
Adaptive changes in zebrafish brain in dominant–subordinate behavioral context

Available online at ww
w.sciencedirect.comANIMAL BEHAVIOUR, 2008, 75, 879e886doi:10.1016/j.anbehav.2007.07.009
Subordinate wasps are more aggressive in colonies with
low reproductive skew
D. FANELLI*†, J . J . BOOMSMA† & S. TURILLAZZI*
*Dipartimento di Biologia Animale e Genetica, University of Florence
yDepartment of Population Biology, Institute of Biology, University of Copenhagen
(Received 15 January 2007; initial acceptance 19 March 2007;
final acceptance 10 July 2007; published online 18 October 2007; MS. number: 9237)
The small societies of primitively eusocial wasps have provided interesting testing grounds for reproductiveskew theory because all individuals have similar reproductive potential, which is unusual in social insectsbut common in vertebrate societies. Aggression is a key parameter in testing the theory, but empirical stud-ies have seldom quantified aggression together with the entire array of other relevant variables. The fewstudies that have done so were recently criticized for failing to control for the overall level of social activity.We analysed behaviour and reproductive partitioning patterns in the stenogastrine wasp Parischnogastermellyi. We used aggression of the subordinate (b) breeder as key variable and analysed how relatedness,body size, number of breeders and productivity affect the interaction between the reproductive skewand the aggression while controlling for nest activity and actual interaction time between a and b. Weshowed that (1) more even reproductive partitioning (lower reproductive skew) is associated with higherlevels of aggression initiated by the b subordinate independent of colony activity and (2) none of the cur-rently available reproductive skew models is convincingly supported. Comparison of our results with ear-lier studies suggests that a common aggression-based mechanism for reproductive partitioning may applyacross all primitively eusocial wasps despite complications arising from variable activity levels; however,currently available models may not apply.
� 2007 The Association for the Study of Animal Behaviour. Published by Elsevier Ltd. All rights reserved.
Keywords: microsatellites; Parischnogaster mellyi; primitiv
CorrespoGenetica(email: dof Popul2100 Co
0003e3
ely eusocial; reproductive transactions; social behaviour;Stenogastrinae; tug of war; wasps
Elucidating why natural selection often favours individ- and the dominant is in control of group composition,
uals that give up reproduction to rear the brood of othersis a fundamental problem in evolutionary biology (Leigh1999). A series of models have produced predictions abouthow reproduction could be partitioned without becomingevolutionarily unstable, but their empirical support hasremained controversial.Optimal skew models (OSMs) assume that the level ofreproductive partitioning is ‘optimal’ for all members ofa society and cooperation is made possible by reproductive‘transactions’. Provided that both a dominant and a sub-ordinate are able to monitor their share of reproduction
ndence: Daniele Fanelli, Dipartimento di Biologia Animale e, Universita di Firenze, Via Romana 17, 50125 Florence, [email protected]). J. J. Boomsma is at the Departmentation Biology, Institute of Biology, University of Copenhagen,penhagen, Denmark.
879472/08/$32.00/0 � 2007 The Association for the Stu
OSMs predict a minimum ‘staying incentive’ that thedominant must concede to keep the subordinate in thegroup and an upper limit of reproductive sharing that thesubordinate can claim, above which the dominant shouldprefer to evict the subordinate (Johnstone 2000; Reeve2000; Reeve & Keller 2001). Between these lower and up-per limits there should be a ‘window of selfishness’ (WOS)where breeders are expected to compete directly overreproduction leading to a compromise situation (Reeveet al. 1998; Johnstone 2000; Reeve 2000). The reproduc-tive transactions paradigm is usually contrasted with‘pure’ compromise models, which assume that neitherdominants nor subordinates have complete control overreproductive partitioning and never peacefully concedeshares of reproduction. The most widely discussed com-promise model is the ‘tug of war’ (TOWM), which assumesthat the degree of reproductive skew in a society is the re-sult of a compromise between the conflicting optima of its
dy of Animal Behaviour. Published by Elsevier Ltd. All rights reserved.

ANIMAL BEHAVIOUR, 75, 3880
members (Reeve et al. 1998; Cant 2006) and formalizesthe most widespread view of the evolutionary significanceof reproductive sharing (Reeve & Keller 2001). Critics ofthese approaches have pointed out that the level of repro-ductive skew may depend on additional factors andassumptions that are not explicitly addressed in mostOSMs and the TOWM, such as the phase of the colony-and/or life-cycle of the individuals concerned and their(in)ability to monitor reproduction, group compositionand other relevant parameters (e.g. Strassmann 1993;Beekman et al. 2003; Kokko 2003; Fanelli et al. 2005;Hart & Ratnieks 2005).
Aggression is the clearest expression of conflict; it isbelieved to play an important role in the partitioning ofresources in animal societies and to be the key parameterfor testing reproductive skew theory (Reeve 2000; Cant2006; Cant et al. 2006). Several skew models have pro-duced testable predictions about how aggression shouldbe correlated with reproductive skew and associated vari-ables. Most OSMs predict that the frequency of aggressionshould be positively correlated with the width of the WOSwhen aggression allows individuals to directly increasetheir share of reproduction. This implies that aggressionshould increase with increasing reproductive skew, relat-edness, productivity and severity of ecological constraints,while it may be positively or negatively correlated withthe fighting ability of subordinates (Reeve 2000). How-ever, if aggression is a not a means to increase reproductiveskew but instead allows a subordinate only to evict or sub-jugate the dominant, optimal skew theory predicts eithera positive or a negative correlation between aggressionand reproductive skew, depending on the value of theother parameters (Cant & Johnstone 2000). If theTOWM applies, then under most conditions aggressionshould be negatively correlated with reproductive skewand relatedness, be positively correlated with the relativefighting ability of subordinates and have no correlationwith productivity and the severity of ecological con-straints (Reeve 2000).
Primitively eusocial wasps are good invertebrate modelsystems for testing reproductive skew predictions becausethey potentially match the assumptions of completecontrol over reproduction and group composition (Reeve& Keller 2001). Genetic colony structure and patterns ofaggression have therefore been intensively investigatedin several species (Reeve & Nonacs 1992; Field et al.1998; Queller et al. 2000; Reeve et al. 2000; Tibbetts &Reeve 2000; Seppa et al. 2002; Sumner et al. 2002; Nonacset al. 2004; Fanelli et al. 2005; Liebert et al. 2005; Liebert &Starks 2006). Most of these studies provided no conclusivesupport for any of the current models of reproductiveskew and cross-species comparison suggests that none ofthe key model parameters (i.e. relatedness, size differencebetween individuals, colony size, and productivity) isa universal predictor of reproductive skew in wasps.However, the only two studies that simultaneously ana-lysed aggression and reproductive skew showed thatthey were negatively correlated; that is, the proportionof aggression by the b female was negatively correlatedwith reproductive skew in Polistes carolina (Seppa et al.2002) and smaller colonies had both lower skew and
higher levels of aggression from the subordinate towardsthe dominant in Polistes bellicosus (Field et al. 1998).
The fundamental importance of aggression for repro-ductive skew has recently been challenged by the sugges-tion that behavioural studies measuring aggression mayhave been flawed. In Polistes fuscatus, Nonacs et al. (2004)found that the proportion of aggression initiated by a sub-ordinate was not significantly correlated with relatedness,size difference, colony size or productivity (in contrast topredictions made by OSMs) and that wasps were morelikely to meet aggression when they were moving andactive on the nest. Since the frequency of aggressive inter-actions between dominant and subordinates is stronglyaffected by the time they spend together on the nest andby the overall level of activity in the colony, Nonacs et al.(2004) hypothesized that the negative correlation betweenreproductive skew and aggression observed previously (inparticular in P. bellicosus) could also have been due to theper capita work load and number of landings being higherin the smaller, low-skew colonies. Unfortunately, this hy-pothesis could not be tested in P. fuscatus, because geneticanalyses were available for only very few colonies, whichdid not vary in reproductive skew (Nonacs et al. 2004).
The aim of the present study was to provide a compre-hensive analysis of aggression and reproductive skew inthe primitively eusocial stenogastrine wasp Parischnogastermellyi while controlling for relevant covariables and possi-ble biases due to differences in activity and incompletedata. We analysed 15 field colonies for which all markedand video-recorded females were recovered upon nest col-lection and for which we had information on virtually allrelevant variables. To our knowledge no previous study onreproductive partitioning in wasps has been as complete.
METHODS
Natural History
The Stenogastrinae or hover wasps are a subfamily ofprimitively eusocial Vespidae living in Southeast Asia(reviews in Turillazzi 1991, 1996). In P. mellyi all adultfemales are potential reproductives, and often multiplefemales in a colony have developed ovaries and are in-seminated (Hansell 1983; Fanelli et al. 2005). Nestmatefemales form well-defined hierarchies with division oflabour, but alternative behavioural strategies are availableto subordinates, which frequently move to vacant nestsor join nests of unrelated colonies to become dominantsor helpers (Hansell 1983; Yamane et al. 1983). Geneticanalyses of natural colonies have shown that (1) within-colony relatedness is highly variable but quite low onaverage (ca. 0.33 � 0.05), (2) the a female monopolizesreproduction in most colonies, although subordinatesoccasionally lay eggs that are raised to maturity, and (3)queen turnover occurs, so that a subordinate sometimessupersedes the a female (Fanelli et al. 2005).
Sampling
All colonies used in the experiment were collected inFebruary 2003 around barracks near the field station of the

FANELLI ET AL.: AGGRESSION AND REPRODUCTIVE SKEW IN WASPS 881
University of Malaya at Ulu Gombak, Peninsular Malaysia.The maximum distance between the colonies was lessthan 200 m. All adult female and male wasps were cap-tured at dawn (before 0700 hours) and individuallymarked. Colonies were subsequently left undisturbed for24e72 h to resume their normal levels of activity. Wevideotaped 32 colonies with multiple females (range 2e7)for 6 h, during the same day, from 1000 to 1300 h andfrom 1500 to 1800 h. Video recordings took place exclu-sively on sunny days. Colonies were then left undisturbeduntil the evening and collected at night. To obtain com-plete information and ensure equal treatment for all colo-nies involved in the study, we performed genetic andbehavioural analyses (see below) only on a subset of 15colonies, which had all marked females present at thetime of collection.
Adult females were dissected and measured undera binocular microscope. The maximum width of thehead was measured with an optical ruler, which alloweda maximum error of 1%. We inspected all brood to classifydevelopmental stages. The dissections and measurementsof adult females, the genetic data and the behaviouralobservations were obtained blindly to one another.
Behavioural Analysis
We scored the position of every female on the nest (orthe time she spent away from it) every 30 s. The hierarchi-cal status of each adult female was assessed according tothe proportion of time spent on the nest, in particularon the upper part of the nest (Hansell 1983). The fre-quency of submissive behaviour (upon encounters, a sub-ordinate female will usually yield place to a dominant)and the direction of aggressive acts were also taken intoaccount to rank the females.
We scored all interactions involving intense antenna-tions, mounting, biting, lunging and fighting as aggressiveand subdivided them on the basis of whether they weredirected towards a moving individual or towards a restingindividual, as suggested by Nonacs et al. (2004).
Genetic Analysis
We extracted DNA from adults and brood using a 5%Chelex and proteinase K solution. The DNA of all in-dividuals was amplified in polymerase chain reactions forfive microsatellite loci developed for other stenogastrinespecies: Lf9, Lf18, Ef92, Ef98, and Ef99 (Zhu et al. 2000;Sumner & Field 2001). There were 4e13 alleles per locusand the observed heterozygosity per locus was 0.63e0.84. Products were separated on 6% polyacrylamide gelsusing an ABI automated sequencer. Populationwide allelefrequencies were obtained from the total sample of 88adult females.
We estimated relatedness using the Relatedness 5.0.8software (Goodnight & Queller 2000), weighting coloniesequally and obtaining standard errors (SE) by jackknifingover colonies. Female kin groups were identified usingthe program Kinship 1.5.1b (Goodnight & Queller 1999),setting full sisters as the primary hypothesis and first
cousins as the null hypothesis and accepting a �0.05 sig-nificance level. DNA from sperm in the spermatheca wasextracted using a 5% Chelex solution and amplified usingthe same protocol as for adults but with a higher DNAconcentration. When successfully genotyped (N ¼ 18),the genetic identity of sperm from the spermatheca gaveadditional information to confirm the assignment orexclusion of putative mothers.
We excluded from the analysis any individual (adult orbrood) whose DNA had failed to amplify at more thanthree loci (3.3% of individuals). The probability fora female to be homozygous for the most common alleleat all five loci was 0.00018, and the maximum probabilityfor a female to be homozygous for the two least poly-morphic loci was 0.08. All brood that exhibited a singleband at every locus was therefore considered to be haploid(i.e. male). Skew was measured as the proportion of eggslaid by the a female. To obtain a more reliable estimate ofreproductive skew among male eggs, we used maximumlikelihood statistics that evaluate the most likely pro-portion laid by the dominant by testing all possibleproportions between 0 and 1 at increments of 0.01 (Tothet al. 2002). However, other measures of skew (simplycounting the eggs that could/could not be attributed tothe dominant as well as calculating any of the availableskew indexes) did not affect our results in any substantialway because we used a nonparametric ranked correlationtest for the univariate analyses (Spearman rS). For the mul-tiple regression analyses, we simply classified the coloniesinto two groups on the basis of whether they had any eggslaid by a nondominant (see also Results).
RESULTS
The average within-colony relatedness was 0.45 � 0.05(N ¼ 15) with colony-level estimates ranging between0.07 and 1.00, that is somewhat higher than those in a pre-vious study, which may be due to unrelated floaters beingmore likely to leave nests at even the slightest disturbance(see Fanelli et al. 2005). Maximum likelihood analysis in-dicated that one colony was a full-sister association, 10colonies were motheredaughter associations with themother still being the dominant and four colonies weremixed groups where at least one female was unrelated tothe others. The estimate of mean relatedness betweena and b females was 0.39 � 0.08 and ranged from 0.03to 1.00, that is from being unrelated to full-sister related-ness. (Haplodiploid full sisters are related on average by0.75, but estimates may produce extreme values of 1.0when all paternal and maternal alleles are identical bychance. Our sample had one such case in a pair of nest-mates). The average proportion of eggs laid by the domi-nant was 0.95 � 0.13 (N ¼ 15), which represents thehigh overall reproductive skew that is typical for this spe-cies (Fanelli et al. 2005) and other stenogastrine wasps(Sumner et al. 2002; Landi et al. 2003). Reproductiveskew was incomplete in three colonies, in which someof the male eggs were not compatible with the genotypeof the dominant. In two of these colonies, the eggs laidby the subordinate were compatible only with the geno-type of the b female and in one of these colonies a single
0
0.02
0.04
0.06
0.08
0.1
0.12
0.14
0.16
Recipient moving Recipient resting
Agg
ress
ion
fre
qu
ency
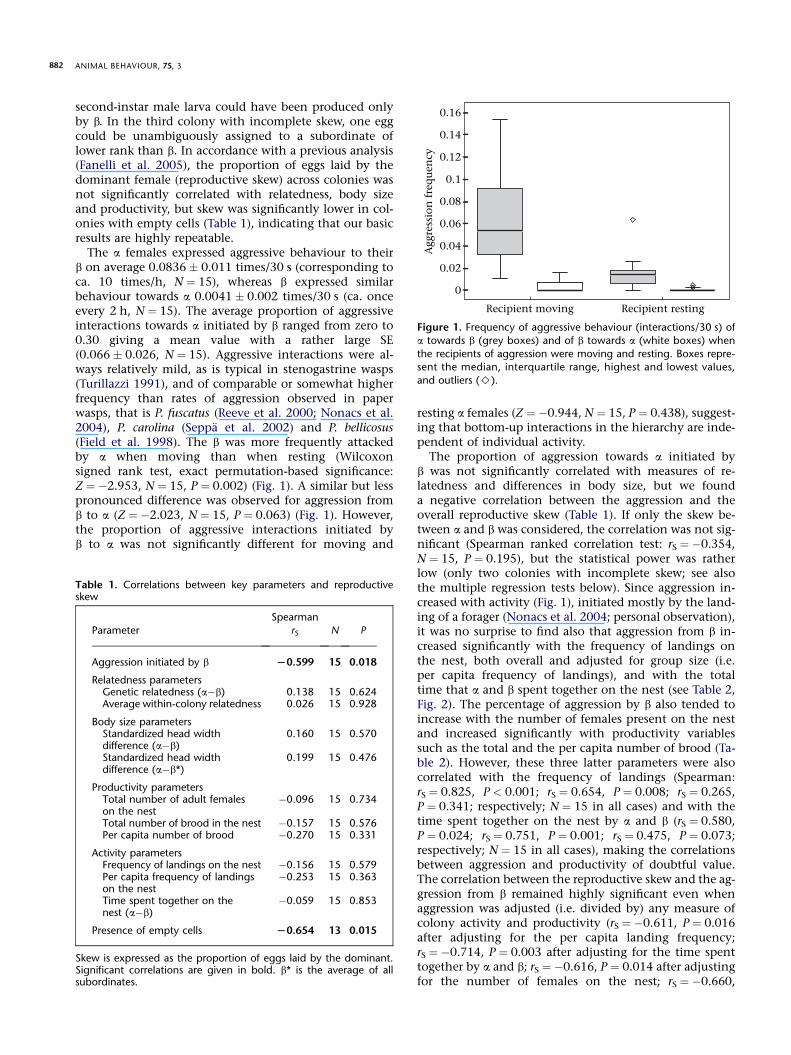
Figure 1. Frequency of aggressive behaviour (interactions/30 s) of
a towards b (grey boxes) and of b towards a (white boxes) when
the recipients of aggression were moving and resting. Boxes repre-sent the median, interquartile range, highest and lowest values,
and outliers (>).
ANIMAL BEHAVIOUR, 75, 3882
second-instar male larva could have been produced onlyby b. In the third colony with incomplete skew, one eggcould be unambiguously assigned to a subordinate oflower rank than b. In accordance with a previous analysis(Fanelli et al. 2005), the proportion of eggs laid by thedominant female (reproductive skew) across colonies wasnot significantly correlated with relatedness, body sizeand productivity, but skew was significantly lower in col-onies with empty cells (Table 1), indicating that our basicresults are highly repeatable.
The a females expressed aggressive behaviour to theirb on average 0.0836 � 0.011 times/30 s (corresponding toca. 10 times/h, N ¼ 15), whereas b expressed similarbehaviour towards a 0.0041 � 0.002 times/30 s (ca. onceevery 2 h, N ¼ 15). The average proportion of aggressiveinteractions towards a initiated by b ranged from zero to0.30 giving a mean value with a rather large SE(0.066 � 0.026, N ¼ 15). Aggressive interactions were al-ways relatively mild, as is typical in stenogastrine wasps(Turillazzi 1991), and of comparable or somewhat higherfrequency than rates of aggression observed in paperwasps, that is P. fuscatus (Reeve et al. 2000; Nonacs et al.2004), P. carolina (Seppa et al. 2002) and P. bellicosus(Field et al. 1998). The b was more frequently attackedby a when moving than when resting (Wilcoxonsigned rank test, exact permutation-based significance:Z ¼ �2.953, N ¼ 15, P ¼ 0.002) (Fig. 1). A similar but lesspronounced difference was observed for aggression fromb to a (Z ¼ �2.023, N ¼ 15, P ¼ 0.063) (Fig. 1). However,the proportion of aggressive interactions initiated byb to a was not significantly different for moving and
Table 1. Correlations between key parameters and reproductiveskew
Parameter
Spearman
rS N P
Aggression initiated by b L0.599 15 0.018
Relatedness parametersGenetic relatedness (a�b) 0.138 15 0.624Average within-colony relatedness 0.026 15 0.928
Body size parametersStandardized head widthdifference (a�b)
0.160 15 0.570
Standardized head widthdifference (a�b*)
0.199 15 0.476
Productivity parametersTotal number of adult femaleson the nest
�0.096 15 0.734
Total number of brood in the nest �0.157 15 0.576Per capita number of brood �0.270 15 0.331
Activity parametersFrequency of landings on the nest �0.156 15 0.579Per capita frequency of landingson the nest
�0.253 15 0.363
Time spent together on thenest (a�b)
�0.059 15 0.853
Presence of empty cells L0.654 13 0.015
Skew is expressed as the proportion of eggs laid by the dominant.Significant correlations are given in bold. b* is the average of allsubordinates.
resting a females (Z ¼ �0.944, N ¼ 15, P ¼ 0.438), suggest-ing that bottom-up interactions in the hierarchy are inde-pendent of individual activity.
The proportion of aggression towards a initiated byb was not significantly correlated with measures of re-latedness and differences in body size, but we founda negative correlation between the aggression and theoverall reproductive skew (Table 1). If only the skew be-tween a and b was considered, the correlation was not sig-nificant (Spearman ranked correlation test: rS ¼ �0.354,N ¼ 15, P ¼ 0.195), but the statistical power was ratherlow (only two colonies with incomplete skew; see alsothe multiple regression tests below). Since aggression in-creased with activity (Fig. 1), initiated mostly by the land-ing of a forager (Nonacs et al. 2004; personal observation),it was no surprise to find also that aggression from b in-creased significantly with the frequency of landings onthe nest, both overall and adjusted for group size (i.e.per capita frequency of landings), and with the totaltime that a and b spent together on the nest (see Table 2,Fig. 2). The percentage of aggression by b also tended toincrease with the number of females present on the nestand increased significantly with productivity variablessuch as the total and the per capita number of brood (Ta-ble 2). However, these three latter parameters were alsocorrelated with the frequency of landings (Spearman:rS ¼ 0.825, P < 0.001; rS ¼ 0.654, P ¼ 0.008; rS ¼ 0.265,P ¼ 0.341; respectively; N ¼ 15 in all cases) and with thetime spent together on the nest by a and b (rS ¼ 0.580,P ¼ 0.024; rS ¼ 0.751, P ¼ 0.001; rS ¼ 0.475, P ¼ 0.073;respectively; N ¼ 15 in all cases), making the correlationsbetween aggression and productivity of doubtful value.The correlation between the reproductive skew and the ag-gression from b remained highly significant even whenaggression was adjusted (i.e. divided by) any measure ofcolony activity and productivity (rS ¼ �0.611, P ¼ 0.016after adjusting for the per capita landing frequency;rS ¼ �0.714, P ¼ 0.003 after adjusting for the time spenttogether by a and b; rS ¼ �0.616, P ¼ 0.014 after adjustingfor the number of females on the nest; rS ¼ �0.660,

Table 2. Correlation between key parameters and proportion ofaggressive interactions initiated by subordinate (b) females
Parameter
Spearman
rS N P
Relatedness parametersGenetic relatedness (a�b) �0.185 15 0.508Average within-colony relatedness �0.121 15 0.668
Body size parametersStandardized head widthdifference (a�b)
�0.167 15 0.553
Standardized head widthdifference (a�b*)
�0.181 15 0.517
Productivity parametersTotal number of adult femaleson the nest
0.498 15 0.059
Total number of brood in the nest 0.710 15 0.003Per capita number of brood 0.540 15 0.038
Activity parametersFrequency of landings on the nest 0.657 15 0.008Per capita frequency of landingson the nest
0.649 15 0.009
Time spent together on thenest (a�b)
0.661 15 0.007
Presence of empty cells 0.137 13 0.655
Significant correlations are given in bold. b* is the average of allsubordinates.
0 100 200 300
0
0.05
0.1
0.15
0.2
0.25
0.3
Time together (α − β)
(b)
0.002 0.004 0.006 0.008 0.010
0
0.05
0.1
0.15
0.2
0.25
0.3
Per capita frequency of landingsA
ggre
ssio
n o
f β
tow
ard
s α
(a)
0.012
Figure 2. Proportion of aggressive behaviour of b towards a as a func-
tion of the per capita frequency of landings (landings per 30 s di-vided by the number of individuals in the colony) (a) and the time
spent together (min) on the nest (b). (B): complete-skew colonies;
(C): incomplete-skew colonies; (:): a colony where the subordi-
nate egg layer was not the b female. Fitted lines are based on linearregression analysis.
FANELLI ET AL.: AGGRESSION AND REPRODUCTIVE SKEW IN WASPS 883
P ¼ 0.007 after adjusting for the total number of brood;rS ¼ �0.636, P ¼ 0.011 after adjusting for the per capitanumber of brood; N ¼ 15 in all cases).
We used univariate nonparametric correlation analysesbecause the data deviated from normal distributions andbecause this allowed direct comparisons with previousstudies of reproductive skew in wasps (e.g. Field et al.1998; Seppa et al. 2002; Nonacs et al. 2004). However,multiple regression analysis with stepwise backward elim-ination of predictor variables corroborated our conclu-sions. Due to the many colonies with skew ¼ 1 and withaggression ¼ 0, the linearity assumption of multiple linearregression could not be matched. Therefore, we useda more robust logistic regression model, which requiresthe dependent variable to be categorical and the distribu-tion of errors to be normal. To satisfy these assumptions,we transformed the aggression and skew variables intobinary variables: colonies where b was never aggressiveto a versus colonies where at least some aggression hadbeen observed and incomplete-skew colonies versus com-plete-skew colonies. The logistic regression model usedaggression as the independent variable and reproductiveskew, relatedness, body size difference and an activity/pro-ductivity variable (one at a time because they were highlycorrelated: either the number of females, or the number ofbrood, or the per capita number of brood, or the timespent together on the nest by a and b, or the frequencyof landings or the per capita frequency of landings) as pre-dictors. In all cases, the stepwise analysis eliminated therelatedness and body size difference predictor variablesand kept the binary reproductive skew parameter plusany productivity/activity parameters as significant predic-tors of aggression (significances for skew: chi-square test:
c2 ¼ 7.111, P ¼ 0.008 with the number of females;c2 ¼ 10.1515, P ¼ 0.001 with the number of brood;c2 ¼ 6.069, P ¼ 0.014 with the per capita number ofbrood; c2 ¼ 13.387, P < 0.001 with the time together onthe nest; c2 ¼ 6.130, P ¼ 0.013 with the frequency oflandings; c2 ¼ 6.196, P ¼ 0.013 with the per capitafrequency of landings; for all tests N ¼ 15, df ¼ 1). This

ANIMAL BEHAVIOUR, 75, 3884
analysis was powerful enough to show that reproductiveskew between a and b (that is, ignoring the reproductionof g subordinates) was also a significant predictor of subor-dinate’s aggression: in all but one case, the binary variableseparating the two colonies where the b female had laideggs from all other colonies had a statistically significanteffect (c2 ¼ 5.908, P ¼ 0.015 with the number of females;c2 ¼ 7.743, P ¼ 0.005 with the number of brood; c2 ¼3.627, P ¼ 0.057 with the per capita number of brood;c2 ¼ 13.387, P<0.001 with the time together on thenest; c2 ¼ 5.702, P ¼ 0.017 with the frequency of land-ings; c2 ¼ 4.854, P ¼ 0.024 with the per capita frequencyof landings; for all tests N ¼ 15, df ¼ 1).
DISCUSSION
This study is apparently the first based only on a homoge-neous subsample of colonies where not a single memberwas lost as a result of marking and where completeinformation on nearly all parameters had been obtained(see Methods). This selection procedure has significantlyreduced the sample size. However, small sample sizewould have invalidated our conclusions only if we hadobtained exclusively nonsignificant correlations, whichwas not the case. Most correlations had nonsignificant Pvalues, and a few correlations were highly significant.Since the sample size is the same in all but one case (seeTables 1 and 2), the P values depend only on the effectsize, which is expressed by the steepness of the correla-tion. Therefore, our results clearly show that a few para-meters are much more tightly linked to aggression andreproductive skew than other parameters.
Our data corroborate recent results for P. fuscatus byNonacs et al. (2004), showing that aggression by b towardsa is significantly affected by the overall activity on thenest and the time spent together with a but not signifi-cantly correlated with many of the key parameters thatare relevant in the TOWM and OSMs (relatedness, sizedifference, group size and productivity once the level ofcolony activity is taken into account). We have focusedmainly on estimating bottom-up aggression in the nest-specific hierarchies to facilitate comparison of our resultswith those of previous studies (e.g. Field et al. 1998; Seppaet al. 2002; Nonacs et al. 2004), but we obtained very sim-ilar results when we measured the individual frequenciesof aggressive behaviour (data not shown).
However, our data do not confirm the hypothesis ofNonacs et al. (2004) that the negative correlation betweenthe aggression and the reproductive skew is a spuriousresult due to differences in activity or time spent together.Univariate and multivariate analyses showed that inP. mellyi aggression from subordinates was significantlycorrelated with both overall activity on the nest and sub-ordinate egg laying. The latter result corroborates resultsfor Polistes wasps (Field et al. 1998; Seppa et al. 2002),whereas the former result corroborates results by Nonacset al. (2004) while demonstrating simultaneously thatsignificant effects of overall activity do not preclude thepossibility that reproductive skew and subordinate aggres-sion are directly linked.
The patterns of aggression and reproductive partition-ing observed in P. mellyi are inconsistent with expecta-tions based on current OSMs but consistent withpredictions about direct reproductive conflicts between in-dividuals that have similar reproductive potential andshort tenure as dominant on the nest (e.g. Hart & Ratnieks2005). If aggression allows individuals to overthrow (i.e.evict or subjugate) same-sex breeders, supersede them(replace dominant breeders that are failing) or become ad-ditional or ‘sneaky’ breeders, then expression of conflictshould generally increase when relatedness is lower andimmediately before and after the replacement of a domi-nant breeder. In addition, supersedure should be morelikely among close kin, while overthrow should predomi-nate among more distantly related individuals (Hart &Ratnieks 2005). These expectations were partly confirmedin P. mellyi: we did not observe any correlation betweenaggression and relatedness but an earlier independentgenetic study showed that dominants remain on thenest as subordinates only when they are replaced by closekin (Fanelli et al. 2005). The Hart & Ratnieks (2005) sce-nario generally predicts that aggression from a subordinateshould be negatively correlated with reproductive skew, aswas observed in the present study.
The Hart & Ratnieks (2005) scenario is consistent withthe compromise paradigm, so it is conceptually closer tothe TOWM situation. However, the TOWM of Reeveet al. (1998) is not strongly supported in P. mellyi for atleast two reasons: (1) this model makes a number of otherpredictions that were not met, including a negative corre-lation between skew and relatedness, no or a negativecorrelation between skew and fighting ability, a negativecorrelation between aggression and relatedness, and a pos-itive correlation between aggression and subordinates’fighting ability, none of which was observed, and (2) thepotential for subordinates to evict or subjugate the domi-nant has also been analysed in the framework of OSMs,leading to the prediction that aggressive dominance test-ing by subordinates should be negatively correlated to re-productive skew when ecological constraints are constantand relatedness varies among colonies (Cant & Johnstone2000). Indeed, we recreated these latter conditions in oursample. Within-colony relatedness was variable and, byvideotaping and collecting all colonies from the samerestricted area and during a short period of time, we keptecological constraints relatively constant. Therefore, thepattern of aggression and skew we observed can in princi-ple be explained by transactional as well as compromiseparadigms, and neither OSMs nor the TOWM are conclu-sively supported by our data. However, we agree with Cant(2006) that OSMs can be falsified with a good degree ofcertainty, while ‘rigorous testing of the tug of war modelis difficult because of the nondiscriminating nature of itspredictions’.
Aggression and egg laying from subordinate waspsmight increase in response to specific cues indicatingthat the dominant is failing, such as the presence ofempty cells in a nest (the ‘weak queen’ hypothesis byStrassmann 1993; see also Liebig et al. 2005). Our presentdata indicate that empty cells were correlated to reproduc-tive skew (Table 1) as expected under this scenario.

FANELLI ET AL.: AGGRESSION AND REPRODUCTIVE SKEW IN WASPS 885
However, the P. mellyi data do not strongly support the‘weak queen’ hypothesis because empty cells are oftenfound in large nests that have recently been reoccupiedby a number of unrelated individuals (Fanelli et al.2005), which implies that open conflict over dominanceand egg laying is expected no matter how fit the dominantis. There is thus no conclusive evidence that empty cellsare being used by P. mellyi subordinates as a cue for weak-ness of the dominant.
In conclusion, most behavioural and genetic studies ofprimitively eusocial wasps have not provided strongsupport for OSMs in which social animals exchangereproductive incentives in return for cooperation becausethe partitioning of reproduction and the level of aggres-sion were not correlated in the predicted ways with therelevant model parameters (Field et al. 1998; Queller et al.2000; Sumner et al. 2002; Seppa et al. 2002; Nonacs et al.2004; Liebert et al. 2005; Fanelli et al. 2005; Liebert &Starks 2006; this study). Support for the TOWM has alsoremained inconclusive (see Cant 2006 for criticism ofclaims to the opposite by Langer et al. 2004). On the otherhand, a negative correlation between the aggression fromsubordinates and the reproductive skew has now beenfound in three species (Field et al. 1998; Seppa et al.2002; this study), suggesting that this may be a commonpattern in primitively eusocial wasps and other taxa. Itseems therefore that very general scenarios in which ag-gression helps subordinates to become dominant breedersappear at least as efficient as the current TOWM and OSMsin explaining patterns of reproductive skew in wasps.
Acknowledgments
We thank Peter Nonacs and anonymous referees forcomments on previous versions of the manuscript, Dr.Davide Croci for assistance in the field, Professor RosliHashim for providing material and logistic support inMalaysia and Sylvia Mathiasen for invaluable help duringthe genetic analyses. Funding of this work was obtainedthrough the Research Training Network INSECTS, financedby the European Commission under the ImprovingHuman Potential Programme (HPRN-CT-2000-00052),the Italian MIUR (COFIN 2001), and the University ofFlorence.
References
Beekman, M., Komdeur, J. & Ratnieks, F. L. W. 2003. Reproductive
conflicts in social animals: who has power? Trends in Ecology andEvolution, 18, 277e282.
Cant, M. A. 2006. A tale of two theories: parenteoffspring conflict
and reproductive skew. Animal Behaviour, 71, 255e263.
Cant, M. A. & Johnstone, R. A. 2000. Power struggles, dominance
testing, and reproductive skew. American Naturalist, 155, 406e417.
Cant, M. A., Llop, J. B. & Field, J. 2006. Individual variation in social
aggression and the probability of inheritance: theory and a fieldtest. American Naturalist, 167, 837e852.
Fanelli, D., Boomsma, J. J. & Turillazzi, S. 2005. Multiple repro-ductive strategies in a tropical hover wasp. Behavioral Ecology
and Sociobiology, 58, 190e199.
Field, J., Solis, C. R., Queller, D. C. & Strassmann, J. E. 1998. Social
and genetic structure of paper wasp cofoundress associations:
tests of reproductive skew models. American Naturalist, 151,545e563.
Goodnight, K. F. & Queller, D. C. 1999. Computer software for per-forming likelihood tests of pedigree relationship using genetic
markers. Molecular Ecology, 8, 1231e1234.
Goodnight, K. F. & Queller, D. C. 2000. Relatedness 5.0.8 Good-
night Software. Available at: http://www.gsoftnet.us/GSoft.html.
Hansell, M. H. 1983. Social behaviour and colony size in the wasp
Parischnogaster mellyi (Saussure), Stenogastrinae (Hymenoptera,
Vespidae). Entomological Proceedings C, 86, 167e178.
Hart, A. G. & Ratnieks, F. L. W. 2005. Crossing the taxonomic
divide: conflict and its resolution in societies of reproductively to-
tipotent individuals. Journal of Evolutionary Biology, 18, 383e395.
Johnstone, R. A. 2000. Models of reproductive skew: a review and
synthesis. Ethology, 106, 5e26.
Kokko, A. 2003. Are reproductive skew models evolutionarily stable?
Proceedings of the Royal Society of London, Series B, 270, 265e270.
Landi, M., Queller, D. C., Turillazzi, S. & Strassmann, J. E. 2003.
Low relatedness and frequent queen turnover in the stenogastrinewasp Eustenogaster fraterna favor the life insurance over the haplo-
diploid hypothesis for the origin of eusociality. Insectes Sociaux, 50,
262e267.
Langer, P., Hogendoorn, K. & Keller, L. 2004. Tug-of-war over
reproduction in a social bee. Nature, 428, 844e847.
Leigh, E. G. J. 1999. Levels of selection, potential conflicts, and their
resolution: the role of the ‘common good’. In: Levels of Selection in
Evolution (Ed. by L. Keller), pp. 15e30. Princeton, New Jersey:Princeton University Press.
Liebert, A. E. & Starks, P. T. 2006. Taming of the skew: transac-tional models fail to predict reproductive partitioning in the paper
wasp Polistes dominulus. Animal Behaviour, 71, 913e923.
Liebert, A. E., Nonacs, P. & Wayne, R. K. 2005. Solitary nesting andreproductive success in the paper wasp Polistes aurifer. Behavioral
Ecology and Sociobiology, 57, 445e456.
Liebig, J., Monin, T. & Turillazzi, S. 2005. Direct assessment of
queen qualita and lack of worker suppression in a paper
wasp. Proceedings of the Royal Society of London, Series B, 272,1339e1344.
Nonacs, P., Reeve, H. K. & Starks, P. T. 2004. Optimal reproduc-
tive-skew models fail to predict aggression in wasps. Proceedingsof the Royal Society of London, Series B, 271, 811e817.
Queller, D. C., Zacchi, F., Cervo, R., Turillazzi, S., Henshaw, M.,Santarelli, L. & Strassmann, J. E. 2000. Unrelated helpers in
a social insect. Nature, 405, 784e787.
Reeve, H. K. 2000. A transactional theory of within-group conflict.
American Naturalist, 155, 365e382.
Reeve, H. K. & Keller, L. 2001. Tests of reproductive-skew models in
social insects. Annual Review of Entomology, 46, 347e385.
Reeve, H. K. & Nonacs, P. 1992. Social contracts in wasp societies.
Nature, 359, 823e825.
Reeve, H. K., Emlen, S. T. & Keller, L. 1998. Reproductive sharing in
animal societies: reproductive incentives or incomplete control by
dominant breeders? Behavioral Ecology, 9, 267e278.
Reeve, H. K., Starks, P. T., Peters, J. M. & Nonacs, P. 2000. Genetic
support for the evolutionary theory of reproductive incentives in
social wasps. Proceedings of the Royal Society of London, Series B,267, 75e79. doi:10.1098/rspb.2000.0107.
Seppa, P., Queller, D. C. & Strassmann, J. E. 2002. Reproduction infoundress associations of the social wasp, Polistes carolina: conven-
tions, competition, and skew. Behavioral Ecology, 13, 531e542.
Strassmann, J. E. 1993. Weak queen of social contract? Nature, 363,
502e503.

ANIMAL BEHAVIOUR, 75, 3886
Sumner, S. & Field, J. 2001. Highly polymorphic microsatellite loci in
the facultatively eusocial hover wasp, Liostenogaster flavolineata and
cross-species amplification. Molecular Ecology Notes, 1, 229e231.
Sumner, S., Casiraghi, M., Foster, W. & Field, J. 2002. High re-
productive skew in tropical hover wasps. Proceedings of the RoyalSociety of London, Series B, 269, 179e186.
Tibbetts, E. & Reeve, H. K. 2000. Aggression and resource sharingamong foundresses in the social wasp Polistes dominulus: testing
transactional theories of conflict. Behavioral Ecology and Sociobiol-
ogy, 48, 344e352.
Toth, E., Strassmann, J. E., Nogueira-Neto, P., Imperatriz-Fonseca,V. L. & Queller, D. C. 2002. Male production in stingless bees:
variable outcomes of queeneworker conflict. Molecular Ecology,11, 2661e2667.
Turillazzi, S. 1991. The Stenogastrine. In: The Social Biology of Wasps
(Ed. by K. G. Ross & R. W. Matthews), pp. 74e98. Ithaca, New
York: Cornell University Press.
Turillazzi, S. 1996. Polistes in Perspective: Comparative Social Biol-
ogy in Belonogaster and Stenogastrinae. In: Natural history andevolution of paper-Wasps (Ed. by S. Turillazzi & M. J. West-
Eberhard), pp. 235e248. Oxford: Oxford University Press.
Yamane, S., Sakagami, S. F. & Ohgushi, R. 1983. Multiple behav-
ioural options in a primitively social wasp, Parischnogaster mellyi.
Insectes Sociaux, 30, 412e415.
Zhu, Y., Landi, M., Queller, D. C., Turillazzi, S. & Strass-mann, J. E. 2000. Polymorphic microsatellite loci for primi-
tively eusocial Stenogastrinae wasps. Molecular Ecology, 9,2203e2205.
Copyright © 2022 FDOKUMEN




















