptide (MDP) detection. J. Biol.Chem. 2003. 278: 8869–8872.
9 Wolfert, M. A., Murray, T. F., Boons, G. J. and Moore, J. N., The origin ofthe synergistic effect of muramyl dipeptide with endotoxin and peptidogly-can. J. Biol. Chem. 2002. 277: 39179–39186.
10 Takada, H., Yokoyama, S. and Yang, S., Enhancement of endotoxin activityby muramyldipeptide. J. Endotoxin Res. 2002. 8: 337–342.
11 Traub, S., Kubasch, N., Morath, S., Kresse, M., Hartung, T., Schmidt, R.R. and Hermann, C., Structural requirements of synthetic muropeptides tosynergize with lipopolysaccharide in cytokine induction. J. Biol. Chem. 2004.279: 8694–8700.
12 Parant, M., Parant, F., Vinit, M. A., Jupin, C., Noso, Y. and Chedid, L.,Priming effect of muramyl peptides for induction by lipopolysaccharide oftumor necrosis factor production in mice. J. Leukoc. Biol. 1990. 47: 164–169.
13 Takada, H. and Galanos, C., Enhancement of endotoxin lethality andgeneration of anaphylactoid reactions by lipopolysaccharides in muramyl-dipeptide-treated mice. Infect. Immun. 1987. 55: 409–413.
14 Parant, M. A., Pouillart, P., Le Contel, C., Parant, F. J., Chedid, L. A. andBahr, G. M., Selective modulation of lipopolysaccharide-induced death andcytokine production by various muramyl peptides. Infect. Immun. 1995. 63:110–115.
15 Annane, D., Bellissant, E. and Cavaillon, J. M., Septic shock. Lancet 2005.365: 63–78.
Eur. J. Immunol. 2005. 35: 2459–2470 Innate immunity 2469
f 2005 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.eji.de

16 Chamaillard, M., Hashimoto, M., Horie, Y., Masumoto, J., Qiu, S., Saab,L., Ogura, Y. et al., An essential role for NOD1 in host recognition ofbacterial peptidoglycan containing diaminopimelic acid. Nat. Immunol.2003. 4: 702–707.
17 Uehara, A., Yang, S., Fujimoto, Y., Fukase, K., Kusumoto, S., Shibata, K.,Sugawara, S. and Takada, H.,Muramyldipeptide and diaminopimelic acid-containing desmuramylpeptides in combination with chemically synthe-sized Toll-like receptor agonists synergistically induced production ofinterleukin-8 in a NOD2- and NOD1-dependent manner, respectively, inhuman monocytic cells in culture. Cell. Microbiol 2005. 7: 53–61.
18 Ogura, Y., Inohara, N., Benito, A., Chen, F. F., Yamaoka, S. andNunez, G.,Nod2, a Nod1/Apaf-1 family member that is restricted to monocytes andactivates NF-kappaB. J. Biol. Chem. 2001. 276: 4812–4818.
19 Gutierrez, O., Pipaon, C., Inohara, N., Fontalba, A., Ogura, Y., Prosper,F., Nunez, G. and Fernandez-Luna, J. L., Induction of Nod2 inmyelomonocytic and intestinal epithelial cells via nuclear factor-kappa Bactivation. J. Biol. Chem. 2002. 277: 41701–41705.
20 Kalinski, P., Schuitemaker, J. H., Hilkens, C. M. and Kapsenberg, M. L.,Prostaglandin E2 induces the final maturation of IL-12-deficientCD1a+CD83+ dendritic cells: the levels of IL-12 are determined duringthe final dendritic cell maturation and are resistant to further modulation. J.Immunol. 1998. 161: 2804–2809.
21 Martinon, F., Agostini, L., Meylan, E. and Tschopp, J., Identification ofbacterial muramyl dipeptide as activator of the NALP3/cryopyrin inflam-masome. Curr. Biol. 2004. 14: 1929–1934.
22 Martinon, F., Burns, K. and Tschopp, J., The inflammasome: a molecularplatform triggering activation of inflammatory caspases and processing ofproIL-beta. Mol. Cell 2002. 10: 417–426.
23 Nagao, S., Akagawa, K. S., Yamada, K., Yagawa, K., Tokunaga, T. andKotani, S., Lack of response of murine peritoneal macrophages to in vitroactivation bymuramyl dipeptide (MDP). I. Macrophage activation byMDP isspecies dependent. Microbiol. Immunol. 1990. 34: 323–335.
24 Zeilhofer, H. U. and Schorr, W., Role of interleukin-8 in neutrophilsignaling. Curr. Opin. Hematol. 2000. 7: 178–182.
25 Rah, S. Y., Park, K. H., Han, M. K., Im, M. J. and Kim, U. H., Activation ofCD38 by interleukin-8 signaling regulates intracellular Ca2+ level andmotility of lymphokine-activated killer cells. J. Biol. Chem. 2005. 280:2888–2895.
26 Inngjerdingen, M., Al-Aoukaty, A., Damaj, B. and Maghazachi, A. A.,Differential utilization of cyclic ADP-ribose pathway by chemokines toinduce the mobilization of intracellular calcium in NK cells. Biochem.Biophys. Res. Commun. 1999. 262: 467–472.
27 Hess, C., Means, T. K., Autissier, P., Woodberry, T., Altfeld, M., Addo, M.M., Frahm, N. et al., IL-8 responsiveness defines a subset of CD8 T cellspoised to kill. Blood 2004. 104: 3463–3471.
28 Takata, H., Tomiyama, H., Fujiwara, M., Kobayashi, N. and Takiguchi,M., Cutting edge: expression of chemokine receptor CXCR1 on humaneffector CD8+ T cells. J. Immunol. 2004. 173: 2231–2235.
29 Guermonprez, P., Valladeau, J., Zitvogel, L., Thery, C. and Amigorena,S., Antigen presentation and T cell stimulation by dendritic cells. Annu. Rev.Immunol. 2002. 20: 621–667.
30 Levy,M.M., Fink, M. P., Marshall, J. C., Abraham, E., Angus, D., Cook, D.,Cohen, J., Opal, S. M., Vincent, J. L. and Ramsay, G., 2001 SCCM/ESICM/ACCP/ATS/SIS International Sepsis Definitions Conference. Crit. Care Med.2003. 31: 1250–1256.
31 Martin, G. S., Mannino, D. M., Eaton, S. and Moss, M., The epidemiologyof sepsis in the United States from 1979 through 2000. N. Engl. J. Med. 2003.348: 1546–1554.
32 Moss, M. and Martin, G. S., A global perspective on the epidemiology ofsepsis. Intensive Care Med. 2004. 30: 527–529.
33 Caroff, M., Tirsoaga, A., El Hamidi, A., Hussein, A., Novikov, A., Adib-Conquy, M., Werts, C. and Cavaillon, J.-M., New methods for theextraction, purification and quick characterization of bacterial endotoxins.8th Biennial Conference of the International Endotoxin Society, Kyoto,Japan, November 15–18, 2004. Abstracts. J. Endotoxin Res. 2004. 10:287–388.
J�rg H. Fritz et al. Eur. J. Immunol. 2005. 35: 2459–24702470
f 2005 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.eji.de
Copyright © 2022 FDOKUMEN




















