
Favretto et al 2016 dryland ecosystem services tradeoffs.pdf
Upload
independentCategory
view
1download
0
a p p l i e d s o i l e c o l o g y 3 6 ( 2 0 0 7 ) 1 6 4 – 1 7 5
Synchronizing nitrogen availability through application oforganic inputs of varying resource quality in a tropicaldryland agroecosystem
Sonu Singh *, Nandita Ghoshal, K.P. Singh
Center of Advanced Study, Department of Botany, Banaras Hindu University, Varanasi 221005, India
a r t i c l e i n f o
Article history:
Received 20 June 2006
Received in revised form
19 December 2006
Accepted 30 January 2007
Keywords:
Dryland agroecosystem
N-mineralization rate
Available-N
Soil microbial biomass N
Mixed input
a b s t r a c t
A 2-year field experiment was conducted to evaluate the impact of management practices
involving manipulation of quantity and quality of exogenous inputs on soil N-mineraliza-
tion rate, N availability and microbial biomass in a rice–barley rotation in a tropical dryland
agroecosystem. At the beginning of each annual cycle an equivalent amount of N was added
through chemical fertilizer and three organic inputs: Sesbania shoot (high quality resource,
C/N 16, lignin/N 3.2, polyphenol + lignin/N 4.2), wheat straw (low quality resource, C/N 82,
lignin/N 34.8, polyphenol + lignin/N 36.8) and Sesbania + wheat straw (high and low quality
resources mixed). The N-mineralization rate was dominated by ammonification in this
dryland agroecosystem. N-mineralization exhibited a distinct seasonal pattern, decreasing
from the rice period through the summer fallow period, except in Sesbania + wheat straw
and wheat straw treatments which showed a slight increase during the early stages of barley
period. The rate of N-mineralization showed a significant relationship with soil moisture
and microbial biomass N. During the rice period, N-mineralization rate and available-N was
highest in the fertilizer treatment followed by Sesbania > Sesbania + wheat straw > wheat
straw. During the barley period, highest N-mineralization rate and available-N was observed
in Sesbania + wheat straw followed by wheat straw > Sesbania > fertilizer. Adding Sesba-
nia + wheat straw resulted in consistently higher levels of microbial biomass N, N-miner-
alization rate and available-N through the annual cycle compared to single application of
Sesbania and wheat straw, indicating synergy between the two inputs, favoring more
efficient utilization of N. It is suggested that mixed application of high and low quality
resources can modulate N release, resulting in relatively higher synchronization which can
help in minimizing N loss from agroecosystem.
# 2007 Published by Elsevier B.V.
avai lable at www.sc iencedi rec t .com
journal homepage: www.e lsev ier .com/ locate /apsoi l
1. Introduction
Nitrogen (N) is one of the major nutrient required by crops in an
agroecosystem. Large reserves of N are present in soil organic
matter, but its availability to plants is influenced by several
competing processes which include mineralization, immobili-
zation (by microorganisms and/or plants), nitrification and
* Corresponding author. Tel.: +91 9415811843.E-mail address: [email protected] (S. Singh).
0929-1393/$ – see front matter # 2007 Published by Elsevier B.V.doi:10.1016/j.apsoil.2007.01.007
denitrification. Only few studies have examined the kinetics of
gross N-mineralization, immobilization, and nitrification rates
in soil at temperatures above 15 8C (i.e., under tropical
conditions) (Hoyle et al., 2006). Agroecosystems in general
and tropical dryland agroecosystem in particular invariably
require replenishment of N through exogenous N sources.
Chemical fertilizers are most widely used as supplemental N.

a p p l i e d s o i l e c o l o g y 3 6 ( 2 0 0 7 ) 1 6 4 – 1 7 5 165
However, questions have been raised about the long-term
sustainability of such systems because rate of release of N in
soils often does not match crop demand with fertilizer
applications (Robertson et al., 2000; Campbell et al., 1995;
Groffman et al., 1987). Moreover, highly concentrated inorganic
N inputs can have detrimental environmental impacts (Ven-
terea and Rolston, 2000; Matson et al., 1997). As an alternative N
source, use of organic materials (crop residue, compost or green
manures) has been advocated (Tilman, 1998). Organic amend-
ments are considered to have the potential for conservation of
soil moisture, one of the major constraints to crop productivity
in dryland, besides providing soil nutrients.
Organic resources like crop residues offer sustainable and
ecologically sound alternatives for meeting the N requirement
of crops. Moreover effective management of postharvest crop
residues remains an important issue in many grain-producing
regions of the world (Beare et al., 2002). There is need to
develop and adopt economical and efficient N management
strategies that conserve natural resources while minimizing
adverse environmental impacts. Many of the present manage-
ment practices do not meet these criteria (Giller et al., 2004).
The suitability of crop residues as a source of N depends on the
mineralization of N in synchrony with crop demand. Thus, to
use crop residues efficiently as a source of N we need to predict
more accurately their rates of N release. Further, we need to
know more accurately how much N flows through agroeco-
systems and how much N is susceptible for loss (Janzen et al.,
2003).
An important question pertinent to the efficient use of
residues is whether their rates of nutrient release can be
effectively managed to coincide with crop demand (Tilman
et al., 2002). If rates of N release exceed plant demands, then
the N becomes susceptible to various pathways of loss
(Goulding, 2004; Peoples et al., 2004). In some instances,
organic amendments can cause excess accumulation of NO3-
N in soil (Khalil et al., 2005) with potentially detrimental effects
on the environment. Conversely, if rates of N release are too
slow, then crop yields may be constrained.
The rate of N-mineralization, which regulates the avail-
ability of N in soil is governed by microbial biomass (Hadas
et al., 2004). Tillage and residue (straw) management have
been shown to affect microbial biomass, N-mineralization rate
and available-N in rice–barley based dryland agroecosystems
(Kushwaha and Singh, 2005). The environmental hazards
associated with N in agroecosystems stem from various
specific microbial N transformations in soils as well as the
behaviour of mineral N in relation to soil physico-chemical
properties (Peoples et al., 1995). Hence study of relations
between N availability and microbial biomass is of great
importance. This study, therefore, was conducted to examine
the effect of resource quality on N-mineralization rate, N
availability, N synchrony and soil microbial biomass N in a
field trial under dry tropical conditions. The aim was to
increase the temporal resolution of N dynamics. Detailed
information on the temporal resolution of mineralization is
important to achieve synchrony between nutrient release
from added materials and the nutrient requirements of the
crop (Myers et al., 1994). This field study examined three
diverse organic inputs (high quality Sesbania shoot, low quality
wheat straw, and a combination of these (Sesbania + wheat
straw) and fertilizer, applied at equivalent rates of N (80 kg N
ha�1). The objectives of the present study were to evaluate the
effect of resource qualities through the annual cycle on: (i) the
N-mineralization rate (ammonification + nitrification) and
microbial biomass N, and (ii) the seasonal availability of
NH4-N and NO3-N in soil.
2. Materials and methods
2.1. Study site
The experiments were conducted for two annual cycles (2002–
2003, 2003–2004) in the cultivated plots of the Botanical Garden
of the Department of Botany, Banaras Hindu University at
Varanasi (258180N lat. and 83810E long., 76 m above mean sea
level). This region has a dry tropical climate, characterized by
strong seasonal variations in temperature and precipitation,
including a warm rainy season (July–September), a cool winter
(November–February), and a hot summer (April–June). The
average annual rainfall is about 1100 mm, of which about 80%
is received during the rainy season. High temperature
(24–34 8C) and high relative humidity (70–80%) prevail during
the rainy season. In the winter season the temperature range
is 4–25 8C. The summer is dry and hot with a temperature
range of 30–48 8C. Crop production occurs in two seasons: the
rainy season and the winter season, and fields are left fallow
during the summer. The soil of the experimental site (order
Inceptisol, suborder Orchrepts, subgroup Udic Ustocrepts) has
pale brown colour and sandy loam texture.
2.2. Experimental design
The crop sequence studied was rice (Oryza sativa var. NDR 97)-
barley (Hordeum vulgare var. Lakhan)-summer fallow. Rice was
grown from day 0 (day of sowing of first rice crop) to day 115 in
the first annual cycle and from day 367 to 475 in the second
annual cycle; barley was grown from day 130 to 265, and day
490 to 645; summer fallow extended from day 266 to 335 and
from day 646 to 700.
The study compared residues of contrasting quality:
Sesbania aculeata shoot (N 3.03%, C/N 16.4, lignin/N 3.2, and
polyphenols + lignin/N 4.2) and wheat straw (N 0.61%, C/N
81.8, lignin/N 34.8, and polyphenols + lignin/N 36.8), each
applied to deliver equivalent N. The experimental design
included five treatments: (1) control (no inputs), (2) chemical
fertilizer (80 kg N ha�1 from urea), (3) Wheat straw
(80 kg N ha�1), (4) Sesbania shoot (80 kg N ha�1), and (5) Sesbania
shoot (40 kg N ha�1) + wheat straw (40 kg N ha�1). The experi-
ment was designed to vary the quality of exogenous soil inputs
having an equivalent amount of N application by using high
quality organic input in form of S. aculeata shoot and low
quality input in form of wheat straw. These treatments were
applied to 3 m � 3 m plots, arranged in a random block design
with three replicates, and a 1 m strip between blocks.
The inputs were applied once each year, 1 or 2 days before
sowing of the rice crop, and incorporated into the soil to a
depth of 0–10 cm. Fresh Sesbania shoots were cut into about 2–
3 cm pieces before incorporation. Wheat straw was air-dried
and then incorporated. In the Sesbania shoot + wheat straw

a p p l i e d s o i l e c o l o g y 3 6 ( 2 0 0 7 ) 1 6 4 – 1 7 5166
treatment both organic inputs were mixed thoroughly and
then incorporated. The N fertilizer was surface-broadcast,
without incorporation. No exogenous inputs were applied to
the barley crop.
2.3. Soil collection
For estimation of available-N, N-mineralization rate and
microbial biomass N soil samples (0–10 cm depth) were
collected eleven times, once during seedling, pre-grain-
forming, grain-forming, post-grain-forming and maturity
stages of rice as well as barley and once during summer
fallow. Three sub-samples of soil were collected from each
replicate plot and composited. After removing visible plant
debris and fauna, the soil was sieved through a 2 mm mesh
screen. The soil was analyzed for available-N (NO3-N and NH4-
N), microbial biomass N, and N-mineralization rate.
2.4. Estimation of available-N and N-mineralization rate
Field moist and sieved soil samples were analyzed for nitrate-
N (NO3-N) by the phenol disulphonic acid method, using CaSO4
as the extractant (Jackson, 1973) and for ammonium-N (NH4-
N) by the phenate method (APHA, 1995), using 2 M KCl as the
extractant. Available-N was calculated as the sum of NO3-N
and NH4-N. N-mineralization rate was measured using the
buried bag technique (Eno, 1960). Two sub-samples of soil
(about 150 g each), with coarse roots and large organic debris
removed to minimize immobilization, were enclosed in sealed
polyethylene bags and buried at 5–10 cm depth in each plot.
Soil NO3-N and NH4-N were determined, as previously
described, before and after field incubation for 30 days.
Ammonification was estimated from the increases in NH4-N
concentration; nitrification was estimated from the increase
in NO3-N concentration; and net N-mineralization was
estimated from the sum of increases in NH4-N and NO3-N
concentrations, during the field incubation. All results were
expressed on an oven dry soil (105 8C) basis.
2.5. Microbial biomass N estimation
Field moist soil samples were pre-conditioned for 7 days at
room temperature in a container with 100% humidity and an
alkali trap to remove CO2. The container was opened for
aeration a few minutes every day. Microbial biomass N (MBN)
was estimated by the chloroform fumigation–extraction
method (Brookes et al., 1985), using a purified CHCl3 treatment,
followed by extraction of both fumigated and non-fumigated
soils with 0.5 M K2SO4, which was then analyzed for total N by
Kjeldahl digestion. The MBN was estimated as: MBN = EN/0.54,
where EN is the difference between the amounts of N extracted
from fumigated and non-fumigated soil (mg N g�1 oven dry
soil); and 0.54 is the fraction of biomass N extracted after
chloroform fumigation.
2.6. Determination of crop N-uptake and apparent N-recovery
In each plot, plants were harvested at ground-level from two
randomly located subplots (each 25 cm � 25 cm) and dried at
80 8C. To estimate root biomass, a soil monolith
(10 cm � 10 cm, 10 cm deep) was excavated from harvested
sub-plot, and washed with a fine jet of water over twin sieves
(2 mm mesh above and 0.5 mm mesh below). Roots recovered
were oven dried at 80 8C and weighed. Above- and below-
ground plant materials were finely ground and analyzed for N
content by a micro-Kjeldahl method (Jackson, 1973). Nitrogen
uptake was calculated by multiplying the biomass of plant
parts for both crops by their N concentration.
Apparent N-recovery (ANR) of added N was calculated as
described by Dilz (1988):
ANR ð%Þ
¼ N-uptake in treatment�N-uptake in controlN applied in treatment
� 100
2.7. Soil moisture content estimation
Soil moisture content was estimated as gravimetric soil water
at 0–10 cm depth. Fresh soil (10 g) was sampled in triplicate
from each plot, dried at 105 8C for 24 h and weighed. Soil
moisture content was calculated as:
soil moisture content ð%Þ
¼ weight of fresh soil�weight of dry soilweight of dry soil
� 100
2.8. Statistical analysis
SPSS package was used for the following statistical analyses:
two-way analysis of variance (ANOVA), correlation and least
significant difference (LSD) test. Two-way ANOVA was used to
detect significant differences between the effects of input
(treatments) and sampling date on rate of N-mineralization.
Treatment means were compared using the LSD test at
P < 0.05. The five treatments were regarded as distinct
strategies. N-mineralization rate, available-N (NH4-N and
NO3-N) and MBN measured through the annual cycle were
averaged to assess the variations among three ecologically
distinct phases (two cropping seasons and summer fallow).
3. Results
3.1. Soil N-mineralization rate
Two-way ANOVA indicated that the impact of treatments and
crop stages on the rate of N-mineralization was significant
(P < 0.05). The N-mineralization rate decreased consistently
from rice period to summer fallow through barley period.
However, a slight increase was observed during the early stage
of barley cropping in Sesbania + wheat straw and wheat straw
treatments (Fig. 1). In treatments receiving Sesbania and
fertilizer, a steep decline in N-mineralization rate was
observed from rice period to barley period whereas it was
gradual in Sesbania + wheat straw and wheat straw treat-
ments. The rate of N-mineralization increased throughout the
annual cycle due to application of various soil amendments
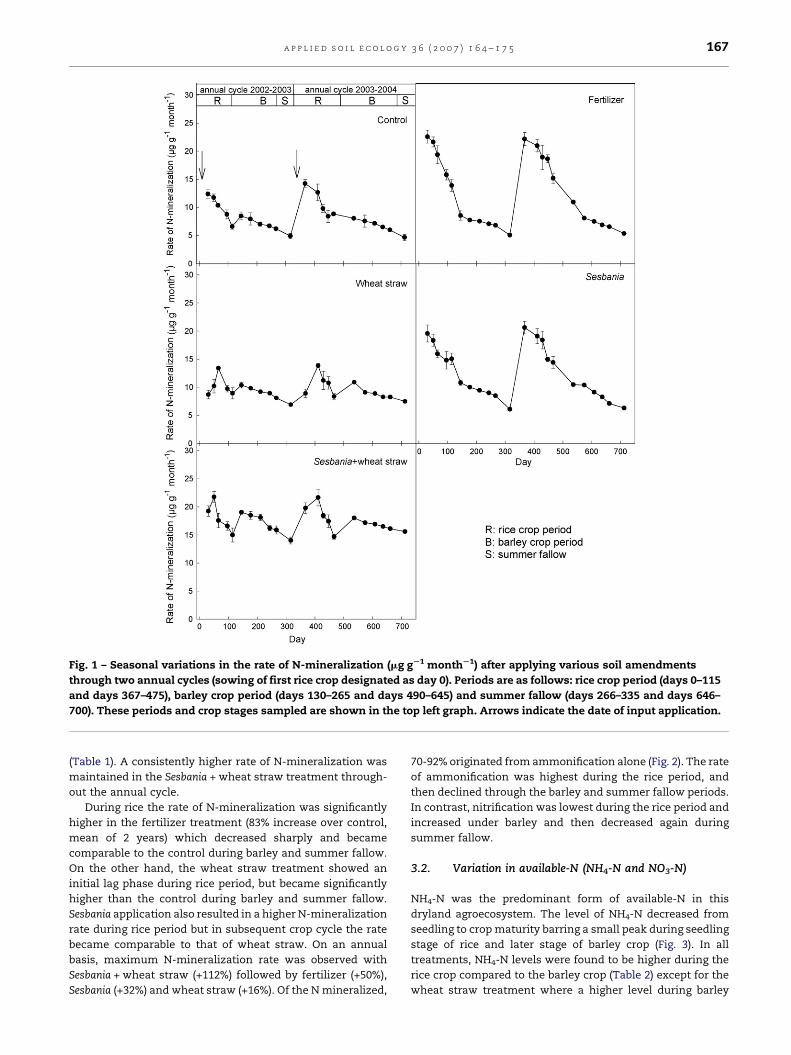
Fig. 1 – Seasonal variations in the rate of N-mineralization (mg gS1 monthS1) after applying various soil amendments
through two annual cycles (sowing of first rice crop designated as day 0). Periods are as follows: rice crop period (days 0–115
and days 367–475), barley crop period (days 130–265 and days 490–645) and summer fallow (days 266–335 and days 646–
700). These periods and crop stages sampled are shown in the top left graph. Arrows indicate the date of input application.
a p p l i e d s o i l e c o l o g y 3 6 ( 2 0 0 7 ) 1 6 4 – 1 7 5 167
(Table 1). A consistently higher rate of N-mineralization was
maintained in the Sesbania + wheat straw treatment through-
out the annual cycle.
During rice the rate of N-mineralization was significantly
higher in the fertilizer treatment (83% increase over control,
mean of 2 years) which decreased sharply and became
comparable to the control during barley and summer fallow.
On the other hand, the wheat straw treatment showed an
initial lag phase during rice period, but became significantly
higher than the control during barley and summer fallow.
Sesbania application also resulted in a higher N-mineralization
rate during rice period but in subsequent crop cycle the rate
became comparable to that of wheat straw. On an annual
basis, maximum N-mineralization rate was observed with
Sesbania + wheat straw (+112%) followed by fertilizer (+50%),
Sesbania (+32%) and wheat straw (+16%). Of the N mineralized,
70-92% originated from ammonification alone (Fig. 2). The rate
of ammonification was highest during the rice period, and
then declined through the barley and summer fallow periods.
In contrast, nitrification was lowest during the rice period and
increased under barley and then decreased again during
summer fallow.
3.2. Variation in available-N (NH4-N and NO3-N)
NH4-N was the predominant form of available-N in this
dryland agroecosystem. The level of NH4-N decreased from
seedling to crop maturity barring a small peak during seedling
stage of rice and later stage of barley crop (Fig. 3). In all
treatments, NH4-N levels were found to be higher during the
rice crop compared to the barley crop (Table 2) except for the
wheat straw treatment where a higher level during barley
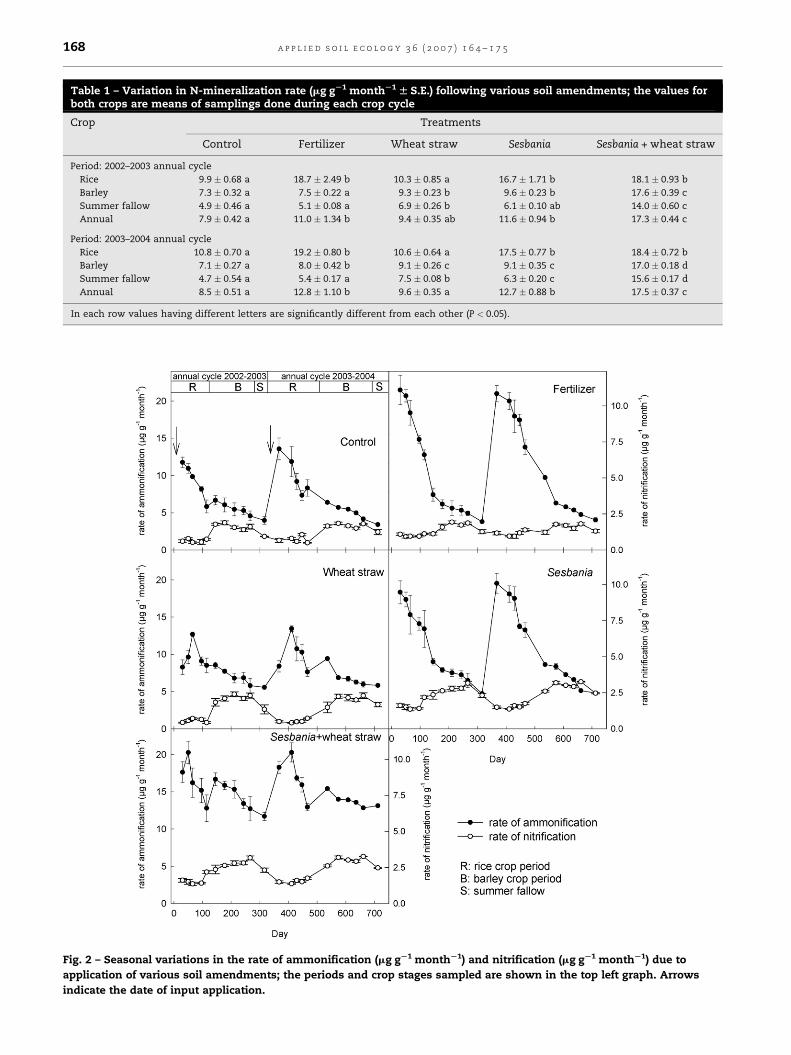
Table 1 – Variation in N-mineralization rate (mg gS1 monthS1 W S.E.) following various soil amendments; the values forboth crops are means of samplings done during each crop cycle
Crop Treatments
Control Fertilizer Wheat straw Sesbania Sesbania + wheat straw
Period: 2002–2003 annual cycle
Rice 9.9 � 0.68 a 18.7 � 2.49 b 10.3 � 0.85 a 16.7 � 1.71 b 18.1 � 0.93 b
Barley 7.3 � 0.32 a 7.5 � 0.22 a 9.3 � 0.23 b 9.6 � 0.23 b 17.6 � 0.39 c
Summer fallow 4.9 � 0.46 a 5.1 � 0.08 a 6.9 � 0.26 b 6.1 � 0.10 ab 14.0 � 0.60 c
Annual 7.9 � 0.42 a 11.0 � 1.34 b 9.4 � 0.35 ab 11.6 � 0.94 b 17.3 � 0.44 c
Period: 2003–2004 annual cycle
Rice 10.8 � 0.70 a 19.2 � 0.80 b 10.6 � 0.64 a 17.5 � 0.77 b 18.4 � 0.72 b
Barley 7.1 � 0.27 a 8.0 � 0.42 b 9.1 � 0.26 c 9.1 � 0.35 c 17.0 � 0.18 d
Summer fallow 4.7 � 0.54 a 5.4 � 0.17 a 7.5 � 0.08 b 6.3 � 0.20 c 15.6 � 0.17 d
Annual 8.5 � 0.51 a 12.8 � 1.10 b 9.6 � 0.35 a 12.7 � 0.88 b 17.5 � 0.37 c
In each row values having different letters are significantly different from each other (P < 0.05).
Fig. 2 – Seasonal variations in the rate of ammonification (mg gS1 monthS1) and nitrification (mg gS1 monthS1) due to
application of various soil amendments; the periods and crop stages sampled are shown in the top left graph. Arrows
indicate the date of input application.
a p p l i e d s o i l e c o l o g y 3 6 ( 2 0 0 7 ) 1 6 4 – 1 7 5168

Fig. 3 – Seasonal variations in the levels of available ammonium-N and nitrate-N (mg gS1 soil) due to application of various
soil amendments; the periods and crop stages sampled are shown in the top left graph. Arrows indicate the date of input
application.
a p p l i e d s o i l e c o l o g y 3 6 ( 2 0 0 7 ) 1 6 4 – 1 7 5 169
period compared to rice period was found. During summer
fallow, the lowest level of NH4-N was recorded in all
treatments. Seasonal variations were very distinct in case of
Sesbania and fertilizer treatments whereas in case of control,
Table 2 – Variation in NH4-N levels (mg gS1 soil W S.E.) followinmeans of samplings done during each crop cycle
Crop
Control Fertilizer W
Period: 2002–2003 annual cycle
Rice 11.9 � 0.62 a 19.6 � 0.85 b
Barley 9.2 � 0.23 a 9.7 � 0.32 a 1
Summer fallow 6.0 � 0.35 a 7.4 � 0.17 b 7
Annual 10.1 � 0.43 a 14.0 � 0.99 b 1
Period: 2003–2004 annual cycle
Rice 12.0 � 0.56 a 19.7 � 0.86 b 1
Barley 9.6 � 0.27 a 10.0 � 0.66 a 1
Summer fallow 6.4 � 0.28 a 7.7 � 0.25 bc
Annual 10.5 � 0.41 a 14.6 � 1.00 b 1
In each row values having different letters are significantly different fro
wheat straw and Sesbania + wheat straw treatments the
variations were moderate.
During the rice period higher levels of NH4-N were observed
in the treatments receiving fertilizer (+65%), Sesbania (+36%)
g various soil amendments; the values for both crops are
Treatments
heat straw Sesbania Sesbania + wheat straw
9.7 � 0.33 c 16.0 � 0.75 d 15.7 � 0.5 d
1.3 � 0.49 b 10.6 � 0.65 ab 14.0 � 0.59 c
.76 � 0.27 b 6.9 � 0.32 ab 10.5 � 0.38 c
0.2 � 0.33 a 12.7 � 0.71 b 14.5 � 0.43 b
0.5 � 0.47 a 16.5 � 0.72 c 16.0 � 0.41 c
1.4 � 0.42 b 10.6 � 0.50 ab 14.6 � 0.38 c
8.7 � 0.36 c 7.2 � 0.35 ab 11.0 � 0.42 d
0.7 � 0.31 a 13.3 � 0.70 b 15.0 � 0.34 b
m each other (P < 0.05).
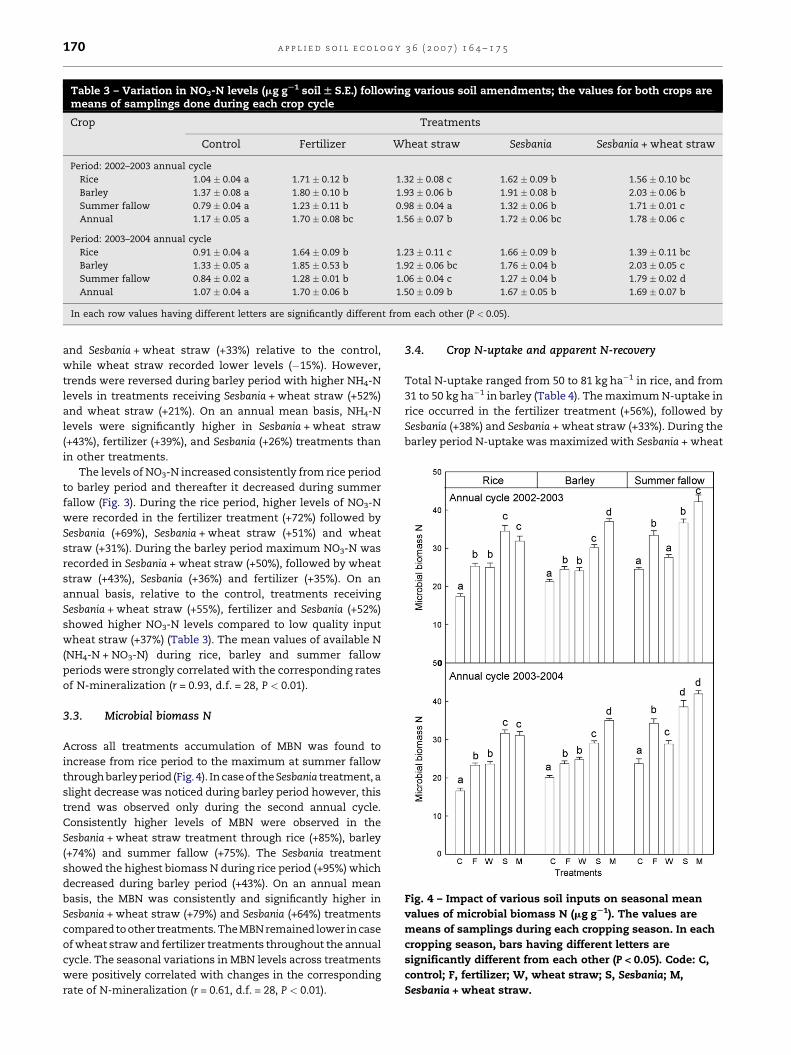
Table 3 – Variation in NO3-N levels (mg gS1 soil W S.E.) following various soil amendments; the values for both crops aremeans of samplings done during each crop cycle
Crop Treatments
Control Fertilizer Wheat straw Sesbania Sesbania + wheat straw
Period: 2002–2003 annual cycle
Rice 1.04 � 0.04 a 1.71 � 0.12 b 1.32 � 0.08 c 1.62 � 0.09 b 1.56 � 0.10 bc
Barley 1.37 � 0.08 a 1.80 � 0.10 b 1.93 � 0.06 b 1.91 � 0.08 b 2.03 � 0.06 b
Summer fallow 0.79 � 0.04 a 1.23 � 0.11 b 0.98 � 0.04 a 1.32 � 0.06 b 1.71 � 0.01 c
Annual 1.17 � 0.05 a 1.70 � 0.08 bc 1.56 � 0.07 b 1.72 � 0.06 bc 1.78 � 0.06 c
Period: 2003–2004 annual cycle
Rice 0.91 � 0.04 a 1.64 � 0.09 b 1.23 � 0.11 c 1.66 � 0.09 b 1.39 � 0.11 bc
Barley 1.33 � 0.05 a 1.85 � 0.53 b 1.92 � 0.06 bc 1.76 � 0.04 b 2.03 � 0.05 c
Summer fallow 0.84 � 0.02 a 1.28 � 0.01 b 1.06 � 0.04 c 1.27 � 0.04 b 1.79 � 0.02 d
Annual 1.07 � 0.04 a 1.70 � 0.06 b 1.50 � 0.09 b 1.67 � 0.05 b 1.69 � 0.07 b
In each row values having different letters are significantly different from each other (P < 0.05).
Fig. 4 – Impact of various soil inputs on seasonal mean
values of microbial biomass N (mg gS1). The values are
means of samplings during each cropping season. In each
cropping season, bars having different letters are
significantly different from each other (P < 0.05). Code: C,
control; F, fertilizer; W, wheat straw; S, Sesbania; M,
Sesbania + wheat straw.
a p p l i e d s o i l e c o l o g y 3 6 ( 2 0 0 7 ) 1 6 4 – 1 7 5170
and Sesbania + wheat straw (+33%) relative to the control,
while wheat straw recorded lower levels (�15%). However,
trends were reversed during barley period with higher NH4-N
levels in treatments receiving Sesbania + wheat straw (+52%)
and wheat straw (+21%). On an annual mean basis, NH4-N
levels were significantly higher in Sesbania + wheat straw
(+43%), fertilizer (+39%), and Sesbania (+26%) treatments than
in other treatments.
The levels of NO3-N increased consistently from rice period
to barley period and thereafter it decreased during summer
fallow (Fig. 3). During the rice period, higher levels of NO3-N
were recorded in the fertilizer treatment (+72%) followed by
Sesbania (+69%), Sesbania + wheat straw (+51%) and wheat
straw (+31%). During the barley period maximum NO3-N was
recorded in Sesbania + wheat straw (+50%), followed by wheat
straw (+43%), Sesbania (+36%) and fertilizer (+35%). On an
annual basis, relative to the control, treatments receiving
Sesbania + wheat straw (+55%), fertilizer and Sesbania (+52%)
showed higher NO3-N levels compared to low quality input
wheat straw (+37%) (Table 3). The mean values of available N
(NH4-N + NO3-N) during rice, barley and summer fallow
periods were strongly correlated with the corresponding rates
of N-mineralization (r = 0.93, d.f. = 28, P < 0.01).
3.3. Microbial biomass N
Across all treatments accumulation of MBN was found to
increase from rice period to the maximum at summer fallow
through barley period (Fig. 4). In case of theSesbania treatment, a
slight decrease was noticed during barley period however, this
trend was observed only during the second annual cycle.
Consistently higher levels of MBN were observed in the
Sesbania + wheat straw treatment through rice (+85%), barley
(+74%) and summer fallow (+75%). The Sesbania treatment
showed the highest biomass N during rice period (+95%) which
decreased during barley period (+43%). On an annual mean
basis, the MBN was consistently and significantly higher in
Sesbania + wheat straw (+79%) and Sesbania (+64%) treatments
compared to other treatments. The MBN remained lower in case
of wheat straw and fertilizer treatments throughout the annual
cycle. The seasonal variations in MBN levels across treatments
were positively correlated with changes in the corresponding
rate of N-mineralization (r = 0.61, d.f. = 28, P < 0.01).
3.4. Crop N-uptake and apparent N-recovery
Total N-uptake ranged from 50 to 81 kg ha�1 in rice, and from
31 to 50 kg ha�1 in barley (Table 4). The maximum N-uptake in
rice occurred in the fertilizer treatment (+56%), followed by
Sesbania (+38%) and Sesbania + wheat straw (+33%). During the
barley period N-uptake was maximized with Sesbania + wheat

Table 4 – Crop N-uptake (kg N haS1) of rice and barley under different soil inputs; the values are mean W S.E.
Treatments 2002–2003 annual cycle 2003–2004 annual cycle
Rice Barley Rice Barley
Control 50.4 � 0.70 a 30.5 � 1.35 a 51.5 � 1.59 a 30.8 � 1.51 a
Fertilizer 76.4 � 1.90 b 35.2 � 1.30 a 81.0 � 1.20 b 37.3 � 1.02 b
Wheat straw 55.0 � 2.70 a 45.0 � 1.75 b 57.6 � 2.76 a 45.7 � 2.13 c
Sesbania 70.1 � 3.38 c 36.1 � 1.89 a 71.1 � 0.58 c 36.9 � 0.74 b
Sesbania + wheat straw 66.9 � 0.93 c 48.4 � 2.79 b 69.0 � 4.78 c 49.9 � 1.18 c
In each column values having different letters are significantly different from each other (P < 0.05).
a p p l i e d s o i l e c o l o g y 3 6 ( 2 0 0 7 ) 1 6 4 – 1 7 5 171
straw (+60%) followed by wheat straw (+48%). Apparent N-
recovery of added N by the crops from the soil was found to be
higher in Sesbania + wheat straw and fertilizer treatments
than in the wheat straw treatment (Table 5).
3.5. Soil moisture content
Soil moisture content followed a seasonal pattern influenced
by precipitation with higher soil water content (13.4–14.9%)
during rice period (rainy season), thereafter decreasing to the
minimum (6.5–7.9%) during summer fallow (Fig. 5). Soil
moisture was not affected (P < 0.05) by treatments throughout
the annual cycle. The seasonal variations in moisture content
across treatments were positively correlated with the corre-
sponding rate of N-mineralization (r = 0.76, d.f. = 28, P < 0.01).
4. Discussion
4.1. Soil N-mineralization rate and microbial biomass N
A marked seasonal variation was observed in soil N-miner-
alization rate, which was higher in the wet season crop (rice)
relative to the dry season crop (barley). During the wet season
the higher soil moisture level, among several factors, seems to
be the chief determinant responsible for greater N-miner-
alization rates. Throughout the annual cycle, across all the
treatments, N-mineralization was found to be influenced
strongly by soil moisture (r = 0.76). After the rainy season, due
to drop in soil moisture during the rest of the annual crop
cycle, there was a decrease in N-mineralization in winter and
summer. Ghoshal (2002) also found strong correlation
between N-mineralization rate and soil moisture content
(r = 0.86, P < 0.01). Available-N concentrations in tropical soils
Table 5 – Annual apparent N-recovery (%) in rice andbarley (combined) under different soil inputs; the valuesare mean W S.E.
Treatments 2002–2003annual cycle
2003–2004annual cycle
Fertilizer 40.8 � 2.4 a 45.1 � 0.1 a
Wheat straw 23.9 � 5.6 b 26.4 � 6.1 b
Sesbania 31.6 � 2.3 ab 32.2 � 0.9 ab
Sesbania + wheat straw 43.2 � 3.3 a 45.4 � 7.3 a
In each column values having different letters are significantly
different from each other (P < 0.05).
fluctuate considerably with seasonal changes in soil water
potential (Wong and Nortcliff, 1995). Rewetting of a dry soil has
been noticed as a major factor for the acceleration of N-
mineralization especially under semi-arid and subtropical
conditions (Dalal and Mayer, 1986). Inaccessible soil organic
matter and dead N-rich microbial cells become accessible to
the microorganisms as a new pool of readily mineralizable N
due to drying and rewetting (Campbell and Biederbeck, 1982).
The N-mineralization rate during the rice period may also
have been enhanced by tillage, which disrupts soil aggregates,
exposing the native soil organic matter to microbes.
The rate of N mineralization varied among treatments,
especially in the early phase of the annual cycle, even though
Fig. 5 – Variation in soil moisture content (%) during various
crop and fallow periods through two annual cycles. Values
are means of samplings during each cropping season.
Seasonal means were not significantly different from each
other (P < 0.05). Code: C, control; F, fertilizer; W, wheat
straw; S, Sesbania; M, Sesbania + wheat straw.

a p p l i e d s o i l e c o l o g y 3 6 ( 2 0 0 7 ) 1 6 4 – 1 7 5172
an equivalent amount of N was applied through all the
exogenous inputs. In the plots receiving Sesbania and fertilizer
the pattern of N-mineralization followed the classical pattern
of initial rapid release followed by a slower linear release
(Fig. 1). Such progress of N-mineralization with time was also
recorded by El-Harris et al. (1983) and Bonde and Rosswall
(1987). Fertilizer application significantly increased the N-
mineralization rate in this study, in agreement with early
studies (Singh and Singh, 1994) in a dryland agroecosystem
(24–66% increase relative to the control); probably due to ready
availability of the added nutrients. According to Woods et al.
(1987), this increased rate of N-mineralization may reflect the
priming effect of fertilizer N on indigenous soil N. Sesbania
having a low C:N ratio decomposed rapidly (instantaneous
decay constant, k = 0.028 day�1, t95 = 109 days; Singh et al.,
2007) after application resulting in a high rate of N-miner-
alization during rice period. Sesbania decomposed completely
in around 120 days which resulted in a low rate of N-
mineralization during barley period. Aulakh et al. (2000) also
observed an increase in N-mineralization rate when Sesbania
was applied in a rice–wheat crop rotation.
The wheat straw treatment showed an initial lag phase in N
release, reflecting early immobilization of N by the microbial
biomass. During the later phase of the annual cycle the
immobilized N was re-mineralized and resulted in a shift of
dominance from immobilization to mineralization. Mubarak
et al. (2001) similarly reported N-immobilization shortly after
incorporating crop residue with high C:N ratio; when soil was
incubated with straw alone, about 30% of the straw N was held
in the microbial biomass within 5 days. When inorganic N was
added with the straw, the consumption of straw N by the
biomass decreased slightly. Azmal et al. (1996) observed net
immobilization throughout the incubation period both at 25
and 35 8C, after adding rice straw (C/N Ratio 60) to upland soils.
Residue with less than 1–1.2% N usually immobilizes mineral
N (Vigil and Kissel, 1991). De Neve et al. (2004) observed initial
immobilization followed by re-mineralization of wheat straw
after 50 days of incubation. Khalil et al. (2005) found that,
irrespective of soil type, chicken manure (C:N 10.6) resulted in
net N-mineralization whereas wheat residue (C:N 75.4)
resulted in net N-immobilization.
Palm et al. (2001) suggested that mixing low quality and
high quality organic inputs generally results in a mineraliza-
tion pattern reflecting the weighted average of the two
separate materials. However, in some cases there have been
non-additive nutrient availability patterns from mixes of low
and high quality materials but the results are difficult to
predict and again do not necessarily result in a synchronous
curve of nutrient availability (Mafongoya et al., 1998; Palm and
Rowland, 1997). If a poor quality residue has high amounts of
available C, the capacity for N immobilization may exceed the
amounts of N available from the soil, thereby immobilizing the
N from the accompanying N-rich residue. Thus, for example,
Sakala et al. (2000) observed that N-mineralization from poor
quality maize residues mixed with leaves of pigeonpea was
much less than that which would have been predicted from
the individual amendments.
The effect of mixed application of high and low quality
resources on N-mineralization rate under dryland condi-
tions has rarely been investigated. Mixed application of
Sesbania + wheat straw resulted in a consistently higher rate
of N-mineralization throughout the annual cycle, probably
due to an interactive effect of Sesbania and wheat straw,
which modulated the N-mineralization rate and showed
synergistic behaviour with reference to N-mineralization.
Our data suggest that mixed application resulted in a
gradual release of inorganic N. Moreover, the greater C
availability in the organic system apparently supports a
more active microbial biomass with greater N demand,
thus promoting immobilization and mineralization of NO3
(Burger and Jackson, 2003) in accordance with crop require-
ment. The challenge in managing high soil organic
matter input systems is to balance C and N inputs to
minimize accumulations of NO3, yet avoid high rates of
microbial N immobilization during peak periods of crop N
demand.
The size and activity of the microbial biomass are key
factors controlling the rates of N-mineralization (Paul and
Voroney, 1984; Azmal et al., 1996). In the present study across
all the treatments through the two annual cycles, positive
correlation was observed between the MBN and the N-
mineralization rate (r = 0.61). Similar relations between N in
soil microbial biomass and N-mineralization rate were
recorded by Kushwaha et al. (2000). Bremer and Kessel
(1992) reported that the relationship of N-mineralization to
microbial biomass dynamics depended on the phase of
decomposition and concluded that the microbial N was the
main source of mineralized N. The amount and temporal
distribution of rainfall strongly influences soil microbial
activity and the fluctuation of mineralizable N in tropical
soils. Amounts of N released from crop residue depend on
microbial immobilization/mineralization as influenced by
crop residue type, placement, and degree of incorporation
into the soil (Aulakh et al., 1991). Nyberg et al. (2002) suggested
that high quality residues, like Sesbania and Crotolaria,
supplying easily accessible C to the microbial community,
can deplete soil oxygen, and create local anaerobic conditions,
stimulating denitrification (Aulakh et al., 2000).
Management practices can influence biological nitrogen
dynamics by their effects on microbial populations. Soils
receiving long-term application of manufactured N fertilizers
often have lower biological activity relative to soils that have
received repeated additions of organic materials (Collins et al.,
1992; Dick et al., 1988; McGill et al., 1986). The diversity of
microbial biomass determines ecological stability and sus-
tained soil productivity in a system; mixed application of
Sesbania + wheat straw, where both low and high quality
resources are available simultaneously, may lead to increased
diversity of the microbial biomass. Low quality resource
application favors slow growing K-strategists over r-strate-
gists for longer duration, however, application of Sesbania may
facilitate rapidly growing r-strategist species on nutrient rich
substrate. In the mixed input treatment presumably both, r-
and K-strategists remain active for longer time periods,
thereby maintaining higher microbial biomass throughout
the annual cycle (Singh et al., 2007; Fontaine et al., 2003). This
could help in build up of SOM and increased cation exchange
capacity and erosion resistance (Young, 1997), minimizing the
risk of losses of the remaining N through leaching, volatiliza-
tion and erosion.

a p p l i e d s o i l e c o l o g y 3 6 ( 2 0 0 7 ) 1 6 4 – 1 7 5 173
4.2. Available-N in soil and apparent N-recovery
In these agroecosystems the dominance of NH4-N was
probably due to the predominance of diverse ammonifying
microbes. In contrast to nitrifying bacteria these microbes can
be retarded by low water potential present in dryland soils,
resulting in high NH4-N levels (Dommergues et al., 1978).
During the rainy season, due to higher soil moisture content,
the rate of biological activity and decomposition are decreased
due to the creation of anaerobic conditions. These anaerobic
conditions can enhance the rate of denitrification resulting in
a lower rate of nitrification during rice cropping compared to
barley cropping (Fig. 2). Aulakh et al. (2000) also reported
higher denitrification under anaerobic soil conditions created
due to high soil moisture content at nearly-saturation and
flooded soil environment. Moreover, under anaerobic condi-
tions, decomposition depends only on anaerobic bacteria
which are less efficient than aerobic organisms (Campbell,
1978; Patrick, 1982). They can function at much lower energy
levels and require low N, resulting in a more rapid release of
NH4+ ions (Patrick, 1982). After the rainy season, with gradually
drying soils, nitrification is favored resulting in a decreased
NH4-N relative to NO3-N. The other possible factor for this
contrast in the seasonal pattern between NO3-N and NH4-N
was probably due to the marked difference in their mobility.
NH4-N is derived from the mineralization of soil organic N,
added organic materials, or addition of urea or ammonium
fertilizer. In contrast to the NH4-N, there is little tendency for
the NO3-N anion to be adsorbed by the soil colloids, which
commonly possess a net negative charge. Nitrate is thus
susceptible to diffusion and mass transport with soil water
(Cameron and Haynes, 1986). Nitrate losses are often larger in
agricultural ecosystems (Morisot, 1981) and leaching is the
most important channel of N loss from cultivated field soils
other than accounted for in crop uptake (Allison, 1973; Legg
and Meisinger, 1982). Dominance of NH4-N is expected to
reduce N losses from these dryland agroecosystems.
The largest proportion of N is found in the soil organic
matter and its availability to plants is dependent on its rate of
mineralization. Rate of N-mineralization largely governed the
availability of N (NH4-N + NO3-N) in the agroecosystem
studied, as evident from the strong relationship between N-
mineralization rate and available-N (r = 0.93). Among the
treatments, available-N level was enhanced in fertilizer,
Sesbania and Sesbania + wheat straw treatments during rice
period, which was in accordance with the enhanced rate of N-
mineralization. Munoz et al. (2003) reported that fertilizer and
manure additions tended to increase soil available-N (parti-
cularly NO3-N). They observed that NO3-N from inorganic
fertilizer was soluble and therefore more susceptible to
downward movement; in comparison, less NO3-N was found
in the lower depths in a manure treatment. Dey and Jain (1997)
reported increased availability of N during the initial period,
due to application of Sesbania, which declined 45 days after
application.
In contrast to fertilizer and Sesbania treatments, low
availability of N in wheat straw treated plots in the early
phase of rice cropping was due to an initial lag of N-
mineralization. During the later phase (barley crop) N became
available due to re-mineralization in the soil resulting in
significantly higher levels of available-N. Ocio et al. (1991)
found no significant immobilization after wheat straw
application. Aulakh et al. (2001) observed a tendency of early
immobilization of straw N which mineralized thereafter later
in the season. They observed that plots receiving Sesbania
showed high mineral N levels for a longer duration, compared
to plots receiving fertilizer where initial low levels and later on
higher levels were found.
The mixed application of Sesbania + wheat straw main-
tained higher levels of available-N throughout the annual
cycle, suggesting that C-rich inputs favor a gradual release of
available-N. Asynchrony occurs when nutrient availability
exceeds plant requirements, often because release occurs at a
time when plant demand is restricted or non-existent, as in
summer fallow or after crop maturity. Apparent N-recovery is
influenced by weather conditions, management practices and
availability of N in soil (Bellido and Bellido, 2001). Mixed
application of Sesbania + wheat straw resulted in higher
apparent N-recovery and N-uptake compared to application
of wheat straw or Sesbania alone, suggesting lower loss of N
through the system in the mixed treatment and thereby
increased utilization of N by the crop. Sesbania mixed with
wheat straw and fertilizer can be effective in replacing
fertilizer, thereby mitigating environmental problems asso-
ciated with its use. However Aulakh et al. (2000) observed
increased apparent N-recovery in Sesbania treatment com-
pared to fertilizer and attributed these results to the rapid
mineralization of Sesbania.
Achieving N synchrony and mitigating the problems of
leaching or denitrification requires a strategy to avoid periods
of nutrient deficiency and excess. The issue of synchrony in
cropping systems is at the root of most environmental hazards
associated with excess N in the atmosphere, terrestrial and
marine ecosystems (Peoples et al., 2004). Few studies have
carefully compared synchrony over a range of cropping
systems. One recent study conducted by Xu et al. (2005)
observed, in a pot experiment with Chinese cabbage as test
crop, that the amount of mineral N was initially highest when
fertilizer was used. Mineral N concentration decreased there-
after to the lowest level in the later stages of cropping. When
untreated rice straw was combined with fertilizer the levels of
mineral N were initially lower but increased later on and were
maintained at a higher level throughout the crop period.
5. Conclusions
This study shows that it is possible to manage the N
immobilization/mineralization process of soil by application
of mixed treatment of Sesbania + wheat straw so as to adjust
the release of available-N in accordance with plant demands.
Lack of synchrony, i.e., asynchrony, occurred in the plots
receiving fertilizer and also with either high or low quality
organic resources. The additional gain in N-mineralization, N-
uptake and apparent N-recovery in the mixed treatment
demonstrated synergy between the two N sources. Consistent
and prolonged availability of N in combined application of
Sesbania + wheat straw indicated a step towards achieving
synchronization which may lead to better nutrient conserva-
tion and reduced losses from the agroecosystem.

a p p l i e d s o i l e c o l o g y 3 6 ( 2 0 0 7 ) 1 6 4 – 1 7 5174
Acknowledgements
We thank the Head and the Programme Co-ordinator, Centre
of Advanced Study in Botany, Department of Botany, for
providing laboratory facilities. University Grants Commission,
New Delhi, India, provided financial support in form of a major
research project which included a Project Fellowship to Mr. S.
Singh. Dr. H.H. Janzen, Agriculture and Agri-Food Canada and
Dr. Kathrin Franzluebbers, Texas A&M University gave helpful
suggestions on the manuscript, for this we are grateful.
r e f e r e n c e s
Allison, F.E., 1973. Soil Organic Matter and its Role in CropProduction. Elsevier, Amsterdam, p. 637.
American Public Health Association (APHA), 1995. StandardMethods of Examination of Water and Wastewater, 19thed. American Public Health Association, Washington, DC,USA.
Aulakh, M.S., Doran, J.W., Walters, D.T., Mosier, A.R., Francis,D.D., 1991. Crop residue type and placement effects ondenitrification and mineralization. Soil Sci. Soc. Am. J. 55,1020–1025.
Aulakh, M.S., Khera, T.S., Doran, J.W., 2000. Mineralization anddenitrification in upland, nearly saturated and floodedsubtropical soil II. Effect of organic manures varyingin N content and C:N ratio. Biol. Fertil. Soils 31,168–174.
Aulakh, M.S., Khera, T.S., Doran, J.W., Bronson, K.F., 2001.Denitrification, N2O and CO2 fluxes in rice–wheat system asaffected by crop residues, fertilizer N and legume greenmanure. Biol. Fertil. Soils 34, 375–389.
Azmal, A.K.M., Marumoto, T., Shindo, H., Nishiyama, M., 1996.Mineralization and changes in microbial biomass in water-saturated soil amended with some tropical plant residues.Soil Sci. Plant Nutr. 42, 483–492.
Beare, M.H., Wilson, P.E., Fraser, P.M., Butler, R.C., 2002.Management effects on barley straw decomposition,nitrogen release, and crop production. Soil Sci. Soc. Am. J.66, 848–856.
Bellido, R.J.L., Bellido, L.L., 2001. Efficiency of nitrogen in wheatunder Mediterranean conditions: effect of tillage, croprotation and N fertilization. Field Crops Res. 71, 31–46.
Bonde, T.A., Rosswall, T., 1987. Seasonal variations ofpotentially mineralizable nitrogen in four croppingsystems. Soil Sci. Soc. Am. J. 51, 1508–1514.
Bremer, E., Kessel, C.V., 1992. Plant available nitrogen fromlentil and wheat residues during a subsequent growingseason. Soil Sci. Soc. Am. J. 56, 1155–1160.
Brookes, P.C., Landman, A., Pruden, G., Jenkinson, D.S., 1985.Chloroform fumigation and release of soil N: a rapid directextraction-method to measure microbial biomass nitrogenin soil. Soil Biol. Biochem. 17, 837–842.
Burger, M., Jackson, L.E., 2003. Microbial immobilization ofammonium and nitrate in relation to ammonification andnitrification rates in organic and conventional croppingsystems. Soil Biol. Biochem. 35, 29–36.
Cameron, K.C., Haynes, R.J., 1986. Retention and movement ofnitrogen in soils. In: Haynes, R.J. (Ed.), Mineral Nitrogen inthe Plant–Soil System. Academic Press Inc., pp. 166–241.
Campbell, C.A., 1978. Soil organic carbon, nitrogen and fertility.In: Schnitzer, M., Khan, S.U. (Eds.), Soil Organic Matter. Am.Elsevier, New York, pp. 173–271.
Campbell, C.A., Biederbeck, V.O., 1982. Changes in mineral Nand numbers of bacteria and actinomycetes during two
years under wheat fallow in southwestern Saskatchewan.Can. J. Soil Sci. 62, 125–137.
Campbell, C.A., Myers, R.J.K., Curtin, D., 1995. Managingnitrogen for sustainable crop production. Fertil. Res. 42,277–296.
Collins, H.P., Rasmussen, P.E., Douglas, C.L., 1992. Crop rotationand residue management effects on soil carbon andmicrobial biomass dynamics. Soil Sci. Soc. Am. J. 56,783–788.
Dalal, R.C., Mayer, R.J., 1986. Long-term trends in fertility of soilsunder continuous cultivation and cereal cropping insouthern Queensland. III. Distribution and kinetics of soilorganic carbon in particle size fractions. Austr. J. Soil Res.24, 293–300.
De Neve, S., Gaona Saez, S., Daguilar, B.C., Sleutel, S., Hofman,G., 2004. Manipulating N mineralization from high N cropresidues using on- and off-farm organic materials. Soil Biol.Biochem. 36, 127–134.
Dey, P., Jain, J.M., 1997. Mineralization and nitrification in soilamended with urea and enriched green manures insubmerged soil system. J. Ind. Soc. Soil Sci. 45, 249–255.
Dick, R.P., Rasmussen, P.E., Kerle, E.A., 1988. Influence of long-term residue management on soil enzyme activities inrelation to soil chemical properties of a wheat-fallowsystem. Biol. Fertil. Soils 6, 159–164.
Dilz, K., 1988. Efficiency of uptake and utilization of fertilizernitrogen by plants. In: Jenkinson, D.S., Smith, K.A. (Eds.),Nitrogen Efficiency in Agricultural Soils. Elsevier AppliedScience, London, pp. 1–26.
Dommergues, R.Y.R., Belser, L.W., Schwidt, E.L., 1978. Limitingfactors for microbial growth and activity in soil. In:Alexander, M. (Ed.), Advances in Microbial Ecology, vol. 2.Plenum Press, New York, pp. 49–104.
El-Harris, M.K., Cochran, V.L., Elliott, L.F., Bezdicek, D.F., 1983.Effect of tillage, cropping and fertilizer management on soilnitrogen mineralization potential. Soil Sci. Soc. Am. J. 47,1157–1161.
Eno, C.F., 1960. Nitrate production in the field by incubating thesoil in polyethlene bags. Soil Sci. Soc. Am. Proc. 24, 277–279.
Fontaine, S., Mariotti, A., Abbadie, L., 2003. The priming effect oforganic matter: a question of microbial competition? Soil.Biol. Biochem. 35, 837–843.
Ghoshal, N., 2002. Available pool and mineralization rate of soilN in a dryland agroecosystem: effect of organic soilamendments and chemical fertilizer. Trop. Ecol. 43, 363–366.
Giller, K.E., Chalk, P., Dobermann, A., Hammond, L., Heffer, P.,Ladha, J.K., Nyamudeza, P., Maene, L., Ssali, H., Freney, J.,2004. Emerging technologies to increase the efficiency ofuse of fertilizer nitrogen. In: Mosier, A.R., Syers, K.J., Freney,J.R. (Eds.), Agriculture and the Nitrogen Cycle, The ScientificCommittee on Problems of the Environment (SCOPE). IslandPress, Covelo, California, USA, pp. 35–51.
Goulding, K., 2004. Pathways and losses of fertilizer nitrogen atdifferent scales. In: Mosier, A.R., Syers, K.J., Freney, J.R.(Eds.), Agriculture and the Nitrogen Cycle, The ScientificCommittee on Problems of the Environment (SCOPE). IslandPress, Covelo, California, USA, pp. 209–219.
Groffman, P.M., Hendrix, P.F., Crossley D.A.Jr., 1987. Nitrogendynamics in conventional and no-tillage agroecosystemswith inorganic fertilizer or legume nitrogen inputs. PlantSoil 97, 315–332.
Hadas, A., Kautsky, L., Goek, M., Kara, E.E., 2004. Rates ofdecomposition of plant residues and available nitrogen insoil, related to residue composition through simulation ofcarbon and nitrogen turnover. Soil Biol. Biochem. 36,255–266.
Hoyle, F.C., Murphy, D.V., Fillery, I.R.P., 2006. Temperature andstubble management influence microbial CO2-C evolution

a p p l i e d s o i l e c o l o g y 3 6 ( 2 0 0 7 ) 1 6 4 – 1 7 5 175
and gross N transformation rates. Soil Biol. Biochem. 38,71–80.
Jackson, M.L., 1973. Soil Chemical Analysis. Prentice Hall ofIndia Limited, New Delhi.
Janzen, H.H., Beauchemin, K.A., Bruinsma, Y., Campbell, C.A.,Desjardins, R.L., Ellert, B.H., Smith, E.G., 2003. The fate ofnitrogen in agroecosystems: an illustration using Canadianestimates. Nutr. Cycl. Agroecosys. 67, 85–102.
Khalil, M.I., Hossain, M.B., Schmidhalter, U., 2005. Carbon andnitrogen mineralization in different upland soils of thesubtropics treated with organic materials. Soil Biol.Biochem. 37, 1507–1518.
Kushwaha, C.P., Tripathi, S.K., Singh, K.P., 2000. Variations insoil microbial biomass and N availability due to residue andtillage management in a dryland rice agro-ecosystem. SoilTillage Res. 56, 153–166.
Kushwaha, C.P., Singh, K.P., 2005. Crop productivity and soilfertility in a tropical dryland agro-ecosystem: impact ofresidue and tillage management. Expl. Agric. 41, 39–50.
Legg, J.O., Meisinger, J.J., 1982. Soil nitrogen budgets. In:Stevenson, F.J. (Ed.), Nitrogen in Agricultural Soils.American Society of Agronomy, Madison, Wisconsin, pp.503–557.
Mafongoya, P.L., Nair, P.K.R., Dzowela, B.H., 1998. Mineralizationof nitrogen from decomposing leaves of multipurpose treesas affected by their chemical composition. Biol. Fertil. Soils27, 143–148.
Matson, P.A., Parton, W.J., Power, A.G., Swift, M.J., 1997.Agricultural intensification and ecosystem properties.Science 227, 504–508.
McGill, W.B., Cannon, K.R., Robertson, J.A., Cook, F.D., 1986.Dynamics of soil microbial biomass and water-solubleorganic C in Breton L after 50 years of cropping to tworotations. Can. J. Soil Sci. 66, 1–19.
Morisot, A., 1981. Erosion and nitrogen losses. In: Clark, F.E.,Rosswall, T. (Eds.), Terrestrial Nitrogen Cycles: Processes,Ecosystem strategies and management Impacts. EcologicalBulletin, Stockholm, pp. 353–361.
Mubarak, A.R., Rosenani, A.B., Zayyah, D.S., Anuar, A.R., 2001.Nitrogen mineralization in tropical soils amended with cropresidues. Trop. Agric. 78, 165–173.
Munoz, G.R., Powell, J.M., Kelling, K.A., 2003. Nitrogen budgetand soil N dynamics after multiple applications ofunlabeled or 15nitrogen-enriched dairy manure. Soil Sci.Soc. Am. J. 67, 817–825.
Myers, R.J.K., Palm, C.A., Cuevas, E., Gunatilleke, I.U.N.,Brossard, M., 1994. The synchronisation of nutrientmineralization and plant nutrient demand. In: Woomer,P.I., Swift, M.J. (Eds.), The Biological Management ofTropical Soils. Wiley-Saye, Chichester, New York, USA, pp.81–116.
Nyberg, G., Ekblad, A., Buresh, R., Hogberg, P., 2002. Short-termpatterns of carbon and nitrogen mineralization in a fallowfield amended with green manures from agroforestry trees.Biol Fertil Soils 36, 18–25.
Ocio, J.A., Brookes, P.C., Jenkinson, D.S., 1991. Fieldincorporation of straw and its effects on soil microbialbiomass and soil inorganic N. Soil Biol. Biochem. 23, 171–176.
Palm, C.A., Giller, K.E., Mafongoya, P.L., Swift, M.J., 2001.Management of organic matter in the tropics: translatingtheory into practice. Nutr. Cycl. Agroecosys. 61, 63–75.
Palm, C.A., Rowland, A.P., 1997. A minimum dataset forcharacterization of plant quality for decomposition. In:
Cadisch, G., Giller, K.E. (Eds.), Driven by Nature: Plant LitterQuality and Decomposition. CAB International, Wallingford,pp. 379–393.
Patrick, W.H., 1982. Nitrogen transformations in submergedsoils. In: Stevenson, F.J. (Ed.), Nitrogen in Agricultural Soils.American Society of Agronomy, Madison, Wisconsin, pp.449–466.
Paul, E.A., Voroney, R.P., 1984. Biomass and activitymeasurements. In: Klug, M.J., Reddy, C.A. (Eds.), CurrentPerspectives in Microbial Ecology, Proceedings of the ThirdInternational Symposium on Microbial Ecology. MichiganState University, American Society for Microbiology,Michigan, USA, pp. 509–514.
Peoples, M.B., Boyer, E.W., Goulding, K.W.T., Heffer, P., Ochwoh,V.A., Vanlauwe, B., Wood, S., Yagi, K., Van Cleemput, O.,2004. Pathways of nitrogen loss and their impacts onhuman health and the environment. In: Mosier, A.R., Syers,K.J., Freney, J.R. (Eds.), Agriculture and the Nitrogen Cycle,The Scientific Committee on Problems of the Environment(SCOPE). Island Press, Covelo, California, USA, pp. 53–69.
Peoples, M.B., Freney, J.R., Mosier, A.R., 1995. Minimizinggaseous losses of nitrogen. In: Bacon, P.E. (Ed.), NitrogenFertilization in the Environment. Marcel Dekker Inc., NewYork, pp. 565–602.
Robertson, G.P., Paul, E.A., Harwood, R.R., 2000. Greenhousegases in intensive agriculture: contributions of individualgases to the radiative forcing of the atmosphere. Science289, 1300–1922.
Sakala, W.D., Cadisch, G., Giller, K.E., 2000. Interactions betweenresidues of maize and pigeonpea and mineral N fertilizersduring decomposition and N mineralization. Soil Biol.Biochem. 32, 679–688.
Singh, H., Singh, K.P., 1994. Nitrogen and phosphorusavailability and mineralization in dryland reduced tillagecultivation: effects of residue placement and chemicalfertilizer. Soil Biol. Biochem. 26, 695–702.
Singh, S., Ghoshal, N., Singh, K.P., 2007. Variations in soilmicrobial biomass and crop roots due to differing resourcequality inputs in a tropical dryland agroecosystem. Soil Biol.Biochem. 39, 76–86.
Tilman, D., 1998. The greening of the green revolution. Nature396, 211–212.
Tilman, D., Cassman, K.G., Matson, P.A., Naylor, R., Polasky, S.,2002. Agricultural sustainability and intensive productionpractices. Nature 418, 671–677.
Venterea, R.T., Rolston, D.E., 2000. Nitric and nitrous oxideemissions following fertilizer application to agriculturalsoil: biotic and abiotic mechanisms and kinetics. J. Geophys.Res. 105, 15117–15129.
Vigil, M.F., Kissel, D.E., 1991. Equations for estimating theamount of nitrogen mineralized from crop residues. SoilSci. Soc. Am. J. 55, 757–761.
Wong, M.T.F., Nortcliff, S., 1995. Seasonal fluctuations of nativeavailable N and soil management implications. Fertil. Res.42, 3–26.
Woods, L.E., Cole, C.V., Porter, L.K., Coleman, D.C., 1987.Transformation of added and indigenous nitrogen ingenobiotic soil: a comment on the priming effect. Soil Biol.Biochem. 19, 673–678.
Xu, Y., Zhang, Y., Shen, Q., Xu, Y., Zhang, J., 2005. An innovativemethod for the treatment of rice straw to improve nitrogenuptake efficiency. Biol. Fertil. Soils 41, 291–294.
Young, A., 1997.In: Agroforestry for soil management. 2nd ed.CAB, Wallingford, UK, p. 320.
Copyright © 2022 FDOKUMEN




















