
Study of programmed cell death in bovine herpesvirus 1 infected mdbk cells and the possible role of...
11
Acta Veterinaria Hungarica 52 (3), pp. 287–297 (2004) 0236-6290/$ 20.00 © 2004 Akadémiai Kiadó, Budapest STUDY OF PROGRAMMED CELL DEATH IN BOVINE HERPESVIRUS 1 INFECTED MDBK CELLS AND THE POSSIBLE ROLE OF NITRIC OXIDE IN THIS PROCESS Z. YAZICI 1 , Yasemin BASKIN 2 , H. BASKIN 3* , Ozlem GECER 4 , I. H. BAHAR 3 and A. ÖZKUL 5 1 Department of Virology, Faculty of Veterinary Medicine, Ondokuz Mayis University, Kurupelit, 55139, Samsun, Turkey; 2 Oncology Institute and 3 Department of Microbiol- ogy and Clinical Microbiology, School of Medicine, Dokuz Eylul University, Inciralti, Izmir, Turkey; 4 Department of Virology, Regional Institute of Hygiene, Ministry of Health, Göztepe, 35350 Izmir, Turkey; 5 Department of Virology, Faculty of Veterinary Medicine, Ankara University, Diskapi, 06110, Ankara, Turkey (Received September 6, 2003; accepted March 2, 2004) Bovine herpesvirus 1 (BHV-1) is the aetiological agent of many disease types and may predispose infected animals, possibly through immunosuppression, to secondary bacterial infections. Immunosuppression may directly be associated with the induction of programmed cell death (PCD) in some virus-infected cells. Nitric oxide (NO) has an important mediating role against fungal, bacterial, pro- tozoal, viral pathogens and tumours. BHV-1 induced apoptosis between 0.5–3 h postinfection (PI) in MDBK cells; however, between 3 and 6 h PI the PCD re- sponse was found to be decreased. It was interesting to see that BHV-1 inhibited staurosporin-induced PCD after 1 h. These results showed similarities with those obtained from herpes simplex type 1 infections in human epithelial cells. PCD re- sponse decreased 1 h following caspase-3 inhibitor applications, whereas NO re- sponse increased 3 h following infection in the presence of caspase-8 and -9 in- hibitory peptides. In conclusion, BHV-1 inhibited the staurosporin-induced apoptotic response and also the NO response. We propose that this inhibition is caspase-3 dependent. Key words: Bovine herpesvirus 1, nitric oxide (NO), programmed cell death Bovine herpesvirus 1 (BHV-1), a member of the Alphaherpesvirinae, is the aetiological agent of infectious bovine rhinotracheitis, infectious pustular vulvovaginitis, abortion, and fatal neonatal systemic infections as described ear- lier (Roizman et al., 1992; Tikoo et al., 1995). In addition to initial respiratory in- fections, BHV-1 may predispose infected animals, possibly through immunosup- * Corresponding author: Huseyin Baskin, MD, PhD; E-mail: [email protected]; Fax: +90 (232) 259 0541
-
Upload
independent -
Category
Documents
-
view
4 -
download
0
Transcript of Study of programmed cell death in bovine herpesvirus 1 infected mdbk cells and the possible role of...

Acta Veterinaria Hungarica 52 (3), pp. 287–297 (2004)
0236-6290/$ 20.00 © 2004 Akadémiai Kiadó, Budapest
STUDY OF PROGRAMMED CELL DEATHIN BOVINE HERPESVIRUS 1 INFECTED MDBK CELLS ANDTHE POSSIBLE ROLE OF NITRIC OXIDE IN THIS PROCESS
Z. YAZICI1, Yasemin BASKIN2, H. BASKIN3*, Ozlem GECER4, I. H. BAHAR3
and A. ÖZKUL5
1Department of Virology, Faculty of Veterinary Medicine, Ondokuz Mayis University,Kurupelit, 55139, Samsun, Turkey; 2Oncology Institute and 3Department of Microbiol-ogy and Clinical Microbiology, School of Medicine, Dokuz Eylul University, Inciralti,
Izmir, Turkey; 4Department of Virology, Regional Institute of Hygiene, Ministry ofHealth, Göztepe, 35350 Izmir, Turkey; 5Department of Virology, Faculty of Veterinary
Medicine, Ankara University, Diskapi, 06110, Ankara, Turkey
(Received September 6, 2003; accepted March 2, 2004)
Bovine herpesvirus 1 (BHV-1) is the aetiological agent of many diseasetypes and may predispose infected animals, possibly through immunosuppression,to secondary bacterial infections. Immunosuppression may directly be associatedwith the induction of programmed cell death (PCD) in some virus-infected cells.Nitric oxide (NO) has an important mediating role against fungal, bacterial, pro-tozoal, viral pathogens and tumours. BHV-1 induced apoptosis between 0.5–3 hpostinfection (PI) in MDBK cells; however, between 3 and 6 h PI the PCD re-sponse was found to be decreased. It was interesting to see that BHV-1 inhibitedstaurosporin-induced PCD after 1 h. These results showed similarities with thoseobtained from herpes simplex type 1 infections in human epithelial cells. PCD re-sponse decreased 1 h following caspase-3 inhibitor applications, whereas NO re-sponse increased 3 h following infection in the presence of caspase-8 and -9 in-hibitory peptides. In conclusion, BHV-1 inhibited the staurosporin-inducedapoptotic response and also the NO response. We propose that this inhibition iscaspase-3 dependent.
Key words: Bovine herpesvirus 1, nitric oxide (NO), programmed celldeath
Bovine herpesvirus 1 (BHV-1), a member of the Alphaherpesvirinae, isthe aetiological agent of infectious bovine rhinotracheitis, infectious pustularvulvovaginitis, abortion, and fatal neonatal systemic infections as described ear-lier (Roizman et al., 1992; Tikoo et al., 1995). In addition to initial respiratory in-fections, BHV-1 may predispose infected animals, possibly through immunosup-
*Corresponding author: Huseyin Baskin, MD, PhD; E-mail: [email protected];Fax: +90 (232) 259 0541

288 YAZICI et al.
Acta Veterinaria Hungarica 52, 2004
pression, to secondary bacterial infections (Filion et al., 1983; Bielefeldt andBabiuk, 1985).
In a variety of acute viral infections, immunosuppression may directly beassociated with the induction of programmed cell death (PCD) (apoptosis) in in-fected cells (Razvi and Welsh, 1995). It is known that BHV-1 induces PCD inmany cell types including peripheral blood mononuclear cells (Hanon et al.,1998a), activated CD4 lymphocytes (Eskra and Splitter, 1997), and Madin-Darby bovine kidney (MDBK) cells (Hanon et al., 1998b).
Nitric oxide (NO) is a free radical gaseous molecule that is produced inmany cell types from molecular oxygen and guanidino nitrogen from L-arginine,which is converted to L-citrulline (Moncada et al., 1991; Nathan, 1992; Bredtand Snyder, 1994). It has also been shown that NO has an important mediatingrole against fungal, bacterial, protozoal, viral pathogens and tumours (Hibbs etal., 1987; Green et al., 1990; Adams et al., 1991; Alspaugh and Granger, 1991;Croen, 1993; Boockvar et al., 1994; Baskin et al., 1997).
In the acute phase of viral infections, a possible relation between nitricoxide response and PCD has been studied in many viruses including bovine viraldiarrhoea virus (Adler et al., 1997), poliovirus (Lopez-Guerrero et al., 2000), ra-bies virus (Ubol et al., 2001), Ebola virus (Hensley et al., 2002), and Dengue vi-rus (Lin et al., 2002). In different cell types, viruses have shown distinct relationsbetween NO and PCD.
The aim of this work is the study of the PCD process in the acute phase ofBHV-1 infection in cultured epithelial cells and, subsequently, to investigate itsrelation with a possible nitric oxide responses in MDBK cells infected with BHV-1. MDBK cells were used in the study as a model because of their epithelial origin.This might be used as a model for BHV-1 infected epithelial cells in bovines.
Materials and methods
Cell line
Madin-Darby Bovine Kidney (MDBK) cells were cultivated in Dulbecco’sMinimal Essential Medium (DMEM, Sigma GmbH, Germany) supplementedwith 10% fetal calf serum (FCS), 100 U/mL of penicillin, 100 µg/mL of strepto-mycin, at 37 °C in CO2 incubator.
Virus strain
The Cooper strain of BHV-1 (ATCC Cat. No.: VR-864) was used in thestudy. The virus was propagated and titrated in MDBK cells under the mainte-nance medium containing 2% FCS and incubated at 37 °C in a CO2 incubator(5% CO2 atmosphere). Cells were monitored by daily microscopic examinationuntil the cytopathic effect (cpe) approached 100%, then infected cells were fro-

STUDY OF PROGRAMMED CELL DEATH IN BHV-1 INFECTED MDBK CELLS 289
Acta Veterinaria Hungarica 52, 2004
zen and thawed twice and aliquots of 1 mL amount were stored at –80 °C forfurther use. Infectivity of the virus was determined using the microtitre tech-nique. For this purpose, serial tenfold dilutions of virus were prepared and 100 µlof each dilution was placed into 4 wells of a 96-well tissue culture plate (Greiner,Germany). Subsequently, 50 µL of a MDBK cell suspension (300,000 cells/mL)was added to the virus dilutions. The plates were then incubated in humidifiedand CO2-containing (5%) atmosphere at 37 °C. The viral titre was calculated onthe basis of cpe readings as described by Reed and Muench (Reed and Muench,1938) as 50% tissue culture infective dose (TCID50)/0.1 µL.
Experimental design
In the first stage, the morphological (with and without staurosporin) andbiochemical changes caused by virus-induced PCD in MDBK cells were investi-gated. Morphological assessment of PCD was performed using Hoechst 33342nuclear staining and fluorescence microscopic evaluation.
In the second phase of the study, the induction of PCD with staurosporin(SS) (alone or with BHV-1 addition) and apoptotic route of BHV-1 infections(with/without SS) were analysed by applying caspase-1, -3, -8 and -9 inhibitors(R&D, Germany).
BHV-1 infection of MDBK cells
MDBK cells were grown in 6-well tissue culture plates (Costar) at a105 cells/mL starting density for 24 h. Following cell confluence, BHV-1 was in-oculated onto MDBK cells with infectivity of TCID50/0.1 mL, 1:1 multiplicity ofinfection (m.o.i). Staurosporin (Sigma GmbH, Germany), diluted in MEM, wasadded to MDBK cells at a final concentration of 1 µM/mL for 24 h. After the 24-h SS incubations, the cells were infected and the culture medium was poured offfor nitrite determinations 30 min, 1 h, 3 h and 6 h after the infection with andwithout SS, and the cells were incubated with Hoechst 33342 dye (Sigma GmbH,Germany; 5 µg/mL in DMEM) for 15 min at 37 °C. Following nuclear staining,alterations in the cell morphology were visualised using a fluorescence micro-scope with an excitation wavelength of 330 nm and a barrier filter of 420 nm.Cell morphologies were scored as apoptotic cells had blue stained nuclei withmultiple bright specks of condensed chromatin. At least 200 cells for each treat-ment were counted and indexed (Lee and Schacter, 1997).
Assays for caspase inhibitions
For inhibition of caspase-dependent apoptosis, the caspase-1 inhibitorypeptide N-acetyl-Tyr-Val-Ala-Asp-aldehyde (WEHO-fmk), the caspase-3 in-hibitory peptide Z-Asp-Glu-Val-Asp-fluoromethyl ketone (Z-DEVD-fmk), thecaspase-8 inhibitory peptide (IEID-fmk), and the caspase-9 inhibitory peptide

290 YAZICI et al.
Acta Veterinaria Hungarica 52, 2004
(LEHD-fmk) (R&D, Germany) were used. MDBK cells were seeded in 24-wellplates at a density of 105 cells/mL. The cells were treated with caspase-1, -3, -8and -9 inhibitors at a final concentration of 50 µM per mL for 2 h before induc-tion and then cells were infected with BHV-1 at TCID50/0.1 mL for various times(0.5, 1, 3, 6 h). Subsequently, MEM containing SS was added to the cells (finalconcentration of 1 µM/mL) and the cultures were maintained at 37 °C, 5% CO2for 24 h. Following this process, the DNA-binding dye Hoechst 33342 stainingmethod was applied to cells as described above.
Nitric oxide determinations
Nitrite is generated by the rapid oxidation of nitric oxide (NO). To assaynitrite, we used a modification of a previously published method (Green et al.,1982). Aliquots of 100 µL samples were mixed with 100 µL of equal volumes ofGriess reagent mixture (A: naphthylethylene-diamine dihydrochloride, 0.1%,B: sulphanilamide in o-phosphoric acid, 1%) in a 96-well microtitre plate (Max-isorb Immunoplate, NUNC). After 10 min of incubation at room temperature, theabsorbance was measured at 540 nm in a microplate reader (Reader Model 230S;Organon Technica, Microwell System, Holland). A range of 2-fold dilutions ofsodium nitrite (0–128 µM) in PBS was run in each assay to generate a standardcurve.
Results
Virus infectivity in MDBK cells
BHV-1 grew rapidly in MDBK cells with its characteristic cpe starting at6 h PI with syncytium formation, rounding and ballooning of the infected cellsand terminated by cellular lysis at day 3 PI. The virus infectivity was determinedto be TCID50 10–6/0.1 mL (1:1 m.o.i) at 72 h PI (Fig. 1).
Morphological changes of apoptosis
Characteristic morphological changes of apoptosis were observed in BHV-1infected cells between 0.5–3 h PI (Fig. 2). We examined the morphologicalchanges of infected cells with DNA-binding dye Hoechst 33342 and determinedapoptotic cell percentages from 1.5 to 4% between 0.5–3 hours PI. Apoptoticcells decreased to 1% following 3 h PI. These results indicated that apoptosiswas slightly induced by BHV-1 during infection.
The effect of SS on the induction of apoptosis was also examined in BHV-1infected and uninfected MDBK cells. After 24 h of SS treatment, the percentagesof apoptotic cells were 25, 10.2, 2.5 and 2.5% in BHV-1 infected MDBK cells 0.5,1, 3 and 6 h PI, respectively (Fig. 2). At the same time, after 24 + 6 h of SS treat-ment, the ratio of apoptotic cells was 80% in uninfected cells. When uninfected

STUDY OF PROGRAMMED CELL DEATH IN BHV-1 INFECTED MDBK CELLS 291
Acta Veterinaria Hungarica 52, 2004
MDBK cells were observed in the absence of SS, the percentages of the apoptoticcells were 1–2% (Fig. 2). Nuclear fragmentation and chromatin condensation wereobserved in all experiments by microscopic examinations (Fig. 3). These resultsindicated that BHV-1 induced apoptosis weakly in MDBK cells 0.5–3 h PI, but 3 hafter infection the number of apoptotic cells was reduced. When apoptosis was in-duced with SS, the apoptotic cell percentage increased by 0.5 h PI, then it was de-creased by the virus. In both situations the virus inhibited apoptosis 3 h PI.
0
1
2
3
4
5
6
7
12 24 36 48 60 72Hours postinoculation
Log
TCID
50
0
10
20
30
40
50
60
70
80
90
100
Prog
ress
of c
ellu
lar d
amag
emm
Titre CPE (%)
Fig. 1. Growth kinetics of bovine herpesvirus 1 (BHV-1) in MDBK cells in terms of percentage ofcytopathic effect production and infectivity titre
To investigate the apoptotic route of BHV-1 infection, caspase-1, caspase-3, caspase-8 and caspase-9 inhibitory peptides were used. We observed that allinhibitors reduced the number of apoptotic cells at 3 h after BHV-1 infection ofMDBK cells (Fig. 4).
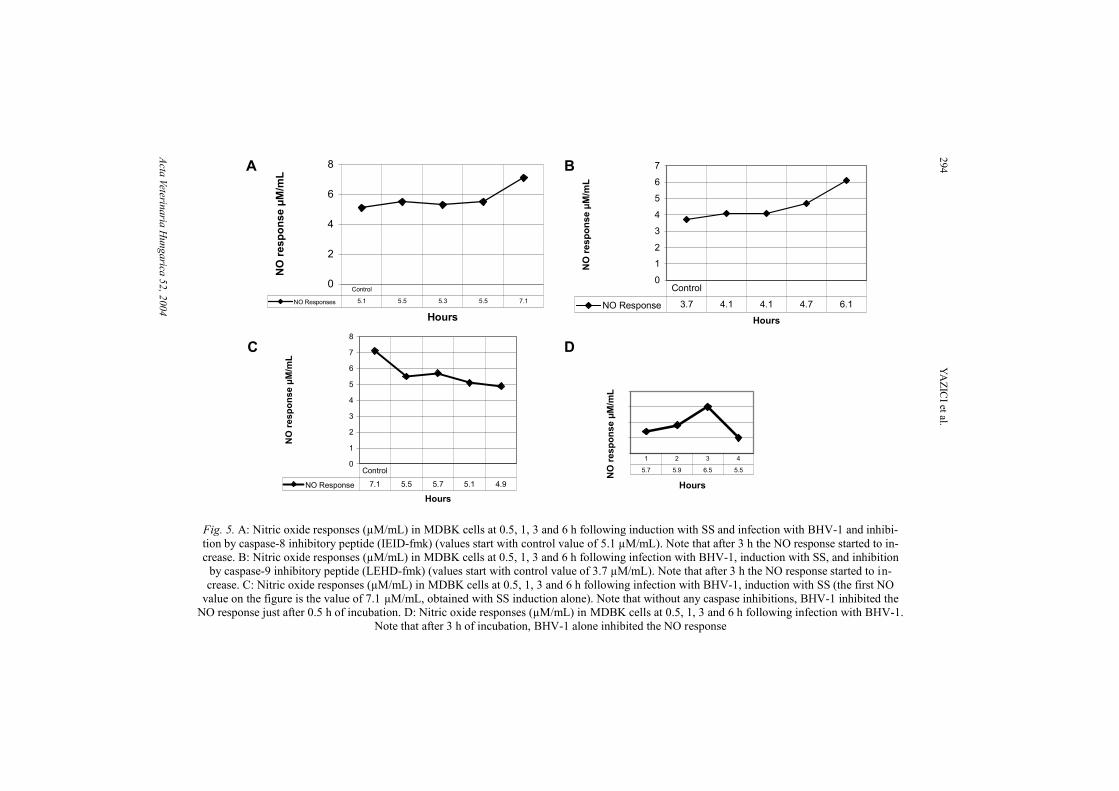
To determine the possible relation between apoptosis and NO, we haveanalysed the nitrite content of all culture supernatants after pouring off them formorphological assessment of cells for PCD. The results presented in Fig. 5Ashowed that by the inhibition of the caspase-8 pathway, NO response increased3 h PI. By inhibiting the caspase-9 pathway, the NO response also increased 3 hPI (Fig. 5B). Instead of being induced by SS at the starting point, without anycaspase inhibitions, BHV-1 inhibited the NO response after 0.5 h of incubation(Fig. 5C). Similarly, without SS induction, BHV-1 inhibited the NO responsecontinuously from the starting point up to 6 h PI (Fig. 5D).

292 YAZICI et al.
Acta Veterinaria Hungarica 52, 2004
0
10
20
30
40
50
60
70
80
90
0 0.5 1 1.5 2 2.5 3 3.5 4 4.5 5 5.5 6
Postinfection (PI) time (hours)
Apo
ptot
ic c
ells
, %
BHV-1(+), ss(+)BHV-1(+), ss(-)BHV-1(-), ss(-)BHV-1(-), ss(+)
Fig. 2. Antiapoptotic effect of BHV-1 in staurosporin- (SS-) induced MDBK cells. MDBK cellmonolayers were infected with BHV-1 at 0.5, 1, 3, 6 hours in different groups and SS was added toBHV-1 infected cultures at a final concentration of 1 µM/mL and incubated for 24 h at 37 °C, 5%
CO2. Cells on coverslips were then fixed and stained with Hoechst 33342 and the apoptotic indexeswere determined using fluorescence microscopy. Note that following 1 h of incubation BHV-1 inhib-
ited apoptotic response while SS reached its uppermost values after 3 h of incubation
0
5
10
15
20
25
30
0 1 2 3 4 5 6 7Time PI (h)
Apo
ptot
ic c
ells
(%)
Caspase-1Caspase-3Caspase-8Caspase-9BHV - 1
Fig. 4. Caspase-specific inhibitory peptides inhibited SS-induced apoptosis slightly further than BHV-1 alone 0.5 h postinfection (PI). Subsequently, caspase-8 and -9 specific inhibitory peptides did not
inhibit SS-induced apoptosis any further than did BHV-1 alone. However, in this experimental set-up,caspase-1 and -3 inhibitory peptides appear to interfere with the inhibition of apoptosis by BHV-1
starting 1 h PI
Apo
ptot
ic c
ells
(%)
Apo
ptot
ic c
ells
(%)

Acta Veterinaria Hungarica 52, 2004
STUD
Y O
F PRO
GR
AM
MED
CELL D
EATH IN
BH
V-1 IN
FECTED
MD
BK
CELLS
293
Fig. 3. Morphological changes in BHV-1 infected cells induced by staurosporin (SS). A: Control cells (BHV-1: negative; SS: negative),B: SS induction for 24 h (BHV-1: negative), C: 0.5 h PI (SS: negative), D: 0.5 h PI (SS: positive). (The square shows nuclear packaging,
the arrow shows micronucleus forming, and the circle shows nuclear condensation, all with a magnification of × 20)(Olympus BHZ-RFCA microscope with a C-35AD-4 photograph attachment)
A B
C D
Acta Veterinaria Hungarica 52, 2004
294YA
ZICI et al.
0
2
4
6
8
Hours
NO
resp
onse
µM
/mL
NO Responses 5.1 5.5 5.3 5.5 7.1
Control0
1
2
3
4
5
6
7
Hours
NO
resp
onse
µM
/mL
NO Response 3.7 4.1 4.1 4.7 6.1
Control
0
1
2
3
4
5
6
7
8
Hours
NO
resp
onse
µM
/mL
NO Response 7.1 5.5 5.7 5.1 4.9
Control
Hours
NO
resp
onse
µM
/mL
5.7 5.9 6.5 5.5
1 2 3 4
Fig. 5. A: Nitric oxide responses (µM/mL) in MDBK cells at 0.5, 1, 3 and 6 h following induction with SS and infection with BHV-1 and inhibi-tion by caspase-8 inhibitory peptide (IEID-fmk) (values start with control value of 5.1 µM/mL). Note that after 3 h the NO response started to in-crease. B: Nitric oxide responses (µM/mL) in MDBK cells at 0.5, 1, 3 and 6 h following infection with BHV-1, induction with SS, and inhibition
by caspase-9 inhibitory peptide (LEHD-fmk) (values start with control value of 3.7 µM/mL). Note that after 3 h the NO response started to in-crease. C: Nitric oxide responses (µM/mL) in MDBK cells at 0.5, 1, 3 and 6 h following infection with BHV-1, induction with SS (the first NO
value on the figure is the value of 7.1 µM/mL, obtained with SS induction alone). Note that without any caspase inhibitions, BHV-1 inhibited theNO response just after 0.5 h of incubation. D: Nitric oxide responses (µM/mL) in MDBK cells at 0.5, 1, 3 and 6 h following infection with BHV-1.
Note that after 3 h of incubation, BHV-1 alone inhibited the NO response
A B
DC

STUDY OF PROGRAMMED CELL DEATH IN BHV-1 INFECTED MDBK CELLS 295
Acta Veterinaria Hungarica 52, 2004
Discussion
The lytic cycle of BHV-1 infection shows three consecutive phases termedimmediate-early, early and late (Wirth et al., 1989). There is a very rapid pene-tration and viral replication following interaction of the virus with the appropri-ate host cell receptors. The presence of viral antigen on the surface of infectedcells becomes evident within 3–4 h PI. Complete assembly and extracellularspread start within 7–8 h PI (Ludwig and Letchworth, 1987).
Viruses have progressed strategies to induce or prevent apoptosis in hostcells. In general, virus-induced apoptosis in the acute phase is believed to en-hance the release of virus progeny from infected cells. Therefore, viral inhibitionof apoptosis may help to prolong the infection (Tschopp et al., 1998). Our resultssupport all these data: shortly after penetration into the cell, BHV-1 showed itscytopathic effect (cpe) and it was prolonged to 72 h of infection in an increasedmanner (Fig. 1).
Studying the probable apoptosis in cells infected with BHV-1, it was in-teresting to see that BHV-1 inhibited SS-induced apoptosis in MDBK cells 1 hPI. Staurosporin is a protein kinase inhibitor, which acts through a caspase-3pathway (Jacobsen et al., 1996). In our experiments, SS itself induced apoptosisin uninfected MDBK cells (Fig. 2).
In this study, it was shown that BHV-1 slightly induced apoptosis alone0.5–3 h PI in bovine epithelial cells; however, 3 and 6 h PI, the apoptotic re-sponse was found to be decreased. These results showed similarities with thoseobtained from herpes simplex type 1 (HSV-1) infections in human epithelial cells(Aubert and Blaho, 2001). Like HSV-1, BHV-1 slightly induced apoptosis justfollowing penetration into epithelial cells (0.5–3 h) and then inhibited the apop-totic process (3–6 h), perhaps for the prevention of PCD. Moreover, it was inter-esting to see that in that period BHV-1 also inhibited the NO response between 3and 6 h PI (Fig. 5D).
For studying the modulation of apoptosis following BHV-1 infection,caspase inhibitory peptides were applied in our experiments. Apoptotic responsesdecreased following application of caspase-1, -3, -8 and -9 inhibitors after 1 h asexpected, whereas NO responses increased 3 h PI with the application of inhibi-tory peptides caspase-1, -3, -8 and -9 (Figs 5A and 5B).
In conclusion, inhibition of caspase-3 did not inhibit apoptosis any furtherthan did BHV-1 alone, which suggested that BHV-1 and caspase-3 might act viathe same pathway: BHV-1 inhibited the apoptotic response in a caspase-3-dependent way (Fig. 4). The kinetic part of the experiments with initiator and ef-fector caspases showed that the virus inhibited apoptosis whereas caspase in-hibitors 8 and 9 did not affect the apoptotic response. On the other hand, BHV-1inhibited the nitric oxide responses when given alone or together with SS,whereas NO responses increased 3 h PI when caspase inhibitors 8 and 9 were

296 YAZICI et al.
Acta Veterinaria Hungarica 52, 2004
given. The results obtained from this study also suggest that BHV-1 may modu-late both the NO response and apoptosis in the same way through the caspase-3pathway. This interesting relationship deserves further study in explaining thepathogenesis of BHV-1 infection in epithelial cells.
Acknowledgement
We thank Christopher Embleton for language editing.
References
Adams, L. B., Franzblau, S. G., Vavrin, Z., Hibbs, J. B. Jr. and Krahenbuhl, J. L. (1991): L-arginine-dependent macrophage effector functions inhibit metabolic activity of Mycobacte-rium leprae. J. Immunol. 147, 1642–1646.
Adler, A., Adler, H., Pfister, H., Jungi, W. T. and Peterhans, E. (1997): Macrophages infected withcytopathic bovine viral diarrhoea virus release a factor(s) capable of priming uninfectedmacrophages for activation-induced apoptosis. J. Virol. 71, 3255–3258.
Alspaugh, J. A. and Granger, D. L. (1991): Inhibition of Cryptococcus neoformans replication bynitrogen oxides supports the role of these molecules as effectors of macrophage-mediatedcytostasis. Infect. Immun. 59, 2291–2296.
Aubert, M. and Blaho, A. J. (2001): Modulation of apoptosis during herpes simplex virus infectionin human cells. Microbes Infect. 3, 859–866.
Baskin, H., Ellerman-Eriksen, S., Lovmand, J. and Mogensen, C. S. (1997): Herpes simplex virustype 2 synergizes with interferon-gamma in the induction of nitric oxide production inmouse macrophages through autocrine secretion of tumour necrosis factor-alpha. J. Gen.Virol. 78, 195–203.
Bielefeldt, O. H. and Babiuk, L. A. (1985): Viral-bacterial pneumonia in calves: Effect of bovineherpesvirus on immunologic functions. J. Infect. Dis. 151, 937–947.
Boockvar, K. S., Granger, D. L., Poston, R. M., Maybodi, M., Washington, M. K, Hibbs, J. B. Jr.and Kurlander, R. L. (1994): Nitric oxide produced during murine listeriosis is protective.Infect. Immun. 62, 1089–1100.
Bredt, D. S. and Snyder, S. H. (1994): Nitric oxide: A physiologic messenger molecule. Ann. Rev.Biochem. 63, 175–179.
Croen, K. D. (1993): Evidence for an antiviral effect of nitric oxide. Inhibition of herpes simplexvirus type 1 replication. J. Clin. Invest. 91, 2446–2452.
Eskra, L. and Splitter, A. G. (1997): Bovine herpesvirus 1 infects activated CD4 lymphocytes. J.Gen. Virol. 78, 2159–2166.
Filion, L. G., McGuire, R. L. and Babiuk, L. A. (1983): Non specific suppressive effect of bovineherpesvirus type 1 on bovine leukocyte functions. Infect. Immun. 42, 106–112.
Green, S. J., Crawford, R. M., Hockmeyer, J. T., Meltzer, M. S. and Nacy, C. A. (1990): Leishma-nia major amastigotes initiate the L-arginine dependent killing mechanism of IFN-gamma-stimulated macrophages by induction of tumour necrosis factor-alpha. J. Immunol. 145,4290–4297.
Green, L. C., Wagner, D. A., Glogowski, J., Skipper, P. L., Wishnok, J. S. and Tannenbaum, S. R.(1982): Analysis of nitrate, nitrite and 15N-nitrate in biological fluids. Anal. Biochem. 126,131–138.

STUDY OF PROGRAMMED CELL DEATH IN BHV-1 INFECTED MDBK CELLS 297
Acta Veterinaria Hungarica 52, 2004
Hanon, E., Lambot, M., Hoornaret, S., Lyaku, J. and Pastoret, P. P. (1998a): Bovine herpesvirusinduced apoptosis: phenotypic characterization of susceptible peripheral blood mononu-clear cells. Arch. Virol. 143, 441–452.
Hanon, E., Meyer, G., Vanderplasschen, A., Dessy-Doize, C., Thiry, E. and Pastoret, P. P. (1998b):Attachment but not penetration of bovine herpesvirus 1 is necessary to induce apoptosis intarget cells. J. Virol. 72, 7638–7641.
Hensley, E. L., Young, H. A., Jahrling, P. B. and Geisbert, T. W. (2002): Proinflammatory re-sponse during Ebola virus infection of primate models: possible involvement of the tumournecrosis factor receptor superfamily. Immunol. Letters 80, 169–179.
Hibbs, J. B. Jr., Taintor, R. R. and Vavrin, Z. (1987): Macrophage cytotoxicity: role for L-argininedeiminase and iminonitrogen oxidation to nitrite. Science 235, 473–476.
Jacobsen, M. D., Weil, M. and Raff, M. C. (1996): Role of Ced-3/ICE family proteases in stauro-sporin-induced programmed cell death. J. Cell Biol. 133, 1041–1051.
Lee, Y. and Schacter, E. (1997): Bcl-2 does not protect Burkitt’s lymphoma cells from oxidant-induced cell death. Blood 89, 4480–4492.
Lin, C. F., Lei, H. Y., Shiau, A. L., Liu, H. S., Yeh, T. M., Chen, S. H., Liu, C. C., Chiu, S. C. andLin, Y. S. (2002): Endothelial cell apoptosis induced by antibodies against Dengue virusnon-structural protein 1 via production of nitric oxide. J. Immunol. 169, 657–664.
Lopez-Guerrero, A. J., Alonso, M., Martin-Belmonte, F. and Carrasco, L. (2000): Poliovirus in-duces apoptosis in the human U937 promonocytic cell line. Virology 272, 250–256.
Ludwig, G. V. and Letchworth, G. J. (1987): Temporal control of bovine herpesvirus 1 glycopro-tein synthesis. J. Virol. 61, 3292–3298.
Moncada, S., Palmer, R. M. J. and Higgs, E. A. (1991): Nitric oxide: physiology, pathophysiology,and pharmacology. Pharmacol. Rev. 43, 109–142.
Nathan, C. (1992): Nitric oxide as a secretory product of mammalian cells. FASEB J. 6, 3051–3064.Razvi, E. S. and Welsh, R. M. (1995): Apoptosis in viral infections. Adv. Viral Res. 45, 1–60.Reed, L. J. and Muench, H. (1938): A simple method of estimating fifty percent endpoints. Am. J.
Hyg. 27, 493–497.Roizman, B., Desrosiers, R. C., Fleckenstein, B., Lopez, C., Minson, A. C. and Studdert, M. J.
(1992): The family of herpesviridae: an update. Arch. Virol. 123, 425–449.Tikoo, S. K., Campos, M. and Babiuk, L. A. (1995): Bovine herpesvirus 1 (BHV-1): biology,
pathogenesis, and control. Adv. Virus Res. 45, 191–223.Tschopp, J., Thome, M., Hofman, K. and Meinl, E. (1998): The fight of viruses against apoptosis.
Curr. Opin. Genet. Dev. 8, 82–87.Ubol, S., Sukwattanapan, C. and Maneerat, Y. (2001): Inducible nitric oxide synthase inhibition
delays death of rabies virus-infected mice. J. Med. Microbiol. 50, 238–242.Wirth, U. V., Gunkel, K. and Engels, M. S. (1989): Spatial and temporal distribution of bovine
herpesvirus 1 transcripts. J. Virol. 63, 4882–4889.




















