
A Vineyard Agroecosystem: Disturbance and Precipitation Affect Soil Respiration under Mediterranean...
9
SSSAJ: Volume 74: Number 1 • January–February 2010 231 Soil Sci. Soc. Am. J. 74:231–239 Published online 13 Nov. 2009 doi:10.2136/sssaj2008.0346 Received 29 Oct. 2008. *Corresponding author ([email protected]). © Soil Science Society of America, 677 S. Segoe Rd., Madison WI 53711 USA All rights reserved. No part of this periodical may be reproduced or transmitted in any form or by any means, electronic or mechanical, including photocopying, recording, or any information storage and retrieval system, without permission in writing from the publisher. Permission for printing and for reprinting the material contained herein has been obtained by the publisher. A Vineyard Agroecosystem: Disturbance and Precipitation Affect Soil Respiration under Mediterranean Conditions Soil & Water Management & Conservation T he soil organic carbon (SOC) pool is the third largest global C pool and is strongly influenced by agricultural management practices (Amundson, 2001). As compared with undisturbed systems, agricultural soils have lost up to 50% of C from the plow layer (e.g., Carlisle et al., 2006), and for this reason provide an opportunity for C sequestration. Management of agricultural soils using practices such as growing cover crops and replacing conventional tillage with minimum tillage and no-till practices has been shown to lead to enhanced soil C sequestra- tion (Kern and Johnson, 1993; Six et al., 1999; Duiker and Lal, 2000; Grandy and Robertson, 2007). e application of such practices to increase sequestration of C in the SOC pool has received increasing attention with the aim of mitigating greenhouse gas emissions (Smith et al., 1998; Martens et al., 2005), but informa- tion is limited with respect to perennial cropping systems such as grape, and for arid and semiarid climates such as those of the Mediterranean climatic zones. Soil respiration is a key component of the terrestrial C budget and is com- posed of the oxidation of organic matter by microorganisms, and root and rhizo- sphere respiration (Hanson et al., 2001). Production of CO 2 during respiration is the ultimate oxidative fate of the SOC pool and represents the primary loss of C from terrestrial ecosystems (Schlesinger and Andrews, 2000). e loss of CO 2 from the SOC pool through respiration is typically much greater than soil C losses from other processes such as leaching and erosion (Trumbore, 1997). Agricultural Kerri L. Steenwerth* USDA-ARS Crops Pathology and Genetics Res. Unit Dep. of Viticulture and Enology Univ. of California, Davis 595 Hilgard Ln. Davis, CA 95616 Danielle L. Pierce Eli A. Carlisle Dep. of Viticulture and Enology Univ. of California, Davis 595 Hilgard Ln. Davis, CA 95616 Robert G. M. Spencer Dep. of Plant Sciences Univ. of California, Davis One Shields Ave Davis, CA 95616 David R. Smart Dep. of Viticulture and Enology Univ. of California, Davis 595 Hilgard Ln. Davis, CA 95616 We investigated impacts of agricultural management practices on soil respiration (R s ) in a Cabernet Sauvignon (Vitis vinifera) vineyard (Oakville, CA; November 2003–December 2005). We determined (i) response of R s to cover cropping, mowing and tillage; (ii) environmental factors controlling R s ; and (iii) total annual C lost through R s . A winter cover crop was either mown (CC+mow), or mown and tilled (CC+Till), and resident vegetation was tilled (RV+Till). Precipitation amount and pattern differed between years, and low R s rates occurred during summer drought and high rates during wet periods. Total CO 2 emissions differed only in Year 2 (RV+Till: 10.99 ± 0.30, CC+Till: 10.11 ± 0.49, CC+mow: 8.57 ± 0.54 Mg CO 2 –C ha −1 ). Aſter tillage or mowing, R s increased five- to six-fold in tilled treatments and two-fold in the mown treatment (Year 1). In Year 2, R s increased two- to three-fold aſter tillage only. R s increased aſter ‘post-management’ rainfall in spring and was 1.5- to 2-fold greater in tilled than mown treatments in both years due to prior incorporation of plant biomass during tillage. Aſter fall rainfall, R s was 1.7-fold greater in mown than tilled treatments (Year 1). Our findings suggest that the interaction of management practice with climate and soil conditions before disturbance (i.e., management and rainfall) influenced R s . Polynomial regressions of R s on GWC and soil temperature indicated that R s increased until gravimetric water content (GWC) reached 14 to 15% in tilled treatments and 20% in the mown treatment, subsequently declining, indicating thresholds of GWC at which soil temperature more strongly influences R s . Abbreviations: CC, cover crop; CO 2 , carbon dioxide; GWC, gravimetric water content, R s , soil respiration; RV, resident vegetation; SOC, soil organic carbon.
Transcript of A Vineyard Agroecosystem: Disturbance and Precipitation Affect Soil Respiration under Mediterranean...

SSSAJ: Volume 74: Number 1 • January–February 2010
231
Soil Sci. Soc. Am. J. 74:231–239Published online 13 Nov. 2009 doi:10.2136/sssaj2008.0346Received 29 Oct. 2008.*Corresponding author ([email protected]).© Soil Science Society of America, 677 S. Segoe Rd., Madison WI 53711 USAAll rights reserved. No part of this periodical may be reproduced or transmitted in any form or by any means, electronic or mechanical, including photocopying, recording, or any information storage and retrieval system, without permission in writing from the publisher. Permission for printing and for reprinting the material contained herein has been obtained by the publisher.
A Vineyard Agroecosystem: Disturbance and Precipitation Affect Soil Respiration
under Mediterranean Conditions
Soil & Water Management & Conservation
The soil organic carbon (SOC) pool is the third largest global C pool and is strongly infl uenced by agricultural management practices (Amundson, 2001).
As compared with undisturbed systems, agricultural soils have lost up to 50% of C from the plow layer (e.g., Carlisle et al., 2006), and for this reason provide an opportunity for C sequestration. Management of agricultural soils using practices such as growing cover crops and replacing conventional tillage with minimum tillage and no-till practices has been shown to lead to enhanced soil C sequestra-tion (Kern and Johnson, 1993; Six et al., 1999; Duiker and Lal, 2000; Grandy and Robertson, 2007). Th e application of such practices to increase sequestration of C in the SOC pool has received increasing attention with the aim of mitigating greenhouse gas emissions (Smith et al., 1998; Martens et al., 2005), but informa-tion is limited with respect to perennial cropping systems such as grape, and for arid and semiarid climates such as those of the Mediterranean climatic zones.
Soil respiration is a key component of the terrestrial C budget and is com-posed of the oxidation of organic matter by microorganisms, and root and rhizo-sphere respiration (Hanson et al., 2001). Production of CO2 during respiration is the ultimate oxidative fate of the SOC pool and represents the primary loss of C from terrestrial ecosystems (Schlesinger and Andrews, 2000). Th e loss of CO2 from the SOC pool through respiration is typically much greater than soil C losses from other processes such as leaching and erosion (Trumbore, 1997). Agricultural
Kerri L. Steenwerth* USDA-ARSCrops Pathology and Genetics Res. UnitDep. of Viticulture and EnologyUniv. of California, Davis595 Hilgard Ln.Davis, CA 95616
Danielle L. PierceEli A. Carlisle
Dep. of Viticulture and EnologyUniv. of California, Davis595 Hilgard Ln.Davis, CA 95616
Robert G. M. SpencerDep. of Plant Sciences Univ. of California, DavisOne Shields AveDavis, CA 95616
David R. SmartDep. of Viticulture and Enology Univ. of California, Davis595 Hilgard Ln.Davis, CA 95616
We investigated impacts of agricultural management practices on soil respiration (Rs) in a Cabernet Sauvignon (Vitis vinifera) vineyard (Oakville, CA; November 2003–December 2005). We determined (i) response of Rs to cover cropping, mowing and tillage; (ii) environmental factors controlling Rs; and (iii) total annual C lost through Rs. A winter cover crop was either mown (CC+mow), or mown and tilled (CC+Till), and resident vegetation was tilled (RV+Till). Precipitation amount and pattern diff ered between years, and low Rs rates occurred during summer drought and high rates during wet periods. Total CO2 emissions diff ered only in Year 2 (RV+Till: 10.99 ± 0.30, CC+Till: 10.11 ± 0.49, CC+mow: 8.57 ± 0.54 Mg CO2–C ha−1). Aft er tillage or mowing, Rs increased fi ve- to six-fold in tilled treatments and two-fold in the mown treatment (Year 1). In Year 2, Rs increased two- to three-fold aft er tillage only. Rs increased aft er ‘post-management’ rainfall in spring and was 1.5- to 2-fold greater in tilled than mown treatments in both years due to prior incorporation of plant biomass during tillage. Aft er fall rainfall, Rs was 1.7-fold greater in mown than tilled treatments (Year 1). Our fi ndings suggest that the interaction of management practice with climate and soil conditions before disturbance (i.e., management and rainfall) infl uenced Rs. Polynomial regressions of Rs on GWC and soil temperature indicated that Rs increased until gravimetric water content (GWC) reached 14 to 15% in tilled treatments and 20% in the mown treatment, subsequently declining, indicating thresholds of GWC at which soil temperature more strongly infl uences Rs.
Abbreviations: CC, cover crop; CO2, carbon dioxide; GWC, gravimetric water content, Rs, soil respiration; RV, resident vegetation; SOC, soil organic carbon.

232 SSSAJ: Volume 74: Number 1 • January–February 2010
CO2 emissions are thought to contribute a signifi cant propor-tion of the global anthropogenic CO2 fl ux (approximately 26%; Zhang et al., 2002). Th us, the utilization of cover crops and/or no-till practices to increase soil C-sequestration will depend on how such practices infl uence SOC oxidation.
Most studies addressing the eff ects of agricultural manage-ment practices on SOC and Rs have been conducted in annual cropping systems of continental climates where irrigation is sel-dom used (e.g., Six et al., 1999; Halvorson et al., 2002; Grandy and Robertson, 2007). Less is known about C cycle processes in Mediterranean climates like that of California and for peren-nial crops, such as grape (Carlisle et al., 2006). Reports from Mediterranean climates that currently exist have focused primar-ily on C cycle processes in annual agroecosystems or natural sys-tems (e.g., Andrews et al., 2002; Rey et al., 2002; Jackson et al., 2003; Asensio et al., 2007; Veenstra et al., 2007; Álvaro-Fuentes et al., 2008). Nonetheless, soil C content associated with spe-cifi c ecosystems may infl uence Rs emissions. Comparatively high CO2 emissions occurred in cover cropped vineyards, and these emissions were associated with higher total soil C and labile C content as compared with frequently tilled soils (Steenwerth and Belina, 2008). Disturbance by tillage and rainfall cause sharp in-creases in Rs, and these responses can vary by management prac-tice and land use (Reicosky et al., 1997; Calderón et al., 2000, 2001; Calderón and Jackson, 2002; Rey et al., 2005; Steenwerth et al., 2005). For example, grasslands with greater soil C content exhibit greater respiration rates aft er precipitation or tillage than their agricultural counterparts with depauperate soil C content (Calderón et al., 2000; Steenwerth et al., 2005). Soil water con-tent before disturbance can likewise infl uence Rs rate post-dis-turbance (Fierer and Schimel, 2002; Rey et al., 2005).
Th e area supporting perennial crops increased in California from 1950–2000 by 0.4 × 106 ha while area in annual crop pro-duction decreased. Th is increase may provide greater opportu-nity for carbon sequestration as management of perennial crops disturbs soils less than annual crop management (NASS, 1950–2000). Additionally, with more than 375,000 ha of vineyards, 393, 000 ha of nuts, and nearly 250,000 ha of non-citrus peren-nial orchards currently under cultivation in California (USDA–NASS, 2009), and only an estimated 16% of grape acreage sown to cover crops (Ingels and Klonsky, 1998), there is much promise for enhancing regional C storage in these soils.
Due to the need to establish baseline information concern-ing the relationships between management practices and Rs in perennial agricultural systems in Mediterranean climates, ef-fects of vineyard fl oor management practices on Rs were inves-tigated in response to disturbance (i.e., mowing and/or tillage) and cover crops over 2 yr in a Cabernet Sauvignon vineyard in Napa County, California. Th e onset of mandatory monitoring of CO2 emissions is anticipated for California in 2012 as set forth in the California Global Warming Solutions Act of 2006 (State Assembly Bill 32), and lends greater practical credence to characterizing these processes. Our objectives in the current in-vestigation were to (i) compare the magnitude of the response of
Rs to mowing and tillage in vineyard alleys, (ii) determine envi-ronmental drivers of CO2 emissions, and (iii) estimate the total annual C lost through Rs among treatments. We hypothesized that Rs would increase in response to management practices (Hypothesis 1) and aft er rainfall (Hypothesis 2), but that these responses could vary with respect to management practice and the soil conditions under which the disturbance (i.e., manage-ment practice and rainfall) occurred (Hypothesis 3).
MATERIALS AND METHODSExperimental Design and Site Description
Th e vineyard site was located at the University of California Davis Oakville Research Station in Napa Valley, California (latitude 38° 25’ 55” N, longitude 122° 24’ 48” W). It was planted in 1994 to Cabernet Sauvignon (Clone 8, Vitis vinifera). Vines were placed at 1.8 m by 2.4 m spacing, with 30 vines per row. Th e experimental design was a ran-domized complete block, with three blocks and one replicate of each vineyard fl oor treatment per block (n = 3), with two subsamples mea-sured per replicate. Each vineyard fl oor treatment per block was there-fore composed of four adjacent alleys (518 m2). Th e outer alleys served as guard rows while the inner alleys were used for data collection. Two of the three vineyard fl oor treatments consisted of a winter cover crop, short stature barley (cv. UC603, Hordeum vulgare), that was mown, leaving residues on the soil surface (CC + mow) or mown and disked to a 30-cm depth (CC + till) in spring. Before drill seeding the barley (78 kg ha−1 in 2003, 156 kg ha−1 in 2004), the soil was disked (upper 5 cm) and rolled in late fall (November 2003 and 2004). Th e third vine-yard fl oor treatment was composed of resident vegetation (RV+Till), which was dominated by non-native annual grasses and forbs common to California grasslands, including Capsella bursa-pastoris (L.) Medik., Bromus hordeaceus L. ssp. hordeauceus, Vulpia myuros (L.) C.C. Gmel., Brassica nigra (L.) W.D.J. Koch, and Bromus diandrus Roth ssp. rigidus (Roth) Lainz. Th e RV + till was mown and tilled at the same time as CC + mow and CC + till (29 Mar. 2004; 1 Apr. 2005). Tilling and mow-ing operations spanned the width of the cover crop or resident vegeta-tion (1.9 m wide), while bare ground remained under the vine row (0.5 m wide). Measurements for all variables were only taken in the region where the vegetation grew between the vine rows and not within the vine row.
Northern California has a Mediterranean climate that consists of hot, dry summers and cool, moist winters. Mean monthly maximum and minimum temperatures for the last 15 yr were 22.8 and 7.0°C, respectively (Oakville Station, California Irrigation Management Information System [CIMIS], 2005). In 2004 and 2005 the mean annu-al temperature was 15.9 and 16.0°C, respectively. Th e soil was classifi ed as a Bale loam (Fine-loamy, mixed, thermic Cumulic Ultic Haploxerolls; Lambert, 1978). General soil characteristics (0–20 cm) were 33% sand, 42% silt, and 25% clay, pH 5.6 (saturated paste method), and soil bulk density of 1.22 ± 0.023 g cm−3 (n = 9, bulk density only).
Weather MeasurementsDaily and monthly precipitation, and mean daily air temperatures,
and maximum and minimum air temperatures by month were reported from CIMIS (Oakville Station) (Table 1). Although Rs data were col-
SSSAJ: Volume 74: Number 1 • January–February 2010 233
lected from January 2004 through December 2005, these weather data are reported from August 2003 to December 2005 as annual precipitation amounts in this region are nor-mally cataloged beginning August fi rst of each year.
Field MeasurementsTo develop an estimate of annual soil C lost through Rs,
soil CO2 emissions were measured in situ every 2 to 4 wk from January 2004 through December 2005 (i.e., 44 dates) using a nondispersive infrared gas analyzer with an attached Rs chamber (Model LI-6400/6400–09; LI-COR, Lincoln, NE). Soil CO2 emissions were measured by fi tting the Rs chamber to polyvinyl chloride (PVC) rings (10 cm high by 10 cm in diam.), which were placed in the alley of each treat-ment replicate at the beginning of the experiment to a depth of 5 cm and remained in place throughout sampling (n = 3 per treatment). Th e only exception to PVC ring permanency occurred when rings were removed during mowing and till-ing and then gently returned to their original positions at least 24 h before sampling. Th is allowed us to measure emis-sions directly following these events while minimizing, as much as possible, artifacts caused by soil disturbance during ring placement. During Rs measurements, soil temperature was measured (0–20 cm) using the soil probe connected to the LI6400/6400–09 soil CO2 fl ux system (LI-COR, Inc., Lincoln, NE). Gravimetric soil water content was collected from the 0- to 20-cm depth at the time of Rs measurements, mixed, and at least 50 g (wet weight) was dried at 105°C for 72 h. Plant biomass was collected periodically from quad-rats (25 cm by 167 cm) placed across the width of the alley. Total soil C content (0–20 cm) was measured by combustion (Pella, 1990).
Statistical AnalysisTh e two subsamples within a treatment replicate
on a given date were averaged before analysis of variance (ANOVA). Data for soil temperature, water content, and respiration were analyzed by ANOVA using a mixed model for repeated measures (PROC MIXED; SAS v. 8.2, Cary, NC) with the covariance structure autoregressive averaging (AR1). Eff ects of year, management, year × management, date (year), and date × management (year) were test-ed. Multiple comparisons were conducted to test a priori hypotheses (see above) and corrected using the Bonferroni correction (P < 0.05). Nonlinear multiple regression (Sigmaplot v. 9.0, Systat Soft ware, San Jose, CA) was used to identify relationships among soil temperature, moisture, and respiration. To determine whether a full or reduced re-gression model was required, we used a log likelihood ratio test and de-termined signifi cance using Pearson’s chi-square distribution (P < 0.05). Aft er determining that a reduced model was necessary to incorporate both date and treatment eff ects, the same approach was employed to compare individual treatments to each other. Type I error resulting from multiple comparisons was controlled by using the Bonferroni correction (P < 0.05). Total CO2 emissions from each treatment were determined by graphing each treatment replicate separately, integrating
the area under these curves, analyzing diff erences by ANOVA (general linear model for repeated measures, n = 3) and using Tukey’s test for mean separation (P < 0.05). Comparisons between monthly and annual rainfall follow Álvaro-Fuentes et al. (2008).
RESULTSPrecipitation, Soil Water Content, Soil Temperature, and Cover Crop Biomass
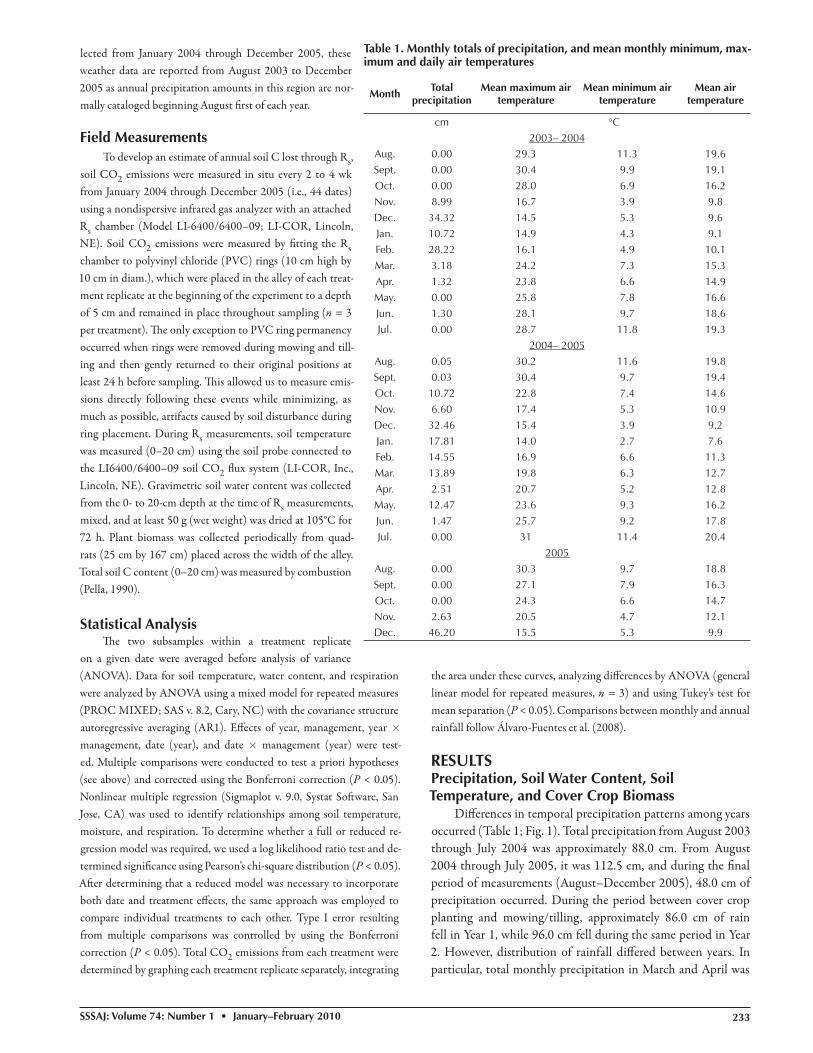
Diff erences in temporal precipitation patterns among years occurred (Table 1; Fig. 1). Total precipitation from August 2003 through July 2004 was approximately 88.0 cm. From August 2004 through July 2005, it was 112.5 cm, and during the fi nal period of measurements (August–December 2005), 48.0 cm of precipitation occurred. During the period between cover crop planting and mowing/tilling, approximately 86.0 cm of rain fell in Year 1, while 96.0 cm fell during the same period in Year 2. However, distribution of rainfall diff ered between years. In particular, total monthly precipitation in March and April was
Table 1. Monthly totals of precipitation, and mean monthly minimum, max-imum and daily air temperatures
MonthTotal
precipitation Mean maximum air
temperature Mean minimum air
temperature Mean air
temperature
cm °C2003– 2004
Aug. 0.00 29.3 11.3 19.6
Sept. 0.00 30.4 9.9 19.1
Oct. 0.00 28.0 6.9 16.2
Nov. 8.99 16.7 3.9 9.8
Dec. 34.32 14.5 5.3 9.6
Jan. 10.72 14.9 4.3 9.1
Feb. 28.22 16.1 4.9 10.1
Mar. 3.18 24.2 7.3 15.3
Apr. 1.32 23.8 6.6 14.9
May. 0.00 25.8 7.8 16.6
Jun. 1.30 28.1 9.7 18.6
Jul. 0.00 28.7 11.8 19.3
2004– 2005
Aug. 0.05 30.2 11.6 19.8
Sept. 0.03 30.4 9.7 19.4
Oct. 10.72 22.8 7.4 14.6
Nov. 6.60 17.4 5.3 10.9
Dec. 32.46 15.4 3.9 9.2
Jan. 17.81 14.0 2.7 7.6
Feb. 14.55 16.9 6.6 11.3
Mar. 13.89 19.8 6.3 12.7
Apr. 2.51 20.7 5.2 12.8
May. 12.47 23.6 9.3 16.2
Jun. 1.47 25.7 9.2 17.8
Jul. 0.00 31 11.4 20.4
2005
Aug. 0.00 30.3 9.7 18.8
Sept. 0.00 27.1 7.9 16.3
Oct. 0.00 24.3 6.6 14.7
Nov. 2.63 20.5 4.7 12.1Dec. 46.20 15.5 5.3 9.9
234 SSSAJ: Volume 74: Number 1 • January–February 2010
four- and two-fold greater in Year 2 than Year 1, respectively, and mean monthly, maximum, and minimum air temperatures in these months were all 2 to 4°C higher in Year 1 than Year 2. No rain fell in May of Year 1 whereas 12.5 cm fell in Year 2. In other words, nearly 93% of the total annual rainfall in Year 1 had oc-curred by the end of February, whereas only 70% had occurred at this same time in Year 2. Aft er the summer drought period, the fi rst month with signifi cant rainfall was October in Year 1 and November in Year 2. Furthermore, October (Year 1) had greater mean monthly air temperatures than in November (Year 2).
Temporal patterns in soil water content and tempera-ture were similar among treatments (Fig. 1 and 2). However, CC+mow tended to have soil water contents that were 1 to 2% (g H2O g−1 × 100) lower than the tilled treatments aft er im-posing tillage and mowing practices (Fig. 2; Table 2). Th e lower soil water contents in CC+mow corresponded to regrowth and tillering by the mown cover crop (personal observation). Soil temperature in the spring was 2 to 6°C greater in the mown than tilled treatments aft er imposing the respective management prac-tices in Year 1 but not in Year 2 (Fig. 1; P < 0.05).
Th e cover crops and particularly resident vegetation ac-quired more biomass earlier in Year 2 than Year 1 (Table 3). Peak aboveground biomass was greater in Year 2 than in Year 1 in all treatments including resident, non-native vegetation in RV+Till (P < 0.05). Two days before mowing or tilling in Year
1, above ground biomass in CC+mow and CC+till was four-fold greater than in RV+till (P < 0.05). However, aboveground biomass in RV+Till had decreased by half during the preceding 3 wk (P < 0.05). Annual grasses and forbs, such as those in RV+Till, will set seed and senesce with decreasing soil water content (Pitt and Heady, 1977). In Year 2, in which rainfall occurred later in the spring than in Year 1, peak aboveground biomass was greatest in CC+Till, but did not diff er between CC+mow and RV+Till (P < 0.05). Total soil C did not diff er between years or treatments. Hence, the grand mean of percentage of soil C was 1.64 ± 0.08 g C g−1 dry soil (n = 36) (data not shown).
Effects of Management Practice on Soil Respiration (Hypothesis 1)
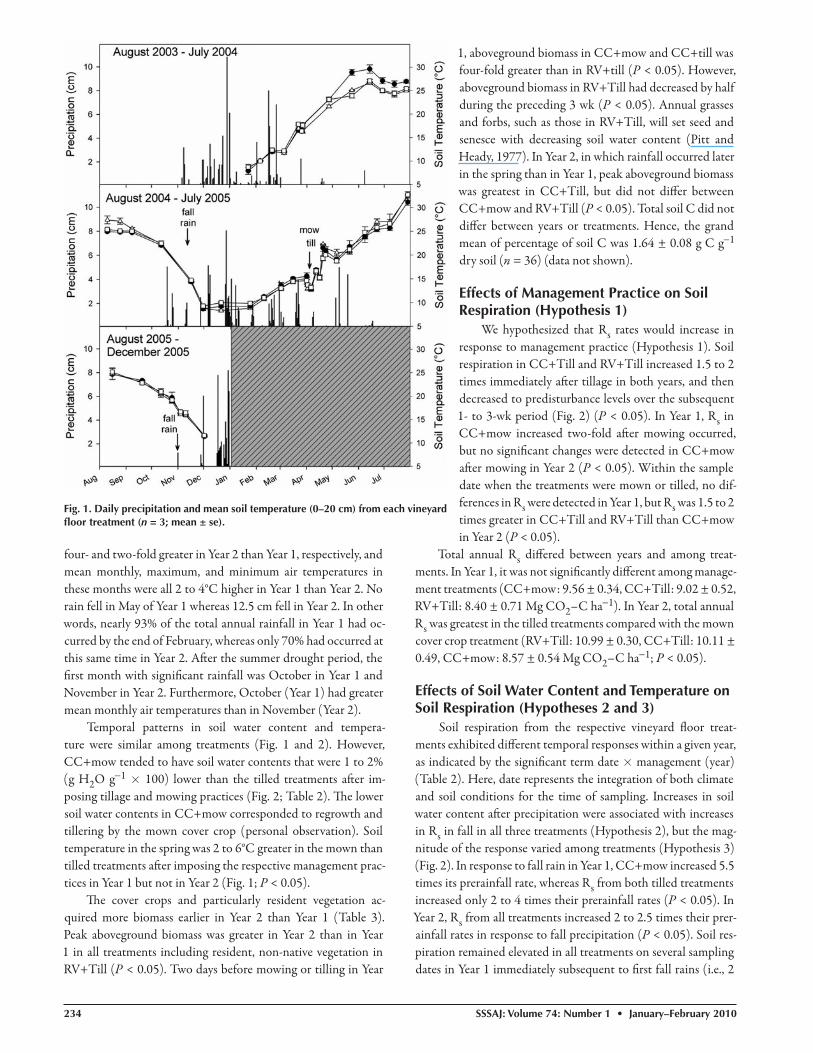
We hypothesized that Rs rates would increase in response to management practice (Hypothesis 1). Soil respiration in CC+Till and RV+Till increased 1.5 to 2 times immediately aft er tillage in both years, and then decreased to predisturbance levels over the subsequent 1- to 3-wk period (Fig. 2) (P < 0.05). In Year 1, Rs in CC+mow increased two-fold aft er mowing occurred, but no signifi cant changes were detected in CC+mow aft er mowing in Year 2 (P < 0.05). Within the sample date when the treatments were mown or tilled, no dif-ferences in Rs were detected in Year 1, but Rs was 1.5 to 2 times greater in CC+Till and RV+Till than CC+mow in Year 2 (P < 0.05).
Total annual Rs diff ered between years and among treat-ments. In Year 1, it was not signifi cantly diff erent among manage-ment treatments (CC+mow: 9.56 ± 0.34, CC+Till: 9.02 ± 0.52, RV+Till: 8.40 ± 0.71 Mg CO2–C ha−1). In Year 2, total annual Rs was greatest in the tilled treatments compared with the mown cover crop treatment (RV+Till: 10.99 ± 0.30, CC+Till: 10.11 ± 0.49, CC+mow: 8.57 ± 0.54 Mg CO2–C ha−1; P < 0.05).
Effects of Soil Water Content and Temperature on Soil Respiration (Hypotheses 2 and 3)
Soil respiration from the respective vineyard fl oor treat-ments exhibited diff erent temporal responses within a given year, as indicated by the signifi cant term date × management (year) (Table 2). Here, date represents the integration of both climate and soil conditions for the time of sampling. Increases in soil water content aft er precipitation were associated with increases in Rs in fall in all three treatments (Hypothesis 2), but the mag-nitude of the response varied among treatments (Hypothesis 3) (Fig. 2). In response to fall rain in Year 1, CC+mow increased 5.5 times its prerainfall rate, whereas Rs from both tilled treatments increased only 2 to 4 times their prerainfall rates (P < 0.05). In Year 2, Rs from all treatments increased 2 to 2.5 times their prer-ainfall rates in response to fall precipitation (P < 0.05). Soil res-piration remained elevated in all treatments on several sampling dates in Year 1 immediately subsequent to fi rst fall rains (i.e., 2
Fig. 1. Daily precipitation and mean soil temperature (0–20 cm) from each vineyard fl oor treatment (n = 3; mean ± se).
SSSAJ: Volume 74: Number 1 • January–February 2010 235
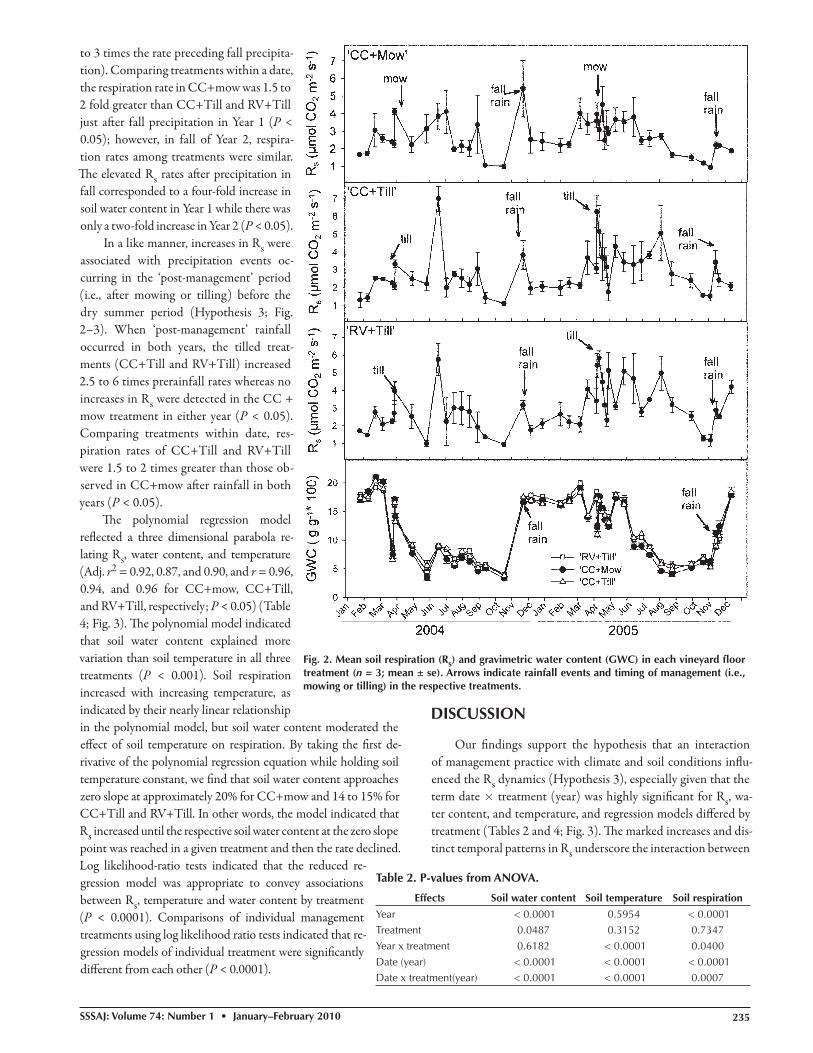
to 3 times the rate preceding fall precipita-tion). Comparing treatments within a date, the respiration rate in CC+mow was 1.5 to 2 fold greater than CC+Till and RV+Till just aft er fall precipitation in Year 1 (P < 0.05); however, in fall of Year 2, respira-tion rates among treatments were similar. Th e elevated Rs rates aft er precipitation in fall corresponded to a four-fold increase in soil water content in Year 1 while there was only a two-fold increase in Year 2 (P < 0.05).
In a like manner, increases in Rs were associated with precipitation events oc-curring in the ‘post-management’ period (i.e., aft er mowing or tilling) before the dry summer period (Hypothesis 3; Fig. 2–3). When ‘post-management’ rainfall occurred in both years, the tilled treat-ments (CC+Till and RV+Till) increased 2.5 to 6 times prerainfall rates whereas no increases in Rs were detected in the CC + mow treatment in either year (P < 0.05). Comparing treatments within date, res-piration rates of CC+Till and RV+Till were 1.5 to 2 times greater than those ob-served in CC+mow aft er rainfall in both years (P < 0.05).
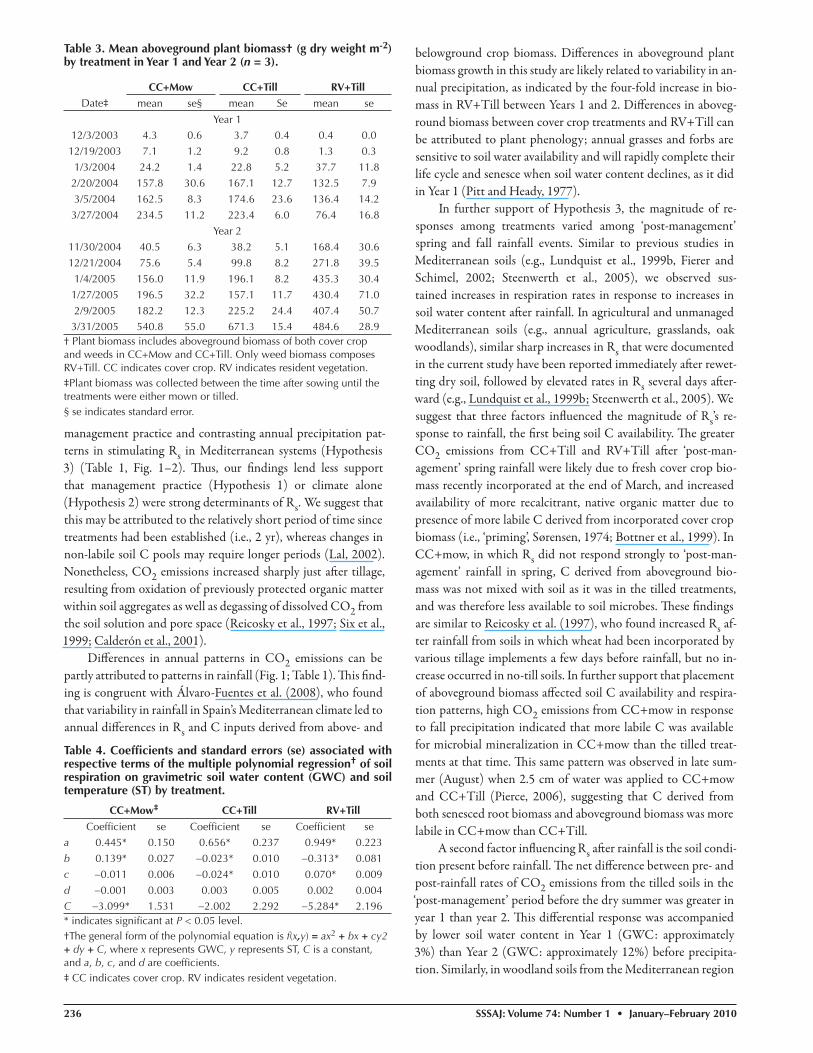
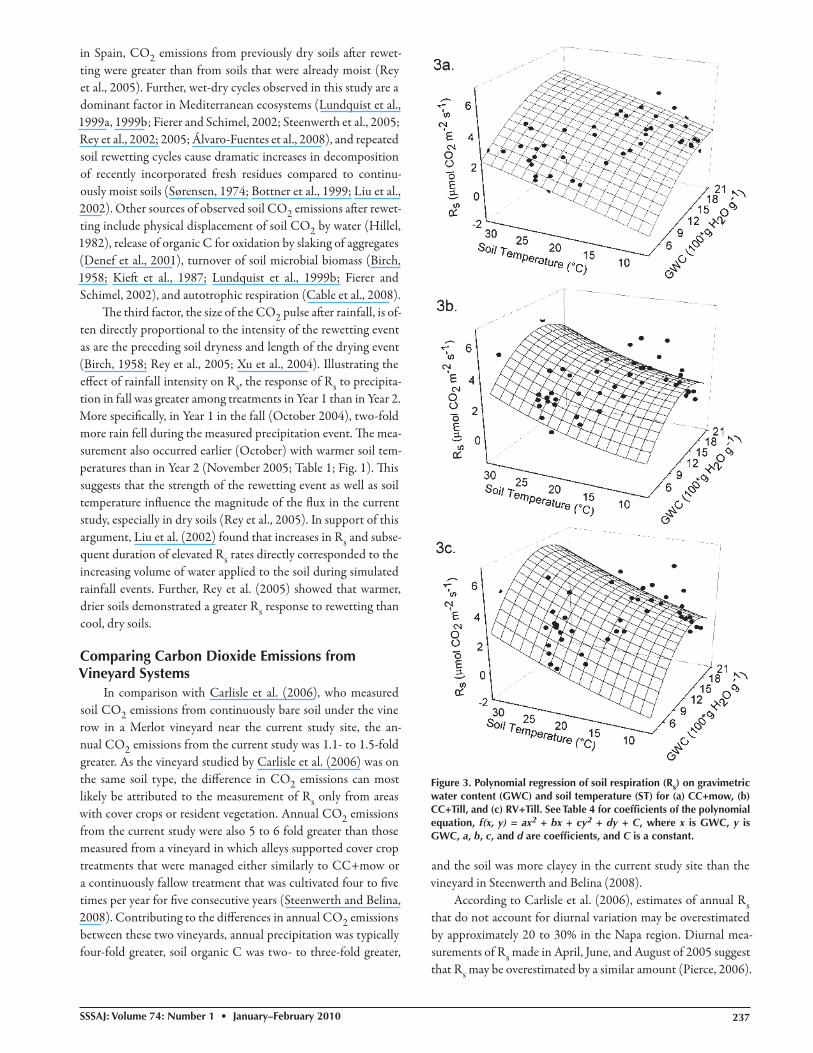
Th e polynomial regression model refl ected a three dimensional parabola re-lating Rs, water content, and temperature (Adj. r2 = 0.92, 0.87, and 0.90, and r = 0.96, 0.94, and 0.96 for CC+mow, CC+Till, and RV+Till, respectively; P < 0.05) (Table 4; Fig. 3). Th e polynomial model indicated that soil water content explained more variation than soil temperature in all three treatments (P < 0.001). Soil respiration increased with increasing temperature, as indicated by their nearly linear relationship in the polynomial model, but soil water content moderated the eff ect of soil temperature on respiration. By taking the fi rst de-rivative of the polynomial regression equation while holding soil temperature constant, we fi nd that soil water content approaches zero slope at approximately 20% for CC+mow and 14 to 15% for CC+Till and RV+Till. In other words, the model indicated that Rs increased until the respective soil water content at the zero slope point was reached in a given treatment and then the rate declined. Log likelihood-ratio tests indicated that the reduced re-gression model was appropriate to convey associations between Rs, temperature and water content by treatment (P < 0.0001). Comparisons of individual management treatments using log likelihood ratio tests indicated that re-gression models of individual treatment were signifi cantly diff erent from each other (P < 0.0001).
DISCUSSION
Our fi ndings support the hypothesis that an interaction of management practice with climate and soil conditions infl u-enced the Rs dynamics (Hypothesis 3), especially given that the term date × treatment (year) was highly signifi cant for Rs, wa-ter content, and temperature, and regression models diff ered by treatment (Tables 2 and 4; Fig. 3). Th e marked increases and dis-tinct temporal patterns in Rs underscore the interaction between
Fig. 2. Mean soil respiration (Rs) and gravimetric water content (GWC) in each vineyard fl oor treatment (n = 3; mean ± se). Arrows indicate rainfall events and timing of management (i.e., mowing or tilling) in the respective treatments.
Table 2. P-values from ANOVA.
Effects Soil water content Soil temperature Soil respiration
Year < 0.0001 0.5954 < 0.0001Treatment 0.0487 0.3152 0.7347
Year x treatment 0.6182 < 0.0001 0.0400
Date (year) < 0.0001 < 0.0001 < 0.0001Date x treatment(year) < 0.0001 < 0.0001 0.0007
236 SSSAJ: Volume 74: Number 1 • January–February 2010
management practice and contrasting annual precipitation pat-terns in stimulating Rs in Mediterranean systems (Hypothesis 3) (Table 1, Fig. 1–2). Th us, our fi ndings lend less support that management practice (Hypothesis 1) or climate alone (Hypothesis 2) were strong determinants of Rs. We suggest that this may be attributed to the relatively short period of time since treatments had been established (i.e., 2 yr), whereas changes in non-labile soil C pools may require longer periods (Lal, 2002). Nonetheless, CO2 emissions increased sharply just aft er tillage, resulting from oxidation of previously protected organic matter within soil aggregates as well as degassing of dissolved CO2 from the soil solution and pore space (Reicosky et al., 1997; Six et al., 1999; Calderón et al., 2001).
Diff erences in annual patterns in CO2 emissions can be partly attributed to patterns in rainfall (Fig. 1; Table 1). Th is fi nd-ing is congruent with Álvaro-Fuentes et al. (2008), who found that variability in rainfall in Spain’s Mediterranean climate led to annual diff erences in Rs and C inputs derived from above- and
belowground crop biomass. Diff erences in aboveground plant biomass growth in this study are likely related to variability in an-nual precipitation, as indicated by the four-fold increase in bio-mass in RV+Till between Years 1 and 2. Diff erences in aboveg-round biomass between cover crop treatments and RV+Till can be attributed to plant phenology; annual grasses and forbs are sensitive to soil water availability and will rapidly complete their life cycle and senesce when soil water content declines, as it did in Year 1 (Pitt and Heady, 1977).
In further support of Hypothesis 3, the magnitude of re-sponses among treatments varied among ‘post-management’ spring and fall rainfall events. Similar to previous studies in Mediterranean soils (e.g., Lundquist et al., 1999b, Fierer and Schimel, 2002; Steenwerth et al., 2005), we observed sus-tained increases in respiration rates in response to increases in soil water content aft er rainfall. In agricultural and unmanaged Mediterranean soils (e.g., annual agriculture, grasslands, oak woodlands), similar sharp increases in Rs that were documented in the current study have been reported immediately aft er rewet-ting dry soil, followed by elevated rates in Rs several days aft er-ward (e.g., Lundquist et al., 1999b; Steenwerth et al., 2005). We suggest that three factors infl uenced the magnitude of Rs’s re-sponse to rainfall, the fi rst being soil C availability. Th e greater CO2 emissions from CC+Till and RV+Till aft er ‘post-man-agement’ spring rainfall were likely due to fresh cover crop bio-mass recently incorporated at the end of March, and increased availability of more recalcitrant, native organic matter due to presence of more labile C derived from incorporated cover crop biomass (i.e., ‘priming’, Sørensen, 1974; Bottner et al., 1999). In CC+mow, in which Rs did not respond strongly to ‘post-man-agement’ rainfall in spring, C derived from aboveground bio-mass was not mixed with soil as it was in the tilled treatments, and was therefore less available to soil microbes. Th ese fi ndings are similar to Reicosky et al. (1997), who found increased Rs af-ter rainfall from soils in which wheat had been incorporated by various tillage implements a few days before rainfall, but no in-crease occurred in no-till soils. In further support that placement of aboveground biomass aff ected soil C availability and respira-tion patterns, high CO2 emissions from CC+mow in response to fall precipitation indicated that more labile C was available for microbial mineralization in CC+mow than the tilled treat-ments at that time. Th is same pattern was observed in late sum-mer (August) when 2.5 cm of water was applied to CC+mow and CC+Till (Pierce, 2006), suggesting that C derived from both senesced root biomass and aboveground biomass was more labile in CC+mow than CC+Till.
A second factor infl uencing Rs aft er rainfall is the soil condi-tion present before rainfall. Th e net diff erence between pre- and post-rainfall rates of CO2 emissions from the tilled soils in the ‘post-management’ period before the dry summer was greater in year 1 than year 2. Th is diff erential response was accompanied by lower soil water content in Year 1 (GWC: approximately 3%) than Year 2 (GWC: approximately 12%) before precipita-tion. Similarly, in woodland soils from the Mediterranean region
Table 3. Mean aboveground plant biomass† (g dry weight m-2) by treatment in Year 1 and Year 2 (n = 3).
CC+Mow CC+Till RV+Till
Date‡ mean se§ mean Se mean se
Year 112/3/2003 4.3 0.6 3.7 0.4 0.4 0.0
12/19/2003 7.1 1.2 9.2 0.8 1.3 0.3
1/3/2004 24.2 1.4 22.8 5.2 37.7 11.8
2/20/2004 157.8 30.6 167.1 12.7 132.5 7.9
3/5/2004 162.5 8.3 174.6 23.6 136.4 14.2
3/27/2004 234.5 11.2 223.4 6.0 76.4 16.8
Year 2
11/30/2004 40.5 6.3 38.2 5.1 168.4 30.6
12/21/2004 75.6 5.4 99.8 8.2 271.8 39.5
1/4/2005 156.0 11.9 196.1 8.2 435.3 30.4
1/27/2005 196.5 32.2 157.1 11.7 430.4 71.0
2/9/2005 182.2 12.3 225.2 24.4 407.4 50.73/31/2005 540.8 55.0 671.3 15.4 484.6 28.9
† Plant biomass includes aboveground biomass of both cover crop and weeds in CC+Mow and CC+Till. Only weed biomass composes RV+Till. CC indicates cover crop. RV indicates resident vegetation.‡Plant biomass was collected between the time after sowing until the treatments were either mown or tilled.§ se indicates standard error.
Table 4. Coeffi cients and standard errors (se) associated with respective terms of the multiple polynomial regression† of soil respiration on gravimetric soil water content (GWC) and soil temperature (ST) by treatment.
CC+Mow‡ CC+Till RV+Till
Coeffi cient se Coeffi cient se Coeffi cient sea 0.445* 0.150 0.656* 0.237 0.949* 0.223
b 0.139* 0.027 −0.023* 0.010 −0.313* 0.081
c −0.011 0.006 −0.024* 0.010 0.070* 0.009
d −0.001 0.003 0.003 0.005 0.002 0.004C −3.099* 1.531 −2.002 2.292 −5.284* 2.196* indicates signifi cant at P < 0.05 level.†The general form of the polynomial equation is f(x,y) = ax2 + bx + cy2 + dy + C, where x represents GWC, y represents ST, C is a constant, and a, b, c, and d are coeffi cients.‡ CC indicates cover crop. RV indicates resident vegetation.
SSSAJ: Volume 74: Number 1 • January–February 2010 237
in Spain, CO2 emissions from previously dry soils aft er rewet-ting were greater than from soils that were already moist (Rey et al., 2005). Further, wet-dry cycles observed in this study are a dominant factor in Mediterranean ecosystems (Lundquist et al., 1999a, 1999b; Fierer and Schimel, 2002; Steenwerth et al., 2005; Rey et al., 2002; 2005; Álvaro-Fuentes et al., 2008), and repeated soil rewetting cycles cause dramatic increases in decomposition of recently incorporated fresh residues compared to continu-ously moist soils (Sørensen, 1974; Bottner et al., 1999; Liu et al., 2002). Other sources of observed soil CO2 emissions aft er rewet-ting include physical displacement of soil CO2 by water (Hillel, 1982), release of organic C for oxidation by slaking of aggregates (Denef et al., 2001), turnover of soil microbial biomass (Birch, 1958; Kieft et al., 1987; Lundquist et al., 1999b; Fierer and Schimel, 2002), and autotrophic respiration (Cable et al., 2008).
Th e third factor, the size of the CO2 pulse aft er rainfall, is of-ten directly proportional to the intensity of the rewetting event as are the preceding soil dryness and length of the drying event (Birch, 1958; Rey et al., 2005; Xu et al., 2004). Illustrating the eff ect of rainfall intensity on Rs, the response of Rs to precipita-tion in fall was greater among treatments in Year 1 than in Year 2. More specifi cally, in Year 1 in the fall (October 2004), two-fold more rain fell during the measured precipitation event. Th e mea-surement also occurred earlier (October) with warmer soil tem-peratures than in Year 2 (November 2005; Table 1; Fig. 1). Th is suggests that the strength of the rewetting event as well as soil temperature infl uence the magnitude of the fl ux in the current study, especially in dry soils (Rey et al., 2005). In support of this argument, Liu et al. (2002) found that increases in Rs and subse-quent duration of elevated Rs rates directly corresponded to the increasing volume of water applied to the soil during simulated rainfall events. Further, Rey et al. (2005) showed that warmer, drier soils demonstrated a greater Rs response to rewetting than cool, dry soils.
Comparing Carbon Dioxide Emissions from Vineyard Systems
In comparison with Carlisle et al. (2006), who measured soil CO2 emissions from continuously bare soil under the vine row in a Merlot vineyard near the current study site, the an-nual CO2 emissions from the current study was 1.1- to 1.5-fold greater. As the vineyard studied by Carlisle et al. (2006) was on the same soil type, the diff erence in CO2 emissions can most likely be attributed to the measurement of Rs only from areas with cover crops or resident vegetation. Annual CO2 emissions from the current study were also 5 to 6 fold greater than those measured from a vineyard in which alleys supported cover crop treatments that were managed either similarly to CC+mow or a continuously fallow treatment that was cultivated four to fi ve times per year for fi ve consecutive years (Steenwerth and Belina, 2008). Contributing to the diff erences in annual CO2 emissions between these two vineyards, annual precipitation was typically four-fold greater, soil organic C was two- to three-fold greater,
and the soil was more clayey in the current study site than the vineyard in Steenwerth and Belina (2008).
According to Carlisle et al. (2006), estimates of annual Rs that do not account for diurnal variation may be overestimated by approximately 20 to 30% in the Napa region. Diurnal mea-surements of Rs made in April, June, and August of 2005 suggest that Rs may be overestimated by a similar amount (Pierce, 2006).
Figure 3. Polynomial regression of soil respiration (Rs) on gravimetric water content (GWC) and soil temperature (ST) for (a) CC+mow, (b) CC+Till, and (c) RV+Till. See Table 4 for coeffi cients of the polynomial equation, f(x, y) = ax2 + bx + cy2 + dy + C, where x is GWC, y is GWC, a, b, c, and d are coeffi cients, and C is a constant.

238 SSSAJ: Volume 74: Number 1 • January–February 2010
Likewise, Parkin and Kaspar (2003) found that a temperature-re-lated hysteresis in diurnal Rs rates could result in under- or over-estimating annual emissions by 30 to 40%, depending on when daily measurements were collected. Recent fi ndings by Vargas and Allen (2008) indicate that hysteresis can also vary by sea-son and vegetation type, and that in herbaceous cover, seasonal means of respiration can diff er as ±15% from the annual mean. Together, fi ndings from these studies suggest that estimates of annual CO2 emissions in the current study are slightly infl ated.
Polynomial Regression Models and Environmental Drivers of Carbon Dioxide Emissions
Among all treatments, soil water content explained more variation in CO2 emissions than soil temperature. Th is fi nding is consistent with previous studies of Rs in California vineyards and grasslands (Xu et al., 2004; Carlisle et al., 2006; Steenwerth and Belina, 2008). Rey et al. (2005) utilized a polynomial model to predict Rs from soil water content and temperature in a Mediterranean oak forest, and found that soil water con-tent and temperature together explained a substantial amount of variation in soil CO2 emissions. Th eir model was based on data generated during controlled incubations, and its predictive power was limited when applied to fi eld data until pulses of CO2 released during summer wet-dry cycles were accounted for (Rey et al., 2002, 2005). Th is suggests that the poor predictive power of soil temperature in the polynomial model in the current study may be related to pulses of CO2 released during specifi c manage-ment practices and wet-dry cycles, which occurred in the cool, wet winter. Th e poor relationship between Rs and temperature has been shown to be caused by a 2- to 3-h temporal lag between soil CO2 emissions and soil temperature in a number of previ-ous investigations (Eriksen and Jensen, 2001; Parkin and Kaspar, 2003; Gaumont-Guay et al., 2006). Nonetheless, Paul et al. (1999) observed a stronger correlation between Rs and air tem-perature than Rs and soil temperature, suggesting that our overall understanding of the biophysical processes controlling soil CO2 emissions are not well understood.
We suggest that Rs in this vineyard was limited by soil water content in summer, and soil temperature in winter (see Fig. 3). Similar patterns have been documented in a California vineyard in the same region as the current study (Carlisle et al., 2006), as well as Mediterranean forests and semiarid steppe in Spain and Italy (Casals et al., 2000; Rey et al., 2002; Maestre and Cortina, 2003). In in situ measurements in an oak forest under Mediterranean conditions, Rey et al. (2002) found that soil tem-perature played a signifi cant role in controlling Rs when volu-metric water content increased above a threshold value of 20%, but that soil moisture controlled Rs below that value. In the cur-rent study, the regression models suggest that thresholds also ex-ist for this system, but such a threshold may vary with respect to management, as well as predisturbance conditions.
CONCLUSIONSWe have demonstrated that the interaction of interannual
diff erences in climate, the soil conditions before disturbance, and management regime strongly infl uenced Rs in the fi rst 2 yr since treatments were established. In particular, annual patterns in Rs varied with diff erences in precipitation timing and magni-tude, soil water content before rewetting, management events, as well as aboveground biomass growth, underscoring the impor-tance of incorporating these factors into models predicting pat-terns of Rs. We also fi nd that soil water content is a main factor controlling Rs in this vineyard agroecosystem, and that thresh-olds in soil water content at which soil temperature becomes a stronger determinant of Rs exist, a result which is consistent with other studies in Mediterranean ecosystems. Interestingly, the similarity in Rs between the two tilled treatments suggests that it did not matter whether a cover crop was planted or not, but that the presence of tillage did infl uence observed Rs pat-terns. Nonetheless, cover crop selection may become important in situations where certain tolerances to environmental condi-tions (e.g., drought, temperature), as well as specifi c functional attributes such as rooting depth, belowground biomass produc-tion, and phenology are desired.
ACKNOWLEDGMENTSWe thank Jason Benz, Brian Strand, Geoff Dervishian, Josh Hunt, and Sharon Schnabel for their assistance with fi eld work. We thank Dr. David Block and Dr. Bruce Mackey for assistance with statistics exercises, and the two anonymous reviewers for their thoughtful comments. Th is research was funded by the Kearney Foundation of Soil Science and the California Department of Food and Agriculture.
REFERENCESAmundson, R. 2001. Th e carbon budget in soils. Annu. Rev. Earth Planet. Sci.
29:535–562.Álvaro-Fuentes, J., M.V. López, J.L. Arrúe, and C. Cantero-Martínez. 2008.
Management eff ects on soil carbon dioxide fl uxes under semiarid Mediterranean conditions. Soil Sci. Soc. Am. J. 72:194–200.
Andrews, S.S., J.P. Mitchell, R. Mancinelli, D.L. Karlen, T.K. Hartz, W.R. Horwath, G.S. Pettygrove, K.M. Scow, and D. Munk. 2002. On-farm assessment of soil quality in California’s Central Valley. Agron. J. 94:12–23.
Asensio, D., J. Peñuelas, R. Ogaya, and J. Llusià. 2007. Seasonal soil and leaf CO2 exchange rates in a Mediterranean holm oak forest and their responses to drought conditions. Atmos. Environ. 41:2447–2455.
Birch, H.F. 1958. Th e eff ect of soil drying on humus decomposition and nitrogen availability. Plant Soil 10:9–31.
Bottner, P., M. Pansu, and Z. Sallih. 1999. Modelling the eff ect of active roots on soil organic matter turnover. Plant Soil 216:15–25.
Cable, J.M., K. Ogle, D.G. Williams, J.F. Weltzin, and T.E. Huxman. 2008. Soil texture drives responses of soil respiration to precipitation pulses in the Sonoran Desert: Implications for climate change. Ecosystems 11:961–979.
Calderón, F.J., L.E. Jackson, K.M. Scow, and D.E. Rolston. 2000. Microbial responses to simulated tillage in cultivated and uncultivated soils. Soil Biol. Biochem. 32:1547–1559.
Calderón, F.J., L.E. Jackson, K.M. Scow, and D.E. Rolston. 2001. Short-term dynamics of nitrogen, microbial activity and phospholipid fatty acids aft er tillage. Soil Biol. Biochem. 65:118–126.
Calderón, F.J., and L.E. Jackson. 2002. Rototillage, Disking, and Subsequent Irrigation: Eff ects on Soil Nitrogen Dynamics, Microbial Biomass, and Carbon Dioxide Effl ux. J. Environ. Qual. 31:752–758.
Carlisle, E.A., K.L. Steenwerth, and D.R. Smart. 2006. Eff ects of land use on soil respiration: Conversion of oak woodlands to vineyards. J. Environ. Qual. 35:1396–1404.

SSSAJ: Volume 74: Number 1 • January–February 2010 239
Casals, P., J. Romanyà, J. Cortina, P. Bottner, M.-M. Coûteaux, and V. Ramon Vallejo. 2000. CO2 effl ux from a Mediterranean semi-arid forest soil. I. Seasonality and eff ects of stoniness. Biogeochemistry 48:261–281.
Denef, K., J. Six, H. Bossuyt, S.D. Frey, E.T. Elliot, R. Merckx, and K. Paustian. 2001. Infl uence of dry-wet cycles on the interrelationship between aggregate, particulate organic matter, and microbial community dynamics. Soil Biol. Biochem. 33:1599–1611.
Duiker, S.W., and R. Lal. 2000. Carbon budget study using CO2 fl ux measurements from a no till system in central Ohio. Soil Tillage Res. 54:21–30.
Eriksen, J., and L.S. Jensen. 2001. Soil respiration, nitrogen mineralization and uptake in barley in following cultivation of grazed grasslands. Biol. Fertil. Soils 33:139–145.
Fierer, N., and J.P. Schimel. 2002. Eff ects of drying-rewetting frequency on soil carbon and nitrogen transformations. Soil Biol. Biochem. 34:777–787.
Gaumont-Guay, D., T.A. Black, T.J. Griffi s, A.G. Barr, R.S. Jassal, and Z. Nesic. 2006. Interpreting the dependence of soil respiration on soil temperature and water content in a boreal aspen stand. Agric. For. Meteorol. 140:220–235.
Grandy, A.S., and G.P. Robertson. 2007. Land-use intensity eff ects on soil organic carbon accumulation rates and mechanisms. Ecosystems 10:58–73.
Hanson, P., N. Edwards, C.T. Garten, and J.A. Andrews. 2001. Separating root and soil microbial contributions to soil respiration: A review of methods and observations. Biogeochemistry 48:115–146.
Halvorson, A.D., B.J. Wienhold, and A.L. Black. 2002. Tillage, nitrogen, and cropping system eff ects on soil carbon sequestration. Soil Sci. Soc. Am. J. 66:906–912.
Hillel, D. 1982. Introduction to soil physics. Academic Press, Inc. San Diego, CA.Ingels, C., and K. Klonsky. 1998. Historical and current uses. p. 3–7. In Cover
cropping in Vineyards. University of California, DANR Publication No. 3338. Univ. of California, Oakland, CA
Jackson, L.E., F.J. Calderón, K.L. Steenwerth, K.M. Scow, and D.E. Rolston. 2003. Responses of soil microbial processes and community structure to tillage events and implications for soil quality. Geoderma 114:305–317.
Kern, J.S., and M.G. Johnson. 1993. Conservation tillage impacts on national soil and atmospheric carbon levels. Soil Sci. Soc. Am. J. 57:200–210.
Kieft , T., E. Soroker, and M.K. Firestone. 1987. Microbial biomass response to a rapid increase in water potential when dry soil is wetted. Soil Biol. Biochem. 19:119–126.
Lal, R. 2002. Soil carbon dynamics in cropland and rangeland. Environ. Pollut. 116:353–362.
Lambert, G. 1978. Soil survey of Napa County, California. USDA-Natural Resources Conservation Service, Washington, DC.
Liu, X., S. Wan, B. Su, D. Hui, and Y. Luo. 2002. Response of soil CO2 effl ux to water manipulation in a tallgrass prairie ecosystem. Plant Soil 240:213–223.
Lundquist, E.J., L.E. Jackson, and K.M. Scow. 1999a. Wet-dry cycles aff ect dissolved organic carbon in two California agricultural soils. Soil Biol. Biochem. 31:1031–1038.
Lundquist, E.J., K.M. Scow, L.E. Jackson, S.L. Uesugi, and C.R. Johnson. 1999b. Rapid response of soil microbial communities from conventional, low input, and organic farming systems to a wet/dry cycle. Soil Biol. Biochem. 31:1661–1675.
Martens, D.A., W. Emmerich, J.E.T. McLain, and T.N. Johnsen. 2005. Atmospheric carbon mitigation potential of agricultural management in the southwestern USA. Soil Tillage Res. 83:95–119.
Maestre, F.T., and J. Cortina. 2003. Small-scale spatial variation in soil CO2 effl ux in a Mediterranean semiarid steppe. Appl. Soil Ecol. 23:199–209.
Parkin, T.B., and T.C. Kaspar. 2003. Temperature controls on diurnal carbon dioxide fl ux: Implications for estimating soil carbon loss. Soil Sci. Soc. Am. J. 67:1763–1772.
Paul, E.A., D. Harris, H.P. Collins, U. Schulthess, and G.P. Robertson. 1999. Evolution of CO2 and soil carbon dynamics in biologically managed, row-crop agroecosystems. Appl. Soil Ecol. 11:53–65.
Pella, E. 1990. Combustion gas analyzer method for total carbon and total nitrogen. Am. Lab. 22:16.
Pierce, D.L. 2006. Soil carbon as a function of tillage and climate in a California vineyard. Ph.D. Th esis. University of California, Davis, CA.
Pitt, M.D., and H.F. Heady. 1977. Responses of annual vegetation to temperature and rainfall patterns in northern California. Ecology 59:336–350.
Reicosky, D.C., W.A. Dugas, and H.A. Torbert. 1997. Tillage-induced carbon dioxide loss from diff erent cropping systems. Soil Tillage Res. 41:105–118.
Rey, A., E. Pegoraro, V. Tedeschi, I. De Parri, P.G. Jarvis, and R. Valentini. 2002. Annual variation in soil respiration and its components in a coppice oak forest in Central Italy. Glob. Change Biol. 8:851–866.
Rey, A., C. Petsikos, P.G. Jarvis, and J. Grace. 2005. Eff ect of temperature and moisture on rates of carbon mineralization in a Mediterranean oak forest soil under controlled and fi eld conditions. Eur. J. Soil Sci. 56:589–599.
Schlesinger, W.H., and J.A. Andrews. 2000. Soil respiration and the global carbon cycle. Biogeochemistry 48:7–20.
Six, J., E.T. Elliot, and K. Paustian. 1999. Aggregate and soil organic matter dynamics under conventional and no-tillage systems. Soil Sci. Soc. Am. J. 63:1350–1358.
Smith, P., D.S. Powlson, and M.J. Glendining. 1998. Preliminary estimates of the potential for carbon mitigation in European soils through no-till farming. Glob. Change Biol. 4:679–685.
Sørensen, L.H. 1974. Rate of decomposition of organic matter in soil as infl uenced by repeated air drying-rewetting and repeated additions of organic material. Soil Biol. Biochem. 6:287–292.
Steenwerth, K.L., L.E. Jackson, F.J. Calderón, K.M. Scow, and D.E. Rolston. 2005. Response of microbial community composition and activity in agricultural and grassland soils aft er a simulated rainfall. Soil Biol. Biochem. 37:2249–2262.
Steenwerth, K.L., and K.M. Belina. 2008. Cover crops enhance soil organic matter, carbon dynamics and microbiological function in a vineyard agroecosystem. Appl. Soil Ecol. 40:359–369.
Trumbore, S.E. 1997. Potential responses of soil organic carbon to global environmental change. Proc. Natl. Acad. Sci. USA 94:8284–8291.
USDA-NASS (National Agriculture Statistics Service). 2009. Noncitrus fruits and nuts 2008 summary. (http://www.nass.usda.gov)
Vargas, R., and M.F. Allen. 2008. Environmental controls and the infl uence of vegetation type, fi ne roots and rhizomorphs on diel and seasonal variation in soil respiration. New Phytol. 179:460–471.Veenstra, J.J., W.R. Horwath, and J.P. Mitchell. 2007. Tillage and cover cropping
eff ects on aggregate-protected carbon in cotton and tomato. Soil Sci. Soc. Am. J. 72:362–371.
Xu, L., D.D. Baldocchi, and J. Tang. 2004. How soil moisture, rain pulses, and growth alter the response of ecosystem respiration to temperature. Global Biogeochem. Cycles 18:1–10.
Zhang, Y., C.S. Li, and X.J. Zhou. 2002. A simulation model linking crop growth and soil biogeochemistry for sustainable agriculture. Ecol. Modell. 151:75–108.




















