
Synergistic Anti-Cancer Effects of Grape Seed Extract and Conventional Cytotoxic Agent Doxorubicin...
12
Breast Cancer Research and Treatment 85: 1–12, 2004. © 2004 Kluwer Academic Publishers. Printed in the Netherlands. Report Synergistic anti-cancer effects of grape seed extract and conventional cytotoxic agent doxorubicin against human breast carcinoma cells Girish Sharma 1,∗ , Anil K. Tyagi 1,∗ , Rana P. Singh 1 , Daniel C.F. Chan 2,3 , and Rajesh Agarwal 1,3 1 Department of Pharmaceutical Sciences, School of Pharmacy, 2 Department of Medicine, Division of Medical Oncology; 3 University of Colorado Cancer Center, University of Colorado Health Sciences Center, Denver, CO, USA Key words: apoptosis, breast cancer, cell-cycle, doxorubicin, grape seed extract Summary With an approach to enhance the efficacy of chemotherapy agents against breast cancer treatment, here, we inves- tigated the anti-cancer effects of grape seed extract (GSE) and doxorubicin (Dox), either alone or in combination, in estrogen receptor-positive MCF-7 and receptor-negative MDA-MB468 human breast carcinoma cells. GSE (25– 200 µg/ml) treatment of cells resulted in 16–72% growth inhibition and 9–33% cell death, in a dose- and a time- dependent manner. In other studies, Dox (10–100nM) treatment showed 23–96% growth inhibition and 10–55% cell death. Based on these results, several combinations of GSE (25–100 µg/ml) with Dox (10–75 nM) were next assessed for their synergistic, additive and/or antagonistic efficacy towards cell growth inhibition and death. In both MCF-7 and MDA-MB468 cells, a combination of 100 µg/ml GSE with 25–75 nM Dox treatment for 48 h showed a strong synergistic effect [combination index (CI) < 0.5] in cell growth inhibition, but mostly an additive effect (CI ∼ 1) in cell death. In cell-cycle progression studies, GSE plus Dox combination resulted in a moderate increase in G1 arrest in MCF-7 cells compared to each agent alone. GSE plus Dox combination showed a very strong and significant G1 arrest in MDA-MB468 cells when compared with Dox alone, however, it was less than that observed with GSE alone. In quantitative apoptosis studies, GSE and Dox alone and in combination showed comparable apoptotic death of MCF-7 cells, however, a combination of the two was inhibitory to Dox induced apoptosis in MDA-MB468 cells. This was further confirmed in another estrogen receptor-negative MDA-MB231 cell line, in which GSE and Dox combination strongly inhibited cell growth but did not show any increase in apoptotic cell death caused by Dox. Together, these results suggest a strong possibility of synergistic efficacy of GSE and Dox combination for breast cancer treatment, independent of estrogen receptor status of the cancer cell. Introduction Breast cancer is the most commonly diagnosed invasive non-skin malignancy and second leading cause of cancer-related deaths in the US women [1]. Epidemiological data suggest that environmental and behavioral factors are more important than ge- netic factors in determining overall cancer frequency among populations [2]. In addition, both epidemiolo- gical and laboratory studies show that diet plays a major role in the pathogenesis as well as in the pro- These authors contributed equally to this work. motion, progression and metastasis of breast cancer. Although there are many therapeutic strategies in- cluding chemotherapy to treat clinical breast cancer, high systemic toxicity and drug resistance limit the successful outcomes in most cases. Accordingly, sev- eral new strategies are being developed to control and treat clinical breast cancer. One such approach could be a combination of non-toxic effective phytochem- icals with chemotherapy agents, which could enhance the efficacy together with reducing toxicity to normal tissues. In present era, the notion has evolved that natural products frequently exert a valuable role in broaden-
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Synergistic Anti-Cancer Effects of Grape Seed Extract and Conventional Cytotoxic Agent Doxorubicin...

Breast Cancer Research and Treatment 85: 1–12, 2004.© 2004 Kluwer Academic Publishers. Printed in the Netherlands.
Report
Synergistic anti-cancer effects of grape seed extract and conventionalcytotoxic agent doxorubicin against human breast carcinoma cells
Girish Sharma1,∗, Anil K. Tyagi1,∗, Rana P. Singh1, Daniel C.F. Chan2,3, and Rajesh Agarwal1,3
1Department of Pharmaceutical Sciences, School of Pharmacy, 2Department of Medicine, Division ofMedical Oncology; 3University of Colorado Cancer Center, University of Colorado Health Sciences Center,Denver, CO, USA
Key words: apoptosis, breast cancer, cell-cycle, doxorubicin, grape seed extract
Summary
With an approach to enhance the efficacy of chemotherapy agents against breast cancer treatment, here, we inves-tigated the anti-cancer effects of grape seed extract (GSE) and doxorubicin (Dox), either alone or in combination,in estrogen receptor-positive MCF-7 and receptor-negative MDA-MB468 human breast carcinoma cells. GSE (25–200 µg/ml) treatment of cells resulted in 16–72% growth inhibition and 9–33% cell death, in a dose- and a time-dependent manner. In other studies, Dox (10–100 nM) treatment showed 23–96% growth inhibition and 10–55%cell death. Based on these results, several combinations of GSE (25–100µg/ml) with Dox (10–75 nM) were nextassessed for their synergistic, additive and/or antagonistic efficacy towards cell growth inhibition and death. Inboth MCF-7 and MDA-MB468 cells, a combination of 100 µg/ml GSE with 25–75 nM Dox treatment for 48 hshowed a strong synergistic effect [combination index (CI) < 0.5] in cell growth inhibition, but mostly an additiveeffect (CI ∼ 1) in cell death. In cell-cycle progression studies, GSE plus Dox combination resulted in a moderateincrease in G1 arrest in MCF-7 cells compared to each agent alone. GSE plus Dox combination showed a verystrong and significant G1 arrest in MDA-MB468 cells when compared with Dox alone, however, it was less thanthat observed with GSE alone. In quantitative apoptosis studies, GSE and Dox alone and in combination showedcomparable apoptotic death of MCF-7 cells, however, a combination of the two was inhibitory to Dox inducedapoptosis in MDA-MB468 cells. This was further confirmed in another estrogen receptor-negative MDA-MB231cell line, in which GSE and Dox combination strongly inhibited cell growth but did not show any increase inapoptotic cell death caused by Dox. Together, these results suggest a strong possibility of synergistic efficacy ofGSE and Dox combination for breast cancer treatment, independent of estrogen receptor status of the cancer cell.
Introduction
Breast cancer is the most commonly diagnosedinvasive non-skin malignancy and second leadingcause of cancer-related deaths in the US women[1]. Epidemiological data suggest that environmentaland behavioral factors are more important than ge-netic factors in determining overall cancer frequencyamong populations [2]. In addition, both epidemiolo-gical and laboratory studies show that diet plays amajor role in the pathogenesis as well as in the pro-
�These authors contributed equally to this work.
motion, progression and metastasis of breast cancer.Although there are many therapeutic strategies in-cluding chemotherapy to treat clinical breast cancer,high systemic toxicity and drug resistance limit thesuccessful outcomes in most cases. Accordingly, sev-eral new strategies are being developed to control andtreat clinical breast cancer. One such approach couldbe a combination of non-toxic effective phytochem-icals with chemotherapy agents, which could enhancethe efficacy together with reducing toxicity to normaltissues.
In present era, the notion has evolved that naturalproducts frequently exert a valuable role in broaden-

2 G Sharma et al.
ing the scope of disease intervention strategies usedby drug designers. Fruits, vegetables, common bever-ages, several herbs and plants with diversified phar-macological properties are known to be rich sourcesof micro-chemicals or ’non-nutritive minor dietarychemical constituents’ that may have potential for theprevention and/or treatment of several human malig-nancies including breast cancer [3–10]. Grapes (Vitisvinifera) are one of the most widely consumed fruits inthe world. Various naturally occurring flavonoid anti-oxidants are known to be present in grape seeds com-monly known as procyanidins [11, 12]. Commercialpreparations of grape seed polyphenols are marketedin the United States as ‘grape seed extract’ (GSE),with 95% standardized procyanidins as a dietary sup-plement because of its health benefits, particularlythe strong antioxidant activity. In addition to grapeseed, procyanidins are a diverse group of polyphen-olics widely distributed in fruits and vegetables [13].Recent studies have shown that procyanidins possessanti-inflammatory, antiarthritic, and antiallergic activ-ities and prevent heart attack and skin aging [14–16].GSE has been shown to act as a potent scavenger ofboth reactive oxygen and nitrogen species. Further,it has also been shown that GSE significantly inhib-its the activities of free-radical producing enzymes inbiological systems [17]. Wine consumption has beenreported to have many beneficial health effects; winemay also be a source of procyanidins [18, 19]. Withregard to its anti-cancer effects, recent cell culturestudies have shown that treatment of human breastcarcinoma MCF-7, human lung cancer A-427, andhuman gastric adenocarcinoma CRL-1739 cells withGSE results in a dose- and a time-dependent inhibitionof cell growth [20]. In animal models, oral feeding of1% (w/w) GSE in the diet has been shown to inhibitACP mutation-associated intestinal adenoma forma-tion in MIN mice [21]. More recent studies have alsoshown that tropical application of GSE significantlyprevents tumor promotion in mouse skin [22, 23]. Re-garding epidemiology, a case-control study showedthat increased consumption of grapes is associatedwith reduced cancer risk [24].
Several cytotoxic chemotherapeutic agents havebeen reported to be effective, alone or in combi-nation in the treatment of different cancers. Studieshave also shown that breast cancer is sensitive to mul-tiple cytotoxic compounds. It has been demonstratedthat combination chemotherapy produces higher over-all and complete remission rates than sequentialsingle agents, and that Dox-containing combinations
are more effective than other regimens [25]. To-gether, based on above studies and rationale, weassessed the effect of GSE and Dox either alone orin combination on estrogen-dependent (MCF-7) andestrogen-independent (MDA-MB468 and/or MDA-MB231) human breast carcinoma cell growth, death,cell-cycle progression and apoptosis. The data ob-tained show strong synergism between GSE and Doxin both cell growth inhibition and death independentof functional estrogen receptor level in human breastcancer cells.
Material and methods
Cell lines and other reagents. Human breast car-cinoma cell lines MCF-7, MDA-MB468 and MDA-MB231 were obtained from the American TypeCulture Collection (Manassas, VA). Cells were grownin Dulbecco’s modified Eagle medium (DMEM;Gibco BRL, Grand Island, NY) with 10% fetal bovineserum (Hylone, Logan, UT) and 1% penicillin–streptomycin (Gibco BRL) at standard culture condi-tions. The GSE preparation used was obtained fromTraco Labs (Champaign, IL). For all studies, GSEwas dissolved in dimethyl sulfoxide (DMSO) as a200 mg/ml stock solution and diluted as desired di-rectly in the medium. Unless specified otherwise,the final concentration of DMSO in the culture me-dium during GSE treatments did not exceed 0.1%(v/v), and therefore, the same concentration of DMSOwas present in control dishes. Dox was from CetusOncology (Emeryville, CA) and dissolved in double-distilled water.
Cell growth and viability assays. Cells were plated at5000 cells/cm2 in 60 mm dishes under the culture con-ditions described above. After 24 h, cells were fed withfresh medium and treated with either 0.1% DMSO(v/v) alone (control) or different concentrations (25,50, 75, 100, and 200 µg/ml) of GSE in DMSO or (10,25, 50, 75 and 100 nM) Dox. After 48 and 72 h of thesetreatments, cells were collected by trypsinization, andcounted in duplicate with a hemocytometer. Trypanblue dye exclusion was used to determine viable anddead cells.
Combination index analysis. The CI-isobologramby Chou and Talalay [26] was used to analyze thedrug combination assays. Briefly, variable ratios ofdrug concentrations (25–100µg/ml GSE and 10–75 nM Dox) were used in several different combi-

Efficacy of GSE plus doxorubicin in breast cancer 3
nations for the treatment of both MCF-7 and MDA-MB468 cells for 48 h. Thereafter, cells were collect-ed by trypsinization, and counted in duplicate witha hemocytometer. Trypan blue dye exclusion wasused to determine viable and dead cells. The cellgrowth and death data obtained were analyzed us-ing mutually exclusive equations to determine the CI[26]. Each CI was calculated from the mean affectedfraction at each drug ratio concentration (triplicate).CI > 1, CI = 1, and CI < 1 indicate antagonistic, ad-ditive and synergistic effect, respectively. Cell growthand viability was also assessed in MDA-MB231cells with selected GSE and Dox doses and theircombination.
Cell-cycle analysis. MCF-7 and MDA-MB468 cellswere treated either with DMSO alone or GSE(100 µg/ml) and Dox (25 or 50 nM) either alone or incombination. After 48 h of these treatments, cells weretrypsinized and cell pellets were collected. Approxi-mately 0.5 × 106 cells in 0.5 ml of saponin/propidiumiodide (PI) solution (0.3% saponin [w/v], 25 µg/ml PI[w/v], 0.1 mM EDTA, and 10 µg/ml RNase [w/v] inPBS) were left at 4◦C for 24 h in the dark. Cell-cycledistribution was then analyzed by flow cytometry us-ing the FACS Analysis Core Services of the Universityof Colorado Cancer Center, Denver.
Quantitative apoptosis analyses. MCF-7, MDA-MB468 and MDA-MB231 cells were grown in 60-mmdishes and treated with either DMSO alone or GSE(100 µg/ml) and Dox (25 or 50 nM) either alone orin combination. After 48 h of treatments, cells werecollected followed by 7-amino actinomycin D (7AAD,Calbiochem, USA) staining and flow cytometric anal-ysis [27]. Briefly, a solution of 7AAD (dissolved inacetone at 1 mg/ml and diluted five times in PBS) wasadded to cell suspension in PBS (∼20 µg 7AAD/106
cells in 1.1 ml) and mixed well. The cells were stainedfor 20 min at 4◦C while protected from light and thenwere pelleted by centrifugation. Scattergrams weregenerated by combining forward light scatter with7AAD fluorescence, and regions were drawn aroundclear-cut populations having negative, dim, and brightfluorescence. Typical FACS scattergrams show threeregions defined by 7AAD staining: 7AAD-negative(live cells), 7AAD-dim (apoptotic cells), and 7AAD-bright (late-apoptotic or dead cells). Cells within theseregions were sorted, first in Enrich mode and sub-sequently in normal-R mode.
Statistical analysis. The data were analyzed usingthe Jandel Scientific SigmaStat 2.03 software. Stu-dent’s t-test was employed to assess the statisticalsignificance of difference between control and differ-ent treatment groups as well as among different drugtreatment groups. A statistically significant differencewas considered to be present at P < 0.05.
Results
Effect of GSE and Dox alone on MCF-7and MDA-MB468 cell growth and death
As shown in Figure 1, GSE treatment of cellsresulted in a moderate to highly significant cellgrowth inhibition and cell death in a dose- and atime-dependent manner. In case of MCF-7 cells,25–200µg/ml doses of GSE caused 16–69%(P < 0.05–0.01) cell growth inhibition (Figure 1(A))and 9–33% (P < 0.05–0.001) cell death (Figure 1(C))following 48 and 72 h of treatments. Similar cellgrowth inhibitory and cell death effects of GSE werealso evident in case of MDA-MB468 cells account-ing for 18–72% (P < 0.05–0.01) growth inhibition(Figure 1(B)) and 9–30% (P < 0.05–0.01) cell death(Figure 1(D)).
In other studies assessing the effect of Dox,treatment of MCF-7 cells with 10, 25, 50, 75and 100 nM Dox for 48 and 72 h resulted in23–92% (P < 0.01–0.001) cell growth inhibition(Figure 2(A)) and 10–55% (P < 0.01–0.001) celldeath (Figure 2(C)) in a dose- and a time-dependentmanner. Similar Dox treatments of MDA-MB-468cells resulted in 26–96% (P < 0.05–0.001) growth in-hibition (Figure 2(B)) and 10–42% (P < 0.01–0.001)cell death (Figure 2(D)). Together, the data shownin Figures 1 and 2 clearly demonstrate the efficacyof both GSE and Dox in human breast carcinomaMCF-7 and MDA-MB468 cell growth inhibition andcell death independent of functional estrogen receptorstatus.
Effect of GSE–Dox combination on MCF-7and MDA-MB468 cell growth and death
Based on the data showing efficacy of GSE and Doxin breast carcinoma cells, next we assessed their ef-fects in combinations on MCF-7 and MDA-MB468cell growth and death, and analyzed the data for apossible synergistic, additive or antagonistic effect.

4 G Sharma et al.
Figure 1. Growth inhibitory effect of GSE on human breast carcinoma cells. For studies assessing the effect of GSE on exponentially growingMCF-7 and MDA-MB468 cell growth (A and B) and cell death (C and D), respectively, 0.5 × 105 cells were plated in 60 mm dishes and 24 hafter plating, cultures were fed with fresh medium with vehicle (DMSO 0.1%, v/v) alone or GSE (25–200 µg/ml) in medium. After 48 and 72 hof these treatments, the total numbers of cells were counted; including both viable and dead cells. Cell viability was determined by trypan bluedye exclusion method. The cell growth and cell death data shown are the mean ± S.E. of two independent plates; each sample was counted induplicate. The representative data shown in the figures were reproducible in an additional independent experiment. GSE, grape seed extract;∗P < 0.05; #P < 0.01; $P < 0.001.
As shown in Figure 3(A), in MCF-7 cells, a com-bination of 100 µg/ml GSE and 25 nM Dox for 48 hresulted in 88% growth inhibition as compared toeither agent alone showing 47% (GSE) and 43% (Dox)growth inhibition. In case of cell death, however,similar GSE plus Dox combination showed 31% celldeath as compared to 27 and 21% by GSE and Doxalone, respectively (Figure 3(C)). It is important tomention here that the comparisons discussed abovewith regard to GSE and Dox alone are those at their∼50% inhibitory concentrations in cell growth. An in-creased cell growth inhibition (Figure 3(A)) and celldeath (Figure 3(C)) in case of MCF-7 were also ob-served in several other GSE plus Dox combinations
ranging from 25, 50, 75 and 100 µg/ml GSE and10, 25, 50 and 75 nM Dox, as compared to similarGSE and Dox doses alone. When similar combina-tion studies were performed in MDA-MB468 cells,100 µg/ml GSE plus 50 nM Dox treatment for 48 hresulted in 82% growth inhibition as compared to 56%(GSE) and 57% (Dox) inhibition by each agent alone(Figure 3(B)), and 33% cell death by the combinationof GSE plus Dox as compared to 25% cell death byeach agent alone (Figure 3(D)). Several other com-binations of GSE plus Dox also showed an increasedgrowth inhibition and death of MDA-MB468 cells ascompared to each single agent alone (Figure 3(B) and(D)).

Efficacy of GSE plus doxorubicin in breast cancer 5
Figure 2. Growth inhibitory effect of Dox on human breast carcinoma cells. For studies assessing the effect of Dox on exponentially growingMCF-7 and MDA-MB468 cell growth (A and B) and cell death (C and D), respectively, 0.5 × 105 cells were plated in 60 mm dishes and 24 hafter plating, cultures were fed with fresh medium with vehicle (DMSO 0.1%, v/v) alone or Dox (25–100 nM) in medium. After 48 and 72 hof these treatments, the total numbers of cells were counted; including both viable and dead cells. Cell viability was determined by trypan bluedye exclusion method. The cell growth and cell death data shown are the mean ± S.E. of two independent plates; each sample was countedin duplicate. The representative data shown in the figures were reproducible in an additional independent experiment. Dox, doxorubicin;∗P < 0.05; #P < 0.01; $P < 0.001.
All the data shown in Figure 3 were next ana-lyzed for a possible synergism between different drugcombinations of GSE and Dox using CI-isobologramby Chou and Talalay [26]. As shown in Figure 4(A)and (B), most of the GSE plus Dox combinationsshowed synergistic effect in terms of MCF-7 andMDA-MB468 cell growth inhibition, respectively. Thestrongest synergistic effects (CI < 0.5), however, wereevident at a GSE dose of 100 µg/ml plus 25–75 nMDox, in both MCF-7 and MDA-MB468 cells. In termsof cell death, most of the CIs for GSE–Dox combina-tions suggested an additive effect in both the cell lines(Figure 4(C) and (D)). In terms of cell death, most
of the CIs for GSE–Dox combinations suggested ad-ditive but mostly weak antagonistic effect (CI > 1.05)in both cell lines (Figure 4(C) and (D)). To furthersupport our observations, normalized isobologramsfor GSE/Dox at various non-fixed ratios in MCF-7and MDA-MB468 cells are shown in Figure 5. Ac-cordingly, data points located below (left) the line ofadditivity indicate synergism, above the line of addi-tivity (right) indicate antagonistic, and on the line ofadditivity indicate additive effects [26]. At various ra-tios indicated in Figure 5, synergistic and antagonisticeffects were obtained for GSE/Dox in both MCF-7 andMDA-MB468 cell lines.

6 G Sharma et al.
Figure 3. Enhanced growth inhibitory effect of GSE–Dox combination on human breast carcinoma cells. For the studies assessing the effectof GSE–Dox combination on exponentially growing MCF-7 and MDA-MB468 cell growth (A and B) and cell death (C and D), respectively,0.5 × 105 cells were plated in 60 mm dishes and 24 h after plating, cultures were fed with fresh medium with vehicle (DMSO 0.1%, v/v) aloneor GSE (25, 50, 75 and 100 µg/ml) or Dox (10, 25, 50 or 75 nM) or combination of GSE and Dox (in both the cell lines). After 48 h of thesetreatments, the total numbers of cells were counted, including viable and dead cells; cell viability was determined by trypan blue dye exclusionmethod. The cell growth and cell death data shown are the mean ± S.E. of two independent plates; each sample was counted in duplicate. Therepresentative data shown in the figures were reproducible in an additional independent experiment. GSE, grape seed extract; Dox, doxorubicin.
Effect of GSE–Dox combination on cell-cycleprogression of MCF-7 and MDA-MB468 cells
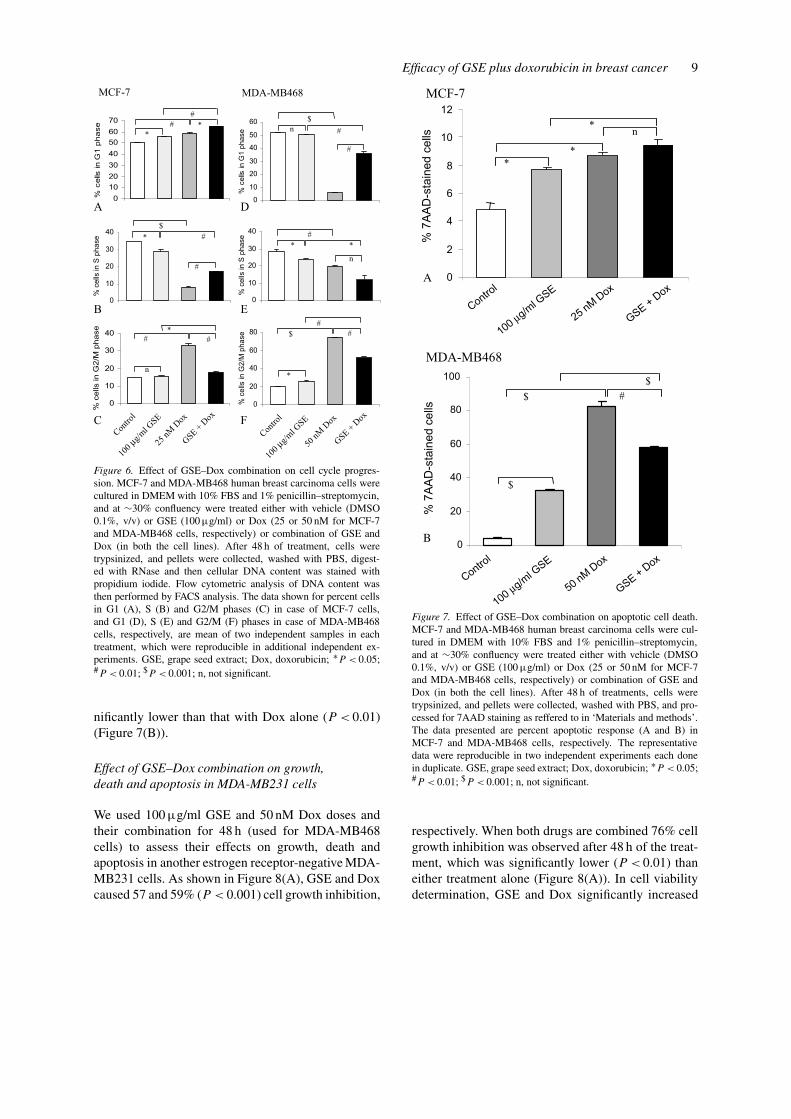
Based on an increased effect of GSE plus Dox combi-nation on cell growth inhibition, next we investigatedthe effect of similar doses (100 µg/ml GSE and 25 or50 nM Dox either alone or in combination) on cell-cycle progression in both MCF-7 and MDA-MB468cell lines. As shown in Figure 6, compared to vehicletreated control cells, a marked difference in cell-cycleprogression was observed in both the cell lines withGSE plus Dox combination as compared to single
agent alone. In MCF-7 cells, as shown in Figure 6(A)–(C), GSE (100 µg/ml) treatment showed a G1 arrest(56% v.s. 50% in control, P < 0.05), whereas Dox(25 nM) caused both G1 arrest (59% v.s. 50% in con-trol, P < 0.01) as well as G2/M arrest (33% v.s. 15%in control; P < 0.01) after 48 h of treatment. The ob-served cell-cycle arrests by GSE and Dox alone werelargely due to a strong decrease (P < 0.05–0.001) inS phase cells. In contrast to single agent, when MCF-7 cells were treated with a combination of GSE plusDox at identical doses and treatment time, it switchedthe Dox induced G2/M and G1 arrests as well as GSE-

Efficacy of GSE plus doxorubicin in breast cancer 7
Figure 4. Combination index (CI) for MCF-7 and MDA-MB468 cell growth (A and B) and death (C and D), respectively. The data shown inFigure 3 were analyzed for the CIs using the index-isobologram method based on the median-principle developed by Chou and Talalay [26].Variable ratios of drug concentrations and mutually exclusive equations were used to determine the CI. The CI equal to 1 indicates an additiveeffect; CI < 1 indicates synergy between the two drugs; and CI > 1 indicates antagonism between the two drugs. GSE, grape seed extract; Dox,doxorubicin.
induced G1 arrest to a much stronger G1 arrest (65%v.s. 50% in control) which was significantly higherthan the either treatment alone (P < 0.05–0.01) againat the expense of S phase population (Figure 6(A)–(C)). In case of MDA-MB468 cells, as shown inFigure 6(D)–(F), 48 h of GSE (100 µg/ml) or Dox(50 nM) treatment showed a moderate (26% in GSEv.s. 19% in control, P < 0.05) or very strong (75%in Dox v.s. 19% in control, P < 0.001) G2/M arrest,respectively, which was accompanied by a markeddecrease in both G1 as well as S phase populations.
A combination treatment of MDA-MB468 cells withGSE plus Dox at same doses for 48 h, also resultedin an alteration in cell-cycle progression phases ascompared to single agent alone. In this case, how-ever, combination showed more G1 arrest (P < 0.01)than that observed with Dox alone, but it was lessthan GSE alone (P < 0.01). In addition, there wasalso an increase in G2/M population when comparedto GSE alone (P < 0.01), but it was less than thatobserved with Dox alone (P < 0.01) (Figure 6(D)–(F)).

8 G Sharma et al.
Figure 5. Normalized isobologram for GSE–Dox combinations at various non-fixed ratios in MCF-7 and MDA-MB468 cell growth (A andB) and death (C and D). Data points located below (left) line of additivity indicated synergism, above line of additivity (right) indicatedantagonistic, and on line of additivity indicated additive effects of the drug combinations. GSE, grape seed extract; Dox, doxorubicin.
Effect of GSE–Dox combination on apoptoticdeath of MCF-7 and MDA-MB468 cells
Employing a quantitative apoptotic cell death assay,we next assessed whether the observed cell death byGSE plus Dox combination is accompanied by an ap-optotic death? MCF-7 and MDA-MB468 cells weretreated with GSE and Dox either alone or in com-bination for 48 h and then stained with 7AAD. Thedoses of GSE and Dox, and treatment time were sameas those for cell-cycle progression studies. In MCF-7 cells, as shown in Figure 7(A), the apoptotic cellpopulation increased from 5% in control to 8 and9% (P < 0.05) following treatments with 100 µg/ml
GSE and 25 nM Dox doses for 48 h, respectively.When cells were treated simultaneously with GSEplus Dox combination for 48 h, it resulted in compar-able apoptosis (10% apoptotic cells) to single agentsalone, however it was significantly higher than theGSE treatment alone (P < 0.05) (Figure 7(A)). In caseof MDA-MB468 cells, as shown in Figure 7(B), theapoptotic cell population increased from 4% in con-trol to 32 or 83% (P < 0.001) following treatmentswith 100 µg/ml GSE or 50 nM Dox for 48 h. How-ever, when cells were treated with the combination ofthese two agents, it caused 59% apoptotic cell death,which was significantly higher than that observed withGSE alone (P < 0.001) but at the same time was sig-
Efficacy of GSE plus doxorubicin in breast cancer 9
Figure 6. Effect of GSE–Dox combination on cell cycle progres-sion. MCF-7 and MDA-MB468 human breast carcinoma cells werecultured in DMEM with 10% FBS and 1% penicillin–streptomycin,and at ∼30% confluency were treated either with vehicle (DMSO0.1%, v/v) or GSE (100 µg/ml) or Dox (25 or 50 nM for MCF-7and MDA-MB468 cells, respectively) or combination of GSE andDox (in both the cell lines). After 48 h of treatment, cells weretrypsinized, and pellets were collected, washed with PBS, digest-ed with RNase and then cellular DNA content was stained withpropidium iodide. Flow cytometric analysis of DNA content wasthen performed by FACS analysis. The data shown for percent cellsin G1 (A), S (B) and G2/M phases (C) in case of MCF-7 cells,and G1 (D), S (E) and G2/M (F) phases in case of MDA-MB468cells, respectively, are mean of two independent samples in eachtreatment, which were reproducible in additional independent ex-periments. GSE, grape seed extract; Dox, doxorubicin; ∗P < 0.05;#P < 0.01; $P < 0.001; n, not significant.
nificantly lower than that with Dox alone (P < 0.01)(Figure 7(B)).
Effect of GSE–Dox combination on growth,death and apoptosis in MDA-MB231 cells
We used 100 µg/ml GSE and 50 nM Dox doses andtheir combination for 48 h (used for MDA-MB468cells) to assess their effects on growth, death andapoptosis in another estrogen receptor-negative MDA-MB231 cells. As shown in Figure 8(A), GSE and Doxcaused 57 and 59% (P < 0.001) cell growth inhibition,
Figure 7. Effect of GSE–Dox combination on apoptotic cell death.MCF-7 and MDA-MB468 human breast carcinoma cells were cul-tured in DMEM with 10% FBS and 1% penicillin–streptomycin,and at ∼30% confluency were treated either with vehicle (DMSO0.1%, v/v) or GSE (100 µg/ml) or Dox (25 or 50 nM for MCF-7and MDA-MB468 cells, respectively) or combination of GSE andDox (in both the cell lines). After 48 h of treatments, cells weretrypsinized, and pellets were collected, washed with PBS, and pro-cessed for 7AAD staining as reffered to in ‘Materials and methods’.The data presented are percent apoptotic response (A and B) inMCF-7 and MDA-MB468 cells, respectively. The representativedata were reproducible in two independent experiments each donein duplicate. GSE, grape seed extract; Dox, doxorubicin; ∗P < 0.05;#P < 0.01; $P < 0.001; n, not significant.
respectively. When both drugs are combined 76% cellgrowth inhibition was observed after 48 h of the treat-ment, which was significantly lower (P < 0.01) thaneither treatment alone (Figure 8(A)). In cell viabilitydetermination, GSE and Dox significantly increased

10 G Sharma et al.
Figure 8. Effect of GSE and Dox and their combination on cellgrowth, death and apoptosis in MDA-MB231 cells. Cells were cul-tured in DMEM with 10% FBS and 1% penicillin–streptomycin,and at ∼30% confluency were treated either with vehicle (DMSO0.1%, v/v) or GSE (100 µg/ml) or Dox (50 nM) or combination ofboth. After 48 h of treatments, cells were collected, and (A) totalcell number, (B) dead cells and (C) apoptosis were determined asdescribed in ‘Materials and methods’. The data are presented asmean ± SE of triplicate samples for each treatment. GSE, grapeseed extract; Dox, doxorubicin; #P < 0.01; $P < 0.001; n, notsignificant.
cell death from 10% in control to 24% (P < 0.01) and41% (P < 0.001), respectively (Figure 8(B)). How-ever, in combination the increase in cell death wasnot significantly different from Dox treatment alone.
Similarly, in apoptosis study, a significant increase ap-optotic death was observed by GSE (21%, P < 0.01)and Dox (45%, P < 0.01) as compared to control(5.7%) (Figure 8(C)). The apoptotic effect of com-bination treatment was only significantly (P < 0.01)higher than the GSE treatment alone, and was slightlylower (not significant) than the Dox treatment alone.These results showed a similar trend in all these threeparameters when compared with MDA-MB468 cells.
Discussion
The central finding of the present investigation is thatGSE increases the efficacy of Dox in human breastcarcinoma MCF-7, MDA-MB468 and MDA-MB231cells. The significance of this finding lies in the factthat Dox and other anthracyclines are well-knowncancer therapeutic agents, but cause high toxicity tothe normal tissues during cancer therapies includingbreast cancer [28]. Several studies have shown that be-sides their harmful effects on health, such as immuno-suppression and cardiomyopathy, these agents lead tothe development of primary and secondary drug res-istance in tumor cells thereby limiting their clinicalsuccess in the cancer chemotherapy [29, 30]. In thisregard, recent reports show that combination chemo-therapy is a superior modality for cancer treatmentthan single-agent, and that naturally occurring dietarysupplements with known anti-cancer activity could beone of the agents in combination chemotherapy thatmight also reduce systemic toxicity of the chemothera-peutic agents [31, 32]. Consistent with these notions,GSE is a non-toxic agent (unpublished observation),which is widely consumed as a dietary supplement andpossesses anticancer activity against various epithelialcancers [22, 23, 33, 34]. Accordingly, based on thedata shown in the present study, GSE could be usefulin synergizing the efficacy of cancer chemotherapeuticagents in breast cancer treatment.
Overall, we observed that in vitro therapeutic ef-fect of Dox in terms of cell growth inhibition at 75and 100 nM doses could be achieved at its nearlyhalf concentration in combination with 100 µg/mldose of GSE in MCF-7 and MDA-MB468 cells,respectively. The results of our studies, showingincreased therapeutic effect of Dox in combina-tion with GSE in both estrogen-dependent (MCF-7)and estrogen-independent (MDA-MB468 and MDA-MB231) human breast carcinoma cells, stronglysupport our hypothesis that combination of a diet-

Efficacy of GSE plus doxorubicin in breast cancer 11
ary supplement like GSE, along with reducing thedosage of a conventional breast cancer chemothera-peutic agent such as Dox, may retain the benefitstogether with minimizing the normal cytotoxic effectsand thus may enhance the therapeutic efficacy. In otherstudies, we found that GSE induces Dox-caused G1(MCF-7 cells) and G2/M (MDA-MB468 cells) arrestin cell cycle progression. Therefore, the regulation ofG1 and G2/M transition points could be an effectivetarget to check the growth and proliferation of breastcancer cells. Various studies have shown that growthsignal determines whether the cells will be in a rest-ing phase or will enter the G1 phase and go throughcell-cycle progression for proliferation [35]. The dataobtained clearly indicate that GSE–Dox combinationcauses the accumulation of MCF-7 cells in G1 phase.Therefore, this combination might have inhibitory ef-fect on G1-S transition in cell-cycle progression ofbreast cancer cells.
In apoptosis studies, we observed that GSE causesapoptotic death of MCF-7 cells, and that it hascomparable effect in presence of Dox dose usedin this study. Future studies are needed to assesswhether other GSE plus Dox dose combinations,which showed synergistic and additive effects ingrowth inhibition and death of MCF-7 cells, re-spectively, result in an enhanced apoptotic death.With regard to MDA-MB468 cells, we observed thatGSE treatment causes a strong apoptotic death, andthat in presence of Dox, apoptotic activity of GSEis significantly increased. Although this combina-tion showed a reduced apoptotic effect as comparedto Dox alone, this might still be useful keepingin view the toxicity of Dox to the normal tissues.Further, these finding in MDA-MB468 cells wereconfirmed in another estrogen receptor-negative cellline MDA-MB231, in which similar trends in cellgrowth inhibition and apoptosis were observed. Ourdata also show that MCF-7 cells are more resistantto apoptosis when compared with MDA-MB468 orMDA-MB231 cells for the same doses of GSE andDox.
In summary, our results clearly demonstrate a bet-ter effect of GSE–Dox combination in human breastcarcinoma MCF-7, MDA-MB468 and MDA-MB231cells than either agent alone, suggesting that studiesare warranted to assess in vivo efficacy of GSE–Doxcombination in pre-clinical breast cancer models to-gether with mechanistic rationale for such efficacy. Apositive outcome of such studies could be useful toincrease the efficacy of existing chemotherapy in the
treatment of breast cancer with a reduced toxicity tothe normal tissues.
Acknowledgement
This study was supported in part by P30 CA46934-14S1 from the National Cancer Institute, Bethesda,MD, USA.
References
1. Jemal A, Thomas A, Murray T, Thun M: Cancer statistics,2002. CA Cancer J Clin 52: 23–47, 2002
2. Fournier DB, Erdman Jr JW, Gordon GB: Soy, its components,and cancer prevention: a review of the in vitro, animal, and hu-man data. Cancer Epidemiol Biomarkers Prev 7: 1055–1065,1998
3. Agarwal R, Mukhtar H: Chemoprevention of photocarcino-genesis. Photochem Photobiol 63: 440–444, 1996
4. Dorai T, Cao YC, Dorai B, Buttyan R, Katz AE: Thera-peutic potential of curcumin in human prostate cancer. III.Curcumin inhibits proliferation, induces apoptosis, and in-hibits angiogenesis of LNCaP prostate cancer cells in vivo.Prostate 47: 293–303, 2001
5. Lippman SM, Lee JJ, Sabichi AL: Cancer chemoprevention:progress and promise. J Natl Cancer Inst 90: 1514–1528, 1998
6. Kelloff GJ: Perspectives on cancer chemoprevention researchand drug development. Adv Cancer Res 78: 199–334, 2000
7. Hill MJ: Nutrition and human cancer. Ann NY Acad Sci 833:68–78, 1997
8. Wargovich MJ: Nutrition and cancer: the herbal revolution.Curr Opin Clin Nutr Metab Care 2: 421–424, 1999
9. Dragsted LO: Natural antioxidants in chemoprevention. ArchToxicol Suppl 20: 209–226, 1998
10. Mukhtar H, Agarwal R: Skin cancer chemoprevention. J InvestDermatol Symp Proc 1: 209–214, 1996
11. Singleton VL: Tannins and the qualities of wines. In: Laks PE,Hemingway RW (eds) Plant Polyphenols. Plenum Press, NewYork, 1992, pp 859–880
12. Bartolome B, Hernandez T, Bengoechea ML, Quesada C,Gomez-Cordoves C, Estrella I: Determination of some struc-tural features of procyanidins and related compounds byphotodiode-array detection. J Chromatogr A 723: 19–26, 1996
13. Escribano-Bailon MT, Gutierrez-Fernandez Y, Rivas-GonzaloJC, Santos-Buelga C: Characterization of procyanidins of Vitisvinifera variety Tinta del Paris grape seeds. J Agric FoodChem 40: 1794–1799, 1992
14. Bagchi D, Garg A, Krohn RL, Bagchi M, Bagchi DJ, BalmooriJ, Stohs SJ: Protective effects of grape seed proanthocyan-idins and selected antioxidants against TPA-induced hepaticand brain lipid peroxidation and DNA fragmentation, andperitoneal macrophage activation in mice. Gen Pharmacol 30:771–776, 1998
15. Maffei Facina R, Carini M, Aldini G, Berti F, Rossoni G,Bombardelli E, Morazzoni P: Procyanidins from Vitis viniferaseeds protect rabbit heart from ischemia/reperfusion injuri:antioxidant intervention and/or iron and copper sequesteringability. Planta Med 62: 495–502, 1996
16. Bagchi D, Garg A, Krohn RL, Bagchi M, Tran MX, StohsSJ: Oxygen free radical scavenging abilities of vitamin C and

12 G Sharma et al.
E, and a grape seed proanthocyanidin extract in vitro. ResCommun Mol Pathol Pharmacol 95: 179–189, 1997
17. Packer L, Rimbach G, Virgili F: Antioxidant activity andbiologic properties of a procyanidin-rich extract from pine(Pinus maritima) bark, pycnogenol. Free Radic Biol Med 27:704–724, 1999
18. Halpern MJ, Dahlgren AL, Laakso I, Seppanen-Laakso T,Dahlgren J, McAnulty PA: Red-wine polyphenols and in-hibition of platelet aggregation: possible mechanisms, andpotential use in health promotion and disease prevention. J IntMed Res 26: 171–180, 1998
19. Soleas GJ, Diamandis EP, Goldberg DM: Wine as a biologicalfluid: history, production, and role in disease prevention. J ClinLab Anal 11: 287–313, 1997
20. Ye X, Krohn RL, Liu W, Joshi SS, Kuszynski CA, McGinnTR, Bagchi M, Preuss HG, Stohs SJ, Bagchi D: The cytotoxiceffects of a novel IH636 grape seed proanthocyanidin ex-tract on cultured human cancer cells. Mol Cell Biochem 196:99–108, 1999
21. Arii M, Miki M, Hosoyama R, Ariga H, Yamaji N, KataokaS: Chemopreventive effect of grape seed extract on intestinalcarcinogenesis in the APCMin mouse. Proc Am Assoc CancerRes Annu Meet 39: Abstract 20, 1998
22. Bomser JA, Singletary KW, Wallig MA, Smith MA: Inhibitionof TPA-induced tumor promotion in CD-1 mouse epidermisby a polyphenolic fraction from grape seeds. Cancer Lett 135:151–157, 1999
23. Zhao J, Wang J, Chen Y, Agarwal R: Anti-tumor-promotingactivity of a polyphenolic fraction isolated from grape seedsin the mouse skin two-stage initiation-promotion protocol andidentification of procyanidins B5-3′-gallate as the most effec-tive antioxidant constituent. Carcinogenesis 20: 1737–1745,1999
24. Zheng T, Boyle P, Willett WC, Hu H, Dan J, Evstifeeva TV,Niu S, MacMahon BA: A case-control study of oral cancer inBeijing. People’s Republic of China. Associations with nutri-ent intakes, foods and food groups. Eur J Cancer Biol OralOncol 29B: 45–55, 1993
25. Hortobagyi GN: Chemotherapy of breast cancer: a historicalperspective. Semin Oncol S17: 1–4, 1997
26. Chou TC, Talalay P: Analysis of combined drug effects: a newlook at a very old problem. Trends Pharmacol Sci 4: 450–454,1983
27. Philpott NJ, Turner AJ, Scopes J, Westby M, Marsh JC,Gordon-Smith EC, Dalgleish AG, Gibson FM: The use of 7-amino actinomycin D in identifying apoptosis: simplicity ofuse and broad spectrum of application compared with othertechniques. Blood 87: 2244–2251, 1996
28. Chen ST, Pan TL, Tsai YC, Huang CM: Proteomics revealsprotein profile changes in doxorubicin-treated MCF-7 humanbreast cancer cells. Cancer Lett 181: 95–107, 2002
29. Von Hoff DD, Layard MW, Basa P, Davis Jr HL, Von Hoff AL,Rozencweig M, Muggia FM: Risk factors for doxorubicin-induced congestive heart failure. Ann Int Med 91: 710–717,1979
30. Raghavan D, Koczwara B, Javle M: Evolving strategies ofcytotoxic chemotherapy for advanced prostate cancer. Eur JCancer 33: 566–574, 1997
31. Hortobagyi GN: Progress in systemic chemotherapy ofprimary breast cancer: an overview. J Natl Cancer Inst Monogr72–79, 2001
32. Tyagi AK, Singh RP, Agarwal C, Chan DCF, AgarwalR: Silibinin strongly synergizes human prostate carcinomaDU145 cells to doxorubicin-induced growth inhibition, G2-Marrest and apoptosis. Clin Cancer Res 8: 3512–3519, 2002
33. Agarwal C, Sharma Y, Agarwal R: Anticarcinogenic effect ofa polyphenolic fraction isolated from grape seeds in humanprostrate carcinoma DU145 cells: modulation of mitogenicsignaling and cell-cycle regulators and induction of G1 arrestand apoptosis. Mol Carcinog 28: 129–138, 2000
34. Agarwal C, Sharma Y, Zhao J, Agarwal R: A polyphen-olic fraction from grape seeds causes irreversible growthinhibition of breast carcinoma MDA-MB468 cells by in-hibiting mitogen-activated protein kinases activation and in-ducing G1 arrest and differentiation. Clin Cancer Res 6:2921–2930, 2000
35. Singh RP, Dhanalakshmi S, Agarwal R: Phytochemicals ascell cycle modulators: a less toxic approach in halting humancancers. Cell Cycle 1: 156–161, 2002
Address for offprints and correspondence: Rajesh Agarwal, PhD,Department of Pharmaceutical Sciences, School of Pharmacy,University of Colorado Health Sciences Center, 4200 East 9thAvenue, Box C238, Denver, CO 80262, USA; Tel.: +1-303-315-1381; Fax: +1-303-315-6281; E-mail: [email protected]




















