
Studies on reactivated ciliaII. Reactivation of ciliated cortices from the oviduct of anolis...
10
Printed in Sweden Copyright ~ 1977by Academic Press, Inc. All rights of reproduction in anyform reserved ISSN 0014-4827 Experimental Cell Research 108 (1977) 311-320 STUDIES ON REACTIVATED CILIA II. Reactivation of Ciliated Cortices from the Oviduct of Anolis Cristatellus L. D. TORRES, F. L. RENAUD and C. PORTOCARRERO Department of Biology, University of Puerto Rico, Rio Piedras, PR 00931, USA SUMMARY Reactivated ciliated cortices from the oviduct of the lizard Anolis cristatellus show symplectic metachrony, and resemble reactivated sea urchin sperm flagella in several characteristics, such as pH optimum and Km~, but differ in other, such as optimal ionic strength. Cilia isolated from the same source give similar results, but show a poorer reactivation. Purified ciliated cortices have adenylate kinase activity and an ATPase activity with a Km value identical with the K~, suggesting a tight coupling between ATP hydrolysis and movement in this system. The chemical fine structure of cell move- ments involving microtubules is one of the most intriguing problems in contemporary cell biology. It has been known for a long time that microtubular structures, such as cilia and flagella, axostyles and the mitotic spindle, utilize ATP as the source of energy for movement. Recently, the chemical ultrastructure of these organelles has begun to be elucidated [3, 14, 17, 18]. However, a lot of gaps still remain in our knowledge of how the macromolecules that compose these organelles interact in order to trans- late the hydrolysis of ATP into the physical displacement of their structural elements. In addition we know very little about how this movement is regulated. A useful ap- proach to the solution of these problems has been the study of reactivated models, where the cell membrane has been dis- rupted or solubilized by extraction with glycerol or with the non-ionic detergent Triton X-100 [4, 13]. This permits the re- initiation of movement by making exo- genous ATP accessible to the motile ap- paratus, and in addition allows for manipu- lation of the environment in which the reactivation occurs. The best results have been obtained with Triton X-100 [3, 13, 18]; sea urchin sperm treated with this detergent become reactivated with a movement very similar to that of living, free-swimming sperm [13]. This has permitted a correlation to be made between the chemical properties of the flagellar axoneme and the quality of the reactivation, and to extrapolate these results to the in vivo situation. However, this approach has been used only scantily for the study of ciliary movement [10, 21, 25, 26]. Cilia and flagella are nearly identi- cal in terms of ultrastructure, but in general differ in length, number and beat pattern [2]. Therefore, it is of interest to determine how similar these organelles are in their Exp Cell Res 108 (1977)
-
Upload
independent -
Category
Documents
-
view
2 -
download
0
Transcript of Studies on reactivated ciliaII. Reactivation of ciliated cortices from the oviduct of anolis...

Printed in Sweden Copyright ~ 1977 by Academic Press, Inc. All rights of reproduction in any form reserved ISSN 0014-4827
Experimental Cell Research 108 (1977) 311-320
S T U D I E S ON R E A C T I V A T E D C I L I A
II. Reactivation of Ciliated Cortices from the Oviduct
of Anolis Cristatellus
L. D. TORRES, F. L. RENAUD and C. PORTOCARRERO
Department o f Biology, University o f Puerto Rico, Rio Piedras, PR 00931, USA
SUMMARY
Reactivated ciliated cortices from the oviduct of the lizard Anolis cristatellus show symplectic metachrony, and resemble reactivated sea urchin sperm flagella in several characteristics, such as pH optimum and Km~, but differ in other, such as optimal ionic strength. Cilia isolated from the same source give similar results, but show a poorer reactivation. Purified ciliated cortices have adenylate kinase activity and an ATPase activity with a Km value identical with the K~ , suggesting a tight coupling between ATP hydrolysis and movement in this system.
The chemical fine structure of cell move- ments involving microtubules is one of the most intriguing problems in contemporary cell biology. It has been known for a long time that microtubular structures, such as cilia and flagella, axostyles and the mitotic spindle, utilize ATP as the source of energy for movement. Recently, the chemical ultrastructure of these organelles has begun to be elucidated [3, 14, 17, 18]. However, a lot of gaps still remain in our knowledge o f how the macromolecules that compose these organelles interact in order to trans- late the hydrolysis of ATP into the physical displacement of their structural elements. In addition we know very little about h o w this movement is regulated. A useful ap- proach to the solution of these problems has been the study of reactivated models, where the cell membrane has been dis- rupted or solubilized by extraction with glycerol or with the non-ionic detergent
Triton X-100 [4, 13]. This permits the re- initiation of movement by making exo- genous ATP accessible to the motile ap- paratus, and in addition allows for manipu- lation of the environment in which the reactivation occurs. The best results have been obtained with Triton X-100 [3, 13, 18]; sea urchin sperm treated with this detergent become reactivated with a movement very similar to that of living, free-swimming sperm [13]. This has permitted a correlation to be made between the chemical properties of the flagellar axoneme and the quality of the reactivation, and to extrapolate these results to the in vivo situation. However, this approach has been used only scantily for the study of ciliary movement [10, 21, 25, 26]. Cilia and flagella are nearly identi- cal in terms of ultrastructure, but in general differ in length, number and beat pattern [2]. Therefore, it is of interest to determine how similar these organelles are in their
Exp Cell Res 108 (1977)

312 Torres, Renaud and Portocarrero
response to various chemical changes. Re- naud & Ramirez [20] have studied the re- activation of ciliated Tetrahymena ghosts obtained by homogenizing the cells with Triton X-100. The movement observed in this system was not coordinated and showed great variability in the percentage of motile ghosts. We have also studied the reactivation of ciliated cortical fractions from the oviduct of the tropical lizard Ano- lis cristateUus. The movement observed in this system is coordinated and the results are highly reproducible. Therefore we have utilized it to study the effect on ciliary movement of changes in the ionic environ- ment in which the reactivation takes place. In addition, we have done preliminary measurements of some of the enzymatic properties of these fractions. A comparison is made between these results and those re- ported previously for sea urchin sperm flagella and other systems. A preliminary account of this work had been presented be- fore [25].
Isolation of cilia Oviduct cilia were isolated by a modification of the method of Gibbons [ 10]. The transected oviducts were resuspended in glycerination solution (60 % glycerol, 25 mM KC1, 0.1% 2-mercaptoethanol and 20 mM Tris- HC1, pH 8.3 at &C) and incubated in a salt-ice mixture at - 2 & C for 10 min. The cilia were then de- tached from the tissue by vigorous agitation with a Vortex mixer for 30 sec, and then incubated in the salt-ice bath for 30 additional seconds. This agitation- incubation alternation was repeated three times. The tissue slices were then removed from the preparation, and the supernatant containing the cilia was used in the reactivation experiments described in the fol- lowing section.
Reactivation of ciliated cortices and isolated cilia The preparations were reactivated by diluting 1 : 1 with reactivating solution (0.8 mM ATP, 75 mM KC1,
MATERIAL AND METHODS
Isolation of the ciliated cortices The method reported here is a modification of the procedure reported previously by Anderson [1] for the isolation of ciliated cortices from rabbit oviduct, and by Renaud & Ramirez [20] for the isolation of ciliated Tetrahymena ghosts. Adult females of the tropical lizard Anolis cristatellus was decapitated and their oviducts dissected and transferred to a small volume of Tyrode's solution in an ice bath. The tissue was then sliced longitudinally to expose the ciliated epithelium and resuspended in SET solution (0.25 M sucrose, 1.0 mM EDTA, 25 mM KCI, 0.05% Triton X-100, 0.1% 2-mercaptoethanol and 20 mM Tris-HC1, pH 8.8 at &C). The suspension was then agitated vigorously with a Vortex mixer for 30 see and incubated in an ice bath for 30 sec. This alternation of agitation and chill- ing was repeated four times, and the preparation was then centrifuged for 5 rain at 1000 g. The sediment, consisting mostly of nuclei and isolated cortices, was resuspended in SE solution (identical with the SET solution, but omitting the Triton X-100), centrifuged for 5 min at 1000 g and resuspended in fresh SE solu- tion.
Exp Cell Res 108 (1977)
Fig. 1. Cross section of the ciliated epithelium of the Anolis oviduct. Note the cilia (arrow) lining the apical surface of the cell N, nucleus, x 2 500. Fig. 2. Detached ciliated cortices from the Anolis oviduct (arrows). Note that the nuclei (N) are the main visible contaminant of the preparation, x 1600.

316 Torres, Renaud and Portocarrero
Table 2. The effect o f ADP on the reactiva- tion o f oviduct ciliated cortices
Beat % of Duration of ADP frequency reactiva- movement conc. (Hz) tion (min)
0.4 0 0 0 0.8 0.2 15 15 1.0 0.3 20 15
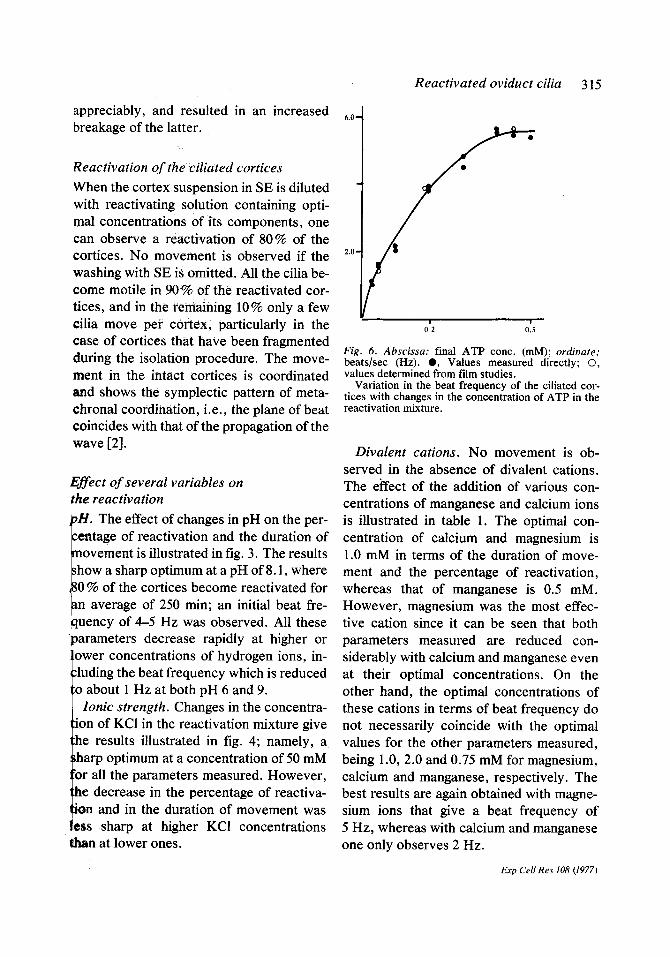
Nucleotide concentration. The minimal concentration of ATP tested that enabled the reactivation to take place was 0.025 mM. Increasing the concentration of the nu- cleotide gives a concomitant increase in the parameters measured, with an optimal con- centration of 0.5 mM (fig. 5). Further in- creases in the concentration of the nucleo- tide result in an inhibition of the reactiva- tion. The changes in beat frequency at dif- ferent ATP concentrations are illustrated in fig. 6. Again we can see that optimal results are obtained at a concentration of 0.5 mM, giving a beat frequency of 5 Hz. Higher con- centrations of ATP decreased the amplitude of the wave, and the movement observed was more vibratory than undulatory. Notice that the values obtained from film studies coincide very nicely with those obtained by visual estimation. A double reciprocal plot of the beat frequency and the ATP con- centration gives a straight line (fig. 7). Ex- trapolation of this line serves to measure a constant, the so called Kmy, which in this case has a value of 0.13 mM.
When the cortices are diluted with a re- activating solution containing ADP instead of ATP, no movement is observed if the nu- cleotide concentration is 0.4 mM or lower. If the ADP concentration is increased to 0.8 mM or higher, a reactivation is ob- served, but only after a lag period of 10 min (table 2). The type of movement is similar to that observed previously with ATP, but
Exp Cell Res 108 (1977)
the reactivation is inferior in terms of all three parameters measured. For example, the beat frequency is less than one Hz as opposed to 4-5 Hz with ATP under optimal conditions.
Reactivation of isolated cilia
The optimal reactivation of isolated cilia was obtained by mixing 1 vol of the gly- cerinated suspension with 2 vol of the reac- tivating solution. The movement observed consists of sinusoidal waves that move from one end to the other of the cilium. How- ever, roughly 10% of the cilia become at- tached to the slide by one end, and their movement is of the whiplash type, as in the cortices.
Variations in the components of the reac- tivation solution gave s o m e results tha t closely parallel those obtained for the cili- ated cortices. The optimal pH in terms of the duration of movement is 7.7, which is very close to 8.1, the optimal pH for the cortices (fig. 8). In addition, the optimal concentration of KCI is 50 mM in both cases (fig. 9). The cilia could also be reactivated with calcium and manganese, in addition to magnesium, and the optimal concentrations of these cations were very similar to the optimal ones for the reactivation of the cor- tices (L. D. Torres, unpublished results).
1.6"
0.8=
,e' J
Fig. 7. Abscissa: reciprocal values of the ATP conc.; ordinate: reciprocal values of the beat frequency.
Double reciprocal plot of the beat frequency of the ciliated cortices at different ATP concentrations, the extrapolated value of the K,~t is 0.13 raM.
Reactivated oviduct cilia 315
appreciably, and resulted in an increased breakage of the latter.
Reactivation o f the ciliated cortices
When the cortex suspension in SE is diluted with reactivating solution containing opti- mal concentrations of its components, one can observe a reactivation of 80 % of the cortices. No movement is observed if the washing with SE is omitted. All the cilia be- come motile in 90% of the reactivated cor- tices, and inthe remaining 10% only a few cilia move per cortex, particularly in the case of cortices that have been fragmented during the isolation procedure. The move- ment in the intact cortices is coordinated and shows the symplectic pattern of meta- chronal coordination, i.e., the plane of beat coincides with that of the propagation of the wave [2].
Effect o f several variables on the reactivation
pH. The effect of changes in pH on the per- centage of reactivation and the duration of novement is illustrated in fig. 3. The results ;how a sharp optimum at a pH of 8.1, where 30 % of the cortices become reactivated for m average of 250 min; an initial beat fre- quency of 4-5 Hz was observed. All these ~arameters decrease rapidly at higher or ower concentrations of hydrogen ions, in- cluding the beat frequency which is reduced :o about 1 Hz at both pH 6 and 9.
Ionic strength. Changes in the concentra- ion of KCI in the reactivation mixture give he results illustrated in fig. 4; namely, a )harp optimum at a concentration of 50 mM 'or all the parameters measured. However, he decrease in the percentage of reactiva- ion and in the duration of movement was ess sharp at higher KC1 concentrations han at lower ones.
6.0-
2.0
0~2 0'.s
Fig. 6. Abscissa: final ATP conc. (mM); ordinate: beats/sec (Hz). 0 , Values measured directly; ©, values determined from f'flm studies.
Variation in the beat frequency of the ciliated cor- tices with changes in the concentration of ATP in the reactivation mixture.
Divalent cations. No movement is ob- served in the absence of divalent cations. The eiTect of the addition of various con- centrations of manganese and calcium ions is illustrated in table 1. The optimal con- centration of calcium and magnesium is 1.0 mM in terms of the duration of move- ment and the percentage of reactivation, whereas that of manganese is 0.5 mM. However, magnesium was the most effec- tive cation since it can be seen that both parameters measured are reduced con- siderably with calcium and manganese even at their optimal concentrations. On the other hand, the optimal concentrations of these cations in terms of beat frequency do not necessarily coincide with the optimal values for the other parameters measured, being 1.0, 2.0 and 0.75 mM for magnesium, calcium and manganese, respectively. The best results are again obtained with magne- sium ions that give a beat frequency of 5 Hz, whereas with calcium and manganese one only observes 2 Hz.
Exp Cell Res 108 (1977)

314 Torres, Renaud and Portocarrero
Table 1. The effect of various divalent cations on the reactivation of oviduct cili- ated cortices
% of Duration of Conc. reactiva- movement
Cation (mM) tion (min)
Mg z+ 0.5 20 150 1.0 80 280 2.0 20 100
Ca ~+ 0.5 0 0 1.0 50 125 2.0 35 100
Mn 2+ 0.5 20 140 1.0 0 0 2.0 0 0
fuge. The nuclei sediment to the bottom of the tube, whereas the ciliated cortices can be collected at the interphase between the KEM and the sucrose solu- tions.
Enzymatic activity of the ciliated cortices The ATPase and adenylate kinase activity of the puri- fied cortices was determined by measuring the release of inorganic phosphate by the technique of Fiske & Subbarow [8]. One volume of the suspension of puri- fied cortices was mixed with 1 vol of reactivation solu- tion in a series of test tubes in a water bath at 25°C, and the reaction stopped at different time intervals by the addition of molybdate and Fiske-Subbarow reagents. The absorbance at 660 nm was measured with a Zeiss PM Q II spectrophotometer. The protein concentration was measured by the technique of Lowry et al. [16] using bovine serum albumin as a standard.
R E S U L T S
Isolation of ciliated cortices
A cross sect ion o f the ciliated epi thel ium o f
the Anolis oviduc t is i l lustrated in fig. 1. The cilia are approx. 10--15 /zm long; and
are located at the apical end o f the epithelial
cells, lining the lumen of the organ, par- t icularly in its anter ior region. W h e n the
ov iduc t slices are agi tated in the Vor t ex
mixer , the apical bo rde r o f the ciliated cell b e c o m e s de t ached as a discrete unit (fig. 2), These isolated borders will be re fer red to as cor t ices [1]. These s t ructures seem to con-
sist main ly o f the ciliary r o w s a t t ached to the basal bodies . N o ul t ras t ruc tura l s tudies
were pe r fo rmed , therefore it is no t k n o w n if they conta in ciliary root le ts and r emnan t s
o f the cell m e m b r a n e , as is the case with rabbit ov iduc t cor t ices [1]. The main visible con t aminan t s p resen t in the p repa ra t ion are
the nuclei (fig. 2), wh ich are res is tant to solubil izat ion by Tr i ton X-100. The cor tex
yield var ied depend ing on severa l fac tors . The opt imal de te rgen t concen t r a t i on was
b e t w e e n 0.02 and 0.06 %. Highe r concen t ra -
t ions caused a dis integrat ion o f the cor t ices , and lower ones r educed the n u m b e r o f cor-
tices l iberated. Omiss ion o f the chela t ing agent E D T A also r educed the co r t ex yield, and those that were p r o d u c e d seemed to be con tamina t ed with coagu la ted cy top lasm. A more p ro longed t r ea tmen t with the Vor t ex
mixer did no t increase the yield o f cor t ices
2 2 5 " ~ a
125-
i 75-
b 25
0% ~'o
Fig. 5. Abscissa: final ATP cone. (mM); ord/nate: (a) % reactivation; (b) time (rain).
The duration of movement (a) min; (b) the per, centage of reactivation of the ciliated cortices at dif ferent concentrations of ATP.
Exp Cell Res 108 (1977)

40-
6.0 9.0
Fig. 3. Abscissa: pH; ordinate: (a) % reactivation; (b) time (rain).
The effect of changes in the pH of the reactivation medium on (a) the percentage of reactivation; (b) the duration of movement of the ciliated cortices.
2.5 mM Mg SO4, 0.1% 2-mercaptoethanol and 20 mM Tris-HC1, pH 8.5). In some experiments the reacti- vating solution was altered in order to determine the optimal concentration of its components by observing the effect of concentration changes on the quality of the reactivation. The Tris-HC1 buffer was substituted by imidazole-HC1 for pH values below 7. Magnesium ions were sometimes substituted by calcium and man- :anese, and ATP by ADP. All observations were made
with a Zeiss phase-contrast microscope with Neofluar 9bjectives at a total magnification of 800×. Each ex- periment was repeated at least twice and the results were plotted tracing the best line by eye; only one ~aint is illustrated if the results for that particular ~oint coincided in duplicate experiments. The para- tinters measured were the percentage of reactivation, '.he duration of movement and the beat frequency. We l¢fine percentage of reactivation as the percentage of solated cilia or ciliated cortices showing movement in madomly selected fields. Each sample consisted of ~ rox . 50 cortices or 25-30 cilia. The end point of the
mbetivation period was arbitrarily chosen as the time which 80 % of the isolated cilia or the ciliated cor-
bees had ceased to move. The beat frequency was ~ u r e d by visual estimation, and in the case of the iliated cortices, by analysis of films of the reactivation
R e a c t i v a t e d ov iduc t cilia 3 ! 3
projected at slow motion. The movies were taken with a Beaulieu 2008-S camera, using Kodak Plus-X Super 8 film.
Pur i f i ca t ion o f c i l ia ted cor t i ces
These organelles were purified by a modification of the technique developed by Forstner et al. [9] for the puri- fication of isolated brush borders. After this proce- dure, the cortices could still be reactivated and were suitable for enzymatic analysis. The suspension con- sisting of nuclei and ciliated cortices was diluted five- fold with EM solution (2.5 mM EDTA, 0.1% 2-mer- captoethanol, pH 7.0) and then centrifuged at 1000 g for 5 min. The sediment was then resuspended in KEM solution (100 mM KCI, 0.8 mM EDTA, 0.1% 2-mer- captoethanol, pH 7.0), and incubated in an ice bath for 15 min. Under these conditions, the nuclei aggre- gate in small clumps, whereas the majority of the ciliated cortices are not affected. The suspension is then filtered through glass wool to remove the nu- clear aggregates, and the filtrate is then chased with a small volume of KEM solution. Any remaining nuclei are then removed from the preparation by layering the filtrate on top of a buffered sucrose solution (1.8 M sucrose, 0.1 M KCI, 0.1% 2-mercaptoethanol and 20 mM imidazole-HCl, pH 7.0) and centrifuging at 53000 g for 2 h, using a Beckman L3-50 Ultracentri-
8
11
b
2 0 0 a ~ ~ s I00"
50 ~00 Fig. 4. Abscissa: final KCI conc. (raM); ordinate: (a) % reactivation; (b) time (rain).
The effect of changes in the concentration of KCI in the reactivation medium on (a) the percentage of reac- tivation, and (b) the duration of movement (min).
Exp Cell Res 108 (1977)

Reactivated oviduct cilia 317
Fig. 8. Abscissa: pH; ordinate: time (min). O, Free- swimming; A, slide-attached cilia.
The duration of the reactivation of isolated oviduct cilia at different pH values.
However, the beat frequency and the dura- tion of movement is in all cases inferior to that obtained for the ciliated cortices. For example, under optimal conditions ciliated cortices will move for over 2 h, whereas free-swimming isolated cilia will do so for only 25 rain. It is of interest to note that when the cilia become attached to the slide the duration of the movement is dupli- cated (fig. 8). However, this did not result in an increased beat frequency.
Isolated cilia differ from ciliated cortices in their response to changes in the concen- tration of ATP in the reactivation medium. No movement was observed at a nucleo- tide concentration of 0.025 mM, in contrast with the ciliated cortices that are capable of movement at this concentration. Reactiva- tion was observed at ATP concentrations of 0.05 mM or higher, with optimal results at 0.2 mM. Concentrations higher than this value inhibited movement; thus, the opti- mal concentration for the reactivation of cortices inhibits the reactivation of isolated cilia.
Enzymatic activity o f the ciliated cortices
At an ATP concentration of 0.5 mM, the specific activity of the ATPase of the puri- fied cortices was 0.25/zmoles of inorganic phosphate min -1 mg -1. The results were not
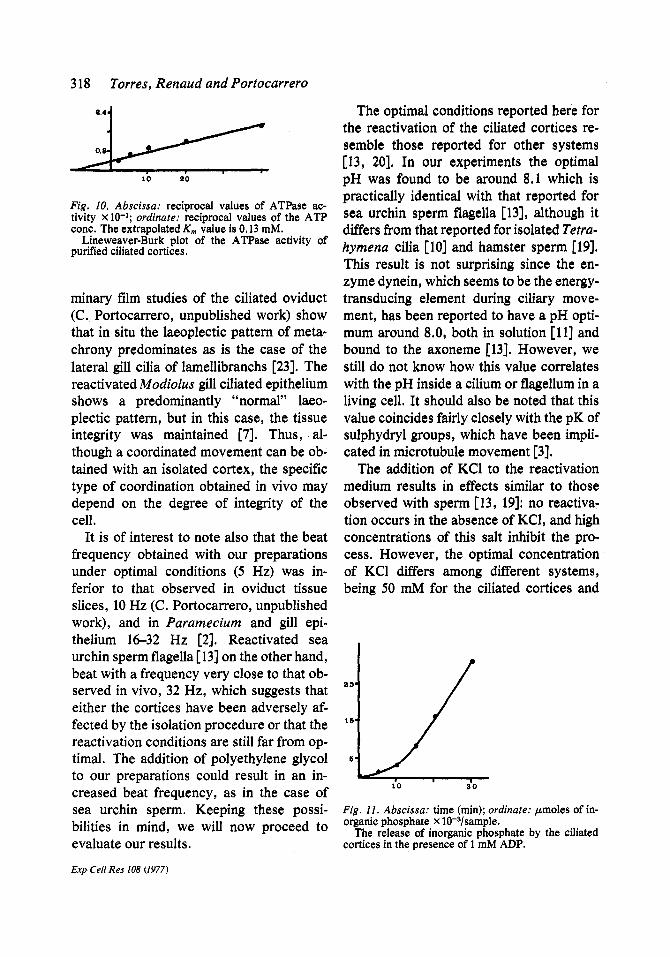
altered by the addition of various con- centrations of ouabain or oligomycin to the reactivation medium. A Lineweaver-Burk plot of the ATPase activity is illustrated in fig. 10, giving an extrapolated Km of 0.13 mM. This value coincides with that of the K,e mentioned previously.
The liberation of inorganic phosphate by the ciliated cortices in the presence of 1 mM ADP is illustrated in fig. 11. Notice that the rate is slow during the first 10 min, increas- ing dramatically afterwards. The time pe- riod of the slow initial rate coincides with the lag period before movement is observed in the cortices.
DISCUSSION
There are two things worth noticing when comparing the reactivation of ciliated cor- tices with the observations made in situ and in other systems. First of all, the fact that the movement of the cortices is coordi- nated, showing symplectic metachrony, suggests that the mechanism of coordina- tion is independent of the integrity of the cell, and is possibly due to viscous inter- actions through the medium among adjacent cilia. A similar conclusion has been reached by other workers [2], although this theory cannot explain the results obtained with the reactivated flagellar apparatus of Chlamy- domonas [5]. Observations made in preli-
2 5 "
Fig. 9. Abscissa: final KCI conc.; ordinate: time (rain). $, Free-swimming; A, slide-attached cilia.
The duration of the reactivation of isolated oviduct cilia at various concentrations of KCI.
21-771802 Exp Cell Res 108 (1977)
318 Torres, Renaud and Portocarrero
B.4
0.8
10 '~0
Fig, 10, Abscissa: reciprocal values of ATPase ac- tivity x 10-1; ordinate: reciprocal values of the ATP cone. The extrapolated Km value is 0.13 raM.
Lineweaver-Burk plot of the ATPase activity of purified ciliated cortices,
minary film studies of the ciliated oviduct (C. Portocarrero, unpublished work) show that in situ the laeoplectic pattern of meta- chrony predominates as is the case of the lateral gill cilia of lameUibranchs [23]. The reactivated Modiolus gill ciliated epithelium shows a predominantly "normal" laeo- plectic pattern, but in this case, the tissue integrity was maintained [7]. Thus, al- though a coordinated movement can be ob- tained with an isolated cortex, the specific type of coordination obtained in vivo may depend on the degree of integrity of the cell.
It is of interest to note also that the beat frequency obtained with our preparations under optimal conditions (5 Hz) was in- ferior to that observed in oviduct tissue slices, 10 Hz (C. Portocarrero, unpublished work), and in Paramecium and gill epi- thelium 16--32 Hz [2]. Reactivated sea urchin sperm flagella [13] on the other hand, beat with a frequency very close to that ob- served in vivo, 32 Hz, which suggests that either the cortices have been adversely af- fected by the isolation procedure or that the reactivation conditions are still far from op- timal. The addition of polyethylene glycol to our preparations could result in an in- creased beat frequency, as in the case of sea urchin sperm. Keeping these possi- bilities in mind, we will now proceed to evaluate our results.
Exp Cell Res 108 (1977)
The optimal conditions reported here for the reactivation of the ciliated cortices re- semble those reported for other systems [13, 20]. In our experiments the optimal pH was found to be around 8.1 which is practically identical with that reported for sea urchin sperm flagella [13], although it differs from that reported for isolated Tetra- hymena cilia [10] and hamster sperm [19]. This result is not surprising since the en- zyme dynein, which seems to be the energy- transducing element during ciliary move- ment, has been reported to have a pH opti- mum around 8.0, both in solution [11] and bound to the axoneme [13]. However, we still do not know how this value correlates with the pH inside a cilium or flagellum in a living cell. It should also be noted that this value coincides fairly closely with the pK of sulphydryl groups, which have been impli- cated in microtubule movement [3].
The addition of KC1 to the reactivation medium results in effects similar to those observed with sperm [13, 19]: no reactiva- tion occurs in the absence of KC1, and high concentrations of this salt inhibit the pro- cess. However, the optimal concentration of KCI differs among different systems, being 50 mM for the ciliated cortices and
2 5 "
1 5 "
5 "
I '0 ' 3O
Fig. 11. Abscissa: time (min); ordinate: g, moles of in- organic phosphate x 10-a/sample.
The release of inorganic phosphate by the eiliated cortices in the presence of 1 mM ADP.

Tetrahymena cilia, and 0.1-0.25 M for sperm flagella. In addition the spectrum of optimal activity is broader for sea urchin ~perm flagella (0.05-0.25 M) than for the ¢iliated cortices; for the latter the quality of the reactivation falls dramatically with slight changes in the KC1 concentration. These differences may reflect biochemical adaptations to the ionic environment in which the organisms live and suggest that although the basic structural pattern of all cilia and flagella is basically identical, there may be important chemical dif- ferences in the axonemal components of different species.
Another resemblance between the cili- ated cortices and the sea urchin sperm fla- gella is that both systems can be reactivated with magnesium, calcium, and manganese ions, the best results being obtained with magnesium. On the other hand, Tetrahy- mena cilia cannot be reactivated with cal- cium ions, and only very poorly with man- p n e s e . Thus, although ATPCa -2 and ATPMn -~ complexes are good substrates for the dynein in vitro [10], they are not as effective as ATPMg -2 in motility.
The response of ciliated cortices to varia- tions in ATP concentrations resembles that
~ sea urchin sperm flagella in that para- eters such as beat frequency are directly
proportional to ATP concentration until
elateau levels are reached and that the mplitude of the wave is reduced at the igher concentrations [13]. However, con- ntrations higher than 0.5 mM inhibit the
frequency and the duration of movement of the ciliated cortices, whereas with sea urchin sperm the frequency increases even at a concentration of 4 mM. Nevertheless, ~h¢ K,¢ of 0.13 mM obtained in our experi- ments is very close to the value of 0.2 mM reported for sea urchin sperm flagella. It is also of interest to note that the Kms value
Reactivated oviduct cilia 319
reported here coincides with the Km ob- tained for the ATPase activity of the cili- ated cortices. This would suggest that under our conditions the hydrolysis of ATP is tightly coupled to movement. The total spe- cific activity in our system is comparable to that obtained with sea urchin sperm, but we have not made any attempt to detect if there is any non-coupled ATPase activity which in the latter amounts to 30 % of the total ac- tivity [13]. The fact that the hydrolysis of ATP in our system is not altered by addition of ouabain or oligomycin suggests that most if not all the activity is due to the dynein enzyme, and not to contaminating mito- chondrial or membrane ATPases.
The reactivation of the ciliated cortices with ADP suggests that, as in other sys- tems, the enzyme adenylate kinase is pres- ent in them, and catalyses the formation of ATP from ADP. The results resemble those obtained for sperm flagella in that the re- activation is inferior to that obtained with ATP, and in that there is a lag period before movement is observed [5]. The latter may represent the time necessary for enough ADP to be converted to ATP to allow movement to occur. This correlates very well with the liberation of inorganic phos- phate during the process, which occurs very slowly initially and increases rapidly after 10 min.
The response of isolated cilia to varia- tions in the reactivation medium resembled very closely those reported for the cor- tices. However, the fact that both the beat frequency and the duration of movement are reduced when compared with the cor- tices suggests that they have been damaged by the shearing forces that were applied in order to detach them from the cortex. This hypothesis is backed by two observations. First of all the isolated cilia show a nar- rower spectrum of ATP concentration in
Exp Cell Res 108 (1977)

320 Torres, Renaud and Portocarrero
which motility can be observed when com- pared with the ciliated cortices. This sug- gests that either the dynein or the mechan- ism that couples the hydrolysis of ATP to motility has been damaged. In addition, if cilia are isolated by treatment of the tissue slices with Triton X-100 instead of glycerol, only about 20-30 % of the cilia become re- activated (C. Portocarrero, unpublished results). Since under these conditions, a more prolonged treatment with the Vortex mixer is necessary in order to detach the cilia, this suggests that this treatment is indeed deleterious for the cilia. However, it is of interest to note that under all condi- tions, if the cilia become attached to the slide the duration of movement is pro- longed, suggesting that attachment to a sub- strate is important for ciliary stability. The evidence so far indicates that ciliary move- ment occurs by the sliding of axonemal doublets [24]. Perhaps the shearing action among adjacent doublets results in a dis- organization of the axonemal structure in an isolated cilium, unless this organelle is an- chored to a substrate. In an intact cell this stabilizing role would be played by the basal body.
This work was supported by NIH grant RR-8102.
REFERENCES
1. Anderson, R G W, J cell biol 60 (1974) 393. 2. Blake, J R & Sleigh, M A, Biol rev 49 (1974) 85. 3. Bloodgood, R A, Cytobios 14 (1975) 101. 4. Brokaw, C J, Exp cell res 22 (1961) 151. 5. Brokaw, C J & Gibbons, I R, J cell sci 13 (1973) 1. 6. Child, F M, Exp cell res 8 (1961) 47. 7. Child, F M & Tamm, S, Biol bull 125 (1963) 373. 8. Fiske, C H & Subbarow, Y, J biol chem 66 (1925)
375. 9. Forstner, G G, Sabesin, S M & Isselbacher, K J,
Biochemj 106 (1968) 381. 10. Gibbons, I R, J cell biol 25 (1965) 400. 11. - - J biol chem 241 (1966) 5590. 12. Gibbons, I R & Fronk, E J, J cell biol 54 (1972)
365. Gibbons, B H & Gibbons, I R, J cell biol 54 (1972) 75.
14. Gibbons, I R, Molecules and cell movement (ed S Inou6 & R E Stephens) p. 207. Raven Press, New York (1975).
15. Hyams, J S & Borisy, G G, Science 189 (1975) 891. 16. Lowry, O H, Rosebrough, N J, Farr, A L & Ran-
dall, R J, J biol chem 193 (1951) 265. 17. Mazia, D, Petzelt, C, Williams, R O & Meza, I,
Exp cell res 70 (1972) 325. 18. Mclntosh, J R, Cande, W Z & Snyder, J A, M01e-
cules and cell movement (ed S Inou6 & R E Step-, hens) p. 31. Raven Press, New York (1975).
19. Morton, B, Exp cell res 79 (1973) 106. 20. Renaud, F L & Ramirez, T, J cell biol 63 (1974)
282a. 21. Saavedra, S & Renaud, F, Exp cell res 90 (1975)
439. 22. Satir, P & Child, F M, Biol bull 125 (1963) 390. 23. Sleigh, M A, Int rev cytol 25 (1969) 31. 24. Summers, K E & Gibbons, I R, Proc natl acad sci
US 68 (1971) 3092. 25. Torres, L D & Renaud, F L, J cell biol 67 (1975)
433a. 26. Winicur, S, J cell biol 35 (1967) c7.
Received March 16, 1977 Accepted April 14, 1977
13.
Exp Cell Res 108 (1977)




















